Embed Size (px)
Citation preview

Photochernistrv and Phofobiology Vol. 50, No. 2 , pp. 259-262. 1989 003 1 -X655/89 $03 .oo + 0.00 Copyright 0 19x9 Maxwell Pergamon Macmillan plc Printed in Great Britain. All rights reserved
DUAL EFFECTS OF TRYPTOPHAN IN THE HORSERADISH PEROXIDASE SYSTEM THAT
GENERATES TRIPLET ACETONE
EDUARDO S I L V A ~ * and GIUSEPPE CILENTO? ‘Facultad de Quimica, Pontificia Universidad Cat6lica de Chile, Casilla 6177, Santiago, Chile and
‘Departamento de Bioquimica, Instituto de Quimica, Universidade de Sio Paulo, C.P. 30780, 01498, SBo Paulo, Brazil
(Receiivd 21 November 1988; accepted 24 February 1989)
Abstract-When tryptophan is added to the horseradish peroxidaseihydrogen peroxide system, it markedly increases the rate of ‘spontaneous’ conversion of peroxidase-compound 11 to the native form without undergoing any alteration by spectral criteria. The o-isomer is more efficient than the L- isomer. The latter binds to the enzyme compound 11, although at a site not very favourable for reduction. The accelerating effect of tryptophan is also seen when isobutyraldehyde or its enol added to the system, is converted into triplet acetone. Again the o-isomer ia more efficient than the L- isomer. The o-isomer is also more efficient in quenching the acetone phosphorescence. These results indicate that, at least in the case of 1.-tryptophan where complexation with compound I1 occurs, the interaction of tryptophan with triplet acetone bound to the enzyme takes place in a ternary complex.
INTRODUCTION
The interaction of D- and L-tryptophan with enzyme- generated and shielded triplet carbonyls shows a marked chiral discrimination as inferred from the quenching of the carbonyl phosphorescence (Rivas- Suarez and Cilento, 1981; Adam ef nl., 1986). Con- comitantly, trytophan is converted to oxidation products, as occurs photochemically (Rivas et al., 1984). The existence of chiral discrimination has permitted us to infer that tryptophan ‘sees’ triplet acetone still bound to the enzyme and that triplet acetone transfers energy to suitable acceptors while still on the enzyme (Adam et al., 1986).
The present note concerns the detailed role of tryptophan
MATERIALS AND METHODS
Horseradish peroxidase (HRP; type VI)?, L-kynuren- ine, o-tryptophan and L-tryptophan were obtained from Sigma Chemical Co. (St Louis, MO, USA); isobutyral- dehyde from Carlo Erba Ind. Quim. (SLo Paulo, Brazil) was purified ’by distillation under nitrogen. Horseradish peroxidase ( E , , ~ ) ~ = 1.02 Y 105 M - ’ cm-l (Ohlsson and Paul, 1976) was determined spectrophotometrically and HzOz by the peroxidase assay (Cotton and Dunford, 1973). Oxygen consumption was followed with a Yellow Springs Instruments Model 53 Oxygen Monitor. Absorp- tion spectra were recorded and kinetic studies performed on a Zeiss DMR 31 spectrometer.
The standard system for triplet acetone generation was prepared as follows: 10 p 8 of an aqueous solution of 0.5 mM HRP was added to a mixture of 20 pC of 0.1 M EDTA, 40 pt of 3.1 lo-’ M HzOz, 1 m8 of 2.1 M L- or o-tryptophan or kynurenine, and 1.8 mt of 1.0 M phosphate buffer, pH 7.4. The reaction was initiated by addition of 0.1 m t of a stock solution of 2.2 M isobutyraldehyde in ethanol.
* To whom correspondence should bc addressed. t Abbrnliutions: EDTA, ethylenediaminetetraacetate;
~~~ ~ .. ~~~ ~~~~~~~
HRP, horseradish peroxidase; Trp, tryptophan.
RESULTS
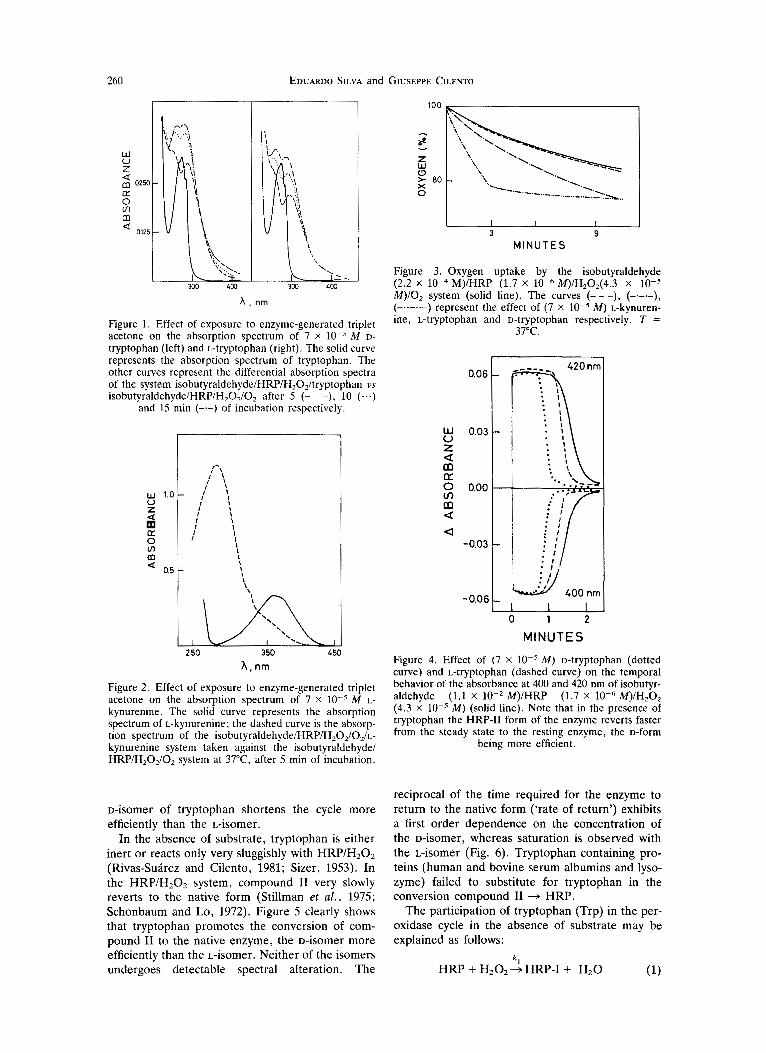
The ‘dark photochemical degradation’ of D - and L-tryptophan, when subjected to the action of triplet acetone generated during the H R P catalyzed aero- bic oxidation of isobutyraldehyde, produces spectral modifications on these indole compounds (Fig. 1). In line with the observed chiral discrimination (Riv- as-Suarez and Cilento, 1981; Adam et al . , 1986), larger alterations were found in the presence of D-
tryptophan. The generation of N-formylkynurenine and kynurenine, as some of the reaction products, was detected on the basis of the corresponding fluo- rescence spectra (Fukunaga et al., 1982). Figure 2 shows that kynurenine itself is altered when exposed to the isobutyraldehyde HRP/HzOz/Oz system. It is likely that the kynurenine alteration is the result of interaction with triplet acetone; thus, the phosphor- escence of the latter is quenched with a Stern-Volmer constant of 5.0 ). lo4 M-I . More- over, kynurenine was not altered when exposed to HRP/H202 during at least 50 min. Figure 3 shows that oxygen uptake by D-tryptophan is faster than by the L-isomer, a chiral discrimination in line with Stern-Volmer data.
For a better understanding of the behavior of tryptophan in the enzymatic system that generates triplet acetone, it is necessary to examine its effect on the peroxidase cycle, both in presence and absence of isobutyraldehyde. Figure 4 presents the effect of both isomers when the substrate was pre- sent. It is well known that changes in A400 reflect changes in the concentration of native HRP; those in A42,, monitor changes in the concentration of compound 11, whereas 411 nm represents an isosb- estic point for the HRPIHRP-I1 conversion. Com- pound I arises from reaction of H R P with H202 and is rapidly transformed into compound 11. The
259
260 EDUARDO SILVA and GIUSEPPE CILENTO
- 1
A , nm
Figure 1. Effect of exposure to enzyme-generated triplet acetone on the absorption spectrum of 7 x M D- tryptophan (left) and L-tryptophan (right). The solid curve represents the absorption spectrum of tryptophan. The other curves represent the differential absorption spectra of the system isobutyraldehydelHRP/H20z/tryptophan vs isobutyraldehyde/HRP/H202/Oz after 5 (- - -), 10 (...)
and 15 min (--) of incubation respectively.
w 1.0 u z a a: 0 Y,
a 0.5 m
I \
I 1 !
h,nm
Figure 2. Effect of exposure to enzyme-generated triplet acetone on the absorption spectrum of 7 x M L- kynurenine. The solid curve represents the absorption spectrum of L-kynurenine; the dashed curve is the absorp- tion spectrum of the isobutyraldehyde/HRP/H20z/0,/L- kynurenine system taken against the isobutyraldehyde/ HRP/H,O,/O, system at 37"C, after 5 min of incubation.
D-isomer of tryptophan shortens the cycle more efficiently than the L-isomer.
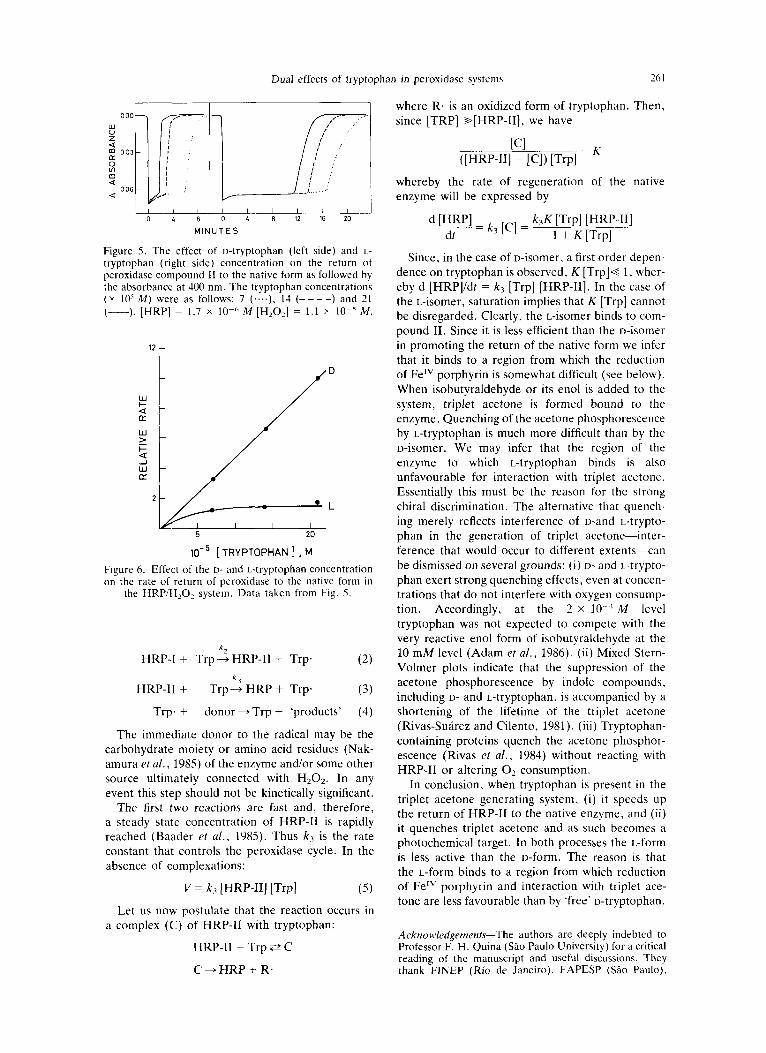
In the absence of substrate, tryptophan is either inert o r reacts only very sluggishly with HRP/H202 (Rivas-Sufirez and Cilento, 1981; Sizer, 1953). In the HRP/H202 system, compound I1 very slowly reverts to the native form (Stillman et al., 1975; Schonbaum and Lo, 1972). Figure 5 clearly shows that tryptophan promotes the conversion of com- pound I1 to the native enzyme, the D-isomer more efficiently than the L-isomer. Neither of the isomers undergoes detectable spectral alteration. The
- 3 9
M I N U T E S
Figure 3. Oxygen uptake by the isobutyraldehyde
M)lO, system (solid line). The curves (---), (---), (-. .-. .-) represent the effect of (7 x M ) L-kynuren- ine, L-tryptophan and D-tryptophan respectively. T =
37°C.
(2.2 x 10-4 M)/HRP (1.7 x 10-6 M)/H,o,(~.~ x 10-5
W 0 z
[12 0 m
a m
m a a
0.03 -
. . \ 0.00
-0.03 -
-0.06 - I I
0 1 2
MINUTES
Figure 4. Effect of (7 X lo-' M ) D-tryptophan (dotted curve) and L-tryptophan (dashed curve) on the temporal behavior of the absorbance at 400 and 420 nm of isobutyr- aldehyde (1.1 X lo-' M)/HRP (1.7 x lo-' M)IH,O, (4.3 x lo-' M ) (solid line). Note that in the presence of tryptophan the HRP-I1 form of the enzyme reverts faster from the steady state to the resting enzyme, the D-form
being more efficient.
reciprocal of the time required for the enzyme to return to the native form ('rate of return') exhibits a first order dependence on the concentration of the D-isomer, whereas saturation is observed with the L-isomer (Fig. 6). Tryptophan containing pro- teins (human and bovine serum albumins and lyso- zyme) failed to substitute for tryptophan in the conversion compound I1 -+ HRP.
The participation of tryptophan (Trp) in the per- oxidase cycle in the absence of substrate may be explained as follows:
kl H R P + H20, -+ HRP-I + HzO (1)
Dual effects of tryptophan in peroxidase systems 26 1
I l l I J 0 4 8 0 4 8 1 2 1 6 2 0
M I N U T E S
Figure 5. The effect of D-tryptophan (left side) and L- tryptophan (right side) concentration on the return of peroxidase compound I1 to the native form as followed by the absorbance at 400 nm. The tryptophan concentrations ( y 10’ M ) were as follows: 7 (....), 14 (- - - -) and 21 (-). [HRP] = 1.7 x lo-” M [H202] = 1.1 lo-’ M .
12 - I
W I-
CL
W
I-
W CT
a
L
4
2
[TRYPTOPHAN I , M
Figure 6. Effect of the D- and L-tryptophan concentration on the rate of return of peroxidase to the native form in
the HRP/H,O, system. Data taken from Fig. 5.
k2 HRP-I + Trp + HRP-I1 + Trp. (2)
HRP-I1 + Trp+ H R P + Trp. (3) k 3
Trp. + donor + Trp + ‘products’ (4)
The immediate donor to the radical may be the carbohydrate moiety or amino acid residues (Nak- amura e ta / . , 1985) of the enzyme and/or some other source ultimately connected with H z 0 2 . In any event this step should not be kinetically significant.
The first two reactions are fast and, therefore, a steady state concentration of HRP-I1 is rapidly reached (Baader et a / . , 1985). Thus k3 is the rate constant that controls the peroxidase cycle. In the absence of complexations:
V = k3 [HRP-IIJ [Trp] ( 5 )
Let us now postulate that the reaction occurs in
HRP-I1 + Trp Fz C
C - H R P t R .
a complex (C) of HRP-I1 with tryptophan:
where R. is an oxidized form of tryptophan. Then, since [TRP] +[HRP-111, we have
[CI ~ ~~
([HRP-II] - [C]) [Trp] = ‘ whereby the rate of regeneration of the native enzyme will be expressed by
k3K [Trp] [HRP-11] = k3 [(‘I = L- d [HRP] ~.
dt 1 + K[Trp]
Since, in the case of D-isomer, a first order depen- dence on tryptophan is observed, K [Trp]< 1, wher- eby d [HRPIidt = k3 [Trp] [HRP-111. In the case of the L-isomer, saturation implies that K [Trp] cannot be disregarded. Clearly, the L-isomer binds to com- pound 11. Since it is less efficient than the D-isomer in promoting the return of the native form we infer that it binds to a region from which the reduction of Fe’” porphyrin is somewhat difficult (see below). When isobutyraldehyde or its enol is added to the system, triplet acetone is formed bound to the enzyme. Quenching of the acetone phosphorescence by L-tryptophan is much more difficult than by the D-isomer. We may infer that the region of the enzyme to which L-tryptophan binds is also unfavourable for interaction with triplet acetone. Essentially this must be the reason for the strong chiral discrimination. The alternative that quench- ing merely reflects interference of D-and L-trypto- phan in the generation of triplet acetone-inter- ference that would occur to different extents--can be dismissed on several grounds: ( i ) D- and I>-trypto- phan exert strong quenching effects, even at concen- trations that do not interfere with oxygen consump- tion. Accordingly, at the 2 X M level tryptophan was not expected to compete with the very reactive enol form of isobutyraldehyde at the 10 mM level (Adam et al . , 1986). ( i i ) Mixed Stern- Volmer plots indicate that the suppression of the acetone phosphorescence by indole compounds, including D- and L-tryptophan, is accompanied by a shortening of the lifetime of the triplet acetone (Rivas-Suarez and Cilento, 1981). (iii) Tryptophan- containing proteins quench the acetone phosphor- escence (Rivas et a/ . , 1981) without reacting with HRP-I1 or altering O2 consumption.
In conclusion, when tryptophan is present in the triplet acetone generating system, (i) it speeds up the return of HRP-I1 to the native enzyme, and (ii) it quenches triplet acetone and as such becomes a photochemical target. In both proces3es the L-form is less active than the D-form. The reason is that the L-form binds to a region from which reduction of Fe” porphyrin and interaction with triplet ace- tone are less favourable than by ‘free’ D-tryptophan.
AcknoM’ledgements-The authors are deeply indebted to Professor F. H. Quina (SBo Paulo University) for a critical reading of the manuscript and useful discussions. They thank FINEP (Rio de Janeiro), FAPESP (Sio Paulo),

262 EDUARDO SILVA and GIUSEPPE CILENTO
CODAC University of Slo Paulo, CNPq (Brasilia), the Volkswagen Foundation (Hanover) and GTZ (Eschborn) for financial support.
REFERENCES Adam, W., W. J. Baader and G. Cilento (1986) Enols of
aldehydes in the peroxidaseioxidase-promoted gener- ation of excited triplet species. Biochim. Biophys. Acta 881, 330-336.
Baader, W. J., C. Bohne, G . Cilento and H. B. Dunford (1985) Peroxidase-catalyzed formation of triplet acetone and chemiluminescence from isobutyraldehyde and mol- ecular oxygen. J . Biol. Chem. 260, 10217-10225.
Cotton, M. L. and H. B. Dunford (1973) Studies on horseradish peroxidase. XI. On the nature of com- pounds I and I1 as determined from the kinetics of the oxidation of ferrocyanide. Can. J . Chem. 51, 582-587.
Fukunaga, G., G. Katsuragi, T. Izumi and F. Sakiyama (1982) Fluorescence characteristics of kynurenine and N’-formylkynurenine. Their use as reporters of the environment of tryptophan 62 in hen egg-white lyso- zyme. J . Biochem. 22, 129-141.
Nakamura, M., I. Yamazaki, T. Kotani and S. Ohtaki (1985) Thyroid peroxidase selects the mechanism of
either 1- or 2-electron oxidation of phenols, depending on their substituents. 1. Biol. Chem. 260, 13546-13552.
Ohlsson, P.4. and K.-G. Paul (1976) The molar absorp- tivity of horseradish peroxidase. Acta Chem. Scand. 830, 373-375.
Rivas, E. I., A. Paladini, Jr. and G . Cilento (1984) Phot- ochemical-like destruction of tryptophan in serum albu- mins induced by enzyme-generated triplet species. Pho- tochem. Photobiol. 40, 565-568.
Rivas-Subrez, E. and G. Cilento (1981) Quenching of enzyme-generated acetone phosphorescence by indole compounds: Stereospecific effects of D- and L-trypto- phan. Photochemical-like effects. Biochemistry 20, 7329-7333.
Schonbaum, G. R. and S. Lo (1972) Interaction of peroxi- dases with aromatic peracids and alkyl peroxides. J . Biol. Chem. 247, 3353-3360.
Sizer, I . W. (1953) Oxidation of proteins by tyrosinase and peroxidase. In Advances in Enzymology (Edited by F. F. Nord), Vol. 14, pp. 129-161. Interscience Publishers, New York.
Stillman, J. S., M. J. Stillman and H. B. Dunford (1975) Horseradish peroxidase. XIX. A photochemical reac- tion of compound I at 5°K. Biochem. Biophys. Res. Commun. 63, 32-35.


![VIII lezione.ppt [modalità compatibilità] · DNA as a template Nanobiotecnologie The enzymes glucose oxidase (GOx) and horseradish peroxidase (HRP) were modified with nucleic acid](https://img.pdfslide.us/doc/110x75/5f0575097e708231d4130ed4/viii-modalit-compatibilit-dna-as-a-template-nanobiotecnologie-the-enzymes.jpg)


























