Embed Size (px)
Citation preview

THE JouRNAL OF INVESTIGATIVE DERMATOLOGY
Copyright© 1970 b y The Williams & Wilkins Co.
Vol. 55, o. 3 Printed in U.S.A.
ENZYMES OF THE URONIC ACID PATHWAY IN THE HUMAN EPIDERMIS*
KIYOMI FUKUI, M.D. ArD KENNETH M. HALPRIN, M.D.t
ABSTRACT
EYidence has been obtained for the presence of four of the enzym of the ur·onic acid
pathway of carbohydrate metabolism in bomogenates of human epiclermi .
The existence of mucopolysaccharides within
the human epidermis and especially in the
intercellular spaces has been well documented
histochemically (1, 2, 3, 4). The biochemical
ob ervation of Fle ch (5) and Flesch and
E oda (6, 7, 8) demonstrated their existence
in normal and abnormal stratum corneum.
Also the uptake of radioactive sulfate (pre
sumably into acid mucopolysacchariden) by
human epidermis has been demonstrated (9,
10) . The importance of these compounds in
providing the intercellular " cement" substance
between epidermal cells has recently been
beautifully demon trated using newer tech
niques of histochemistry combined with elec
tron microscopy (11, 12).
Biochemical tudies of the enzymatic ma
chinery necessar~' for the production of this
''biological glue" by the epidermis have not
appeared. Except for histochemical (13, 14)
and biochemical (15) evidence of ,8-glucuroni
dase actiYit.y in epidermis, the literature is
confined to the demonstration of uridine di
phosphoglucose dehydrogenase (UDPGDH) both histochemically (16) and biochemically (17,
1 ) . Thi present work is an effort to expand
this field and provides quantitative determi
nation and kinetic data on four of the en
zymes of the "uronic acid" pathway of car
bohydrate metabolism in the kin. Thi path
way proYide the uronic acids which com
bine with the amino sugars to make the poly
saccharide chain and also leads directly into
the pento._ e pathway of carbohydrate metabo
lism (Fig. 1).
Received November 26, 1969; accepted for publication April 16, 1970.
This -work supported in part by The John A. Hartford Foundation, Inc.
*From the University of Miami D epartment of Dermatology and the Dermatology Service of the Veterans Administration Hospital, 1201 Northwest 16th Street. Miami, Florida 33125t. Reprint requests to Dr. Halprin.
MATERIALS AND METHODS
Six normal male adults ·were used. Epidermis from the upper back was removed using a Castroviejo electric keratome without anesthesia after the skin was prepared with alcohol. The removed epidermal strip measured 2 em X 5 em and were approximately 0.2 mm thick. These strip contained 5% to 20 % of dermis. In addition pure epidermal sheets were obtained from the abdominal wall using a uction blister device for derma-epidermal separation ( 19. 20). Ti sue \vere frozen in liquid
r2 within 5- 10 econds after removal. Frozen trips of epidermis were weighed while still frozen,
homogenized for one minute at ooc in 0.05 M t ris buffer pH 7.4 which contain d 0.003 M MgCh and 0.001 M EDTA1 (100 mg of epidermis/! ml of buffer) using a motor-driven glass homogenizer.
The enzyme activities were measured usinu a fluorornetric method which depends on the formation or disappearance of reduced nicotinamide adenine dinucleotide phosphate (N ADPH) or of reduced nicotinamide adenine dinucleotide ( N ADH) (22). These nucleotides are co-factors in many enzymatic reactions and fluoresce at approximately 460 m,u when irradiated by light of 340 m,u. The amount of fluorescence is directly proportional to the amount of reduced nucleotide present in the olution. The oxidized form of these nucleotides, i .. N ADP+ or N AD+ do not absorb or fluore ce at these wavelengths.
The optimum incubation mixtures for demonstrating the various enz~·me actiYities are given in Table I. All reactions wer carried out at room temperature. In all cases 20 ,ul of epidermal homogenate (corresponding to 2 mg of fresh tissue) were added to 1 ml of .05 M tris buffer containing 3 mM MgCL and 1 mM EDTA. Nucleotides were then added in the indicated amounts. After stabilization of the fluorescence occurred, reactions were started by adding the appropriate substrate. Initial velocities were recorded continuously and used to calculate the maximal reaction velocities2
• NAD+
1 Because of the contradictory information concerning the importance of metal ions with these enzymes (21 , 29) EDTA was added to chelate possible inhibitoDr metals while an excess of Mg++ wa added to allow metal requiring enzymes to retain activity.
2 Using a Turner Model 111 fluorometer equipped with a high sensitivity door the lowest enzyme values recorded (0.01 nm/rninjmg) con·e-
179

180 THE JOURNAL OF INVESTIGATIVE DERMATOLOGY
Uronic Acid Pothwoy Pentose Shunt ( sugars for
3-K eto l-gulonic Acid - L-xylulose..ili..Xylitol nucleos ide l 1 formation )
l -gulon ic Acid - Ascorbic D 1 <•It D-ribose-5-P t Ac id -xy u ose l r~ l
D 1 · A 'd D-xylulose-5-P - D-ribulose-5-P
-g ucurontc Ct 1 I
I Glycogen 1 Glucose
I /1/ UD;,t;lucose - Giucose-1 -P- /cos\~-P .
j Fructose -6 -P \ Glycolysts
UDPGiucuronic Acid* I \ 1
Glucosomine-6-P ~ / lactate
* -· UDP lduro'nic Ac id *uDP-Amino Sugars
""Mucopolysocchorides : Chains of repeating units of uronic octds end omino sugars . The UDP is split off
when the sugar is transferred . If the mucopo lys accharide is linked to o peptide or protein : glycoprotein .
**Enzymes essayed in present work ;
(1) UDP glucose dehydrogenase (2) D-glucuronote reductase (3) l -xylulose reductase (4) Xylitol dehydrogenase.
FIG. 1
is r duced to NADH by epidermal homogenates in the absence of any added substrates . It is, therefore, essential in all reactions requiring this coenzyme that the formation of N ADH by the ti ue be allowed to go to completion (about 10 minutes) prior to adding the substrate3 for the particular enzyme activity to be measured. This "nothing" dchydrogena e which reduces NAD+ but not NADP ,+ has also been found on starch gel electrophoresis experiments and seems to be due to lactic dehydrogenase (23) . Dialysis for a long as 48 hours do s not remove the activity (24) .
RESUL'l'S
1. In VIYO enzyme activities. Table II lists th maximal measurable activities of these enzyme m epidermal homogenates made from keratome lice (5-20% dermal contamination) and from uction blister tops (100%
pid rmi ) (FiO'. 3). All nzyme activities are extremely low
(approximately 100 to 1000 times les, active than the enzym of glycolysis for example). Activitie in th pure epidermal sheet were uniformly le than the corre ponding activi-
pond to a change of 3/ 100ths of the recording paper during a thirty minute incubation. Reaction rates are linear for the thirty minute period. Reaction r tes ar proportional to the amount of enz. me (homogenat ) added over the range of 1-4 mg (1 0-40 ,ul) .
8 All sub trat s ga single spots on thin layer chromatography u ing a butanol ( 40): acetone (50): water (50) ystem.
ties measured in homogenates made from keratome slices containing some dermis.
2. In vitro kinetic studies. (A) pH optima for the four enzymes are shown in Figure 2. All have sharp peaks in the alkaline range (pH .6-9.6).
(B) The effects of different substrate and coenzyme concentrations on enzyme activity were determined over a wide range of concentrations for each enzyme activity. The results were plotted according to the method of Lineweaver and Burke and the "apparent" Michaelis con tants (Km) determined (i.e. the concentration of substrate or coenzyme required for half -maximal enzyme activity) . These re ult together with the pH optima are shown in Table III. Relatively high concentrations of sub trate are necessary for optimal velocity of all four enzymatic activities when tudied in crude homogenates.
The pH optimum of .7 for UDPGDH agree well with those of rabbit skin (25) and calf liver (26) a does the "apparent" Km for N . ..AJ)+ (26). The "apparent" Km for UDPG varies widely depending on buffer and condition of as ay ha. ving been reported over a range of 2 X 10-5 M for calf liver (27) and rabbit (25) to 2.5 X 10-<1 again for calf liver (26). Our rrapparent" Km is some 4-5
times higher. Pig kidney has been report~d to have similar
values for D-glucuronate reductase (28) even when mea ured in the rever ·e direction. Guinea pi()' liver has been reported to have similar pH optima and rrapparent" Km's for substrate in t he case of both L-xylulose reductase and xylitol DH v .. ·hen mea ured in the same direction that we have used (29).
DI CUSSION
Evidence has been obtained for the presence of four of the enzymes of the uronic acid pa.thway of carbohydrate metabolism in homogenates of human epidermis. Since this pathway is involved in the formation of mucopoly-accharide and might be expected to be quite
active in the dermis (25) it is important to tre the fact that these activities were found
in homogenates of pure epidermis as well as in homogenate of keratome slices.
The activitie were lower in the pure epidermis a compared to the keratome slice. This can be interpreted a indicating that the

URONIC ACID PATHWAY ENZYMES 1 1
TABLE I
Optimal reaction mixtures for enzymes of the uronic acid pathway substrate concentration (micromoles / milliliter)
Enzyme pH* of UDPG D-glu- Xylitol NAD+ ADP+ NADPH buffer curonate
UDPGDH 8 .7 4.5 2
D-glucUJ·onate reductase 8.8 40 0.03
L-xylulose reductase 9.6 100 0 .1
Xylitol"'dehydrogenase 8 .8 50 0.5
*-Buffer is 0.05 M Tris buffer containing MgCl 2 (3mM) and EDTA (1mM).
dermis is more active than the epidermis but
it may also be due to damage to the epidermis during the process of blister formation.
UDPGDH, the first enzyme of the pathway has also been demonstrated histochemically m
TABLE II
Enzyme activities in nor·mal human epidermis
Epidermis with 5-20% Pure dermal contamination epidermis
Enzyme
Range Aver- Range Aver-age age
-- - -
UDPGDH 0.053-0.113* 0.084 0 .013- 0.020 0.017 D-glucuron- 0 .042-0 .056 0 .049 0.03Q-0 .035 0.033
ate reduc-tase
L-xyl ulose 0.017-0.030 0.025 0 . 010- 0 .013 0.012 reductase
Xylitol DH 0 .035-0 .058 0.047 0.033-0 .040 0.037
* Activities in nanomoles of substrate turned
over / min/ mg fresh tissue.
>.....
100
> 10
..... u 70 -c(
0 30
10
pH ACTIVITY CU RV E
~ 0- glu curo nolll reductase
•- L - xyl ulose reductase
o---o Xylitol DH
0
o ~-r----~-----r-----r----~------7.0 ap 9.0 101) n.o pH
0 .05 M Tr is buffer containing MgCI 2 (3mM)
ond EDTA (lmM)
FIG. 2
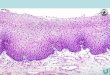
FIG. 3. Photomicrograph of suction induced blister to show complete epidermal separation.~
human epidermis (16). Uridine dephosphogluco-·e (UDPG) which initiate the pathway i
also the substrate for glycogen synthetase leading to the formation of glycogen. It would be useful to know the relative amount of UDPG going in each direction. The mea urable enzyme activit~' of glycogen syntheta e is four times that of DPGDH, however, one cannot as ume from thi~· in vitro measurement that glycogen formation is four times fa;-ter than mucopoly
saccharide formation in vivo. Conditions tn vivo are probably quite different from those in vitro; e.g. there may be two substrate pools of UDPG within the ce.ll rather than one, pH conditions may be quite different, or since
mucopolysaccharide probably do not turn
over as fast as glycogen even low activities of
l Courtesy of Doctor James Fulton.

182 THE JOURNAL OF INVESTIGATIVE DERMATOLOGY
TABLE III
Kinetic data for each enzyme (at room temperature)
"Apparent" Km (Michaelis Constant)
Enzyme pH
optimum For substrate For coenzyme
UDPGDH 8.7 1.5 X 10- 3M (UDPG) 7 . 2 X 10-4M (N AD+)
D-glucuronate reductase 8.8 1.3 X 10-2M (D-glucuronate) 1 .0 X 10-6M (NADPH)
L-xylulose reductase 9.6 2. 5 X 10-3M (xylitol) 8. 7 X 10-6M (NADP+)
Xylitol dehydrogenase 8.8 3.5 X 10-3M (xylitol) 1. 3 X 10-4M (N AD+)
formation may lead to considerable amounts in
the ti sue and perhaps inhibition of further
formation. The formation of mucopolysaccharides re
quires not only the formation of UDP glu
ronic acid from UDPG, but also the transfer
of this bound glucuronate to the growing
mucopolysaccharide chain by the enzyme UDP
glucuronyl transferase and no information is
available on the activity of this enzyme.
It is also unknown whether the transferase
which adds glucurona te to its normal substrate
in mucopoly accharide formation is the same
enzyme as that which transfers glucuronate to
simple chemicals; e.g. addition of glucuronate
to bilirubin. Once t ran ferred to its sub t rate molecule
from UDP-glucuronate the glucuronate is at
tached by ,8-linkage and is susceptible to the
action of ,8-glucuronidase releasing free glu
curonate (30). Glucuronate may also be formed
dir ctly by the breakdown of UDP-glucuronate
to glucuronate-1-phosphate and then to free
glucuronate (21). The format ion of L-gulonate from D-glu
curonate by D-glucuronate reductase provides
a ubstance which in most animals (except
man, other primates, and guinea pigs) is con
v rted to a corbic acid (vitamin C). From the
tudi -s of Baker, et al. (31) it would seem that
th immedia t pr cur or of ascorbic acid are
not D-O'lucuronate and L-gulonate but the
la tone of th e compound .- These workers
hawed that 25% of admini tered C14 labeled
D-glucuronolactone appeared in the urine as
ascorbic acid in human while no activity
could be found after the administration of cu lab lied O'lucuronic acid. Presumably man can
not make the lactone from the free acid (28).
mce a corbic acid i not made by man it
ma.y m uperfluou to have enzymes which
metabolize compounds having no known func
tion. Perhaps a new role for these carbohy
drates will be found or as suggested by
Touster (21) perhaps they somehow function
to regulate the relative proportions of the
coenzymes NAD and NADP in their reduced
and oxidized states.
REFERENCES
1. Wislocki, G. B., Bunting, H. and Dempsey, E. W.: Metachromasia in mammalian tissues and its relationship to mucopolysaccharides. Amer. J. Anat., 81: 1, 1947.
2. Braun-Falco, 0.: Histochemical and morphological studies of normal and pathological human skin. Arch. Derm. Syph., 198: 111, 1954.
3. Braun-Falco, 0 .: The histochemistry of psoriasis. Ann. N.Y. Acad. Sci., 73: 936 1958.
4. Steigleder, G. K. and Weakley, D. R.: Mucopolysaccharides in human epidermis. Brit. J. Derm .. 73: 171, 1961.
5. Flesch, P.: Mucopolysaccharides in human epidermis. J. Soc. Cos. Chemists, 10: 154, 1959.
6. Flesch, P. and Esoda, E. C. J.: Mucopolysaccharides in human epidermis. J. Invest. D erm .. 35: 43. 1960.
7. Flesch, P. and Esoda, E. C. J.: Isolation of a glycoproteolipid from human horny layers. J. Invest. Derm., 39: 409, 1962.
8. Flesch, P. and Esoda, E. C. J.: Further studies of epidermal mucopolysaccharides. Arch. Derm., 88 : 706, 1963.
9. Braun-Falco. 0., Christophers. E. and Kurban, A.: On the uptake of S36 sulfate by psoriatic kin in vitro. Arch. Klin. Exp. Derm., 224:
310. 1966. 10. Barker, S. A .. Cruickshank, C. N. and Webb,
T.: The effect of vitamin A. Hydrocortisone and citral upon sulphate metaboli m in skin. Exp. Cell Res., 35: 255, 1964.
11. Mercer, E. H., John, R. A. and Maibach, H. I.: Surface coats containing pol~·saccharides on human epidermal cells. J. Invest. D erm., 51 : 204. 1968.
12. M ercer, E. H. and Maibach. H. I.: Intercellular adhesion and surface coats of epidermal cell in p oriasi . J. Inve t . Derm., 51: 215, 1968.
13. Montagna. W.: Histology and cytochemistry of human skin XI. The di t ribution of beta-

URONIC ACID PATHWAY ENZYMES 183
glucuronidase. J. Biophys. Biochem. Cytol., 3: 343, 1957.
14. Mesirow, S. and Stoughton, R.: Demonstration of beta-glucuronidase in human skin. J. Invest. Derm., 23: 315, 1954.
15. Gibbs, Gordon, E. and Griffin, G. D.: Quantitative determination of B-glucuronidase in sweat gland and other skin components in cystic fibrosis. J. Invest. Derm., 51: 200, 1968.
16. Kamihata, S.: Histochemical study on uridine diphosphoglucose dehydrogenase in certain dermatological applications: Part III The histochemical demonstration of UDPG-DH activity in the lesions of cutaneous diseases. Jap. J. Derm. (Ser. B), 78: 170, 1966.
17. Halprin, K. and Ohkawara, A.: Glucose and glycogen metabolism in the human epidermis. J. Invest. Derm., 46: 43, 1966.
18. Halprin, K. and Ohkawara, A.: Carbohydrate metabolism in psoriasis: An enzymatic study. J. Invest. Derm., 46: 51, 1966.
19. Kiistala, U. and Mustakallio, K. K.: Dermaepidermal separation with suction. Electron microscopic and histochemical study of initial events of blistering on human skin. J. Invest. Derm., 48: 466, 1967.
20. Kiistala, U.: Suction blister device for separation of viable epidermis from dermis. J . Invest. Derm., 50: 129, 1968.
21. Touster, 0. and Shaw, D. R. D.: Biochemistry of the acyclic polyols. Physiol. Rev., 42: 181, 1962.
22. Lowry, 0. H., Roberts, N. R. and Kapphahn, J. I.: The fiuorometric measurement of pyridine nucleotides. J. Biol. Chern., 224 : 1047, 1957.
23. Ohkawara, A., Halprin, J., Barber, P. and Halprin, K. M.: Human epidermal isoenzyme . Arch. Derm. 95: 412, 1967.
24. Fukui, K. and Halprin, K. M.: Unpublished data.
25. Jacobson, B. and Davidson, E. A.: Biosynthesis of uronic acids by skin enzymes I. Uridine diphosphate-D-glucose dehydrogenase of rabbit skin. J. Bioi. Chern., 237: 635. 1962.
26. Bergmeyer, H. U.: Methods of Enzymatic Analysis. Verlag Chemie Academic Press, N.Y. and London, 1963.
27. Maxwell, E. S., Kalckar, H. M. and Strominger, J. L.: Some properties of uridine diphosphoglucose dehydrogenase. Arch. Biochem. Biophys., 65: 2, 1956.
28. York, J. L., Grollman, A. P. and Bublitz , C.: TPN -L-gulonate dehydrogenase. Biochim. Biophys. Acta, 47: 298, 1961.
29. Hickman, J. and Ashwell, G.: A sensitive and stereospecific enzymatic assay for xylulose. J. Biol. Chern., 234: 758, 1959.
30. Levy, G. A., Conchie, J., Colowick, S. P. and Kaplan, N.D.: Mammalian glycosidoses and their inhibition by aldonolectones, Methods in Enzymology VIII. Complex Carbohydrates. Academic Press, New York and London, 1966.
31. Baker. E. M., Sauberlich, H. E., Wolfskill, S. J .. Wallace, W. T. and Dean, E. E.: Tracer studies of vitamin C utilization in men: Metabolism of D-glucuronolactone-6-C14
, Dglucuronic-6-C14 acid and L-ascorbic-l-C14
acid. Proc. Soc. Exp. Biol. Med., 109: 737, 1962.