Embed Size (px)
Citation preview

UNIVERSIDAD COMPLUTENSEDE MADRIDFACULTAD DE VETERINARIA
DEPARTAMENTO DE PATOLOGíA ANIMAL 1
ESTUDIO DE LA INFLUENCIA DE LA DEFICIENCIA EN SELENIOY VITAMINA E EN LA LISTERIOSIS MURINA.
JOSEANTONIO GARCíA CABRERA
MEMORIA PRESENTADA PARA OPTAR AL TITULO DEDOCTOR EN VETERINARIA
Directoresdc a tesisDr. D. Lucas Do,ní,,gueiRodríguezDr. 1). Victor BrionesDiesteDr. 1). JoséFranciscoFernández-Garayzábal
Madrid, 1995

úD~ [OS ¿icffes y fiunzifdesp uede.n salir [ossantos,pocasveces[ossabios.
Santiago£Pyanióny Cajal

~$1Ana
A mis paifres

AGRADECIMIENTOS
Cuando uno llega al final de un camino tan sólo se recuerdan los buenos momentos, y se
tiende a olvidar todas las dificultades y malos tragos que se han tenido que pasar. Sin embargo, no
quisiera olvidarlos sin recordar antes a todos los que, de alguna manera, han colaborado a que este
trabajo haya podido ver la luz.
A los tres directores de la presente Tesis:
Al Dr. Lucas Domínguez por numerosas razones imposibles de enumerar, por la paciencia que
ha demostrado conmigo, su comprensión ante mi desánimo y su preocupación por mí, que ha
ido más lejos de lo exigible a un director por su pupilo.
Al Dr. Víctor Briones por compartir tantos sacrificios, así como por su amistad.
Al Dr. José Francisco Fernández-Garayzábal, porsus acertados consejos y enfoques durante
la realización del presente trabajo.
A D~. Ana Doménech, porque si valoro en mucho su capacidad de trabajo y rigor en el
laboratorio, valoro aún más su voluntad y capacidad de sacrificio fuera de él.
Al Dr. Guillermo Suárez por sus consejos, y su ejemplo de dedicación a la Universidad.
Al Dr. Joaquín Goyache, porque siempre ha estado ahi, incluso en los momentos más críticos.
A los Drs. José Luis Blanco y Marta García, por tanto tiempo perdido conmigo.
A las Dras. María Teresa Cutu¡i, Ana Mateos y M~ Jesús Payé, por ofrecerme sus consejos
y su ayuda desinteresada.
A la Dra. Esperanza Gómez-Lucía, por su carácter jovial y por tener siempre una palabra
amable.
Al resto del equipo listerias, Dra. Mar Blanco, compañera de fatigas durante todo este tiempo:
Dr. José Antonio Vázquez-Boland, ejemplo de combatitividad, incluso en inferioridad aquí y en Berlín;
Dras. Alicia Gibello y Mónica Suárez, que aunque han sido las últimas incorporaciones no han
regateado esfuerzos para ponerse a la altura del resto; D~. Mayte Ripio y D. Gustavo Domínguez, claro
ejemplo de que se puede disfrutar de la vida aunque sea en el laboratorio; D~. Belén López, toda una

Al resto de compañeros de Microbiología del Departamento de Patología Animal 1, Dr. Miguel
Angel Moreno, porque ha sabido trasmitir su carácter organizativo a todo aquello en lo que interviene;
0. Ignacio Septién, porpresentarme al “Orfeón Burgalés”; D8 Alicia Aranaz y D. Ernesto Liébana, “tanto
monta, monta tanto...”; D. Louie LLames, por haber sabido apreciar lo mejor de las costumbres
españolas; 08. Mercedes García, por saber aclimatarse a cualquier situación; D. Félix Fernández y 08.
Pilar Capón, porque siempre han estado prestos a ayudar.
Al Dr. Ricardo de la Fuente, por su ejemplar actitud.
A los Dra. José Antonio Orden y José Antonio Ruiz-Santa Quiteria, compañeros de “cubículo,
dispuestos a cualquier cosa para mejorar la convivencia.
Al resto de integrantes de Patología Infecciosa del Departamento de Patología Animal 1, en la
que me encuentro perfectamente integrado.
A los restantes miembros del Departamento de Patología Animal 1, con los que la convivencia
diaria es todo un placer.
Al Dr. Javier Cañón, del Departamento de Producción Animal, por su inestimable ayuda en
el estudio estadístico.
A los Drs. Mariano Domingo y Alberto Marco, así como a D~. Neus Prats ya D. Jaume Altimira,
del Departamento de Anatomía Patológica e Histología de la Facultad de Veterinaria de la Universidad
Autónoma de Barcelona, sin cuya colaboración la parte histológica de este trabajo hubiera tenido
mayores dificultades. A los tres últimos, además por una cena “inolvidable” en Madrid.
A todos aquellos que, por un imperdonable error, han quedado fuera de esta relación, muchas
gracias.

INDICE

Indice
INTRODIJCCION
1.1- BREVE RESEÑA HISTORICA
1.11- TAXONOMíA
lila- TAXONOMÍA INTERGENERICA
l.II.b- TAXONOMíA INTRAGENERICA
1.111- ESTRUCTURA ANTIGENICA
¡.1V- EPII)EMIOLOGIA DE LA LISTERIOSIS
IIVa- RESERVORIOS
1.IV,b- VíAS DE TRANSMISION
1 IVe- MARCADORES EPIDEMIOLOGICOS
1V- ENFERMEDAD
IVa- PENETRACION EN EL HOSPEDADOR
1.V,b- CINETICA DE LA INFECCION Y RESPUESTA
[VI- FACTORESPREDISPONENTES
IVII- FORMAS CLíNICAS
IViIa- MENINGITIS Y ENCEFALmS
I.VII.b- INFECCION MATERNO-FETAL
1 ,VII,c- INFECCIONESSEPTICEMICAS
1.VII.d- OTRAS FORMAS CLíNICAS
1 VIII- DEFICIENCIAS NUTRICIONALES Y ENFERMEDAD
1 IX- IMPORTANCIA BIOLOGICA DEL SELENIO
1 IXa- ACCIONES BIOLOGICAS. ENZIMAS.
1 .IX.b- TOXICIDAD DEL SELENIO
1,IX.c- DEFICIENCIA EN SELENIO
I.IX.d- SELENIO Y SISTEMA INMUNE
INMUNE
l.IX.d.1- Influencia sobrela función fagocítica
1.IX.d.2- Influencia sobrela producciónde
anticuerpos
1.IX.d.3- Influencia sobrela función secretora
de las célulasdel sistemainmune
1.IX.d.4- Influencia sobrela capacidad
proliferativadc los linfocitos T y E
1.X- SELENIO Y RESISTENCIAA ENFERMEDADES INFECCIOSAS
2. OBJETIVOS
3. MAtERIAL Y METODOS.,,.,
3.1- MATERIAL
3.11- MEDIOS DE CIJETIVO
3.111- 501.1ICIONES
3.1V- CEPAS
2
3
6
9
12
12
14
15
18
¡8
21
24
28
28
29
30
31
32
36
36
39
41
43
45
47
49
50
52
55
58
59
59
60
62

Indice
3.V- ANIMALES DE EXPERIMENTACION
3.VI- ALIMENTO Y AGUA DE BEBIDA
3.VI,a- RATONES NORMALES
3.VI.b- RATONES DEHCIENTESEN SELENIO Y VITAMINA E
3.VII- OBTENCION DEL INOCULO
3.VIII- INOCIJLACIONES
3 VIlla- INOCULACIONES ENDOVENOSAS
3,VIII.b- INOCULACIONES ORALES
3.IX- SACRIFICIOS Y OBTENCION DE LOS ORGANOS
3.X- CALCULO DE LA DOSIS LETAL 50 (DL5O) PORVIA
ENDOVENOSA
3.XI- ESTUDIOS DE MORTAnDAD POR VíA ORAL CON DOSIS UNICA
3.XII- ESTUDIODE MORTALIDAD POR VíA ORAL CON DOSIS
REPETIDAS
3.XIU- SEGUIMIENTO DE LA INFECCION POR VíA ENDOVENOSA..
3.XIV- ESTUDIO DE LA INFECCION PORVíA ORAL
3,XIV.a- SEGUNLA VíA DE INOCULACION EMPLEADA
3.XIV.b- SEGUN LA CEPA EMPLEADA
3.XIV.c- SEGUNDOSIFICACION UNICA O REPETIDA
3.XV- ESIUDIO DE LA INFECCION POR VíA ORAL EN RATONES
DEFICIENTES/NODEFICIENTES
3.XV.a- SEGUIMIENTO DE LA INFECCION POR VíA ORAL
CON DOSIS UNICA
3,XV.h- SEGUIMIENTODE LA INFECCION POR VíA ORAL
CON DOSIS REPETIDAS
3.XVI- ESTUDIO IIISTOPATOLOGICO
3.XVI.a- PREPARACION DE LAS MUESTRAS
3.XVI.b- ESTUDIO HISTOLOGICO
3.XVII- DETERMINACION DE LA CONCENTRACION DE SELENIO
3.XVIII- VALORACION DE LA DEFICIENCIA EN SEI.,ENIO EN
FUINCION DEL GRADO DE HEMOLISIS
3.XIX- ESTUDIOESTADíSTICO
.4 RESULTADOSY DISCIJSION
4,1- VALORACION DE LAS CONCENTRACIONESDE SELENIO EN
1-lIGADO
4.11- ESTUDiOSDF. MORTALIDAD
4.II,a- DL50 VíA ENDOVENOSA
4.II,b- MORTALIDAD POR VíA ORAL CON DOSIS UINICA Ql
63
63
63
63
64
66
66
66
67
71
72
73
73
76
76
76
78
78
78
81
81
81
83
83
84
84
86
87
91
Ql
II

Li’dice
4.II.c- MORTALIDAD PORVíA ORAL CON DOSIS REPETIDAS. 94
4.111- SEGUIMIENTO DE LA INFECCION POR VíA ENDOVENOSA 96
4.lli.a- DOSIS io4-5 .,... 96
4mb- DOSIS i0~5 101
4.1V- SEGUIMIENTO DE LA INFECCION POR VíA ORAL CONDOSIS
UNICA 105
4.IV.a- CON Listeria ,oonocvtogenes4b ..... 105
4.IV.h- CON Listeria monocytcgenes112a 119
4.IV.c- COMPARACION DE AMBAS SEROTIPOS 119
4V- SEGUIMIENTODE LA INFECCION POR VTA INTRAGASTRICA , 123
4.VI- COMPARACION DE LAS VIAS ORAL-INTRAGASTRICA 127
4.VII- SEGUIMIENTODE LA INFECCION POR VíA ORAL CON
100 7 DíAS ..,............ 128
4.VIII- DEHCIENCIA EN SELENIO Y VITAMINA E Y
SUSCEPTIBILIDAD A LA LISTERIOSIS 132
5. CONCLUSIONES 137
6. RESUMEN 140
7. BIBLIOGRAFIA 142
III

1. INTRODUCCION

1 utroducción
1.1- BREVE RESEÑAHISTORICA
La historia del género Listeria es relativamentereciente, ya que arrancade la
descripciónque Murray, Webb y Swann hacendel microorganismoresponsablede una
epizootiaentre los conejosy cobayosde su laboratorioen 1924 (Murray et aL, 1926). Al
considerarque la bacteriaaisladaporellosno habíasido descritaaúndecidenbautizaríacomo
“Bacteriurn monocytogenes’,al cursarel procesoinfeccioso,como signo máscaracterístico,
con una intensamonocitosis.
Sin embargo,parecequeestosautoresno fueron los primerosen encontrarsecon L.
monocytogenes.En 1911 GustavHtilphers habíaaisladoun microorganismomuy similar a
partir de lesionesnecróticasde hígadode conejo; desgraciadamentesus cultivos no fueron
conservadosy, por tanto, no pudieronsercontrastadoscon los de Murray, Webb y Swann.
Porotra parte.en 1921 Dumonty Cotonidescribenun microorganismoparecidoa “Bacterium
rnonocvtogenes”,aisladoa partir de un casode meningitishumana.Estos autoresunicamente
señalaronquesetratabade unabacteriamuyparecidaa la productoradel Mal Rojo del cerdo.
Este descubrimientopemanecióignorado durantemuchos años, al no ser colTectamente
interpretado,por lo queseconsiderancomoauténticosdescubridoresdel géneroa los autores
citadosen primer lugar.
Posteriormente,Pide aislauna bacteriagram-positivaa partir de un broteepizoótico
en gerbos(Tatera lobenguiae)duranteel cursode unasinvestigacionessobrela pestehumana
en Sudáfrica. Al no encontrarreferenciaalguna sobre la enfermedad,cuya lesión mas
importanteconsistíaen la existenciade focosnecróticosen hígado,ni sobreel agentecausal
propuso denominarlasrespectivamente“Tiger River Disease’ atendiendoa la localizacion
geográficaen que seproduce(zonadel río Tiger, en el estadolibre de Orange),y “Listerella
hepatolytica’ (Pirie, 1927) en honor a Lord Lister.
A partir de ese momento se suceden las aportacionesque involucran a este
microorganismoen la etiologíade diversosprocesosinfecciosos.Así, Nyfeldt (1932) la aísla
a partir de sangrede individuosenfermoscon un cuadroclínico de mononucleosis(‘Listerella
monocvtogeneshominis”), relacionándolapor primeravezcon un cuadroinfecciosohumano.
PocosañosdespuésBurn establecelos descuadrosprincipalesde la listeriosishumana,al
2

Introduccion
aislarlaen 1933 de casosde septicemianeonatal,y en 1935 de cuadrosde meningoencefalitis
fatal en adultos.Sin embargo,los trabajosde Burn (1935, 1936) fueron prontoolvidados,ya
que en 1951 Reiss, Potel y Krebs publican sus hallazgos sobre la “granulomatosis
infantiseptica” de la que responsabilizana “Corynebacrerium infanriseprica”, que. sin
embargo,no eraotra que L. rnonocytogenes tal y como demostróSeeligerposteriormente
(Seeliger, 1952). Peroestosautoresno fueron los primerosen confundir L. nionocvtogenes
con unacorinebacteria,en 1934 Schultz et al. describencomoagenteresponsablede cuadros
de meningoencefalitishumanaa “Corynebacreriumparvulum”.
Con lo expuestohasta ahora, debemosestar de acuerdocon las afirmacionesde
Vázquez-Boland(1989),segúnlas cualesla listeriosis neonatalen fonria de “granulomabosís
infantiseptica” habría sido observadacon anterioridad a la descripción de Burn ya
mencionada.Estos cuadroshabrían sido recogidos bajo la denominaciónde “septicemia
neonatal’ y se habríaresponsabilizadode ella a “bastoncillosgram-positivoso argentófilos’.
sin que sellegasea establecersu correctaetiología.
Las primerasaportacionesen relación con la listeriosis dentro del mundo de la
Veterinaria,son debidasa GuI (1933, 1937). Esteinvestigadorfue el primeroen relacionar
una afeccióndel sistemanerviosode las ovejas,la llamada“circling disease’o torneo (hoy
por extensióntodaafeccióndel sistemanerviosocentralde los rumiantespor Listeria) con
un microorganismoal que denomina“Ltnerella ovis”.
Por fin, en 1940Pirie proponecomo nuevonombredel géneroel de Listeria ya que
el de ‘Listerella” ya sehabíaempleadocon anterioridadparadesignara un génerovegetal.
1.11- TAXONOMIA
1.I1.a- TAXONOMIA INTERGENERICA
Listeria es un género formado por bacilos gram-positivos,mesófilos, aerobioso
anaerobiosfacuhativos,no capsuladosni formadoresde esporos,catalasapositivos, no
miceliaresy productoresdel ácidoL(-i-)láctico en el metabolismofermentativode la ghícosa.
Por todo esto,esfácil suponerquelas relacionestaxonómicasintergenéncasde ListeRohayan
3

Introducción
sido productode gran controversiaduranteaños.
En la 7~ edición del Bergey’s Manual (Breed et aL, 1957) se incluyen dentro de la
familia Corynebacteriaceae,con el géneroCorynebacteriurncomo génerotípico, atendiendo
a sus característicasmorfológicas. Dentro de esa misma familia se incluía el género
Erysipelothrix,con el que algunosautoresconsiderabanmuy relacionadasa Listerio (Jones,
1975). Posteriormente,y atendiendoa estudios de taxonomíanumérica(Davis y Newton,
1969; Davis er aL, 1969; Stuarty Pease,1972; citadospor Vázquez-Boland,1989; Stuarty
Welshimer,1973, 1974)y de composiciónquímicade la pared(Cumminsy Harris, 1956)se
demostróque las listeriaserandistintas a las corinebacterias,y que incluso no estabantan
relacionadascon Erysipelorhrixcomo secreíaen un principio. Segúnestosestudios,parecían
másemparentadascon Staphylococcus,Racillus,Kurthia y, sobretodo, Brochotrix.Por todo
ello, en la 8~ edición del Bergey’sManual (Seeligery Welshimer, 1974) el géneroListeria
apareceseparadode la familia Corynebacteriaceaey esincluido entrelos génerosde filiación
inciertajunto con Erysipelothrix, Ca.ryophanony la familia Lactobacillaceae,mientrasque
BroChotrix (poraquelentoncesMicrobacteriurnrherrnophactum)y Kurthia sí semantuvieron
dentrode la familia Corynebacteriaceae.
Esta separaciónde las corinebacteriasfue confirmadapor posterioresestudiosde
taxonomía numérica (Jones, 1975; Wilkinson y Jones, 1977; Jones et aL, 1986),
quimiotaxonomia(Tadayony Carrol, 1971; Srivastavay Siddique, 1973; Kamisangoet aL,
1982; Collins y Jones,1981; Fiedíery Seger,1983; Fiedíeretat, 1984;citadosporVázquez-
Bolanó, 1989; Joneset al., 1979, 1986), composicióndel AUN e hibridación AUN-AUN
(Stuarty Welshimer, 1973, 1974, citadosporVázquez-Boland,1989; Rocourt et al., 1982).
Estos estudiosconfirmaron a su vez la estrecharelación entre Listeria y Lactobacillus,
Streptococcus,Enterococcus,Lactococcus,Staphylococcus,Racillusy Kurthia, observándose
la máximaproximidadconBrochotrix(Kandlery Weiss,1986; Seeligery Jones,1986;Jones,
1988). Géneroscaracterizadostodos ellos por poseerun bajocontenidode G+C en su AUN
(36-42%),por no tenerácidosmicólicospero sí lipoteicoicos,por tenerpeptidoglicanode la
variedad Al gammaasociadoa ácidosteicoicos del tipo polirribitol-fosfato, y por poseer
comomenaquinonaprincipal la MK 7 (Joneset al., 1972; Ruhlandy Fiedler, 1987; Fiedler,
1988,; citadospor Vá-zque-z-Boland,1989; Seeligery Jones,1986).
4

~ete~ia—
ac,:cac:¡Ius:7:u:
— vrerc::z
e05
oo51
6-05o5
e’o
eaoe’c
Mee:otasr¿S—
•ar:ctjs 2 •. ¡res—
o
oa‘eo
.3-.-
c<~o
.0’‘Ú~ </7/7 <‘
‘5 ,/ -¼,.,
- - . 3..
Figura 1- Dendrogramasimplificado, enel quese puedenobservarlas relacionesfilogenéticasdelgéneroListeria conotrasbacteriasgranipositivas(tomadodeJones,1988>.
NiotO. úxvgen USOSVTS 13.05 STO.COuire meno , - .. ase
5.; ,~ duwcn
AC:a15005
ose
2~550. NíainoiePtao. Níasor
,%~3fl .a5’&0t05, !V.5tV¿-
3010 2tnone
Srorhorhrsx — Faculsarove — -r — — A meso-DA? XIX- 7 S.l. -
zr’sioesothr:x — Facuitauve — — — — E L-Lvs¡ne — 5. A. ¡ L o—-OPacuicarive — — — — ..-. meso-DA? XIX-’ 5. A. 5 n 5
— Pacuirarove -- — — — A. Lvssneor — t.
‘sieso-DA?
or orojíhine
Xurjo:a — Aerob,c — — — — A L-LVSiaC XIX- 7 5. A. s 6.. .9
Tabla 1- Caracteresdiferencialesde los génerosBrnchorhrix, Erysipelothrix,Lactobacillus,Kurtl,ia y
Listeria (tomadode Seeligery Jones,1986)
.55<00

Introducción
Todos estos estudios concuerdancon los de base filogenética (catalogación de
secuenciasde oligonucleótidosdelARN 16s) segúnlos cualesListeria seencuadradadentro
de la rama Cíostridium-Lactobacilius-Bacillus(porcentaje molecular de G+C < 55%)
(Stackerbrandty Woese,1981 , citados porVázquez-Boland,1989),con una relación más
estrechacon Brochotrix, y ocupandocon ésteuna posición intermediaentreBacillus y el
grupoLoctobaciliuslStreptococcus(Stackerbrandtet aL, 1983; Ludwig et aL, 1984; citados
por Vázquez-Boland,1989; Rocourtci aL, 1982; Kandler y Weiss, 1986).
1.II.b- TAXONOMLX INTRAGENERICA
Durantemuchosañosel géneroListeria estuvoconstituido poruna únicaespecie,L.
~nonocytogenes,en la que seencontrabanincluidastodaslas cepasaisladas.Estoconlíevóque
cepashemolíticasy patógenasseencuadraranjunto con cepasno hemolíticasy apatógenas,
con la consiguienteconfusión a la hora de intentarextraerconclusionesde tipo ecológico-
epidemiológico de muestras no clínicas (Seeliger, 1976). Además se observó que L.
rnonocytogenes‘sensu lato” tal y como estabadescritaen la ga edición del Bergey’sManual
(Seeligery Welshimer,1974) resultabaun taxónheterogéneo,no sólo desdeel punto de vista
de poder hemolítico y patógeno,sino incluso en cuanto al patrón de acidificación de
determinadosazúcares,llegándosea demostrarque estas diferencias no se limitaban a
aspectosfenotípicos,sino que existíandiferenciasgenómicas(Stuart y Welshimer, 1973).
Así, de las que actualmentecomponenel género,la primera especiedistinta dc L.
monocvtogenes propuestafue L. innocua(Seeliger, 1981) en la que se incluyeron todas las
cepasno hemolíticasy no patógenasque presentaranlos factoresantigénicosO-XI y O-XV.
y que hubieransido aisladasa partir de portadoressanoso de fuentesambientales(Bojsen-
Moller, 1967; citadosporVázquez-Boland,1989;Welshimer,1968;Ralovich, 1975; Seeliger,
1975, 1976; Seeligery SchooÑ,1979). Por otro lado, se propusola inclusión de las cepas
aisladaspor Ivanov en Bulgariaen 1955 a partir de ovejascon listeriosis (Ivanov, 1962)en
otraespeciequesedenominada“L. bulgarico” (Ivanov,1975)o “L perhaemolvtica” (Seeliger
ci aL. 1982, citado por Vá-zquez-Boland,1989).Estascepasse caracterizabanpor pertenecer
específicamentea un único serovar,el 5, por presentarun marcadoefecto hemolíticobizonal
en agar sangre,y por ser patógenasexclusivamentepara el ganadoovino (lvanov, 1957,
1962).Estascepasfueron caracterizadasposteriormentepor el mismo autor (Ivanov, 1975).
6

Introducción
Posteriormente,Rocourt et aL (1982) propusieronla división de L. rnonocvtogenes
‘sensu lato” en cinco grupos genómicosdistintos, medianteensayosde hibridación AUN-
AUN. Estoscinco gruposserían:
- grupo 1- constituidopor las cepas“clásicas’ de L. nwnocvtogeneshemolíticas y
patógenas(L. ¡nonocytogenes‘sensu stricto”) en el que se incluirían cepasde los
serovares112a, 1/2b, l/2c, 3a, 3b, 3c, 4a, 4ab, 4b, 4c, 4d y 7;
- grupo 2- en estegrupo se incluirían las cepashiperhemolíticasy patógenasdel
serovar5;
- grupo3- aquíseencuadraríanlas cepasno hemolíticasy apatógenasde los serovares
4ab, 6a y 6b;
- grupo4- en estegrupoquedaríanencuadradaslas cepas“americanas’de L. inflOCua
descritaspor primeravezen Norteamérica,y quese diferenciande las ‘europeas’por
presentarreacciónpositiva a la xilosa (Grovesy Welshimer,1977; Rocouí’t y Seeligeí;
1985);
- grupo 5- este grupo quedaría reservadopara cepas debilmente hemolíticas y
apatógenasde distintos serovares.
Al observarque cadauno de estosgrupospodíanserconsideradoscomo una especie
distinta sc ha consideradola división de L. mcnocytogenes“sen9u lato’ en cínco especies
distintas:
- grupo 1- L. inonocvtogenes
- grupo2- L. ivanovil (nombredefinitivo de 1. bulgañea’ (Seeligeret aL, 1984))
- grupo 3- L. innocua
- grupo4- L. welshimer,
- grupo5- L.seelige¡-i
A estascinco especieshabríaqueañadirotrasespeciesdescritasindependientemente,
L. gravi (Larsen y Seeliger, 1966, citados por Vázquez-Boland,1989) y L. ,nurrav,
(Welshirnery Meredith, 1971),que a pesarde la controversiaque su inclusión dentrode este
géneí-o despenéen un principio, llegándoseincluso a pretendersu exclusión del mismo
(Stuart y Welshimer, 1974), hoy son consideradascomo miembros de un mismo taxón
homogéneo,queconfirmalassimilaridadesfenotípicasy genotípicasencontradas,y querecibe
7

00
o>e’
‘oe->cee->e->-e:
o‘e00—
v+
++
+
ce
+‘S
S
ce00o+
+u
+
ce•~
ce
‘Oc
s~
‘ece-e00e’o>---.0+
.~
‘ece‘e00e
’
e’-—
o)+
te)=
-_
‘ece‘e00e
’
e’
.0‘0)‘e
“-.500
e.>
e’
+
-4-
+
e’
ceC
iD++
uu
eee’e’-o.’.
e)
o.)~,
~
00o>e)
0.0ooE2
e’
.~2
¡¡.t002
e)-~
Q.e
’00~.>
<2
‘05
ce
Ee).0‘ee)ceQee
’c~
-o—
-uceo.>
—Q
e)‘e
’
0)
n-t
—e)
-u~
Qe‘O
LZ
ct
--o
-
e)
ooo00
5
-OC
-
00
5
0~
>E
‘eoe
E-
o-t
oe
s
NO
0>0
-

Ini roducción
el nombrede L. grayi (Rocourtu al., 1992).
Mención apartemerecela primeraespeciedescritadespuesde L. nwnocvtogenes,L.
denitrificans. Su inclusión dentro del géneroListeria fue cuestionadadurantemuchosaños,
hastaque recientesestudiosde secuenciacióndel ARNr lás demostraronsu pertenencia
filogenética a la subdivisióncorineformes-actinomicetos,bacteriasgrarn-positivasde alto
contenidoen G+C (>55%), y ha sido transferidaa un nuevo génerocon la denomínaciónde
Jonesiadenitlficans(Rocourtu aL, 1987; Rocourt, 1988).
Paraconcluir estecapítulo,y resumiendotodo lo expuestohastaaquí, debereseñarse
queel géneroListeria constituyeun grupohomogéneoquimiotaxonómicamente,formadopor
7 especiesquetienenen comúnun contenidoen G÷Cde 36-42%,poseenun peptidoglicano
Aly asociadoa ácidosteicoicosdel tipo polirribitol-fosfato, carecende ácidos micólicos,
poseenácidos lipoteicoicos y presentancomo menaquinonaprincipal la MK 7. Estas7
especiespuedendividirse en dos gruposdesdeel punto de vista filogenético, en vino se
encuadraríanL. grayi (incluyendo a L. rnurravi), y en el otro las cinco especiesrestantes
(Rocourt, 1988).
1.111- ESTRUCTURA ANTIGENICA
Los estudiosserológicoshan sido una herramientaampliamenteutilizada en la
clasificaciónde numerososgruposbacterianos,no siendoListeuia una excepcióna esto. Es
más, la estructuraantigénicaen estegéneroha sido el origen de una de las más aceptadas
clasificacionesbasadasen criterios serológicos.
El primeroen realizarestudiosserológicoscon distintascepasde Listenafue Seastone
(citado por Domínguez,1982), quien encontróuna estrecharelación entrecepasde origen
humano y cepasde origen animal, a excepciónde la cepa tipo original de Murray que
reaccionabade formadistinta. Tambiénencontraronuna gransimilitud en el comportamiento
serológicoentrelas distintascepasde Listería Webh y Barberen 1937 (citadosporVázquez-
Boland, 1984).
Utilizando un gran númerode cepasprocedentesde GranBretañay EstadosUnidos
9

lníroduccion
Seastoneintentó estableceruna relación entre las diferenciasantigénicasde los grupos
serológicosy el origen de las mísmas.Deestamanera,seguíael ejemplode lo realizadocon
el géneroSainwnella,dondesehabíaencontradouna relaciónserotipo-hospedador,relación
que hoy se sabees una excepciónen estegénero.Desgraciadamente,las cepasbritánicas
proveníande roedoresy las americanasde rumiantesen su mayoría. Así, dadoque las cepas
europeaspertenecíana un grupo serológico(serovar1) y las americanasa otro (serovaí~4),
Schultzpor un ladoy Julianelley Ponspor otro (citadospor Vázquez-Boland,1984) llegaron
a la conclusiónde que las listeriasdebíandividirse en dos gruposserológicosdistintos,que
estarían relacionadoscon el origen de la cepa (grupo Roedoresy grupo Rumiantes).
estableciendo,por tanto, una clasificación sero-ecológica.Las cepas de Listeria de
procedenciahumanaperteneceríanindistintamentea uno o a otro grupo. Sin embargo,
Patersonen 1939 y 1940 (Domínguez,1982) consiguióidentificar serologicamentedistintos
modelos antigénicossomáticos (O) y flagelares (H) usando los mismos métodos que
Kauffman y White habíanutilizado parala clasificacióndel géneroSalnwnella, paralo que
empleó54 cepasde listerias. De estamaneraestablecióla base de la actual clasificación
serológicadel géneroListeria. Con los resultadosobtenidos,esteautordividió la especieL.
rnonocytogenes en cuatroserotipos,demostrandoademásla no existenciade relaciónalguna
entreserotipoy hospedador.De los cuatroserotiposencontrados,el 1, 3 y 4 sediferenciaban
por sus antígenossomáticos;mientrasque el 2 tenía la misma composiciónde antígenos
somáticosque el 1, del que sólo sediferenciabapor sus antígenosflagelares.
Aunqueel modelo de Patersonsigue todavíahoy vigente,seha tenido que ampliar al
demostrarsela existenciade otros factoresantigénicos.En 1958 Seeliger(citado por Seeliger,
1961) subdividió el serotipo4 en otros dos serotipos,que denominó4a y 4b, basándoseen
diferenciasde antígenosO. Donker-Voet(1957y 1958, citado por Vázquez-Boland,1984)
añadiónuevosserotipos,4c y 4d atendiendoa diferenciasen antígenosO, y la y 3a por
diferenciasen los antígenosH.
En 1962 Ivanov describióel serotipo5 al estudiarla listeriosis ovina. Este nuevo
serotipopresentaun factorantigénicoqueno existeen los serotipos1, 2, 3 y 4a al 4e (Cooper
et aL, 1973), y difiere del restode las listeriaspor su granpoderhemolítico.En 1966Donker-
Voet describiólos serotipos3c, 6 y 7, y en 1974 Stuarty Welshimerdescribieronel serotipo
de L. inurrayL
1<>

ji
~2.
xl
>1
xixxi
¡
xi
xxxi
xi
xi
xi
xi
¡1¡
•¡
¡—
——
—‘o
¡—
——
o-
>>
>:-
0-0>
>>
>‘o>e
)
.‘Oo
0~
ce—
—e
)
-o)ce0-~-é
-o.>05
o’te
00
1
o‘o
.N
0-0.
e:0.’~
.—
050t

1ntroduccion
Seeligery Donker-Voetdescribieronlos serotipos4f, 4g y 6 (citado por Vázquez-
Boland, 1984). Estos poseíanuna estrecharelaciónserológicay correspondíana cepasno
hemolíticasy no patógenasdelisterias;en 19>79Seeligery Hóhneencuadraronestosserotipos,
que nunca habíansido aisladosde materialpatológico alguno y sí de hecesy del medio
ambiente,en la nuevaespecieL. innocua.Más tardeSeeligery Schoofs(1979)revisaronla
designación de estos serotipos no hemolíticos encuadrándolosen el serotipo 6, que
subdividieronen 6a y 6b (que corresponderíana los antiguos4f y 4g) puestoque el factor
VIII presenteen el 6 y no en 4f y 4g no volvió a encontrarseal volver a analizarestascepas.
Sin embargo,Ralovich en 1984 pusode manifiestola complejidadserológicade la especie
L. innocua, ya que los serotipos6a y 6b puedendividirse a su vez en vanossubserotipos.
Todos estoscambiosestánreflejadosen la 905 edicióndel Bergey’s Manual(Seeligery Jones.
1986).
Nuestro grupo ha contribuido al establecimientode la clasificación antigénicade
Listeria, así,en 1988 Vázquez-BolandetaL describieronunanuevaestructuraantigénicapara
L. grayi y L. murrayi, segúnla cualestosdos serotiposcompartiríantres lactoresantigénicos
comunes,pero cadauno de ellos presentaríaademásal menosun factor propio. Igualmente,
Garcíaet al. (199<)) encontraronvariacionescon respectoa la estructuraantigénicasomática
admitida hastaese momentoparaListeria. Estasvariacionessecircunscribena la aparición
del factor IX en cepasde serotiposque aparentementeno lo poseían(4ab y 6b), a la no
aparicióndel mismo factoren todas las cepasdel serotipo4b, y a la no aparicióndel factor
VIII en todaslas cepasdel serotipo6b. Segúnesto la estructuraantigénicadelgéneroListeria
seríacomo apareceen la tabla 3.
1.1V- EPIDEMIOLOGíA DE LA LISTERIOSIS
I.IV.a- RESERVORIOS
Laenormeubicuidadde L. monocytogenesha sido puestade manifiestocon numerosas
referenciasal aislamientode estemicroorganismoa partir de una gran cantidadde sustratos.
Así, se han descrito aislamientosde sueloscultivados,barbechos,lodos, etc. (Blenden y
Szatalowich,1967; Weisy Seeliger,1975; Weis, 1975; Kampelmachery Janssen,1975; Mero
y Steucr, 1981; Watkins y Sleath, 1981); de vegetales,como pastos, cultivos, hierbas,
12

mt roducciSn
ensiladosmal conservados,hojasde arbustos(Welshimer,1968; Welshimery Donker-Voet,
1971; Weis y Seeliger, 1975; Vázquez-Bolandet aL, 1992); y, de aguasresidualestratadas
y sin tratar, aguas fluviales, lagos, efluentes de mataderose industrias transformadoras
(Dijkstra, 1981; Watkins y Sleatb, 1981); todo lo cual constituyeel llamado reservorio de
origen telúrico.
También los animales constituyen un importante reservorío. De hecho L.
rnonocyrogenesha sido aisladaa partir de másde 42 especiesde mamíferos,22 de aves,30
de peces, anfibios, crustáceos,arácnidose insectos (Gray y Killinger, 1966; Hird y
Genigeorgis,1990).En estesentido,seconsiderancomo los más importantesreservoriosde
Listeria a las personasy los animalessanos;llegándosea estimarque un 40% de la población
humanason portadoresfecales de L. rnonocytogenes(Lovett, 1989, como media de 29
estudiosdistintos), aunqueentre la población sometidaa riesgo de carácterprofesional
(matarifes,veterinarios,ganaderos,etc,) estosporcentajespuedendispararsehastaun
(Kampelmachery Van Noorle-Janssen,1968, citadospor Briones, 1990; McLauchlin, 1987).
El porcentajedeportadoresfecalesentrelos animalessanostambiénesenormementevariable,
oscilandoentre un 14% (Ralovich, 1984a) y un 52% (Skovgaardy Morgen, 1988). Es
importantesubrayarel importantepapeldesempeñadopor los portadoresfecalesdesdeel
punto de vistaepidemiológico,puescontribuyenenormementea la diseminaciónde Liste,-ia,
al constituirun elementobásicoen la transmisiónde la enfermedadbien de forma directao
biende formaindirectaa travésde sus productos(carne,leche,etc.) o contaminandocon sus
excrementosdistintosproductosde consumohumano,comoson las verdurasy hortalizas.En
estesentido,caberecordarel papel atribuido a las aves,que actúancomo portadoresfecales
contaminandocon sus excrementoslos ensilados(Penlon, 1985). Sin embargo,la principal
fuente de contagiola constituyenlos animalesenfermosque diseminanla bacteriaa través
de su ovina, hecesy secreciones(Cray y Killinger, 1966).
A pesarde todo lo expuestohastaaquí, debemosrealizar algunasmatizaciones.No
podemosolvidar la existenciade especiesno patógenasdentrodel género(Listeria innocua
especialmente)que también puedenser aisladasde las mismasfuentes que las patógenas
(Seeliger,1976). Por otro lado,tambiéndebemosseñalarqueel numero de individuosen los
quesedeteetaL. rnvnocytogenesen tracto gastrointestinalesmuy superioral de aquellosque
llegan a desarrollaralgún tipo de manifestaciónclínica; en estesentido,nuestro grupo ha
13

1 n roducción
descrito la excreción fecal de L. rnonocytogenespor parte de ratones inoculados
experimentalmenteporvía subcutáneaque no desarrollaronSignosclínicosy quepresentaron
muy escasaslesiones(Brioneset aL, 1992), lo quesugiereotro posibleorigen de portadores
fecalesapartede la ingestión.
1.IV.b- VíAS DE TRANSMISION
Pesea la gran ubicuidadde Listeria y su aislamientoa partir de tan amplio espectro
de substratos,la verdaderaimportanciasanitariade estosmicroorganismosradicaen que la
listeriosisesunaenfermedadtransmitidaprincipalmentepor los alimentos(GómezMampaso,
1993). Es importanteseñalarque el númerode casosen 1990 (Rocourt, citadapor Gómez
Mampaso,1993)seencontrabaentre0,22 y 7,4/millón de habitantessiendoEE.UU.,Nueva
Zelanda,Australia, Franciay Dinamarcalos paísescon mayor incidencia,señalándosepara
España,en esteestudio,una incidencia del 0,6/millón de habitantes.
La contaminaciónde los alimentosha sido temade bastantecontroversiadurantelos
añosen quela patogeniade la listeriosiseraun tema“oscuro”. Perola existenciade una vía
de transmisión indirecta a través de los alimentos fue puestade manifiesto de manera
indiscutible medianteel aislamientode LineRa a partir dc alimentosdestinadosa] consumo
humano.Entre los alimentosimplicadosde formaespecialen estatransmisiónseencuentran
los productos lácteos, carnesy verduras,responsablesde los últimos brotes de listeríosís
(Schlechet al., 1983; Fleming et aL, 1985; Jameset al., 1985; Ho et aL, 1986; Bille, 1988).
En este sentido, cabe recordar que L. nwnocyrogenesha sido aislada de leche cruda y
pasterizada(Lovetí et aL, 1987; Fernández-Garayzábalu aL, 1986), quesos frescos y
madurados(Marth. 1988b),en carnesde poiio, corderoy ternera (Lee y MeClain, 1986;
Lowry y Tiong, 1988), salsaspreparadas(Farber, 1987, citado por Bracket, 1988), frutas,
verdurasy hortalizas(Schlechet aL, 1983; Ho etal., 1986),asícomode pescadosy mariscos
(Lennon et aL, 1984), sin olvidar el evidente riesgo de contaminaciónde los alimentos
durantesu manipulacióny procesamientoindustrial. De estamanera,los lugareshúmedosy
con restos de materia orgánica, sumideros, etc. constituyen impol-tantes fuentes de
contaminaciónexógenade los alimentos (Lowry y Tiong, 1988; Cox, 1989; Skovgaardy
Morgen, 1988),asícomomanipuladoresdelos alimentoscon unalisteriosislatente(Ralovich,
14

Introducción
1984b).Otro importantehechoque debeser tenido en cuentaes la capacidadde Listeria de
crecera bajastemperaturas,lo que permitiría su proliferaciónen alimentosalmacenadosen
refrigeración.
De lo señaladoanteriormente,sedesprendeel carácterzoonósicode la listeriosis y la
importanciade la transmisióna travésde los alimentos(Hird, 1987). Sin embargo,algunos
autores(GómezMampaso,1993)señalanla alta frecuenciacon que aparecealgunapatología
subyacente:70% en Franciaó 82% en nuestropaís.
Ademásde la transmisiónporlos alimentosno sedebendescartarotrasvías, como la
transmisión directa de los animales al hombre, caso de la listeriosis cutáneade los
veterinarios,contraídaal manipularprotegidosde maneraindebidafetos abortadoso el tracto
genital de hembrasenfermas(Blendenci aL, 1987).Es tambiénposiblela transmisióndirecta
hombre-hombre,como ha ocurrido en algunosbrotesintrahospitalarios(McLauchlin, 1987;
Ho et aL, 1986).
Por último, la listeriosis de los recién nacidos puede tener dos orígenes: una
transmisióndirecta madre-hijo(hematógenao producidaen el momento del parto), o una
transmisiónnosocomial,de aparicióntardía,debidaal deficientemanejode material clínico
(termómetros,sondas,etc.) (McLauchlin, 1987).
En cuantoa la listeriosis animal, el carácterubicuitariode Listeria, y su presenciaen
el suelo,la vegetación,aguassuperficiales,etc. permitensu apariciónen alimentosdestinados
a los animales,en especialensiladosy forrajes en mal estadoo con un pH alto, existiendo
unaestrechaasociaciónentreel consumode estosproductosy la apariciónde listeriosis(Gray
y Killinger, 1966; Fenlon, 1985; Giner et al., 1986; Dijkstra, 1987). A su vez, los animales
infectadosjueganun importantepapelen la transmíslona otros animalesy al hombre,tanto
de forma directacomo indirecta.
1.IV.c- MARCADORES EPIDEMIOLOGICOS
En los últimos brotesde listeriosisocurridosen EstadosUnidos y Canadáseprocedió
a una encuestaepidemiológicaexhaustiva,con el fin de determinarlas circunstanciasy la
¡5

Introducción
fuentede contagio (Schlecha aL, 1983; Fleming a aL,1985;Jamesci al., 1985). En este
sentidose utilizaron los datos concernientesa alimentación,pero también otros como los
relativosal desarrollode actividadesal aire libre Gardinería),viajes,contactocon enfermos,
educación,etc. Se tuvo en cuenta,parala definición de listeriosis,unicamenteaquelloscasos
en los queel aislamientode L. rnonocyeogenesfue positivo,aúnen los casosen queno estuvo
acompañadode sintomatología(Schlechet aL, 1983; McLauchlin, 199<)).
Enla mayoríade los brotessepudodeterminarel origencomúnde los mismosgracias
al empleo de marcadoresepidemiológicos.Entre las técnicas utilizadas se mostraron
especialmenteútiles la fagotipia y la serotipia, aunqueel empleode técnicasbasadasen la
hibridaciónde AUN (sondas,patronesde restricción,PCR) y las técnicasínmunoenzímáticas
presentanuna gran proyecciónde futuro (Blancoci aL, 1993). Sin embargo,por el momento
presentancomo inconvenientespara su utilización de forma general un elevadocoste, la
necesidadde personalespecializado,y el queno hayandemostradounamayorefectividadcon
respectoa los métodosmicrobiológicosclásicos(Blancoci al., 1993).
La fagotipia fue propuesta,al ampliar la capacidadde establecerrelacionesentre
distintas cepas,como una nuevaaportaciónal estudioepidemiológicode la listeriosis. En
1985 Auduriere, al. describieronun sistemade tipificación de L. rnonocvtogenes de 27 fagos.
Con los datos obtenidosen un brote en 1975. este grupo sugirió la posibilidad de que
existieseuna estrecharelación entrelas estructurasutilizadaspor los fagoscomo receptores
y los antígenossomáticos,es decir, entrelos fagovaresy los serovares.
La fagotipiapresentaunaelevadaespecificidad,no produciéndosereacciónde lisis con
otras 2<> especiesbacterianastaxonómicamentepróximasa Listeria (Lovett. 1989; Ortel,
1989b;Rocourty Catimel, 1989;Loessnery Busse,1990). La fagotipiay laserotipiahan sido
empleadasconjuntamentecon resultadosque puedencalificarsecomo excelentes(Espazeet
aL, 1989). Sin embargo,algunosserovares,el 1/2 en especial,presentanuna bajacapacidad
de fagotipia (es decir, reaccióncon al menosun fago), que en ocasionesalcanzatan solo un
29% de cepasfagotipables,lo que anula, en cierta medida, la validez de estastécnicas
(McLauchlin, 1987; Saunderset aL, 1989).
El análisisserológicode casosaisladosy brotesde listeriosis ha probadoqueel título
16

Introducción
de anticuerposfrente a las listerias aumentacomo consecuenciade la infección, no
pudiéndosenegaruna correlaciónentrela incidenciade la enfermedady la elevaciónde las
tasasde anticuerpos(Larsen y Jones, 1972; López et aL. 1992). aunqueel estudio de la
presenciade aglutininasanti-Listeria en la poblaciónsanatanto humanacomo animal, pone
en evidenciala existenciade un alto númerode individuosque presentantítulos séricoscon
valoressimilares a los encontradosen los individuos enfermos(Jasinska,1969, citado por
Hyslop, 1975; Domínguez,1982).Varias hipótesishantratadode explicarestefenómeno,una
de ellas implica a las listeriaspresentesen el intestinode los individuos comoresponsables
de estastasasde anticuerpos(Domínguez,1982; Seeliger, 1987).Por otro lado, tambiénse
ha responsabilizadoa la existenciade reaccionescruzadasentre los microorganismosdel
géneroListeria con Staphylococcusaureus(Seeligery Sulzbacher,1956). Pero el hechode
queen muchasocasioneslos serovaresaisladosde portadoresfecalesno se correspondancon
los aisladosde procesospatológicosllamapoderosamentela atención.En estesentido,nuestro
grupo ha realizadoestudioscon especiesrelacionadascon Listeria que apuntanque la
existenciade anticuerposespecíficosfrentea Listeria sólo seaposible por un contactoprevio
con este agente,y no por reaccíonescruzadascon otros aeentesbacterianos(López et al.,
1993).
Los serovaresmás frecuentementeaisladosde individuos enfermossonel 4b y el l/2a
(Farber y Peterkin, 1991; Suárez, 1991). SegúnMcLauchlin (199<)), el serovar4b es el
responsablede la mayoríade los casosde listeriosis registradospor su laboratoriode 1967
a 1988 (64%), seguidopor l/2a (15%) y l/2b (10%). Estascifras coincidencon las señaladas
porGouletetaL (1989)en un estudiollevado a caboen Franciaentre 1983 y 1986, en el que
los porcentajesson respectivamente62%, 22% y 13%. En cuanto al resto de serovares,
McLauchlin (199<)) les concedevalores del 6% de casos de listenosísproducidos por
serovares4 distintos al 4b, un 4% para l/2c y, por último, un 1’X para los serovares3.
GómezMampaso(1993)afirmaque en nuestropaíshasta1986 los serotiposdel grupo4 eran
responsablesdel 92% de los casosde listeriosis,mientrasque los del 1/2 lo eran del 8%
restante;esteautor cita, asimismo,un estudiode Rocourtde 1990, en el queel 54% de los
casossondebidosal serogrupo4, y el 43% al 1/2.
17

1nt roduccwn
1.V- ENFERMEDAD
Tal y comose ha señaladoen el capítulode epidemiología,el númerode casosde
listeriosis resulta pequeñocon respectoa la ubicuidad de estos microorganismos.Parece
obvio, por tanto,que el desarrollode la listeriosisprecisade unacombinaciónde factoresde
patogenicidadde Listeria y de susceptibilidaddel hospedador(Kampelmachery Janssen,
1969), lo que parecedemostrarlo acertadode la calificación de “agentes oportunistas’
realizada por Simpson en 1971 sobre estos microorganismos(citado por Fernández-
Garayzábal,1984). De tal forma que hoy se consideramás importante la existenciade
factores predisponentespara el desarrollo de la enfermedadque la mera entrada del
microorganismoen un posible hospedador.Los factorespredisponentesmás conocidosson
la depresiónde la inmunidad de basecelular durantela gestación,inmadurezdel Sistema
Inmune en los recién nacidos, inmunosupresiónterapéuticaen trasplantados,linfomas,
tratamientoscon antiácidos,etc. (Siddiqueci aL, 1985; Stammet aL, 1982; Briones, 1993).
Sin embargo,todavíaexistencasosque no puedenserexplicadospor la existenciade estos
factores,lo queha llevado alos investigadoresal convencimientode que no seconocentodos
los factorespredisponentesque puedenintervenir en la listeríosís.
Afortunadamenteel modelo experimentalmurino permite observartodas las formas
de listeriosis que podemosencontrar en el hombre y en nuestrasespeciesdomésticas
(Domingo et aL, 1993). Este modelo tambiénpermiteel estudiode la respuestainmune de
basecelular, así como los mecanismosque utiliza Listeria para producir la infección, y se
consideraválido paraextrapolarlos resultadosal hombre(Stelmaet aL. 1987).
1.V.a- PENETRACION EN EL HOSPEDADOR
La listeriosis presentatres cuadros clínicos principales (abortos, septicemiasy
encefalitis),así como otra seriedc cuadrosque suelennecesitardel establecimientode una
septicemiaprevia.De formageneral,tal y comoconfirmanlos últimos brotesapaíecidosentre
la población humana,a excepcióndel brotefrancésde 1992 que no ha podido relacionarse
claramentecon alimentoalguno (Dehaumont,1993). todos ellos se encuentranasociadosal
consumode productoslácteos,carnesy verdurascontaminadas(Schlecbet aL, 1983;Fleming
eaaL, 1985; Jameset aL, 1985; Ho ci aL, 1986; Bule, 1988), y en los animalescasi siempre
lx

1 ntroduccio
seencuentreasociadaal consumodeensilado en mal estado(Grayy Killinger, 1966;Fenlon,
1985; Audurieret aL, 1985;Gitter, 1985;Dijkstra, 1987),por tanto, podemosafirmar que la
vía de entradade Listeria enel hospedadores la oral (Domingoa al., 1993),si exceptuamos
la formaencefálicaen los rumiantesque trataremosposteriormente.Sin embargo,aúnexisten
dificultadespara conocerel mecanismoexacto de la infección por vía oral, como son la
necesidad de emplear altísimas dosis infectantes para reproducir la enfermedad
experimentalmente,y, de forma especial,la variabilidad en la respuestade individuos en
condicionessimilares.
Dos son las vías de penetraciónpropuestasa partir de la luz intestinal. Raczy col.
(citadospor Gaillard et al.,1987; y por Briones, 1990), medianteestudios de microscopia
electrónica,describenla invasión, en una primera fase, de los enterocitosdel intestino
delgado,y la multiplicación de L. nwnocyrogenesen el interior de esascélulas.Las listerias
libres en el citoplasmapodríanpasara enterocitosadyacentes,o bien salir al exteriordonde
seríancapturadaspor células fagocíticas.Estateoríapermitea L. monocytogeneseludir en
ciertamedidala acciónbactericidade los macrófagosde la lámina propia,convirtiéndolaen
un parásitointracelularcapazde invadir célulasno fagocíticas.
Estahipótesisse ve reforzadaen primer lugar por el hecho de que otros epitelios
(conjuntiva, epitelio nasal) puedenservir como lugaresiniciales de la infección (Gray y
Killinger, 1966); y, ensegundolugarpor la capacidadde Listeria de penetrary multiplicarse
en célulasepitelialesin vitro (Gaillard u aL, 1987; Cossarty Mengaud, 1989; Meyer et al.,
1992). En esta fase la hemolisina desempeñaun importante papel en la diseminación
extraintestinalde Listeria (Rolí y Czuprynski,1990),ya queuna vezen el interior de la célula
hospedadoraL. monocytogenesdependede ella para su multiplicación, al intervenir en su
liberacióndel fagosoma(Gaillard et aL, 1987; Kuhn et aL, 1988; Portnoy, 1992).
La segundahipótesis(MacDonaldy Carter,1980; Marco et aL, 1992b)sitúala puerta
de entradaen unascélulasespecializadasde las placasde Peyer.denominadascélulas M. a
partir de las que Listeria sería rápidamentefagocitadapor los macrófagosíntimamente
asociadosa estascélulas,sin que seproduzcamultiplicación en las célulasM. Por tanto, en
esta hipótesis la difusión de las listerias en el hospedadordependeúnicamente de su
capacidadde multiplicarsedentrode los macrófagos.Es conocidala capacidadde las células
19

Listeria
Teoría de McDonald/Carter Teoría de Racz
Figura 2- Esquemade penetración deListeria en el hospedador.
Enterocito
Macrófagos

Introducción
M para procesarantígenossolublesintestinalesy de fagocitar procariotas;de hecho los
Reovirusutilizan estavía paraatravesarla barreraintestinal (Wolf et al., 1981, citado por
Briones, 1990).
Sin embargo,puedenconsiderarseno excluyentes,y es posible que Listeria utilice
ambasvías parapenetrara travésdel epitelio intestinal (Czuprynski, 1994).Una vez en el
citoplasmaListeria es capaz de inducir la polimerización de filamentos de actina a su
alrededor,lo quele sirve paradesplazarseporel citoplasma,e inclusoparapenetrarencélulas
adyacentessin salir al medio externo(Tielney y Tielney, 1993; Czuprynski, 1994) (Fig. 2).
Porúltimo, esprecisoreseñarotra posiblevía de infecciónsustentadaen la ideade
que Léteria puedeescapara la accióndel SistemaInmunesi se encuentracon una mucosa
o epitelio alterados.Barlow y McGorumdemostraronque las ovejasen épocade cambiode
denticióny sometidasa factoresde alto riesgo,comoesla alimentaciónmedianteensilado,
presentabanuna mayorprevalenciade encefalitis por listerias (Barlow y McGorum, 1985).
Listeria rnonocyrogenes alcanza las terminaciones nerviosas de los pares craneales,
principalmentetrigémino,facial y vestibulococlear(DomingoetaL, 1993),y avanzade forma
centrípetahacia el cerebro por el interior de los axones (Otter y Blakemore, 1989),
pudiéndoseproducir lesionesde tipo inflamatorio. Una vez en el cerebro,continúa su
migración a través de las fibras nerviosas,produciéndosela apariciónde fuertesreacciones
inflamatoriasen cuyo inicio predominanpolimorfonuclearesneutrófilos,que posteriormente
son sustituidospor macrófagosactivados. Esta fuerte reacción inmunopatológicaque se
produce en el cerebro puede ocasionar graves alteraciones tisulares (microabscesos,
demielinización,necrosis)responsablesa su vez de los síntomasclínicos apreciadosen este
cuadro(Charíton y García, 1977; Domingo et aL, 1993). Tambiénse ha sugeridoque las
heridasde cabezay cuello podrían actuar de igual forma (Pohjanviruay Huttunen, 1985),
afirmación que se ha visto apoyadapor los resultadosde nuestrogrupo, al demostrarla
aparición de meningitis en animalesinoculadospor vía subcutánea(Prats et aL, 1992a,
1 992b).
I.V.b- CINETiCA DE LA INFECCION Y RESPUESTA INMUNE
Los estudiosrelativosa la cinéticade la enfermedad,la diseminaciónintraorgánica,
21

Introducción
histopatología,etc, han necesitadoel desarrollode un modelo experimentalen el que se
pudieraninvestigar éstos y otros extremos.En estesentido son numerososlos estudios
realizadossobre la listeriosis experimentalen el ratón desde los primeros trabajos de
Mackanessen 1962(citado porBriones, 1990).El carácterde parásitointracelularde Li steria
y la inmunidad de base celular como respuestaa la infección confieren a la listeríosís
experimentalcaracterísticaspeculiaresen lo que a sus mecanísmospatogénicos,mediadores
del SistemaInmune,modulaciónde la respuestainmune,etc., serefiere.
La cinética de la infección sigue un patrón ampliamenteaceptadopor los diversos
autores(Wilkinson y Hall, 1971; Cheerset aL, 1978; Kongshaveny Skamene,1984).Ue los
estudiosrealizadosen ratonesseconocela existenciade varias fasesen la resistenciaa la
enfermedad.A los 10 minutosde unainoculaciónendovenosacon una dosissubletal.el 9<1%
de las células han sido capturadaspor los macrófagosresidentesdel hígado (células de
Kupffer) y la mayoríade las restantespor los del bazo(Kongshaveny Skamene,1984); a las
6 horasse ha producidouna destrucciónde un 9<1% de las bacteriasviablesexistentesen
estosórganosimputablea los macrófagostisulares(Mainou-Fowleret al., 1988).
En el caso de la vía intraperitonealla infección sigue una pautamuy similar a la
reseñadaparala vía endovenosa,si bien en estecaso los macrófagosperitonealescapturan
una cierta cantidadde las bacteriasinoculadas(Archinal y Wilder, 1988b). Cuandola vía
empleadaesla oral, sonlos macrófagosde la láminapropiadel intestinolos responsablesde
la fagocitosisde las listeriasque hanconseguidoatravesarel epitelio, independientementede
la rutaseguidaparahacerlo.Tambiénesésteel patrón aceptadoparaotrasvías de infección
a través de epitelios, comoel conjuntival y el nasal (Kautterea aL, 1963; Miller y Burns,
1970; Pohjanvirtay Huttunen, 1985).
Trasestaprimerafase, las listeriascomienzana multiplicarsede formalogarítmicaen
cl interior de los macrófagosdel hígado y bazo, alcanzandoel máximo de población de
listeriasa los 2-3 díasdel comienzode la infección, segúnla vía. Se ha podido comprobar
que estos macrófagosinfiltrados en los tejidos provienende la médulaósea a través del
torrentecirculatorio,ya queel usoen estafasede sustanciasincrementadorasde la respuesta
macrofágicao de la monocitopoyesis(RX, vinblastina, carragenina,ciclofosfamida,etc.)
aumentadc maneraconsiderableel número de células macrofágicasy, por tanto, el de
22

Introduccion
bacteriaspor aumentodel númerode lugaresen los que puedenmultiplicarse.
Otra seriede mediadoresde la respuestamonociticaque intervienenen estafase son
el CSF (Colony Stimulating Factor)(Cheersy Stanley, 1988; Mielke et aL, 1993), y el TNF
(Tumor NecrosisFactor) (Nakaneet aL, 1988a;Miclke er aL, 1993). En resumen,podemos
afirmar que se produce una respuestainmune no específica mediada por fagocitos
profesionales,entrelos quepredominanlos monocitos y dondepodemosencontrar,a su vez,
numerosospolimorfonuclearesneutrófilos.Esteaflujo de célulasinflamatoriasda lugara los
granulomasque constituyenla lesiónespecíficade la listeriosis.
A partir de estemomento,comienzauna tercerafase,en la que el desarrollode una
inmunidad de basecelular, basadaen macrófagosactivadospor linfoquinas originadaspor
linfocitos T sensibilizadosprovocauna rápida inactivaciónde las listerias (Kongshaveny
Skamene,1984; Mainou-Fowleret aL, 1988). Esta activaciónde los macrófagosdestruye
rápidamentea las bacteriaspor un incrementode su poderbactericida.La sensibilizacióny
proliferaciónde los linfocitos T exige,previamente,el contactoentreL. rnonocvtogenesy los
macrófagosresponsablesde su presentacióna las célulasT (Goosenset al., 1988). En este
procesojueganun importantepapel las moléculasla superficiales(Deschryver-Kecskemeti
et aL, 1988).Unavezactivadoslos linfocitos T inducenunaactivamonocitopoyesismediada
por linfoquinas como el CSF, 1L2 (interleuquina2), a, f3 y y interferón, etc. (Kratz y
Kurlander, 1988; Nakaneci’ aL, 1989; Mielke ci’ aL, 1993).
La secreciónde IFN-gammaimpide lasalidade los microorganismosde los fagosomas
y, por ende,su multiplicación en el citoplasma(Czuprynski, 1994; Fujiki y Tahaka, 1988;
Higginbotham er al, 1992). Al parecereste efecto del IFN-gammase debe a una mayor
producciónde peróxidos(Higginbothamci al, 1992; Tielney y Tielney, 1993).Sin embargo,
estafunción del lEN-gammano pareceestartan clara,ya queMielkeci aL (1993)afirma que.
basadosen experimentosin vitro, sí pareceactuarde la forma indicada,esto es. impidiendo
la salidade listerias al citoplasmacelular;pero, por otra parte,su apariciónno coincidecon
la etapade desapariciónde las listerias,por lo que pareceque su acción seríamásresultado
de las funciones proinflamatoriasdurantela infección, que un efecto sobrela capacidad
bacteriostáticacelular. Este mismo autorafirma que esel TNF-alfa el que puedeestimular
o coeslimularla producciónde estoscompuestosde oxígeno,actuandoasísobrela capacidad
23

Introducción
antibacterianade los fagocitos.
Llegadoestepunto,el hospedadorpuedecontrolardefinitivamentela infección,reducir
el númerode listerias y, en el plazo de aproximadamenteuna semanaeliminarías,o bien
sufrir unasepticemia(Briones, 1990).A esterespectoparecequela limitación del crecimiento
bacterianoen los primerosestadiosde la infección sedebea la presenciade un alto número
de linfocitos T reactivos(Goosensci aL, 1988).
La inmunidad de tipo humoral no es predominanteen el establecimientode la
resistenciaa la infecciónporListeria, lo cualsedemuestraporel hechode que no seaposible
conferir una inmunidad de tipo pasivo medianteel empleo de suerohiperinmune(Miki y
Mackaness,1964).
l.VI- FACTORESPREDISPONENTES
La resistenciao susceptibilidada la infecciónpor L. monocvtogenesen el ratónestá
íeguladapor un genautosomaldominante(genLr, Listeriaresistance)queseexpresadurante
la multiplicación logarítmica manteniéndolabajo control (Mandel y Cheers, 1980). Esta
resistenciapareceresidir en la capacidadde desarrollaruna inmunidad no específicaque
apareceríaen la 20> fase de la infección, originándoseuna cierta inmovilización de los
macrófagosintegrantesde los infiltrados inflamatorios,o una disminución de su capacidad
paraeliminar las listerias (Czuprinsky et aL, 1985).La ausenciao disminuciónde respuesta
en este momentopuedeoriginar una septicemiacon posible muerte del animal (Mandel y
Cheers,1980).
La mayoría de los casosde listeriosis humanaparecenocurrir en individuos que
presentanalgún tipo de factor subyacenteque conducea una alteración de la respuesta
inmune mediadapor células1 (Farber y Peterkin, 1991). Aunque todavía parecenexistir
algunaslagunasen estaafirmación,ya que ello implicaría una gran incidencia de listeriosís
entre los enfermosde SIDA, y esto no ocurreasí (Schuchatei’ al., 1991; Deckeret aL, 1991;
Berengueret aL. 1991; Farber y Peterkin, 1991), aunque parece que la incidencia de
meningitisy septicemiaporLisrerio entreestosenfermosseencuentraen ascenso(Juradoet
aL, 1993); ademásesteautorseñalala existenciade casosen individuos que no presentaban
24

Intr(xIucci(Sn
ningún tipo de factor predisponente.
Es fácil deducir la importanciaque tiene en la listeriosis la capacidadde respuesta
inmunecelular,atendiendoalmecanismopatogénicode L. rnonon’togenes. Así,el tratamiento
con inmunomoduladoresmodifica la susceptibilidadde éstea la infección, incrementándola
o disminuyéndola.De forma general,podemosafirmar quecualquierinmunomoduladorque
estimule la respuestamacrofágicay que sea inoculado antesde la infección aumentala
resistenciaa la misma;por tanto, la acciónde inmunoestimulantesreduceconsiderablemente
la susceptibilidaddel ratóna la infecciónporL. rnonocytogenes.A esterespecto1-a acciónde
las citoquinas,comoel IFN-’y (Nakaneet aL, 1988b),o el TNF (Nakaneet aL, 1 988a;Mielke
et aL, 1993),o las ILI, 1L2 ó 1L12 (Mielke et aL, 1993) tienen un efectoprotector;y. de
hecho, el tratamiento previo con inmunoglobulinasanti-IFN-y y anti-TNF aumenta la
susceptibilidada la infección por L. rnonocytogenesen ratonesnormales (Nakaneet al.,
1989); sin embargo,Mielke eraL (1993)afirma quela accióndel IFN-y, esmásun resultado
susfuncionesproinflamatoriasqueuna accióndirectasobrelos mecanismosbacteriostáticos
de la célula.
Por el contrario,y tal comocabríaesperartras lo expuestoen el párrafoanterior, el
tratamientocon inmunosupresores,como la carragenina(Stelmau aL, 1987) o ciclosporina
(Hanger al., 1987),permitela producciónde la enfermedadcon dosisde L. rnonocytogenes
considerablementebajas. En este sentido, diversosautores(Stamm ea al.. 1982; Gómez
Mampaso et al., 1987; Gómez Mampaso, 1993) han alertado sobre las posibles
complicaciones que L. monocvwgenes puede ocasionar en enfermos sometidos a
inmunosupresiónterapéutica(trasplantadosrenaleso cardíacospor ejemplo).Asimismo, los
individuosquepresentanun cuadrode inmunodeficienciaadquirida(Ridgwayy Brown, 1989,
GómezMampaso,1993), con la salvedadya comentadaen el casodel SIDA, o congénita
(Ueschryvert-Kecskemetiy Bancroft, 1988) también presentanuna mayor susceptibilidad
frente a la listeriosis. Asimismo, otras enfermedadescomo la diabeteso las hepatopatias
puedenpredisponera padecerla listeriosis (Gómez Mampaso. 1993; Ribiere ci aL, 1990:
Schuchatet aL, 1991).
Ademásde los inmunomoduladores,tambiénexistenotros factoresde tipo fisiológico
que influyen en la presentaciónde la enfermedad.Es de sobraconocidala susceptibilidadque
25
F

introducción
presentanlas hembrasgestantes(Luft y Reminglon,1982; Siddiqueet aL, 1985; Lallemand
et aL, 1992; Briones, 1993),y que sedebeaunadisminuciónde la inmunidadde basecelular
provocadaporel crecientenivel de progesteronageneradopor la placenta(Pungci aL, 1985).
Asimismo, hay autores(Kos et aL, 1984) que afirman queratonesalimentadoscon una dieta
rica en colesterolsufrenuna disminuciónde la actividadmacrofágica,y, por tanto, son más
susceptiblesa la enfermedad;mientrasque otros (Pohjanvirtay Huttunen. 1985) mantienen
quelos animalessometidosa dietaparecensermásresistentesa la infección.A pesarde esta
controversia,pareceinnegableque determinadoshábitosalimentarioscontribuyena facilitar
la apariciónde la enfermedad(Briones, 1993).
Otros factoresrelacionadoscon la apariciónde la listeriosis han sido (Briones, 1993):
las variacionesbruscasde la temperatura,aunqueno existenestudios concluyentesa este
respecto;el estrés,que provocaun aumentode corticoidesque conlíevauna disminuciónde
la inmunidad de basecelular; la existenciade heridas,principalmenteen encíasy zonasde
cuello y cabeza;la existenciade tumores,principalmentelinfomas (Schuchatci aL, 1991);
y, la edad,no olvidemos que, aunquetodas las edadesson susceptibles.existe una mayor
incidenciaen individuos jóvenesy ancianos.
Finalmente, no queremos,dentro de esta relación de factoresque tienen alguna
incidencia sobrela susceptibilidada la infecciónpor L. monocytogenes,olvidar el papeldel
pH gástrico.En estesentido, se ha asociadoel uso de cimetidina con la adquisiciónde la
enfermedaden uno de los brotes ocurridos en EstadosUnidos (Ho et aL, 1986). En ratas
tratadascon cimetidina,parecensernecesariasdosisde L. rnonocytogenessignificativamente
menorespara provocar la enfermedadque las necesítadasen ratas no sometidasa este
tratamiento(Schlechet aL, 1986, 1990). Ello demostradael papel protectorque ejerceel pH
gástricoen la infecciónpor vía oral. Aunqueestefactor no pareceser importantea la hora
de estableceruna respuestahumoral(LópezeaaL, 1992).
A pesarde lo expuestohastael momentoacercade los factorespredisponentes,existen
casosque no parecenpoder explicarsepor los expuestosaquí, lo que hacepensaren la
existenciade otros factoresdistintos.
26

Serovares Significaciónpatógena
t. monocytogenes
1<. ivannovií
lb. innocua
l/2a 1/2b l!2cSa 3b Sc4a 4ab 4c 4d 4e7
5
4ab 6a 6bindeterminados
Abortos,rneningoencefalitis,septicemias
fundamentalmenteabortos en ovejas
no patógenas
lb. weishimeri 6a 6b practicamenteapatógenas
lb. seeilígeri 1/2b 4c 4d 6bindeterminados
1 caso descrito
no patógenas
Tabla 4— Significación patógena de lasy serovares de Listaría.
distintas especies
FORMASCLINICOPATOLOGICAS ESPECIES
Septicémica Hombre, animales jóvenes deespecies domésticas, roedores
y lagomorfosMeningítica
Abortos Hombre, rumiantes y roedores
Romboencefálica Hombre y rumiantes
Miocárdica (postsepticémica) Aves domésticas
Cutánea H0mbre
Queratoconjuntival Hombre, rumiantes, roedores ylagomarf os
Mastitis Vaca
Endocarditis Hombre
Tabla 5- Principales cuadros clínicos de la listeriosis (tomadode Domingo et al., 1993)
Especie
lb. grayí

Iniroduccion
l.VII- FORMAS CLINICAS
Comohemosexpuestoyaanteriormente,el númerodepersonasy animalesquepueden
serinfectadospor L. nzonocytogeneses muy elevado,aunqueen la mayoríade ellos no se
desarrollarála enfermedad.Sin embargo,en los casosen quesilo hacen,la listeriosis puede
presentaruna gran diversidadde cuadrosclínicos, tantoen el hombrecomo en los animales,
aunquelos más frecuentesson las formas meningiticas o encefalíticas,materno-fetalesy
septicémicas(tablas4 y 5), que estudiaremosbrevementeen esteapartado.
1.VII.a- MENINGITIS Y ENCEFALITIS
Es la forma más frecuente en los animales domésticos, entre los que afecta
principalmentea los pequeñosrumiantes,en los que cursacomo encefalitis;pero también lo
es en las personasadultasy en individuos inmunocomprometidos,en las que cursacomo
meningitis (Farber y Peterkin, 1991; Schuchates al., 1991; Domingo es al., 199i~). Las
manifestacionesclínicassecorrespondencon las de unameningoencefalitiscon afectacióndel
romboencéfalo,tallo cerebral y médula oblonga. Esto se traduce en la aparición de
opistótonos.ceguera,nistagmo,parálisisfacial, ptosis parpebral,sialorrea,y el típico andar
en círculosque le dio nombrea la enfermedaden el ganadoovino: “circling disease.Esto
último seproduceal afectarselos núcleosfacial y vestíbulococlear(Briones, 199<)). En las
personasadultas,estaforma sueleestarasociadacon una alta mortalidad(Farber y Peterkin,
1991), entreel 13 y el 34% duranteel año 1989 en todo el mundo.
Segúnha sido señaladopor investigadoresde nuestrogrupode trabajo (Pratses aL,
1992a,1992b), una de las vías de producciónde meningitisseríala llegadade las listerias al
sistemanerviosocentral a partir de monocitos infectadosque ingresanen él a través de los
plexos coroideos.De esta forma se produce una inflamación de los mismos que acaba
desembocandoen meningitis.
En la oveja y cabra esta forma de listeilosis puede ser extremadamentegrave,
pudiendoproducir la muertedel animal entrelas 4 y 48 horassiguientesa la apariciónde los
primerossignos.En el casode la vaca,sin embargo,el cuadroesmenosagudo,sobreviviendo
y reponiéndosela mayoríade los animalestras 4-14 días de la apariciónde los primeros
28

InI ¡oclucci,in
síntomas.La recuperaciónespontáneaes relativamentefrecuente(Gitter, 1985; Dijkstra,
1987).
En el caso del hombre, y aunquepuededarse en niños, en cuyo caso tiene un
desenlacefatal, tiene una apariciónpredominanteen adultos.Normalmentecomienzacon un
cuadropseudo-gripaly secontinúacon cefaleas,artralgias,conjuntivitis, rinitis, fiebre,rigidez
creciéntedel cuello, náuseas,vómitos y fotofobia (Farbery Peterkin,1991; Schuchatet aL,
1991). Estafase de excitaciónno sueledurar más de unos 10 días,y se continúacon una
somnolenciaqueseincrementahastallegar al comacon episodiosconvulsivos;en esteestado
se produce la muerte tras 2-3 días desde su aparición sí no se instaurael tratamiento
adecuado,que consisteen ampicilina (sola o en combinacióncon un aminoglucósido),
penicilinaG, y másreciéntementecotrimoxazol(Carvajales al., 1989).En sangrecirculante
se detectauna marcadaleucocitosiscon predominanciade granulocitos,mientras que en
líquido céfalo-raquídeoseapreciapresiónelevada,valoresde glucosamuy bajos, turbideze
inclusoaspectopurulento.La apariciónde monocitosen LCR esindicativo de una meningitis
producidapor L. monocvtogenes.En este caso es posible el aislamiento a partir de éste
(Seeliger,1961; Emile y Bazin, 1975; Ralovich, 1984a;Domingo esaL, 1986; Allerbergery
Guggenbichler,1989; Ortel. 1989b: Carvajalet aL, 1989).
1.VII.b- INFECCION MATERNO-FETAL
Esta forma no es sino una consecuenciade una infección materna que acaba
provocandoabortoo septicemiaperinatal(Schuchatet al., 1991), aunquela infección de la
madre no implica necesariamentela infección del feto (Farber y Peterkin, 1991).
Fundamentalmentese presentaen oveja, vaca y la mujer. En estaúltima puede cursarde
forma inaparenteo bien producir un cuadro inespecíficocon cefalea, fiebre, en ocasiones,
faringitis y pielitis, e incluso síntomasgastrointestinales(SchuchatesaL, 1991). Estecuadro
pseudogripal(Farbery Peterkin,1991)coincidecon la bacteriemia(Briones, 1990; Domingo
u aL, 1993), y duranteel mismo puedenalsíarselisteriasde sangre.loquios.mucus vaginal,
orina y tejido de placenta (Briones, 1990). El endometrio gestantese infecta por vía
hematógena,pasandola infeccióna placentade forma muy rápida (Domingo es al., 1993).
A la fasede bacteriemiale sigueun periodoen el cuallas listeriasseencuentranen el liquido
amniótico, de donde es aspiradopor el feto, provocandola implicación de los tractos
29

Introducción
digestivoy respiratoriode ésteen los quesedesarrollanlesionesapreciablesen la necropsia
(Briones, 1990). Tambiénesfactible que la diseminaciónorgánicay apariciónde Listeria en
los distintostejidosseaconsecuenciade labacteriemia(Seeliger,1961).Trasla infeccióndel
feto, éstemuereproduciéndoseel aborto, lo más frecuenteen vaca y oveja, o si llega a
términoel neonatomuererápidamentedebidoa una septicemia(Domingo u al., 1993).
Estecuadrosepticémicodel neonatocursacon disnea,cianosis,vómitos,adipsia,etc.
(Lorber, 1990; Farber y Peterkin, 1991), siendo más propio de la especiehumana.En los
rumiantesel cuadroesmásatenuado,pasandoprácticamentedesapercibido(Dijkstra, 1987).
En el hombrese produceuna marcadaleucocitosiscon apariciónde célulasrojas y blancas
inmaduras,y, en algunos individuos, pueden apreciarsepequeñosgranulomascutaneos,
llamados listeriomas, que constituyen la denominada ‘granulomatosis infantiséptica’
(Ralovich, 1984a;Gitter, 1985; SchuchatesaL, 1991; Farbery Peterkin, 1991). En algunos
casos,fundamentalmenteen el hombre,el reciénnacidopuededesarrollarun cuadro tardío,
de apariciónentre varios días y varias semanasposterioresal parto de tipo meningítico
(Schuchates aL, 1991; Farbery Peterkin, 1991), que puedeestaracompañadode trastornos
gastrointestinales,conjuntivitis y neumonía.Esta forma clínica suele estar asociadacon
infeccioneshospitalariasde tipo nosocomial,aunquetambiénseha demostradola existencia
de contagioen el momentodel parto (McLauchlin, 1987; Lorber, 1990; Farbery Peterkin,
1991). Tras el parto o aborto la hembrapuedeseguirportandoListeria en su tracto genital
o eliminaríasporhecesduranteun tiempo variable,lo que esrelativamentefrecuenteen los
rumiantes(Grayy Killinger, 1966; Clegg, 1975; Larsson,1979; Gitter, 1979; Dilkstra, 1987;
Goulet ti’ aL, 1989; Marth, 1988a).
1.VII.c- INFECCIONES SEPTICEMICAS
Los individuos adultos tambiénpuedenverse afectadospor una forma septicemíca.
Estaformaincluyeaproximadamenteal 25 % de los casos(Lorber, 1990). Normalmentesólo
son afectados por esta forma los individuos adultos cuando sufren algún tipo de
inmunodepresión;porestemotivo, esunagravecomplicaciónde cualquiercuadropatológico
que produzcaalgún gradode inmunodepresión(linfomas,enfermedadde Hodgkin, diabetes,
alcoholismo,lupus eritematoso,SIDA, etc.) (Ortel, 1989a;Carvajal ea aL, 1989; Gómez
Mampaso. 1993), así como de pacientessometidos a tratamiento inmunomodulador
30

Intr’>duccion
(trasplantados,quimioterapia,radioterapia)(GómezMampaso,1993).
Desdeel puntode vistaanatomo-patológico,estecuadroafectaa numerososórganos,
en los queprovocala apariciónde nódulosde distintostamaños.En cualquiercaso,el hígado
es el órgano diana por excelencia,y en él se desarrollannumerososgranulomas; otios
órganosque también pueden verse afectadosson el bazo, glándulas adrenales,pulmon.
esófago,paredfaríngea,tonsilas,nódulos linfáticos, timo, médulaósea,miocardio,testículos
y músculosesqueléticos.La localizaciónde estosgranulomasen tejidos epitelialespuede
originar la apariciónde áreasnecróticasen éstos.
Estaforma esmuy infrecuentementeobservadaen los animalesdomésticos(Hird y
Genigeorgis,1990).
1.VII.d- OTRAS FORMAS CLíNICAS
Existen un buen númerode formas clínicas, de menor importanciaen cuanto a su
frecuencia,como son las infeccionesmixtas en las que concurrenotras infeccionescon la
listeriosis,aunqueno seconocenporel momentolas circunstanciasen las queseproduceesta
concurrencia(Gitter, 1985). A consecuenciageneralmentede un cuadrosepticémicopuede
desarrollarseunaneumoníay endocarditisagudao subaguda,lo quepodríamosdescribircomo
listeriosisneumónica,y que sueleconstituirunagravecomplicación(Marth, 1988a;Schuchat
et aL, 1991). Los pacientesafectadospor una listeriosis septicémicasuelendesarrollarcon
frecuenciaendocarditisy un cuadrode hepatitis(Armstrong,1985). Las infeccionesfocales
puedentraducirseen artritis, osteomielitisy abscesosespinaleso cerebrales(Schuchatci al.,
1991).
Sin embargo,podríamosdecirque las másimportantesde entretodasellasson las que
brevementedescribiremosa continuación.
- Queratoconjuntivitis:En humanossueleserfrecuentecomo una secuelade la forma
septicémica,aunquetambiénpuedeapareceraisladamentepudiendollegar a provocar
el cuadromeningíticodebidoa una diseminaciónnerviosacentrípeta(Ortel, 1989a).
En los bóvidos es relativamentefrecuentela apariciónde Listeria asociadaa otros
31

Introduccion
microorganismoscomo Mycoplasrnabovoculi, Moraxella bovis. etc, másque como
único agenteresponsabledel cuadro(Gitter, 1985).
- Lisíeriosis cutánea:es una forma muy rara en los animales. Suele apareceren
ganaderosy veterinariosque han tenido contactocon tejidos infectados.Las lesiones
que puedenobservarseson nódulos cutáneosque devienenen pústulas,de cuyo
líquido puedeaislarseL. monocytogenes (Cain y McCann, 1986). Estas lesiones
cutáneaspodríanser causade la apariciónde portadoresfecales,tras una fase de
bacteriemia,colonizaciónhepáticay posteriorexcreciónbiliar (Brionesei’ aL. 1992).
- Mastitis: Debido a la ubicuidadde Listeria seriade esperarque estecuadrofuese
más frecuentede lo que esen realidad.Es un cuadroque carecede especialrelevancia
desdeel punto de vistaclínico, sin embargodesdeel punto de vistaepidemiológico
y de SaludPública si quetiene unamayorimportancia,ya que seríael responsablede
unacontaminaciónendógenade la leche por L. rnonocvtogenes. En estalínea Gitter
(1985)consiguiódemostrarla presenciade Listeria en leche de vacasvarios meses
despuésde haberdesaparecidola sintomatología.Sin embargo,el principalriesgo,más
que los animales que llegan a desarrollarsintomatologíaclínica, lo constituirían
aquellosque padecenuna mastitis subelínica,al serdifícil su detecciónporquese ha
de demostrarla presenciade un elevadonúmero de leucocitosen la leche, o bien
aislarseL. monocytogenesen cantidadesde aproximadamenteí03 UFC¡ml (Errebo-
Larsen y Jensen,1979; Dijkstra, 1987; Bryner et al., 1989). Sin embargo,en la
actualidad se han puesto a punto medios lo suficientementeselectivoscomo para
poder llegar a detectarla presenciade hasta 10’ UFC Listeria /ml (Blanco et al.,
1989).
1.VIII- DEFICIENCIAS NUTRICIONALES Y ENFERMEDAD
Los seresvivos necesitaningerir una serie de sustanciasparapoder mantenersus
funciones biológicas en perfecto estado. Estas sustanciasson, de forma general, agua,
prótidos, lípidos, glúcidos, vitaminas y elementosminerales. La ingestión insuficientede
algunosde estosprincipios podríaproducir un estadocarencialque puedeprovocaralgún
estadopatológicoper se,o bien puedefacilitar la instauraciónde éste.Sin embargo,además
32

Introducción
de la ingestióninsuficientehemosde teneren cuentaque,en ocasiones,a pesarde haberuna
cantidadóptima en la dietade estoselementos,la existenciade circunstanciasque alteransu
normal absorciónpuedeconllevar los mismos efectosque si no lo estuviesen;entreestas
alteracionescabecitar alteracionesdentarias,del tubo digestivo y glándulasanejas(Blood et
al., 1986).
Otros factoresque puedenprovocarla apariciónde deficienciasnutricionalesen un
individuo es la existenciade sustanciasque destruyandeterminadoselementosabsorbidoso
sus metabolitos, o bien que impidan a alguno de ellos realizar sus funciones;cualquier
alteraciónen el transportede metabolitostiene tambiénel mismo efecto; y, porúltimo, la
falta de apetito o anorexia,bien sea patológicao voluntaria, producetambién los mismos
efectos(Blood et aL, 1986).
La resistenciafrente a las enfermedadesse halla íntimamenterelacionadacon la
ingestadel individuo, relación que se ha visto confirmada por estudios epidemiológicos
llevadosa cabo, principalmente,en el TercerMundo, aunqueno debemosolvidar que las
deficienciasnutricionalesno son rarasen los paisesdesarrollados,en los queesrelativamente
frecuenteencontrarestadosde ligeras deficiencias.Por ejemplo en Norteaméricaexiste
deficienciade hierro en al menosun 9% de la población.En casosde malnutriciónproteica
y energéticapuedenapareceralteracionesde la respuestainmune, que, además,pueden
apareceren casosde deficienciasde un único elemento(Chandra,1985, 1989).
Sin embargo,existenpocosdatos sobrela significacióneconómicay la incidencia de
estosproblemasa los que poderreferirnos.La mayoríade los trabajospublicadossobrelas
enfermedadescarencialesmás conocidasno dan en su mayoría datos cuantitativos que
permitanconocersu verdaderaincidencia así como su impacto económicoreal. A pesarde
estasdeficienciasinformativas,el libro de SanidadAnimal de la FAO/OMS (citadopor Blood
et aL, 1986) revela que una gran mayoría de los casosen los que se pudo identificar las
causasfueron debidasa problemasnutricionales,bien deficienciaso bien excesos.
Otro problemasobreañadidoa la hora de valorar la importanciaeconómicade estos
procesosse encuentraen que muchasveces la deficiencia nutricional provoca un mayor
susceptibilidada otras enfermedadesque son a las que se achacanesaspérdidas.Como
33

Introducción
ejemplo digamosque en el Reino Unido, a pesarde la política de suplementarlas dietas,
aparecensignosde deficienciaen cobreanualmenteen el 0,9% de la poblaciónvacuna,pero
las pérdidasdebidasa estadeficienciapodríanestarmuy infravaloradasdebido una mayor
susceptibilidadde los animalesdeficientesen cobrea las enfermedadesinfecciosas(Blood et
aL, 1986).
Podemosafirmar, por tanto, que las deficienciasmineralesseencuentranampliamente
distribuidas, pero el conocimiento de su incidencia e importancia real se encuentra
infravaloradodebidoa quelas formassubclínicasde deficienciasuelenpasardesapercibidas.
En los paísesdesarrolladoslas deficienciasde los oligoelementossuelenconfundirse
con otrascarenciasmáscomunescomosonlas deficienciasenergéticas,proteicas,de fósforo
y aguaque afectanal desarrollopostnataly a la función reproductora.De formageneral,se
consideraa la malnutricióncomola causamás frecuentede inmunodeficienciasecundariaen
el mundo.Podemosafirmar quela malnutriciónprovocaalteracionesen los tejidos linfoides,
que presentanun menordesarrolloque en condicionesnormales.Son numerososlos factores
involucradosen estemenor desarrollo,pero podemoscitar, entreotros, un descensoen la
síntesis proteica y, por tanto, en la proliferación celular, así como un aumentode los
fenómenosde citolisis debido a un aumentode los niveles de glucocorticoides(Chandra.
1985).
Porotraparte, las deficienciasmineralesy los desequilibriosen el sueloy forrajeshan
sido factoresadmitidoscomocausasdebajaproduccióny problemasreproductivos.Dehecho,
el ganado de zonas con bajas cantidadesde cobre, cobalto y fósforo en los terrenosse
encuentranmás limitados incluso por la falta de estoselementosque por deficienciasde
energíao proteínas(Blood et aL, 1986).
De forma generalpodemosafirmarquelos elementosesencialesparalos seresvivos
son 15, que puedendividirse en dos grandesgrupos:
a) macronutrientes:calcio, cloro, fósforo, sodio, potasio,magnesioy azufre;
b> micronutrientes:cobre,selenio,zinc,cobalto,hierro,lodo, manganesoy molibdeno.
34

introducción
Estosmicronutrienteso elementostrazaseencuentraninvolucradoscomocomponentes
de muchostejidosy algunosson indispensablesparael correctofuncionamientode unao más
enzimas.Su carenciaconduciríaa una ampliaseriede consecuenciaspatológicasy defectos
metabólicos(Blood et aL, 1986).
La susceptibilidad a enfermedadespodría encontrarseinfluida por el estado de
desarrollofisiológico, diferenciasgenéticase interrelacionescon otros elementos;por tanto,
dos animalescon niveles parecidosde un elementono tendrían por qué responderigual
(Blood esal., 1986).
Unadeficiencianutricionalno implica la aparicióndeenfermedad,existenfactoresque
predisponenel individuo parasu aparicióncomo la edada la que aparecela deficiencia,
determinadoscambiosen el medioambiente,el contactocon determinadosagentesinfecciosos
y/o parasitarios,la respuestaindividual a la deficiencia, la posibilidad de utilizar rutas
alternativaspor partedel organismo,y la existenciay tamañode reservasde ese elemento
(Blood e, aL. 1986).
Los tres principios quedeberíanpresidirla interpretaciónde los criterios bioquímicos
acercadel estadode los elementosmineralesson:
1- su relación con la ingesta;
2- tiempo que dura la deficiencia;y,
3- función del elementodeficiente.
Cuandoun elementodesaparecede la dieta, seproduceun aportedel mineral a partir
de los distintoscompartimentosdel organismo,lo que permiteque durantealgún tiempo los
nivelessanguíneosdel mineralencuestiónno varien.~Sinembargo,si la deficienciapersiste,
los mecanismosencargadosde mantenerla homeostasisen el organismono puedenmantener
esosnivelesplasmáticos;así se llega aun momentoen el cualla actividadde la(s) enzima(s)
que necesitana esemineral comienzaa disminuir, apareciéndouna fasede disfunción. Esta
fase,de duraciónvariable,suelecoincidir con el periodo subclinicode la deficiencia,pci-o sí
finalmente la carenciamineral en la dieta no se recupera,aparecenuna serie de cambios
celularesresponsablesde las manifestacionesclínicas (Blood es aL, 1986).
35

1 uir,,ducción
1.IX- IMPORTANCIA BIOLOGICA DEL SELENIO
1.IX.a- ACCIONES BIOLOGICAS. ENZIMAS.
La importanciaen los seresvivos de esteelementofue debidaen un principio a sus
efectostóxicos (Spallholzet aL, 1990; Turnery Finch, 1991).En estesentido,estosúltimos
autoresafirmanque de los escritosde MarcoPolo puedeinferirsequealgunosmercaderesen
viaje haciael LejanoOrientepudieranperdersus bestiasde cargadebidoal usocomo forraje
de hierbacon altoscontenidosen selenio.
Durantemuchosañospredominósu consideracióncomo tóxico (Gyanget aL, 1984;
Spallholzci aL, 1990; Bedwal et aL, 1993),hastaque en 1957 Schwarz(citado porBackall
y Scholz, 1979; Gyang ci aL, 1984; Miller, 1985; Spallholz ei’ aL, 1990; Turner y Finch,
1991; Zacharaet aL, 1993) publica los resultadossobresus investigacionesacercade la
degeneraciónhepáticanutricionalen la ratay la influenciade unadieta pobreen vitaminaE.
En éstos afirma que el llamado “Factor 3’ aislado de hígado porcino prevenía esta
enfermedad.Este “Factor 3” conteníaen su composiciónselenio,y poco tiempo despuésse
demostróque esteelementoeracapazde prevenirla degeneraciónhepáticanutricionalen la
iata. Desdeese momento se le reconocíasu papel como un nutriente esencial para el
organismo.
A partirde estehecholos descubrimientosrelacionadoscon el seleniose multiplicaron
(Miller. 1985; Zacharaci aL, 1993). Especial relevanciatuvo el descubrimientode que
lormabaparteesencialde la enzimaglutation-peroxidasa(GSHPx),llevado a cabo en 1973
por John Rotruck (citado por Backall y Scholz, 1979; Miller, 1985; Spallholzet al. 199<);
Turner y Finch 1991). Estaenzima,junto con la vitamina E, forma partede los sistemasde
detoxicación de determinadosmetabolitos activos de oxígeno que se producen durante
numerosasfuncionesmetabólicasde las células (Fig. 3). El fallo de estossistemascausaun
dañocelular e incluso la muertecelular (Rotruck, 1972; Morris, 1984; citadospor Boyne y
Arthur, 1990).
La glutation-peroxidasano esla únicaenzimadependientede selenioen los animales
superiores.En 1985 Ursini (Spallholz. ei’ aL, 1990) describela existenciade una enzimade
36

Introducción
peso molecular inferior a la primera (20.00<) y 84.000 respectivamente), la
fosfolipidihidroperóxido glutation peroxidasa(PLGSHPx). Esta enzima parece tener las
mismas acciones que la anterior, además de intervenir en la reducción de los
fosfolipidihidroxiperóxidos.El selenioestambiénun componentede otrasproteínascomoson
la selenoproteinadel músculo,la selenoflagelina,las proteínastransportadorasde selenioy
las enzimasbacterianasformato-dehidrogenasay glicina-reductasa(Koller y Exon, 1986).
Asimismo, el selenioparticipaen el metabolismode numerosasdrogasy xenobióticos;por
ejemplo, el selenio contrarrestala toxicidad de determinadosmetales pesadoscomo el
arsénico,cadmio,mercurio,cobre,plata y plomo (Hill, 1975; Koller y Exon, 1986).
El papelbioquímicoesencialdel seleniosedebe,pues,aque formapartede la enzima
glutation-peroxidasa(GSHPx)(Braun ei’ aL, 1991;Bedwalet aL, 1993).La actividad de esta
enzimaen los eritrocitosha sido directamenterelacionadacon la concentraciónhemáticade
selenioen vacuno,ovejas,caballospoílos, ratasy cerdos(Backall y Scholz, 1979), y es un
dato muy útil tantoel diagnósticode la deficienciaen selenio,existiendouna relacióndirecta
entrelos nivelesde GSHPxy tasade selenioen sangrey tejidos de estosanimales(Backall
y Scholz, 1979). Estaenzimacontiene4 moles de seleniocomo selenocisteinaen su región
activa(Bedwal et aL, 1993).
La glutation peroxidasaprotegela fracción lipídica de las membranascelularesy
orgánulosdel daño por peroxidaciónmedianteinhibición y destrucciónde los peróxidos
endógenos,actuandoen conjunción con la vitamina E paramantenerla integridadde estas
membranas(Backall y Scholz, 1979; Koller y Exon, 1986). El peróxido de hidrógeno y
peróxidoslipídicos producenla desnaturalizaciónirreversiblede proteínasesencialesde la
célula lo que conduciría a la degeneracióny necrosis,por alteración de las membranas
biológicas,e incluso,a la destruccióndel tejido conectivo(Braun eta!., 199]). La GSHPx
catalizala degradaciónde peróxido de hidrógenoy determinadoshidroperóxidosorgánicos
producidospor el glutation duranteel ciclo redox (Koller y Exon, 1986) (Fig. 4). Esta
dependenciade la actividadGSHPxcon la presenciade selenioexplica la interrelaciónentre
selenio,vitamina E y los aminoácidossulfuradosen los animales.Estosaminoácidospueden
actuarcomo precursoresdel glutation que, a su vez, aauai-íacomo substratode GSHPx y
mantienelos grupossulfidrilos de la célula.
37
Fi
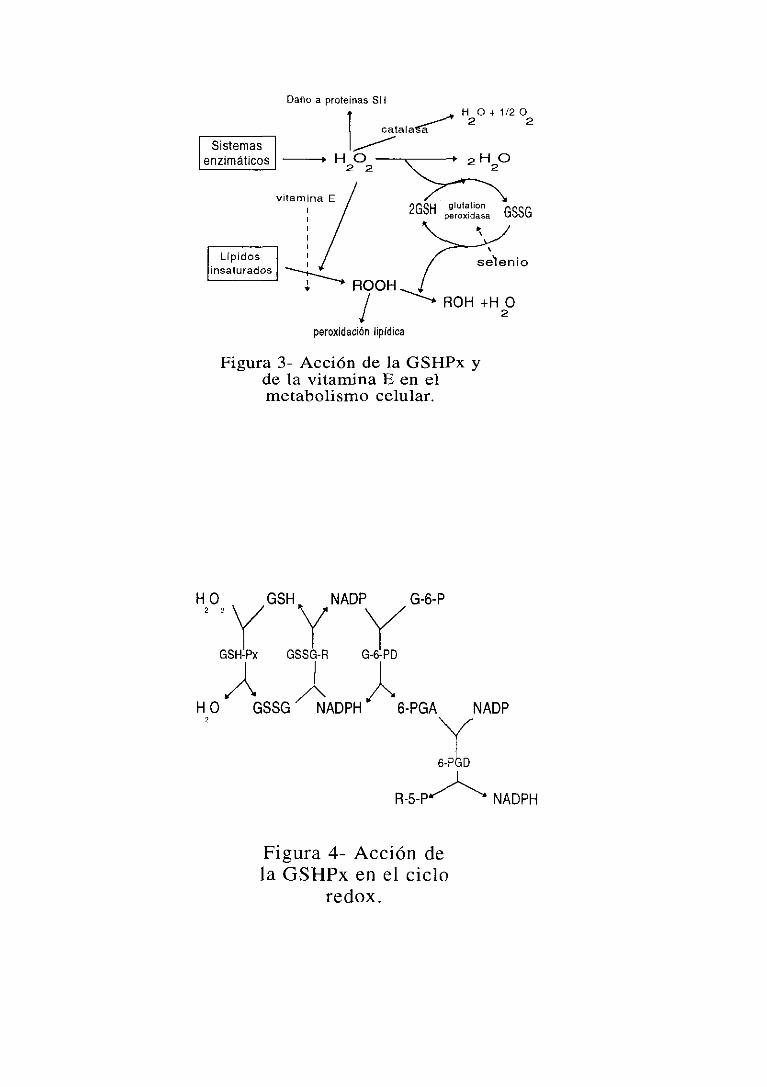
Daño a proteinas SHHO+ 1120
2estala a
2H022 2
Vitamina 2GSH glutation
peroxidasa GSSG
s¿len ¡o
ROH +HO2
peroxidación ¡ipidica
Figura 3- Acción de la GSHPx yde la vitamina E en elmetabolismo celular.
H O GSSG NADPH
G-6-P
G-6-PD
6-PGA NADP
6-P D
R-5-P NADPH
Figura 4-la GSHPx
Acción deen el ciclo
‘1Sistemasenzimáticos
Lípidosinsatu radas
E
4 ROOH
4/
HO22
GSH NADP
GSH-Px GSSG-R
redox.

Introducción
El papel bioquímico de la vitamina E es el de actuar como un antioxidante,
previniendoel dañooxidativo de los lípidossensiblesde las membranascelularesal disminuir
la formación de hidroperóxidos (Braun et aL, 1991). Esta vitamina presentaun papel
importanteen la protecciónde las membranascelularesdel dañoporlipoperoxidación(Turner
y Finch, 1991; Braun et aL, 1991).
Existe una importante interrelación entreselenio, vitamina E y los aminoácidos
sulfuradosen la prevenciónde algunasde las enfermedadesnutricionalescausadaspor su
deficiencia.Si la vitamina E previenela formaciónde hidroperóxidosde ácidosgrasos,y los
aminoácidossulfurados(como precursoresde la GSHPx)y el seleniosehallan involucrados
en la destrucciónde los peróxidos,estosnutrientespodríanproducir un efectobioquímico
muy similar, reduciendola concentracióntanto de peróxidoscomo de compuestosinducidos
por éstosen los tejidos (Blood et aL, 1986>. La protecciónfrente al daño oxidativo de las
proteínasno de membranasusceptibles,porel seleniode la dieta, pero no por la vitamina E,
podríaexplicarpor qué algunasenfermedadesnutricionalesrespondenal selenio,pero no a
la vitamina E. Por otro lado, determinadostejidos o componentesa nivel subcelularpodrían
no estar correctamenteprotegidos debido a que presentande forma inherente menores
cantidadesde GSHPx, inclusocon dietasadecuadas.El dañoproducido a estos tejidos se
veríaagravadopor dietascon alto contenidoen ácidosgrasosinsaturados,siendode esperar
que respondieranpositivamentea la administraciónde vitamina E. pero no a la de selenio.
Las variacionesen la actividadde GSHPxentredeterminadosórganoscomo hígado,corazón
y músculosesqueléticoy cardíacopodrían explicar las variacionesen la gravedadde las
lesionesentreespecies.
1.IX.h- TOXICIDAD DEL SELENIO
Como ya hemosreseñadoanteriormente,el seleniofue conocidoantespor sus efectos
tóxicos que por ser un nutrienteesencial(Gyang ci aL, 1984; Spallholz ti’ aL, 1990). Los
efectostóxicos en los animalesestánasociadosa lugarescon una alta concentraciónde
selenioen el suelo.Podemosdistinguir un efecto agudo,con sintomatologíaprincipalmente
nerviosa,y un efectocrónico, caracterizadoporemaciación,miopatíasy alopecias(Blood et
al., 1986; Koller y Exon, 1986).
39

introduccion
La selenotoxicosisestáligada a la existenciade una seriede plantasacumuladorasde
esteelemento.La existenciade estasplantas,pertenecientesa los génerosAstrogalus y
Oxyi’ropis, debenalertamossobrela probableapariciónde estapatología.Sonplantasque no
suelencreceren sueloscon bajoscontenidosen selenio,y, además,tienenla relativaventaja
de no serexcesivamenteatractivasparael ganadodebidoa sufuerte olor (Blood ci aL, 1986;
Bedwal ti’ aL. 1993).
Otras vías de intoxicación por selenio las constituirían la ingestión de residuos
industriales,asícomo unaposiblevía yatrogénicapor la costumbreactual de inyectarselenio
a los animalesparaprevenirenfermedadescomo la distrofiamuscularenzootica(Blood ei’ al.,
1986; BedwalesaL, 1993).
Independientementede la vía de entradadel excesode selenioen el organismo,el
mecanismode su efecto tóxico sedebea que esincorporadoen los amínoacidossulfurados
en lugar de azufre,y por tanto interfiereen los complejosenzimáticosen los queintervienen
estos aminoácidos(Bedwal et aL, 1993). El selenioreduce,además,las concentraciones
hepáticasde azufre y proteínasen ovejas; en estesentido,dietas con alta concentración
proteicaposeenun efecto protectorfrentea estaintoxicación (Blood ci aL, 1986).
En las intoxicacionesagudassuelenaparecerhalitosisaliácea,diarrea acuosa,fiebre,
taquicardia,posturasanormales,postracióny muerte de los individuos afectadospor fallo
respiratorio(Koller y Exon, 1986; Bedwal ti’ aL, 1993). Porsuparte, la forma crónicacursa
con adinamia, emaciación,pérdida de vitalidad, rigidez y cojeras. Otro síntoma de la
intoxicación crónica es la alopeciaparcial o total que puede aparecer.Las lesionesmás
frecuentesson congestiónhepáticay renal, con aparición de focos necróticosen hígado,
endocarditis,miocarditis y hemorragiasepicárdicas(Blood eta!., 1986; Koller y Exon, 1986;
Bedwal ti’ aL, 1993).
Aunque el selenio puede detectarsetanto en sangrecomo en leche y pelo de los
animalesafectados,no hemosencontradodemasiadosdatossobreestosúltimos parámetros.
Sin embargo,nivelessanguíneosde 3 ppm de seleniosuponenla apariciónde enfermedad
clínica.Por otraparte, tambiénpuedeencontrarseunaleve anemiatantoen los casosagudos
como crónicos;en estesentido,valoresde hemoglobinapróximos a 7 g/dl suelenaparecer
40
ji

Introducción
asociadosa estaintoxicación.
En lo querespectaal hombre,las cantidadesnecesariasparaproducirintoxicaciónhan
sido durantemuchos años objeto de debate (Koller y Exon, 1986). Así, existen datos
contradictorioscomo son que en zonas dondese ingieren cantidadesde 4,99 mg/día la
selenosises endémica,y por otra partese han realizadoestudiosdondese han llegado a
ingerir 1,510 mg/díasin que aparecieransignosde la intoxicación (Koller y Exon, 1986), e
incluso existendatos sobreun hombrede 62 añosque tomó 1 mg de selenioen forma de
selenito sódicodurantedosañosantesde que aparecieranligeros síntomasde intoxicación
(Koller y Exon, 1986).
En cualquier caso, los síntomasde la intoxicación por selenioen el hombre son
halitosisaliácea,engrosamientoy fragilidad de las uñascon aparición de puntosblancoso
líneas longitudinales,pelo seco y frágil que se rompe con facilidad, enrojecimientoe
inflamaciónde la piel en la que puedenformarseampollaso incluso úlceras,y alteraciones
de tipo nerviosocomoconvulsionesy parálisis(Koller y Exon, 1986; Bedwal ti’ aL. 1993).
La apariciónde estossignosdependede la intensidadde la selenosis.
1.IX.c- DEFICIENCIA EN SELENIO
Debido a que es más fácil que se produzcaestaalteración, se conocenmejor las
alteracionesde los animalesdomésticosdebidaso, al menosasociadas,a una deficienciade
selenio y/o vitamina E. Normalmenteestas manifestacionessuelen estar asociadascon
determinadosfactorespredisponenteso desencadenantes,como por ejemplo consumode
ácidosgrasosinsaturadoscon la dieta, ejercicio inhabitual y crecimientodemasiadorápido
en los animalesjóvenes(Blood ti’ aL, 1986). Estasenfermedadessuelen agruparsebajo el
epígrafe de “enfermedadesasociadasa carenciasde selenio-vitaminaE”, puestoque en su
mayoría pueden ser prevenidasmediante una con~ecta administraciónde uno o ambos
elementosen la dieta (Bínod ci al.. 1986).
Como sedijo, un correctoaportede selenio-vitaminaE es importantey su deficiencia
puedeprovocardiversasalteraciones(Braunet al., 1991)de entrelas que podríamoscitar las
siguientes(Blood ci a1, 1986; Koller y Exon, 1986; Hassanti’ aL. 1990), aunquepa’a su
41

Introduccion
estudiomás detalladodebamosremitirnos a un tratadode patología:
1-distrofiasmusculares:son,probablemente,los problemaspatológicosmásconocidos
debidosa las deficienciasen selenio/vitaminaE en la dieta. Puedenafectara distintas
especies,y tanto a los músculoscardiacoscomo a los esqueléticos.En algunoscasos
puedenproducirparálisispor lesionessimétricasde la espinadorsal.La másconocida,
con casi todaseguridad,es la ‘enfermedaddel músculoblanco’ en los rumiantes;
2- hepatosisdietética: esta forma suele apareceren cebaderosde cría intensiva de
ganadoporcino, en los que se utilizan piensoscon escasasconcentracionesen estos
elementos;
3- alteraciones de la respuesta inmune: esta es la alteración de más reciente
conocimiento.Serátratado con cierta profundidaden el siguientecapítulo; y,
4- otrasalteraciones:problemasen la reproducción,retenciónde membranasfetales
y mayor fragilidad eritrocitaria.
Estasalteracionesestáncausadaspordietasdeficientesen selenioo selenio-vitamina
E, junto con la posiblepresenciade factorescondicionantes,como una excesivacantidadde
ácidosgrasospoliinsaturadosen la dieta. Prácticamentela totalidadde estasenfermedadesque
aparecende forma naturalhan podido serreproducidasde formaexperimentalusandodietas
deficientesen selenio y/o vitamina E; y a la inversa,las lesionespuedenser prevenidas
mediantela suplementacióncon selenioy vitamina E (Blood er aL, 1986).En determinadas
circunstanciasseha podidocomprobarquela incorporaciónde cantidadesexcesivasde ácidos
grasospoliinsaturadosha resultadoser un factor importanteen la enfennedadexperimental.
Parecequela presenciade estosácidosgrasosen la dietacausaríaunadeficienciaen vitamina
E, por la actuaciónantioxidantede ésta.
Puedenexistir interaccionesentreel selenioy otros elementostraza (Blood et al.,
1986; Van Vleet et aL, 1977, 1981; Hill, 1975). Las lesionesdebidasa las deficienciasen
selenioy vitaminaE puedenserreproducidasen lechonesy patosalimentadoscon cantidades
excesivasde cobalto,plata,telu¡-o,zinc y vanadio(Van Vleet eraL, 1981).La suplementación
42

lntroduccion
con selenio-vitaminaE puedeconferir protección frente a la cardiomiopatiainducida por
cobalto (Van Vleet ti’ aL, 1977).
En el hombre,una deficienciaen seleniotambiénproducealteraciones,siendola más
importante la enfermedadde Keshan (Koller y Exon, 1986; Bedwal ci aL, 1993). Esta
enfermedades una cardiomiopatíaque puede acabarcon la vida del enfermo,pero que
respondebien a la terapiacon selenio.De forma general,cursacon alteracionesmusculares,
así como aumento de las actividadesGOT, GPT y CK en sangre. Este mismo autor,
basándoseen estudiosepidemiológicos,afirma que la deficienciaen selenioes un factor a
tener en cuentaen las enfermedadescardiovascularesdel hombre, ya que pareceque la
incidenciade hipertensiónesmás alta en zonascon deficienciaen selenioque en las que su
concentraciónes la adecuada(Koller y Exon, 1986). Porotra parte, igual que pareceocurrir
en los animales,la deficienciaen seleniono seexpresaa no ser que exista algún factor
predisponenteo desencadenante,como el estrés(Koller y Exon, 1986), y debeser tenida en
cuentaen casode SIDA, síndromede Down y enfermedadde Crohn (Bedwalet al., 1993).
Los datos disponibleshastael momento,sugierenque tanto el seleniode la dieta,
como los aminoácidossulfuradosy la vitamina E actuaríande forma sinérgicaprotegiendo
los tejidos de un daño oxidativo. Es evidenteque la GSHPx,quedependedel seleniode la
dieta, juegaun papelimportanteen la detoxicaciónde los peróxidoslipídicos reduciéndolos
a compuestoshidroxi de los ácidosgrasosno tóxicos,mientrasque la vitamina E prevendría
de la formaciónde hidroperóxidosde ácidosgrasos.La presenciade altos nivelesde ácidos
grasosinsaturadosen la dieta, aumentaríalos requerimientosde vitamina E, produciéndose
la oxidación tisular con la consiguientedegeneracióny necrosistisular, sobretodo en los
casosen que coexistaun escasoaportede seleniocon la dieta.
1.IX.d- SELENIO Y SISTEMA INMUNE
Esteapartadode las funcionesbiológicasdel selenioes el de descripciónmásreciente
y el que centra, en la actualidad,el gruesode las investigaciones,hastael punto de que
Spallholz ci aL (1990) llega a citar 82 referenciasacerca del papel del selenio en la
inmunidad entre 1972 y 1989. Todos los autores parecen estar de acuerdoen que la
suplementaciónde la dietacon seleniotiene efectosbeneficiosossobrela respuestainmune,
43

Introducción
y que, por el contrario, la deficiencia de este elemento provoca alteraciones en ésta (Koller
y Exon, 1986; Spallholz ti’ aL, 1990; Turner y Finch, 1991)
La primera referencia de que el selenio jugaba algún papel en la respuesta inmune data
del año 1959 (Spallholz ti’ al?, 1990) cuando McConnell observa como el te inoculado
subcutáneamente en perros era incorporado en la fracción proteica de sus linfocitos. A éste
le siguieron varios descubrimientos en este sentido, entre los que podemos citar (Spallholz
ci al., 1990) la descripción por primera vez del efecto antiinflamatorio de una fracción, que
posteriormente se vio estaba formada por selenio; el hallazgo de la GSHPxen los leucocitos
humanos; y, la disminución de la actividad de esta enzima en mujeres con la enfermedad
crónica granulomatosa.
Sin embargo, la primera referencia específica acerca de selenio y respuesta inmune,
se produce en 1972 (Spallholz et aL, 1990) cuando Berenshtein observa cómo la producción
de anticuerpos en conejos inoculados con toxina diftérica fue mayor tras una segunda o,
incluso, una tercera inoculación en los animales tratados con selenito sódico que en los
respectivos controles no tratados. A partir de este descubrimiento los trabajos que relacionan
selenio y sistema inmune se han visto aumentados en gran número tal como puede verse en
la excelente revisión de Spallholz ti’ aL (1990). En este sentido, la disponibilidad de selenio
en la dieta afecta a todos los componentes del sistema inmune (Kiremidjian-Schumacher ti’
aL, 1992), tanto a la función de los polimorfonucleares neutrófilos, como a la proliferación
linfocita¡-ia, mecanismos de citotoxicidad y de producción de anticuerpos (Reffert Stabel ci
aL, 1989; Sordillo et aL, 1993) entre otros.
Dado que existe una relación directa entre la tasa de selenio y actividad de GSHPx,
es preciso hacer referencia a la actuación de ésta respecto al Sistema Inmune. Se ha podido
comprobar que la GSHPxse encuentra en la práctica totalidad de las células que derivan de
la médula ósea, esto es, eritrocitos, linfocitos, monocitos, e incluso en las plaquetas, así como
en los órganos principales del sistema inmune como médula ósea, timo, hígado, bazo,
glándulas linfoides, e incluso en el intestino. No debemos olvidar que este último es uno de
las principales puertas de entrada de elementos extrañosa~ organismo.
Las funciones de esta enzima con respecto al sistema inmune son:
44

InIn ducciin
1- reducción de los peróxidos orgánicos e inorgánicos que se forman durante el
metabolismo general;
2- metabolismo de los hidroperóxidos que se forman en las rutas de la lipoxigenasa
y de la cicloxigenasa a partir del ácido araquidónico; y,
3- reducción del peróxido de hidrógeno, así como del anión (Y formados durante la
explosión oxidativa en la fagocitosis.
l.IX.d.1- Influencia sobre la función fagocitica
Una vez llegadosal foco inflamatorio o infeccioso los monocitos y macrófagos
eliminan todo aquelloque puedaresultardañinoal hospedador.Paraello englobanal agente
extrañoincluyéndolodentrode unavacuolaensucitoplasma,vacuolaque esconocidacomo
fagosoma. Las células fagocíticas presentan una gran riqueza en lisosomas en su citoplasma.
orgánulosque fusionansus membranascon las del fagosomaliberandosu contenidoen su
interior, que pasa a denominarse fagolisosoma. De esta manera en el interior del fagolisosoma
existeun ambienteácido, íico enenzimasquedestruyeneseagenteextraño,peroademásese
ambienteesrico en unaseriede metabolitosde oxigeno(H,02 y O; entreotros)que facilitan
esalabor tóxica del fagolisosoma(Fig. 5>.
Vemos,pues,cómoexistegran cantidadde compuestosde oxígenoque actúancomo
antimicrobianos. Sin embargo estos productos son también tóxicos para la célula, y, por tanto,
si ésta fuera incapaz de detoxicar estos productos se produciría un daño a sí misma. Entre los
responsables de evitar esos daños a las membranas se encuentran tanto la GSHPxcomo la
vitamina E. Es de destacarque en los leucocitosla actividadGSHPx es mayorque en los
eritrocitos, lo que indica que esta enzima actúa también en los fenómenos de fagocitosis
(Erskine et aL, 1987).
Una deficiencia en selenio produce una menor actividad GSHPx y un fallo en la
fagocitosis de Candida (Erskine et aL, 1987; Turner y Finch, 1991), fallo que parece ser
debido a una disminución en la capacidad fungicida más que a una alteración en la capacidad
45

macrófago
REORGANIZACIONTI SU LAR
elastasa, colagenasa,hialuronidasa, factor ________________
estimulante fibroblastos
Figura 5- Resumendel papel del
macrófago en larespuesta inmune.
tomado de Roitt,I.;Brostoff,J.; Male,d.
“Immunology” (1985).Gower Medical Publishing.
Londres.
INFLAMACIONFIEBREILt, Pg
ACTIVAC IONLIN FOGITA A ¡A
presentaciónAgIL1
MICROBICIDA
reactivos de oxígeno
DAÑO TISULAR
reactivos deoxígeno
ANTITU MORAL
TNF, proteasas

Introducción
de ingeriria (Serfass y Gauther, citado por Miller, 1985). Aunque estos mismos autores no
parecenponersede acuerdoen cuantoa si la actividadbactericidaseencontraríaafectada,los
primeros afirman que se encontraría disminuida ante determinadas bacterias como ocurre con
S. aureus y neutrófilosbovinosdeficientes,mientrasquefrenteaProteusmirabilis no se vería
afectada. Por el contrario, Turner y Finch (1991), basándose en los trabajos de diversos
autores, llegan a afirmar que la actividad bactericida podría no verse afectada en animales
deficientesen selenioy vitaminaE, lo queen nuestrojuicio es,cuantomenos,una afirmación
comprometida a la vista de la bibliografía existente sobre el tema. En cualquier caso los fallos
funcionalesde los neutrófilosde animalesseleniodeficientespodríanrestaurarsemediantela
administración de pequeñas cantidades de selenito sádico (citado por Turner y Finch, 1991),
duranteun escasoperíodode tiempo.
La fagocitosis produce un aumento en el consumo de O, que es reducido por la
NADPH oxidasaa otras formasque resultanmásbactericidas,pero que también resultan
perjudicialesparalas membranascelularesal producir la peroxidaciónlipídica de los ácidos
grasospoliinsaturados.La deficienciaen selenio implicaría unas cantidadesmenoresde
OSHPx,lo que influiría sobrela capacidadcelular de detoxicarH,0,, lo que conllevaríaun
aumentode los peróxidoslipídicos, que inhibirían la ruta de las hexosasmonofosfato,que
inhibiría, a su vez, la regeneración de la NADPHa través de esta ruta y por tanto acabaría
produciendo una limitación en la capacidad bactericida de los neutrófilos (Gyang ci al., 1984;
Erskine et al., 1989). Pero ademásel NADPH es necesariopara reducir nuevamentela
glutation peroxidasaoxidada al detoxicar los peróxidos de hidrógeno,que en estaforma
oxidadaesinactivaa estosefectos(Gyangti’ al., 1984).
Por otra parte, algunos autores afirman que las rutas de la ciclooxigenasa y de la
lipoxigenasaen el metabolismodel ácidoaraquidónicosonseleniodependientes,y, por tanto,
podríanapareceralteracionesen la función reguladorade los macrófagospor alteraciónde
determinadosmetabolitosderivadosdel ácidoaraquidónicocomopor ejemplolaPOE,,la F,~,
y el leucotrienoB4 (Erskinees al., 1989; Eskewes al., 1989).
IJX.d.2- Influenciasobrela producción de anticuerpos
Existen descripciones sobre cómo el aporte de selenio extra en la dieta de ratones llega
47
ji

ini roduccitn
a provocar un aumento de entre 7 y 30 veces (según cantidad suplementada) en la producción
de anticuerpos(Miller, 1985>. Sin embargo,estasalteracionesen la producciónde anticuerpos
parecen estar condicionadas por la edad del individuo (citado por Turner y Finch, 1991). Así,
pollos de dos semanas mantenidos con una dieta deficiente en selenio o selenio-vitamina E
mostraron títulos menores frente a eritrocitos de oveja, sin embargo, si los animales tienen
una semana más, sólo los mantenidos con deficiencia en selenio-vitamina E mantuvieron esa
menor producción de anticuerpos. Otros autores citados por Turner y Finch (1991) han
empleado ratones y han observado que si son mantenidos con deficiencias en selenio se
produce disminución de la respuesta con IgG. pero no con IgM, lo cual indicaría que lo que
resulta particularmente sensible a esa deficiencia serían los linfocitos T-colaboradores. Para
poder observar una disminución de la producción de la IgM se tendría que mantener esa dieta
hasta la segunda generación (Turner y Finch, 1991), aunque si se utiliza una dieta deficiente
en selenio-vitamina E esa producción se ve alterada con mayor rapidez (Turner y Finch,
1991).
Por otra parte, las cantidades de selenio suplementadas en la dieta que producen una
mayor producción de anticuerpos, también están condicionadas a la especie del receptor, edad,
sexo y naturaleza del antígeno. Por ejemplo, Turner y Finch (1991) citan que las cantidades
de selenioen la dietaque producenun aumentode los títulos frentea eritrocitos de ovejaen
ratones jóvenes son inhibitorias en machos, pero no en hembras; asimismo, afirman que esa
elevaciónque seobservaen ratonesy cerdosjóvenesno seve en animalesmayores,y que
una dosis de selenito sódico en la dieta de corderos, que había dado los mejores resultados
frente a la toxina tetánica, dio unos resultados muy pobres cuando el antígeno empleado fue
el virus de la parainfluenza-3o Corynebactcrium.pseudotuberculosis.En este sentido estos
mismosautoresafirmanqueuna microdeficienciaen seleniono implica una menorrespuesta
de anticuerpossiemprequelos nivelesde vitaminaE semantengannormales(Turner y Finch,
1991). Otros autores (Blodgett es aL, 1986) afirman que el aumento en la producción de
anticuerposen animalessuplementadoscon selenioen su dieta, se produceúnicamentehasta
una determinada cantidad de este elemento, que ellos cuantifican en 0,9 mg/Kg. por encima
de la cual la tasa de anticuerpos desciende.
Sin embargo,existenautoresqueafirman no haberencontradodiferenciasen las tasas
de anticuerpos de animales alimentados con una dieta deficiente en selenio y animales
48

ini roduccion
alimentadoscon cantidadesadecuadasde esteelemento(ReffertStabel ti’ aL, 1989). Aunque
estos mismos autoresafirman que esto podría ser debido a que la deficiencia en selenio
provocadano fuera lo suficientementeintensapara ello. Por otra parte, estos autoressi
encuentrandiferenciasen las tasasde anticuerposel día 17 postinfecciónentre animales
deficientes y los controles, debido posiblemente a que los primeros tendrían una peor
eliminación del patógeno (en este caso P. haemolytica),y, portanto, sefavoreceríaunamayor
estimulación antigénica de los linfocitos B.
Estos mismos autores mezclan animales deficientes y no deficientes inoculados y no
inoculadoscon P. ltatrnoíytica, y observancómo los títulos de anticuerposen los animales
deficientes son superiores que los observados en los animales suplementados. Lo cual podría
ser debido a que los animalesdeficientespresentaríanmayor susceptibilidadfrente a una
infección bacteriana secundaria que los suplementados, o bien, a que la suplementación con
selenio acaba provocandouna disminución de la respuestainmune. Esto último está de
acuerdo con lo afirmado por otros autores (Murray y Murray, 1985), quienes llegan a la
conclusión que la deficiencia en selenio aumenta la supervivenciade ratones frente a
Plasmodiumbergeii, Listeria monocytogcneso frenteal virus de la pseudorrabia.
También Droke ti’ al? (1989) menciona la posibilidad de que se produzca algún tipo
de inmunosupresión en animales suplementados con selenio, ya que encuentran niveles muy
bajos de seroconversión en sus experiencias con P. hae¡nolvtica,aunqueno seextiendensobre
este panicular. Sí encuentran un aumento de las tasas de IgG frente a este agente aunque no
aprecian mejoría en el estado sanitario de los terneros tratados, lo cual podría ser debido a
las interaccionesde otros agentesresponsablesdel complejorespiratoriobovino.
1.IX.d.3- Influencia sobre la función secretora de las células del sistemainmune
Tanto los linfocitos como los macrófagos secretan una serie de sustancias que actúan
comomediadoresquímicosde la respuestainmune(Fig. 5). De entreéstoslos másconocidos
son las interleuquinas1 y 2. Las cantidadesde estassustanciassecretadaspor células de
animales delicientes y normales no presenta diferencias importantes (Kiremidjian-Schumacher
esal., 1990, 1992; Sordillo es al., 1993), porlo que estosautoresafirman que la secreción de
estos mediadores no es dependiente de los niveles de selenio existentes en el Organismo.
49

Introduccion
Por otra parte, parece existir un aumento en la producción de TNF-a en animales
suplementados (Kiremidjian-Schumacher er al., 1992), aunque estos autores responsabilizan
del mismo al aumento de macrófagoscitotóxicos en los animales suplementados.Sin
embargo,no parecehaberdatos del efecto de la deficienciaen seleniosobremediadoresel
TNF o las interleuquinas6 y 8 (Sordillo tt aL, 1993). Estas sustancias, con un importante
papel en los fenómenos de inflamación, podrían desempeñar también funciones quimiotácticas
sobre los neutrófilos, y podrían verse afectados por los niveles de selenio en el organismo.
En definitiva, no parecenexistir datos concluyentesy, por tanto, ésto necesitaaún ser
estudiadocon mayor profundidad.
1.IX.d.4- Influencia sobre la capacidad proliferativa de los linfocitos T y B
Una vezeí antígenoesprocesadopor los macrófagos,seunea las moléculasde clase
1 ó II del Complejo Mayor de Histocompatibilidad existentesen la superficie de los
macrófagos y son presentados a los linfocitos T, lo cual constituye la primera señal para la
activación de estasúltimascélulas(Fig. 5). Estemismoefectolo puedenrealizar,entreotras,
determinadaslectinas mitogénicas vegetales como la concavalina-A (Con-A) y la
fitohemaglutinina (PHA). Sin embargo, para que se produzca la estimulación máxima es
necesariala presenciade un factor de co-estimulacióncomo la ILí. Como respuestaa esa
estimulación,una subpoblaciónde linfocitos T produce 1L2, mientrasotra subpoblación
expresa receptores para esa última. Finalmente, la unión entre la 1L2 y sus receptores provoca
unaseñalparaque las célulasque poseenestosreceptorescomiencena proliferar.
Utilizando mutágenos específicos para los linfocitos T, como la concavalina A (Con
A) o la fitohemaglutinina(PHA), seha comprobadoqueen poííosy animalesmonogástricos
la deficienciaen selenioprovocaunamenorrespuestade las célulasT (Turnery Finch, 1991;
Kiremidjian-Schumacheret aL, 1992). Esta menor respuestade los linfocitos T seve mucho
másdisminuidasi la deficienciaesen selenioy vitaminaE, e incluso esmásevidenteen este
casoque la alteraciónqueseproduceen la respuestade los linfocitos T a causade unamenor
ingestade alimento (citado por Turner y Finch, 1991).
Como los niveles de ILI y 2 no se ven afectadosde forma diferente por una
deficienciao suplementacióncon selenio,y ademásla función presentadorade antígenosde
50

Introducción
los macrófagosno seencuentraalteradatampoco(Kiremidjian-Schumacherti’ al., 1990),esta
menorrespuestapuedeserachacadaalas célulasaccesorias.Estos autores(Ro>’ et al., 1992)
han observadocómo, efectivamente,esta alteración de la respuestablastogénicade los
linfocitos T sedebea que la expresiónde los receptoresde 1L2 seencuentraalterada,hasta
el punto de que en caso de deficiencia estaexpresiónsc lentifica en gran medida. Esta
lentificación sedebea que en casode deficienciaen seleniose producenun menornúmero
de receptoresde alta afinidad (Roy ti’ aL, 1993), primandolos de baja afinidad ya que el
númerode receptoressemantieneconstanteindependientementedel selenio(Roy esal., 1993;
Sordillo ni al?, 1993), por lo que se necesita un tiempo mayor para alcanzar la concentración
umbralnecesariaparacomenzarla proliferaciónde los linfocitos T.
También se han utilizado mutágenos específicos de los linfocitos B como el
lipopolisacárido (LPS) de E. cali, con el que seha podido observaruna menorrespuestaen
animalesdeficientesen selenio-vitaminaE (citado por Turner y Finch, 1991).
Sin embargo,pareceque esta respuestaa mutágenospor parte de los linfocitos
dependede la edad,ya que mientrasque ovejasadultasmantenidascon una dieta deficiente
en selenioy vitaminaE presentanuna buenarespuestaa PHA, no ocurrede igual maneraen
corderos,en los que sí seproduceun fallo en la respuestade linfocitos T (citado porTurner
y Finch, 1991).Estosmismosautoresexponenunahipótesisa esterespecto:en ovejasadultas
los microorganismosexistentesen el rumenposeenunascantidadesdeseleniomuy altas,que
podrían ser puestasa disposición de las células inmunocompetentescuando ésto ft¡ese
necesario.
Esta alteración en la proliferación de los linfocitos, puede ser detenidae incluso
invertidamediantela inyecciónde selenito sódicoal animal o la adición de ésteal medio de
cultivo de las células.Esto indicaríaque el fallo radicaríaen el ambientecelular másque en
la célula en sí (citado por Turner y Finch, 1991). Pero otros autoreshan advertido de la
posibilidad de que aparezcanaberracionescromosómicascuandose administraselenio en
cantidadessuperioresa las necesarias;las cantidadesnecesariaspara que los linfocitos
presentenuna mayor respuestason de 1 ng/ml de medio de cultivo, con cantidadesde 10
ng/ml sealcanzauna meseta,y con 1 pg/ml se produceuna espectacularcaída(Citado por
Turner y Finch, 1991). Incluso se señalaque la adición de 6 ng Se/ml al medio de cultivo
51

introducción
provocala apariciónde un 13% de células aberrantes,porcentajeque llega al 40% si las
cantidades añadidas son de 600 ng/ml.
1.X- SELENIO Y RESISTENCIA A ENFERMEDADES INFECCIOSAS
Hemosvisto como la deficienciaen selenioprovocala disminuciónde la respuesta
inmune en cadauno de sus aspectos,pero veremosahoralo que ocurrecuandotodos estos
mecanismosactúanconjuntamente.
La suplementacióncon selenio-vitaminaE durante los periodos de gestación y
lactación en cerdaspodría disminuir las pérdidaspor el síndromemastitis-metritis-agalaxia
porcino (Whitehair,citado porMiller, 1985).Erskine ti’ al? (1987) describeque la leche de
los animales suplementadoscon selenio y vitamina E presentabanrecuentoscelulares
menoresque los no suplementados,y que estosanimalesresistíanmejor a las infecciones
mamariaspor Streptococcusagalactiae y Si’aphylacoccusaureus. Además,las mamitis en
estos animales tenían una menor duración, así como una menor gravedad de los signos
clínicos. Estos autoresafirman que estehechose debe a que en los animalesdeficientesse
produceunamenoractividadGSHPx,y esto implicaríaunamenoreficaciabactericida,sobre
todo si la bacteriaingeridapresentaalgún mecanismoparadegradarlos peróxidosformados
durante la fagocitosis.
Lo expuestoanteriormentesevio confirmadocon estudiossobremamitis provocadas
por E. ccli (Erskine ti’ aL, 1989).Estosautorescomprobaroncómo,nuevamente,los animales
suplementados con selenio y vitamina E presentaban menos problemas de mamitis por este
agentepatógenoque los no suplementados,siendo menor la frecuenciade apariciónde la
mamitis y también menor la duración de la misma. Atribuyen este hecho a una mejor
movilización de los neutrófilos desdeel torrente circulatorio, y a una mayor capacidad
fagocitica de los neutrófilos observadain vitro. Los neutrófilos de vacas deficientes
demostraronuna menor capacidadbactericidatanto frentea CarnUda albicanscomo frente
a 8. aureus.Sin embargo,en estudiosrealizadospor el mismo autor con posterioridadno
llega a estamisma conclusiónsobre el efecto del selenio en las mamitis por S. aureus
(Erskine eS al., 199<)), ya que en esteestudiono vio diferenciasentreel grupodeficiente y
el suplementado,aunquejustifica esto en que mientrasque en las mamitis por E. ccli son
52

Introducción
bastantefrecuenteslas formasagudasde la enfermedad,en las producidaspor S. aureusno
lo son tanto, y ademásesteúltimo presentauna mejor adaptacióna la mama.
Porotro lado, Braun ci aL (1991) confirma la mayorgravedadde las mamitis en los
animales selenio deficientes, ya que la suplementación con selenio produce un aumento del
númerode leucocitosque llegan a la glándula, y, por tanto, a un aumentode la fagocitosis
queproduceun descensoen el númerode bacteriasviables.
Tambiénen otrasenfermedadestransmisiblesseapreciauna mejor respuesta,como
ocurre en los poííos con la coccidiosis (Colnago er al., 1984a). Estos autoresdescriben
cambiosen el númerode leucocitoscirculantesen poííos inmunizadosinfectadoscon distintas
especiesde Eirneria cuandoselessuplementala dietacon selenio.Asimismo observancómo
una suplementaciónconselenioy vitamina E produceun aumentoen la gananciade pesode
poííos no inmunizados frente a E. tenella, así como una disminución de la mortalidad
(Coinagoesal., 1984b).Estamejorade la respuestainmune pareceafectara los mecanismos
de defensageneralfrentea infestacionesprimarias.Llegan incluso a exponersu sospechade
que estasmejorías sondebidasa un aumentode la fagocitosis.
Sin embargo,la concentraciónde seleniono tiene incidenciasobrela infestaciónpor
Haernanchuscantari’us en oveja(Jelinek eS al?, 1988), aunqueel mismo autor lo atribuyea
que las ovejasselenio-deficientesutilizadasen su experienciapresentaronamplias reservas
de a-tocoferol,que podría habercompensadola deficienciaen selenio. Por otra parte, la
utilización de dietassuplementadascon seleniosí parecentenerinfluenciasobrela infección
por Seipulina hyodysenteriae, anteriormenteincluida en el géneroTi-eponerna,(Teige ci al.,
1984), ya que la resistenciaa esteagentefue mayoren los grupossuplementadosque en el
no suplementado.Si bienhemosde señalarla apariciónde un resultadocuandomenoscurioso
muy similar al descritopor Blodgett ti’ aL (1986) en producciónde anticuerpos,ya que el
grupo al que se administró la cantidad más alta de selenio no presentó diferencias
significativas con el no suplementado, lo que nos confirma la sospecha de que el aporte de
selenioporencimade determinadascantidades(0,8-0,9mg/Kg) no producebeneficioalguno.
Con lo expuestohastael momentoparececlaro que la deficienciaen selenioafecta
a 1-a función de los neutrófilospolimorfonuclearesasociadaa los cambiosfisiológicos en los
53

ini roduccion
niveles de GSHPx. Los neutrófilos de vacas mantenidasexperimentalmentecon dietas
deficientes en selenio presentan un menor consumo de oxigeno, así como menores actividades
de GSHPxqueen el casode neutrófilosde vacascon una dietanormal (Erskine ti’ aL, 1987,
1989). Estemismoautorha comprobadoquela deficienciaen selenioprovocala disminución
en la producción de leucotrieno B4, un metabolito de la lipooxigenación del ácido
araquidónicopor los netrófilosqueademásactúacomoun potentequimiotácticoy estimulante
quimiocinético paralos neutrófilos,lo que provocauna disfunción de éstos (Erskine ti’ al?.
1989).
Estoscambiospuedenhacerqueun animal deficienteseamássensiblea enfermedades
infecciosas,aunquepor el momento no pareceexistir pruebaalgunaacercade una mayor
incidenciao gravedadde enfermedadesinfecciosasentreanimalesnaturalmentedeficientes
en selenioo vitaminaE. Los neutrófilosde animalesdeficientespierdenpartede su capacidad
parafagocitaralgunosmicroorganismos,aunqueno esconocidala relevanciaque éstopueda
teneren las infeccionesnaturales.
En resumen,podemosafirmar que unadeficienciaen seleniosetraduceen una menor
capacidaddel sistema inmune, y. por tanto, en una mayor susceptibilidadpara padecer
enfermedades.
54

2. OBJETIVOS

Objetivos
2.OBJETIVOS
Ya hemosapuntado anteriormente que la listeriosis precisa de una serie de factores
predisponentes.Estos factoresconducena un estadode inmunodeticienciamás o menos
acusado,bien por la existenciade un estadopatológicosubyacente,por actuaciónmédicao
de forma fisiológica. Sin embargo,estosfactoresconocidosno soncapacesde explicar todos
los casosque seproducen,esdecir, debenexistir otros factorespredisponentesdesconocidos
por el momento como tales. Entreestosúltimos cabría incluir los estadosde deficiencias
nutricionales tanto las que cursan en forma manifiesta, que haría fácil estableceruna
correlacióncausa-efecto,como las microdeficiencias,es decir, todo aquél estadoen el que
existeuna deficienciade un determinadoelementoque no essuficientecomoparaprovocar
alteracionesvisibles.
A pesarde los puntososcurosexistentestodavíaen la patogeniade la listeriosis,existe
un convencimientogeneralde que la principal vía de transmisiónes la oral, ya que en los
últimos brotes ocurridos en el hombre y los animales la infección se ha producido a través
de alimentos.Sin embargo,la mayoríade los autoresprefiereutilizar la vía intragástricacomo
vía experimentalde infecciónal requerirdosisalgo inferioresque la vía oral. Consideramos
como vía oral aquellaen que el individuo ingiere las listerias con el aguao el alimento,
permitiendoel contactode las bacteriascon todo el tracto digestivo.
Además,los datosepidemiológicosexistentesmuestranuna mayor prevalenciadel
serovar4h que del 112a en los brotes, aunqueno existendatos que confirmen una mayor
frecuencia de aislamientos de 4b que de l/2a de muestras ambientales.
Por todo lo expuesto,decidimosinvestigar,utilizandoel modeloexperimentalmuríno:
1- las repercusionesqueunamicrodeficienciaen selenioy vitaminaE podríateneren
la susceptibilidada la listeriosis,paralo que comparamosanimalesdeficientesy no
deficientes;
2- la dinámica de la infección y la mortalidad por vía oral, en comparación con la vía
endovenosacomoreferenciay con la vía intragástrica;
56
.1,

Ohjcti <O,s
3- la repercusiónen la infecciónque podríatenerla inoculaciónoral con una única
dosiso con dosisrepetidasduranteun cierto períodode tiempo;
4- la existenciade diferenciasen la infecciónpor vía oral con los dos serotiposmás
frecuentesen los brotesde listeriosis (4b y l/2a).
5?

3. MATERIAL Y METODOS

Material y nétodos
3.1- MATERIAL
Para la realizacióndel presentetrabajo utilizamos el material generalpropio de un
laboratoriode microbiología,esdecir, placasde petri, tubos de ensayode distintostamaños,
tubos estériles de polipropileno, matraces,mecheros,asasde platino, gradillas, pipetas
ordinariasde distintasmedidas,pipetasPasteur,pipetasautomáticasdosificadorasde volumen
variable (de (1 a 1 mí, y de 0 a 200 pl) con sus puntasdeshechablesrespectivas,frascosde
vidrio, probetas,filtros estérilesde 22 pm de un solo uso, etc. Asimismo, hemosutilizado
jeringasestérilesde 1 ce (insulina)con sus agujasparalas inoculacionesendovenosas.
En cuanto a los aparatos empleados,también han sido los de uso rutinario en un
laboratoriode microbiología,comoautoclaves,estufasde incubación,cámarade refrigeración,
arconescongeladoresde -20 y -800C, centrífugas,baños-maría,balanzas,microscopioóptico,
agitadores-calentadoresmagnéticos,etc.Entretodosellos,debemosresaltarel usode una lupa
estereoscópicaWild 376788, básicaa la hora de reconocerlas presumiblescolonias de
listerias, así como el de un homogeneizadorStomacherLUL Tipo 0480 que nos facilitó la
labor de homogeneizaciónde las muestras.
3.11- MEDIOS DE CULTIVO
- Caldo para la obtención del inóculo
Brain Heart Infusion (BW) (Difeo) 37 g
Agua destilada 1000 ml
Una vez disuelto el medio se separanunos 100 ml a otro matraz,y seesterilizana
1210C 15 minutosambosmatraces.
- BHI Agar (medio de recuento)
BHI (Difeo) 37 g
Aga 15 g
Agua destilada 1000 ml
59

Material y métodos
Se disuelvenperfectamentetanto el BRI corno el agar en el agua y seesterilizan a
1210C 15 minutos.Trasla esterilizaciónsedejaenfriarel medio hastaunos 45-500C
y sedistribuye en placasde petri.
- Listeria SelectiveAgar Medium modifled (LSAMm)
BHI (Difco) 37 g
Cloruro de litio 15 g
Esculina 0,75 g
Citrato amónico-férrico 0,5 g
Acriflavina 5 mg
Polimixina 10 mg
Ceftazidima 20 mg
Telurito potásico 40 mg
Agar 15 g
Agua destilada 1000ml
En 90<) ml de agua se disuelvenlas cantidadesindicadasde BRI, cloruro de litio,
esculina,acriflavina y agar; mientras que en los 100 ml restantesse disuelveel citrato
amónico-férricoen otro matraz.Estosmatracesseesterilizana 1210Cdurante15 minutos,tras
lo cual sedejanenfriarhastaunos500C, semezclany seañadenlos restantescomponentes.
A continuaciónseprocedea su distribuciónen placasde petri.
3.111-SOLUCIONES
- Solución salina
Cloruro sódico 9 g
Agua destilada 1000 ml
- Solución de polimixina
Sulfato de Polimixina B (Sigma) 1 g
60

Material y métodos
Agua destilada 100 ml
De estasolución seañaden1 mIlL de LSAMm.
- Solución de ceftazidima
Ceftazidima sódica pentahidratada (Glaxo) 2 g
Agua destilada 100 ml
Para preparar1 L de LSAMm sele añaden1 ml de estapreparación.
- Solución de telurito potásico
Telurito potásico 4 g
Agua destilada 100 ml
Se utilizaba 1 ml para preparar 1 L de LSAMm.
- Solución tamponada de formalina
Formol 37-40% (Panreac) 10<) ml
Fosfato sódico dibásico anhidro (Merck) 0.8 g
Fosfato disódico anhidro (Fluka) 1,3 g
Agua destilada 900 ml
- Solución de gelatina-alumbre
Solución A: Gelatina (Panreac) 2,25 g
Agua destilada 50<) ml
Solución B: Timol (Merck) un cristal
Detergente (Mistol) dos gotas
Alumbre crómico ¡ KCr(504t. 1 2H20]
61

Material y métodos
(Merck) 0,733 g
Agua destilada 19,3 ml
Se preparala solución A a 55-600C y, una vez disuelta la gelatina, se añadela
soluciónB.
- Solución de Hematoxilina de Mayer
Hematoxilina (Merck) 1 g
Agua destilada 1000 ml
Yodato sódico(Merck) 0,2 g
Alumbre potásico[(S04)2KAl.1 2H20] (Merck) 50 g
Acido cítrico (C. Erba) 1 g
Hidrato de cloral (C. Erba) 50 g
Disolver el alumbre en el agua,añadir y disolver la hematoxilina.A continuación
añadir el yodato sódico, el ácido cítrico y el hidrato de cloral y agitar hastasu total
disolución.La soluciónresultantepresentaunacoloraciónrojo-violeta,y semantieneestable
durantevarios mese.Es importanteseguirel ordendado en su elaboración.
- Solución de eosinaal 1%
Eosinaamarillenta 1 g
Agua destilada 100 ml
Acido acético 2-3 gotas
3.1V- CEPAS
Para la realizacióndel presente trabajo hemosutilizado, básicamente,una cepa de L.
monacytagenesde origen clínico denominadaP-14B,pertenecienteal serovar4b. Estacepa
fue aisladaa partir de un enfermode listeriosis duranteun pequeñobrote humanoocurrido
en Valencia en 1989 (Vázquez-Bolandti’ aL, 1991).
62

Matecial y i~tétodos
Para los estudiosde patogenicidadcomparadase utilizó, además,la cepatipo de L.
nwnocyi’ogenes 1/2a NCTC 7973.
3.V- ANIMALES DE EXPERIMENTACION
Los animales utilizados para la presenteexperiencia fueron ratones de tipo Swiss de
la estirpeCD- 1, criadosen nuestrolaboratorio.Las edadesde estosanimalesoscilaronentre
las 10 y las 12 semanasal inicio de cada experimento, lo que significa un pesoaproximado
entre 20 y 25 gramos/ratón.
3.VI- ALIMENTO Y AGUA DE BEBIDA
3.VI.a- RATONES NORMALES
El pienso empleado para la alimentación de los animales normales fue un pienso
comercialelaboradopor la casaInterfaunaIbérica S.A., cuyacomposición,facilitada por el
fabricantees:
Cenizas 58 g/Kg
Grasa bruta 21 »
Proteínabruta 145 »
Fibra bruta 50 »
Carbohidratos 59 »
Energíadigestibleen MJ/Kg 123
En cuantoa las cantidadesde selenioy vitamina E. éstasfueron de 0,09 mg/g y de
9<) UIJKg respectivamente.
El aguaempleadaparaestosratonesfue aguacorriente.
3.VI.b- RATONES DEFICIENTES EN SELENIO Y VITAMINA E
El piensoutilizado paraobtenerratonesdeficientesen selenioy vitamina E fue un
63

Malerial y métodos
pienso especialsuministradopor PanlabS.L., con menosde 0,03 mg/Kg de selenioy no más
de 8 mg/Kg de vitaminaE.
El aguaempleadaparaestosanimalesfue aguaultrapura (MilliQ) suplementadacon
cloruro sódico(2,8 g/]) y glucosa(5 g/l).
Para saberel tiempo necesariopara obtener animales deficientesserealizaron análisis
de selenioen hígado durantevarias semanas.De esta manera,observamosque tras seis
semanas con esta dieta las concentraciones de selenio én este órgano eran muy inferiores a
la de los animalescontrol, sin que se observaransíntomaalguno de la deficiencia, ni se
pudieraobservaralteraciónhistológicaalguna.Tambiénserealizaroncontrolesconsistentes
en compararel tiempo necesarioparaqueseprodujerahemólisis evidenteen una muestrade
sangre.Estetiempo fue siemprede 48-72horasmenosen los deficientesque en los controles
utilizados.
3.VII- OBTENCION DEL INOCULO
La cepa utilizada era sembrada en BHI Agar y se incubaba a Y/0C durante 18-24
horas,tras estaincubaciónse comprobabala purezadel crecimientomedianteobservación
macroscópicay tinción de Gram. A continuación,seinoculabaun matrazcon 100ml de BHI
que se incubabaen agitaciónorbital otras 18-24horasa 370C.
Estematrazera utilizado parainocularotro con 9<30 ml de BHI, despuésde comprobar
medianteuna tinción de Gram su pureza.También este matraz se incubabaa 370C en
agitaciónorbital durante18 horas.Transcurridoestetiempo se centrifugabaa 10000 g. De
estamanerapretendíamosobtenerlas bacterias,por lo que despreciábamosel sobrenadante.
El sedimentoeraresuspendidoen soluciónsalinaestérily nuevamentecentrifugadoa lOOOOg
paraeliminar todo el medio.
Nuevamentedespreciábamosel sobrenadanteconservandoel sedimento,que era
resuspendidoen 80 ml de solución salinaestéril. Una vez resuspendidoel sedimentoera
alicuotado en tubos estériles de polipropileno (2 ml/tubo) y almacenado en congelación a -
800C hastael momentode serutilizado.
64

1
rg 9~o
~ci
EN
Ococ\J
4?
<-5clE
NO
a
<~
,~
EI~
O)oo
LO
(tOr
(~x
..JC\1
1
oe’E
NO
o,1
oo’o
1-
a,0c2
a)0
6oNoO
1-
-EoooO-o0ÑO
1-Ea)oE

Material y actodOs
Antesde su congelaciónserealizó un recuentode las bacteriasasí obtenidas,paralo
cual serealizabandiluciones decimalesde la suspensiónde bacterias~,~sembtandose100 iaL
de las que seestimabanadecuadasen BRI Agar. Estosrecuentosdieron siempreun resultado
muy similar, que osciló entre lxlO’0 y i3xlO~0 UFC/ml. Paraasegurarnosque la congelación
a -800C no afectabade forma importantea nuestrasbacterias,realizamoslos recuentos
despuésde la congelaciónsiguiendo la misma metodologíaya reseñada.Los resultados
obtenidosno variaronsustancialmentede los obtenidosantesde la congelación.
De forma esquemáticapuede verse el proceso en la figura 6.
3.VIII- INOCULACIONES
3.VIII,a- INOCULACIONES ENDOVENOSAS
Dos díasantesde la inoculaciónse descongelabaun tubo de inóculo y se realizabaun
recuentodel númerode bacterias/mí,mediantedilucionesdecimalesy siembrade 10<) pl en
BHI Agar. De estaforma podíamossaberel númerode bacteriasque habíaen los tubos
congelados, y preparar el inóculo cada vez con la dosis lo más exacta posible.
El día de la inoculación se descongelabael númerode tubos necesariossegúnel
recuentorealizadocon anterioridad,y se hacíanlas diluciones necesariasparaobtenerlas
dosisrequeridascon gran exactitud.
Para poder inocular las concentracionesrequeridas se inocularon 100 pL de una
suspensióncon una concentraciónde bacterias10 vecesmayor a la requeridaen la vena
dorsaldel apéndicecaudal de los ratones(figura 7).
Porotra, paite, los ratonescuya inoculaciónno fue perfectafueron desechados,lo que
aumentala fiabilidad del experimento.
3.VIII.b- INOCULACIONES ORALES
Tambiénen este caso se descongelabaun tubo de inóculo vados días antes de la
66

Material y métodos
inoculación,parapoderhacerun recuentodel númerode bacteriaspor ml existentesen el
inóculo. La metodologíaempleadafue la misma que la expuestaen el apartadoanteriorde
inoculacionesendovenosas,esdecir, mediantedilucionesdecimalesy siembrade 100 pl en
BHI Agar.
El día de la inoculación sedescongelabael número de tubos necesariosy sepreparaba
la cantidad necesariade inóculo, administrándosecomo agua de bebida. El inóculo se
mantuvoduranteunanoche,tiempo tras el cual secambiabanlos biberones,siemprevacíos,
porlos deaguanormal.En los casosde inoculacionesrepetidasdurantevariosdías,el inóculo
era renovadodiariamente(figura 8).
Para calcular la cantidad de inóculo por ratón y día medimos la cantidad de agua
bebida por un ratón durante vados días en distintas épocasdel año, según nuestras
estimacionesesacantidadfue de unos5 ml/ratón y día.
3.IX- SACRIFICIOS Y OBTENCION DE LOS ORGANOS
Los ratonesfueron sacrificadosmedianteinhalación de eter etílico, procediéndose
inmediatamentea la recogidade los órganosa estudiar.Paraello en primer lugarseabríala
piel de la zonaventral desdela unión de las ramasde la mandíbulahastala región inguinal,
a continuaciónseabríala musculaturaabdominalpor la línea blancahastala última costilla.
Una vez abierta la cavidadabdominalseretirabael paqueteintestinal haciaun lado, y, con
ayudade una jeringa de insulinaen ocasionesheparinizada,serecogíahasta1 ml de sangre
de la vena cavainferior (imagen 1). Estasangrenos servíaparacomprobarsi los animales
deficienteslo eran realmenteal compararel tiempo de hemólisisde éstosy los controles.
Con la sangreya recogida,procedíamosa la obtencióndel resto de muestrasen el
siguienteorden.
- Bazo;
- Hígado;
67

o
Figura 7-Inoculaciónintravenosa
(4b).
0.1 ml 0.1 ml 0.1 ml 0.1 mJ
5 mI/ratón ‘
Figura 8-Inoculaciónpor vía oral.
Sol, madre lOx dilución
deseada
0.1 ml
Sol, madre Diluciónc~’ deseada‘4~~•~

5 ratones/grupo
de muestras.
Figura 10- Cálculode la OLSO por vía
intravenosa (4b).D= de fici enles
bazohígadoa mesenlérteosp. digestivog, submandibularorofaríngeesófagoencéfalo
9- Sacrificio yFiguratoma
lo
lo
DOSIS7
lo6,5
6
5,5lo
5
lo4,5lo4lo a,s
lo3
lo
102.5
E


Material y métodos
- Ganglio mesentérico;
- Aparatodigestivo: en primer lugarse cortabaa la altura de la ampollarectal y del
cardias,extrayendotodo eí estómagoe intestinoconjuntamentea unaplacade petri
estéril, y, a continuación,serecogíacadamuestrade forma independiente.
1- estómago;
2- intestinodelgado,que eracortadoen trozosmáspequeñosparafacilitar su
homogeneización;
3-intestino grueso,al igual que el delgado,también se cortó en trozos mas
pequeños.
- Gangliosubmandibular,conjuntamentecon las glándulassalivaressituadasen esa
región.
- Región orofaríngea:para recogeresta zonacortábamoslos músculosde la zona
inferiordel cuelloa la alturade susinsercionesen las ramasmandibulares,extrayendo
la lenguahastallegar a la faringe. De estaforma se pretendíatomar unaporción de
las formacioneslinfoides del anillo orofaringeo.
- Esófago.
3.X- CALCULO DE LA DOSIS LETAL 5<> (DL50) POR VíA ENDOVENOSA
Paraestecálculoseinocularon,porvíaendovenosa,diez. lotes de 5 ratonesdeticientes
y 5 no deficientescadauno, con distintas dosis.Las dosisempleadasfueron: l0~, 106.5, 106,
~ lOt ~ 10~, ~ í<V y 1025 UFC/ml (figura 10).
Estoslotes fueron observadosdurante10 días,anotándoseen cadamomentoel número
de muertospor dosis.En los animalesmuertosseprocedióa realizaraislamientode listerias
a partir del hígado,bazoy encéfalo,ya que sólo considerábamoscomomuertospor listeriosís
>71

Material y métodos
aquellosen los que seconsiguióaislamientode algunode éstosórganos.
La extracción de los órganos se realizó tal como se explica en el apartado
correspondiente.
Los órganos una vez extraídoseran introducidos en bolsas de homogeneización
estériles.Estasbolsashabíansido pesadaspreviamente,con lo que con una nuevapesaday
medianteuna sencilla operaciónaritméticapodíamosconocerel pesodel órgano. Una vez
pesadose homogeneizabaen un Homogeneizadortipo Stomacher,y se diluían los órganos
al 1:10 (p:v) en soluciónsalinaestéril. A partir de estaprimeradiluciónserealizabannuevas
diluciones decimalesy se sembraban100 microlitros en placasde LSAMm que eran
incubadasa 370C durante48 horasantesde procederal recuentode las colonias.De esta
sencillamanerapodíamosconocerla cantidadde listeriasexistentesen un gramodel órgano
en cuestión.
Utilizamoscomo medio de aislamientoLSAMm, ya que al existir la posibilidadde
tenerquerealizaraislamientosa partir de animalesmuertosla probabilidadde quecrecieran
bacteriascontaminanteserabastantealtasi utilizábamosmediosgenerales.Porotraparte,esto
tambiénevitó el crecimientode bacteriascontaminantesen los cultivos a partir de intestino.
Unavez conocidoel númerode animalesmuertospor dosisseprocedióal cálculo de
la DL5O, esdecir, aquelladosis en la que moriríanel 50% de los animalesinoculados.Para
estecálculoutilizamosel métodode Karber (Schmidt y Emmons,1989). Estemétodoutiliza
la siguientefórmula:
<I.og de la dilución más aila utilizada)-
suíía del ‘4 dc. mortalidad
en cada diiución iog del faclor
0.5 ) x ( óe dilución 1loo
3.XI- ESTUDIOS DE MORTALIDAD POR VíA ORAL CON DOSIS UNICA
Paraestosestudiosseinoculabancuatrogruposde 48 ratonescon las dosisa estudiar,
22

Matehal y ¿lodos
100 y ío~ UEC/ml, incluyendodos grupo/dosis,uno de deficientesy otro no deficiente.Una
vez inoculadosse les dabasu piensoy aguacorrespondiente,y se les observabadurante10
días.Con los muertosseprocediótal y comoserefleja en el apartadoanterior (figura II).
3.XII- ESTUDIO DE MORTALIDAD POR VIA ORAL CON DOSIS REPETIDAS
En estecaso seutilizaron seis gruposde 48 ratonescadauno, de los cualestres eran
de deficientesy los otros tresde no deficientes,que fueron inoculadosde la siguientemanera
(figura 12):
lote 1- ut UFC/ml 4 días;
lote 2- ío9 UF~/ml 7 días; y,
lote 3- ío~ UFC/ml 10 días.
En todos los casosel inóculo era preparadodiariamente,de tal maneraque en los
bebederossólo seponíala cantidadnecesariaparaun día
El estudioseprolongó durantediez días desdela última toma del inóculo. Sólo se
tomaronen consideraciónaquellosratonesmuertosque dieron un cultivo positivo a partir de
hígado.
3.XIII- SEGUIMIENTO DE LA INFECCION POR VIA ENDOVENOSA
Con estosexperimentosse pretendíaconocerla dinámicade la infección en los dos
tipos de ratones(deficientesen selenioy vitamina E y no deficientes)ante una infección
experimental.Paraello se inocularonlotes de 24 ratonesde cadatipo con las DL50 de cada
tipo (l0~~ y ío~~ UFC/ml respectivamente).Tras la inoculaciónsesacrificaronlos ratonesa
los siguientestiempospost-inoculación:30 minutos, 12 y 24 horas,y 2, 3, 4, 5, 6, 7, 8, 9 y
1<) días (figura 13). El númerode ratonessacrificadoen cadatandafue de dos,de los cuales
se realizaroncultivos a partir de hígadoy bazo.
Los órganoseranextraídosde forma aséptica,e introducidos en bolsasde plástico
estérilesquehabíansido pesadaspreviamente.De nuevosepesabanlas bolsas,ahoracon el
73

DOSIS
1O
1O a
Figura 11-Mortalidad por vía
oral con dosis única(4b).
D= ótIicie,~tes
DOSIS YTIEMPO
10 4 días
10 7díasa
10 lOdías
D
48 ratones/grupo
48 ratones/grupo
Figura 12- Mortalidadpor vía oral con dosis
repetidas (4b).D= deficientes

4,5lo 5,5lo30 mm.
12 h24 h2d3d4d5d6d7d8d9d10 d
Figura
D
24 ratones/grupo
13- Seguimientode la infección
por vía endovenosa.0= deficientes.

Material y métodos
órganoen cuestiónen suinterior, y medianteuna simple operaciónaritméticacalculábamos
el pesodel órgano extraído.Una vez conocidoestedatose homogeneizabacon ayudadel
Stomachery sediluía al 1:10(p:v), realizándosesiembrasapartir de dilucionesdecimalesdel
homogeneizado,paraver, no sólo la presenciao no de listerias, sino podercuantificarésta
presencia.
3.XIV- SEGUIMIENTO DE LA iNFECCION POR VIA ORAL
3.XIV.a- SEGUN LA VIA DE INOCULACION EMPLEADA
Paraesteestudioseutilizaronlas vías oral e intragástrica.Parala primeraseprocedió
exactamenteigual acomosehadescritoen la experienciadel seguimientode la infecciónpor
vía oral, esto es, se administró el inóculo con el agua de bebida. En el caso de la vía
intragástricase utilizó una sondade extremoromo paradepositarel inóculo en el estómago.
Como control de la correctainoculaciónse realizaroncultivos de pulmónde los animales
sondados.
El inóculo empleado fue de io~ UFC Lm/ml, en amboscasos,y las muestrasfueron
recogidasa las 6, 12, 24 h, 2, 3, 4, 5, 6, 7, 8, 9 y 10 díaspost-inoculación.Estasmuestras
fueron hígado, bazo, ganglios mandibular y mesentérico, región orofaríngea,esófago,
estómago,intestinodelgadoy grueso.
El número de ratones fue de 44 animalespor grupo, no se emplearonanimales
deficientes (figura 14). En cada momento se sacrificaron 4 animales/grupomediante
inhalación de eter etílico, de los cuales2 se destinaronal estudio microbiológico y 2 al
estudiohistopatológico.
IXIV.h- SEGUN LA CEPA EMPLEADA
En estecasoseinocularondos cepasdeL. monocysogenes(P-14B y l/2a)porvía oral
únicamente.Como en los casosanterioresla inoculación se realizó medianteel aguade
bebida,y la dosisinoculadafue de iO~ UFC/ml (figura 15).
76

9
10
6h1224k oral2d3d4dBd3d7d3d9dlo d
Figura 14-Comparación de lainfección por las
vías oral eintragástrica (112a).
o10
6h1224k 4b2d3d4d5dCd 44 ratones/grupo7d3d9d 1/2a10 d
Figura 15-Comparación de lainfección con los
servares 112a y 4b.
44 ratones/grupo

Mater4al y métodos
Seutilizaron44 ratones/cepa,tampocoen estecasoseemplearonanimalesdeficientes.
Tanto los tiempospost-inoculacióna los que sesacrificaron,como el métodode sacrificio,
como las muestrasrecogidasfueron los mismosque en el caso antenor.
3.XIV.c- SEGUNDOSIFICACION: UNICA O REPETIDA
Para estaexperienciase emplearondos lotes de 48 animalesdivididos en cuatro
gruposde 24 ratones,dos de deficientesy dos de controles.Los cuatrogruposbebieronun
inóculo de IO~ UFCImI, pero mientrasqueel lote 1 lo hizo únicamenteduranteun día,el lote
2 lo hizo durante7 días.Los tiempos a los que seprocedióal sacrificiode animalesfueron
1, 3, 6,11 y 16 días desdela última inoculación(figura 16).
Se sacrificaron5 animalescadavez, aexcepciónde la última en que se sacrificaron
4. De los 5 animales3 fueronmuestreadosparamicrobiologíay 2 parahistología,mientras
queen el último sacrificiosólo se muestrearon2 paramicrobiologíamanteniéndoselos otros
2 parahistología.Las muestrastomadasy su procesamientofue el mismo que en los casos
anteriores.
3.XV- SEGUIMIENTO DE LA INFECCION POR VIA ORAL EN RATONES
DEFICIENTES/NO DEFICIENTES
3.XV.a- ESTUDIO DE LA INFECCION POR VIA ORAL CON DOSIS UNICA
En estecaso empleamosdos grupos de 54 ratones,uno de deficientesy otro de
control, que bebieronun inóculo de l0~ UFC/ml duranteun día. Los sacrificiosserealizaron
a los siguientestiempos post-inoculación:30 mm, 6, 12 y 24 h, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11
y 16 días (figura 17).
Se sacrificaroncuatroanimalesporgrupocadavez,de éstosdos fueronutilizadospara
microbiología y los dos restantespara histopatologiae inmunohistoquimica.El muestreo
realizadofue más amplio que en el caso anterior, tomándosemuestrasde bazo, hígado,
gangliomesentérico,gangliosmandibulares,región orofaríngea,esófago,estómago,intestino
delgadoe intestino grueso.
78
II

9
lo idía9
10 7 días
Días desdela última toma
ola infección
según el tiempode bebida del
inóculo.
1361116
24 ratones/grupo
Figura 16-C mparación de
D= deficientes

10 54 ratones/grupo
30 mmCh
24h N2d D3d4dCdCd7dAd9d
10 dlid Figura 17- SeguimientolSd de la infecciónpor vía
oral con dosisúnica(4b).
0= deficieíítes
o
10 7d~as
1Días desde
6 ¡a úhíma toma1116
Figura 18-Seguimiento dela infección por
vía oral condosis repetidas
(4b).0= def¡ceííles
24 ratones/grupo

Maledal y métodos
Una vezrecogidaslas muestrasfueronprocesadasde la misma maneraqueen el caso
anterior.
3.XV.b- SEGUIMIENTO DE LA INFECCION POR VíA ORAL CON DOSIS
REPETIDAS
La mecánica seguida para esta experiencia fue similar a la empleadapara el
seguimientocon dosisúnica.Seutilizaron dosgruposde ratones,uno de ratonesdeficientes
y otro de no deficientes,de 24 animalescadauno. Cada grupo recibió un inóculo de l0~
UFC/ml durante7 días.
Los tiempos de sacrificios fueron 1, 3, 6, 11, 16 díascontadosdesdela última toma
de inóculo (figura 18). Se sacrificaron5 animalesde cadagrupocadavez, tres de los cuales
semuestrearonparamicrobiologíay los dos restantesparahistopatologia.
Las muestrasrecogidasy su procesamientofue idéntico al realizadocon dosisúnica.
3.XVI- ESTUDIO HISTOPATOLOGICO
3.XVI.a- PREPARACION DE LAS MUESTRAS
Paralelamenteal estudio microbiológico serealizó un estudio histopatológico de las
muestras. Para lo cual los órganos destinados a este efecto eran fijados en formalina
tamponada.
A continuación,los órganosfuerontalladosparaorientardefinitivamentesu superficie
de corte.Paralo cual, seobtuvieronbloquesde tejidos de 0,5 cm de espesormediantehojas
de bisturí. Tras el tallado de las muestrasse colocaronen cassettesespecialesen las que
pasaron el proceso de inclusión.
Las cassettesconteniendolas muestrasseintrodujeronen un procesadorde tejidospara
su inclusión en parafina.La pautaseguidafue:
81.

Material y métodos
Alcohol etílico 50% 3 h
Alcohol etílico 70% 2 h
Alcohol etílico 80% 1 h
Alcohol etílico 96% 2 h
Alcohol etílico 100% 1 1 h
Alcohol etílico 100% II 1 h
Metil benzoato 30 mm
Benceno1 45 mm
BencenoII 30 mm
Parafina1 2 h
Parafina11 4 h
ParafinaIII 1 h
El pasofinal de la inclusión consistió en la formación de bloquesagrupandolas
muestí~asy orientándolaspara obtenerlos mejorescortes. Este procesose realizó en una
unidad formadorade bloquesmedianteel empleo de moldes metálicosconteniendolas
muestrasya orientadasen parafinalíquida a 600C (parafina III) y que se depositaronsobre
una placa fría sobrela que la parafina solidifica rápidamente.El molde se separó y los
bloquesde tejidos pasarona ser procesadosen el microtomo.
Antes de efectuarlos corteshistológicoslos portasfueron tratadoscon una solución
de gelatina-alumbreparaevitarel desprendimientode la muestradurantela realizaciónde las
técnicasinmunohistológicas.Paraello:
1- se sumergieronlos portaobjetosen NaOH lN (solución acuosaal 4%)
duranteuna nochea 600C;
2- a continuaciónselavaronen aguacorriente15 minutos;
3- sesumergieronen ClH lN 5 minutos;
4- volvieron a serlavadosen aguacorriente 15 minutos;
5- se introdujeron en la solución de gelatina-alumbrea 600C durante 15
minutos; y,
6- sesecarony dejarona cubierto.
82
III

Material y métodos
Los bloques fueron cortados en un microtomo de rotación y cuchillas de acero
desechables(Biocut 3100). Se obtuvieroncortesde 3 pm de grosorque fueron depositados
en un bañode aguacaliente(370C)que permitierala dilataciónde la parafinahastaalcanzar
la superficieque teníaen el bloque.Estoscortesse recogieronen portastratadoscon gelatina-
alumbre,y sedejaronsecaruna nochea 370C.
3.XVI.b- ESTUDIO HISTOLOGICO
La valoración de las lesiones en las muestrasse realizó con el método de la
hematoxilina-cosinasegúnel siguienteprotocolo:
1- desparafinare hidratar;
2- tinción con la soluciónde hematoxilina2-10 minutos;
3- lavadoen aguacorriente20 minutos;
4- tinción con la soluciónde eosina1 minuto;
5- alcohol de 96’ 1 minuto;
6- deshidratarcon un cambio rápidoen alcoholde 960, y dos de 5 minutosen
alcohol absoluto;
7- aclararen xileno, doscambiosde 5 minutos; y,
8- montaren Eukitt.
Con esta técnicase consiguenobservar las membranascon un color azul, y los
citoplasmasen tonos de rosa
3.XVII- DETERMINACION DE LA CONCENTRACIONDE SELENIO
Las determinacionesde las concentracionesde selenio,tantoen los ratonesdeficientes
comoen los controles,serealizaronen muestrasde hígadotomadasal azara distintostiempos
de sacrificios. La concentraciónde selenio fue cuantificada por espectrofotometríade
absorciónatómicaporgeneraciónde hidruros,trasunadigestióncompletade las muestrasen
reactor a presión en horno microondas.Estos análisis fueron realizaron por ITSEMAP
SEGURIDAD AMBIENTAL S.A. en sus laboratorios.
83

Malerial y rn6iodos
3.XVIII- VALORACION DE LA DEFICIENCIA EN SELENIO EN FUNCION DEL
GRADO DE HEMOLISIS
La deficienciaen selenioprovoca una mayor fragilidad en las membranasde los
eritrocitos,lo que se traduceen una mayor facilidad parasu ruptura(Koller y Exon, 1986;
Boyney Arthur, 1990;Bedwal et al. 1993).Segúnesto, la sangrede animalesdeficientesse
hemolizaráantesque la de animalesno deficientes.Conesto ideamosun sistemaparavalorar
la existenciade deficienciasegúnel gradode hemólisisobtenidodurantelos 4 primerosdías
desdesu obtención,tal y como puedeapreciarseen la tabla 6. Paralo cual seextrajo sangre
de la venacavainferior de los ratones,con ayudade una jeringade insulina heparinizada,y
se introdujo en tubos que se mantuvierona 40C durante4 días,observándoloscadadía.
3.XIX- ESTUDIO ESTADíSTICO
Se realizó un estudio de significaciónestadísticade los resultadosobtenidoscon
animalesdeficientesy no deficientes,para lo cuál se realizó un análisis multivariantede
medidasrepetidascon el programaestadísticoSAS.
84

Días 1 2 3 4
Deficientes + ++/±±± +++/±+++
No deficientes + ++
ausencia toia~ de heinólisis
+ hemólisis débil
++ ligera henótisis
I-+± hen,óiisis evidente
++++ hemólisis total
Tabla6- Valoración de la deficienciaen selenioen función del gradode hemólisis,

4. RESULTADOS Y DISCUSION

Resultados y discusión
4.1- VALORACION DE LAS CONCENTRACIONES DE SELENIO EN HíGADO
Para valorar la posible deficiencia en selenio se realizó la determinaciónde su
concentraciónen el hígado de los ratones muestreadosmedianteespectofotometríade
absorciónatómica.Se utilizó la determinaciónen hígadoporqueconsideramosquelos valores
hepáticosde selenioson másestablesque los séricos,no estandosometidosa fluctuaciones
dependientesdel estadode hidratacióno ingesta.Los resultadosde estasdeterminaciones
puedenverseen la tabla7. En ella sepuedeapreciarcómo estosvalorestuvieronun máximo
de 3,93 mg/Kg, un mínimo de 0,94 mg/Kg con una mediade 2,151 mg/Kg paralos grupos
control; y 0,606,menosde 0,015 y 0,192 mg/Kg respectivamenteparalos alimentadoscon
la dieta deficiente.
La mayoríade los autoresconsultadosutilizan comovalor de referenciade seleniosu
concentraciónen suero,lo que permitela repeticióndel muestreo.Sin embargo,Blood ci al.
(1986) señalaunosvaloresde selenioen hígadoparael ganadovacunode 0,9 a 1,75 mg/Kg
en el caso de animalesnormales(animalescon cantidadesfisiológicasde selenio),y de 0,07
a 0,6 mg/Kg en el caso de animales deficientes.Estos intervalos concuerdancon los
resultadosobtenidospor otros autores(JelinekeS aL, 1988; ZacharaeS al., 1993)que han
llevadoa cabosusexperimentosen la especieovina.Estosautoresencuentranvaloresde 1,06
mg/Kg en el casode los animalescontrol y de 0,12 mg/Kg en los deficientes,en el casodel
primero; y de 0,9 a 1,2 mg/Kg y, aproximadamente,0,2 mg/Kg respectivamenteen el caso
de los deficientes,en el caso del segundoautor citado.
Porel contrario,los resultadosexpuestospor autoresquehan estudiadolos efectosde
la deficienciaen selenioen cerdos(Van Vleet eSal?, 1981; TeigeeSal., 1984) no concuerdan
con los descritospor los autoresanteriores.Probablementeello sedebaal uso de unaespecie
animal distinta. El primer autor encontróvalores de 0,15 mg/Kg en el hígado de cerdos
mantenidoscon un piensonormal; por su parte,el segundoencontróvaloreshepáticosde 0,05
mg/Kg en el casode animalesdeficientes.
A pesarde las diferenciasexpuestas,se puedeobservarque las concentracionesde
selenio en hígadoen los asiimalesdeficientesen estenutriente,independientementede la
especieutilizada, seencuentraentreel 10 y el 15% de la concentraciónhalladaen los hígados
87

¡ Concentracioneshepáticasde selenio(mgfKg)
Deficientes No deficientes
Máximo 0,606 3,93
Mínimo <0,015 0,94
Media 0,192 2,151
Tabla 7- Concentracioneshepáticasdetectadaspor espectofotometríade absorción atómica (ITSEMAPAMBIENTAL SA.)


.1.
Resultados y discusion
de los animalesutilizados como control, no superandoestos niveles en ningún caso. En
nuestrosexperimentosse obtienenproporcionessimilares,puestoque las concentracionesde
selenioen hígadoen el casode los animalesalimentadoscon la dietadeficientees,en todos
los casos,inferior al 10% de las concentracionesobtenidasen los controles.Porestemotivo
creemosquesepuedenconsiderarcomo deficientesen selenioa los animalesmantenidoscon
la dieta deficiente en selenioy vitamina E siguiendola pautaestablecida.
Estosvaloresseobtuvieronde maneraconstantetras 6 semanasde alimentacióncon
dieta deficiente en selenioy vitamina E, momentoen el que se dabainicio a los distintos
experimentos.A modo de valoracióncualitativa, y de fácil realización,establecimosuna
relaciónentrela concentraciónde selenioenel sueroy la rapidezcon quesehemolizabanlos
hematíes,tal y comosedescribeenel apartado3.XVIII.
Con estaspremisasseestablecióla duracióndel aportemínimo de pienso deficiente
en selenio-vitaminaE paraobtenerde forma seguraratonesdeficientesen seis semanas,lo
que seconfirmó cualitativamente(hemólisis) y cuantitativamente(determinaciónmediante
espectofotometríade absorción atómica) en cada experimento.El aporte de esta dieta
deficientecontinuódurantecadaexperimentosin queseprodujeranvariacionessignificativas
respectoa los valoresobtenidostras las 6 primerassemanas,por lo que podemosconsiderar
quela tasade seleniose mantuvoprácticamenteuniformea lo largo de todo el estudio.
Es importanteseñalarque la valoración histológica no reveló ninguna alteración
muscular,ni en músculoesqueléticoni en miocardio,como tampocoen ningúnotro tejido de
los animalesdeficientes.No seapreciaron,en consecuencia,diferenciasentrelos deficientes
y no deficientesa estenivel.
En definitiva, los animales sometidos a dieta deficiente en selenio-vitaminaE
presentaronuna deficienciade seleniode tipo subelínico,únicamentedetectablemediante
valoracióninstrumentalespecífica,y que, por tanto, puedecalificarsecomo microdeficiencía.
90

Resultadosy discusión
4.11- ESTUDIOS DE MORTALIDAD
4.ILa- DL50 VtA ENDOVENOSA
Los resultados obtenidos se muestranen la tabla 8. Como puede apreciarsela
mortalidadpor vía endovenosaentrelos animalesdeficientesesmayorque en el caso de los
controles.En estosúltimos esnecesarioalcanzardosisde ío~~ parapoderllegar a observar
algún muerto (40%); sin embargo,a partir de esa dosis se alcanzacon cierta rapidezuna
mortalidaddel 100% (106.5). Porsu parte,en los deficientesseobservamortalidaddesdedosis
relativamentebajas(10~~).
Con los datosrecogidos,y aplicandoel métodode K~irber (Schmidty Emmons.1989).
podemoscalcular una DL50 para los animales deficientesde ~ y de 105.65 para los
controles.EstaDL50, enel casode los controles,no difiere de la encontradaporotros autores
(Vázquez-Bolandy Berche,resultadosno publicados)paraestamisma cepade Listeria y de
ratones,por lo que la consideramosuna buenareferencia.
Los resultadosobtenidosestablecenuna diferenciaentrela DL50 paradeficientesy
no deficientesde cercade un logaritmo,lo queconsideramossuficienteen cuantoaestablecer
un criterio de mayoro menorsusceptibilidadpor estavía entreambosgrupos.
Las DL50 obtenidasparacadagrupo se usaronparalos estudiosde seguimientode la
infecciónpor vía endovenosa.
4.II.b- MORTALIDAD POR VtA ORAL CON DOSIS UNICA
Los resultadosde esteexperimentosepuedenobservaren la tabla9a. Enella se puede
apreciar cómo la mayor mortalidad se produce con la dosis de 1010 Lm/ml, tanto en
deficientescomo en no deficientes(56,25y 18,75%respectivamente),mientrasque cuando
se utiliza una dosis de io~ Lm/ml las mortalidadesdesciendenentre un 59,25% en los
deficientes(22,92%) y un 66,7% en los no deficientes(6,25%). La influenciade la dosis
sobrela mortalidades menor en el caso de los animalesdeficientesque en el de los no
deficientes,debidoa queen los primeroslos mecanismosresponsablesde la respuestainmune
91

Dosis/ratón N0 ratones N0 muertos % muertos
102.5
Deficientes 5 0 0
Controles 5 0 0
10~Deficientes 5 0 0
Controles 5 0 0
lo”Deficientes 5 1 20
Controles 5 0 0
ío’Deficientes 5 0 0
Controles 5 0 0
Deficientes 5 3 60
Controles 5 0 0
Deficientes 5 2 40
Controles 5 0 0
Deficientes 5 4 80
Controles 5 2 40
106Deficientes 5 5 100
Controles 5 4 80
106.5Deficientes 5 5 100
Controles 5 5 100
10’Deficientes 5 5 100
Controles 5 5 100
Deficientes:7-{ [(500/I00)~0,5]x0,51= 4.55 ~ DL5O= l0<’~
Controles: 74 [(320¡100)-05]xO,5}=5,65 ~ OLSO= io~~
Tabla8- Mortalidadesobtenidaspor vía endovenosacon Lm 4b en ratonesdeficientesy no deficientes.

Dosis N0 ratones N0 muertos % muertos
lotoDeficientes 48 27 56.25
Controles 48 9 18,75
í o9Deficientes 48 11 22,92
Controles 48 3 6,25
Tabla9a- Mortalidadpor vía oral con dosisúnicade Lm 4b.
Dosis/T.b. N0 ratones N0 muertos % muertos
l0~ 4 díasDeficientes 48 16 33,33
Controles 48 7 14,58
l0~ 7 díasDeficientes 48 19 39,58
Controles 48 10 20,83
Deficientesío~
10 días Controles
48 12 25
48 8 16,67
Tabla9b- Mortalidadpor vía oral con dosisrepetidasde Lm 4b.

Resultadosy d,scus,on
seencuentranalterados,tal y cómoseexpusoen el apartado1.IiX.d.
En cuantoa las diferenciasentrelastasasdemortafldadobtenidasen deficientesy no
deficientes,tambiénsepuedenapreciardiferenciasimportantesindependientementedeladosis
utilizada. Así, la inoculaciónde dosisde io~ bm/ml suponenunatasade mortalidadinferior
en un 72,7% en los no deficientes(6,25%) con respectoa la obtenidaen los deficientes
(22,92%);y con ío’~ LmIml esareducciónesdel 66,7% (18,75 y 56,25%respectivamente).
Todo ello parece representaruna posible influencia de la deficiencia en selenio en la
susceptibilidada la listeriosis, tal y comosediscutiráen el apanado4.X.
4.H.c- MORTALIDAD POR VIA ORAL CON DOSIS REPETIDAS
Esteexperimentofue enfocadoaestudiarel efectode dosisrepetidasen la mortalidad
por vía oral. Como puedeapreciarseen la tabla 9b, existe una relación directa entre la
mortalidady el númerode repeticionesde una misma dosis.Así, la mayormortalidad,tanto
en deficientescomo en no deficientes,seobtiene dosisde iO~ Lm/ml durante7 días, 39,58
y 20,83%respectivamente.Cuandola misma dosisse administradurante4 días seproduce
una reducciónde la mortalidadhastavaloresde 33,33%en los deficientesy 14,58% en los
no deficientes.El empleode una dosismenor(lOa LmIml) duranteun períodode tiempode
10 días, produce una menor mortalidad en ambos grupos,pero si en los deficientes la
mortalidadpermanecepordebajode la obtenidacon í0~ Lm/ml durante4 y 7 días (25, 33,33
y 39,58%respectivamente),no ocurreigual en el casode los no deficientes,en los que esta
mortalidad es superior a la encontradacon 1(9 Lm/ml durante4 días (16,67 y 14,58%
respectivamente),aunqueestamínimadiferencia(aproximadamente2 logaritmos)no parece
serrelevante.
A nuestrojuicio, estosresultadossugierenla existenciade un efectoacumulativo,en
el sentidode que una dosis de 108 Lm/ml durante10 días producemayor mortalidaden
ambosgruposque una única de ío~ Lm/ml, siendoidénticala dosis total inoculada.Esto es
especialmentemarcadoen el caso de los no deficientes,con mortalidadesde 6,25% (1x109
Lm/ml) y 16,67 (10x108 LmIml), lo que representaun incrementode la mortalidadde un
166,72%.
94

Resultadosy discusión
En el caso de las dosisrepetidasen el tiempo, las diferenciasentredeficientesy no
deficientesdesciendena medidaque aumentael númerode repeticiones,así con 1(9 Lm/ml
durante4 días se obtiene un incremento de la mortalidad de 128,6% en el caso de los
deficientes,si aumentamosa 7 díasel periodode inoculacióneseincrementoesde tan sólo
un 90%, y si utilizamos 108 Lm/ml durante10 días el incrementono superael 50%.
Los estudiosexperimentalessobrelisteriosisque utilizan la inoculaciónpor vía oral
sesirven por lo generalde dosismuy elevadas,del ordende itl’0 ó i09 Lm/ml (Golnazarian
a aL, 1989; Notermansy Chakraborty,1992),en tomasúnicascomopautamás extendida.
Este sistemapermite reproducir, de forma más o menos constante,la infección e incluso
producirlaenfermedadclínica.Nuestrosresultadosconfirmanlaproducciónde laenfermedad
medianteel empleode dosisaltaspor vía oral, pero además,sugierenla posibleexistencia
de un efectoacumulativocon el que seobtienenmortalidadessuperioresutilizando la misma
dosistotal pero fraccionadaen varias tomas.
Estefenómenoacumulativoya ha sido descritopor nuestrogrupo en referenciaa la
respuestaserológicafrente a inoculacionescon L. nwnocytogenespor vía oral a dosis de
incluso í0~ Lm/ml (Lópeza aL, 1992).En esteestudiosedemostróel desarrollode unatasa
detectablede aglutininasmedianteinoculacionesrepetidasde la citadadosis. Estarespuesta
inmunerevelaun contactoentreL. nwnocytogenesy el sistemainmune del hospedador,y, por
tanto, de una ciertadiseminaciónextraintestinal,controladapor el hospedador,en razóndel
bajo númerode L. rnonocytogenesque lo logran.
La posibleexistenciade un efecto acumulativo podría tenerimportancia desdeun
punto de vistaepidemiológicoy experimental.En el primercaso seríarelevanteen cuantoa
que en condicionesnaturalesla infección se producea partir de alimentoscontaminadosen
los que las tasasde L. monocytogenesraramentealcanzanlos valoresempleadosen estudios
experimentales(l0~ ensilado). Sin embargo,un aporte reiteradopodría traducirseen el
desarrollode la enfermedadaunque el número de L. monocvtogenesen el alimento sea
relativamentebaja.En cuantoa las repercusionesen el ámbitoexperimental,podríaplantearse
la necesidadde establecerpautasde inoculaciónen basea aportesdiarios reducidos,pero
prolongadosduranteun periodo de tiempo relativamentelargo.
95

Resultadosy discusion
Podríaespecularsecon la posibilidadde que aportessucesivosde listerias pudieran
ayudara dicho microorganismoa soslayarel efecto competitivo de las restantesbacterias
intestinales.Dehecho,L. monocytogenesha sidodescritacomouna pobrecompetidoraa este
respecto(Zachary Savage,citado por Golnazariana al. 1989; y porMarco et al., 1992b),
e incluso seha comprobadocómoL. nwnocytogenesúnicamenteproducelesionesintestinales
tras un bruscocambiodel ambientegastrointestinal(Ráczcitado por Marco et al.. 1992b).
De igual forma el posible efecto negativo del pH gástrico (Schlech, 1990) podría verse
disminuido en basea aportesrepetidosde L. mrnwcyrogenes.
La influenciareal de estosfactoresdebe,en cualquiercaso,sercontrastadamediante
nuevosestudiosencaminadosespecíficamentea valorarestoshechos,ya que nuestrosdatos
no sondeterminantesa esterespecto.
Parafinalizaresteapartado,queremoshacerreferenciaal hecho deque en ningunode
los experimentosde dosisúnicalos animalesmostraronmanifestacionesclínicasantesde una
semanapost-inoculación,lo quepareceapoyarla teoríade que,porvía oral, Listeria necesita
un cierto tiempoduranteel cualsemultiplicaríaen epitelio intestinal hastaalcanzarel número
necesarioparaatravesarla barreraintestinal y llegar a sus órganosdianas;esetiempo podría
verseafectadopor la dosis utilizada; así, la apariciónde signosclínicos es anterioren 24
horascon 1010 que con 1(9 Lm/ml, lo que pareceestaren consonanciacon que la infección
por listeriasporvía oral es dosis dependiente(Schlech,1990).
4.111-SEGUIMIENTO DE LA INI~’ECCION POR VIA ENDOVENOSA
4.III.a- DOSIS iO~~
Estadosisfue utilizadaporser la másaproximadaa la DL50 de los ratonesdeficientes
que pudimos obtener, ya que la pauta seguidatan sólo nos permite ajustarnosa medio
logaritmo. En la tabla 10 puedenapreciarselos resultadosobtenidosde los recuentosde L.
rnonocytogenesen hígadoy bazo, tanto en los animalesdeficientescomo en los controles.
Podemosobservarcómo los recuentosen bazo son muy similaresen ambosgrupos
a lo largo de casi todael experimento.En primer lugar seproduceun aumentode los
96

TPI
Deficientes No deficientes
Hígado Bazo Hígado Bazo
30 ni l,12x10’ 2,SSxJO’ tino’ 4,2x10’
12 h 2x102 2,3lxlO’ IxIO’ 4,25x10~
1 d 2,73x10’ 3,45xI0~ 3,07x10’ 323x10’
2 d 7,82x10’ 3,lSxIO’ 1,02x10’
3 d í,57x10’ 7,54x10~ 6x10’ 9,12x10’
4 d 5,SxIO’ 1,97x10~ 21102 8,3x10’
5 d 1,32x10’ l,45x10’ O 5,65x10’
6 d 4,4x10’ S,55x10 0 2,37x10’
7 d o I4x104 O I,65x10’
Sd 0 0 0 0
9d o O 0 0
lOd 0 0 0 0
Tabla 10- Resultadode los recuentosen hígadoy bazotras inoculaciónintravenosacon i0~ Lni

o
oNxm
oO
O~
~e
~C
iCM
—e14
«‘4
\¡/“
¡¡
te¿
ji~
1.KY
-{zbe
¡
.2u
•¡¡m
’I)
¡
OQ
bO
i~-
CCCD
—C
iCM
—
o,
fi~=
oIi
CCoz
7)
LO
A,/4.
7,
>7
,
‘o,
//
1‘o4)o4)ci
Ci
CMo
fioeoe.0EO)
.5eoO)
fioeO)
aO)
ao=O)
fioeO)
O)
O)
fioO)
O)
oII
a
fioO)cio)te
ozCe
‘ocioo)O-
CCO

Resultadosy discusión
recuentoshastael día2 pi, y a partir de esemomentoseproduceun descensoen los mismos
hastadesaparecerel día 8 en amboscasos(gráfica lb).
En líneasgenerales,nuestrosresultadoscoincidencon los expuestospor Goosenset
al. (1988),segúnlos cualeslos recuentosobtenidosa partir del bazo de ratonessusceptibles
y resistentesa la listeriosis eranprácticamenteiguales.Este y otros autorescitadospor él,
opinanque esto se debeaque en el bazono seexpresaunarápidaresistenciaa la listeriosis,
apareciendoéstaa partir del día4; opiniónque parececonfirmarsepor los trabajosde Conlan
y North (1994),en los que se afirmaque los neutrófilosno lisan las célulasinfectadasen el
bazocomo parecenhaceren el hígado,quedandoen esteórganolas listerias a resguardode
las células fagocíticas.Esto coincide con por lo expuestopor otros autores(Archinal y
Wilder, 1988a;Hof y Hefner, 1988;Cossarty Mengaud,1989),quienesafirmanque tras una
inoculaciónintraperitonealo intravenosaseproduceun aumentode los recuentosen bazo
hastael día 3 pi, produciéndosea continuaciónun descensode esosrecuentosque llevan
hastael control de la infección por parte del hospedador.La aparición más lenta de esa
resistenciaparece,asimismo,confirmarseporel hecho de que en nuestrosdos grupos los
recuentosde listeriasen esteárganono comienzaa descenderhastadespuésdel día 2.
En el hígado,el númerode bacteriasaisladasa los 30 minutospi fue mayor en los
controlesqueen los deficientes,a continuaciónseproduceun descensoenel númerode los
recuentosobtenidosen ambos grupos a las 12 horas pi, para a partir de ese momento
aumentarel día 1 pi; esteaumentoes mayoren el caso de los deficientesque en el de los no
deficientes (graf. la). Desde el día 1 pi los recuentoscomienzana descenderhasta
desapareceren los no deficientesel día 5 pi; por su parte,en los deficienteslos recuentos
desciendenlevementehasta producirse un nuevo incrementoel día 6 pi, tras el que
desaparecenel día 7 pi. Estosresultadosson semejantesa los expuestospor otros autores
(Cossarty Mengaud,1989), segúnlos que tras una inoculaciónintravenosaseproduceun
aumentodel númerode listeriasen hígadohastael día 2 6 3 pi, a partir de los queseproduce
una inactivaciónde las bacteriasy la recuperacióndel hospedador.
En los deficientesse observanmayoresrecuentosen hígado, así como un mayor
tiempode permanenciade los microorganismosen éste,probablementedebido a un fallo en
la respuestainmune tal y como discutiremosen el apartadoIX. Por otra parte, esemayor
99

Resultadosy discusion
tiempo de permanenciaaumentalas posibilidadesde generalizacióny de mortalidad,lo que
podría explicar las diferencias de mortalidad observadascon esta dosis, 60 y 0% en
deficientesy no deficientesrespectivamente(tabla 8).
Otropuntoa destacares la disminuciónde listeriasobservadoa las 12 horaspi en los
dosgrupos.Consideramosqueestedescensosedebea la actuaciónde los neutrófilosdurante
las primerashoras de la infección hastala llegadade los macrófagosy linfocitos T, tal y
como apuntanalgunosautores(Conlany North, 1991, 1993, 1994). Estedescensoes más
enérgicoen el casode los animalescontrolesqueen los deficientes,lo cual, probablemente,
seadebidoa una menoreficacia de estascélulasen los últimos a la hora de poderinstaurar
una correctarespuestaala infección.Menoreficaciaquedebemosasumirseadebidaal único
factor quediferenciaambosgrupos,estoes la deficienciaen selenioy vitamina E, tal como
sereflejaen 4.X.
En cuantoal aumentoobservadoen los recuentosa partir de hígadode los animales
deficienteslos días 5 y 6 (gráfica la), y a la luz de los resultadosobtenidoscon la dosis de
1 0~’, y que se exponenen el siguienteapartado,consideramosque podrían debersea una
segundageneralizacióna partir de los focosproducidosen la infección inicial.
En líneas generales,nuestros resultadosen el hígado también parecenestar en
consonanciacon los expuestospor Goosenset al. (1988),quien afirma que los recuentosen
hígado fueron mayoresen el caso de los ratonessensiblesa la listeriosis que en los
resistentes.Además, este autor también señala que en los animales resistentesla
sensibilizaciónde los linfocitos T se produceen un tiempo menorque en los sensibles.La
desapariciónmás rápida de las listerias en los no deficientespodría debersea esta
sensibilizaciónmás tempranade dichoslinfocitos.
Observandolas gráficasla y b, podemosver cómo las listerias tardanmás tiempo en
desaparecerdel bazoque del hígado,observaciónrealizadaasimismopor otros autores(Hof
y Hefner, 1988;Cossarty Mengaud,1989),debidoa unamayorrapidezen el establecimiento
de los mecanismosde defensaen el hígadoque en el bazo(Goosenset al., 1988; Cossarty
Mengaud, 1989), a un mayor tropismo de L. monocytogenespor el bazo (Hof y Hefner,
1988), o a que los neutrófilosno actúan de la misma forma en ambosórganos(Conlan y
1 0(.)

Resultadosy discusión
North, 1994).
Los resultadosestadísticosconfirmanla existenciade diferenciassignificativasen los
recuentosobtenidosa partir de hígadoentrelos deficientesy no deficientes(P<0,05).
4.III.b- DOSIS iO~~
Estadosisseescogióen virtud de ser la másaproximadaa la DL50 calculadaparalos
ratonesno deficientesque podíamosobtenermediantelas pautasutilizadas.La tabla II
muestralos resultadosde los recuentosde L. monocytogenesa partir hígadoy bazo de los
gruposcontrol y deficiente,inoculadoscon estadosis.
Tal y como puedeverse en la citada tabla, a los 30 minutos pi los recuentosson
ligeramentesuperioresen el bazo de los controlesqueen el de los deficientes.Sin embargo,
a partir de esemomentoexistendiferenciasentreambosgrupos,al producirseun aumentode
las listeriasaisladashastael día 1 pi, tal y como ocurrieracon la dosisde l0~, siendoéste
mayor en el caso de los deficientes.El aumentode los recuentosen el bazo durantelas
primeras24 horaspareceestaren consonancia,asimismo,con lo expuestopor Archinal y
Wilder (1988a).
Entrelos días2 y 4 pi seproduceun levedescensoen los recuentosde ambosgrupos,
aunquelas diferenciasentreellosson de casiun logaritmo (gráfica2b). El día 5 pi seproduce
un segundoaumento,muchomás acusadoen los deficientesque en los no deficientes,para
a partir de entoncesdescenderhastadesapareceren amboscasosel día 8 pi. Estesegundo
aumento parececonfirmar una reactivaciónde la infección inicial, que seríacontrolada
muchomásrápidamente.Es importanteseñalarquela existenciade esesegundoaumentode
los recuentosno aparecereflejadaen ningunade las citasbibliográficasconsultadas,ya que
la mayoríade los estudiossuelenacabarel día 4 pi. Asimismo, essorprendentela aparición
de estepico cuandolos recuentosseencuentrandescendiendo,sin que hayamosencontrado
otra explicaciónque una posibleliberaciónde listerias a partir de focosdondesehubiesen
acantonadodurantela infección inicial.
Estosresultadosdifieren de los observadospor Goosensaal. (1988), quienesno
101

Deficientes No deficientesHígado Bazo Hígado Bazo
30 m 5,25x10’ 4.75x10’ 1.36x10’ 7,SSxlO’
12 h 1,78,10’ 1,5~10’ 72h10’ 1.07x10’
1 d 3,32x10~ 4,36x10~ 2h10’ l,29x10’
2 d 1,9x10’ 1,24x10’ 2,62,10’ 2,09,10’
3 d 1,36,10’ 3,59,10’ 5,12x10’ 4,95x104
4 d i,io’ R,65x10’ 2,10’ I.34x10’
5 d 3,2x10’ 4.52,10’ 8,77,10’ 2,35,10’
6 <1 s,ssxio’ 9,2,10’ 0 3,10’
7 d 0 1,37,10’ 0 9í10’
o O o o
9d o 0 o o
lOd o O O O
‘fabla 11- Resultadode los recuentosen hígadoy bazotras inoculaciónintravenosacon itt’ Lm
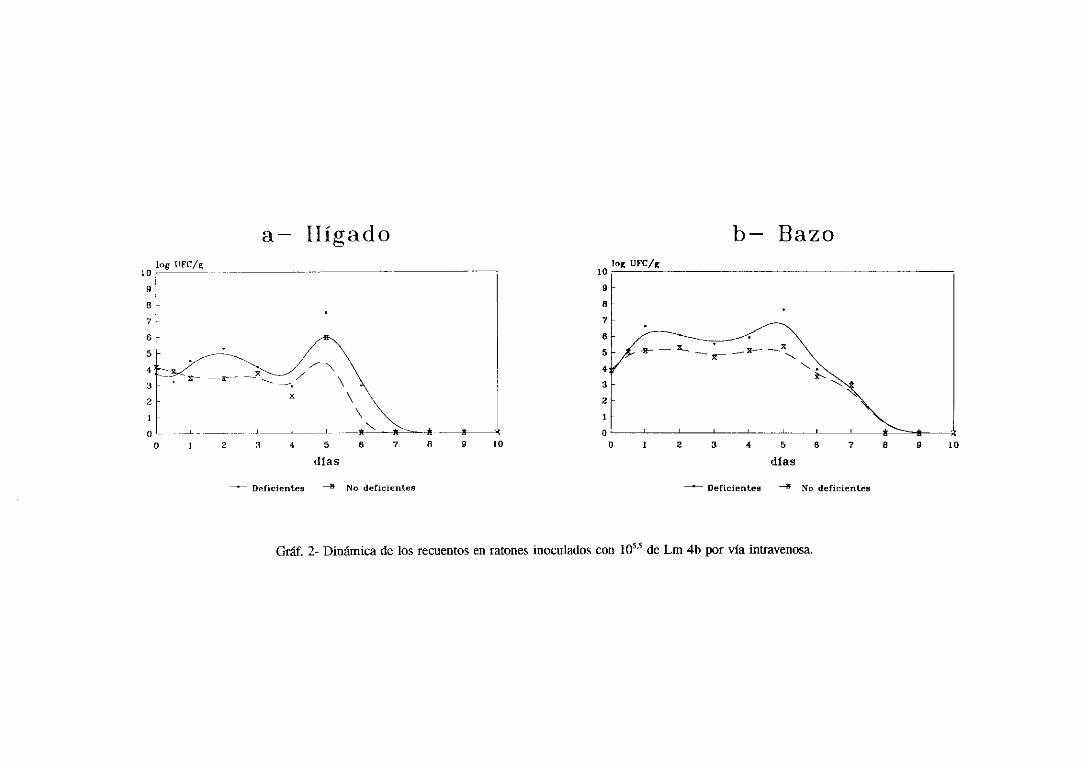
oN.0o-ebL
‘ouCC‘oo
oo
o’o
‘ooz
‘oof
‘eo’
4
o.uC
io’
o’
eo’~
0‘o
o
u,~
5’oO’~.0
Ci
uts.
CC‘oo
OO
i‘o
‘oib
4C
iCQ
—O
o
51~o4
)
‘oSn
ÚEO)
‘e‘oao)>CCo‘e‘oO
)o‘oC1fl4
)‘oO
-‘oO
)CCo‘o4
)‘ooO)
O-
CCoO)
O)
oCci
OO
Ci
‘oO
.CO
ti4
Ci
03—
O

Resultadosy discusión
encontrarondiferenciasen ratonessensiblesy resistentesentrelos recuentosdurantelas 72
primerashorastras inocular hasta9x105 L. monocytogenes/ratónpor esta vía. Quizásesa
diferenciaseadebido,tal y comoreferimosanteriormente,a unamayor resistenciade la cepa
de ratonesutilizada por nosotrosa la listeriosis.
En el hígadoserepitela observacióninicial hechaen el bazo,esto es,el númerode
bacteriasaisladasa los 30 minutos es mayor en los controles que en los deficientes.
Asimismo, seobservaunareduccióndel númerode L. monocytogenesaisladoalas 12 horas;
reducción ésta, aunquemuy similar, ligeramentesuperior en los deficientesque en los
controles.A las 24 horasesenúmeroes ya un logaritmomayor en los segundos(graL 2a).
Por otra parte, mientrasque en los controles los recuentosse mantienenprácticamente
constanteshastael día 3, e incluso desciendenligeramenteel día 4, en el caso de los
deficientestodavíaaumentanun logaritmo antes de iniciar ese descenso.El día 5 puede
observarsecómo seproduceun aumentode los recuentosde listeriasaisladas,muy similar
al observadoen el bazo, y que podría responderde igual forma a una reactivaciónde la
infeccióninicial. Esteaumentoparecesermásimportantequeel observadoenel bazo,puesto
que en ambosgrupos el aumentoen el caso del hígadoes de alrededorde 4 logaritmos,
mientrasque en el bazoes aproximadamente2 logaritmosen los deficientesy de menosde
uno en los controles(graL 2a y b).
Por último, hemosde destacarque los deficientestardanúnicamente24 horasmás
que los controlesen eliminar las listeriasde hígado, lo que podría tal vez serdebido a que
a dosis próximas a la DL50 las diferenciasentre deficientes y no deficientes no sedan
apreciables,y el comportamientode ambosgruposseriamuy similar, lo que implicaríaque
existe un número de listerias lo suficientementeelevado para rebasarla capacidadde
respuestainmune,estéo no alterada.
La reactivación de la infección observada,parece ser dependientede la dosis,
apareciendoa partir de dosisletalesy no siendoobservadaen dosissubletales(tabla 10). Esta
seríala causapor la queno seobservaen los no deficientescon la dosis de ~ que para
estegrupo es subletal, y si en cambio al utilizar la dosis de ~ lo que también parece
confirmar las diferenciasde mortalidadencontradasen amboscasos(tabla 8).
104

Resulíadosy discusión
La existenciade estareactivacióny las diferenciasde recuentosde casi2 logaritmos,
podría relacionarsedirectamentecon las diferenciasen cuantoa mortalidadobservadascon
estadosisen animalesdeficientesy no deficientes,quesondel 80 y el 40% respectivamente.
Ademásello implicada,en nuestraopinión,quelos estudiossobrela listeriosisdeberíantener
una duraciónsuficiente(en nuestrocaso 10 díaspi) parapodercontemplarestefenómeno.
Tal y comoreflejamosen el casoanterior,seobservacómoexisteunamayortardanza
en desaparecerlas listerias del bazo que del hígado, y que lógicamentetendría la misma
explicaciónexpuestaen el apartadoanterior.
Nuevamente,los resultados estadísticosconfirman la existenciade diferencias
significativasentreambosgruposde animales(P<0,05)con la dosisutilizadade ío~’~ porvía
iv.
4.1V- SEGUIMIENTO DE LA INFECCION POR VTA ORAL CON DOSIS UNICA
4.IV.a- CON LISTERIA MONOCYTOGENES4b
Los resultadosobtenidosdel aislamientoen órganosde listerias inoculadasporvíaoral
en ratonesdeficientesy no deficientesson los expuestosen las tablas12 y 13. En ellas se
puedeobservarun retrasode 24 horascon respectoa la vía intravenosaparapoderaislarlas
de hígadoy bazo; ese tiempo podría ser el necesariopara traspasarla barrera intestinal,
alcanzary, eventualmente,multiplicarseen los órganoslinfoides regionales,a partir de donde
llegan a hígadoy bazo, tal y como seexplicó en el apartado4.II.c.
En la mucosadeltractodigestivoúnicamenteseencontraronlesionesen las placasde
Peyer. En éstasse observó,tanto lesión evidentecomo bacteriashastael día 6 pi en los
deficientes.Las lesiones más graves mostraron, de forma predominante,una reacción
inflamatoriapurulenta,que afectabasiemprea las áreascúpulade las placasde Peyer,entre
el epitelio y los folículos linfoides (imágenes7, 8,9. 10 y 11). En algunoscasosesteepitelio
de las Placasde Peyerse presentabaulcerado.Las lesionesencontradasen los animalesno
deficientesfueron muy similaresa las descritasen los deficientes,aunquela inflamación fue
más ligera, y secircunscribióúnicamentea los días2 y 3 pi.
105

ti).4z
oCCC
rree
---4CC
44CC
C
44
rre
uu,e
j
00<u~
o
e
ee-
000<o,00
000<o’O
N
OOOZ
VVVi
>>><U~
‘o
S
SS
~-
000<
....4
..~>~
>>>><
>>>4
‘oO
CCCoe
a
-u
,
00
—.~
——
00<ej
sssC
<50<uyVi
ee>4
444
0
00
00
<5000
o~
,
2—
~~~
00<0C
..4
<5>4‘o.< 0N
00<U
No
,-e
j
00<U
Nre—
00
00
0000’
OOO<<<4
444
ttt~ay<5.~o
uuu,
Oc
—~oo
uy
<5~eju
,0<o,
00
00
t’o
Z
‘‘oSC
<52o’—
<504‘oe-
<504‘oo,
<50<ej
‘o<‘o
e~0<
Vi-
33<O
OO
OO
O
Eo
<<
¿2
00<
Vi
ay
—o
,
CC
oo,
—
000
00
0
..0~
<52~4
<5~CC0’
<5t~5
ffl2
0<—>4re
00
‘‘eCiSn
e~
.4,;
oc
e~
<¿>4CC
<5fleieej
——eej
~o
a
eeen
~-cti—
.c~
~e
~
‘et
‘e‘fl
-e~O
‘efto
‘e00
‘eQN
‘eO—
——‘-O—
E.5CCo
•‘e
•C
iO
)e‘C
iC
iC
iC
ieoO
)
¡C
i
O-
oo-CC
o‘eCi
Ci
O)
oCiCC
O)
O)
O)
teO)
‘eCC
O)
Ci
oCi
O-
O)
‘eoCi
CitÉO-
‘oCCoCiCC
itoCiO)
CC
oCiO)
Ci
O)O)
O-
CC
oo)‘eC
CO
)
tiCi
.0Ci
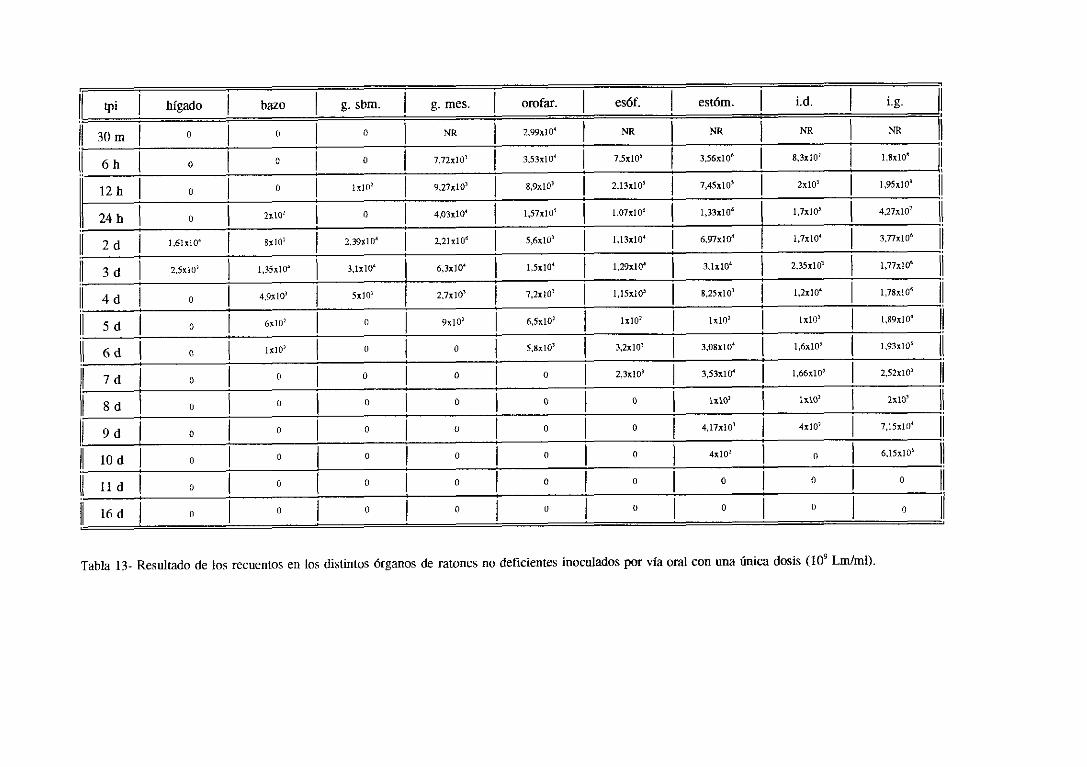
OO
Oa
OO
aa
Oe
.a
aa
tÉ—
0<>4
0<>4
0<
0<
0<
>4
0<
><O
o-,
e-
e-
oc
ze
>e
-u
,O
~V
ie
>—
——
4¿
—-~
——
ej
e-
‘o
a<
5<
5b
a—o
aa
~—
.4—
—>4
0<>4
SC
0<0<
0<0<
-<
4-
--
.4re
~e>-
—SC>
SC
—t
oc—
—
<<
5<
5<
5<
5<
¿<
5<
5a
o—
——
o—
—O
—‘O
~0<
>4
0<0<
~—
.4o<
20
—Z
~a
ej
—O
UN
—C
CV
N—
.V
i>4
u,
0<e
-.4
4)
ree
-—
jre
.4re
4
0
<5<5
ea
ao
aa
>4>4
•<><
o<o,
,;,;
,;a
Oca
O)—
ej.e-
ej4
—re
eja
~~
é<5~
<5<
5<5
oa
——
OO
aa
aa
oe>
reVN
O-SC
VN
<5<5
<5<5
<5<5
E~
-z‘~
—2
aa
oa
ao
Ze
je-re
—<
ne-
>4o’e-
e>SC
<4ej
0’~
ejtÉ
a
a,;
oa
oa
~<5
<5
aa
ao
aa
oa
—re
-V
ire
tÉe
>
oa
o5
O—
—~
oa
aa
a0<
0<0<
.0e>
00re
QC
C—
-0~
a
Oa
<5‘e
ao
a—
aO
OO
OO
OCC
Ci
3<0<
tÉCC
O
.0
E.0
.0-~
‘e‘e
‘e‘e
-~‘e
‘e‘e
‘e‘e
o~
ti—
—en
—~
tiCt
t50—
~QN
O—
‘e—So
Eo
CC
CCO‘eE‘Ci
Ci
CiCi
Ci
OO)
oCi
O-
oo-CC
o‘eCi
CiO)OCi
CC‘uCiO)
O)
SnO)
‘eoCiCe
O)
CioL.oO)
‘eCCoCi
Ci
tÉO-
‘oitoCi‘o-‘E.2Ci
O)
LooCici
CiO)
0)
O-
o0)
‘eo‘e5CiO)
enCi
.0Ci
Fi

SCibCi
03
5—
o2
4>o
’
04
¡SC
4,e-
-CO
~1:103
-.•
YuCC
‘s~—
[..~ll±>il~ffl~wil~
-~—
Oo’
u,
e-O
ib—
ev
ibib
1;42o,
u,
04
e-
Á-
Cci
05Ci
‘e
oo
ou0
CCrn
ozofs
u’O
6
~0-e
ibIt
‘e
a
ce
-eo’CC
ceC
Cos
of
oO’u
-—-4
--4bEceoci
<o-E
Ci
—~
oó
03—
—5’4>
.0—
O
EO
o’
—.~
4)
o‘e
o’
zCCC
=Ci
-0
=‘e
itO’
CCo<o
‘eCi
.2o1
Ci
03
ito‘eCi
O
O)
o‘oO)
Ci
oO-
4)
‘e‘-1o4)
Ci
CC
o4>
0o)
O-
‘oOo>
Cici
O)CC
CCo
Ci
zO)C
i
of‘e
CC
o4>
4)
CoCCC
ciO
)4.
.—‘ID
O’
en
03
-r
e-
ON-cO-ecebE0
-4
ce
OO
’S
CID~
Coib
‘C
i03
—O
o
uCCo’
OO
O’
CC
O-
CO
ib~
Ci
Da—
00
u1’-o01o
Oo
,o>
e-
<oib
—eD
03—
o

o‘oUD
a)
tÉ-
oza
—A
—o>
eo,
Ao,
t45
•o,
5
oU)a)¡04
a)oa)obLs•0ti)o
—5
o>o,
ci
~4
—5
oo’
Co
A
o>4
—A
oo,
o,
¡
-N-4.0-4cf,o‘0CioO)
Ci‘OO
)
CiO)
ooCC4)bE
-eO)
“eotÉOCi
CC
oCiE)
Ci
O)
O)
O-
CCoO)
‘eCiO)







Resultadosy discusión
Es de destacarque aunqueel númerode listeriasque consiguenatravesarla barrera
intestinal es relativamenteescaso(Marco et aL, 1992b), pero suficiente para infectar el
epitelio intestinal y las placasde Peyer,tal y como afirman distintosautores(Gaillard et al.,
1987a;Domingo et aL, 1992; Marco et al., 1992b;Czuprynski, 1994),donde realizaríauna
primeramultiplicación que le permitiría adquirir un númerosuficienteparaque la acciónde
los macrófagosno seadefinitiva (Gaillard etaL, 1987a,1987b),pasara los ganglioslinfáticos
regionalesy, desdeahí, alcanzarel hígadoy el bazo (Golnazarianet al., 1989),dondebien
seríacontroladala infección,o bien, tras un nuevoperíodode multiplicación,sediseminaría
por el hospedadordando lugar a nuevosfocos en órganos distintos a los mencionados
(Schlech, í990; Marco a aL, 1992b).Esta primeramultiplicación en célulasepitelialesha
sido observadapor otros autoresutilizando cultivos celularesde la líneaCaco-2(Gaillard a
aL, 1987b; Bercheu aL, 1988).
Una vez atravesadala mucosadigestivalas listeriaspodríanseguirvarios caminos:o
bien generalizarsepor vía sanguínea,bien porvía linfática, o bien por vía linfohematógena.
Nuevamente,la rápidaapariciónde listerias en los gangliosmesentéricos,con respectoal
hígadoy bazo (graf. 3d), parececonfirmar lo expuestopor otros autores(Golnazarianni al.,
1989; MarcoaaL, 1991, 1992a;Low y Donachie,1991)en el sentidodequela víautilizada
es la segunda.Llama poderosamentela atenciónla desapariciónde las listerias másrápida
de los gangliosquede sus tramosrespectivosde tubo digestivo (gral’. 3c y d, 4a, b. c, d y e).
En el caso de los ratonesdeficientes,la infección, entendidacomo la presenciay
multiplicación de listerias en el hospedador,seprolongaen el tiempo, apareciendoantesen
hígado,y tardandomásen desaparecer,a lo que se añadela obtenciónde recuentosmayores
en hígadoy bazo. Por otra parte,en estesegundocaso,se puedeobservarun leve aumento
de los recuentoshaciael día 7 en el hígadoy 8 en el bazo,que podría corresponderse,al
igual que sucedieraen la vía iv, con una posiblereactivación(gral’. 3).
Además,debemosreseñarcómo la cinéticade la infección (grál 3 y 4) essimilar en
ambosgmpos,si bien los valoresde los deficientesseencuentran,por lo general,porencima
de los de los controles,lo que esespecialmentevisible en las gráficasde hígadoy bazo (3a
y b). Por otra parte,la recuperaciónde L. rnonocytogenesa partir de órganosinternos y tubo
digestivocesaantesen el casode los no deficientes,excepciónhechadel estómago.
Y16

Resultadosy discusión
Es de destacarla rápidaapariciónde listeriasen los ganglioslinfáticos muestreados,
incluyendo el ganglio submandibular(graf 3), ya que el que aparezcanenel mesentéricoa
las 6 horas,y posiblementeantesdeesetiempo,no deberíaextrañarnosal utilizar el intestino
como puertade entrada.Esteresultadodel gangliosubmandibular,podríaindicar laexistencia
de otrapuertade entradaen el tractosuperiordel aparatodigestivo,posiblementelas tonsilas,
algo ya sugeridopor nuestrogrupode trabajoen otrosestudios(Briones,1990; Lópezetal.,
1993), ya que consideramosdescartableque esto se deba a la posibilidad de una
generalizaciónde la infecciónporapareceren estelugarantesqueen hígadoy bazo,además
de que estosgangliosrecogenel drenajede toda la zonaorofaríngea.Lamentablementeno
sepudoevidenciarhistologicamentelas tonsilasde los ratones,por lo que nosfue imposible
confirmarla utilizaciónde estasformacioneslinfoides comopuertade entradaal hospedador.
Sin embargo,se realizó un pequeñoexperimentoadicional con la intención de
compararla dinámica de la infección de L. monocytogenesen ganglio submandibularen
animalesinoculadospor vía iv con la observadapor vía oral. Siguiendola misma patita que
la utilizadaen los estudiosde seguimientoporvía iv (4.Ill.b), se muestrearonlos ganglios
submandibularesde ratonesinoculadospor estavía con dosisde 1V Lmlratón hastael día
5 pi. Los resultadosobtenidos(no mostrados)reflejabanuna dinámicay tasade recuperación
a partir de dicho órganosimilar a la retiejadaen la gráfica 3c, pero con un retrasode 48
horas hastaobtenerlos primeros aislamientos.A nuestrojuicio, este retraso nos permite
sustentar,suficientemente,la idea de que en la inoculaciónpor vía oral podría haberuna
penetracióna nivel del tracto digestivo superior,ya que aparecenantes en estaestructura
cuandola inoculaciónespor vía oral que cuandoserealizapor via xv.
Otros autores(Raez,citado porGaillard et al., 1987a)han señaladola posibilidad de
que las listeriaspenetrena travésde otrasmucosascomo la corneal,bronquial o conjuntival.
por lo que no seríaextrañola utilización de la mucosadigestiva a diferentesniveles. Sin
embargo,a la vista de nuestrosresultados,esta, llamemos,puerta superiorpareceser de
menorimportanciacuantitativaque la mucosaintestinal,ya quetanto los valoresencontrados
comoel tiempo que tardanen desaparecerlas listerias de esteganglio es menorque en el
caso del ganglio mesentérico.Sin embargo,es de gran trascendenciacualitativa, ya que
suponesoslayarla acciónde los ácidosgástricos,lo que podríarestarimportanciaal posible
efectodel pH gástricorespectoal establecimientode la infección. Además,representauna
117
:1

Resultadosy diseusson
grave carenciaen aquellosestudiosque se sirven de la vía intragástrica(ig) como vía de
inoculaciónfrentea la vía oral (Lópeza al., 1993).
Desdeel punto de vistahistológico las lesionesencontradasen los ganglioslinfáticos
mesentéricos,en ambosgrupos,fueronligeraslesionesmultifocalesde tipo piogranulomatoso
(PMNSy macrófagos)desdeel día2 hastael 4 pi. Estaslesionesseencontrarondistribuidas
de forma constantepor las áreasT-dependientesde la cortezadel ganglio. La mayoría de
estaslesionesse encontraronasociadasa la presenciade listerias. Hay que notarque, de
formageneral,el gradode inflamaciónen las placasde Peyerno secorrespondiócon cambio
histopatológiconotableen hígado,bazoo ganglioslinfáticos mesentéricos.
En lineasgenerales,nuestrosresultadosen bazoy gangliosmesentéricosseasemejan
bastantea los expuestospor Audurier et al. (1981)durantelos 3 primerosdías. Este autor
utilizó la vía oral con tres cepasde L. nwnocyrogenes4b obteniendoun aumentode los
recuentosen esosdías; sin embargo,nosesimposiblehacercomparacionesmásallá de ese
tiempo al interrumpir sus resultadosen ese punto. En nuestrocaso, los aumentosen los
recuentosson muy similares,aunquecon valoresalgo inferiores a los encontradospor este
autor. Estasdiferenciaspodríanserdebidasa la utilización decepasdistintas.Los resultados
obtenidosen el bazoconcuerdan,también,con los ya citados(Archinal y Wilder. 1988a;
Cossarty Mengaud,1989),y en estecasomásexactamentequeen la vía iv. Así, por vía oral
el incrementodel númerode listeriasen bazoseproducehastael día3, e incluso4 en el caso
de los deficientes.Esto parececonfirmarunamismadinámicade infecciónseacualseala vía
de inoculaciónempleada,aunquecadavía sediferenciapor el tiempoquetardaen controlarse
esainfección. En estesentido,porvía oral el númerode listerias en hígadoy bazo escero
hastael día 1 pi, mientrasqueen intravenosaesya alto en las primerashorasdebidoa la falta
de filtro intestinal.
En cuanto a los resultadosobtenidos del hígado, el hecho de que las listerias
desaparezcan4 díasantesen los controlesqueen los deficientes,podríaestarrelacionadocon
una másprontasensibilizaciónde los linfocitos T, tal y comoseñalanotros autores(Goosens
ce aL, 1988), esta activación debe suponersealteradaen los deficientes,tal y como se
estudiaráen el apartado4.X.
118
.1 E

Resultadosy discusión
Paradójicamente,aunqueseobtuvieronrecuentosen todos los órganosmuestreados,
tan sólo se detectaronlesiones histológicas significativas en el hígado de los animales
deficientes.Las lesionesobservadasconsistieronen microabscesosdistribuidos de forma
aleatoriapor el parénquima,en algunasde las cualesse pudieron observarlisterias. Las
lesionesdescritasse observaronlos días 2 y 3 pi. El resto de los órganosestudiadosno
presentólesiónalgunadurantela expenencia.
Finalmente debe reseñarseque las diferencias observadasen las gráficas fueron
estimadascomo significativastras el estudioestadísticorealizado,presentandoun valor de
p<o,05.
4.IV.b- CON LISTERIA MONOCYTOGENES112a
Los resultadosde la infeccióncon l/2a porvía oral puedenobservarseen la tabla 14.
Los valoresobtenidosen tubo digestivo son realmentebajos,no superandolos ío~ Lm/g en
el intestinogrueso,de donde, además,desapareceen 48 horas; respectoa otros tramosdel
tubo digestivo,exceptuandoesófago,los recuentossonprácticamentetestimoniales,e incluso
no selograronrecuperarlisteriasapartir de estómagoni intestinodelgado.Nuestrosdatosno
nos permitenexplicar la razónde estehecho,peroen cualquiercasodemuestranuna escasa
capacidadde supervivenciade esteserovaren tracto digestivo.
Pese a todo ello, existe una cierta diseminaciónextraintestinal.reflejada en los
recuentosobtenidosa partir de gangliosy órganosinternos.Segúnestosresultados,la cepa
de l/2a utilizadapresentaun poder infectivo muy bajo,ya que su tiempo de permanenciaen
los órganosno es superiora los 4-5 díasen ningún caso.
4.IV.c- COMPARACION DE AMBOS SEROTIPOS
Si comparamoslos resultadosobtenidoscon los dos serotiposutilizados en nuestros
experimentos(tablas13 y 14, y gráf. 5 y 6), podemosobservarcómo la cepade 4b escapaz
de permanecermás tiempo tantoen órganosinternoscomo en el tubo digestivo,mientrasque
la presenciade l/2a en esteúltimo esprácticamentetestimonial; asimismo,los recuentosen
todoslos casosson superiorescon 4b quecon la cepade 1/2a empleada.Todo lo cual, en el
119
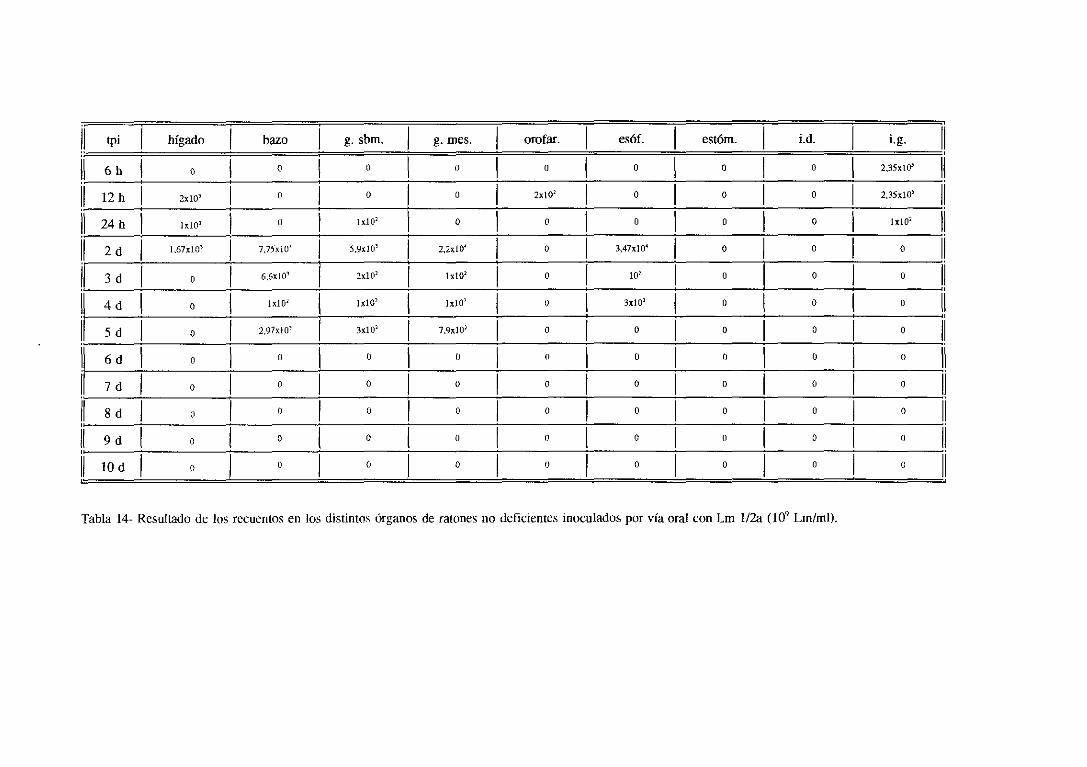
ti)VVVNree
j
>>><re<
4
<5—
~O
OO
OO
Oo
a
‘ea
aa
aa
aa
aa
aa
o
‘‘o‘oo)a
aa
aa
aa
aa
aa
O
<oa
aa
eee-re
.—a—
000<rea
aa
aa
a
ooa
>>>4ej
aa
ao
ao
aa
aa
‘oO)
EtÉ
ao
..4ejej
004—
..4—
00<e.
oa
aa
..0CCtÉ0
0——
<5
<5
o’e>
——rre
aO
aa
a
.0
OO
O>><rs
<5,;CC
<5——ee-o,e
j
Oa
aa
a
>>4e‘o
‘‘eCitúSn
aeej
<.
22
—
aa
aa
ao
aa
.Ci
.0Q
‘ecg
“een“e
“e‘e‘.0
‘et~-‘e00
‘e~
‘e2
E06EoO
)
Ci
o<CiO-
o00
CCo‘eCiCi
ooCiLoo)C
iO
)O
)O
)
‘eoCi
CC
O)
oO-
O)
“eCC
oCi
túO-
‘o‘ooCi
VN
-‘E¡
itoCi
O)
oCiO)
Ci
O)
¡O
)5
-itoO
)
‘eo‘eCi
‘oO)
0±
Ci
.0Ci

<o
ib
Oc
03-o)
orn
—Itcg
o’
c~
t‘e
e-.0
0
<o
f-4
CO
ce0
st
LI,ej03o0,o,It
‘Co
50ib
‘7—
<»1<’03oib
e-s
oCO
—C
i0
3-o
c
ce
-4-o-eceIt03
s-oCI).0
.2--4ceO0
<o
o’,
ev4-t
03>
0
Ci
tiE
o’—
oO)
St
CoO
e-.0
5-
<ot
LIDC
i‘OO
)—
Ci
5.,C
i
03
‘oOo
O)
Ci
<o
~ib
Cj
.40
-4.50e-
CIDCi
-Co
<o“-5
CCOCiCitÉ<oc‘OC
iO
)‘oOC
iO
)C
i
CCOO)
‘eCi
O)
o,03
-‘e
•C
i
O)
“e
.0C
i‘OO
)
EOuIt
0
a[.
01
O)5.
CC01o
Oo
’<o
e-
st
ibC
jcg
—o
03O
O)o
OO
bt~
<oib
c’b0
30
oce
-oo-4ce
O)4>.CCSCo
Co
’
01
O)
CL.CC01o
O0
,50
t-<o
CO—
C03
—a

oo’-
•0ti)
,0a)so
tÉ-
o5-o
N4>¡
~5.
oU)a)
a)
—o
4
4Y
0.5•0
.5
>0Citi.0
-.
oCi
-‘oLoa0
0
Ci
‘O‘uCi
Ci
O)
oCiCCCi
o‘oO)
tú
‘eo.0Ci
Ci
O)
CCoCi
O)
O)
O)
O-
‘oo0)
‘e‘ECi
0)
‘eCi
‘OO)
EouSo
oa)
-4
Y
SÉ‘oti)o
‘0
ID
1~~~

Resultadosy d,scus,on
caso de 4b. aumentaríalas posibilidadesde producir una infección y, en consecuencia,el
desarrollode la enfermedady eventualmentela muertedel hospedador.
En estesentidono debemosolvidarunagrandiferenciaentrelos serotiposempleados,
y expuestapor Kaufmann(citado por Mainou-Fowleru al., 1988),segúnel cual la cepade
l/2a NCTC 7973 resultaser un serotipocon infección no persistente,mientrasque 4b silo
es; las cepaspersistentesserianaquellasque trasinoculaciónintravenosapodríanseraisladas
de bazoa los 6 día pi, mientrasque las no persistentessedanrápidamenteeliminadasy no
podríandetectarseen bazoa los 6 días pi. Aunqueesteestudiose realizó utilizando la vía
endovenosa,nuestrosresultadosconfirman estasobservacionesporotra vía.
Paraconcluir,nuestrosresultadosparecenconfirmarla ideaexistentesobreuna mayor
virulenciade el serotipo 4b, basadaen datos epidemiológicos,expresadapor otros autores
(Barbouru aL, 1992; Broschu al., 1992; Tabouretu al., 1991). Esto podríaexplicar la
mayorfrecuenciade apariciónde esteserotipofrentea l/2a en los brotes de listeriosis tal y
como se expusoen el apartado1.JV.c,dondeseseñalaronporcentajesde apariciónde entre
un 62-64%y un 13-15%respectivamente(Gouletet al., 1989; McLauchlin, 1990), e incluso,
más ampliamente,de un 92% para los serovaresdel grupo4 frente al 8% restantepara el
grupo 1/2 en nuestropaís (GómezMampaso,1993).
4.V- SEGUIMIENTO DE LA INFECCION POR VTA INTRAGASTRICA
Estavía ha sido consideradapor algunosautores(Barbouru al., 1992)como la más
natural para la infección con Listeria, ademásde haber sido utilizada en numerosas
infeccionesexperimentalespor vía <‘oral (Schlech, 1990). En nuestrocasoúnicamentese
realizó con el serovarl/2a, puestoque la gran mayoríade los estudiospor estavía sehan
realizadocon esteserovar,y al haberencontradodiferenciasen el comportamientoentrelos
serovaresutilizados en la vía oral se pretendíaver la posible existenciade diferenciasal
utilizar distintasvías de inoculacióncon estacepa,que ademásesla cepatipo de la especie.
Los resultadosobtenidospor estavía puedenobservarseen la tabla 15. Entreéstos
llamala atenciónla presenciade listeriasen la región orofaringeay en esófagodesdelas 6
horaspi, asícomo su persistenciaen pequeñascantidadesen estaszonashastalos días 6 y
123
Iii

tÉ
o’a~—
000<~cl
000<oco,re
~a
aa
aa
aa
O
‘ea
a000<
aa
aa
aa
aa
a
‘oO)
aa
aa
>>4—a
aa
aa
aa
‘0O)
Ci>4
.j
<5C
i<5‘~
<500
<5>>>4e
aa
aa
a
o
<5—
<5<5
oo’<5
<5<5
<5e-a
aa
a
itEtú
aa
aa
accl
aa
aa
aa
CCtú
aa
aa
aa
<Oa
aa
aa
.0
aa
—
<5—o
eee-SC
<5-—
000<rl
aa
0a
a
‘‘eCitú.0
aa
———..4—
<5e-a
aa
aa
aa
e.0
,.~
ti
-c~0±
“eti“een
‘e‘eLtN
‘eSo
-eho‘eOC
‘eQN“e
E06Ci
cgoO)
Ci
O)
túCi
Ci
Ci
O-
oo--
o-CoCi
“EO)
oCi
‘oO)
CiO)
O)
O)
‘eoCi
CCO)
Ci
oCi
O-
0)
‘eCCoCiCitú5-
‘0itoCi‘o‘oCis
O)”’
‘o44
0
Ci’
Ci—
~O-
CCo
02
0)
e
Co
)“en
fCiR
CC0)0
.
Sr
.2ID
E-.>

st
oo‘O)
.5O)
o(IDa)
O>
St
a..O
~<o
~—
ibej03--e
St~
‘e
Ito4>
e-‘It01It0.
<oib—
ce.0-e~ce
.0UDO—4
-4ce
oO
01‘0<O
)0.>CC01O
oo’
<o
e-<o
oj
01‘~
0O)O’-a
Ob<o
a.-<o
ib
soibev03o
<o
’
St<o
It“ee
-
‘<o
41•1
y,
ySL
O
/4</>
5<‘0<
oNce
.0
01-o
.-O
)0.-CC01O
o¡
—¡ o
o’<o
e-st
ibev
03—
oo
0~~~~
:ev
>0
3
—o.’
4,
-
—C
iev
cg
Ci
tiEOIt
Ci
oO)
¶4>’.
It‘C
i
<ItCi
00It
1-
ItOC
i‘OO)C
i
CiCCOCCOCiCitÉO-
‘OCiO)
‘ooCiO)
Ci
O)O)
O-
‘o—
OIt
—0.
0)
O‘e-‘E
5<C
i
-~O
)0<
‘e•0<
Ci
‘0o
00Eou4>.‘C
i
<oibejcgoo’
<o
¡eva.-
aso
y¡
00
<->
it
4’—
4c.D
y‘0
~.’
laej
Da—
O
St
It
“e
5<‘~0O
)0.-CC0>
¡ao~
<oe-
st
it—
ev03
—o
oev03o

O
‘0o>.
0.
<~
0—
orl)a)a)
U.5
1.E
oa)
‘e
Itso
5
o‘oUDEz~
yo
‘0
.7<
y.,,
i
O
‘y
O’--
O~00.O
otÉ44
‘oUD
y
a)
e-o5-
o
5
tiECi
oO)O)
oCi‘OO)C
iC
iO)oCiCCC
i
o‘oO)
túo.0CoCi
E)‘o2CiO
)C
i
CCoO)
“e1-‘6Ci
O)
-eCi‘OO)C
i
CoEouOC
~0It
00
15
0<SL

Resultadosy discusión
5 respectivamente.Estapresenciapodríaser debidaa la propia técnicade inoculación,que
suponeen muchoscasos,unacasiinapreciable,peroexistenteerosiónde la mucosaesofágica,
y a que la retiradade la sondaimplica el depósitode una ciertacantidadde inóculo en el
tracto digestivo superior, cuyo epitelio, levementealterado,permite a Listeria una mejor
colonización,hechodemostradohistológicamentepor Domingo (comunicaciónpersonal).No
seobtuvieronrecuentosde pulmonesa partir de ninguno de los animalesmuestreados,datos
no reflejadosen la tabla 15 ni en las gráficascorrespondientes,por lo quepodemosconsiderar
quela inoculaciónserealizócorrectamente.
Por otra parte, también llama la atenciónel aislamientode listerias a partir de
estómagoe intestinodelgado,los días 3 y 1 respectivamente.A esterespectodebemosdecir
que estosaislamientosfueron realizadosúnicamenteen uno de los ratonessacrificadosese
día, y podría debersea la existenciade factoresindividuales, o a una mala prácticade la
inoculación,sin que podamosexplicarestehecho.
Sin embargo,los recuentosde intestinogruesosonsimilaresa los que seobtienencon
esta misma cepa por vía oral, lo que confirma el bajo poder de colonización intestinal
señaladopor otros autores(Marco et al., 1992b).
Segúnnuestrosresultados,se obtienen recuentosmayoresen bazo que en hígado,
siendo ademásla persistenciamayor en el primeroque en el segundo(graf. 7). Además,los
recuentosmayoresseobservanentrelos días3 y 4 en ambosórganos,lo que concuerdacon
otros autores(Barbour a al., 1992; Bracegirdlea al., 1992; Domingo et aL, 1992). Sin
embargo,algunosde estosautores(BarbouretaL, 1992;Bracegirdleu aL, 1992)afirmanque
no seaíslancantidadesimportantesde Listeria a partir de hígadoy bazo hastalos 3-4 días
de unainoculaciónintragástrica,aunquesegúnnuestrosresultadospodemosencontrarlisterias
en hígadoy bazo a las 24-48 horas post-inoculación,lo que parececonfirmar resultados
anterioresobtenidospor nuestrogrupo(Domingo et al., 1992)
4.VI- COMPARACION DE LAS VIAS ORAL-INTRAGASTRICA
Los resultadosobtenidosporambasvíasson muy similaresen hígado,bazoeintestino
grueso,tal como sepuedeapreciaren las gráficas7a, 7b y 8e; sin embargo,los resultados
127
II

Resultadosy diseusion
obtenidosen ganglios,estómagoe intestinodelgadodifierenengranmedida(gráficas7c, 7d,
8c y Sd). Por otra parte,los resultadosobtenidosen orofaringey esófago(gráficas8a y 8b)
puedendebersea la posible erosiónocasionadapor la sonday al depósito de inóculo
producido al retirarla, tal como se explicaen el apartadoanterior. En definitiva la vía ig
ofreceresultadosmenosconsistentesquelos obtenidosporvía oral y, en consecuencia,la vía
ig no pareceseruna elecciónacertadacomovía de inoculaciónya que:
- la utilización de sondalesionael epitelio esofágico;
2- seproduceun depósitode inóculo en la zonaerosionadaal retirar la sonda;lo que,
unido a lo anterior,implica una colonizacióndel epitelio de la orofaringey esófago;
3- presentauna mayorcomplejidadque la vía oral;
4- requiererealizarcontrolesde pulmónparaasegurarla correctainoculación;
5- no esequivalenteal modo naturalde infección.
Nuestrosresultadosconfirman lo ya expuestopornuestrogrupode trabajo (Lópezet
al., 1993), al no encontrarmejoraalgunaal utilizar la vía ig con respectoa la vía oral, ni
siquieraparaobteneruna mejor respuestahumoral.Por tanto, contrariamentea lo expuesto
pordiversosautores(Barboura al., 1992)que considerana la vía intragástricacomola más
naturalen todos los sentidos,porquealterael contactode Listeria con las zonaslinfoides del
tracto digestivo superior,queintervienen,segúnnuestrosdatos,en el procesode penetración
de estosmicroorganismosen el hospedador.Además, de no presentarmejora algunacon
respectoa la vía oral, esde mayorcomplejidadal tenersequeadministrarel inóculo con una
sondaindividualmentea cadaanimal, y depositade hecholisteriasen orofaringe,ademásde
lesionar el epitelio esofágico,falseandolos resultados.Otro factor que desaconsejasu
utilización es la necesidadde realizarrecuentosde los pulmonesde los animalesinoculados
paraasegurarnosla correctainoculación(Marco a aL, 1992b>.
4.VII- SEGUIMIENTO DE LA INFECCION POR VIA ORAL CON 1O~ 7DIAS
Las tablas16ay 16b nos muestranlos resultadosobtenidoscon la dosisde ío~ Lm/ml
administradasdurante7 días. En estastablas puedeobservarsecómo los valores de los
deficientesson ligeramentesuperioresa los obtenidosen el caso de los controles,si bien
pareceque el comportamientoen amboscasosfuera muy similar (gráficas9 y 1<)). En este
128
¡ ¡¡ ¡ ¡
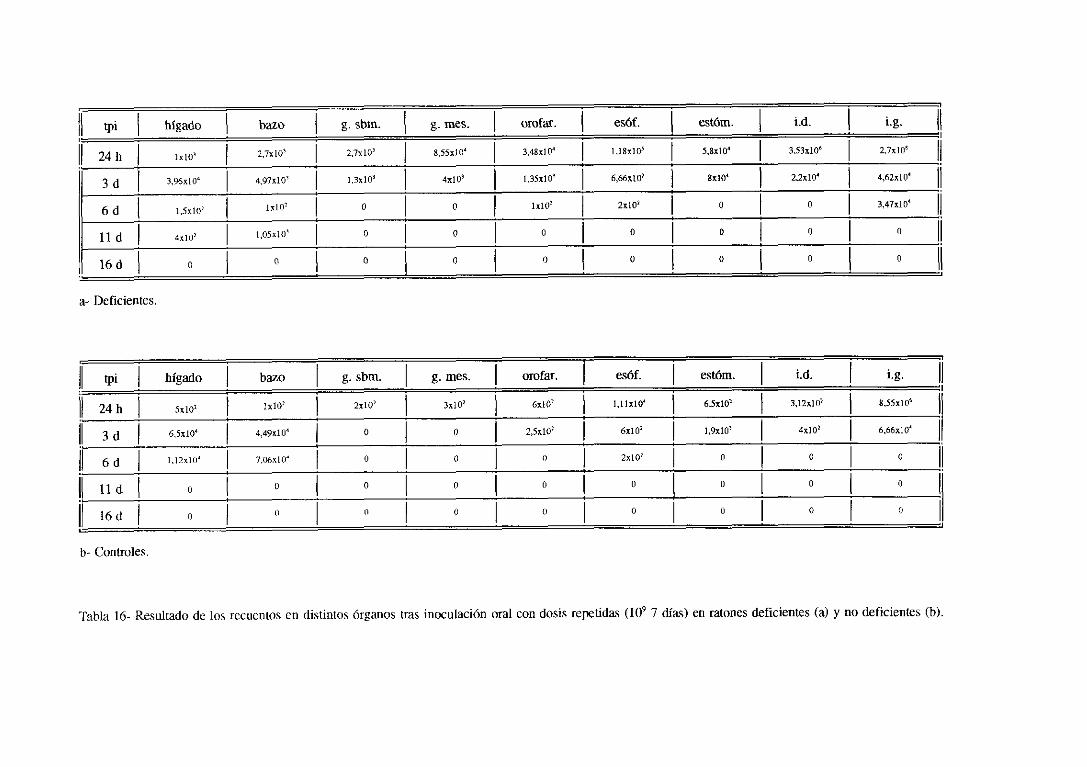
.4
<5ze-e>
000*
ee-
0*re
aa
.4
UUNr
<5qej
aa
a
‘‘0~E)0<UN
00
E)
——
SC<O<O
>4e
j
.4<
L.0
EO
.4~re
re
00
‘oE)
Etú
<5—0<VNoc
00*a
a
..0‘otú
<5
<5
~‘~
e-re
cl—
oa
..0
<500
’0
*
<5<50—
a
““eCitú.0
<5
2—
0<.?~
Ore
VVVN—
<5—0<0*
o
sso
Sn
-tcg
“e~“eS
o“e—
‘e
~i)
—
UUUNVN00
..~SCSCSC
aa
a
“e000<e
j—re
...40*~
~a
‘‘oSC
—
‘0
<5>4<<O
eej
aa
~<
5<5~
‘~E
<Oo
aa
a
CC~E~i)
22a
aa
o
‘otú
ej
~
o<5
0*
e
aa
SSn
5,~VN
~<O
20
0.4
‘e‘C
ie
Sn
~
‘oO)
eO)
O)
SnO)Ci
,0CCO)
CiO)
O)S
t0)
‘eoCi>0
Ci
‘oO)
Ci
O)
O)S
tO
)“e‘oO
)C
ioCiO-
Ci
O)
i06O-
CCCoo‘eC
ioO
)
Ci
O-
oCi
‘oO)CiCitúO-
‘oCCoCi
CC‘EC
iO
)CC2C
iO
)C
iO)O
)UcCCoO
)‘eo5‘oO
)‘oO
)
‘0C
i—
o.00$
.0

<o‘a
Oe,
‘04=
,-
0.
-0
aLo
-O
CIDz
r1~o’
Srz
<oC
i
“e‘e0.
e-o
0.•—
st
-4b0Uo”
itt
O
o<oibce
-4
-ev‘-4
03oc0.
—It
ce
o,o
ct
rO<o
‘Co
CID
so---‘e
1’—Ci
0*‘ee)
2É.o
2o
—0.
<4—’
0O
)Ci
-eo
0.—
‘0.0
OE
<o
~1-
“e“e
e-06
o<o
0.o
‘2‘a
‘oev
CiO)
oCi
a‘ooO)Ci
<Oso-1—
.ev
Io,<o
e-0.
¡—e
-
<oo
½0*
-ev
ce
/1~
½“
1I4
~Q
Un
.2¡
00
00
,<o
e-<o
00
*—
‘ootÉCi
E)
CCoCi
O)
Ci
2>O)
OsO
)O
-0.
ito‘e
0)
‘0‘eC
iO)
“e0.
Ci
O)
‘eCi
a—O>EouQ
N
+
01¾
.O
)rL.CC01o
.9’..cg
OO
b<o
5..so
ib0’
03—
O
oNce-ootce‘-4ce
oo’
st
e-
<oib
0*ev
03—
a
-t.
be‘0<u0..CCU
.aa
<oa..
<o‘a
0*c.D
cg—
a

(O
¡O
‘O
4/
-O
~0~
“ea4
E)O-
.0¡
.5:
O<4~o
‘o0±
EE)“e
06Ci
OO)
Ci
‘oO)
Ci
Co
O)
oCi
YCC-E)tú
“esCi
E)CCOCiE)
CCoE)
o“e
U.—“eC
io
o>-E)-“e
.04
Ci
.0.5
‘0e
—O)
o>CCO
>
O‘e
OUo
.0.5‘0
c.5so
~.0
OUDa)a)
zY1
o‘eCCo)‘e‘e
zY1
oCC5.-
‘oU)a)1-CC
5-‘o>-
LooCC
‘OO
O
SÉ‘oUD0
FI

Resultadosy discuston
sentido,pareceque en los deficientesla penetraciónde listeriassealargaalgomásque en los
controles,como indicanlos recuentosobtenidosde los ganglios,en los que no desaparecen
las listeriashastadespuésdel día 3; mientrasqueen los controlesla penetraciónacabaantes
de ese día.
Otro hechoque pareceavalar la tesis de un mayor tiempode permanenciaen el caso
de los deficientes,esque los recuentosno desaparecendel intestinogruesohastadespuésdel
día 6 en éstos,y antesde esedíaen el casode los controles.
Si comparamoslos resultadosobtenidosconunaúnicainoculación(gráficas3 y 4) con
los obtenidoscon la inoculaciónrepetidadurante7 días (gráficas9 y 10), podemosobservar
que los animalesinoculadosparecencontrolar la infecciónentrelos días5 y Y pi en ambos
casos.Estosresultadosno secorrespondencon los criterios expuestospor diversosautores
(Galsworthy, 1987; Marco et aL, 1991), quienesproponenuna posible alteración de la
capacidadde la respuestainmune del hospedadorderivadade la existenciade lesionesen los
órganoslinfoides y de un efecto inmunosupresorde la propiaL. monocvrogenes.Nuestros
datos demuestranque, en amboscasos,la infecciónes controladaen el mismo periodode
tiempo, lo que revelaque el estatusinmunitario de los animalesno seencuentraalterado.Sin
embargo,esnecesarioresaltarque la ingestiónrepetidade listeriasdurante7 díasaumentaría
las posibilidadesde que se produjera infección, lo que podría concordarcon la mayor
mortalidadobservadacon estapautade inoculación,aunquea diferenciade lo que ocurríaen
la vía iv no podemosestableceruna relación directa entreestehecho y la cinética de la
infecciónobservada.
4.VIII- DEFICIENCIA EN SELENIO Y VITAMINA E Y SUSCEPTIBILIDAD A LA
LISTERIOSIS
Laexistenciade mortalidadesmayoresen el casode los animalesdeficientes,asícómo
que, en general,los recuentosobtenidosde órganosinternosseanmayoresen éstosy quelas
lesioneshistológicasseantambiénmás intensasen los deficientes,nosinclina a pensaren una
mayor susceptibilidada la listeriosisen el caso de estosanimales.Dicha susceptibilidadse
deberíaa los siguientesfactores:
¡32
II

Resultadosy discusión
- alteraciónde la función fagociticade neutrófilosy macrófagos;
2- menorexpresiónde receptoresde 1L2 en los linfocitos;
3- menorpresentaciónde antígenopor partede las célulascorrespondientes.
Estosresultadosse encuentranen clara contradiccióncon los expuestospor Murray
y Murray (1985),segúnlos cualesla deficienciaen selenioconferidauna mayorresistencia
a la listeriosis, disminuyendo la mortalidad en ratones. Sin embargo, estos autores
suplementaronla dieta con vitamina E; que, comoya hemosapuntado,puedeactuarcomo
sustitutodel selenio,suavizandolos efectosde su deficiencia.Además,no secomentani la
cantidadde selenioen la dietadeficiente,ni el tiempoquetardaronlos ratonesenconvertirse
en deficientes.Por tanto, podríahaberocurridoque los ratonesno fuerandeficientesen el
momentode la experiencia,y la mayor resistenciafueradebidaa otro fenómenodistinto a
la deficienciaen selenio.
Los mecanismosde resistenciaa la listeriosis son muy complicados,tal y cómo
apuntamosen el apartado1.V.b de la introducción.Debemosrecordarque la resistencia
naturala la listeriosisradicaenla capacidadde estableceruna respuestainflamatoriaasociada
acélulas1 sensibilizadas(Archinal y Wilder, 1988a),cuyo fallo encualquierade susestadios
previosseriael responsablede un aumentode la susceptibilidada padecerestaenfermedad.
Al inicio de la infección, las primerascélulasfagociticasen llegar al hígadoson los
neutrófilos,enestatempranallegadade los neutrófilospareceestarinvolucradael TNF-alfa
(Mielke eral., 1993).Estostienenla misión de evitar la propagaciónde la infección,paralo
cual destruiríanlos hepatocitosinfectados,dejandolibres las listeriasque seencontrabanen
su interior, y que así quedanexpuestasa la acción de los macrófagosy monocitos(Conlan
y North, 1993, 1994). Por tanto la acción de estascélulas es de vital importancia parael
hospedador,ya que limitan el crecimientoinicial en los hepatocitos,ademásde dejar las
listerias expuestasa la acción de los macrófagosy monocitos. Duranteuna deficienciaen
selenioy vitamina E, las funcionesfagociticasde los neutrófilosse encuentrantan alteradas
como las de los macrófagos(Gyanga aL, 1984; Erskine a aL, 1989), por lo que es muy
posible que la acción limitante realizadapor ellos se encuentredisminuida. Esto último
133

Resultadosy discusión
facilitaría la formaciónde focosinfecciososprincipalmenteen hígadoy bazo,en los cuales
las listerias semultiplicaríanantesde la llegadade los monocitos/macrófagosa partir de las
24-48horas post-infección, lo que pareceestar en total acuerdo con los resultados expuestos
en la tabla 11.
Una vez englobadaslas listerias en el fagosomase produceuna acidificación del
mismo (Chastelliery Berche, 1994). Estaacidificaciónresultaindispensableparaque pueda
actuarla Listeriolisina O (LLO), cuyo pH óptimo de actuaciónes5,5 (Bercheet al., 1988;
Cossarty Mengaud, 1989). Además,el fagosomaes un medio carentede hierro, lo que
estimulaaún más la secreciónde estacitolisina (Bercheet al., 1988).Mediante la acciónde
éstala bacteriaconsigueliberarsedel fagosomaantesde que resultedañada,y multiplicarse
en el citoplasma,dondeel ambienteresultamenosdañinoparaestasbacterias,y dondepor
añadiduraexistedisponibilidadde hierro (Bercheetal., 1988;Cossarty Mengaud,1989).Esta
multiplicación resultaesencialparala inducciónde una inmunidadmediadapor linfocitos T
(Bercheet al., 1987; Brunt et al., 1990; Chastelliery Berche, 1994),y explicapor qué esta
inmunidadno seobtienecon bacteriasmuertas.
El númerode bacteriasque consigueescapardel fagosomaantesde la fusión de éste
con el lisosomaresulta muy pequeño(Chastelliery Berche, 1994), pero suficiente para
producir las alteracionesposteriores.Estasbacterias,unavez multiplicadasen el citoplasma,
son capaces de moverse mediante un mecanismoactina-dependientee infectar células
adyacentes(Tielney y Tielney, 1993; Chastelliery Berche, 1994) sin necesidadde salir al
medio extracelular,lo que las expondríaal sistemainmune del hospedador.
Conjuntamentecon la acidificacióndel fagosomase producela explosiónoxidativa
en el fagosoma,mediantela que seaumentanconsiderablementelos compuestosde oxígeno
(peróxidos principalmente)que sirven como mecanismobactericida general, aunque no
parecentenerdicho efectoen la listetiosis(Harrington-Fowleret al, 1981; Higginbothamu
al, 1992).
Por tanto, al liberarsedel fagosomaes muy posibleque estasbacteriasdesencadenen
la muerte del macrófagopor un mecanismosimilar a la apoptosis (Greenspany Aruoma,
1994; Zychlinski u aL, 1994),quele imposibilitaríapararealizarsu misióncomo presentador
134
II

Resultadosy discusión
de antígenos.Pero la presentaciónde antígenostambiénpareceverseafectadapor la LLO
(Cluff a al., 1990); según este autor, la LLO puede inhibir esta función, evitando la
estimulaciónde los linfocitos T, y, por tanto, la activaciónmacrofágica.En el caso de la
deficienciaen selenioy vitamina E ya hemosexpuestoque las membranasde lascélulasse
encuentranafectadaspor defecto de GSHPx, y que ello implica una mayor facilidad de
Listeria parasalir del fagosoma.Estasalidaestáacompañadapor liberaciónde LLO, lo que
finalmenteimplicaríauna mayorcantidadde LLO en la célula,que podríainhibir la función
presentadorade antígenos.
Por otra parte,se ha descritoque en los animalesdeficientesen seleniola expresión
de receptorespara la 1L2 en los linfocitos T se encuentradisminuida (Kiremedjian-
Schumacherez’ aL, 1990, 1992; Roy a al., 1992, 1993).Estadisminuciónen la expresiónde
estosreceptoresconllevaríauna menoractivaciónde linfocitos T, y estamenor activación
linfocitaria implicaríauna menorproducciónde linfoquinasy, porello, unamenoractivación
de macrófagos.No debemosolvidarqueson los macrófagosactivadoslos responsablesde la
eliminación de las listerias (Harrington-Fowleret aL, 1981), y un retrasoen su activación
permitidauna mayormultiplicación de las listerias,lo quetendríacomo consecuenciael que
el individuo tuvieraque enfrentarsea un númeromayor de éstasal queinicialmentetendría
que haberhechofrente,y que, por tanto, el procesoadquirieseuna mayor gravedad.
Además, en el caso de una deficienciaen seleniose produce una disminución de
GSHPx,y, por tanto, sedisminuyela detoxicacióndel H202 cuyascantidadesaumentan.Esta
menorconcentraciónde GSHPxconlíevaunamenorcantidadde glutationoxidado,que para
su reducciónnecesitabael pasode NADPH a NADP, lo queimplica unareduccióndel NADP
disponible.En líneasgeneralesuna deficienciaen selenionos conducidaal mismopunto de
partida que los mecanismosdescritos para la apoptosis (Greenspany Aruoma, 1994;
Zychlinski u aL, 1994) ya que el excesode productosde oxigeno conlíevala alteraciónde
la membranamediantesu peroxidacióny acciónde fosfolipasas,y, como resultadode ello,
a la alteracióndel equilibrio del Ca2, rotura de las membranas,alteracióndel ADN celular,
y, finalmente, la muertecelular. La alteracióndel equilibrio del Ca2~ puedeverse agravada
en los casosde deficienciaen a- tocoferol (Eskewa aL, 1989), por lo que en nuestrocaso
la propiadeficienciacolaboradacon los fenómenosconducentesa la muertecelular.
135

Resultadosy discusión
Vemos,pues,cómo la deficienciaen selenioproduceuna alteraciónen la respuesta
inmune de base celular por un fallo en la presentaciónde antígenospor parte de los
macrófagos,y por un fallo en la activación de los mismosal estardisminuidala expresión
de los receptoresparala 1L2. Hemosseñaladoanteriormentequelos factoresque influyen en
la resistenciaa la listeriosis sebasanen la capacidadde establecerunarespuestainflamatoria
efectiva, la capacidadde los macrófagospara evitar el crecimiento bacterianoen los
momentosinicialesde la infeccióny el tiempoque setardeen instaurarunarespuestainmune
adquiridamediadapor células (Archinal y Wilder, 1988a, 1988b); todos estosmecanismos
se encuentranalteradosen los individuosseleniodeficientes,y, poreste motivo, resultarían
mássusceptiblesa la listeriosis,puestoqueestospatógenosencontraríanmayoresfacilidades
para su viabilidad en estoshospedadores.
136
¡¡ i

5. CONCLUSIONES
.1 L~ Iii

Conclusiones
PRIMERA
En el modelo experimentalmurino, los animalesdeficientesen selenioy vitamina E
presentanmayorsusceptibilidada la listeriosis,valorableen indicessuperioresde mortalidad
y en mayoresrecuentosde bacteriasa partir de órganosinternosque los gruposcontroles.
SEGUNDA
La deficienciaen selenioy vitamina E, incluso inaparente(no valorableclínica ni
histopatológicamente),podría ser consideradacomo un factor predisponenterespectoa la
listeriosis.
TERCERA
La presencia de Listeria monocytogenes, cuando es inoculada por vía oral,
simultaneamenteen los gangliossubmandibularesy mesentéricos,y con anterioridada su
recuperacióna partir de bazoe hígado,sugierela existenciade variaspuertasde entradade
estemicroorganismoen el tracto digestivo.
CUARTA
La mortalidadregistradacon la misma dosistotal inoculadaporvía oral esmayor si
ésta se fraccionaen varias tomas que si se administra en una única toma. Puestoque el
desarrollode los casosde listeriosisestáasociadoal consumode alimentoscontaminadoseste
efecto acumulativo podría tener relevancia desdeun punto de vista epidemiológico y
experimental.
OUINTA
Los recuentosde bacteriasobtenidosa partir de bazoe hígadomuestranun segundo
incrementoentre el 50 y 80 día post-inoculación,lo que sugiere la existencia de una
reactivacióntardíade la infección,cuyo tiempo de aparicióny magnitudseríadependiente
de la dosisy de la vía de inoculaciónempleada.
138

Conclusiones
SEXTA
El seguimientode la infección tras inoculaciónintragástricaofreceresultadosmenos
consistentesquelos obtenidosmedianteinoculaciónporvía oral, porlo queéstaúltimaparece
másadecuadaparareproducirexperimentalmentela enfermedad,ya quereproducemejor las
condicionesnaturalesde la infección.
SEPTIMA
Cuandoseinoculanlos serovares4b y l/2a a las mismasconcentracionesporvía oral,
con el serovar4b seobtienenrecuentosmásaltos y prolongadosen intestino, bazoe hígado.
Estehechopodríajustificar la mayor frecuenciade casosde listeriosis de origen alimentario
debidosal serovar4b.
139
¡ ¡ ¡• L ¡

6. RESUMEN

Rcsurnen
6. RESUMEN
En el presentetrabajosehaestudiadoen el modeloexperimentalmurino la influencia
de la deficienciaen selenioy vitaminaE en la susceptibilidada la listeriosis.Paraello sehan
realizadoestudiosde mortalidad,estableciéndoselas tasasde mortalidad por vía oral con
dosis únicasy repetidas,asícomo las DL50 por vía intravenosa.Asimismo, seha estudiado
lacinéticade la infecciónporvía orale intravenosa,medianterecuentosde L. rnanocytogenes
a partir de órganosinternosy tracto digestivo; se ha comparadoademás,la cinética de la
infección utilizando las vías de inoculaciónoral e intragástrica,el efectode dosisrepetidas
frente a dosis únicaspor vía oral, y la infección por vía oral entrelos serovares4b y 1/2a.
De los distintos experimentos,se han obtenido resultadosque indican una mayor
susceptibilidada la listeriosis en el caso de la citada deficiencia; que se recuperaL.
rnonocytogenesa partir de gangliosmesentéricosy submandibularesen las primerasfasesde
la infección, lo que sugiere distintos puntos de entradaa partir del tracto digestivo,
probablementeestructuraslinfoides (placasde Peyer,tonsilas,etc.); y que en la listeriosis
experimentalpareceexistir unareactivacióntardía,cuyo tiempode apariciónseríadependiente
de la dosis y la vía de inoculación.Asimismo, se ha podido comprobarque la vía oral
presentaevidentesmejorasrespectoa la intragástricaa la hora de realizarestudios~~sobrela
listeriosis;así como que la administraciónde una misma dosis fraccionadaen variastomas
produce una mayor mortalidad que si se administra en una única toma. Finalmente, la
inoculaciónpor vía oral de los serovares4b y l/2a demostróque con el serovar4b se
obtienenrecuentosmás altos y prolongadosen intestino, bazoe hígadoque en el caso del
serovarl/2a.
141

7. BIBLIOGRAFIA

I3íbliografía
ALLERBERGER,E. y GUGGENBICHLER,1. P. (1989).Lisíeriosisin Austria: reponof anoutbreakin 1986.ActaMicrobiol. Hung. 36, 149-152.
ARCHINAL, W. A. y WILDER, M. 8. (1988a).Susceptibilityof HRS/Jmice lo listeriosis: macrophageactivity.Inilecí. lmmun. 56< 6113-MS.
ARCHINAL, W. A. y WILDER, M. 5. (1988b).Susceptibilityof HRS/Jmice to listeriosis:Dynainics of infection.lnfect. linmun. 56, 607-612.
ARMSTRONG, 0. (1985).L. monocvtogenes.En ‘<Principies andpracticesof infectiousdiseases”(eds.Mandel, O.L., Douglas.R. O. y Bennet,J. E.), pp. 1177-1 186. Ed. Wiley & Sons,Nueva York.
AIOJDURJER,A., PARDON, P., MARLY, J., LANTIER, F. y LOULEGUE, J. (1981). Mesurede la virulencechezla sourisde dilférentesbactériesappartenantau genreListeria. Ann. Inst. Past. Immunol. 1320, 191-200.
AUDURIER, A., TAYLOR, A. O., CARI3ONNELLE, B. y MCLAUCHLIN, J. (1985), A phageíyping systembrListeria monocyíogenesami lis usein epidemiologicalsiudies.Clin. Invest, Med. 7, 229-232.
BACKAIL, K. A. y SCHOLZ, R. W. (1919). Referentevalueslot a fleid tesílo esúmateinadequateglutaihioneperoxida.seactiviry andseleniuni statusin theblood of cattle. Ant 3. Vet. Res.40, 733-738.
BARBOUR, A., RAMPLINO, A. y HORMAECHE,C. (1992). The infectivity of Listeriamonocyto~enesfollowinginíragastricinoculation into mice.Proc.Xliii InternationalSymposiumon dic Problemsof Listeriosis(Copenhagen)235-236.
BARLOW, R. M. y MCGORUM,E. (1985).Ovinelisterial encephalitis:analysis,hypoíbesisandsynthesis.Va. Rec.116.233-236.
BEDWAL, R. 8., NAIR, N., SHARMA, M. P. y MATEUR, R. 8. (1993>. Selenium-iísbiologicalperspectives.Mcd.lIyp. 41, 150-159.
BERCHE, P., GAILLARD, J. L. y RICHARD, 8. (1988). Invasivenessand intracellular growth of Listeriamonocytogenes.tnfect. Suppl.2, 145-148.
BERCHE, P., GAILLARO, J. L., SANSONETI”I, P., GEOFFROY, C. y ALOUF, J. E. (1987>. Towardsa belterunderstandingof dic molecularmechanismsof intracellulargrowth of Listeria monocyro~=enes.Ann. Insí. Past.Microbiol. 138, 242-245.
BERENGUER, J., SOLERA, .1., DIAZ, M. D., MORENO, 5., LOPEZ-HERCE,1. A. y BOUZA, E. (1991).Listeriosisin patienísinfeaedwith human immunodeficiencyvirus. Rey. tufecí. fis. 13,115-1190
BILLE, J. (1988). Epide¡niology of human listexiosis in Europe.Conferenciaexpuestaen el SIM. California. USA.2-5 Octubre.
BLANCO, M., FERNANDEZ-GARAYZABAL, 1. E., DOMINGUEZO L., BRIONES, V., VAZQUEZ. J. A.,BLANCO, J. L., GARCíA, .1. A. y SUAREZ, 0. (1989>. A technique for dic direcí identiticadon ofhaernoly¡ic-paíhogenlisíeria on selectiveplating media.Leí. Appl. Microbiol. 9, 125-128.
BLANCO, M. M., DOMíNGUEZ, L. y VAZQUEZ, j. A. (1993). Listeria: Deteccióne identificación.En “Listeriaen alúnentos.ConferenciaConsenso”,PP.61-80. Ed. M0 Sanidady Consumo.León.
BLENDEN, D. C., KAMPELMACHER, E. 1-1. y TORRES-ANJEL,M. . 1. (1987), Zoonosisupdate.Listeriosis. J.Am. Va. Mcd. Assoc. 191, 1546-1552.
143

B <él Lognsfía
BLENDEN, D. C. y SZATALOWICH, E. T. (1967).Ecologicaspectsof listeriosis.1. Am. Vet. Mcd, Assoc. 151,1761-1766.
BLODGEYI?, Di,, SCHURIG, 6. 6. y KORNIEGAY, E. T. (1986). linmunornodulationin weanling swine witlidietaryselenium. Am. J. Vet. Res. 47, 1517.1519.
BLOOD, O. C., RADOSTITS,O, M. y HENDERSON,J. A, (1986). “Veterinarymedicine”, Di. BailhiéreTindail.Londres.
BOYNE, R. y ARTHUR, J. R. (1990).Anaemiaami changesin erythrocytemorphologyassociatedwith copperandseleniumdeficienciesand dietaryrestriction iii rats. Res. Vet ,Sci, 49, 151-156.
BRACEGIROLE, P., FIIZGEORGE, R. B., WEST, A. A. y BASKERVILLE, A, (1992). A study of thepathogenesisof virulent and aviruíent strainsof Listeria monocyto2eneswbcn infectedvia intragastricor aerosolroutes.Proc. XIth InternationalSymposiumon the Problemsof Listeriosis(Copenhagen)241-242.
BRACKET, R. E. (1988), Presenceaudpersistanceof L. monocytogenesin food ami water. FoodTechuol. April,162-164.
BRAUN, U., FORRER,R., FURER,W. y LUIZ, H. (1991). Seleniumandvitainin E in blood seraof cowsfromfarmswith increasedincidenceof disease.Vet. Rec. 128, 543-547.
BREED, R. S., MURRAY< E. 6. 0. y SMITh, N. T. (1957),En “Bergey’sManualof DeteminativeBacteriology”,PP. s~~-sgg~Ed, The Wihliams & Wilkins Co. Baltimore.
BRIONES, V. (1990), Estudio de un método indirectoparael diagnóstico de la listeriosis,Tesis Doctoral, Fac,Veterinaria,13CM.
BRIONES, V. (1993). Factoresdel hospedadorquedeterminanla susceptibilidada la listeriosis.En “Listeria enalimentos,ConferenciaConsenso”,PP. 95-100.Ed. M” Sanidady Consumo.León.
BRIONES, y., BLANCO, M. M., MARCO, A,, PRATS,N., EERNANDEZ-GARAYZABAL, 1. E., SUAREZ,6.,DOMINGO, M. y DOMíNGUEZ,L. (1992).Biliary exeretionas possibleorigin of Listeriamonocytogenesiii fecalcarricrs. Am. 1. Vet. Res. 53, 191-193.
BROSCH.R., CATIMEL, B., BUCHRIESER,C. y ROCOURT,1. (1992).Viruleneeevaluadonof L. monocytogeness¡rains, isolatedfrom humans,foodandanimais in immunocompetentmice,Proc.XIth InternationalSymposiuinonLhe Prohlemsof Lísteriosis(Copenhagen>95-96.
BRIJNT, L. M., PORTNOY,D. A. y UNANNE, E. R. (1990). Presentationof Listerialnonoeytogenestu CD8+ Tcelis requiressecretionof hemolysinanó intracellularbacterial growth. J. linmunol. 145, 3540-3546.
BRYNER, J., WIlEY, 1. y VAN DER MAMEN, M. (1989>. Researchof listeriosis in rnilk cows willxintramammaryinoculation of Listeriamonocyto~enes.Acta Microbiol. Hung. 36< 137.140.
BURN, C. 6. (1935). Characteristiesof a new speciesof dic genusListerella obtainedfroin human sources.1.Bacteriol. 30, 373-
BURN, C. G. (1936). Chinical asid paihological featuresof an infection causedby a new pathogenof the genusListerella. Am. i, Pathol. 12, 341-348.
CAIN, D, B. y MCCANN, V. L. (1986). An unusualcaseof cutaneoushisteriosis.J. Clin. Microbiol. 23, 916-977.
JzJ4
II

Biblk¡gr$fi¡’.
CARVAJAL, A., SAMUELSSON,5,, ROTHGARDT<N. 1>. yFREDERIKSEN,W. (1989).TheIrcatmncurof Listeriamonocytogenesinfectionsin Che CentralNervousSystem.Acta Microbiol. Hung. 36. 159-164.
CHANDRA, R. K. (1985>.Effeet cfmacro-ami micronutrientdeficienciesami excesseseninnunneresponse.FeodTechnol.Fcb,, 91-93.
CHANDRA, R. K. (1989>. Nutritional regulationof immunity and risk of illness. Indian J. Pediatr.56, 607-611.
CHARLTON, K. M. y GARCíA, M. M. (1977). Spontaneouslisteric encephalitisand neuritis iii shecp.Lightmicroscopicstudies.Vet. Pathol. 14, 297-313,
CHASTELLIER, C. 14. y BERCHE, 1’. (1994).Pateof Listeria monocvto2enesin murine macrophages:evidencefor simultaneouskilling and surviva] of intracellularbacteria.Infect. Immun. 62, 543-553.
CHEERS,C., MCKENZIE, 1. E. C., PAVLOV, H., WAID, P. y YORK, J. (1978). Resistanceami suscep¡ibilityofmice to bacterialinfection: courseof listeriosisin resistantor susceptiblemice. Infect. Irnmun. 19, 763-770.
CHEERS,C. y STANLEY, E. R. (1988). Macropliageproductionduring murine listeriosis: Colony-StimutatingFactor 1 (CSF-l) and CSF-I binding celís in genetically resistaintand susceptiblemice. Infect. Irnmun. 56<2972-2978.
CLEGO,E. 6. (1975).Listerieinfectionand thegenitalsystem.Proc.VItb InternationalSymposiumen the Problemsof Listeriesis, (Nottingham)145-155.
CLUFE, C. N., GARCíA, M. y ZIEGLER, H. K. (1990). Intracellularbemolysin-producingListeriarnonocvto2enesstrains inhibit macrophage-mediatedantigenprocessing.Infect. imniun. 58, 3601-3612
COLNAGO, 6. L., JENNSEN,L. 5. y LONG, P. L. (1984a).Effect of seleniumon periphemlhicod lcucocytesofchikensinfeeted with Eimcria. Poultry Sci. 63, 896-903.
COLNAGO, 6. L., JENNSEN<L. 5. y LONG, P. L. (1984b).Effect of seleniumandvitamin E en Che developmentof inimunity te coccidiosisin chikcns. Poultry Sci. 63, 1136-1143.
CONLAN, J. W. y NORTH, R. J. (1991).Neutrophil-mediateddissoíutionof infectedhos¡celís asadefensestrategyagainsta facultative intracellularbacteriuro.J. Exp. Mcd. 174, 741-744.
CONLAN, J. W. y NORTH, R. J. (1994).Neutrophilsare essentialfor earlyanti-Listeriadefensein the liver. butnot in dic spleenor peritonealcavity, as revealedby a granulocyte-depletingmonoclenalantibody.J. Exp. Mcd. 179,259-268.
CONLAN, W. y NORTE,R. (1993).Neutrophil-mediatedlysis of infectedhepatocyíes.MM News 59. 563-567.
COOPER,R. E., DENNIS, 5. M. y HARRIS, J. 0(1973).Fractionatienof Listeriamonocytogcnesserotype5. Am.1. Vet. Res. 34, 1093-1099.
COSSART, P. y MENGAUD, J. (1989). Listeria rnonocytogenesa niodel systemfor the molecullar study ofintracellular parasnxsmMcl Exol Mcd. 6, 463-424
COX, L. i. (1989). Preventionaudcontrol of Listeriain dic foed industry.En “Feotibornelisleriosis”, PP. 100-120.Ed. Behr’s Verlag GmbH & Co. RFA.
CIJMMINS, C. 5. y HARRIS, E. (1956).The chemicalcompositionof dic ccli wall in sornegrarn-pcsilivebacteria
145

Bibliografía
asid jis pessiblevalueasa (axonemiecharacter.J. Gen. Microbiol. 14, 583-600.
CZUPRYNSKI, C. J. (1994). Host defenseagainstListeria monecytoaenes:implications for foed safety. FeodMicrobiol. II, 131-147.
CZIJPRYNSKI, C. J., CANONO, E. P., HENSON, E M. y CAMPBELL, P. A. (1985). Genetically deterniinedresiscancete listeriosisis asseciatedwith increasedaccumulaúenof inflamatoryneutrophylsaudmacrophageswhichhaveenhancedlistericidal activity. lmmunol, 55, 511-518.
DECKER, C. F., SIMON, O. L., DIGIOLA, R. A. y TUAZON, C. U. (1991).Listeria rnonocytogenesinfections iii
patienis with AIDS: reportof Vive casesami review. Rey. Infecí. Dis. 13(3), 413-417.
DEHAUMONT, PO (1993).Listeriamenecytogenesel alimentationenFrance.En Listeriaen alimentos.ConferenciaConsenso,PP. 39-53. Ed. M0 Sanidady Consumo.León.
DESCHRYVER-KECSKEMETI,K.; BANCROFT,G. it; BOSMA, O. C.;BOSMA, M. J. y UNANUE, E. R. (1988).Pathologyof Listeriainfectionin murineseverecembinedimmunodefmcieney:a studyby immunohistochemistryandelectrenmicroscopy.Lab. Invesí. 58, 698-705.
DIJKSTRA, R. O (1987).Listeriosisiii animals:clinical sigus,diagnosisandtreatmení.En Listeriosis. Reunióndela OMS, Veí. Med. Hefte 5/1987,
DIJKSTRA, R. G. (1981).The ocurrenteof L. monocytegenesin surfacewater of canaisand lakes, in ditchesofenelarge polderaud in dic effluentsandcanalsof sewagetrealmeníplant. Proc. VIIIth International Symposiumen te Problernsof Lisíeriosis (Madrid>.
DOMINGO, M., MANCEBO, J., BLANCH, L., COLL, P., NET, A. y NOLLA, J. (1986).Meningitis por Listeriamonecyíogenesen adultos.Estudiode 10 casos.Rey. Esp. Microbiol. Clin. 178, 158-163.
DOMINGO, M., MARCO, A,, PRATS,N., ALTIMIRA, 1., BRIONES, V., BLANCO. A. y DOMINGUEZ, L.(1992). Intragastrieinoculationof Listeriamonocytogenesin Che meuse.Proc.Xfth IntemationajSyrnposiu¡nen dicProblemsof Listeriosis(Copenhagen)247-248.
DOMINGO, M., MARCO, A, J., PRATS, N. y ALTIMIRA, .1. (1993). Patogeniay patologíacomparadade lalisteriosis.En Lisíeria en alimentes.ConferenciaConsenso,PP. 157-162.Ed. M0 Sanidady Consumo.León.
DOMíNGUEZ, L. (1982>.Aislamientoe identificaciónbacterianaenel géneroLisíeria(Pirie 1940).TesisDoctoral,Fac. Veterinaria,U.C.M.
DROKE. E. A. y LOERCH, S. C. (1989).Effects of parenteralseleniumasid vitainin E en perfonnance.hcalrhandhumoralimmuncresponseof steersnew te te feedlotenvironmení.J. Anim. Sel. 67, 1350-1359.
DUMONT. J. y COTONI, L. (1921). Bacille semblableá celui du reugeldo pote rencontrédaus le LCR. dunméningitiquc.Ann. Insí. Pasí.35, 625-633.
EMILE, 1. y BAZIN, C. (1975).Listeria neuromeningitisin Che adulÉ. Proc. VIth InternationalSympcsiuínen theProblemsof Listeriesis(Nottingharn)pp. 164-172.
ERREBO-LARSEN,1-1. y JENSEN,1. (1979>.Ocurrenceof hovine mastistiscausedby L. monoc’vte2enes.Proc.VIItli InternationalSymposiumen te Preblcmsof Listeriosis(Sofia), pp. 211-219.
ERSKINE, R. 1., EBERHART, R. 1., GRASSO, P. i. y SCI-IOLZ, R. W. (1989). Induction of Eseherichiaccli
146

Bibliografía
mastitis in cows fed selenium-deficientor selenium-supplememeddiesis. Am. 1. Vet. Res. 50, 2093-2100.
ERSKINE, R. J., EBERHART, Ii. J., HUTCIIINSON, L, J. y SCHOLZ, R. W. (1987). Bleod seleniumconcentradonsand glutadiioneperoxidaseactivities in dairy herdswith high and10w somaticcelí ceunts.JAVMA190, 1417-1421.
ERSKINE, R. J., EBERHART, R. J. y SCHOLZ, R. W. (1990). ExperimentallyinducedStanhvlococcusaureusmastitis in selenium-deficientaud selenium-supplementeddairy cows. Am. J. Vet. Res. 51, 1107-1111.
ESKEW, M. L., ZARKOWER, A., SCHEUCHENZUBER, W. J., BURGESS, J. R., SCHOLZ, R. W.,HILDENBRANDT, 6. y REDDY, C. C. (1989).Effects of inadequatevitamin E and/erseleniumnutritionen sihereleaseof arachidonieacid metabolitesin rat alveolarmnacrophages.Prostaglandins38, 79-89.
ESPAZE,E. P., GAIJTREAU, D., CATIMEL, B., MIEGEVILLE, A. E., ROCOURT,J. y CORTWU,A. L. (1989).An epidemiologica]survey of humanlisteñosisiii Franceduring 1987 using serotyping amI phage-typing.ActaMicrobiol. IIung. 36, 231-234.
FARBER, J. M. y PETERKIN< P, 1. (1991). Listeriamenocvtegenesa foed-bemepaChegen.Micrebiel. Rey. 55,476-511.
FENLON. D. R. (1985). Wild birds and silageas reserveirsof Listeria in Che agricultura] environment.J. ApplBacteriol. 59, 537-543.
FERNANDEZ-GARAYZABAL, J. F. (1984). Microorganismosdel géneroListeria (Pirie 1940) en leche naturalypasterizada.TesisDoctoral.Fac. Veterinaria,U.C.M.
FERNANDEZ-GARAYZABAI, J. F., DOMINGUL?, L., VAZQUEZ,J, A., BLANCO, J. L. y SUAREZ,6. (1986).Listeriamonecvtogenesdansle lail pasteurisé.Can.1, Micrebiol. 32, 149.
FLEMINO, D. W., COCHI, 8. L., MACDONALD, K. L., BRONDUM, 1., RAYES, P. 5., PLIKAYTIS, B. O.,HOLMES, M. O,, AUDURIER, A., BROOME, C. y. y REINGOLO, A. L. (1985). Pasteurizedmilk as a vehicleof infectien iii an eutbreakof listeriosis.New Eng. J. Med. 312, 404.
GAILLARO, J. L., BERCHE, P., MOUNIER, J., RICHARD, 5. y SANSONETTI, P, (1987a). In vitre model ofpenetrationasidintracellulargrowth of Listeriamonocytogenesin the humanenterocyte-likecelí line Caeo-2.Infecí.Immun. 55, 2822-2829.
GAILLARD, j. L., BERCHE,P., MOUNIER, J., RICHARD, 8. y SANSONETTI, P. (1987b).Penetrationof Listeriamonocvtogenesinto Che bosea crocial stepof die infectiousprocess.Ann. Inst. Pasí. Microbiel. 138. 259-264.
GALSWORTHY, 5. B. (1987). Role of the eclI surfacein virulence of Listeria menocytegenes.Ann. InsÉ. Pas!.Microbiol. 138, 241.284.
GARCíA, 1. A., DOMINGTJEZ, L., BRIONES, y., BLANCO, M.. FERNANDEZ-GARAYZABAL, J. E. ySUAREZ. G. (1990). Revisionof Che ntigenic stn~ctureof genusListeria. FEMS Micrebiol. Let. 67, 113-120.
GILL, D. A. (1933).Circling disease:a meningeenceplialitisof sheepin New Zealand.Notes en a new specicsofpatbogenicorganisrn.Austral. Vet. J. 89, 258-270.
GILL, O. A. (1937). Ovinebacterialencephalitis(Circling disease)anódic bacteria]genusListerella. Austral. Vel.3. 13, 46-56.
GITTFR, M. (1979).Listeriamenecvte~enesinfectionin bovine aborden.Proc. VIlth lnternatioiíalSymposiunien
147

Bibliografía
Che Problemsof Listeriosis (Sofia) pp. 193-204.
GITTER, M. (1985).Listeriosisin farmn animalsin GreatBritain. En <‘Isolation and identification of miaoorganismsof medical andveterinary importante” (eds. Collins, C. E. y Grange,LI. M.), Pp. 191-200. Ed. AcadeiniePress.Londres.
GITIER, M.< STEBBINGS< R. Sil., MORRIS,1. A., I-IANNAM, D. y HARRIS, C. (1986y Relantioshipbetweenovine listeriosis andsilage feeding. Vet. Rec. 118, 207-208.
GOLNAZARIAN, C. A., DONNELLY, C. W., PINTAURO, 5. J. y HOWARD, D. B. (1989). Comparisonofinfectiousdoseof Listeria monocytogenesF5817as determinantfor normal versuscompromisedCS7Bl/6j mice.1. FeodProt. 52, 696-701.
GOOSENS,P. L., MARCHAL, G. y MILON, 6. (1988).Early influx of Listeria-reactiveT lympbocytesin liverof mice geneticallyresistantte listeriosis,J. Inununol. 141, 2451-2455.
GOULET, y., ESPAZE, E., BASTIDE, 1. y REVIERE, 1. (1989). Human listeriosis in France in 1987. ActaMicrobiol. Hung.36, 173-176.
GOMEZ MAMPASO, E, (1993). Listeriesis humana:grupos de riesgo y portadores.En “Listeria en alimentos.ConferenciaConsenso’,PP. 141-144.114. M” Sanidady Consume.León.
GOMEZ MAMPASO, E., TERUEL, J. L., CASCALES, P., GARCíA MARTIN, F., BAQLJERO, F. y ORTUÑO,J. (1987).Portadoresfecalesde Listeria y transplanterenal.Rey. Esp. Microbiol. Clin. 2, 99-101.
GRAY, M. L. y KILLINGER, A. U. (1966). L. monocvtogenesandlisterie infections.Bacteriol. Rey. 30. 309-373.
GREENSPAN, H. C. y ARUOMA, 0. 1. (1994). Oxidative sterss andapoptosisin HIV infecrion: a role for
plant-derivedmetabeliteswidi synergisticantioxidant activity. Immunol. Today 15, 209-213.CTROVES,R. O. y WELSI-IIMER, 1-1. 1. (1977>.Separationof padiogenicfrom apathogenicListeriamenocvtogenesby diree in vitre reactions..1. Clin. Microbiol. 5, 559-563.
GYANG, E. O., STEVENS, 1? B., OLSON, W. G., TSITSAMIS, 5. D. y USENIK, E. A. (1984). Effects ofseleniuxn-vilaminE injection en bovinepolyrnorphonucleatedleukocytesphagocytosisandkilling of Ssiaphylococcusaureus.Am. j. Vet. Res.45, 175-177.
HANG< C. E., GILL, R. G., BABCOCK, 5. K., LAFFERTY, K. J., BELLGRAU< 1). y WEIL, R. (1987).Cyclosporine-inducedtolerance requires aníigenscapableof initiating an immune response.J. Immunol. 139,2947-2949.
HARRINGTON-FOWLER, L., HENSON. P. M. y WWDER, M. 5. (1981). Fate of Listeria monocvto~enesinresidentandactivatedmacropliages.Infect. linmun. 33, 11-16.
HASSAN, 5., EAKKARA]NEN, J., JONSON,L. y TYOPPONEN,J. (1990). Histopathologicaland biochemicalehangesassociaedwith seleniumaud vitaminE defíciencyin chieks. LI. Vet. Mcd. 37, 708-720.
IIIGGINBOTHAM, J. N., LIN, T. L. y PRUETT,5. B, (1992). Effcct of macrophageactivationenkilling of Listeriamonocvtogenes.Rolesof reactiveoxygenor nitrogeninterniediates,ratesof phagocytosis.aud relentionof bacteriain eridosomes.Clin. Exp. linmunol. 88, 492-498.
HILL, C. II. (1975). Interrelationshipsof selenim witb other trace elcments.Fed. Proc. 34, 2096-2100.
148

Bibliografía
l-iLRD, O. W. (1987>. Reviewof evidenceter zeonotielisteriosis.3. of FoedProt. 50, 429-433.
HIRO, D. W. y GENIGEORGIS,C. (1990).Listeriosisin foedanimals:clinical signsaixd livestock as a potentialseurceof direct (nenfoodborne)infection ter humans.En ‘<FoodbomeListeriosis” (eds.Miller, A. J <Smith, J. L,y Somkusii, G. A.), pp. 31-39.lid. Elsevier. Amsterdam.
HO, J. L., SHANDS, K. N., FRIEDLAND, O., ECKIND, P. y FRASER,D. W. (1986). An outbreakof 4b-íypeLmonocytegenesinfection involving patiensisfroin cigiit Bostonhespitals.Ascii. Intern. Mcd. 146, 520.
HOF, E. y HEFNER, P. (1988). Pathogenicityof Listeria monecvto~enesin comparisonte odierListeria species.Infect. Suppl.2, 141-144.
HÚLPHERS,0. (1911).Lefvernekrosheskanin ersakadal en ej fórnt beskrifvenbakterie.Sven.Vot. Tidskrift. 2,265-273.
HYSLOP,N. 5. 0. (1975).Epidemielogieaudirnmunelegicfactorsin listeriosis.Proc.VICh InternationalSymposiumen die Problemsof Listeriesis ~Nottingham).
IVANOV, 1. (1957>.Listériosechezles ovins el les eaprins.Buil. Oíl. mí. Epiz. 48, 511-583.
IVANOV. 1. (1962). Untersuchungenaher dic ListeriosederSehafeinBulgarien. Monaths. Veterinaenned.17,729-736.
IVANOV. 1. (1975). Establishmentof non-motile swains of Listeria monecvto~enestype 5. En “Preblemsoflisteriesis” (ed. Woedbine,M.), PP. 18-26.LeicesterUniversity Press
JAMES, 5. M., FANNIN, 5. L., AGEE, B. A., GALL, B., PARKER,E., VOGT, LI., RUN, O., WILLIAMS, J., LIEB,J.,PRISDERGAST,T., WERNER,5. E. y CHIN, J. (1985).Listeriosisoutbreakasseciatedwith Mexican-style-cheeseiii California. Morbid. Mortal. Weekly Rept. 34, 357.
JELINEK, P. D., ELLIS, T., WROTH, R. E.. SUTHERLAND, 5. 5., MASTERS. H. O. y PETTERSON,D. 5.(1988).Theeffectof seleniumsupplementationen ixnmunity, andsitie establishmentof an experimentalHaemonchuscontortusinfection, in weanerMerino sheepfed a low seleniumdiet. Austral. Vet. 1. 65. 214-217.
JONES,D. (1975).A numericalsiaxonomic studyof coryneformandrelatedbacteria.1. Gen. Microbiol. 87, 52-96.
JONES, D. (1988). Re placeof Listeria amonggrain-positivebacteria.Infect, Suppl.2, 85-88.
JONES, O., COLLINS, M. O., GOODFELLOW, M. y MINNIKIN, D. E. (1979). Chemical studies in dicclassifieasiionof Che genusLisíeria ami pxobably related bacteria. Proc. VIlth International Symposiumen CheProblemsof Listeriosis(Sofia) PP. 17-24.
JONES,O., FERESIJ,5. E. y COLLINS, M. 0. (1986). Classificationandidentification of Listeria, Brochodirix audErysinelothrix, En Listeria-Listeriosis” (eds.Courtieu, A. L., Espaze,E. P. y Reynaud,A. 119, PP. 29-34. Ed.Universidadde Nantes.Nantes.
JITRADO, R. L., FARLEY, M. M., PEREIRA, E., HARVEY, R. C., SCHUCHAL A.. WENGER, j. ~. ySTEPHENS,D. 5. (1993). Increasedrisk of meningitisandbacteremiadue te Listeria monocytogenesin patieníswitt humanimmunodefxciencyviros. Clin. Infect. 1)is. 17, 224-227.
KAMPELMACHER, E. H. y JANSEN, N. (1975). Ocurrenceof L. monecvtoeenesin effluents. Proc. VIChInternationalSymposiumen Problemsof Listeriosis (Nottingham).
149

Bib~iografís
KAMPELMACI-IER, E. E. y NOORLE JANSEN, E. M, (1969). Iselation of L. menocytogenesfroin fecesofctinicalty healtbyhumausandanimais.Zbl. Bát. 1 Att Orig. 221, 353-359.
KANDLER, O. y WEISS,N.(1986).Regular,nonsporinggram-positiverods.En “Bergey’sManualof DeterminativeBacsieriology” (eds. Sueath,P. U. A., Muir, N. 8., Sharpe,M. E. y Holt, 1. Ci.), Pp. 1208-1209.Ed. The Wittiams& Wilkins Co. Baltimnere.
KAUTTER, O. A,, SILVERMAN, 5. 5., ROESSLER,W. G. y DRAWDY, J. F. (1963). Virulence of L
.
rnonocytogenesfor experimentalaniimals.5. of Infect. Dis. 112< 167-180.
KIREMIDJIAN-SCHUMACHER, L., ROY, M., WISHE, E. 1., COHEN, M. W. y STOTZKY, G. (1990). Seleniumand immnuneccli functions.I.Effect en lymphocyte proliferationasid productionof interleukin 1 and interleukin 2.P.S,E.B.M.193, 136-142.
KIREMIDJIAN-SCHUMACHER,E., ROY, M,,WISHE< 11.1.,COHEN,M. W. y STOTZKY, Ci. (1992).Regulationof cellular immuneresponsesby seleuium.Biot. TraceEtemeutsRes. 33,23-35.
KOLLER, L. D. y EXON. J. H. (1986).TIte wo facesof selenium-defxciencyand texieity- aresimilar iii animaisami mart. Can. 1. Vet. Res. 50, 297-306.
KONGSHAVEN, P. A. L. y SKAMENE, E. (1984).Tiie roleof naturalresistancein protectionof Che murine hostfrom listeriosis. Clin. Invest. Med. 7, 253.
KOS, W. L., KOS, K. A. y KAPLAN, A. M. (1984). Impaired function of inimune reactivity te Listeriainonecytegenesin diet-fed mice. Infect. Immun. 43, 1094-1096.
KRATZ, 5. 5. y KIJRLANDER, R. J. (1988).Characterizationof Che pasiternof infkunatoryeclI influx andcyíokineproduction during tIte murine iiest responsete Listeria monocytegenes.J, Immunol. 141, 598-606.
KUHN, M., KATHARIOU, 5. y GOELLIEL. W. (1988).Hernolysin supportssurvivalhutnot entryof tIte intsacellularbacteriumListeria monocvto~enes.Infect. Immun, 56, 79-82.
LALLEMANO, A. y., GAILLARO, D. A,, PARADIS, P. E. y CHIPPAUX, C. G. (1992).Fetal listeriosisduringChe secondtrimesterof gestation.Pediatr.PaChel.12, 665-671.
LARSEN, 5. A. y SONES,W. (1972).Evaluationandstandardizationof un agglutinationtestterhuman listeriosis.Appl. Microbiol. 24, 101-107.
LARSON,8. (1979).Cíinicaldifferencesin neenatallisteriosisacquiredin uteroversuseflramaternally.Proc.VllthInternationalSympesiumen tIte Problemsof Listeriosis (Sofia) PP. 170.172.
LEE,W. IR. y MCLAIN, 0. (1986). ImprovedlIsteriamonoeyyogenesselectiveagar.Appl. Environ.Microhiol. 52.1215-1217.
LENNON, D., LEWIS, B., MANTELE, C., BECROFT, 0., DOVE, E., FARMER, K.. TONKIN, 5., YEATS. N..STAMP, R. y MICKELSON, K. (1984). Epidemicperinatallisteriosis.Pediat. Infeo. Dis. 3, 30.
LOESSNEROM. j. y BIJSSE,M. (1990). Bacteriophagetyping of Listeria species.Appl Enviren. Microbiol. 56,1912-1918.
LOPEZ, M. B.; BRIONES, y.; FERNANDEZ~GARAYZABAL, J. F.; BLANCO. M. M.; GARCíA, J. A.;DELGADO.M. C.; DOMINGO, M. y DOMIIÑGUEZ, L. (1992). Serologicalresponsein rabbits te oral inoculationwitii Listeria monocytogenes.Proc. XIth InternationalSymposiumen tIte Problenmsof Listeriosis (Copenhagen)
150

bibLiografía
LOPEZ, M. B,; BRIONES,V.; FERNANDEZ-GARAYZABAL, J. E.; VAZQUEZ-BOLAND, J. A.; GARCíA, LI.A.; BLANCO, M. M.; SUAREZ, G. y DOMINGUEZ, L. (1993). Serelegical responsein rabbits te Listeriamonocvtogenesafter oral er intragasuleineculation.FEMS Immunol. Med. Microbiel. 7, 131.134.
LORBER B (1990>. Clin. listeriosis- implicationsfor padiogenesis.En ‘Feedbornelisteriosis” (eds.Miller, A. J.,Smidi, 5. L. y Semkuti,Ci. A.), pp. 41-49. Ed. ElsevierAmsterdam.
LOVETY, 1. (1988). Isolatienand identificatienof Listeriamonocvtogenesin dairyproducis.5. Assoc.0ff. Anal.Ciiem. 71, 658-660.
LOVETT, 5. (1989).Listeria monocvto2enes.En “Feedbernebacterialpathegens”(cd. Doyle, M. 1’.). Ed. MarcelDekker Inc. NuevaYork.
LOW, J. C. y DONACHIE, W. (1991).Clinical andserumantibody responsesof lambs te infection by Listeriamonocysiogenes.Res. Vet. Sci. 51, 185-192.
LOWRY, P. D. y TIONG, 1. (1988).The incidenceof Listeriamonocvtogenesin meatandmeatproducís: factorsaffeetingdistrubinien,Proc. 34xb InternationalCongressof Meat SejenceandTechnology,Brisbane, Australia.
LUFT, B. 1. y REMIINGTON, 5. 5. (1982). Effect of pregnancyen resistancete Listeria menoeviogenesandTexoplasmagondii infectionsin mice. Infect. Iimmun. 38, 1164-1171
MACDONALD, T. T. y CARTER,P. E. (1980).Celí-mediatedimmunity te intestinalinfection. Infect. linmun. 28,5 16-523.
MAINOU-FOWLER, T., MACGOWAN, A. P. y POSTLEHWAITE, R. (1988). Virulence of Lisíeria spp.:CeuNeof infection in resistantandsusceptiblemice. J. Mcd.Microbiel. 27, 131-140.
MANDEL, T. E. y CHEERS,C. (1980).Resistanceandsusceptibility of mice te bacterialinfectien:histopathologyof listeriosis in resissiantandsusceptiblestrains.Infect. Immun. 30, 851-861.
MARCO, A., DOMINGO, M.. PRATS, N., BRIONES, y., PUMAROLA, M. y DOMíNGUEZ, L. (1991).Pathogcnesisof lympiioid lesionsin murine experimentallisteriosis.J. Comp. PaChol. 105, 1-15.
MARCO, A., DOMINGO, M., RUBERTE, J., CARRETERO,A., BRIONES, V. y DOMíNGUEZ, L. (1992a).LymphaticdrainageofLisíeria monocyto~enesandindian ink inoculatedin Che peritonealcavity of tIte meuse.Lab.Anim. 26. 200-205.
MARCO, A., PRATS,N., RAMOS, II. A.. BRIONES, V., BLANCO, M., DOMíNGUEZ, L. y DOMINGO, M.(1992b).A inicrobiological,iiistepadiologicalandimnmunohisíolegicalstudyof Che intragastrieinoculationof Lisíeriamonocytogenesin mice. 1. Comp. Padiol. 107, 1-9.
MARTE, E. U. (1988a).Diseasecharactetisriesof L. monocytonenes.FoedTeciinol. April. 165-168.
MARTH, E. H. (1988b).Ocurrenceof Listeriain foods: milk anddairyproducts.Conferenciaexpuestaen el SIM.Red Lien Inn, California, USA. 2-5 Octubre.
MCLAUCHLIN, J. (1987).Listeriamonecytebenes,recentadvancesin dic taxonomyandepidexniologyof listeriosisin humans.J. Appl. Bacteriol. 63, 1-11.
MCLAUCHLIN, 1. (1990).Distributionof serovarsof Listeriamonecvie2encsiselatedfrom diffcrent categoriesofpaticntsw¡Ch listeriesis.Eurep. 3. Comp.Microbiol. Infect. Dis. 9, 210-213.
151

II,
Bibliografía
MERO, E. y STEUER, W. (1981).Ocurrenceof L. monocyto~enesin effluent of sewagetreatmentplantsand inreceiving body river water. Proc. VIIIth InternationalSymposiumen Problemsof Listeriosis (Madrid>.
MEYER, 1). 11., BUNDUKI, M., BELIVEAN, C. M. y DONNELLY, C. W. (1992). Differencesin invasionamiadherenceof Listeriamonocytogeneswidi mammaliangut celís. FoedMicrobiol. 9, 115-126.
MIELKE, M. E. A., EHLERS,S.y IIAUN, 11. (1993). Re roleof cytoklixes in experimentallisteriosis.Linmunobiol.189, 285-315.
MIKI, K. y MACKANESS, G. B. (1964). TIte passivetransferof acquircdresistancete Listeriamonocysio2enes.J.Exp. Med. 120, 93-103.
MILLER, E. R. (1985). Mineral x diseaseinteractions.1. Anim. Sci. 60, 1500-1507.
MILLER, J. K. y BURNS, J. (1970). Histopathologyof Listeriamonocytogenesafteroral feeding te mice. Appl.Microbiol. 19, 772-775.
MURRAY, E. Ci. D., WEBB, R. A. y SWANN, M. B. R (1926). A diseaseof rabbits characterizedby a largemononuclearleucocytosiscausedby Ititiierto undescribedbacillus Bacteriummonoeytoízenes.J, PatItel. Bacteriol.29, 407-439.
MURRAY, J. M. y MURRAY< A. B. (1985). Re effects of seleniumdeficiencyand repletion en bosÉ resistancelo infection. En “Trace elementsin man and animals”, Pp. 244-247.Ed. CommonwealChAgricultural Bureau.Cambridge.
NAKANE, A., MINAGAWA, T. y KATO, K. (1988a>.Endogeneustumor necrosisfactor (Cachectin)is essentialte host resistanceagainsíListeria monocysioQenesinfection. Infect. Immun. 56, 2563-2569.
NAKANE, A., MINAGAWA, T., YASULIA, 1., YU, C. y KATO, K. (19881,). Preventionby gammainterferenoffatal infection witii Listeria monocvto2encsin mice treatedwith cyclosporinA. Infect. lmmun. 56, 2011.2015.
NAKANE, A., MINAGAWA, T., KOHANAWA, M., CHEN, Y., SATO. H., MORIYAMA. M. y TSURUOKA, N.(1989). Interactionsbetweenendogeneusgammainterferon and tumor necrosisfactor in host resistanceagainsíprimary andsecendaryListeria monocytogenesinfections.Infect. Inimun. 57, 3331-3337.
NOTERMANS, 5. y CHAKRABORTY, T. (1992). Pasibogenicityof Listeria monocvto2enes:Protectionacquiredhy infection. Proc. Xliii InternationalSymposiumen tIte Problemsof Listeriesis (Copenhagen)78-79.
NYFELIYr, A. (1932>. Klinishe und experimentelleuntersuchungenUber die Mononucleosisinfectiosa. FoliaHaematol.Leipzig 47. 1-44.
ORTEL, 5. (1989a).Listeria-meningitisand -septicaemiain immunocompromisedpatients.Acta Microbiol. 1-Jung.36, 153-158.
ORTEL, 5. (1989b). Furrher results aud expenenceswith phage-iypingof Listeria. Acta Microbiel. 1-Jung. 36,2 19-224.
(JTTER, A. y BLAKEMORE. W. F. (1989).Observationsen tiie preseiiccof Listeriarnonocytouenesin axeus.ActaMicrebiol. líung. 36, 125-132.
PIRIE, J. 11. H. (1927). A new diseaseof veldt rodents.<‘Tiger River Disease”.Pubí. 5. African Insí. Mcd. Res. 3,163-186.
152

Bibliografía
PIRIE, J. II. H. (1940). Listeria: changeof naine for a genusof bacteria.Nature 145, 264.
POHJANVIRTA, R. y HUTTUNEN, T. (1985).Someaspectsof murineexperimentallisteriosis.Acta Vet. Scand.26, 563-580.
PORTNOY, D. A. (1992). Molecular determinansisof Listeria monocytogenespasitogenesis.Infect. Inunun. 60,1263-1267.
PRATS,N., BRIONES, y., BLANCO, M. M., ALTIMIRA, J., RAMOS, J. A., DOMINGUEZ, L. y MARCO, A.(1992a).Choroiditis andmeningitisin experimentalmurine infection witii Listeriamenocytoueues.Europ.J. Comp.Microbiol. Infecí. Dis. 11, 5-8.
PRATS, N., BRIONES,V., MARCO, A., DOMINGO,M., DOMINGUEZ, L., BLANCO, M. M. y ALTIMIRA, J.(1992b). Choreiditisandmeningitisare late tesleusin experimentalmurine infectionwiíb Listeriamenoeytogenes
.
Proc.XIth InternationalSymposiumen tIte Problemsof Listeriosis (Copenhagen)253-254.
PUNG, O. J., TUCKER, A. N.. VORE, 5. J. y LUSTER, M. (1985). Influence of estrogenen hostresistance:increasedsusceptibility of mice te Listeria monoevtonenescorrelatesválh depressedproductienof interleukin-2.Infect. Imniun. 50, 91-96.
RALOVICH, B. (1975). Seleetiveand enrichmentmedia te iselate Listeria. Lii “Preblemsof listeriesis’, ed.WOODBINE, M., Pp. 286-294.LeicesterIlniversity Press.
RALOVICH, B. (1984a).Listeriosis research.Presentsituatienandperspective.AkademiaiKiado, Budapest.
RALO VICH, B. (1984b).Wiiat is Cherole of food-stuffsin transportationof Listeria, En ‘Microbial associationnandinteractionsiii foed’, Pp. 99. Ed. AkadcmiaiKiado. Budapest.
REI-TERT STABEL, .1., SPEARS,J. W., BROWN, T. T. y BRAKE, 5. (1989). Scleniumeffects en glutathioncperexidaseandChe irnmuneresponseof stressedcalvesciiallengedwitb Pasteurellaheniolvtica,J. Anim. Sci. 67,557-564.
REISS, 1-1. LI.; POTEL, J. y KREBS, A. (1951). Granulomatosisinfantiseptica.Z. Gesantz.lun. Med. 6,1-7.
RIBIERE. O., COUTAREL, P., JARLIER, V., BOUSQUET,O., BALDERACCEI, U., LECOIJTURIER,J. 1’. yí’HERVET, F. (1990). Abeesdu foie a Listeria monocytoízenes.Chez une maladediabetique. PresseMed. 19<1538-1540.
RIDGWAY, E. .1. y BROWN, 1. M. (1989>.Listeriamonocyto~enesmeningitis1w tIte acquiredimmunodet0icieneysyndrome:iimitatiens of conventionaltyping methodsin tracinga foodborneseurce.LI. Infect 19, 167-171.
ROCOURT, LI. (1988). Taxonomyof die genusListeria. Infect. Suppl.2 16, 89-91.
ROCOURT, J., GRIMONT, F., GRIMONT, P. A. D. y SEELIGER,H. P. R. (1982). [)NA relatednessamoneserovaisof Listeriamonocvsiogenessensulato. Curr. Microbiol. 7, 383-388.
ROCOURT, LI. y SEELIGER, U. P. R. (1985). La listériose: une infectien hospitaliére?Méd. Mal. Infect. 12,72h725.
ROCOtJRT,LI. WEI-IMEYER, V. y STACKEBRANDT, E. (1987).Transferof Listeriadenitrificansteanewgenuslenesiagen.nev.as LIonesiadenitrifxcanscomb.nov. Ini. J. Syst. Bacteriol. 37, 266-270.
153

BLbLiografía
ROCOURT, J. y CATIMEL, u. (1989). International PhageTyping Centerfer Listeria: report for 1987. ActaMicrobiol. Hung. 36, 225-230.
ROCOURT,5., I3OERLIN, P., GRIMONT, F., JACQUET,C. y PIFFARETTI, J. C. (1992).Assignrnentof Listeriagravi audlissieriamurravite a singlespoecies,Listeriatzrayi, with areviseddescriptionof LisItria 2rayi. lnt. LI. Syst.Bacteriol.42, 171-174.
ROLL, J. T. y CZUPRYNSKI, C. 1. (1990). Hemolysin is requiredfor extraintestinaldisse¡ninationof Listeriaínonocytogenesin intragastrieallyinoculatedmice. Infect. lmmun. 58, 3147-3150.
ROY, M., KIREMIDJL4N-SCHUMACHER,L., WISI-IE, H. 1., COHEN,M. W. y STOTZKY, Ci. (1992).Lifecí ofseleniumen te expressionof high affinliy interleukin 2 receptors.PSOE.BOM. 200. 36-43.
ROY, M., KIREMIDHAN-SCHIJMACHER,L,, WISHE, 14.1.,COItEN,M. W. y STOTZKY, Ci. (1993).Seteniumsupplementationenhanceste expressionnof interleukin 2 receptorsubunitsand intemalizationof interleukin 2.P.S.E.130M.202, 295-301.
SAUNDERS, N. A., RIDLEY, A. M. y TAYLOR, A. Ci. (1989). Typing of Listeria monocyÉo~enesforepidemiologicalstudiesusingDNA probes.ActaMicrebiol. Hung. 36, 205-209.
SCHLECH, W. F. (1990).Listerb’ animalsandman: aspectsof virulence. En ‘Foodbornelisteriosis” (eds. Miller,A. J., Smit, 1. L. y Somkuti, G. A.>. Ed. Elsevier.Amsterdam.
SCHLECH< W. F., LAVIGNE, P. M., BORTOLUSSI, R. A., ALLEN, A. C., HALDANE, E. y., HIGHTOWER,A. W., JOHNSON,5. E., KING, 5. U.,NICHOLLS, E. S.y BROOME, C. V. (1983).Epidemielisteriosis;evidenteof transmisionby foed. New Eng. .1. Med. 308, 203.
SCHLECH, W. F., CHASE, D. P. y BADLEY, A. (1986).A mt model of Listeria monocytotienesinfection via Cheoral reute: developmentandeffect of tIte gastric acidity en infectivedose.Proc. 26th Anual ASM Meeting,NewOrícaus.
SCHMIDT, Ni, y EMMONS, R.W. (1989).GeneralprincipIesof laboratorydiagnosticmethodsfer viral. rickettsialandchlamydialinfections.En ‘Diagnostie proceduresfor viral, rickettsialandchlamydialinfections” (eds. Setimidt.N.J. y Ennnons,R.W.), Ed. AmericanPublic Healtii Association,WashingtonDC.
SCHUCHAT< A., SWAMINATHAN, B. y BROOME, C. V. (1991). Epidemielogy of human listeriesis. Cliii.Microbiol. ltev. 4,169-183.
SCHULTZ, E. W.; TERRY, M. C.; BRICE, A. T. y GEBHARDT, L. P. (1934). Bacteriologicalobservatiensen acaseof meningo-encepliajitis.PSEBM 3,1021-1023.
SEELIGERR, H. P. R. (1952). Zur áetiologie der Cimnulomatosis infantiseptica und pseudetuherkulóser.Erkrankungen.Dtsch.Med. Wochenschr.77, 587-
SEELIGER.H. P. R. (1961).Listeriesis.Ed. HatberPub. Co. NuevaYork.
SEELIGER, IR. PO R. <1975). TIte contenof listeriosis.En ‘Problemsof listeriesis” (cd. Weedbine,M.). PP. 1-3.LeicestertJniversity Press.
SEEIIGER,1-1. P. R. (1976).Notion actuellesur l<epidemiolegiede la listériose.Med. Mal. Infect. 8-Qbis, 6-14.
SEELIGER, 1-1. P. R. (1981>. Apatliogenelisterien: Listeria innocuasp. nov. (SeeligercÉ Scoofs 1977).Zeniralbí.Bakteriol. Mikrobiol. Hyg. Abt.l Orig. ReiheA 249, 487493.
154

BibLiografía
SEELIGER, II. P. R. (1987). Classifxcationandpathogenicityof Listeria. In Listeriosis, Reuniónde la OMS, Vet.Med. Hefte 5/1987,
SEELIGER,H. P. R. y SULZBACHER, F. (1956). Antigenic relationshipsbetweenListeria menocvte~enesaudStanhyloceccusaureus.Can, J. Microbiol. 2, 220-231.
SEELIGER, 14. P. R. y WELSHIMER, 14. 5. (1974). Genus Listeria. En ‘Bergey’s Manual of DeterminativeBacteriology’ (eds. Buchanan,R. E. y Gibbons,N. E.), PP. 593-596.Ed. TIte Williams & Wilkius Co. Baltirnore.
SEELIGER,14. P. R. y HÓIRNE, K. (1979). Seotypingof L. monocvtogenesand related species.En ‘Methods inMicrobiology’ vol. 13, Pp. 31-49. lid. AcademiePress.Londres.
SEELIGER,14. P. R. y SCHOOFS,M. (1979). Serologicalanalysisof non-iiemolyzingListeria-strainbelongingtea speciedifferentfrom Listeriamonocvtogenes.Proc. VtIth InternationalSymposiumen sihe problemsof Listeriosis(Sofia) 24-28.
SEELIGER,H. P. R., ROCOURT, J., SCHRE’ITENBRUNNER, A., GRIMONT, P. A. D. y SONES,D. (1984).Listeria ivanovii sp. nov. mt. J. Syst. Bacteriol. 34, 336-337
SEELIGER, 14. P. R. y JONES, D. (1986). Genus Listeria Pirie 1940. En “Bergey’s Manual of SystematieBacteriology’ vel.2, PP. 1235-1245.Ed. Tiie Williams & Wilkins Co. Baltimore.
SERFASS,R. E. y GAUTHER, 14. E. (1975). Defectivemicrobicida] activity in glutathioneperoxidase-deficientneutropiiils of selenium-deficiensimts.Nature 255, 640-641.
SIDDIQUE, C. 14., MCKENZIE, B. E., SAPPI,W. J. y RICH, 1’. (1985). Ligiit nial electronmicroscopicstudy oftIte livers of pregnantmice infectedwitii Listeria monocvtogenes.Am. J. Vet. Res. 39, 887-892.SKOVGAARD, N. y MORGEN, C. A. (1988). Deteetionof Listeria spp. in faecesfroro animals, in feedsand inraw feodof animal origin. Int. J. FeodMicrobiol. 6, 229-242.
SORDILLO, L. M., HICKS, C. R., WIiLSON, R. y MADDOXO 5. (1993). Effects of seleniumstatusen bovinemononuclearcelí funetion.J. Vet. Mcd. 40, 615-623.
SPALLHOLZ, J. E., BOYLAN, L. M. y LARSEN, H. 5. (1990).Advancesin understandingseleniunisrole in tIteImmuneSystem.Annais of dic New York Academy of Sciences587, 123-139.
STAMM, A. M., DISMUKES, W. E., SIMMONS, B. P., COBBS, C. Ci., ELLIOT. A., BUDRICI-I, P. y HARMAN,14. (1982).Listeriosisin renal siransplantrecipiensis:repofl of aix oubreakaud review of 102 cases.Rey. lnfect. Dis.4, 665-684.
STELMA, Ci. N., REYES. A. L., PEELER,1. T., FRANCIS,D W, HUNL ¡ M., SI’AULDING, P. L., JOHNSON.LI. L. y LOVETr, J. (1987).Patiiogenicitytestfor Listeriarnonocvtogenesusing immunocomprornisedmice.5. Cliii.Mierobiol. 25, 2085-2089
STUART. 5. E. y WELSHIMER, 11. 1. (1973). Intagenerierelaednessof Listeria Pirie. mt. J. Syst. Bacteriol. 23.8-14.
STUART, 5. E. y WELSHIMER, J. 5. (1974). Taxonotnicreexanñnationof Listeria Pirie and transferof L. ~rayiand L. muxTayi te a new genusMurraya. mt. J. Syst. Bacteriel. 24, 177-185.
SUAREZ, Ci. (1991). Epidemiologíade la listeriosis.Analesde la Real AcademiaNacional de Medicina, lomoCVIII, pp=~
155

Bibtiogcafta
TABOURET, M., DE RYCKE, J., AUDURIER, A. y POUTREL, B. (1991). Padiogenicity of Listeriamonocvtogenesisolatesin immunecompromisedmice in relation te listeriolysin producrion.J. Med. Microbiol. 34,13-18.
TEIGE, LI., LARSEN, 14. J. y TOLLERSRUD, S. (1984). Swine dysentery:die influence of dietary seleniuxnenclinical and pathologicaleffectsof Treponemahyodysenteriaeinfection. Acta Vet. Scand.25, 1-9.
TILNEY, L. Ci. y TILNEY, M. 5. (1993). The wily waysof a parasite:inductien of actin asemblyby Listeria
.
TrendsMicrobiol. 1, 25-31.TURNER, R0 J. y HNCH, 5. M. (1991). Seleniumand site immuneresponse.Proc.Nut. See.50, 275-285.
VAN VLEET, 5. F., BOON,Ci. D. y FERRANS,V. J. (1981).Inductienof lessionsof seleniuin-vitaminEdeficiencyin weanling swinefed silver, cobalt, tellurium, zinc, cadmiun,andvanadium.Am. J. VeÉ. Res.42, 789-799.
VAN VLEET, J. F., REBAR, A. 14. y FERRANS,y. 5. (1977). Acute cobaltaud isoproterenolcardiotoxicity inswine: proteetionby selenium-vitaminE supplementationandpotentiationby stress-susceptiblephenotype.Am. J.Vet. Res. 38, 991-1002.
VAZQUEZ, J. A. (1989). Citolisinasde losmicroorganismosdel géneroListeria. TesisDoctoral.Fac. Veterinaria,1.1CM.
VAZQUEZ, J. A., DOMíNGUEZ, L., FERNANDEZ,J. F., BLANCO, J. L., GOMEZ-LUCIA, E., BRIONES. V.y SUAREZ, Ci. (1988). Serologicalstudiesen L. gravi and L. murravi. J. Appl. Bacteriol. 64, 371-378.
VAZQUEZ-BOLAND,J.A.;DOMIINGUEZ, L.; BLANCO, M.; ROCOURT,J.; FERNANDEZ-GARAYZABAL, j.F.;GUTIERREZ, CH.; TASCON, RL.; RODRIGUEZ-FERRI,E.?. (1992). Epidemielogic investigationof a silage-associatedepizeotieof ovine listeric encephalitis,using a new Listeria-selectiveenumerationmediumanépiiagetyping. Am. JO Vel. Res. 53, 368-371.
VAZQUEZ-BOLAND, J. A. (1984).Anticuerposproducidosenconejosinoculadoscondistintasespeciesdel géneroListeria. Tesinade Licenciatura.Fac.Veterinaria,U.C.M.
VAZQUEZ-BOLAND, 5. A., FERRER, D. y ROCOURT, J. (1991). Heterogeneidadde las cepasde Listeriamonocytogenesaisladasde un brotedelisteriosisbumanaocurridoenValenciaen 1989.Enf.Infec.yMicrobiol.Clin.9, 442-444.
WATKINS, LI. y SLEATH, K. P. (1981). Isolation and enumerationof L. menec~qogenesfroin sewage.sewagesludge andriver water.J. Appl. Bacteriol. 50, 1-9.
WEIS, LI. (1975). The incidenceof L. monocytogenesen plantsaud soil. Proc. VICh InternationalSympesiumenProblemsof Listeriosis(Nottiugham>.
WEIS, J. y SEELIGER. II. P. R. (1975). Incidenceof L. monocysiogenesin nature. AppI. Microbiol. 30. 29-32.
WELSI-IIMER, 1-1. J. (1968). Iselatienof L. monecytogenesfrom vegetation.J. Bacteriel. 95, 300-303.
WFLSI-IIMER. U. .1. y DONKER-VOET, .1. (1971).L. monocysio~enesin ¡‘ature. Appl. Microbiol. 21, 516-519.
WELSIIIMER, fi. LI. y MEREDITH. A. L. (1971).Listeriamurrayi sp.nov.: a nitrate-reducingmanniiol-fenneníingListeria. ¡¡u, 1. Sys¡. Bacteriol. 21. 3-7.
WILKINSON, E. LI. y JONES,0. (1977). A numericaltaxonemicsurveyof Listeria and relatedbacteria.LI. Gen.
156

BibIio¡’rafja
Microbiol. 98, 399-421.
WILKIINSON, T. R. y HALL, E. R. (1971). Survival of L. monocytogenesin experimentallyinfected mice. Appl.Microbiel. 21, 108-111.
ZACHARA, B. A., MIKOLAOJCZA, J. y TRAHKOWSKA, U. (1993).Effec of variousseleniumn(Se) inlakes OiltissueSe levelsaud glutathioneperoxidaseactivities in lambs.5. Vet. Med. 40, 310-318.
ZYCHLINSKY, A., KENNY, E., MÉNARD, R., PR VOST, M. C., HOLLAND, 1. E. y SANSONE’fTl, P. J.(1994). lpaB mediatesmacrophageapoptosisinducedby ShWellaflexneri.Mcl. Microbiel. 11, 619-627.
157





























