Embed Size (px)
Citation preview

UNIVERSIDADE ESTADUAL DE CAMPINAS
INSTITUTO DE BIOLOGIA
Camila Lubaczeuski
ENVOLVIMENTO DO SISTEMA NERVOSO PARASSIMPÁTICO
SOBRE O CONTROLE GLICÊMICO E FUNÇÃO DAS ILHOTAS
PANCREÁTICAS EM CAMUNDONGOS DESNUTRIDOS
SUBMETIDOS À DIETA HIPERLIPÍDICA
INVOLVIMENT OF THE PARASYMPATHETIC NERVOUS SYSTEM
ON GLYCEMIC CONTROL AND PANCREATIC ISLETS FUNCTION
IN MALNOURISHED MICE SUBMITTED TO THE HIGH-FAT DIET
CAMPINAS
2017

Camila Lubaczeuski
ENVOLVIMENTO DO SISTEMA NERVOSO PARASSIMPÁTICO SOBRE O
CONTROLE GLICÊMICO E FUNÇÃO DAS ILHOTAS PANCREÁTICAS EM
CAMUNDONGOS DESNUTRIDOS SUBMETIDOS À DIETA HIPERLIPÍDICA
INVOLVEMENT OF THE PARASYMPATHETIC NERVOUS SYSTEM ON
GLYCEMIC CONTROL AND PANCREATIC ISLETS FUNCTION IN
MALNOURISHED MICE SUBMITTED TO THE HIGH-FAT DIET
Tese apresentada ao Instituto de Biologia da
Universidade Estadual de Campinas como parte
dos requisitos exigidos para a obtenção do título de
Doutora em Biologia Funcional e Molecular, na
área de Fisiologia.
Thesis presented to the Institute of Biology of the
University of Campinas in partial fulfillment of the
requirements for the degree of PhD in Functional
and Molecular Biology in the area of Phisiology.
Orientador: Prof. Dr. Everardo Magalhães Carneiro
ESTE EXEMPLAR CORRESPONDE À
VERSÃO FINAL DA TESE DEFENDIDA
PELA ALUNA CAMILA LUBACZEUSKI E
ORIENTADA PELO PROF. DR.
EVERARDO MAGALHÃES CARNEIRO
CAMPINAS
2017


Campinas, 31 de julho de 2016.
COMISSÃO EXAMINADORA
Prof.(a) Dr. Everardo Magalhães Carneiro (Presidente)
Prof.(a). Dr. Paulo Cézar de Freitas Mathias
Prof.(a) Dra. Sandra Lucinei Balbo
Prof.(a) Dr. Claudio Chrysostomo Werneck
Prof.(a) Dr. Bruno Geloneze
Os membros da Comissão Examinadora acima assinaram a Ata de Defesa, que se encontra no
processo de vida acadêmica do aluno

AGRADECIMENTOS
Agradeço aos meus pais e meu irmão, por serem meus exemplos de caráter e honestidade e
por “tudo” sempre.
Ao professor Everardo Magalhães Carneiro, pela oportunidade de trabalhar com um ótimo
grupo de pesquisa, pela orientação e todos os ensinamentos.
Ao professor Antonio Carlos Boschero por ser um grande exemplo de cientista e professor.
Aos amigos do laboratório, pela ajuda nos experimentos, pelas discussões, pelo apoio diário e
toda a colaboração durante o desenvolvimento deste trabalho.
Agradeço à todos os meus amigos, os de perto e de longe, que sempre me incentivam e
estarem presentes nos melhores momentos.
Ao apoio dos técnicos do laboratório pelo indispensável esforço e contribuição fundamental
para o melhor andamento dos projetos do nosso grupo.
Agradeço às professoras Maria Lúcia Bonfleur e Sandra Balbo, por ter me apresentado à
pesquisa.
Agradeço ao meu noivo, Leandro, pelo apoio incondicional durante essa jornada e
contribuição para eu ser uma pessoa melhor todos os dias.
Agradeço à FAPESP e à UNICAMP pelo apoio financeiro e estrutural.

Somos resultados dos livros que lemos,
das viagens que fazemos e das pessoas
que amamos... (Airton Ortiz – escritor
brasileiro)

RESUMO
A restrição proteica em camundongos, durante a infância, promove a reprogramação do
metabolismo, causando prejuízo na tolerância à glicose e na ação da insulina. O sistema
nervoso parassimpático (SNP) parece estar envolvido nessas alterações, visto que em modelos
de programação metabólica ocorre aumento do tônus parassimpático e redução do simpático.
Considerando isso, nosso objetivo foi avaliar o papel da vagotomia sobre o controle glicêmico
através da secreção e ação dos principais hormônios pancreáticos: insulina e glucagon, em
camundongos desnutridos precocemente e posteriormente submetidos à dieta hiperlipídica
(HF). Após o desmame, camundongos C57Bl/6 foram submetidos à restrição proteica, sendo
alimentados com dieta hipoproteica (6% de proteína). Após 4 semanas, estes camundongos
foram divididos em três grupos: grupo LP, o qual continuou a receber a dieta hipoproteica;
grupo LP+HF, o qual passou a receber dieta HF (35% de lipídios); e grupo LP+HFvag, o qual
foi submetido ao procedimento de vagotomia no mesmo momento em que foi oferecido a
dieta HF. Ambas as dietas foram ofertadas durante 8 semanas. Após este período, foram
avaliados os seguintes parâmetros: secreção de insulina e glucagon estimulada por glicose;
tolerância à glicose; sensibilidade à insulina e ao glucagon; concentração plasmática de
insulina, peptídeo-c e glucagon; expressão proteica da enzima que degrada a insulina (IDE); e
conteúdo de glicogênio hepático. A vagotomia melhorou a tolerância à glicose e reduziu a
secreção de insulina, sem alterar a adiposidade e a sensibilidade à insulina nos animais
submetidos à reprogramação metabólica. Estes efeitos foram acompanhados pelo aumento da
insulinemia, provavelmente devido a redução do clearence de insulina observado no grupo
LP+HFvag. Além disso, os animais LP+HF e LP+HFvag foram mais sensíveis ao glucagon e
ao piruvato, contribuindo para a redução dos estoques de glicogênio hepático. Os animais
LP+HF apresentaram aumento da secreção de glucagon em ilhotas isoladas em resposta à
glicose, tanto em condições estimulatória (0,5mM) quanto inibitória (11,1mM). A vagotomia
melhorou a supressão de glucagon, em resposta a glicose, em ilhotas isoladas e in vivo,
contribuindo para a tolerância à glicose. Em conclusão, animais reprogramados
metabolicamente e submetidos à vagotomia apresentam melhora da tolerância à glicose com
aumento da insulinemia provavelmente devido a redução do clearance deste hormônio e
também pela redução da glucagonemia.
Palavras-chave: vagotomia; reprogramação metabólica; obesidade.

ABSTRACT
Protein restriction in mice during early life, promotes metabolic programming in adulthood,
impairing their glucose tolerance and insulin action. Parasympathetic nervous system
disruption seems to be involved in this process, since increased in parasympathetic tonus and
decreased in sympathetic activity was observed in metabolic programmed mice. Thus, the aim
of this study was to investigate the effect of subdiaphragmatic vagotomy on glucose
homeostasis through insulin and glucagon secretion and action, in metabolic programmed
mice, induced by a low-protein diet early in life, followed by high fat diet (HF) in adulthood.
After weaning, C57BL/6 mice were received low protein diet with 6 % of protein, composing
LP group. After 4 weeks, LP group was divided in LP+HF, which started to receive high-fat
diet (HF) (35% of lipids), for 8 weeks, and LP+HFvag, was submitted to the vagotomy
procedure at the same time as the diet was offered. Glucose-stimulated insulin and glucagon
secretion; glucose tolerance; insulin and glucagon sensitivity; plasma insulin, c-peptide and
glucagonconcentration; liver insulin-degrading enzyme (IDE) protein expression and hepatic
glycogen content, were evaluated. Vagotomy improved glucose tolerance and reduced insulin
secretion, but did not alter adiposity and insulin sensitivity in metabolic programmed mice,
compared to metabolic programmed mice. These effects were accompanied by increased in
insulinemia, probably due to a diminished insulin clearance, observed in LP+HFvag group.
Furthermore, LP+HF and LP+HFvag mice were more sensitive to glucagon and pyruvate,
contributing to lower hepatic glycogen content. The LP+HF mice presented increased
glucose-stimulated glucagon secretion in isolated islets, both in stimulatory (0.5 mM) or
inhibitory (11.1 mM) conditions. Vagotomy was able to improve glucagon suppression by
glucose in isolated islets and in vivo, improving glucose tolerance. In conclusion, the
metabolic programmed mice, when submitted to vagotomy, showed improved glucose
tolerance, associated with an increase of plasma insulin concentration as result of insulin
clearance reduction and reduction on glucagonemia.
Key-words: vagotomy; metabolic programming; obesity.

LISTA DE FIGURAS E TABELAS
Tabela 1: Composição das Dietas ........................................................................................... 20
Figura 1: Delineamento experimental para caracterização do modelo. .................................. 19
Figura 2: Delineamento experimental para indução da reprogramação metabólica. .............. 20
Artigo 1
Table 1: Body weight, fat pads and stomach weight, followed by glucose and insulin
measured in plasma in LP, LP+HF and LP+HFvag mice. ...... 3Erro! Indicador não definido.
Figure 1: Intraperitonial (ip) Glucose tolerance test (ipGTT) (A) and area under curve (AUC)
of ipGTT (B), insulin tolerance test (ITT) (C) and % glucose disappearance rate (KITT) (D)
in LP, LP+HF and LP+HFD mice.. .......................................................................................... 36
Figure 2: Glucose stimulation insulin secretion (A) and total insulin content (B) of islets from
LP, LP+HF and LP+HFvag group............................................................................................ 37
Figure 3: Effects of vagotomy on plasma c-peptide:insulin ratio during ipGTT in LP+HF
mice. AUCs of Plasma insulin (A), plasma c-peptide (B), and c-peptide:insulin ratio (C) at 0,
15, and 60 min during ipGTT.. ................................................................................................. 38
Figure 4: IDE protein expression in liver of LP, LP+HF and LP+HFvag mice.. .................... 39
Supplemental figure 1: Body Weight (A) and serum total protein (B) , of NP an LP groups.
.................................................................................................................................................. 40
Supplemental figure 2: Body Weight (A), perigonadal (B) and retroperitoneal fat pad (C), of
NP, NP+HF and NP+HFvag groups. ........................................................................................ 41
Supplemental figure 3: Glucose tolerance and insulin sensitivity evaluated by ipGTT (A),
AUC of piGTT (B), ipiTT (C), kITT (D) in NP, NP+HF and NP+HFvag mice.. ................... 42
Artigo 2
Table 1. Body weight, perigonadal and retroperitoneal fat pads, glycemia and insulinemia in
the fasted state of NP, LP, LP+HF, LP+HFvag mice. .............................................................. 48
Figure 1. Vagotomy improves glucose tolerance but not insulin resistance in LP + HFVag
mice.. ........................................................................................................................................ 49
Figure 2. Vagotomy reduces glucagonemia and glucagon secretion in LP + HFVag mice. ... 50
Figure 3.Vagotomy did not alter glucagon and pyruvate sensitivity in LP + HFVag mice.. .. 51
Figure 4. Hepatic glycogen content is reduced in LP + HF and LP + HFVag mice.. ............. 52

SUMÁRIO
1. Introdução.......................................................................................................................... 12
1.1 Desnutrição proteica e reprogramação metabólica ......................................................... 12
1.2 Secreção, ação e clearance de insulina ........................................................................... 13
1.3 Secreção e ação do glucagon .......................................................................................... 15
1.4 Sistema nervoso parassimpático (SNP) e controle glicêmico pelos hormônios
pancreáticos .......................................................................................................................... 17
2. Objetivo ............................................................................................................................. 19
3. Materiais e Métodos .......................................................................................................... 19
4. Artigo 1 ............................................................................................................................. 22
VAGOTOMY REDUCES INSULIN CLEARANCE IN OBESE MICE PROGRAMMED BY
LOW PROTEIN DIET IN THE ADOLESCENCE ................................................................. 22
Abstract ............................................................................................................................... 23
Introduction ........................................................................................................................ 23
Materials and Methods ...................................................................................................... 24
Results .................................................................................................................................. 27
Discussion ............................................................................................................................ 28
References............................................................................................................................ 30
5. Artigo 2 ............................................................................................................................. 43
VAGOTOMY REDUCES GLUCAGON SECRETION IN MALNOURISHED MICE
SUBMITTED TO HIGH FAT DIET ....................................................................................... 43
Abstract ............................................................................................................................... 44
5.1 Introduction .................................................................................................................. 44
5.2 Material and Methods .................................................................................................. 45
5.3 Results ............................................................................................................................ 47
5.4 Discussion ...................................................................................................................... 52
5.5 References...................................................................................................................... 58
6. Considerações Finais ......................................................................................................... 61
7. Referências ........................................................................................................................ 62
ANEXO 1- ................................................................................................... 72_Toc491953446
ANEXO 2 ................................................................................................................................. 73

12
1. Introdução
1.1 Desnutrição proteica e reprogramação metabólica
A desnutrição proteica é um termo genérico que associa uma série de doenças
relacionadas à falta de nutrientes na alimentação. Ela é caracterizada como uma desordem
nutricional de maior prevalência entre crianças de países em desenvolvimento (Food and
Agriculture Organization of the United Nations, 2004).
Estudos têm demonstrado que a restrição proteica em camundongos modifica o
mecanismo de secreção de insulina alterando a resposta secretória para nutrientes, agentes
despolarizantes e potencializadores da secreção, através da alteração do influxo e mobilização
intracelular de íons Ca2+
, e também da expressão de genes e proteínas fundamentais para a
secreção de insulina (Bradford, 1976; Latorraca et al., 1998; Ferreira et al., 2003; Araujo et
al., 2004; Delghingaro-Augusto et al., 2004; Ferreira et al., 2004; Filiputti et al., 2008;
Amaral et al., 2010; Da Silva et al., 2010; Filiputti et al., 2010). De forma adaptativa, apesar
de apresentarem insulinopenia, os animais submetidos à restrição proteica são
normoglicêmicos e mais tolerantes à glicose (Batista et al., 2012). Este efeito deve-se ao
aumento da sensibilidade periférica à insulina evidenciada no músculo esquelético desses
animais (Reis et al., 1997).
O organismo pode ajustar seu balanço energético de acordo com fatores ambientais e
genéticos. Esses dados podem ser observados em um estudo conduzido com indivíduos filhos
de mães que estavam grávidas durante o final da II guerra mundial e passaram por período
longo de fome, “Dutch Famine” (1944–1945). Os pesquisadores observaram que esses
indivíduos eram mais susceptíveis a obesidade, diabetes e doenças cardiovasculares. Essa
programação metabólica pode ser explicada pela hipótese do “Fenótipo Econômico”, o qual
propõe que uma nutrição pobre durante o desenvolvimento neonatal, pode resultar em uma
reprogramação fisiológica no intuito de permitir que a prole maximize a capacidade do corpo
de estocar energia, quando fora do útero (Hales e Barker, 1992). Barker e colaboradores,
demonstraram que indivíduos nascidos com baixo peso corporal podem desenvolver, na vida
adulta, hipertensão, obesidade, e Diabetes mellitus tipo 2 (DM2) (Barker et al., 1993).
Durante os primeiros estágios de desenvolvimento há uma janela de plasticidade
neuronal, período no qual o organismo tem um grande potencial de adaptação ao ambiente
nutricional (Remmers e Delemarre-Van De Waal, 2011). Camundongos adultos provenientes
de mães submetidas à desnutrição proteica durante o período gestacional apresentam várias

13
características que podem pré-dispor ao diabetes incluindo intolerância à glicose, aumento na
adiposidade e alterações na fisiologia do ciclo circadiano relacionado à atividade física e ao
gasto energético (Hales e Barker, 1992; Sutton et al., 2010). Essas observações sugerem que
uma crescente pandemia das doenças metabólicas ocorra por consequência de uma transição
nutricional, onde uma nutrição pobre durante a gestação ou na infância é substituída por uma
dieta hipercalórica durante a fase adulta, como pode ser comumente observado nos países em
desenvolvimento (Barker, 1995; Hales, 1997; Vickers et al., 2000). Atualmente vários estudos
buscam conhecer a origem do processo saúde-doença, baseado na hipótese do fenótipo
econômico, para ajudar explicar o aumento da prevalência da síndrome metabólica (Fewtrell
et al., 2000; Petry et al., 2000).
1.2 Secreção, ação e clearance de insulina
A glicose é o principal estimulador da secreção de insulina nas células β-pancreáticas.
Os mecanismos responsáveis pela secreção de insulina estimulada pela glicose iniciam-se com
o transporte deste açúcar para dentro das células β-pancreáticas, através de um transportador
específico denominado de GLUT2. A glicose é fosforilada à glicose-6-fosfato pela enzima
glicoquinase e metabolizada gerando ATP. O resultado é o aumento da razão ATP/ADP, que
provoca o fechamento do canal de K+ sensível ao ATP (K
+ATP), presente na membrana da
célula β. A redução do efluxo de K+ das células leva à despolarização da membrana que, por
sua vez, provoca a abertura dos canais de cálcio (Ca2+
) sensíveis à voltagem e influxo deste
cátion (Yang e Berggren, 2006; Hiriart e Aguilar-Bryan, 2008), culminando com a exocitose
dos grânulos que contêm insulina.
A metabolização da glicose nas células β e a subsequente elevação da concentração
intracelular de Ca2+
([Ca2+
]i), podem ativar enzimas que produzirão outros mensageiros
intracelulares que contribuem para a amplificação do sinal iniciado pela glicose. Uma destas
enzimas é a adenilato ciclase (AC) que, ao clivar o ATP, produz adenosina monofosfato
cíclico (AMPc) que, por sua vez, ativa a proteína quinase A (PKA) (Delmeire et al., 2003;
Dyachok et al., 2008). Além disso, a metabolização da glicose e o aumento da [Ca2+
]i também
estimulam a hidrólise de fosfoinositídeos de membrana, através da ativação da fosfolipase C
(PLC) (Thore et al., 2005; Thore et al., 2007). Isto resulta na formação do inositol-1,4,5-
trifosfato (IP3) e diacilglicerol (DAG), que induz a liberação de Ca2+
de estoques
intracelulares e ativa a proteína quinase C (PKC), respectivamente. Portanto, aumento da

14
[Ca2+
]i e ativação da PKA, PLC e PKC, culminam com a exocitose dos grânulos de insulina
(Nesher et al., 2002; Seino e Shibasaki, 2005; Tengholm e Gylfe, 2009). A secreção de
insulina, além de ser regulada por outros nutrientes circulantes, tais como os aminoácidos
arginina e leucina, também pode ser regulada pelo sistema nervoso autônomico (SNA). O
sistema nervoso parassimpático (SNP) potencializa a secreção de insulina, via nervo vago,
através de receptores muscarínicos na célula β, enquanto o sistema nervoso simpático (SNS),
por meio de receptores α-adrenérgicos, inibe a secreção de insulina (Ahrén, 1999).
A ação da insulina ocorre principalmente no músculo e tecido adiposo estimulando a
captação da glicose, e no fígado inibindo a liberação de glicose (Whiteman et al., 2002;
Youngren, 2007). A ação desse hormônio nos tecidos alvos ocorre por meio da sua ligação
com a subunidade α do receptor de insulina (IR), membro da família de receptores de tirosina
quinases. O resultado da interação entre o hormônio e o IR, é uma mudança conformacional,
que induz a autofosforilação de vários residuos de tirosina nas subunidades β do IR, e a
fosforilação em tirosina dos substratos do IR (IRS), os quais servem de ancoragem para
proteínas que possuem um sítio específico para o acoplamento com outros sinalizadores
proteicos que apresentam o domínio SH2 (assim denominados devido à homologia com o
produto do oncogene src). A ligação dos domínios SH2 da fosfatidilinositol-3-quinase (PI3-K)
aos resíduos fosfotirosinas do IRS-1, ativa esta enzima que produz fosfolipídeos de
fosfatidilinositol (PI), dentre eles o PI (3,4,5)P3, o qual ativa a proteína quinase dependente de
PI (PDK1), e subsequentemente a proteína serina/treonina quinase (Akt). Esta proteína
quinase fosforila várias proteínas, como por exemplo, a AS160 (proteína substrato da Akt de
160 kDa), a qual participa da translocação de GLUT4 para a membrana plasmática. Além
disso, a Akt modula a atividade dos componentes da tradução, contribuindo para a síntese
protéica, fosforila e inativa a enzima glicogênio sintase quinase 3 (GSK3), estimula a
transcrição de enzimas relacionadas à lipogênese, e, em contrapartida, inibe a lipólise por
ativar fosfodiesterases inibindo a lipase hormônio sensível (Saltiel e Kahn, 2001; Whiteman et
al., 2002; Youngren, 2007).
A degradação de insulina pode ocorrer em todos os tecidos sensíveis à insulina, sendo
o fígado o principal órgão responsável pelo clearance de insulina. Aproximadamente 50% da
insulina secretada é removida na primeira passagem pelo fígado. O clearance de insulina
ocorre, basicamente, em três etapas: 1) ligação da insulina ao seu receptor (IR); 2)
internalização do complexo insulina-IR; e 3) degradação do hormônio pela IDE (insulin-
degrading enzyme), que é a principal enzima responsável por esse processo. A IDE é uma

15
zinco metaloprotease com peso molecular de 110kDa, sendo encontrada, principalmente, no
citosol das células (Duckworth et al., 1998). Apesar de ser uma enzima expressa em todos os
tipos celulares, a IDE é altamente expressa nas células hepáticas. Inicialmente, quando a IDE
foi descoberta, recebeu esse nome devido sua habilidade em degradar a insulina rapidamente.
Entretanto, hoje sabemos que a IDE também é capaz de degradar outros hormônios, como por
exemplo, o glucagon e a amilina (Maianti et al., 2014). Portanto, prejuízos na função da IDE
pode impactar o controle glicêmico, favorecendo o desenvolvimento de patologias, tal como
DM2.
Camundongos knockout para IDE apresentam um fenótipo diabético, com intolerância
à glicose e resistência à insulina, em consequência da hiperinsulinemia crônica induzida pela
deficiência dessa enzima (Abdul-Hay et al., 2011). Além disso, sabe-se que um modelo de
DM2 muito utilizado, o rato Goto-Kakizaki (Goto et al., 1976), o qual apresenta mutações no
gene da IDE, também são hiperinsulinêmicos, intolerantes à glicose e resistentes à insulina
(Farris et al., 2004). Em conjunto, estes dados apontam a IDE como um importante alvo
terapêutico para o tratamento e/ou preveção do DM2.
Apesar dos prejuízos na função da IDE representar um risco para o desenvolvimento
do DM2, alguns estudos sugerem que a sua inibição poderia atuar no tratamento dessa
patologia (Mirsky e Perisutti, 1955; Leissring et al., 2010; Maianti et al., 2014).
Recentemente, foi demonstrado que o tratamento com um inibidor da IDE potencializa a ação
da insulina, melhorando assim a tolerância à glicose em camundongos. Neste estudo, os
autores sugerem que uma inibição rápida e curta, no período pós-prandial, poderia ser a
melhor abordagem terapêudica para inibição da IDE (Maianti et al., 2014). Entretanto, ainda é
incerto se essa abordagem seria a mais apropriada, uma vez que em outro estudo a inibição da
IDE, apesar de melhorar a sinalização da insulina, reduziu a tolerância à glicose em
camundongos (Deprez-Poulain et al., 2015).
1.3 Secreção e ação do glucagon
O glucagon é secretado pelas células α-pancreáticas em resposta à redução da
glicemia. O mecanismo pelo qual ocorre a secreção de glucagon, ainda não está
completamente elucidado (Quesada et al., 2008). Muitos dos componentes envolvidos na
secreção de glucagon pelas células α-pancreáticas são semelhantes aos da secreção de insulina
pelas células β (Quesada et al., 2008).

16
A secreção desse hormônio é estimulada quando as concentrações de glicose estão
baixas, através de um transportador de glicose específico (GLUT-1), o qual apresenta um Km
baixo (1-2 mM), sendo muito eficiente em fazer o transporte de glicose em baixas
concentrações desse nutriente. A glicose então é transportada para o interior da célula e
metabolizada, aumentando a razão ATP/ADP, que provoca o fechamento dos canais de KATP,
promovendo alteração da voltagem da membrana plasmática das células α, levando a abertura
de canais de Ca2+
do tipo T. O influxo de Ca2+
aumenta a voltagem da célula, promovendo
abertura dos canais de sódio (Na+) e Ca
2+ do tipo N. Isto aumenta ainda mais o influxo de
Ca2+
, contribuindo para a ativação da maquinaria de exocitose dos grânulos contendo
glucagon (Nadal et al., 1999; Quesada et al., 2006; Quesada et al., 2008).
A contribuição do glucagon na manutenção da glicemia ocorre devido ao aumento da
produção hepática de glicose durante o jejum, pois estimula a glicogenólise e a
gliconeogenese (Ramnanan et al., 2011). O glucagon promove sua ação biológica através da
ligação e ativação do seu receptor (GcGR), o qual é expresso principalmente no fígado, mas
também pode ser encontrado em outros tecidos como o tecido adiposo, renal, pancreático,
adrenal e gastrointestinal (Svoboda et al., 1994). No pâncreas, o GcGr é expresso tanto nas
células α como nas β (Kedees et al., 2009; Müller et al., 2017). O GcGR é um receptor
acoplado à proteína G estimulatória (Gs), então quando o glucagon se liga ao seu receptor a
proteína é ativada, estimulando a adenilato ciclase a produzir cAMP, o qual inicia os eventos
de sinalização na célula. O aumento dos níveis intracelulares de cAMP, ativa a PKA,
promovendo a fosforilação da CREB (cAMP response element-binding protein), que migra
para o núcleo e induz a expressão de genes chave para a regulação da gliconeogenese e
glicogenólise, como a pro-hormonio convertase (PC), fosfoenolpiruvato carboxiquinase
(PEPCK) e a glicose 6-fosfatase (G6Pase) (Quinn e Granner, 1990; Herzig et al., 2001;
Henquin e Nenquin, 2014).
Elevadas concentrações de glucagon e secreção de insulina insuficiente, são os
marcadores fisiopatológicos do diabetes (Perley e Kipnis, 1967; Raskin e Unger, 1978) essa
condição é conhecida como hipótese bi-hormonal (Unger e Orci, 1975). A falta de insulina ou
a resistência a este hormônio, somado ao aumento da secreção glucagon, causa aumento da
produção hepática de glicose, contribuindo para a manutenção da hiperglicemia (Dunning e
Gerich, 2007; Quesada et al., 2008). As alterações na resposta secretória do glucagon,
observados no diabetes, têm sido atribuídos a defeitos na regulação e na sensibilidade à

17
glicose das células α, redução da função das células β, resistência à insulina e alterações do
SNA .
1.4 Sistema nervoso parassimpático (SNP) e controle glicêmico pelos hormônios
pancreáticos
As ilhotas pancreáticas são invervadas pelos SNP e pelo SNS. As terminações
nervosas dos SNP liberam neurotrasmissores, como aceticolina, e outros neuropeptidos, tais
como: peptídeo vasoativo intestinal (VIP), peptídeo ativador da adenilato-ciclase pituitária
(PACAP) e o peptídeo liberador de gastrina (GRP), os quais interagem com receptores
específicos nas células α ou β pancreáticas. O principal neurotramissor liberado pelas
terminações do SNS é a noraepinefrina, além dela libera também neuropeptído Y (NPY) e
galanina. As terminações do SNA intervam tanto as células α como as β pancreáticas,
controlando assim a secreção de glucagon e insulina, respectivamente. Enquanto o SNP
potencializa a secreção de insulina, via nervo vago, através de receptores muscarínicos
presentes na célula β, o SNS, por meio de receptores α-adrenérgicos, inibe a secreção de
insulina (Ahrén, 1999). Além disso, tanto a acetilcolina como a noraepinefrina, também
desempenham função importante no controle da secreção de glucagon, em resposta a
hipoglicemia (Ahrén, 2000).
A obesidade é caracterizada por um desbalanço do SNA. Em humanos obesos e em
modelos animais, ocorre aumento da atividade parassimpática com redução do tônus
simpático. Além disso, animais com aumento do tônus parassimpático, causado por lesão no
hipotálamo ventro medial (VMH), por exemplo, apresentam aumento da proliferação e da
massa das células β, sugerindo que a proliferação e secreção de insulina podem ser reguladas
pelo SNP (Berthoud e Jeanrenaud, 1979; Mitrani et al., 2007; Lubaczeuski et al., 2015).
Sabe-se que o nervo vago tem inúmeras funções fisiológicas relacionadas com o
consumo alimentar, metabolismo energético e controle da glicemia (Pardo et al., 2008).
Muitos estudos são conduzidos para avaliar o papel do nervo vago e do SNP na instalação da
obesidade, pesquisas da década de 80 demonstram que a vagotomia é capaz de reverter ou
prevenir o desenvolvimento de obesidade em roedores com lesão no hipotálamo ventromedial
(VMH) (Inoue e Bray, 1977; Cox e Powley, 1981)
A vagotomia promove redução do peso corporal, aumento da saciedade, redução da
secreção de insulina e melhora da homeostase glicêmica em modelos animais de obesidade

18
(Edvell e Lindström, 1998; Balbo et al., 2000; Kral et al., 2009). Reforçando a hipótese que o
SNP tem envolvimento central na instalação da obesidade.
Recentemente, a organização governamental dos Estados Unidos, Food and Drug
Administration (FDA), aprovou uma terapia que bloqueia o nervo vago com impulsos
elétricos em voltagens capazes de inibir a estimulação vagal, conhecida como terapia VBLOC.
Um dispositivo elétrico é colocado na região abdominal, ao redor do nervo vago, através de
uma laparoscopia, e então impulsos intermitentes bloqueiam o tráfico neural dos ramos vagais
(Ikramuddin et al., 2014). Essa terapia melhora o controle glicêmico e reduz o peso corporal
em pacientes obesos (Camilleri et al., 2008; Shikora et al., 2013). Esse tratamento apresenta
os mesmo efeitos observados pela vagotomia.
Condições de desnutrição proteica, seja no perído uterino ou nas primeiras fases de
desenvolvimento, também estão associadas com o desbalanço do SNA. Entretanto, ao
contrário da obesidade, em roedores desnutridos ocorre aumento do tônus simpático e redução
do parassimpático (Leon-Quinto et al., 1997; Leon-Quinto et al., 1998; Malta et al., 2014).
Isso está de acordo com a secreção de insulina, que é reduzida em modelos de desnutrição.
Por outro lado, quando roedores são submetidos à reprogramação metabólica, através
da restrição proteica durante a infância, seguido da exposição de dieta controle na fase adulta,
estes apresentam inversão do tônus autonômico, com aumento da atividade parassimpática e
redução do tônus simpático (Malta et al., 2014). Essas alterações foram acompanhadas por
intolerância à glicose e hiperinsulinemia, observadas nesses roedores.
O alto índice de obesos e sobrepeso está intimamente associado com a reprogramação
metabólica promovida por uma pobre nutrição nas fases iniciais da vida, levando uma carga
genética que propcia, na vida adulta, o acúmulo de gordura e prejuízos na homeostase
glicemia devido a alterações da secreção de insulina e glucagon. Acreditamos que o SNA
tenha importante participação nesse processo, visto que desempenha um papel importante no
controle da secreção desses hormônios e é profudamente alterado em condições de obesidade
ou desnutrição. Nossa hipótese é que na reprogramação metabólica o aumento do tônus
parassimpático seja o responsável, pelo menos em parte, pelos prejuízos na homeostase
glicêmica e função das células α e β pancreáticas.

19
2. Objetivo
Artigo 1: Analisar as repercussões da vagotomia subdiafragmática sobre a
sensibilidade, secreção e degradação da insulina.
Artigo 2: Analisar as repercussões da vagotomia subdiafragmática sobre a
sensibilidade e secreção glucagon.
3. Materiais e Métodos
3.1 Delineamento experimental
Nessa sessão de matériais e métodos, apresento o delinemento experimental que foi
utilizado para o desenvimento dos artigos, para facilitar o entendimento dos grupos
experimentais. Os matériais e métodos específicos estão descritos nas sessões de metodologia
dos respectivos artigos.
Utilizamos camundongos C57BL/6J, provenientes do Biotério Central da
Universidade Estadual de Campinas (CEMIB/UNICAMP) e mantidos no biotério setorial do
departamento de biologia funcional e estrutural do Instituto de Biologia da UNICAMP sob
condições de temperatura e ciclo claro-escuro (de 12h) controlados, com livre acesso à água e
ração (Protocolo CEUA/UNICAMP: 3379-1). Ao desmame (30º dia de vida), os
camundongos foram separados em 6 grupos.
Para a caracterização do modelo de desnutrição e da vagotomia, primeiramente
utilizamos animais que receberam dieta normoproteica (NP). Para isso utilizamos 3 grupos
(Figura 1) que receberam dieta normoproteica (NP), contendo 14% de proteína, durante as 12
semanas de tratamento. Os grupos NP+HF e NP+HFvag, com 60 dias passaram receber dieta
hiperlipídica (HF), por 8 semanas, até os 120 dias de vida. Os animais NP+HFvag foram
submetidos à vagotomia aos 60 dias de vida, enquanto os animais NP+HF foram submetidos a
falsa cirurgia (sham). De acordo com o desenho experimental abaixo:
Figura 1: Delineamento experimental para caracterização do modelo.
20
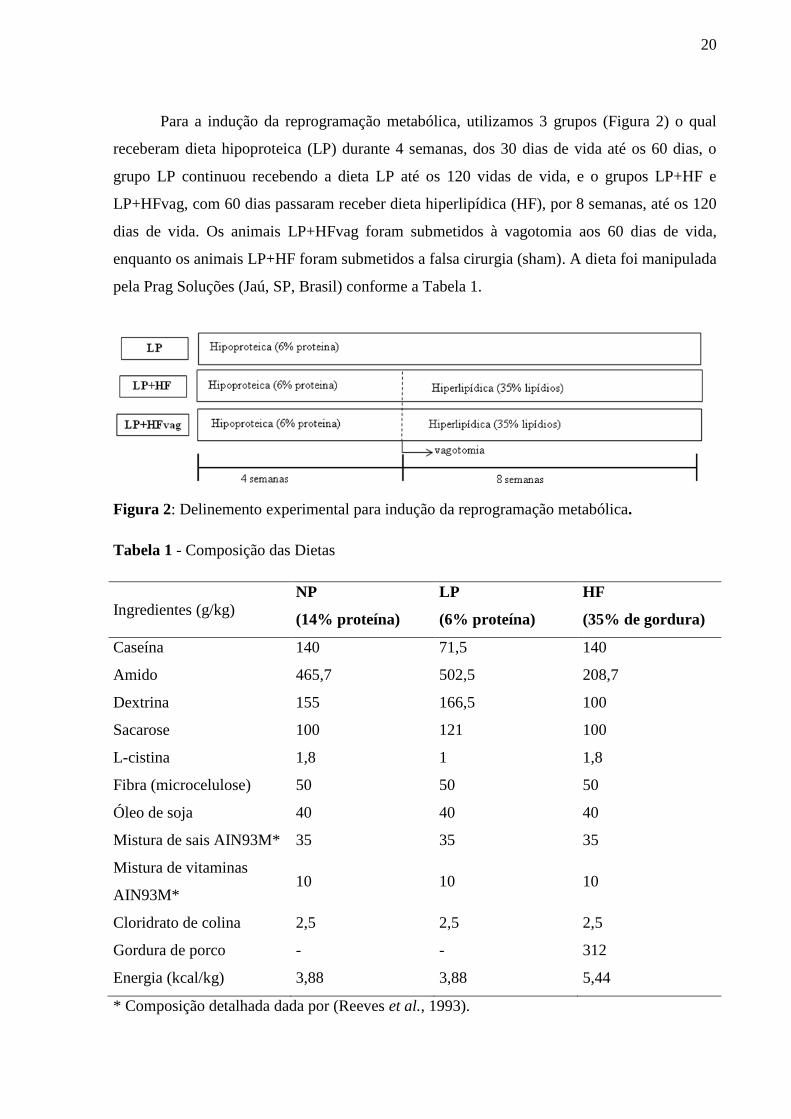
Para a indução da reprogramação metabólica, utilizamos 3 grupos (Figura 2) o qual
receberam dieta hipoproteica (LP) durante 4 semanas, dos 30 dias de vida até os 60 dias, o
grupo LP continuou recebendo a dieta LP até os 120 vidas de vida, e o grupos LP+HF e
LP+HFvag, com 60 dias passaram receber dieta hiperlipídica (HF), por 8 semanas, até os 120
dias de vida. Os animais LP+HFvag foram submetidos à vagotomia aos 60 dias de vida,
enquanto os animais LP+HF foram submetidos a falsa cirurgia (sham). A dieta foi manipulada
pela Prag Soluções (Jaú, SP, Brasil) conforme a Tabela 1.
Figura 2: Delinemento experimental para indução da reprogramação metabólica.
Tabela 1 - Composição das Dietas
Ingredientes (g/kg) NP
(14% proteína)
LP
(6% proteína)
HF
(35% de gordura)
Caseína 140 71,5 140
Amido 465,7 502,5 208,7
Dextrina 155 166,5 100
Sacarose 100 121 100
L-cistina 1,8 1 1,8
Fibra (microcelulose) 50 50 50
Óleo de soja 40 40 40
Mistura de sais AIN93M* 35 35 35
Mistura de vitaminas
AIN93M* 10 10 10
Cloridrato de colina 2,5 2,5 2,5
Gordura de porco - - 312
Energia (kcal/kg) 3,88 3,88 5,44
* Composição detalhada dada por (Reeves et al., 1993).

21
3.2 Vagotomia subdiafragmática bilateral
A vagotomia foi realizada aos 60 dias de vida com o objetivo de cortar a conexão
vagal. Os animais foram anestesiados com cloridrato de quetamina (Dopalen®, Vetbrands,
Paulínia, SP, BR) associado com cloridrato Xilazina (Anasedan®, Vetbrands, Paulínia, SP,
BR) (0,06mg/g de peso corporal e 0,02mg/g de peso, respectivamente). A vagotomia
realizada de acordo com (Balbo et al., 2000; Dixon et al., 2000). Os animais foram
posicionados em decúbito dorsal, foi realizada uma incisão inferior ao esterno na metade
superior do abdômen. O fígado foi afastado e os dois troncos do nervo vago e seus respectivos
ramos foram dissecados (vago anterior, o qual se divide em ramo hepático e gástrico anterior;
e vago posterior, o qual se divide acima do hiato esofágico em ramo celíaco e gástrico
posterior). O nervo vago e seus ramos foram ligados com fio de algodão e cortado. Nos
animais sham foi realizada a laparotomia exploratória.

22
4. Artigo 1
Title page
VAGOTOMY REDUCES INSULIN CLEARANCE IN OBESE MICE
PROGRAMMED BY LOW PROTEIN DIET IN THE ADOLESCENCE
Camila Lubaczeuski1, Luciana Mateus. Gonçalves
1, Jean Franciesco Vettorazzi
1, Mirian
Ayumi Kurauti1, Junia Carolina Santos-Silva1, Maria Lúcia Bonfleur
2, Antonio Carlos
Boschero1, José Maria Costa-Júnior
1, Everardo M. Carneiro
1*.
1University of Campinas (UNICAMP) – Campinas – São Paulo – Brazil
2State University of Western Paraná (UNIOESTE) – Cascavel – Paraná – Brazil
*Correspondent author:
Everardo M. Carneiro
Department of Structural and Functional Biology, Institute of Biology, University of
Campinas (UNICAMP), Monteiro Lobato Street, 13083-970, Campinas, SP, Brazil.
Email: [email protected]

23
Abstract
The aim of this study was to investigate the effect of subdiaphragmatic vagotomy on insulin
sensitivity, secretion and degradation in metabolic programmed mice, induced by a low-
protein diet early in life, followed by exposure to a high fat diet in adulthood. Weaned 30-day
old C57Bl/6 mice were submitted to a low protein diet (6% protein). After 4 weeks, the mice
were distributed into three groups: LP group, which continued receiving low protein diet;
LP+HF group, which started to receive a high fat diet; and LP+HFvag group, which
underwent vagotomy and also was kept at high fat diet. Glucose-stimulated insulin secretion
(GSIS) in isolated islets, ipGTT, ipITT, in-vivo insulin clearance, and liver expression of the
insulin-degrading enzyme (IDE) were accessed. Vagotomy improved glucose tolerance and
reduced insulin secretion, but did not alter adiposity and insulin sensitivity in the LP+HFvag,
compared with LP+HF group. Improvement in glucose tolerance was accompanied by
increased insulinemia, probably due to a diminished insulin clearance, as judged by the lower
c-peptide:insulin ratio, during the ipGTT. Finally, vagotomy also reduced liver IDE
expression in this group. In conclusion, when submitted to vagotomy, the metabolic
programmed mice showed improved glucose tolerance, associated with an increase of plasma
insulin concentration as result of insulin clearance reduction, a phenomenon probably due to
diminished liver IDE expression.
Introduction
It has been proposed that low or high calorie intake by mothers and fathers is
associated with disruption of glucose-insulin homeostasis in their offspring [1]. The mice kept
on a low protein diet early in life and fed a control diet, during adulthood, also display a
catch-up growth associated with glucose intolerance [2]. Indeed, economic improvements in
developing countries, during recent decades, have placed human subjects into similar
conditions. In these subjects, the intake of a normal or high-calorie diet in adulthood, after a
period of calorie restriction early in life, increases the risk to develop metabolic diseases [3,
4]. These early environmental situations are known as predictive adaptive response or Thrifty
Phenotype Hypothesis, postulated by Hales and Barker at 1992 [1].
We have shown that mice fed on a low protein diet in adolescence, followed by a
high fat diet during adulthood, develop glucose intolerance, insulin resistance and reduced
insulin secretion, compared to those fed on high fat diet during the whole experimental period

24
[5]. This indicates that the metabolic programming, induced by malnutrition in early life,
impairs insulin-glucose homeostasis to a greater extent than obesity per se. In addition,
malnourished and obese mice, may display injuries to the hypothalamic neurons, which
control energy intake and expenditure [6]. Glucose intolerant mice, exposed to a low protein
diet early in life and a control diet in adulthood, also show increased vagal activity, suggesting
the participation of the parasympathetic nervous system upon glucose homeostasis [2].
Metabolic programming can be explained by Developmental Origins of Health
and Diseases (DOHaD) concept, that describes through several studies how early
environmental factors , such as nutrition, which can induce physiological changes in fetal,
neonatal, adolescence and adult individuals, leading to a program to long-term postnatal
consequences [7-9].
Thus, we sought to explore the effect of subdiaphragmatic vagotomy on insulin
sensitivity, secretion, and degradation in metabolic programmed mice, induced by a low-
protein diet early in life, followed by exposure to a high fat diet in adulthood.
Materials and Methods
Animals
All animal experiments were carried out in accordance with the protocols approved by the
Animal Care and Use Committee of University of Campinas (UNICAMP) (approval number:
3379-1). Male C57Bl/6 mice were obtained from UNICAMP and maintained at 22 ± 1° C in a
12-h light–dark cycle. Thirty-day-old mice were fed on normal protein diet (14% protein) (NP
group) or low protein diet (6% protein) (LP group) during 4 weeks. After, LP mice were
distributed into three groups: LP, which was kept with low protein diet; LP+HF, which started
to receive a high fat diet (35% fat) during 8 weeks; and LP+HFvag which was submitted to
vagotomy and also started to receive a high fat diet during 8 weeks. The diets compositions
were described in a previous study [10].
Subdiaphragmatic vagotomy procedure
At 4 weeks after low protein diet consumption, LP+HF mice were submitted to
subdiaphragmatic truncal vagotomy (LP+HFvag group) or sham operation (LP+HF). For this
procedure, 12-h fasted mice were anesthetized with a mixture of ketamine and xylazine (0.06
and 0.02 mg/g via i.p., respectively; Vetbrands®, Paulínia, SP, BRA). Subsequently, the

25
stomach and esophagus were exteriorized from the peritoneal cavity, and both, dorsal and
subdiafragmatic vagal trunk, were separated from the esophagus and cut off. Sham-operation
mice underwent the same procedures, but the vagus nerve was kept intact. At the end of the
experimental period, to confirm subdiaphragmatic vagotomy, stomach food retention from all
groups of mice was evaluated by the ratio between the stomach weight per body weight
(BW), according to previous study [11-13].
Intraperitoneal glucose and insulin tolerance test
For the intraperitoneal (ip) glucose tolerance test (ipGTT), mice were fasted overnight (12 h)
and a basal blood sample was harvested from the tail tip (t=0 min). Mice received an ip
administration of 2g/kg glucose (Labsynth, Sao Paulo, Brazil) dissolved in saline solution
(0.9% NaCl wt/vol), and additional blood samples were recorded at 15, 30, 60, and 120 min.
Glucose was recorded using a handheld glucometer (Accu-Chek Performa II, Roche
Diagnostics, Switzerland). For the ip insulin tolerance test (ipITT), mice were fasted for 2
hours and an ip insulin (Humulin R, Eli Lilly, Indianapolis, USA) load (1 U/kg) was
administered. Blood was taken immediately before insulin injection (t=0 min) and at the times
3, 6, 9, 12, 15, 18 and 21 min via tail snip using a handheld glucometer. Glucose
disappearance rate (KITT) was calculated as previously described [14, 15].
Insulin clearance
During the ipGTT, blood samples were collected from the tail tip, before glucose load (t=0)
and 15 and 60 min after glucose administration, and placed into microtubes containing
anticoagulant heparin. The tubes were centrifuged at 1100g, 15min, 4°C, and the plasma was
collected and stored at −80°C. Insulin and c-peptide was measured by Rat/Mouse Insulin or c-
peptide 2 ELISA Kit (Cat. EZRMI-13K and EZRMCP2-21K, EMD Millipore, USA,
respectively), according to the manufacturer’s instructions. Insulin clearance was evaluated by
c-peptide:insulin ratio, as previously described [16].
Islet isolation and GSIS
Islets were isolated by collagenase digestion of the pancreas, as described (Boschero, et al.
1995). For static incubations, groups of five islets were pre-incubated for 30 min at 37 °C
with 500 μl of Krebs buffer (KBB) with the following composition: 115 mM NaCl, 5 mM
KCl, 2.56 mM CaCl2, 1 mM MgCl2, 10 mM NaHCO3, 15 mM HEPES, supplemented with

26
5.6 mM glucose, 3 g bovine serum albumin (BSA) per liter, and equilibrated with a mixture of
95% O2-5% CO2 to provide pH 7.4. After, this medium was replaced with fresh buffer, and
the islets were incubated for 1-h with 1 mL of KBB containing 5.6, 11.1 or 16.7 mM glucose.
At the end of the incubation period, the supernatants were collected and maintained at –20 °C.
For islet insulin content, groups of five islets were collected, transferred to tubes containing 1
mL deionized water and homogenized using a sonicator (Brinkmann Instruments, USA). The
insulin was measured by RIA using human insulin radiolabelled with 125
I as tracer, rat insulin
as standard (Crystal Chem Inc., USA), and rat insulin antibody (donated by Dr Leclerq-
Meyer, Free University of Brussels, Belgium). The charcoaldextran method was used to
separate free insulin from antibody-bound 125
I insulin.
Western blot
For Western blot analysis, liver samples from the mice were collected, snap-frozen in liquid
nitrogen and stored at −80°C, for subsequent protein extraction using a lysate buffer
(10mmol/L EDTA, 100mmol/L Tris base, 100mmol/L sodium pyrophosphate, 100mmol/L
sodium fluoride, 10mmol/L sodium orthovanadate, 2mmol/L phenylmethylsulphonyl fluoride,
1% Triton X-100, and 1μg/mL aprotinin). The Bradford method was performed to determine
the protein concentration, using BSA as a standard. After, 50 μg of the protein samples was
homogenized with Laemmli buffer and boiled at 100 ºC during 5 min. These samples were
resolved using 10% SDS-PAGE and electroblotted into nitrocellulose membranes, These
membranes were blocked in 10mmol/L Tris base, 150mmol/L NaCl, and 0.25% (vol/vol) of
Tween 20 (TBS buffer) containing 5% (wt/vol) BSA for 1h at room temperature. Membranes
were then incubated with primary antibodies (IDE, Abcam cat. ab32216; anti-GAPDH, Sigma
cat. G9545) overnight at 4°C. The detection was performed by enhanced chemiluminescence
(SuperSignal West Fento, Pierce Biotechnology Inc., Rockford, IL, USA) after incubation
with horseradish peroxidase-conjugated secondary antibody. The bands were visualized using
an Amersham Imager 600 (GE Healthcare Biosciences) and the intensities were quantified
using ImageJ software (National Institutes of Health, Bethesda, MD, USA).
Statistical Analysis
The data are presented as the means ± SEM, and the differences were considered significant
when p < 0.05. Comparisons were performed using ANOVA one-way followed by Tukey’s
test. Tests were carried out using GraphPad Prism, version 5.0 for Windows (GraphPad

27
Software, Inc., San Diego, CA, USA). Sample size was determined taking into account the
size effect. Bilateral statistic with a significance level of 5 % and potency of 0.98 was used to
rule out type II errors. Under these conditions, the recommended sample size required would
be n = 5, however, we opted for a size of n = 6 as a safety measure.
Results
Diets and vagus denervation characterization
First of all, we characterized the malnourished model, which showed reduced body weight
and serum total proteins (Supplemental figure 1). Then, we confirmed the efficiency of the
high fat diet used, since the mice fed on this diet became obese with augmented adiposity. We
also confirmed that vagotomy reduced body weight and fat pads (Supplemental figure 2) in
addition to improved glucose tolerance and insulin sensitivity in obese mice induced only by
high fat diet (Supplemental figure 3), a well-known effect of this surgery. Surprisingly,
vagotomy did not alter the body weight and adiposity in the LP+HFvag mice (Table 1). The
stomach weight was higher in LP+HFvag, compared with LP+HF mice, confirming the
efficiency of the vagotomy. The fasting glycemia and insulinemia were higher in the LP+HF
compared with LP mice. The fasting insulinemia, but not glycemia, was reduced in
LP+HFvag, compared with LP+HF mice (Table 1). However, we did not observe difference
in the fed insulinemia comparing the LP+HF with the LP+HFvag group.
Vagotomy improved glucose tolerance but not insulin sensitivity
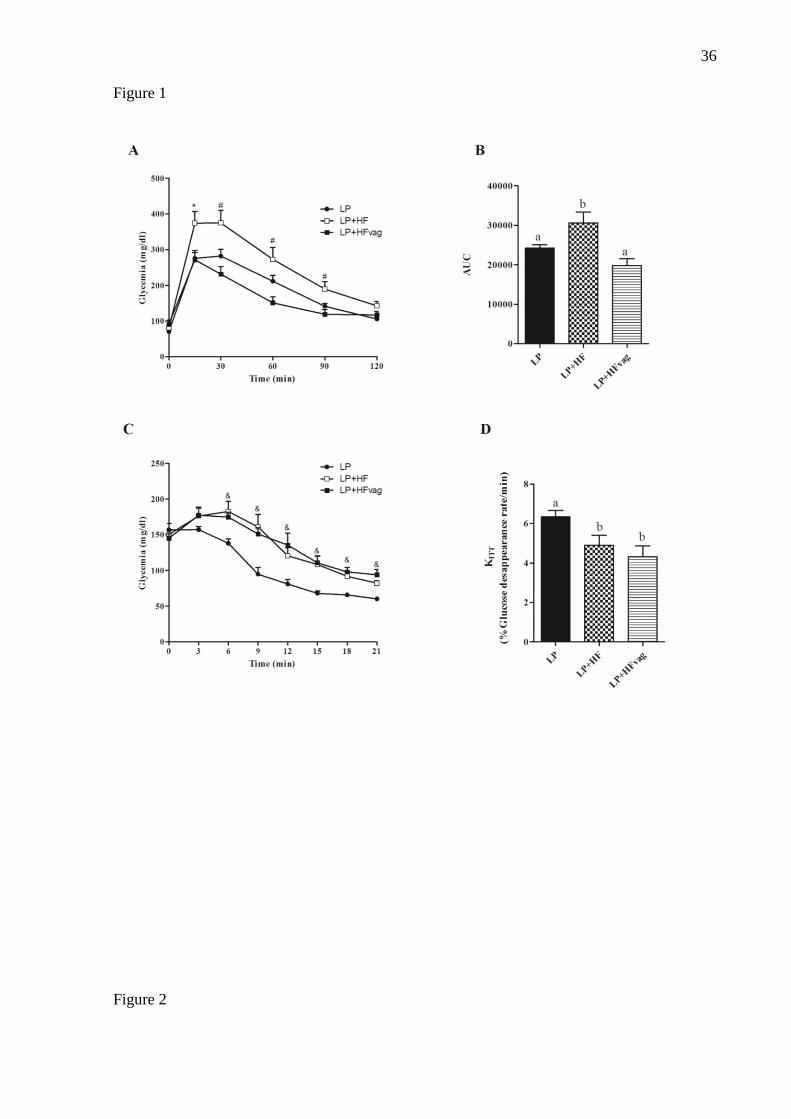
During ipGTT, LP+HF mice had an increased glycemia (Figure 1A), indicating an
impairment on glucose tolerance compared with LP mice, as judged by the AUC (Figure 1B).
Interestingly, vagotomy restored glucose tolerance in the LP+HFvag mice to the levels of
those observed in LP group, as observed in the AUC graph (Figure 1B). During ipITT (Figure
1C), LP+HF mice displayed impairment on insulin sensitivity, compared with LP group, as
demonstrated by the KITT (Figure 1D). Although the vagotomy did not alter the insulin
sensitivity, LP+HFvag mice had an increased fed insulinemia (Table 1), which could explain
the improved glucose tolerance in these mice.
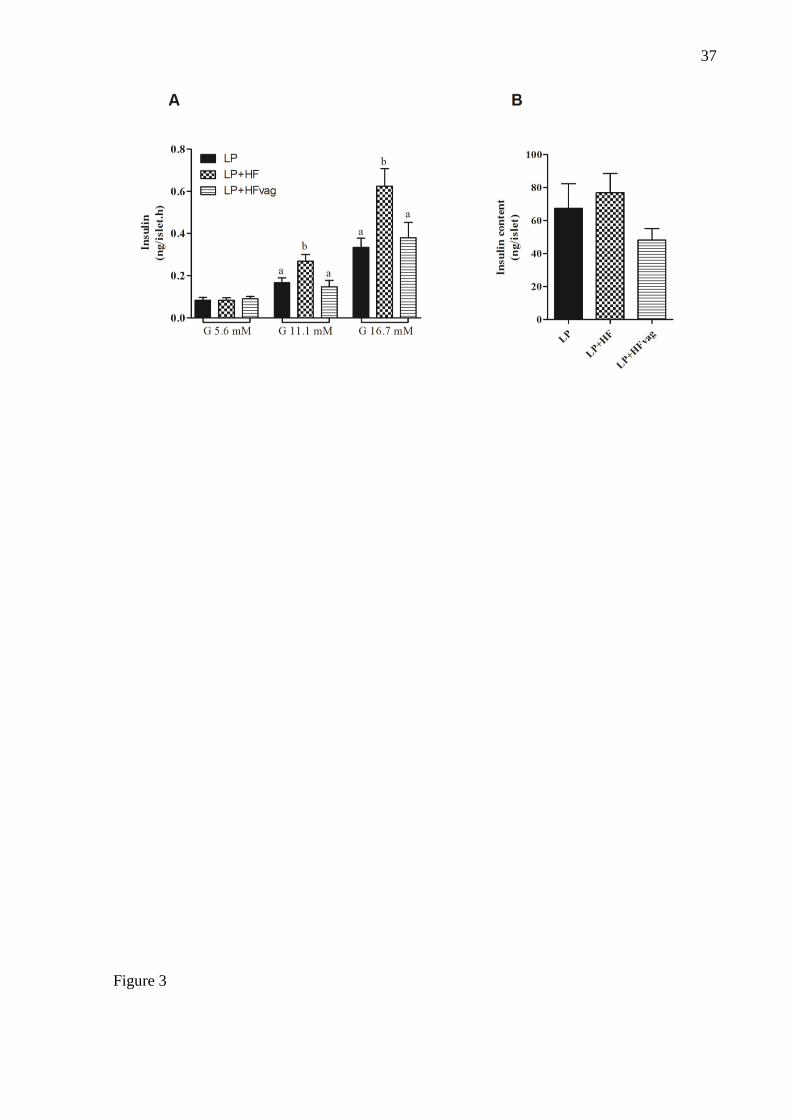
Vagotomy reduced GSIS in isolated pancreatic islets
To explain the higher insulinemia observed during ipGTT of LP+HFvag mice, we accessed

28
the GSIS in isolated pancreatic islets. At low glucose concentration (5.6 mM), insulin
secretion of all groups was similar. However, at high glucose concentrations (11.1 and 16.7
mM), an increased insulin secretion in the islets from LP+HF was observed, compared with
LP mice. The insulin secretion was lower in the islets from LP+HFvag mice, reaching similar
levels of those observed for the LP group (Figure 2A). The total insulin content of the islets in
all groups was not significantly different (Figure 2B).
Vagotomy reduced insulin clearance
The lower GSIS of LP+HFvag mice did not justify the higher insulinemia found in these mice
during the ipGTT (Figure 3A). Thus, we also evaluated the insulin clearance of these mice
(measuring the c-peptide:insulin ratio). It is known that pancreatic β cells co-secrete insulin
and c-peptide in a 1:1 ratio, however the half-time of c-peptide is longer than insulin. Thus an
augmentation in the c-peptide:insulin ratio indicates an increased insulin clearance, as
observed in the LP+HF, compared with LP mice (Figure 3C). Interestingly, insulin clearance
was reduced in LP+HFvag mice, with a decreased c-peptide:insulin ratio, compared with
LP+HF group (Figure 3C), explaining the higher insulinemia of those mice during the ipGTT.
Vagotomy reduced IDE expression in the liver of LP mice
IDE is the most important protein involved in insulin clearance, a phenomenon that
occurs mainly in the liver. Therefore, we evaluated IDE protein expression in the liver of
mice. Corroborating the insulin clearance data, LP+HF mice displayed higher IDE expression,
compared with LP group (Figure 4). The expression of this enzyme, in the liver of LP+HFvag
mice, was reduced, returning its values similar to those found in LP mice (Figure 4).
Discussion
Previous studies have demonstrated that metabolic programmed mice, fed a low
protein diet during childhood followed by a control diet in adulthood, developed glucose
intolerance, associated with augmented vagal activity [2]. Here, we performed vagotomy in
mice kept on a low protein followed by high fat diet to verify a possible role of the
parasympathetic nervous system on their insulin-glucose homeostasis. We observed that
vagotomy improved glucose tolerance of metabolic programmed mice by decreasing insulin
clearance, which probably occurs through reduced expression of the liver IDE.
It is known that diet-induced obesity, in mice, also provokes glucose intolerance

29
accompanied by increased insulin secretion, which compensates for the peripheral insulin
resistance [17]. This phenomenon has also been detected in metabolic programmed mice fed a
low protein diet in early life followed by regular diet [2] or high fat diet [5, 18] during
adulthood. Here, we confirmed these results with the LP+HF showing impaired insulin
sensitivity and glucose tolerance, as well as an increased insulin secretion.
Increased vagal activity in obese and metabolic programmed mice has been associated
with weight gain and higher insulin secretion, during the fed state [2, 19]. In addition,
vagotomized obese rodents showed reduced body weight, due to decreased fat pads,
associated with improved glucose tolerance, insulin sensitivity and secretion. In fact, we
confirmed that vagotomy was efficient to induce all these benefits, in the diet-induced obese
mice (figure S3). However, body weight and fat pads, as well as insulin action and secretion,
were similar between LP+HF and LP+HFvag mice. These results suggest that the
improvement observed in glucose homeostasis, in LP+HFvag mice, was independent of
alteration in body composition. These findings confirmed that metabolic programming
induced obesity is more problematic than obesity per se, since the programmed mice did not
display the well-known benefits from vagotomy, such as reducing in adiposity and body
weight [13, 20-22]. The same scenario has been seen when metabolic programmed mice
received taurine supplementation in an attempt to improve their glucose-insulin homeostasis
[18].
Contrary to our findings, previous reports have demonstrated that vagotomy did not
alter insulin clearance in lean pigs [23]. This suggests that vagotomy-induced reduction in
insulin clearance is a phenomenon observed only in an obese mice model, probably because
they experienced malnutrition during early life. In fact, in non-programming obese rats,
vagotomy also reduced insulin secretion but in contrast with our data, the insulinemia was
reduced [24], which reinforces our idea that insulin clearance reduction induced by vagotomy
may be dependent on metabolic programming. Although not evaluated, it seems that mice fed
a low protein diet early in life followed by a regular diet in adulthood, also develop decreased
insulin clearance. This assumption is based on the observation that they display augmented
plasma insulin concentration without an increase in glucose stimulated insulin secretion [2].
However, the mechanism by which vagotomy reduced insulin clearance and liver IDE
expression remains unclear.
Although reduced insulin clearance and liver IDE expression has been associated with
insulin resistance and the development of glucose intolerance [25, 26], in early life, IDE KO

30
mice showed higher plasma insulin concentration and improved glucose tolerance. However,
maintaining high plasma insulin concentrations for an extended period, led to insulin
resistance and glucose intolerance in these mice at 6 months old [27]. This phenomenon
seems to be caused by a negative feedback of the insulin pathway, orchestrated by the
overstimulation of the proximal insulin cascade. Thus, the improvement of glucose tolerance,
observed in LP+HFvag mice, could be a transient effect, and these mice are susceptible to
develop glucose intolerance and consequently T2D.
In conclusion, subdiaphragmatic vagotomy improves glucose tolerance in metabolic
programmed mice fed a low-protein diet early in life followed by exposure to a high fat diet in
adulthood. Vagotomy increases their plasma insulin concentration by reducing insulin
clearance, a phenomenon probably due to diminished expression of liver IDE. However, it is
necessary to keep in mind that strategies toward vagal activity inhibition, for the control of
metabolic diseases, may jeopardize glucose tolerance over time.
Acknowledgments
The authors thank Marise Brunelli, Julia Agulhari and Jheynifer Souza for technical
assistance and Bridgett A Bollin for English editing. This work was supported by the
Fundação de Amparo e Pesquisa do Estado de São Paulo (FAPESP; process number
2013/27847-6, 2013/07607-8, 2014/01717-9) and Conselho Nacional de Pesquisa (CNPq).
Conflict of Interest
The authors declare that there is no conflict of interest regarding the publication of this article.
.
References
1. Hales CN, Barker DJ: Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty
phenotype hypothesis. Diabetologia 1992, 35:595-601.
2. Malta A, de Oliveira JC, Ribeiro TA, Tófolo LP, Barella LF, Prates KV, Miranda RA,
Elmhiri G, Franco CC, Agostinho AR, et al: Low-protein diet in adult male rats has
long-term effects on metabolism. J Endocrinol 2014, 221:285-295.
3. Abdullah A: The Double Burden of Undernutrition and Overnutrition in
Developing Countries: an Update. Curr Obes Rep 2015, 4:337-349.
4. Sandovici I, Hammerle CM, Ozanne SE, Constância M: Developmental and
environmental epigenetic programming of the endocrine pancreas: consequences
for type 2 diabetes. Cell Mol Life Sci 2013, 70:1575-1595.

31
5. Leite NC, de Paula F, Borck PC, Vettorazzi JF, Branco RC, Lubaczeuski C, Boschero
AC, Zoppi CC, Carneiro EM: Protein malnutrition potentiates the amplifying
pathway of insulin secretion in adult obese mice. Sci Rep 2016, 6:33464.
6. Camargo RL, Batista TM, Ribeiro RA, Branco RC, Da Silva PM, Izumi C, Araujo TR,
Greene LJ, Boschero AC, Carneiro EM: Taurine supplementation preserves
hypothalamic leptin action in normal and protein-restricted mice fed on a high-
fat diet. Amino Acids 2015, 47:2419-2435.
7. Armitage JA, Khan IY, Taylor PD, Nathanielsz PW, Poston L: Developmental
programming of the metabolic syndrome by maternal nutritional imbalance: how
strong is the evidence from experimental models in mammals? J Physiol 2004,
561:355-377.
8. Howie GJ, Sloboda DM, Kamal T, Vickers MH: Maternal nutritional history
predicts obesity in adult offspring independent of postnatal diet. J Physiol 2009,
587:905-915.
9. de Oliveira JC, Lisboa PC, de Moura EG, Barella LF, Miranda RA, Malta A, Franco
CC, Ribeiro TA, Torrezan R, Gravena C, Mathias PC: Poor pubertal protein
nutrition disturbs glucose-induced insulin secretion process in pancreatic islets
and programs rats in adulthood to increase fat accumulation. J Endocrinol 2013,
216:195-206.
10. Batista TM, Ribeiro RA, da Silva PM, Camargo RL, Lollo PC, Boschero AC,
Carneiro EM: Taurine supplementation improves liver glucose control in normal
protein and malnourished mice fed a high-fat diet. Mol Nutr Food Res 2013,
57:423-434.
11. Balbo SL, Mathias PC, Bonfleur ML, Alves HF, Siroti FJ, Monteiro OG, Ribeiro FB,
Souza AC: Vagotomy reduces obesity in MSG-treated rats. Res Commun Mol
Pathol Pharmacol 2000, 108:291-296.
12. Dixon KD, Williams FE, Wiggins RL, Pavelka J, Lucente J, Bellinger LL, Gietzen
DW: Differential effects of selective vagotomy and tropisetron in aminoprivic
feeding. Am J Physiol Regul Integr Comp Physiol 2000, 279:R997-R1009.
13. Lubaczeuski C, Balbo SL, Ribeiro RA, Vettorazzi JF, Santos-Silva JC, Carneiro EM,
Bonfleur ML: Vagotomy ameliorates islet morphofunction and body metabolic
homeostasis in MSG-obese rats. Braz J Med Biol Res 2015, 48:447-457.
14. Protzek AO, Rezende LF, Costa-Júnior JM, Ferreira SM, Cappelli AP, de Paula FM,
de Souza JC, Kurauti MA, Carneiro EM, Rafacho A, Boschero AC:
Hyperinsulinemia caused by dexamethasone treatment is associated with reduced
insulin clearance and lower hepatic activity of insulin-degrading enzyme. J
Steroid Biochem Mol Biol 2016, 155:1-8.
15. Rafacho A, Abrantes JL, Ribeiro DL, Paula FM, Pinto ME, Boschero AC, Bosqueiro
JR: Morphofunctional alterations in endocrine pancreas of short- and long-term
dexamethasone-treated rats. Horm Metab Res 2011, 43:275-281.
16. Kurauti MA, Costa-Júnior JM, Ferreira SM, Dos Santos GJ, Protzek AO, Nardelli TR,
de Rezende LF, Boschero AC: Acute exercise restores insulin clearance in diet-
induced obese mice. J Endocrinol 2016, 229:221-232.
17. Kahn SE, Hull RL, Utzschneider KM: Mechanisms linking obesity to insulin
resistance and type 2 diabetes. Nature 2006, 444:840-846.
18. Cappelli AP, Zoppi CC, Barbosa-Sampaio HC, Costa JM, Protzek AO, Morato PN,
Boschero AC, Carneiro EM: Taurine-induced insulin signalling improvement of
obese malnourished mice is associated with redox balance and protein
phosphatases activity modulation. Liver Int 2014, 34:771-783.

32
19. Kiba T, Tanaka K, Numata K, Hoshino M, Misugi K, Inoue S: Ventromedial
hypothalamic lesion-induced vagal hyperactivity stimulates rat pancreatic cell
proliferation. Gastroenterology 1996, 110:885-893.
20. Barella LF, Miranda RA, Franco CC, Alves VS, Malta A, Ribeiro TA, Gravena C,
Mathias PC, de Oliveira JC: Vagus nerve contributes to metabolic syndrome in
high-fat diet-fed young and adult rats. Exp Physiol 2015, 100:57-68.
21. Johannessen H, Revesz D, Kodama Y, Cassie N, Skibicka KP, Barrett P, Dickson S,
Holst J, Rehfeld J, van der Plasse G, et al: Vagal Blocking for Obesity Control: a
Possible Mechanism-Of-Action. Obes Surg 2017, 27:177-185.
22. Shikora S, Toouli J, Herrera MF, Kulseng B, Zulewski H, Brancatisano R, Kow L,
Pantoja JP, Johnsen G, Brancatisano A, et al: Vagal blocking improves glycemic
control and elevated blood pressure in obese subjects with type 2 diabetes
mellitus. J Obes 2013, 2013:245683.
23. Blat S, Malbert CH: The vagus is inhibitory of insulin secretion under fasting
conditions. Am J Physiol Endocrinol Metab 2001, 281:E782-788.
24. Balbo SL, Ribeiro RA, Mendes MC, Lubaczeuski C, Maller AC, Carneiro EM,
Bonfleur ML: Vagotomy diminishes obesity in cafeteria rats by decreasing
cholinergic potentiation of insulin release. J Physiol Biochem 2016, 72:625-633.
25. Duckworth WC, Bennett RG, Hamel FG: Insulin degradation: progress and
potential. Endocr Rev 1998, 19:608-624.
26. Groves CJ, Wiltshire S, Smedley D, Owen KR, Frayling TM, Walker M, Hitman GA,
Levy JC, O'Rahilly S, Menzel S, et al: Association and haplotype analysis of the
insulin-degrading enzyme (IDE) gene, a strong positional and biological
candidate for type 2 diabetes susceptibility. Diabetes 2003, 52:1300-1305.
27. Abdul-Hay SO, Kang D, McBride M, Li L, Zhao J, Leissring MA: Deletion of
insulin-degrading enzyme elicits antipodal, age-dependent effects on glucose and
insulin tolerance. PLoS One 2011, 6:e20818.

33
Table
Table 1: Body, fat pads and stomach weight, followed by blood glucose and plasma insulin
concentration in LP, LP+HF and LP+HFvag mice.
LP LP+HF LP+HFvag
Body Weight, BW (g) 23.6 ± 0.7b 27.6 ± 1.0
a 27.4 ± 0.7
a
Retroperitoneal fat pad (% BW) 0.6± 0.1a 1.2 ± 0.1
b 0.9 ± 0.2
b
Perigonadal fat pad (% BW) 1.5 ± 0.1a 2.3 ± 0.3
b 1.9 ± 0.3
ab
Stomach weight (% BW) 1.5 ± 0.2a 0.9 ± 0.1
a 2.5 ± 0.4
b
Serum total Protein (g/dL) 4.0 ± 0.7b 5.7 ± 0.3
a 5.4 ± 0.3
ab
Fasting glycemia (mg/dL) 55 ± 1b 70±4
a 67 ± 6
a
Fasting insulinemia (ng/mL) 0.09 ± 0.01a 0.28 ± 0.06
b 0.14 ± 0.07
a
Fed insulinemia (ng/ml) 1.62 ± 0.13a 2.56 ± 0.26
b 2.70 ± 0.24
b
Data are means ± SEM from LP (n= 6-13); LP+HF (n= 6-13); LP+HFvag (n= 5-10). Different
letters over the numbers indicate significant differences (P < 0.05).

34
Figure legends
Figure 1: Vagotomy improved glucose tolerance but not insulin sensitivity. Blood glucose
during the ipGTT (A) and its respective area under curve (AUC) (B). Blood glucose during
ipITT (C) and glucose disappearance rate (KITT) (D) in LP, LP+HF and LP+HFD mice. Data
are means ± SEM from LP (n= 11-13); LP+HF (n= 8-10); LP+HFvag (n= 5-6). The symbols
* indicate significant differences between LP and LP+HFvag compared to LP+HF; # between
LP+HF and LP+HFvag and & between LP+HF and LP+HFvag compared to LP. Different
letters over the bars indicate significant differences. ANOVA one-way followed by Tukey’s
test. P < 0.05.
Figure 2: Vagotomy reduced GSIS in isolated pancreatic islets. Glucose-stimulated insulin
secretion (A) and total insulin content (B) of islets from LP, LP+HF and LP+HFvag mice.
Data are means ± SEM from LP (n= 13); LP+HF (n= 8); LP+HFvag (n= 8). Different letters
over the bars indicate significant differences. ANOVA one-way followed by Tukey’s test. P <
0.05.
Figure 3: Vagotomy reduced insulin clearance. Plasma concentration of insulin (A), c-
peptide (B) and c-peptide:insulin ratio (C) at 0, 15, and 60 min during ipGTT. Data are means
± SEM from LP (n= 3-6 ) LP+HF (n= 3-5 ) LP+HFvag (n= 4-6 ). Different letters over the
bars indicate significant differences. ANOVA one-way followed by Tukey’s test. P < 0.05.
Figure 4: Vagotomy reduced IDE expression in the liver of LP mice. IDE protein
expression in the liver of LP, LP+HF and LP+HFvag mice. Data are means ± SEM from LP
(n= 6); LP+HF (n= 5); LP+HFvag (n= 4). Different letters over the bars indicate significant
differences. ANOVA one-way followed by Tukey’s test. P < 0.05.
Supplemental data
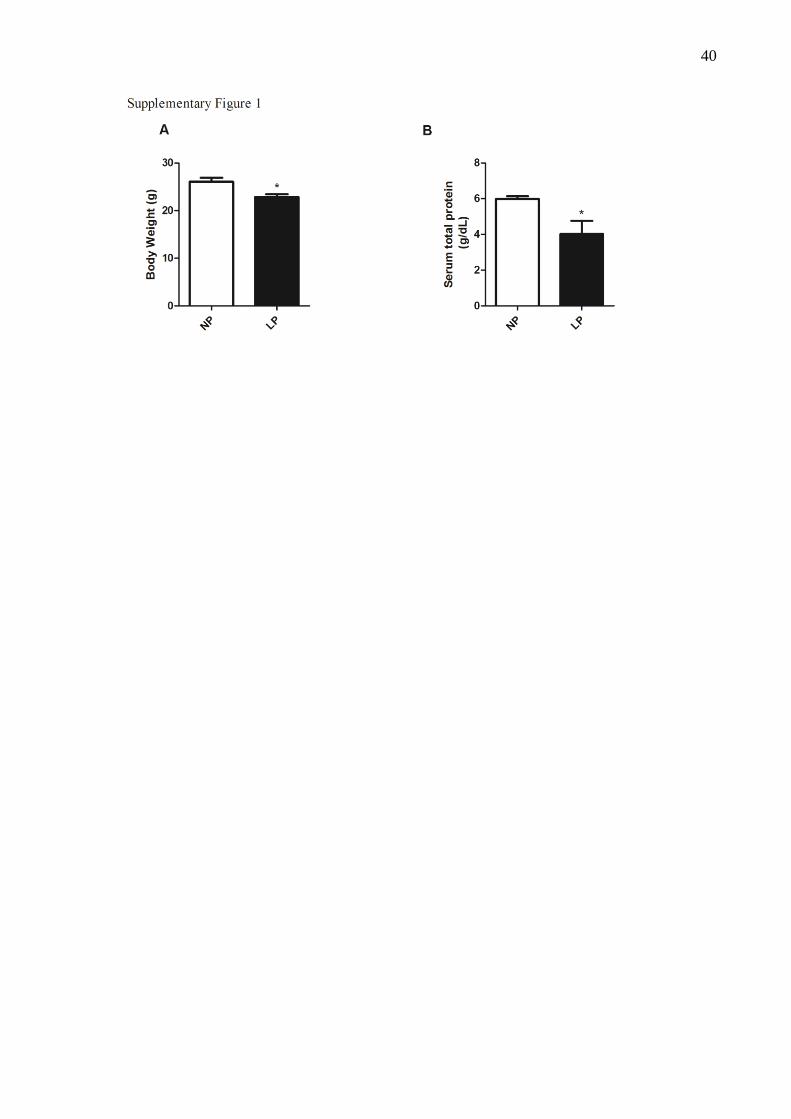
Supplemental figure 1: Body Weight (A) and serum total protein (B) , of NP an LP groups.
Data are means ± SEM from n = 6-8. The symbol * indicate significant differences (P < 0.05).
Student’s t-test.
Supplemental figure 2: Body Weight (A), perigonadal (B) and retroperitoneal fat pad (C), of
NP, NP+HF and NP+HFvag groups. Data are means ± SEM from n = 6-8. Different letters
over the bars indicate significant differences (P < 0.05). ANOVA one-way followed by
Tukey’s test.

35
Supplemental figure 3: Glucose tolerance and insulin sensitivity evaluated by ipGTT (A),
AUC of piGTT (B), ipiTT (C), kITT (D) in NP, NP+HF and NP+HFvag mice. Data are
means ± SEM from n = 6-20. Different letters over the numbers indicate significant
differences (P < 0.05). ANOVA one-way followed by Tukey’s test.
36
Figure 1
Figure 2
37
Figure 3
38
Figure 4

39
40

41

42

43
5. Artigo 2
VAGOTOMY REDUCES GLUCAGON SECRETION IN
MALNOURISHED MICE SUBMITTED TO HIGH FAT DIET
Camila Lubaczeuski1, Jean Franciesco Vettorazzi
1, Luciana Mateus Gonçalves
1, Maria Lucia
Bonfleur2, Antonio Carlos Boschero
1, José Maria Costa-Junior
1 and Everardo Magalhães
Carneiro1*
1 - Department of Structural and Functional Biology, Institute of Biology, University of
Campinas (UNICAMP), 13083-970 Campinas, SP, Brazil
2 - Human Physiology Division, Center for Biological Sciences and Health, UNIOESTE,
Cascavel-PR, Brazil.
*Corresponding author: Everardo Magalhães Carneiro, Department of Structural and
Functional Biology, Institute of Biology, University of Campinas (UNICAMP), Monteiro
Lobato Street, 13083-970, Campinas, SP, Brazil. Phone: +55 1935216203.
Email: [email protected].

44
Abstract
The goal of this study was to investigate the effect of vagotomy on glucagon secretion and
glucose output in metabolic programmed mice, induced by a low-protein diet in early life,
followed by exposure to obesity by high fat diet (HF) in adulthood. After weaning, C57BL/6
mice were received low protein diet with 6 % of protein, composing LP group. After 4 weeks,
LP group was divided in LP+HF, which started to receive high-fat diet (HF) (35% of lipids),
for 8 weeks, and LP+HFvag, was submitted to the vagotomy procedure at the same time as
the diet was offered. We evaluated ipGTT, ipITT; glucose release by glucagon and pyruvate;
glucose-stimulated glucagon secretion in vivo and in isolated islets; hepatic glycogen content.
Metabolic programmed mice showed impaired glucose homeostasis, as a result of
hyperglucagonemia. Furthermore, it showed disruption on glucagon suppression in response
to higher glucose, both in vivo and isolated islets. The vagotomy improved glucose tolerance,
without modifying adiposity and insulin sensitivity. The glucose tolerance was according to
reduced glucagon secretion both in vivo and in isolated islets. The hepatic glucose output,
after a glucagon and pyruvate challenge, was higher in metabolic programmed mice compared
to malnourished mice, and the vagotomy does not alter this. Hepatic glycogen content
analysis showed lower amount in metabolic programmed mice compared to LP mice, without
alterations after vagotomy. In conclusion, metabolic programming induces glucose
intolerance associated with an impaired glucagon-induced glucose suppression in-vivo, and in
isolated pancreatic islets. The vagotomy reduced glucagonemia and ameliorated glucagon
suppression by glucose, being efficient to the improvement of glucose tolerance.
5.1 Introduction
Protein malnutrition during the perinatal period of life is associated with increased risk
of obesity, insulin resistance, and development of type 2 diabetes mellitus (type 2 DM) later
in life. According to the thrifty phenotype hypothesis, the reduced availability of nutrients
during developmental stages favors metabolic programming that may disrupt glycemic
control after increased nutrient intake (Hales e Barker, 1992; Duque-Guimarães and Ozanne,
2013). Indeed, economic improvements in developing countries during recent decades have
placed human being into similar conditions where the intake of a normal or high-calorie diet
in adulthood increases the risk to develop metabolic diseases, after a period of calorie
restriction early in life (Sandovici et al., 2013; Abdullah, 2015).
The pancreatic islet dysfunction appers during metabolic programming in humans

45
(Hales and Barker, 1992), and rodents (Malta et al., 2014; Leite et al., 2016). This
phenomenon occurs through malnourished inheritance and also during postnatal low protein
intake, if these individuals or rodents were exposure to a regular or high fat diet at adulthood
(Malta et al., 2014; Leite et al., 2016). In this context, the majority of the studies have been
focused on the effect of metabolic programming upon pancreatic beta cell function and insulin
homeostasis, where the beta cell disruption is triggered by increased insulin secretion,
mitochondrial dysfunction (Leite et al., 2016) and parasympathetic nervous system over-
activation (Malta et al., 2014).
The islet of Langerhans is highly innervated by parasympathetic and sympathetic
nerves (Ahrén, 2000). The increased vagal activity is associated with pancreatic beta cell
dysfunction during obesity (Balbo et al., 2000; Barella et al., 2015). In fact, we demonstrated
that the vagotomy procedure was efficient to abrogate the glucose intolerance of metabolic
programed mice (Lubaczeuski, et al., submitted data). However, the involvement of
pancreatic α-cell, as well as the implications of parasympathetic denervation on glucagon
homeostasis in metabolic programed mice has not been studied yet.
Thus, we sought to evalute the effect of parasympathetic nervous system on glucagon
homeostasis in protein malnourished mice exposed to high fat diet, through subdiaphargmatic
vagotomy.
5.2 Material and Methods
Animals
All the experimental animals were carried out in accordance with the protocols approved by
the Animal Care and Use Committee of University of Campinas (UNICAMP) (approval
number: 3379-1). Male C57Bl/6 mice were obtained from UNICAMP and maintained at 22 ±
1° C in a 12-h light–dark cycle. Thirty-day-old mice were fed on normal protein diet (14%
protein) (NP group) or low protein diet (6% protein) (LP group) during 4 weeks. After, LP
mice were distributed into three groups: LP, which was kept with low protein diet; LP+HF,
which started to receive a high fat diet (35% fat) during 8 weeks; and LP+HFvag which was
submitted to vagotomy and also started to receive a high fat diet during 8 weeks. The diets
compositions were described in a previous study (Batista et al., 2013) and manipulated by
Prag Soluções Biociências (Jaú – Brazil).

46
Subdiaphragmatic vagotomy procedure
After for 4 weeks receiveing low protein diet, LP+HF mice were submitted to
subdiaphragmatic truncal vagotomy (LP+HFvag group) or sham operation (LP+HF). For this
procedure, 12-h fasted mice were anesthetized with a mixture of ketamine and xylazine (0.06
and 0.02 mg/g via i.p., respectively; Vetbrands®, Paulínia, SP, BRA). Subsequently, the
stomach and esophagus were exteriorized from the peritoneal cavity, and both dorsal and
subdiafragmatic vagal trunk was separated from the esophagus and cut off. Sham-operation
mice were undergone to the same procedures, but the vagus nerve was kept intact.
Intraperitoneal glucose (ipGTT), insulin (ipITT), glucagon (ipGlTT), and pyruvate (ipPTT)
tolerance tests
At the end of treatment, the mice were subjected to 12-h fasting to perform the ipGTT. The
fasting blood glucose level were measured by a glucometer (time 0). Then the mice received
an i.p. glucose load of 2 g/kg body weight and the glycemia was measured at 15, 30, 60 and
120 min after the glucose load. For glucagon measurements, the blood was collected at 0 and
30 min before glucose load. For the ipITT, the mice were subjected to a 2-h fasting and the
glycemia was measured before the i.p. administration of 1 U/kg insulin load (Humulin® R,
Lilly, Indianapolis, USA) and at 3, 6, 9, 12, 15 and 18 min after the application. For ipGluTT,
the glycemia was measured after 12 h fasting, then the mice received an i.p. glucagon load
(GlucaGen® HypoKit, Novo Nordisk, Gentofte, Dinamarca) of 100 μg/kg body weight, and
the glycemia was measured again at 15, 20, 40, 60 min. For ipPTT, the fasted mice (14-16 h)
were injected with 2 g/kg BW of sodium pyruvate (Merck Millipore; Darmstadt, Germany).
Blood glucose was determined before in fasting (time 0) and 15, 30, and 60 min after
pyruvate injection.
Corporal parameters
At 120 days of age, after 12h-fasting, all mice were weighed and the glucose was
measured using a glucose analyzer (Accu-Chek Performa, Roche Diagnostic, USA).
Subsequently, the mice were euthanized by decapitation and the total blood samples were
collected in heparinized tubes. Plasma was used for insulin or glucagon measurement by RIA
and the perigonadal and retroperitoneal fat pads were collected and weighed.

47
Islet isolation and Glucose induced glucagon secretion
Islets were isolated by collagenase digestion of the pancreas, as described (Boschero, et al.
1995). For static glucagon incubations, groups of 15 islets were preincubated for 1 hr at 37 °C
with 500 μM of Krebs buffer (KBB) with the following composition: 115 mM NaCl, 5 mM
KCl, 2.56 mM CaCl2, 1 mM MgCl2, 10 mM NaHCO3, 15 mM HEPES, supplemented with
0.5 mM glucose, 3 g bovine serum albumin (BSA) per liter, and equilibrated with a mixture of
95% O2-5% CO2 to provide pH 7.4. This medium was then replaced with fresh buffer, and
the islets were incubated for 1 hour with 300uL of KBB containing 0.5 or 11.1 mM glucose.
At the end of the incubation period, the supernatants were collected and maintained at –20 °C.
The glucagon was measured by RIA using human glucagon radiolabelled with 125
I as tracer
(GL-32K, Millipore Sigma, St. Charles, Missouri, USA).
Liver Glycogen content
Liver samples (100mg) were hydrolyzed in 500 ul of 30% KOH (1h at 98 ° C), and then the
glycogen was precipitated with 50 μl of Na2SO4 and 1.5 ml of absolute alcohol (15 sec at 98 °
C). The supernatant was discarded and the pellet was washed three times with 500 μl of
distilled water (60°C) and 1.5 ml of alcohol was added. The glycogen concentration was
determined through a colorimetric reaction, adding 15 μl phenol, 2 mL H2SO4, distilled water
and boiled for 15 min in the water bath. Then absorbance was read at 490 nm.
Statistical Analysis
The data are presented as the means ± standard error of mean (SEM), and the differences were
considered significant when p < 0.05. Comparisons were performed using ANOVA one-way
followed by Tukey’s test or between NP and LP using t-test. Tests were carried out using
GraphPad Prism, version 5.0 for Windows (GraphPad Software, Inc., San Diego, CA, USA).
5.3 Results
Model characterization
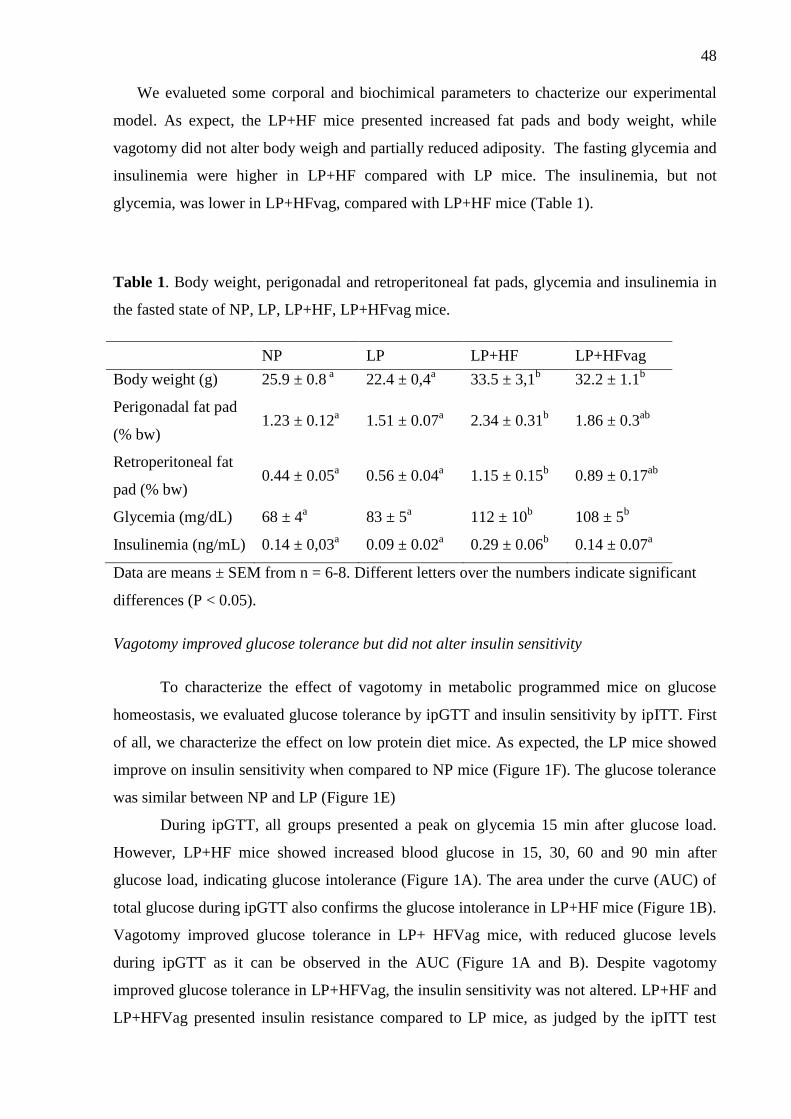
48
We evalueted some corporal and biochimical parameters to chacterize our experimental
model. As expect, the LP+HF mice presented increased fat pads and body weight, while
vagotomy did not alter body weigh and partially reduced adiposity. The fasting glycemia and
insulinemia were higher in LP+HF compared with LP mice. The insulinemia, but not
glycemia, was lower in LP+HFvag, compared with LP+HF mice (Table 1).
Table 1. Body weight, perigonadal and retroperitoneal fat pads, glycemia and insulinemia in
the fasted state of NP, LP, LP+HF, LP+HFvag mice.
NP LP LP+HF LP+HFvag
Body weight (g) 25.9 ± 0.8 a 22.4 ± 0,4
a 33.5 ± 3,1
b 32.2 ± 1.1
b
Perigonadal fat pad
(% bw) 1.23 ± 0.12
a 1.51 ± 0.07
a 2.34 ± 0.31
b 1.86 ± 0.3
ab
Retroperitoneal fat
pad (% bw) 0.44 ± 0.05
a 0.56 ± 0.04
a 1.15 ± 0.15
b 0.89 ± 0.17
ab
Glycemia (mg/dL) 68 ± 4a 83 ± 5
a 112 ± 10
b 108 ± 5
b
Insulinemia (ng/mL) 0.14 ± 0,03a 0.09 ± 0.02
a 0.29 ± 0.06
b 0.14 ± 0.07
a
Data are means ± SEM from n = 6-8. Different letters over the numbers indicate significant
differences (P < 0.05).
Vagotomy improved glucose tolerance but did not alter insulin sensitivity
To characterize the effect of vagotomy in metabolic programmed mice on glucose
homeostasis, we evaluated glucose tolerance by ipGTT and insulin sensitivity by ipITT. First
of all, we characterize the effect on low protein diet mice. As expected, the LP mice showed
improve on insulin sensitivity when compared to NP mice (Figure 1F). The glucose tolerance
was similar between NP and LP (Figure 1E)
During ipGTT, all groups presented a peak on glycemia 15 min after glucose load.
However, LP+HF mice showed increased blood glucose in 15, 30, 60 and 90 min after
glucose load, indicating glucose intolerance (Figure 1A). The area under the curve (AUC) of
total glucose during ipGTT also confirms the glucose intolerance in LP+HF mice (Figure 1B).
Vagotomy improved glucose tolerance in LP+ HFVag mice, with reduced glucose levels
during ipGTT as it can be observed in the AUC (Figure 1A and B). Despite vagotomy
improved glucose tolerance in LP+HFVag, the insulin sensitivity was not altered. LP+HF and
LP+HFVag presented insulin resistance compared to LP mice, as judged by the ipITT test
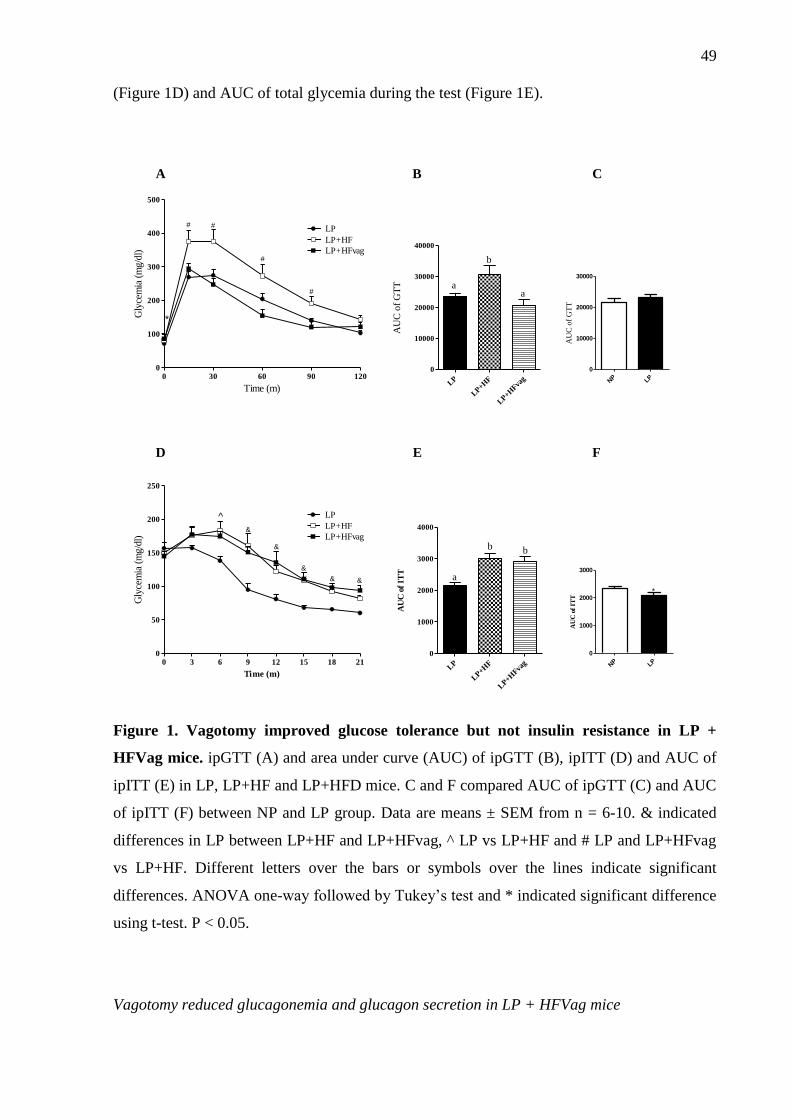
49
(Figure 1D) and AUC of total glycemia during the test (Figure 1E).
0 30 60 90 1200
100
200
300
400
500
LP
LP+HF
LP+HFvag
*
# #
#
#
Time (m)
Gly
cem
ia (
mg/d
l)
LP
LP+H
F
LP+H
Fvag
0
10000
20000
30000
40000
a
b
a
AU
C o
f G
TT
A B
0 3 6 9 12 15 18 210
50
100
150
200
250
LP
LP+HF
LP+HFvag
^
&
&
&
& &
Time (m)
Gly
cem
ia (
mg/d
l)
D
LP
LP+H
F
LP+H
Fvag
0
1000
2000
3000
4000
a
b b
AU
C o
f IT
T
E
NP LP
0
10000
20000
30000
AU
C o
f G
TT
C
NP LP
0
1000
2000
3000
*
AU
C o
f IT
T
F
Figure 1. Vagotomy improved glucose tolerance but not insulin resistance in LP +
HFVag mice. ipGTT (A) and area under curve (AUC) of ipGTT (B), ipITT (D) and AUC of
ipITT (E) in LP, LP+HF and LP+HFD mice. C and F compared AUC of ipGTT (C) and AUC
of ipITT (F) between NP and LP group. Data are means ± SEM from n = 6-10. & indicated
differences in LP between LP+HF and LP+HFvag, ^ LP vs LP+HF and # LP and LP+HFvag
vs LP+HF. Different letters over the bars or symbols over the lines indicate significant
differences. ANOVA one-way followed by Tukey’s test and * indicated significant difference
using t-test. P < 0.05.
Vagotomy reduced glucagonemia and glucagon secretion in LP + HFVag mice
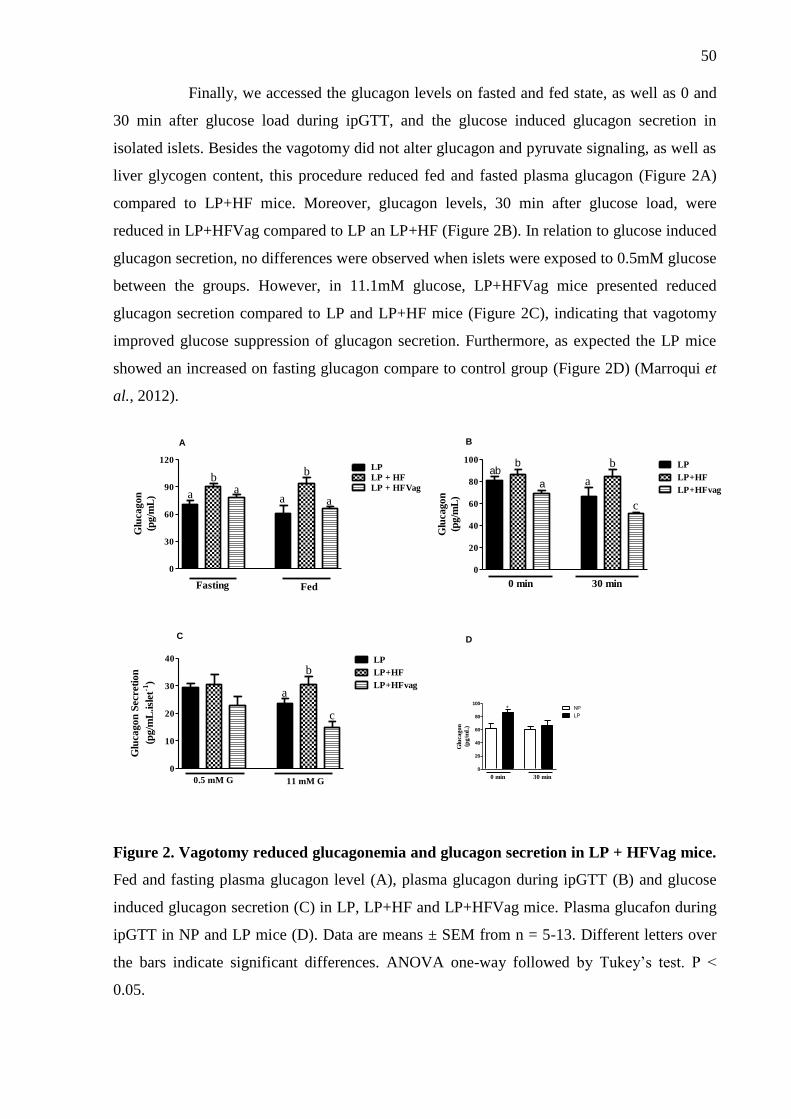
50
Finally, we accessed the glucagon levels on fasted and fed state, as well as 0 and
30 min after glucose load during ipGTT, and the glucose induced glucagon secretion in
isolated islets. Besides the vagotomy did not alter glucagon and pyruvate signaling, as well as
liver glycogen content, this procedure reduced fed and fasted plasma glucagon (Figure 2A)
compared to LP+HF mice. Moreover, glucagon levels, 30 min after glucose load, were
reduced in LP+HFVag compared to LP an LP+HF (Figure 2B). In relation to glucose induced
glucagon secretion, no differences were observed when islets were exposed to 0.5mM glucose
between the groups. However, in 11.1mM glucose, LP+HFVag mice presented reduced
glucagon secretion compared to LP and LP+HF mice (Figure 2C), indicating that vagotomy
improved glucose suppression of glucagon secretion. Furthermore, as expected the LP mice
showed an increased on fasting glucagon compare to control group (Figure 2D) (Marroqui et
al., 2012).
0
20
40
60
80
100LP
LP+HF
LP+HFvag
0 min 30 min
a
b
c
abb
a
Glu
cagon
(pg/m
L)
0
30
60
90
120LP
LP + HF
LP + HFVaga a
b
a a
b
Fasting Fed
Glu
cagon
(pg/m
L)
0
10
20
30
40 LP
LP+HF
LP+HFvag
0.5 mM G 11 mM G
a
b
c
Glu
cago
n S
ecre
tion
(pg/m
L.isl
et-1
)
A B
C
0
20
40
60
80
100
0 min 30 min
NP
LP
*
Glu
cagon
(pg/m
L)
D
Figure 2. Vagotomy reduced glucagonemia and glucagon secretion in LP + HFVag mice.
Fed and fasting plasma glucagon level (A), plasma glucagon during ipGTT (B) and glucose
induced glucagon secretion (C) in LP, LP+HF and LP+HFVag mice. Plasma glucafon during
ipGTT in NP and LP mice (D). Data are means ± SEM from n = 5-13. Different letters over
the bars indicate significant differences. ANOVA one-way followed by Tukey’s test. P <
0.05.
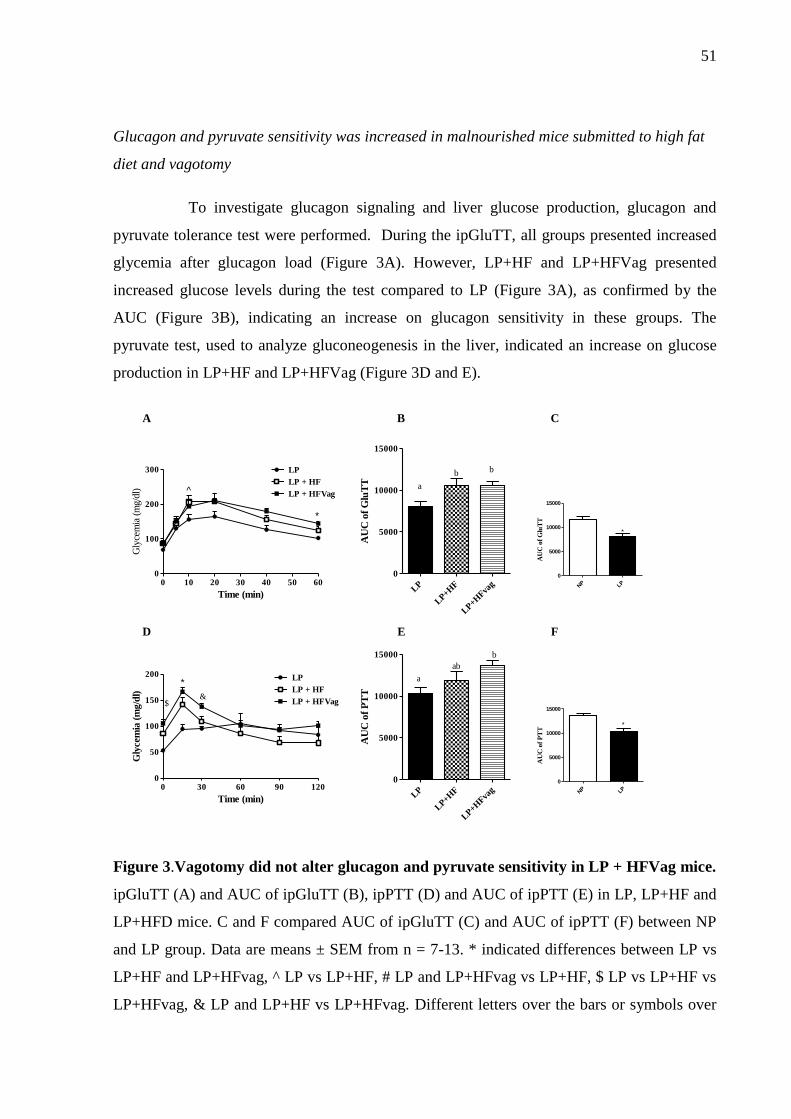
51
Glucagon and pyruvate sensitivity was increased in malnourished mice submitted to high fat
diet and vagotomy
To investigate glucagon signaling and liver glucose production, glucagon and
pyruvate tolerance test were performed. During the ipGluTT, all groups presented increased
glycemia after glucagon load (Figure 3A). However, LP+HF and LP+HFVag presented
increased glucose levels during the test compared to LP (Figure 3A), as confirmed by the
AUC (Figure 3B), indicating an increase on glucagon sensitivity in these groups. The
pyruvate test, used to analyze gluconeogenesis in the liver, indicated an increase on glucose
production in LP+HF and LP+HFVag (Figure 3D and E).
0 10 20 30 40 50 600
100
200
300 LP
LP + HF
LP + HFVag^
*
Time (min)
Gly
cem
ia (
mg/d
l)
LP
LP+H
F
LP+H
Fvag
0
5000
10000
15000
a
b b
AU
C o
f G
luT
T
0 30 60 90 1200
50
100
150
200 LP
LP + HF
LP + HFVag$
*
&
Time (min)
Gly
cem
ia (
mg/d
l)
LP
LP+H
F
LP+H
Fvag
0
5000
10000
15000
a
ab
b
AU
C o
f P
TT
A B
D E
NP LP
0
5000
10000
15000
*A
UC
of
Glu
TT
NP LP
0
5000
10000
15000
*
AU
C o
f P
TT
C
F
Figure 3.Vagotomy did not alter glucagon and pyruvate sensitivity in LP + HFVag mice.
ipGluTT (A) and AUC of ipGluTT (B), ipPTT (D) and AUC of ipPTT (E) in LP, LP+HF and
LP+HFD mice. C and F compared AUC of ipGluTT (C) and AUC of ipPTT (F) between NP
and LP group. Data are means ± SEM from n = 7-13. * indicated differences between LP vs
LP+HF and LP+HFvag, ^ LP vs LP+HF, # LP and LP+HFvag vs LP+HF, $ LP vs LP+HF vs
LP+HFvag, & LP and LP+HF vs LP+HFvag. Different letters over the bars or symbols over
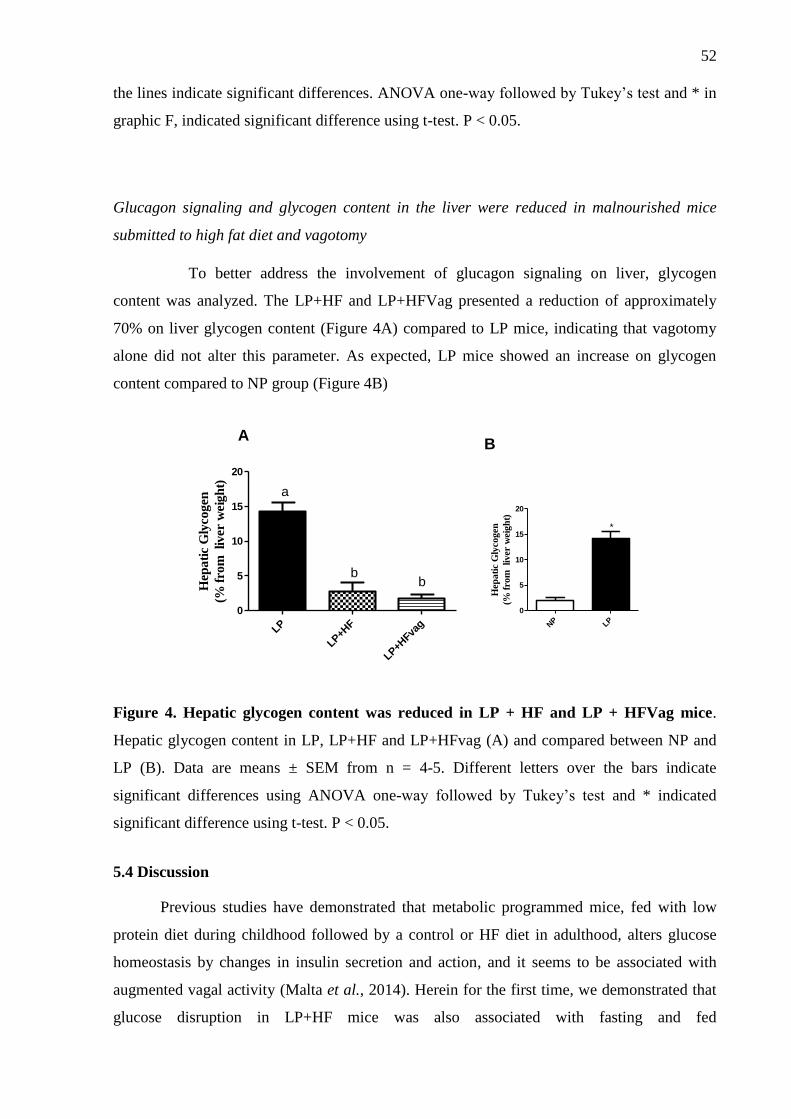
52
the lines indicate significant differences. ANOVA one-way followed by Tukey’s test and * in
graphic F, indicated significant difference using t-test. P < 0.05.
Glucagon signaling and glycogen content in the liver were reduced in malnourished mice
submitted to high fat diet and vagotomy
To better address the involvement of glucagon signaling on liver, glycogen
content was analyzed. The LP+HF and LP+HFVag presented a reduction of approximately
70% on liver glycogen content (Figure 4A) compared to LP mice, indicating that vagotomy
alone did not alter this parameter. As expected, LP mice showed an increase on glycogen
content compared to NP group (Figure 4B)
LP
LP+H
F
LP+H
Fvag
0
5
10
15
20
a
bbH
epa
tic
Gly
cogen
(% f
rom
liver
wei
gh
t)
NP LP
0
5
10
15
20
*
Hep
ati
c G
lyco
gen
(% f
rom
liv
er w
eig
ht)
AB
Figure 4. Hepatic glycogen content was reduced in LP + HF and LP + HFVag mice.
Hepatic glycogen content in LP, LP+HF and LP+HFvag (A) and compared between NP and
LP (B). Data are means ± SEM from n = 4-5. Different letters over the bars indicate
significant differences using ANOVA one-way followed by Tukey’s test and * indicated
significant difference using t-test. P < 0.05.
5.4 Discussion
Previous studies have demonstrated that metabolic programmed mice, fed with low
protein diet during childhood followed by a control or HF diet in adulthood, alters glucose
homeostasis by changes in insulin secretion and action, and it seems to be associated with
augmented vagal activity (Malta et al., 2014). Herein for the first time, we demonstrated that
glucose disruption in LP+HF mice was also associated with fasting and fed

53
hyperglucagonemia and glucagon hypersecretion in response to glucose. These alterations are
due, in part, to parasympathetic nervous system, since truncal vagotomy normalizes
hyperglucagonemia and reduces glucagon secretion in the presence of glucose.
First, we characterized our model of LP. As described in the literature, LP mice
present enhanced peripheral insulin sensitivity, increase on glucagon secretion and glycogen
content, when compared to NP mice (Marroqui et al., 2012).
Our previous study showed that vagotomy procedure might be helpful to correct
glucose intolerance from metabolic programmed mice, increasing insulinemia through a
reduction in insulin clearance (Lubaczeuski et al., submitted data).
In the same way than insulin, glucagon secretion and action is also impaired by obesity
(Borghi et al., 1984; Santos-Silva et al., 2015). Although glucose concentrations fluctuations
are the main regulator of glucagon secretion, other factors, as autonomic innervation, also
impact the α -cells hormone release. Parasympathetic and sympathetic tonus work together to
increase glucagon secretion in order to reestablish normal glucose level during a
hypoglycemic challenge. The efficiency of such system is necessary to provide enough
glucose to the brain. Obese mammals present parasympathetic over-activation, which in turn,
may exacerbates their high plasmatic glucagon concentration (Larsson and Ahrén, 2000;
Kahn et al., 2006; Zhang et al., 2013; Santos-Silva et al., 2015). However, the role of vagus
nerve has not been explored in the context of metabolic programming focusing on glucagon
homeostasis.
We characterized our model of metabolic programming, since the LP+HF mice
presented higher body weight, fat pads storage, insulinemia and insulin resistance, resulting in
hyperglycemia and glucose tolerance compared to LP mice. In this sense, we performed a
vagotomy surgery in an attempt to improve the glycemic control of these obese mice. As
expected, LP+HFvag improved glucose tolerance without alterations in insulin sensitivity. In
fact, vagotomy seems to improve glucose-insulin homeostasis of metabolic programmed mice
by decreasing insulin clearance through decreased liver IDE expression (Lubaczeuski et al.,
submitted data).
In terms of glucagon secretion, the LP+HF mice also demonstrated a metabolic
syndrome profile, and this might occur due to a disruption in α-cell responsiveness to glucose,
since LP+HF islets hypersecrete glucagon at 0.5 and 11.1 mM of glucose compared to LP
mice. Also, no suppression of glucagon release in these mice was observed after an ip
injection of glucose, an effect that closely matches to those observed during T2D (Zhang et
al., 2013).

54
The vagotomy reduced glucagonemia in LP+HF mice and induced a reduction in
glucagon secretion in response to glucose, contributing to improve glucose tolerance in these
group. Our findings make sense with the paradigm that vagal tonus contributes to increase
glucagon secretion during fast and fed state, once we observed diminished glucagon secretion
in both, low and high glucose concentration in-vivo and in-vitro experiments (Plamboeck et
al., 2013; Wewer Albrechtsen et al., 2014).
We suspected that some other hormone could be differentially secreted after
vagotomy, driving such phenomenon. In this sense the glucagon like peptide -1 (GLP-1)
emerges as a potential candidate, due to its action upon glucagon suppression, and by the fact
that vagotomized humans present higher GLP-1 secretion (Plamboeck et al., 2013). GLP-1
may bind to GLP-1 receptor (GLP1R) in α- cells membrane, increasing the intracellular cyclic
adenosine monophosphate (cAMP) content, which in turn increases protein kinase A (PKA)
activation, that augments the phosphorylation of N-type Ca2+
channels resulting in a strong
inhibition of their activity and a reduction in exocytosis (De Marinis et al., 2010). However,
further experiments are necessary to prove that similar mechanism could be acting in our
vagotomized model of metabolic programed mice.
Glucagon actions are mainly performed in the liver, stimulating gluconeogenesis and
glycogenolysis through the increment of hepatic glucose production and release (Quesada et
al., 2008; Ramnanan et al., 2011). In this sense, we measured the capacity of hepatic glucose
output after a pyruvate and glucagon challenge, and as expected, the LP+HF mice presented
higher glucose after these challenges and vagotomy did not improve it. We speculate that
vagal inhibition was unable to reestablish these hepatic parameters in metabolic programmed
mice because this surgery may induce a hyper-sympathetic activation, increasing
glyconeogeneses.
Keeping it in mind, we evaluated hepatic glycogen content. We observed that LP+HF,
as well as LP+HFvag mice, decreased their hepatic glycogen storage compared to LP mice.
Such phenomenon could be attributed to a possible higher catecholamine release due to vagal
inhibition or to a hepatic insulin resistance. However, we need to deep into investigations to
conclude such statement.
In conclusion, metabolic programming induces glucose intolerance associated with a
damaged on glucagon-induced glucose suppression in-vivo, and in isolated pancreatic islets.
The vagotomy reduced glucagon secretion and ameliorated glucagon suppression by glucose,
being efficient to the improvement of the glucose tolerance. Although, other hormones, as

55
GLP-1, seem to be involved. Further studies are necessary to figure out which mechanism are
implicated in vagal control of glucagon secretion in metabolic programmed mice.
Acknowledgements
We thank M. Carlenossi for her expert technical assistance. This work was supported by
grants from Fundação de Amparo à Pesquisa do Estado de São Paulo (FAPESP –
2013/25847-6), Conselho Nacional para o Desenvolvimento Científico e Tecnológico (CNPq
200030/2014-0), Instituto Nacional de Obesidade e Diabetes (CNPq/FAPESP) and
Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Conflict of interest
All contributing authors report no conflict of interest.

56
List of Figures
Figure 1: Vagotomy improves glucose tolerance but not insulin resistance in LP +
HFVag mice. ipGTT (A) and area under curve (AUC) of ipGTT (B), ipITT (D) and AUC of
ipITT (E) in LP, LP+HF and LP+HFD mice. C and F compared AUC of ipGTT (C) and AUC
of ipITT (F) between NP and LP group. Data are means ± SEM from n = 6-10. & indicated
differences in LP between LP+HF and LP+HFvag, ^ LP vs LP+HF and # LP and LP+HFvag
vs LP+HF. Different letters over the bars or symbols over the lines indicate significant
differences. ANOVA one-way followed by Tukey’s test and * indicated significant difference
using t-test. P < 0.05.
Figure 2: Vagotomy reduces glucagonemia and glucagon secretion in LP + HFVag mice.
Fed and fasting plasma glucagon level (A), plasma glucagon during ipGTT (B) and glucose
induced glucagon secretion (C) in LP, LP + HF and LP + HFVag mice. Data are means ±
SEM from n = 5-13. Different letters over the bars indicate significant differences. ANOVA
one-way followed by Tukey’s test. P < 0.05.
Figure 3: Vagotomy did not alter glucagon and pyruvate sensitivity in LP + HFVag
mice. ipGluTT (A) and AUC of ipGluTT (B), ipPTT (C) and AUC of ipPTT (D) in LP,
LP+HF and LP+HFD mice. C and F compared AUC of ipGluTT (C) and AUC of ipPTT (F)
between NP and LP group. Data are means ± SEM from n = 5-6. Data are means ± SEM from
n = 5-13.* indicated differences in LP between LP+HF and LP+HFvag, ^ LP vs LP+HF, # LP
and LP+HFvag vs LP+HF, $ LP vs LP+HF vs LP+HFvag, & LP and LP+HF vs LP+HFvag.
Different letters over the bars or symbols over the lines indicate significant differences.
ANOVA one-way followed by Tukey’s test and * indicated significant difference using t-test.
P < 0.05.
Figure 4. The hepatic glycogen content is reduced in LP + HF and LP + HFVag mice.
Hepatic glycogen content (A), obtained as the % of liver weight in LP, LP + HF and LP +

57
HFVag mice, and Hepatic glycogen content compared between NP and LP (B). Data are
means ± SEM from n = 5-13. Different letters over the bars indicate significant differences.
ANOVA one-way followed by Tukey’s test. P < 0.05.

58
5.5 References
ABDULLAH, A. The Double Burden of Undernutrition and Overnutrition in Developing
Countries: an Update. Curr Obes Rep, v. 4, n. 3, p. 337-49, Sep 2015. ISSN 2162-4968.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26627492 >.
AHRÉN, B. Autonomic regulation of islet hormone secretion--implications for health and
disease. Diabetologia, v. 43, n. 4, p. 393-410, Apr 2000. ISSN 0012-186X. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/10819232 >.
BALBO, S. L. et al. Vagotomy reduces obesity in MSG-treated rats. Res Commun Mol
Pathol Pharmacol, v. 108, n. 5-6, p. 291-6, 2000 Nov-Dec 2000. ISSN 1078-0297.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/11958282 >.
BARELLA, L. F. et al. Vagus nerve contributes to metabolic syndrome in high-fat diet-fed
young and adult rats. Exp Physiol, v. 100, n. 1, p. 57-68, Jan 2015. ISSN 1469-445X.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25398717 >.
BATISTA, T. M. et al. Taurine supplementation improves liver glucose control in normal
protein and malnourished mice fed a high-fat diet. Mol Nutr Food Res, v. 57, n. 3, p. 423-34,
Mar 2013. ISSN 1613-4133. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/23280999 >.
BORGHI, V. C.; WAJCHENBERG, B. L.; CESAR, F. P. Plasma glucagon suppressibility
after oral glucose in obese subjects with normal and impaired glucose tolerance. Metabolism,
v. 33, n. 12, p. 1068-74, Dec 1984. ISSN 0026-0495. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/6390086 >.
DE MARINIS, Y. Z. et al. GLP-1 inhibits and adrenaline stimulates glucagon release by
differential modulation of N- and L-type Ca2+ channel-dependent exocytosis. Cell Metab, v.
11, n. 6, p. 543-53, Jun 2010. ISSN 1932-7420. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/20519125 >.
DUQUE-GUIMARÃES, D. E.; OZANNE, S. E. Nutritional programming of insulin
resistance: causes and consequences. Trends Endocrinol Metab, v. 24, n. 10, p. 525-35, Oct
2013. ISSN 1879-3061. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23791137 >.
EndNote. 2013. Disponível em: <
http://www.myendnoteweb.com/EndNoteWeb.html?func=downloadInstallers&amp; >.
HALES, C. N.; BARKER, D. J. Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty
phenotype hypothesis. Diabetologia, v. 35, n. 7, p. 595-601, Jul 1992. ISSN 0012-186X.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/1644236 >.

59
KAHN, S. E.; HULL, R. L.; UTZSCHNEIDER, K. M. Mechanisms linking obesity to insulin
resistance and type 2 diabetes. Nature, v. 444, n. 7121, p. 840-6, Dec 2006. ISSN 1476-4687.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/17167471 >.
LARSSON, H.; AHRÉN, B. Glucose intolerance is predicted by low insulin secretion and
high glucagon secretion: outcome of a prospective study in postmenopausal Caucasian
women. Diabetologia, v. 43, n. 2, p. 194-202, Feb 2000. ISSN 0012-186X. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/10753041 >.
LEITE, N. C. et al. Protein malnutrition potentiates the amplifying pathway of insulin
secretion in adult obese mice. Sci Rep, v. 6, p. 33464, Sep 2016. ISSN 2045-2322. Disponível
em: < https://www.ncbi.nlm.nih.gov/pubmed/27633083 >.
MALTA, A. et al. Low-protein diet in adult male rats has long-term effects on metabolism. J
Endocrinol, v. 221, n. 2, p. 285-95, May 2014. ISSN 1479-6805. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/24599936 >.
MARROQUI, L. et al. Functional and structural adaptations in the pancreatic alpha-cell and
changes in glucagon signaling during protein malnutrition. Endocrinology, v. 153, n. 4, p.
1663-72, Apr 2012. ISSN 1945-7170 (Electronic)
0013-7227 (Linking). Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/22334714 >.
PLAMBOECK, A. et al. The effect of exogenous GLP-1 on food intake is lost in male
truncally vagotomized subjects with pyloroplasty. Am J Physiol Gastrointest Liver Physiol,
v. 304, n. 12, p. G1117-27, Jun 2013. ISSN 1522-1547. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/23599045 >.
QUESADA, I. et al. Physiology of the pancreatic alpha-cell and glucagon secretion: role in
glucose homeostasis and diabetes. J Endocrinol, v. 199, n. 1, p. 5-19, Oct 2008. ISSN 1479-
6805. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18669612 >.
RAMNANAN, C. J. et al. Physiologic action of glucagon on liver glucose metabolism.
Diabetes Obes Metab, v. 13 Suppl 1, p. 118-25, Oct 2011. ISSN 1463-1326. Disponível em:
< https://www.ncbi.nlm.nih.gov/pubmed/21824265 >.
SANDOVICI, I. et al. Developmental and environmental epigenetic programming of the
endocrine pancreas: consequences for type 2 diabetes. Cell Mol Life Sci, v. 70, n. 9, p. 1575-
95, May 2013. ISSN 1420-9071. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/23463236 >.
SANTOS-SILVA, J. C. et al. Taurine supplementation ameliorates glucose homeostasis,
prevents insulin and glucagon hypersecretion, and controls β, α, and δ-cell masses in genetic
obese mice. Amino Acids, v. 47, n. 8, p. 1533-48, Aug 2015. ISSN 1438-2199. Disponível
em: < https://www.ncbi.nlm.nih.gov/pubmed/25940922 >.

60
WEWER ALBRECHTSEN, N. J. et al. Hyperglucagonaemia analysed by glucagon sandwich
ELISA: nonspecific interference or truly elevated levels? Diabetologia, v. 57, n. 9, p. 1919-
26, Sep 2014. ISSN 1432-0428. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/24891019 >.
ZHANG, Q. et al. Role of KATP channels in glucose-regulated glucagon secretion and
impaired counterregulation in type 2 diabetes. Cell Metab, v. 18, n. 6, p. 871-82, Dec 2013.
ISSN 1932-7420. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/24315372 >.

61
6. Considerações Finais
Neste trabalho realizamos a vagotomia subdiafragmática nos camundongos que receberam
dieta hipoproteica na infância seguido da indução da obesidade na vida adulta, para avaliar o
possível papel do SNP na homeostase glicêmica, através do controle da secreção e da ação da
insulina e do glucagon. Nossos achados demonstram que os efeitos da vagotomia em animais
obesos que foram submetidos à reprogramação metabólica, não são os mesmos que ocorrem
em animais obesos que não foram submetidos a essa reprogramação. Em animais obesos, a
vagotomia reverte a obesidade, melhora a sensibilidade à insulina e a função das células β-
pancreáticas, sendo um fator protetor do desenvolvimento do DM2 (Berthoud e Jeanrenaud,
1979; Balbo et al., 2000). Por outro lado, em animais obesos que receberam dieta
hipoproteica na infância, não observamos esse fenômeno, sugerindo que a reprogramação
metabólica parece responder de maneira diferente às intervenções que visam melhorar a
obesidade.
Neste estudo, a vagotomia melhorou a tolerância à glicose em camundongos que foram
submetidos à reprogramação metabólica e promoveu aumento da insulinemia no estado pós-
prandial, devido a redução do clearance de insulina, provavelmente em função da redução da
expressão da IDE no fígado. Os animais obesos reprogramados apresentaram aumento da
secreção de glucagon, tanto in vivo como em ilhotas isoladas. A vagotomia normalizou a
secreção de glucagon nesses animais, melhorando a tolerância à glicose independente da
sensibilidade à insulina.
Em conclusão, embora a vagotomia não altere o peso corporal e a sensibilidade à insulina
em animais obesos reprogramados, a redução do tônus vagal reestabelece a tolerância a
glicose, através de dois mecanismos: aumento da insulinemia, devido à redução do clearance
de insulina, e redução da secreção de glucagon em resposta à glicose. Isto sugere a vagotomia
como uma ferramenta terapêutica no tratamento da obesidade na reprogramação metabólica.
Entretanto, é importante considerar que o aumento da insulinemia induzida pela vagotomia,
associado à resistência à insulina, pode eventualmente contribuir para o desenvolvimento do
DM2. Portanto, estudos adicionais são necessários para estabelecer a função do nevo vago em
condições de reprogramação metabólica a longo-prazo.

62
7. Referências
ABDUL-HAY, S. O. et al. Deletion of insulin-degrading enzyme elicits antipodal, age-
dependent effects on glucose and insulin tolerance. PLoS One, v. 6, n. 6, p. e20818, 2011.
ISSN 1932-6203. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/21695259 >.
ABDULLAH, A. The Double Burden of Undernutrition and Overnutrition in Developing
Countries: an Update. Curr Obes Rep, v. 4, n. 3, p. 337-49, Sep 2015. ISSN 2162-4968.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26627492 >.
AHRÉN, B. Regulation of insulin secretion by nerves and neuropeptides. Ann Acad Med
Singapore, v. 28, n. 1, p. 99-104, Jan 1999. ISSN 0304-4602. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/10374034 >.
______. Autonomic regulation of islet hormone secretion--implications for health and disease.
Diabetologia, v. 43, n. 4, p. 393-410, Apr 2000. ISSN 0012-186X. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/10819232 >.
AMARAL, A. G. et al. Leucine supplementation augments insulin secretion in pancreatic
islets of malnourished mice. Pancreas, v. 39, n. 6, p. 847-55, Aug 2010. ISSN 1536-4828.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/20697208 >.
ARAUJO, E. P. et al. Restoration of insulin secretion in pancreatic islets of protein-deficient
rats by reduced expression of insulin receptor substrate (IRS)-1 and IRS-2. J Endocrinol, v.
181, n. 1, p. 25-38, Apr 2004. ISSN 0022-0795. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/15072564 >.
BALBO, S. L. et al. Vagotomy reduces obesity in MSG-treated rats. Res Commun Mol
Pathol Pharmacol, v. 108, n. 5-6, p. 291-6, Nov-Dec 2000. ISSN 1078-0297 (Print)
1078-0297 (Linking). Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/11958282 >.
BARELLA, L. F. et al. Vagus nerve contributes to metabolic syndrome in high-fat diet-fed
young and adult rats. Exp Physiol, v. 100, n. 1, p. 57-68, Jan 2015. ISSN 1469-445X.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25398717 >.
BARKER, D. J. Intrauterine programming of adult disease. Mol Med Today, v. 1, n. 9, p.
418-23, Dec 1995. ISSN 1357-4310. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/9415190 >.
BARKER, D. J. et al. Growth in utero and serum cholesterol concentrations in adult life.
BMJ, v. 307, n. 6918, p. 1524-7, Dec 1993. ISSN 0959-8138. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/8274921 >.

63
BATISTA, T. M. et al. Taurine supplementation restores glucose and carbachol-induced
insulin secretion in islets from low-protein diet rats: involvement of Ach-M3R, Synt 1 and
SNAP-25 proteins. J Nutr Biochem, v. 23, n. 3, p. 306-12, Mar 2012. ISSN 1873-4847.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/21543213 >.
______. Taurine supplementation improves liver glucose control in normal protein and
malnourished mice fed a high-fat diet. Mol Nutr Food Res, v. 57, n. 3, p. 423-34, Mar 2013.
ISSN 1613-4133. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/23280999 >.
BERTHOUD, H. R.; JEANRENAUD, B. Acute hyperinsulinemia and its reversal by
vagotomy after lesions of the ventromedial hypothalamus in anesthetized rats.
Endocrinology, v. 105, n. 1, p. 146-51, Jul 1979. ISSN 0013-7227. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/446404 >.
BORGHI, V. C.; WAJCHENBERG, B. L.; CESAR, F. P. Plasma glucagon suppressibility
after oral glucose in obese subjects with normal and impaired glucose tolerance. Metabolism,
v. 33, n. 12, p. 1068-74, Dec 1984. ISSN 0026-0495. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/6390086 >.
BRADFORD, M. M. A rapid and sensitive method for the quantitation of microgram
quantities of protein utilizing the principle of protein-dye binding. Anal Biochem, v. 72, p.
248-54, May 1976. ISSN 0003-2697. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/942051 >.
CAMILLERI, M. et al. Intra-abdominal vagal blocking (VBLOC therapy): clinical results
with a new implantable medical device. Surgery, v. 143, n. 6, p. 723-31, Jun 2008. ISSN
1532-7361. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18549888 >.
COX, J. E.; POWLEY, T. L. Prior vagotomy blocks VMH obesity in pair-fed rats. Am J
Physiol, v. 240, n. 5, p. E573-83, May 1981. ISSN 0002-9513. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/7235009 >.
DA SILVA, P. M. et al. Preliminary report: leucine supplementation enhances glutamate
dehydrogenase expression and restores glucose-induced insulin secretion in protein-
malnourished rats. Metabolism, v. 59, n. 6, p. 911-3, Jun 2010. ISSN 1532-8600. Disponível
em: < http://www.ncbi.nlm.nih.gov/pubmed/20015523 >.
DE MARINIS, Y. Z. et al. GLP-1 inhibits and adrenaline stimulates glucagon release by
differential modulation of N- and L-type Ca2+ channel-dependent exocytosis. Cell Metab, v.
11, n. 6, p. 543-53, Jun 2010. ISSN 1932-7420. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/20519125 >.
DELGHINGARO-AUGUSTO, V. et al. A low protein diet alters gene expression in rat
pancreatic islets. J Nutr, v. 134, n. 2, p. 321-7, Feb 2004. ISSN 0022-3166. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/14747667 >.

64
DELMEIRE, D. et al. Type VIII adenylyl cyclase in rat beta cells: coincidence signal
detector/generator for glucose and GLP-1. Diabetologia, v. 46, n. 10, p. 1383-93, Oct 2003.
ISSN 0012-186X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/13680124 >.
DEPREZ-POULAIN, R. et al. Catalytic site inhibition of insulin-degrading enzyme by a
small molecule induces glucose intolerance in mice. Nat Commun, v. 6, p. 8250, Sep 2015.
ISSN 2041-1723. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/26394692 >.
DIXON, K. D. et al. Differential effects of selective vagotomy and tropisetron in aminoprivic
feeding. Am J Physiol Regul Integr Comp Physiol, v. 279, n. 3, p. R997-R1009, Sep 2000.
ISSN 0363-6119. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/10956259 >.
DUCKWORTH, W. C.; BENNETT, R. G.; HAMEL, F. G. Insulin degradation: progress and
potential. Endocr Rev, v. 19, n. 5, p. 608-24, Oct 1998. ISSN 0163-769X. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/9793760 >.
DUNNING, B. E.; GERICH, J. E. The role of alpha-cell dysregulation in fasting and
postprandial hyperglycemia in type 2 diabetes and therapeutic implications. Endocr Rev, v.
28, n. 3, p. 253-83, May 2007. ISSN 0163-769X. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/17409288 >.
DUQUE-GUIMARÃES, D. E.; OZANNE, S. E. Nutritional programming of insulin
resistance: causes and consequences. Trends Endocrinol Metab, v. 24, n. 10, p. 525-35, Oct
2013. ISSN 1879-3061. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23791137 >.
DYACHOK, O. et al. Glucose-induced cyclic AMP oscillations regulate pulsatile insulin
secretion. Cell Metab, v. 8, n. 1, p. 26-37, Jul 2008. ISSN 1932-7420. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/18590690 >.
EDVELL, A.; LINDSTRÖM, P. Vagotomy in young obese hyperglycemic mice: effects on
syndrome development and islet proliferation. Am J Physiol, v. 274, n. 6 Pt 1, p. E1034-9,
Jun 1998. ISSN 0002-9513. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/9611153
>.
EndNote. 2013. Disponível em: <
http://www.myendnoteweb.com/EndNoteWeb.html?func=downloadInstallers&amp; >.
FARRIS, W. et al. Partial loss-of-function mutations in insulin-degrading enzyme that induce
diabetes also impair degradation of amyloid beta-protein. Am J Pathol, v. 164, n. 4, p. 1425-
34, Apr 2004. ISSN 0002-9440. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/15039230 >.

65
FERREIRA, F. et al. Decreased insulin secretion in islets from rats fed a low protein diet is
associated with a reduced PKAalpha expression. J Nutr, v. 134, n. 1, p. 63-7, Jan 2004. ISSN
0022-3166. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/14704294 >.
______. Decreased cholinergic stimulation of insulin secretion by islets from rats fed a low
protein diet is associated with reduced protein kinase calpha expression. J Nutr, v. 133, n. 3,
p. 695-9, Mar 2003. ISSN 0022-3166. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/12612139 >.
FEWTRELL, M. S. et al. Effects of size at birth, gestational age and early growth in preterm
infants on glucose and insulin concentrations at 9-12 years. Diabetologia, v. 43, n. 6, p. 714-
7, Jun 2000. ISSN 0012-186X. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/10907116 >.
FILIPUTTI, E. et al. Impaired insulin secretion and decreased expression of the nutritionally
responsive ribosomal kinase protein S6K-1 in pancreatic islets from malnourished rats. Life
Sci, v. 82, n. 9-10, p. 542-8, Feb 2008. ISSN 0024-3205. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/18234235 >.
______. Augmentation of insulin secretion by leucine supplementation in malnourished rats:
possible involvement of the phosphatidylinositol 3-phosphate kinase/mammalian target
protein of rapamycin pathway. Metabolism, v. 59, n. 5, p. 635-44, May 2010. ISSN 1532-
8600. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/19913855 >.
GOTO, Y.; KAKIZAKI, M.; MASAKI, N. Production of spontaneous diabetic rats by
repetition of selective breeding. Tohoku J Exp Med, v. 119, n. 1, p. 85-90, May 1976. ISSN
0040-8727. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/951706 >.
HALES, C. N. Metabolic consequences of intrauterine growth retardation. Acta Paediatr
Suppl, v. 423, p. 184-7; discussion 188, Nov 1997. ISSN 0803-5326. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/9401569 >.
HALES, C. N.; BARKER, D. J. Type 2 (non-insulin-dependent) diabetes mellitus: the thrifty
phenotype hypothesis. Diabetologia, v. 35, n. 7, p. 595-601, Jul 1992. ISSN 0012-186X.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/1644236 >.
HENQUIN, J. C.; NENQUIN, M. Activators of PKA and Epac distinctly influence insulin
secretion and cytosolic Ca2+ in female mouse islets stimulated by glucose and tolbutamide.
Endocrinology, v. 155, n. 9, p. 3274-87, Sep 2014. ISSN 1945-7170. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/24977470 >.
HERZIG, S. et al. CREB regulates hepatic gluconeogenesis through the coactivator PGC-1.
Nature, v. 413, n. 6852, p. 179-83, Sep 2001. ISSN 0028-0836. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/11557984 >.

66
HIRIART, M.; AGUILAR-BRYAN, L. Channel regulation of glucose sensing in the
pancreatic beta-cell. Am J Physiol Endocrinol Metab, v. 295, n. 6, p. E1298-306, Dec 2008.
ISSN 0193-1849. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/18940941 >.
IKRAMUDDIN, S. et al. Effect of reversible intermittent intra-abdominal vagal nerve
blockade on morbid obesity: the ReCharge randomized clinical trial. JAMA, v. 312, n. 9, p.
915-22, Sep 2014. ISSN 1538-3598. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/25182100 >.
INOUE, S.; BRAY, G. A. The effects of subdiaphragmatic vagotomy in rats with
ventromedial hypothalamic obesity. Endocrinology, v. 100, n. 1, p. 108-14, Jan 1977. ISSN
0013-7227. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/830533 >.
KAHN, S. E.; HULL, R. L.; UTZSCHNEIDER, K. M. Mechanisms linking obesity to insulin
resistance and type 2 diabetes. Nature, v. 444, n. 7121, p. 840-6, Dec 2006. ISSN 1476-4687.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/17167471 >.
KEDEES, M. H. et al. Differential expression of glucagon and glucagon-like peptide 1
receptors in mouse pancreatic alpha and beta cells in two models of alpha cell hyperplasia.
Mol Cell Endocrinol, v. 311, n. 1-2, p. 69-76, Nov 2009. ISSN 1872-8057. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/19647035 >.
KRAL, J. G.; PAEZ, W.; WOLFE, B. M. Vagal nerve function in obesity: therapeutic
implications. World J Surg, v. 33, n. 10, p. 1995-2006, Oct 2009. ISSN 1432-2323.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/19618240 >.
LARSSON, H.; AHRÉN, B. Glucose intolerance is predicted by low insulin secretion and
high glucagon secretion: outcome of a prospective study in postmenopausal Caucasian
women. Diabetologia, v. 43, n. 2, p. 194-202, Feb 2000. ISSN 0012-186X. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/10753041 >.
LATORRACA, M. Q. et al. Protein deficiency and nutritional recovery modulate insulin
secretion and the early steps of insulin action in rats. J Nutr, v. 128, n. 10, p. 1643-9, Oct
1998. ISSN 0022-3166. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/9772130 >.
LEISSRING, M. A. et al. Designed inhibitors of insulin-degrading enzyme regulate the
catabolism and activity of insulin. PLoS One, v. 5, n. 5, p. e10504, May 2010. ISSN 1932-
6203. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/20498699 >.
LEITE, N. C. et al. Protein malnutrition potentiates the amplifying pathway of insulin
secretion in adult obese mice. Sci Rep, v. 6, p. 33464, Sep 2016. ISSN 2045-2322. Disponível
em: < https://www.ncbi.nlm.nih.gov/pubmed/27633083 >.

67
LEON-QUINTO, T.; ADNOT, P.; PORTHA, B. Alteration of the counterregulatory
hormones in the conscious rat after protein-energy restriction. Diabetologia, v. 40, n. 9, p.
1028-34, Sep 1997. ISSN 0012-186X. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/9300239 >.
LEON-QUINTO, T.; MAGNAN, C.; PORTHA, B. Altered activity of the autonomous
nervous system as a determinant of the impaired beta-cell secretory response after protein-
energy restriction in the rat. Endocrinology, v. 139, n. 8, p. 3382-9, Aug 1998. ISSN 0013-
7227. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/9681486 >.
LUBACZEUSKI, C. et al. Vagotomy ameliorates islet morphofunction and body metabolic
homeostasis in MSG-obese rats. Braz J Med Biol Res, v. 48, n. 5, p. 447-57, May 2015.
ISSN 1414-431X. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/25714886 >.
MAIANTI, J. P. et al. Anti-diabetic activity of insulin-degrading enzyme inhibitors mediated
by multiple hormones. Nature, v. 511, n. 7507, p. 94-8, Jul 2014. ISSN 1476-4687.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/24847884 >.
MALTA, A. et al. Low-protein diet in adult male rats has long-term effects on metabolism. J
Endocrinol, v. 221, n. 2, p. 285-95, May 2014. ISSN 1479-6805. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/24599936 >.
MARROQUI, L. et al. Functional and structural adaptations in the pancreatic alpha-cell and
changes in glucagon signaling during protein malnutrition. Endocrinology, v. 153, n. 4, p.
1663-72, Apr 2012. ISSN 1945-7170 (Electronic)
0013-7227 (Linking). Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/22334714 >.
MIRSKY, I. A.; PERISUTTI, G. Effect of insulinase-inhibitor on hypoglycemic action of
insulin. Science, v. 122, n. 3169, p. 559-60, Sep 1955. ISSN 0036-8075. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/13255892 >.
MITRANI, P. et al. Autonomic involvement in the permanent metabolic programming of
hyperinsulinemia in the high-carbohydrate rat model. Am J Physiol Endocrinol Metab, v.
292, n. 5, p. E1364-77, May 2007. ISSN 0193-1849. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/17227957 >.
MÜLLER, T. D. et al. The New Biology and Pharmacology of Glucagon. Physiol Rev, v. 97,
n. 2, p. 721-766, Apr 2017. ISSN 1522-1210. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/28275047 >.
NADAL, A.; QUESADA, I.; SORIA, B. Homologous and heterologous asynchronicity
between identified alpha-, beta- and delta-cells within intact islets of Langerhans in the
mouse. J Physiol, v. 517 ( Pt 1), p. 85-93, May 1999. ISSN 0022-3751. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/10226151 >.

68
NESHER, R. et al. Beta-cell protein kinases and the dynamics of the insulin response to
glucose. Diabetes, v. 51 Suppl 1, p. S68-73, Feb 2002. ISSN 0012-1797. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/11815461 >.
PARDO, J. V. et al. Chronic vagus nerve stimulation for treatment-resistant depression
decreases resting ventromedial prefrontal glucose metabolism. Neuroimage, v. 42, n. 2, p.
879-89, Aug 2008. ISSN 1095-9572. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/18595737 >.
PERLEY, M. J.; KIPNIS, D. M. Plasma insulin responses to oral and intravenous glucose:
studies in normal and diabetic sujbjects. J Clin Invest, v. 46, n. 12, p. 1954-62, Dec 1967.
ISSN 0021-9738. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/6074000 >.
PETRY, C. J. et al. Effects of early protein restriction and adult obesity on rat pancreatic
hormone content and glucose tolerance. Horm Metab Res, v. 32, n. 6, p. 233-9, Jun 2000.
ISSN 0018-5043. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/10898553 >.
PLAMBOECK, A. et al. The effect of exogenous GLP-1 on food intake is lost in male
truncally vagotomized subjects with pyloroplasty. Am J Physiol Gastrointest Liver Physiol,
v. 304, n. 12, p. G1117-27, Jun 2013. ISSN 1522-1547. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/23599045 >.
QUESADA, I. et al. Glucose induces opposite intracellular Ca2+ concentration oscillatory
patterns in identified alpha- and beta-cells within intact human islets of Langerhans. Diabetes,
v. 55, n. 9, p. 2463-9, Sep 2006. ISSN 0012-1797. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/16936194 >.
______. Physiology of the pancreatic alpha-cell and glucagon secretion: role in glucose
homeostasis and diabetes. J Endocrinol, v. 199, n. 1, p. 5-19, Oct 2008. ISSN 1479-6805.
Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/18669612 >.
QUINN, P. G.; GRANNER, D. K. Cyclic AMP-dependent protein kinase regulates
transcription of the phosphoenolpyruvate carboxykinase gene but not binding of nuclear
factors to the cyclic AMP regulatory element. Mol Cell Biol, v. 10, n. 7, p. 3357-64, Jul 1990.
ISSN 0270-7306. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/2141384 >.
RAMNANAN, C. J. et al. Physiologic action of glucagon on liver glucose metabolism.
Diabetes Obes Metab, v. 13 Suppl 1, p. 118-25, Oct 2011. ISSN 1463-1326. Disponível em:
< https://www.ncbi.nlm.nih.gov/pubmed/21824265 >.
RASKIN, P.; UNGER, R. H. Hyperglucagonemia and its suppression. Importance in the
metabolic control of diabetes. N Engl J Med, v. 299, n. 9, p. 433-6, Aug 1978. ISSN 0028-
4793. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/683275 >.

69
REEVES, P. G.; NIELSEN, F. H.; FAHEY, G. C. AIN-93 purified diets for laboratory
rodents: final report of the American Institute of Nutrition ad hoc writing committee on the
reformulation of the AIN-76A rodent diet. J Nutr, v. 123, n. 11, p. 1939-51, Nov 1993. ISSN
0022-3166. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/8229312 >.
REIS, M. A. et al. Glucose-induced insulin secretion is impaired and insulin-induced
phosphorylation of the insulin receptor and insulin receptor substrate-1 are increased in
protein-deficient rats. J Nutr, v. 127, n. 3, p. 403-10, Mar 1997. ISSN 0022-3166. Disponível
em: < http://www.ncbi.nlm.nih.gov/pubmed/9082023 >.
REMMERS, F.; DELEMARRE-VAN DE WAAL, H. A. Developmental programming of
energy balance and its hypothalamic regulation. Endocr Rev, v. 32, n. 2, p. 272-311, Apr
2011. ISSN 1945-7189. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/21051592 >.
SALTIEL, A. R.; KAHN, C. R. Insulin signalling and the regulation of glucose and lipid
metabolism. Nature, v. 414, n. 6865, p. 799-806, Dec 2001. ISSN 0028-0836. Disponível em:
< http://www.ncbi.nlm.nih.gov/pubmed/11742412 >.
SANDOVICI, I. et al. Developmental and environmental epigenetic programming of the
endocrine pancreas: consequences for type 2 diabetes. Cell Mol Life Sci, v. 70, n. 9, p. 1575-
95, May 2013. ISSN 1420-9071. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/23463236 >.
SANTOS-SILVA, J. C. et al. Taurine supplementation ameliorates glucose homeostasis,
prevents insulin and glucagon hypersecretion, and controls β, α, and δ-cell masses in genetic
obese mice. Amino Acids, v. 47, n. 8, p. 1533-48, Aug 2015. ISSN 1438-2199. Disponível
em: < https://www.ncbi.nlm.nih.gov/pubmed/25940922 >.
SEINO, S.; SHIBASAKI, T. PKA-dependent and PKA-independent pathways for cAMP-
regulated exocytosis. Physiol Rev, v. 85, n. 4, p. 1303-42, Oct 2005. ISSN 0031-9333.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/16183914 >.
SHIKORA, S. et al. Vagal blocking improves glycemic control and elevated blood pressure
in obese subjects with type 2 diabetes mellitus. J Obes, v. 2013, p. 245683, 2013. ISSN
2090-0716. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/23984050 >.
SUTTON, G. M.; CENTANNI, A. V.; BUTLER, A. A. Protein malnutrition during
pregnancy in C57BL/6J mice results in offspring with altered circadian physiology before
obesity. Endocrinology, v. 151, n. 4, p. 1570-80, Apr 2010. ISSN 1945-7170. Disponível em:
< https://www.ncbi.nlm.nih.gov/pubmed/20160133 >.
SVOBODA, M. et al. Relative quantitative analysis of glucagon receptor mRNA in rat
tissues. Mol Cell Endocrinol, v. 105, n. 2, p. 131-7, Nov 1994. ISSN 0303-7207. Disponível
em: < https://www.ncbi.nlm.nih.gov/pubmed/7859919 >.

70
TENGHOLM, A.; GYLFE, E. Oscillatory control of insulin secretion. Mol Cell Endocrinol,
v. 297, n. 1-2, p. 58-72, Jan 2009. ISSN 0303-7207. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/18706473 >.
THORE, S. et al. Feedback activation of phospholipase C via intracellular mobilization and
store-operated influx of Ca2+ in insulin-secreting beta-cells. J Cell Sci, v. 118, n. Pt 19, p.
4463-71, Oct 2005. ISSN 0021-9533. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/16159958 >.
THORE, S.; WUTTKE, A.; TENGHOLM, A. Rapid turnover of phosphatidylinositol-4,5-
bisphosphate in insulin-secreting cells mediated by Ca2+ and the ATP-to-ADP ratio.
Diabetes, v. 56, n. 3, p. 818-26, Mar 2007. ISSN 0012-1797. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/17327453 >.
UNGER, R. H.; ORCI, L. The essential role of glucagon in the pathogenesis of diabetes
mellitus. Lancet, v. 1, n. 7897, p. 14-6, Jan 1975. ISSN 0140-6736. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/46337 >.
VICKERS, M. H. et al. Fetal origins of hyperphagia, obesity, and hypertension and postnatal
amplification by hypercaloric nutrition. Am J Physiol Endocrinol Metab, v. 279, n. 1, p.
E83-7, Jul 2000. ISSN 0193-1849. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/10893326 >.
WEWER ALBRECHTSEN, N. J. et al. Hyperglucagonaemia analysed by glucagon sandwich
ELISA: nonspecific interference or truly elevated levels? Diabetologia, v. 57, n. 9, p. 1919-
26, Sep 2014. ISSN 1432-0428. Disponível em: <
https://www.ncbi.nlm.nih.gov/pubmed/24891019 >.
WHITEMAN, E. L.; CHO, H.; BIRNBAUM, M. J. Role of Akt/protein kinase B in
metabolism. Trends Endocrinol Metab, v. 13, n. 10, p. 444-51, Dec 2002. ISSN 1043-2760.
Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/12431841 >.
YANG, S. N.; BERGGREN, P. O. The role of voltage-gated calcium channels in pancreatic
beta-cell physiology and pathophysiology. Endocr Rev, v. 27, n. 6, p. 621-76, Oct 2006.
ISSN 0163-769X. Disponível em: < http://www.ncbi.nlm.nih.gov/pubmed/16868246 >.
YOUNGREN, J. F. Regulation of insulin receptor function. Cell Mol Life Sci, v. 64, n. 7-8,
p. 873-91, Apr 2007. ISSN 1420-682X. Disponível em: <
http://www.ncbi.nlm.nih.gov/pubmed/17347799 >.
ZHANG, Q. et al. Role of KATP channels in glucose-regulated glucagon secretion and
impaired counterregulation in type 2 diabetes. Cell Metab, v. 18, n. 6, p. 871-82, Dec 2013.
ISSN 1932-7420. Disponível em: < https://www.ncbi.nlm.nih.gov/pubmed/24315372 >.

71

72
ANEXO 1-
Certificado da comissão de ética no uso de animais

73
ANEXO 2
Direitos autorais