Embed Size (px)
Citation preview

APPLIED AND ENVIRONMENTAL MICROBIOLOGY, Jan. 1994, p. 167-1730099-2240/94/$04.00+0Copyright ( 1994, American Society for Microbiology
Quantitative Centrifugation To Extract Benthic Protozoafrom Freshwater Sediments
MATHIEU STARINK,l* MARIE-JOSE BAR-GILISSEN,1 ROLF P. M. BAK,2AND THOMAS E. CAPPENBERG'
Centre for Limnology, Netherlands Institute of Ecology, 3631 AC Nieuwersluis, 1 and NetherlandsInstitute for Sea Research, 1790 AB Den Burg, Texel,2 The Netherlands
Received 8 June 1993/Accepted 3 November 1993
Two methods for extracting protists from freshwater sediment are described: (i) an adapted isopycniccentrifugation technique for sandy and gyttja-like sediments and (ii) a rate zonal centrifugation technique forsediments rich in particulate organic material (litter-like sediments). The recoveries of protists during isopycniccentrifugation in media of several densities were compared. No significant losses in sodium diatrizoate andPercoll were recorded. After known amounts of nanoflagellates were added to azoic sediments, the protistswere extracted and counted. For sandy sediments, we found 100%o recovery, and for the gyttja-like sedimentswe found a maximum recovery of 94%. The recovery of protozoa extracted from litter-like sediments,characteristic of littoral systems, depends on a given centrifugal force, on time, and on the dimensions of theflagellates. A recovery model which takes into account cell dimensions and centrifugation characteristics givesthe minimum expected recovery.
In pelagic marine and freshwater systems, heterotrophicnanoflagellates are major consumers of bacteria (7, 9, 16) andplay an important role in enhancing the mineralization rate oforganic matter (5, 15). Less information is available on therole of marine benthic flagellates within sediment (4, 11), andthe possible impact of heterotrophic nanoflagellates on themicrobial processes in freshwater sediments has been ne-glected completely to our knowledge. We have begun inves-tigating the role of heterotrophic nanoflagellates in the mi-crobial processes in these sediments as part of a long-termstudy of the carbon cycling in a littoral and in a profundalregion.Up to now, there has been no description of a method for
separating nanoflagellates quantitatively from freshwatersediments, which is necessary for counting and investigatingthe protists without masking effects by sediment particles.For marine sediments, various extraction techniques forlarger protists (mostly ciliates) are available (1, 14, 18).Alongi (1) has compared methods for extracting larger pro-tists and suggested that methods in which silica sols are
used, particularly Percoll-sorbitol, were superior to the othertechniques tested. Subsequent counting of the extractedprotozoa was performed with dissecting microscopes. How-ever, to include the smallest flagellates in the observations itis necessary to use a large magnification and epifluorescencetechniques (2, 3). A reliable method for extracting smallprotists from sediments, comprising subsampling of super-natant and epifluorescence microscopy, was developed byBak and Nieuwland (3). However, using their techniques wefailed to extract flagellates from our sandy-detritus-richfreshwater sediments. Gyttja sediment is difficult to separatefrom the nanoflagellates by the cited methods because of itsfine structure. To surmount this problem, we have adaptedand improved the Percoll method for the sandy and gyttjasediments of freshwater systems. In this isopycnic centrifu-
* Corresponding author. Mailing address: Centre for Limnology,Netherlands Institute of Ecology, 3631 AC Nieuwersluis, TheNetherlands. Phone: +31 2943 3599. Fax: +31 2943 2224.
gation technique, particles are separated solely on the basisof differences in buoyant density, irrespective of size. If thedensity of the gradient is appropriate, no pelleting of protistswill occur. Some locations in freshwater habitats, such as
reed belts, carry high loads of fresh particulate organicmatter. Isopycnic centrifugation extraction techniques are
not suitable for these sediments, and therefore we developeda rate zonal centrifugation technique to extract protists fromthese sediments. In this type of separation, size differencebetween particles is the parameter that determines theseparation. Relatively large particles pellet and are separatedfrom the relatively small ones that stay in suspension andwill be recovered.The efficiency of both methods was tested by adding
known amounts of benthic nanoflagellates to azoic sedi-ments. The recovery from our Percoll method was between94 and 100%, depending on the sediment type but irrespec-tive of the size of the protists. The recovery from the ratezonal centrifugation technique depends on the size of theflagellates and varied in our study between 71 and 100%. Wepresent a theoretically based recovery model that will givethe minimum recovery as a function of flagellate size for agiven centrifugation force and time.
MATERIALS AND METHODS
Samples. Sediment samples were collected from a littoralzone and from a profundal region. The littoral study sites are
located on the southern shore of Lake Gooimeer near thetown of Naarden in the central part of the Netherlands. LakeGooimeer is one of the surface waters created during thedamming and reclamation of the Zuiderzee, now Ijsselmeer.This littoral zone is a former sandbank at the southern shoreof the lake, which is mainly colonized by Phragmites aus-
tralis over a length of about 4 km and a surface of ca. 0.3km2. The profundal region is located in Lake Vechten. Thislake was formed by excavation of sand and is also situated inthe central part of the Netherlands, near the city of Utrecht.
Counting. Protist cultures as well as sediment sampleswere fixed with 1% glutaraldehyde (final concentration). The
167
Vol. 60, No. 1
on April 14, 2020 by guest
http://aem.asm
.org/D
ownloaded from

APPL. ENVIRON. MICROBIOL.
TABLE 1. The mean recovery of cells from different sediments, extracted with the rate zonal and the isopycniccentrifugation techniques
Sediment Location Species Protist density ESD Method Recoverytype (cell~~~~~~Potst densit ESD etho (% ± SD)
Sand Gooimeer Mixed 1.19 x 105 NDa Isopycnic 101 + 7.0; n = 6Gyttja Vechten Monas sp. 1.48 x 105 5.0 Isopycnic 94 + 3.4; n = 3Litter Gooimeer Mixed 1.19 x 105 5.3 Rate zonal 86 ± 5.4; n = 2Litter Gooimeer Mixed 4.37 x 104 5.7 Rate zonal 80 + 8.5; n = 2Litter Gooimeer Mixed 1.08 x 105 7.9 Rate zonal 71 - 3.9; n = 4Litter Gooimeer Bodo sp. 1.35 X 105 4.8 Rate zonal 102 + 2.2; n = 2a ND, not determined.
cultured and the extracted protists were stained with prim-ulin (Janssen Chemica, Geel, Belgium) and counted directlyon 1-p.m membrane filters (Nuclepore Corp., Pleasanton,Calif.) by epifluorescence microscopy, at x 1,250 magnifica-tion (6). Protists were counted on randomly selected fields,until either 200 fields or 250 protists were counted. Forbiovolume estimates, 250 fixed organisms per sample weremeasured by eyepiece micrometer and volumes were calcu-lated from length and width, assuming a spherical or cylin-drical shape. Equivalent spherical diameters (ESDs) werecalculated from these volumes.
Centrifugational extracting techniques. (i) Testing gradientsolutions. The chemical-physical influence of some gradientmedia on protists was tested in order to choose the optimumgradient solution for the isopycnic centrifugation extractiontechnique. Ficoll, sodium diatrizoate, and sorbitol (SigmaChemical Co., St. Louis, Mo.) were mixed with a knowndensity of fixed protists so that the final density of thesolution was 1.13 g cm-3. The final protist concentration inthese solutes ranged from 1.6 x 105 to 2.7 x 105 cells cm-3.Percoll (Pharmacia Fine Chemicals, Uppsala, Sweden) wasmixed with a similar volume of fixed protists to obtain a 50%Percoll solution with a final cell concentration of 2.7 x 105cells cm-3. After different time intervals, the gradient-protistsolutions were subsampled and the recovery of the protistswas determined.
(ii) Isopycnic centrifugation. Nonlinear 50% Percoll gradi-ents were made from 4.5 ml of 0.2-p.m (pore-size)-mem-brane-filtered (Schleicher & Schull, Inc., Keene, N.H.)Percoll and 4.5 ml of Milli-Q water in 15-ml polycarbonatecentrifuge tubes. The centrifuge tubes were placed in an SS34 angle rotor and centrifuged at 38,800 x g for 30 min in aSorvall SR5C centrifuge (Sorvall Instruments, du Pont,Wilmington, Del.). Fixed gyttja sediment samples werediluted lx to 40x with Milli-Q water. After diluting, thesediment slurry was allowed to settle for 15 s before analiquot was taken with a pipette. Samples (2 ml) werecarefully loaded onto the top of the Percoll gradient. Sandysediments were processed in the same way without diluting.The loaded gradients were centrifuged at 4,300 x g for 15min in a Sigma 3E-1 centrifuge. A swing-out head (Sigma;type 11117) was used to reduce wall effects appearing infixed-angle rotors. Supernatants were decanted from thecentrifuge tubes and diluted in 30-ml volumetric flasks. Fromthis, a subsample was filtered on a membrane filter andrinsed with 2 ml of water before the actual staining proce-dure was carried out. Rinsing with Milli-Q water is necessaryto remove the Percoll from the filter, which tends to gel whenthe pH decreases below 5.5 during the staining procedure.
(iii) Rate zonal centrifugation. Fixed sediment samples richin particulate organic matter were diluted (1:8) with a 0.05 MTrizma (Sigma) buffer, pH 4.0. Glass centrifugation tubes
were filled with 10 ml of the diluted sediment samples. Thesamples were centrifuged at 100 x g from 0.5 to 5 min in aSigma 3E-1 centrifuge with a swing-out head. Supernatantswere decanted from the centrifuge tubes. The pellets wereresuspended with the diluent and centrifuged again. In total,three centrifugation steps were performed; the supernatantswere collected in 30-ml volumetric flasks. From this, asubsample was used for the staining procedure.
Subsampling of sediment. (i) Effect of sediment dispersion.To test the effect of sediment dispersion on the amount ofextracted cells, 10 ml of fixed sediment (reed litter) contain-ing the natural protist population was diluted with 80 ml of adiluent. Three different diluents were tested: 0.05 M Trizmabuffer (pH 4.0), 0.05 M Trizma buffer (pH 7.0), and a 1%glutaraldehyde solution. After dilution, the sediment sam-ples were shaken on a wrist-action shaker (Gallenkamp,Loughborough, England) at speed 7 (about 180 oscillationsmin-') for 0, 15, 30, and 60 min. Then, the protozoa wereextracted from the sediment by the rate zonal centrifugationtechnique. After extraction of the protists, slides wereprepared and protists were counted.
(ii) Subsampling technique. With some adaptations, weprocessed the sediment samples as described by Bak andNieuwland (3). Instead of washing the sediment in sterileseawater fixative-proflavine, we used Milli-Q water withglutaraldehyde (at a final concentration of 1%) and a 0.05 MTrizma buffer, pH 4.0. The samples were allowed to standfor 30 or 60 min (see Results) before subsampling the centerof the 12-cm-high column. Samples (5 ml) of the supernatantwere filtered and stained as described above.
Recovery. (i) Recovery experiments. The efficiency of theextraction methods of both centrifugation techniques wastested by adding a protozoan culture of known density andsize frequency to azoic sediments (Table 1). In our experi-ments, we used three types of sediment: quartz-carbonatesand, gyttja sediment, and a muddy sediment rich in partic-ulate organic matter from a reed (P. australis) belt. Thesediments were made azoic by drying at 80°C for 12 h.Blanks were run to check if the sediments were azoic. Afterdrying, a volume of a protist suspension equal to the amountof evaporated water was added. The protists were added liveto the sediments, and these were subsampled after 5 min.
(ii) Recovery model. To investigate the relation betweenparticle size (ESD) and recovery value, a size-dependentrecovery model for the rate zonal extraction technique wasconstructed. The base of this model was formed by theequation of Stokes for ideally spherical particles: v = d2 (p.- Pl) x g x (18&r)-1, where v is the sedimentation rate(meters per second), d is the diameter of the particle (equalspherical diameter) (meters), pp is the particle density (kilo-grams per cubic meter),p, is the liquid density (kilograms percubic meter), 9 is the viscosity of the medium (newtons per
168 STARINK ET AL.
on April 14, 2020 by guest
http://aem.asm
.org/D
ownloaded from
EXTRACTION OF BENTHIC PROTOZOA FROM SEDIMENTS 169
second per square meter), and g is the centrifugal force(meters per second per second).
In order to obtain the buoyant density (p,) of a spherical,fixed Monas sp. culture with known cell diameter, thesedimentation rate in a centrifugal field was measured, andfrom this the buoyant density (p,) was calculated. For thispurpose, 10 centrifuge tubes were each filled with 1.25 ml ofa fixed Monas sp. suspension and 8.75 ml of 0.05 M Trizmabuffer (pH 4.0). The tubes were centrifuged at 100 x g in aSigma 3E-1 centrifuge with a swing-out head. After 0, 5, 10,15, and 20 min of centrifugation, the cell concentration (inthe duplicate tubes) was determined in the upper 5 ml (a2.8-cm-high column) of the supernatant. To compare thesedimentation rates of nonspherical flagellates and the spher-ical Monas sp., the sedimentation rate of a flagellate popu-lation with different length and width ratios (ranging from 1to 4 with a mean [+ standard deviation] ESD of 5.5 + 2.1,um) was determined in the same way. The nonsphericalflagellates were cultured on bacteria-enriched fresh reedlitter (P. australis).
(iii) Comparison of the centrifugational extracting tech-niques. Because the yield of the rate zonal extraction tech-niques is size dependent, in contrast to that of the isopycnictechnique, an experiment to compare the extraction efficien-cies of both techniques was carried out. An expected sizefrequency distribution for cells extracted by the rate zonaltechnique was constructed on the basis of the predictedrecovery values from the model and the size frequencydistribution of cells extracted by the isopycnic technique.For this purpose, a glutaraldehyde-fixed and homogenized
sandy-sediment sample was split into five equal subsamples.From two samples, the protists were extracted by theisopycnic centrifuge technique, and from the other threethey were extracted by the rate zonal technique. The sam-ples were processed as described above. The differencesbetween the ESD frequency distributions were statisticallytested with the Kolmogorov-Smirnov two-sample test (17).
RESULTS
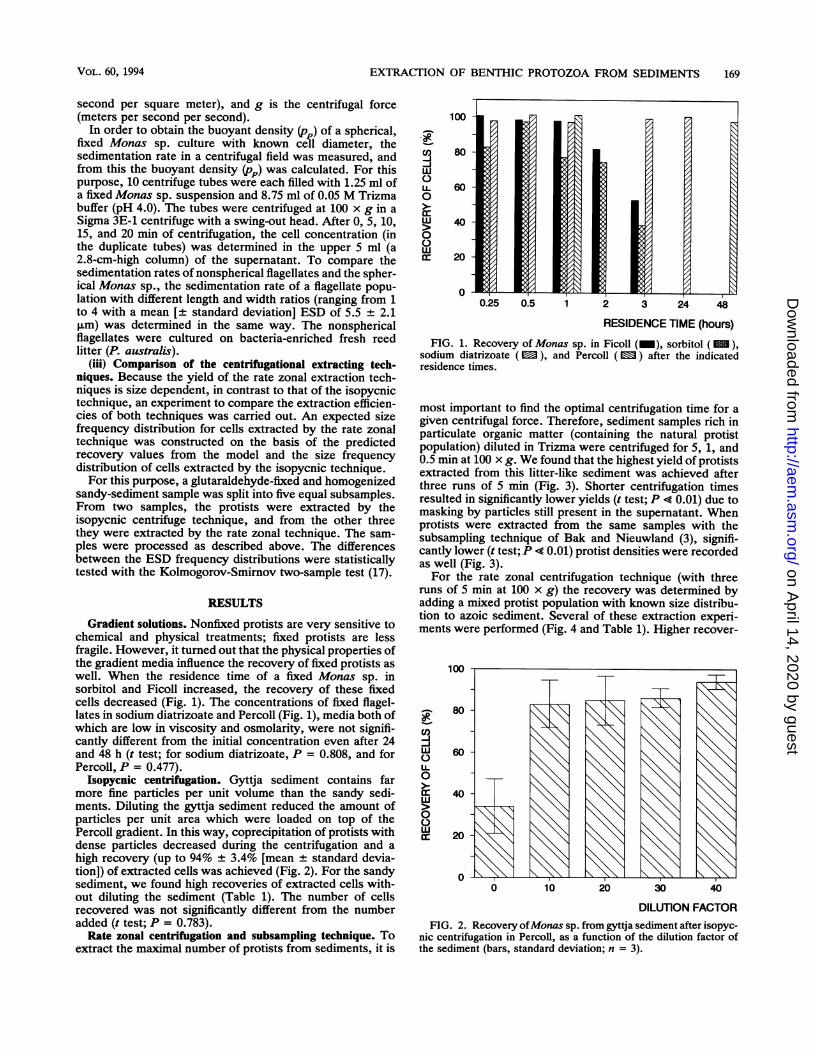
Gradient solutions. Nonfixed protists are very sensitive tochemical and physical treatments; fixed protists are lessfragile. However, it turned out that the physical properties ofthe gradient media influence the recovery of fixed protists aswell. When the residence time of a fixed Monas sp. insorbitol and Ficoll increased, the recovery of these fixedcells decreased (Fig. 1). The concentrations of fixed flagel-lates in sodium diatrizoate and Percoll (Fig. 1), media both ofwhich are low in viscosity and osmolarity, were not signifi-cantly different from the initial concentration even after 24and 48 h (t test; for sodium diatrizoate, P = 0.808, and forPercoll, P = 0.477).
Isopycnic centrifugation. Gyttja sediment contains farmore fine particles per unit volume than the sandy sedi-ments. Diluting the gyttja sediment reduced the amount ofparticles per unit area which were loaded on top of thePercoll gradient. In this way, coprecipitation of protists withdense particles decreased during the centrifugation and ahigh recovery (up to 94% ± 3.4% [mean + standard devia-tion]) of extracted cells was achieved (Fig. 2). For the sandysediment, we found high recoveries of extracted cells with-out diluting the sediment (Table 1). The number of cellsrecovered was not significantly different from the numberadded (t test; P = 0.783).Rate zonal centrifugation and subsampling technique. To
extract the maximal number of protists from sediments, it is
100
-
10U-LJIL0
LLJ0C.)w
80
60
40
20
0 WpA rOfA ,A w mA r A
0.25 0.5 1 2 3 24 48
RESIDENCE TIME (hours)
FIG. 1. Recovery of Monas sp. in Ficoll (_), sorbitol (M),sodium diatrizoate ( 1 ), and Percoll ( 1 ) after the indicatedresidence times.
most important to find the optimal centrifugation time for agiven centrifugal force. Therefore, sediment samples rich inparticulate organic matter (containing the natural protistpopulation) diluted in Trizma were centrifuged for 5, 1, and0.5 min at 100 x g. We found that the highest yield of protistsextracted from this litter-like sediment was achieved afterthree runs of 5 min (Fig. 3). Shorter centrifugation timesresulted in significantly lower yields (t test; P < 0.01) due tomasking by particles still present in the supernatant. Whenprotists were extracted from the same samples with thesubsampling technique of Bak and Nieuwland (3), signifi-cantly lower (t test;P < 0.01) protist densities were recordedas well (Fig. 3).For the rate zonal centrifugation technique (with three
runs of 5 min at 100 x g) the recovery was determined byadding a mixed protist population with known size distribu-tion to azoic sediment. Several of these extraction experi-ments were performed (Fig. 4 and Table 1). Higher recover-
100
-J
wU-
LIJ0
w00wa:
80 -
60 -
40 -
20 -
00 10 20 30 40
DILUTION FACTORFIG. 2. Recovery ofMonas sp. from gyttja sediment after isopyc-
nic centrifugation in Percoll, as a function of the dilution factor ofthe sediment (bars, standard deviation; n = 3).
VOL. 60, 1994
-T-
-FN\
1\
-N-- N N
on April 14, 2020 by guest
http://aem.asm
.org/D
ownloaded from
APPL. ENVIRON. MICROBIOL.
ECe
.bCD)
o1-
z0
z
0z00
80
60
40
20
0
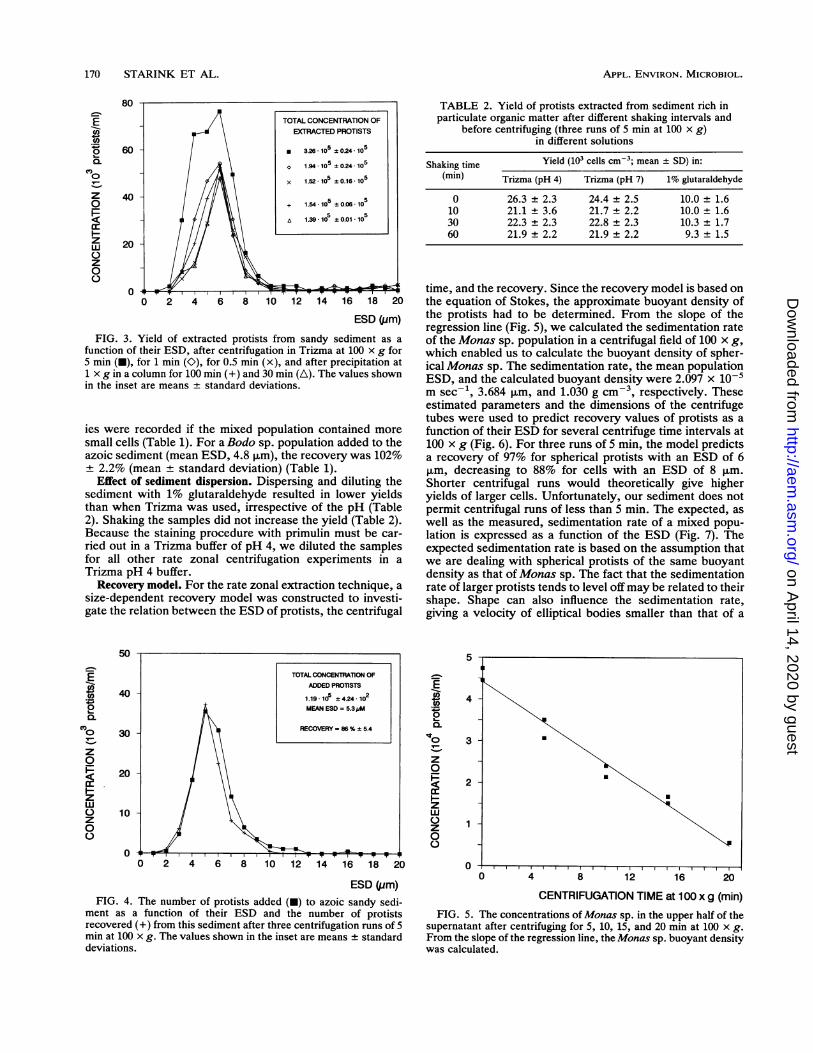
FIG. 3. Yield of extracted protists from sandy sediment as afunction of their ESD, after centrifugation in Trizma at 100 x g for5 min (G), for 1 min (O), for 0.5 min (x), and after precipitation at1 x g in a column for 100 min (+) and 30 min (A). The values shownin the inset are means ± standard deviations.
ies were recorded if the mixed population contained moresmall cells (Table 1). For a Bodo sp. population added to theazoic sediment (mean ESD, 4.8 p,m), the recovery was 102%+ 2.2% (mean + standard deviation) (Table 1).
Effect of sediment dispersion. Dispersing and diluting thesediment with 1% glutaraldehyde resulted in lower yieldsthan when Trizma was used, irrespective of the pH (Table2). Shaking the samples did not increase the yield (Table 2).Because the staining procedure with primulin must be car-ried out in a Trizma buffer of pH 4, we diluted the samplesfor all other rate zonal centrifugation experiments in aTrizma pH 4 buffer.
Recovery model. For the rate zonal extraction technique, asize-dependent recovery model was constructed to investi-gate the relation between the ESD of protists, the centrifugal
50
40
30
20
10
0
ESD (,jm)FIG. 4. The number of protists added (-) to azoic sandy sedi-
ment as a function of their ESD and the number of protistsrecovered (+) from this sediment after three centrifugation runs of 5min at 100 x g. The values shown in the inset are means standarddeviations.
TABLE 2. Yield of protists extracted from sediment rich inparticulate organic matter after different shaking intervals and
before centrifuging (three runs of 5 min at 100 x g)in different solutions
Shaking time Yield (103 cells cm-3; mean ± SD) in:(mm) Trizma (pH 4) Trizma (pH 7) 1% glutaraldehyde
0 26.3 ± 2.3 24.4 + 2.5 10.0 ± 1.610 21.1 ± 3.6 21.7 ± 2.2 10.0 ± 1.630 22.3 ± 2.3 22.8 ± 2.3 10.3 ± 1.760 21.9 + 2.2 21.9 + 2.2 9.3 ± 1.5
time, and the recovery. Since the recovery model is based onthe equation of Stokes, the approximate buoyant density ofthe protists had to be determined. From the slope of theregression line (Fig. 5), we calculated the sedimentation rateof the Monas sp. population in a centrifugal field of 100 x g,which enabled us to calculate the buoyant density of spher-ical Monas sp. The sedimentation rate, the mean populationESD, and the calculated buoyant density were 2.097 x 10-m sec-1, 3.684 ,um, and 1.030 g cm-3, respectively. Theseestimated parameters and the dimensions of the centrifugetubes were used to predict recovery values of protists as afunction of their ESD for several centrifuge time intervals at100 x g (Fig. 6). For three runs of 5 min, the model predictsa recovery of 97% for spherical protists with an ESD of 6p,m, decreasing to 88% for cells with an ESD of 8 ,um.Shorter centrifugal runs would theoretically give higheryields of larger cells. Unfortunately, our sediment does notpermit centrifugal runs of less than 5 min. The expected, aswell as the measured, sedimentation rate of a mixed popu-lation is expressed as a function of the ESD (Fig. 7). Theexpected sedimentation rate is based on the assumption thatwe are dealing with spherical protists of the same buoyantdensity as that ofMonas sp. The fact that the sedimentationrate of larger protists tends to level offmay be related to theirshape. Shape can also influence the sedimentation rate,giving a velocity of elliptical bodies smaller than that of a
5
0.
z
z
z
0=1
4
3
2
1
0
CENTRIFUGATION TIME at 100 x g (min)
FIG. 5. The concentrations of Monas sp. in the upper half of thesupernatant after centrifuging for 5, 10, 15, and 20 min at 100 x g.From the slope of the regression line, the Monas sp. buoyant densitywas calculated.
E.to4-
0-
zoz0IL
zw00
170 STARINK ET AL.
on April 14, 2020 by guest
http://aem.asm
.org/D
ownloaded from
EXTRACTION OF BENTHIC PROTOZOA FROM SEDIMENTS 171
100
80
60
40
20
0
30
=c)00
w
LL00ll-Jw
20
10
02 4 6 8 10 12 14 16 18 20
ESDQm)
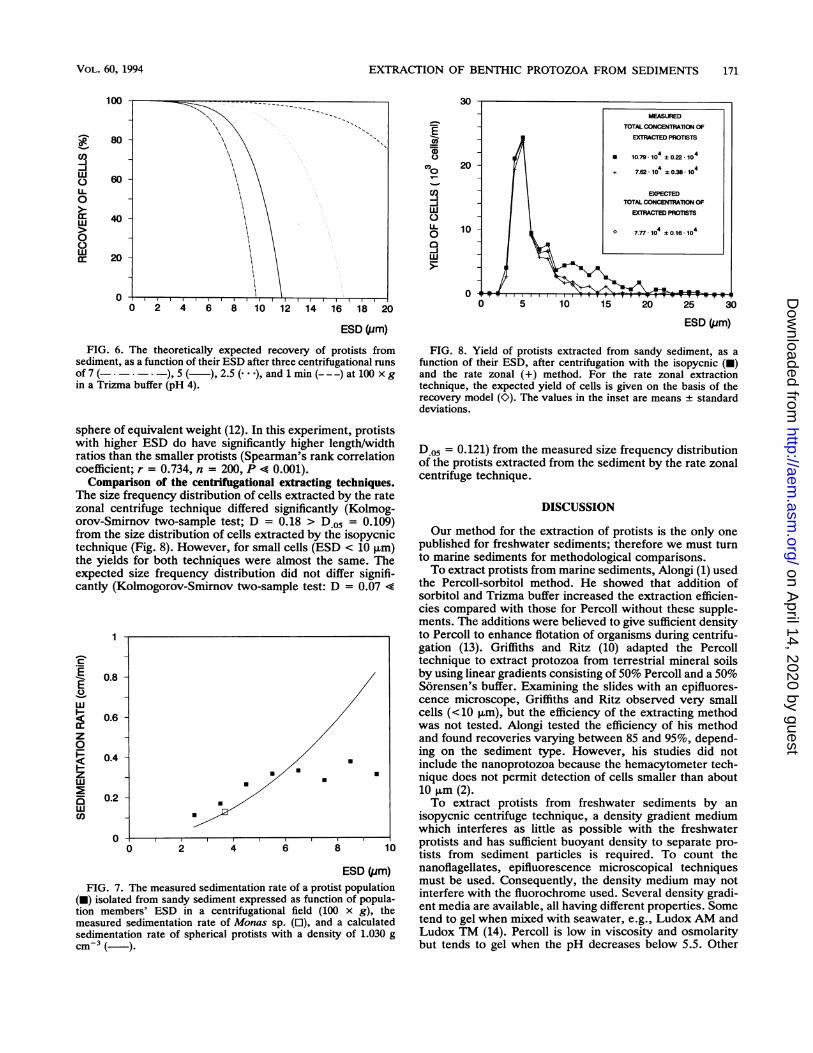
FIG. 6. The theoretically expected recovery of protists fromsediment, as a function of their ESD after three centrifugational runsof 7 ( ), 5 ( ,2.5 ( ,and 1 min (---)at 100 x gin a Trizma buffer (pH 4).
sphere of equivalent weight (12). In this experiment, protistswith higher ESD do have significantly higher length/widthratios than the smaller protists (Spearman's rank correlationcoefficient; r = 0.734, n = 200, P < 0.001).Comparison of the centrifugational extracting techniques.
The size frequency distribution of cells extracted by the ratezonal centrifuge technique differed significantly (Kolmog-orov-Smirnov two-sample test; D = 0.18 > D.05 = 0.109)from the size distribution of cells extracted by the isopycnictechnique (Fig. 8). However, for small cells (ESD < 10 ,um)the yields for both techniques were almost the same. Theexpected size frequency distribution did not differ signifi-cantly (Kolmogorov-Smirnov two-sample test: D = 0.07 4
1
-
EE
0I-
w
z
w
5wC,)
0.8 -
0.6 -
0.4 -
0.2 -
0
0 2 4 6 8
ESD (pm)FIG. 7. The measured sedimentation rate of a protist population
(-) isolated from sandy sediment expressed as function of popula-tion members' ESD in a centrifugational field (100 x g), themeasured sedimentation rate of Monas sp. (O), and a calculatedsedimentation rate of spherical protists with a density of 1.030 gcm-3( )
ESD (um)
FIG. 8. Yield of protists extracted from sandy sediment, as afunction of their ESD, after centrifugation with the isopycnic (-)and the rate zonal (+) method. For the rate zonal extractiontechnique, the expected yield of cells is given on the basis of therecovery model (K). The values in the inset are means ± standarddeviations.
D.05 = 0.121) from the measured size frequency distributionof the protists extracted from the sediment by the rate zonalcentrifuge technique.
DISCUSSION
Our method for the extraction of protists is the only onepublished for freshwater sediments; therefore we must turnto marine sediments for methodological comparisons.To extract protists from marine sediments, Alongi (1) used
the Percoll-sorbitol method. He showed that addition ofsorbitol and Trizma buffer increased the extraction efficien-cies compared with those for Percoll without these supple-ments. The additions were believed to give sufficient densityto Percoll to enhance flotation of organisms during centrifu-gation (13). Griffiths and Ritz (10) adapted the Percolltechnique to extract protozoa from terrestrial mineral soilsby using linear gradients consisting of 50% Percoll and a 50%Sorensen's buffer. Examining the slides with an epifluores-cence microscope, Griffiths and Ritz observed very smallcells (<10 ,um), but the efficiency of the extracting methodwas not tested. Alongi tested the efficiency of his methodand found recoveries varying between 85 and 95%, depend-ing on the sediment type. However, his studies did notinclude the nanoprotozoa because the hemacytometer tech-nique does not permit detection of cells smaller than about10 ,um (2).To extract protists from freshwater sediments by an
isopycnic centrifuge technique, a density gradient mediumwhich interferes as little as possible with the freshwaterprotists and has sufficient buoyant density to separate pro-tists from sediment particles is required. To count thenanoflagellates, epifluorescence microscopical techniquesmust be used. Consequently, the density medium may notinterfere with the fluorochrome used. Several density gradi-ent media are available, all having different properties. Sometend to gel when mixed with seawater, e.g., Ludox AM andLudox TM (14). Percoll is low in viscosity and osmolaritybut tends to gel when the pH decreases below 5.5. Other
-J
w
LL0a:w0wa:
.
VOL. 60, 1994
on April 14, 2020 by guest
http://aem.asm
.org/D
ownloaded from

172 STARINK ET AL.
media, such as the sucrose polymer Ficoll, are low inosmolarity but highly viscous. Sucrose and CsCl are highlyosmotic, which may result in dehydration of protists andincorrect estimates of their densities. The recoveries of cellsin Ficoll and sorbitol were in the same low order of magni-tude. High recoveries of cells were recorded in Percoll aswell as in sodium diatrizoate. We decided to use Percollbecause of its property of forming self-generating gradients.A disadvantage of Percoll is that fine crystals form when
the solution dries. These crystals tend to interfere withmicroscopic observations of the primulin-stained protists.Although no interference occurred if the protists werestained with DAPI (4',6-diamidino-2-phenylindole), wechose to use primulin. This facilitates distinguishing autotro-phic from heterotrophic forms (8). Working with cleanequipment (i.e., free of Percoll) reduced the amount ofcrystals significantly.The isopycnic extraction method we present is optimized
for sandy and gyttja-like sediments of freshwater systems.High recoveries of up to 100% extracted nanoflagellateswere recorded after the first extraction procedure. Thistime-saving one-step extraction procedure was made possi-ble by the use of nonlinear gradients in combination with ahigh centrifugal speed. At low centrifugal forces (500 x g for20 min), fine sediment particles did not sediment to theirequilibrium position in the gradient. Optimal separation ofparticles is only achieved when all particles have reachedtheir isopycnic position.
Percoll is often used because it is nontoxic and causes noappreciable osmotic damage to various organisms. Thisallows extraction of living cells from sediments as describedby Alongi (1). However, preliminary experiments showedthat up to 20% fewer nanoflagellates were recovered whenunfixed samples of a Monas sp. culture were loaded on aPercoll gradient even at the low centrifugal force of 500 x g.Schwinghamer (14) found a decreasing efficiency of theextraction procedure when the sediment/Percoll-sorbitolvolume ratio exceeded 0.25. We also found decreasingefficiencies when gradients were overloaded with gyttjasediment.As mentioned previously, sorbitol is commonly added to
impart sufficient density to Percoll. Because such supple-ments also increase the osmolarity of the gradient, weomitted this additive without observing lower yields. How-ever, for certain applications one might wish to increase thedensity of Percoll. This can be done by mixing Percoll withsodium diatrizoate without increasing the osmolarity signif-icantly.The rate zonal centrifugation technique is closely related
to the subsampling method of Bak and Nieuwland (3). Weenhanced the extraction procedure by centrifuging the sam-ples instead of using a normal gravitational field. The highestyield was obtained by centrifuging the samples for 5 min at100 x g three times. Pilot studies (unpublished) showed thatcentrifugal runs of 7 min and longer resulted in extremelylow yields; this can be expected from the recovery model.As shown (Fig. 3), the subsampling technique of Bak andNieuwland (3) resulted in lower yields than the centrifuga-tion technique because of masking of nonsedimented parti-cles. Calculations suggested that centrifuging samples for 5min at 100 x g three times should give results similar tothose from the subsampling method, in which samples areallowed to stand for about 8 h.The yield from the rate zonal centrifugation techniques
was highly dependent on dimensions of the cells, centrifugalforce, and the length of centrifugational time. We found a
recovery of 71% for a protist population with a mean ESD of7.9 ,um and of 100% for a population with a mean ESD of 4.8,um. The recovery model for the rate zonal extractiontechnique predicts such decreasing extraction yields. Therecovery of cells decreases when the ESD of the cellsincreases, and cells with an ESD of >12 ,um will not beextracted at all. However, because elliptical protists havelower sedimentation rates than spherical ones, we can con-sider the predicted recovery values as minimum values.Correcting the amount of extracted protists with the recov-ery values from the model is possible, but considering thelength/width ratio-dependent sedimentation rate and thenonextractable fraction of organisms with an ESD of >12,um, the corrected yields will still be underestimates.Although the performed recovery experiments have given
us insight into the properties of the extraction techniques,some remarks have to be made. In natural samples, someprotists may, even after fixation, still be attached to particlesand therefore they will not be extracted from the sediment.This would result in underestimation of the real protistdensities in the sediment. However, our Percoll method gavevery high yields of protists extracted from natural samplesfor which other techniques failed, and the rate zonal tech-nique appeared superior to the subsampling method of Bakand Nieuwland (3). Therefore, we believe that both methodsare reliable for the study of the role of protists in benthicenvironments.
In comparing the rate zonal and isopycnic extractiontechniques, the total yield for the former technique was 30%lower, indeed mainly because of the nonextractable fractionof larger protists (ESD >12 p,m). This decreased extractionefficiency is in agreement with the predictions of the model.
Studies on tidal flats (3) showed that approximately 55% ofthe protists in the sediment water interface are smaller than5 ,um (after fixation). Deeper in the sediment, this percentageincreased to 70%. Our research in freshwater sandy sedi-ments indicates that the majority (ca. 65%) of protists (afterfixation) were smaller than 5 ,um and about 88% were smallerthan 10 p,m in the upper part of the sediment (0 to 0.5 cm).Although the size-dependent recovery of the rate zonalmethod is a disadvantage, in practice one measures reliabledensities of protists because of the dominance of the smallerprotists.
In conclusion, we have two methods. (i) There is theisopycnic centrifuge technique with Percoll, a reliablemethod for extracting protists from sandy and gyttja-likesediments, which permits the detection of small flagellates.The yield is independent of the size distribution of theprotists. (ii) For sediments rich in fresh particulated organicmaterial, there is the rate zonal centrifuge technique. Itshould only be applied when extraction of protists with theisopycnic centrifuge technique fails, because of the size-dependent yield.
ACKNOWLEDGMENTS
We thank F. M. Ellenbroek and two anonymous reviewers fortheir comments on an earlier version of the manuscript and S. Visserfor collecting the field samples.
REFERENCES1. Alongi, D. M. 1986. Quantitative estimates of benthic protozoa
in tropical marine systems using silica gel: a comparison ofmethods. Estuarine Coastal Shelf Sci. 23:443-450.
2. Alongi, D. M. 1990. Abundances of benthic microfauna inrelation to outwelling of mangrove detritus in a tropical coastalregion. Mar. Ecol. Prog. Ser. 63:53-63.
APPL. ENvIRON. MICROBIOL.
on April 14, 2020 by guest
http://aem.asm
.org/D
ownloaded from

EXTRACTION OF BENTHIC PROTOZOA FROM SEDIMENTS 173
3. Bak, R. P. M., and G. Nieuwland. 1989. Seasonal fluctuations inbenthic protozoan populations at different depths in marinesediments. Neth. J. Sea Res. 24:37-44.
4. Bak, R. P. M., F. C. Van DuyI, G. Nieuwland, and A. J. Kop.1991. Benthic heterotrophic nanoflagellates in North Sea field/mesocosm bottoms and their response to algal sedimentation.Ophelia 33:187-196.
5. Bloem, J., C. Albert, M.-J. B. Bar-Gilissen, T. M. Berman, andT. E. Cappenberg. 1989. Nutrient cycling through phytoplank-ton, bacteria and protozoa, in selectively filtered Lake Vechtenwater. J. Plankton Res. 11:119-131.
6. Bloem, J., M.-J. B. Bair-Gilissen, and T. E. Cappenberg. 1986.Fixation, counting, and manipulation of heterotrophicnanoflagellates. Appl. Environ. Microbiol. 52:1266-1272.
7. Bloem, J., F. M. Ellenbroek, M.-J. B. Bar-Gilissen, and T. E.Cappenberg. 1989. Protozoan grazing and bacterial productionin stratified Lake Vechten estimated with fluorescently labeledbacteria and by thymidine incorporation. Appl. Environ. Micro-biol. 55:1787-1795.
8. Caron, D. A. 1983. Technique for enumeration of heterotrophicand phototrophic nanoplankton, using epifluorescence micros-copy, and comparison with other procedures. Appl. Environ.Microbiol. 46:491-498.
9. Fenchel, T. 1982. Ecology of heterotrophic microflagellates. IV.Quantitative occurrence and importance as bacterial consum-ers. Mar. Ecol. Prog. Ser. 9:35-42.
10. Griffiths, B. S., and K. Ritz. 1988. A technique to extract,
enumerate and measure protozoa from mineral soils. Soil Biol.Biochem. 20:163-173.
11. Hondeveld, B. J. M., R. P. M. Bak, and F. C. van Duyl. 1992.Bacterivory by heterotrophic nanoflagellates in marine sedi-ments measured by uptake of fluorescently labeled bacteria.Mar. Ecol. Prog. Ser. 89:63-71.
12. Kunkel, W. B. 1948. Magnitude and character of errors pro-duced by shape factors in Stokes' law estimates of particleradius. J. Appl. Physiol. 19:1056-1058.
13. Price, C. A., E. M. Reardon, and R. R. L. Guillard. 1978.Collection of dinoflagellates and other marine microalgae bycentrifugation in density gradients of a modified silica sol.Limnol. Oceanogr. 23:548-553.
14. Schwinghamer, P. 1981. Extraction of living meiofauna frommarine sediments by centrifugation in a silica sol-sorbitol mix-ture. Can. J. Fish. Aquat. Sci. 38:476-478.
15. Sherr, B. F., E. B. Sherr, and T. Berman. 1982. Decompositionof organic detritus: a selective role for microflagellate protozoa.Limnol. Oceanogr. 27:765-769.
16. Sieburth, J. M. 1984. Protozoan bacterivory in pelagic marinewaters, p. 405-444. In J. E. Hobbie and P. J. L. Williams (ed.),Heterotrophic activity in the sea. Plenum Publishing Corp.,New York.
17. Sokal, R. S., and F. J. Rohlf. 1981. Biometry. Freeman and Co.,San Francisco.
18. Uhlig, G. 1968. Quantitative methods in the study of interstitialfauna. Trans. Am. Microsc. Soc. 87:226-232.
VOL. 60, 1994
on April 14, 2020 by guest
http://aem.asm
.org/D
ownloaded from