Embed Size (px)
Citation preview

Journal of Insect Physiology 47 (2001) 1197–1204www.elsevier.com/locate/jinsphys
Drought acclimation confers cold tolerance in the soil collembolanFolsomia candida
M. Bayley a,*, S.O. Petersenb, T. Knigge c, H.-R. Kohler c, M. Holmstrupd
a Department of Zoology, Institute of Biological Sciences, University of Aarhus, Building 135, Dk-8000 Aarhus, Denmarkb Danish Institute of Agricultural Sciences, Department of Crop Physiology and Soil Science, P.O. Box 50, DK-8830 Tjele, Denmark
c Cell Biology Section, Zoological Institute, University of Tubingen, Auf der Morgenstelle 28, D-72076 Tubingen, Germanyd National Environmental Research Institute, Department of Terrestrial Ecology, Vejlsøvej 25, P.O. Box 314, Dk-8600 Silkeborg, Denmark
Received 6 February 2001; accepted 3 May 2001
Abstract
It has been noted that both summer drought and sub-zero winter temperatures induce the synthesis of sugars and polyols ininvertebrate tissues. This has led several authors to suggest that many of the adaptations, previously viewed as a response to cold,might be part of a more universal desiccation tolerance mechanism. Here we show that acclimation of the soil dwelling collembolanFolsomia candida to a sublethal desiccation stress confers tolerance to cold shock and a significant increase in the molar percentof membrane fatty acids with a mid-chain double bond. These changes in membrane fatty acids are interpreted as conferring asignificant reduction in the transition temperature of cell membranes, as would be expected in acclimation to cold, and these changesare therefore interpreted as contributing to the cross-tolerance. Drought acclimation was also shown to trigger the synthesis of the70 kDa family of heat-shock proteins (Hsp70). This group of heat shock proteins is implicated in the reestablishment of the normalthree-dimensional structure of partially unfolded proteins and therefore are also likely to contribute to the observed cross-tolerance.This study provides evidence that the stresses exerted by desiccation and cold at the cellular level have sufficient similarities toinduce overlapping adaptations. 2001 Elsevier Science Ltd. All rights reserved.
Keywords: Cross-tolerance; Desiccation tolerance; Cold tolerance; Sugars; Phospholipids; Hsp70
1. Introduction
During the winter, many cold hardy organisms syn-thesise high concentrations of low molecular weightorganic molecules such as sugars and polyols(collectively SPs henceforth), often regarded as cryopro-tectants (Sømme and Block, 1982; Zachariassen, 1985;Storey, 1990). The realisation that SPs are also syn-thesised to high concentrations by insects in summerdiapause, where temperatures are well above zero butwhere desiccation is likely to occur because of drought,led Ring and Danks (1994) to suggest that many of theover-wintering adaptations previously explained asresponses to cold, might in fact be part of a more univer-sal desiccation tolerance mechanism. Holmstrup andWesth (1994), showed that cocoons of the earthworm
* Corresponding author. Tel.:+45-89-42-27-20.E-mail address: [email protected] (M. Bayley).
0022-1910/01/$ - see front matter 2001 Elsevier Science Ltd. All rights reserved.PII: S0022-1910 (01)00104-4
Dendobaena octaedra dehydrate at subzero temperaturesto an extent that their melting point equilibrates with theenvironmental temperature. Ice formation is therefore nolonger possible in these cocoons, and this provides aneffective over-wintering strategy in animals with highintegumental permeability, as is the case with many soilorganisms. This mechanism has since been shown tooperate in an arctic collembolanOnychiurus arcticus(Holmstrup and Sømme, 1998; Worland et al., 1998).Its existence in two entirely unrelated soil-animal groupssharing the same high integumental permeability towater loss lends credence to the dehydration mechanismproposed by Holmstrup and Westh (1994). One way oftesting the importance of dehydration tolerance in thesurvival of sub-zero temperatures would be to show anincrease in cold shock tolerance in soil organismsacclimated to summer desiccating conditions (e.g.drought at 20°C).
This approach has not been previously attempted inthese organisms because, until recently, most of the soil-

1198 M. Bayley et al. / Journal of Insect Physiology 47 (2001) 1197–1204
dwelling organisms such as collembolans were con-sidered as having no physiological or metabolic meansof preventing water loss (Noble-Nesbitt, 1963; Harrisonet al., 1991). In other words, it has not been consideredprobable that these organisms are capable of acclimatingto drought, but have rather been expected to rely exclus-ively on a behavioural response by migrating to deepersoil layers. In a recent study we showed, on the contrary,that a widespread soil-living collembolan, Folsomia can-dida (Willem), is well adapted to the desiccating forceslikely to occur in the root zone during drought (Bayleyand Holmstrup, 1999). This animal can actively increasethe osmolality of its body fluids in response to drought,regaining water lost during the initial drought exposureby passive water vapour absorption, and thus remainactive in soil dried to below the permanent wilting per-centage of plants at �15 bar (Gardner, 1960). Thismechanism involves the synthesis of high concentrationsof glucose and myo-inositol (in combination up to 8%of their dry weight), contributing to the increased bodyfluid osmolality which, after seven days of drought stressat 98.2% RH (�25 bars), reached 1250 mOsm/kg. Thislaboratory culture of F. candida has not previously beenconsidered by us as cold tolerant because massive mor-tality is observed at above zero temperatures (2–3°C).However, the high body fluid osmolality attained duringdrought is congruent with a melting point of �2.3°C,and in a pilot study we indeed found that animalsacclimated to 98.2% RH at 20°C for seven days couldsurvive a sudden and prolonged temperature drop to�3°C.
The fact that these animals had survived a sudden fallin temperature of more than 20°C led us to hypothesisethat drought acclimation in these organisms must involveother physiological adaptations than the synthesis of SPs.Two major candidates include alterations in membranephospholipid composition contributing to the mainte-nance of membrane fluidity in desiccating or coldenvironments (Hazel and Williams, 1990; Hazel, 1995),and the expression of chaperone proteins such Hsp70swhich are triggered by a whole variety of stresses (Federand Hofmann, 1999). The aims of the present study weretherefore, firstly, to test the importance of dehydrationtolerance mechanisms in the survival of sub-zero tem-peratures by investigating this cross-tolerance in summerdrought-acclimated F. candida and, secondly, to shedsome light on the underlying mechanisms by screeningfor changes in membrane phospholipid composition andthe expression of Hsp70s.
2. Materials and methods
2.1. Stock animals and drought acclimation
The animals used in all experiments were between oneand two months of age and originated from a laboratory
stock culture maintained in our laboratory in 10 cmdiameter petri dishes containing a 5 mm substrate ofwater-saturated plaster of Paris:charcoal mix (8:1). Theculture was kept at 20±1°C in a 12:12 light:dark regimeand fed a diet of dried baker’s yeast ad libitum.
Animals used in all the experiments were removedfrom this stock culture and placed in water-saturated cul-ture petri dishes without food for two days. The droughtacclimation group was then exposed to air maintained at98.2% RH using 31.6 g NaCl/L H20 salt solution,whereas controls were kept at 100% RH using purewater as described by Holmstrup and Westh (1995). Forthe cold tolerance measurements, groups of ten individ-uals were placed in plastic sample vials (4 cm high, 2 cmdiameter) with a perforated lid, each of which wereplaced in the center of a 200 ml plastic cup containing25 ml of either the salt solution or pure water. This largercontainer was sealed with a tight fitting plastic lid. Theanimals destined for measurement of lipids or Hsp70swere placed in groups of 200 in 10 cm diameter petridishes with lid but without the plaster of Paris substrate.These petri dishes were placed in tightly sealed plasticboxes (25×15×8 cm3) containing either the salt solutionor pure water, to a depth of 4 cm. Thus each of the plas-tic boxes contained three petri dishes with collembolafloating on the surface of the salt solution or water. Allcontainers were placed in a constant temperature roommaintained at 20±0.5°C for seven days.
At this time the animals for measurement of Hsp70swere grouped 100 at a time, and for lipids 200–300 ata time, placed in 0.6 ml Eppendorf tubes and transferredimmediately to a �80°C freezer where they were storeduntil transportation to the appropriate collaborating lab-oratory. Samples were shipped over dry ice (�78.5°C).
2.2. Cold tolerance measurements
After the seven-day drought acclimation period, thesample vials each containing a group of ten F. candidawere removed from the 200 ml plastic constant-humiditycups. The vials’ perforated lids were replaced with ident-ical intact lids and the vials placed in either +5, +1.6, 0,�1, �2, �3, �4 or �5°C surroundings for 48 h. Fourdrought-acclimated and four control samples wereplaced at each temperature. The sub-zero exposure wasperformed in cooling chambers accurate to ±0.1°C; the0°C exposure was performed in a crushed-ice/water bathin a 40×40×40 cm3 polystyrene box; and the positivetemperatures in constant temperature rooms accurate to±0.5°C. After these 48 h each vial was emptied on to acorresponding 5 cm petri dish containing water-saturatedplaster of Paris:charcoal mix (8:1), maintained at 5°Cand kept at this temperature for a further 24 h prior todetermination of mortality.

1199M. Bayley et al. / Journal of Insect Physiology 47 (2001) 1197–1204
2.3. Hsp70 analysis
Pools of F. candida (n=11 for positive controls of aGerman strain which has been selected for heat toleranceat 23°C for seven years and is characterised by a highconstitutive Hsp70 level), n=10 for drought-stressed F.candida from Denmark, (n=9 for non-stressed ones ofthe identical Danish strain) were mechanically homogen-ised in ice-cold extraction buffer composed of 80 mMpotassium acetate, 4 mM magnesium acetate and 20 mMHepes (N-[2-hydroxyethyl] piperazine N-2-ethanesul-fonic acid) at pH 7.5. After homogenisation and centri-fugation of the samples (15 min at 19,100g), total proteinconcentration in the supernatant was determined accord-ing to the method of Bradford (1976). Constant proteinweights (30 �g of total protein/lane) were analysed byminigel SDS-PAGE (12% acrylamide, 0.12% bisacryla-mide (w/v), 15 min at 80 V, 90 min at 120 V). Proteinwas transferred to nitrocellulose by semi-dry blottingand the filter blocked for 2 h in 50% horse serum in TBS(50 mM Tris pH 5.7, 150 mM NaCl). After washing inTBS, monoclonal antibody (mouse anti-human Hsp70,Dianova, Hamburg, Germany, dilution 1:5,000 in 10%horse serum/TBS) was added and the sample was thenincubated at room temperature overnight. After repeatedwashing in TBS for 2 min, the nitrocellulose filter wasincubated in secondary antibody goat anti-mouse IgG(H+L) coupled to peroxidase (Dianova, Hamburg, Ger-many, dilution 1:1000 in 10% horse serum/TBS) at roomtemperature for 2 h. After subsequent TBS washing, theantibody complex was detected by 1 mM 4-chloro(1)naphthol and 0.015% H2O2 in 30 mM Tris pH8.5 containing 6% methanol. Grey value (averageintensity×area) quantification of the Western blot proteinbands took place with the densitometric image analysissystem E.A.S.Y. (Herolab, Wiesloch, Germany) afterbackground subtraction. If Hsp70 fragments weredetected (which was the case for drought-stressedanimals), the bands within one lane were analysed separ-ately and the grey values added. The mean grey valueof the bands obtained for positive control specimens wasset arbitrarily to 1.00 as a standard reference. Based onthis, relative grey values±standard deviations were cal-culated for drought-stressed and non-stressed collem-bola.
2.4. Lipid analysis
Profiles of ester-linked membrane lipid fatty acids(PLFA) derived from F. candida were based on a modi-fied Bligh–Dyer single-phase lipid extraction (White etal., 1979), except that the extraction was carried out intwo steps as described by Kates (1986). Thirty milli-grams portions of material, each consisting of two tothree hundred animals, were extracted; three samplesfrom the control group were included, but only two
samples from the population of drought-acclimated ani-mals due to limitations in the amount of material avail-able. Following extraction in chloroform:methanol:phos-phate buffer (50 mM K2HPO4, pH 7.4), phases were splitby addition of more chloroform and buffer. The chloro-form phase was transferred to a test tube and evaporatedto dryness under N2.
Phospholipids were isolated from the crude lipidextract by solid phase extraction (100 mg silicic acid;Varian, Harbor City, CA). Lipids of low and intermedi-ate polarity were eluted with 1.5 ml chloroform and 6 mlacetone, respectively, and then polar lipids (mainlyphospholipids) were eluted with 1.5 ml methanol (Kates,1986) and collected in a test tube, dried under N2, andtrans-methylated (Dowling et al., 1986). Fatty acidmethyl esters were extracted in hexane, dried and re-dissolved in a small volume of hexane for gas chromato-graphic analysis on a Hewlett–Packard 6890 GC systemwith auto-sampler. The column was an HP-5 (Hewlett–Packard; 50 m fused silica, ID, 0.32 mm) and the ovenwas programmed to increase from 60 to 300°C(Frostegard et al., 1993). Helium was used as the car-rier gas.
Identification of individual fatty acids was based on acombination of retention times and parallel analyses ona similar analytical system interfaced with a mass spec-trometer (Frostegard et al., 1993). Fatty acids are desig-nated as X:YωZ, where X indicates the number of Catoms, Y the number of double bonds, and Z the positionof the first double bond counting from the methyl (ω)end of the molecule. Branching is indicated by prefixesi (iso), a (anteiso) or br (un-specified), and the configur-ation around double bonds by suffixes c (cis) or t (trans)where known. The prefix cy indicates a cyclopropylbonding.
Areas of identified peaks were quantified, correctedfor molecular weight, and the mol% distribution calcu-lated. Pair-wise comparisons between treatments werecarried out for each fatty acid; the sequential Bonferronitest with an initial a level of p�0.05/k (k=number oftests) was used to control the overall error rate (Rice,1989). The data were also used for a principal compo-nent analysis following log(n+1) transformation, andusing the covariance matrix (Bøe and Gjerde, 1980).
3. Results
3.1. Cold tolerance
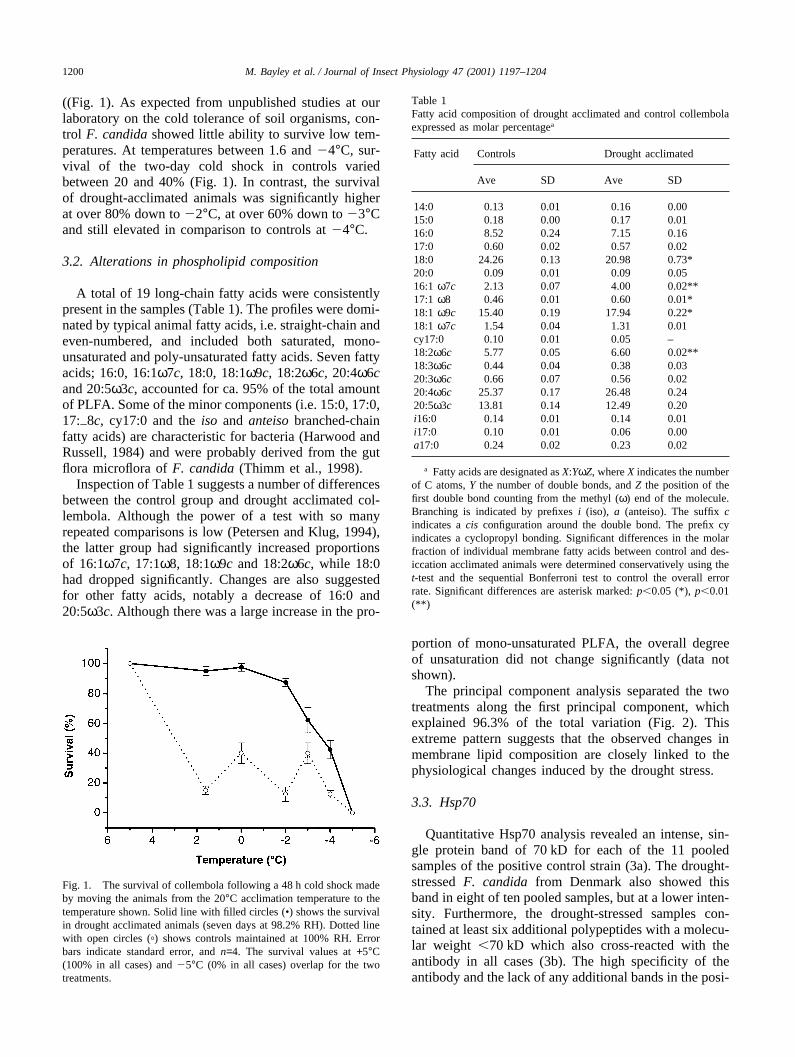
All animals, both the drought acclimated and the con-trols, survived the sudden drop in temperature from 20to 5°C, whereas none survived the shock of moving from20 to �5°C. However, temperature shifts between thesetwo extremes showed that drought acclimation conferredsignificant increases in cold tolerance in F. candida
1200 M. Bayley et al. / Journal of Insect Physiology 47 (2001) 1197–1204
((Fig. 1). As expected from unpublished studies at ourlaboratory on the cold tolerance of soil organisms, con-trol F. candida showed little ability to survive low tem-peratures. At temperatures between 1.6 and �4°C, sur-vival of the two-day cold shock in controls variedbetween 20 and 40% (Fig. 1). In contrast, the survivalof drought-acclimated animals was significantly higherat over 80% down to �2°C, at over 60% down to �3°Cand still elevated in comparison to controls at �4°C.
3.2. Alterations in phospholipid composition
A total of 19 long-chain fatty acids were consistentlypresent in the samples (Table 1). The profiles were domi-nated by typical animal fatty acids, i.e. straight-chain andeven-numbered, and included both saturated, mono-unsaturated and poly-unsaturated fatty acids. Seven fattyacids; 16:0, 16:1ω7c, 18:0, 18:1ω9c, 18:2ω6c, 20:4ω6cand 20:5ω3c, accounted for ca. 95% of the total amountof PLFA. Some of the minor components (i.e. 15:0, 17:0,17:F8c, cy17:0 and the iso and anteiso branched-chainfatty acids) are characteristic for bacteria (Harwood andRussell, 1984) and were probably derived from the gutflora microflora of F. candida (Thimm et al., 1998).
Inspection of Table 1 suggests a number of differencesbetween the control group and drought acclimated col-lembola. Although the power of a test with so manyrepeated comparisons is low (Petersen and Klug, 1994),the latter group had significantly increased proportionsof 16:1ω7c, 17:1ω8, 18:1ω9c and 18:2ω6c, while 18:0had dropped significantly. Changes are also suggestedfor other fatty acids, notably a decrease of 16:0 and20:5ω3c. Although there was a large increase in the pro-
Fig. 1. The survival of collembola following a 48 h cold shock madeby moving the animals from the 20°C acclimation temperature to thetemperature shown. Solid line with filled circles (•) shows the survivalin drought acclimated animals (seven days at 98.2% RH). Dotted linewith open circles (�) shows controls maintained at 100% RH. Errorbars indicate standard error, and n=4. The survival values at +5°C(100% in all cases) and �5°C (0% in all cases) overlap for the twotreatments.
Table 1Fatty acid composition of drought acclimated and control collembolaexpressed as molar percentagea
Fatty acid Controls Drought acclimated
Ave SD Ave SD
14:0 0.13 0.01 0.16 0.0015:0 0.18 0.00 0.17 0.0116:0 8.52 0.24 7.15 0.1617:0 0.60 0.02 0.57 0.0218:0 24.26 0.13 20.98 0.73*20:0 0.09 0.01 0.09 0.0516:1 ω7c 2.13 0.07 4.00 0.02**17:1 ω8 0.46 0.01 0.60 0.01*18:1 ω9c 15.40 0.19 17.94 0.22*18:1 ω7c 1.54 0.04 1.31 0.01cy17:0 0.10 0.01 0.05 –18:2ω6c 5.77 0.05 6.60 0.02**18:3ω6c 0.44 0.04 0.38 0.0320:3ω6c 0.66 0.07 0.56 0.0220:4ω6c 25.37 0.17 26.48 0.2420:5ω3c 13.81 0.14 12.49 0.20i16:0 0.14 0.01 0.14 0.01i17:0 0.10 0.01 0.06 0.00a17:0 0.24 0.02 0.23 0.02
a Fatty acids are designated as X:YωZ, where X indicates the numberof C atoms, Y the number of double bonds, and Z the position of thefirst double bond counting from the methyl (ω) end of the molecule.Branching is indicated by prefixes i (iso), a (anteiso). The suffix cindicates a cis configuration around the double bond. The prefix cyindicates a cyclopropyl bonding. Significant differences in the molarfraction of individual membrane fatty acids between control and des-iccation acclimated animals were determined conservatively using thet-test and the sequential Bonferroni test to control the overall errorrate. Significant differences are asterisk marked: p�0.05 (*), p�0.01(**)
portion of mono-unsaturated PLFA, the overall degreeof unsaturation did not change significantly (data notshown).
The principal component analysis separated the twotreatments along the first principal component, whichexplained 96.3% of the total variation (Fig. 2). Thisextreme pattern suggests that the observed changes inmembrane lipid composition are closely linked to thephysiological changes induced by the drought stress.
3.3. Hsp70
Quantitative Hsp70 analysis revealed an intense, sin-gle protein band of 70 kD for each of the 11 pooledsamples of the positive control strain (3a). The drought-stressed F. candida from Denmark also showed thisband in eight of ten pooled samples, but at a lower inten-sity. Furthermore, the drought-stressed samples con-tained at least six additional polypeptides with a molecu-lar weight �70 kD which also cross-reacted with theantibody in all cases (3b). The high specificity of theantibody and the lack of any additional bands in the posi-

1201M. Bayley et al. / Journal of Insect Physiology 47 (2001) 1197–1204
Fig. 2. The composition of membrane fatty acids (PFLA) in control(�) and drought acclimated (�) collembola were analysed using prin-ciple component analysis which summarises the variability of a multi-variate data (in this case the molar fatty acid concentrations) set into asmaller number of principle components. The plot shows the calculatedscores for the first two principal components explaining 96.3% (PC1)and 2.1% (PC2) of the variation. Component loadings for the individ-ual fatty acids (not shown) confirmed the trends discussed in the text.
tive controls justified the interpretation of these smallerpolypeptides as fragments of Hsp70 and hence the sum-mation of all detectable bands per sample into one ‘valueof intensity’ for the Hsp70 level of each sample. Theaverage Hsp70 level of the drought-stressed collembol-ans reached 42.3 (SD: 25.1) % of the Hsp70 level of thepositive controls. For the non-drought stressed collem-bola, the analysis of 30 �g total protein failed to detectany Hsp70 band in all nine samples (3c) and, thus, theHsp70 levels in these samples were at most equivalentto the background level of 2.3% of the Hsp70 level ofthe positive controls (5.4% of that of the drought-stressed collembola of the identical F. candida strain).
4. Discussion
The fact that drought acclimation of F. candida at20°C conferred cross-tolerance and the ability to survive
Fig. 3. Standardised Western blots of three representative pools of animals showing Hsp70 levels in: (a) positive controls; (b) drought-stressedanimals; (c) non-stressed animals. Intact Hsp70 is marked with the filled triangle (�), numbers adjacent to open triangles (�) refer to molecularsize of marker proteins, slashes (-) indicate the presence of Hsp70 fragments of various molecular sizes occurring in drought stressed collembola.
a sudden drop in temperature down to �1.6°C providescompelling evidence of the importance of desiccationtolerance mechanisms to the winter survival of terrestrialarthropods. Despite the connection between cold anddesiccation tolerance mechanisms found in many invert-ebrate species including nematodes (Forge and MacGu-idwin, 1992), earthworms (Holmstrup and Zachariassen,1996), tardigrades (Sømme, 1996), the arctic collem-bolan O. arcticus (Holmstrup and Sømme, 1998) andseveral insects (Ring and Danks 1994, 1998), few studieshave addressed the existence of cross-tolerance betweenthese two stresses in invertebrates.
Tammariello et al. (1999) showed that a 24 h acclim-ation of the flesh fly Sarcophaga crassipalpis in 50 or0% RH at 25°C reduced, rather than increased, theability of these animals to survive a cold shock of�10°C lasting more than 60 min. One possible expla-nation of the difference in conclusions between thisstudy and our results may be found in the time allowedfor acclimation; the flesh flies were acclimated for 24 h,whereas our animals were acclimated for seven days.Our reason for choosing this relatively long acclimationperiod was that the SPs, synthesised during droughtstress in F. candida (Bayley and Holmstrup, 1999), con-tinued to increase and only reached a stable concen-tration after five to seven days. At least some aspects ofdrought acclimation seem, therefore, to require morethan 24 h to reach full effect. Indeed in the botanicalliterature, which contains numerous reports of frost toler-ance induced by drought acclimation going back as earlyas 1872 (Chen et al., 1975), there is a tendency toacclimate plants to drought for weeks to months beforetesting for cross-tolerance (Chen et al. 1975, 1977; Bur-rows et al., 1989; Thomas and James, 1993; Anisko andLindstrom, 1995).
Membrane integrity and function is obviously anessential prerequisite for all living organisms, and alter-ations in the phase behaviour of membrane phospholipidfatty acids (PLFA) may pose a serious challenge to

1202 M. Bayley et al. / Journal of Insect Physiology 47 (2001) 1197–1204
physiological functions in poikilotherms. There is con-siderable evidence that tailoring of PLFA composition,by regulating fatty acid length and degree of unsatu-ration, is important in minimising the risk of phase tran-sition occurring at ambient temperature (Hazel, 1995).These adaptations make an important contribution indefining the thermal limits for the growth and functionof living organisms (Hazel, 1995).
Only one reference to lipid composition of a collem-bolan species was found in the literature, namely Stran-sky et al. (1986), who isolated various fractions of sur-face and internal lipids of Tetrodontophora bielanensis(Waga). The phospholipid fraction was not analysed intheir study, but all major (�1 mol%) fatty acids ident-ified in the present study, except for 20:5ω3c, were alsoidentified in the free fatty acid fraction by Stransky etal. (1986). In this study, drought acclimation caused asignificant increase in the mol% of membrane lipidmono-unsaturated fatty acids and 18:2ω6c, while theother polyunsaturated fatty acids showed only minorincreases or even a decrease resulting in a net status quoin the overall degree of unsaturation. Regulation ofmembrane phase transition temperature via changes inlipid fatty acid composition is not simply a matter ofincreasing the overall degree of unsaturation; the numberand position of cis–cis double bonds in long-chain fattyacids has a pronounced influence on the properties ofmembrane lipid bilayers (Hazel, 1995; Wang et al.,1999). In general, the gel to liquid phase transition tem-perature (Tm), decreases with increasing number of dou-ble bonds, and with increasing distance from a doublebond to the ends of the molecule. Also, the effect of thefirst double bond introduced is stronger than the effectof the second, and so on. Hence, the observed shifttowards a higher proportion of mono-unsaturated fattyacids with a mid-chain double bond in this study can beexpected to correspond to a relatively strong decrease ofTm in the drought acclimated animals, and to make animportant contribution to the subsequent cross-toleranceto cold shock.
However, these adaptations occurred as a result ofdrought stress and not low temperature stress. Surpris-ingly, there is very little literature concerning the effectsof desiccation on membrane integrity despite droughtand water availability being one of the major factorsshaping terrestrial ecosystems. In plant and bacterialstudies, desiccation stress alone appears to induce areduction in the level of membrane PFLA unsaturationcorresponding to an increase in Tm (Liljenberg andKates, 1985; Kieft et al., 1994). Higher values of Tm
correspond to a higher level of order in the packing ofmembrane lipids (Separovic and Gawrisch, 1996). Con-versely, decreasing Tm values will reflect increasing dis-order, and this has been shown to increase the waterpermeability of artificial polyunsaturated membranes(Huster et al., 1997). The reduced level of PLFA unsatu-
ration, or increasing Tm, observed in plant and bacterialmembranes during drought (Liljenberg and Kates, 1985;Kieft et al., 1994) has thus been interpreted as an adapt-ive response to water stress.
In F. candida, drought results in an initial water-lossfollowed by an active increase in body fluid osmolalitythrough the synthesis of high concentrations of glucoseand myo-inositol, resulting in passive water vapourabsorption from the soil pore air. In studies on the phasebehaviour of lipids in water, it has been shown that notonly decreasing temperatures, but also drying of themedium, can induce membrane phase transition from thenormal liquid phase to the more ordered gel phase asso-ciated with membrane dysfunction (Small, 1986; New-man et al., 1993). In this light it would appear that theincrease in mono-unsaturated PLFAs and 18:2ω6c meas-ured in this study can be considered adaptive both to thedesiccating regime during acclimation and to the sub-sequent cold shock.
Drought acclimation of F. candida using an identicalprocedure to this study resulted in an accumulation ofhigh concentrations of glucose and myo-inositol (Bayleyand Holmstrup, 1999). Several studies have shown thatat sufficient concentrations, a variety of carbohydratescan stabilise membrane structure and lower Tm (Newmanet al., 1993). Sugars and polyols, at molarities similar tothose expected in the animals in this study, have alsobeen shown to exert a direct influence on membrane flu-idity. Thus, Furtula et al. (1990) found that a sterol probediffused four times faster within protoplast membranesin a solution of 0.45 M mannitol or trehalose comparedto the diffusion rate in 0.9 M solutions, and the obser-vation was intepreted as a result of decreased membranefluidity at the higher osmotic potential. A similar effectwas not observed for a phospholipid probe, however, afact that the authors could not explain. It would thereforeappear, that both the alterations in PLFAs measured inthis study and the high concentrations of SPs reportedpreviously (Bayley and Holmstrup, 1999), might con-tribute to the animals’ tolerance to drought and to theircross-tolerance to cold shock.
Drought acclimation in this study also induced theexpression of Hsp70 in F. candida. Proteins in the Hsp70family are induced by a wide variety of stressors includ-ing, in addition to the obvious heat shock, cold shock(Joplin et al., 1990; Goto et al., 1998), return fromanoxia (Ananthan et al., 1986), exposure to desiccationstress (Tammariello et al., 1999) and a wide variety ofchemical contaminants (Ananthan et al., 1986; Kam-menga et al., 2000). Whilst there is clear evidence onthe importance of Hsp70 induction to a variety of stresstolerances in cell lines, their effect on acquired tolerancein whole organisms is less clear. Dahlgaard et al. (1998)showed that the expression of Hsp70 in adult fruit fliesDrosophila melanogaster reached a peak after 2 h butsubsequently fell and approached background levels 32 h

1203M. Bayley et al. / Journal of Insect Physiology 47 (2001) 1197–1204
after the heat stress. In contrast, the acquired toleranceto a subsequent heat treatment reached a maximumbetween 8 and 32 h after the first heat shock. The authorsconcluded that whilst whole body measurements werepositively associated with survival at all times they wereinadequate as a full explanation of acquired tolerance.Similarly, Goto et al. (1998) showed that diapausingindividuals of D. triauraria had consistently higher toler-ance to heat, cold and desiccation than non-diapausingindividuals. However, diapausing individuals consist-ently accumulated lower levels of Hsp70 mRNA thanthose not in diapause. Goto et al. (1998) concluded, thatthe extra tolerance in diapausing animals was inde-pendent of the transcriptional regulation of Hsp70.Reported functions of Hsp70 proteins are numerous andapparently diverse but all include the basic process ofbinding and subsequent release of partially unfolded pro-teins in an ATP-dependant cycle whose end result iseither the reestablishment of their normal 3D structureor their degradation (Forreiter and Nover, 1998). Thisfunction fits well with the model for their induction pro-posed by Ananthan et al. (1986) who showed that thecommon factor in the diversity of stresses causing tran-scription of the Hsp70 gene, was the presence of par-tially denatured proteins within the cell. There are there-fore strong mechanistic arguments that Hsp70 inductionmakes an important contribution to the overall reactionof an organism to stress and that its expression in thepresent study contributed both to this animal’s toleranceto the stress encountered during the drought acclimationperiod, and to its survival of the subsequent cold shock.
The direct adaptive value of this cross-tolerance wheredrought adaptation confers subsequent frost tolerance ishard to imagine for this temperate species. There areprobably few climatic regions where winter frosts aredirectly preceded by periods of severe drought, althoughthis type of climatic event does frequently occur in higharctic tundra regions. Despite this, cross-tolerance of thistype has been noted on numerous occasions in the plantliterature (Chen et al. 1975, 1977; Burrows et al., 1989;Thomas and James, 1993; Anisko and Lindstrom, 1995).The existence of this kind of cross-tolerance sharingsimilar adaptations is probably indicative of similaritiesin the nature of these two, superficially different, stresstypes. Both sub-zero temperatures and summer droughtare environmental stresses that cause dehydration andhence an increase in cellular solute concentrations. Theseelevations in solute concentration present challenges toboth protein and membrane structural integrity requiringinduction of a variety of adaptations within the organismsuch as those found in this study. The effect of des-iccation on membrane integrity and the molecularenvironment within the cell appears to have been spar-sely investigated in terrestrial arthropods and is an areaworthy of future study.
Acknowledgements
This study received funding from the Danish NaturalScience Research Council (Grant No. 9700177).
References
Ananthan, J., Goldberg, A.L., Voellmy, R., 1986. Abnormal proteinsserve as eukaryotic stress signals and trigger the activation of heatshock genes. Science 232, 522–524.
Anisko, T., Lindstrom, O.M., 1995. Reduced water supply induces fallacclimation of evergreen azaleas. Journal of the American Societyfor Horticultural Science 120, 429–434.
Bayley, M., Holmstrup, M., 1999. Water vapor absorption in arthro-pods by accumulation of myoinositol and glucose. Science 285,1909–1911.
Bradford, M.M., 1976. A rapid and sensitive method for the quantifi-cation of microgram quantities of protein utilizing the principle ofprotein-dye binding. Analytical Biochemistry 72, 248–254.
Burrows, R.L., Waters, L. Jr., Markhart, A.H. III, 1989. Cold acclim-ation of asparagus seedlings subjected to low temperatures or waterstress. Hortscience 24, 812–814.
Bøe, B., Gjerde, J., 1980. Fatty acid patterns in the classification ofsome representatives of the families Enterobacteriaceae and Vib-rionaceae. Journal of General Microbiology 116, 41–49.
Chen, P.M., Li, P.H., Weiser, C.J., 1975. Induction of frost hardiness inred-osier dogwood stems by water stress. Hortscience 10, 372–374.
Chen, P.M., Li, P.H., Burke, M.J., 1977. Induction of frost hardinessin stem cortical tissues of Cornus stolonifera Michx. by waterstress. Plant Physiology 59, 236–239.
Dahlgaard, J., Loeschcke, V., Michalak, P., Justesen, J., 1998. Inducedthermotolerance and associated expression of the heat-shock pro-tien Hsp70 in adult Drosophila melanogaster. Functional Ecology12, 786–793.
Dowling, N.J.E., Widdel, F., White, D.C., 1986. Phospholipid ester-linked fatty acid biomarkers of acetate-oxidizing sulphate-reducersand other sulphide-forming bacteria. Journal of General Micro-biology 132, 1815–1825.
Feder, M.E., Hofmann, G.E., 1999. Heat-shock proteins, molecularchaperones, and the stress response: evolutionary and ecologicalphysiology. Annual Review of Physiology 61, 243–282.
Forge, T.A., MacGuidwin, A.E., 1992. Effects of water potential andtemperature on survival of the nematode Meloidogyne hapla infrozen soil. Canadian Journal of Zoology 70, 1553–1560.
Forreiter, C., Nover, L., 1998. Heat induced stress proteins and theconcept of molecular chaperones. Journal of Biosciences 23,287–302.
Frostegard, A., Tunlid, A., Bath, E., 1993. Phospholipid fatty acidcomposition, biomass, and activity of microbial communities fromtwo soil types experimentally exposed to different heavy metals.Applied Environmental Microbiology 59, 3605–3617.
Furtula, V., Khan, I.A., Nothnagel, E.A., 1990. Selective osmoticeffect on diffusion of plasma membrane lipids in maize protoplasts.Proceedings of the National Academy of Sciences of the USA 87,6532–6536.
Gardner, W., 1960. Dynamic aspects of water availability to plants.Soil Science 89, 63–73.
Goto, S.G., Yoshida, K.M., Kimura, M.T., 1998. Accumulation ofHsp70 mRNA under environmental stresses in diapausing and non-diapausing adults of Drosophila triauraria. Journal of Insect Physi-ology 44, 1009–1015.
Harrison, P.M., Rothery, P., Block, W., 1991. Drying processes in theantarctic Collembolan Cryptopygus antarcticus (Willem). Journalof Insect Physiology 37, 883–890.

1204 M. Bayley et al. / Journal of Insect Physiology 47 (2001) 1197–1204
Harwood, J.L., Russell, N.J. 1984. Lipids in Plants and Microbes.George Allen and Unwin, London.
Hazel, J.R., 1995. Thermal adaptation in biological membranes: ishomeoviscous adaptation the explanation? Annual Review ofPhysiology 57, 19–42.
Hazel, J.R., Williams, E.E., 1990. The role of alterations in membranelipid composition in enabling physiological adaptation of organ-isms to their physical environment. Progress in Lipid Research 29,167–227.
Holmstrup, M., Westh, P., 1994. Dehydration of earthworm cocoonsexposed to cold: a novel cold hardiness mechanism. Journal ofComparative Physiology Series B 164, 312–315.
Holmstrup, M., Westh, P., 1995. Effects of dehydration on waterrelations and survival of lumbricid earthworm egg capsules. Journalof Comparative Physiology Series B 165, 377–383.
Holmstrup, M., Zachariassen, K., 1996. Physiology of cold hardinessin earthworms. Comparative Biochemistry and Physiology SeriesA 115, 91–101.
Holmstrup, M., Sømme, L., 1998. Dehydration and cold hardiness inthe arctic collembolan Onychiurus arcticus Tullberg 1876. Journalof Comparative Physiology Series B 168, 197–203.
Huster, D., Jin, A.J., Arnold, K., Gawrisch, K., 1997. Water per-meability of polyunsaturated lipid membranes measured by 17ONMR. Biophysical Journal 73, 855–864.
Joplin, K.H., Yocum, G.D., Denlinger, D.L., 1990. Cold shock elicitsexpression of heat shock proteins in the flesh fly, Sacophaga crassi-palpis. Journal of Insect Physiology 36, 825–834.
Kammenga, J.E., Dallinger, R., Donker, M.H., Kohler, H.-R.,Simonsen, V., Triebskorn, R., Weeks, J.M., 2000. Biomarkers interrestrial invertebrates for ecotoxicological soil risk assessment.Reviews of Environmental Contamination and Toxicology 164,93–147.
Kates, M., 1986. Techniques of Lipidology, 2nd ed. Elsevier, Amsterd-am.
Kieft, T.L., Ringelberg, D.B., White, D.C., 1994. Changes in ester-linked phospholipid fatty acid profiles of subsurface bacteria duringstarvation and desiccation in a porous medium. Applied Environ-mental Microbiology 60, 3292–3299.
Liljenberg, C., Kates, M., 1985. Changes in lipid composition of oatroot membranes as a function of water deficit stress. Canadian Jour-nal of Biochemistry and Cell Biology 63, 77–84.
Newman, Y., Ring, S., Colaco, C., 1993. The role of trehalose andother carbohydrates in biopreservation. In: Tombs, M. (Ed.),Biotechnology and Genetic Engineering Reviews, vol. 11. Inter-cept, Andover, pp. 263–294.
Noble-Nesbitt, J., 1963. Transpiration in Podura aquatica L.(Collembola, Isotomidae) and the wetting properties of its cuticle.Journal of Experimental Biology 40, 681–700.
Petersen, S.O., Klug, M.J., 1994. Effects of sieving, storage, and incu-bation temperature on the phospholipid fatty acid profile of a soil
microbial community. Applied Environmental Microbiology 60,2421–2430.
Rice, W.R., 1989. Analyzing tables of statistical tests. Evolution 43,223–225.
Ring, R., Danks, H., 1994. Desiccaion and cryprotection: overlappingadaptations. Cryo-Letters 15, 181–190.
Ring, R., Danks, H., 1998. The role of trehalose in cold-hardiness anddesiccation. Cryo-Letters 19, 275–282.
Separovic, F., Gawrisch, K., 1996. Effect of unsaturation on the chainorder of phosphatidylcholines in a dioleoylphosphatidylaminematrix. Biophysical Journal 71, 274–282.
Small, D.M., 1986. Handbook of Lipid Research 4. The PhysicalChemistry of Lipids: from Alkenes to Phospholipids. Plenum Press,New York.
Storey, K.B., 1990. Biochemical adaptation for cold hardiness ininsects. Philosophical Transactions of the Royal Society of LondonSeries B Biological Sciences 326, 635–654.
Stransky, K., Trka, A., Budesınsky, M., Streibl, M., 1986. Lipidic com-pounds from the extract of the springtail Tetrodontophora biel-anensis (Waga). Collection of Czechoslovak Chemical Communi-cations 51, 948–955.
Sømme, L., 1996. Anhydrobiosis and cold tolerance in tardigrades.European Journal of Entomology 93, 349–357.
Sømme, L., Block, W., 1982. Cold hardiness of collembola at SignyIsland, maritime Antarctic. Oikos 38, 168–176.
Tammariello, S.P., Rinehart, J.P., Denlinger, D.L., 1999. Dessicationelicits heat shock protein transcription in the flesh fly, Sarcophagacrassipalpis, but does not enhance tolerance to high or low tem-peratures. Journal of Insect Physiology 45, 933–938.
Thimm, T., Hoffmann, A., Borkott, H., Munch, J.C., Tebbe, C.C.,1998. The gut of the soil microarthropod Folsomia candida(Collembola) is a frequently changeable but selective habitat anda vector for soil microorganisms. Applied and EnvironmentalMicrobiology 64, 2660–2669.
Thomas, H., James, A.R., 1993. Freezing tolerance and solute changesin contrasting genotypes of Lolium oerenne L. acclimated to coldand drought. Annals of Botany 72, 249–254.
Wang, G., Li, S., Lin, H., Brumbaugh, E.E., Huang, C., 1999. Effectsof various numbers and positions of cis double bonds in the sn-2acyl chain of phosphatidylethanolamine on the chain-melting tem-perature. Journal of Biological Chemistry 274, 12289–12299.
White, D.C., Davis, W.M., Nickels, J.S., King, J.D., Bobbie, R.J.,1979. Determination of the sedimentary microbial biomass byextractable lipid phosphate. Oecologia 40, 51–62.
Worland, M.R., Gruborlajsic, G., Montiel, P.O., 1998. Partial des-iccation induced by sub-zero temperatures as a component of thesurvival strategy of the arctic collembolan Onychiurus Arcticus(Tullberg). Journal of Insect Physiology 44, 211–219.
Zachariassen, K.E., 1985. Physiology of cold tolerance in insects.Physiological Review 65, 799–831.





























