-
BIOTECHNOLOGICALLY RELEVANT ENZYMES AND PROTEINS
Expression of enzymes for
3′-phosphoadenosine-5′-phosphosulfate(PAPS) biosynthesis and their
preparation for PAPSsynthesis and regeneration
Payel Datta1,2 & Li Fu 1,2 & Wenqin He1,2 & M. A. G.
Koffas1,2,3 & J. S. Dordick1,2,3,4 & R. J.
Linhardt1,2,3,4
Received: 24 April 2020 /Revised: 22 May 2020 /Accepted: 31 May
2020# Springer-Verlag GmbH Germany, part of Springer Nature
2020
AbstractThe synthesis of sulfated polysaccharides involves the
sulfation of simpler polysaccharide substrates, through the
actionsulfotransferases using the cofactor,
3′-phosphoadenosine-5′-phosphosulfate (PAPS). Three enzymes are
essential for the in vitrosynthesis of PAPS, namely,
pyrophosphatase (PPA), adenosine 5′-phosphosulfate kinase (APSK),
and ATP sulfurylase (ATPS).The optimized enzyme expression ratio
and effect on PAPS synthesis were evaluated using ePathBrick, a
novel synthetic biologytool that assemble multiple genes in a
single vector. The introduction of multiple promoters and stop
codons at different locationenable the bacterial system to fine
tune expression level of the genes inserted. Recombinant vectors
expressing PPA (U39393.1),ATPS (CP021243.1), and PPA (CP047127.1)
were used for fermentations and resulted in volumetric yields of
400–1380 mg/Lwith accumulation of 34–66% in the soluble fraction.
The enzymes from soluble fraction, without any further
purification, wereused for PAPS synthesis. The PAPS was used for
the chemoenzymatic synthesis of a heparan sulfate polysaccharide
and coupledwith a PAPS-ASTIV regeneration system. ASTIV catalyzes
the regeneration of PAPS. A recombinant vector expressing
theenzymeASTIV (from Rattus norvegicus) was used for fermentations
and resulted in volumetric yield of 1153mg/L enzymewithaccumulation
of 48% in the soluble fraction. In conclusion, we have successfully
utilized a metabolic engineering approach tooptimize the overall
PAPS synthesis productivity. In addition, we have demonstrated that
the ePathBrick system could be appliedtowards study and improvement
of enzymatic synthesis conditions. In parallel, we have
successfully demonstrated anautoinduction microbial fermentation
towards the production of mammalian enzyme (ASTIV).
Key points• ePathBrick used to optimize expression levels of
enzymes.• Protocols have been used for the production of
recombinant enzymes.• High cell density fed-batch fermentations
with high yields of soluble enzymes.• Robust fermentation protocol
successfully transferred to contract manufacturing and research
facilities.
Keywords Heparin . 3′-phosphoadenosine-5′-phosphosulfate (PAPS)
. PAPS-ASTIV recycling system . Fed-batchfermentation . High cell
density autoinduction . ePathBrick platform
Introduction
Sulfation of biomolecules is ubiquitous in eukaryotic cellsand
plays a pivotal role in various biological processesinvolving
homeostasis and hormone metabolism; inactiva-tion and drug
metabolism; activation and inactivation ofmutagens and xenobiotics;
and post-translational modifica-tion of proteins, carbohydrates,
and polysaccharides(Günal et al. 2019; Hale et al. 2020; Italia et
al. 2020;Kamiyama et al. 2003; Klaassen and Boles 1997;Koprivova
and Kopriva 2016; Lin 2004; Piecewicz and
Electronic supplementary material The online version of this
article(https://doi.org/10.1007/s00253-020-10709-6) contains
supplementarymaterial, which is available to authorized users.
* M. A. G. [email protected]
* J. S. [email protected]
* R. J. [email protected]
Extended author information available on the last page of the
article
https://doi.org/10.1007/s00253-020-10709-6
http://crossmark.crossref.org/dialog/?doi=10.1007/s00253-020-10709-6&domain=pdfhttp://orcid.org/0000-0003-2219-5833https://doi.org/10.1007/s00253-020-10709-6mailto:[email protected]:[email protected]:[email protected]
-
Sengupta 2011; Xiong et al. 2014). The sulfation of
bio-molecules is catalyzed by tissue-specific
sulfotransferasesfound in the Golgi and cytosol of eukaryotes
(Coughtrie2016; Goettsch et al. 2006; Kamiyama et al. 2003).
Thecofactor, 3′-phosphoadenosine 5′-phosphosulfate (PAPS)is nearly
universally used as a sulfate donor in the sulfationof a target
biomolecule (Badri et al. 2019; Burkart et al.2000; Zhou et al.
2011). Therefore, the synthesis and re-generation of PAPS are
critical for the action ofsulfotransferases and their use in
biosynthesis. PAPS issynthesized in the cytosol, through the
sequential actionof adenosine-triphosphate (ATP) sulfurylase (ATPS,
ATPsu l f a t e adeny ly l t r ans fe ra se ) and adenos ine -5 ′
-phosphosulfate (APS) kinase (APSK, ATP
adenylylsulfate3′-phosphotransferase) (Burkart et al. 2000; Zhou et
al.2011). ATPS catalyzes the sulfation of ATP with inorganicsulfate
to yield (APS) and inorganic pyrophosphate (PPi)(Fig. 1). The APS
is then converted to PAPS by APSK inthe presence of ATP (Burkart et
al. 2000; Zhou et al.2011). The resulting PAPS is then either
utilized by cyto-solic sulfotransferases or translocated into the
Golgi(Schwarz et al. 1984) for use by
Golgi-associatedsulfotransferases (Coughtrie 2016; Goettsch et al.
2006;Kamiyama et al. 2003).
Understanding the biochemistry and activity of enzymesinvolved
in PAPS synthesis is critical for myriad applications,including the
cost-efficient chemoenzymatic synthesis of sulfat-ed
polysaccharides like HS and heparin (Badri et al. 2018;
Zhang et al. 2009). HS is important in signaling in
developmen-tal biology, physiology, and pathophysiology (Lindahl et
al.1998; Soares da Costa et al. 2017; Zhang et al. 2009).Heparin is
important pharmacologically as an anticoagulantdrug (Linhardt
2003). The biosynthesis of HS/heparin biosyn-thesis takes place in
the Golgi of mammalian mast cells andinvolves the chemical
N-deacetylation, N-sulfation ofheparosan to afford
N-sulfoheparosan, followed byepimerization of the glucuronic acid
residues to iduronic acid,catalyzed by C5-epimerase, and a series
of HS-sulfotransferase-catalyzed reactions at the 2, 6, and 3-OH
positions in the pres-ence of PAPS (Liu and Linhardt 2014; Zhang et
al. 2009). Thisapproach has been used in vitro in the
chemoenzymatic synthe-sis of HS/heparin (Fu et al. 2017). An
efficient one-potchemoenzymatic synthesis of PAPS has been reported
that in-volves APSK, ATPS, and PPA (Fig. 1a) (Burkart et al.
2000;Zhou et al. 2011). PAPS can then be used in sulfation
reactionsand subsequently recycled using aryl sulfotransferase
IV(ASTIV) (Fig. 1b) (Fu et al. 2017; Paul et al. 2012).Applications
of the ASTIV-PAPS recycling system have beenused successfully
towards production of biosynthetic heparin,and for enhancing the
anticoagulant activity of bovine intestinalheparin (Fu et al.
2017).
An effective chemoenzymatic process requires the suf-ficient
production of the three enzymes involved inPAPS synthesis as well
as ASTIV involved in PAPSrecycling. To this end, we have focused on
enzyme pro-duction using high cell density fed-batch
fermentations
Fig. 1 Enzymes in PAPSsynthesis and regenerationsystem. a PAPS
is synthesizedthrough the addition of inorganicsulfate to ATP and
the addition ofphosphate to APS (Burkart et al.2000; Zhou et al.
2011). b Oncegenerated, PAPS serves as adonor for sulfotransferases
andcan be regenerated in vitro usingp-nitrophenol sulfate (PNPS) as
asacrificial sulfate donor
-
(Tables 1 and 2). In addition, we demonstrate that theseenzymes
can be used in the chemoenzymatic synthesis ofdefined HS
polysaccharides.
Materials and methods
Materials, bacterial strains, and plasmid constructs
Chemicals for media formulation, enzyme purification, en-zyme
quantification, and PAPS production were purchasedfrom
Sigma-Aldrich (St. Louis, MO). Antibiotics and IPTG
were purchased from Gold Biotechnology (St. Louis, MO).The
bacterial strains used in the study are listed in Table 1.
High cell density non-autoinduction fed-batch fer-mentation of
PAPS synthesis enzyme
Enzyme fermentations were performed in 5-L BioFlo 320Eppendorf
or 1-L Applikon bioreactors. The fermentationsconsisted of a batch
phase, followed by a fed-batch phase.The batch media and feed
compositions for each strain areshown in Table 2. The batch phase
was initiated with theinoculation of seed culture in batch media
(starting OD600 =
Table 1 Strain, gene sequenceaccession numbers, and
plasmidinformation (see Table S1 forprimer information and
cloningstrategy for the genes ATPS,APSK and PPA)
Strain Description Source
DH5α General cloning host Invitrogen
BL21Star™ (DE3) F-ompT hsdSB (rB-, mB
-) galdcmrne131 (DE3) NovagenRosetta-gami B (DE3) F– ompT hsdSB
(rB
– mB–) gal dcm lacY1 ahpC
(DE3) gor522::Tn10 trxBpRARE (CamR, KanR, TetR)
Novagen
Gene Organism Accession numbers
APSK Penicillium chrysogenum U39393.1ATPS Kluyveromyces lactis
CP021243.1PPA Kluyveromyces lactis CP047127.1
Plasmid Description Reference
pETM6 T7 promoter, ColE1 ori. AmpR (Xu et al. 2012)
pETM6_ATP
pETM6_ATPSH
pETM6 carrying gene atps
pETM6 carrying gene atps fused with Histag in N-terminus
This study
This study
pETM6_APSK
pETM6_APSKH
pETM6 carrying gene apsk
pETM6 carrying gene apsk fused with His tag in N-terminus
This study
This study
pETM6_PPA
pETM6_PPA
pETM6 carrying gene ppa
pETM6 carrying gene ppa fused with His tag in N-terminus
This study
This study
pETM6_PSKP pETM6 carrying genes in the order of atps,apsk ppa in
pseudo-operon configuration
This study
pETM6_PSPK pETM6 carrying genes in the order of atps,ppa and
apsk in pseudo-operon configuration
This study
pETM6_PKSP pETM6 carrying genes in the order of apsk,atps, and
ppa in pseudo-operon configuration
This study
pETM6_PKPS pETM6 carrying genes in the order of apsk,ppa and
atps in pseudo-operon configuration
This study
pETM6_PPSK pETM6 carrying genes in the orderof ppa, atps and
apsk in pseudo-operonconfiguration
This study
pETM6_PPKS pETM6 carrying genes in the order of ppa,apsk and
atps in pseudo-operon configuration
This study
pCDFDuet-1_PPA pCDFDuet-1 carrying gene ppa This study
pETDuet-1_ATPS_APSK
pETDuet-1 carrying genes ATPS and APSK This study
pET28a(+)_ASTIV pET28a(+) carrying gene ASTIV fromR.
norvegicus
Gift from Pr. Jian Lu,UNC
pMalc2x_6OST3 pMalc2x carrying the catalytic domainof mouse
6OST-3 (Pro121–Pro450)
(Chen et al. 2005)
-
0.25 ± 0.1). The fermentation and bioreactor conditions
areprovided in supporting information Table S2. The fed-batchphase
was initiated when OD600 reached 3.5 ± 2.0. The cellswere fed at an
initial feed rate of 2.5 mL/h/L and the feedingrate was adjusted
based on fluctuations in pH and dissolvedoxygen (DO) levels. The
cells were induced at 22 °C at anOD600 of 6.5 ± 2.0. Overnight
induction at 22 °C was per-formed with 0.5 mM IPTG. Post-induction,
the cells wereharvested by centrifugation at 5000×g for 20 min at 4
°C.The cell paste was weighed and stored at − 80 °C.
High cell density autoinduction fed-batch fermenta-tion of ASTIV
enzyme
Enzyme fermentations were performed in 5-L BioFlo 320Eppendorf
or 1-L Applikon bioreactors. The fermentationsconsisted of a batch
phase, followed by a fed-batch phase.The batch media and feed
compositions for each strain areshown in Table 2. The batch phase
was initiated with theinoculation of seed culture in batch media
(starting OD600 =0.25 ± 0.1). The fermentation and bioreactor
conditions areprovided in supporting information Table S2. The
fed-batchphase was initiated when OD600 reached 3.5 ± 2.0 and
tem-perature was reduced to 22 °C. The cells were fed at an
initialrate of 2.5 mL/h/L and the feeding rate was adjusted, based
onfluctuations in pH and dissolved oxygen (DO) levels. Thelactose
in the feed served as the inducer and overnight induc-tion at 22 °C
was performed. Post-induction, the cells wereharvested by
centrifugation at 5000×g for 20 min at 4 °C. Thecell paste was
weighed and stored at − 80 °C.
Quantification of enzyme yield using SDS-PAGEanalysis
Enzyme concentrations were determined using semi-quantitative
SDS-PAGE analysis with bovine serum albumin
(BSA) used as a standard. Briefly, the cell pellet (5 g
wetweight) was dispersed in 25 mL of purification buffer(25 mM
Tris-HCl (Bio-Rad, USA), 500 mM NaCl (Sigma,USA), pH 7.5)
containing 8000 kU/LDNAse I (Sigma, USA),and protease inhibitors
(SIGMAFAST™ Protease InhibitorCocktail Tablets, EDTA-Free, Sigma,
USA). The cells werelysed using sonication, followed by
centrifugation at13,500×g for 40 min at 4 °C. The cell lysate
(soluble enzymefraction) was carefully transferred to a clean tube,
and the cellpellet (insoluble fraction) was dissolved in 2.5 mL
lysis buffer.Both the undiluted and the diluted fractions were
evaluatedusing SDS-PAGE performed using a MiniProtean Tetra sys-tem
(Bio-Rad, Hercules, CA). Denatured enzymes (10 μL of a1:1 (v:v)
boiled enzyme solution and sample buffer) and pro-tein ladder
(Precision Plus Protein™ Kaleidoscope™Prestained Protein Standards,
Bio-Rad) were resolved usinga 4–15% precast gel (Mini protean TGX
gels, Bio-Rad) at 220V. Gels were washed with deionized water,
stained withGelCode™ Blue Safe Protein Stain (ThermoFisher,
City,State), and destained with deionized water.
PAPS production using APSK, ATPS, and PPA
PAPS synthesis was performed using a modified protocol,based on
a previously described method (Burkart et al.2000; Zhou et al.
2011). Briefly, following fermentation, cellswere harvested by
centrifugation at 5000×g for 20min at 4 °C.The supernatant was
discarded, and the cell paste was storedin − 80 °C. On the day of
the experiment, PPA expressingcells and ATPS+APSK expressing cells
were thawed in lysisbuffer (20 mM tris(hydroxymethyl) amino
(Tris)–HCl,500 mM NaCl, pH 8.0) and lysed using a microfluidizer at
4°C. After centrifugation at 12,000×g for 20 min at 4 °C, thelysate
was filtered using a 0.45-μm filter. The filtered celllysate was
utilized for PAPS synthesis. Briefly, the cell lysate(0.3 mg/mL PPA
and 0.6 mg/ml APSK+ATPS) was added to
Table 2 Batch media and feed composition
Strain Description Composition
Recombinant E. coli BL21 Star™ (DE3)expressing ASTIV (kanamycin,
50 μg/ml)ASTIV
Autoinduction batch media Ammonium chloride (31 mM), Glycerol (7
g/L),KH2PO4 (25 mM), K2HPO4 (25 mM), lactose (2 g/L),MgSO4.7H20
(1.8 g/L), sodium chloride (5 g/L),tryptone (10 g/L), yeast extract
(12 g/L)
Autoinduction feed 40% Glycerol, supplemented with 67 g/L
NH4Cland 100 g/L lactose
Recombinant E. coli BL21 Star™ (DE3)expressing PPA (streptomycin
50 μg/ml)andecombinant E. coli BL21 Star™ (DE3)expressing ATPS and
APSK (carbenicillin, 50 μg/ml)
Non-autoinduction batch media Glycerol (7 g/L), MgSO4.7H20 (1.8
g/L),sodium chloride (5 g/L), tryptone (10 g/L),yeast extract (12
g/L)
Non-autoinduction feed 50% glycerol, supplemented with 67 g/L
NH4Cland 100 g/L yeast extract
-
a reaction mixture that contained variable concentrations ofATP
sodium (0 mM to 22.5 mM), 100 mM Na2SO4, 10 mMMgCl2, 10 mM LiCl2,
50 mM Tris-HCl (pH 8.0), and 0.02%(w/v) sodium azide. The reaction
was performed for 16 h at 30°C. Following the reaction, the
reaction mixture was centri-fuged at 12,000×g for 20 min. The
supernatant was removed,clarified, and subjected to ion exchange
chromatography,wherein PAPS was eluted with 5 mM sodium phosphate,
pH8.0, and 500 mM NaCl. PAPS was desalted using size exclu-sion
chromatography (Sephadex G-10 resin) using high-performance liquid
chromatography (HPLC) grade water asthe mobile phase. The desalted
PAPS was lyophilized andconcentrated in HPLC grade water,
aliquoted, and stored at− 80 °C. The final product was assessed for
yield, PAPSpurity, and concentration.
Analysis of PAPS by HPLC-MS
Liquid chromatography mass spectrometry (LC-MS) anal-yses were
performed on an Agilent 1200 LC/MSD instru-ment (Agilent
Technologies, Inc. Wilmington, DE). Theinstrument was equipped with
a 6300 ion-trap and a binarypump and a Poroshell 120 EC-C18 column
(2.0 × 100 mm,2.7 μm, Agilent, USA) was used for the
experiments.Eluent A contained water/acetonitrile (85:15, v/v) and
elu-ent B contained water/acetonitrile (35:65, v/v). Both elu-ents
were supplemented with 12 mM tributylamine (TrBA)and 38 mM NH4OAC.
For separation of ATP and PAPS, alinear gradient from 0 to 18 min
(0–80% of solution B) wasused at 80 μL/min. For continuous
detection by MS, thecolumn effluent entered the electrospray
ionization MSsource. The electrospray interface was set in negative
ion-ization mode with a skimmer potential of − 40.0 V, a cap-illary
exit of − 40.0 V, and a source temperature of 350 °C,to obtain the
maximum abundance of the ions in a full-scanspectrum (100–900 Da).
Nitrogen (8 L/min, 40 psi) wasused as a drying and nebulizing
gas.
Purification and immobilization of ASTIV enzymes
The cell pellet (5 g wet weight) of His6-tagged-ASTIV
wasdispersed in 25 mL of purification buffer (25 mM
Tris-HCl(Bio-Rad, USA), 500 mM NaCl (Sigma, USA), and 30
mMimidazole, pH 7.5) containing 8000 kU/L DNAse I (Sigma,USA), and
protease inhibitors (SIGMAFAST™ ProteaseInhibitor Cocktail Tablets,
EDTA-Free, Sigma, USA). Thecells were lysed using sonication,
followed by centrifugationat 13,500×g for 40 min at 4 °C. The
enzymes from the solublefraction were purified using 5 mL of Ni-NTA
Sepharose (GEHealthcare, Sweden) according to manufacturer’s
instruc-tions. The purity and MW of the enzyme was assessed
usingSDS-PAGE gel analysis.
ASTIV activity assay
The ASTIV activity assay was performed as previously de-scribed
(Paul et al. 2012) with some modification using aShimadzu
UV-1650PC. Briefly, the enzyme was purifiedusing Ni-NTA Sepharose
and quantified using nanodrop spec-troscopy at 280 nm. The activity
was determined by incubat-ing 0.1 mg of purified ASTIV with 5 mM
PNPS, 0.05 mM 3′adenosine-5′ phosphate (PAP), in phosphate-buffered
saline(pH 7.0) (15 min, 37 °C, absorbance at 400 nm). The
specificactivity was calculated using an extinction coefficient of
1.05× 10−2 M−1 cm−1.
Production, purification, and immobilization of
MBP-tagged-6OST-3 enzyme
Recombinant E. coli Rosetta-gami B (DE3) strain
(Novagen,Cambridge, MA) expressing the plasmid pMalc2x_6OST3(the
catalytic domain of mouse 6OST-3, Pro121–Pro450)was used for
production of MBP-tagged-6OST-3 (Chenet al. 2005). The
non-autoinduction fermentation protocolfor PAPS biosynthetic
enzymes was used for production ofthe MBP-tagged-6OST3 enzymes. The
cell pellet (5 g wetweight) of MBP-tagged-6OST-3 was dispersed in
25 mL ofpurification buffer (25 mM Tris-HCl (Bio-Rad, USA),500 mM
NaCl (Sigma), pH 7.5) containing 8000 kU/LDNAse I (Sigma), and
protease inhibitors (SIGMAFAST™Protease Inhibitor Cocktail Tablets,
EDTA-Free). The cellswere lysed using sonication, followed by
centrifugation at13,500×g for 40 min at 4 °C. The enzymes from the
solublefraction were purified using 5 mL of amylose resin (NEB,USA)
according to manufacturer’s instructions. The purityand MW of the
enzyme was assessed using SDS-PAGE gelanalysis.
6-O-sulfation of N-sulfo heparosan using ASTIV andPAPS recycling
system
NSH was prepared using a method previously described(Wang et al.
2011).N-sulfoheparosan (NSH) (2mg) was treat-ed with 6OST-3, using
a modified protocol, based on a previ-ously described method (Xiong
et al. 2013). The sulfationreaction was coupled with the PAPS
recycling system thatconsisted of PNPS, PAPS, and ASTIV. The
detailed reactionconditions were as follows: 0.1 mg/mL NSH, 0.5
mg/mL6OST-3, and 10 mM and 250 μM of PNPS and PAPS, re-spectively.
Reactions were incubated at 37 °C for 40 h in50 mM
2-(N-morpholino)ethanesulfonic acid buffer (pH7.2). After the
reaction was complete, the mixture containingN-sulfo, 6-O-sulfo
heparosan (NS6S) was boiled for 10 minfollowed with 5000×g
centrifugation (10 min) to spin downprecipitated free enzymes. The
supernatant was collected,
-
desalted, and freeze-dried for further analysis, including
disac-charide analysis and NMR.
Disaccharide compositional analysis
NSH and NS6S samples (200 μg each) were completelydigested using
a mixture of heparinase I, II, and III (10 mUeach) in 200 μL 50 mM
ammonium acetate buffer (pH 7.2) at37 °C for 12 h. The resulting
disaccharides were recoveredusing 10 KDa molecular weight cut-off
(MWCO) ultrafiltra-tion units (Millipore, MA) and washed with
deionized water.The ultrafiltrates containing disaccharides were
collected andfreeze-dried for disaccharide analysis.
Disaccharide analysis was carried out with high-pressure liquid
chromatography (HPLC)–ultraviolet spec-trometry (UV). The analyses
were performed with aShimadzu LC-20 AD pump, Shimadzu CBM-20A
con-troller, Shimadzu SIL-20AHT auto-sampler, ShimadzuCTO-20 AC
column oven, and a Shimadzu SPD-20AVUV detector (Shimadzu, Kyoto,
Japan). A Spherisorb-SAX chromatography column (4.0 × 250 mm, 5.0
μm,Waters) was equilibrated with 1.8 mM monobasic sodiumphosphate
(eluent A, pH 3.0) and followed with ingredi-ent elution after
injection using 1.8 mM monobasic sodi-um phosphate with sodium
perchlorate (eluent B, pH 3.0).Disaccharide standards were
purchased from Iduron(Manchester, UK). The weight/weight (w/w)
percentagefor each disaccharide was calculated. Peak areas wereused
for quantification.
NMR spectroscopy
NSH and NS6S samples (each 1.5 mg) were analyzed with1D 1H
nuclear magnetic resonance (NMR). The 1D 1H-NMR experiments were
performed on a Bruker Advance II600 MHz spectrometer (Bruker Bio
Spin, Billerica, MA)with Topsin 2.1.6 software (Bruker). Samples
were eachdissolved in 0.5 mL D2O (99.996%, Sigma-Aldrich)
andfreeze-dried repeatedly to remove the exchangeable pro-tons. The
samples were dissolved in 0.4 mL D2O and trans-ferred to NMR
microtubes (outside diameter, 5 mm, Norell(Norell, Landisville,
NJ)). All NMR experiments were car-ried out with the conditions as
previously reported. (Fuet al. 2014; Fu et al. 2016)
Data analysis
All data analysis was performed using Bio-rad Image Labsoftware,
Microsoft Excel, and GraphPad (GraphPad Prism,San Diego, CA,
USA).
Results
Expression of PAPS synthesis enzymes and PAPSePathbrick
screening platform
ATPS, APSK, and PPA are used for the in vitro synthesis ofPAPS.
The enzymes were cloned into the pETM6 vector,expressed as
His-tagged enzymes in recombinant E. coli cells.All three enzymes,
ATPS (57.3 KDa), APSK (24.6 KDa), andPPA (20.5 KDa) were expressed
as His-tagged proteins inE. coli (Fig. 2).
PAPS expression analysis was evaluated usingePathBrick platform
(He 2017; Xu et al. 2012). TheePathBricks is a versatile platform
used for rapid designand optimization of metabolic pathways in E.
coli; in thecurrent study, the ePathbrick platform has been
success-fully used as a tool to study gene expression of
PAPSsynthesis enzymes. The PAPS ePathbrick platform dem-onstrated
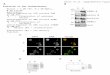
that the ratio of PPA, APSK, and ATPS impactPAPS production.
Increasing ATPS concentration nega-tively impacted PAPS production
(Fig. 3). Two ap-proaches were evaluated: (1) three genes in a
single plas-mid pETM6_PSKP used for expression and (2) threegenes
placed in a pseudo-operon configuration in differentgene orders to
optimize PAPS production (He 2017). Thisexpression system
demonstrated the feasibility of crude
Fig. 2 Shake flask experiments and SDS-PAGE gel of the
expression ofPAPS synthesis related genes. Lane 0, protein ladder;
lane 1, solublefraction of ATPS; lane 2, purified ATPS (57.3 KDa);
lane 3, solublefraction of APSK; lane 4, purified APSK (24.6 KDa);
lane 5, solublefraction of PPA; lane 6, purified PPA (20.5 KDa)
-
cell lysate in successful in vitro PAPS production(Figure S2)
and the need to balance the expression of
the three enzymes involved in vitro PAPS synthesis. ThePAPS
ePathbrick system was used to evaluate the impactof the expression
levels of these three enzymes on PAPSproduction. Optimal PAPS
production required the threegenes (PPA, APSK, ATPS) to be placed
in a pseudo-operon configuration (He 2017). The reactions were
car-ried out without supplementing ATP after cell lysis. Theoptimal
strains contained plasmid pETM6_PSPK, whereAPSK was most highly
expressed, followed by PPA, andthen ATPS (Fig. 3b). An optimal
strain should be able tocovert most of the ATP to PAPS. Based on
these results,two recombinant E. coli strains were engineered;
namely,(1) E. coli BL21 Star™ (DE3) strain expressing PPA and(2) E.
coli BL21 Star™ (DE3) strain expressing APSKand ATPS. These two
strains were used for scale-upfed-batch fermentation
experiments.
High cell density non-autoinduction fed-batch fer-mentation of
PAPS synthesis enzyme
High cell density, fed-batch fermentation of PAPS
synthesisenzymes without autoinduction may be broadly classified
intothree growth phases: (1) seed inoculation and cell growth
inbatch media; (2) initiation of feed and growth of cells at
theexponential growth rate; and (3) induction with IPTG. Twostrains
expressing the three PAPS synthesis enzymes wereexpressed using the
same fermentation protocol. The fermen-tation profiles for the two
strains are shown in Figure S3a(E. coli BL21 Star™ (DE3) strain
expressing PPA), andFigure S4a (E. coli BL21 Star™ (DE3) strain
expressingAPSK and ATPS). The yield and MW of the enzymes
fromsoluble fraction were analyzed and quantified. Enzymes
werepresent in both soluble and insoluble fractions (Figures S3band
S4b). The high cell density fed-batch fermentation of
Fig. 3 In vitro PAPS synthesis reaction with 6 different molar
ratios ofATPS, APSK, and PPA. a Pure enzyme with the same molar
ratio frompETM6 pseudo-operon configuration with 0.09mMof ATP. b
The crudelysate contained six different pETM6 plasmids with genes
in differentorder. The intensity of the protein expression is
indicated by the color:black is the highest, dark gray is the
medium, and light gray is the lowest
Table 3 Fermentation and enzyme yield (see Figure S1 for
schematic diagram and experimental plan; see Table S3 for
calculations; # semi-quantitativeSDS-PAGE analysis)
Strains ASTIV PPA ATPS and APSKFermentation strategy Fed-batch
lactose induction IPTG induction (0.5 mM) IPTG induction (0.5
mM)
Total fermentation time 23 to 27 h 26 h 24 h
Total induction time 19 h 21 h 19 h
Harvest OD600 57 40 34
Soluble enzyme yield# (mg/g cell paste) 5.75 14.45 ATPS:
4.65APSK: 6.44
Volumetric enzyme yield in soluble fraction(mg/L fermentation
broth)
555 908 ATPS: 199APSK: 276
Volumetric total enzyme yield in solubleand insoluble fraction
(mg/L fermentation broth)
1153 1383 ATPS: 579APSK: 419
% Enzyme in soluble fraction 48 % 66 % ATPS: 34%APSK: 66%
Enzyme purification and utilization method Ni-NTA resin
purification Soluble fraction Soluble fraction
-
strain expressing PPA resulted in a volumetric yield of 1380mg/L
of fermentation broth (Table 3); and 34% of the PPAenzymes
accumulated in the insoluble fraction (misfoldedproteins or
inclusion bodies) and the volumetric yield ofPPA in the soluble
fraction (correctly folded active proteins)was 910 mg/L (Table 3).
The high cell density fed-batch fer-mentation of strain expressing
both APSK andATPS varied involumetric yield and the percentage of
enzyme in the solublefractions (Figure S4b and Table 3). The high
cell density fed-batch fermentation of APSK resulted in a
volumetric yield of420 mg/L of fermentation broth (Table 3) and 34%
of theAPSK enzymes accumulated in the insoluble fraction andthe
volumetric yield of APSK in the soluble fraction was280 mg/L (Table
3). The high cell density fed-batch fermen-tation of ATPS resulted
in a volumetric yield of 580 mg/L offermentation broth (Table 3)
and 66% of the ATPS enzymesaccumulated in the insoluble fraction
and the volumetric yieldof ATPS in the soluble fraction was 200
mg/L (Table 3).Current PAPS synthesis involves the quantification
of thePAPS enzymes from the soluble fraction and directly addingthe
filtered soluble fractions (that contain the PAPS synthesisenzymes)
into the PAPS synthesis reaction mixture.
High cell density fed-batch autoinduction fermenta-tion of
ASTIV
High cell density fed-batch autoinduction fermentation ofASTIV
can be broadly classified into two growth phases: (1)seed
inoculation and cell growth in a batch media and (2)initiation of
feed and growth of cells at exponential growthrate (Fig. 4a). The
yield and MW of the enzymes from solublefraction were analyzed and
quantified. Results demonstratethe presence of ASTIV in both
soluble and insoluble fractions(Fig. 4b). The high cell density
autoinduction fed-batch fer-mentation resulted in a volumetric
ASTIV yield of 1150mg/Lof fermentation broth (Table 3). However,
approximately 52%of the enzyme accumulated in the insoluble
fraction (inactiveenzyme) and the volumetric yield of ASTIV in the
solublefraction (active enzyme) was 560 mg/L and most of the
solu-ble ASTIV was immobilized using Ni-NTA resin (Figs. 4b, 5,and
Figure S5).
Utilization of PAPS towards generation of
definedoligosaccharides, using HS-6OST3 sulfotransferaseand ASTIV
regeneration system
PAPS produced using the PAPS synthesis enzymes can
besuccessfully used in the synthesis of defined HS polysaccha-rides
in the presence of a ASTIV-coupled PAPS regenerationsystem. To this
end, the ASTIV was prepared as a His-taggedfusion protein. The HS
biosynthetic enzyme, 6OST-3 was
a
b
Fig. 4 High cell density fermentation of ASTIV and production
ofsoluble enzymes. a Fermentation was performed in 1-L
Applikonbioreactor, equipped with a BugLab cell growth probe. The
primary y-axis denotes cell growth, using arbitrary Buglab units
(black line). Thesecondary y-axis denotes pH (set at 6.9 ± 0.1,
dashed gray line),temperature (°C, gray line), and offline OD600
measurement (blackspots/dots). The spots are offline OD600
measurements (usingspectrophotometer). The OD600 is shown in
secondary Y-axis. The cellgrowth was monitored using BUGLAB) and
the units are arbitraryBUGLAB units (straight line, primary
Y-axis). The graph demonstratesthat the BUGLAB accessory could be
successfully used to assess the cellgrowth and is comparable with
the offline OD600 measurements. bFollowing fermentation, the cells
were harvested and analyzed forinsoluble and soluble enzymes. Lane
1, protein ladder; lane 2, enzymesin the undiluted insoluble
fraction; lane 3, enzymes in the undilutedsoluble fraction; lane 4,
enzymes in the 1/10 diluted insoluble fraction;lane 5, enzymes in
the 1/10 diluted soluble fraction; lane 6, 62.5 μg/mLBSA standard;
lane 7, 125μg/mLBSA standard; lane 8, 250 μg/mLBSAstandard; lane 9,
500 μg/mL BSA standard; lane 10, 1000 μg/mL BSAstandard. For each
lane, 12 uL of sample was loaded. Results demonstratepresence of
the 37 KDa ASTIV enzyme in both soluble and insolublefractions
-
prepared as a maltose binding protein (MBP) fusion.
Theseproteins were immobilized on Ni-NTA Sepharose and amy-lose
resins, respectively. NSH was treated with immobilized6OST3
overnight at 37 °C. The sulfation reaction was coupledwith PAPS
recycling system consisting of (1) ASTIV, (2)PNPS as a sacrificial
sulfo-donor, and (3) PAPS. After thereaction was complete, the
reaction mixture was filtered toremove the beads and desalted using
1 KDa MWCO centrif-ugal ultrafiltration. The purified mixture was
lyophilized forfurther disaccharide analysis and NMR analysis. The
disac-charide compositions of NSH and NSH treated with 6OST3are
shown in Table 4. After treatment with 6OST3, in pres-ence of PAPS
recycling system, NSH affords 6-O-sulfated,N-sulfoheparosan (NS6S).
The formation of NS6S was verifiedusing 1D 1H-NMR (Fig. 6).
Discussion
The scalable production of enzymes using high cell
densityfermentation generally focuses on the cost-effective
produc-tion of active biocatalysts. Upstream fermentation
parameters,such as media design, can impact downstream
purificationand utilization of the enzymes. The current work
demonstratestwo different enzyme fermentation and utilization
strategies:
(1) fed-batch fermentation of PAPS biosynthetic enzymeswithout
autoinduction, followed by utilization of the enzymespresent in the
soluble fraction; and (2) autoinduction fed-batchfermentation of
ASTIV, followed by utilization of purifiedenzymes in a PAPS
regeneration system. In both strategies,the first step requires
minimizing feed and reagents in thefermentation. For example, trace
metals were omitted andthe batch medium was prepared using
tryptone, yeast extract,and glycerol. The batch medium for
autoinduction was de-signed to use ammonium chloride, sodium
chloride, phos-phate buffer, and lactose. Minimizing the number of
ingredi-ents ensures better fermentation control, reduces errors
relatedto media formulation, scale-up, and technology transfer,
andreduces the costs and inventory management. Fed-batch
fer-mentation without autoinduction for the production of thePAPS
synthesis enzymes, PPA, APSK and ATPS, relies onstandard IPTG
induction. This fermentation strategy is robustand has been widely
used towards the production of manyother enzymes with minor
modifications for IPTG concentra-tion, duration of induction, and
induction temperature. A fed-batch fermentation strategy using
lactose autoinduction for theproduction of ASTIV was inspired based
on the shake flaskexperiments. Protein expression in shake flasks
using com-plex or defined media supplemented with phosphate
buffer,glucose, glycerol, lactose, and trace metals has been
reported(Studier 2005). The fed-batch fermentation strategy
forASTIV utilized this strategy with modifications includingthe
elimination of glucose and trace metals and an optimizedratio of
tryptone and yeast extract.
Enzyme fermentations are best evaluated using the volu-metric
yield of active enzymes required to generate the desiredproduct.
The current study utilizes two approaches of enzymeutilization: (1)
purification of enzymes using specific resin andusing immobilized
enzymes in the reaction of ASTIV-coupledPAPS regeneration system
and (2) directly adding the solublefraction to the reactionmixture
in PAPS synthesis. The currentHS polysaccharide synthesis involves
purification of ASTIVfrom the soluble fraction using a Ni-NTA
Sepharose resin,and the immobilized enzymes utilized for the
ASTIV-coupled PAPS regeneration system. The high cell
densityautoinduction fed-batch fermentation of the strain
expressingASTIV resulted in a volumetric yield of 560 mg/L of
fermen-tation broth, which comprises 48% of the total
enzymesexpressed in the soluble and insoluble fraction (Table
3).The current approach for PAPS synthesis involves quantifica-tion
of the PAPS enzymes from the soluble fraction and di-rectly adding
filtered soluble fractions containing PAPS syn-thesis enzymes in
the PAPS synthesis reaction mixture. Afterthe fermentation to
produce PAPS biosynthetic enzymes, theenzyme yield was calculated
using a semi-quantitative SDS-PAGE (Table 3). The PAPS biosynthetic
enzymes wereexpressed in two strains, E. coli BL21 Star™ (DE3)
strainexpressing PPA and E. coli BL21 Star™ (DE3) strain,
Fig. 5 Purified ASTIV on Ni-NTA Sepharose resin. Lane 1,
proteinladder; lane 2, 250 μg/mL BSA standard; lane 3, 125 μg/mL
BSAstandard; lane 4, 62.5 μg/mL BSA standard; lane 5, purified
ASTIV(37 KDa) enzyme
Table 4 Disaccharide compositional analysis of NSH and NS6S
0S NS 6S 2S NS6S NS2S 2S6S TriS
NSH 0.8 99.2 0.0 0.0 0.0 0.0 0.0 0.0
NS6S 0.7 13.1 0.0 0.0 86.2 0.0 0.0 0.0
-
expressing APSK and ATPS. The high cell density
fed-batchfermentation of the strain expressing PPA resulted in a
volu-metric yield of 910 mg/L of fermentation broth, which
com-prises 66% of the total enzymes expressed in the soluble
andinsoluble fraction (Table 3). Fermentation of the strain
co-expressing APSK and ATPS demonstrated variable expres-sion of
APSK and ATPS. The high cell density fed-batchfermentation for APSK
resulted in a volumetric yield of 280mg/L of fermentation broth
with 66% of the total enzymesexpressed in the soluble (active) and
insoluble (inactive) frac-tion (Table 3). The high cell density
fed-batch fermentation ofATPS resulted in a volumetric yield of 200
mg/L of fermen-tation broth, which comprises 34% of the total
enzymesexpressed in the soluble and insoluble fraction (Table
3).The expression of APSK and ATPS are desirable featuresbecause,
preliminary experiments showed that decreasedATPS expression
increased APSK expression favor PAPSproduction (Fig. 3).
We hypothesize that enzymes in the insoluble fraction con-sist
primarily of unfolded, partially processed, or inactive en-zymes
and that these enzymes may be scavenged with addi-tional
purification steps, including enzyme refolding. Theremay be various
factors that impact the accumulation of en-zymes in the insoluble
fraction. In analyzing PAPS biosyn-thetic enzymes and their
fermentations, we speculate thatsmaller enzymes can easily fold and
accumulate in the soluble
(active) fraction (Palmer and Wingfield 2012). Both APSK(25 KDa)
and ATPS (57 KDa) are produced in the samefermentation. However,
there are differences in the volumetricyield of APSK (66% soluble)
and ATPS (34% soluble).Similarly, PPA is smaller (21 KDa) and 66%
of the enzymeis in the soluble fraction. The production of enzymes
is oftennot straightforward and many factors can impact the
produc-tion of the volumetric yield of active enzymes, including
(1)expression of chaperones, (2) post-translational
modifications,and (3) strain metabolism (Belval et al. 2015;
Brylski et al.2019; Ellis 2013; Jhamb and Sahoo 2012; Thomas et
al.1997). In this context, we have observed with other enzymesthat
under similar fermentation conditions, and either induc-tion at 30
°C for 4 h or higher cell growth rate during induc-tion, the
majority of enzymes expressed accumulate in theinsoluble fraction.
Additionally, the carbon and nitrogen mo-lar ratio during
fermentation can impact enzyme production.Future research is geared
towards evaluating strategies to-wards improving soluble enzymes
utilization, including strainengineering, fermentation
optimization, and purification ofenzymes from insoluble
fractions.
In conclusion, the paper demonstrated that the
ePathbrickplatform that had previously been used for metabolic
engineer-ing, could be successfully utilized to study in vitro
enzymaticsynthesis conditions. This application of ePathbrick
platformcould be applied for one-pot synthesis of polysaccharides,
such
Fig. 6 1D 1H-NMR of NSH before and after 6OSTs treatment (NS6S).
Assignment, A, glucosamine; G, glucuronic acid; *, residual PAPS
and PAP
-
as heparin and heparan sulfate. Heparin and heparan sulfate
aresulfated polysaccharides with tissue-specific sulfation
domains.Tissue-specific sulfotransferases (and their isoforms)
dictate thesulfation patterns on heparin and heparan sulfate.
TheePathbrick tool could assemble seven-gene pathways (Xuet al.
2012) and could be utilized to evaluate effect ofsulfotransferase
expression ratio on sulfation patterns on hepa-rin and heparan
sulfate. In parallel, we have successfully dem-onstrated a
fed-batch fermentation protocol (IPTG induction ornon-autoinduction
protocol) that could be utilized for expres-sion of various
enzymes, including sulfotransferases (e.g.,6OST-3). The protocol
has been successfully utilized for fer-mentation, scale-up (100-L)
and technology transfer (to indus-trial partners) of heparin
biosynthetic enzymes (e.g., 2OST, C5epimerase, 6-OST-1, 6OST-3, and
3OST-1). The media andfeed composition are similar for all these
fermentations, withvariations in induction parameters (e.g., IPTG
concentration,induction temperature, and induction time). Adapting
all thebiosynthetic enzymes in similar fermentation protocol
enablesus to control inventory management and cost. In addition,
thepaper successfully demonstrates an autoinduction
microbialfed-batch fermentation strategy towards the production of
theenzyme ASTIV. The successful fermentation strategies and
re-duction in cost of the production of enzymes (related to
up-stream) has enabled the successful pilot scale production
andutilization of the enzymes.
Authors’ contributions RJL and JSD provided the funding for this
study.RJL and PD planned the study. PD, RJL, and JSD wrote the
manuscript.PD performed the fermentations. WH andMAGK designed and
preparedthe recombinant organisms. LF was responsible for the
studies.
Funding information This work was supported by the US
NationalScience Foundation award CBET-1604547 awarded to MAGK and
byNational Institutes of Health grants CA231074 and DK111958 to
RJL.
Compliance with ethical standards This article does notcontain
any studies with human participants or animals performed byany of
the authors.
Conflict of interest The authors declare that they have no
conflict ofinterest.
References
Badri A, Williams A, Linhardt RJ, Koffas MAG (2018) The road
toanimal-free glycosaminoglycan production: current efforts and
bot-tlenecks. Curr Opin Biotechnol 53:85–92.
https://doi.org/10.1016/j.copbio.2017.12.018
Badri A, Williams A, Xia K, Linhardt RJ, Koffas MAG (2019)
Increased3′-phosphoadenosine-5′-phosphosulfate levels in
engineeredEscherichia coli cell lysate facilitate the in vitro
synthesis of chon-droitin sulfate A. Biotechnol J 14(9):e1800436.
https://doi.org/10.1002/biot.201800436
Belval L, Marquette A, Mestre P, Piron MC, Demangeat G,
MerdinogluD, Chich JF (2015) A fast and simple method to eliminate
Cpn60
from functional recombinant proteins produced by E. coli
ArcticExpress. Protein Expr Purif 109:29–34.
https://doi.org/10.1016/j.pep.2015.01.009
Brylski O, Ebbinghaus S, Mueller JW (2019) Melting down protein
sta-bility: PAPS synthase 2 in patients and in a cellular
environment.Front Mol Biosci 6:31.
https://doi.org/10.3389/fmolb.2019.00031
Burkart MD, Izumi M, Chapman E, Lin CH, Wong CH
(2000)Regeneration of PAPS for the enzymatic synthesis of sulfated
oli-gosaccharides. J Org Chem 65(18):5565–5574.
https://doi.org/10.1021/jo000266o
Chen J, Avci FY, Muñoz EM, McDowell LM, Chen M, Pedersen
LC,Zhang L, Linhardt RJ, Liu J (2005) Enzymatic redesigning of
bio-logically active heparan sulfate. J Biol Chem
280:42817–42825.https://doi.org/10.1074/jbc.M504338200
Coughtrie MWH (2016) Function and organization of the human
cyto-solic sulfotransferase (SULT) family. Chem Biol Interact
259:2–7.https://doi.org/10.1016/j.cbi.2016.05.005
Ellis RJ (2013) Assemwbly chaperones: a perspective. Philos
Trans RSoc B Biol Sci 368(1617):20110398.
https://doi.org/10.1098/rstb.2011.0398
Fu L, Zhang F, Li G, Onishi A, Bhaskar U, Sun P, Linhardt RJ
(2014)Structure and activity of a new low-molecular-weight heparin
pro-duced by enzymatic ultrafiltration. J Pharm Sci
103(5):1375–1383.https://doi.org/10.1002/jps.23939
Fu L, Sun X, HeW, Cai C, Onishi A, Zhang F, Linhardt RJ, Liu Z
(2016)Keratan sulfate glycosaminoglycan from chicken egg
white.Glycobiology 26(7):693–700.
https://doi.org/10.1093/glycob/cww017
Fu L, Li K,Mori D, HirakaneM, Lin L, Grover N, Datta P, Yu Y,
Zhao J,Zhang F, Yalcin M, Mousa SA, Dordick JS, Linhardt RJ
(2017)Enzymatic generation of highly anticoagulant bovine
intestinal hep-arin. J Med Chem 60:8673–8679.
https://doi.org/10.1021/acs.jmedchem.7b01269
Goettsch S, Badea RA, Mueller JW, Wotzlaw C, Schoelermann
B,Schulz L, Rabiller M, Bayer P, Hartmann-Fatu C (2006) HumanTPST1
transmembrane domain triggers enzyme dimerisation andlocalisation
to the golgi compartment. J Mol Biol
361:436–449.https://doi.org/10.1016/j.jmb.2006.06.023
Günal S, Hardman R, Kopriva S, Mueller JW (2019) Sulfation
pathwaysfrom red to green. J Biol Chem 294:12293–12312.
https://doi.org/.https://doi.org/10.1074/jbc.REV119.007422
Hale AT, Brown RE, Luka Z, Hudson BH, Matta P, Williams CS,
YorkJD (2020)Modulation of sulfur assimilationmetabolic toxicity
over-comes anemia and hemochromatosis in mice. Adv Biol Regul
76:100694. https://doi.org/10.1016/j.jbior.2020.100694
He W (2017) Metabolic engineering and applied enzymology for
thepreparation of nutraceutical/ pharmaceutical chondroitin
sulfate.Rensselaer Polytechnic Institute
Italia JS, Peeler JC, Hillenbrand CM, Latour C, Weerapana E,
ChatterjeeA (2020) Genetically encoded protein sulfation in
mammalian cells.Nat Chem Biol 16:379–382.
https://doi.org/10.1038/s41589-020-0493-1
Jhamb K, Sahoo DK (2012) Production of soluble recombinant
proteinsin Escherichia coli: Effects of process conditions and
chaperone co-expression on cell growth and production of xylanase.
BioresourTechnol 123:135–143.
https://doi.org/10.1016/j.biortech.2012.07.011
Kamiyama S, Suda T, Ueda R, Suzuki M, Okubo R, Kikuchi N, Chiba
Y,Goto S, Toyoda H, Saigo K, Watanabe M, Narimatsu H, Jigami
Y,Nishihara S (2003) Molecular cloning and identification of
3′-phosphoadenosine 5′-phosphosulfate transporter. J Biol
Chem278(28):25958–25963. https://doi.org/10.1074/jbc.M302439200
Klaassen CD, Boles JW (1997) The importance of
3′-phosphoadenosine5′-phosphosulfate (PAPS) in the regulation of
sulfation. FASEB J11:404–418.
https://doi.org/10.1096/fasebj.11.6.9194521
https://doi.org/10.1016/j.copbio.2017.12.018https://doi.org/10.1016/j.copbio.2017.12.018https://doi.org/10.1002/biot.201800436https://doi.org/10.1002/biot.201800436https://doi.org/10.1016/j.pep.2015.01.009https://doi.org/10.1016/j.pep.2015.01.009https://doi.org/10.3389/fmolb.2019.00031https://doi.org/10.1021/jo000266ohttps://doi.org/10.1021/jo000266ohttps://doi.org/10.1074/jbc.M504338200https://doi.org/10.1016/j.cbi.2016.05.005https://doi.org/10.1098/rstb.2011.0398https://doi.org/10.1098/rstb.2011.0398https://doi.org/10.1002/jps.23939https://doi.org/10.1093/glycob/cww017https://doi.org/10.1093/glycob/cww017https://doi.org/10.1021/acs.jmedchem.7b01269https://doi.org/10.1021/acs.jmedchem.7b01269https://doi.org/10.1016/j.jmb.2006.06.023https://doi.org/10.1074/jbc.REV119.007422https://doi.org/10.1016/j.jbior.2020.100694https://doi.org/10.1038/s41589-020-0493-1https://doi.org/10.1038/s41589-020-0493-1https://doi.org/10.1016/j.biortech.2012.07.011https://doi.org/10.1016/j.biortech.2012.07.011https://doi.org/10.1074/jbc.M302439200https://doi.org/10.1096/fasebj.11.6.9194521
-
Koprivova A, Kopriva S (2016) Sulfation pathways in plants. Chem
BiolInteract 259:23–30.
https://doi.org/10.1016/j.cbi.2016.05.021
Lin X (2004) Functions of heparan sulfate proteoglycans in cell
signalingduring development. Development 131:6009–6021.
https://doi.org/10.1242/dev.01522
Lindahl U, Kusche-Gullberg M, Kjellen L (1998) Regulated
diversity ofheparan sulfate. J Biol Chem 273:24979–24982.
https://doi.org/10.1074/jbc.273.39.24979
Linhardt RJ (2003) 2003 Claude S. Hudson award address in
carbohy-drate chemistry. Heparin: Structure and activity. J Med
Chem 46:2551–2564
Liu J, Linhardt RJ (2014) Chemoenzymatic synthesis of heparan
sulfateand heparin. Nat Prod Rep 31:1676–1685.
https://doi.org/10.1039/c4np00076e
Palmer I, Wingfield PT (2012) Preparation and extraction of
insoluble(inclusion-body) proteins from Escherichia coli. Curr
Protoc ProteinSci:Unit 6.3.
https://doi.org/10.1002/0471140864.ps0603s70
Paul P, Suwan J, Liu J, Dordick JS, Linhardt RJ (2012) Recent
advancesin sulfotransferase enzyme activity assays. Anal Bioanal
Chem403(6):1491–1500. https://doi.org/10.1007/s00216-012-5944-4
Piecewicz S, Sengupta S (2011) The dynamic glycome
microenviron-ment and stem cell differentiation into vasculature.
Stem Cells Dev20:749–758. https://doi.org/10.1089/scd.2010.0454
Schwarz JK, Capasso JM, Hirschberg CB (1984) Translocation of
aden-osine 3′-phosphate 5′-phosphosulfate into rat liver Golgi
vesicles. JBiol Chem 259(6):3554–3559
Soares da Costa D, Reis RL, Pashkuleva I (2017) Sulfation of
glycosami-noglycans and its implications in human health and
disorders. AnnuRev Biomed Eng 19:1–26.
https://doi.org/10.1146/annurev-bioeng-071516-044610
Studier FW (2005) Protein production by auto-induction in high
densityshaking cultures. Protein Expr Purif 41(1):207–234.
https://doi.org/10.1016/j.pep.2005.01.016
Thomas JG, Ayling A, Baneyx F (1997) Molecular chaperones,
foldingcatalysts, and the recovery of active recombinant proteins
fromE. coli: to fold or to refold. Appl Biochem Biotechnol Part
AEnzym Eng Biotechnol 66(3):197–238.
https://doi.org/10.1007/BF02785589
Wang Z, Li J, Cheong S, Bhaskar U, Akihiro O, Zhang F, Dordick
JS,Linhardt RJ (2011) Response surface optimization of the
heparosanN-deacetylation in producing bioengineered heparin. J
Biotechnol156:188–196.
https://doi.org/10.1016/j.jbiotec.2011.08.013
Xiong J, Bhaskar U, Li G, Fu L, Li L, Zhang F, Dordick JS,
Linhardt RJ(2013) Immobilized enzymes to convert N-sulfo,
N-acetylheparosan to a critical intermediate in the production
ofbioengineered heparin. J Biotechnol 167:241–247.
https://doi.org/10.1016/j.jbiotec.2013.06.018
Xiong A, Kundu S, Forsberg-Nilsson K (2014) Heparan sulfate in
theregulation of neural differentiation and glioma development.
FEBS J281:4993–5008. https://doi.org/10.1111/febs.13097
Xu P, Vansiri A, Bhan N, Koffas MAG (2012) ePathBrick: a
syntheticbiology platform for engineering metabolic pathways in E.
coli.ACS Synth Biol 1:256–266.
https://doi.org/10.1021/sb300016b
Zhang F, Zhang Z, Linhardt RJ (2009) Glycosaminoglycans.
In:Cummings RD, Pierce M (eds) Handbook of Glycomics:Glycoconjugate
Structural Analysis, vol Chapter 3. AcademicPress/Elsevier, pp
59–80
Zhou X, Chandarajoti K, Pham TQ, Liu R, Liu J (2011) Expression
ofheparan sulfate sulfotransferases in Kluyveromyces lactis and
prep-aration of 3′-phosphoadenosine-5′-phosphosulfate.
Glycobiology21(6):771–780.
https://doi.org/10.1093/glycob/cwr001
Publisher’s note Springer Nature remains neutral with regard to
jurisdic-tional claims in published maps and institutional
affiliations.
Affiliations
Payel Datta1,2 & Li Fu 1,2 &Wenqin He1,2 &M. A. G.
Koffas1,2,3 & J. S. Dordick1,2,3,4 & R. J.
Linhardt1,2,3,4
1 Department of Chemical and Biological Engineering, Center
forBiotechnology and Interdisciplinary Studies, Rensselaer
PolytechnicInstitute, Troy, NY 12180, USA
2 Department of Chemistry and Chemical Biology, Center
forBiotechnology and Interdisciplinary Studies, Rensselaer
PolytechnicInstitute, Troy, NY 12180, USA
3 Department of Biomedical Engineering, Center for
Biotechnologyand Interdisciplinary Studies, Rensselaer Polytechnic
Institute,Troy, NY 12180, USA
4 Department of Biology, Center for Biotechnology
andInterdisciplinary Studies, Rensselaer Polytechnic
Institute,Troy, NY 12180, USA
https://doi.org/10.1016/j.cbi.2016.05.021https://doi.org/10.1242/dev.01522https://doi.org/10.1242/dev.01522https://doi.org/10.1074/jbc.273.39.24979https://doi.org/10.1074/jbc.273.39.24979https://doi.org/10.1039/c4np00076ehttps://doi.org/10.1039/c4np00076ehttps://doi.org/10.1002/0471140864.ps0603s70https://doi.org/10.1007/s00216-012-5944-4https://doi.org/10.1089/scd.2010.0454https://doi.org/10.1146/annurev-bioeng-071516-044610https://doi.org/10.1146/annurev-bioeng-071516-044610https://doi.org/10.1016/j.pep.2005.01.016https://doi.org/10.1016/j.pep.2005.01.016https://doi.org/10.1007/BF02785589https://doi.org/10.1007/BF02785589https://doi.org/10.1016/j.jbiotec.2011.08.013https://doi.org/10.1016/j.jbiotec.2013.06.018https://doi.org/10.1016/j.jbiotec.2013.06.018https://doi.org/10.1111/febs.13097https://doi.org/10.1021/sb300016bhttps://doi.org/10.1093/glycob/cwr001http://orcid.org/0000-0003-2219-5833