Embed Size (px)
Citation preview

Department of Clinical Veterinary Sciences Faculty of Veterinary Medicine
University of Helsinki Finland
Department of Veterinary Clinical Sciences
Faculty of Veterinary Science University of Sydney
Australia
Endocrinological basis of seasonal infertility in pigs
Anssi Tast
ACADEMIC DISSERTATION
To be presented, with the permission of
the Faculty of Veterinary Medicine, University of Helsinki, for public criticism in Auditorium Maximum,
Hämeentie 57, Helsinki, on September 9th, 2002 at 12 noon
HELSINKI 2002

ISBN 952-91-5024-5ISBN (PDF) 952-10-0655-2
Helsinki 2002Yliopistopaino

1
CONTENTS
ABSTRACT 3
LIST OF ORIGINAL PAPERS 6
ABBREVIATIONS 7
1. INTRODUCTION AND REVIEW OF LITERATURE 9
1.1. INTRODUCTION 9 1.2. MANIFESTATIONS OF SEASONAL INFERTILITY 10
1.2.1. Reduced farrowing rate 10 1.2.2. Delayed puberty 11 1.2.3. Prolonged weaning to oestrus interval 12
1.3. ENVIRONMENTAL FACTORS CONTRIBUTING TO SEASONAL INFERTILITY 12 1.3.1. Photoperiod 12 1.3.2. Temperature 13 1.3.3. Feeding level 14 1.3.4. Other environmental factors 15
1.4. PATTERN OF MELATONIN SECRETION SYNCHRONIZES BREEDING WITH
SEASON 16 1.4.1. Circadian rhythm in melatonin secretion 16 1.4.2. Pineal secretion of melatonin 18 1.4.3. The role of melatonin in regulation of hypothalamo – pituitary – gonadal axis 18 1.4.4. The melatonin pattern in the pig 20 1.4.5. Effects of light intensity on melatonin 22
1.5. RECOGNITION OF PREGNANCY 23 1.5.1. PGF2α secretion during the oestrous cycle 24 1.5.2. Embryonic signals 24 1.5.3. Oestrogen signals alter PGF2α secretion during pregnancy 27 1.5.4. Conceptus secretory proteins (CSP) 28
1.6. MATERNAL RESPONSE 29 1.6.1. Formation of CL and luteotrophic role of LH 29 1.6.2. Progesterone in early pregnancy 31
2. THE AIMS OF THE STUDY 33
3. MATERIALS AND METHODS 34
3.1. ANIMALS 34 3.2. BLOOD COLLECTION 35
3.2.1. Jugular vein catheters (I, II, III, V) 35 3.2.2. Single blood samples (IV, V) 35 3.2.3. Saphenous arterial catheters (I) 36
3.3. GNRH-IMMUNIZATION 37

2
3.3.1. Active immunization (V) 37 3.3.2. Passive immunization (V) 37
3.4. HORMONE AND ANTIBODY ASSAYS 37 3.4.1. Melatonin (I, II, III) 37 3.4.2. Progesterone (IV, V) 40 3.4.4. GnRH-antibody (V) 42
3.5. PREGNANCY TESTING 42 3.6. LIGHTING PROGRAMS 43 3.7. STATISTICAL ANALYSES 43
4. RESULTS 44
4.1. SEASONAL ALTERATIONS IN MELATONIN SECRETION (I) 44 4.2. EFFECTS OF ARTIFICIAL LIGHTING PROGRAMS ON MELATONIN SECRETION
(II, III) 45 4.3. EARLY DISRUPTION OF PREGNANCY (IV, V) 47
5. DISCUSSION 49
6. CONCLUSIONS 55
ACKNOWLEDGEMENTS 56
REFERENCES 58

3
ABSTRACT
Seasonal infertility of the domestic pig has different manifestations, but reduced farrowing rate has
been found to be of highest economic significance. In other seasonally breeding non-tropical
mammals, the seasonal effects on reproduction are mediated by the pineal hormone melatonin. In
the domestic pig, there has been considerable controversy about the existence and role of a
circadian melatonin pattern.
The aims of the study were to investigate the melatonin secretion of the wild boar and compare it
with that of the domestic pig. The effects of different lighting regimens and light intensities on the
circadian melatonin response of pig were also studied. The association between the seasonally
reduced farrowing rate and early disruption of pregnancy was clarified in a field trial. The
endocrinological mechanism of early disruption of pregnancy was investigated by active and
passive GnRH-immunization trials, the aim being to develop an experimental model of this
mechanism.
To determine seasonal alterations in melatonin profiles of the European wild boar and the domestic
pig, the wild boars and domestic gilts were sampled at two-hour intervals for 48 hours four times a
year. The wild boars were under the natural lighting and the gilts under the typical indoor piggery
light environment, where artificial lighting was provided for 12 hours a day and the sunlight
through the windows was not controlled. Both pigs expressed a clear seasonal variation in the
duration of melatonin secretion. There was no difference in lighting – melatonin transduction
between wild and domestic pigs.
To test the effect of different lighting regimens on the secretion pattern of melatonin an experiment
was set up, where young prepubertal boars in four different temperature- and lighting-controlled
climate rooms were exposed to short day or long day lighting conditions. Following a two-week
acclimatizing period, lighting regimens were changed to be opposite in each room. Blood samples
were collected at two-hour intervals for 48 hours spanning the changeover and assayed for
melatonin. The procedure was repeated three times so that the pigs ended up with the same lighting
with which they started. This experiment demonstrated existence of a circadian melatonin pattern
under both lighting regimens and in every animal sampled. Furthermore, the experiment showed

4
that pigs were able to respond to abrupt and extreme changes in lighting within two weeks in terms
of adjustment of the circadian melatonin pattern to a new lighting regime.
The effects of the light intensity during the photophase on the subsequent scotophase melatonin
response were also investigated. Three groups of four gilts were accommodated in three lighting-
and temperature-controlled climate rooms. In each room the light-dark cycle was 12 h : 12 h and the
light intensity was either 40 lx, 200 lx or 10000 lx during the photophase. Following a one-week
acclimatizing period, blood samples were collected at two-hour intervals for 24 hours and assayed
for melatonin. All the groups went through all the treatments (light intensities). This experiment
showed that pigs are able to recognize a light intensity as low as 40 lx. The experiment also
demonstrated no difference in the scotophase melatonin response to the different light intensities.
This experiment suggests that a light-dark cycle is important in generating the melatonin signal,
whereas the light intensity is of little importance.
The association of lowered farrowing rate with early disruption of pregnancies was studied in a
commercial 160-sow unit with a known history of seasonally lowered farrowing rate. Every
sow/gilt in production was included in the study for four months periods in the winter – spring and
in the summer – autumn. The pregnancies were followed for six weeks by serum progesterone
concentrations and ultrasound scanning. An increase in disruption of pregnancies was associated
with a seasonal decrease in the farrowing rate.
The mechanism of early disruption of pregnancy was studied by active and passive GnRH-
immunization. Sows were immunized for the first time on the day of farrowing and the second time
either on day 10 or on day 20 post mating. The passive immunization of gilts was carried out on day
12 post mating. None of the sows actively immunized on day 10 was detectably pregnant (real time
ultrasound) on day 18 post mating. The sows immunized for the second time on day 20 aborted with
a mean vaccination-abortion interval of 10 ± 1.5 days. The passive immunization on day 12 had a
similar effect on establishment of pregnancy as active immunization on day 10 post mating.
Progesterone samples showed that in both active immunization groups the regression of CL took
place within eight days following the second immunization. These results suggest that two different
mechanisms can cause disruption of pregnancy depending on the timing of the immunization. The
immunization on day 20 caused a regression of CL within eight days and abortion followed two
days later whereas the immunization on day 10 caused a failure in establishment of pregnancy
before a regression of CL took place. These findings suggest that seasonally decreased progesterone

5
secretion of CL, because of inadequate LH-support, may interfere with development of embryos
and their ability to produce oestrogen signals for recognition of pregnancy.
It is concluded that the basis of lighting-melatonin transduction in the pig is similar to other
mammals and that simple artificial lighting programs can manipulate the circadian rhythm of
melatonin secretion. The seasonally decreased farrowing rate can partly be explained by an
increased number of disrupted pregnancies. It is suggested that seasonal early disruption of
pregnancy is mediated through seasonally decreased progesterone concentrations, which interfere
with development of embryos. The seasonal decrease in progesterone may be caused by inadequate
LH-support for CL which results from changed melatonin secretion pattern that is inhibitory for
GnRH-secretion.

6
LIST OF ORIGINAL PAPERS
The present thesis comprises the following original papers. In the text, the papers are referred to by
their Roman numerals. They were reprinted by the kind permission of the journal concerned.
I) Tast, A., Hälli, O., Ahlström, S., Andersson, H., Love, R.J. and Peltoniemi, O.A.T.
Seasonal alterations in circadian melatonin rhythms of the European wild boar and
domestic gilt. J. Pineal Res. 2001, 30: 43-49.
II) Tast, A., Love, R.J., Evans, G., Telsfer, S., Giles, R., Nicholls, P., Voultsios, A. and
Kennaway, D.J. The pattern of melatonin secretion is rhythmic in the domestic pig and
responds rapidly to changes in daylength. J. Pineal Res. 2001, 31:294-300.
III) Tast, A., Love, R.J., Evans, G., Andersson, H., Peltoniemi, O.A.T. and Kennaway, D.J.
The photophase light intensity does not affect the scotophase melatonin response in the
domestic pig. Anim. Reprod. Sci. 2001, 65: 283-290.
IV) Tast, A., Peltoniemi, O.A.T., Virolainen, J.V. and Love, R.J. Early disruption of
pregnancy as a manifestation of seasonal infertility in pigs. Anim. Reprod. Sci. (in press)
V) Tast, A., Love, R.J., Clarke, I.J. and Evans, G. Effects of active and passive
gonadotrophin-releasing hormone immunization on recognition and establishment of
pregnancy in pigs. Reprod. Fertil. Dev. 2000, 12: 277-282.

7
ABBREVIATIONS
Ab antibody
BSA bovine serum albumin
bTP-1 bovine trophoplast protein-1
C18 active charcoal
CL corpus luteum
cm centimeter
CSP conceptus secretory proteins
CV coefficient of variation
EDP early disruption of pregnancy
EDTA ethylenediaminetetraacetic acid
EMAI Elizabeth McArthur Agricultural Institute
FSH follicle stimulating hormone
g grams
GnRH gonadotrophin releasing hormone
I125 radioactive iodine
IFN interferon
kg kilograms
L liters
LH luteinizing hormone
LPP long pseudo pregnancy
MB maximal binding
min minutes
MJ megajoules
ml milliliters
mm millimeters
NaCl natrium chloride
ng nanograms
NSB non specific binding
oTP-1 ovine trophoplast protein-1
pg picograms
PGF2α prostaglandin F2α

8
PGE prostaglandin E
registered trade mark
RBP retinol binding protein
RIA radioimmunoassay
rpm rounds per minute
SCN suprachiasmatic nucleus
SPP short pseudo pregnancy
µg micrograms
µl microliters

9
1. INTRODUCTION AND REVIEW OF LITERATURE
1.1. INTRODUCTION
The Western domestic pig breeds are derived from the European wild boar (Sus scrofa scrofa),
which has a breeding season in late autumn - early winter. Piglets of the wild boar are born in the
spring when climatic conditions and feed sources maximise the chances of survival. After giving
birth to piglets, a wild boar sow remains in anoestrus until the next winter. Initially this anoestrus is
associated with lactation, but after weaning the sow stays in anoestrus until the next breeding
season. The photoperiod is the primary factor determining the onset of the breeding season, while
availability of feed has a modifying effect. Changing photoperiod affects the pattern of melatonin
secretion from the pineal gland, and it is thought to be the most important factor that synchronizes
breeding with the preferred season.
Although the domestic pig is not a distinct seasonal breeder, it is clear that it has a tendency to a
lowered fertility in the autumn. Seasonal infertility appears in late summer and early autumn, same
time when the European wild boar experiences total anoestrus. A reduced farrowing rate, delayed
puberty in gilts and a prolonged weaning to oestrus interval manifest seasonal infertility. Gilts and
primiparous sows are most severely affected (Love, 1978; Peltoniemi et al., 1999b).
Seasonal effects on reproductive performance of the pig and its physiological and endocrinological
mechanisms are discussed in the present review of the literature. In the first section, different
manifestations of seasonal infertility are described. In addition, environmental factors, which
contribute to seasonal infertility, are briefly reviewed. In the second section, attention is paid to the
endocrinology of seasonal breeding and some particular features of melatonin secretion patterns in
pigs are pointed out. The third section discusses the effect of changing photoperiod on the
recognition and establishment of pregnancy.

10
1.2. MANIFESTATIONS OF SEASONAL INFERTILITY 1.2.1. Reduced farrowing rate
Farrowing rate is defined as the proportion of mated sows that farrow (Love, 1981). Repeated
matings/inseminations to the same oestrus are calculated as a single mating event. This may be
modified by removing sows that have died or been culled for some reason other than fertility
problems to give an adjusted farrowing rate.
Reduced farrowing rate is typical following matings in late summer and early autumn (Love et al.,
1978). In another study by Love (1981), more delayed returns to oestrus after mating were found
during summer-autumn than winter-spring. Using oestrone sulphate as an indicator of pregnancy, it
was found that 25-30 % of sows detected to be pregnant did not farrow in the autumn where as in
the spring only 4 % of sows detected to be pregnant did not farrow (Mattioli et al., 1987). These
studies provide strong evidence for the important role of early disruption of pregnancy in seasonal
infertility, and it appears to be important in reducing farrowing rate seasonally.
A reduction in the farrowing rate following matings in autumn has been revealed in many studies,
but the severity of the problem has been variable. Stork (1979) reported only a 3-5 % reduction in
farrowing rates caused by the season in Britain, and Lucia et al. (1994) found season to cause
approximately a 3 % reduction in farrowing rates in North American herds. In a retrospective study
on the seasonal effect on fertility, a 5-10 % reduction in the farrowing rate following matings from
August to October was found in Finland (Peltoniemi et al., 1999a). In an Australian study (Fig. 1.)
farrowing rates following autumn matings dropped down to 50 % in most severe cases and 10-15 %
reductions were commonly seen in this country (Love et al., 1993). In Italy, Enne et al. (1979)
reported that only 40 % of sows mated in the summer farrowed.
Variation in the severity of seasonal infertility is at least partly explained by the different
management and environmental factors (Hancock, 1988). It is also typical for seasonal infertility to
have a great variation between years, weeks, piggeries and even within the same piggery amongst
different groups of pigs (Love et al., 1978; 1993). The great variation in severity and an
unpredictable appearance of the problem make it difficult to control. Early disruption of pregnancy
causes irregular return to oestrus, as discussed earlier. The traditional method of detecting non-
pregnant sows by boar at three weeks after mating is unlikely to detect sows, which have had an

11
early disruption of the pregnancy. If any other pregnancy test methods are not used the risk of larger
numbers of sows being non-pregnant near the expected farrowing date increases following summer
and autumn matings.
F a r ro w in g ra te
6 46 66 87 07 27 47 67 88 08 28 48 6
s p r in g s u m m e r a u tu m n w in te r
N = 7 2 1 3
Figure 1: Seasonal effects on farrowing rates 1983-1991. Data from Love et al. (1993).
1.2.2. Delayed puberty
Most of the studies support the opinion that gilts reach puberty at an older age during the seasonal
infertility period compared with the rest of the year (Hughes, 1982; Peltoniemi et al., 1999a). In an
Australian study, 53 % of gilts reached puberty at 225 days’ age, when kept in short day lighting
conditions around the expected time of the puberty and isolated from boars, whereas only 13 % of
gilts reached puberty by that age when kept in long day lighting conditions and isolated from boars
(Paterson et al., 1991). The effect of season on puberty is dependent partly on other environmental
factors, especially on boar contact. However, the seasonal delay of puberty in gilts seems to remain
to some extent even under the influence of boar contact (Paterson et al., 1991). This seasonally
delayed puberty is typical for the European wild boar in its natural environment. After threshold
values in age and weight are achieved, the occurrence of puberty depends on season; if the right age
and weight are reached late in the spring, the attainment of the puberty will be delayed until the next
winter (Mauget, 1987).
In a Finnish study, (Peltoniemi et al., 1997a) puberty of domestic gilts was delayed by
approximately 10 days during the autumn compared with the rest of the year. This delayed
attainment of puberty has economic consequences in commercial piggeries, especially because it

12
occurs simultaneously with the period of reduced farrowing rate and prolonged weaning to oestrus
interval in sows.
1.2.3. Prolonged weaning to oestrus interval
Effects of seasonal infertility on the first oestrus after weaning seem to vary. Many authors have
reported severe adverse effects of season on weaning to oestrus interval. In these reports, weaning
to oestrus interval in primiparous sows was prolonged significantly during the period of seasonal
infertility (Hurtgen and Leman, 1980; Mattioli et al., 1987; Prunier et al., 1996). These sows
experienced a total anoestrus after weaning, analogous to the situation in the wild boar. In Finland,
a prolonged weaning to oestrus interval seems not to be a severe problem according to data
collected from the database of the Finnish Animal Breeding Association (Peltoniemi et al., 1999b).
There is not a clear explanation why prolonged weaning to oestrus interval is not such a severe
problem in Finland as in many other countries. One explanation could be the small average herd
size in Finland, where average number of sows per farm is only about 50. The oestrus detection
might be more efficient in small farms than in larger herds and, thus, weak oestrus signs might be
more easily recognized. It has to be taken into consideration that collecting data from this kind of
database may lead to an underestimation of some particular problem depending on the methods of
data collection and classification. However, in general, a prolonged weaning to oestrus interval is
still considered an important and economically significant manifestation of seasonal infertility.
1.3. ENVIRONMENTAL FACTORS CONTRIBUTING TO SEASONAL INFERTILITY
1.3.1. Photoperiod
Photoperiod synchronizes the beginning of the breeding season with the preferred season in most
non-tropical seasonally breeding mammals. For short day breeders such as the sheep, the long
duration of high nocturnal plasma melatonin concentrations leads to an increased frequency of
GnRH-pulses through complicated neural pathways and hormonal feed back mechanisms, which
stimulate the release of LH from the pituitary gland and thereby activates the gonads. In pigs,
effects of the photoperiod on melatonin and on subsequent hypothalamic activity are not as well
documented as in the sheep. Some authors have found extraordinary melatonin profiles like

13
increased melatonin concentration during the photophase in pigs (Peacock, 1991), and others seem
to think that the neuro-endocrinological basis of the pig is similar to sheep and that confusing
results in melatonin profiles may be due to technical difficulties in assays used (Paterson et al.,
1992). Some fundamental details in melatonin patterns in pigs under different lighting conditions
have remained unclear.
Confusing results of effects of photoperiod on reproduction (and how the photoperiodic information
is transduced into endogrinological form) in domestic pigs have led to the recommendation of a 16-
hours’ photophase all year round in many countries. For example, in Scandinavian countries where
pigs are kept indoors and lighting is controlled artificially, recommendation is to provide sows with
16-17 hours of light daily. However, the latest experiments have provided strong evidence that the
domestic pig still has a tendency towards seasonal breeding similar to European wild boar, which is
a distinct short day breeder. One could expect that short day lighting regimens might be beneficial
in commercial piggeries at least a couple of weeks before and after moving sows or gilts into a
breeding unit. The optimal lighting regimens might demand different lighting conditions in different
units but these programs are yet to be determined.
1.3.2. Temperature
It has been suggested that high ambient temperatures have importance in seasonal infertility
(Wetteman and Bazer, 1985). However, high temperatures do not correlate with the time of the year
when seasonal infertility is seen, namely late summer and early autumn. If the high temperature was
the main reason for seasonal infertility, one would expect to see reduced litter size rather than
disruption of the whole pregnancy. However, a reduced litter size is not a typical feature of seasonal
infertility (Love, 1978). Use of air conditioning and cooling showers has not solved the problem,
which speaks against high ambient temperature as the underlying factor in seasonal infertility
(Hurtgen and Leman, 1980). High ambient temperatures may, under some circumstances, have an
indirect adverse effect on fertility by reducing the voluntary feed intake of lactating sows leading to
energy imbalance and fertility problems (Prunier et al., 1996). Low ambient temperatures without
compensatory energy feeding in the autumn are more likely to have an adverse effect on fertility
than high temperatures. Low temperatures play an important role in the “autumn abortions
syndrome“ (Almond et al., 1985).

14
1.3.3. Feeding level
According to the conventional feeding recommendations pregnant sows have received a restricted
feeding level (2.0-2.2 kg/day, 13 MJ/kg diet) for the whole dry period. The rationale for restricted
feeding level after mating is based on findings, which have demonstrated less early embryonic
deaths taking place in primiparous sows when held under low feeding levels compared with high
feeding levels (Aherne and Kirkwood, 1985). The conventional feeding recommendations for sows
have been same throughout the year.
Effects of feeding level and the type of housing during early pregnancy (individual stalls vs. group
housing) have been studied in two large piggeries in Australia (Love et al., 1995). In group housing
system, the low feeding level (1.6-2.0 kg, 13 MJ/kg) increased the weaning to oestrus interval and
reduced farrowing rate (50 %) during summer and autumn (Table 1.). In the winter and spring, the
same level of feeding did not have any adverse effect on weaning to oestrus interval and the
farrowing rate remained high (> 85 %). Higher amount of feed (2.5 kg and over 3.6 kg) had a
positive effect on the farrowing rate in summer and autumn. Feeding level did not affect the litter
size. Seasonal infertility was not detected when sows were kept in individual stalls and fed 2.5 kg
per day. This is the best study available, because the comparison between groups was done in the
same piggery, whereas many other studies have compared housing systems and feeding levels
between piggeries, thereby confounding the data.
Table 1. Effect of type of housing and feeding level on farrowing rates (Love et al., 1995). Level of feeding: low = 1.6-2.0 kg/day, moderate = 2.5 kg/day, high > 3.6 kg/day (13 MJ/kg). These feeding levels were applied for the first four weeks of pregnancy. Thereafter a feeding level of 2.5-3.2 kg / day was given for the remainder of pregnancy.
Season Farrowing rate
Group housing Individual stalls
Feeding Low Moderate High Moderate
Summer-Autumn 50.0 69.0 74.1 84.0
Winter-Spring 87.0 - 87.5 87.6

15
It has been hypothesized that the protective effect of higher feeding against seasonal infertility
might be because of increased pituitary LH support to the CL. It has been documented that pituitary
LH-pulses are of lower amplitude and irregular under long day lighting, in other words during the
seasonal infertility period (Paterson et al., 1992; Peltoniemi et al., 1997b). Increased feeding level
might compensate for this seasonal decrease in LH-pulses. Peltoniemi et al. (1997c) studied the
effect of different energy levels and gut fill on LH-secretion in early pregnant gilts, and they found
that the gilts on the highest energy intake had higher LH-pulse amplitude compared with gilts on
lower energy intake. However, there was no effect of energy intake on plasma LH-pulse frequency,
mean concentration, area under the curve or mean nadir. They concluded that the protective effect
of higher feeding level against seasonal disruption of pregnancy appears to be mediated by a
mechanism other than an alteration in LH-secretion. In that study, the energy content of the feed
was increased by adding a fat supplement (soybean oil) to a commercial ration. It did not mimic the
situation in normal farm conditions, where the energy increase is achieved by increasing the total
amount of the basic commercial ration. Increasing the total energy intake through carbohydrates has
more effects on insulin-regulated glucose metabolism than increasing energy intake through fat.
The glucose metabolism, in particular, is known to affect the hypothalamic-pituitary-ovarian
function in the pig (Booth, 1990). More investigations of the mechanism are needed to determine
how higher feeding level in early pregnancy is able to protect against adverse effects of season on
establishment of pregnancy.
1.3.4. Other environmental factors
Pigs are known to have a very sensitive olfactory system. Pheromones secreted by the boar have
been documented to have a strong positive effect on oestrous signs in sows. It has also been shown
that oestrous sows have effects on other sows in the group leading to synchronization of oestrus in
the group (Pearce and Pearce, 1992). Similar synchronization has been reported in the European
wild boar sows (Delcroix et al., 1990). In some circumstances pheromones may have a negative
effect on oestrus, especially in young sows or gilts. In the summer and autumn, when sows appear
more sensitive to all kinds of adverse effects on fertility, negatively effecting pheromones may
abolish oestrus and even disrupt pregnancy. Too many animals in too small a pen can cause stress,

16
and sows or gilts that are of a lower social ranking may lose the pregnancy. In one study, mixing
older pregnant sows in the same pen with younger sows lowered fertility in the young sows (Wilson
and Love, 1990).
In many studies, group housing has led to lower fertility during the seasonal infertility period
compared with individual stalls or pens (Hurtgen and Leman, 1980; Love et al., 1995). This has at
least partly been explained by social stress related to competition for feed and a need to defend
social rank in a group housing situation. However, in the study carried out by Peltoniemi et al.
(1999a), where management and seasonal effects on fertility were examined in 1298 pig farms over
a four-year period in 1992-1996, a clear reduction in farrowing rate was evident during late summer
and early autumn despite 70.2 % of the sows being housed in individual stalls. This study did not
support the earlier studies where individual stalls have been found to protect against reduced
farrowing rate during the seasonal infertility period. It is obvious that some management strategies
essential for achieving good fertility in loose housing system differ from those conducted in
piggeries where animals are individually housed. However, if these differences in management,
especially in a feeding strategy during early pregnancy, are recognized, there is little evidence that
individual housing per se provides better fertility.
1.4. PATTERN OF MELATONIN SECRETION SYNCHRONIZES BREEDING WITH SEASON
1.4.1. Circadian rhythm in melatonin secretion
Seasonality in breeding is a characteristic of animals that inhabit the higher latitudes, where
variations in temperature and food availability are much greater than at lower latitudes. During
evolution, many mammals have opted to use changing photoperiod as the signal to provide
appropriate timing of the breeding season. Photoperiod is a very reliable signal of the season,
compared e.g., with the ambient temperature, which can vary considerably at a particular time of
the year. Light entering the eye stimulates retina and the pulses generated pass to the
suprachiasmatic nuclei (SCN) in the hypothalamus. This synchronizes endogenous melatonin
secretion of the pineal gland. Melatonin secretion increases in the dark and is inhibited by light. The
secretion rhythm is synchronized by ambient lighting conditions, but the melatonin secretion itself

17
is an endogenous circadian rhythm and this rhythm is generated by the SCN (Ebling and Hastings,
1992).
The pattern of circadian rhythm of melatonin secretion synchronizes the beginning of the breeding
season in seasonal breeders, irrespective of whether they are long day or short-day breeders (Turek
and Campbell, 1979). The photoperiod - pineal - hypothalamic pathway in rams has been reviewed
extensively by Lincoln (1992), and the principle features are discussed here. In short day breeders
such as the sheep, the long duration of high melatonin concentrations in the systemic blood
circulation causes an increase in gonadotrophin secretion, which leads to the onset of the breeding
season. The importance of the duration of melatonin secretion has been well documented in the
sheep. Daily 8-hour infusions of melatonin to pinealectomized sheep induce a long day response,
while 16-hour infusions induce a short day response. The daily pattern of melatonin secretion is a
cue in synchronizing the breeding season also in the hamster, which is a long day breeder. The
amount of melatonin, which induced gonadal regression, administered over 10 hours a day to
pinealectomized hamsters had an opposite effect if it was administered over a period of 4 hours
(Carter and Goldman, 1983). The function of the pineal gland is much more complicated than only
to give the information about absolute daylength or nightlength. It also has to provide information
to responsive organs if the days are getting longer or shorter. In this way a given organ can begin its
physiological adjustments in advance, so that it is able to change its function at the time required
(Reiter, 1991). The unusual melatonin profiles of the domestic pig under different lighting reported
in the literature have lead to suggestions that domestication has eliminated or attenuated the
circadian melatonin pattern of the pig (Green et al., 1996). However, this hypothesis has not been
proven.
A change in circadian melatonin secretion is not the only factor synchronizing the breeding season.
It is well known that, for example in the wild boar, the availability of feed affects the onset of the
breeding season. Social interactions are also known to be important in the regulation of seasonality.
The early studies in pinealectomized sheep failed to demonstrate the role of the pineal gland in
synchronizing seasonal breeding because the pinealectomized sheep were kept together with intact
sheep (Roche et al., 1970). Pinealectomized animals were able to respond to the social signals from
intact male and/or female sheep in the absence of photoperiodic information (Wayne et al., 1989).
Melatonin is an extraordinary hormone because its physiological effects do not depend simply on its
presence or absence or the total concentration secreted at a particular time. It seems that the pattern

18
of secretion over a longer period is more important, as discussed earlier. Another interesting feature
of melatonin is that it is able to cause different responses in different species, depending on whether
the animal is a long- or short day breeder. Many details of the action of melatonin are yet to be
clarified but it seems to be a key hormone in synchronizing the beginning of the breeding season
with preferred season along with some modifying factors.
1.4.2. Pineal secretion of melatonin
It has now been clearly documented that a pineal indoleamine melatonin is the substance that gives
the chemical indicator of darkness to the hypothalamus through a complex neural and hormonal
feed back mechanism. The pineal gland secretes quite a few different indoleamines and
neuropeptides, but the treatment of pinealectomized animals with the appropriate pattern of
melatonin induces seasonal effects on reproduction. It has been shown conclusively that melatonin
is the key hormone in interpreting photoperiods (Ebling and Hastings, 1992). SCN works as the
"inner clock", which produces the circadian rhythm of melatonin secretion. This endogenous
circadian rhythm is synchronized by light-dark cycles, and light also directly suppresses melatonin
secretion, provided that the light intensity is high enough or the duration of the light is long enough
(Aoki et al., 1998). The combination of these two factors ensures that nocturnal melatonin secretion
follows the nightlength (Ebling and Hastings, 1992).
1.4.3. The role of melatonin in regulation of hypothalamo – pituitary – gonadal axis
The role of melatonin in synchronizing the breeding season is dependent upon whether the animal is
a long day or a short day breeder. The regulating mechanism is complicated, since the basis of the
melatonin secretion seems to be the same, irrespective of whether the animal is a long day breeder,
a short day breeder or does not express seasonality in breeding. The interpretation of the melatonin
signal must be different in different species. In the short day breeders such as the sheep, the
breeding season is synchronized to the right time of the year by interpreting the nocturnal melatonin
secretion (together with secondary cues). When the duration of melatonin secretion is long enough,
it alters the pulsatile GnRH-secretion in hypothalamus, which in turn leads to increased pulsativity
(or higher amplitude pulses) in LH-secretion from the pituitary. However, the altered melatonin
secretion can only synchronize the timing of the breeding season within a certain timeframe,

19
because seasonal breeding follows a circannual rhythm. This means that even without any
photoperiodical cues reproductively active and passive (refractory) periods follow after certain
interval (for review see Ebling and Hasting, 1992; Lincoln, 1992).
The site of action of melatonin has been localised within the intracranial central nervous system
using active immunisation or systemic passive immunisation against melatonin (Arendt et al.,
1981), and comparing those results with intraventricular injection of an anti-melatonin serum
(Bonnefond et al., 1989). The melatonin antibodies in the systemic circulation have no access to
intracranial areas, and so systemic immunisation failed to have any effects on photoperiodic
responses, whereas the intraventricular injection of an anti-melatonin serum abolished the
photoperiodic response. The locations of melatonin receptors in the brain are different from those
exhibiting GnRH-neurones. The action of the melatonin is thought to be mediated by interneurones,
rather than being directly on GnRH-neurones. In the sheep, the neurotransmitters involved in
carrying the message have been thought to be catecholamines, endogenous opioids and glutamate
(for review see Ebling and Hastings, 1992; Lincoln, 1992).
The frequency of GnRH-controlled LH-pulses in the sheep is much lower in the non-breeding
season as compared with the breeding season (for review see Ebling and Hastings, 1992). There is
no difference in the number of immunocytochemically identified GnRH-neurons between the
breeding and the non-breeding season in the sheep (Lehman et al., 1986) and the amount of stored
GnRH in the hypothalamus is greater in the reproductively inactive state than in breeding season
(Ebling et al., 1987). Thus, inadequate synthesis of GnRH is not the reason for the inactive breeding
state.
There is also evidence for involvement of oestradiol in the control of GnRH-secretion. Karsch et al.
(1984) extensively reviewed the role of oestradiol in seasonal breeding. Some parts of the review
are discussed in this section. In this model GnRH-pulses are thought to be controlled by the GnRH-
pulse generator, the activity of which depends on inhibitory feedback of oestradiol. The change in
the photoperiod and in the melatonin secretion is thought to affect the sensitivity of the GnRH-pulse
genarator to the oestradiol feedback. In short day breeders, the long duration of melatonin secretion
reduces the negative feedback of oestradiol on the pulse generator and allows increased GnRH-
secretion, which leads to the commencement of the breeding season.

20
It is likely that the seasonality of reproduction in the pig is dependent on photoperiodism and
melatonin secretion, which in turn controls release of GnRH either through neurotransmitters and
activation/inhibition of particular neural pathways, as discussed earlier, or neurotransmitters control
the sensitivity of GnRH-pulse generator to oestradiol feedback. Most likely, both mechanisms are
involved in controlling GnRH-secretion (Robinson et al., 1985). The work by Peacock (1991)
demonstrated that the oestradiol feedback theory could not alone explain the seasonal effects on
reproduction. The photoperiod altered pituitary LH-secretion even if the oestradiol was absent
(ovariectomised gilts).
1.4.4. The melatonin pattern in the pig
The daily melatonin pattern is well established in many species. However, the melatonin profiles in
the pig have remained unclear, despite many studies. Results of the first experiments were
controversial, and it is impossible to draw any common conclusion from them. Most of the studies
were not able to demonstrate any changes between melatonin concentrations during the scotophase
and photophase (Minton et al., 1989; Diekman et al., 1992; Diekman and Green, 1997). Some
authors have reported a nocturnal rise in melatonin only in a small propotion of the animals studied
or only under particular lighting conditions (McConnell and Ellendorff, 1987; Griffith and Minton,
1992; Green et al., 1996; Bollinger et al., 1997). There are also some reports of an exceptional rise
in melatonin during illumination (Peacock, 1991). There is only one study, which has shown a
consistent nocturnal rise in melatonin in domestic pigs under different lighting regimens (Paterson
et al., 1992).
Minton et al. (1989) was not able to demonstrate a nocturnal rise in melatonin concentrations in
pubertal crossbred boars exposed to a 16-hour photoperiod in controlled climate rooms. It was
concluded that melatonin was not secreted in a rhythmic fashion under long photoperiods.
However, the basal melatonin concentrations detected in Minton’s study during the photophase
ranged between 10 and 50 pg/ml, which is a high basal concentration compared with other studies
(Paterson et al., 1992; Mack and Unshelm, 1997). Higher basal concentrations are probably due to a
crossreaction of the antiserum (Guildhay®) with some component(s) other than melatonin or non-
specific serum/plasma effects commonly observed in direct RIAs (Klupiec et al., 1997). Another
study carried out by Diekman et al. (1991) reported extremely high basal melatonin concentrations
in prepubertal gilts without any changes between the dark and light period. In that study,

21
crossreactivity (antisera R1055) was a possible explanation for the high basal concentrations and for
a lack of the circadian melatonin profile, even though the assay was an extraction assay. The study
in question employed chloroform extraction, and it is possible that chloroform also extracted the
substance(s) causing the crossreaction. Furthermore, in Diekman’s study, or at least in some parts of
it, one reason for the lack of nocturnal increase in melatonin might have been a relatively low light
intensity (80-100 lx) used during photophase. Mack and Unshelm (1997) have shown that 50 lx
intensity during illumination did not lead to an increase in melatonin levels during the scotophase
(measured from saliva). A similar problem with crossreactivity seems evident in other experiments
reported on another occasion by the same group (Diekman and Green, 1996). McConnell and
Ellendorf (1987) reported a nocturnal rise in melatonin in three sows out of four under a 12 L : 12 D
lighting regimen. In three sows during the dark phase, concentrations in individuals increased two-
to fivefold over the peak light phase values. When the lighting regimen was changed to 16 L : 8 D
or 8 L : 16 D, the melatonin surge was abolished. The assay was a direct RIA and lacked sensitivity.
The sensitivity of the assay was 16 pg / ml, which is higher than average nocturnal melatonin
concentrations in pig serum or plasma (Paterson et al., 1992). Green et al. (1996) found an increase
in nocturnal melatonin concentrations only in some of the animals and only under certain (close to
12 L : 12 D) lighting regimens. They used a direct RIA utilising an antiserum (Guildhay®) that led
to the high basal melatonin concentrations. Crossreactivity and insensitivity (5 pg/ml) most
obviously also affected the results published by Bollinger et al. (1997). They found a nocturnal rise
in melatonin in only 16 % of the gilts included in the experiment, although they detected a nocturnal
rise in 86.2 % of the ewes housed in the same room. They used an extraction assay but the assay
utilised an antiserum (Guildhay®), which has been shown to crossreact with some small molecular
weight compound(s) present in pig plasma or serum (Klupiec et al., 1997). Measuring melatonin in
the sheep serum does not require such a sensitive assay as does measuring melatonin in pig serum
because the scotophase melatonin levels are much higher in sheep than in pigs.
Crossreaction of the antiserum, non-specific plasma/serum effects in direct RIAs, inappropriate
extraction in extraction based RIAs and insensitivity of the assay are likely to be the most common
reasons for failure to reveal a melatonin profile in the pig. Klupiec et al. (1997) have shown that a
commonly used antiserum (Guildhay®) has some limitations in direct RIAs because of its
crossreactivity with some plasma substance(s) other than melatonin and a non-specific
plasma/serum effect. It seems to be questionable that direct RIAs are adequate for measuring
melatonin in pig plasma or serum because of relatively low melatonin concentrations present
demanding a high sensitivity of the assay. Furthermore, non-specific plasma/serum effect varies

22
considerably between samples from the same individual making correction of plasma/serum effect
difficult (Klupiec et al., 1997). The direct RIAs usually measure higher baseline melatonin
concentrations than extraction assays even if the assay is utilizing highly specific antiserum (G280).
This indicates that the non-specific plasma/serum effect does not exist only with Guildhay
antiserum but also to some extent with other antisera. However, it may be possible to measure
reliably pig melatonin also by employing a direct RIA, if correction of non-specific plasma/serum
effect is done properly and if the validation of the assay includes comparing results with an
extraction assay and/or mass-spectrometry, as shown by Paterson et al. (1992). It seems that an
extraction assay where a plasma or serum effect is eliminated by extraction and reconstituting of the
samples, together with a highly specific antiserum, is the most reliable RIA-method to analyze
melatonin in pigs.
1.4.5. Effects of light intensity on melatonin
The intensity of light has a decisive effect on melatonin secretion. If the intensity of light is not high
enough during the photophase, a rise in plasma melatonin will not take place during the dark phase.
In other words, the intensity of the light during the photophase has to be sufficient to suppress the
melatonin secretion, before there can be any nocturnal rise in melatonin. Griffith and Minton (1992)
showed that 113 lx during the photophase was not sufficiently intense to cause a rise in plasma
melatonin during the dark phase, whereas 1783 lx during the photophase led to a clear rise in
plasma melatonin during the dark phase. These results have to be interpreted with care because of
the difficulties in the assays discussed in the previous section. The exact limit of light intensity
required during the light period to cause high melatonin concentrations during the dark period in the
pig is not known, but some experiments have suggested that it might be somewhere between 200
and 300 lx (Love et al., 1993). If there is not enough light during the photophase, sows might be
unable to recognize the difference between night and day, which is known to be an essential part of
melatonin signals in regulating reproduction (Karch et al., 1988). In addition, it has been suggested
that pigs might be unable to react to rapid changes in lighting conditions (Paterson et al., 1992). It
seems evident that both naturally changing lighting conditions and insufficient artificial lighting
regimens can detrimentally affect reproductive performance of the pig.
Early studies in humans demonstrated that relatively high light intensities were needed to acutely
suppress melatonin secretion. Lewy et al. (1980) reported that 500 lx was not sufficient to suppress

23
melatonin secretion in humans, whereas 1500 lx caused an intermediate suppression and 2500 lx
caused a total suppression. However, Hashimoto et al. (1996) concluded that the threshold of the
light intensity to acutely suppress the nocturnal melatonin secretion in humans was somewhere
between 200 and 500 lx. Humans have been considered to be very insensitive to changes in lighting
conditions but the latest reports of the effect of the illumination on melatonin in humans have
demonstrated that humans are much more sensitive to light than was suspected initially. Boivin et
al. (1996) revealed that light intensity of 180 lx significantly shifts the human circadian pacemaker.
These values are quite similar to the light intensity needed in pigs to acutely suppress melatonin
secretion.
There seems to be a relationship between the light intensity and the illumination time in suppressing
melatonin secretion both in humans and in pigs. In indoor piggeries, controlled by artificial lights,
light intensity and the duration of photophase are the critical factors to consider when planning the
lighting regimens of the different units in piggeries. This is also one of the basic questions to be
considered when trying to prevent seasonal infertility problems in outdoor piggeries. Paterson and
Pearce (1990) showed that if gilts remained isolated from the boars, the mean age of the gilts at
puberty was 209 days when kept under short day lighting conditions and 214 days when kept in
long day regimens. Diekman et al. (1991) revealed that daily oral administration of 3 mg of
melatonin caused a reduction in age of puberty in gilts compared with control gilts. These studies
clearly demonstrated that the photoperiod and melatonin had an effect on reproduction in pigs.
However, details of the role of photoperiod (and melatonin) in controlling the reproductive
performance in pigs needs to be elucidated before different lighting regimens can be introduced to
commercial piggeries or melatonin supplementation can be used.
1.5. RECOGNITION OF PREGNANCY
The following part of the review will concentrate on the recognition and establishment of
pregnancy, since disruption of early pregnancy leading to a reduced farrowing rate seems to be the
most significant manifestation of seasonal infertility, as discussed in earlier sections. Events related
to the luteolysis in cyclic animals, embryos’ ability to prevent luteolysis and the maternal response
to the embryonic signals will be discussed in the following sections.

24
1.5.1. PGF2α secretion during the oestrous cycle
It was shown in the seventies that PGF2α is the hormone causing luteolysis in the sow during the
normal oestrous cycle. The role of PGF2α in controlling the oestrous cycle in the pig has been
reviewed extensively by Bazer et al (1982). The endometrium is the source of PGF2α since its
destruction or hysterectomy before day 14 of the oestrous cycle extends the function of CL beyond
30 days (Melampy and Anderson, 1968). In the cyclic sow, PGF2α secretion starts to increase after
day 12 of the oestrous cycle, and at about day 15 PGF2α concentration reaches its peak value when
measured in the utero-ovarian vein (Moeljono et al., 1977). PGF2α concentration falls to its basal
level between days 18 and 19 of the oestrous cycle. The corpora lutea of the sow appear to be
refractory to the luteolytic effects of PGF2α before day 11 of the cycle as demonstrated by Hallford
et al. (1975). They injected gilts intramusculary with PGF2α at 12-hour intervals on days 4 and 5 of
the cycle. The total dose of 80 mg of PGF2α did not affect oestrous cycle length. However, when the
same treatment protocol was used on days 12 and 13, the cycle length was reduced. Krzymowski et
al. (1976) infused a total of 2 mg of PGF-2α into the anterior uterine vein of sows over a 10-hour
period on either day 6, 8, 10, 12, 14 or 15 of the cycle. The PGF2α did not affect CL weight or
plasma progesterone levels when given on days 6, 8 or 10, but luteolysis was initiated in sows
treated on days 12, 14 or 15. These data clearly demonstrate that CL of the pig are refractory to the
luteolytic effects of PGF2α during the first eleven days of the oestrous cycle. Increased CL
sensitivity to PGF2α after day 11 has been associated with an increase in the number of PGF2α
receptors in the luteal cells (Gadsby et al., 1988). This increase corresponds to the time when LH
begins to dissociate from its membrane receptors (Henderson and McNatty, 1975) and CL become
dependent on a continual pituitary LH support (Anderson et al., 1967).
1.5.2. Embryonic signals
After fertilization has occurred in the oviduct of the sow, embryos move into the uterine horns at the
4-cell stage, about 60-72 hours after the onset of oestrus. Embryos reach the blastocyst stage by day
5 and hatching from the zona pellucida occurs between days 6 and 7. Thereafter, embryos grow to
2-6 mm diameter on day 10 and rapid elongation to a threadlike organism, 10 – 100 cm in length
(Anderson, 1978; Geisert and Yelich, 1997), takes place on days 12 – 16. These filamentous

25
organisms lie in a close contact with the endometrium and are able to give a local signal of their
presence by oestrogen secretion. Van der Meulen et al. (1988) has examined the critical time for the
presence of viable embryos in the uterus in the pig. They flushed blastocysts from the both uterine
horns of gilts on days 10, 11, 12 or 13 and observed effects on the interval to the subsequent oestrus
and progesterone profile. In mated non-pregnant control gilts flushing did not affect progesterone
profile or the cycle length. Flushing of the blastocysts on day 10 resulted in a cycle with a normal
progesterone profile and a normal length. However, flushing on days 11, 12 or 13 resulted in a
normal cycle or in maintenance of the CL for 3-13 days longer than a normal lifespan of CL (in
only one of 6 gilts flushed on day 11). The elevated progesterone levels and prolonged oestrous
cycles were 22.0 ± 1.2 (day 11), 24.8 ± 1.4 (day 12) and 26.3 ± 2.3 days (day 13). There was a large
variation in the stage of the development between embryos flushed on the same day. Only those
gilts in which the blastocysts were ≥ 8 mm or filamentous showed the prolonged oestrous cycle.
The authors concluded from these results that the first signal for maternal recognition of the
pregnancy is generated on day 12 and that blastocysts ≥ 8 mm or filamentous are required for
prolongation of CL function for three or more days. They also concluded that a second signal seems
necessary to maintain CL for the whole pregnancy. It is also an interesting finding that development
of embryos during the first twelve days of pregnancy varied considerably and it contributed to the
embryos’ capability to produce the signal. It has been well documented that endometrial secretions
(histotrophe), which are necessary to nourish embryos, are progesterone-induced. Abnormally low
progesterone concentrations in early pregnancy might interfere with the growth of embryos, and
hence their ability to signal their presence early enough. During the first twelve days of pregnancy
when CL are still working autonomously, subnormal progesterone concentrations may be related to
the feeding level provided to sows and especially to gilts. The energy level of feeding has effects on
progesterone concentrations through metabolic clearance of progesterone rather than on
progesterone secretion from CL (for review see Foxcroft, 1997). After day 12 when CL become
increasingly dependent upon pituitary LH (luteinizing hormone), inadequate LH secretion might
cause subnormal progesterone concentrations and failure of embryos to produce the second signal
needed for continuation of pregnancy. The absence of the second signal might play an important
role in the early disruptions of pregnancy seen typically during the seasonal infertility period, since
season has been shown to affect LH secretion (Paterson et al., 1992; Peltoniemi, 1994).
Several investigators have suggested that embryos give the signal, discussed earlier, by producing
oestrogen. Perry et al. (1973) first described the physiological role of oestrogen in the maintenance

26
of pregnancy in pigs and these results have been confirmed and expanded later by others (Gadsby et
al., 1980; Fischer et al., 1985). Conceptuses of the pig produce oestrogens between days 11 and 18
in a biphasic manner. Uterine concentration of oestrogen increases for the first time on day 11 or
12. At this same time conceptuses go through rapid elongation from round shaped blastocysts to
long filaments. This stage is followed by a decrease in uterine oestrogen content between days 13
and 14 before a new increase after day 14 (Stone and Seamark, 1985). An increase in uterine
content of oestrogen does not occur before day 15 in non-pregnant gilts or sows. Since the capacity
of endometrium to synthesize oestrogen in the early pregnancy is low, the uterine content of
oestrogen reflects the embryos’ oestrogen production (Fischer et al., 1985). It has been
demonstrated that pig blastocysts are very efficient in synthesizing oestrone and oestradiol from
different precursors including progesterone (Fischer at al., 1985). These “free oestrogens” produced
by embryos are most likely acting only locally in the endometrium, because they are conjugated to
biologically inactive sulphated forms in the endometrium as they move into systemic circulation
(Heap and Perry, 1974), but the possibility of the systemic effects can not be rejected.
Experiments where oestrogen has been injected into cyclic gilts have provided further evidence of
oestrogen being the signal that embryos produce to ensure prolonged CL function. Pusateri et al.
(1996a) carried out on experiment where they determined minimal requirement for exogenous
oestradiol-17β to induce either a short or long pseudopregnancy in cycling gilts. They carried out
five experiments where oestradiol-17β was injected into the cyclic gilts in different combinations
between days 11 and 25 post oestrus. The results demonstrated that short pseudo pregnancy (SPP)
can be induced in gilts by oestrogen treatment that is effective over a 2-day period between days 11
and 15 of the oestrous cycle. The most consistent induction occurred when the 2-day period
encompassed day 12 or 13, or both. The long pseudo pregnancy (LPP) was most effectively induced
by the injection regimen which begun on day 12 and continued with daily injections to between
days 17 and 19. None of the gilts that received daily injections from day 12 to day 16 exhibited
LPP. The duration of oestradiol treatment required to induce pseudopregnancy was clearly
demarcated between SPP and LPP, being 23-35 days in SPP and over 50 days in LPP. The authors
therefore suggested that maternal recognition of pregnancy might occur in two phases with
continuous exposure to oestradiol being required from day 12 to 17-19 of pregnancy. In this
experiment they used oestradiol-17β because of its shorter half-life compared with oestradiol
valerate or oestradiol benzoate. Intramuscular injection of oestradiol-17β at dosages of 4 to 17
µg/kg resulted in elevated serum oestradiol concentrations for 16 to 18 hours whereas peripheral

27
oestradiol remained elevated for at least 72 hours after a single injection of 5 mg oestradiol valerate.
After daily injections of 5 mg of oestradiol valerate for five days, serum oestradiol concentrations
remained above the preinjection baseline for five to seven days. Earlier studies done with oestradiol
valerate or benzoate did not provide accurate indications of the exact time when the oestrogen
signal has to be initiated to cause pseudopregnancy because of the long residual effects of these
compounds.
1.5.3. Oestrogen signals alter PGF2α secretion during pregnancy
There is common agreement that embryonic oestrogen signals are the factor causing the
maintenance of CL, but there is not unanimity of opinion on the mechanism of action. Several
mechanisms have been proposed and they include both systemic and local effects of oestrogen. It
appears that oestrogen is capable of stimulating progesterone secretion of CL possibly through an
increase in luteal LH (Garverick et al., 1982). However, the inability of embryos to maintain the
pregnancy when a large portion of the uterus is unoccupied or when the pregnancy appears only in
another horn of the uterus does not support the theory of systemic effects (see Dziuk, 1985). It has
also been proposed that oestrogens exert their effects by reducing endometrial release of PGF2α
(Guthrie and Rexroad, 1981). Although several studies have indicated that in vitro PGF2α
production of pregnant gilts’ endometrium or oestrogen treated cyclic gilts’ endometrium is
diminished compared with non-treated cyclic gilts’ endometrium, the luminal concentration of
PGF2α is greater on days 14-20 in oestrogen treated than in cyclic gilts (Frank et al., 1977). The
PGF2α concentrations are even greater in pregnant animals during this time (Zavy et al., 1980) that
might reflect embryonic prostaglandin production. The elevated PGF2α concentrations found in the
uterus of pregnant animals support the most widely accepted theory, proposed by Bazer and
Thatcher (1977), based on reorientation of endometrial PGF2α secretion. The secretion of PGF2α
during the mid and late luteal phase in the cyclic animals is mainly toward the uterine venous
drainage (endocrine direction) causing the regression of CL and allowing follicular development to
progress and oestrus to occur. In pregnant sows or gilts, oestrogens produced by the embryos on
days 12 and 18 alter the PGF2α secretion toward the uterine lumen (exocrine direction) without
reaching the systemic circulation, and the regression of CL does not take place. There are a number
of studies which have shown that PGF2α concentration in the utero-ovarian vein is higher in cyclic
animals between days 12 and 17 compared with pregnant or oestrogen treated animals (Frank et al.,

28
1977; Moeljono et al., 1977). It has also been shown that peripheral concentrations of PGF-
metabolites are higher in cyclic gilts than in pregnant gilts on days 12-13 and 15-17 reflecting the
reorientated PGF2α secretion into the uterine lumen in pregnant gilts (Shille et al., 1979).
The exact mechanism by which oestrogen is able to reorientate the PGF2α secretion is not known
but endometrial calcium release would appear to be involved. There is an increase in uterine
calcium concentration about twelve hours after the embryonic oestrogen signal (or after exogenous
oestrogen) and a re-uptake of calcium by the endometrium occurs about twelve hours after the
concentration of calcium in the uterine lumen reaches the maximum level. This oestrogen induced
endometrial calcium secretion and re-uptake has been associated with the reorientation of PGF2α
secretion. More evidence of calcium being involved in alteration of endometrial prostaglandin
release has been provided by Gross et al. (1990). This study demonstrated that treatment with
calcium ionophore shifts orientation of PGF2α secretion to the luminal side during in-vitro
perfusion. Reorientation of PGF2α secretion to the luminal side also involves interaction between
oestrogen and prolactin, since neither oestrogen nor prolactin alone causes the reorientation (Gross
et al., 1990). There is no increase in prolactin concentration in early pregnancy, but the number of
prolactin receptors increases between days 14 and 30 (Dehoff et al., 1984). There are also some
other factors, like embryonic prostaglandin-E (PGE) and conceptus secretory proteins (CSP), which
might have an effect on the reorientation of endometrial PGF2α secretion.
It appears that two embryonic oestrogen signals are needed between days 11 and 18 to extend the
function of CL until the end of the normal pregnancy period. Pusateri et al (1996b) demonstrated
that in all oestrogen treated cyclic gilts serum prostaglandin-metabolite concentrations peaked
always 4-6 days before the post-treatment oestrus, independent of the treatment response (SPP or
LPP). This indicates that the second signal is needed around pregnancy day 18 to ensure the
continuation of the endometrial PGF2α exocrine secretion pattern. It seems that no further signals
are needed after that, since two appropriately timed exogenous oestrogen signals are able to cause
the LPP of the normal pregnancy length (Pusateri et al., 1996b).
1.5.4. Conceptus secretory proteins (CSP)
In sheep and cattle, maternal recognition of pregnancy is thought to be initiated by the conceptus
secretory proteins, ovine trophoblast protein-1 (oTP-1) and bovine trophoplast protein-1 (bTP-1),

29
which are members of interferon-alpha (IFN-α) family. In these species, trophoplast proteins seem
to prevent the ability of oestrogen and oxytocin to stimulate endometrial PGF-2α secretion that
initiates the regression of CL. Cross and Roberts (1989) showed that IFN production by early
conceptuses occurs also in pig, between days 11 and 17, but the role of this IFN is unknown. A
large body of evidence suggests that in the pig the events related to prevention of luteolysis are
triggered by embryonic oestrogen secretion, as discussed earlier, and not by the IFN-secretion as in
sheep and cattle. However, it is likely that IFN plays an important role in establishment of
pregnancy also in the pig, perhaps independent on effects on luteal life span. Other potential roles of
conceptus IFN include regulation of the local maternal immune system, regulation of endometrial
secretions and induction of antiviral protection against infection (Cross and Roberts, 1989; La
Bonnardière, 1993). However, at this stage the potential roles of the IFN in recognition and
establishment of pregnancy in the pig are speculative and further investigations are needed.
1.6. MATERNAL RESPONSE
1.6.1. Formation of CL and luteotrophic role of LH
After ovulation the newly formed corpora lutea continue to develop autonomously independent of
pituitary support until day 15, when PGF2α secretion reaches its peak value, and if pregnancy has
not been established, regression of CL are initiated. Sammelwitz et al. (1961) showed that large
amounts of progesterone injected into pregnant pigs from the time of ovulation until days 10 to 13
of gestation did not prevent the formation of CL. Injections of progesterone beyond days 12 to 16 of
pregnancy resulted in the rapid and complete regression of CL. Large amounts of exogenous
progesterone are thought to cause a complete blockade of pituitary LH secretion through the
negative feed-back mechanism. Brinkley et al. (1964) reported similar results where formation of
CL after ovulation was not affected by the pituitary LH-blockade caused by exogenous
progesterone lasting from seven to ten days. These two experiments provide evidence that CL of
pigs are able to function autonomously until day 15, but after that the pituitary LH-support appears
to be necessary for continuation of pregnancy.
The work of Anderson et al. (1967) provides further evidence that establishment of pregnancy
needs firstly the reorientation of endometrial PGF2α secretion and secondly a certain requirement
for pituitary secretion of LH. In their study hypophysial stalk transection or hypohysectomy was

30
performed on pigs on the 1st day after oestrus or mating. Corpora lutea developed following the
hypophysial stalk transection, but by day 12 they were smaller in unmated stalk-sectioned animals
than those in mated stalk-sectioned, as well as being smaller than those in day 13 unmated or mated
control pigs. By day 20 CL regressed in both mated and unmated stalk-sectioned animals. Living
embryos were present in the uterus of mated stalk-sectioned pigs at days 12 and 16, but only
necrotic embryos were found at day 20. In hysterectomised animals with persisting CL, luteal
regression begun within four days after hypophysial stalk transection and was completed by day 12.
Oestrogen maintained the CL in intact control animals but not in stalk-sectioned pigs. Desiccated
porcine anterior pituitary or human chorionic gonadotrophin maintained CL in hypophysectomised
and hysterectomised pigs. Collectively these data confirm the autonomous function of the CL about
15 days following the ovulation and the essential role of pituitary LH in the maintenance of CL and
continuation of pregnancy after CL have lost their autonomous function.
The autonomous function of CL appears to disappear gradually at the same time as the PGF2α
secretion increases. Pituitary LH has to take a supporting role for CL and this support has to
continue until around pregnancy day 29 as demonstrated by GnRH-agonist implantation of gilts in
different stages of pregnancy (Peltoniemi, 1994). GnRH-agonist implants abolish the LH-surges
and if the implantation occurred on day 14 or 21 of pregnancy all the gilts aborted. However, if
implantation occurred on day 29, 50 % of the gilts were able to maintain the pregnancy. These
results indicate increasing independence of CL on pituitary LH when pregnancy progresses.
In the early stage of pregnancy, prolactin concentrations measured in pregnant gilt serum are very
low (Peltoniemi, 1994). However, prolactin is known to be the primary luteotrophin beyond day 60
of pregnancy in pigs (Szafranska and Tilton, 1993). It is not well understood how maintenance of
CL are ensured between days 30 and 60, when pituitary LH is no longer critical to the continuation
of pregnancy and before prolactin has taken a role of primary luteotrophin. This might be some kind
of transient period that is not extremely sensitive to the lack of either of these luteotrophins.
In conclusion, pituitary LH appears to be essential to the continuation of pregnancy between days
15 and 29. At that time, corpora lutea gradually lose the autonomous function and become
dependent on pituitary luteotrophin. After day 29, prolactin gradually takes its place as a primary
luteotrophin.

31
1.6.2. Progesterone in early pregnancy
Corpora lutea are the main source of progesterone in pigs throughout pregnancy, and functional CL
are needed for the entire length of pregnancy. The maintenance of CL is ensured by the prevention
of PGF-2α secretion into ovarian circulation and pituitary luteotrophic support as discussed earlier.
Progesterone induces the secretion of different kind of proteins like uteroferrin and retinol binding
proteins (RBP), which act by transporting vitamins and minerals to the developing conceptuses.
There is a wide consensus that progesterone is the key hormone inducing the secretion of these
proteins essential for development of conceptuses, but the effect of embryonic oestrogen on
regulation of this secretion is unsure. Trout et al. (1992) reported that while progesterone
concentration is sufficient to allow the secretion of RBP, oestrogens secreted by embryos have a
major influence on the amount and timing of RBP secretion. In contrast to this study, Vallet et al.
(1998) concluded from their experiments that conceptuses do not influence secretion of either total
protein or RBP during pregnancy and that the onset of secretion of these proteins is controlled by
progesterone alone. However, progesterone concentration seems to be critical during and after the
time CL begin to be dependent on pituitary LH. This might be because of the effect of progesterone
on secretion of nutrients to embryos, effect of progesterone and embryonic oestrogens together on
uterine secretion or effect of progesterone on embryos’ ability to produce an oestrogen signal to
prevent the luteolysis.
In normal pregnancy, there is even 30 % decrease in maternal progesterone around day 14 of
pregnancy, and this decrease has been proposed to be due to metabolism of conceptuses to produce
the oestrogen signal (see Bazer et al., 1982). Vallet et al. (1998) found that increase in progesterone
induced protein secretion always preceded the change in embryonic oestrogen secretion. They
proposed that the uterine protein secretion (induced by progesterone) might control conceptus
oestrogen secretion. Down-regulation of epithelial progesterone receptors in uterine epithelia occurs
after day 10 of gestation but receptors remain in deeper layers and might control endometrial
secretion throughout the pregnancy (Geisert et al., 1994). Even if all details of the role of
progesterone in early pregnancy are not clear it seems to be related to the recognition of the
pregnancy through the embryonic oestrogen signal. Progesterone may be the primary precursor of
oestrogen produced by embryos or progesterone may control embryos’ growth by protein secretion
and their ability to produce oestrogens when needed. However, the changes in maternal
progesterone concentrations at the same time as down-regulation of uterine epithelial progesterone

32
receptors and need for the embryonic oestrogen signal, are all part of critical events associated to
establishment of pregnancy.

33
2. THE AIMS OF THE STUDY
The primary aim of the study was to investigate the endocrinological basis of seasonal infertility
concentrating on the photoperiodic – melatonin transduction. The secondary aim of the work was to
study association between early disruption of pregnancy (EDP) and lowered farrowing rate in the
summer – autumn as well as the mechanism of EDP.
1) The first experiment where European wild boars (gilts and young boars) and domestic gilts
were studied in the same trial was aimed to give a basis to all the rest lighting experiments,
as there was a fundamental lack of information regarding the lighting – melatonin
transduction in the pig.
2) The other two lighting trials were aimed to give more detailed information of the ability of
the pig to respond to abrupt changes in lighting regimens and effects of the light phase light
intensity on the dark phase melatonin response.
3) The association between EDP and seasonally decreased farrowing rate was studied by a
field trial in a commercial herd having a constant history of seasonal infertility.
4) The endocrinological mechanism of the early disruption of pregnancy was investigated. The
aim was also to develop an experimental model of the mechanism for further studies. This
was done by active and passive GnRH-immunization trials.

34
3. MATERIALS AND METHODS
The experiments of the study were divided into two main categories; lighting – melatonin
transduction trials (papers I, II, III ) and trials concerning early disruption of pregnancy (papers IV,
V). This division was justified by the fact that seasonal changes in the circadian secretion pattern of
melatonin are behind the seasonally altered reproduction performance, whereas early disruption of
pregnancy appears to be the most important single manifestation of seasonal infertility in practise.
Both areas of the study were further divided into a field trial and strictly controlled laboratory
investigations.
Materials and methods that were used only in one of the studies or were similar in all the studies are
described here in details. Those materials and methods that varied between studies (number of
animals, management, feeding) are described in details in the paper concerned, and only general
guidelines are given in this section.
3.1. ANIMALS
The trial where melatonin secretion of wild boars was compared with that of domestic pigs (paper I)
used pure bred European wild boars (young boars and gilts) and crossbred (Yorkshire x Landrace)
non-pregnant gilts. The European wild boars were from a wild boar farm where the animals lived in
a semi natural environment. The population of the wild boars originated from animals captured
from the nature.
In the light intensity experiment (paper III), eight-month old Yorkshire-Landrace and Yorkshire-
Landrace-Duroc hybrid non-pregnant gilts were used.
Pre-pubertal male grower pigs were used in the study where ability of pigs to respond to abrupt
changes in lighting was investigated (paper II). The body weight of 40 – 45 kg was used as an
inclusion criterion.
The field trial investigating association between seasonally lowered farrowing rate and early
disruption of pregnancy (paper IV) used all the animals (gilts and sows) in production in a

35
commercial 160-sow unit. Most of the animals were hybrids (Yorkshire x Landrace) but
occasionally there were some purebred Yorkshire or Landrace gilts/sows.
The immunization experiments (paper V) used pregnant multiparous crossbred (Yorkshire x
Landrace) sows and pregnant crossbred (Yorkshire x Landrace) gilts.
3.2. BLOOD COLLECTION
3.2.1. Jugular vein catheters (I, II, III, V)
The frequent blood samples from domestic pigs were collected via jugular vein catheters inserted
non-surgically via an ear-vein (Peacock, 1991). Pigs were restrained using a soft rope snare and the
surface of the ear was cleaned and disinfected. A 14-gauge intra-venous catheter placement unit
(Jelco®, Johnson & Johnson, Chiyoda-Ku, Tokyo) was inserted into the ear-vein and approximately
50 cm of vinyl tube was threaded through the catheter into the vein and another 50 cm remained
externally. The vinyl tube (Dural Plastic and Engineering, Auburn, NSW) had an inner diameter of
1.0 mm and an outer diameter of 1.5 mm. A blunted 18-gauge needle hub was fitted into the end of
the vinyl tube to allow blood collection and a stopper was used in the needle hub to prevent a
backflow. To ensure the patency of the catheter, it was filled with sterile EDTA-saline (9 g NaCl
and 9 g di-potassium EDTA in 1 l of sterile water). A zippered pouch placed on the pig's neck was
used to store the external part of the vinyl tube and the stoppered needle hub. The catheter was
bandaged on the ear using elastic adhesive bandage (Elastoplast®, Smith and Nephew, Clayton,
Vic.) and the ear was then strapped to the neck to prevent the pig from dislodging the catheter. Prior
to collecting a blood sample, the first 2 ml drawn up was discarded to prevent contamination of the
sample with saline. The sample (approximately 8 ml) was drawn into a 10 ml syringe and
immediately emptied either into 10 ml plastic tubes containing 150 µl EDTA solution (250 mg/ml)
and 0.25 g spinning granules (Kwik Spin®, Disposable Products, Ridleyton, S.A.) or into plain
glass tubes.
3.2.2. Single blood samples (IV, V)

36
When single blood samples were needed, they were taken as a vein puncture of V. jugularis or V.
saphena medialis.
3.2.3. Saphenous arterial catheters (I)
The frequent blood samples of the wild boars were collected by the non-surgical method described
by Tast et al., 2000. Animals were sedated with a combination of medetomidin (Domitor® 1 mg /
ml, Orion-Farmos, Finland), zolazepam and tiletamine (Zoletil forte vet® 50 mg / ml + 50 mg / ml,
Virbac Laboratories, France). The dose of Domitor® was 0.025 mg / kg and of Zoletil® 5 (2.5 +
2.5) mg / kg. An air-pressure sedation gun with self-emptying shooting syringe and needle (both
from Dan Inject®, Denmark) was used to inject the sedatives intramuscularly. With the animal in
lateral recumbancy, the hair was cut from the medial surface of the hind leg (Musculus gracilis) and
the Arteria saphena medialis was palpated. The skin was washed with water and disinfected with
clorhexidin (Hibiscrub®, Zeneca Pharma, Norway). A small incision (2 mm) was made in the skin
with an injection needle. An Intraflon® 14 G catheter (Intraflon 2® 14 G, Vygon, France) was
inserted into the Arteria saphena medialis, and the inner metallic needle removed. A teflon tube
(Prevett, Finland, inner diameter 0.9 mm and outer diameter 1.5 mm.) was placed into the artery
through the Intraflon® 14 G catheter. Approximately 40 cm of this tube was inserted into the artery,
and the Intraflon® 14 G catheter was withdrawn. The Intraflon® 14 G catheter was attached by a
plastic clue to the outer end of the tube, where it supported the connection between the tube and the
19 G injection needle that had earlier been attached to the end of the tube. To ensure that the tube
was functioning properly, 10 ml of blood was aspirated, and 1 ml of diluted heparin (Heparin 5000
IE / ml®, Lövens, Denmark) was injected into the tube to prevent coagulation. The hair was cut
from the ventral side of the hind limb up to the back, and the tube was covered by tape
(Elastoplast® 6 cm, Beiersdorf AG, Spain). Interrupted skin sutures (Suturamid® 0, Ethicon,
Germany) at 5 to 10 cm intervals secured the tape to the skin .A blunted hypodermic needle hub (19
G) was fitted to the end of the tube, with a stopper to prevent back flow. The tape covered the
needle hub until the sampling begun. When sampling started, a small hole was made in the tape
allowing the 19 G injection needle to exit. The blood sample was drawn into a 10-ml syringe and
emptied immediately into a plain glass tube. The samples were refrigerated immediately and serum
was separated by centrifugation on the following day. The serum was stored at -20°C until analysis.

37
3.3. GNRH-IMMUNIZATION
3.3.1. Active immunization (V)
A commercially available vaccine (Improvac®, CSL Limited, Australia) was used to actively
immunize sows against GnRH. The first immunization was done on the day of farrowing injecting 2
ml of the vaccine subcutaneously. The booster was given 10 or 20 days after mating.
3.3.2. Passive immunization (V)
A number of cull sows and finisher pigs were immunized with Improvac® two or three times at
four-week intervals with the last vaccination given 14 days before culling to obtain GnRH-immune
pig serum. Blood was collected at slaughter into plastic containers and allowed to clot at room
temperature over night. On the next day, blood was centrifuged and serum harvested. Serum was
stored at -20ºC until used. Before use, serum was warmed to +38ºC. On day 12 post mating, the
gilts were infused intravenously with 1 L of pig serum containing GnRH-antibodies.
3.4. HORMONE AND ANTIBODY ASSAYS
3.4.1. Melatonin (I, II, III)
Melatonin concentrations of plasma/serum samples were determined by a commercially available
double-antibody radioimmunoassay (Bühlmann Laboratories AG, Switzerland) based on the
Kennaway G280 anti-melatonin antibody (Kennaway et al., 1982). Klupiec et al. (1997) have
demonstrated specificity of G280 antiserum for melatonin extracted from porcine plasma. The
sensitivity of the assay was 0.3 pg/ml.
As a modification for manufacturer’s instructions the ether extraction was used rather than column
extraction in the wild boar study (I). Before assay, controls and samples were extracted twice with
4.5 ml of diethyl ether. The tubes were shaken for 1 minute and put into a freezing bath. The

38
supernatant was decanted and the solvent was evaporated to dryness in a 37 ºC water bath. The
residue was dissolved in 1 ml of incubation buffer and proceeds with assay according to
manufacturer’s instructions with some minor modifications (described on the next page).
The samples of lighting program trials (II, III) were assayed in the Department of Obstetrics and
Gynaecology, University of Adelaide. This laboratory has originally provided the antiserum used in
the commercial Bühlmann Melatonin radioimmunoassay and the laboratory currently continues its
co-operation with Bühlmann Laboratories. The assay described in following paragraphs was
basically the same as the commercial one, but some components, which normally are included in
the commercial kit, were made in the laboratory following the manufacturer’s directions and with
the manufacturer’s permission. There were also some minor modifications in the extraction and
assay procedures. The components of the assay were as follows:
1. Extraction columns: The extraction columns (RK-MEL2, Bühlmann Laboratories,
Switzerland) containing charcoal (C18) were provided with the assay kit. All columns were
used up to five times if blockage of the columns did not occur earlier.
2. Melatonin buffer: Melatonin buffer is included to the commercial assay kit. Because of the
great number of samples analyzed the buffer was made up in the laboratory according to the
manufacturer’s directions, and contained di-Sodium Hydrogen Orthophosphate dihydrate
(14.2 g), Sodium Dihydrogen Orthophosphate (2.9 g), Sodium chloride (8.7 g), BSA (5 g)
and Sodium Azide (0.2 g) in one litre of distilled water.
3. Standards: One set of five vials, each containing different amount of melatonin, was
provided with the assay kit. The standards were reconstituted prior assaying adding 5 ml of
melatonin buffer into each vial, vortexed and incubated at room temperature for 1 hour and
vortexed again. After reconstitution, the standards contained 0.5, 1.5, 5, 15 and 50 pg/ml of
melatonin respectively.
4. Controls: One set of two vials containing 4.5 ml of human serum with lot-specific amounts
of melatonin (LOW and HIGH concentrations) were included in the assay kit. The quality
controls (LOW and HIGH) were included in the beginning and end of each assay to
calculate intra and inter assay coefficients of variation (CV).

39
5. Antiserum: The antiserum was made up in the laboratory adding 6.6 µl of stock antibody to
100-ml of melatonin buffer. In the commercial assay kit, antiserum is provided in ready-to-
use form.
6. Melatonin tracer: The tracer (125I γ-iodomelatonin) was diluted in the laboratory adjusting
total counts to be about 10 000 counts in 100 µl of the diluted tracer. The needed amount of
the tracer was approximately 75 µl in 100-ml of melatonin buffer. In the commercial assay
kit tracer is provided in ready to use form.
7. Second antibody: The solid phase bound anti-goat second antibody (RK-MEL2) was
provided in ready-to-use form.
Extraction and assay procedure:
All the samples were thawed the day before assay, stored in the refrigerator and taken to room
temperature one hour before extraction. Prior to extraction, samples were centrifuged for 10 min at
500 x g to remove any small clots in the plasma/serum that can block the extraction columns. The
extraction columns were inserted into the extraction tubes (Pyrex® 16x125 mm, Corning
Laboratory Science Company, USA) and conditioned two times adding 1 ml of methanol to each
tube and centrifuging for 1 min. at 200 x g. Columns were washed twice by adding 1 ml of water
and centrifuging for 1 min at 200 x g before loading columns with standards and samples. One
millilitre of melatonin buffer was added to columns representing NSB (non-specific binding) and
MB (maximum binding) tubes and 1 ml of standards and samples into respective columns and
centrifuged for 1 min at 200 x g. Columns were washed twice with 10 % methanol and once with
hexane centrifuging for 1 min at 500 x g between washings to remove everything but melatonin
from the columns. After washing, columns were transferred into borosilicate glass tubes (Kimble®
13x100 mm, ASG Inc., USA) and 1 ml of methanol was added into the columns and centrifuged for
1 min at 200 x g to remove the extracted melatonin from the columns. Methanol was evaporated
from the tubes using airflow and heating blocks. Once evaporated, the extracted samples were
reconstituted by adding 1 ml of melatonin buffer into the tubes, vortexed and allowed to equilibrate
30 min at room temperature. Once the reconstituted samples were equilibrated, the assay proceeded
by adding 500 µl extracted buffer into the NSB tubes, 400 µl extracted buffer into MB tubes and
400 µl extracted standards and samples into the respective tubes. Melatonin antibody (100 µl) and

40
tracer (100 µl) were added into each tube except into the NSB tubes, where only tracer was added.
Into the total count tubes 100 µl tracer was added. All the tubes were vortexed and incubated
overnight in a cold room. On the next day 25 µl of the second antibody was added into all the other
tubes but total counts, vortexed and incubated 30 min in a cold room. One millilitre of cold water
was added into all the tubes except the total counts and centrifuged for 15 min at 2000 x g. The
supernatant was decanted and radioactivity of the pellets was counted by γ-counter (Wallac®,
Turku, Finland).
Table 2. The cross-reactivities of the melatonin antiserum shown above were found at 50 % binding (Bühlman Laboratories, 1998).
C om p ound C rossreactivity
M elatonin 100 %
Seroton in < 0.001 %
5-H ydrox y-in doleacetic acid < 0.001 %
N -A setylseroton in 0.027 %
5-M eth oxytryptam in e 0 .003 %
5-M eth oxytryptoph an 0 .001 %
6-Sulfa toxym ela tonin 0 .002 %
5-M eth oxytryptoph ol 0 .001 %
3.4.2. Progesterone (IV, V)
Plasma progesterone concentrations were determined using a commercially available, solid-phase
progesterone RIA (Spectria®, Orion Diagnostica, Finland), which has previously been validated to
measure progesterone in pig plasma (Peltoniemi, 1994). In the assay, 50 µl of plasma sample and
500 µl of buffered I125-label were added to the antibody-coated tubes. The tubes were incubated for
two hours at room temperature and the supernatant was poured out. Radioactivity of the tubes was

41
counted by a γ-counter (Wallac®, LKB-Wallac, Turku, Finland). Sensitivity of the assay was 0.09
ng/ml.
Table 3. Cross-reactivity of the progesterone antiserum (Orion Diagnostica, 1999)
Compound Crossreactivity
progesterone 100 %
Pregnenolone 3.9 %
Corticosterone 0.9 %
5β-Dihydroprogesterone 0.75 %
11-Deoxycorticosterone 0.38 %
5α-Dihydroprogesterone 0.22 %
20β-Hydroxyprogesterone 0.045 %
17α-Hydroxyprogesterone 0.017 %
3.4.3. Luteinizing Hormone (V)
Plasma LH concentrations were determined using a direct double-antibody RIA as previously
reported (Niswender et al., 1970), with modifications reported by Peacock (1991). Purified porcine
LH (LER-786-3) was supplied Professor L.E. Reichert Jr. and radio-iodinated with I125 (Amersham
Australia Pty.Ltd.). In the iodination procedure, the Chloramine-T method described by Greenwood
et al. (1963) was used. After iodination, the label was stored at 20°C and used within six weeks.
0.01 M phosphate buffer with EDTA was used to dilute the label to give approximately 10 000
counts.
The antiserum used was rabbit anti-porcine LH (Niswender No. 566), donated by Professor G.
Niswender. It was frozen in a final dilution of 1:17 000 in the buffer. A solid-phase second antibody
coated cellulose suspension (Sac-Gel®, A-SAC1, Immunodiagnostics, Waverley, Australia) raised
in donkeys against rabbit serum was used to separate bound and unbound label. An assay buffer

42
(0.133 g NaH2PO4.2H2O, 1.299 g Na2HPO4, 9.000 g NaCl, 1.000 g BSA, 1.000 g NaN3 added to 1
L of distilled water) with additional 1 % Bovine Serum Albumine (BSA) was used to prepare the
standards. One hundred micro liters of the purified LH was added to 3.9 ml of the buffer. Two
milliliters of the previous standard was serially diluted into 2 ml of the buffer to obtain standards
from 16 to 0.125 ng/ml.
Two hundred microlitres of sample or standard were mixed with 600 µl of the buffer. Antiserum
(100 µl) was added and the tubes were incubated for 24 hours at 4°C. After the incubation, 100 µl
of the labeled LH was added and incubated for 48 hours at 4°C. The second antibody was then
added (100 µl), incubated for 30 min at room temperature and centrifuged for 8 min at 2000 rpm.
The supernatant was decanted and the tubes were counted on a gamma counter (Wallac®, LKB-
Wallac, Turku, Finland).
3.4.4. GnRH-antibody (V)
GnRH-antibodies from immunized animals and from the antiserum pool were analyzed using RIA
described by Carson et al., 1997. Iodinated GnRH (Amersham International Limited, Amersham,
England) was diluted to approximately 10 000 counts per minute per 100 µl of assay buffer (50 mM
sodium phosphate, 10 mM EDTA and 0.3 % BSA; pH7.2). One hundred microliters of diluted
samples were incubated at 4°C overnight in test tubes (75 x 12 mm; GS Ross Limited,
Macclesfield, England) with 100 µl of the labeled GnRH and 300 µl of assay buffer. Then 250 µl of
dextran (Pharmacia Fine Chemicals, Uppsala, Sweden) coated charcoal (Sigma) was added to
separate free from bound labeled GnRH. The tubes were centrifuged at 4°C for 15 min at 2000 rpm.
and the supernatant decanted and counted on a gamma counter (Wallac®, LKB-Wallac, Turku,
Finland). Titres were calculated by expressing binding of labeled GnRH minus non-specific bound
counts as the percentage of the total available labeled GnRH.
3.5. PREGNANCY TESTING
The sows and gilts were pregnancy tested trans-cutaneously (IV, V) using a real time ultrasound
(Hondex®, Honda Electronics Co. Ltd., Japan) with a linear probe (5 MHz).

43
3.6. LIGHTING PROGRAMS
The lighting experiments (II, III) were done in temperature and lighting controlled climate rooms,
except the wild boar trial (I), in which the wild boars were under the natural lighting and domestic
gilts under a typical light environment of an indoor piggery where the artificial lighting was
controlled but the natural light was allowed through the windows. In the climate rooms, the
scotophase light intensity was less than 7 lx (dim red lights) measured at pig eye-level. The light
intensity was measured using a digital light meter (Topcon® IM-2D, Japan) in the studies presented
in papers II and III. Another digital light meter (Gossen Mavolux digital®, Gossen, Germany) was
used in the wild boar trial (I).
3.7. STATISTICAL ANALYSES
Statistical analyses were carried out using Stata Intercooler, version 5.0 statistical package (Stata
Corporation, Texas, U.S.A).
An analysis of variance with repeated measures was used to study variation in melatonin and
progesterone profiles (I, III, IV, V). The method of linear mixed models for log-transformed data
was used to study effects of lighting programs on melatonin secretion (II). Fischer’s exact
probability test was used to test seasonal effect on rate of pregnancy disruption (IV). Where
appropriate, Student’s t-test was used for further analyses (I, IV, V).
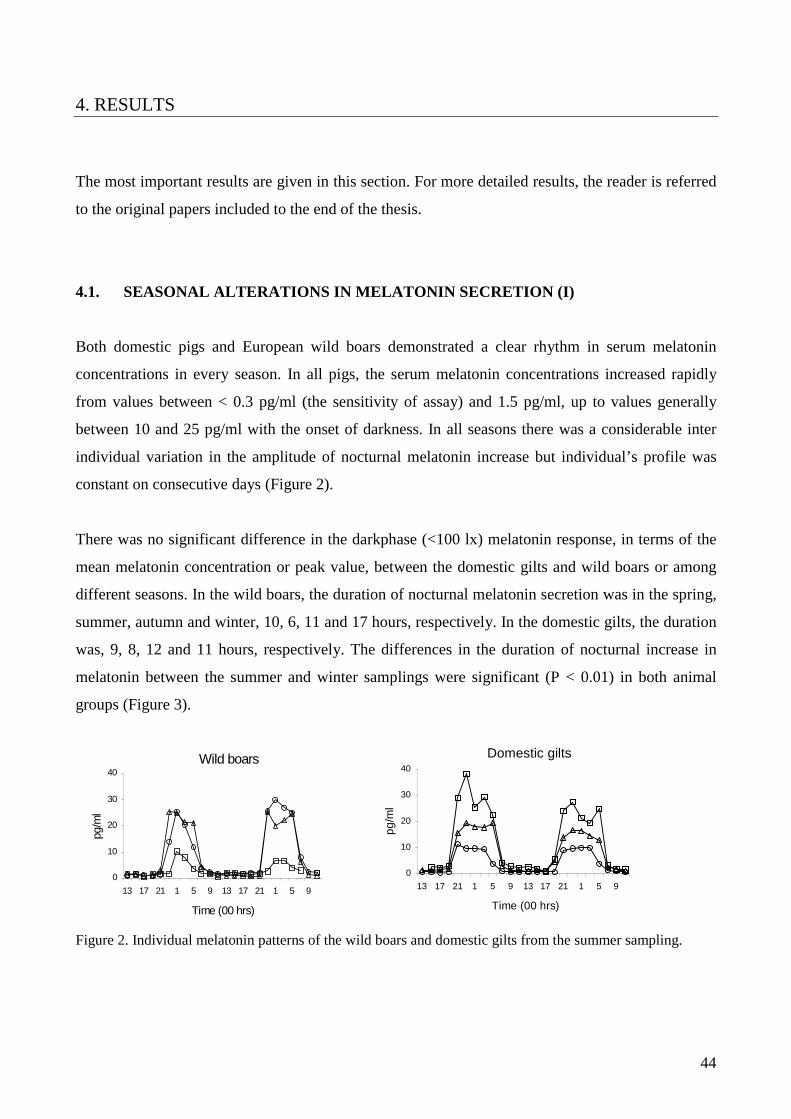
44
4. RESULTS
The most important results are given in this section. For more detailed results, the reader is referred
to the original papers included to the end of the thesis.
4.1. SEASONAL ALTERATIONS IN MELATONIN SECRETION (I)
Both domestic pigs and European wild boars demonstrated a clear rhythm in serum melatonin
concentrations in every season. In all pigs, the serum melatonin concentrations increased rapidly
from values between < 0.3 pg/ml (the sensitivity of assay) and 1.5 pg/ml, up to values generally
between 10 and 25 pg/ml with the onset of darkness. In all seasons there was a considerable inter
individual variation in the amplitude of nocturnal melatonin increase but individual’s profile was
constant on consecutive days (Figure 2).
There was no significant difference in the darkphase (<100 lx) melatonin response, in terms of the
mean melatonin concentration or peak value, between the domestic gilts and wild boars or among
different seasons. In the wild boars, the duration of nocturnal melatonin secretion was in the spring,
summer, autumn and winter, 10, 6, 11 and 17 hours, respectively. In the domestic gilts, the duration
was, 9, 8, 12 and 11 hours, respectively. The differences in the duration of nocturnal increase in
melatonin between the summer and winter samplings were significant (P < 0.01) in both animal
groups (Figure 3).
Figure 2. Individual melatonin patterns of the wild boars and domestic gilts from the summer sampling.
Wild boars
0
10
20
30
40
13 17 21 1 5 9 13 17 21 1 5 9
Time (00 hrs)
pg/m
l
Domestic gilts
0
10
20
30
40
13 17 21 1 5 9 13 17 21 1 5 9
Time (00 hrs)
pg/m
l

45
Figure. 3. Mean (-sem) serum melatonin concentrations (�) for the wild boars and domestic gilts and light intensity ({) over 48 hours. The season and animal group (wild boar/domestic pig) are indicated in the figure.
4.2. EFFECTS OF ARTIFICIAL LIGHTING PROGRAMS ON MELATONIN SECRETION
(II, III)
There were no significant effects of lighting regimen (16 L : 8 D vs. 8 L : 16 D) on scotophase mid-
interval mean melatonin concentrations (II). When the lighting regimen was changed from long day
to short day, there was an immediate change in the pattern of melatonin profile toward alignment
with the new lighting conditions. When the lighting regimen was changed from short day to long
day there was only a small change in the melatonin profile (Figure 4). At 1800, 2000 and 0600
hours when lights were on mean melatonin concentration continued to significantly exceed the
basal level (P<0.001, 0.001, 0.01 respectively) but at 0800 hours the mean was at basal level. After
one week there was further progression towards a long day melatonin pattern. After two weeks,
melatonin profiles were adjusted to a new light environment excepting of some variation in the first
and the last sample of the scotophase.
Dom es tic (S um m er)
05
1 01 52 02 53 03 5
1 3 1 7 2 1 1 5 9 1 3 1 7 2 1 1 5 9
Tim e (00 hrs )
pg
/ml
0
1 0 0
2 0 0
3 0 0
4 0 0
lux
W ild B oar (S um m er)
05
1 01 52 02 53 03 5
1 3 1 7 2 1 1 5 9 1 3 1 7 2 1 1 5 9
Tim e (00 hrs )
pg/m
l
0
1 00 00
2 00 00
3 00 00
4 00 00
lux
W ild boar (W inter)
05
1 01 52 02 53 03 5
1 9 2 3 3 7 1 1 1 5 1 9 2 3 3 7 1 1 1 5
Tim e (00 hrs )
pg/m
l
0
1 0 0 0
2 0 0 0
3 0 0 0
4 0 0 0
lux
Domestic (Winter)
05
101520253035
19 23 3 7 11 15 19 23 3 7 11 15Time (00 hrs)
pg/m
l
0
100
200
300
400
lux

46
Figure 4. Mean (- SEM) plasma melatonin concentrations of young boars. Pigs were allowed to adjust to the lighting (16 L : 8 D) for two weeks before the lighting was changed to the opposite (8 L : 16 D). A 48-hour sampling spanning the changeover with samples collected at two-hour intervals was performed at the time the lighting was changed. Black bar under axis illustrates the scotophase.
When the light-dark cycle was 12 L : 12 D, the photophase light intensity (40 lx, 200 lx or 10000
lx) did not affect the scotophase mean melatonin concentration (III). There was no difference in the
basal melatonin concentrations between the lighting regimens within or between the groups (Figure
5).
Figure 5. The mean (+SD) melatonin concentration during the photophase at all treatments (basal concentration) and the mean (+SD) melatonin concentrations during the scotophase at different treatments. The treatments are indicated under the x-axis.
Long day->Short day
0 2 4 6 8
10 12 14 16
12 16 20 24 4 8 12 16 20 24 4 8 12 Time
pg/m
l
Short day->Long day
0 2 4 6 8
10 12 14 16
12 16 20 24 4 8 12 16 20 24 4 8 12
Time
pg/m
l
0
2
4
6
8
10
12
14
Basal 40 lx 200 lx 10000 lx
pg/m
l
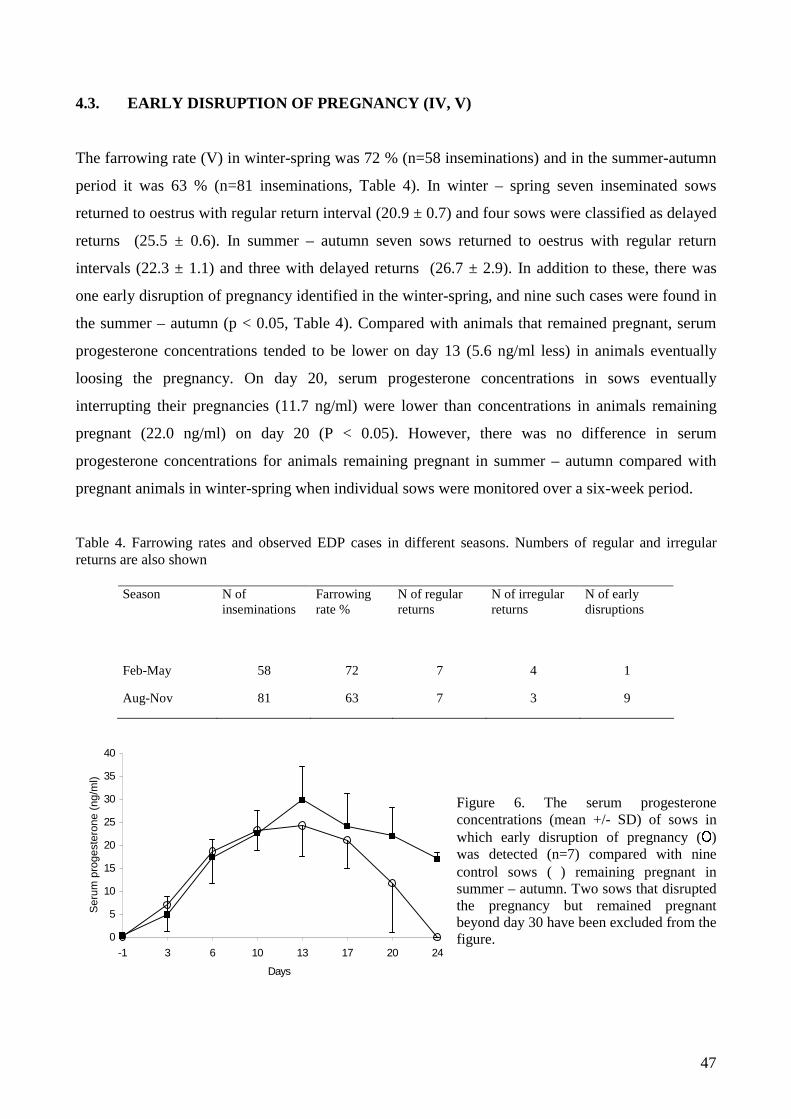
47
0
5
10
15
20
25
30
35
40
-1 3 6 10 13 17 20 24
Days
Ser
um p
roge
ster
one
(ng/
ml)
4.3. EARLY DISRUPTION OF PREGNANCY (IV, V)
The farrowing rate (V) in winter-spring was 72 % (n=58 inseminations) and in the summer-autumn
period it was 63 % (n=81 inseminations, Table 4). In winter – spring seven inseminated sows
returned to oestrus with regular return interval (20.9 ± 0.7) and four sows were classified as delayed
returns (25.5 ± 0.6). In summer – autumn seven sows returned to oestrus with regular return
intervals (22.3 ± 1.1) and three with delayed returns (26.7 ± 2.9). In addition to these, there was
one early disruption of pregnancy identified in the winter-spring, and nine such cases were found in
the summer – autumn (p < 0.05, Table 4). Compared with animals that remained pregnant, serum
progesterone concentrations tended to be lower on day 13 (5.6 ng/ml less) in animals eventually
loosing the pregnancy. On day 20, serum progesterone concentrations in sows eventually
interrupting their pregnancies (11.7 ng/ml) were lower than concentrations in animals remaining
pregnant (22.0 ng/ml) on day 20 (P < 0.05). However, there was no difference in serum
progesterone concentrations for animals remaining pregnant in summer – autumn compared with
pregnant animals in winter-spring when individual sows were monitored over a six-week period.
Table 4. Farrowing rates and observed EDP cases in different seasons. Numbers of regular and irregular returns are also shown
Figure 6. The serum progesterone concentrations (mean +/- SD) of sows in which early disruption of pregnancy (³) was detected (n=7) compared with nine control sows ( ) remaining pregnant in summer – autumn. Two sows that disrupted the pregnancy but remained pregnant beyond day 30 have been excluded from the figure.
Season N of inseminations
Farrowing rate %
N of regular returns
N of irregular returns
N of early disruptions
Feb-May 58 72 7 4 1
Aug-Nov 81 63 7 3 9

48
In the active GnRH-immunisation trial (IV), none of the 7 sows receiving the booster immunisation
10 days after mating were pregnant as detected by real time ultrasound 18 days post mating. There
was no visible evidence of abortion at any time. The effect of treatment on progesterone
concentrations was significant (Figure 7A; P=0.05). Plasma progesterone levels in immunised sows
started to decrease compared with pregnant controls after day 16, and by day 18 progesterone levels
were similar in immunised and non-pregnant sows. All sows that received the booster immunisation
on day 20 of pregnancy (n=4) aborted between 7 and 14 days later with a mean immunisation to
abortion interval of 10 ± 1.5 (SEM) days (Figure 7B). In these sows progesterone levels decreased
gradually until day 26 followed by a steep decrease between days 26 and 28. After progesterone had
reached a basal concentration, it took approximately two days before abortion occurred.
In the passive GnRH-immunisation trial, none of the gilts infused with antiserum (n=6) 12 days
post mating was pregnant 18 days after mating, whereas 5 out of 6 controls were pregnant on day
18 and remained pregnant until the end of the trial. The non-pregnant gilts on day 18 typically had a
small amount of fluid in the uterus with distribution of fluid distinguishably different from a typical
uterine oestrous fluid. The fluid was in a small isolated area rather than being distributed throughout
the uterus and between the endometrial folds. Despite daily close monitoring of animals there was
no visible evidence of abortion.
0
5
10
15
20
25
10 12 14 16 18 20 22 24 26 28
days after mating
plas
ma
prog
este
rone
ng/
ml
vaccinated preg contr non-preg contr
0
5
10
15
20
25
20 22 24 26 28 30 32 34 36 38
days after mating
plas
ma
prog
este
rone
ng/
ml
vaccinated preg contr
average abortion time
Figure 7. The mean (±SEM) plasma progesterone concentrations after active immunisation by a commercial GnRH-vaccine (Improvac®). The first immunisation was done on the day of farrowing and the second immunisation was done on day 10 (A) (n=7) or on day 20 (B) (n=4) post mating. The average abortion time after active immunisation on day 20 post mating is also shown (B).
A) B)

49
5. DISCUSSION
This study demonstrated that pigs secrete melatonin in rhythmic fashion similar to melatonin
secretion in other mammals studied. There is no difference in lighting – melatonin transduction
between wild boars and domestic pigs. Both the wild boar and the domestic pig express seasonal
variation in the circadian secretion pattern of melatonin provided that the artificial lighting in the
piggery is constant and the natural light is allowed through the windows. The seasonal variation in
melatonin secretion is directly related to the changes in the day length. It is justified to assume,
according to studies with other mammals, that also wild boars synchronize their breeding season
with a preferred season using the changing melatonin profile to anticipate the right time of the year.
It seems more than a coincidence that an increased number of pregnancy failures in the domestic
pig appears at the time when the European wild boar is in a sexual arrest (Peltoniemi et a., 1999). In
addition, when it is evident now that seasonal alterations in the melatonin secretion of these two
types of pigs are similar, it is obvious that changes in the day length contribute to the seasonal
infertility of the domestic pig.
These results provide further evidence that earlier confusion about the existence of a circadian
melatonin rhythm in the domestic pig related to inadequacies in the RIAs used, as demonstrated by
Klupiec et al. (1997) and confirmed later by Andersson et al (2000). The present study, together
with some earlier studies (Paterson et al., 1992), has conclusively demonstrated a circadian
melatonin rhythm in pigs under different lighting conditions, and further studies should be directed
towards the applicability of artificial lighting programs for commercial piggeries.
The circadian melatonin secretion can be entrained by artificial lighting. Pigs are able to respond
within relatively short acclimatization periods to abrupt and extreme changes in ambient lighting.
Indeed, pigs are able to respond immediately to an extension of the dark phase as demonstrated by
entrained melatonin profiles after the change from the eight-hour dark phase to the 16-hour dark
phase. It seemed to be more difficult for pigs to adjust to a change in the other direction (from short
day to long day). This was demonstrated by almost unchanged melatonin profiles during the first 24
hours after the changeover. However, in this group melatonin profiles analysed one week after the
changeover showed further entrainment to the new lighting regimen and the profiles were
practically adjusted within two weeks. The melatonin profiles were evident under all lighting
regimens and also during the first 24-hours after every changeover, even if they were not always

50
immediately adjusted to the ambient lighting. There was no evidence that extreme and abrupt
changes in lighting could abolish the circadian rhythm of melatonin secretion as proposed by
McConnell and Ellendorff (1987). It is evident that the response in circadian rhythms can be
achieved using simplified lighting regimens instead of step-wise changes in lighting (Paterson et al.,
1992). This has importance for application of artificial lighting programs to commercial piggeries.
Another important aspect of lighting regimens in commercial piggeries is the light intensity during
the photophase and its effects on melatonin secretion during the darkphase. Earlier studies implied
that pigs need relatively high light intensities during the photophase to entrain their melatonin
rhythms (Minton et al., 1989; Mack and Unshelm, 1997). It is important to use sufficient light
intensity during the phoptophase so that pigs can differentiate between the light- and darkphases. It
was demonstrated in the present study that the light intensity needed for differentiation between
photo- and scotophase is much lower in pigs than previously thought. As low intensity as 40 lx is
enough to entrain melatonin secretion so that high melatonin concentrations occur in the systemic
circulation during the darkphase. It appears that there is no difference in the darkphase melatonin
response when the lightphase light intensity varies between 40 and 10 000 lx. This suggests that
extremely high light intensities (10 000 lx) during the photophase do not provide any advantage
over a normal comfortable light intensity (200 lx) in attempts to manipulate reproduction by
artificial lighting regimens. It can be concluded that manipulation of circadian rhythms is possible
by simple artificial lighting programs and that most attention should be paid to the light-dark cycle
rather than the absolute light intensity, which is of little importance.
There are different manifestations of seasonal infertility; delayed puberty in gilts, prolonged
weaning to oestrus interval in sows and a reduction in the proportion of mated sows that farrow.
The last of these is of the greatest economic significance (Love et al. 1993). A major contribution to
the reduced farrowing rate is the increased number of irregular returns to oestrus 25 to 35 days after
mating (Love 1981). Early disruption of pregnancy, because of early embryonic deaths (Xue et al.
1994) or because of regression of corpora lutea (CL) (Wrathall et al. 1986), has been suggested to
cause these irregular returns in summer-autumn. The delayed return to oestrus, although commonly
reported in the literature and known as a manifestation of seasonal infertility, has never before been
shown to result from the disruption of an established pregnancy. This study demonstrated for the
first time a clear association between the seasonally lowered farrowing rate and early disruption of
pregnancies. Wrathall et al. (1986) have demonstrated a clear seasonal variation in the serum
progesterone levels that contradicts with the present study. However, they compared progesterone

51
levels at a later stage of pregnancy (between days 40 and 90), whereas in the present study the
comparison was at the early stage that is a more relevant time for studying this phenomenon. They
did compare progesterone also on days 25 – 30 in one single farm (250 sows), but the comparison
was made between single samples of animals rather than longer profiles as was done in the present
study. Interestingly, in the present study progesterone concentrations in animals eventually loosing
the pregnancy were somewhat lower as early as day 13 after mating. This implies that
endocrinological events leading to disruption of pregnancy in summer-autumn are demonstrable
only in “problem sows” and not in the general population. The endocrinological mechanisms of
early disruption of pregnancy are yet to be confirmed, but this study implies that something happens
to the normal development of embryos first and after that the CL regress.
A number of earlier studies have shown that pituitary LH-secretion is subject to seasonal variation
in the pig, even if there is some controversy about seasonal effects on LH-pulsatility. However,
most studies have found that LH-pulses are more regular and of higher amplitude in the winter (or
under short day lighting) when compared with summer (or long day lighting) (Peacock, 1991;
Paterson, 1992; Peltoniemi, 1997b). The LH-support for CL is lower during the time LH-pulses are
irregular and of lower amplitude, i.e. during the summer – autumn. It has been proposed that this
lowered pituitary support might cause regression of CL, when they are no longer autonomous after
day 12 of pregnancy, and this leads to early disruption of pregnancy typical for seasonal infertility
(Wrathall et al., 1986; Love et al., 1993). However, the studies by Anderson et al. (1967) and
Peltoniemi et al. (1995) have demonstrated that a prolonged complete blockade of LH-pulses is
needed to initiate regression of CL. Those studies together with the present work demonstrate that
from four to nine days’ complete LH-deprivation is needed to cause a destruction of CL. It is
unlikely that there could be such a complete seasonal LH-blockade.
The study also showed two different outcomes of active GnRH-immunization depending on the
timing of the second immunization. None of the sows immunized for the second time on day 10
post mating was detectably pregnant 18 days after mating, and there were no outer signs of
abortion. The passive immunization of gilts on day 12 post mating had a similar effect. The second
immunization on pregnancy day 20 resulted in abortion with normal fetuses expelled (Knight et al.,
1977). The mean interval between the second immunization and abortion was 10 ± 1.5 days. In both
actively immunized groups the progesterone decreased to the basal level eight days after the second
immunization. This means that in the 10-day immunization group the recognition and establishment
of pregnancy had already failed before the regression of CL, whereas in the 20-day vaccination

52
group the regression of CL caused the abortion. This is similar to the observation in the field trial,
where the normal development of embryos was first disturbed and the regression of CL was
initiated after that. This study implies that early disruption of pregnancy is caused by a mechanism
other than by a direct regression of CL because of inadequate pituitary LH-support.
The progesterone concentrations after pregnancy day 12 depend on pituitary support whereas before
day 12 the number of formed CL and a metabolic clearance of progesterone mainly affect the
progesterone concentrations. It is well known that progesterone controls a number of events
affecting embryos’ development rate during early pregnancy that is critical for signaling
commenced pregnancy and preventing the luteolysis (Geisert and Yelich, 1997). This study implies
that decreased progesterone concentration between days 14 and 18 post mating can disturb
development of embryos and prevent establishment of pregnancy before regression of CL occurs
(because of deprivation of LH-pulses). The recognition of pregnancy takes place in two phases
between days 12 and 18 as demonstrated by Pusateri et al. (1996a). This means that the first
oestrogen signal from the embryos occurs during the period of inadequate pituitary LH-support
(summer) because the pituitary LH does not affect progesterone and subsequently the development
rate of embryos before day 12. However, the seasonally decreased pituitary LH-support can affect
progesterone levels after day 12 and could interfere with the development of embryos, which in turn
could result in an inadequate or missing oestrogen signal on pregnancy day 18. If that was the case,
as suggested in the present study, it would lead to reorientation of PGF-2α secretion back to an
endocrine direction, luteolysis of CL, early disruption of pregnancy and a delayed return to oestrus
with interval of 25-30 days. A schematic figure of the proposed mechanism of early disruption of
pregnancy is presented in Figure 6.
It is concluded that the basis of lighting - melatonin transduction in the pig is similar to other
mammals. There is no difference with the melatonin secretion in the European wild boar and the
domestic pig. The circadian pattern of melatonin secretion can be modified by simple lighting
regimens. A light-dark cycle itself appears to be more important in producing a scotophase
melatonin response than the light intensity during the photophase. The lowered farrowing rate
typical for seasonal infertility is associated with early disruptions of pregnancy. It is possible that
the seasonal early disruption of pregnancy is mediated by decreased progesterone concentrations
leading to underdeveloped embryos at the time of recognition of pregnancy. This seasonal decrease
in progesterone appears to be under the influence of seasonally changing melatonin secretion

53
through the GnRH- and LH-secretion. Further studies are required to ensure the mechanism of
seasonal early disruption of pregnancy.

54
LONG DAY SCN SHORT DAY PINEAL GLAND
Nocturnal melatonin secretion
HYPOTHALAMUS
Effects on GnRH-neurons (sensitivity to oestradiol feed-back) Direct effects of neurotransmitters
GnRH-secretion
PITUITARY GLAND
LH-pulses CORPORA LUTEA ESTABLISHED
Progesterone secretion PREGNANCY beyond pregnancy day 12
UTERUS Secretion of histotrophes
UNDER-DEVELOPED OR DEAD NORMAL EMBRYOS EMBRYOS ON PREGNANCY DAY 18 ON PREGNANCY DAY 18
Figure 6. The schematic figure of the proposed mechanism of seasonal early disruption of pregnancy.
shor
t dur
atio
n
long
dur
atio
n
cate
chol
amin
e en
dog.
opi
oids
glut
amat
ergi
c ne
uron
s
decr
eas
incr
eas
low amplitude irregular
high amplitude regular
decr
ease
d
nor
mal
Oestrogen signal on day 18
PGF-2α secretion back to endocrine direction beyond day 18
Continuation of PGF-2α secretion to exocrine direction beyond day 18
decreased normal
EARLY DISRUPTION OF PREGNANCY

55
6. CONCLUSIONS
1. The basis of lighting – melatonin transduction in the pig is similar to other mammals.
2. There is no attenuation of melatonin secretion in the domestic pig due to altered genotype or
reduced light environment.
3. Simplified artificial lighting programs can modify the circadian pattern of melatonin
secretion. Adjustment of the secretion pattern takes place within two weeks even after
abrupt and extreme changes in lighting.
4. The photophase light intensity has no effect on the scotophase melatonin response, once the
threshold light intensity to differentiate the dark and light phases is exceeded. This threshold
is less than 40 lx.
5. The seasonally lowered farrowing rate is associated with an increased number of early
disruptions of pregnancies. There is/are factor(s) interfering first with the normal
development of embryos and the regression of CL initiates after that.
6. This retarded development of embryos might be caused by seasonally altered LH-secretion
decreasing the progesterone secretion of CL affecting the secretion of histotrophes of
endometrium.

56
ACKNOWLEDGEMENTS
The project has been financially supported by Finnish Ministry of Agriculture, Finnish
slaughterhouses and University of Helsinki, which are gratefully acknowledged.
I had no plans for PhD project when I came to ambulatory clinic of Saari to work as a clinic
veterinarian in 1997. However, Olli Peltoniemi, PhD, had a great vision of importance of
seasonality studies for Finnish pig production. Olli talked me into the project and ever since he has
been a principal supervisor of my PhD project and the closest fellow worker. His enthusiasm and
support has been irreplaceable, for which I will always be grateful.
I have got a lot of support and friendly guidance also from Professor Terttu Katila, the leader of the
PhD project, for which I will be indebted. Professor Robert Love was my supervisor when I spent
an unforgettable research year in Australia at University of Sydney. Ever since, Professor Love has
had a great contribution to my PhD project, of which I want to express my warmest thanks. I would
like to extend my gratitude to Professor Gareth Evans and Professor David Kennaway for their help
and cooperation. Professor Hannu Saloniemi and Professor Magnus Andersson have been my
supervisors giving me an incredible amount of encouragement during the course of years. They
both are gratefully acknowledged.
Håkan Andersson, PhD, my co-author, has always been ready for cooperation and stimulant
scientific discussions, for which I am grateful. My co-author, Outi Hälli, DVM was a great help
especially in the most demanding wild boar studies, for which I want to thank her. During my
project, Professor Satu Pyörälä, Erkki Pyörälä, DVM and Juhani Taponen, DVM, have all shown
their support to my PhD project in a number of ways for which I will always be indebted. I have
had a privilege to work with swine specialists like Mari Heinonen, PhD, Juha Virolainen, DVM,
Jonna Oravainen, DVM, Veikko Tuovinen, PhD, Taneli Tirkkonen, DVM, Tapio Laurila, DVM
and a number of others. They all are acknowledged with gratitude. I want to extend my gratitude to
all the staff of Saari Unit. I am particularly grateful to Reijo Jokivuori for his kindness.
I am also indebted to the pre-examiners of the dissertation, Professor Stig Einarsson and Professor
Erik Ropstad for their constructive criticism at the final stage of the present work. I also would like
to acknowledge the opponent, Professor Gerald Lincoln.

57
Finally, I would like to thank my beloved wife Kaija for her support and love. I am grateful for our
two sons, Julius and our beautiful baby. I also want to thank my parents and my brothers, who have
always supported me with love.
Anssi Tast

58
REFERENCES
Aherne, F.X. and Kirkwood, R. Nutrition and sow prolificacy. J. Reprod. Fertil. Suppl. 33, 1985:
169-183.
Almond, G.W., Friendship, R.M. and Bosu, W.T.K. Autumn abortions in sows. Can.Vet. J. 26,
1985: 162-163.
Andersson, H., Lillpers, K., Rydhmer, L. and Forsberg, M. Influence of light environment and
photoperiod on plasma melatonin and cortisol profiles in young domestic boars, comparing two
commercial assays. Domest. Anim. Enocrinol. 19, 2000: 261-274.
Anderson, L.L., Dyck, G.W., Mori, H., Henricks, D.M. and Melampy, R.M. Ovarian function in
pigs following hypophysial stalk transection or hypohysectomy. Am. J. Physiol. 1967: 1188-1194.
Anderson, L.L. Growth, protein content and distribution of early pig embryos. Anat. Rec. 190,
1978: 143-153.
Aoki, H., Yamada, N., Ozeki, Y., Yamane, H. and Kato, N. Minimum light intensity required to
suppress nocturnal melatonin concentration in human saliva. Neurosci. Lett. 252, 1998: 91-94.
Arendt, J., Symons, A.M. and Laud, C. Pineal function and photoperiod in the ewe. In:
Photoperiodism and Reproduction in Vertebrates. Colloq INRA 6, 1981: 219-229.
Bazer, F.W. and Thatcher, W.W. Theory of maternal recognition of pregnancy in swine based on
estrogen controlled endocrine versus exocrine secretion of prostaglandin F2alpha by the uterine
endometrium. Prostaglandins 14, 1977: 397-400.
Bazer, F.W., Geisert, R.D., Thatcher, W.W. and Roberts, R.M. The establishment and maintenance
of pregnancy. In: D.J.A. Cole and C.R. Foxcroft (eds.) Control of Pig Reproduction. Butterworths,
London, 1982: 227-252.
Boivin, D.B., Duffy, J.F., Kronauer, R.E. and Czeisler, C.A. Dose-response relationships for
resetting of human cicardian clock by light. Nature 379, 1996: 540-544.

59
Bollinger, A.L., Wilson, M.E., Pusateri, A.E., Green, M.L., Martin, T.G. and Diekman, M.A. Lack
of a nocturnal rise in serum concentrations of melatonin as gilts attain puberty. J. Anim. Sci. 75,
1997: 1885-1892.
Bonnefond, C. Walker, A.P., Stutz, J.A., Maywood, E., Juss, T.S., Herbert, J. and Hastings, M.H.
The hypothalamus and photoperiodic control of FSH secretion by melatonin in the male Syrian
hamster. J. Endocrinol. 122, 1989: 247-254.
Booth, P.J. Metabolic influences on hypothalamic-pituitary-ovarian function in the pig. J. Reprod.
Fertil. Suppl. 40, 1990: 89-100.
Brinkley, H.J., Norton, H.W. and Nalbandov, A.V. Role of a hypophysial luteotrophic substance in
the function of porcine corpora lutea. Endocrinology 74, 1964: 9-13.
Carson, A.F., Mc Caughey, W.J., Mackie, D.P., Higgins, H.C. and Steen, R.W.J. Production and
biological activity of Murine monoclonal antibodies against GnRH. Theriogenology 48, 1997:193-
207.
Carter, D.S. and Goldman, B.D. Antigonadal effects of timed melatonin ifusion in pinealectomized
male Djungarian hamsters (Phodopus sungorus sungorus): Duration is the critical parameters.
Endocrinology, 1983.
Cross, J.S. and Roberts, R.M. Porcine conceptuses secrete an interferon during the preattachment
period of early pregnancy. Biol. Reprod. 40, 1989: 1109-1118.
Dehoff, M.H., Bazer, F.W. and Collier, R.J. Ontogeny of prolactin receptors in porcine uterine
endometrium during pregnancy. In Proceedings: 4th International prolactin congress,
Charlottesville. 1984: 95 (abstr.).
Delcroix, I., Mauget, R. and Signoret, J.P. Existence of synchronization of reproduction at the level
of the social group of the European wild boar (Sus scrofa). J. Reprod. Fert. 89, 1990: 613-617.
Diekman, M.A., Clapper, J.A., Green, M.L. and Stouffer, D.K. Reduction in age of puberty in gilts
consuming melatonin during decreasing or increasing daylength. J. Anim. Sci. 69, 1991: 2524-
2531.
Diekman, M.A., Brandt, K.E., Green, M.L., Clapper, J.A. and Malayer, J.R. Lack of a nocturnal rise
of serum melatonin in prepubertal gilts. Domest. Anim. Endocrinol. 2, 1992: 161-167.

60
Diekman, M.A. and Green, M.L. Serum concentrations of melatonin in prepubertal or postpubertal
gilts exposed to artificial lighting or sunlight. Theriogenology, 47, 1997: 923-928.
Dziuk, P.J. Effect of migration, distribution and spacing of pig embryos on pregnancy and fetal
survival. J. Reprod. Fert. Suppl. 33, 1985: 57-63.
Ebling, F.J., Lincoln, G.A., Martin, G.B. and Taylor, P.L. LHRH and beta-endorphin in the
hypothalamus of the ram in relation to photoperiod and reproductive activity. Dom. Anim.
Endocrinol. 4, 1987: 149-156.
Ebling, F.J. and Hastings, M.H. The neural basis of seasonal reproduction. Ann Zootech. 41, 1992:
239-246.
Enne, G., Beccaro, P.V. and Tarocco, C. A note on the effect of climate on fertility in pigs in the
Padana Valley of Italy. Animal Production. 28, 1979: 115-117.
Fischer, H.E., Bazer, F.W. and Fields, M.J. Wsteroid metabolism by endometrial and conceptus
tissues during early pregnancy and pseudopregnancy in gilts. J. Reprod. Fert. 75, 1985: 69-78.
Foxcroft, G.R. Mechanisms mediating nutritional effects on embryonic survival in pigs. J. Reprod.
Fert. Suppl. 52, 1997: 47-61.
Frank, M., Bazer, F.W., Thatcher, W.W. and Wilcox, C.J. A study of prostaglandin F2alpha as the
luteolysin in swine: III effects of estradiol valerate on prostagalndin F, progestins, estrone and
estradiol concentrations in the utero-ovarian vein of nonpregnant gilts. Prostaglandins 14, 1977:
1183-96.
Gadsby, J.E., Heap, R.B. and Burton, R.D. Oestrogen production by blastocyst and early embryonic
tissue of various species. J. Reprod. Fert. 60, 1980: 409-17.
Gadsby, J.E., Fitz, T.A, Balapure, A.K. and Britt, J.H. PGF-2α receptors on pig luteal cells during
the oestrus cycle. Biol. Reprod. 38 Suppl.1, 1988: 181 (abstr.).
Garverick, H.A., Polge, C. and Flint, A.P. Oestradiol administration raises luteal LH receptor levels
in intact and hysterectomized pigs. J. Reprod. Fert. 66, 1982: 371-7.

61
Geisert, R.D., Pratt, T.N., Bazer, F.W., Mayes, J.S. and Watson, G.H. Immunocytochemical
localization and changes in endometrial progestin receptor protein during the porcine oestrus cycle
and early pregnancy. Reprod. Fertil. Dev. 6, 1994: 749-760.
Geisert, R.D. and Yelich, J.V. Regulation of conceptus development and attachment in pigs. J.
Reprod. Fert. Suppl. 52, 1997: 133-149.
Green, M.L., Clapper, J.A., Andres, C.J. and Diekman, M.A. Serum concentrations of melatonin in
prepubertal gilts exposed to either constant or stepwise weekly alteration in scotophase. Domest.
Anim. Endocrinol. 13, 1996.
Greenwood, F.C., Hunter, W.M. and Glover, J.S. The preparation of 131-I labeled human growth
hormone and high specific radioactivity. Biochemical Journal 89, 1963: 114-123.
Griffith, M.K. and Minton, J.E. Effect of light intensity on circadian profiles of melatonin, ACTH,
and cortisol in pigs. J. Anim. Sci. 70, 1992: 492-498.
Gross, T.S., Mirando, M.A., Young, K.H., Beers, S., Bazer, F.W. and Thatcher, W.W.
Reorientation of prostaglandin F secretion by calcium ionophore, estradiol and prolactin in
perifused porcine endometrium. Endocrinology 127, 1990: 637-42.
Guthrie, H.D. and Rexroad, C.E.Jr. Endometrial prostaglandin F release in vitro and plasma 13, 14-
dihydro-15-keto-prostaglandin F2alpha in pigs with luteolysis blocked by pregnancy, oestradiol
benzoate or human chorionic gonadotropin. J. Anim. Sci. 52, 1981: 330-9.
Hallford, D.M., Wettemann, R.P., Turman, E.J. and Omtvedt, I.T. Luteal function in gilts after
prostaglandin F2alpha. J. Anim. Sci. 41, 1975: 1706-10.
Hancock, R.D. Clinical observations on seasonal infertility in sows in Cornwall. Vet. Rec. 123,
1988: 413-416.
Hashimoto, S., Nakamura, K., Honma, S., Tokura, H. and Honma, K. Melatonin rhythm is not
shifted by lights that suppress nocturnal melatonin in humans under entrainment. Am. J. Physiol.
270, 1996: 1073-1077.
Heap, R.P. and Perry, J.S. The maternal recognition of pregnancy. Br. J. Hosp. Med. 12, 1974: 8-
14.

62
Henderson, K.M. and McNatty, K.P. A biochemical hypothesis to explain the mechanism of luteal
regression. Prostaglandins 9, 1975: 779-97.
Hughes, P.E. Factors affecting the natural attainment of puberty in the gilt. In: D.J.A. Cole and C.R.
Foxcroft (eds.) Control of Pig Reproduction. Butterworths, London. 1982: 117-138.
Hurtgen, J.P. and Leman, A.D. Seasonal influence on the fertility of sows and gilts. J. Am Vet.
Med. Ass. 177, 1980: 631-635.
Karch, F.J., Bittman, E.L., Foster, D.L., Goodman, R.L., Legan, S.J. and Robinson, J.E.
Neuroendocrine Basis of seasonal Reproduction. Recent Prog. Horm. 40, 1984: 185-232.
Karsch, F.J., Malpaux, B., Wayne, N.L. And Robinson, J.E. Characteristics of the melatonin signal
that provide the photoperiodic code for timing seasonal reproduction in the ewe. Repr. Nutr. Dev.
28,1988: 459-472.
Kennaway, D.J., Gilmore, T.A. and Seamark, R.F. Effect of melatonin feeding on serum prolactin
and gonadotropin levels and the onset of seasonal oestrus cyclicity in the sheep. Endocrinology 110,
1982: 1766-1772.
Klupiec, C., Evans, G., Love, R.J. and Kennaway, D.J. Clarifying plasma melatonin profiles in
domestic pigs: A critical and comparative evaluation of two radioimmunoassay systems. J. Pineal
Res. 22, 1997: 65-74.
Knight, J.W., Bazer, F.W., Thatcher, W.W., Franke, D.E. and Wallace, H.D. Conceptus
development in intact and unilaterally hysterectomized – ovariectomized gilts: interrelations among
hormonal status, placental development, fetal fluids and fetal growth. J. Anim. Sci., 44, 1977: 620-
637.
Krzymowski, T., Kotwicka, J., Okrasa, S. and Doboszynska, T. The function and regression of
corpora lutea during the sow’s oestrus cycle after 10 hours of prostaglandin F-2α infusion into the
anterior uterine vein. In Proceedings: VIII Int. Congr. Anim. Reprod. A. I., Krakow. 1976: 143.
La Bonnardière, C. Nature and possible functions of interferons secreted by the preimplantation pig
blastocyst. J. Reprod. Fert. Suppl. 48, 1993:157-170.

63
Lehman, M.N., Robinson, J.E., Karch, F.J. and Silverman, A.J. Immunocytochemical localization
of luteinizing hormone-releasing hormone (LHRH) pathways in the sheep brain during anestrus and
the mid-luteal phase of the estrous cycle. J. Comp. Neurol. 244(1), 1986: 19-35.
Lewy, A.J., Wehr, T.A. and Goodwin, F.K. Light suppresses melatonin secretion in humans.
Science. 210, 1980:1267-1269.
Lincoln, G.A. Photoperiod-pineal-hypothalamic relay in sheep. Anim. Reprod. Sci. 28, 1992: 203-
217.
Love, R.J. Definition of a seasonal infertility problem in pigs. Vet. Rec.104, 1978: 238-240.
Love, R.J. Seasonal infertility in pigs. Vet.Rec. 109,1981: 407-409.
Love, R.J., Evans, G. and Klupiec, C. Seasonal effects on fertility in gilts and sows. J. Reprod.
Fertil. Suppl. 48, 1993: 191-206.
Love, R.J., Klupiec, C., Thornton, E.J. and Evans, G. An interaction between feeding rate and
season affects fertility of sows. Anim. Reprod. Sci. 39, 1995: 275-284.
Lucia, T., Dial, G.D., Marsh, W.E. and P.R. Davies. Long-term breeding patterns in commercial
swine farms. In Proceedings: 13th International Pig Veterinary Society Congress, Bangkok,
Thailand. 1994.
Mack, A.C. and Unshelm, J. Cicardian rhythm of the light-dependant hormone melatonin as an
indicator for the photicrequirement of domestic pigs. In Proceedings: 9th International Congress in
Animal Hygiene, Helsinki, Finland. 1997: 216-220.
Mattioli, M., Prandi, A., Camporesi, A., Simoni, A. and Seren, E. Investigations on swine summer
infertility in Italy. In: E. Seren and M. Mattioli (eds.) Definition of the Summer Infertility Problem.
Commission of the European Communities, Brussels. 1987: 33-43.
Mauget, R. Reproductive biology of the European wild boar. In: E. Seren and M. Mattioli (eds.)
Definition of the Summer Infertility Problem. Commission of the European Communities, Brussels.
1987: 107-119.
McConnell, S.J. and Ellendorff, F. Absence of nocturnal plasma melatonin surge under long and
short artificial photoperiods in the domestic sow. J. Pineal. Res. 4, 1987: 201-210.

64
Melampy, R.M. and Anderson, L.L. Role of the uterus in corpus luteum function. J. Anim. Sci.
Suppl. 27, 1968: 77-96.
Meulen, J., Helmond, F.A., Oudenaarden, C.P.J. and Van der Meulen, J. Effect of flushing of
blastocysts on days 10-13 on the life-span of the corpora lutea in the pig. J. Reprod. Fert. 84, 1988:
157-162.
Minton, J.E., Davis, D.L. and Stevenson, J.S. Contribution of the photoperiod to cicardian
variations in serum cortisol and melatonin in boars. Domest. Anim.Endocrinol. 6, 1989: 177-181.
Moeljono, M.P., Thatcher, W.W., Bazer, F.W., Frank, M., Owens, L.J. and Wilcox, C.J. A study of
prostaglandin F2alpha as the luteolysin in swine: Characterization and comparison of prostaglandin
F, oestrogens and progestin concentrations in utero-ovarian vein plasma of nonpregnant and
pregnant gilts. Prostaglandins 14, 1977: 543-555.
Niswender, G.D., Reichert, L.E. and Zimmerman, D.R. Radioimmunoassay of serum levels of
luteinising hormone throughout the estrus cycle in pigs. Endocrinology 87, 1970: 576-580.
Paterson, A.M. and Pearce, G.P. Attainment of puberty in domestic gilts reared under long-day or
short-day artificial light regimens. Anim. Reprod. Sci. 23, 1990: 135-144.
Paterson , A.M., Pearce, G.P. and Antuono, M.F.D. Seasonal variation in attainment of puberty in
isolated and boar-exposed domestic gilts. Anim. Reprod. Sci. 24, 1991: 323-333.
Paterson, A.M., Martin, G.B., Foldes, A., Maxwell, C.A. and Pearce, G.P. Concentrations of plasma
melatonin and luteinizing hormone in domestic gilts reared under artificial long or short days. J.
Reprod. Fertil. 94, 1992: 85-95.
Peacock, A.J. Environmental and social factors affecting seasonal infertility of pigs. PhD-thesis.
University of Sydney, 1991.
Pearce, G.P. and Pearce A.N. Contact with sow in oestrus or a mature boar stimulates the onset of
oestrus in weaned sows. Vet. Rec. 130, 1992: 5-9.
Peltoniemi, O.A.T. Seasonal and Nutritional Effects on Gonadotrophins in the Early Pregnant Gilts.
MVet.Sc-thesis. University of Sydney, 1994.

65
Peltoniemi, O.A.T., Easton, B.G., Love, R.J., Klupiec, C. and Evans, G. Effect of chronic treatment
with a GnRH agonist (Goserelin) on LH secretion and early pregnancy in gilts. Anim. Reprod. Sci.
40, 1995: 121-133.
Peltoniemi, O.A.T., Heinonen, M., Tuovinen, V., Saloniemi, H. and Love, R.J. Characteristics of
seasonal infertility of the sow in Finland. In Proceedings: 9th International Congress in Animal
Hygiene, Helsinki, Finland. 1997: 208-211.(a)
Peltoniemi, O.A.T., Love, R.J., Klupiec, C. and Evans, G. Effect of feed restriction and season on
LH and prolactin secretion, adrenal response, insulin and FFA in group housed pregnant gilts.
Anim. Reprod. Sci. 49, 1997: 179-190.(b)
Peltoniemi, O.A.T., Love, R.J., Klupiec, C., Revell, D.K. and Evans, G. Altered secretion of LH
does not explain seasonal effects on early pregnancy in gilts. Anim. Reprod. Sci. 49, 1997: 215-
224.(c)
Peltoniemi, O.A.T., Love, R.J., Heinonen, M., Tuovinen, V. and Saloniemi, H. Seasonal and
management effects on fertility of the sow: a descriptive study. Anim. Reprod. Sci. 55, 1999: 47-61.
(a)
Peltoniemi, O.A.T., Heinonen, M., Leppävuori, A. and Love, R.J. Seasonal effects on reproduction
in the domestic sow in Finland – A herd record study. Acta Vet. Scand. 40, 1999: 133-144. (b)
Perry, J.S., Heap, R.B. and Amoroso, E.C. Steroid hormone production by pig blastocysts. Nature
245, 1973: 45-47.
Prunier, A., Quesnel, H., Messias de Braganca, M., Kermabon, A.Y. Environmental and seasonal
influences on the return-to-estrus after weaning in primiparous sows: a review. Liv. Prod. Sci. 45,
1996: 103-110.
Pusateri, A.E., Smith, J.M., Smith, J.W., Thomford, P.J. and Diekman, M.A. Maternal recognition
of pregnancy in swine. I. Minimal requirement for exogenous estradiol-17β to induce either short or
long pseudopregnancy in cycling gilts. Biol. Reprod. 55, 1996: 582-589.(a)
Pusateri, A.E., Wilson, M.E. and Diekman, M.A. Maternal recognition of pregnancy in swine. II.
Plasma concentrations of progesterone and 13,14-dihydro-15-keto-prostaglandin F-2α during the

66
estrous cycle and during short and long pseudopregnancy in gilts. Biol. Reprod. 55, 1996: 590-
597.(b)
Reiter, R.J. Melatonin: the chemical expression of darkness. Molecular and cellular endocrinology.
79, 1991: C153-C158.
Robinson, J.E., Radford, H.M. and Karsch, F.J. Seasonal changes in pulsatile luteinizing hormone
(LH) secretion in the ewe: Relationship of frequency of LH pulses to day length and response to
oestradiol negative feedback. Biol. Reprod. 33, 1985: 324-334.
Roche, J.F., Karch, F.J., Foster, D.L., Takagi, S. and Dzuik, P.J. Effect of pinealectomy on estrus,
ovulation and luteizing hormone in ewes. Biol. Reprod. 2, 1970: 251-256.
Sammelwitz, P.H., Aldred, J.P. and Nalbandov, A.V. Mechanism of maintenance of corpora lutea
in pigs and rats. J. Reprod. Fert. 2, 1961: 387-393.
Shille, V.M., Karlbom, I., Einarsson, S., Larsson, K., Kindahl, H. and Edqvist, L.E. Concentrations
of progesterone and 15-keto-13,14-dihydroprostaglandin F2alpha in peripheral plasma during the
estrous cycle and early pregnancy in gilts. Zentralbatt Für Veterinärmedizin - Reihe A. 26A, 1979:
169-81.
Stone, B.A. and Seamark, R.F. Steroid hormones in uterine washings and in plasma of gilts between
days 9 and 15 after oestrus and between days 9 and 15 after coitus. J. Reprod. Fert. 75, 1985: 209-
21.
Stork, M.G. Seasonal reproductive inefficiency in large pig breeding units in Britain. Vet. Rec. 104,
1979: 49-52.
Szafranska, B. and Tilton, J.E. Prolactin as a luteotrophin during late pregnancy in pigs. J. Reprod.
Fert. 98, 1993: 643-8.
Tast, A., Hälli, O., Peltoniemi, O.A.T. A Practical method for non-surgically inserting intra-arterial
catheters in European wild boars (Sus scrofa). Vet. Res. Comm. 24, 2000: 25-30.
Trout, W.E., Hall, J.A., Stallings-Mann, M.L., Galvin, J.M., Anthony, R.V. and Roberts, R.M.
Steroid regulation of the synthesis and secretion of retinol-binding protein by the uterus of the pig.
Endocrinology 130, 1992: 2557-2564.

67
Turek, F.W. and Campbell, C.S. Photoperiodic regulation of neuroendocrine-gonadal activity. Biol.
Reprod. 20, 1979: 32-50.
Vallet, J.L., Christenson, R.K., Trout, W.E. and Klemcke, H.G. Conceptus, progesterone, and breed
effects on uterine protein secretion in swine. J. Anim. Sci. 76, 1998: 2657-2670.
Van der Meulen, J., Helmond, F.A and Oudenaarden, C.P.J. Effect of flushing of blastocysts on
days 10-13 on life-span of the corpora lutea in the pig. J. Reprod. Fert. 84, 1988: 157-162.
Wayne, N.L., Malbaux, B. and Karsch, F.J. Social cues can play a role in timing onset of the
breeding season of the ewe. J. Reprod. Fertil. 87, 1989: 707-7 Frank et al., 197713.
Wettemann, R.P. and Bazer, F.W. Influence of environmental temperature on prolificacy of pigs. J.
Reprod. Fertil. Suppl. 33, 1985: 199-208.
Wilson, M.R. and Love, R.J. The effect of the presence of older females on the maintenance of
pregnancy in gilts in the seasonal infertile period. In Proceedings: The International Pig Veterinary
Society, Scientific Committee of 11th International Pig Sciences Society Fritz Weibel, Thun 1990:
442.
Wrathall, A.E., Wells, D.E., Jones, P.C. and Foulkes, J.A. Seasonal variations in serum
progesterone levels in pregnant sows. Vet. Rec. 118, 1986: 685-687.
Zavy, M.T., Bazer, F.W., Thatcher, W.W. and Wilcox, C.J. A study of prostaglandin F2alpha as the
luteolysin in swine: V. Comparison of prostaglandin F, progestins, estrone and estradiol in uterine
flushings from pregnant and nonpregnant gilts. Prostaglandins 20, 1980: 837-51.
Xue, J.L., Dial, G.D., Marsh, W.E. and Davies, P.R. Multiple manifestations of season on
reproductive performance of commercial swine. J. Am. Vet. Med. Ass. 204, 1994: 1486-1489.





























