Embed Size (px)
Citation preview

Effect of Exotic Tree PlantationsEffect of Exotic Tree PlantationsEffect of Exotic Tree PlantationsEffect of Exotic Tree PlantationsEffect of Exotic Tree Plantationson Plant Diversity andon Plant Diversity andon Plant Diversity andon Plant Diversity andon Plant Diversity andBiological Soil Fertility inBiological Soil Fertility inBiological Soil Fertility inBiological Soil Fertility inBiological Soil Fertility inthe Congo Savanna:the Congo Savanna:the Congo Savanna:the Congo Savanna:the Congo Savanna:
With Special Reference to EucalyptsWith Special Reference to EucalyptsWith Special Reference to EucalyptsWith Special Reference to EucalyptsWith Special Reference to Eucalypts
Editor
F. Bernhard-Reversat

Effect of Exotic Tree Plantationson Plant Diversity and Biological Soil Fertility
in the Congo Savanna:With Special Reference to Eucalypts
Editor
F. Bernhard-Reversat
halaman depan 12/4/01, 5:14 PM1
Black

Cover and inside photos by C. Cossalter (July 2001)Understorey development in a mature clonal plantation of Eucalyptus hybrid,Pointe-Noire, Congo.
© 2001 by Center for International Forestry ResearchPublished in December 2001Printed by SMK Grafika Desa Putera, Indonesia
ISBN 979-8764-72-2
Bernhard-Reversat, F. (Editor) 2001. Effect of exotic tree plantations on plant diversityand biological soil fertility in the Congo savanna: with special reference to eucalypts.Center for International Forestry Research, Bogor, Indonesia, 71p.
Published byCenter for International Forestry ResearchMailing address: P.O. Box 6596 JKPWB, Jakarta 10065, IndonesiaOffice address: Jl. CIFOR, Situ Gede, Sindang Barang, Bogor Barat 16680, IndonesiaTel.: +62 (251) 622622; Fax: +62 (251) 622100E-mail: [email protected] site: http://www.cifor.cgiar.org
halaman depan 12/4/01, 5:14 PM2
Black

IntroductionGeneral objectives and sites: J.P. Bouillet and F. Bernhard-Reversat 1
Part 1. Improvement of biodiversity in plantations
Chapter 1. Effect of exotic tree plantations and site management on plantdiversity.C. Huttel and J.J. Loumeto 9
Chapter 2. Effect of exotic tree plantations on vertebrate fauna.A.Brosset 19
Part 2. Biological factors of fertility related to organic matter dynamics
Chapter 3. Litterfall, litter quality and decomposition changeswith eucalypt hybrids and plantation age.F. Bernhard-Reversat, J.J. Loumeto and J.P. Laclau 23
Chapter 4. Soil fertility changes with eucalypt hybridsand plantation age: soil organic matter.J.J. Loumeto and F. Bernhard-Reversat 31
Part 3. Biological factors of fertility related to the diversity and densityof soil biota
Chapter 5. Asymbiotic nitrogen fixationin savanna and eucalypt plantations.J. Le Mer and P. A. Roger 39
Chapter 6. Effect of exotic tree plantationson free living and plant parasitic soil nematodes and populationchanges with eucalypt hybrids and plantation age.P.M. Loubana and G. Reversat 43
Chapter 7. Effect of exotic tree plantationson invertebrate soil macrofauna and populationchanges with eucalypt hybrids and plantation age.I.M. Mboukou-Kimbatsa and F. Bernhard-Reversat 49
ConclusionsSoil biological fertility undergoes fundamental changeswhen fast growing exotic trees are planted on a poor savanna soil.F. Bernhard-Reversat and C. Huttel 57
References 61
Table of Contents
halaman depan 12/4/01, 5:14 PM3
Black

his report is the result of the collaboration of scientists from IRD (Institut de Recherche
pour le Développement, previously ORSTOM, France), UR2PI (Unité de Recherche sur la
Productivité des Plantations Industrielles, Congo), University of Brazzaville, DGRST,
(Direction de la Recherche Scientifique, Congo), CIRAD (Centre de Coopération Internationale en
Recherche Agronomique pour le Dévelopement), and MNHN (Museum National d’Histoire Naturelle,
France).
Financial support for this research and its publication provided by the CIFOR/JAPAN Research
Project ‘Rehabilitation of Degraded Tropical Forest Ecosystems’ and the involvement of Mr. C. Cossalter
(CIFOR) in planning the projects are acknowledged. ECO S.A. and the Congolese Service National de
Reboisement (S.N.R.) are acknowledged for welcoming the field work in their plantations. Directors
of the Centre IRD of Pointe-Noire, Mr. A. Joseph and Mr. J. Geoffroy, are acknowledged for supplying
field and laboratory facilities for the research team. Soil and plant nutrient analyses were carried out
at the Analysis Laboratory of the Centre IRD of Pointe-Noire, under the direction of P. Esmard and L.
Veysseyre. Technical assistance from Mrs. V. Teixeira and S. Sid Saya (IRD Bondy), Mr. M. Tsatsa
and Mr. J.J. Kibondzi (IRD Pointe-Noire) is acknowledged.
The authors are greatly indebted to Dr. J.A. Parrotta (USDA Forest Service) and Dr. J.W. Turnbull
for reviewing and improving the manuscript. We also thank Mr. C. Cossalter, Mr. G. Applegate,
Dr. R. Nasi and Dr. T. Toma (CIFOR) for their valuable comments.
Acknowledgements
TTTTT
halaman depan 12/4/01, 5:14 PM4
Black

General Objectives and SitesJ.P. Bouillet1 and F. Bernhard-Reversat2
1 UR2PI, B.P. 1120, Pointe-Noire, Congo.2 Laboratoire d’Ecologie des Sols Tropicaux, Centre IRDd’Ile de France, 32 avenue Henri Varagnat, 93143 Bondy,France.
Introduction
Introduction
F ast-growing tree plantations, mainlyEucalyptus spp., Pinus spp. and Acacia spp.,are widespread in tropical countries.
Among these species eucalypts are continuouslybeing planted and planted areas have increasedtwofold every ten years since 1970, reaching 14million hectares in 1997 (FAO 1982; Pandey 1997;Bouvet 1999). In 1977, eucalypts accounted for18% of the forest plantation areas in tropicalregions compared to 4% for pines and 7% foracacias (Pandey 1997). Their main use is for pulpproduction and eucalypt pulp production in theworld increased from 22 to 31 million tonsbetween 1979 and 1995 (Wilson et al. 1995).
Environmental Impact ofFast-growing Tree Plantationsin the TropicsThis has been a controversial topic, some arguingthat these species exhaust soil water and nutrientresources, and prevent understorey vegetationgrowth, thus resulting in decreased biodiversityand in further soil erosion and loss of fertility(Poore and Fries 1985; Abbasi and Vinithan 1997;Bouvet 1998). Several regional meetings (India,Thailand) have debated these issues, and anabundant literature has emerged from the 1991,1995, 1997 meetings of the IUFRO ‘Silvicultureand breeding of eucalypts’ group (IUFRO workingparty 2.08.03). The IUFRO work focussed onassessing the environmental impact of eucalypt
plantation compared to other ecosystems, nativeor planted. Four main issues were considered.
The first issue, concerning biodiversity, wasaddressed during the Word Bank BiodiversityRehabilitation project (Parrotta 1995b), whichexamined the hypothesis that tree plantation cancatalyse biodiversity rehabilitation on degradedtropical lands (Parrotta et al. 1997) Mainlyconducted on sites with degraded soils andvegetation, these studies found fast-growing treeplantations (eucalypts and other species) helpedregeneration of undergrowth plants fromsurrounding forests, thus increasing theirbiodiversity and fertility (Geldenhuys 1997;Harrington and Ewel 1997). A few studies reportedthe similar results for animal populations (Brosset1997; Mboukou-Kimbatsa et al. 1998).
The second issue, dealt with nutrientrequirement and nutrient cycling. It was shownthat nutrient requirements of eucalypt and pinestands were of the same order of magnitude andwere generally higher than those of nativeecosystems (Baker and Attiwill 1985; Poore andFries 1985; Cortez 1996; Grove et al. 1996).However nutrient outputs resulting from harvest

2 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
varied greatly according to species, even amongeucalypts (Morais et al. 1990; O’Connell andGrove 1996; Gonçalves et al. 1997; Sankaran etal. 2000), and according to harvest practices forthe previous stand (Bouillet et al. 2000; Gonçalveset al. 1999; Laclau et al. 2000a). Nutrients neededfor stand growth were provided primarily throughnutrient cycling by litterfall and internal fluxes(Turner and Lambert 1983, 1996; Attiwill et al.1996; Cortez 1996). These processes allowed asatisfactory tree growth on nutrient-poor soils.
The third issue dealt with changes in thephysico-chemical properties of the soils.Comparisons were made between monospecificeucalypt stands with monospecific stands of othertree species (Bernard-Reversat 1987; Brent et al.1996; Cortez 1996; Negi and Sharma 1996;Harmand 1998; Dambrine et al. 1999), mixedstands eucalypts and nitrogen fixing trees (Binkley1997; Parrotta 1999) or the previous vegetation(Herbert 1996; Binkley and Resh 1999). Thegeneral trend in these studies was a decrease ofsoil Ca and N under eucalypt or pine monoculture.However the introduction of N fixing speciesimproved N balance and tree growth.
The fourth issue concerned the water needs ofeucalypt plantations. It was shown that water useefficiency depended on water availability(Kallarackal and Somen 1997; Chunyang 1999)and on genetic origin of the planted trees (Bondand Stock 1990; Dye 1996; Chunyang 1999).Hydrological studies carried out in South Africashowed a decrease in streamflows when treeplantations were grown in the watersheds, and thisdecrease was greater with eucalypt than with pine,although pines exhibit a lower water-use efficiency(Dye 1996; Scott and Smith 1997). Such resultscould guide the choice of the species to be grown.
Background and GeneralObjectives of the StudyIn the southern coastal region of Congo nearPointe-Noire, savanna covers large areas on verypoor sandy soils, and only small areas close tohuman settlements are used for agriculturalpurposes. These savannas have been used since1978 for plantations of fast-growing trees, mainlyeucalypts, which are grown commercially. Thecommon management practice is a first seven-year
rotation established from rooted-cutting clonesfollowed by a second and sometimes a thirdcoppice rotation of the same duration.Experimental plantations of several fast-growingspecies, among which are eucalypts, Australianacacias and tropical pines, were also establishedin this area, and 100 km west of it, near Loudimain the Niari valley on clay soils (Fig. 0.1).
In the Congo, due to nutrient poor soils andlow rainfall climate, sustainability is a priority areafor scientific research on eucalypt productionsystems (Bouillet et al. 1997) and multidisciplinarystudies focusing on this goal have been conductedsince 1997 (Fig. 0.2). The first topic deals withbiogeochemical cycles, for which nutrient balancestudies are conducted in a first rotation eucalyptcrop and in a native savanna (Laclau et al. 2000b,2000c). The second topic is specially concernedwith logging residue management, the objectiveof the CIFOR network ‘Site Management andProductivity in Tropical Plantation Forests’(Bouillet et al. 2000). The third topic is theimportance of the biological factors in fertility,which has received little attention in mostmanagement studies, and is the purpose of thepresent report. Although not being exhaustive, a
EQUA.GUINEA
CAMEROON
CENTRAL AFRICANREPUBLIC
G A B O N
100 200 km
0
0
100 200 ml
DEMOCRATICREPUBLIC
OF THECONGO
Owando
Djambala
Oyo
ANGOLA(Cabinda)
Mossendjo
Loudima
PointeNoire
OuessoImpfondo
SouthAtlanticOcean
Congo
BRAZZAVILLE
Figure 0.1 Map of the Republic of Congo showingthe study sites
adapted from :http://www. lib.utexas.edu/libs/pcl/map_collection/africa.html

3Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
set of biotic processes was investigated in relationto plantation age and in particular the effectsuccessive rotations on biotic changes was studied.The World Bank Biodiversity Rehabilitationproject with CIFOR support gave an opportunityto study the development of native vegetationtogether with vertebrates and soil fauna withinplantations. Following these studies, the CIFOR/Japan project ‘Rehabilitation of Degraded TropicalEcosystems’ supported a second phase to furtherinvestigate the biological factors of fertility.
Because plant biodiversity controls soil biotaand is known to influence the whole trophic chain(Lamotte 1979; Lavelle et al. 1997), it is oftremendous importance in soil and ecosystemfunctioning (Schulze and Mooney 1993). Aparticular feature of the Congolese plantations wastheir establishment on savanna with poor soil andvegetation. Although young plantations of fast-growing species (eucalypt, acacia and pine) exhibitsparse understorey vegetation, undergrowth startsinvading plantations and building up thickets afterabout ten years. The effect of both savanna andforest on undergrowth colonisation, and the trendsof understorey vegetation changes with age,received special attention (Part 1). Therelationships between vegetation and fertility areclosely linked to litter and soil organic matterquality and quantity; these were studied in an ageseries of eucalypt hybrids and clones (Part 2). Thedensity of the main taxa of macrofauna andnematodes, which act either through organic matter
decomposition, soil biogenic structures, andparasitism, was assessed according to the age ofplantation and logging. Nitrogen fixation ability,related to the dramatic N shortage which isexpected to occur in aging plantations, was alsostudied (Part 3).
Location and Site DescriptionThe study areas were located near Pointe-Noire(4° 48’ S, 11° 54’ E) and near Loudima (4° 07’ S,13° 51’ E). At both sites, annual precipitationaverages 1250 mm, with four dry months fromJune to September and the mean annualtemperature is 25°C.
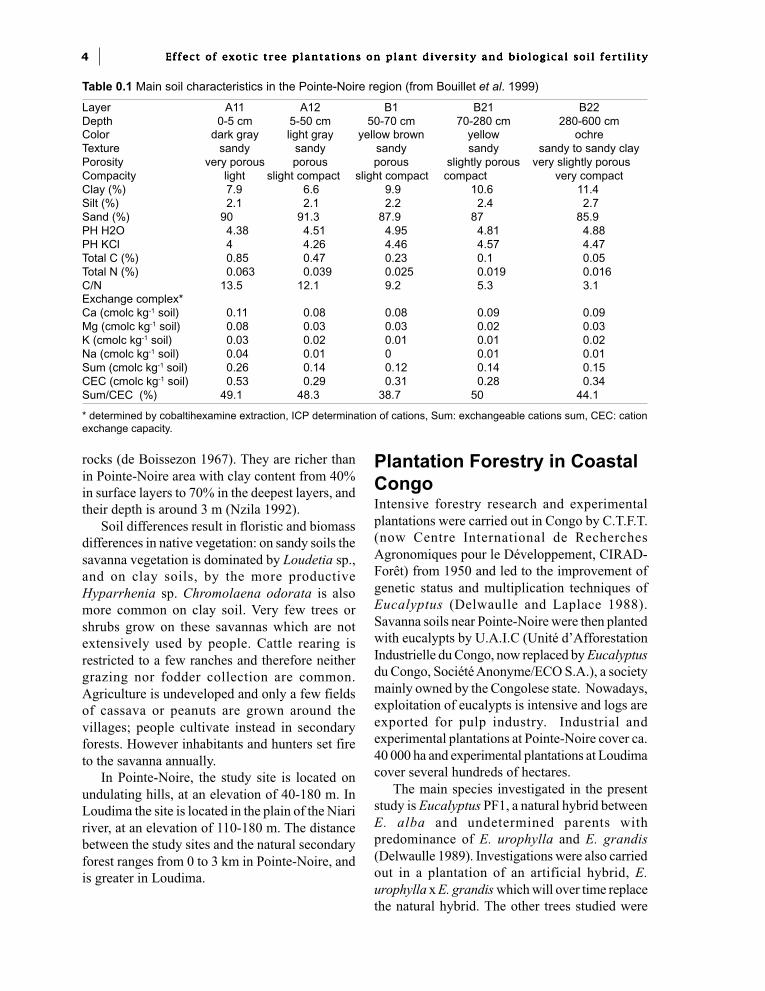
Soils are highly desaturated ferrallitic soils(French classification). According to the FAO/UNESCO classification they are FerralicArenosols at Pointe-Noire and Xanthic ferralsolsat Loudima. In Pointe-Noire, where most of thestudy was carried out, soils are sandy in texture(clay content less than 5% in the top layer). Theywere developed from thick Plio-Pleistocenecontinental sandy deposits. Soils in this series varyfrom 80 to 300 m depth, and are underlaid by adolomitic sandstone clay series from theCretaceous period (Jamet 1975). They areparticularly poor in nutrients and organic matter(the exchange capacity is 0.4 to 0.7 cmol kg-1 andthe C content 0.7% in the 0-10 cm layer) (Table0.1).
In Loudima the soils were developed onsediments (schisto-calcareous) from Precambrian
Figure 0.2 Representation of the coordinated research programme on plantation sustainability in the Congo
UR2PI-CIRADEXPERIMENT
Nutrient cycle, water andnutrient budget in savanna,and on 1st rotation planted
crop
CORE GOALSustainability of
eucalypt plantations
CIFOR/UR2PI/IFSEXPERIMENT
Impact of slash management andsoil preparation on plantation
productivity and soil characteristics
CIFOR/IRD/UR2PI EXPERIMENTEffect of plot age and rotations
on biological fertility
4 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
rocks (de Boissezon 1967). They are richer thanin Pointe-Noire area with clay content from 40%in surface layers to 70% in the deepest layers, andtheir depth is around 3 m (Nzila 1992).
Soil differences result in floristic and biomassdifferences in native vegetation: on sandy soils thesavanna vegetation is dominated by Loudetia sp.,and on clay soils, by the more productiveHyparrhenia sp. Chromolaena odorata is alsomore common on clay soil. Very few trees orshrubs grow on these savannas which are notextensively used by people. Cattle rearing isrestricted to a few ranches and therefore neithergrazing nor fodder collection are common.Agriculture is undeveloped and only a few fieldsof cassava or peanuts are grown around thevillages; people cultivate instead in secondaryforests. However inhabitants and hunters set fireto the savanna annually.
In Pointe-Noire, the study site is located onundulating hills, at an elevation of 40-180 m. InLoudima the site is located in the plain of the Niaririver, at an elevation of 110-180 m. The distancebetween the study sites and the natural secondaryforest ranges from 0 to 3 km in Pointe-Noire, andis greater in Loudima.
Plantation Forestry in CoastalCongoIntensive forestry research and experimentalplantations were carried out in Congo by C.T.F.T.(now Centre International de RecherchesAgronomiques pour le Développement, CIRAD-Forêt) from 1950 and led to the improvement ofgenetic status and multiplication techniques ofEucalyptus (Delwaulle and Laplace 1988).Savanna soils near Pointe-Noire were then plantedwith eucalypts by U.A.I.C (Unité d’AfforestationIndustrielle du Congo, now replaced by Eucalyptusdu Congo, Société Anonyme/ECO S.A.), a societymainly owned by the Congolese state. Nowadays,exploitation of eucalypts is intensive and logs areexported for pulp industry. Industrial andexperimental plantations at Pointe-Noire cover ca.40 000 ha and experimental plantations at Loudimacover several hundreds of hectares.
The main species investigated in the presentstudy is Eucalyptus PF1, a natural hybrid betweenE. alba and undetermined parents withpredominance of E. urophylla and E. grandis(Delwaulle 1989). Investigations were also carriedout in a plantation of an artificial hybrid, E.urophylla x E. grandis which will over time replacethe natural hybrid. The other trees studied were
Table 0.1 Main soil characteristics in the Pointe-Noire region (from Bouillet et al. 1999)
Layer A11 A12 B1 B21 B22Depth 0-5 cm 5-50 cm 50-70 cm 70-280 cm 280-600 cmColor dark gray light gray yellow brown yellow ochreTexture sandy sandy sandy sandy sandy to sandy clayPorosity very porous porous porous slightly porous very slightly porousCompacity light slight compact slight compact compact very compactClay (%) 7.9 6.6 9.9 10.6 11.4Silt (%) 2.1 2.1 2.2 2.4 2.7Sand (%) 90 91.3 87.9 87 85.9PH H2O 4.38 4.51 4.95 4.81 4.88PH KCl 4 4.26 4.46 4.57 4.47Total C (%) 0.85 0.47 0.23 0.1 0.05Total N (%) 0.063 0.039 0.025 0.019 0.016C/N 13.5 12.1 9.2 5.3 3.1Exchange complex*Ca (cmolc kg-1 soil) 0.11 0.08 0.08 0.09 0.09Mg (cmolc kg-1 soil) 0.08 0.03 0.03 0.02 0.03K (cmolc kg-1 soil) 0.03 0.02 0.01 0.01 0.02Na (cmolc kg-1 soil) 0.04 0.01 0 0.01 0.01Sum (cmolc kg-1 soil) 0.26 0.14 0.12 0.14 0.15CEC (cmolc kg-1 soil) 0.53 0.29 0.31 0.28 0.34Sum/CEC (%) 49.1 48.3 38.7 50 44.1
* determined by cobaltihexamine extraction, ICP determination of cations, Sum: exchangeable cations sum, CEC: cationexchange capacity.

5Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Table 0.2 Characteristics of plots in first project
Code* Species Age Type** Treatment(yr)
Kouilou Region (Pointe-Noire) on sandy soil
Ea Eucalyptus PF 1 6 E Age seriesEb 11 E Age seriesEc 13 E Age seriesEd 16 E Age seriesEe 18 E Age seriesEf 20 E Age seriesEg 15 E Spacing 4.8 x 5.4 mEh 15 E Spacing 2.5 x 2.8 mEi 13 C Distance to forest 10 mEj 13 C Distance to forest 50-100 mEk 12 C Control no herbicide or fireEl 12 C Herbicide applied 1 year beforeEm 12 C Herbicide applied 3 months beforeEn 12 C Fires in Nov. 1993 and Nov. 1994Eo 12 C Fire in Nov. 1993A Acacia auriculiformis 16 EP Pinus caribaea 15 CSa Savanna Native vegetationF Secondary forest Native vegetation
Niari Region (Loudima) on clay soil
EL Eucalyptus PF 1 26 EAL Acacia auriculiformis 12 EPL Pinus caribaea 26 ESL Savanna Native vegetation
*Codes identify plots in this report. **E = experimental plot, C = commercial plot.
Acacia auriculiformis, for which only smallexperimental plots were available, and Pinuscaribaea, for which commercial plantations areavailable on small areas (1000 ha in the Pointe-Noire region and 1500 ha in Loudima).The meansize of 7-year-old Eucalyptus (age of harvest)when growing well is 28 m in height, 14-16 cm indiameter at breast height, 16 m2 ha-1 in basal area.This gives an average stand volume ofapproximately 175 m3 per hectare. Total biomassis approximately 100-120 t ha-1 in both acacia andeucalypts at 7 years of age (Loumeto 1986,Bernhard-Reversat et al. 1993).
Trees were planted after tillage to a depth of20 cm and fertilisation in the plantation hole(150 g tree-1 NPK 13-13-21). There was nosubsequent fertilisation. Manual or mechanicalweeding took place for two or three years afterplanting. Trees were planted at various spacings,with a trend toward closer spacing, ranging from5.0 x 5.0 m to 3.5 x 3.5 m. There was no thinning.
Trees were clear-felled and harvested whenplantations were 7 years old, except in plotspreserved for experimental purposes. One to twosprouts were then allowed to coppice from thestumps in order to establish a new plantation. Thestands were fertilised (200 kg ha-1 NPK 13-13-21)between 1 and 2 years old and harvested when 5to 7 years old. Fires in the plantations areaccidental but occur at an increasing rate, and nowaffect 8-17% of plantations, depending on the area.
The Studied PlotsDuring the first project (World Bank BiodiversityRehabilitation project), first rotation stands werechosen for study, mainly in the experimentalplantations. In the Pointe-Noire region (sandysoils) the studies were conducted in an age seriesof PF1 eucalypt and in plots chosen to evaluatethe effect of tree spacing, distance to forest,herbicide and fire on understorey regeneration and

6 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
biodiversity. Studies were also conducted on oneplot of each of A. auriculiformis and P. caribaea.In the Loudima region (clay soils) one plot of eachspecies, Eucalyptus PF1, A. auriculiformis, and P.caribaea, was studied (Table 0.2)
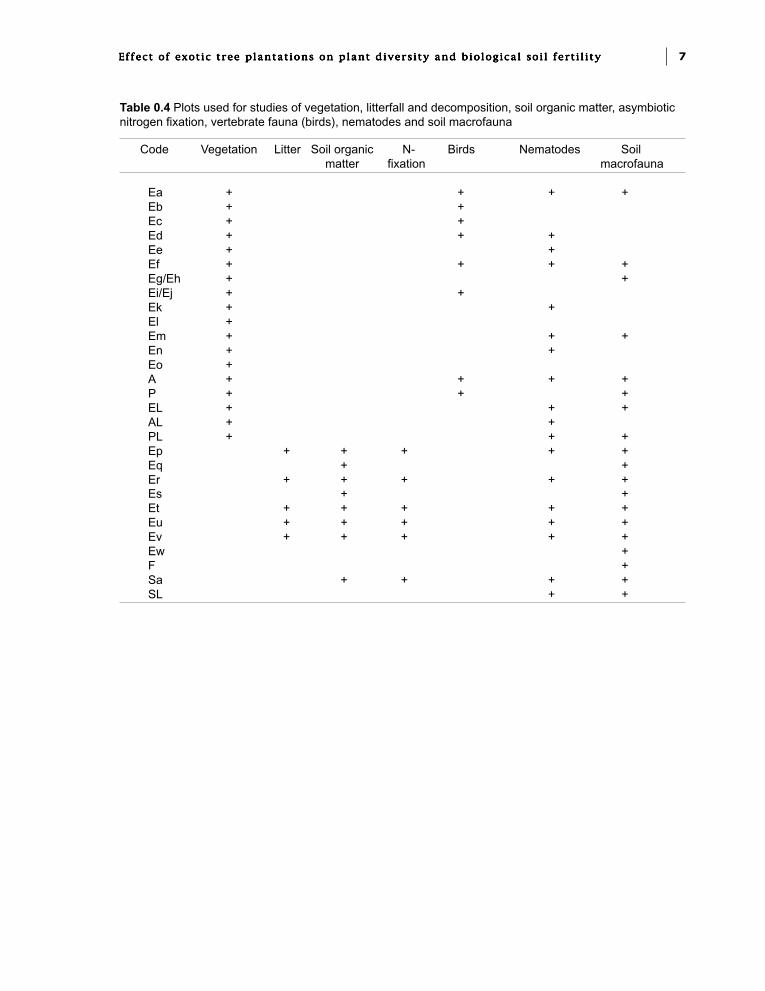
The second project (Biological FertilityProject) involved detailed studies of vegetation,litterfall and decomposition, soil organic matter,Nitrogen asymbiotic fixation, vertebrate fauna(mainly birds), nematodes and soil microfauna.
Research sites were chosen in the commercialeucalypt plantations of ECO S.A. near Pointe-Noire. These included an age series of EucalyptusPF1 comprised of first rotation stands and coppicestands, two of which were just clear-felled, andby one first rotation plot of E.urophylla x E.grandis (Table 0.3). One multiclonal plot of E.urophylla x E. grandis and one multiclonal plot ofE. PF1 were used for clone comparisons. Moststudies were carried out in a subset (Table 0.4).
Table 0.3 Characteristics of plots in second project in Pointe-Noire region
Hybrid Clone Plot Tree Plantation Present Previousage (yr) age (yr) type* state** exploitation
Ep E. PF 1 1-41 6 6 1R no 0Eq 6 0 1R CF 0Er 13 6 Coppice no 1Es 14 0 Coppice CF 1Et 19 7 Coppice no 2Eu 19 19 1R no 0Ev E. PF1 and various 8 8 1R no 0
urograndisEw E. PF 1 various 5 6 1R no 0Sb Savanna no
* 1R = first rotation, **CF = clear felled.
150
100
50
0
300
200
100
0
precipitation
temperature
prec
ipita
tion
(mm
)
tem
pera
ture
(o C
)
months (Jan - Dec)
Figure 0.3 Mean annual precipitation and temperature at Pointe-Noire (1990-1996)
7Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Table 0.4 Plots used for studies of vegetation, litterfall and decomposition, soil organic matter, asymbioticnitrogen fixation, vertebrate fauna (birds), nematodes and soil macrofauna
Code Vegetation Litter Soil organic N- Birds Nematodes Soilmatter fixation macrofauna
Ea + + + +Eb + +Ec + +Ed + + +Ee + +Ef + + + +Eg/Eh + +Ei/Ej + +Ek + +El +Em + + +En + +Eo +A + + + +P + + +EL + + +AL + +PL + + +Ep + + + + +Eq + +Er + + + + +Es + +Et + + + + +Eu + + + + +Ev + + + + +Ew +F +Sa + + + +SL + +

Improvement of Biodiversity in Plantations
Part 1

1 Le Caillou, Valojoux, 24290 Montignac, France.2 Laboratoire de Biologie Végétale, Université deBrazzaville, BP 69, Brazzaville, Congo.
Effect of Exotic Tree Plantationsand Site Management on Plant DiversityC. Huttel1 and J.L. Loumeto2
Chapter 1.
Introduction
T he ability of plantations to improvebiodiversity must be taken into accountby foresters when they have to address the
problem of plantation sustainability, especially ina poor environment like that of the Congoleseeucalypt plantations. Species’ richness undernatural eucalypt forests can vary considerably anddiversified understoreys are well-known(Kirkpatrick 1997). The catalytic effect of forestplantations on the regeneration of biodiversity andthe possibility of restoration of degraded lands hasbeen documented (Parrotta 1993, 1995; SilvaJunior et al. 1995; Lamb 1998; Parrotta andKnowles 1999) and is relevant to the ‘Eucalyptuscontroversy’ where criticism related to biodiversityissues has prevailed over a long period (IFS 1989;Calder et al. 1992; see also the introduction to thisreport). Although most exotic tree plantations areestablished for wood production, and forestvegetation reconstitution is only a secondary andoften undesirable effect, a less obvious componentof biodiversity, that of soil organisms, mightbenefit from plant diversity and improve soilquality.
Studies on understorey vegetation in eucalyptplantations have been reported by Basanta et al.1989; Madeira et al. 1989; Lugo et al. 1990;Parrotta 1995a; Silva et al. 1995; and Michelsenet al. 1996. Some data are available on theunderstorey vegetation of Congolese eucalyptplantation (Nongamani 1988; Soler 1994).Loumeto and Huttel reported in 1997 preliminary
observations within the framework of a multi-sitestudy which involved several countries (Parrota1995b). This study adopted a common surveymethodology for vegetation studies in order toobtain comparable results. In the present study,species richness, species behaviour and standparameters were analysed under fast-growing treeplantations and compared with the characteristicsof nearby secondary forest patches.
Study Sites and MethodsThe general characteristics of the study plots aregiven above in the ‘General objectives and sites’(Table 0.2) and also in Loumeto and Huttel (1997).The investigated planted species were mainly aeucalypt hybrid (PF1), and secondarily Acaciaauriculiformis and Pinus caribaea. Surveys weremore intensive on the sandy site near Pointe-Noire,due to easier accessibility and because moreplantations were grown there than on the clay soilsite near Loudima, which was added in order toassess the effect of soil. In both places, the originalsavanna was also investigated, and a secondaryforest plot was surveyed at Pointe-Noire site. Mostobservations were carried out in experimentalunexploited plantations, but some observations incommercial plantations allowed evaluation of theeffects of logging, fire or herbicide application (cf.

10 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
the introduction and Table 0.2.). These disturbedplots were chosen with similar age and history,first planted in 1983 or 1984 and logged in 1990or 1991. Therefore all selected commercial plotswere coppice with trees from 5 to 6.5 years old.The sampling method took into account the levelof heterogeneity within each type of plots. Incommercial plots, survey circles were locatedexclusively on plantation lines where undergrowthhad grown undisturbed. In doing so, the surveyavoided areas where undergrowth was depletedby vehicle traffic and areas showing excessivegrowth due to increased nutrient supplies throughdecomposing material in swaths. Plants werecounted according to height classes (0-30 cm, 30-200 cm and over 200 cm height) in concentriccircles of 1, 2 and 5 m radius respectively. Thenumber of replicates in each plot (from 4 to 11)was chosen to fit with vegetation heterogeneity.The height and diameter at breast height (dbh) ofwoody plants, if relevant, were measured, whereasthe presence and cover percentages of herbaceousspecies, mainly Poaceae and Cyperaceae, wereestimated. Planted tree characteristics, density,height and dbh, were also measured.
The plants listed in the undergrowth wereseparated in two main groups, the planted timbertrees and the native species. The second group wasdivided according to their usual habitat (forest or
savanna), or usual behaviour for ruderal speciescommon in disturbed habitats such as roadsidesor burned areas. Further in the text, the expressions‘preference groups’ and ‘preferring species’ willrefer to the most usual habitat of the species.
Results and Discussion
Heterogeneity in production plotsUndergrowth was relatively homogeneous inexperimental undisturbed plots. In commercialplots, silvicultural practices added further spatialheterogeneity to the effects of other parameters.During the two first years of plantation growth,undergrowth was weeded using a cover-cropbetween plantation lines, and manually within thelines when necessary. So undergrowth wasgenerally more developed on the plantation linesthan between them (Fig. 1.1).
Eucalypts are clear fallen and harvested when7 years old. When logs are taken out the remainingwood is collected for charcoal or fuelwood, andthe residues, eucalypt and undergrowth leaves andtwigs are stacked in swath lines. The generalpattern of these disturbances and their effects onundergrowth distribution (Fig. 1.1) showed regularalternating areas of dense and high woodyundergrowth in the swath interlines, mainly
Plantation linesSwaths between plantationlines
Untouched spaces betweenplantation lines
Haulage tracks
Planted eucalypts
Increasingundergrowthdensity
Locationof the samplingcircles
Figure 1.1 Undergrowth heterogeneity in eucalypt production plots
11Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
herbaceous cover on the haulage tracks, and stripsof undisturbed undergrowth on the lines.
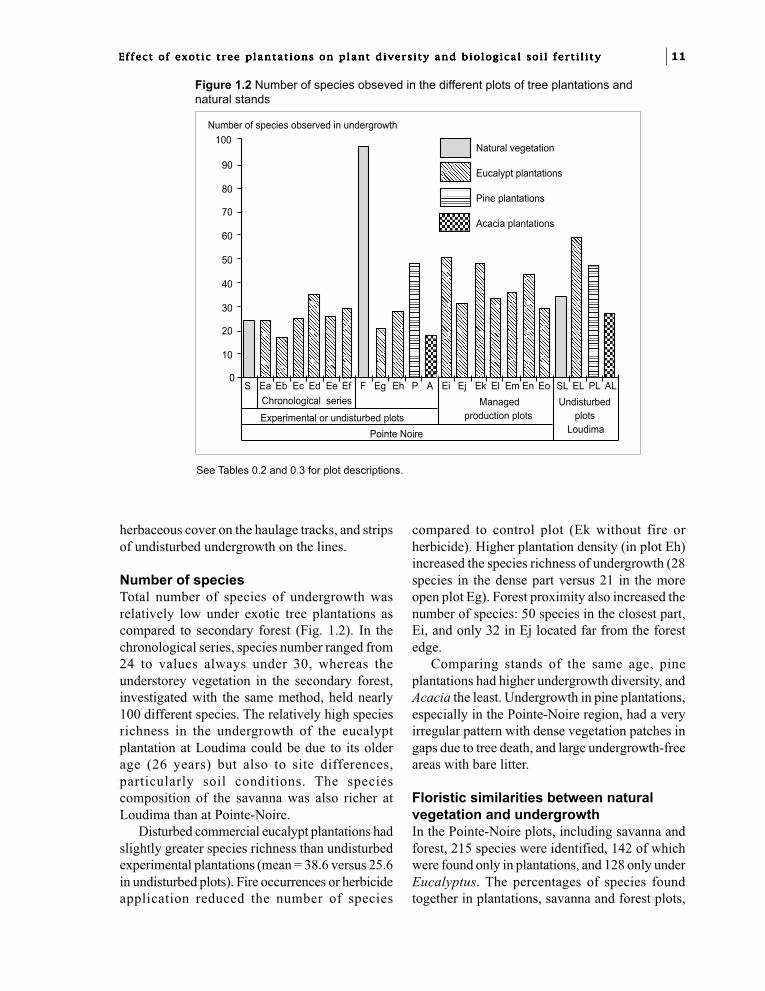
Number of speciesTotal number of species of undergrowth wasrelatively low under exotic tree plantations ascompared to secondary forest (Fig. 1.2). In thechronological series, species number ranged from24 to values always under 30, whereas theunderstorey vegetation in the secondary forest,investigated with the same method, held nearly100 different species. The relatively high speciesrichness in the undergrowth of the eucalyptplantation at Loudima could be due to its olderage (26 years) but also to site differences,particularly soil conditions. The speciescomposition of the savanna was also richer atLoudima than at Pointe-Noire.
Disturbed commercial eucalypt plantations hadslightly greater species richness than undisturbedexperimental plantations (mean = 38.6 versus 25.6in undisturbed plots). Fire occurrences or herbicideapplication reduced the number of species
compared to control plot (Ek without fire orherbicide). Higher plantation density (in plot Eh)increased the species richness of undergrowth (28species in the dense part versus 21 in the moreopen plot Eg). Forest proximity also increased thenumber of species: 50 species in the closest part,Ei, and only 32 in Ej located far from the forestedge.
Comparing stands of the same age, pineplantations had higher undergrowth diversity, andAcacia the least. Undergrowth in pine plantations,especially in the Pointe-Noire region, had a veryirregular pattern with dense vegetation patches ingaps due to tree death, and large undergrowth-freeareas with bare litter.
Floristic similarities between naturalvegetation and undergrowthIn the Pointe-Noire plots, including savanna andforest, 215 species were identified, 142 of whichwere found only in plantations, and 128 only underEucalyptus. The percentages of species foundtogether in plantations, savanna and forest plots,
Figure 1.2 Number of species obseved in the different plots of tree plantations andnatural stands
See Tables 0.2 and 0.3 for plot descriptions.

12 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
or found only in plantations for each kind ofplantation was calculated (Fig.1.3).
Species found exclusively in plantations madethe largest species’ group ranging from 59 to 67%of the total number of species. An importantproportion of species was found only in oneplantation type, including mainly Anthocleistaschwenfurtii in undisturbed eucalypt plantationsand Maprounea africana in the disturbed ones.There were no outstandingly frequent species inpine and acacia undergrowth. Taking into accountthe species present in more than one kind ofplantation, the highest similarity was foundbetween undisturbed and disturbed eucalyptplantations, and the most frequent among thesespecies were three Clerodendron spp., Bartieranigritiana, Macaranga monandra orPsorospermum tenuifolium. Species found onlyunder plantation belonged mainly to the forest-preferring group (over 60%), and 25% wereruderal species.
The percentage of species found in bothsavanna and plantations varied inversely with thepercentage of species found in both forest andplantations. The first were savanna species(Asystasia gangetica or Annona arenaria) and alsoruderal species (Scleria boivinii, Hypoxis cfangustifolia). The second were exclusively forest-preferring species and the most frequent were theerect shrub Chaetocarpus africanus, the climbingshrub Premna lucens and the shrub-like forest edgespecies Psychotria cf peduncularis.
The savanna and forest plots contained somespecies that were not found under plantations. Onlytwo species, Loudetia arundinacea and Vernoniapotamophylla, were exclusively present in thesavanna plot. Most savanna plot species were
found under eucalypt plantations (19 from 24 werepresent in undisturbed eucalypt plots and 21 indisturbed plots). This number was much reducedunder the other tree species: one species (theruderal Scleria boivinii) under acacia and threespecies under pine.
The pattern was different for the species foundin the forest plots, and 71 out of 96 were neverfound outside the forest. Some of these specieshad high frequencies in the forest plot, amongwhich were emergent tree species like Carapaprocera or Hymenocardia ulmoides, although mostwere shrub-like or woody understorey species:Microdesmis puberula, Cola sp.1, Rhabdophyllumwelwitschii, Cnestis urens, or the strictly forestvine Gnetum africanum. Numerous species (41)were found at low frequency (only in one surveycircle).
Disturbed eucalypt plantation plots shared 21species with the forest plot, 11 with undisturbedplots, 13 in pine plantation and 5 in acaciaplantation. Three forest-preferring shrubs specieswere present in all types of plantation and in theforest plot, Chaetocarpus africanus, Psychotria cfpeduncularis and Premna lucens. Two forestpioneers, Alchornea cordifolia and Macarangaspinosa are frequent in the forest and eucalyptplantations but absent under pine or acacia; twomore species were found only in forest andeucalypt undergrowth, Anthocleista schwenfurtiiin undisturbed plantations and Maprouneaafricana in commercial areas. There were noespecially frequent species shared with the forestin pine or acacia undergrowth.
Figure 1.3 Percentages of species shared with savanna, forest or exclusiveof plantation undergrowth
13Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
EoEi EjEg Ek El Em En SL EL PL ALEa Eb Ec Ed Ee Ef FS
Pointe-Noire
Eh0
10
20
30
40
50
60
70
80
90
%100
P
LoudimaUndisturbed
plots
Savanna Ruderal Planted ForestPreferencegroups
Chronological series
Undisturbed plotsDisturbed plots
A
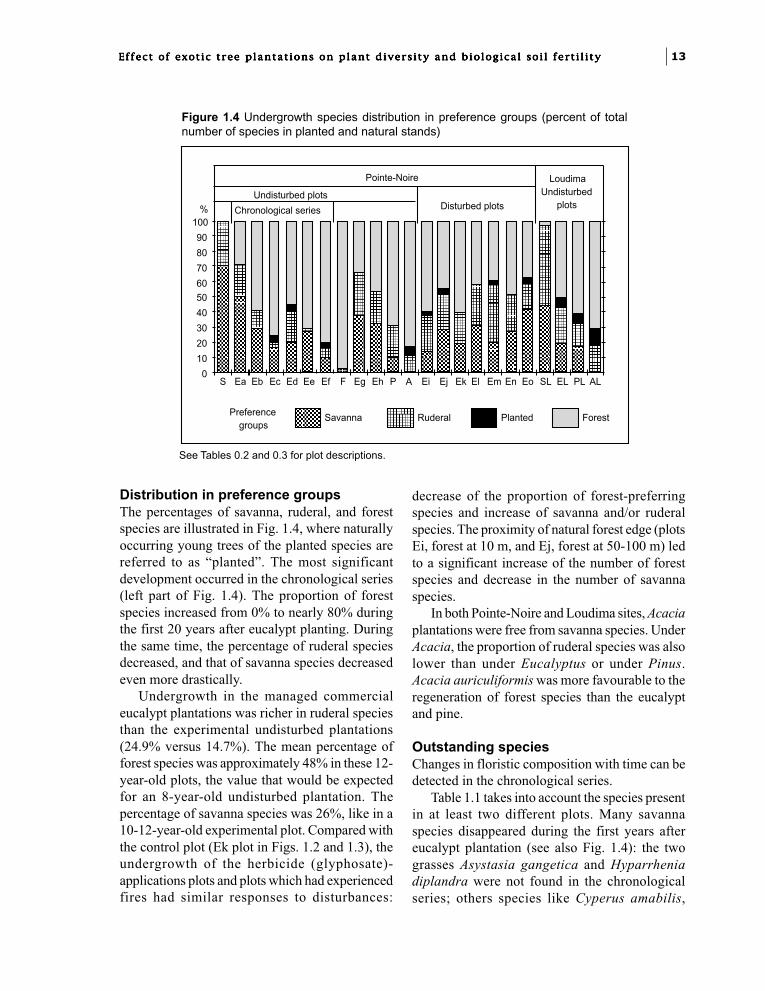
Distribution in preference groupsThe percentages of savanna, ruderal, and forestspecies are illustrated in Fig. 1.4, where naturallyoccurring young trees of the planted species arereferred to as “planted”. The most significantdevelopment occurred in the chronological series(left part of Fig. 1.4). The proportion of forestspecies increased from 0% to nearly 80% duringthe first 20 years after eucalypt planting. Duringthe same time, the percentage of ruderal speciesdecreased, and that of savanna species decreasedeven more drastically.
Undergrowth in the managed commercialeucalypt plantations was richer in ruderal speciesthan the experimental undisturbed plantations(24.9% versus 14.7%). The mean percentage offorest species was approximately 48% in these 12-year-old plots, the value that would be expectedfor an 8-year-old undisturbed plantation. Thepercentage of savanna species was 26%, like in a10-12-year-old experimental plot. Compared withthe control plot (Ek plot in Figs. 1.2 and 1.3), theundergrowth of the herbicide (glyphosate)-applications plots and plots which had experiencedfires had similar responses to disturbances:
decrease of the proportion of forest-preferringspecies and increase of savanna and/or ruderalspecies. The proximity of natural forest edge (plotsEi, forest at 10 m, and Ej, forest at 50-100 m) ledto a significant increase of the number of forestspecies and decrease in the number of savannaspecies.
In both Pointe-Noire and Loudima sites, Acaciaplantations were free from savanna species. UnderAcacia, the proportion of ruderal species was alsolower than under Eucalyptus or under Pinus.Acacia auriculiformis was more favourable to theregeneration of forest species than the eucalyptand pine.
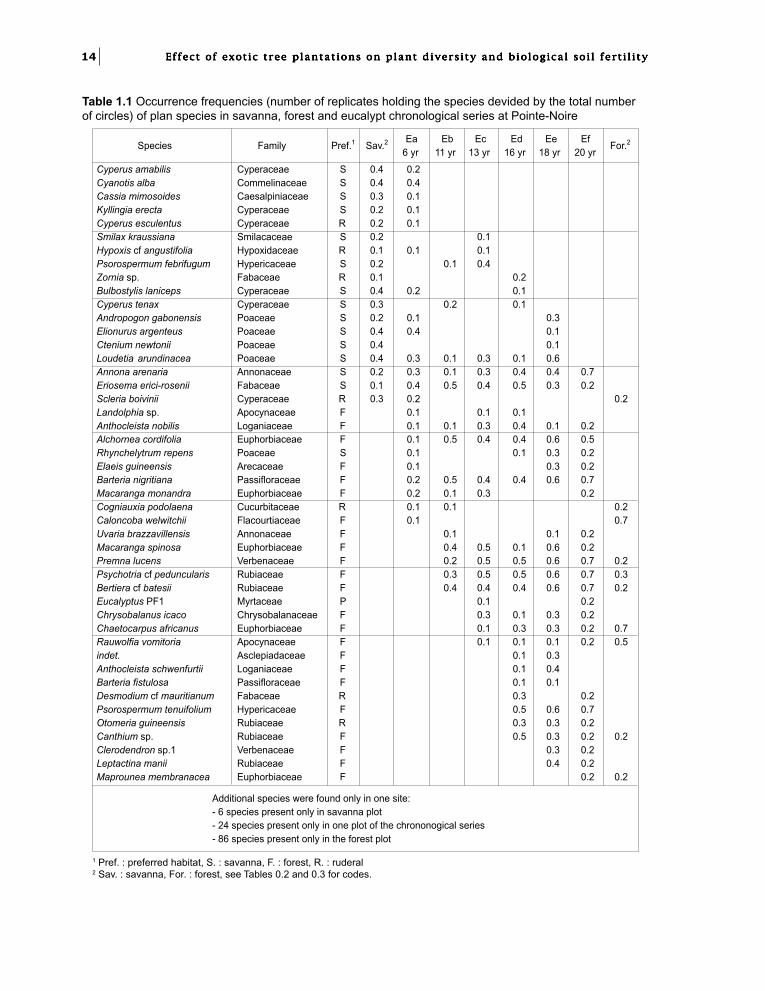
Outstanding speciesChanges in floristic composition with time can bedetected in the chronological series.
Table 1.1 takes into account the species presentin at least two different plots. Many savannaspecies disappeared during the first years aftereucalypt plantation (see also Fig. 1.4): the twograsses Asystasia gangetica and Hyparrheniadiplandra were not found in the chronologicalseries; others species like Cyperus amabilis,
Figure 1.4 Undergrowth species distribution in preference groups (percent of totalnumber of species in planted and natural stands)
See Tables 0.2 and 0.3 for plot descriptions.
14 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Cyperus amabilisCyanotis albaCassia mimosoidesKyllingia erectaCyperus esculentusSmilax kraussianaHypoxis cf angustifoliaPsorospermum febrifugumZornia sp.Bulbostylis lanicepsCyperus tenaxAndropogon gabonensisElionurus argenteusCtenium newtoniiLoudetia arundinaceaAnnona arenariaEriosema erici-roseniiScleria boiviniiLandolphia sp.Anthocleista nobilisAlchornea cordifoliaRhynchelytrum repensElaeis guineensisBarteria nigritianaMacaranga monandraCogniauxia podolaenaCaloncoba welwitchiiUvaria brazzavillensisMacaranga spinosaPremna lucensPsychotria cf peduncularisBertiera cf batesiiEucalyptus PF1Chrysobalanus icacoChaetocarpus africanusRauwolfia vomitoriaindet.Anthocleista schwenfurtiiBarteria fistulosaDesmodium cf mauritianumPsorospermum tenuifoliumOtomeria guineensisCanthium sp.Clerodendron sp.1Leptactina maniiMaprounea membranacea
CyperaceaeCommelinaceaeCaesalpiniaceaeCyperaceaeCyperaceaeSmilacaceaeHypoxidaceaeHypericaceaeFabaceaeCyperaceaeCyperaceaePoaceaePoaceaePoaceaePoaceaeAnnonaceaeFabaceaeCyperaceaeApocynaceaeLoganiaceaeEuphorbiaceaePoaceaeArecaceaePassifloraceaeEuphorbiaceaeCucurbitaceaeFlacourtiaceaeAnnonaceaeEuphorbiaceaeVerbenaceaeRubiaceaeRubiaceaeMyrtaceaeChrysobalanaceaeEuphorbiaceaeApocynaceaeAsclepiadaceaeLoganiaceaePassifloraceaeFabaceaeHypericaceaeRubiaceaeRubiaceaeVerbenaceaeRubiaceaeEuphorbiaceae
Species Family
SSSSRSRSRSSSSSSSSRFFFSFFFRFFFFFFPFFFFFFRFRFFFF
0.40.40.30.20.20.20.10.20.10.40.30.20.40.40.40.20.10.3
Pref.1 Sav.2Ea
6 yr
0.20.40.10.10.1
0.1
0.2
0.10.4
0.30.30.40.20.10.10.10.10.10.20.20.10.1
0.1
0.2
0.10.10.5
0.10.5
0.50.10.1
0.10.40.20.30.4
Eb11 yr
Ec13 yr
0.10.10.4
0.30.30.4
0.10.30.4
0.40.3
0.50.50.50.40.10.30.10.1
Ed16 yr
0.20.10.1
0.10.40.5
0.10.40.40.1
0.4
0.10.50.50.4
0.10.30.10.10.10.10.30.50.30.5
Ee18 yr
0.30.10.10.60.40.3
0.10.60.30.30.6
0.10.60.60.60.6
0.30.30.10.30.40.1
0.60.30.30.30.4
0.70.2
0.20.50.20.20.70.2
0.20.20.70.70.70.20.20.20.2
0.20.70.20.20.20.20.2
Ef20 yr
For.2
0.2
0.20.7
0.20.30.2
0.70.5
0.2
0.2
Additional species were found only in one site:- 6 species present only in savanna plot- 24 species present only in one plot of the chrononogical series- 86 species present only in the forest plot
Table 1.1 Occurrence frequencies (number of replicates holding the species devided by the total numberof circles) of plan species in savanna, forest and eucalypt chronological series at Pointe-Noire
1 Pref. : preferred habitat, S. : savanna, F. : forest, R. : ruderal2 Sav. : savanna, For. : forest, see Tables 0.2 and 0.3 for codes.
15Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Annona arenariaAnthocleista nobilisBarteria nigritianaBulbostylis lanicepsCyperus amabilisElionurus argenteusLoudetia arundinaceaOldenlandia affinisAlchornea cordifoliaDissotis cf rotundifoliaMacaranga monandraPremna lucensPsorospermum febrifugumAsystasia gangeticaBertiera cf batesiiChromolaena odorataChrysobalanus icacoCyperus esculentusHypoxis cf angustifoliaMaprounea membranaceaPanicum brevifoliumRhynchelytrum repensAndropogon gabonensisChaetocarpus africanusDesmodium cf mauritianumEucalyptus PF1Macaranga spinosaMaprounea africanaPanicum cogoenseSmilax kraussianaVernonia brazzavillensisAdenia cissampeloidesindet.Cogniauxia podolaenaElaeis guineensisEriosema erici-roseniiLandolphia spPsychotria cf peduncularisScleria boivinii
AnnonaceaeLoganiaceaePassifloraceaeCyperaceaeCyperaceaePoaceaePoaceaeRubiaceaeEuphorbiaceaeMelastomataceaeEuphorbiaceaeVerbenaceaeHypericaceaeAcabthaceaeRubiaceaeAsteraceaeChrysobalanaceaeCyperaceaeHypoxidaceaeEuphorbiaceaePoaceaePoaceaePoaceaeEuphorbiaceaeFabaceaeMyrtaceaeEuphorbiaceaeEuphorbiaceaePoaceaeSmilacaceaeAsteraceaePassifloraceaeAsclepiadaceaeCucurbitaceaeArecaceaeFabaceaeApocynaceaeRubiaceaeCyperaceae
Species Family
SFFSSSSRFRFFSSFRFRRFRSSFRPFFSSFFFRFSFFR
0.40.30.30.40.10.20.40.40.30.10.10.40.40.10.40.10.1
0.30.2
0.10.10.2
0.30.10.10.20.30.10.1
0.10.10.10.3
Pref. Ek El
0.40.60.40.10.30.10.60.30.10.10.1
0.10.30.40.10.10.3
0.10.10.1
0.30.1
0.3
0.1
0.1
0.1
0.40.40.40.40.10.30.60.40.40.10.30.60.30.10.40.60.10.40.10.1
0.3
0.6
0.3
0.10.4
Em En
0.60.60.40.60.30.30.40.1
0.40.1
0.10.30.10.10.30.1
0.10.3
0.10.10.3
0.10.6
0.1
Eo
0.50.10.40.50.10.50.40.50.20.10.10.50.20.10.2
0.5
0.10.30.40.30.1
0.10.2
0.10.3
0.1
Ei
0.40.60.80.60.20.80.60.60.80.60.40.8
0.40.60.20.4
0.6
0.40.8
0.40.20.40.8
0.20.40.20.2
0.2
0.6
0.40.60.60.60.30.40.60.30.40.10.10.30.4
0.10.10.30.60.30.10.10.10.3
0.1
0.10.4
0.10.1
0.3
Ej
Additional species were found only in few sites:-15 species present only in one plot-18 species present only in two plots
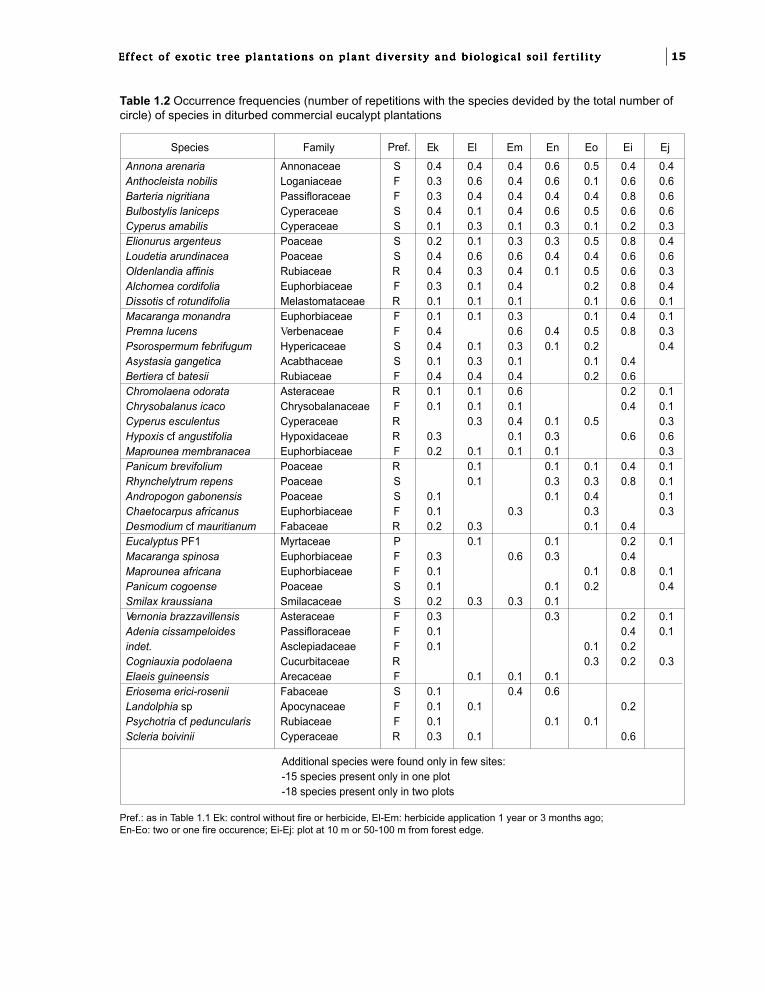
Table 1.2 Occurrence frequencies (number of repetitions with the species devided by the total number ofcircle) of species in diturbed commercial eucalypt plantations
Pref.: as in Table 1.1 Ek: control without fire or herbicide, El-Em: herbicide application 1 year or 3 months ago;En-Eo: two or one fire occurence; Ei-Ej: plot at 10 m or 50-100 m from forest edge.
16 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
SavannaE. PF1 6-yr-oldE. PF1 11-yr-oldE. PF1 13-yr-oldE. PF1 16 yr-oldE. PF1 18-yr-oldE. PF1 20-yr-oldSecondary forestE. PF1 low densityE. PF1 high densityE. PF1 10 m from forestE. PF1 50-100 m from forestE. PF1 without herbicide or fireE. PF1 herbicide 1 year agoE. PF1 herbicide 3 months agoE. PF1 two fire occurrencesE. PF1 one fire occurrenceacacia plantationpine plantationsavanna (Loudima)eucalypt (Loudima)acacia (Loudima)pine (Loudima)
Treatment
- 6.25.75.37.85.57.2-
3.810.8
8.26.78.47.59.67.55.69.37.5-
14.07.2
10.2
Density (a)Codes
SaEaEbEcEdEeEfFEgEhEiEjEkElEmEnEoAPSLELALPL
planted
Soil Cov. (b) Height (c) Basal area (d)
und. (e)
08.0
11.728.559.0
162.9106.0177.8
33.139.450.325.424.180.212.321.212.8
374.337.5
4.991.1
° 118.2
herbac. woody
521112++232243344125212
21234455333332323532454
planted und.(e)
- 23.824.628.923.623.928.2
- 26.823.423.521.919.922.221.618.921.414.322.1
- 25.116.426.1
° 1.86.29.2
19.019.012.817.5
3.43.74.62.73.74.35.92.74.55.74.44.8
22.6°
15.9
planted und.(e)
- 13.012.813.715.8
9.420.2
- 8.9
16.115.411.811.3
9.614.610.0
8.514.134.5
- 28.510.048.5
00
0.10.51.32.50.8
14.10.10.10.1e
0.20.20.1
00.10.3e
1.44.5°
3.2
a: number of woody plants over 30 cm high in 100 m2
b: soil cover according to the Braun-Blanquet scalec: in metres, mean height for planted trees, maximum values for undergrowth°: missing datae: value < 0.1
Cyanotis lanata or Kylingia erecta were presentonly in the first stages of plantation growth. Onthe other hand, some savanna species persistedeven in older stands, among which were Loudetiaarundinacea and the savanna characteristic shrub,Annona arenaria.
Forest species invaded the undergrowth insuccessive waves. The trees Barteria nigritianaand Anthocleista nobilis, and the shrub Alchorneacordifolia were representative of the first forestpioneers. A second wave, colonising the 10-year-old plantations, comprised mainly shrubs, erectspecies like Bartiera cf batesii, Psychotria cfpeduncularis or climbing species like Premnalucens. The last wave in the 20 years chronologicalseries comprised shrubs like Canthium sp,Maprounea membranacea and Psorospermumtenuifolium.
In the disturbed commercial plantations (Table1.2), some savanna species were present in mostplots. Besides those which persisted in theundisturbed experimental plots of the age seriesmentioned above, the most frequent savannaspecies were herbs like Bulbostylis laniceps,Elyonurus argenteus, Psorospermum febrifugum.The sedge Cyperus amabilis, which disappearedrapidly in the chronological series, was present inall the disturbed plots, while Cyanotis lanata waspresent only in two plots.
The ruderal species were numerous in thecommercial plantation plots. The most frequentincluded mainly forbs, like Oldenlandia affinis orDissotis cf rotundifolia. It was noticeable that theweed Chromolaena odorata was more widelyspread in commercial plantations than in theundisturbed experimental plots.
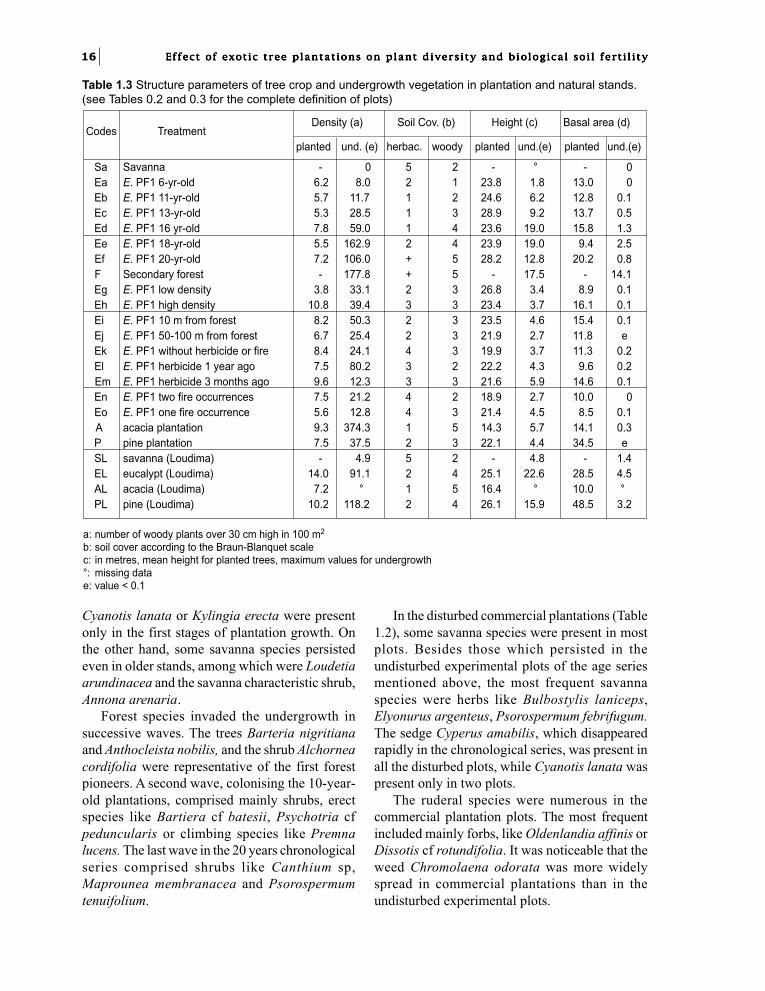
Table 1.3 Structure parameters of tree crop and undergrowth vegetation in plantation and natural stands.(see Tables 0.2 and 0.3 for the complete definition of plots)

17Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
The most frequent forest species present incommercial plantations were trees, Anthocleistanobilis, Barteria nigritiana, or erect shrubs,Alchornea cordifolia, Bertiera cf batesii,Macaranga monandra , Maprouneamembranacea, or the climbing shrub Premnalucens. Undergrowth in commercial plantationsincluded few species of the last invading wave,which was observed in experimental undisturbedplots: only Maprounea membranacea was foundwith noticeable frequency.
Plot ordinationA factorial correspondence analysis (FCA) of thesurveyed plots at Pointe-Noire, taking into accountthe presence/absence of species in undergrowth(Fig. 1.5) and including both Pointe-Noire andLoudima plots, resulted in a strong separationbetween the two sites, and a lack of discriminationamong Pointe-Noire plots; consequently the laterwere analysed separately. The species present inat least two plots were used for FCA calculationsto reduce the excessive weight of very uncommonspecies. The points representing the plots aredistributed within a triangle whose vertices aresavanna, forest and acacia plantation. Axis 1 canbe assumed to represent the increasing frequencyof woody species or forest-preferring species. Thedisturbed plantations were close to each other andsituated between the 6 and 10-year-old undisturbedplantation; the undergrowth of the forest proximityplot (Ei) and the control plot without herbicide orfire (Ek) were the most advanced towards forestregeneration. The undisturbed eucalypt plots(chronological series and spacing tests from Ea toEh) showed a classification according to plot ageand showed a trend to floristic similarity with theacacia plantation. The pine plantation was situatedin an intermediate place. The points located closeto savanna point corresponded to very disturbedsituations (recent herbicide application or two fireoccurrences, Em and En).
When the FCA was drawn with speciescoordinates instead of plot ordinates, the resultsled to similar conclusions. The FCA computationsmade with Loudima plots (not shown here)confirmed the different undergrowth in acaciaplantations, the similarity of eucalypt undergrowthwith savanna floristic list, and the pine plot positionclose to eucalypt plots. The similarity of plot
respective positions in Pointe-Noire and Loudimacould help the interpretation of axis 2, andsuggested that axis 2 represented an effect of exotictimber plantation introducing a floristic pattern,different from native vegetation.
Woody undergrowth parametersThe density of woody plants in undergrowthincreased with age in the chronological series(Table 1.3). Two woody forest-preferring species,Psychotria cf peduncularis and Psorospermumtenuifolium had particularly high densities. In thecommercial eucalypt plantations (5 to 6.5 years),woody undergrowth density was approximatelythe same than that of undisturbed 11/13-year-oldplantations. Undergrowth density in the 15-year-old pine plantation was comparable to that ofeucalypt plantations of the same age, whereas the16-year-old acacia plantation showed a higherdensity value, 80% of which resulted from onlytwo forest species, Psychotria cf peduncularis andTarenna soyauxii.
Undergrowth plant height increased with standage in undisturbed plots. Plantation managementin commercial plantations, including undergrowthcontrol practices, reduced the height ofspontaneous vegetation to approximately that ofthe undergrowth of an 8-year-old undisturbed plot.
6 yr (Ea)
high density plantation (Eh)
20 yr (Ef)
savanna (S)
forest (F)
acaciaplantation (A)
pineplantation (P)
1,0
1,0
Axis 1
Axis 2
Natural vegetation
Chronological series
Others undisturbed plots
Disturbed production plots
16 yr (Ed)
EmEk
forest nearestplot (Ei)
En
low density plantation (Eg)
Ek: control eucalypt plot no fire or herbicideEm: herbicide application 3 months beforeEn: two fire occurrences
Figure 1.5 Ordination of the planted and naturalstands at Pointe-Noire

18 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
plantations, undergrowth in undisturbed plotstended to be different from that of the forest andsavanna, as in acacia plantations, whereasdisturbances resulted in a vegetation nearer tosavanna. Effects of nurse species on undergrowthfloristics reported in the literature are variable,significant in Parrotta (1995a) and Keenan et al.(1997), but of little importance in Lugo et al.(1993). The proximity of natural forest mayinfluence vegetation as far as 100 m (Parrotta1993) or 300 m (Keenan et al. 1997) whereas inthe Pointe-Noire site, the influence of forestproximity was important only at shorter distances.
A good understanding of the complexity ofvegetation processes requires long-termmonitoring of vegetation change. Further studiesare needed. They could be focused on seeddispersion mechanisms, including biotic factors(birds and bats, see chapter 2) and abiotic factors(e.g. wind), and on specific heterogeneity ofundergrowth with the presence of dense low-diversity patches. More attention has to be paid tothe variability of the floristic composition anddensities in the undergrowth of the natural forest,which might affect undergrowth composition innearby plantations, and to management practices.
Heights of undergrowth under pine and acaciawere lower than in undisturbed eucalyptplantations of similar age, and were also low inthe Loudima plots.
The basal area of undergrowth was low. In thechronological series, undergrowth basal areavaried from 0 to 10% of total basal area, althougha higher percentage of 21% was found in the 18-year-old plot because of the high mortality ofeucalypts. In the commercial plantation plots,undergrowth basal area was lower than 1% of thetotal.
Discussion and ConclusionsExotic timber plantations initiated an importantfloristic change from the grass-dominatedvegetation of native savanna toward undergrowthcharacterised by the importance of woody plants.The changing rate of the vegetation was sloweddown by management practices or accelerated byhigh plantation density or by forest proximity. Thebasal area of undergrowth was low in all thestudied plots, and far less than that of the plantedtrees.
Floristic change was characterised by theincrease of total species number and the proportionof forest-preferring species, with increasing plotage. An increase in the number of species withplantation age is a common observation (Parrotta1993, 1999; Abbasi and Vinithan 1999).Nevertheless, important differences in floristiccomposition still remained between naturalsecondary forest and plantation undergrowth. Onone hand, many forest-preferring species observedunder plantation were not found in the forest plotand, on the other hand, most species identified inthe forest plot were not found in the plantationundergrowth. Undergrowth vegetation could becharacterised as typical of a pioneer stage of forestsuccession. Nevertheless, the important proportionof ruderal species could be an indicator of plantcommunity fragility.
Floristic change trends were differentaccording to the planted tree species. Undergrowthin acacia plantations was very different to that offorest or other tree plantations, whereas under pinethe spontaneous vegetation showed a trend towardsthat of the forest vegetation. In eucalypt

Effect of Exotic Tree Plantationson Vertebrate FaunaA. Brosset1
1 Laboratoire d’Ecologie Végétale, Muséum Nationald’Histoire Naturelle, 4 avenue du Petit Château 91800Brunoy, France.
Chapter 2.
Introduction
T he diversity and abundance of thevertebrate fauna was studied as an indexof the environment quality. A study carried
out in the Kouilou forest, 70 km north of Pointe-Noire, showed a rich diversity in the birdpopulation (Dowsett-Lemaire and Dowsett, 1991).Although a comparable study is not available forthe nearby savanna, an increased diversity of birdpopulations was hypothesised when forestplantations are grown on savanna soil. There areindications in the literature that the abundance ofbirds in old plots with dense undergrowth, mightcontribute to the dispersal of seeds, resulting inthe maintenance and the increasing diversity anddensity of the undergrowth, as observed by Duncanand Chapman (1999). Abbasi and Vinithan (1999)found that the bird population was not less ineucalypt plantations (hybrid tereticornis xglobulus) compared to the other planted speciesthey studied. Bird populations in eucalypt, pineand acacia plantations were surveyed to test theseobservations and hypothesis (Brosset 1997).
Study Sites and MethodsAn age series of eucalypts, a 16 year-old acaciaand a 15 year-old pine plantation were selected inPointe-Noire (Table 2.1). Most bird species wereknown from previous extensive studies in Gabon(Brosset 1990, 1997). In Congo, each selected areawas surveyed during six hours, walking along theperimeter, then along transects 50 to 100 m apart,
with regularly spaced ten minute pauses. A list ofthe species encountered was prepared (Brosset1997). Since the experimental plots which wereselected for other measurements were too smallfor an adequate survey of bird populations, theywere extended to a radius of about 500 m.Occasional observations for mammals were madeduring the bird study.
Birds
Bird ecological groupsThe nature of ecological relationships betweenplantations and their avifauna may be understoodconsidering bird assemblages or “guilds”. Sixguilds were defined (Brosset 1997):
1. The guild of savanna ground feeder birdsexploiting young plantations. It comprises twoplovers (genus Charadrius and Vanellus), onecursor (genus Cursorius), two larks (genusMirafra and Macronyx), one pipit (genusAnthus). These birds are invertebrate feeders.
20 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
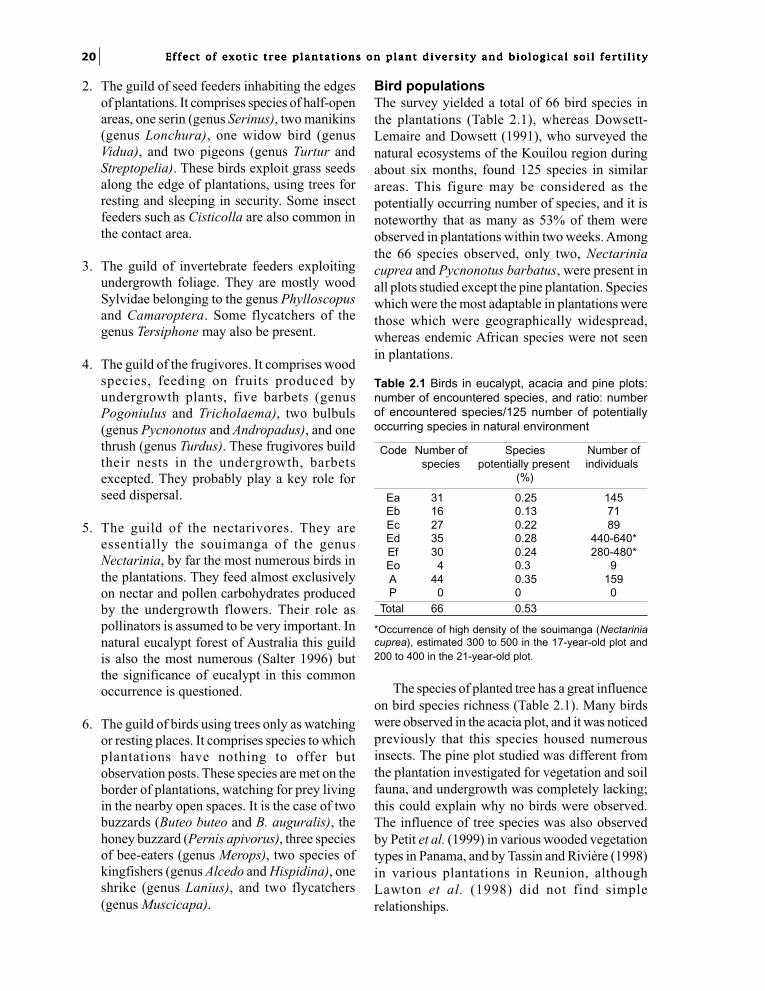
Table 2.1 Birds in eucalypt, acacia and pine plots:number of encountered species, and ratio: numberof encountered species/125 number of potentiallyoccurring species in natural environment
Code Number of Species Number ofspecies potentially present individuals
(%)
Ea 31 0.25 145Eb 16 0.13 71Ec 27 0.22 89Ed 35 0.28 440-640*Ef 30 0.24 280-480*Eo 4 0.3 9A 44 0.35 159P 0 0 0
Total 66 0.53
*Occurrence of high density of the souimanga (Nectariniacuprea), estimated 300 to 500 in the 17-year-old plot and200 to 400 in the 21-year-old plot.
2. The guild of seed feeders inhabiting the edgesof plantations. It comprises species of half-openareas, one serin (genus Serinus), two manikins(genus Lonchura), one widow bird (genusVidua), and two pigeons (genus Turtur andStreptopelia). These birds exploit grass seedsalong the edge of plantations, using trees forresting and sleeping in security. Some insectfeeders such as Cisticolla are also common inthe contact area.
3. The guild of invertebrate feeders exploitingundergrowth foliage. They are mostly woodSylvidae belonging to the genus Phylloscopusand Camaroptera. Some flycatchers of thegenus Tersiphone may also be present.
4. The guild of the frugivores. It comprises woodspecies, feeding on fruits produced byundergrowth plants, five barbets (genusPogoniulus and Tricholaema), two bulbuls(genus Pycnonotus and Andropadus), and onethrush (genus Turdus). These frugivores buildtheir nests in the undergrowth, barbetsexcepted. They probably play a key role forseed dispersal.
5. The guild of the nectarivores. They areessentially the souimanga of the genusNectarinia, by far the most numerous birds inthe plantations. They feed almost exclusivelyon nectar and pollen carbohydrates producedby the undergrowth flowers. Their role aspollinators is assumed to be very important. Innatural eucalypt forest of Australia this guildis also the most numerous (Salter 1996) butthe significance of eucalypt in this commonoccurrence is questioned.
6. The guild of birds using trees only as watchingor resting places. It comprises species to whichplantations have nothing to offer butobservation posts. These species are met on theborder of plantations, watching for prey livingin the nearby open spaces. It is the case of twobuzzards (Buteo buteo and B. auguralis), thehoney buzzard (Pernis apivorus), three speciesof bee-eaters (genus Merops), two species ofkingfishers (genus Alcedo and Hispidina), oneshrike (genus Lanius), and two flycatchers(genus Muscicapa).
Bird populationsThe survey yielded a total of 66 bird species inthe plantations (Table 2.1), whereas Dowsett-Lemaire and Dowsett (1991), who surveyed thenatural ecosystems of the Kouilou region duringabout six months, found 125 species in similarareas. This figure may be considered as thepotentially occurring number of species, and it isnoteworthy that as many as 53% of them wereobserved in plantations within two weeks. Amongthe 66 species observed, only two, Nectariniacuprea and Pycnonotus barbatus, were present inall plots studied except the pine plantation. Specieswhich were the most adaptable in plantations werethose which were geographically widespread,whereas endemic African species were not seenin plantations.
The species of planted tree has a great influenceon bird species richness (Table 2.1). Many birdswere observed in the acacia plot, and it was noticedpreviously that this species housed numerousinsects. The pine plot studied was different fromthe plantation investigated for vegetation and soilfauna, and undergrowth was completely lacking;this could explain why no birds were observed.The influence of tree species was also observedby Petit et al. (1999) in various wooded vegetationtypes in Panama, and by Tassin and Rivière (1998)in various plantations in Reunion, althoughLawton et al. (1998) did not find simplerelationships.

21Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
The presence of many species and individualsfrom the bird community, seems to be linked tothe development of a naturally regeneratingundergrowth vegetation, and the number of birdspecies and individuals was significantly relatedto the density and biodiversity of plants in theundergrowth. Relationships between number ofbirds and age of plot were not observed. Thevariability of plots regarding the number of speciesand the individual number of birds was assumedalso to be partly linked to shape and size of plots:a longer edge compared to the area promote birdpresence, activity and nutrition, and a large blockwith few understorey plants, as encountered atHinda, was almost devoid of birds.
The type of undergrowth present affected birduse. When undergrowth was exclusivelyChromolaena odorata, a spreading weed, no birdswere observed. The plant association mostoccupied by birds combined Psychotria sp.,Alchornea cordifolia, Maprounea and Canthiumspp. Psychotria flowers, as well as those of anotherpioneer plant, Anthocleista sp., provide food formost nectarivorous souimanga, whereas the otherthree fruiting species provide food for the bulbulPycnonotus barbatus and the genera Pogoniulusand Turtur. The only species that built their nestsin eucalypt trees were souimangas, while someundergrowth trees, like Maprounea andMacaranga spp., were used for nesting by severalspecies.
The number of migratory bird species andindividuals from palearctic countries was relativelyhigh. Seven species were observed two of whichwere present at fairly high densities. First, thesebirds prefer a vegetation which is physionomicallysimilar to their native environment (Brosset andErard 1986), and secondly, tropical forests aregenerally saturated with local sedentary species.However migratory birds were encountered inplots where the understorey vegetation was thickand varied.
MammalsMammals are known to scatter seeds and toimprove their germination. In eucalypt plantations,in the short time available for the study, presenceor signs of presence of mammals were noticed.Tracks of jackals, Canis adustus, which eat fruits,were seen everywhere, and individuals were
observed several times during previous field tripsand once during the present study. Tracks of civet,Civectictis civetta, which is known to be efficientin scattering seeds (Pendge 1994), were seen inand nearby eucalypt plots. One small forestbovidae, Cephalophus sp., was observed twice inprevious studies, and tracks were seen once.Antelopes are intensively hunted and are probablyscarce in the region (Wilson and Wilson 1991).Bats were not studied at night and one colonygroup of Micropteropus was observed duringdaytime. This frugivore bat is widely spread inCongo (Brosset 1966), and could play a significantrole in seed dispersal.
ConclusionsUnlike in drier countries, where no undergrowthoccurs in eucalypt plantations and few bird speciesare found (e.g. Smith 1974), higher rainfall inCongo permits development of undergrowthvegetation which favours the occurrence of birdspecies (Loumeto and Huttel 1997 and chapter 1,this report). Bird species which are able to colonisethe plantations are those which utilise shrub-likevegetation, and the abundance of undergrowthdetermined the diversity and density of birdpopulations in the studied plantations. The samerelationship could occur with mammals(Armstrong and Henbergen 1996). Moreover, theedges of the planted parcels along the alleys aremore frequently used than the inner parts, andsmall parcels have higher bird densities than largerones. It is noticeable, however, that the speciesplanted is a main factor of bird colonisation, andtrees like acacia with abundant herbivorous anddetritivorous insects attract a great number of birds.

Biological Factors of Fertility Related to OrganicMatter Dynamics
Part 2

Litterfall, Litter Qualityand Decomposition Changeswith Eucalypt Hybrids and Plantation AgeF. Bernhard-Reversat1 , J.J. Loumeto2 and J.P. Laclau3
1 Laboratoire d’Ecologie des Sols Tropicaux, Centre IRDd’Ile de France, 32 avenue Henri Varagnat, 93143 Bondy,France.2 Laboratoire de Biologie Végétale, Université deBrazzaville, BP 69, Brazzaville, Congo.3 UR2PI, B.P. 1120, Pointe-Noire, Congo.
Chapter 3.
Introduction
Litterfall serves three main functions in theecosystem: energy input for soil microfloraand fauna, nutrient input for plant nutrition,
and material input for soil organic matterdevelopment. The first two functions arecompleted through decomposition andmineralisation, and the third throughdecomposition and humification. Those functionsare related to main soil processes, especiallymicrobiological activity and soil fauna activity,litter quality and quantity. Annual litterfall in theCongolese eucalypt plantations was previouslyshown to be low compared to many natural tropicalforests or to other fast growing trees (Loubelo1990; Bernhard-Reversat 1993), and to have highcontent of phenolic compounds (Bernhard-Reversat 1999) which are known to affect manybiological processes. The hypothesis of the controlof litter quality, quantity and decomposition byplantation age, management, and hybrid wasstudied.
Studied Plots and Methods
Litter measurementsLitterfall was measured in 6, 13, and 19-year-oldplots of Eucalyptus PF1 and an 8-year-old plot ofEucalyptus urophylla x E. grandis hybrid referredto as E. urograndis (plots Ep, Er, Et, Eu, Ev). Asthe plots were logged every 7 years, the trees were6-7 years old in all plots except in the 19-year-old
plot, where half of the area was never logged andthe trees were 19 years old, and the other half wascoppiced. The 8-year-old plot of E. urograndis(clone 18-85) referred to as (plot Ev), was alsosampled to compare its litter production with thatof PF1. In the 6-year-old PF1 (plot Ep), litterfallwas sampled every week in 15 quadrats of 0.56 m2.In the other plots, litterfall was sampled in 10quadrats of 0.25 m2, and sampling was made everyweek during the rainy season and every other weekduring the dry season. The quadrats were coveredwith 1-2 mm mesh plastic screen. Samples wereoven-dried (65-75°C) and weighed. Threefractions were sorted: leaves, twigs and bark, fruitsand flowers. Samples from each four-week periodwere combined for nutrient analysis at the IRDAnalysis Laboratory of Pointe-Noire.Unfortunately, except in 6-year-old plot Ep, thispattern was disturbed because the time lag betweensampling, was occasionally longer and somequadrats, screens were sometimes stolen;consequently it was not possible to calculate theannual litterfall per quadrat for statistical purposes.Statistical analysis was made on the basis of

24 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
litterfall per week, and comparisons took intoaccount the overall results for a sampling time.Annual litterfall was calculated by multiplying themean weekly litterfall for each plot by 52.
Standing litter was sampled with a 6 cmdiameter cylinder, and three cores were taken foreach sample. Three replications were made in eachplot. This rapid method gave good reproducibilityand therefore allowed reliable comparisonsbetween stands, but over-estimated byapproximately 25% the results when comparedwith the usual single sampling of larger areas.
Characterisation of leaf litterFor the organic characterisation of litter, freshlyfallen leaves, recognised by their colour, werepicked up from the soil, and air-dried. Samplingwas made in February 1998 and 1999. Thethickness of the leaf blade was measured. It wasmeasured between the main veins with a thicknessgauge (by 0.01 mm), after re-humidification oflitter leaves in boiling water. Two measurementswere made on each of ten leaves for each clone.
Air-dried and milled litter were chemicallyanalysed and chemical contents were expressedas oven-dried weight (75°C). Fibres were analysedby the method of Van Soest (1963) bydetermination of neutral detergent fibre (NDF),acid detergent fibre (ADF) and acid detergentlignin (ADL) at the CIRAD-AMIS Laboratory.With this proximate method, a variety ofcompounds are included in the so-called“hemicellulose”, “cellulose” and “lignin” fractionsbut it is generally used for comparisons in plantmaterial. Some samples were analysed for nitrogenon 100 mg samples using a LECO FP 428 CHNapparatus in the same laboratory, and in othersamples nitrogen was determined as NH
4 by the
Nessler method after mineralisation inconcentrated sulphuric acid with H
2O
2. Soluble C
and soluble phenolics were extracted by mixing 1or 2 g of litter in 60 ml of cold water during twohours. Total extractable phenolic compounds wereextracted according to the Tropical Soil Biologyand Fertility Programme method (Anderson andIngram 1993) by heating 0.5 g of litter in 50%(v:v) methanol in water at 80°C for one hour. Thedifference between methanol extracted phenolicsand water extracted phenolics was referred to as
“insoluble phenolics”, and the phenoliccompounds which were not extracted by methanolwere not taken into account in this study. Solublecarbon was determined in water extracts by thechemical oxygen demand (COD) with the rapidHACH method (Anonymous 1994) and forconversion of COD to carbon, the assumption waschecked previously that soluble carbon consistedmainly of glucids (Reversat 1981). Phenoliccompounds were determined in water extracts andin methanol extracts by the tyrosine reagent(HACH method, Anonymous 1994) which takesinto account all hydroxyled aromatic compounds.
Iron mobilisation abilityLitter solutions were made with 2 g in 60 mldistilled water and 2 hours shaking. To performiron extraction, 30 ml of litter extracts were shakenfor one hour with 5 g of a standard eucalypt soil(0-5 cm depth) and centrifuged. Iron wasdetermined colorimetrically in the solution by thephenanthroline reactant after organic matter wasmineralised with H2O2. Control extractions weremade with distilled water.
In vitro and in situ decomposition ratesIn vitro litter decomposition was studied by carbonmineralisation measurements of air-dried andmilled fresh litter. Micro-litterbags (50 x 50 mm)were made with 60 mm pore size nylon mesh. Theywere filled with 1 g of litter, which was humidified,with 2 ml of soil solution (20 g of savanna soilmixed with 50 ml of water for 30 minutes anddecanted). Each bag was put in a jar with a beakercontaining 50 ml of 0.1 N NaOH, which trappedthe CO
2. NaOH determinations were made at
intervals with 0.5 N HCl.In situ decomposition rates were measured in
20 cm x 20 cm litter bags, 1.5-2 mm mesh sizeeach, with 10 g of air dried leaf litter, the ovendried weight being measured in another part ofthe sample. Litter bags were put on the soil surfacein the same plot from which the litter came, inMarch, July and October, and were sampled after4 and 12 weeks in the field with 12 replications ateach date (in some cases, a few replications werelost). The decomposed litter was dried at 75°C andweighed.

25Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Results and Discussion
LitterfallAnnual litterfall ranged from 6.9 t ha-1 in theyounger plot to 12.8 t ha-1 in the old first rotationplot (Table 3.1). Total litterfall and leaf litterfallwere significantly different among plots, unlikewood litterfall. Fruits fell only in the 19-year-oldfirst rotation plot. In the young 6-year-old firstrotation plot the result was close to the 6.5 t ha-1
which was found previously in another PF1 plot(clone 1-45) (Bernhard-Reversat 1993). It washigher than in dry tropical or Mediterraneaneucalypt plantations where litterfall was 2-3 t ha-1
yr-1 (Knokaert 1981 in Morocco; Bernhard-Reversat 1987 in Senegal; Abelho and Graça 1996in Portugal). In temperate natural eucalypt forestslitterfall was of the same order of magnitude asfound in Congo at the same age (Polglase andAttiwill 1992). In Puerto Rico, Parrotta (1999)found an annual average of 5.4 t ha-1 in youngertropical plantations. When compared to nitrogenfixing plantations, eucalypt had generally a lowerlitterfall (Parrotta 1999; Bernhard-Reversat andLoumeto, in press).
Litterfall increased with aging of plantations,and was also influenced by harvest operations andbetween hybrids. Although the trees wereapproximately of the same age, leaf litterproduction was significantly lower in the young6-year-old first rotation plot than in the older 13-year-old coppice plot (about 6 years of coppicegrowth). Previous observations (Bernhard-Reversat 1993) reported that the litterfall of 2-year-old E. PF1 coppice plot was the same as that ofthe former 7-year-old first rotation crop in the sameplot. This suggested a rapid growth of the canopyin the coppice, allowed by the expanded rooting
system. The lower litter production in the 19-year-old coppice compared to the 13-year-old coppice,and to the 19-year-old unlogged first rotation crop,might suggest the physiological aging of the stock,the depletion of soil nutrient by export (Stewardet al. 1988, Miranda et al. 1998) or the alterationof the roots by parasitic nematodes (Loubana andReversat, chapter 6), together with the possibledecrease of nutrient availability.
Eucalyptus urograndis had a significantlyhigher leaf litterfall (6.8 t ha-1 yr-1) than the youngfirst rotation plot of E. PF1 (4.3 t ha-1 yr-1 ). Theage of the trees differed by two years and mightexplain the difference in litterfall, although a linearregression with the 6-year-old and the 19-year-oldfirst rotation plot litterfall gave 5.2 t ha-1 for a 8-year-old E. PF1 plot. It was shown previously thatthe leaf biomass of four E. urograndis clones wasabout 30% higher than the leaf biomass of E. PF1,clone 1-41 (Safou-Matondo et al. 1999). Duringthe study of an age series of E. PF1 (clone 1-41)by Laclau (unpublished data), a very low amountof litterfall was collected during the first year afterplanting (0.4 t ha-1 yr-1) and was only leaves.Between the second year and the logging age, leaflitterfall remained stable (about 4 t ha-1 yr-1) whilethe amount of wood and bark litterfall increasedup to the fourth year after planting. Parrotta (1999)also observed a relatively steady litterfall from thesecond to the fourth year of plantation.
Nutrients in litterfallSimilar values of leaf litter nutrient concentrationwere observed for the different plots and neitherplot age nor previous exploitation appeared to havean effect on nutrient contents (Fig. 3.1). Eucalyptusurograndis leaf litter was not different from E. PF1except for a higher P concentration. Laclau et al.
Table 3.1 Annual litterfall in eucalypt plots (g m-2 yr-1 with standard error in brackets)
Plot Stand type Eucalypt Plot age Leaves Twigs and Fruits Totalhybrid bark
(yr) (g m-2 yr-1)
Ep Seedling PF 1 6 431 (75) 256 (80) 0 688 (107)Er Coppice PF 1 13 831 (27) 256 (48) 0 1087 (54)Et Coppice PF 1 19 664 (28) 271 (37) 0 938 (46)Eu Coppice PF 1 19 888 (44) 378 (57) 25 (3) 1290 (76)Ev Seedling urograndis 8 684 (27) 320 (72) 0 1004 (77)
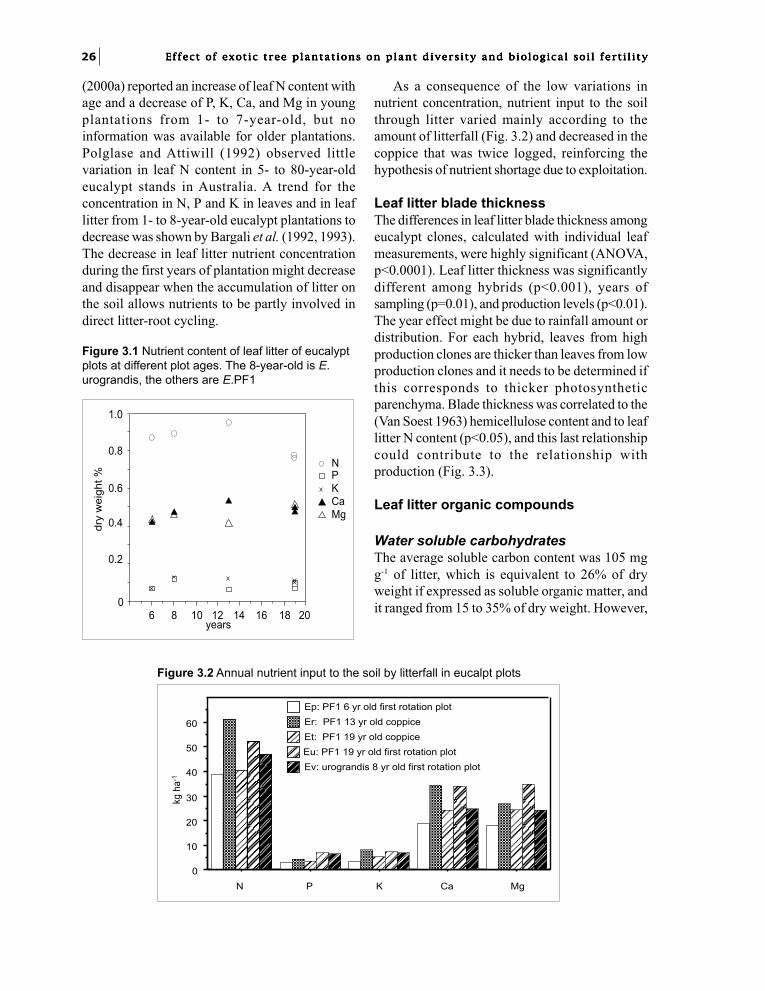
26 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
years
MgCaKPN
0
0.2
0.4
0.6
0.8
1.0
6 8 10 12 14 16 18 20
dry
we
igh
t %
N P K Ca Mg
0
Ev: urograndis 8 yr old first rotation plot
Eu: PF1 19 yr old first rotation plot
Et: PF1 19 yr old coppice
Er: PF1 13 yr old coppice
Ep: PF1 6 yr old first rotation plot
10
20
30
40
50
60
kg h
a-1
(2000a) reported an increase of leaf N content withage and a decrease of P, K, Ca, and Mg in youngplantations from 1- to 7-year-old, but noinformation was available for older plantations.Polglase and Attiwill (1992) observed littlevariation in leaf N content in 5- to 80-year-oldeucalypt stands in Australia. A trend for theconcentration in N, P and K in leaves and in leaflitter from 1- to 8-year-old eucalypt plantations todecrease was shown by Bargali et al. (1992, 1993).The decrease in leaf litter nutrient concentrationduring the first years of plantation might decreaseand disappear when the accumulation of litter onthe soil allows nutrients to be partly involved indirect litter-root cycling.
As a consequence of the low variations innutrient concentration, nutrient input to the soilthrough litter varied mainly according to theamount of litterfall (Fig. 3.2) and decreased in thecoppice that was twice logged, reinforcing thehypothesis of nutrient shortage due to exploitation.
Leaf litter blade thicknessThe differences in leaf litter blade thickness amongeucalypt clones, calculated with individual leafmeasurements, were highly significant (ANOVA,p<0.0001). Leaf litter thickness was significantlydifferent among hybrids (p<0.001), years ofsampling (p=0.01), and production levels (p<0.01).The year effect might be due to rainfall amount ordistribution. For each hybrid, leaves from highproduction clones are thicker than leaves from lowproduction clones and it needs to be determined ifthis corresponds to thicker photosyntheticparenchyma. Blade thickness was correlated to the(Van Soest 1963) hemicellulose content and to leaflitter N content (p<0.05), and this last relationshipcould contribute to the relationship withproduction (Fig. 3.3).
Leaf litter organic compounds
Water soluble carbohydratesThe average soluble carbon content was 105 mgg-1 of litter, which is equivalent to 26% of dryweight if expressed as soluble organic matter, andit ranged from 15 to 35% of dry weight. However,
Figure 3.1 Nutrient content of leaf litter of eucalyptplots at different plot ages. The 8-year-old is E.urograndis, the others are E.PF1
Figure 3.2 Annual nutrient input to the soil by litterfall in eucalpt plots
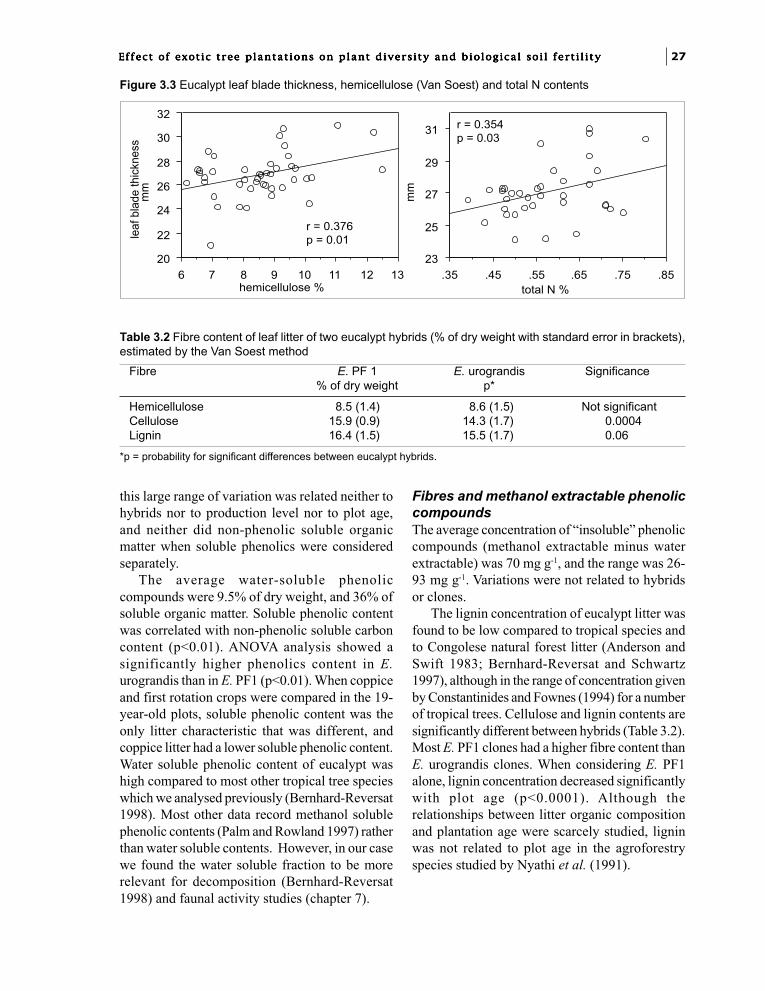
27Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
total N %hemicellulose %
23
25
27
29
31
.35 .45 .55 .65 .75 .85
r = 0.354p = 0.03
20
22
24
26
28
30
32
6 7 8 9 10 11 12 13
r = 0.376p = 0.01
mm
mm
leaf
bla
de th
ickn
ess
Table 3.2 Fibre content of leaf litter of two eucalypt hybrids (% of dry weight with standard error in brackets),estimated by the Van Soest method
Fibre E. PF 1 E. urograndis Significance% of dry weight p*
Hemicellulose 8.5 (1.4) 8.6 (1.5) Not significantCellulose 15.9 (0.9) 14.3 (1.7) 0.0004Lignin 16.4 (1.5) 15.5 (1.7) 0.06
*p = probability for significant differences between eucalypt hybrids.
this large range of variation was related neither tohybrids nor to production level nor to plot age,and neither did non-phenolic soluble organicmatter when soluble phenolics were consideredseparately.
The average water-soluble phenoliccompounds were 9.5% of dry weight, and 36% ofsoluble organic matter. Soluble phenolic contentwas correlated with non-phenolic soluble carboncontent (p<0.01). ANOVA analysis showed asignificantly higher phenolics content in E.urograndis than in E. PF1 (p<0.01). When coppiceand first rotation crops were compared in the 19-year-old plots, soluble phenolic content was theonly litter characteristic that was different, andcoppice litter had a lower soluble phenolic content.Water soluble phenolic content of eucalypt washigh compared to most other tropical tree specieswhich we analysed previously (Bernhard-Reversat1998). Most other data record methanol solublephenolic contents (Palm and Rowland 1997) ratherthan water soluble contents. However, in our casewe found the water soluble fraction to be morerelevant for decomposition (Bernhard-Reversat1998) and faunal activity studies (chapter 7).
Fibres and methanol extractable phenoliccompoundsThe average concentration of “insoluble” phenoliccompounds (methanol extractable minus waterextractable) was 70 mg g-1, and the range was 26-93 mg g-1. Variations were not related to hybridsor clones.
The lignin concentration of eucalypt litter wasfound to be low compared to tropical species andto Congolese natural forest litter (Anderson andSwift 1983; Bernhard-Reversat and Schwartz1997), although in the range of concentration givenby Constantinides and Fownes (1994) for a numberof tropical trees. Cellulose and lignin contents aresignificantly different between hybrids (Table 3.2).Most E. PF1 clones had a higher fibre content thanE. urograndis clones. When considering E. PF1alone, lignin concentration decreased significantlywith plot age (p<0.0001). Although therelationships between litter organic compositionand plantation age were scarcely studied, ligninwas not related to plot age in the agroforestryspecies studied by Nyathi et al. (1991).
Figure 3.3 Eucalypt leaf blade thickness, hemicellulose (Van Soest) and total N contents
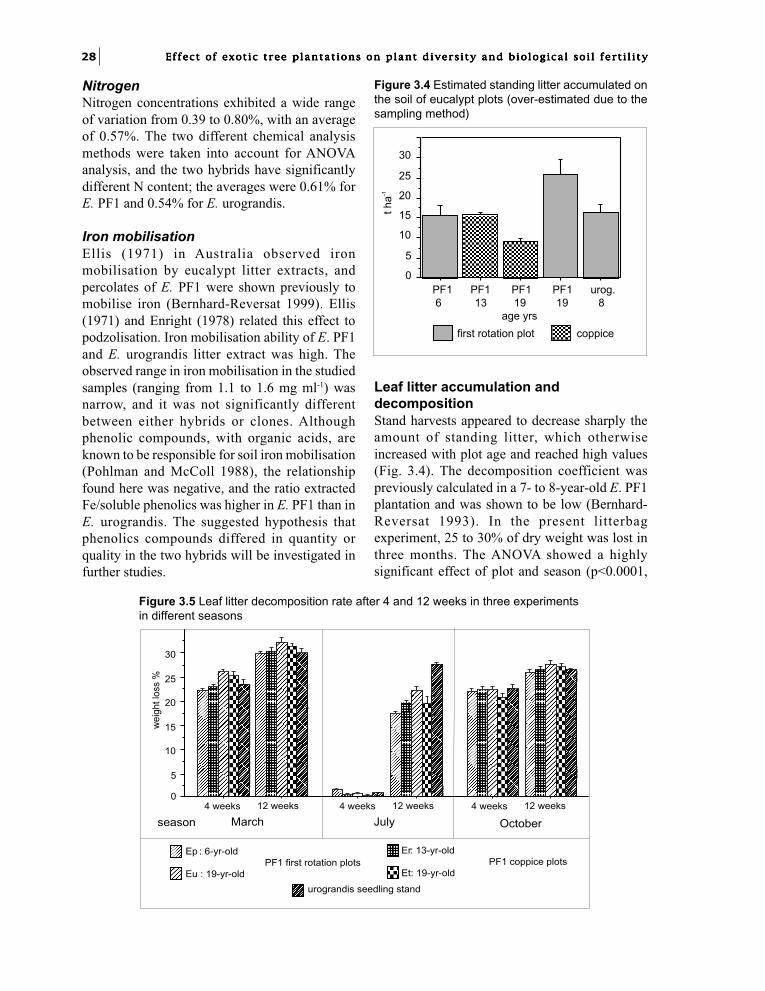
28 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Ep : 6-yr-oldPF1 first rotation plots
Eu : 19-yr-old
urograndis seedling stand
Er: 13-yr-oldPF1 coppice plots
Et: 19-yr-old
0
5
10
15
20
25
30
season March July October
wei
ght l
oss
%
4 weeks 12 weeks 4 weeks 12 weeks 4 weeks 12 weeks
0
5
10
15
20
25
30
PF1 PF1 PF1 PF1 urog.6 13 19 19 8
age yrs
first rotation plot coppice
t ha
-1
NitrogenNitrogen concentrations exhibited a wide rangeof variation from 0.39 to 0.80%, with an averageof 0.57%. The two different chemical analysismethods were taken into account for ANOVAanalysis, and the two hybrids have significantlydifferent N content; the averages were 0.61% forE. PF1 and 0.54% for E. urograndis.
Iron mobilisationEllis (1971) in Australia observed ironmobilisation by eucalypt litter extracts, andpercolates of E. PF1 were shown previously tomobilise iron (Bernhard-Reversat 1999). Ellis(1971) and Enright (1978) related this effect topodzolisation. Iron mobilisation ability of E. PF1and E. urograndis litter extract was high. Theobserved range in iron mobilisation in the studiedsamples (ranging from 1.1 to 1.6 mg ml-1) wasnarrow, and it was not significantly differentbetween either hybrids or clones. Althoughphenolic compounds, with organic acids, areknown to be responsible for soil iron mobilisation(Pohlman and McColl 1988), the relationshipfound here was negative, and the ratio extractedFe/soluble phenolics was higher in E. PF1 than inE. urograndis. The suggested hypothesis thatphenolics compounds differed in quantity orquality in the two hybrids will be investigated infurther studies.
Leaf litter accumulation anddecompositionStand harvests appeared to decrease sharply theamount of standing litter, which otherwiseincreased with plot age and reached high values(Fig. 3.4). The decomposition coefficient waspreviously calculated in a 7- to 8-year-old E. PF1plantation and was shown to be low (Bernhard-Reversat 1993). In the present litterbagexperiment, 25 to 30% of dry weight was lost inthree months. The ANOVA showed a highlysignificant effect of plot and season (p<0.0001,
Figure 3.4 Estimated standing litter accumulated onthe soil of eucalypt plots (over-estimated due to thesampling method)
Figure 3.5 Leaf litter decomposition rate after 4 and 12 weeks in three experimentsin different seasons

29Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Fig. 3.5). The plot effect was due to a slightincrease of decomposition rate with plot age anda higher (p<0.05) one in the first rotation plot thanin coppice when compared in the 19-year-oldplots, suggesting an effect of litter quality. Theslow decomposition in tropical plantationscompared to natural forest, especially in eucalyptand pine plantations, has been observed bynumerous authors and stressed in some reviews(O’Connell and Sankaran 1997; Bernhard-Reversat and Loumeto 2001). The decrease inlignin content with age could account forincreasing decomposition rate. Such an increasein decomposition rate with plot age was observedfor subtropical tree fine roots by Arunachalam etal. (1996) together with lignin content decrease.
The difference between hybrids changedaccording to seasons; during the rainy season(experiments beginning in March, October orJanuary), decomposition rates were only slightlyhigher in E. urograndis compared to E. PF1 (8-and 6-year-old respectively); during the dry seasonthe litter of E. urograndis decomposedsignificantly faster than the litter of PF1.Differences in litter fauna activity might explainthis observation but fauna observations(Mboukou-Kimbatsa et al. 1998) were not madeduring the dry season.
ConclusionsThe litter system is highly dependent on themanagement factors: the results suggest that plotage, exploitation, and hybrid have an effect onlitter quality and decomposition rate. Althoughthese effects are small as measured values, theyare significant and might influence nutrient cyclingand soil organic matter accumulation and couldalso control microbiological and faunal activity.The aging of plots appeared to improve thefunctioning of the litter system. Loggingoperations create major disturbance of litterdecomposition dynamics.

Soil Fertility Changes with Eucalypt Hybridsand Plantation Age: Soil Organic MatterJ.J. Loumeto1 and F. Bernhard-Reversat2
1 Laboratoire de Biologie Végétale, Université deBrazzaville, BP 69, Brazzaville, Congo.2 Laboratoire d’Ecologie des Sols Tropicaux, Centre IRDd’Ile de France, 32 avenue Henri Varagnat, 93143 Bondy,France.
Chapter 4.
Introduction
S oil organic matter in the plantationsoriginates from eucalypt litter and from theprevious savanna vegetation (Trouvé et al.
1994). The amount of organic input in savanna islow and originates mainly from dead roots due toannual burning. Soils planted with eucalypts havea low organic matter content together with lowclay and nutrient contents. Increasing organicmatter improves the nutrient status of soil, and isalso assumed to improve biological activity andsoil structure. Therefore any research andknowledge improvement in this area is oftremendous importance for nutrient conservationin the plantations, especially when grown on poorsoils.
It was shown previously that soil organic mattercontent was the same in savanna soil and in a 7-year-old eucalypt plantation, and that it increasedlater, mainly due to light fraction increase(Bernhard-Reversat 1991). Observations carriedout in other situations have either confirmed theabove trend or shown different patterns of change.Trouvé et al. (1994) and Binkley and Resh (1999)showed that under eucalypt plantations, organicmatter originating from the native savannadecreased whereas organic matter from eucalyptincreased; therefore total C content did not changefor several years. Parrotta (1999) also observedthat no significant change in soil C occurred in7.5-year-old eucalypt plantations established on agrassland soil in Puerto Rico. Jaiyeoba (1998)observed that soil organic matter decreased in
eucalypt plantations grown on savanna soil duringthe first years and increased at a later age. Otherobservations showed organic matter decrease ineucalypt plantations grown on C-poor soils(Bernhard-Reversat 1988; Nandi et al. 1991; Kingand Campbell 1994). In the arboretum studied byLugo et al. (1990) in a rain forest area whereseveral planted species were compared, soilorganic matter content either increased ordecreased according to the species, and organicmatter contents in eucalypt plots were among thehighest. The aim of the present study was toinvestigate changes in soil fertility with a selectedclone, and to assess the effect of plot age,exploitation and hybrids on soil organic matter,nitrogen and cation exchange capacity.
Studied Plots and MethodsThe study was carried out on seven planted plotseither harvested or not (Ep, Eq, Er ,Es, Et, Eu, Ev)and on savanna (Sb) (Table 0.3). Six compositesamples were made in two layers (0-10 cm and10-20 cm) in each plot. Each composite samplecomprised four cores were which were extracted,close to each other, with a 62 mm diameter tube
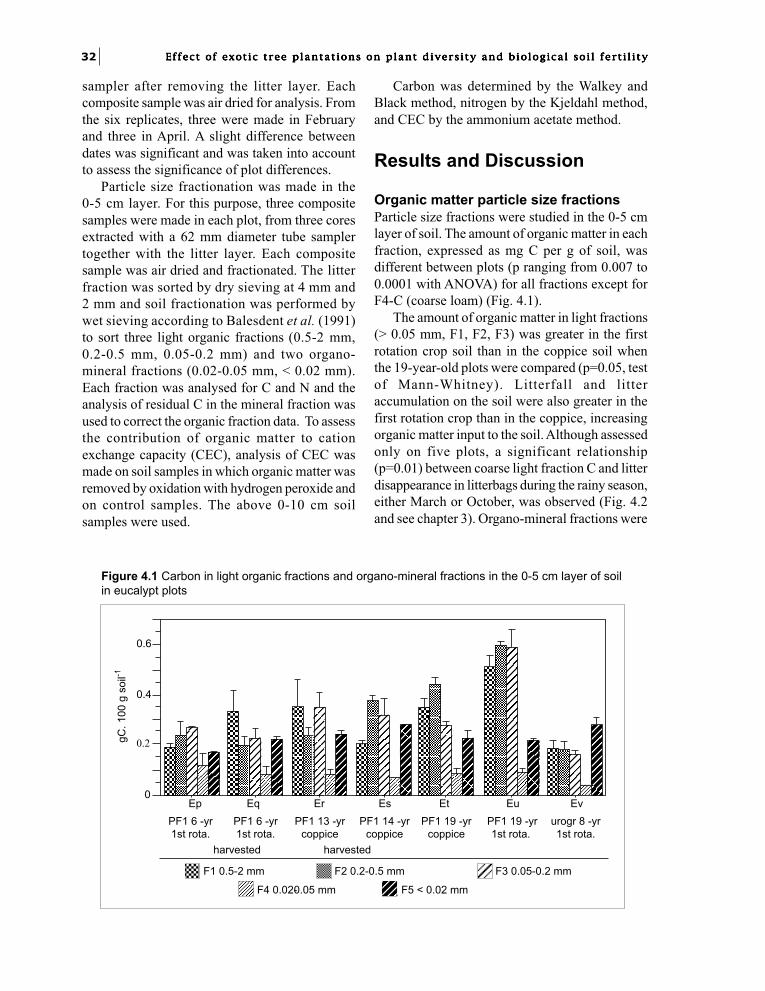
32 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
sampler after removing the litter layer. Eachcomposite sample was air dried for analysis. Fromthe six replicates, three were made in Februaryand three in April. A slight difference betweendates was significant and was taken into accountto assess the significance of plot differences.
Particle size fractionation was made in the0-5 cm layer. For this purpose, three compositesamples were made in each plot, from three coresextracted with a 62 mm diameter tube samplertogether with the litter layer. Each compositesample was air dried and fractionated. The litterfraction was sorted by dry sieving at 4 mm and2 mm and soil fractionation was performed bywet sieving according to Balesdent et al. (1991)to sort three light organic fractions (0.5-2 mm,0.2-0.5 mm, 0.05-0.2 mm) and two organo-mineral fractions (0.02-0.05 mm, < 0.02 mm).Each fraction was analysed for C and N and theanalysis of residual C in the mineral fraction wasused to correct the organic fraction data. To assessthe contribution of organic matter to cationexchange capacity (CEC), analysis of CEC wasmade on soil samples in which organic matter wasremoved by oxidation with hydrogen peroxide andon control samples. The above 0-10 cm soilsamples were used.
Carbon was determined by the Walkey andBlack method, nitrogen by the Kjeldahl method,and CEC by the ammonium acetate method.
Results and Discussion
Organic matter particle size fractionsParticle size fractions were studied in the 0-5 cmlayer of soil. The amount of organic matter in eachfraction, expressed as mg C per g of soil, wasdifferent between plots (p ranging from 0.007 to0.0001 with ANOVA) for all fractions except forF4-C (coarse loam) (Fig. 4.1).
The amount of organic matter in light fractions(> 0.05 mm, F1, F2, F3) was greater in the firstrotation crop soil than in the coppice soil whenthe 19-year-old plots were compared (p=0.05, testof Mann-Whitney). Litterfall and litteraccumulation on the soil were also greater in thefirst rotation crop than in the coppice, increasingorganic matter input to the soil. Although assessedonly on five plots, a significant relationship(p=0.01) between coarse light fraction C and litterdisappearance in litterbags during the rainy season,either March or October, was observed (Fig. 4.2and see chapter 3). Organo-mineral fractions were
F5 < 0.02 mm F4 0.02-0.05 mm
F3 0.05-0.2 mm F2 0.2-0.5 mm F1 0.5-2 mm
Ep Eq Er Es Et Eu Ev
PF1 6 -yr1st rota.
PF1 6 -yr1st rota.
PF1 13 -yrcoppice
PF1 14 -yrcoppice
PF1 19 -yrcoppice
PF1 19 -yr1st rota.
urogr 8 -yr1st rota.
harvested harvested
gC. 1
00 g
soi
l-1
0
0.2
0.4
0.6
Figure 4.1 Carbon in light organic fractions and organo-mineral fractions in the 0-5 cm layer of soilin eucalypt plots
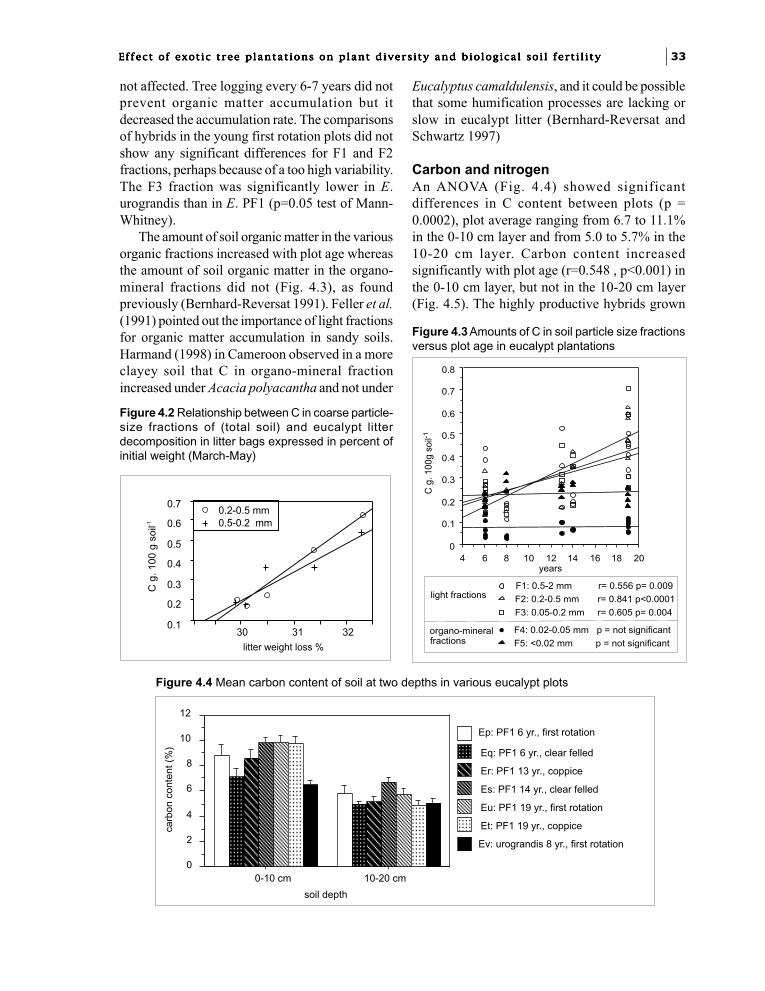
33Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
30 31 320.1
0.2
0.3
0.4
0.5
0.6
0.70.2-0.5 mm0.5-0.2 mm
C g
. 100 g
soil-1
litter weight loss %
Figure 4.2 Relationship between C in coarse particle-size fractions of (total soil) and eucalypt litterdecomposition in litter bags expressed in percent ofinitial weight (March-May)
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
4 6 8 10 12 14 16 18 20
F3: 0.05-0.2 mm r= 0.605 p= 0.004
F2: 0.2-0.5 mm r= 0.841 p<0.0001
F1: 0.5-2 mm r= 0.556 p= 0.009
F5: <0.02 mm p = not significant
F4: 0.02-0.05 mm p = not significant
light fractions
organo-mineralfractions
C g
. 100
g so
il-1
years
Figure 4.3 Amounts of C in soil particle size fractionsversus plot age in eucalypt plantations
0-10 cm 10-20 cm
Ev: urograndis 8 yr., first rotation
Et: PF1 19 yr., coppice
Eu: PF1 19 yr., first rotation
Es: PF1 14 yr., clear felled
Er: PF1 13 yr., coppice
Eq: PF1 6 yr., clear felled
Ep: PF1 6 yr., first rotation
0
2
4
6
8
10
12
carb
on c
onte
nt (
%)
soil depth
Figure 4.4 Mean carbon content of soil at two depths in various eucalypt plots
not affected. Tree logging every 6-7 years did notprevent organic matter accumulation but itdecreased the accumulation rate. The comparisonsof hybrids in the young first rotation plots did notshow any significant differences for F1 and F2fractions, perhaps because of a too high variability.The F3 fraction was significantly lower in E.urograndis than in E. PF1 (p=0.05 test of Mann-Whitney).
The amount of soil organic matter in the variousorganic fractions increased with plot age whereasthe amount of soil organic matter in the organo-mineral fractions did not (Fig. 4.3), as foundpreviously (Bernhard-Reversat 1991). Feller et al.(1991) pointed out the importance of light fractionsfor organic matter accumulation in sandy soils.Harmand (1998) in Cameroon observed in a moreclayey soil that C in organo-mineral fractionincreased under Acacia polyacantha and not under
Eucalyptus camaldulensis, and it could be possiblethat some humification processes are lacking orslow in eucalypt litter (Bernhard-Reversat andSchwartz 1997)
Carbon and nitrogenAn ANOVA (Fig. 4.4) showed significantdifferences in C content between plots (p =0.0002), plot average ranging from 6.7 to 11.1%in the 0-10 cm layer and from 5.0 to 5.7% in the10-20 cm layer. Carbon content increasedsignificantly with plot age (r=0.548 , p<0.001) inthe 0-10 cm layer, but not in the 10-20 cm layer(Fig. 4.5). The highly productive hybrids grown
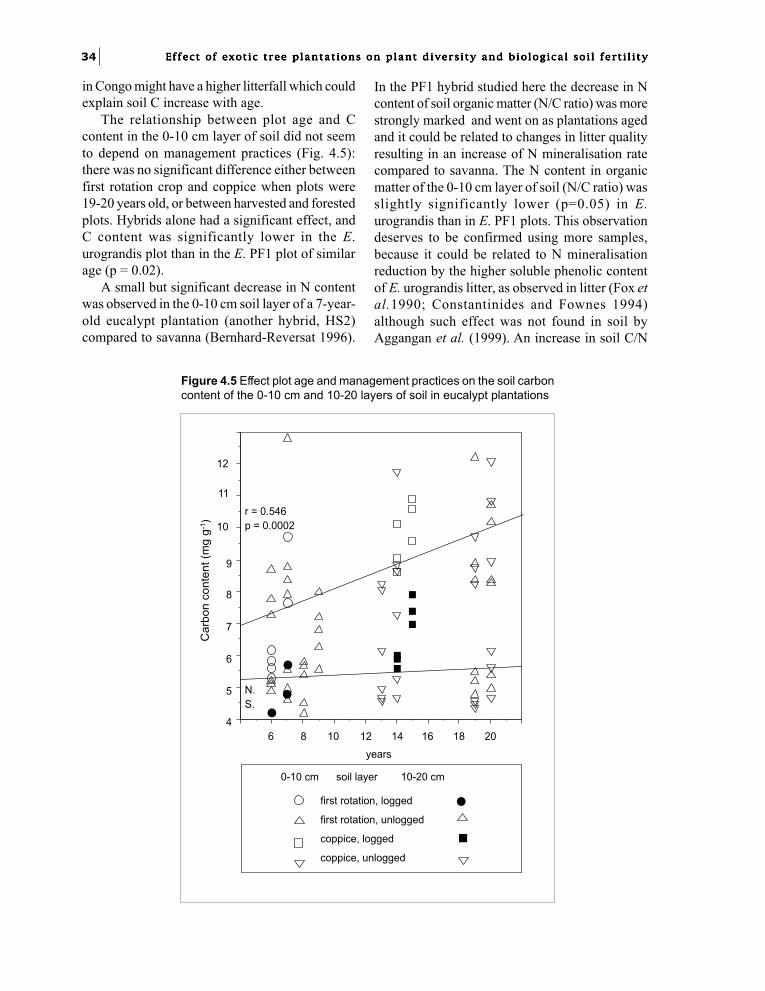
34 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
In the PF1 hybrid studied here the decrease in Ncontent of soil organic matter (N/C ratio) was morestrongly marked and went on as plantations agedand it could be related to changes in litter qualityresulting in an increase of N mineralisation ratecompared to savanna. The N content in organicmatter of the 0-10 cm layer of soil (N/C ratio) wasslightly significantly lower (p=0.05) in E.urograndis than in E. PF1 plots. This observationdeserves to be confirmed using more samples,because it could be related to N mineralisationreduction by the higher soluble phenolic contentof E. urograndis litter, as observed in litter (Fox etal.1990; Constantinides and Fownes 1994)although such effect was not found in soil byAggangan et al. (1999). An increase in soil C/N
in Congo might have a higher litterfall which couldexplain soil C increase with age.
The relationship between plot age and Ccontent in the 0-10 cm layer of soil did not seemto depend on management practices (Fig. 4.5):there was no significant difference either betweenfirst rotation crop and coppice when plots were19-20 years old, or between harvested and forestedplots. Hybrids alone had a significant effect, andC content was significantly lower in the E.urograndis plot than in the E. PF1 plot of similarage (p = 0.02).
A small but significant decrease in N contentwas observed in the 0-10 cm soil layer of a 7-year-old eucalypt plantation (another hybrid, HS2)compared to savanna (Bernhard-Reversat 1996).
first rotation, logged
first rotation, unlogged
coppice, logged
coppice, unlogged
0-10 cm soil layer 10-20 cm
years
10 12 14 16 184
5
6
7
8
9
10
11
12
6 8 20
r = 0.546p = 0.0002
N.S.
Ca
rbo
n c
on
ten
t (m
g g
-1)
Figure 4.5 Effect plot age and management practices on the soil carboncontent of the 0-10 cm and 10-20 layers of soil in eucalypt plantations
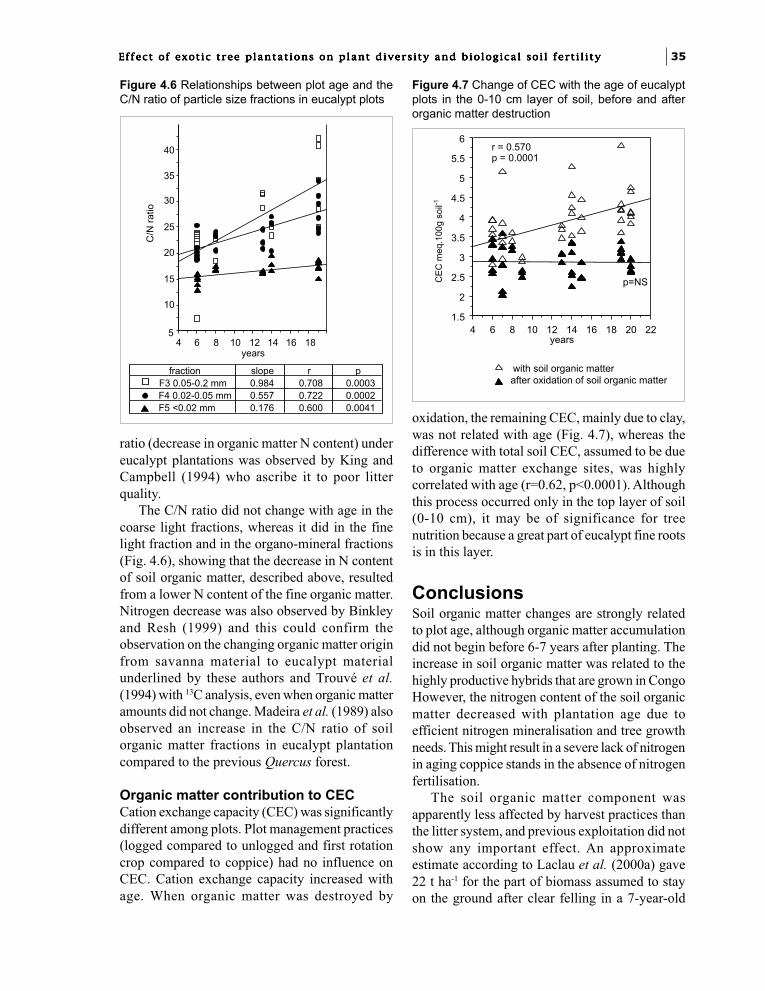
35Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
ratio (decrease in organic matter N content) undereucalypt plantations was observed by King andCampbell (1994) who ascribe it to poor litterquality.
The C/N ratio did not change with age in thecoarse light fractions, whereas it did in the finelight fraction and in the organo-mineral fractions(Fig. 4.6), showing that the decrease in N contentof soil organic matter, described above, resultedfrom a lower N content of the fine organic matter.Nitrogen decrease was also observed by Binkleyand Resh (1999) and this could confirm theobservation on the changing organic matter originfrom savanna material to eucalypt materialunderlined by these authors and Trouvé et al.(1994) with 13C analysis, even when organic matteramounts did not change. Madeira et al. (1989) alsoobserved an increase in the C/N ratio of soilorganic matter fractions in eucalypt plantationcompared to the previous Quercus forest.
Organic matter contribution to CECCation exchange capacity (CEC) was significantlydifferent among plots. Plot management practices(logged compared to unlogged and first rotationcrop compared to coppice) had no influence onCEC. Cation exchange capacity increased withage. When organic matter was destroyed by
oxidation, the remaining CEC, mainly due to clay,was not related with age (Fig. 4.7), whereas thedifference with total soil CEC, assumed to be dueto organic matter exchange sites, was highlycorrelated with age (r=0.62, p<0.0001). Althoughthis process occurred only in the top layer of soil(0-10 cm), it may be of significance for treenutrition because a great part of eucalypt fine rootsis in this layer.
ConclusionsSoil organic matter changes are strongly relatedto plot age, although organic matter accumulationdid not begin before 6-7 years after planting. Theincrease in soil organic matter was related to thehighly productive hybrids that are grown in CongoHowever, the nitrogen content of the soil organicmatter decreased with plantation age due toefficient nitrogen mineralisation and tree growthneeds. This might result in a severe lack of nitrogenin aging coppice stands in the absence of nitrogenfertilisation.
The soil organic matter component wasapparently less affected by harvest practices thanthe litter system, and previous exploitation did notshow any important effect. An approximateestimate according to Laclau et al. (2000a) gave22 t ha-1 for the part of biomass assumed to stayon the ground after clear felling in a 7-year-old
after oxidation of soil organic matterwith soil organic matter
years
r = 0.570p = 0.0001
p=NS
1.5
2
2.5
3
3.5
4
4.5
5
5.5
6
4 6 8 10 12 14 16 18 20 22
CE
C m
eq.1
00g
soil-1
Figure 4.7 Change of CEC with the age of eucalyptplots in the 0-10 cm layer of soil, before and afterorganic matter destruction
years
5
10
15
20
25
30
35
40
4 6 8 10 12 14 16 18
fraction slope r pF3 0.05-0.2 mm 0.984 0.708 0.0003F4 0.02-0.05 mm 0.557 0.722 0.0002F5 <0.02 mm 0.176 0.600 0.0041
C/N
ratio
Figure 4.6 Relationships between plot age and theC/N ratio of particle size fractions in eucalypt plots

36 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
plot. If each harvest brings to the soilapproximately this amount of residue, accountingfor the equivalent of 3 t yr-1, it could make up forthe annual difference of litterfall input betweenthe 19-year-old coppice and the first rotation crop.The branch wood which is left on the soil may beeasily humified because its lignin is lesspolymerised than that of the stem wood, and ithas a higher nutrient content (Lemieux 1996).Although eucalypts are among the less efficientspecies for improving soil nutrient status(O’Connell and Sakaran 1997) when grown on thepoor savanna soils of Congo, their litter contributesto soil fertility increase after 6 to 7 years ofplantations.

Biological factors of fertility relatedto the diversity and density of soil biota
Part 3

Asymbiotic Nitrogen Fixationin Savanna and Eucalypt PlantationsJ. Le Mer1 and P. Roger1
1 Laboratoire IRD de Microbiologie, Université deProvence, ESIL, Case 925, 163 Avenue de Luminy 13288,Marseille Cedex 9, France.
Chapter 5.
Introduction
The obvious importance of nitrogen (N)availability for eucalypt growth in anitrogen-poor environment required
determining if biological nitrogen fixation (BNF)was occurring. Microbial activity in eucalyptplantations was expected to be strongly reducedby the well known allelopathic effect of the litter(Souto et al. 1995; Bernhard-Reversat 1999).
Simple acetylene reducing activity (ARA)measurements provide a qualitative estimation ofthe actual soil N
2 fixing activity (Roger and Ladha,
1992). The addition of glucose is generally usedto estimate their N
2 fixing potential (expressed
when C substrates are not limiting). Indeed ARAin the presence of glucose is also an index of theabundance of free-living N2-fixing micro-organisms in the soil. O’Connell and Grove (1987)found some ARA in the litter of Australian nativeeucalypt forests. Although the amount of N
2 fixed
extrapolated from these measurements was low,BNF could be a relatively significant source ofnitrogen in the nitrogen-poor soil of the eucalyptplantations. Crews et al. (2000), also using ARAmeasurements, reported very low BNF valuesranging from 0.05 to 1.25 kg ha-1 year-1 in the leaflitter of Metrosideros polymorpha in Hawaii. Theaim of this study was to assess the impact of thedifferent methods of exploitation on the N fertilityof the surface of tropical soils planted witheucalypts in Congo. The ARA method (Hardy etal. 1968) was used.
Material and Methods
SamplesA first series of composite core samples (4 sub-samples) was collected in the first 5 cm of theprofile. Four composite samples were collected insavanna and in each of the age series of eucalyptplots (Ep, Er, Et and Eu). Litter samples wereseparated from the soil with forceps, except forthe savanna samples where no litter was present(Table 5.1).
A second series of 12 litter samples originatingfrom the 19-year-old first rotation crop and coppiceplots were used (Et and Eu). Litter alone wassampled because none of the previous soil samplesexhibited ARA.
ARA measurementsAcetylene reducing activity (Hardy et al. 1968)was estimated on the basis of the acetylene/ethylene pic area ratio, assuming no significantchange of the acetylene concentration in the flasks.This method of calculation is valid for aconsumption of less than 1% of the acetyleneintroduced (which was the case in allmeasurements performed) and has the advantageof being independent of the gas pressure within
40 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Table 5.1 First series of samples for ARA measurements
Plot Stand type Plot age Sample Times Number of(yr) harvested sample
Ep Seedling 6 Litter 0 1-4Ep Seedling 6 Soil 0 1-4Er Coppice 13 Litter 1 9-12Er Coppice 13 Soil 1 9-12Eu Seedling 19 Litter 0 13-16Eu Seedling 19 Soil 0 13-16Et Coppice 19 Litter 2 17-20Et Coppice 19 Soil 2 17-20
Savanna - - Soil - 5-8
the flask and from eventual leakages. Acetyleneand ethylene were measured in a FID Varianchromatograph.
Experimental protocols
First experimentSoil and litter incubations were performed in60 ml flasks at 30°C. Soils samples (15 g) andlitter samples (5 g) were brought to water retentioncapacity by the addition of 1.5 ml and 5.8 ml water,respectively. Samples were incubated for one dayin open flasks. Then the flasks were closed with arubber stopper and 10 ml acetylene was injectedwith a syringe in each flask. As ARAmeasurements performed daily for five daysshowed no significant activity, flasks were opened,ventilated to replace the flask atmosphere, 2%glucose was added, then the flasks were closedand 10 ml acetylene was again injected. ARAmeasurements were performed daily for four daysand then after 15 days of incubation.
Second experimentA second series of measurements was performedon all litter samples, which were first incubated inopen flasks for five days, then enriched with 2%glucose before performing daily ARAmeasurement for four days
Third experimentA second series of 12 litter samples originatingfrom the 19-year-old first rotation crop and coppiceplots was tested. Results showed no ARA both inthe absence and in the presence of 2% glucose.
In order to check the potential inhibitory effectof the eucalypt litter on ARA and N
2-fixing micro-
organisms we conducted an experiment with atemperate calcareous garden soil having receivedno agro-chemicals. The soil was treated aspreviously indicated and received the fourcombinations of two treatments : 2% glucose : +and - ; eucalypt litter extract : + and -. The eucalyptlitter extract was prepared by blending 20 g of litter(second set of litter samples) in 20 ml water,magnetic stirring for 1 hour, followed by filtration.The extract was used straight after its preparation.
ResultsIn the first experiment (first set of samples),samples tested daily for five days with no glucoseadded showed no ARA. Six of the 36 sampleswhere glucose was subsequently added, exhibitedARA. All were litter samples. Only data dealingwith samples exhibiting significant ARA arepresented in Table 5.2.
In the second experiment (first set of samples),only four litter samples collected in two of the plotswhich had significant ARA in the first experiment,showed ARA ranging from 5 to 75 µ moles g-1 h-1.Samples exhibiting significant ARA are presentedin Table 5.3.
In the third experiment (second set of samples),none of the 12 litter samples exhibited ARA in theabsence or in the presence of 2% glucose. Samplesof temperate garden soil without glucose additionhad no ARA. Samples of temperate garden soilreceiving 2% glucose exhibited ARA valuesranging from 198 to 367 µ moles C
2H
4 g-1 h-1. The
addition of eucalypt litter extract resulted in asignificant inhibitory effect (33%). Only samplesof the temperate garden exhibiting significant ARAare presented in Table 5.4.
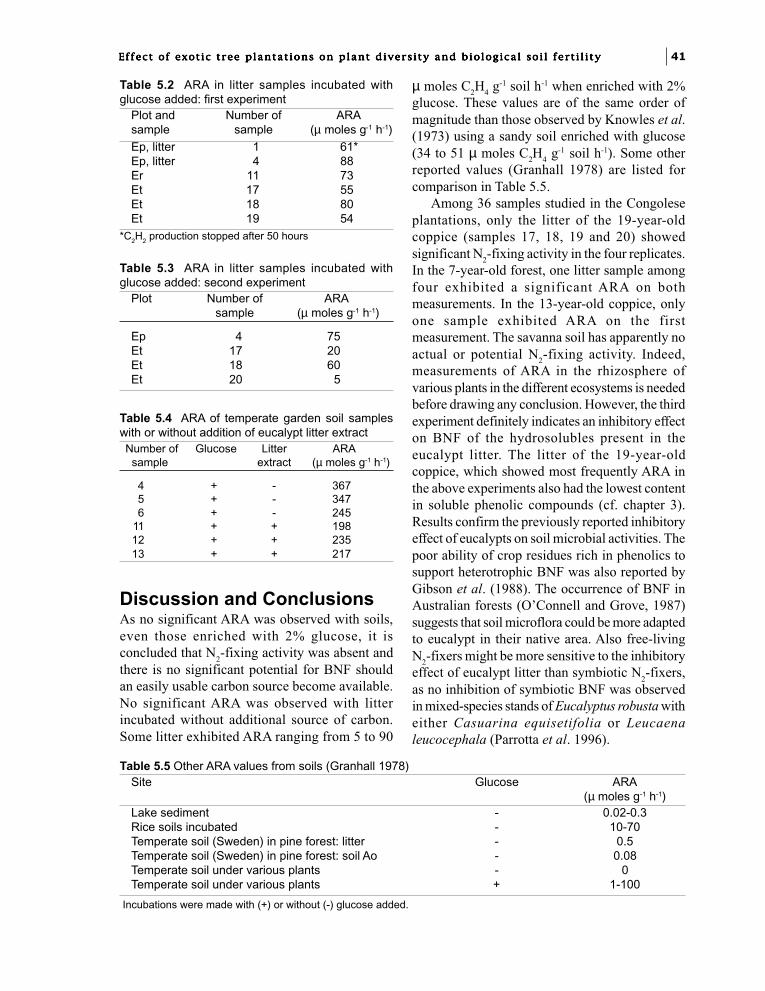
41Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Table 5.5 Other ARA values from soils (Granhall 1978)Site Glucose ARA
(µ moles g-1 h-1)Lake sediment - 0.02-0.3Rice soils incubated - 10-70Temperate soil (Sweden) in pine forest: litter - 0.5Temperate soil (Sweden) in pine forest: soil Ao - 0.08Temperate soil under various plants - 0Temperate soil under various plants + 1-100
Incubations were made with (+) or without (-) glucose added.
Table 5.2 ARA in litter samples incubated withglucose added: first experiment
Plot and Number of ARAsample sample (µ moles g-1 h-1)
Ep, litter 1 61*Ep, litter 4 88Er 11 73Et 17 55Et 18 80Et 19 54
*C2H
2 production stopped after 50 hours
Table 5.3 ARA in litter samples incubated withglucose added: second experiment
Plot Number of ARAsample (µ moles g-1 h-1)
Ep 4 75Et 17 20Et 18 60Et 20 5
Discussion and ConclusionsAs no significant ARA was observed with soils,even those enriched with 2% glucose, it isconcluded that N
2-fixing activity was absent and
there is no significant potential for BNF shouldan easily usable carbon source become available.No significant ARA was observed with litterincubated without additional source of carbon.Some litter exhibited ARA ranging from 5 to 90
µ moles C2H4 g-1 soil h-1 when enriched with 2%
glucose. These values are of the same order ofmagnitude than those observed by Knowles et al.(1973) using a sandy soil enriched with glucose(34 to 51 µ moles C2H4 g
-1 soil h-1). Some otherreported values (Granhall 1978) are listed forcomparison in Table 5.5.
Among 36 samples studied in the Congoleseplantations, only the litter of the 19-year-oldcoppice (samples 17, 18, 19 and 20) showedsignificant N2-fixing activity in the four replicates.In the 7-year-old forest, one litter sample amongfour exhibited a significant ARA on bothmeasurements. In the 13-year-old coppice, onlyone sample exhibited ARA on the firstmeasurement. The savanna soil has apparently noactual or potential N2-fixing activity. Indeed,measurements of ARA in the rhizosphere ofvarious plants in the different ecosystems is neededbefore drawing any conclusion. However, the thirdexperiment definitely indicates an inhibitory effecton BNF of the hydrosolubles present in theeucalypt litter. The litter of the 19-year-oldcoppice, which showed most frequently ARA inthe above experiments also had the lowest contentin soluble phenolic compounds (cf. chapter 3).Results confirm the previously reported inhibitoryeffect of eucalypts on soil microbial activities. Thepoor ability of crop residues rich in phenolics tosupport heterotrophic BNF was also reported byGibson et al. (1988). The occurrence of BNF inAustralian forests (O’Connell and Grove, 1987)suggests that soil microflora could be more adaptedto eucalypt in their native area. Also free-livingN2-fixers might be more sensitive to the inhibitoryeffect of eucalypt litter than symbiotic N
2-fixers,
as no inhibition of symbiotic BNF was observedin mixed-species stands of Eucalyptus robusta witheither Casuarina equisetifolia or Leucaenaleucocephala (Parrotta et al. 1996).
Table 5.4 ARA of temperate garden soil sampleswith or without addition of eucalypt litter extractNumber of Glucose Litter ARA sample extract (µ moles g-1 h-1)
4 + - 3675 + - 3476 + - 245
11 + + 19812 + + 23513 + + 217

Effect of Exotic Tree Plantationson Free Living and Plant ParasiticSoil Nematodes and Population Changeswith Eucalypt Hybrids and Plantation AgeP.M. Loubana1 and G. Reversat2
1 Laboratoire de Nématologie, Centre IRD de Pointe-Noire, B.P. 1286, Pointe-Noire, Congo.2 Laboratoire d’Ecologie des Sols Tropicaux, Centre d’Ilede France, 32 avenue Henri Varagnat, 93143 Bondy,France.
Chapter 6.
Introduction
The effect of changing environment onmicrofauna in plantations compared tosavanna was studied for free-living and
plant parasitic nematode populations. The first areof importance in the micro-food web, and althoughlittle is known about it, change in organic matterquality is assumed to affect their populations. Thepopulations of plant parasitic nematodes, whichare bound to one or several host plants, areexpected to be influenced by the new planted andundergrowth species and by the disappearance ofsavanna species. Thus if planted eucalypts werehost-plants for one of the native plant parasiticnematode species, population densities of thisparasite could increase and their possiblepathogenic effect on the crop should be considered.
Knowledge of nematodes associated with theeucalypts, rhizosphere in natural environments andin plantations is very limited, according to Majeret al. (1997), who mentioned nematodes as themost numerous animal class in the soil but did notquote any references on this topic. In eucalyptplantations in the Congo, a qualitative approachshowed two species of plant parasitic nematodesbelonging to the genus Xiphinema (Family:Longidoridae) as parasites of eucalypt roots: X.parasetariae and X. souchaudi (Baujard et al.1998).
The words “nematode” and “eucalypt” can befound linked very often in bibliographic data bases.This is related to the permanent quest fornematicidal properties of natural substances
among plants, including eucalypts, and the needto reduce the use of synthetic chemical nematicidesfor environmental considerations. However, suchstudies were not concerned with nematodeparasites of eucalypt roots.
The study had two aims: (i) to assess thechanges in species distribution and populationdensities of nematodes associated with changesin plant cover; (ii) to study a possible pathogeniceffect of these changes on the eucalypt crop.
Studied Plots and Methods
Field studiesThe study of nematodes was first carried out inthe age series of the Pointe-Noire savanna andexperimental plantations (Sa, Ea, Ed, Ee, Ef) andthe Loudima savanna and plantations (SL, El, AL,PL, Table 0.2). Three to five composite soilsamples were extracted along one or two transectsacross each plot. Composite samples resulted fromfive soil cores (0 to 15-20 cm depth) taken 15 cmaway from the trunk of every other plantedeucalypt tree. A second sampling series in thecommercial plantations (second project, Table 0.3)

44 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
aimed to study the distribution pattern ofnematodes in savanna (Sb), in E. PF1 plots (Ep,Er, Et, Eu) and in the 8-9-year-old multiclonal plotwhere E. urograndis was studied (Ev). Due to thetransect method, in this last plot, the sampledtransect crossed several clones and hybrids. Soilcores were made on tree lines at every other tree.This resulted in 10 to 20 samples on each sampledtree line, beginning on the plot edge. Samples weretaken 1 m away from the trunks, and cores were15 cm depth, with a soil volume of 424 ml.Nematodes were extracted from soil by the two-flask technique (Seinhorst 1955) and from rootsby the mistifier extraction technique (Seinhorst1950). All individuals of each species of plantparasitic nematodes were counted, and neithergenera nor species were determined for free-livingnematodes. Plant parasitic nematodes extractedfrom roots were very scarce, whereas free-livingnematodes extracted from roots were sometimesvery numerous. According to the short life-cycleof free-living nematodes (4-7 days), and the timerequired for an efficient recovery of plant parasiticnematodes from roots through the mistifier (2weeks), these free-living nematodes were assumedto multiply on the roots during mistifying.Therefore data on free-living nematodes extractedfrom roots were not included in the results. Thefrequency was calculated as the ratio of the numberof samples containing nematodes to the totalnumber of samples expressed as a percentage. Rootweight was measured for each sample.
Laboratory study of eucalypt clonesusceptibility to Xiphinema parasetariaeThe susceptibility of eucalypts to Xiphinemaparasetariae, the major plant parasitic nematodefound in the eucalypt plantations, was assessed inthe laboratory. Young cuttings were planted in PVCvessels (one cutting per vessel) with 500 ml ofheat-sterilised savanna soil (maintained for 10hours at 75°C). Each plant was inoculated with10 nematodes and the vessels were kept at 30°Cin a temperature-controlled box. Nematodes wereextracted from each vessel after three months. Thefinal population was obtained by addition of theXiphinema extracted from soil and from roots, andthe reproduction rate was calculated as the ratiofinal population/initial inoculum. The experimentcomprised four replications for each clone.
Results and Discussion
Two great groups of soil nematodes wereconsidered, free-living and plant parasitic, becauseof the different ways in which their populationscould change. The second group is linked to theoccurrence of host plants although these plants arenot always known. The mean number ofindividuals of the two groups of soil nematodeswas calculated (Table 6.1), and eventuallycompared to the vegetation characteristics.
Free living nematodesFree-living nematodes were more numerous insavannas than in plantations. In plantations asignificant correlation (r = 0.733 for 12 plots) wasobserved with the percentage of savanna plantspecies (as defined in chapter 1) in the vegetationwhich is an index of environment change. Itsuggested that the savanna populations ofnematodes were disappearing while those of theforest had not yet stabilised, and indicated thatthere were no differences due to the planted treespecies. Consequently, although an increase ofnematode number with increasing carbon contentis generally reported (Yeates 1979; Pradhan et al.1988), no significant relationship occurred in thestudied plantations. Taking into account the dataof the first project, a negative relationship betweenthe number of free-living nematodes and themacrofauna biomass (expressed as log, r = -0.915p = 0.1% Fig. 6.1), which is largely dominated byearthworms, could indicate an antagonistic effect.However this hypothesis deserves more studybecause this relationship could also result fromthe effect of litter or organic matter quality. Thedensity of free-living nematodes was lower in thesecond sampling series in commercial eucalyptplantations than observed in the experimentalplots, and in the commercial plantations, theirdensity was higher in the old E. PF1 plots than inthe younger plots (p=0.02). There was nosignificant difference between the first rotationcrop and coppice. The comparison between thesandy soil (Pointe-Noire) and the clay soil(Loudima), showed a trend towards a higherdensity of free-living nematodes in sandy soilunder savanna (Table 6.1), whereas this was lessobvious under eucalypt and acacia plantations
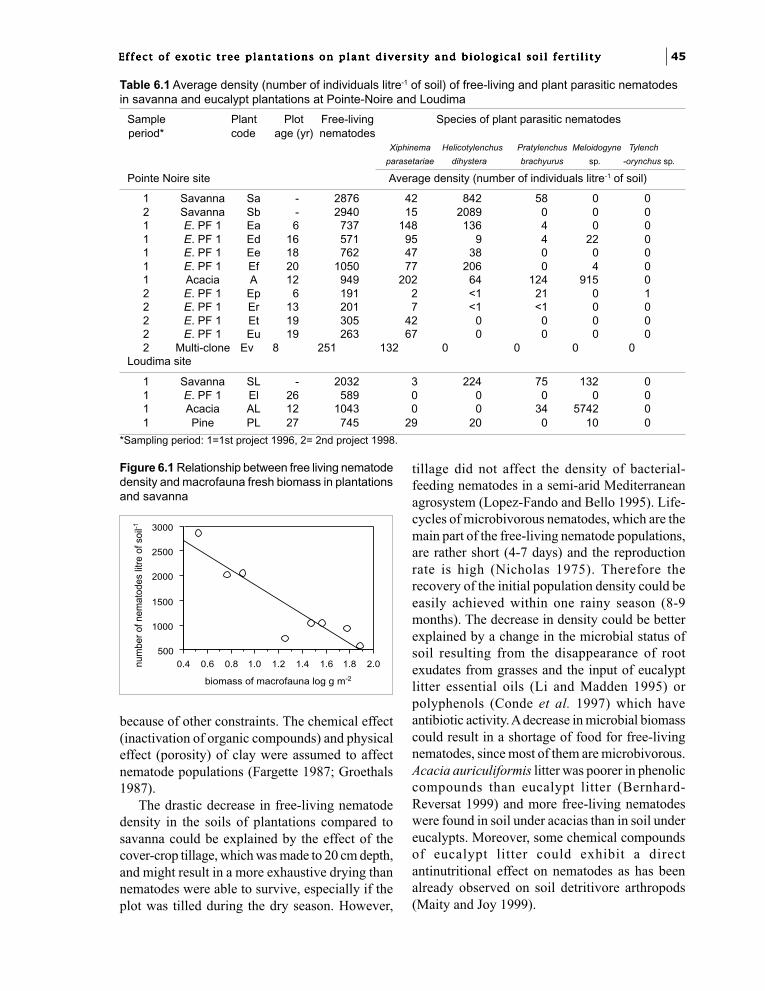
45Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Table 6.1 Average density (number of individuals litre-1 of soil) of free-living and plant parasitic nematodesin savanna and eucalypt plantations at Pointe-Noire and Loudima
Sample Plant Plot Free-living Species of plant parasitic nematodesperiod* code age (yr) nematodes
Xiphinema Helicotylenchus Pratylenchus Meloidogyne Tylench
parasetariae dihystera brachyurus sp. -orynchus sp.
Pointe Noire site Average density (number of individuals litre-1 of soil)
1 Savanna Sa - 2876 42 842 58 0 02 Savanna Sb - 2940 15 2089 0 0 01 E. PF 1 Ea 6 737 148 136 4 0 01 E. PF 1 Ed 16 571 95 9 4 22 01 E. PF 1 Ee 18 762 47 38 0 0 01 E. PF 1 Ef 20 1050 77 206 0 4 01 Acacia A 12 949 202 64 124 915 02 E. PF 1 Ep 6 191 2 <1 21 0 12 E. PF 1 Er 13 201 7 <1 <1 0 02 E. PF 1 Et 19 305 42 0 0 0 02 E. PF 1 Eu 19 263 67 0 0 0 02 Multi-clone Ev 8 251 132 0 0 0 0
Loudima site
1 Savanna SL - 2032 3 224 75 132 01 E. PF 1 El 26 589 0 0 0 0 01 Acacia AL 12 1043 0 0 34 5742 01 Pine PL 27 745 29 20 0 10 0
*Sampling period: 1=1st project 1996, 2= 2nd project 1998.
tillage did not affect the density of bacterial-feeding nematodes in a semi-arid Mediterraneanagrosystem (Lopez-Fando and Bello 1995). Life-cycles of microbivorous nematodes, which are themain part of the free-living nematode populations,are rather short (4-7 days) and the reproductionrate is high (Nicholas 1975). Therefore therecovery of the initial population density could beeasily achieved within one rainy season (8-9months). The decrease in density could be betterexplained by a change in the microbial status ofsoil resulting from the disappearance of rootexudates from grasses and the input of eucalyptlitter essential oils (Li and Madden 1995) orpolyphenols (Conde et al. 1997) which haveantibiotic activity. A decrease in microbial biomasscould result in a shortage of food for free-livingnematodes, since most of them are microbivorous.Acacia auriculiformis litter was poorer in phenoliccompounds than eucalypt litter (Bernhard-Reversat 1999) and more free-living nematodeswere found in soil under acacias than in soil undereucalypts. Moreover, some chemical compoundsof eucalypt litter could exhibit a directantinutritional effect on nematodes as has beenalready observed on soil detritivore arthropods(Maity and Joy 1999).
because of other constraints. The chemical effect(inactivation of organic compounds) and physicaleffect (porosity) of clay were assumed to affectnematode populations (Fargette 1987; Groethals1987).
The drastic decrease in free-living nematodedensity in the soils of plantations compared tosavanna could be explained by the effect of thecover-crop tillage, which was made to 20 cm depth,and might result in a more exhaustive drying thannematodes were able to survive, especially if theplot was tilled during the dry season. However,
biomass of macrofauna log g m-2
500
1000
1500
2000
2500
3000
0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0num
ber
of n
emat
odes
litr
e of
soi
l-1
Figure 6.1 Relationship between free living nematodedensity and macrofauna fresh biomass in plantationsand savanna
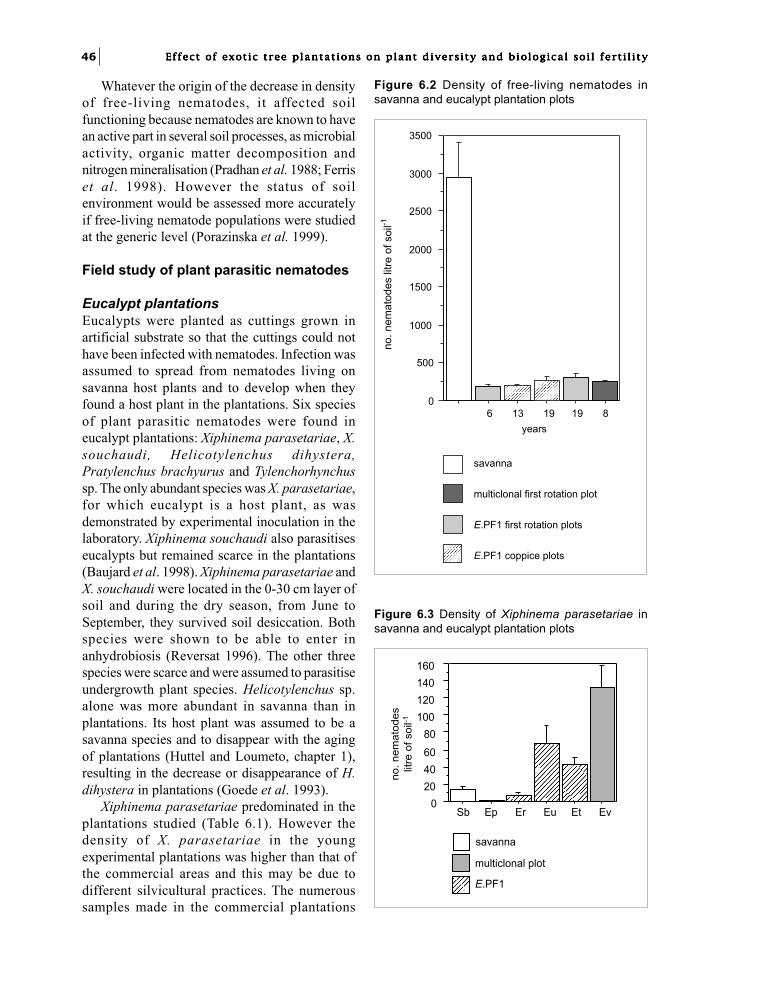
46 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
multiclonal plot
E.PF1
0
20
40
60
80
100
120
140
160
Sb Ep Er Eu Et Ev
no
. n
em
ato
de
s li
tre
of
soil-1
savanna
Whatever the origin of the decrease in densityof free-living nematodes, it affected soilfunctioning because nematodes are known to havean active part in several soil processes, as microbialactivity, organic matter decomposition andnitrogen mineralisation (Pradhan et al. 1988; Ferriset al. 1998). However the status of soilenvironment would be assessed more accuratelyif free-living nematode populations were studiedat the generic level (Porazinska et al. 1999).
Field study of plant parasitic nematodes
Eucalypt plantationsEucalypts were planted as cuttings grown inartificial substrate so that the cuttings could nothave been infected with nematodes. Infection wasassumed to spread from nematodes living onsavanna host plants and to develop when theyfound a host plant in the plantations. Six speciesof plant parasitic nematodes were found ineucalypt plantations: Xiphinema parasetariae, X.souchaudi, Helicotylenchus dihystera,Pratylenchus brachyurus and Tylenchorhynchussp. The only abundant species was X. parasetariae,for which eucalypt is a host plant, as wasdemonstrated by experimental inoculation in thelaboratory. Xiphinema souchaudi also parasitiseseucalypts but remained scarce in the plantations(Baujard et al. 1998). Xiphinema parasetariae andX. souchaudi were located in the 0-30 cm layer ofsoil and during the dry season, from June toSeptember, they survived soil desiccation. Bothspecies were shown to be able to enter inanhydrobiosis (Reversat 1996). The other threespecies were scarce and were assumed to parasitiseundergrowth plant species. Helicotylenchus sp.alone was more abundant in savanna than inplantations. Its host plant was assumed to be asavanna species and to disappear with the agingof plantations (Huttel and Loumeto, chapter 1),resulting in the decrease or disappearance of H.dihystera in plantations (Goede et al. 1993).
Xiphinema parasetariae predominated in theplantations studied (Table 6.1). However thedensity of X. parasetariae in the youngexperimental plantations was higher than that ofthe commercial areas and this may be due todifferent silvicultural practices. The numeroussamples made in the commercial plantations
savanna
multiclonal first rotation plot
E.PF1 first rotation plots
E.PF1 coppice plots
0
500
1000
1500
2000
2500
3000
3500
6 13 19 19 8
years
no. n
emat
odes
litr
e of
soi
l-1
Figure 6.2 Density of free-living nematodes insavanna and eucalypt plantation plots
Figure 6.3 Density of Xiphinema parasetariae insavanna and eucalypt plantation plots
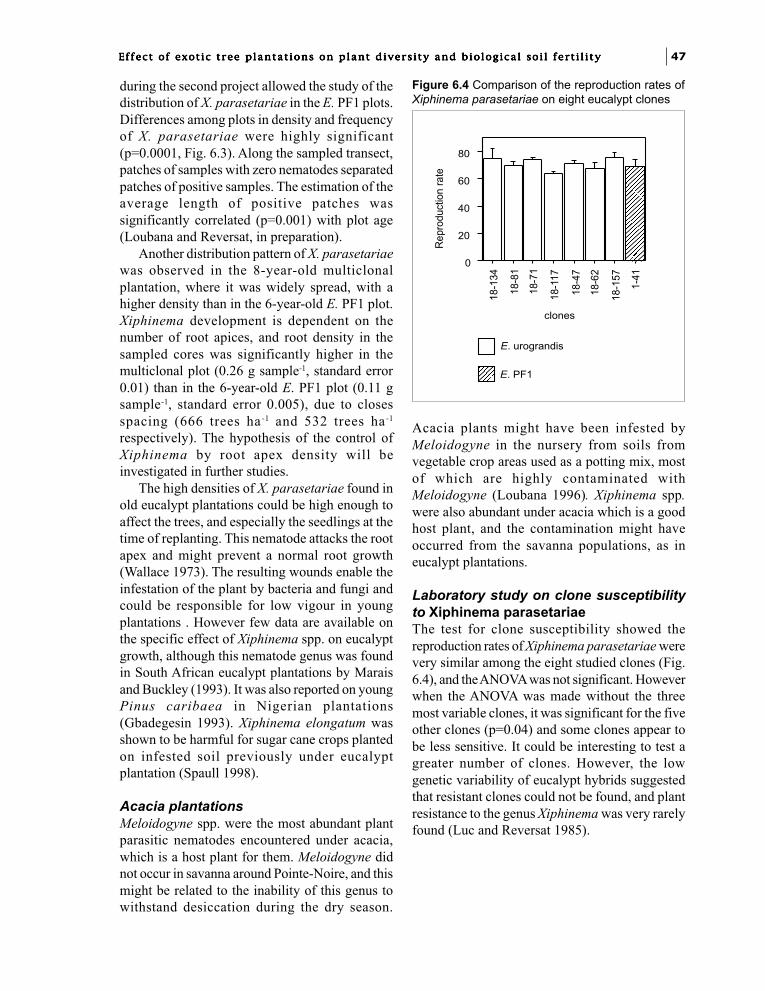
47Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
during the second project allowed the study of thedistribution of X. parasetariae in the E. PF1 plots.Differences among plots in density and frequencyof X. parasetariae were highly significant(p=0.0001, Fig. 6.3). Along the sampled transect,patches of samples with zero nematodes separatedpatches of positive samples. The estimation of theaverage length of positive patches wassignificantly correlated (p=0.001) with plot age(Loubana and Reversat, in preparation).
Another distribution pattern of X. parasetariaewas observed in the 8-year-old multiclonalplantation, where it was widely spread, with ahigher density than in the 6-year-old E. PF1 plot.Xiphinema development is dependent on thenumber of root apices, and root density in thesampled cores was significantly higher in themulticlonal plot (0.26 g sample-1, standard error0.01) than in the 6-year-old E. PF1 plot (0.11 gsample-1, standard error 0.005), due to closesspacing (666 trees ha-1 and 532 trees ha-1
respectively). The hypothesis of the control ofXiphinema by root apex density will beinvestigated in further studies.
The high densities of X. parasetariae found inold eucalypt plantations could be high enough toaffect the trees, and especially the seedlings at thetime of replanting. This nematode attacks the rootapex and might prevent a normal root growth(Wallace 1973). The resulting wounds enable theinfestation of the plant by bacteria and fungi andcould be responsible for low vigour in youngplantations . However few data are available onthe specific effect of Xiphinema spp. on eucalyptgrowth, although this nematode genus was foundin South African eucalypt plantations by Maraisand Buckley (1993). It was also reported on youngPinus caribaea in Nigerian plantations(Gbadegesin 1993). Xiphinema elongatum wasshown to be harmful for sugar cane crops plantedon infested soil previously under eucalyptplantation (Spaull 1998).
Acacia plantationsMeloidogyne spp. were the most abundant plantparasitic nematodes encountered under acacia,which is a host plant for them. Meloidogyne didnot occur in savanna around Pointe-Noire, and thismight be related to the inability of this genus towithstand desiccation during the dry season.
Acacia plants might have been infested byMeloidogyne in the nursery from soils fromvegetable crop areas used as a potting mix, mostof which are highly contaminated withMeloidogyne (Loubana 1996). Xiphinema spp.were also abundant under acacia which is a goodhost plant, and the contamination might haveoccurred from the savanna populations, as ineucalypt plantations.
Laboratory study on clone susceptibilityto Xiphinema parasetariaeThe test for clone susceptibility showed thereproduction rates of Xiphinema parasetariae werevery similar among the eight studied clones (Fig.6.4), and the ANOVA was not significant. Howeverwhen the ANOVA was made without the threemost variable clones, it was significant for the fiveother clones (p=0.04) and some clones appear tobe less sensitive. It could be interesting to test agreater number of clones. However, the lowgenetic variability of eucalypt hybrids suggestedthat resistant clones could not be found, and plantresistance to the genus Xiphinema was very rarelyfound (Luc and Reversat 1985).
0
20
40
60
80
E. urograndis
E. PF1
clones
Rep
rodu
ctio
n ra
te
18-1
34
18-8
1
18-7
1
18-1
17
18-4
7
18-6
2
18-1
57
1-41
Figure 6.4 Comparison of the reproduction rates ofXiphinema parasetariae on eight eucalypt clones

48 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
ConclusionsDramatic changes in nematode populationsoccurred when eucalypt trees were planted onsavanna. Population densities of free-livingnematodes decreased drastically to one tenth oftheir initial values, from several thousandindividuals per litre of soil in savanna to severalhundred of individuals per litre of soil in 6-year-old eucalypt plots. During the same time, thebiodiversity of plant parasitic nematodes wasreduced from savanna (five genera) to eucalyptplots (only one genus, Xiphinema) and the densityof the main species, X. parasetariae, increased inthe eucalypts.
Much remains to be done concerning thefunctional groups of free-living nematodes andtheir role in soil functioning. A more carefulapproach during the early stages (between thebeginning and 5 years) of eucalypt plantationestablishment is needed. The investigations shouldfocus on the variation of the free-living nematodepopulation and an evaluation of the soil microbialbiomass, of which a part is consumed by free-living nematodes.
The plant parasitic nematode populations arebetter known, because their taxonomical status isvery simple. Until now, only two species have beendemonstrated as parasites of eucalypt roots inCongo: Xiphinema parasetariae and X. souchaudi.Their densities were found to be as high as 150individuals per litre of soil in eucalypt plots, andthe possible detrimental consequence of thisinfestation on the functional status of roots andthe growth of trees deserves further study.

Effect of Exotic Tree Plantationson Invertebrate Soil MacrofaunaI.M.C. Mboukou-Kimbatsa1 and F. Bernhard-Reversat2
1 Laboratoire d’Écologie, Centre IRD de Pointe-Noire,B.P. 1286, Pointe-Noire, Congo.2 Laboratoire d’Ecologie des Sols Tropicaux, Centre IRDd’Ile de France, 32 avenue Henri Varagnat, 93143 Bondy,France.
Chapter 7.
Introduction
T he effect of plantations on plantbiodiversity was discussed by Parrotta(1993) and for the studied plantations by
Huttel and Loumeto (see chapter 1). Vegetation isknown to influence the trophic chain and soil faunain particular (Lavelle et al. 1994). Soilinvertebrates carry out essential functions for soilfunctioning, specifically soil organic matterdecomposition, soil aggregate formation, and soilmixing allowing the activation of micro organisms(Lavelle et al. 1997). Vegetation change isexpected to affect macrofauna (Abbott et al.1999;Vohland and Schroth 1999; Gonzales and Seastedt2000) and degraded lands generally undergo adecrease in soil fauna density and diversity (Tian1998). In order to assess the effect of growingexotic trees on savanna soils, soil macrofauna wasstudied among plots of various ages, species,hybrids and management practices.
Studied Plots and MethodsSoil macrofauna was studied in the Pointe-Noireage series plots of the first project (Sa, Ea, Eb, Ec,Ef, F Table 0.2), in Loudima plots (EL, AL, PL,SL, Table 0.2.) and in plots of the second project(Ep, Eq, Er, Es, Et, Eu, Ev, Ew, Sb, Table 0.3).Only E. urograndis was sampled in the two lastmulticlonal plots.
Soil macrofauna was sampled according to theTropical Soil Biology and Fertility Programmemethod (Anderson and Ingram 1993). In each plot,10 samples were taken 5 m apart along a randomly
chosen transect. Each sample was a column of soil30 cm deep and 25 x 25 cm square. Columns wereseparated into four layers: litter, 0-10 cm, 10-20cm and 20-30 cm. Samples were hand-sorted in alarge flat dish, and all visible invertebrates wereplaced in 85° alcohol and then in a 4% formolsolution for conservation. They were counted andweighed. To take into account weight loss due toalcohol or formalin, corrections for fresh weightwere applied (Anderson and Ingram 1993).Organisms were not identified to species and onlythe main taxa were considered. These taxa weregathered together in the following groups:earthworms, termites, ants, and litter inhabitinganimals.
Results and Discussion
Changes with vegetation and soilDensities of earthworms, termites and litteranimals were significantly different between plotswhereas ants were present everywhere and did notshow significant differences (Fig. 7.1). However,because density data are highly variable, therelationships between vegetation and frequency
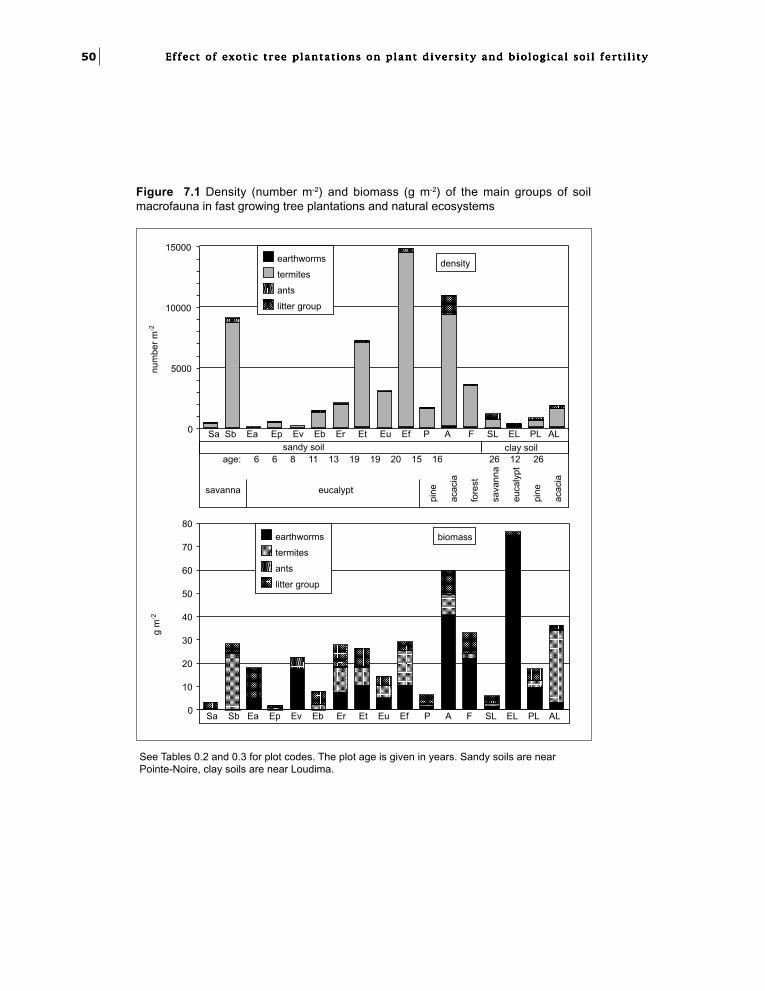
50 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
biomass80
70
60
50
40
30
20
10
0
earthworms
termites
ants
litter group
density
age: 6 6 8 11 13 19 19 20 15 16 26 12 26
0
15000
10000
5000
earthworms
termites
ants
litter group
Sa Sb Ea Ep Ev Eb Er Et Eu Ef P A F SL EL PL AL
savanna eucalypt
sandy soil
Sa Sb Ea Ep Ev Eb Er Et Eu Ef P A F SL EL PL AL
pine
acac
ia
fore
st
sava
nna
euca
lypt
pine
acac
ia
See Tables 0.2 and 0.3 for plot codes. The plot age is given in years. Sandy soils are near Pointe-Noire, clay soils are near Loudima.
clay soil
g m
-2nu
mbe
r m
-2
Figure 7.1 Density (number m-2) and biomass (g m-2) of the main groups of soilmacrofauna in fast growing tree plantations and natural ecosystems
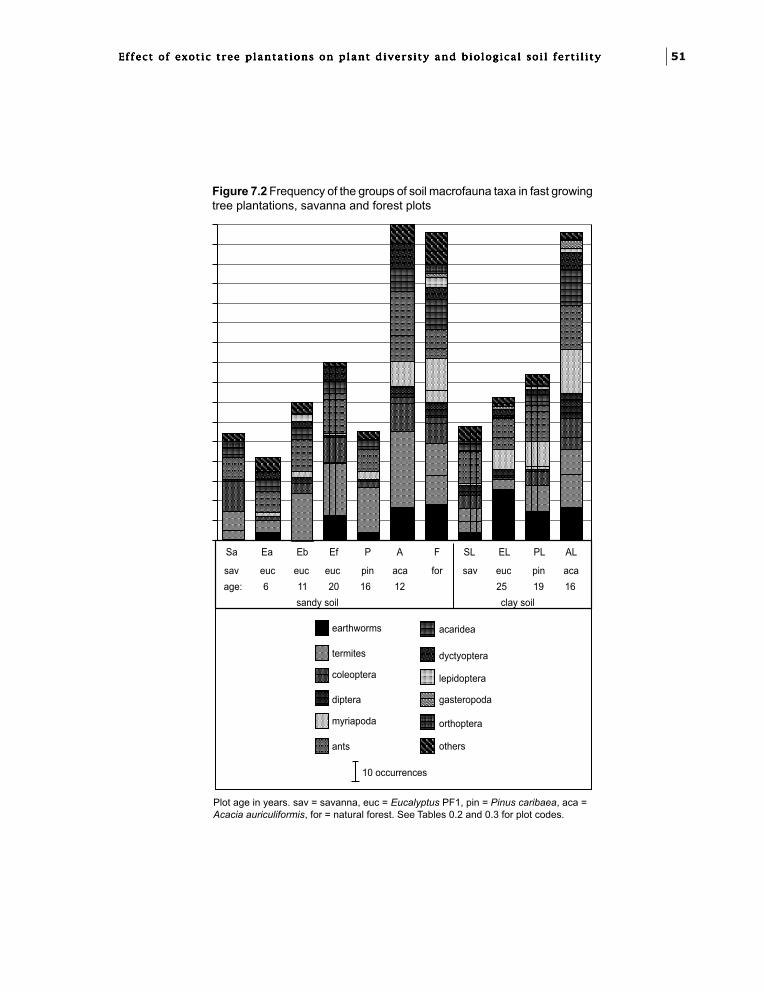
51Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
sav euc euc euc pin aca for
age: 6 11 20 16 12
sandy soil
sav euc pin aca
25 19 16
clay soil
earthworms
termites
coleoptera
diptera
myriapoda
ants
acaridea
dyctyoptera
lepidoptera
gasteropoda
orthoptera
others
10 occurrences
Sa Ea Eb Ef P A F SL EL PL AL
Figure 7.2 Frequency of the groups of soil macrofauna taxa in fast growingtree plantations, savanna and forest plots
Plot age in years. sav = savanna, euc = Eucalyptus PF1, pin = Pinus caribaea, aca =Acacia auriculiformis, for = natural forest. See Tables 0.2 and 0.3 for plot codes.
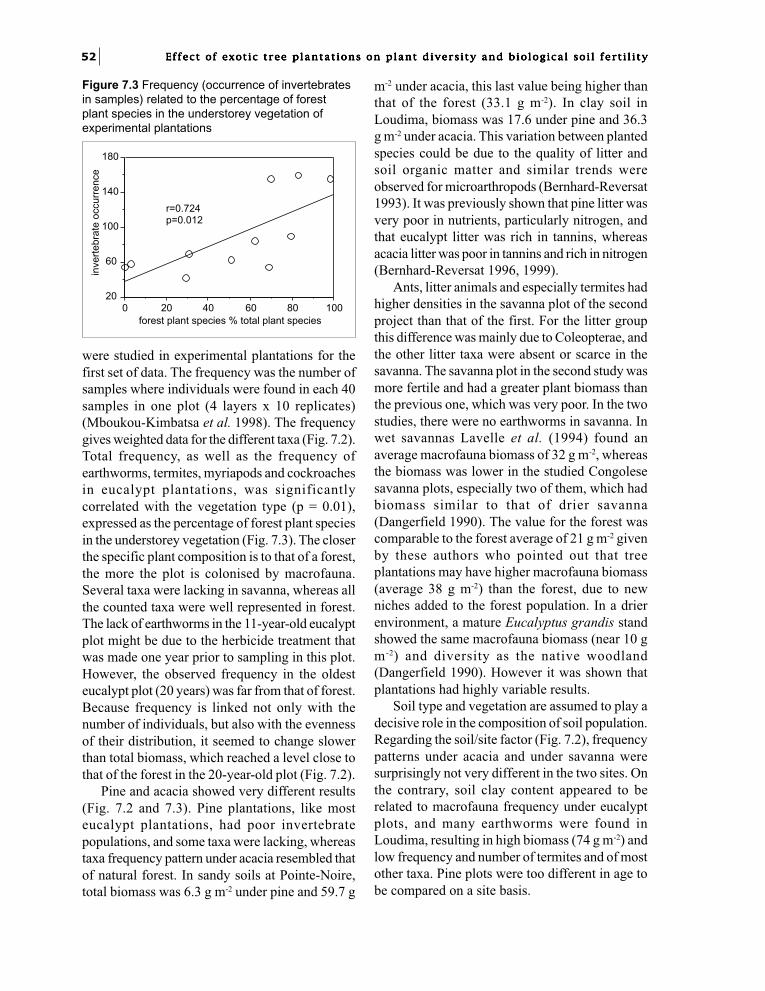
52 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
were studied in experimental plantations for thefirst set of data. The frequency was the number ofsamples where individuals were found in each 40samples in one plot (4 layers x 10 replicates)(Mboukou-Kimbatsa et al. 1998). The frequencygives weighted data for the different taxa (Fig. 7.2).Total frequency, as well as the frequency ofearthworms, termites, myriapods and cockroachesin eucalypt plantations, was significantlycorrelated with the vegetation type (p = 0.01),expressed as the percentage of forest plant speciesin the understorey vegetation (Fig. 7.3). The closerthe specific plant composition is to that of a forest,the more the plot is colonised by macrofauna.Several taxa were lacking in savanna, whereas allthe counted taxa were well represented in forest.The lack of earthworms in the 11-year-old eucalyptplot might be due to the herbicide treatment thatwas made one year prior to sampling in this plot.However, the observed frequency in the oldesteucalypt plot (20 years) was far from that of forest.Because frequency is linked not only with thenumber of individuals, but also with the evennessof their distribution, it seemed to change slowerthan total biomass, which reached a level close tothat of the forest in the 20-year-old plot (Fig. 7.2).
Pine and acacia showed very different results(Fig. 7.2 and 7.3). Pine plantations, like mosteucalypt plantations, had poor invertebratepopulations, and some taxa were lacking, whereastaxa frequency pattern under acacia resembled thatof natural forest. In sandy soils at Pointe-Noire,total biomass was 6.3 g m-2 under pine and 59.7 g
m-2 under acacia, this last value being higher thanthat of the forest (33.1 g m-2). In clay soil inLoudima, biomass was 17.6 under pine and 36.3g m-2 under acacia. This variation between plantedspecies could be due to the quality of litter andsoil organic matter and similar trends wereobserved for microarthropods (Bernhard-Reversat1993). It was previously shown that pine litter wasvery poor in nutrients, particularly nitrogen, andthat eucalypt litter was rich in tannins, whereasacacia litter was poor in tannins and rich in nitrogen(Bernhard-Reversat 1996, 1999).
Ants, litter animals and especially termites hadhigher densities in the savanna plot of the secondproject than that of the first. For the litter groupthis difference was mainly due to Coleopterae, andthe other litter taxa were absent or scarce in thesavanna. The savanna plot in the second study wasmore fertile and had a greater plant biomass thanthe previous one, which was very poor. In the twostudies, there were no earthworms in savanna. Inwet savannas Lavelle et al. (1994) found anaverage macrofauna biomass of 32 g m-2, whereasthe biomass was lower in the studied Congolesesavanna plots, especially two of them, which hadbiomass similar to that of drier savanna(Dangerfield 1990). The value for the forest wascomparable to the forest average of 21 g m-2 givenby these authors who pointed out that treeplantations may have higher macrofauna biomass(average 38 g m-2) than the forest, due to newniches added to the forest population. In a drierenvironment, a mature Eucalyptus grandis standshowed the same macrofauna biomass (near 10 gm-2) and diversity as the native woodland(Dangerfield 1990). However it was shown thatplantations had highly variable results.
Soil type and vegetation are assumed to play adecisive role in the composition of soil population.Regarding the soil/site factor (Fig. 7.2), frequencypatterns under acacia and under savanna weresurprisingly not very different in the two sites. Onthe contrary, soil clay content appeared to berelated to macrofauna frequency under eucalyptplots, and many earthworms were found inLoudima, resulting in high biomass (74 g m-2) andlow frequency and number of termites and of mostother taxa. Pine plots were too different in age tobe compared on a site basis.
Figure 7.3 Frequency (occurrence of invertebratesin samples) related to the percentage of forestplant species in the understorey vegetation ofexperimental plantations
forest plant species % total plant species
20
60
100
140
180
0 20 40 60 80 100
r=0.724p=0.012
inve
rteb
rate
occ
urre
nce
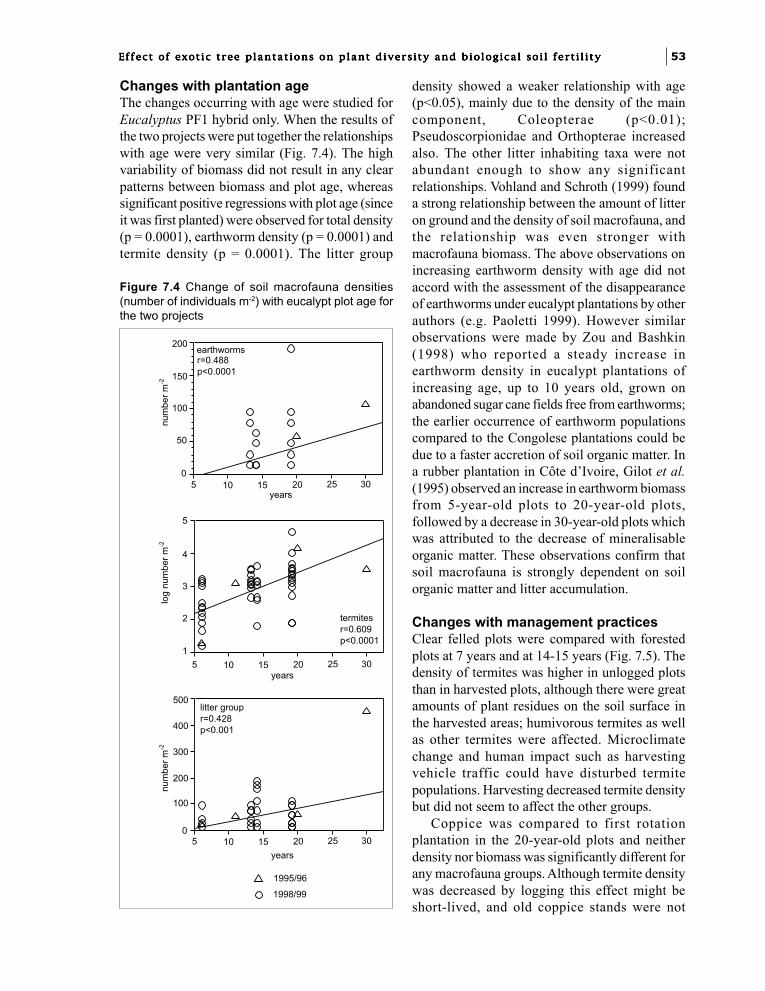
53Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Changes with plantation ageThe changes occurring with age were studied forEucalyptus PF1 hybrid only. When the results ofthe two projects were put together the relationshipswith age were very similar (Fig. 7.4). The highvariability of biomass did not result in any clearpatterns between biomass and plot age, whereassignificant positive regressions with plot age (sinceit was first planted) were observed for total density(p = 0.0001), earthworm density (p = 0.0001) andtermite density (p = 0.0001). The litter group
density showed a weaker relationship with age(p<0.05), mainly due to the density of the maincomponent, Coleopterae (p<0.01);Pseudoscorpionidae and Orthopterae increasedalso. The other litter inhabiting taxa were notabundant enough to show any significantrelationships. Vohland and Schroth (1999) founda strong relationship between the amount of litteron ground and the density of soil macrofauna, andthe relationship was even stronger withmacrofauna biomass. The above observations onincreasing earthworm density with age did notaccord with the assessment of the disappearanceof earthworms under eucalypt plantations by otherauthors (e.g. Paoletti 1999). However similarobservations were made by Zou and Bashkin(1998) who reported a steady increase inearthworm density in eucalypt plantations ofincreasing age, up to 10 years old, grown onabandoned sugar cane fields free from earthworms;the earlier occurrence of earthworm populationscompared to the Congolese plantations could bedue to a faster accretion of soil organic matter. Ina rubber plantation in Côte d’Ivoire, Gilot et al.(1995) observed an increase in earthworm biomassfrom 5-year-old plots to 20-year-old plots,followed by a decrease in 30-year-old plots whichwas attributed to the decrease of mineralisableorganic matter. These observations confirm thatsoil macrofauna is strongly dependent on soilorganic matter and litter accumulation.
Changes with management practicesClear felled plots were compared with forestedplots at 7 years and at 14-15 years (Fig. 7.5). Thedensity of termites was higher in unlogged plotsthan in harvested plots, although there were greatamounts of plant residues on the soil surface inthe harvested areas; humivorous termites as wellas other termites were affected. Microclimatechange and human impact such as harvestingvehicle traffic could have disturbed termitepopulations. Harvesting decreased termite densitybut did not seem to affect the other groups.
Coppice was compared to first rotationplantation in the 20-year-old plots and neitherdensity nor biomass was significantly different forany macrofauna groups. Although termite densitywas decreased by logging this effect might beshort-lived, and old coppice stands were not
Figure 7.4 Change of soil macrofauna densities(number of individuals m-2) with eucalypt plot age forthe two projects
earthwormsr=0.488p<0.0001150
100
50
0
200
5 10 15 20 25 30years
5 10 15 20 25 30years
5 10 15 20 25 30
years
num
ber
m-2
log
num
ber
m-2
num
ber
m-2
1
2
3
4
5
termitesr=0.609p<0.0001
litter groupr=0.428p<0.001
500
400
300
200
100
0
1995/96
1998/99
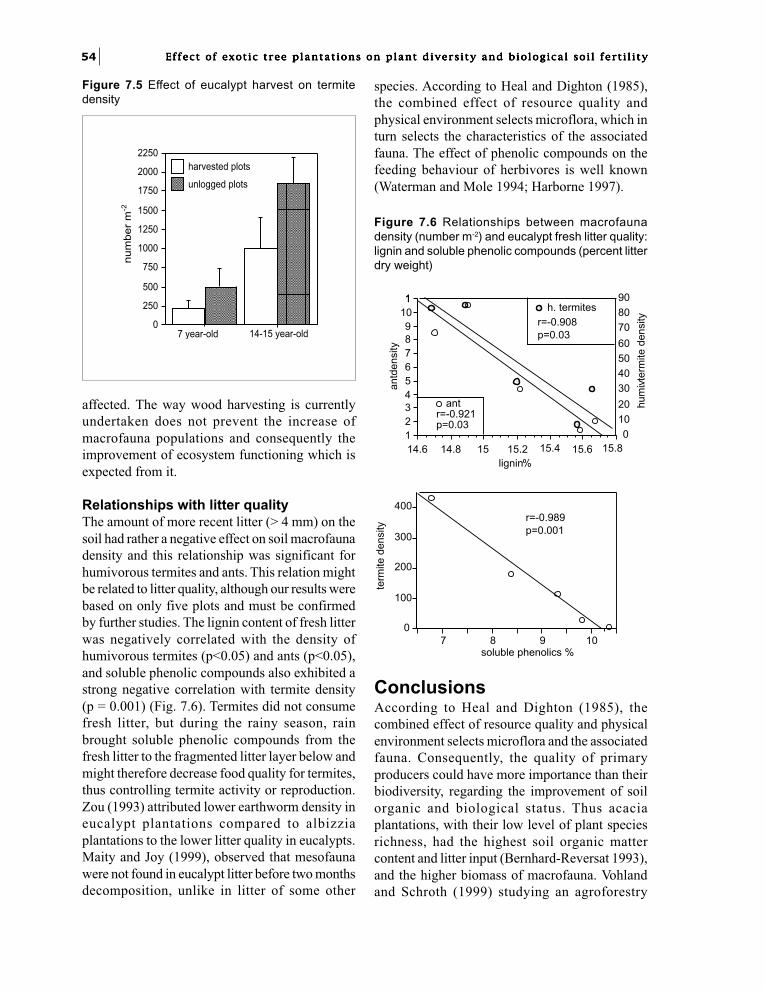
54 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
species. According to Heal and Dighton (1985),the combined effect of resource quality andphysical environment selects microflora, which inturn selects the characteristics of the associatedfauna. The effect of phenolic compounds on thefeeding behaviour of herbivores is well known(Waterman and Mole 1994; Harborne 1997).
affected. The way wood harvesting is currentlyundertaken does not prevent the increase ofmacrofauna populations and consequently theimprovement of ecosystem functioning which isexpected from it.
Relationships with litter qualityThe amount of more recent litter (> 4 mm) on thesoil had rather a negative effect on soil macrofaunadensity and this relationship was significant forhumivorous termites and ants. This relation mightbe related to litter quality, although our results werebased on only five plots and must be confirmedby further studies. The lignin content of fresh litterwas negatively correlated with the density ofhumivorous termites (p<0.05) and ants (p<0.05),and soluble phenolic compounds also exhibited astrong negative correlation with termite density(p = 0.001) (Fig. 7.6). Termites did not consumefresh litter, but during the rainy season, rainbrought soluble phenolic compounds from thefresh litter to the fragmented litter layer below andmight therefore decrease food quality for termites,thus controlling termite activity or reproduction.Zou (1993) attributed lower earthworm density ineucalypt plantations compared to albizziaplantations to the lower litter quality in eucalypts.Maity and Joy (1999), observed that mesofaunawere not found in eucalypt litter before two monthsdecomposition, unlike in litter of some other
ConclusionsAccording to Heal and Dighton (1985), thecombined effect of resource quality and physicalenvironment selects microflora and the associatedfauna. Consequently, the quality of primaryproducers could have more importance than theirbiodiversity, regarding the improvement of soilorganic and biological status. Thus acaciaplantations, with their low level of plant speciesrichness, had the highest soil organic mattercontent and litter input (Bernhard-Reversat 1993),and the higher biomass of macrofauna. Vohlandand Schroth (1999) studying an agroforestry
Figure 7.5 Effect of eucalypt harvest on termitedensity
14-15 year-old7 year-old0
250
500
750
1000
1250
1500
1750
2000
2250harvested plots
unlogged plots
num
ber
m-2
Figure 7.6 Relationships between macrofaunadensity (number m-2) and eucalypt fresh litter quality:lignin and soluble phenolic compounds (percent litterdry weight)
0
100
200
300
400
7 8 9 10soluble phenolics %
r=-0.989
p=0.001
lignin%
15.8
0
10
20
30
40
50
60
70
80
90
1
2
3
4
5
6
7
8
9
10
11
14.6 14.8 15 15.2 15.4 15.6
antr=-0.921p=0.03
h. termites
r=-0.908
p=0.03
an
tde
nsity
hu
miv.
term
ite d
ensity
term
ite d
ensity

55Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
system in Brazil, observed a greater density ofmacrofauna in a tree monoculture of peach palmthan in a tree polyculture including peach palm,because this tree had a better litter quality. In theCongolese plantations which are grown on a verypoor sandy soil, the increase of organic matterresource could be of tremendous importance inchanging savanna habitat to forest habitat, not onlyregarding energy but also nutrient resource andbiochemical quality (Mboukou-Kimbatsa et al.1998). Although eucalypt plantations exhibit lowerdensity and biodiversity than other tree species,they are able to improve degraded soil fertility byenhancing macrofauna populations as they do formesofauna (Day et al. 1999).
Some trends were assessed through the 1996and 1999 studies gathering 11 eucalypt plots inthe Pointe-Noire plantations. Soil macrofaunaspread out in plantations, together with aging andsoil change, at different rates according to theplanted tree species. Forest exploitation had littleimpact on these changes.

Soil Biological Fertility UndergoesFundamental Changes When Fast-growingExotic Trees Are Plantedon a Poor Savanna SoilF. Bernhard-Reversat1 and C. Huttel2
1 Laboratoire d’Ecologie des Sols Tropicaux, Centre IRDd’Ile de France, 32 avenue Henri Varagnat, 93143 Bondy,France.2 Le Caillou, Valojoux, 24290 Montignac, France.
Conclusions
The changing environment from savanna toartificial forests was investigated with amultidisciplinary approach including
vegetation, soil and soil organisms, to understandthe main relationships between plant diversity,management practices and the biological factorsof fertility (Fig. 8.1).
Plantations Improve BiodiversityAn original vegetation, rich in pioneer species,developed under plantations. Although secondaryforest patches occurred among the plantations andcould act as a seed source, the floristic compositionof the undergrowth was very different from thatobserved in a nearby forest: most species presentin the forest were not found in the plantations, andmost species in the plantation undergrowth wereabsent from the forest plot. The plantations studiedwere grown on savannas which are not colonisedby forest species because annual fires destroy theforest tree seedlings, and where natural evolutionfrom grassland to forest seems not to be possible,unlike the situation described in unburntCongolese savannas (de Foresta 1990).Commercial tree plantations in a very poor savannaenvironment bring drastic changes inenvironmental conditions, due to the reduction offire occurrence and to the change toward forestmicroclimate and forest soil. These changes allowplant diversity increase in understorey vegetation,as observed elsewhere in afforested degraded lands
such as mined sites or abandoned pastures (Lugo1997; Parrotta et al. 1997; Powers et al. 1997),although Parrotta and Knowles (1999) showed thatchanges were faster when native species wereplanted instead of exotic species. In unloggedeucalypt plantations, the general trends withplantation age were an increase of species number,forest species percentage, woody cover density,height and basal area. In managed commercialplantations, the species richness was higher,mainly due to the abundance of savanna andruderal species, and height and basal area werelower. Further studies should examine the role ofthe natural forest and the conditions for seeddissemination, germination and seedling growthin plantations, undergrowth. Some results,especially the relationships between soil C, soilpH, and either the number of undergrowth speciesor the percentage of forest species, suggested thatthe changing soil environment through organicmatter accumulation could promote theestablishment of the undergrowth.
This biodiversity, unwanted by foresters, couldimprove soil functioning by increasing thediversity of organic sources for soil biota and bydecreasing the negative effect of eucalypt litter
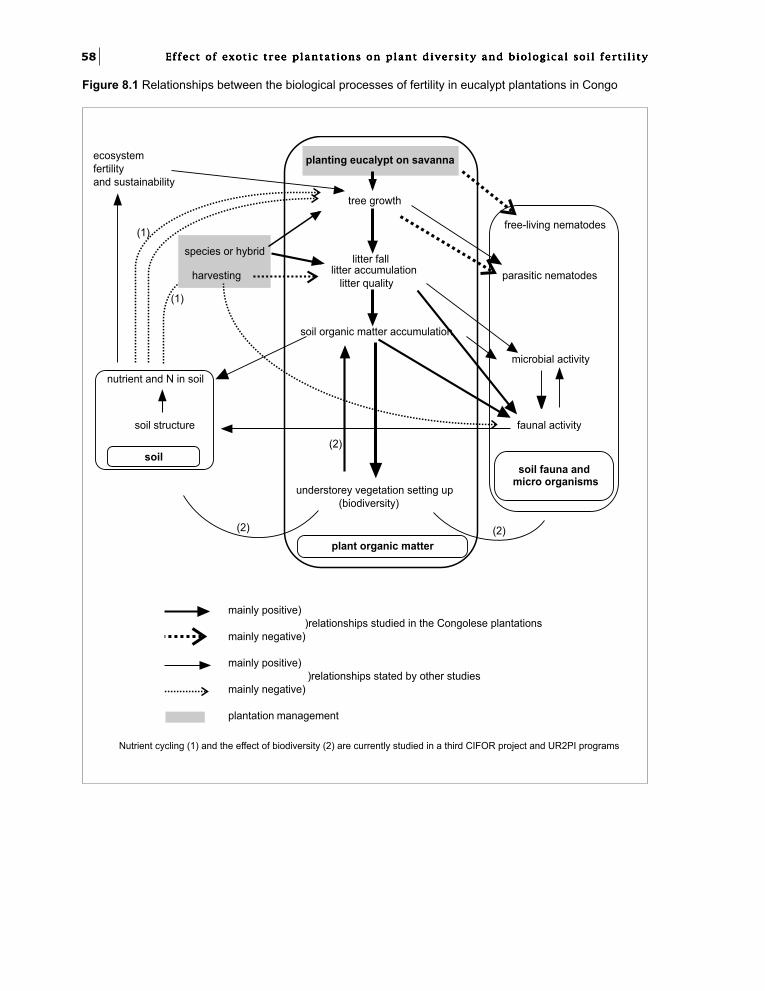
58 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
tree growth
litter falllitter accumulation
soil organic matter accumulation
microbial activity
faunal activitysoil structure
harvesting
understorey vegetation setting up
(biodiversity)
ecosystem
fertility
and sustainability
parasitic nematodes
nutrient and N in soil
planting eucalypt on savanna
free-living nematodes
soil fauna andmicro organisms
plant organic matter
soil
mainly positive)
)relationships studied in the Congolese plantations
mainly negative)
mainly positive)
)relationships stated by other studies
mainly negative)
plantation management
(1)
litter quality
(1)
species or hybrid
(2)
(2)
(2)
Nutrient cycling (1) and the effect of biodiversity (2) are currently studied in a third CIFOR project and UR2PI programs
Figure 8.1 Relationships between the biological processes of fertility in eucalypt plantations in Congo

59Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
biochemicals such as polyphenols and terpenes,although undergrowth vegetation competes forwater and nutrients with the tree crops (Dembi1988). Microclimate, litter quantities and tree litterphysical and chemical quality were assumed tocontrol the abundance and nature of soil biota, anddiffered according to the planted species. Thehypothesis of modified litter influence beneathundergrowth patches on soil biological processeswill be studied during further investigations.
Plantations Improve SoilOrganic Matter Status andBiological ActivityThe present results suggest that litter accumulationand quality, and the resulting soil organic matteraccumulation and quality, were among the mainecological changes when savanna was planted withtrees, and determined the development ofundergrowth and changes in soil characteristics(Wardle and Lavelle 1997). The fact that soil faunawas mostly correlated with percentage of forestor savanna species, rather than with the density ofunderstorey plants, and the fact that changes invegetation and fauna occurred after 7-10 yearswhen soil organic matter content began to increase,suggested that both fauna and understoreyvegetation were mainly affected by the type ofhabitat. Heal and Dighton (1985) observed thatthe increasing importance of fauna and theincreasing dominance of macrofauna were relatedto the improvement of the quality of primaryproduction residues. The great differences in litterquantity and quality occurring between eucalypt,acacia and pine resulted in major differences insoil microfauna and macrofauna density andbiomass, and suggested that litter quality of thetree crop took precedence over plant biodiversityin controlling soil biota. In eucalypt plantations,the trend of fauna distribution to be associated withvariations in eucalypt litter quality also supportsthis conclusion.
Lavelle (1996) suggests three mechanismslinking biodiversity and soil biological processes:the first implies plant diversity as the origin of soilfauna diversity (nested biodiversity), the secondimplies soil structure as mediator betweenbiodiversity and functioning, and the third impliesorganic matter level and quality for changes in
energy resources. In the plantations studied,organic matter resource could be of tremendousimportance in changing savanna environment toforest environment, not only regarding energyresource availability but also nutrient resource andbiochemical quality.
Improved soil processes were observed withthe aging of eucalypt plantations due to changesin the soil-litter system: increased decompositionrates together with increased litterfall in agingplantations is assumed to enhance nutrient cycling.Increased soil organic matter with plot ageenhanced cation exchange capacity and wastherefore expected to improve the retention ofnutrient imputs from rainfall, litter and fertilisers.An increase in soil organic matter is also assumedto be responsible for changes in macrofauna, andan improved soil functioning is expected from theincreasing density of earthworm, termites and litterfauna (Lavelle 1994; Altieri 1999), related mainlyto a faster organic matter turnover and improvedsoil structure.
Assessment of the AdverseEffect of PlantationsAmong fast-growing trees, eucalypt is the only onegrown commercially on large areas in the Congoand consequently their adverse effects on theenvironment must be managed.
Strong evidence for disturbance of microbialprocesses in eucalypt plantations is indicated bythe absence of N fixation (chapter 4), nitrification(Bernhard-Reversat 1996), and white rot fungi(Bernhard-Reversat and Schwartz 1997).Although the lack of nitrification reduces nitrogenleaching, the decreasing nitrogen content of soilorganic matter with plot age leads to mineralnitrogen deficiencies for tree growth, requiringfertiliser use by the second rotation, and this isone of the main problems foresters have to address.The introduction of a nitrogen-fixing understoreycrop could greatly improve litter quality and soilorganic matter content, but could also increase thelevel of tree infestation by plant parasiticnematodes.
Eucalypts are often regarded as damaging forthe environment (Poore and Fries 1985) becauseof their allelopathic effects, which was shown inexperimental conditions for the Congolese

60 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
plantations (Bernhard-Reversat 1999) andnumerous authors in other countries (e.g.Lisanework and Michelsen 1993; Souto et al.1995; Reigosa et al. 1999). Concerningundergrowth biodiversity, eucalypts depressedforest species more than pine or acacia plantations,and the question arises if it is due either toallelopathy or to environmental conditions. Lightintensity is often put forward as an alternativecause, but observations showed a lower canopydensity under eucalypt than under the other plantedspecies (Loumeto, unpublished results), whichshould result in a higher light intensity in eucalyptstands and favour pioneer species. The decreasein soil humidity due to eucalypt is also pointedout, but although comparison with savanna showeda depletion of soil humidity under eucalypt below1 m, no data are available for comparison withother tree species (Nizinski et al., in press). Dembi(1988) showed that in a young eucalypt plantationthe adventitious vegetation competed efficientlyfor water, but in older plantations it might bestressed by water shortage. Nevertheless the needfor weeding and the abundance of undergrowth inthe field after 10-years of eucalypt croppingdemonstrated that some plant species were ableto avoid the adverse effect of eucalypts on plantgrowth.
The increasing density of the eucalypt parasiticnematodes Xiphinema parasetariae with the agingof plantations suggested that continuous eucalyptcultivation should result in an entirely infestedarea, and the effect of some practices, such as treespacing or stump killing, on population decreaseshould be assessed. The effect of nematodes ontree growth is still unknown and should be studiedtogether with the evaluation of the susceptibilityof hybrids and its relationship with easilymeasurable parameters. Eucalypt plantations seemto be safe for eventual further cropping: Xiphinemaparasetariae does not affect most local crops(cassava, groundnut, vegetable crops) which couldreplace the plantations, and the nematodes whichare highly noxious for many food crops (mainlyMeloidogyne spp.), do not develop on eucalyptroots.
The present practices of exploitation ofeucalypt did not affect biological soil fertilityfactors, although the floristic composition of
undergrowth and its abundance were changed.These practices kept the existing litter and residueson the ground, and even though the temporaryincrease of litter after exploitation does not lastmore than one year (Nzila, personalcommunication), it balances the low litterfall inthe young regrowth. Consequently, although thelitter system was affected by exploitation, soilorganic matter was not obviously affected, and thebiological activity in litter and soil was preserved.Silvicultural practices which improve theconservation of plant residue on the ground,besides being recommended to prevent nutrientexport (O’Connell and Sankaran 1997), willenhance soil fauna density and consequently soilfertility (Altieri 1999).
ConclusionsThe debate on growing block plantations of exotictrees is not closed and more factors must be takeninto account than those studied here. Thereplacement by plantations of biological diverseecosystems like rain forests are rightly questioned.In the case of harsh environment like theCongolese savanna, silviculture is a land use thathas numerous positive effects although there aremany problems still to be addressed.
The increase in soil organic matter whichaccounts for many positive changes is however afragile process. All management practices likelyto decrease inputs, such as the burning of harvestresidues, the harvest of twigs and leaves for fuel,and the harvest of logs with the bark, are expectedto counteract environmental improvementresulting from eucalypt growth (Brown et al.1997). The choice of species and hybrids, besidetheir growth and wood qualities, should take intoaccount litter quality and decomposition rate.Further research is needed on the cummulativeeffect of silvicultural practices involved in short-rotation forestry and its relation with soil organicmatter.
This interdisciplinary work emphasisedpositive trends in eucalypt and other fast-growingplantations in relation to plant diversity andbiological soil fertility. The knowledge generatedby this study suggests changes in cultural practiceswhich could have potential for more sustainableproduction in the long-term.

Abbasi, S.A. and Vinithan, S. 1997. Ecologicalimpacts of eucalypts - myths and realities.Indian Forester 123: 710-739.
Abbasi, S.A. and Vinithan, S. 1999. Ecologicalimpacts of Eucalyptus tereticornis-globulus(Eucalyptus hybrid) plantation on a miningarea. Indian Forester 125: 163-186.
Abbott, I., Wills, A. and Burbidge, T. 1999. Theimpact of canopy development on arthropodfaunas in recently established Eucalyptusglobulus plantations in Western Australia.Forest Ecology and Management 121: 147-158.
Abelho, M. and Graça, M.A.S. 1996. Effect ofeucalyptus afforestation on leaf litter dynamicsand macro invertebrate community structureof stream in Central Portugal. Hydrobiologia324: 195-204.
Aggangan, R.T., O’Connell, A.M., McGrath, J.F.and Dell, B. 1999. The effect of Eucalyptusglobulus leaf litter on C and N mineralizationin soil from pasture and native forest. SoilBiology and Biochemistry 31: 1481-1487.
Altieri, M. 1999. The ecological role ofbiodiversity in agroecosystems. Agriculture,Ecosystems and Environment 74: 19-31.
Anderson, J.M. and Ingram, J.S.I. 1993. Tropicalsoil biology and fertility. A handbook of
References
methods. C.A.B. International, Oxford,England. 221p.
Anderson, J.M. and Swift M.J. 1983.Decomposition in tropical forests. In: SuttonS.L., Whitmore T.C. and Chadwick A.C. (eds.)Tropical rain forest: ecology and management,287-309. Special publication 2, BritishEcological Society. Blackwell Scientific,Oxford, England.
Anonymous. 1994. Model DR/700 portablecolorimeter instrument manual. HACHCompany, Loveland. 69.7-69.12.
Armstrong, A. and van Hensbergen, H.J. 1996.Impact of afforestation with pine onassemblages of native biota in South Africa.Suid-Afrikaan Bosbourydskrif 175: 35-42.
Arunachalam, A., Pandeu H.N., Tripathi, R.S. andMaithani, K. 1996. Fine root decompositionand nutrient mineralization patterns in asubtropical humid forest following tree cutting.Forest Ecology and Management 86: 141-150.
Attiwill, P.M., Polglase, P.J., Weston, P.J. andAdams, M.A. 1996. Nutrient cycling in forestsof south-eastern Australia. In: Attiwill P.M. andAdams M.A. (eds.) Nutrition of eucalypts, 191-227. CSIRO Publishing, Collingwood,Australia.

62 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Aweto, O.A. 1995. Organic carbon diminutionand estimates of carbon dioxide release fromplantation soil. The Environmentalist 15: 10-15.
Baker, T.G. and Attiwill, P.M. 1985. Above groundnutrient distribution and cycling in Pinusradiata D. Don and Eucalyptus obliquaL’Herit. forests in southeastern Australia.Forest Ecology and Management 13: 41-52.
Balesdent, J., Pétraud, J.P. and Feller, C. 1991.Effets des ultrasons sur la distributiongranulométrique des matière organiques dessols. Sciences du Sol 29: 95-105.
Bargali, S.S., Singh, S.P. and Singh, R.P. 1992.Structure and function of an age series ofeucalypt plantations in Central Himalaya. IINutrient dynamics. Annals of Botany 69: 413-421.
Bargali, S.S., Singh, S.P. and Singh, R.P. 1993Pattern of weight loss and nutrient release fromdecomposing leaf litter in an age series ofeucalypt plantation. Soil Biology andBiochemistry 25: 1731-1738.
Basanta M., Vizcaino D.E., Casal, M. and Morey,M. 1989. Diversity measurements in shrublandcommunities of Galicia (NW Spain). Vegetatio82: 105-112.
Baujard, P., Luc, M. and Reversat, G. 1998.Description of Xiphinema souchaudi n. sp. andobservations on X. yapoense Luc, 1958 and X.algeriense Luc and Kostadinov, 1982(Nematoda: Longidoridae). Fundamental andApplied Nematology 21: 637-644.
Bernhard-Reversat, F. 1987. Les cycles deséléments minéraux dans un peuplement àAcacia seyal et leur modification en plantationd’Eucalyptus au Sénégal. Acta Oecologica,Oecologia Generalis 8: 3-16.
Bernhard-Reversat F. 1988. Soil nitrogenmineralization under a Eucalyptus plantationand a natural Acacia forest in Senegal. ForestEcology Management 23: 233-244.
Bernhard-Reversat, F. 1991. Evolution of the soillitter interface under Eucalyptus plantation onsandy soil in Congo. Acta Oecologia 12: 825-828.
Bernhard-Reversat, F. 1993. Dynamics of litterand organic matter at the soil-litter interface infast-growing tree plantations on sandy ferraliticsoils (Congo). Acta Oecologia 14: 179-195.
Bernhard-Reversat, F. 1996. Nitrogen cycling intree plantations grown on poor savanna soil inCongo. Applied Soil Ecology 4: 161-172.
Bernhard-Reversat F. 1998. Change in CO2 releaserelationships with initial litter quality duringearly laboratory decomposition of tropical leaflitters. European Journal of Soil Biology 34:117-122.
Bernhard-Reversat, F. 1999. The leaching ofEucalyptus hybrids and Acacia auriculiformisleaf litter: laboratory experiments on earlydecomposition and ecological implications inCongolese tree plantations. Applied SoilEcology 12: 251-261.
Bernhard-Reversat, F. and Loumeto, J.J. In press.The litter system in African forest-treeplantations. In: Reddy, W. (ed.) Tropicalplantation-forest management and the soil littersystem: litter, biota and soil nutrient dynamics.Oxford and IBH Publications, New Delhi.
Bernhard-Reversat, F. and Schwartz, D. 1997.Change in lignin content during litterdecomposition in tropical forests soils (Congo):comparison of exotic plantations and nativestands. C.R. Academie des Sciences, Paris,Sciences de la Terre et des Planetes 325: 427-432.
Bernhard-Reversat, F., Diangana, D. and Tsatsa,M. 1993. Biomasse, minéralomasse etproductivité en plantation d’Acacia mangiumet A. auriculiformis au Congo. Bois et Forêtsdes Tropiques No. 238: 35-44.
Binkley, D. 1997. Bioassays of the influence ofEucalyptus saligna and Albizia falcataria onsoil nutrient supply and limitation. ForestEcology and Management 91: 229-234.

63Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Binkley, D. and Resh, S.C. 1999. Rapid changesin soils following Eucalyptus afforestation inHawaii. Soil Science Society of AmericanJournal 63: 222-225.
Boissezon, (de) P. 1967. Etude pédologique de lavallée du Niari. In: Quinze ans de travaux etde recherches dans les pays du Niari. Tome 3.Les sols de la vallée du Niari: étudespédologiques et agro-pédologiques, 9-48.ORSTOM , Paris.
Bond, W.J. and Stock, W.D. 1990. Preliminaryassessment of the grading of eucalyptus clonesusing carbon isotope discrimination. SouthAfrican Forestry Journal 154: 51-55.
Bouillet, J.P., Nizinski, G., Nzila, J.D. and Ranger,J. 1997. The sustainability of Eucalyptuscommercial plantations: the Congoleseapproach. In: Proceedings of IUFRO meetingSilviculture and Genetic Improvement ofEucalyptus, Salvador, Bahia, Brazil, August1997. Vol. 4: Environmental and social impactsof eucalypt plantations, 232-237. EMBRAPA,Colombo, Brazil.
Bouillet, J.P., Nzila, J.D., Laclau, J.P. and Ranger,J. 2000. Effects of site management onEucalyptus plantations in the Equatorial Zone,on the coastal plains of the Congo. In: Nambiar,E.K.S., Tiarks, A., Cossalter, C. and Ranger, J.(eds.) Site management and productivity intropical plantation forests: a progress report:Workshop proceedings 7-11 December 1999,Kerala, India, 12-20. CIFOR, Bogor, Indonesia.
Bouvet, J.M. 1998. Les plantations d’Eucalyptus.Evolutions récentes et perspectives. InternalNote. CIRAD-Forêt, Montpellier, France. 35p.
Bouvet, J.M. 1999. Les plantations d’Eucalyptus.Evolutions récentes et perspectives. LeFlamboyant 49: 4-14.
Brent, E., Binkley, D. and Bashkin, M. 1996.Influence of adjacent stand on spatial patternof soil carbon and nitrogen in Eucalyptus andAlbizia plantations. Canadian Journal ofForestry Research 26: 1501-1503.
Brosset, A. 1966. Contribution à la faune duCongo (Brazzaville). Mission A. Villiers et A.Descarpentries. XX Chiroptères. Bulletin de l’Institut Fondanental d’ Afrique Noire 28 (A):362-369.
Brosset, A. 1990. A long term study of the rainforest birds in Mpassa (Gabon). In: Keast, A.(ed.) Biogeography and ecology of forest birdcommunities, 259-274. SPB AcademicPublishers bv, The Hague.
Brosset, A.1997. Peuplements en oiseaux desplantations d’Eucalyptus dans la région dePointe-Noire, Congo. Revue d’ Ecolologie 52:133-151.
Brosset, A. and Erard, C. 1986. Les oiseaux desrégions forestières du nord-est du Gabon. Vol.1: Ecologie et comportement des espèces.SNPN, Paris. 289p.
Brown, A.G., Nambiar, E.K.S. and Cossalter, C.1997. Plantations for the tropics - their role,extent and nature. In: Nambiar E.K.S. andBrown A.G. (eds.) Management of soil,nutrients and water in tropical plantationforests, 3-23. ACIAR Monograph No. 43.ACIAR, Canberra.
Calder, I.R., Hall, R.L. and Adlard, P.G. (eds.)1992. Growth and water use of forestplantations. Wiley and Sons, New York. 381p.
Chunyang, L. 1999. Carbon isotope composition,water-use efficiency and biomass productivityof Eucalyptus microtheca under different watersupplies. Plant and Soil 214: 165-171.
Conde, E., Cadahaia, E. and Garcia-Vallejo, M.C.1997. Low molecular weight polyphenols inleaves of Eucalyptus camaldulensis, E.globulus and E. rudis. Phytochemical Analysis8: 186-193.
Constantinides, M. and Fownes, J.H. 1994.Nitrogen mineralization from leaves and litterfrom tropical plants: relationship to nitrogen,lignin and soluble polyphenol concentrations.Soil Biology Biochemistry 26: 49-55.

64 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Cortez, N.R.D.S. 1996. Compartimentos e ciclosde nutrientes em plantações de Eucalyptusglobulus Labill. ssp. globulus e Pinus pinasterAiton. PhD Thesis in Agronomy, Lisboa.
Crews, T.E., Farrington, H. and Vitousek, P.M.2000. Changes in asymbiotic, heterotrophicnitrogen fixation on leaf litter of Metrosiderospolymorpha with long-term ecosystemdevelopment in Hawaii. Ecosystems 3: 386-395.
Dambrine, E., Vega, J.A., Taboada, T., Rodriguez,L., Fernandez, C., Macias, F. and Gras, J.C.2000. Bilans d’éléments minéraux dans depetits bassins versants forestiers de Galice (NWEspagne). Annals of Forest Science 57: 23-38.
Dangerfield, J.M. 1990. Abundance, biomass anddiversity of soil macrofauna in savannawoodland and associated managed habitat.Pedobiologia 34: 141-150.
Day, C., Majer, J. and Mattiessen, J.N. 1999. Leaflitter invertebrates in the Western Australianwheatbelt: recolonisation of revegetation andits implications in restoring ecosystemfunctions and biodiversity. In: Proceedings ofthe 7th Australian Conference on GrasslandInvertebrate Ecology, 184-194. CSIROEntomology, CSIRO Centre for MediterraneanAgricultural Research, Wembley.
Delwaulle, J.C. 1989. Plantations clonales auCongo. Point des recherches sur le choix desclones dix ans après les premières plantations.In: Gibson, G.L., Griffin, A.R. and Matheson,A.C. (eds.) Breeding tropical trees.Proceedings IUFRO Conference, Pattaya,Thailand, November 1988, 431-434. OxfordForestry Institute, Oxford.
Delwaulle, J.C. and Laplace, Y. 1988. La cultureindustrielle de l’Eucalyptus en RépubliquePopulaire du Congo. Bois et Forêts desTropiques no. 216: 35-42.
Dembi, F. 1988. Effet de l’élimination de lavégétation concurente sur l’humidité du sol etsur la croissance initiale d’une plantationmonoclonale d’Eucalyptus 12 ABL x saligna
(Clone L2-109). Mémoire d’étude IDR,Université de Brazzaville. (unpublished).
Dowsett-Lemaire, F. and Dowsett, R.J. 1991. Theavifauna of the Kouilou bassin in Congo. In:Dowsett-Lemaire, F. and Dowsett, R.J. (eds.)Flore et faune du Kouilou et leur exploitation,189-239. Touraco Research Report 4. Touraco,Liège.
Duncan, R.S. and Chapman, C.A. 1999. Seeddispersal and potential forest succession inabandoned agriculture in Africa. EcologicalApplications 9: 998-1007.
Dye, P.J. 1996. Climate, forest and streamflowrelationships in South African afforestedcatchments. Commonwealth Forestry Review75: 31-38.
Ellis, R.C. 1971. The mobilisation of iron byextracts of Eucalyptus leaf litter. Journal of SoilSciences 22: 8-22.
Enright, N.J. 1978. The interrelationship betweenplant species distribution and properties of soilsundergoing podzolization in a coastal area ofS.W. Australia. Australian Journal of Ecology3: 389-401.
Fargette, M. 1987. Use of esterase phenotype inthe taxonomy of genus Meloïdogyne. 2.Esterase phenotype observed in west Africanpopulations and their characterisation. Revuede Nématologie 10: 45-56.
Feller, C., Fritsch, E., Poss, R. and Valentin, C.1991. Effet de la texture sur le stockage et ladynamique des matières organiques dansquelques sols ferrugineux et ferralitiques(Afrique de l’Ouest en particulier). CahiersORSTOM, serie Pedologie 26: 25-36.
Ferris, H., Venette, R.C., van der Meulen, H.R.and Lau, S.S. 1998. Nitrogen mineralizationby bacterial-feeding nematodes: verificationand measurement. Plant and Soil 203: 159-171.
Food and Agriculture Organization (FAO). 1982.Les Eucalyptus dans les reboisements.Collection FAO Forêt. 753p.

65Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Foresta, (de), H., 1990. Origine et évolution dessavanes intramayombiennes (R.P. du Congo).Apports de la botanique forestière. In :Lanfranchi, R. and Schwartz, D. (eds.)Paysages quaternaires de l’Afrique centrale,326-335. ORSTOM, Paris.
Fox, R.H., Myers, R.J.K. and Vallis, I. 1990. Thenitrogen mineralization rate of legume residuesin soil as influenced by their polyphenol, lignin,and nitrogen contents. Plant and Soil 129: 251-259.
Gbadegesin, R.A. 1993. Effect on thedevelopment of Pinus caribaea seedlings ofcertain plant parasitic nematodes from thesavanna area of Nigeria. Forest Ecology andManagement 60: 163-166.
Geldenhuys, C.J. 1997. Native forest regenerationin pine and eucalypt plantations in NorthernProvince, South Africa. Forest Ecology andManagement 99: 101-116.
Gibson, A.H., Halsall, D.M. and Roper, M.M.1988. Nitrogen fixation associated withresidues breakdown. In: Bothe, H., de Bruijn,F.J., Newton, W.E. (eds.) Nitrogen fixation:hundred years after, 753-758. Gustav FischerVerlag, Stuttgart, Germany.
Gilot, C., Lavelle, P., Blanchard, E., Keli, J.,Kouassi, P., Guillaume, G. 1995. Biologicalactivity of soil under rubber plantations in Côted’Ivoire. Acta Zoologica Fennica 196: 186-189.
Goede (de), R.G.M., Georgieva, S.S., Verschoor,B.C. and Kamerman, J.W. 1993. Changes innematode community structure in a primarysuccession of blown-out areas in a drift sandlandscape. Fundamental and AppliedNematology 16: 501-513.
Gonçalves, J.L.M., Barros, N.F., Nambiar, E.K.S.and Novals, R.F. 1997. Soil and standmanagement for short-rotation plantations. In:Nambiar, E.K.S. and Brown, A.G. (eds.)Management of soil, nutrients and water intropical plantation forests, 379- 418. ACIARMonograph 43. ACIAR, Canberra.
Gonçalves, J.L.M., Poggiani, F. and Stape, J.L.1999. Eucalypt plantations in the humidTropics: São Paulo, Brazil. In: Nambiar,E.K.S., Cossalter, C. and Tiarks, A. (eds.) Sitemanagement and productivity in tropicalplantation forests: workshop proceedings, 16-20 February 1998, Pietermaritzburg, SouthAfrica, 5-12. CIFOR, Bogor, Indonesia.
Gonzales, G. and Seastedt, T.R. 2000. Comparisonof the abundance and composition of litterfauna in tropical and subalpine forests.Pedobiologia 44: 545-555.
Granhall, U. 1978. Environmental role of N2-fixing blue-green algae and asymbioticbacteria. Ecological Bulletin no. 26. 391p.
Groethals, M. 1987. Répartition et pante hôte desnématodes à galles (Meloidogyne spp.) auBurundi. In: Séminaire sur les ravageurs desprincipales cultures vivrières d’Afriquecentrale, Bujumbura, 16-20 février 1987, 367-375. CTA, Wageningen, The Netherlands.
Grove, T.S., Thomson, B.D. and Malajczuk, N.,1996. Nutritional physiology of eucalypts:uptake, distribution and utilization. In : Attiwill,P.M. and Adams, M.A. (eds.) Nutrition ofeucalypts, 77-108. CSIRO Publishing,Collingwood, Australia.
Harborne, J.B. 1997. Role of phenolic secondarymetabolites in plants and their degradation innature. In: Cadisch, G. and Giller, K.E. (eds.)Driven by nature: plant litter quality anddecomposition, 67-74. CAB International,Oxford.
Hardy, R.W.F., Holsten, R.D., Jackson, E.K. andBurns, R.C. 1968. The acetylene-ethyleneassay for N2 fixation : laboratory and fieldevaluation. Plant Physiology 43: 1185-1207.
Harmand, J.M. 1998. Rôle des espèces ligneusesà croissance rapide dans le fonctionnementbiogéochimique de la jachère. Effets sur larestauration de la fertilité des sols ferrugineuxtropicaux. (Bassin de la Bénoué au NordCameroun). Thesis in tropical plant biology andecology. Université Paris.

66 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Harrington, R.A. and Ewel, J.J. 1997. Invasibilityof tree plantations by native and non-indigenous plant species in Hawaii. ForestEcology and Management 99: 153-162.
Heal, O.W. and Dighton, J. 1985. Resource qualityand trophic structure in the soil system. In:Fitter, A.H., Atkinson, D., Read, D.J. andUsher, M.B. (eds.) Ecological interaction insoil, 339-354. Blackwell Scientific, Oxford,England.
Herbert, M.A. 1996. Fertilizers and eucalyptplantations in South Africa. In: Attiwill, P.M.and Adams, M.A. (eds.) Nutrition of eucalypts,303-325. CSIRO Publishing, Collingwood,Australia.
International Foundation for Science (IFS). 1989.Proceedings of a regional seminar on trees fordevelopment in sub-Saharan Africa, Nairobi(Kenya). Nairobi, Kenya. 361p.
Jaiyeoba, I.A. 1998. Change in soil propertiesrelated to conversion of savannah woodlandinto pine and eucalyptus plantations, NorthernNigeria. Land Degradation and Development9: 207-215.
Jamet, R. 1975. Evolution des principalescaractéristiques des sols des reboisements dePointe-Noire. Orstom, Cote MC 189, Centrede Brazzaville, décembre 1975. 35p.(unpublished).
Kallarackal, J. and Somen, C.K. 1997. Anecophysiological evaluation of the suitabilityof Eucalyptus grandis for planting in thetropics. Forest Ecology and Management 95:53-61.
Keenan, R., Lamb, D., Woldring, O., Irvine, A.and Jensen, R. 1997. Restoration of plantdiversity beneath tropical tree plantations innorthern Australia. Forest Ecology andManagment 99: 117-132.
King, J.A. and Campbell, B.M. 1994. Soil organicmatter relations in five land cover types in themiombo region (Zimbabwe). Forest Ecologyand Management 67: 225-239.
Kirkpatrick, J.B. 1997. Vascular plant-eucalyptinteractions. In: Williams J.E. and WoinarskiJ.C.Z. (eds.) Eucalypt ecology: individuals toecosystems, 227-245. Cambridge UniversityPress, Cambridge, England.
Knockaert, C. 1981. Production de littiere dansquatre plantations d’Eucalyptus camaldulensiset dans un peuplement naturel de Quercussuber. Annales de la Recheche Forestière auMaroc 21: 351-373.
Knowles, R., Brouzes, R. and O’Toole, P. 1973.Kinetics of nitrogen fixation and acethylenereduction, and effects of oxygen and ofacethylene on these processes, in a soil system.Bulletin of Ecological ResearchCommunications 17: 255-262.
Laclau, J.P., Bouillet, J.P. and Ranger, J. 2000aDynamics of biomass and nutrientaccumulation in a clonal plantation ofEucalyptus in Congo. Forest Ecology andManagement 128: 181-196.
Laclau, J.P., Bouillet, J.P. and Ranger, J. 2000b.Canopy and soil modification of precipitationchemistry in a clonal eucalypt plantation in theCongo. Comparison with an adjacent savannaecosystem. Proceedings of the XXI IUFROWorld Congress, Kuala Lumpur, Malaysia.
Laclau, J.P., Ranger, J., Nzila, J.D. and Bouillet,J.P. 2000c. Nutrient cycling in a short rotationEucalyptus plantation and adjacent savanna inCongo. In: Nambiar, E.K.S., Tiarks, A.,Cossalter, C. and Ranger, J. (eds.) Sitemanagement and productivity in tropicalplantation forests: a progress report: Workshopproceedings 7-11 December 1999, Kerala,India, 95-99. CIFOR, Bogor, Indonesia.
Lamb, D. 1998. Large scale ecological restorationof degraded tropical forest lands: the potentialrole of timber plantations. Restoration Ecology6: 271-279.
Lamotte, M. 1979. Secondary production:consumption and decomposition. In: Tropicalgrazing land ecosystems, 146-208. UNESCONatural Resource Research 16. UNESCO.

67Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Lavelle P. 1996. Diversity of soil fauna andecosystem function. Biology International 33:3-16.
Lavelle, P., Bignell, D., Lepage, M., Wolters, V.,Roger, P., Ineson, P., Heal, O.W. and Dhillion,S. 1997. Soil function in a changing world:the role of invertebrate ecosystem engineers.European Journal of Soil Biology 33: 159-193.
Lavelle, P., Dangerfield, M., Fragoso, C.,Eschenbrenner, V., Lopez-Hernandez, D.,Pashani, B. and Brussaard, L. 1994. Therelationship between soil fauna and tropical soilfertility. In: Woomer, P.L. and Swift, M.J. (eds.)The biological management of tropical soilfertility, 137-169. TSBF, Wiley, Chichester,England.
Lawton, J.H., Bignell D.E., Bolton, B., BloemersG.F., Eggleton, P., Hammond, P.M., Hodda, M.,Holt, R.D., Larsen, T.B., Mawdsley, N.A.,Stork, N.E., Srivastava, D.S. and Watt, A.D.1998. Biodiversity inventories, indicator taxaand effect of habitat modification in tropicalforest. Nature 391: 72-76.
Lemieux, G. 1996. Le bois raméal et lapédogénèse: une influence agricole et forestièredirecte. Publication No.15, 2ième édition. http://forestgeomat.for.ulaval/
Li, H. and Madden, J.L. 1995. Analysis of leafoils from a Eucalyptus species trial.Biochemical Systematic and Ecology 23: 167-177.
Lisanework, N. and Michelsen, A. 1993.Allelopathy in agroforestry systems: the effectsof leaf extracts of Cupressus lusitanica andthree Eucalyptus spp. on four Ethiopian crops.Agroforestry 21: 63-74.
Lopez-Fando, C. and Bello, A. 1995. Variabilityin soil nematode populations due to tillage andcrop rotation in semi-arid Mediterraneanagrosystems. Soil and Tillage Research 36: 59-72.
Loubana, P.M. 1996. Survey of nematodes of thegenus Meloidogyne parasitic on crops inCongo. Nematropica 26: 287.
Loubelo, E. 1990. Etude comparative de quelqueséléments du fonctionnement de deuxpeuplements d’Eucalyptus au Congo. Thesis,Université de Rennes.141p.
Loumeto, J.J. 1986. Contribution à l’étude de ladistribution minérale dans les Eucalyptus duCongo. Thesis, Université de Rennes.
Loumeto, J.J. and Huttel, C. 1997. Understoryvegetation in fast-growing tree plantations onsavanna soils in Congo. Forest Ecology andManagement 99: 65-81.
Luc, M. and Reversat, G. 1985. Possibilités etlimites des solutions génétiqures aux affectionsprovoquées par les nématodes sur les culturestropicales. C.R. des Séances de l’Académied’Agriculture de France 71: 781-791.
Lugo, A.E. 1997. The apparent paradox ofreestablishing species richness on degradedlands with tree monocultures. Forest Ecologyand Management 99: 9-19.
Lugo, A.E., Cuevas, E. and Sanchez, M.J. 1990.Nutrients and mass in litter and top soil of tentropical tree plantations. Plant and Soil 125:263-280.
Lugo, A.E., Parrotta, J. and Brown, S. 1993. Lossin species caused by tropical deforestation andtheir recovery through management. Ambio 22:106-109.
Madeira, M.A.V., Andreux, F. and Portal, J.M.1989. Changes in soil organic mattercharacteristics due to reforestation withEucalyptus globulus in Portugal. The Scienceof the Total Environment 81/82: 481-488.
Maity, S.K. and Joy, V.C. 1999. Impact ofantinutritional compounds of leaf litter ondetritivore soil arthropod fauna. Journal ofEcobiology 11: 193-202.
Majer, J.D., Recher, H.F., Wellington, A.B.,Woinarski, J.C.Z. and Yen, A.L. 1997.Invertebrates of eucalypt formations. In:Williams, J.E. and Woinarski, J.C.Z. (eds.)Eucalypt ecology: individuals to ecosystems,278-302. Cambridge University Press,Cambridge, England.

68 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Marais, M. and Buckley, N.H. 1993. Plantparasitic nematodes in forest soils in theTransvaal and Natal, South Africa. SouthAfrican Forestry Journal 166: 9-16.
Mboukou-Kimbatsa, I.M.C., Bernhard-Reversat,F. and Loumeto, J.J. 1998. Change in soilmacrofauna and vegetation when fast growingtrees are planted on savanna soils. ForestEcology and Management 110: 1-12.
Michelsen, A., Lisanework, N., Friis, I. and Holst,N. 1996. Comparisons of understoreyvegetation and soil fertility in plantations andadjacent natural forests in the Ethiopianhighlands. Journal of Applied Ecology 33: 627-642.
Miranda, G.A. de, Barros, N.F. de, Leite, H.G.,Couto, L., Teixeira, J.L. de, -Miranda, G.A. deand Barros, N.F. 1998. Yield of eucalyptstands managed by coppicing, as influencedby fertilizing and slash disposal in theJequitinhonha Valley, Minas Gerais, Brazil.Revista Arvore 22: 307-314.
Morais, E.J., Barros, N.F., Novais, R.F. and Brandi,R.M. 1990. Biomassa e eficiência nutricionalde espécies de eucalipto em duas regioesbioclimaticas de Minas Gerais. RevistaBrasileira de Ciência do Solo 14: 353-362.
Nandi, A., Basu, P.K. and Banerjee, S.K. 1991.Modification of some soil properties byEucalyptus species. Indian Forester 117: 53-57.
Negi, J.D.S. and Sharma, S.C. 1996. Mineralnutrition and resource conservation inEucalyptus plantations and other forest coversin India. In: Attiwill, P.M. and Adams, M.A.(eds.) Nutrition of eucalypts, 399-416. CSIROPublishing, Collingwood, Australia.
Nicholas, W.L. 1975. The biology of free-livingnematodes. Clarendon Press, Oxford, England.219p.
Nizinski, J., Loumeto, J.J., Ayoma, M.-B.,Mabiala-N’goma, A. and Nsemi, R.N. In press.Bilan d’eau du sol d’une savane et d’une
plantation d’Eucalyptus dans le bassin duKouilou (Congo). Science et ChangementsPlanétaires – Sécheresse.
Nongamani, A. 1988. Dynamique de la végétationet des sols sous forêts d’Eucalyptus et de pins.Cas de Loudima. Mémoire de Diplome del’Institut de Développement rural, Universitéde Brazzaville, 98p. (unpublished).
Nyathi, P., Campbell, B. and Maghembe, J.A.1991. Leaf quality of Sesbania sesban,Leucaena leucocephala, and Brachystegiaspiciformis: potential agroforestry species.Forest Ecology and Management 64: 259-264.
Nzila, J.D. 1992. Etude des transformationsstructurales et physico-chimiques d’un solferrallitique acide de la vallée du Niari (Congo)soumis à la pratique de l’écobuage. Thesis,University of Paris 12.
O’Connell, A.M. and Grove. T.S. 1987. Seasonalvariation in C
2H
2 reduction (N
2-fixation) in the
litter layer of eucalypt forests of south-westernAustralia. Soil Biology and Biochemistry 19:135-142.
O’Connell, A.M. and Grove, T.S. 1996. Biomassproduction, nutrient uptake and nutrient cyclingin the jarrah (Eucalyptus marginata) and karri(Eucalyptus diversicolor) forest of south-western Australia. In: Attiwill, P.M. and Adams,M.A. (eds.) Nutrition of eucalypts, 155-190.CSIRO Publishing, Collingwood, Australia.
O’Connell, A.M. and Sankaran, K.V. 1997.Organic matter accretion, decomposition andmineralisation. In: Nambiar, E.K.S. and Brown,A.G. (eds.) Management of soil, nutrients andwater in tropical plantation forests, 443-480.ACIAR Monograph No. 43. ACIAR, Canberra.
Palm, C.A. and Rowland, A.P. 1997. A minimumdataset for characterization of plant quality fordecomposition. In: Cadisch, G. and Giller, K.E.(eds.) Driven by nature: plant litter quality anddecomposition, 379-392. CAB International,Oxford, England.

69Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Pandey, D. 1997. Hardwood plantations in thetropics and subtropics. Tropical forestplantations area in 1995. CIFOR OccasionalPaper. CIFOR, Bogor, Indonesia. 76p.
Paoletti, M.G. 1999. The role of earthworms forassessment of sustainability and asbioindicator. Agriculture, Ecosystems andEnvironment 74: 137-135.
Parrotta, J.A. 1993. Secondary forest regenerationon degraded tropical lands. The role ofplantations as ‘foster ecosystems’. In: Lieth,H. and Lohman, M. (eds.) Restoration oftropical forest ecosystem, 63-73. KluwerAcademic Publishers, Netherlands.
Parrotta, J.A. 1995a. Influence of overstorycomposition on understory colonisation bynative species in plantations on degradedtropical site. Journal of Vegetation Science 6:627-636.
Parrotta, J.A. 1995b. The catalytic effect of treeplantings on the rehabilitation of native forestbiodiversity on degraded tropical lands: anexploratory analysis. Methodologies, fieldprotocols and schedule. The World Bank,USDA Forest Service, IITF, ODA (UK),CIFOR. 20p (unpublished).
Parrotta, J.A. 1999. Productivity, nutrient cycling,and succession in a single - and mixed-speciesplantations of Casuarina equisetifolia,Eucalyptus robusta and Leucaenaleucocephala in Puerto Rico. Forest Ecologyand Management 124: 45-77.
Parrotta, J.A. and Knowles, O.H. 1999.Restoration of tropical moist forests on bauxite-mined lands in the Brazilian Amazon.Restoration Ecology 7: 103-116.
Parrotta, J.A., Baker. D.D. and Fried, M. 1996.Changes in dinitrogen fixation in maturingstands of Casuarina equisetifolia and Leucaenaleucocephala. Canadian Journal of ForestResearch 26: 1684-1691.
Parrotta, J., Knowles, O.H. and Wunderle, J.M. jr.1997. Development of floristic diversity in 10-
year-old restoration forests on a bauxite minedsite in Amazonia. Forest Ecology andManagement 99: 21-42.
Parrotta, J.A., Turnbull, J.W. and Jones, N. 1997.Catalyzing native forest regeneration ondegraded tropical lands. Forest Ecology andManagement 99: 1-8.
Pendge, G. 1994. Fruit consumation and seeddispersal by the African civet Civectictis civettain Mayombe, Zaïre. Revue d’Ecologie 49: 107-116.
Petit, L.J., Petit, D.R., Christian, D.G. and Powell,H.D.W. 1999. Bird communities in natural andmodified habitats in Panama. Ecography 22:292-304.
Pohlman, A.A. and McColl, J.G. 1988. Solubleorganics from forest litter and their role in metaldissolution. Soil Science Society of AmericaJournal 52: 265-271.
Polglase, P.J. and Attiwill, P.M. 1992. Nitrogenand phophorus cycling in relation to stand ageof Eucalyptus regnans F. Muell. I. Return fromplant to soil in litterfall. Plant and Soil 142:157-166.
Poore, M.E.D. and Fries, C. 1985. The ecologicaleffect of Eucalyptus. F.A.O. Forestry Paper 59.FAO, Rome. 87p.
Porazinska, D.L., Duncan, L.W., McSorley, R. andGraham, J.H. 1999. Nematode communitiesas indicators of status and processes of a soilecosystem inflenced by agriculturalmanagement practices. Applied Soil Ecology13: 69-86.
Powers, J.S., Haggar, J.P. and Fisher, R.F. 1997.The effect of overstory composition onunderstory woody regeneration and speciesrichness in 7-year-old plantations in CostaRica. Forest Ecology and Management 99: 43-54.
Pradhan, G.B., Senapati, B.K. and Dash, M.C.1988. Relationship of soil nematodepopulation to carbon/nitrogen in tropical

70 Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
habitat and their role in the laboratorydecomposition of litter amendments. Revued’Ecologie et de Biologie du Sol 25: 59-76.
Reigosa, M.J., Sanchez-Moreiras, A. andGonzales, L. 1999. Ecophysiological approachin allelopathy. Critical Review in Plant Science18: 577-608.
Reversat, G. 1981. Age related changes in thechemical oxygen demand of second stagejuveniles of Meloidogyne javanica andHeterodera oryzae. Nematologica 27: 220-227.
Reversat, G. 1996. Anhydrobiosis in plantparasitic dorylaimids. Nematropica 26: 218-219.
Roger, P.A. and Ladha, J.K. 1992. Biologicalnitrogen fixation in wetland ricefields:estimation and contribution to nitrogenbalance. Plant and Soil 141: 41-55.
Safou-Matondo, R., Laclau, J.P. and Bouillet, J.P.1999. Etude de biomasse sur quatre clonesd’Eucalyptus urophylla x Eucalyptus grandis:établissement de tarifs pour différentscompartiments aériens et exploration de lavariabilité inter-clonale. UR2PI report. 15p.(unpublished).
Salter, P.J. 1996. The interaction of birdcommunities with vegetation in Brisbane forestPark. Emu 95: 194-207.
Sankaran, K.V. et al. 2000. Effects of sitemanagement on Eucalyptus plantations in themonsoonal tropics. In: Nambiar, E.K.S., Tiarks,A., Cossalter, C. and Ranger, J. (eds.) Sitemanagement and productivity in tropicalplantation forests: a progress report: Workshopproceedings 7-11 December 1999, Kerala,India, 51-60. CIFOR, Bogor, Indonesia.
Schulze, E.D. and Mooney, H.A. 1993.Biodiversity and ecosystem function.Ecological Studies 99. Springer Verlag, Berlin.525p.
Scott, D.F. and Smith, R.E. 1997. Preliminaryempirical models to predict reductions in totaland low flows resulting from afforestation.Water South Africa 23: 135-140.
Seinhorst, J.W. 1950. De betekenis van de toestandvan de grond voor het optreden van aantastingdoor het stengelaaltje (Ditylenchus dipsaci(Kühn) Filipjev). Tijdschrift overPlantenziekten 56: 289-348.
Seinhorst, J.W. 1955. Een eenvoudige methodevoor het afschieden van aaltjes uit grond.Tijdschrift over Plantenziekten 61: 188-190.
Silva (da), M.C. Jr., Scarano, F.R. and SouzaCardel, F. 1995. Regeneration of an atlanticforest formation in the understory of aEucalyptus grandis plantation in south-easternBrazil. Journal of Tropical Ecology 11: 147-152.
Smith, K.D. 1974. The utilization of gum-treesby birds in Africa. Ibis 116: 155-164.
Soler, C. 1994. Etude de la dynamique du recrûforestier en sous-bois d’eucalyptus. Universitédes Sciences Paul Sabatier/Orstom, Rapport deMaîtrise, Université de Toulouse. Unpublishedreport. 53p.
Souto, X.C., Gonzalez, L. and Reigosa, M.J. 1995.Allelopathy in forest environment in Galicia,NW Spain. Allelopathy Journal 2: 67-68.
Spaull,V.W. 1998. A note on the effect of using anematicide on sugar cane planted in a sandysoil previously cropped to Eucalyptus trees.Proceedings of the Annual Congress of theSouth African Sugar Technology Association72: 94-97.
Stewart, H.T.L. and Van der Lingen, S.A. 1988.Soil chemical properties under miombowoodland and eucalypt plantation on Kalaharisand in Zimbabwe. East African Agriculturaland Forestry Journal 52: 171-179.
Tassin, J. and Rivière, J.N. 1998. Évaluation del’impact des plantations forestières surl’avifaune: application au littoral réunionais.Bois et Forêts des Tropiques No. 258: 37-47.
Tian, G. 1998. Effect of soil degradation on leafdecomposition and nutrient release underhumid tropical conditions. Soil Science 163:897-906.

71Ef fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i tyE f fec t o f exot i c t ree p lanta t ions on p lant d ivers i ty and b io log ica l so i l f er t i l i ty
Trouvé, C., Mariotti, A., Schwartz, D. and Guillet,B. 1994. Soil organic carbon dynamics underEucalyptus and Pinus planted on savannas inthe Congo. Soil Biology Biochemistry 26: 287-295.
Turner, J. and Lambert, M.J. 1983. Nutrientcycling within a 27-year-old Eucalyptusgrandis plantation in New South Wales. ForestEcology and Management 6: 155-168.
Turner, J. and Lambert, M.J. 1996. Nutrientcycling and forest management. In: Attiwill,P.M. and Adams, M.A. (eds.) Nutrition ofeucalypts, 229-240. CSIRO Publishing,Collingwood, Australia.
Van Soest, P.J. 1963. Use of detergents in theanalysis of fibrous feeds. II. A rapid methodfor the determination of fibre and lignin.Journal of the Assistant Officers in AgriculturalChemistry 46: 829-835.
Vohland, K. and Schroth, G. 1999. Distributionpatterns of the litter macrofauna in agroforestryand monoculture plantations in centralAmazonia as affected by plant species andmanagement. Applied Soil Ecology 13: 57-68.
Wallace, H.R. 1973. Nematode ecology and plantdisease. Edward Arnold, London. 228p.
Wardle, D. and Lavelle, P. 1997. Linkagesbetween soil biota, plant litter quality anddecomposition. In: Cadisch, G. and Giller, K.E.(eds.). Driven by nature: plant litter quality anddecomposition, 107-125. CAB-International,Wallingford, England.
Waterman, P.G. and Mole, S. 1994. Analysis ofphenolic plant metabolites. Methods inEcology. Blackwell Scientific, London. 238p.
Wilson, J.V. and Wilson, B.L.P. 1991. La chassetraditionnelle et commerciale dans le sud ouestdu Congo. In: Dowsett-Lemaire, F. andDowsett, R.J. (eds.) Flore et faune du Kouilouet leur exploitation. Touraco Research Report4: 279-289. Touraco, Liège.
Wilson, R.A., Astorga, R., Gomez, C. andGonzalez-Rio, F. 1995. Papermaking withDNA. ‘Intelligent Fibre’. In: Potts, B.M.,Borralho, N.M.G., Reid, J.B., Cromer, R.N.,Tibbitts, W.N. and Raymond, C.A. (eds.)Proceedings of the CRCTHF-IUFROConference: Eucalypt plantations: improvingfibre yield and quality, 5-9. CooperativeResearch Centre for Temperate HardwoodForestry, Hobart, Australia.
Yeates, G.W. 1979. Soil nematodes in terrestrialecosystems. Journal of Nematology 11: 213-229.
Zou, X. 1993. Species effect on earthworm densityin tropical tree plantations in Hawaii. Biologyand Fertility of Soils 15: 35-38.
Zou, X. and Bashkin, M. 1998. Soil carbonaccretion and earthworm recovery followingrevegetation in abandoned sugar cane fields.Soil Biology and Biochemistry 30: 825-830.

JAPANOfficial Development Assistance
CIFORCENTER FOR INTERNATIONAL FORESTRY RESEARCH
The environmental impact of eucalypt plantations in particular on soil water and nutrients,understorey vegetation and biodiversity is a controversial topic. Commercial plantations,
mainly eucalypts, have been established on poor sandy soils of savannas in the Congo since1978. This report on the role of biological factors in soil fertility complements studies ofnutrient balance and logging residue management in progress in these plantations. Bioticprocesses in relation to plantation age and biotic changes in successive rotations wereinvestigated. Changes of understorey vegetation with age, received special attention.Relationships between vegetation and soil fertility, which are closely linked to litter and soilorganic matter quality and quantity; were studied in an age series of eucalypt hybrids andclones. The density of the main taxa of macrofauna and nematodes, which act through organicmatter decomposition, soil biogenic structures and parasitism, was assessed according toplantation age and logging history. Nitrogen-fixation by soil organisms was also assessed.Soil organic matter accumulation and quality were among the main ecological changes foundto occur when savanna was planted with trees, and determined the development of undergrowthand changes in soil characteristics. The differences in litter quantity and quality of eucalypt,acacia and pine plantations resulted in major differences in soil microfauna and macrofaunadensity and biomass, and suggested that litter quality of the tree crop took precedence overplant biodiversity in controlling soil biota. Short-rotation plantation forestry is a land usewith numerous positive effects in the Congolese savanna environment, although more researchis needed on longer-term effects of silvicultural practices on soil organic matter.





























