Embed Size (px)
Citation preview

HEATTOLERANCEANDACCLIMATIONINFEMALEATHLETES
JessicaAnneMee
ADoctoralthesissubmittedinfulfilmentoftherequirementoftheUniversityofBrighton
fortheawardofDoctorofPhilosophy

I
Abstract
Thisthesisaimedtodeveloparunningheattolerancetest(RHTT)toassesschangesinheattolerance
andtoinvestigatethemechanismandoptimisationofheatacclimation(HA)forfemaleathletes.
ThefirststudyintroducedaRHTTandassesseditsrepeatability.Resultsdemonstrategoodagreement,
strongcorrelationsandsmalldifferencesbetweenrepeatedtrials.Thetypicalerrorofmeasurevalues
suggested low within-participant variability. Furthermore, the RHTT was effective in differentiating
betweenindividuals’physiologicalresponses,demonstratingthatheattoleranceliesalongacontinuum.
ThesecondstudyexaminedthesensitivityoftheRHTTtochanges inheattoleranceandtoevaluate
individualresponsestoHA.ResultsdemonstratethattheRHTTissensitivetochangesinheattolerance
and that themagnitudeof adaptation is highly individual; supporting theuseof theRHTT in future
investigations.
ReducingthermalstrainthroughHAinnotfullyunderstoodforafemalepopulation.Thethirdstudy
comparedmales’ and females’ temporal patterning to short-termHA (STHA; 5-d) and long-termHA
(LTHA;10-d).TheRHTTwasusedtoquantifychangesinheattolerance.Theresultsconfirmthatwhilst
STHAmaybeeffectiveinachievingpartialadaptationinmalesandfemales,femalesrequireLTHAto
establishreductionsinthermoregulatoryandcardiovascularstrain.
ImprovedthermotolerancefollowingHA,reducesdisruptionstocellularhomeostasisprincipally,butnot
exclusively,byincreasingbasalheatshockprotein72followingtranscriptionofitsgene(Hsp72mRNA)
aspartoftheheatshockresponse(HSR).Thefourthstudycomparedmales’andfemales’Hsp72mRNA
responseduringSTHAandLTHA.ThesimilartranscriptionofHsp72mRNAobservedinallparticipants
suggeststhattherearenodifferencesintheendogenouscriteriatoelicittheHSRbetweensexes.
ThefifthstudyassessedtheeffectivenessofprecedingSTHAwithapassiveheatexposure(HAsauna)in
females.HAsaunaresultedinreductionsinthermoregulatory,cardiovascularandperceptualstrain.The
adaptation pathway was likely mediated in part, by plasma volume expansion and an improved
thermoeffectorandthermosensitivityresponseofthesudomotorfunction.
Together,evidenceinthisthesissupportsthenotionthatspecialconsiderationsneedtobetakenwhen
usingHAtoattenuatethermoregulatorystraininfemaleathletespriortotrainingandcompetinginthe
heat.

II
TableofContents
Abstract......................................................................................................................................I
Listofabbreviations................................................................................................................VIII
Listoftables...............................................................................................................................X
Listoffigures............................................................................................................................XII
Listofequations.......................................................................................................................XV
Acknowledgments...................................................................................................................XVI
Declaration.............................................................................................................................XVII
1. Introduction........................................................................................................................1
2. Literaturereview................................................................................................................6
2.1. Environmentalheatstressandexchange...........................................................................6
2.1.1. Dryheattransferpathways........................................................................................7
2.1.1. Wetheattransferpathways.......................................................................................8
2.1.2. Heatbalance...............................................................................................................8
2.2. Heatstrain........................................................................................................................10
2.2.1. Measurementofcoretemperature.........................................................................11
2.2.2. Measurementofskintemperature..........................................................................13
2.2.3. Bodyheatcontent....................................................................................................14
2.3. Behaviouraltemperatureregulation................................................................................17
2.4. Physiologicaltemperatureregulation..............................................................................17
2.4.1. Adjustablesetpoint.................................................................................................18
2.4.2. Reciprocalinhibition.................................................................................................20
2.4.3. Heatregulation.........................................................................................................21
2.5. Physiologicalresponsestoheatstress.............................................................................22
2.5.1. Sweatingandevaporativeheatloss.........................................................................23

III
2.5.2. Skinbloodflowanddryheatloss.............................................................................24
2.5.3. Cardiovascularsupportofthermoregulation...........................................................26
2.6. Sexdifferencesinphysiologicalresponsestoheatstress................................................27
2.6.1. Sweatrateandevaporativeheatloss......................................................................28
2.6.2. Skinbloodflowanddryheatloss.............................................................................28
2.7. Menstrualcycle................................................................................................................29
2.8. Verificationofthemenstrualcyclephase........................................................................32
2.8.1. Countingdaysfromtheonsetofmenses.................................................................32
2.8.2. Basalbodytemperature...........................................................................................33
2.8.3. Urinaryluteinisinghormoneconcentration.............................................................33
2.8.4. Measurementofoestrogenandprogesterone........................................................33
2.9. Thermoregulatorydifferencesoverthemenstrualcycle.................................................34
2.9.1. Sweatingandskinbloodflow...................................................................................35
2.9.2. Cardiovascular..........................................................................................................36
2.9.3. Exerciseperformanceintheheat............................................................................37
2.10. Individualdifferenceinheattolerance........................................................................38
2.10.1. Aerobicfitness..........................................................................................................38
2.10.2. Anthropometry.........................................................................................................40
2.11. Methodstoassessheattolerance................................................................................41
2.12. Exertionalheatillnessesinrunning..............................................................................42
2.13. Heatacclimation...........................................................................................................44
2.14. Heatacclimationprotocols...........................................................................................45
2.14.1. Fixedintensityheatacclimation...............................................................................45
2.14.2. Self-regulatedheatacclimation................................................................................46
2.14.3. Passiveheatacclimation..........................................................................................46
2.14.4. Controlledhyperthermiaheatacclimation..............................................................46

IV
2.15. Heatacclimationinduction..........................................................................................47
2.15.1. Sweatingandskinbloodflow...................................................................................47
2.15.2. Bloodvolumeandfluidbalance...............................................................................51
2.15.3. Cardiovascularstability............................................................................................54
2.15.4. Thermotolerance......................................................................................................58
2.15.5. Exerciseperformance...............................................................................................60
2.15.6. Perceptualresponses...............................................................................................62
2.16. Timecourseofheatacclimation..................................................................................62
2.17. Heatacclimationdecay................................................................................................63
2.18. Females’responsestoheatacclimation......................................................................64
2.19. Aimofthesis.................................................................................................................65
2.19.1. Researchquestionsarisingfromtheliteraturereview............................................65
2.19.2. Proposedresearchstudiesandhypotheses.............................................................66
3. Generalmethods..............................................................................................................68
3.1. Ethics,healthandsafety..................................................................................................68
3.2. Participants.......................................................................................................................69
3.3. Facilities............................................................................................................................70
3.4. Pre-trialdietandexercisestandardisation......................................................................70
3.5. Anthropometricassessment............................................................................................72
3.6. Menstrualcycle................................................................................................................74
3.7. Environmentalconditions................................................................................................75
3.8. Thermoregulatorymeasures............................................................................................76
3.9. Cardiopulmonarymeasures.............................................................................................78
3.10. Physiologicalstrainindex.............................................................................................79
3.11. Haematologicalmeasures............................................................................................79
3.12. Sweatrate....................................................................................................................80

V
3.13. Perceptualscales..........................................................................................................81
3.14. Cyclingexercisetrials...................................................................................................82
3.15. Runningexercisetrials..................................................................................................85
3.16. Heattolerancetests.....................................................................................................86
3.17. Statisticalanalysis.........................................................................................................91
3.17.1. Poweranalysis..........................................................................................................91
3.17.2. Normalityandsphericityofdata..............................................................................91
3.17.3. Repeatabilitystatistics..............................................................................................91
3.17.4. Statisticaltestsofdifference....................................................................................92
3.17.5. Effectsizes................................................................................................................93
3.17.6. Posthocanalysis......................................................................................................94
4. Repeatabilityofarunningheattolerancetest..................................................................95
4.1. Abstract............................................................................................................................95
4.2. Introduction......................................................................................................................95
4.3. Materialsandmethods....................................................................................................98
4.4. Results..............................................................................................................................99
4.5. Discussion.......................................................................................................................103
4.6. Limitations......................................................................................................................106
4.7. Conclusion......................................................................................................................106
5. Sensitivityofarunningheattolerancetest.....................................................................107
5.1. Abstract..........................................................................................................................107
5.2. Introduction....................................................................................................................107
5.3. Materialsandmethods..................................................................................................109
5.4. Results............................................................................................................................111
5.5. Discussion.......................................................................................................................114
5.6. Limitations......................................................................................................................116

VI
5.7. Conclusion......................................................................................................................117
6. Acomparisonofmaleandfemaletemporalpatterningtoshort-andlong-termheatacclimation.............................................................................................................................118
6.1. Abstract..........................................................................................................................118
6.2. Introduction....................................................................................................................118
6.3. Materialsandmethods..................................................................................................121
6.4. Results............................................................................................................................123
6.5. Discussion.......................................................................................................................131
6.6. Limitations......................................................................................................................134
6.7. Conclusion......................................................................................................................135
7. SexcomparisonofleukocyteHsp72mRNAtranscriptionduringheatacclimation...........136
7.1. Abstract..........................................................................................................................136
7.2. Introduction....................................................................................................................136
7.3. Materialsandmethods..................................................................................................138
7.4. Results............................................................................................................................141
7.5. Discussion.......................................................................................................................146
7.6. Limitations......................................................................................................................147
7.7. Conclusion......................................................................................................................148
8. Passiveheatexposureprecedingshort-termheatacclimationacceleratesadaptationinfemales..................................................................................................................................149
8.1. Abstract..........................................................................................................................149
8.2. Introduction....................................................................................................................149
8.3. Materialsandmethods..................................................................................................151
8.4. Results............................................................................................................................155
8.5. Discussion.......................................................................................................................163
8.6. Limitations......................................................................................................................165
8.7. Conclusion......................................................................................................................165

VII
9. Generaldiscussion..........................................................................................................167
9.1. PrincipleFindings............................................................................................................167
9.1.1. Heattolerancetest.................................................................................................167
9.1.2. Sexcomparisonstoheatacclimation.....................................................................167
9.1.3. Acceleratedheatacclimationresponsesinfemales..............................................168
9.2. MechanisticOverview....................................................................................................171
9.2.1. HeatTolerance.......................................................................................................171
9.2.2. HeatAcclimation....................................................................................................176
9.2.3. Acceleratedheatacclimationinfemales................................................................181
9.3. Directionsforfutureresearch........................................................................................182
9.4. Practicalapplicationoffindings.....................................................................................185
9.5. Conclusion......................................................................................................................186
References.............................................................................................................................188
Appendices............................................................................................................................201
Informedconsent.......................................................................................................................201
Medicalquestionnaire...............................................................................................................202
Menstrualcyclequestionnaire...................................................................................................204
Calculationof65%VbO2peak......................................................................................................205

VIII
Listofabbreviations
BM Bodymass
BSA Bodysurfacearea
CI Confidenceinterval
CO2 Carbondioxide
FSH Folliclestimulatinghormone
HA Heatacclimation
HAsauna HAcombinedwithapassiveheatexposure
HR Heartrate
HspmRNA Heatshockproteinmessengerribonucleicacid
HSP Heatshockprotein
HSR Heatshockresponse
HTT Heattolerancetest
ICC Intraclasscorrelationcoefficient
IDF IsraeliDefenceForce
LH Luteinisinghormone
LTHA Long-termheatacclimation
NaCl SodiumChloride
O2 Oxygen
PV Plasmavolume
RH Relativehumidity

IX
RHTT Runningheattolerancetest
RPE Ratingofperceivedexertion
rpm revolutionspermin
RT-qPCR Reversetranscriptionpolymerasechainreaction
SD Standarddeviation
SWR Sweatrate
SWRBSA Sweatraterelativetobodysurfacearea
STHA Short-termheatacclimation
TEM Technicalerrorofmeasurement
TE(CV%) Technicalerrorasacoefficientofvariation
Tc Coretemperature
Tre Rectaltemperature
TS Thermalsensation
Tsk Meanskintemperature
USG Urinespecificgravity
VbO2max Maximaloxygenuptake
VbO2peak Peakoxygenuptake
WBGT Wetbulbglobetemperature

X
Listoftables
Table2.1Estimatedwholebodyskinbloodflowrequirementsduringprolonged,severerunning
exerciseatdifferentbodycoreandskintemperatures.
Table2.2Commonterminologyandhormoneconcentrationsassociatedwithatypical28-dcycle
withovulationoccurringonD14.
Table2.3Heatillnessesandtheirassociatedsymptoms.
Table2.4Summaryofadaptationstoheatacclimation.
Table2.5Change inwholebodysweat rateobserved following short-and long-termcontrolled
hyperthermiaheatacclimation.
Table 2.6 Change in plasma volume observed following short- and long-term controlled
hyperthermiaheatacclimation.
Table 2.7 Change in resting heart rate observed following short- and long-term controlled
hyperthermiaheatacclimation.
Table2.8Changeinrestingcoretemperatureobservedfollowingshort-andlong-termcontrolled
hyperthermiaheatacclimation.
Table2.9Plateaudaysofphysiologicaladaptations(thepointatwhichapproximately85%ofthe
adaptationoccurs)duringheatacclimatisation.
Table3.1Repeatabilityofurineosmolalityandurinespecificgravitymeasurement.Mean±SD.
Table3.2Repeatabilityofskinfoldmeasurements.Mean±SD.
Table3.3Plasmaconcentrationof17β-estradiolandprogesterone.Mean±SD(range).
Table3.4Repeatabilityofrectalthermistors.Mean±SD.
Table3.5Repeatabilityofskinthermistors.Mean±SD.
Table3.6Repeatabilityofcapillarybloodlactateanalysis.Mean±SD.
Table3.7PhysiologicalandsubjectiveresponsetocyclingatsetpercentagesofVbO2peak.

XI
Table3.8PhysiologyvariablesmeasuredduringtheIDFHTTandtheRHTT.Mean±SD.
Table 4.1Repeatability statistics for physiological variables during repeated trials of the RHTT.
Mean±SD.
Table5.1Participantcharacteristics.Mean±SD(ranges).
Table5.2PeakphysiologicalvariablesduringtheRHTT.Mean±SD(ranges).
Table6.1Participantcharacteristics.Mean±SD.
Table6.2Performanceandphysiologicalresponsesduringshort-termheatacclimationandlong-
termheatacclimation.Mean±SD.
Table 6.3 Physiological variables during baseline testing (RHTT1), following STHA (RHTT2) and
followingLTHA(RHTT3).Mean±SD.
Table7.1Participantcharacteristics.Mean±SD.
Table7.2Hsp72mRNAprimersequences.
Table7.3PhysiologicalandperformancemeasuresonD1,D5andD10ofcontrolledhyperthermia
heatacclimation.Mean±SD.
Table8.1Heat acclimationprotocol andphysiological responsesduringD1andD5ofHACH and
HAsauna.Mean±SD.
Table8.2Physiologicalvariablesduringbaselinetesting(RHTT1),andfollowing5-dHACHorHAsauna
(RHTT2).Mean±SD.
Table9.1Hypothesesforeachstudychapterpresentedinthisthesis.
Table9.2Participantcharacteristics.Mean±SD(Range).
Table9.3Plateaudaysofphysiologicaladaptations(thepointatwhichapproximately85%ofthe
adaptationoccurs)duringheatacclimatisation.
Table9.4Daysofphysiologicaladaptation inmalesandfemalesduringcontrolledhyperthermia
heatacclimation.

XII
Listoffigures
Figure 2.1. Avenues for heat exchange when performing exercise. Adapted from Gisolfi and
Wenger,(1984)
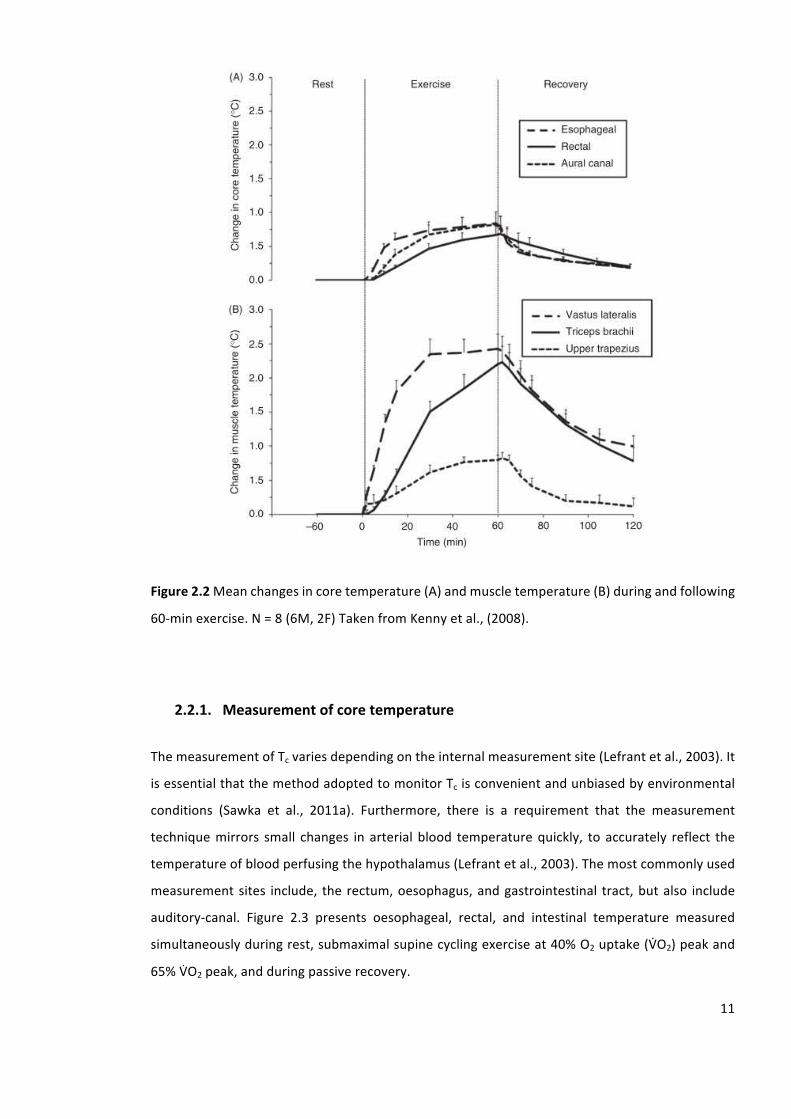
Figure2.2Meanchangesincoretemperature(A)andmuscletemperature(B)duringandfollowing
60-minexercise.N=8(6M,2F)TakenfromKennyetal.,(2008).
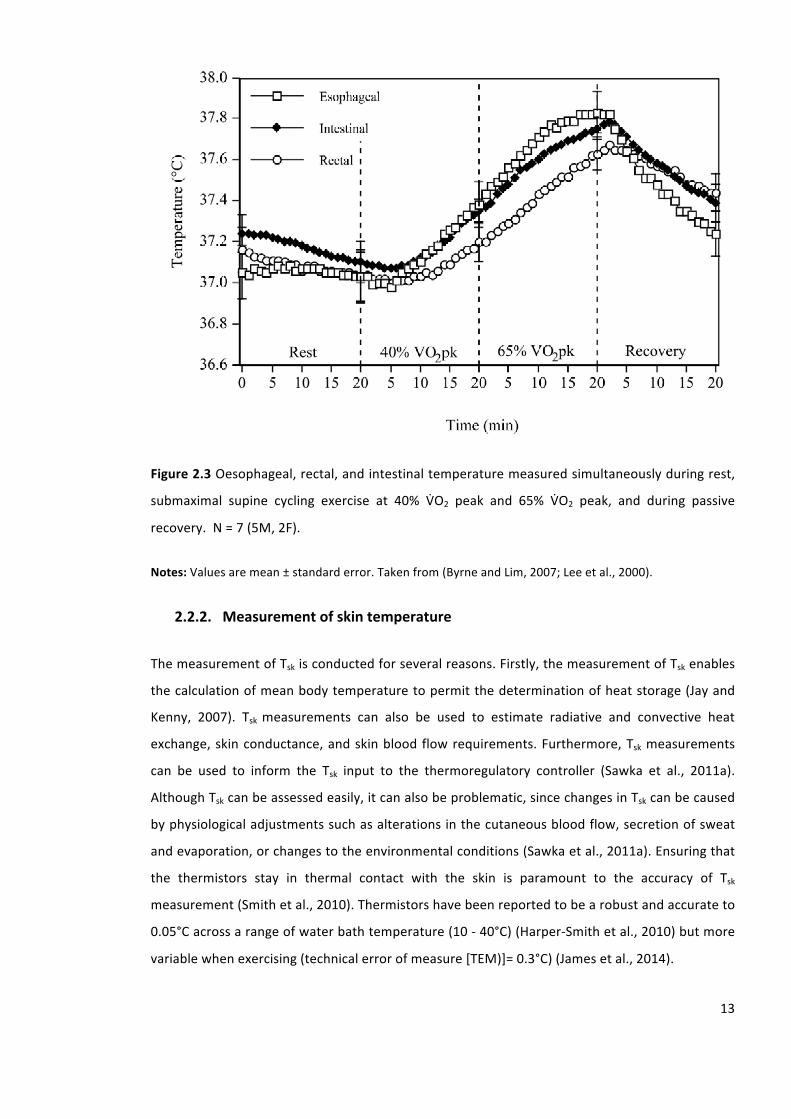
Figure2.3Oesophageal,rectal,andintestinaltemperaturemeasuredsimultaneouslyduringrest,
submaximal supine cycling exercise at 40% VbO2 peak and 65% VbO2 peak, and during passive
recovery.N=7(5M,2F).
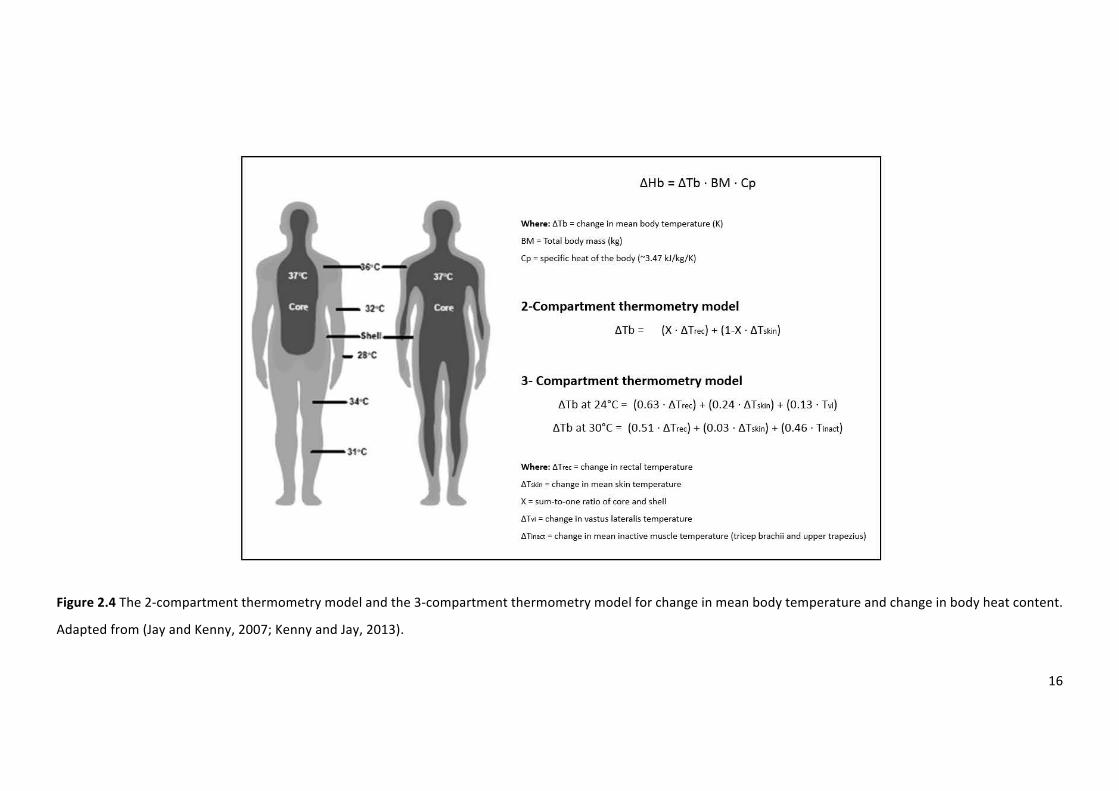
Figure2.4The2-compartmentthermometrymodelandthe3-compartmentthermometrymodel
forchangeinmeanbodytemperatureandchangeinbodyheatcontent.Adaptedfrom(Jayand
Kenny,2007;KennyandJay,2013).
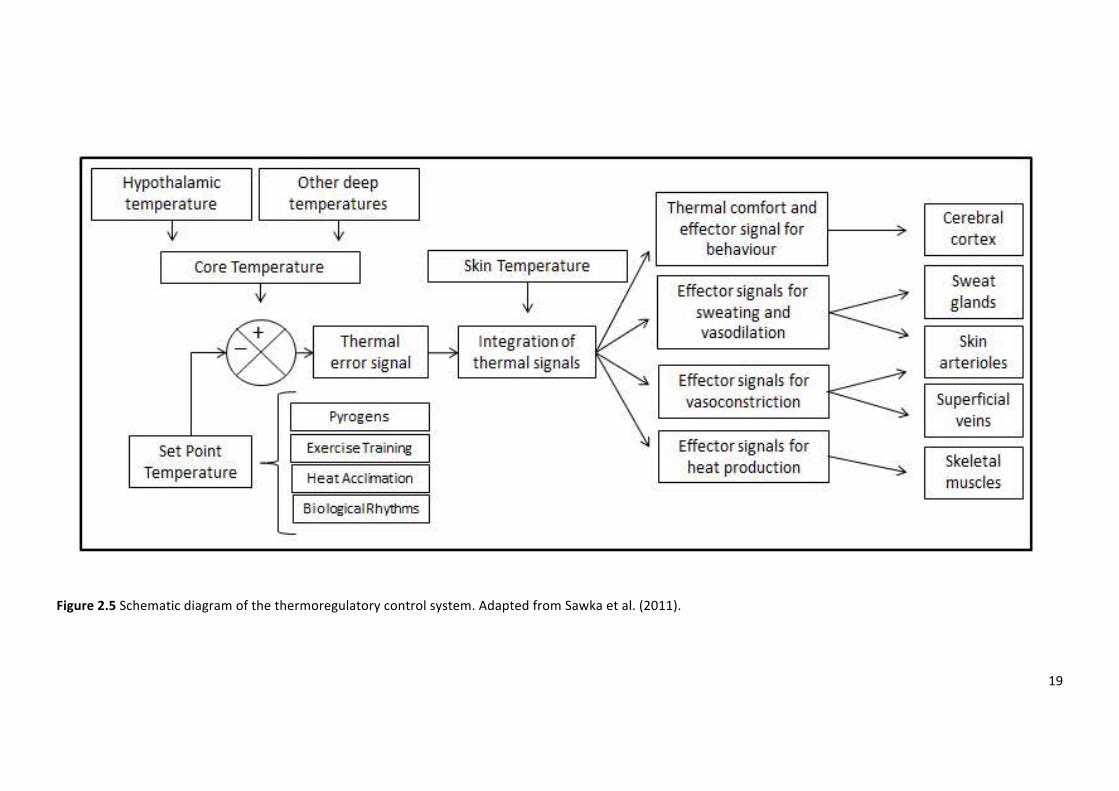
Figure2.5Schematicdiagramofthethermoregulatorycontrolsystem.AdaptedfromSawkaetal.
(2011).
Figure2.6Thereciprocalinhibitionmodelofthermoregulation.Adaptedfrom(Bligh,2006).
Figure2.7Meanwholebodycalorimetrydataforrateoftotalheatproductionandrateofnetheat
loss after60-min rest, 60-min cyclingat70Wand60-minof inactive recovery.N=8 (6M,2F).
AdaptedfromKennyetal.(2008).
Figure2.8Typicalcoretemperature(Tc)andhormonechangesacrossthemenstrualcycle.Noteall
valuesareapproximatevaluessincetheyvarygreatlyacrossindividuals.LH=luteinizinghormone
FSH=folliclestimulatinghormone.AdaptedfromStachenfeld&Taylor,(2014).
Figure2.9Hsp72mRNApre-andpost-sessionsonD1,D5andD10ofcontrolledhyperthermiaHA
inmaleparticipants.Adaptedfrom(Gibsonetal.,2015a).
Figure2.10EffectofheatacclimationonVbO2maxinacool(13°C)andhot(38°C)environmentin
maleparticipants.
Figure3.1Ratingofperceivedexertion(RPE)andthermalsensation(TS)scales.

XIII
Figure3.2.Exampleofrelativeexerciseintensityelicitedduringa90-mincontrolledhyperthermia
heatacclimationsession.
Figure3.3Restingrectaltemperature(Tre)andrestingheartrate(HR)duringtheIsraeliDefence
Forceheattolerancetest(IDFHTT)(x-axis)andrunningheattolerancetest(RHTT)(y-axis)formales
(closedmarkers)andfemales(openmarkers).Dottedlinerepresentslineofequality.N=16(8M,
8F).
Figure3.4Peakrectaltemperature(Tre)andpeakheartrate(HR)duringtheIsraeliDefenceForce
heat tolerance test (IDF HTT) (x-axis) and running heat tolerance test (RHTT) (y-axis) formales
(closedcircles)andfemales(opencircles).Dottedlinerepresentslineofequality.N=16(8M,8F).
Figure4.1 Peak rectal temperature (Tre),peakheart rate (HR), sweat rate (SWR),andpeak skin
temperature(Tsk)duringRHTT1(x-axis)andRHTT2(y-axis),formales(closedmarkers)andfemales
(openmarkers).Dottedlinerepresentslineofequality.N=16(8M,8F).
Figure4.2Bland-Altmanplotswithmeanbias(solidline)and95%limitsofagreement(dottedline)
for peak rectal temperature (Tre), peak heart rate (HR), sweat rate (SWR), and peak skin
temperature(Tsk)formales(closedmarkers)andfemales(openmarkers).N=16(8M,8F).
Figure5.1Individualdataforvaluesofpeakrectaltemperature(Tre),peakskintemperature(Tsk),
peakheatrate(HR)andsweatrateSWRduringRHTT1(xaxis)andRHTT2(yaxis)formales(closed
markers)andfemales(openmarkers).Dottedlinerepresentslineofequality.N=25(12M,13F).
Figure6.1Rectal temperature (Tre)at5-min intervalsduring therunningheat tolerancetest for
males(left)andfemales(right).GreymarkersrepresenttheRHTT1,blackmarkersRHTT2andwhite
markerRHTT3.Mean±SD.N=16(8M,8F).
Figure6.2Heartrate(HR)at5-minintervalsduringtherunningheattolerancetestformales(left)
andfemales (right).Greymarkersrepresent theRHTT1,blackmarkersRHTT2andwhitemarker
RHTT3.Mean±SD.N=16(8M,8F).
Figure7.1Restingrectaltemperature(Tre)(left)andrestingheartrate(HR)(right)onD1,D5and
D10ofheatacclimation.Dottedlinerepresentslineofequality.N=10(5M,5F).
Figure7.2Hsp72mRNApreandpostsessiononD1,D5andD10ofheatacclimationinmalesand
females.Mean±SD.N=10(5M,5F).

XIV
Figure7.3Hsp72mRNApretopostsessiononD1,D5andD10relativetoD1preofheatacclimation
inmales(opencircles)andfemales(closedcircles).N=10(5M,5F).
Figure8.1Schematicoftheexperimentaldesign.
Figure8.2Plasmaconcentrationsof17ß-estradiolandprogesteronepriortoRHTT1andRHTT2in
theHACH intervention(whitebars)andRHTT1andRHTT2intheHAsauna intervention(blackbars)
andD22ofthemenstrualcycle(greybar).Mean±SD.N=9.
Figure8.3Rectal temperature (Tre)at5-min intervalsduringtheRHTTfor theHACH intervention
(left)andtheHAsaunaintervention(right).OpenmarkersrepresentRHTT1andtheclosedmarkers
representRHTT2.Mean±SD.N=9.
Figure8.4Heartrate(HR)at5-minintervalsduringtheRHTTfortheHACHintervention(left)and
theHAsaunaintervention(right).OpenmarkersrepresentRHTT1andtheclosedmarkersrepresent
RHTT2.Mean±SD.N=9.
Figure8.5Sweatrate(SWR)(left),sodiumchloride(NaCl)sweatconcentration(middle),andTreat
onsetofsweating(right)duringRHTT1(xaxis)andRHTT2(yaxis)intheHACHintervention(closed
markers)andHAsaunaintervention(openmarkers).Dottedlinerepresentslineofequality.N=9.
Figure9.1Correlationcoefficientsforassociationbetweenchangesinrectaltemperatureandbody
surfacearea(BSA),sumofskinfolds,relativeVbO2peak,relativeexerciseintensity,absoluteheat
production(Hprod,W),heatproductionrelativetoBSA(Hprod,W.m2),andheatproductionrelative
toBM(Hprod,W.kg-1).
Figure9.2Scatterplotillustratingtherelationshipbetweenthechangeinrectaltemperature(∆Tre)
andabsoluteheatproduction(Hprod,W)duringtheRHTT.N=30.

XV
Listofequations
Equation2.1Calculationforwetbulbglobetemperature(WBGT)(YaglouandMinard,1957)
Equation2.2Heatbalanceequation
Equation2.3Calculationformetabolicenergyexpenditure
Equation2.4Calculationformeanskintemperature(Tsk)
Equation3.1Calculationofbodysurfacearea(BSA)
Equation3.2Calculationofphysiologicalstrainindex(PSI)
Equation3.3Calculationofwholebodysweatrate(SWR)
Equation3.4Calculationofwholebodysweatraterelativetobodysurfacearea(SWRBSA)
Equation3.5Calculationofworkduringcycleergometry
Equation3.6Calculationofpowerfromworkduringcycleergometry
Equation3.7CalculationofrateofTreincrease
Equation3.8Calculationoftypicalerrorofthemeasure(TEM)
Equation3.9Calculationoftechnicalerrorasacoefficientofvariation(TE(CV%))
Equation3.10Calculationofintraclasscorrelation(ICC)
Equation3.11CalculationofCohen’sd(d)
Equation8.1CalculationforchangeinPlasmaVolume(∆PV)

XVI
Acknowledgments
Iwouldliketotakethisopportunitytothankthosewhohavemadethisthesispossibleandtothe
followingindividualsfortheirunconditionalsupportandguidancethroughthevariousstagesofmy
thesis.
Firstly,IwouldliketothankDrNeilMaxwellandProfJoDoustfortheirsupervisionandguidance
inundertakingmyDoctorateofPhilosophyinatopicthatIampassionateabout.Youhaveboth
actedasmentorsformeandsupportedmygrowthasanEnvironmentalPhysiologist.Youradvice
onbothresearchaswellasonmycareerhavebeeninvaluable.
ThankyoufellowTechnicalInstructor’sBen,James,Oli,andRosie,andPhDstudentsAsh,Carl,and
Cat for providing stimulating scientific discussions over extended coffee and lunch breaks.My
gratitudeisalsoextendedtothetechnicalteamattheUniversityofBrightonandLeeandJames
fromBedfordshireUniversityfortheirsupportwiththeHsp72mRNAanalysis.Iwouldalsoliketo
giveaspecialthankstoalloftheparticipantswhovolunteeredtheirtime,energyandcommitment
totheresearchprojects.
Aspecialthankstomyfamily.BothmymumHeatheranddadDavidhavealwayssupportedme
withmydecisiontoundertakeaPhD.Theyprovidedmewithagreatfoundationtolife,teaching
metobecaring,hardworking,dedicatedandindependent.MysisterLizzyforalwaysbeingthere
andofcourseDillanwhoseloveiscompletelyunconditional.
Lastbutnotleast,ImustacknowledgeBenedictforalwayskeepingmegroundedandhelpingme
torealisetheimportantthingsinlife.Benedicthasbeencentraltothecompletionofthisthesisas
hehasgivenmeconfidenceandmotivationwhentheywerelost.

XVII
Declaration
Ideclarethattheresearchcontainedinthisthesis,unlessotherwiseindicatedwithinthetext, is
originalwork of the author. The thesis has not been previously submitted to this or any other
universityforadegree,anddoesnotincorporateanymaterialalreadysubmittedforadegree.
Signed:
JessicaAnneMee
Date:18thMarch2016

XVIII
The following academic publications and conference proceedings have been due to the work
containedwithinthisthesis:
Academicpublications
Mee,J.A.Doust,J.andMaxwell,N.S.(2015).Repeatabilityofarunningheattolerancetest.Journal
ofThermalBiology49-50,91-97.
Mee,J.A.Gibson,O.R.Doust,J.D.Maxwell,N.S.(2015).Sexdifferencesinadaptationtoshort-and
long-termheatacclimation.ScandinavianJournalofMedicineandScienceinSports,25(Suppl.1),
250-258.
Mee,J.A.Peter,S.Doust,J.Maxwell,N.S.(2015).Restrictedsweatevaporationprecedingshort-
termheatacclimationacceleratesadaptationinfemales.ExtremePhysiologyandMedicine4(1),
A112.[Abstractonly].
Conferenceproceedings
Mee,J.A.Peter,S.Doust,J.Maxwell,N.S.(2015).Restrictedsweatevaporationprecedingshort-
term heat acclimation accelerates adaptation in females. International Conference of
EnvironmentalErgonomics(ICEE),Portsmouth,UK,28thJune-3rdJuly2015.
Mee,J.A.Gibson,O.R.Taylor,L.Tuttle,J.A.Watt,P.W.Doust,J.Maxwell,N.S.(2015)Sexcomparison
ofleukocyteHsp72mRNAupregulationduringheatacclimation.EuropeanCollegeofSportScience
(ECSS)Malmo,Sweden,24th–27thJune2015.
Mee,J.A.Gibson,O.R.Doust,J.Maxwell,N.S.(2014)Sexdifferencesinadaptationtoshort-and
long-term isothermicheatacclimation.Trainingandcompeting in theheatconference,Aspetar,
Doha,Qatar,23rd-24thMarch2014.
Mee,J.A.Doust,J.Maxwell,N.S.(2013).Reliabilityandvalidityofashortdurationrunningheat
tolerancetest.BritishAssociationofSportandExerciseScience(BASES)studentconference,Cardiff,
UK.

1
1. Introduction
_______________________________________________________________________________
Moderate ambient temperatures are known to present sufficient heat stress that result in an
elevated core temperature (Tc) and reduce endurance capacity (Galloway andMaughan 1997).
Duringexercise inahotenvironment,activemusclesperformworkcausingan increase inbody
heat content. These changes represent the rate of change in body heat storage,which in turn
reflectsthebalancebetweenmetabolicheatproduction,heatabsorbedfromtheenvironmentand
totalbodyheat loss(JayandKenny,2007).Exertionalheatstrokeischaracterisedbyarise inTc
beyond40°Ccoupledwithcentralnervoussystemdysfunctionandmultipleorgansystemfailure
(Casaetal.,2012).
TheworkbyNielsen(1996)providesdatatosuggestanenduranceathletemayexperienceuptoa
1°CriseinTcevery9-minwhenracinginhighambientconditions.ThisrateofriseinTcwouldresult
inarunnerreachingaTcof40°Cwithin25-30min,withtheimmediatedangersofheatexhaustion.
Dataarelimitedontheincidenceofexertionalheatillnessinlongdistancerunning,particularlythe
reportingofnon-fatalcases.The11.5kmFalmouthRoadRaceintheUSA,typicallyruninwarm
humidconditions(23.3°C,70%RH),reportsonaverage10–20casesper10,000entrantsperyr
(DeMartinietal.,2014).Incontrast,theTwinCitiesMarathoninUSA,runincoolerconditions(16°C,
60%RH)reportsanaverageof1caseofexertionalheatillnessper10,000finishers(Roberts,2000).
WhilehighambienttemperatureandRHareimportantfactorscontributingtowardsexertionalheat
illness, there are also incidences in cooler environments (Hawes et al., 2010; Roberts, 2006;
RobertsonandWalter,2010).TheGreatNorthRunintheUKin2009(18°C)reported55casesof
exertionalheatillnessamongthe55,000participants(Hawesetal.,2010),anincidenceratesimilar
tothatobservedattheFalmouthRoadRacewhereconditionsaremuchwarmer.
Martin (1997) reportedahigher incidencerate in females (36 incidence;3.63%)comparedwith
males(26incidence;2.18%);withfemalesaccountingfor58%ofthetotalnumberofcases(Martin,
1997).Questionably,oneofthemostmemorablemarathonfinisheswasofa39-yrs.old,female
Swissathlete,GabrielaAnderson-Schiess.ItwasapparentGabrielawassufferingwithaseriouscase
ofheatexhaustionduringthelatterstagesofthe1984Olympicinauguralfemales’marathon.She
limpedaroundthefinal400min5-minand44-s,refusingmedicalsupport,eventuallycrossingthe
finish line in2-hr48-minand45-s. The severityof this case is still apparent today,with similar
incidencessuchasthatof34-yr.old,female,Namibianathlete,BeataNaigambo.Duringthe2014
CommonwealthGamesmarathon,Beata staggereddown thehomestraightand faintedmeters

2
fromthefinishline,eventuallycollapsingoverthefinishlineunassistedin2-hr39-min23-s.This
body of evidence emphasises the severity of heat related illnesses in endurance athletes,
specificallyfemales.Further,ithighlightstheneedtodevelopeffectivescreeningproceduresand
heat-alleviating strategies, to support endurance athletes when training and competing in
thermally-challengingenvironments.
Heatintolerancemaybepermanentoracquiredandreferstoasyndromedescribingindividuals
whoareunabletoadaptproperlytoexercisinginahotenvironment.Heattolerancetests(HTT)are
currentlyusedtoevaluatemilitarypersonalthathavehadapreviousexperienceofexertionalheat
illness, to determine whether they are able to return to duty. The Israeli Defence Force (IDF)
developedaHTTwhichinvolves120-minwalkingonatreadmillatapaceof5km.hr-1anda2%
gradientinambientconditionsof40°Cand40%relativehumidity(RH).Inspiteofthesignificant
rolefortheIDFHTTtoscreenmilitarypersonnelforheatintolerance,theapplicationoftheIDFHTT
hasbeenquestionedinreferencetoenduranceathletesduetodifferencesinexerciseintensityand
duration,theclothing,andthecarryingload(Johnsonetal.,2013).Enduranceathletestypically
race forashorterdurationandatahigherexercise intensitycomparedwithmilitarypersonnel,
whomostcommonlyperformlowintensitymarchingforprolongedperiodsoftime.Therefore,the
rateofheatproduction,requirementforevaporativecooling,andchangesinTcdiffersubstantially
betweenthesepopulations.Theincidenceofexertionalheatillnessinenduranceathletes(Martin,
1997) raises the importance of developing a heat tolerance screening procedure that informs
medical personal about an endurance athlete’s current heat tolerance state, and is capable of
monitoringchangesinheattolerance.
Repeated heat exposure to a stressful thermal environment initiates a phenotypic adaptation
known as heat acclimation (HA), an element of which has been identified as thermotolerance
(Moseley,1997).HAimprovesthermalcomfort,submaximalexerciseperformance,andincreases
maximal aerobic capacity in the heat (Lorenzo et al., 2010). The benefits arise from enhanced
sudomotorandskinbloodflowresponses,plasmavolume(PV)expansion,cardiovascularstability
and an improved fluid balance (Poirier et al., 2015). Thermotolerance or acquired cellular
thermotolerance describes the cellular adaptation accompanying systemic changes induced by
successful HA (Horowitz, 2014; Magalhães et al., 2010). As a result, HA is the consensus
recommendation strategy to attenuate the physiological strain associated with training and
competingintheheat(Racinaisetal.,2015).However,HAitisnotroutinelyadoptedbyendurance
athletes; UK athletics currently only offer altitude training camps as part of their world class
performanceprogramme.

3
Controlled hyperthermiaHA ensures equal thermal strain is placedon participants during each
session,asitinvolveselevatingandmaintainingasteadystateTcabovethesweatingthreshold(Fox
et al., 1963a). This method has become increasingly popular since it offers more complete
adaptationdue to theprogressiveoverloadapproach comparedwith traditional, fixed-intensity
protocolswhichuseaconstantoverloadapproach(Gibsonetal.,2015b).However,thereremains
apaucityofdataconcerningbestpractice forHA in females; consequently, femaleathletesare
attendingwarmweather trainingcampsor implementingHAstrategiesbasedondatacollected
frommale participants. The implications of this are that femalesmay be adopting sub-optimal
strategiesthatprovidelittlebenefittoalleviatingphysiologicalstrain.
Sex has traditionally been considered an independent modulator of temperature regulation
(Kaciuba-UscilkoandGrucza,2001;Kenney,1985;Nunneley,1978),howevertheseobservations
may be confounded by inherent physical differences between the sexes. More recently, sex
differences in temperature regulation during exercise have been reported, irrespective of
confounding differences inmetabolic heat production andphysical characteristics (Gagnon and
Kenny, 2012, 2011). Interestingly, these differences only become evident above a certain
requirement for heat loss and are solely attributed to peripherally mediated control of the
sudomotor function. Furthermore, fluctuations in hormonal concentrations associatedwith the
menstrualcyclehaveaknowneffectonthermoregulationinfemales.Duringthelutealphaseof
themenstrual cycle there is anelevation inprogesterone concentrationwhichmodifies central
regulatorymechanisms resulting in alterations in basal Tc, the body temperature threshold for
sweating,andthebodytemperaturethresholdforcutaneousvasodilationontheforearm,chest,
back,forearm,andthigh(Inoueetal.,2005).Thesesexdifferencesinthermoregulatoryresponses
toacuteexerciseheatstress,raisesthequestionofwhetherfemalesadapttochronicheatexposure
differently tomales. Thus, research investigating females’ phenotypic and cellular responses to
controlledhyperthermiaHAovershort-andlong-termtimescalesisrequired.
Recently,novelHAstrategieshavebeeninvestigatedinmaleparticipants.Thesemethodsinvolve
permissivedehydration(Garrettetal.,2012;Nealetal.,2015),saunabathing(Scoonetal.,2007;
Stanley et al., 2015), and hotwater immersion (Zurawlew et al., 2015). Thesemethods aim to
establish an accelerated adaptation via targeting adaptation pathways including, PV expansion,
increased sudomotor function, and altered cutaneous vascular function. Once the temporal
patterningtocontrolledhyperthermiainfemaleshasbeendetermined,furtherresearchisrequired
toestablishnovelHAstrategiesthataccelerateadaptionresponsesinfemales.

4
Thisintroductionhasoutlinedtheincidenceofheatrelatedillnessinrunners,particularlyfemales,
the requirement to develop a running HTT (RHTT), the clear need for research into females
phenotypic and cellular adaptations to controlledhyperthermiaHA, and introduced the ideaof
investigating novel HA strategies to accelerate adaptation in females. This thesis is therefore
presentedinthefollowingchapters.
Chapter2reviewstheliterature,discussingacuteandchronicresponsestoheatstress.Thefirst
partoftheliteraturereviewconsidersthephysiologicalandthermoregulatoryresponsetoanacute
heat exposure, discussing individual differences, with a particular focus on any potential sex
differences.Thesecondpartoftheliteraturereviewfocusesonheatillnessesandtheincidence
rateinrunning.ThethirdpartoftheliteraturereviewdiscussesHAprotocolsandthetimecourse
ofadaption.ThefourthsectionfocusesonthepotentialsexdifferencesinresponsestoHA.The
final sectionof this chapteroutlines the thesis aims, researchquestions andhypothesesof the
experimentalstudies.ThesearchwasrestrictedtofulltextarticlespublishedinEnglishlanguage
peer reviewed journals. Where possible information was limited to articles using human
participants,howeverdue to thenoveltyof someof researchpresented, animaldatahasbeen
used.
Chapter3describesthecommonmethodsusedacrossmultipleexperimentalchapters.
Chapter4presentsthefirstexperimentalchapter(Study1)wherebytherepeatabilityofaRHTT
wasassessed.Thefindingsfromthisstudyinformsfutureexperimentalchaptersbyquantifyingthe
measurement error associated with the RHTT to support accurate conclusions regarding the
magnitudeofadaptationobserved.
Chapter5presentsthesecondexperimentalchapter(Study2)wherebythesensitivityoftheRHTT
toquantifychangesinheattolerancefollowingshort-termHA(STHA)wasassessed.
Chapter6usestheRHTTtoquantifythechangesinphysiologicalandthermoregulatoryresponses
followingcontrolledhyperthermiaHAovershort-andlong-termtimescalesinmalesandfemales
(Study3). This chapterprovidesnovelHAdataon females anddescribes thedifferences in the
temporalpatterningbetweenmalesandfemales.
Chapter7presentsthefourthexperimentalchapter (Study4)describingtheheatshockprotein
generesponses(Hsp72mRNA)tocontrolledhyperthermiaHAovershort-andlong-termtimescales
inmalesandfemales.

5
Chapter 8 describes the findings from the final experimental chapter (Study 5). Following the
identificationthatfemalesrequiredlong-termHA(LTHA)toestablishareductionincardiovascular
andthermoregulatoryresponsesinchapter6(Study3),anovelmethodofcombiningasauna-like
exposure with controlled hyperthermia HA was investigated to establish its effectiveness in
acceleratingadaptationresponsesinfemalesoverashort-termtimescale.
Chapter9discussestheindividualandcumulativefindingsofalltheexperimentalchapterstoform
thegeneraldiscussion.Thefirstpartofthegeneraldiscussionwillreviewtheprinciplefindingsfrom
the experimental chapters presented. The second part of the general discussionwill provide a
mechanisticoverview,focusingspecificallyonheattolerance,thetemporalpatterningofHA,and
acceleratingtheadaptioninfemales;involvingretrospectiveanalysesofpooleddatafromwithin
thethesis.Thischaptercloseswithdiscussingpotentialfuturedirectionsforresearchinthefield,
inadditiontoreviewingthepracticalapplicationofthepresenteddata.

6
2. Literaturereview
________________________________________________________________________________
2.1. Environmentalheatstressandexchange
Heatstress refers toboththeenvironmentalandmetabolicconditions that typically increaseTc
(Sawkaetal.,2011a).Heat stress is categorisedascompensableheat stress,oruncompensable
heatstress.CompensableheatstressreferstoasteadystateTc,wherebytheheatlossmatchesthe
heatproduction(Sawkaetal.,2011a).Uncompensableheatstressreferstoanimbalancebetween
theevaporativerequirementandtheevaporativecapacityofanindividual(Cheungetal.,2000).
Duringuncompensableheatstress,anindividualisunabletodissipatethemetabolicheat,causing
acontinualriseinTc,untileitherexhaustionoccurs,ortheseverityoftheenvironmentalconditions
decreases(Cheungetal.,2000).
To quantify environmental heat stress in occupational, military and sports applications a user-
friendlymeasureknownaswetbulbglobetemperature(WBGT)iscommonlyused(Budd,2008).
WBGTisanempiricalindexofclimaticheatstresswhichcanbeusedtoquantifytheheatstress
(Sawkaetal.,2011a).WBGTcanbecalculatedusingEquation2.1.
Equation2.1Calculationforwetbulbglobetemperature(WBGT)(YaglouandMinard,1957)
WBGT=0.7Tw+0.2Tg+0.1Td
Where:Tw=wetbulbtemperature,Tg=blackglobetemperature,Td=drybulbtemperature
TheWBGT has a number of limitations which questions its validity, accuracy and applicability.
Specifically,theWBGTdoesnotmakeanyconsiderationsforclothingormetabolicrate,asaresult,
it cannot predict heat exchange between a person and the climate (Budd, 2008). Despite its
limitations,theWBGTisthemostwidelyusedindex,sinceitiseasytouseandisareliablemeasure
ofheatstress(Budd,2008).ManygeographicalregionsexperienceWBGTabove29°Cduringthe
summermonths.Thisiscausedbyahighhumidityand/orahighairtemperatureandsolarload,
asreflectedbyahighwetbulbtemperatureandblackglobetemperature,respectively(Sawkaet
al.,2011a).Theuniversal thermalclimate index(Fialaetal.,2012)providesanalternativetothe
WBGT,byintegratingpredictiveformulasofheatstrainbasedonactivitiesconductedindifferent

7
temperaturesatvaryingconstantmetabolicheatproductions.Whiletheuniversalthermalclimate
indexmayultimatelyofferamorecomprehensiveandrobustevaluationoftheriskofphysiological
heatstrain,itismorecomplexandexpensivetoadopt.Toensurethatanyheatstressmonitoring
system is widely adopted across all settings, solutions must be relatively simple to use and
implement, while being affordable (Périard et al., 2016). Additional research is warranted to
developamoreappropriatetooltosupportgoverningbodiesinensuringthehealthandsafetyof
athletesinhotenvironmentalconditions.
Physicalexerciseincreasesmetabolicheatproductionabovetherestingrate(<100W).Depending
uponthetypeofexercise,approximately80to90%ofenergyproduced isreleasedasheatand
thendispersedaroundthebody,mostlythebloodstream(JayandKenny,2007).Itisnecessarythat
the heat is dissipated from the body to avoid heat storage, with a resultant increase in Tc.
Cardiovascularadjustmentsoccurinordertoredirectthebloodflowfromthebody’scoretothe
peripheryforsubsequentheatdissipation(Nyboetal.,2014).Theheatdeliveredtotheperiphery
is transferred to the environment by dry (radiative, conductive and convective) and wet
(evaporative)heattransferpathways.Theeffectivenessoftheheattransferpathwaysisdependent
upon the biophysical properties of the environment. This includes the temperature differences
betweentheskinsurfaceandtheenvironment,theevaporativepotentialandthesolarconditions
(Sawkaetal.,2011a).
2.1.1. Dryheattransferpathways
Thenon-evaporativeavenuesofheatexchangesarecollectivelycalleddryheatexchange;these
includeconduction,convectionandradiation(figure2.1).Conductiveheattransferoccurswhen
twosolidobjectsareindirectcontactwitheachotherandthereisatemperaturegradientbetween
thebody’ssurfaceandacontactingsolidobject.Whenperformingexerciseinastandingposition
such aswalking and running the conductive heat exchanges isminimal since the contact point
betweenthebody’ssurfaceandthegroundisverysmall(KennyandJay,2013).Convectionisthe
heat exchange between the surface of a solid object and a fluid, including air andwater.Heat
transfer via convection to air or fluid occurs when the air or fluid temperature is below the
temperatureofthebody.Anincreaseinwindorfluidcurrentfacilitatesagreaterrateofconvective
heattransfer;furthermore,theheatcapacityofwaterisfargreaterthanairincreasingconvective
heat transfer. Radiation is the heat exchange between a solid object and a large natural, or
manufacturedobjectsuchasthesunoraheater(Sawkaetal.,2011a).

8
2.1.1. Wetheattransferpathways
Evaporativeheat lossoccurswhen liquid changes intowater vapour. Theevaporativeheat loss
capacityisdeterminedprimarilybythewatervapourpressuregradientbetweenthebody’ssurface
andtheenvironment.Thiscanbemodifiedbytheenvironmentandclothing(Sawkaetal.,2011a),
inadditiontophysiologicalalterationinsweatglandactivityandoutput(Taylor,2014).Evaporative
coolingistheprimarymeansofheatremovalduringexercise.Evaporativecoolingistheonlymeans
forheatremovalduringexerciseinanambienttemperaturethatisequalto,orgreaterthanskin
temperature(Tsk).
Figure2.1Avenuesforheatexchangewhenperformingexercise.AdaptedfromGisolfiandWenger,
(1984)
2.1.2. Heatbalance
Theinteractionbetweenfactorsthatincreaseordecreaseheatproductionandheatdissipationand
theresultantimpactuponbodyheatstorageisevidentintheheatbalanceequation(Equation2.2).
Metabolicheatproductionistypically<100Watrestand>1000Wduringhighintensityexercise.
Therateofbodyheatstoragerepresentsheatgainifpositiveandheatlossifnegative.

9
Equation2.2.Heatbalanceequation
S(watts)=M–(±W)±(R+C)±K–E
Where:S=rateofbodyheatstorageinwatts,M=rateofmetabolicenergyproduction,W=mechanical
work,whetherconcentric(positive)oreccentric(negative)exercise,R+C=rateofradiantandconvective
energyexchanges,K=rateofconduction,E=rateofevaporativeloss.
Metabolicheatproductionisthedifferencebetweenmetabolicrateandthefractionofthisenergy
thatisusedtocreateexternalwork.Thehumanbodyisinefficientatperformingexternalworkand
thus80%-100%oftheenergyproducedisreleasedasheatenergyinsidethebody.Forexample,if
aparticipantiscyclingat100W,400Wofthe500Woftotalenergyproductionwillbeheatenergy
insidethebody.
Indirectcalorimetry
Indirectcalorimetry is the traditionalmethod forestimatingmetabolicenergyexpenditure.This
approachrequiresthemeasurementoftherateofoxygen(O2)consumptionandcarbondioxide
(CO2)production.Usingtheenergyequivalentforthefulloxidationofcarbohydrates(21.13kJper
LofO2consumed)andfats(19.63kJperLofO2consumed),metabolicenergyexpenditurecanbe
subsequentlycalculatedusingEquation2.3(KennyandJay,2013).
Equation2.3Calculationformetabolicenergyexpenditure
M=[VbO2((((RER-0.7)/0.3)ec)+(((1.0-RER)/0.3)ef)]/60
Where:VbO2ismeasureinL.min-1;ecisthecaloricequivalentperlitreofoxygenfortheoxidationof
carbohydrates(21130J)andefisthecaloricequivalentperlitreofoxygenfortheoxidationoffat
(19630J).
When direct calorimetry is unavailable, indirect calorimetry is the most precise method of
estimating rate of metabolic energy expenditure and subsequently metabolic heat production
(KennyandJay,2013).However,despiteheatproductionincreasingalmostimmediatelyafterthe
startofexercise,O2consumptionincreasesexponentiallywithatimeconstantof15-30s.Assuch
>1-minofexerciseatthesameexternalworkloadisrequiredtoattainanelevatedsteadystateof

10
O2consumption(AstrandandSaltin,1961;Cheuvrontetal.,2010;WhippandWasserman,1972).
Thus,thedifferencebetweentheactualrateofmetabolicheatproductionatthetissueleveland
itsmeasurementfromanalysingexpiredairshouldbeacknowledged.
Directcalorimetry
Duringboutsofheat imbalance, suchasduring theonsetofexercise,a summationof theheat
exchangeviaconvection,conduction,radiationandevaporationmustbeobtainedandcompared
toconcurrentmeasurementsofmetabolicenergyexpendituretodeterminethequantityofheat
storedinsidethebody(KennyandJay,2013).Themostaccuratemethodforquantifyingtherate
of net heat loss is with direct whole body calorimetry (Jay and Kenny, 2007). Whole body
calorimetry is the simultaneousmeasurementsof the internalheatgeneratedby thebodyasa
resultofanaerobicandoraerobicmetabolismandthetotalheatlosttotheenvironment(Jayand
Kenny,2007).Thus,ifanimbalanceoccursbetweenmetabolicheatproductionandnetheatloss
to the environment, the time dependent differences between thesemeasurements provide an
estimateofthechangeinbodyheatcontent.
2.2. Heatstrain
Heatstrainreferstoanyphysiologicalconsequencesofheatstress(Sawkaetal.,2011b).Measuring
Tc is fundamentaltotheexperimentalstudyoftemperatureregulation inhumans(Sawkaetal.,
1993).Tc istypicallycontrolledat37.0±0.5°Cduringrest.However,Tcvariesbetweendifferent
bodyregionsdependingonthemetabolicrateofthelocaltissues,thesourceandthevolumeof
localbloodflow,inadditiontothetemperaturegradientsbetweenconnectingbodyregions(Sawka
etal.,2011a).
Inarestedstate,theinternalorgansandvisceraproduceapproximately70%ofmetabolicheat,
howeverduringmovement,theskeletalmusclesproduceapproximately80-95%ofmetabolicheat
(Sawkaetal.,2011a).Metabolicheat sourcesarealtered fromrest toexercise.Asa result, the
calculatedchangeinbodytemperaturemaybedisproportionateinonebodyregioncomparedwith
another.Forexample,whenhumansareinarestedstatetheirTc(37.0±0.5°C)istypicallyhigher
than muscle temperature (36.0 ± 0.5°C), however during exercise, the temperature of active
skeletalmuscleusuallyexceedsthatofTc(Kennyetal.,2008)(figure2.2).
11
Figure2.2Meanchangesincoretemperature(A)andmuscletemperature(B)duringandfollowing
60-minexercise.N=8(6M,2F)TakenfromKennyetal.,(2008).
2.2.1. Measurementofcoretemperature
ThemeasurementofTcvariesdependingontheinternalmeasurementsite(Lefrantetal.,2003).It
isessentialthatthemethodadoptedtomonitorTcisconvenientandunbiasedbyenvironmental
conditions (Sawka et al., 2011a). Furthermore, there is a requirement that the measurement
techniquemirrors small changes in arterial blood temperaturequickly, toaccurately reflect the
temperatureofbloodperfusingthehypothalamus(Lefrantetal.,2003).Themostcommonlyused
measurementsites include, therectum,oesophagus,andgastrointestinal tract,butalso include
auditory-canal. Figure 2.3 presents oesophageal, rectal, and intestinal temperature measured
simultaneouslyduringrest,submaximalsupinecyclingexerciseat40%O2uptake(VbO2)peakand
65%VbO2peak,andduringpassiverecovery.

12
Thelocationofrectaltemperature(Tre)isconsideredthemostpracticalandaccurateformeasuring
Tc (Moran andMendal, 2002). Furthermore, Tre is typically very easy to administer and causes
minimaldiscomforttotheparticipants.Heatexchange intherectumisduetotheneighbouring
tissues,ratherthantheregularperfusionofhotblood;thethermalinertiaandpoorheatremoval
results in a stable measurement. However, Tre responds slower to the rapid changes in Tc
experiencedduringtheonsetofexerciseandhasameandifferenceof0.4±1.0°Ccomparedwith
thepulmonaryartery(Robinsonetal.,1998).Oesophagealtemperatureisoftenconsideredtobe
the best non-invasive method to determine Tc because of its deep location, close to the left
ventricle, the aorta, and to the blood flow to the hypothalamus. Furthermore, oesophageal
temperaturerespondsrapidlyandquantitativelytochangesincentralbloodtemperature(Lefrant
etal.,2003)andthepulmonaryarterytemperature(0.0±0.5°C)(Robinsonetal.,1998).However,
oesophageal probes are sometimes deemed as undesirable in many settings because of the
difficultly in inserting the thermistor, irritation to the nasal passage and general participant
discomfort(MoranandMendal,2002).Thegastrointestinalpillprovidescomparablesteadystate
TcmeasurementstoTre.However,thegastrointestinalpillrespondsmoreslowlytothechangesin
Tc than oesophageal temperature (Lee et al., 2000). Furthermore, the gastrointestinal pill
temperaturevalueshavebeenshowntobemorevariableovertimesincethepillistravellingwithin
thegastrointestinaltract(ByrneandLim,2007).
Auditory-canalisanotherestablishedmethodtotrackTc(Cooperetal.,1964;Edwardsetal.,1978;
Haywardetal.,1984)albeitwithanoffsetandaphasedelay(Tayloretal.,2014).Auditory-canal
hasbeenreportedtobecomparabletoTre(Ilsleyetal.,1983).However,surfacetemperaturesare
influencedbyambienttemperatureresultinginareducedaccuracyofauditory-canaltemperature
totrackTc(Lowetal.,2007)
Inconclusion,itappearsthatTre,duetoitsinherentstability,isthepreferredmethodtoaccurately
trackTcwithminimalmeasurementartefact,practicality,andminimalparticipantdiscomfort.
13
Figure2.3Oesophageal,rectal,andintestinaltemperaturemeasuredsimultaneouslyduringrest,
submaximal supine cycling exercise at 40% VbO2 peak and 65% VbO2 peak, and during passive
recovery.N=7(5M,2F).
Notes:Valuesaremean±standarderror.Takenfrom(ByrneandLim,2007;Leeetal.,2000).
2.2.2. Measurementofskintemperature
ThemeasurementofTskisconductedforseveralreasons.Firstly,themeasurementofTskenables
thecalculationofmeanbodytemperaturetopermitthedeterminationofheatstorage(Jayand
Kenny, 2007). Tsk measurements can also be used to estimate radiative and convective heat
exchange,skinconductance,andskinbloodflowrequirements.Furthermore,Tskmeasurements
can be used to inform the Tsk input to the thermoregulatory controller (Sawka et al., 2011a).
AlthoughTskcanbeassessedeasily,itcanalsobeproblematic,sincechangesinTskcanbecaused
byphysiologicaladjustmentssuchasalterationsinthecutaneousbloodflow,secretionofsweat
andevaporation,orchangestotheenvironmentalconditions(Sawkaetal.,2011a).Ensuringthat
the thermistors stay in thermal contact with the skin is paramount to the accuracy of Tsk
measurement(Smithetal.,2010).Thermistorshavebeenreportedtobearobustandaccurateto
0.05°Cacrossarangeofwaterbathtemperature(10-40°C)(Harper-Smithetal.,2010)butmore
variablewhenexercising(technicalerrorofmeasure[TEM)]=0.3°C)(Jamesetal.,2014).

14
MeanTskisoftencalculatedbasedonmultiplemeasurementstobetterreflectthechangesinTsk.
MeanTsk represents the sumof theweighted individualmeasurementsofTsk. Theweightingof
meanTskisestablishedbasedonthepercentageofbodysurfacearea(BSA)thatisrepresentedby
themeasurementsite(MitchellandWyndham,1969).TwelvetofifteenTskmeasurementsiteshave
beenusedtocalculatemeanTsk.However,investigatorsreducedthenumberofTskmeasurement
sitesrequiredtoobtainavalidestimateofmeanTsk.Whenitisproblematictomeasureaccurately
a large number of sites, the equation (Equation 2.4) introduced by Ramanathan should be
considered(Ramanathan,1964).
Equation2.4.Calculationformeanskintemperature(Tsk)
Tsk=0.3·(Tchest+Tarm)+0.2·(Tupperleg+Tlowerleg)
Where:Tchest,themidpointofthepectoralismajor;Tarm,themidpointofthetricepsbrachiilateral
head;Tupperleg,rectusfemoris;Tlowerleg,gastrocnemiuslateralhead.
In contrast, mean Tsk can be calculated based on the regional weighting of the skins thermal
sensitivity. The thermal receptors within the skin are not evenly distributed. As a result, the
warmingofbodyregionswhichhavethemostthermalsensors,ultimatelyhasthelargestinfluence
ontheeffectorresponses(Sawkaetal.,2011a).Thus,itissuggestedthatcalculatingmeanTskusing
thermal sensitivityweightingsmayprovidea superior index,whichmoreaccurately reflects the
peripheral input to the thermoregulatory controller. However, there remains some level of
disagreement as towhether themean Tsk values actually differ if calculated based on regional
thermalsensitivityweightingsorregionalBSAweightings(Libertetal.,1984).
2.2.3. Bodyheatcontent
Intheeventofanimbalancebetweenheatproductionandheatlossthereisarateofchangein
body heat storage and subsequently, a change in body heat content. Body heat content is the
productofmeanbody temperatureand theaverageheat capacityof thebody tissues (Jayand
Kenny, 2007). The determination of body heat content is of fundamental importance when
assessing the exposure of the human body to a hot environment thatmay result in a thermal
imbalance.Themeasurementofbodyheatexchangeusingsimultaneousmeasuresoftotalheat
productionandheatloss,intheoryistheonlymethodwherebybodyheatcontentcanbedirectly
determined(JayandKenny,2007).Therefore,bydefinitionthedifferencebetweenmetabolicheat

15
productionusingindirectcalorimetryandthetotalheatlostfromthebodyusingdirectcalorimetry
is theonlymethodtocalculate the rateofchange inbodyheatstorageandbodyheatcontent
accurately(JayandKenny,2007).Changesinbodyheatcontentarerarelymeasureddirectlyusing
calorimetry since this approach requires very sophisticated and expensive equipment and
instrumentation(JayandKenny,2007).Typically,changesinbodyheatcontentareestimatedusing
thetwocompartmentthermometrymodelwhichinvolvesthemeasurementofchangesinmean
bodytemperatureestimatedfromweightedTcandmeanTskmeasurements(JayandKenny,2007)
(figure2.4).ForthismethodtheTcandmeanTskvaluesareweightedbytheirrelativesizewhich
variesreciprocallywithcutaneousvasodilationandvasoconstrictionwithcore/skinweightingsof
0.90/0.10,0.79/0.21,and0.66/0.34inhot,temperate,andcoolconditions(Sawkaetal.,2011a).
The three-compartment thermometry model (figure 2.4) is reported to be a superior model
comparedwiththetraditional two-compartmentmodelduringexercise intemperateandwarm
conditions(JayandKenny,2007).Thethree-compartmentthermometrymodelusesmeasuresof
Tc, Tsk, and muscle temperature collectively, to predict changes in mean body temperature.
However, even when including invasive muscle temperature measurements, the three-
compartmentthermometrymodelonlyaccountsforapproximately50%ofthevarianceofmean
bodytemperature(JayandKenny,2007).Thisevidencesuggeststhattemperaturemeasurements
of more tissues intermediate to the core and shell, are required to provide a more accurate
thermometricderivationofbodyheatcontent.Subsequently,atwo-compartmentalthermometry
modelwasintroduced(JayandKenny,2007).Thismodelonlyaccountsforapproximately56%of
thevarianceofmeanbodytemperaturefromcalorimetrymeasurements.Thesefindings,suggest
that thermometry provides inaccurate estimates of mean body temperature changes, and
therefore thermoregulatory models to determine body heat content are likely flawed. Thus,
thermometrydeterminationofbodyheatcontentshouldbeusedwithcautionandonlyemployed,
whenusingarepeatedmeasuresdesign.
16
Figure2.4The2-compartmentthermometrymodelandthe3-compartmentthermometrymodelforchangeinmeanbodytemperatureandchangeinbodyheatcontent.
Adaptedfrom(JayandKenny,2007;KennyandJay,2013).

17
2.3. Behaviouraltemperatureregulation
Humans typically regulate their Tc within a narrow range between 35°C and 41°C using a
combination of behavioural and physiological responses. Thermoregulatory behaviour, is the
regulationofbodytemperaturebyacomplexpatternofmuscularskeletalresponsestoheatand
cold,whichmodifytheratesofheatproductionand/orheatloss(BlighandJohnson,1973).
Behavioural temperature regulation is often the first line of defence in maintaining body
temperature and involves a number of conscious alterations in behaviour. Examples of such
behaviours include,butarenot limitedto,modifying theexercise intensity,addingor removing
clothing,alteringbodyposition,andseekingshadeorshelter(Schladeretal.,2011b;Tuckeretal.,
2006).Schladeretal.,(2011a)reportedashortertimetoexhaustion(20.3±3.4minversus23.2±
4.1min),higherpeakTre(39.4±0.3°Cversus39.1±0.4°C),andhigherratingsofperceivedexertion
(18versus16)whenexercisewasperformedatafixedintensitycomparedwithself-pacedexercise,
in an uncompensable environment. Thus, when self-paced exercise is performed in an
uncompensableenvironment,itislessphysiologicallydemandingsincethevoluntaryreductionsin
exerciseintensityreducemetabolicheatproduction,improvethermalcompensability,andincrease
exerciseduration.Thisevidenceissupportedbythehighincidencerateofexerciseinducedheat
strokewhenhighmotivationandexternalfactorsdictateexerciseintensityandalowerincidence
whenpeopleexercisealoneorself-paced(Armstrongetal.,2007;Epsteinetal.,1999).Athletes
oftenignorebehavioursdesignedtosupportthermoregulationduetoahighlevelofmotivation
duringcompetition.
2.4. Physiologicaltemperatureregulation
Physiological temperature regulation is controlled by responses that are independent to our
conscious voluntary behaviours. Examples of such responses include, but are not limited to,
modifyingthebodyheatdistributionviaredistributingbloodflowthroughthecoreandtheskinby
cutaneousvasodilationandvasoconstriction,andalteringtherateofmetabolicheatproduction
through involuntary behaviours such as shivering, and sweating. There are several theories
developedwhichtrytoexplainhumanthermoregulatorycontrol;theseincludetheadjustableset
pointmodel,thereciprocalinhibitionmodel,andtheheatregulationmodel.

18
2.4.1. Adjustablesetpoint
Thetraditionalmodelofthermoregulatorycontrol,knownasthesetpointmodelwasintroduced
byHaroldHamel(figure2.5).Thissystemassumesthatafferentsignalsfromthecoreandperiphery
are combined into a thermal signal transmitted through the central nervous system and likely
withinthepreopticanteriorhypothalamus(Hammeletal.,1963).Thismodelisinpartinformedby
warmandcoldreceptorsunderthehumanskin.Thesereceptorsaretypicallylocatedatadepthof
0.3-0.6mmand0.15-0.17mm,respectively(Boulant,2006).Furthermore,neuralsupportforthis
model is highlighted by the observation that the hypothalamus holdswarm and cold sensitive
neuronswhichaltertheirfiringratebasedoneitherwarmingorcooling.Thisthermalsignalisthen
comparedtoaninternallystoredthermalsignal,resultingintheappropriateactivationofheatloss
orheatgaineffectorresponses(Hammeletal.,1963).
Acommonargumentincontradictionoftheadjustablesetpointmodelisthatitdoesnotexplain
any deviations from the reference temperature stored within the central nervous system. For
example, during the luteal phase of the menstrual cycle, baseline temperature is regulated
approximately 0.3-0.5°C higher than compared with the follicular phase (Coyne et al., 2000).
Similarly,duringafeverbodytemperatureisraisedandgenerally,appropriatelydefended.Theset
pointmodeloftemperatureregulationarguesthatthereisnorequirementforaconstantreference
temperature;instead,thesetpointtemperaturemayvaryoverabroadrangebasedontheactivity
ofwarmandcoldsensitiveneurons(Cabanac,2006).
19
Figure2.5Schematicdiagramofthethermoregulatorycontrolsystem.AdaptedfromSawkaetal.(2011).
20
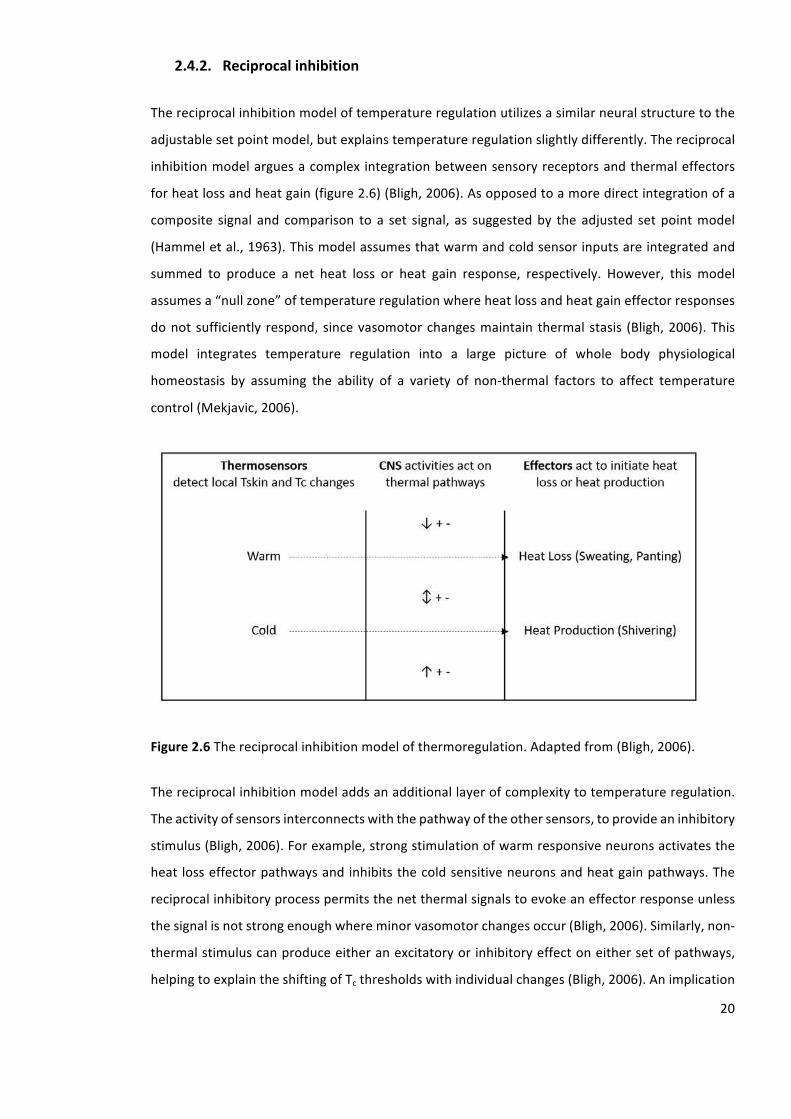
2.4.2. Reciprocalinhibition
Thereciprocalinhibitionmodeloftemperatureregulationutilizesasimilarneuralstructuretothe
adjustablesetpointmodel,butexplainstemperatureregulationslightlydifferently.Thereciprocal
inhibitionmodelarguesacomplex integrationbetweensensoryreceptorsandthermaleffectors
forheatlossandheatgain(figure2.6)(Bligh,2006).Asopposedtoamoredirectintegrationofa
composite signal and comparison to a set signal, as suggestedby the adjusted setpointmodel
(Hammeletal.,1963).Thismodelassumesthatwarmandcoldsensorinputsareintegratedand
summed to produce a net heat loss or heat gain response, respectively. However, this model
assumesa“nullzone”oftemperatureregulationwhereheatlossandheatgaineffectorresponses
donotsufficiently respond, sincevasomotorchangesmaintain thermal stasis (Bligh,2006).This
model integrates temperature regulation into a large picture of whole body physiological
homeostasis by assuming the ability of a variety of non-thermal factors to affect temperature
control(Mekjavic,2006).
Figure2.6Thereciprocalinhibitionmodelofthermoregulation.Adaptedfrom(Bligh,2006).
Thereciprocalinhibitionmodeladdsanadditionallayerofcomplexitytotemperatureregulation.
Theactivityofsensorsinterconnectswiththepathwayoftheothersensors,toprovideaninhibitory
stimulus(Bligh,2006).Forexample,strongstimulationofwarmresponsiveneuronsactivatesthe
heatlosseffectorpathwaysandinhibitsthecoldsensitiveneuronsandheatgainpathways.The
reciprocalinhibitoryprocesspermitsthenetthermalsignalstoevokeaneffectorresponseunless
thesignalisnotstrongenoughwhereminorvasomotorchangesoccur(Bligh,2006).Similarly,non-
thermalstimuluscanproduceeitheranexcitatoryorinhibitoryeffectoneithersetofpathways,
helpingtoexplaintheshiftingofTcthresholdswithindividualchanges(Bligh,2006).Animplication

21
of thismodel, is that there isnocentral comparator signalas impliedby theadjustedsetpoint
modelwhichpermitseasiertheoreticalabilityforTcshifts(Sawkaetal.,2011a).Anatomicalsupport
for this ideacanbeseen inclinical tests inwhich thehypothalamus is impairedorblockedand
thermoregulatorycontrol,althoughcoarser,isstillpossible(Sawkaetal.,2011a).
2.4.3. Heatregulation
Theheatregulationmodelisdifferentinthetheoreticalconstructfromtheadjustedsetpointand
thereciprocalinhibitionmodelsofphysiologicaltemperatureregulation.Theunderlyingprinciple
oftheheatregulationmodelisthattheoverallheatstorage,orthehomeostaticnetbalanceofheat
gainedversusheat lost is theregulatedvariable.Suchthatbodytemperature isnotaregulated
variable, it is the resultof thebody’sattempt tomaintaina stableoverallheatbalance (Webb,
1995).AsopposedtosensingandintegratingperipheraltemperatureandTc,thebodyusesheat
flowandthetemperaturegradientacrosstheskinsurfaceasitsprimaryafferentinputs.Thecentral
integrationof theheatbalanceoperateswith feedback fromheat lossand feedforwardcontrol
fromheatproduction,drivingthephysiologicalresponsesthatdefendbodyheatcontent(Webb,
1995).
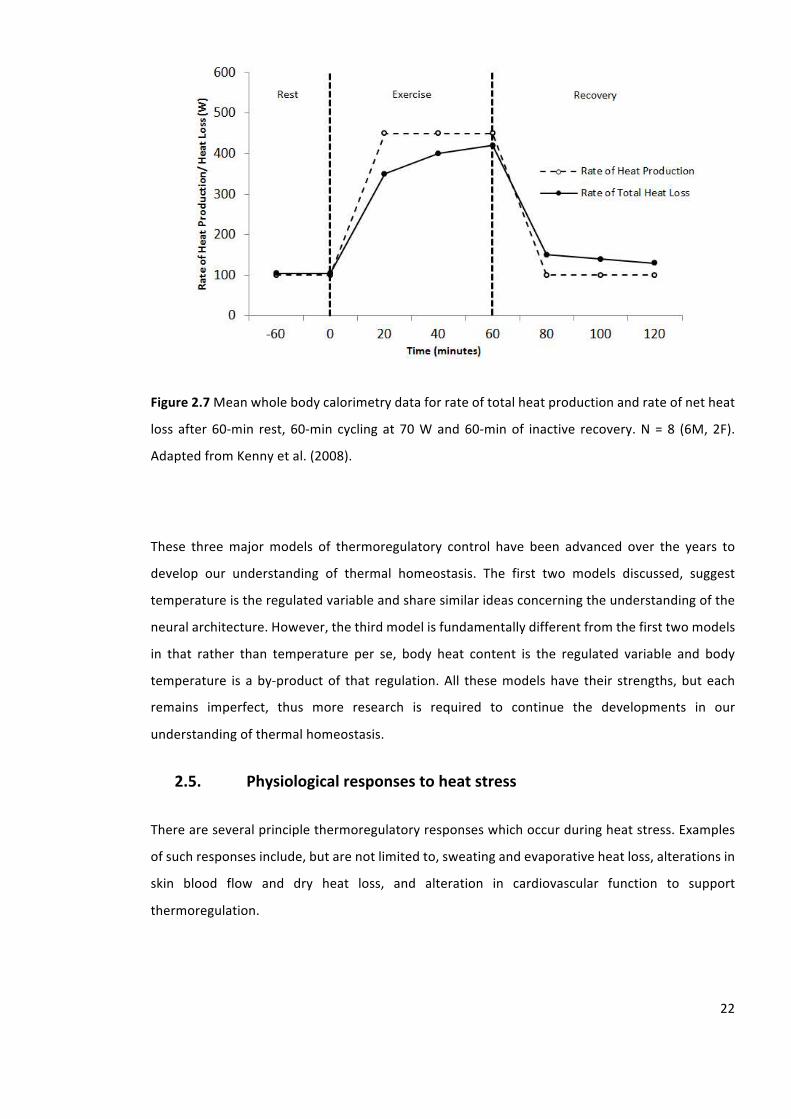
Theprimarystrengthoftheheatregulationmodeloftemperatureregulationisthatitissensitive
tothedeviationsinbodytemperature.Asapracticalexample,whenacyclistbeginsexercisingthere
isaconcurrent increase inTc,this is followedbyastabilisationofTcuntilexercise isterminated
(figure2.7).Heatproductionrespondsmorequicklycomparedtotheheatdissipationmechanism
however,eventuallytheheatlossmechanismsmatchestherateofheatproduction,resultingina
stable elevated Tc.Whenexercise is terminated, Tc remains elevated for a periodof timeeven
though heat dissipating responses are reduced rapidly (Kenny et al., 2008) (figure 2.7). This is
difficulttoaccommodateinthetemperatureregulationmodelssince itwouldassumethatheat
dissipating responseswould remainelevated throughout the recoveryperioduntilTc returns to
resting levels. The primary argument against the heat regulation model is the anatomical
requirement for heat flow sensors which are anatomically difficult to demonstrate, whilst
accountingforthepresenceoftemperaturesensitiveneuronsthroughouttheperipheriesandthe
central nervous system. In response supporters for the heat regulation model argue that the
thermal receptors throughout the body representmultiple thermal sensors, and the difference
betweenthesethermalsignalscanbecomeintegratedasratesofheatflow.
22
Figure2.7Meanwholebodycalorimetrydataforrateoftotalheatproductionandrateofnetheat
loss after60-min rest, 60-min cyclingat70Wand60-minof inactive recovery.N=8 (6M,2F).
AdaptedfromKennyetal.(2008).
These threemajormodels of thermoregulatory control have been advanced over the years to
develop our understanding of thermal homeostasis. The first two models discussed, suggest
temperatureistheregulatedvariableandsharesimilarideasconcerningtheunderstandingofthe
neuralarchitecture.However,thethirdmodelisfundamentallydifferentfromthefirsttwomodels
in that rather than temperature per se, body heat content is the regulated variable and body
temperature is aby-productof that regulation.All thesemodelshave their strengths,buteach
remains imperfect, thus more research is required to continue the developments in our
understandingofthermalhomeostasis.
2.5. Physiologicalresponsestoheatstress
Thereareseveralprinciplethermoregulatoryresponseswhichoccurduringheatstress.Examples
ofsuchresponsesinclude,butarenotlimitedto,sweatingandevaporativeheatloss,alterationsin
skin blood flow and dry heat loss, and alteration in cardiovascular function to support
thermoregulation.

23
2.5.1. Sweatingandevaporativeheatloss
Asambienttemperatureormetabolicheatproductionincreases,thereisincreasingdependence
uponthesudomotorfunctionandevaporativeheatlosstodissipatebodyheat.Therearetypically
twotypesofhumansweatglands,apocrineglandsandeccrineglands(Shibasakietal.,2006).The
eccrinesweatglandareprimarilyresponsibleforthermoregulatorysweatinginhumans(Satoetal.,
1989).
Eccrinesweatglands
Eccrinesweatglandsaredistributedovernearlytheentiresurfaceofthehumanbody.Thenumber
ofhumansweatglandstypicallyrangesfrom1.6to4.0million.Theeccrinesweatglandstructure
consistsofabulboussecretorycoilwhichleadstoaduct.Thissecretorycoilislocatedinthelower
dermisandtheductextendsthroughthedermallayersandopensdirectlyontotheskinsurface.
Theuncoileddimensionofthesecretoryportionoftheeccrinesweatglandisapproximately30-50
µmindiameterand2-5mminlength.Theadultsecretorycoilrangesfrom1to8x10-3
mm3
(Sato
andSato,1983).Thereisapositivecorrelationbetweenthesizeofanindividualsweatglandand
themaximalsweatrate(SWR)ofthatgland(SatoandSato,1983).
Given the challenges of identifying neural tracts in humans, the exact neurological pathways
responsible for thermoregulatory sweating are not entirely understood. Evidence from animal
studiessuggeststhatefferentsignalsfromthepreoptichypothalamustravelviathetegmentumof
theponsandthemedullaryrapheregionstotheintermediolateralcellcolumnofthespinalcord.
In the spinal cord, neurons emerge from the ventral horn, pass through the white ramus
communicans,combinewithperipheralnervesandtraveltosweatglands(Low,2004;Nakamura
etal.,2004).
Sweatrateandevaporativeheatloss
Theinitialincreaseinsweatrate(SWR)duringheatstressisaresultofanincreaseintheactivated
sweatglands;however,additionalincreasesinSWRoccurthroughanincreaseintheproduction
and secretionof sweatpergland (Kondoet al., 2011;Randall, 1946). The recruitmentof sweat
glandsisrapid,withnearmaximalrecruitmentbeingachievedinaslittleas8-minofexerciseand
passiveheatstress.Incontrast,theincreasesinsweatoutputperglandaremoregradual(Kondo
etal.,2011).Theratethatsweatevaporates,isdeterminedbythegradientbetweentheskinand
airvapourpressures,andthecoefficientofevaporativeheattransfer.Thewiderthewatervapour
gradient, the greater the rate of evaporation (Sawka et al., 2011a). Furthermore, warm skin

24
enhancesthesweatresponse(Nadaletal.,1971),whichislikelymediatedbyboththeincreased
localtemperatureandassociatedincreasedskinbloodflow(Wingoetal.,2010).
Heat loss through the evaporation of sweat is an essential process in the thermoregulatory
responsetoexercise.Forexample,whenanindividualperformsexerciseatametabolicrateof600
W,approximately80%oftheenergyconsumedbecomesheat.Assuch,480W(28.8kJ.min−1
)must
bedissipatedtoavoidheatstorage.Thespecificheatofbodytissueapproximates3.5kJ.kg.°C−1
,so
a70kgmanora60kgfemalehasaheatcapacityof245kJ.°C−1
or210kJ.°C−1
,
respectively.Ifsweat
isnotsecreted,Tcincreasesbyapproximately1.0°Cevery8-min30-sforthe70kgmale(245kJ.°C−1
/28.8kJ.min−1
)or7-min20s forthe60kgfemales(210kJ.°C−1
/28.8kJ.min−1
).However,asthe
latent heat of evaporation is 2.43 kJ.g−1
, secretion and evaporation of ∼12 g of sweat (28.8kJ.min
−1
/2.43kJ.g−1
)perminwouldenabletheheattobetransferredtotheenvironmentavoiding
heatstorage(Sawkaetal.,2011a).
Whenperformingexercise inahighambientcondition,humanshave thecapacity toproducea
remarkableamountofsweat.Individualsperformingactivitiesinbothadesertandtropicalclimates
havebeenreportedtoexperienceameanSWRof1.6L.hr-1
whenpartiallyclothed,and2.0L.hr-1
whenwearingfullprotectiveclothing(Montainetal.,1994).Factorssuchasexerciseintensityand
duration,clothing,andenvironmentalconditionsultimatelyaffectanathlete’sSWR.ObservedSWR
inavarietyofathletesduringtrainingandcompetition,rangefrom0.2L.hr-1
to3.4L.hr-1
(Sawkaet
al.,2007).
2.5.2. Skinbloodflowanddryheatloss
In thehumanbody,bloodtransfersheatbyconvection fromthedeepbody tissues to theskin.
Bloodflowtothecutaneouscirculationhasatremendousrange,fromnearlyzeroinextremecold
toapproximately7L.min-1
whenTc ishigh(JohnsonandKellogg,2010).Reflexescontrollingskin
blood flowhave the capacity to use nearly this entire range.When thermoregulatory sweating
begins,skinbloodflowservesasameanstodeliverheattotheskinwhichisthenremovedbysweat
evaporation(Sawkaetal.,2011a).Thus,skinbloodflowandsweatingworktogethertodissipate
theheat.
Vasoconstrictionandvasodilationcontrolofskinbloodblow
SkinbloodflowisultimatelyaffectedbyTsk,whichdirectlyimpactsonthevascularsmoothmuscle.
CutaneouscirculationisaffectedbytheTskandTc,viareflexesoperatingthroughthesympathetic
nervoussystemfromthethermoregulatorycontrolcentres(JohnsonandKellogg,2010).Certain

25
areas of the body, including the palm of the hand, sole of the foot, lips, ears, and nose, are
predominantly innervated by adrenergic vasoconstrictor fibres (Sawka et al., 2011a). The
vasodilatationactivitythatoccursintheseregionsislargelyduetowithdrawingthevasoconstrictor
activity(Tayloretal.,1984).ThereisminimalvasoconstrictoractivitywhenTskexceeds39°Conthe
remaining body regions, including the arms, legs and torso. Active vasodilatation during heat
exposure depends largely on sympathetic innervations (Taylor et al., 1984). Skin blood flow
responses differ between rest and aerobic exercise during heat stress (González-Alonso et al.,
2008).Exercise increasesvasoconstrictoractivitywhich increases the threshold temperature for
vasodilatationandreducesskinbloodflowathighbodytemperaturescomparedtorest(Sawkaet
al.,2011a).Thesemodifiedskinbloodflowresponsesoccurduringexerciseheatstressbecausethe
cardiovascularsystemischallengedtosupportsimultaneouslybothhighskeletalmusclebloodflow
andskinbloodflow.
Coreandskintemperatureeffectsonskinbloodflow
Table 2.1. illustrates the effect of different Tc and Tsk on the skin blood flow requirements to
preserve heat transfer (Kenefick et al., 2007). In the example, if the net heat production is 7.7
kcal.min-1
andTcandTskare38°Cand30°C,respectively,theskinbloodflowrequirementwillbe
~1.1L.min−1
.WhenTskincreasesto34°Cand36°Casaresultofheatstress,theminimalskinblood
flowrequirementtopreservethesameheattransferrateincreasesto~2.2L.min−1
and~4.4L.min−1
,
respectively.However, ifTc rises to39°C theskinblood flowrequirementsare reducedwithout
compromising heat transfer. This example, demonstrates how an elevated Tsk as a result of a
warmer ambient or reduced evaporative cooling, increases skin blood flow requirements. In
addition,theexampledemonstrateshowanelevatedTcmayreduceskinbloodflowrequirements
andcardiovascularstrainassociatedwithexercise-heatstress.
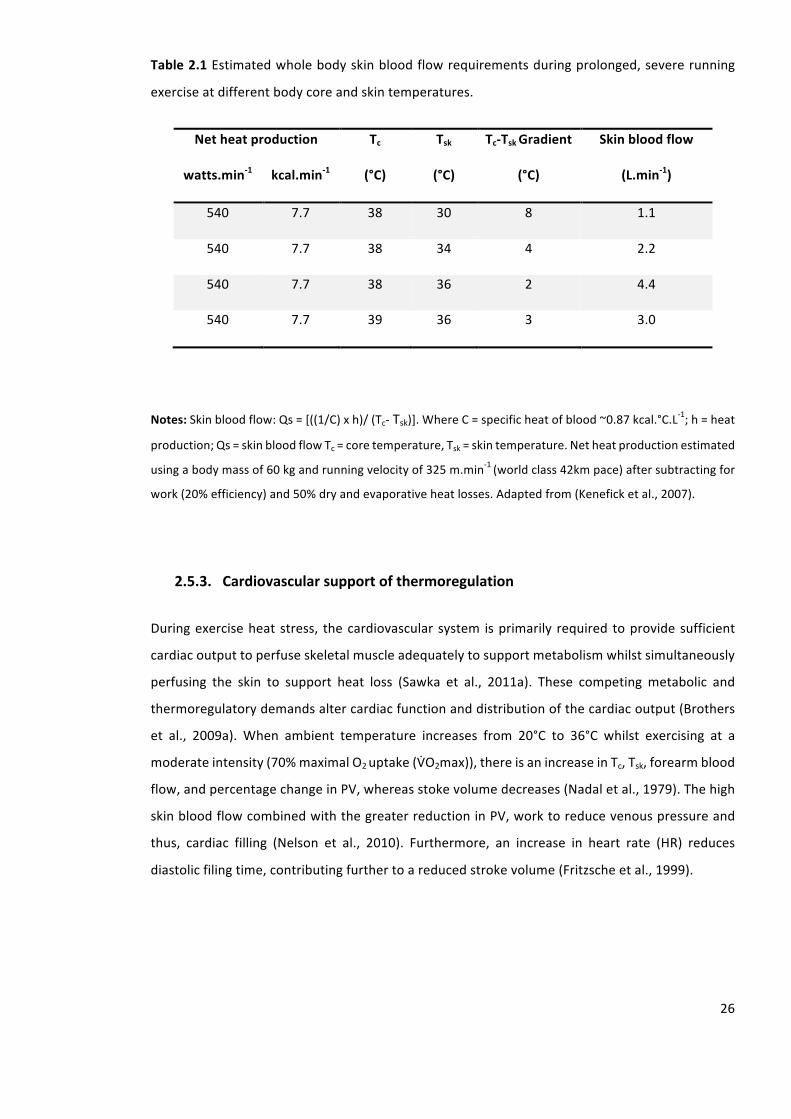
26
Table2.1Estimatedwholebodyskinbloodflowrequirementsduringprolonged,severerunning
exerciseatdifferentbodycoreandskintemperatures.
Netheatproduction Tc
(°C)
Tsk
(°C)
Tc-TskGradient
(°C)
Skinbloodflow
(L.min-1)watts.min-1 kcal.min-1
540 7.7 38 30 8 1.1
540 7.7 38 34 4 2.2
540 7.7 38 36 2 4.4
540 7.7 39 36 3 3.0
Notes:Skinbloodflow:Qs=[((1/C)xh)/(Tc-Tsk)].WhereC=specificheatofblood~0.87kcal.°C.L-1
;h=heat
production;Qs=skinbloodflowTc=coretemperature,Tsk=skintemperature.Netheatproductionestimated
usingabodymassof60kgandrunningvelocityof325m.min-1
(worldclass42kmpace)aftersubtractingfor
work(20%efficiency)and50%dryandevaporativeheatlosses.Adaptedfrom(Keneficketal.,2007).
2.5.3. Cardiovascularsupportofthermoregulation
Duringexerciseheatstress, thecardiovascular system isprimarily required toprovidesufficient
cardiacoutputtoperfuseskeletalmuscleadequatelytosupportmetabolismwhilstsimultaneously
perfusing the skin to support heat loss (Sawka et al., 2011a). These competingmetabolic and
thermoregulatorydemandsaltercardiacfunctionanddistributionofthecardiacoutput(Brothers
et al., 2009a). When ambient temperature increases from 20°C to 36°C whilst exercising at a
moderateintensity(70%maximalO2uptake(VoO2max)),thereisanincreaseinTc,Tsk,forearmblood
flow,andpercentagechangeinPV,whereasstokevolumedecreases(Nadaletal.,1979).Thehigh
skinbloodflowcombinedwiththegreaterreductioninPV,worktoreducevenouspressureand
thus, cardiac filling (Nelson et al., 2010). Furthermore, an increase in heart rate (HR) reduces
diastolicfilingtime,contributingfurthertoareducedstrokevolume(Fritzscheetal.,1999).

27
Skintemperatureandcardiovascularstrain
An increase in Tsk andanarrower Tcto Tsk gradient,which typically occursduringexerciseheat
stress,resultsinagreatercardiovascularstrain.Cheuvrontetal.(2003)demonstratedtheeffectof
analteredTctoTskgradientoncardiovascularstrain.WhenTcremainsconstant(37.5°C),whilstTsk
is increasedfrom32°Cto35°C,HRincreasedbyapproximately26beats.min-1
.Inaddition,when
raisingTskfrom32°Cto36°CHRincreasedbyapproximately49beats.min-1
.Thisevidencesuggests
thatanarrowTctoTskgradientresultsinagreaterstrainonthecardiovascularsystem.
Redistributionofbloodflowwithexerciseheatstress
Duringexerciseheatstress,thecardiovascularsystemsabilitytodeliveradequatebloodsupplyto
theskeletalmuscleandcerebrumcanbecompromised(González-Alonsoetal.,2008;Nyboetal.,
2002;Rasmussenetal.,2010).Although,cardiacoutputissustainedduringlowintensityexercise
in the heat, cardiac output is compromised atmoderate to high intensity exercise in the heat
(Gonzalez-Alonso,2003).Tosupportthedemandsofthecardiovascularsystemduringexerciseheat
stress,bloodisredirectedfromthevisceratotheskinandactiveskeletalmuscles(Sawkaetal.,
2011a).Thevisceralbloodflowreductionsaregradedtotheexerciseintensity,andtheeffectsof
exercise and heat are additive. Secondary to the reduced visceral blood flow, blood can be
mobilizedfromthecompliantsplanchnicbedstohelpmaintaincardiacfillingduringexerciseheat
stress(Sawkaetal.,2011a).
Cardiovascularcomplicationsassociatedwithexerciseheatstress
Thehighdemandsplacedon thecardiovascular systemduringexerciseheat stressmay lead to
syncopeandorthostatic intolerance(KenefickandSawka,2007).Hyperthermiaanddehydration
areoftenrelatedandmaymodifycardiacfilling,bloodpressure,andbaroreceptorreflexregulation,
whichcontributetoorthostaticintolerance(Charkoudianetal.,2003;Crandalletal.,2010;Wilson
etal.,2009).Inaddition,hyperthermiacanreducelocalveno-arteriolarresponses(Brothersetal.,
2009b)andreducecerebralvascularconduction(Wilsonetal.,2006)andaccompanyingcerebral
perfusionduringorthostaticchallenges.
2.6. Sexdifferencesinphysiologicalresponsestoheatstress
Sexhastraditionallybeenconsideredanindependentmodulatoroftemperatureregulation.Ithas
beenwellreportedthatmales’andfemales’thermoregulatoryresponsestoexerciseintheheat
differ(Kaciuba-UscilkoandGrucza,2001;Kenney,1985;Nunneley,1978).Thetwocommonheat

28
lossavenueswhichhavebeencomparedbetweenmalesandfemalesareevaporative(sweating)
anddry(skinbloodflow)heatloss.
2.6.1. Sweatrateandevaporativeheatloss
Females,havepreviouslydemonstratedalowerSWRwhilstmaintainingasimilarTctothatofmales
(Avellinietal.,1980;Keatisuwanetal.,1996).TheprimaryconcernwithcomparingSWRbetween
malesandfemalesisthattypicallyexperimentaldesignsadoptaprotocolusingarelativeVoO2max.
AdministeringexerciseatarelativepercentageofVoO2maxwillresultinsubstantialdifferencesin
absoluteO2consumption(Keatisuwanetal.,1996).Forexample,amaleandfemalewithabody
mass(BM)of70kgand60kgrespectively,bothwitharelativeVoO2maxof50mL.kg-1
.min-1
willboth
havearelativeO2consumptionof25mL.kg-1
.min-1
whenexercisingat50%ofVoO2max.However,
males will have an absolute O2 consumption of 1,750 mL.min-1
and females
1,500 mL.min-1
respectively. This 250 mL.min-1
difference provides a metabolic heat production that is
approximately 85W greater inmales comparedwith females (Gagnon et al., 2008). The lower
resultantrateofheatproductioninfemaleswouldtherefore,havenecessitatedasmalleramount
ofevaporationfromtheskinandsubsequentlyalowersweatproductiontosatisfybiophysicalheat
balance requirements. Thus, the previously reported thermoregulatory differences observed
between males’ and females’ SWR may be confounded by the inherent physical differences
betweenthesexes.
Sexdifferencesintemperatureregulationduringexercisehavebeenshowntoexistirrespectiveof
confoundingdifferencesinmetabolicheatproductionandphysicalcharacteristics.However,these
differencesonlybecomeevidentaboveacertainrequirementforheatlossandaresolelyattributed
toalowersweatglandoutputinfemales(GagnonandKenny,2012).Specifically,femalesexhibita
lowerincreaseinbothlocal(GagnonandKenny,2012)andwholebody(GagnonandKenny,2012,
2011)sudomotoractivityasafunctionofincreasesinTc(i.e.,thermosensitivity),withnodifferences
in the onset threshold of these responses. The lower thermosensitivity of sudomotor activity,
combinedwith a lack of sex differences in the onset threshold suggests that sex differences in
sudomotoractivityduringexercisearemediatedperipherally(GagnonandKenny,2011,2012).
2.6.2. Skinbloodflowanddryheatloss
To date, there is very little data published evaluating the sex differences in the sensitivity of
cutaneous vasodilation using methods that eliminate the inherent limitation associated with
exercise intensityprescribedwhencomparingmalesand females, asdescribed in section2.6.1.

29
Following passive heating, Inoue et al. (2005) reported that females have a relatively greater
cutaneousbloodflowonthethighcomparedwithmales,howevernodifferenceswereobserved
atanyoftheothersites(Inoueetal.,2005).Thehighercutaneousbloodflowwassuggestedto
reflect a greater degree of cutaneous vasodilation on this site, since no sex differences were
observed in themeanarterialpressure(Inoueetal.,2005).Theseresultssuggest, that females’
dependenceuponthecutaneousvasodilationresponseforheatlossisgreaterthanthesweating
response. Furthermore, it was suggested that the regional differences between sexes in
vasodilatationandheatdissipationonthethighmayberelativelygreaterinfemalesthaninmales.
Itislikelythatheatdissipationfromthelimbsismoreeffectivethanheatdissipationfromthetrunk,
becausetheBSA:BMratioofthelimbsisgreater.Conversely,GagnonandKenny(2012)reported
nodifferenceinskinbloodflowresponsesbetweenmalesandfemales,followingexerciseatafixed
requirementofheatloss.TheseobservationsarefurthersupportedbyGagnonetal.(2013)who
reportednodifferencesbetweenmalesandfemales’cutaneousvascularconductancetovarying
dosesofacetylcholineandsodiumnitroprusside.Althoughthereremainsnoconclusiveevaluation
of the skin blood flow differences between males and females, it remains an important
considerationwhencomparingthermoregulatoryresponsesbetweensexes.
2.7. Menstrualcycle
Themenstrualcycleisaseriesofvarioushormonereleasesthatcoordinatethereadinessofthe
femalereproductivesystemforconception(figure2.8).Atypicalmenstrualcycleoccursovera28-
dperiodandisbrokenupintotwophases;thefollicularphase(D1–D13)andthelutealphase(D15
–D28).Othercommonterminologymaybeusedtodescribethedifferentmenstrualcyclephases
(table2.2).Ondayoneofthemenstrualcycle,thehypothalamusreleasesgonadotropin-releasing
hormonewhichcontrolsstimulationandinhibitionofthereleaseoffollicular-stimulatinghormone
(FSH)andluteinisinghormone(LT)fromtheanteriorpituitary.Thesehormonesthenregulatethe
release of oestrogen from the ovarian follicles and progesterone from the corpus luteum,
coordinatingtheprogressionoftheseeventsduringanaveragecycleof28-d(MarshandJenkins,
2002)(figure2.8)
An essential consideration to make when conducting physiological research within a female
populationisthetimingoftestinginrelationtothemenstrualcycle.Thefluctuationsinhormones
associatedwiththemenstrualcyclemayalterthefunctionofanumberofdifferentphysiological
responses associated with thermoregulation later discussed in section 2.9. The most common
methodtominimisethechangesinhormoneconcentrationsassociatedwiththemenstrualcycle,
thatmayinfluencethedependentvariable,istostudyfemalesinthesamephaseoftheirmenstrual

30
cycleacrossparticipants,oracrossstudydaysinlongitudinalstudies.Femalesaremostcommonly
tested during the early follicular phase (D1-D7), when both oestrogen and progesterone
concentrationsareattheirlowestlevels.
Despitetheconvenienceofstudyingfemalesintheearlyfollicularphaseofthemenstrualcycle,the
broaderclinicalrelevanceofthefindingsmaybelimitedsincefemalesareinthispartoftheircycle
foronly~25%oftheirreproductivelives.Although,oestrogenandprogesteroneconcentrationsare
lowrelative tootherphasesof thecycle,hormone levels remainconsiderablyhighercompared
withthoseofmen.Thus,theirimpactwhenmakingcomparisonsbetweenthesexesisnotentirely
eliminated.Researcherscanattempttokeepthecycledaysconsistentacrossfemaleswithinagiven
studytoinvestigatethephysiologicaleffectsindependentofcyclephase,buttheymusttakeinto
account the variability of these hormones across cycle phases within females. Additional
considerationsforhormoneverificationincludethelargevariabilitybetweenindividualswithinthe
same menstrual cycle phase, the diurnal variation in hormone secretion with progesterone
concentrations typically higher in themorning and the known effect of exercise on increasing
oestrogenandprogesteroneconcentrations(SyropandHammond,1987).
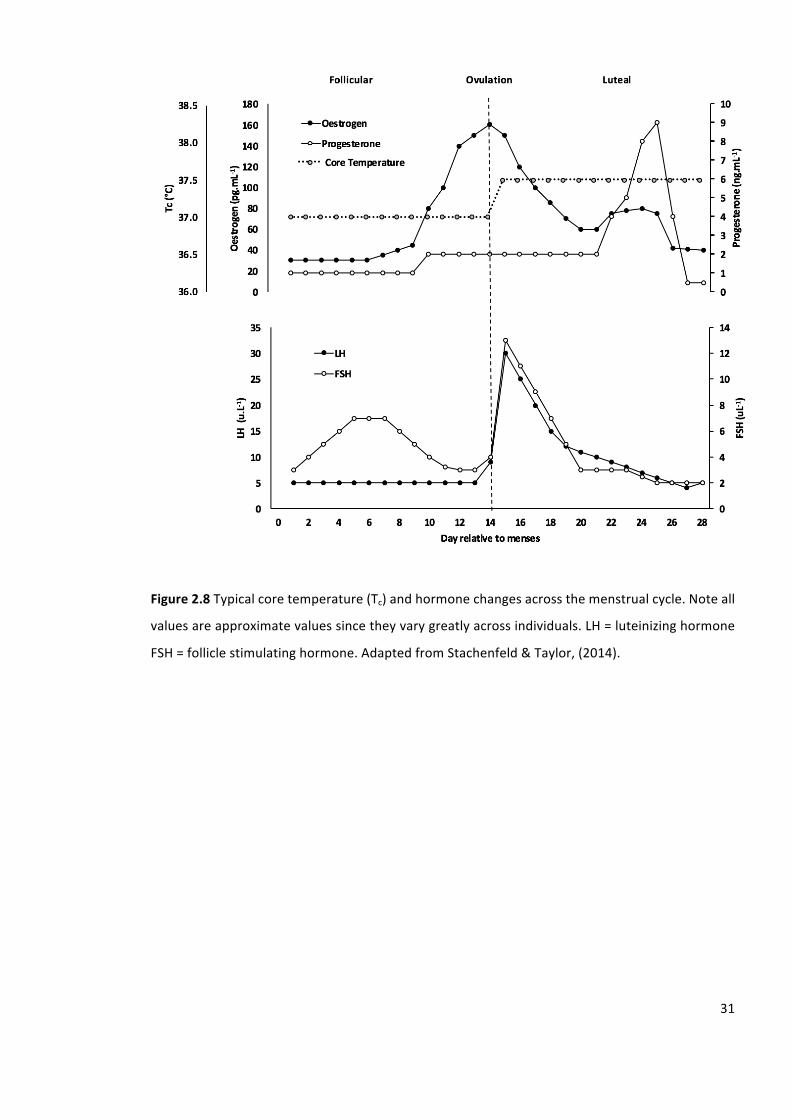
31
Figure2.8Typicalcoretemperature(Tc)andhormonechangesacrossthemenstrualcycle.Noteall
valuesareapproximatevaluessincetheyvarygreatlyacrossindividuals.LH=luteinizinghormone
FSH=folliclestimulatinghormone.AdaptedfromStachenfeld&Taylor,(2014).
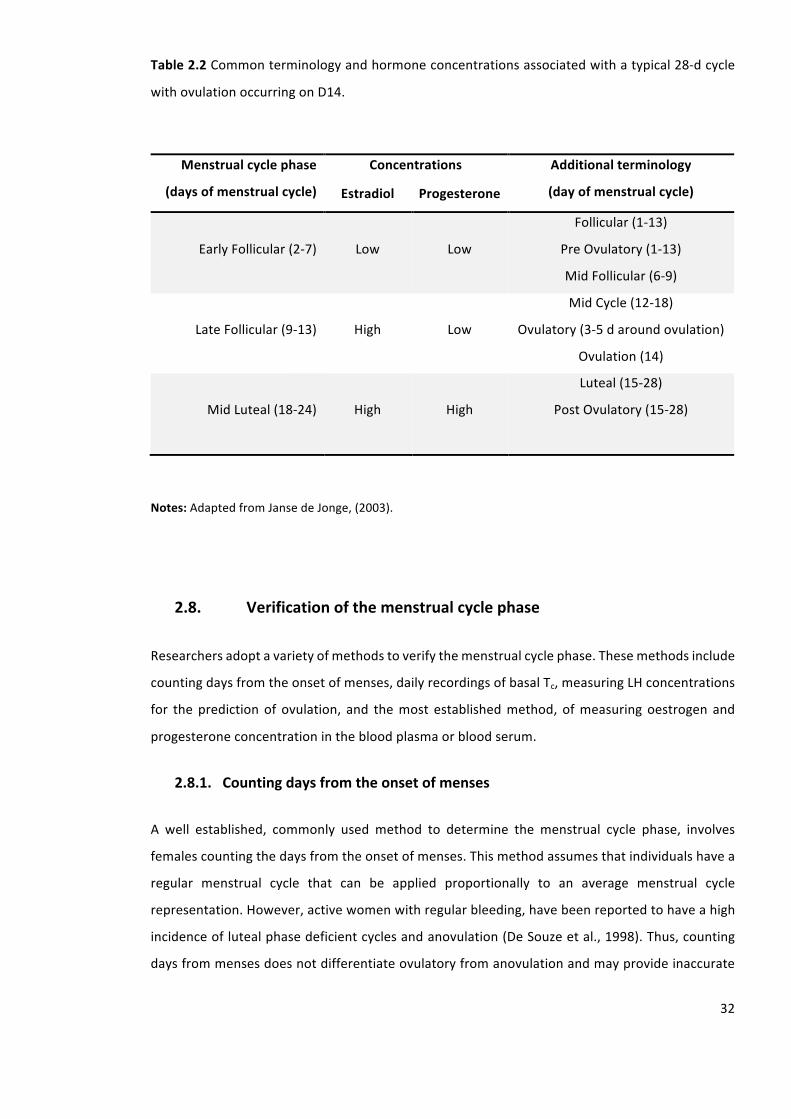
32
Table2.2Commonterminologyandhormoneconcentrationsassociatedwithatypical28-dcycle
withovulationoccurringonD14.
Notes:AdaptedfromJansedeJonge,(2003).
2.8. Verificationofthemenstrualcyclephase
Researchersadoptavarietyofmethodstoverifythemenstrualcyclephase.Thesemethodsinclude
countingdaysfromtheonsetofmenses,dailyrecordingsofbasalTc,measuringLHconcentrations
for the predictionof ovulation, and themost establishedmethod, ofmeasuringoestrogen and
progesteroneconcentrationinthebloodplasmaorbloodserum.
2.8.1. Countingdaysfromtheonsetofmenses
A well established, commonly used method to determine the menstrual cycle phase, involves
femalescountingthedaysfromtheonsetofmenses.Thismethodassumesthatindividualshavea
regular menstrual cycle that can be applied proportionally to an average menstrual cycle
representation.However,activewomenwithregularbleeding,havebeenreportedtohaveahigh
incidenceoflutealphasedeficientcyclesandanovulation(DeSouzeetal.,1998).Thus,counting
daysfrommensesdoesnotdifferentiateovulatoryfromanovulationandmayprovideinaccurate
Menstrualcyclephase
(daysofmenstrualcycle)
Concentrations Additionalterminology
(dayofmenstrualcycle)Estradiol Progesterone
EarlyFollicular(2-7)
Low
Low
Follicular(1-13)
PreOvulatory(1-13)
MidFollicular(6-9)
LateFollicular(9-13)
High
Low
MidCycle(12-18)
Ovulatory(3-5daroundovulation)
Ovulation(14)
MidLuteal(18-24)
High
High
Luteal(15-28)
PostOvulatory(15-28)

33
information regarding the menstrual cycle phase. Furthermore, assumptions are made that
individualscorrectlyreporttheirmenstrualactivity.
2.8.2. Basalbodytemperature
Anothermethod formenstrual cycle determination is daily recording of basal Tc. This method
assumesaregularpatternofactivitythroughoutthecycle.Typically,ovulatorywomenhavea0.3°C
increase in basal Tc subsequent to ovulation, which is sustained throughout the luteal phase
(HorvathandDrinkwater,1982).However, therelationshipbetweenbasalTcandovulationmay
varyconsiderably,sincesomefemalesdonotexperienceanincreaseinbasalTcduringtheluteal
phase(Bauman,1981).Although,anincreaseinbasalTcoftenreflectsanincreaseinprogesterone,
aweakcorrelation(r≤0.302)haspreviouslybeenfound(Formanetal.,1987).Thus,thereliability
ofrecordingdailybasalTctodeterminemenstrualcyclephasesanditsreflectionofprogesterone
levelsshouldbetreatedwithcaution.
2.8.3. Urinaryluteinisinghormoneconcentration
Urinary LH concentration can be determined using ovulation predictor kits, consisting of
colorimetricenzymeimmunoassaysofurinaryLH(JansedeJonge,2003).WhentheurinaryLHsurge
hasbeenshown,itcanbeassumedthatovulationwilltakeplacewithinthenext14-26hr(Miller
andSoules,1996).
2.8.4. Measurementofoestrogenandprogesterone
OnD1ofthemenstrualcyclegonadotropin-releasinghormone issecretedbythehypothalamus
which controls the stimulation and inhibition of FSH and LH (Marsh and Jenkins, 2002). These
hormones regulate the releaseofoestrogenandprogesteroneduringanaveragecycleof28d.
Consequently, the measurement of oestrogen and progesterone can be used to verify the
menstrual cycle phase. The hormone concentrations can be measured in blood serum, blood
plasmaandsaliva.Inaddition,theirmetabolitescanbemeasuredinurine.Measuringoestrogen
and progesterone concentration in blood serum or blood plasma confirms themenstrual cycle
phase,basedonanincreaseinprogesteronefromthefolliculartothelutealphase,indicatingthat
ovulationhasoccurred(JansedeJonge,2003).

34
2.9. Thermoregulatorydifferencesoverthemenstrualcycle
Inwomenwhohavenosignofanymenstrualcycleirregularity,themenstrualcyclecomprisesa
follicularphaseandalutealphaseduringwhichoestrogenandprogesteroneconcentrationsalter.
ChangesintheconcentrationsofoestrogenandprogesteronemodifythebasalTcandthethreshold
forsweatingandcutaneousvasodilationbyalteringtheactivityofcentralregulatorymechanisms
(CharkoudianandJohnson,2000).Specifically,theelevationofprogesteroneconcentrationduring
the lutealphase iscorrelatedwithan increase inthebasalTc (0.3-0.5°C)andthethresholdfor
sweatingandcutaneousvasodilation(CarpenterandNunneley,1988;CharkoudianandJohnson,
2000; Inoue et al., 2005). Whereas, the elevation of oestrogen concentration during the late
follicularphasecausesadecreaseinthesameparameters(StephensonandKolka,1999).Themost
widelyacceptedexplanationforanelevatedTcinthelutealphaseisthatthethermoregulatoryset
point is increased (Forman et al., 1987). Thus, this would imply that the threshold for all
thermoregulatory effector responses are shifted in a similar direction and the increased luteal
phasebasalTcwouldremainelevatedthroughoutexerciseand/orheatstress.
Themechanismsthatcontrolsthesealterationsinthethermoregulatorysetpointassociatedwith
the menstrual cycle are not well understood in females. In animal research, progesterone
administrationhasbeenshowntodecreasetheactivityofthewarmsensitiveneuronsandincrease
theactivityofcoldsensitiveneuronsinthepreopticarea(Nakayamaetal.,1975).Thesefindings
indicateacentraleffectofprogesteroneinthepreopticarea,resultinginanincreasedsetpoint
temperature. In contrast, a decrease in Tc has been associated with oestrogen administration
(Tankersley et al., 1992). Animal research indicates that oestrogen increases the activity of the
warmsensitiveneuronsinthepreopticarea(SilvaandBoulant,1986)causingadecreaseinbody
temperature(SilvaandBoulant,1986).Asaresult,researchershavespeculatedthattheincreased
thermoregulatorysetpointduringthelutealphase,isrelatedtotheratiobetweenoestrogenand
progesterone(Formanetal.,1987).
Alterationsinthethermoregulatorysetpointandeffectorresponsesoverthemenstrualcyclehave
beenexaminedinfemalesusingexerciseand/orheatstress.Someresearchershavereportedno
differences in basal Tc between the follicular and luteal phase (Avellini et al., 1980;Wells and
Horvath,1974).Incontrast,someresearchershavereporteddifferencesinbasalTc,howeverthese
differenceswere not apparent following exercise and/ or heat stress (Horvath andDrinkwater,
1982;Senay,1973).Themajorityofthesestudiesfailedtoverifythemenstrualcyclephasewith
measurements of oestrogen and progesterone concentrations accurately. However, when
hormonalverificationof themenstrual cyclehadbeenconducted, the reduction inbasalTchas

35
been reported to remain elevated throughout exercise and/ or heat stress (Carpenter and
Nunneley,1988;HessemerandBruck,1985;Stachenfeldetal.,2000).Severalofthesestudiesalso
reportedanincreasedTcthresholdforthermoregulatoryeffectorresponsesduringthemid-luteal
phase(HessemerandBruck,1985;Stachenfeldetal.,2000).Accumulatively,thesefindingssupport
theincreasedthermoregulatorysetpointtheory.
2.9.1. Sweatingandskinbloodflow
Itiswellestablished,thatthethresholdforonsetofsweatingisincreasedduringthelutealphase
relative to the follicular phase (Hessemer and Bruck, 1985; Stephenson and Kolka, 1985). In
addition,anincreaseinwholebodySWRhasbeenobservedduringthelutealphasecomparedwith
the follicular phase (Grucza et al., 1993; Hessemer and Bruck, 1985). However, during intense
exercise in hot environmental conditions (35°C and 50°C), where there is a greater strain on
evaporativeheatloss,nodifferencesinSWRhavebeenobservedbetweenthefollicularandluteal
phases (Kolka and Stephenson, 1989; Stephenson and Kolka, 1985). The potentialmechanisms
responsiblefortheincreasedthresholdfortheonsetofsweatingduringthelutealphasecouldbe
associatedwiththeactionsofeitherprogesteroneoroestrogen,asbothareelevatedduringthis
time.Thestimulustoincreasesweatproductionoccurscentrally,adirectresultofprogesterone
actingonthepreoptic/anteriorhypothalamus(StephensonandKolka,1993).
The fluctuations inhormonal concentrationofoestrogenandprogesteroneassociatedwith the
menstrualcyclehavebeenshowntoalteractivecutaneousvasodilatorycontrols(Bruntetal.,2012;
Charkoudian and Johnson, 2000; Charkoudian, 2010). When oestrogen and progesterone
concentrations are elevated during the luteal phase, there is a concurrent increase in the Tc
thresholdforcutaneousvasodilation,relativetothefollicularphase(Charkoudian,2001).However,
beyond the threshold for cutaneous vasodilation, further increases do not incur additional
alterations (Charkoudian, 2001). Horvath and Drinkwater (1982) reported a 25% reduction in
forearmbloodflowatrestduringexposuretoambientconditionsof28°Cduringthefollicularphase
comparedwiththelutealphase.However,whenexercisewasperformedinbothawarm(35°C)
and hot (48°C) environment therewere no differences in forearmblood flowbetweenphases.
Similarthresholdshiftsoccurwithadministrationoforalcontraceptivesthatcontainoestrogenand
progesterone (Charkoudian and Johnson, 2000; Grucza et al., 1993; Stachenfeld et al., 2000;
StephensonandKolka,1993).Thisobservedshiftinthresholdsistheneteffectofareductionin
oestrogenandanelevationofprogesterone,thelatterhavingdominance.Theimpactofoestrogen
andprogesteroneonsweatingandskinbloodflowresponsesarefairlyconclusive.Consequently,

36
consideration of themenstrual cycle phasemust bemadewhen assessing these variables in a
femalepopulation.
2.9.2. Cardiovascular
Onlyafewalterationsincardiovascularresponseshavebeenreportedasaresultofthechangesin
hormoneconcentrationsduringthemenstrualcycle.AnelevationinrestingandexercisingHRhas
previouslybeenreportedduringthelutealphaseofthemenstrualcyclecomparedtothefollicular
phase.Kolka&Stephenson, (1997) reportedan increase inHRwhenexercising at 80%ofpeak
aerobicpowerin35°Cand22%RH,inthemidlutealphase(161±9beats.min-1
)comparedtothe
earlyfollicularphase(150±11beats.min-1
).ThisevidenceisfurthersupportedbytheworkofInoue
et al., (2005) who reported higher resting and exercising HR during the luteal phase (80 ± 4
beats.min-1
, 100 ± 5 beats.min-1
) compared to the follicular phase (70 ± 3 beats.min-1
, 93 ± 5
beats.min-1
)ofthemenstrualcycle.However,Tenaglia,McLellan,&Klentrou,(1999)reportedno
phaserelateddifferencesinHRresponsesduringa300-minexposureinvolving15-minintervalsof
walkingandrestin40°Cand30%RH.Evidenceremainsinconclusiveregardingthemenstrualcycle
effectonHRresponsesatrestandduringexerciseinhotconditions.
It remains unclear whether PV is affected by menstrual cycle related fluctuations in hormone
concentrations.Chapmanetal.(1997)reportednodifferencesinPVbetweenthefollicular(45±3
mL.kg-1
)and the lutealphase (47±3mL.kg-1
)of themenstrual cycle. In contrast,Calzoneetal.
(2001)reportedalowerPVinthelutealphase(2437±113mL)comparedtothefollicularphase
(2529±183mL). Furthermore, thedifferencesobservedmaybedue to theprocedureused to
determinePV;Chapmanetal., (1997)usedamodifiedcarbonmonoxide rebreathing technique
whichdirectlymeasuresbloodvolumeandCalzoneetal.,(2001)usedadyedilutiontechniquewith
Evans bluewhich directlymeasures PV. A lower PV in the luteal phase is further supportedby
Stachenfeldetal.(1999)whoreporteda8.4±2.5%higherPVinthefollicularphasecomparedto
thelutealphase.Thealterationsarepotentiallyduetotheincreasedlevelsofoestrogenassociated
withthe lutealphaseof themenstrualcycle.An increase inoestrogen isknowntoenhancethe
actionsofaldosteroneintheabsorptionofsodiumintherenaltubules(Stachenfeldetal.,1998)
causingareductioninPV.
Generally, no differences are observed in haemoglobin concentrations over the course of the
menstrual cycle (Lebrun et al., 1995;Maughan et al., 1996; Tenaglia et al., 1999). In contrast,
Jurkowskietal.(1981)reportedanincreaseinrestinghaemoglobinduringthelutealphaseofthe
menstrualcycle,althoughthisdidnotresult inan increase inarterialO2content,orO2delivery

37
during exercise. Furthermore, haematocrit levels generally remain unchanged both at rest and
duringexerciseoverthecourseofthemenstrualcycle(Chungetal.,1999;Tenagliaetal.,1999).
However, Stachenfeld et al. (2000) reported an increase inhaematocrit levels during the luteal
phase(39.3±0.5%)comparedtothefollicularphase(38.3±0.7%).Evidenceremainsinconclusive
as to whether consideration of the menstrual cycle phase must be made when investigating
females’cardiovascularandhaematologicalresponsesacrossthemenstrualcycle.
2.9.3. Exerciseperformanceintheheat
Although,femaleathletestrainandcompleteduringallphasesoftheirmenstrualcycle,itisofhigh
importancetounderstandtheeffectsthatthehormonalfluctuationsassociatedwiththemenstrual
cyclemayhaveonexerciseperformance.TheincreasesinTctypicallyobservedduringtheluteal
phaseofthemenstrualcyclemayresultinanincreasedthermoregulatoryandcardiovascularstrain
causing,adecreaseinprolongedexerciseperformance.
Tenaglia et al. (1999) examined the effects of the possible thermoregulatory changes over the
menstrual cycle on exercise time to exhaustion in the heat. A longer time to exhaustion was
reported in the early follicular phase during light intensity intermittent exercise. Tenaglia et al.
(1999) also reported a higher Tc during rest in the luteal phase, however, no differenceswere
observedintherateofTcriseortheTcattheendofthetimetoexhaustion.Thesefindingsassume
thatthereisacriticalTclimitforexerciseperformance,thus,duringthelutealphasewhenresting
Tciselevated,exercisetimetoexhaustionwillbelimited.However,thereremainslimitedliterature
conducted within this area. Future research is warranted to determine whether the
thermoregulatory differences across the course of themenstrual have on effect on endurance
performance.
VoO2max isa strongpredictorofenduranceperformance.Data indicates that there isnophase
relateddifferencesinVoO2orVoO2maxinhotconditions(HorvathandDrinkwater,1982;Stachenfeld
etal.,2000;Tenagliaetal.,1999).However,Williams&Krahenbuhl(1997)reportedhigherresting
VoO2 inthelutealphase(4.8±0.1L.min-1
)comparedtothefollicularphase(3.9±0.2L.min-1
). In
contrast,Lebrunetal.(1995)reportedalowerVoO2maxinthelutealphase(53±1mL.kg-1
.min-1
)
comparedtothefollicularphase(54±1mL.kg-1
.min-1
).WilliamsandKrahenbuhl(1997)reported
lowerventilationrateduringthefollicularphase(10.3±0.8L.min-1
)comparedtothelutealphase
(12.4±0.7L.min-1
)whilstatrest.Furthermore,ahigherventilationratewhilstexercisingat55%
VoO2maxand80%VoO2maxduringthefollicular(42.2±1.4L.min-1
;63.6±2.0L.min-1
)comparedto
thelutealphase(46.2±0.9L.min-1
;68.8±3.0L.min-1
)ofthemenstrualcyclewasalsoreported.An

38
increaseinprogesteroneconcentrationsduringthelutealphaseofthemenstrualcycleincreases
thephrenicnerveactivity.Asthephrenicnerveinnervatesthediaphragm,elevatedprogesterone
duringthelutealphaseisoftenassociatedwithvaryingdegreesofhyperventilation(Denisetal.,
1976).Toconclude,althoughprogesteronehasbeenreportedtohaveamarkedeffectonphrenic
nerve activity, subsequent influences of the menstrual cycle on respiratory function are not
conclusive.Thus,considerationofthemenstrualcyclephasesshouldbeconsideredwhenassessing
thermoregulatoryresponsesinfemales,asalterationsinVoO2willultimatelyaffectthemetabolic
heatproduction.
2.10. Individualdifferenceinheattolerance
Individuals vary in their ability to sustain heat strain, with some demonstrating a decreased
capability todissipateheatandgreaterbodyheatcontentunder thesameexerciseheatstress.
Theseindividualshavebeendescribedasheatintolerantandareoftencharacterisedbyanearlier
andgreater rise inTc, agreater storageofmetabolicheat,haveahigherphysiological strain to
moderateintensityexerciseintheheatandreducedsweatingsensitivity(Epstein,Shapiro,&Shai,
1983;Moran,Heled,Still,Laor,&Shapiro,2004).Aheatintolerancestatecanbeeithertemporary
or permanent. It may stem from transient predisposing factors such as an acute injury to the
thermoregulatorycentre,insufficientHA,dehydration,orinfectiousdisease.Inaddition,alasting
thermoregulatorydysfunctionmay stem fromconditions such as cardiacdisease, diabetes, and
stokeimpairmenttosweatglands(Epstein,1990),orduetodifferencesingeneexpression(Moran
etal.,2006).Congenitalfactorssuchasectodermaldysplasiamayalsocompromiseheattolerance1
insomeindividuals.Forthepastseveraldecades,researchershaveundertakeninvestigationsto
develop a better understanding of the various factors that contribute to the inter-individual
variationsobserved in thehumanheatstress response.Historically,anthropometryandaerobic
fitnesshavebeenconsideredtobetheprimarytransientdeterminantsthatalterthermoregulatory
responses.
2.10.1. Aerobicfitness
Traditionally, itwasbelievedthatTcchangesduringexercisewerecloselyrelatedtotherelative
exerciseintensity(SaltinandHermansen,1966).Tobalancetheincreaseinmetabolicheat,higher
1
Thetermheattolerancewillbeusedthroughoutthisthesistodescribethephenotypicresponses
toheatstress,whereasthetermthermotolerancewillbeusedtodescribethecellularresponsesto
heatstress.

39
aerobically trained individuals would typically increase their heat loss, resulting in similar Tc
responsetountrainedindividuals.Inanattempttoisolatetheinfluenceoftheconfoundingeffects
of aerobic capacity between groups, many studies over the past few decades comparing
thermoregulatory responses between independent groups have consequently administered
exerciseusingpercentageVoO2max.
Mora-Rodriguezetal.(2010)reportedthatwhenexerciseisadministeredrelativetoVoO2max(40%
and60%VoO2max),aerobicallytrainedindividuals(66±6mL.kg-1
.min-1
)experiencesimilarchanges
in Tc to untrained individuals (44 ± 3 mL.kg-1
.min-1
). Enhanced heat dissipation in the trained
participantspermittedsimilarTcchangescomparedtotheuntrainedparticipants,despiteahigher
netmetabolicheatproduction.However,whenexercisingatahigherintensity(i.e.80%VoO2max)
Tcwashigherintrainedindividualscomparedwithuntrainedindividuals.Thus,thesuperiorheat
dissipation of the trained participants did not compensate for the higher net metabolic heat
productionwhenexercisingatahigherpercentageofVoO2max.
Itiswidelyacknowledged,thatthehumanthermoregulatorysystemworkstoattainasteadystate
Tc,suchthattherateofmetabolicheatproductionisbalancedbyanequallyelevatedrateofheat
dissipation (Jay and Kenny, 2007). The increase in heat production during exercise is mostly
determined by the change in absolute rate of O2 consumption, whereas the increase in heat
dissipationoccursmainlyduetoanelevatedrateofsweatevaporationfromtheskin(Gagnonet
al.,2013).Thus,whenexercisingatasimilarpercentageofVoO2max,endurancetrainedparticipants
exerciseatahigherabsoluteworkratethanuntrainedindividualsandgeneratemoremetabolic
heat (Gagnonet al., 2008). In turn, cutaneousblood flow (Fritzsche andCoyle, 2000) and SWR
(Kuwaharaetal.,2005)arehigherintheaerobicallytrainedparticipantscomparedtotheuntrained
participantstocounterbalancetheincreasedheatproduction.
Cramer&Jay(2014)reportedthattherearenodifferencesinwholebodySWRbetweenaerobically
fitindividuals(60mL.kg-1
.min-1
)andrelativelyunfitindividuals(40mL.kg-1
.min-1
)duringexerciseat
a fixed heat production of 275 W.m-2
, despite large differences in percentage VoO2max.
Furthermore, the only fitness-related differences in local SWRwere observed on the forehead,
wheresweatingwaslowerinindividualswithagreateraerobiccapacity.Thisfindingwaspotentially
duetogreatersympatho-adrenalactivityassociatedwithagreatercardiovascular,perceptualand
ventilatorystraininthelessfitindividualsatafixedheatproduction,potentiallyelicitingagreater
sweatproductioninglaborousskinareas(Crameretal.,2012).Thisevidencesuggeststhatprevious
observations that have used relative VoO2max to assess the influence of aerobic fitness on
thermoregulatoryresponses,maybeconfoundedbymethodologicalissues.Thus,whenisolating

40
theindependentinfluenceofphysiologicalfactorsonthethermoregulatoryresponsestoexercise
using a between subject design, exercise intensity should be administered using fixed heat
production.
2.10.2. Anthropometry
AhighbodyfatpercentagemayalterTcchangesduetoaloweraveragespecificheatcapacityof
adiposetissue. Jayetal. (2011)comparedthethermoregulatory responsesof individualswitha
high(60±5mL.kg-1
.min-1
)andlow(40±3mL.kg-1
.min-1
)aerobiccapacity,whilstexercisingatafixed
heatproduction.Theparticipantsinthelowaerobiccapacitygrouphadatwofoldhigherbodyfat
percentage(22.2±7.0%)comparedwiththehighaerobiccapacitygroup(11.9±5.9%).Thelower
specificheatcapacityofadiposetissue(2.97kJ.-1
kg-1
.°C)wouldhaveyieldedaslightlyloweraverage
specificheatofthebody inthe lowaerobiccapacitygroup(3.53kJ.-1
kg-1
.°C)relativetothehigh
aerobicgroup(3.45kJ.-1
kg-1
.°C)(GeddesandBaker,1967).Therefore,asimilarbodyheatstorage
wouldhavetheoreticallyelevatedTcinthelowgroupbyagreatermagnitude.Nevertheless,the
changesinTcwerealmostidenticalbetweenthetwogroups(high:0.87±0.15°C,low:0.87±0.18°C)
when exercising at a fixed heat production. Therefore, it appears that any thermoregulatory
implicationofthedifferenceinadipositybetweenthetwogroupswasinsignificant,likelydueto
any insulation created from the fat layers beingbypassedbyblood flow following theonsetof
exercise (Havenith and Middendorp, 1990). Another implication of a higher adiposity is a
concomitant difference in body volumeand thereforeBSA. The lower density of fat (0.9 kg.L-1
)
relativetomuscle(1.1kg.L-1
)willallowforagreaterevaporativerateinthelowaerobiccapacity
groupduetothehigherbodyfatpercentage.
Cramer&Jay(2014)comparedthethermoregulatoryresponsesofparticipantswithasmall(68±6
kg)andlarge(92±7kg)BMwhilstexercisingatafixedrateofmetabolicheatproduction(500W
and600W).Resultsdemonstratethat largedifferencesinBMsystematicallyalteredTcchanges,
with smaller participants achieving a greater Tc rise. However, when exercise intensity was
prescribedusingafixedheatproductionrelativetoBM(6.5W.kg-1
and9.0W.kg-1
)thesystematic
differencesinTcwereeliminateddespitedifferencesinBM,absoluteheatproduction,andrelative
exercise intensity.Thus, thehigherTc in thesmallergroupat thesame fixedheatproduction is
directlyexplainedbytheinfluenceofBMperseandnotbyanydifferencesinheatdissipationor
bodycomposition.Whenexercisingataheatproductionofboth500Wand600W,nodifferences
inTskandtherefore,dryheatlosswereobservedbetweenparticipantswithasmallandlargeBM.
Thisresultedinasimilarabsoluteevaporativerequirementandtherefore,thesamewholebody
SWRandpresumablyevaporation(CramerandJay,2014).Assuch,itislikelythatthedifferences

41
inbodyfatpercentagebetweenthetwogroupscontributedtotheobserveddifferences inTre.
WhileitmaybepossiblethatmuchlargerdifferencesinbodyfatpercentagealterchangesinTc,
theindependentinfluenceofhighversuslowadiposetissuewhilstcontrollingforheatproduction
and BM has not yet been evaluated and merits further investigation. Thus, when assessing
thermoregulatory responses to exercise, using a between subject design with participants of
varyingbodysizes,exerciseintensityshouldbeadministeredusingfixedheatproduction.
2.11. Methodstoassessheattolerance
ExperimentalprocedureshavebeenemployedtoraiseTcunderrestingandexerciseconditionsto
challengethethermoregulatoryresponsesasamethodofassessingthermoregulatorymechanisms
(Inoueetal.,2005;Kenney&Hodgson,1987;Montain,Sawka,Cadarette,Quigley,&McKay,1994).
AHTTcanprovideinformationtoevaluatetheabilitytoavoidtheriskofexertionalheatstrokeand
aid in decisions about readiness to return to training, competition and/or active duty. The
importance of heat tolerance screening was highlighted by Schutte (2010). Following the
implementationofheattolerancescreening,incidencesofheatstrokeandheatillnessinmining
werereducedto33casesovera2yr.periodcomparedwith88fortheprevious3yrs.(Schutte,
2010).
Exercise HTTs commonly adopt protocols of a long duration (>100 min), low intensity (≤ 50%
VoO2max)inenvironmentalconditionsof35-40°Cand40-50%RH,usingavarietyofexercisemodes
including,runningandbenchstepping(Epstein,1990;Moranetal.,2007).Theextensivevariation
intestparadigmsmakesbetweenstudycomparisonsdifficult.Constructvalidityoftheseprotocols
has been assumed since physiological measures were different between groups of individuals,
althoughtheprotocolsemployedwerenotalwayscapableofdifferentiatingbetweenheattolerant
andheatintolerantindividuals(Moranetal.,2004).Shapiroandcolleagues(1979)developedaHTT
whichconsistedofsteppingfor180-minona30cmhighbenchinacontrolledhotenvironment.
This was adapted by the IDF to be performed on a treadmill. The protocol involves 120-min
exposureto40°Cand40%RHwhilewalkingonatreadmillatapaceof5km.hr-1
anda2%elevation
(Moran et al., 2007). Criteria to discern tolerance and intolerance to exercise heat stresswere
developed.Attheendofthe2-hrexposure,heattolerancewasdefinedasapeakTre≤38.0°C,peak
HR≤120beats·min-1
,andSWR≥780g·h-1
.Heatintolerancewasdefinedas,peakTre≥38.5°Cor
peakHR≥145beats·min-1
(Moranetal.,2007)andapeakphysiologicalstrainindex≥6(Moranet
al.,2004).

42
ThereareseverallimitationsassociatedwithIDFHTT,whicharemoreapparentwhenconducting
thetestonanathleticpopulation(Johnsonetal.,2013).Endurancerunnerstypicallyrunathigher
intensitiesforshorterdurationsthanmilitarypersonnel.TheintensityoftheprotocoloftheHTTis
considerablybelowthatatwhichathleteswouldtrainandcompete.Thus,theHTTmisrepresents
the metabolic heat production endurance runners may experience and potentially may
misdiagnosetheirsusceptibilitytoahyperthermicstate,orworseaheat-relatedillness.Protocols
ofthisdurationwouldrequireasignificantamountoftimeoutfromtrainingforathletes,which
mayresultinacoachand/orathletebeingreluctanttohavetheirheattoleranceassessed.Further
work is required to establish an effective short duration, running based HTT improving the
ecologicalvalidityandapplicationtoendurancerunners.
2.12. Exertionalheatillnessesinrunning
The challenges of training and competing in a hot environment are complex and difficult to
comprehendfully,sinceathletesareaffectedinavarietyofwaysduringhighintensityexercisein
hothumidenvironments.Pathologiceventswhichoccur inhotenvironmentsare referredtoas
exertionalheatillnessesandareoftencategorisedasexertionalheatstroke,heatexhaustionand
heatcramps(table2.3).Exertionalheatillnessescommonlyaffectathletesduringhighintensity,or
long duration exercise and the severity of the illness depends largely upon the degree of
hyperthermiaanditsduration.Ourknowledge,regardingexertionalheatstrokeislimitedtocase
studies of athletes who push beyond the normal physiological limits, since laboratory studies
cannotbeconducted,duetotherisksofseverehyperthermiaareethicallyunacceptableforhuman
research. The survival of these athletes depends largely on prompt recognition and the
implementation of effective cooling strategies to limit tissue exposure due to destructive
hyperthermia(HoweandBoden,2007).
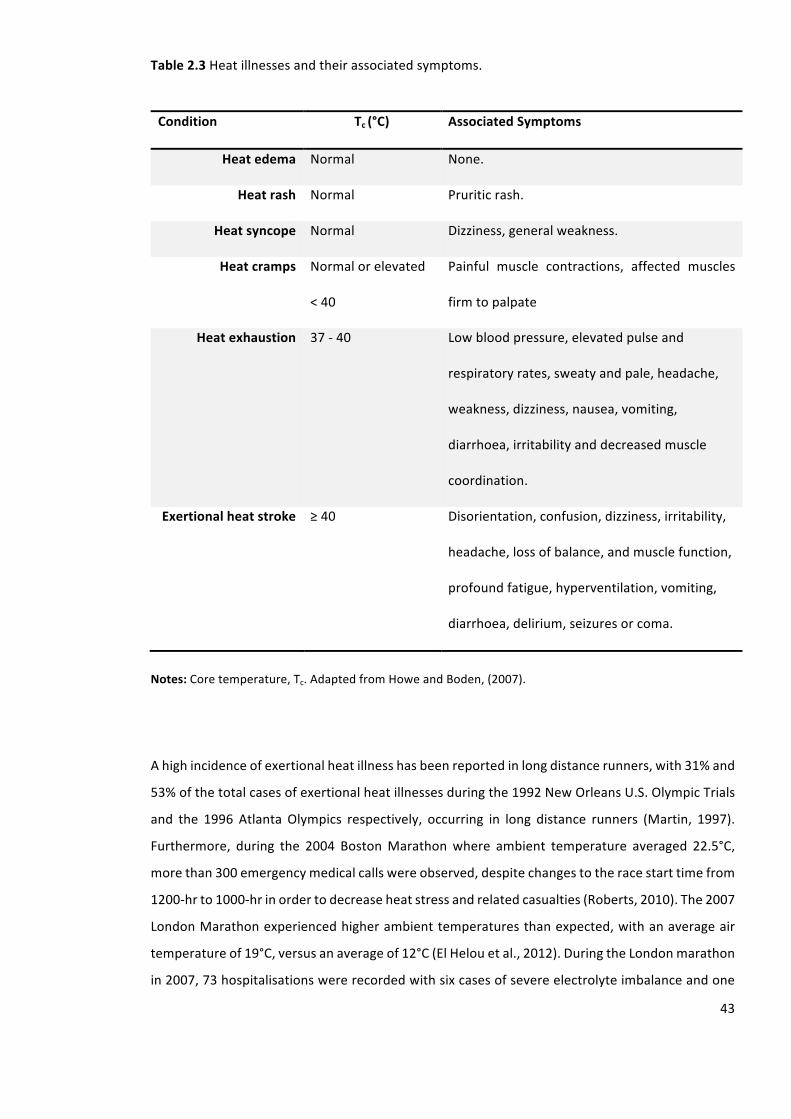
43
Table2.3Heatillnessesandtheirassociatedsymptoms.
Condition Tc(°C) AssociatedSymptoms
Heatedema Normal None.
Heatrash Normal Pruriticrash.
Heatsyncope Normal Dizziness,generalweakness.
Heatcramps Normalorelevated
<40
Painful muscle contractions, affected muscles
firmtopalpate
Heatexhaustion 37-40 Lowbloodpressure,elevatedpulseand
respiratoryrates,sweatyandpale,headache,
weakness,dizziness,nausea,vomiting,
diarrhoea,irritabilityanddecreasedmuscle
coordination.
Exertionalheatstroke ≥40 Disorientation,confusion,dizziness,irritability,
headache,lossofbalance,andmusclefunction,
profoundfatigue,hyperventilation,vomiting,
diarrhoea,delirium,seizuresorcoma.
Notes:Coretemperature,Tc.AdaptedfromHoweandBoden,(2007).
Ahighincidenceofexertionalheatillnesshasbeenreportedinlongdistancerunners,with31%and
53%ofthetotalcasesofexertionalheatillnessesduringthe1992NewOrleansU.S.OlympicTrials
and the 1996 Atlanta Olympics respectively, occurring in long distance runners (Martin, 1997).
Furthermore, during the 2004 BostonMarathonwhere ambient temperature averaged 22.5°C,
morethan300emergencymedicalcallswereobserved,despitechangestotheracestarttimefrom
1200-hrto1000-hrinordertodecreaseheatstressandrelatedcasualties(Roberts,2010).The2007
LondonMarathonexperiencedhigherambienttemperaturesthanexpected,withanaverageair
temperatureof19°C,versusanaverageof12°C(ElHelouetal.,2012).DuringtheLondonmarathon
in2007,73hospitalisationswererecordedwithsixcasesofsevereelectrolyteimbalanceandone

44
death.Incontrast,thenumberofpeopletreatedwas20%lowerattheLondonMarathonayear
laterin2008,whenconditionswerecoolandrainywithanaverageambienttemperatureof10°C
(Roberts,2010).Furthermore,thetotalaveragetimetocompletethemarathonwas17-minslower
in2007comparedtopreviousyears. Inaddition, thepercentageofracewithdrawals inChicago
2007was thehighest (30.74%)amongall60marathonracesanalysedbyElHelouetal., (2012)
when temperature were 13°C higher than the average temperature. Sixty six runners were
admittedtothehospitalwith12reportedtobeintensivecarecaseswithhydrationdisorders,heat
shocksyndromesandonedeath.
Ina12yr. summaryofmarathonmedicalencounters, therewere1.2casesofheatcrampsper
1,000 race entrants; accounting for 6.1% ofmedical encounters (Roberts, 2000). Furthermore,
duringa14kmroadraceinmildconditions(11°C-20°C),14heatexhaustioncaseswerereported
in10,000finishers(RichardsandRichards,1984).Exertionalheatstrokehasbeenobservedmost
commonly during activities that involve continuous high intensity exercise. The twin cities
marathonwhichisrunincoolconditionsaveragesoneexertionalheatstrokepatientper10,000
finishers (Roberts, 2000). Furthermore, during an 11.5 km road race perform in hot humid
conditions there were approximately 10-20 exertional heat stroke cases per 10,000 finishers
(Brodeuretal.,1989).Despitethevolumeofrunnersexperiencinganexertionalheatillnessduring
runningevents,therearecurrentlynoevidencebasedrecommendationsregardingthereturnof
anathletetotrainingandcompetition.TheAmericanCollegeofSportMedicine(HoweandBoden,
2007)recommendthatexertionalheatstrokecasualtiesreturntotrainingandcompetitionwhen
they have re-established heat tolerance. However, there is currently no establishedmethod to
evaluatearunner’sheattolerancetodeterminewhethertheyhavere-establishedheattolerance.
Establishingsuchaprotocolmaypreventathletesfromreturningtotrainingandcompetitiontoo
soonandreducethelikelihoodofexperiencinganotherepisodeofaheatrelatedillness.
2.13. Heatacclimation
Humansareregulators,andassuchstrivetokeepmeanbodytemperaturewithinanarrowrange
(37.0±0.5°C).Accordingly,humansnotonlypossess thecapabilityofenduringawiderangeof
climatic stressors, but also have a considerable capacity to extend this tolerance, through
adaptation.Thephysiologicaladaptationsthatoccurduringexposuretotheheatarereferredtoas
acclimatization responseswhen seen in a naturally occurring environment andHA responses if
observed ina controlledenvironmental setting.HA isachievedbyapersistent challenge to the
thermoregulatorycontrolsystemthatimprovesheatdissipationandthermotolerance,prolonging
endurancetoheatstress.

45
Heatacclimationdevelopsthroughfrequentexposuretohotenvironmentalconditions,whichelicit
responsesthatattenuatethenegativeeffectsofheatstress.HAadaptationarediscussedindetail
insection2.15.Inshort,thebenefitsofHAincludeanimprovedthermoregulatory,cardiovascular,
sweatingandskinbloodflow,cellular,andmetabolicfunctioning,areductioninperceptualstrain
andanimprovedexerciseperformance(Horowitz,2014;Périardetal.,2015;Sawkaetal.,2011a;
Taylor,2014)(table2.4.).ThetimecourseofHAistypicallycategorizedintoSTHA(≤8d)andLTHA
(≥10d)(Garrettetal.,2011;Guyetal.,2015;Périardetal.,2015).Themagnitudeofphysiological
adaptation inducedbyHAdepends largelyonthe intensity,duration,frequency,andnumberof
heatexposures(Sawkaetal.,2011b).
2.14. Heatacclimationprotocols
Heat acclimation is typically achieved through one of three induction pathways; fixed intensity
(Castleetal.,2011;Lorenzoetal.,2010),self-pacedexercise(Armstrongetal.,1986;Sunderlandet
al.,2008)orcontrolledhyperthermia(Gibson,etal.,2015;Pattersonetal.,2004a,2014;Garrettet
al.,2009,2012;Reganetal.,1996).Although,exerciseintheheatisthemosteffectivemethodfor
developingHA,passiveheatexposurehasalsobeenshowntoincursomeadaptation(Beaudinet
al.,2009;Scoonetal.,2007;Stanleyetal.,2015;Takamataetal.,2001).
2.14.1. Fixedintensityheatacclimation
Traditionally,HAaimedtopreparepeopletoperformaspecifictaskwithinaknownsetofclimatic
conditions.Thisinvolvedrepeatedlyapplyingafixedendogenousandexogenousthermalloadto
increaseTcasaresultofanimbalancebetweenheatproductionandheatloss(Castleetal.,2011;
Houmardetal.,1990;Kresfelderetal.,2006;Lorenzoetal.,2010;Sandströmetal.,2008).Thus,
duringeachHA sessionparticipants’workat the sameabsoluteexercise intensity for the same
duration.Thismodeldevelopshabituation, reducesphysiologicalstrain,and increases tolerance
(Foxetal.,1963a).Whilethisapproachappearslogical,therearemajorlimitationsregardingthe
effectivenessandinterpretationofthisprocedure.Itscapacitytoinduceadaptationcontinuallywill
progressivelydeclineasadaptationoccurs,sincethestrengthoftheadaptationstimulusisreduced.
In addition, these protocols require individuals to work at the same absolute intensity; the
physiological strain they experience will therefore, be variable since individuals differ in their
inherent heat tolerance (Taylor, 2014). When undertaking mechanistic research, the resulting
variationsinphysiologicalstrainduringadaptationmakedatainterpretationchallenging.

46
2.14.2. Self-regulatedheatacclimation
Self-regulatedexercise requiresparticipants to followprescribedworkand rest intervals,whilst
self-selecting and modifying the work rate. Although, this method has considerable practical
applicationsinceitcanbeusedtorehearsepacingstrategiesorsimulategameplay,thetechnique
has no standardised endogenous thermal loading, reducing control and possibly constraining
adaptation(TaylorandCotter,2006).
2.14.3. Passiveheatacclimation
Passive HA uses exogenous heat to incur thermoregulatory stress, with a minimal metabolic
contribution.Adaptationisinducedifthecumulativeadaptationimpulseissufficientlylarge,and
thiscanbebroughtabout throughhot-waterbathing (TurkandThomas,1975;Zurawlewetal.,
2015)andbypassiveexposure toanenvironmental chamber, steamroomor sauna (Foxetal.,
1963a; Scoon et al., 2007; Stanley et al., 2015). While passive techniques are beneficial, the
techniquehasbeenreportedtobe lesseffectivethanexercise intheheat (Shapiroetal.,1981;
Wyndham, 1973). The reason is that passive heating may only induce a slight-to-moderate
homeostaticdisturbance,sothecumulativeadaptationimpulseremainssmall.
Foxandcolleaguesdevelopedaveryeffectivepassivetechniquetosolvethislimitation(Foxetal.,
1963a). The method was based on elevating and clamping Tc (controlled hyperthermia).
Participantswereheated in a hot (43°C), humid (100%RH) condition causing a rapid rise in Tc.
Participantsweredressed invapour-barrier suits thatwere continuously ventilatedwithhotair
(35°C)andenteredasecondchamber(38°C),wheretheyrested.ThreeclampedTc(37.3°C,37.9°C,
and38.5°C)andthreeexposuredurations(30,60,and120-min)wereevaluatedfor12-d.Results
suggesteda60-minclampat38.5°Cwas required toestablish fullHA (Foxetal.,1963a).These
specificationshaveledtouseofcontrolledhyperthermiaduringexercise.
2.14.4. Controlledhyperthermiaheatacclimation
ControlledhyperthermiaHAensuresequalthermalstrainisplaceduponparticipantsasitinvolves
elevatingandmaintaininga steadystatebody temperatureabove thesweating thresholdusing
exercise(Garrettetal.,2011).TheuseofthecontrolledhyperthermiaHAhasbecomeincreasingly
popular. Thismethod has consistently been found to reduce thermal strain and increasework
capacityduringbothSTHAandLTHAandmayoffermorecompleteadaptationtofixedintensityHA
(Nealetal.,2015;Gibsonetal.,2015;Garrettetal.,2009,2012;Pattersonetal.,2004a;Reganet
al.,1994).

47
2.15. Heatacclimationinduction
Humanshavearemarkableabilitytoadapttoheatstressandgivenadequatewaterandprotection
fromthesun,ahealthyheatacclimatedindividualcantolerateextendedexposuretovirtuallyany
natural,weather-relatedheatstress.HAresultsinaseriesofadaptationsthatreducetheharmful
effectsofheatstress.Typically,adaptationoccursthroughmorphological,chemical,functional,and
genetic adjustments that decrease physiological strain under stress. HA involves a number of
complexadaptations(table2.4),whichallintegratetoenhanceexerciseperformance.
2.15.1. Sweatingandskinbloodflow
SudomotoradaptationswereamongthefirstdescribedinresponsetoHA.Bytheendofthe1940s,
it was widely accepted that HA increased SWR and decreased sweat sodium and chloride
concentrations(Dilletal.,1933).Theobservedshiftintheonsetthresholdforsweating(Patterson
etal.,2004a)andanalteredSWR(table2.5)andcomposition,areconsideredtheprincipaladaptive
responsestoheatexposure(Buonoetal.,2009a).Theseresponsesareindicativeofbothcentral
andperipheraladaptation.Atthecentrallevel,HAdecreasestheTcatwhichsweatingisinitiated.
Thisadjustmentinonsetthresholdisproposedtocorrespondtoanabsolutechangeinmeanbody
temperature,ratherthantotheattainmentofapredeterminedmeanbodytemperature(Patterson
etal.,2004a).Peripheraladaptations,manifestedbychangesinSWRandsensitivityandoccurat
thelevelofthesweatgland(Buonoetal.,2009a;Inoueetal.,1999).FollowingSTHAandLTHAusing
acontrolledhyperthermiamodel,thereisonaveragea0.23L.hr-1
and0.46L.hr-1
increaseinSWR
observed(table2.5).Thepotentialmechanismsthatexplainthesechangesinclude,anincreased
cholinergicsensitivityoftheeccrinesweatglandandincreaseglandularhypertrophyandefficiency
(Lorenzo&Minson,2010;Buonoetal.,2009;Sato&Sato,1983).Inaddition,eccrinesweatglands
becomeresistanttohidromeiosis,resultingintheabilitytosustainahigherSWRforaprolonged
periodoftime(Foxetal.,1963b).
InadditiontoanenhancedSWR,sweatcompositionchangeswithHA.Electrolytesarereabsorbed
andsweatsodiumconcentrationisreduced,resultinginamoredilutesweat(Chinevereetal.,2008;
Saatetal.,2005).Thepreservationofsodiumappearstostemfromincreasedsodiumconservation
withinthere-absorptiveductofthesweatgland(SatoandDobson,1970;Satoetal.,1971),which
is dependent on aldosterone, a hormone secreted in response to exercise andheat stress that
facilitatesthereabsorptionofsodium.EvaporativeheatdissipationisenhancedforagivenSWR
whenamoredilutesweatissecreted,sinceitismoreeasilyevaporatedbecauseofawideningof
thewatervapourgradientbetweentheskinandambientair(Taylor,2014).
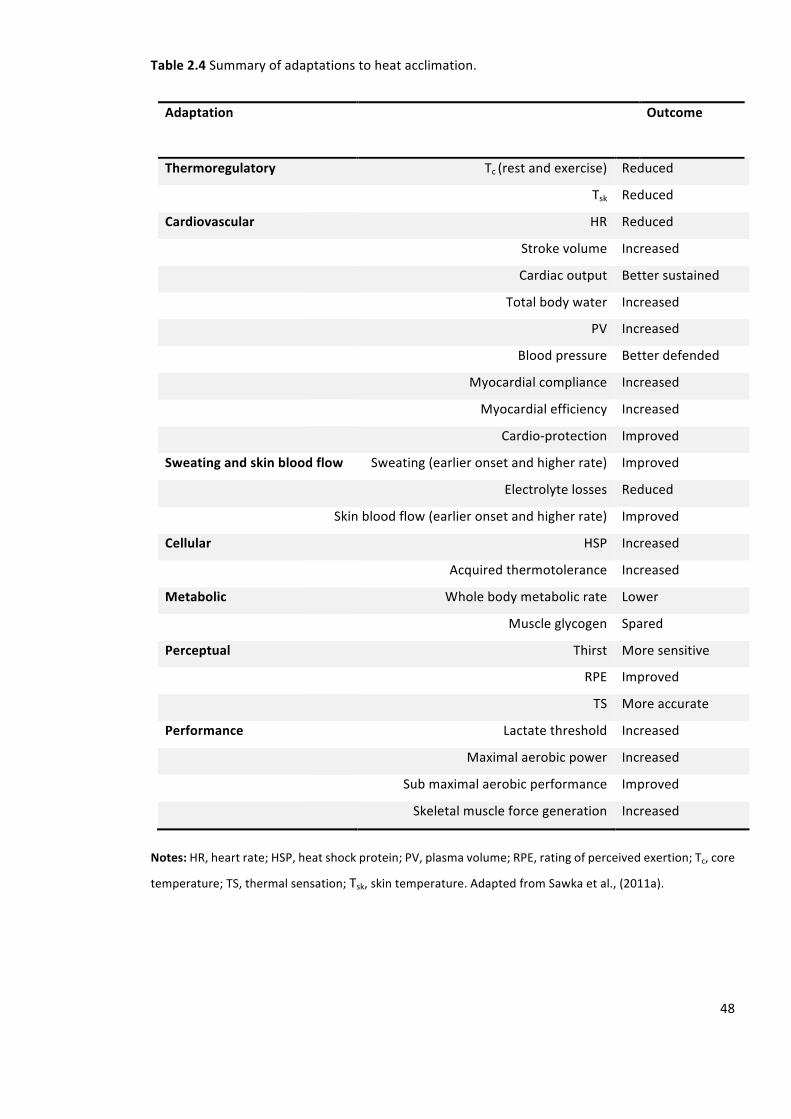
48
Table2.4Summaryofadaptationstoheatacclimation.
AdaptationOutcome
Thermoregulatory Tc(restandexercise) Reduced
Tsk Reduced
Cardiovascular HR Reduced
Strokevolume Increased
Cardiacoutput Bettersustained
Totalbodywater Increased
PV Increased
Bloodpressure Betterdefended
Myocardialcompliance Increased
Myocardialefficiency Increased
Cardio-protection Improved
Sweatingandskinbloodflow Sweating(earlieronsetandhigherrate) Improved
Electrolytelosses Reduced
Skinbloodflow(earlieronsetandhigherrate) Improved
Cellular HSP Increased
Acquiredthermotolerance Increased
Metabolic Wholebodymetabolicrate Lower
Muscleglycogen Spared
Perceptual Thirst Moresensitive
RPE Improved
TS Moreaccurate
Performance Lactatethreshold Increased
Maximalaerobicpower Increased
Submaximalaerobicperformance Improved
Skeletalmuscleforcegeneration Increased
Notes:HR,heartrate;HSP,heatshockprotein;PV,plasmavolume;RPE,ratingofperceivedexertion;Tc,core
temperature;TS,thermalsensation;Tsk,skintemperature.AdaptedfromSawkaetal.,(2011a).

49
AnearlierandgreatersudomotorresponseduringHAimprovesevaporativecooling,assumingthe
climateallowsevaporation,andreducesTskandthus,skinbloodflowrequirements.AlowerTskmay
alsoreducecutaneousvenouscompliance,suchthatthebloodvolumeisredistributedfromthe
peripheraltothecentralcirculation(Rowelletal.,1967).Untilrecently, itwasassumedthatHA
centrally modifies thermoregulatory responses in the skin by reducing the Tc threshold for
vasodilation,withoutalteringtheslopeofthebloodflow–Tcrelationship(Foxetal.,1963a;Roberts
et al., 1977; Yamazaki and Hamasaki, 2003). However, by locally infusing an endothelium-
dependentvasodilator (acetylcholine)viamicrodialysis, Lorenzo&Minson (2010)demonstrated
thatHAimproveslocalcutaneousvascularresponses.Theauthorssuggestedthatthisperipheral
responsemaybeassociatedwithanincreaseinthenumberandsensitivityofmuscarinicreceptors,
adecreaseincholinesteraseactivity,leadingtoanimprovedvascularresponsetoacetylcholine,or
alterations inthepathwayofvasodilationwithinsmoothmusclesortheendothelialcells.Given
thatHAdoesnotaltermaximalskinbloodflow,themodifiedcutaneousvascularresponseappears
to stem from improvements in vascular function (i.e., increased sensitivity of the skin
microvasculature to vasodilate), rather than structural changes that limit maximal vasodilator
capacity(LorenzoandMinson,2010).
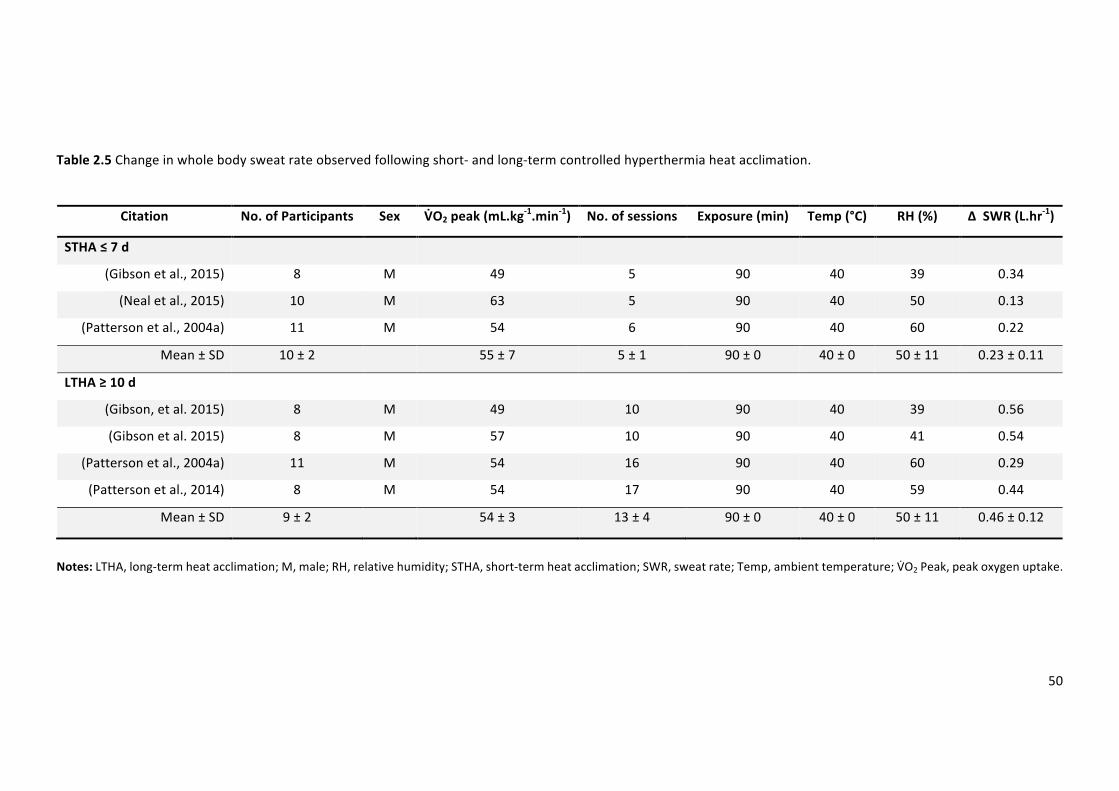
50
Table2.5Changeinwholebodysweatrateobservedfollowingshort-andlong-termcontrolledhyperthermiaheatacclimation.
Citation No.ofParticipants Sex V9O2peak(mL.kg-1.min-1) No.ofsessions Exposure(min) Temp(°C) RH(%) ∆SWR(L.hr-1)
STHA≤7d
(Gibsonetal.,2015) 8 M 49 5 90 40 39 0.34
(Nealetal.,2015) 10 M 63 5 90 40 50 0.13
(Pattersonetal.,2004a) 11 M 54 6 90 40 60 0.22
Mean±SD 10±2 55±7 5±1 90±0 40±0 50±11 0.23±0.11
LTHA≥10d
(Gibson,etal.2015) 8 M 49 10 90 40 39 0.56
(Gibsonetal.2015) 8 M 57 10 90 40 41 0.54
(Pattersonetal.,2004a) 11 M 54 16 90 40 60 0.29
(Pattersonetal.,2014) 8 M 54 17 90 40 59 0.44
Mean±SD 9±2 54±3 13±4 90±0 40±0 50±11 0.46±0.12
Notes:LTHA,long-termheatacclimation;M,male;RH,relativehumidity;STHA,short-termheatacclimation;SWR,sweatrate;Temp,ambienttemperature;VVO2Peak,peakoxygenuptake.

51
2.15.2. Bloodvolumeandfluidbalance
Heatacclimationhasbeenreportedtoincreasetotalbodywaterbyapproximately2–3L,(Basset
al.,1955;Pattersonetal.,2014,2004b).Thisincreaseiswellwithinthemeasurementresolution
(4.9%) for total body water (Maw et al., 1996) and thus, appears to be a real physiological
phenomenon.Thechangesinintracellularfluidandextracellularfluidthatcontributetowardsthe
increaseintotalbodywaterarevariable;sinceextracellularfluidhasbeenreportedtoaccountfor
greater,equal,andsmallerthanitspercentageincreaseintotalbodywaterafterHA(Sawkaand
Coyle,1999).Thereisrelativelyhighvariabilityassociatedwiththemeasurementofextracellular
fluid,andtherefore,trendsforsmallchangesaredifficulttointerpret.Thechangestointracellular
fluidaretypicallycalculatedasthedifferencebetweentotalbodywaterandextracellularfluid,and
thus resultsareunclear since themeasurementvariability inherent inboth these techniques, is
confounded in thecalculationof intracellular fluid.The increase in secretionof fluidconserving
hormones aldosterone and arginine vasopressin, and/or renal sensitivity to a given plasma
concentrationinpartexplaintheincreaseintotalbodywater(Périardetal.,2015).Furthermore,
theconservationofsodiumassistsinthemaintenanceofosmolalityintheextracellularfluidwhich
maintainsorincreasesextracellularfluidvolumeduringheatadaptation(Noseetal.,1988).Asa
result of an increase in total body water and extracellular fluid following HA, youmay expect
expansionofPV.
Following3-4dofheatexposureorasaresultofseasonalchanges,PVexpansionispresent(Sawka
andCoyle,1999).A5%expansioninrestingPVoccursinthehottestmonthsoftheyearanda3%
contraction in the coldestmonths (Sawka and Coyle, 1999). However, individuals have varying
responses,withsomeindividualsnotexperiencinganyexpansioninPV.TheobservedPVexpansion
following HA has previously been described as a transient phenomenon (Shapiro et al., 1981),
howeverthismayhavebeenanexperimentalartefactrelatedtothetraditionalconstantworkrate
modelofHA.Recentfindings,suggestthatbyusingthecontrolledhyperthermiatechnique,which
maintainsa constantadaptationstimulusbyclampingTc throughoutHA,PV iselevatedby15%
following10consecutiveexposures(Gibsonetal.2015)andremainssimilarlyexpanded(∼14%)after8-22dofheatexposure(Pattersonetal.,2004a,2014).Followingcontrolledhyperthermia
HA,PVexpansionisapproximately7%and14%forSTHAandLTHA,respectively(table2.6).The
magnitudeof increase inPV is typicallydependentupon theHAday, thehydration statewhen
measured,Tskandwhethertheindividualisatrestorperformingexercise(Harrison,1985;Kenefick
et al., 2014; Sawka et al., 1983). The magnitude of PV expansion may also depend on the
participants’ fitness level, since endurance training will typically result in an expanded PV. PV

52
expansion offers a thermoregulatory advantage, since it increases vascular filling to support
cardiovascularstability(Sawkaetal.,2011b).
Areductionintotalbodywaterasaresultofdehydrationwilladverselyaffectthermoregulation
andincreasecardiovascularstrain(SawkaandCoyle,1999).Thus,dehydrationwillcounteractthe
adaptationstothermoregulationinitiatedfollowingHAandaerobictraining(Sawkaetal.,1983);
whichthemoreseverethelevelofdehydration,thegreaterthestrainandthegreaterelevationin
Tc.Dehydrationalsoimpairsdryandevaporativeheatlossviathedevelopmentofplasmahyper-
osmolalityandhypovolemia(SawkaandCoyle,1999).However,Takamataetal. (2001)provides
evidencetosuggestthatHAmightattenuatetheadverseeffectsofhyper-osmolalityonimpairing
sweatingandskinbloodflowresponses.Assuch,totalbodywaterexpansionstemmingfromHA
mayconferaprotectivebenefitagainstdehydration.Althoughitisapparentthatdehydrationhas
clear detrimental effects on exercise performance, moderate permissive dehydration during
exerciseHAmayacceleratetheHAprocess,(TaylorandCotter,2006)byincreasingfluidelectrolyte
retention,PVexpansion,andcardiovascularresponsestoheatstress(Garrettetal.,2014,2011;
Nealetal.,2015).
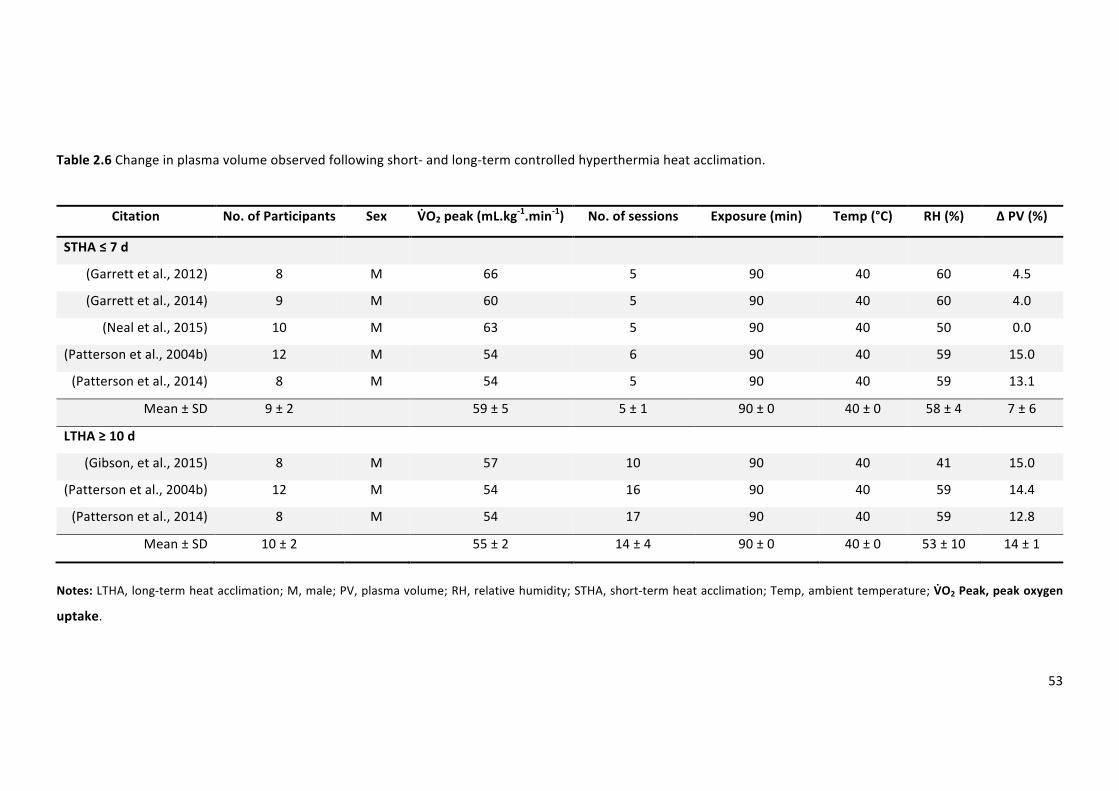
53
Table2.6Changeinplasmavolumeobservedfollowingshort-andlong-termcontrolledhyperthermiaheatacclimation.
Citation No.ofParticipants Sex V9O2peak(mL.kg-1.min-1) No.ofsessions Exposure(min) Temp(°C) RH(%) ∆PV(%)
STHA≤7d
(Garrettetal.,2012) 8 M 66 5 90 40 60 4.5
(Garrettetal.,2014) 9 M 60 5 90 40 60 4.0
(Nealetal.,2015) 10 M 63 5 90 40 50 0.0
(Pattersonetal.,2004b) 12 M 54 6 90 40 59 15.0
(Pattersonetal.,2014) 8 M 54 5 90 40 59 13.1
Mean±SD 9±2 59±5 5±1 90±0 40±0 58±4 7±6
LTHA≥10d
(Gibson,etal.,2015) 8 M 57 10 90 40 41 15.0
(Pattersonetal.,2004b) 12 M 54 16 90 40 59 14.4
(Pattersonetal.,2014) 8 M 54 17 90 40 59 12.8
Mean±SD 10±2 55±2 14±4 90±0 40±0 53±10 14±1
Notes:LTHA,long-termheatacclimation;M,male;PV,plasmavolume;RH,relativehumidity;STHA,short-termheatacclimation;Temp,ambienttemperature;V9O2Peak,peakoxygen
uptake.

54
Eichnaetal.(1950)demonstratedthatHAreducesvoluntarydehydrationasaresultofanimproved
relationshipbetweenthirstandbodywaterneeds.Consequently,heat-acclimatedindividualsare
betterabletomaintainhydrationduringexerciseintheheat,andthusminimizebodywaterdeficits
and voluntary dehydration, provided that access to fluids is not restricted. This is an essential
adaptation since following HA SWR increases and if fluid replacement is not proportionately
increased,greaterdehydrationwilloccur.AlongwiththeexpansionofPVinducedduringHA,the
ability to better maintain fluid balance through thirst represents an adaptive response that
contributestoreducecardiovascularstrainduringsubsequentheatstress.
2.15.3. Cardiovascularstability
Following STHA and LTHA using a controlled hyperthermia model, resting HR reduces by 3
beats.min-1and12beats.min-1respectively(table2.7).Thereareseveralpotentialmechanismsthat
explainthereductionincardiovascularstrain.Thesemechanismsinclude,animprovedskincooling
andredistributionofbloodvolume,PVexpansion, increasedvenousreturnfromcutaneousand
non-cutaneousvascularbedsandreducedTskandTc(table2.8).
FollowingHA,centralcardiacfunctionwasassessedbyeitherwalkingat5.6km.h−1(Rowelletal.,
1967) or cycling at 50% of their maximal aerobic power (Nielsen et al., 1993) until volitional
exhaustion.Duetotheprotocoladministeredtheheatstrainwasvariablebetweensessionsand
between the pre andpostHAheat stress tests.Nevertheless, both groups observed significant
elevationsinstrokevolumealongwithreducedHRfollowingHA.Thesechangessignifyanimproved
cardiacfunctionsinceHRandTcwerelower(Nielsenetal.,1993;Rowelletal.,1967).Wyndham,
(1951)andWyndhametal.,(1976,1968)findingsaredivergenttothatobservedbyRowelletal.
(1967)andNielsenetal.(1993).ThesestudiesalladoptedafixedintensityHAprotocolandthus,
theabsolute stress levelwas identicalbetween thepreandpostheat stress test. FollowingHA
cardiacoutputandHRdeclined,whilestrokevolumeremainedconstant.Theseobservationsmay
inpartbeexplainedbythereductionintheHAstimulustowardstheendofHA,asaresultonthe
fixedintensityprotocol.However,Wyndhametal.(1976)reporteda25%increaseincardiacoutput
following4–7dofHA,howeverthiswasonly8%following10d.Theobservedchangestocardiac
outputmayinpartbemediatedbyPVexpansionwhichwasreportedinacompanionpaperusing
thesameparticipants(Senayetal.,1976).Inthispaperitwasreportedthattherewasagradual
elevationinPVoverthefirst6-dofHA,afterwhichitremainedstable.Interestingly,nochangesin
HR,strokevolumeorcardiacoutputwerereportedduringsubsequentmaximalexerciseintheheat
(40°C,30%RH)following10-dHAintrainedcyclists(Lorenzoetal.,2010).However,whenthesame

55
exercise was performed in cool environment (13°C, 30% RH), both stroke volume and cardiac
outputweresignificantlyhigher.
Gotoetal.(2010)reportedrestingstrokevolumetobeelevatedfollowingHA.Thisispotentially
mediatedbyabettermaintenanceofventricularfillingpressure(GledhillandJamnik,1994),and
blood volume, which support venous return and cardiac preloading. Indeed, during times of
multisystemstress,asmallPVelevationcandictateone’sabilitytoaccommodatethesestresses
and to avoidhypotension and cardiovascular insufficiency.Workingwith a ratmodel,Horowitz
(2003)hasadvancedourunderstandingintothecardiovascularadaptationachievedfollowingHA.
Whilethetranslationofthesemodificationstohumansisspeculativeatpresent,theyhelpexplain
someadaptationspreviouslyexplained.There isrightwardshift inthediastolicpressure-volume
relationship of the heart following chronic heat exposure (Horowitz et al., 1986b), signifying
increased cardiac compliance. This is important, since it indicates the heart can be filledmore
readily and without relying on the relatively small blood volume elevation accompanying heat
adaptation to increase central venous pressure. This observed change in compliance enhances
strokevolume,making theheartmoreeconomicaland lowering themetaboliccostofpumping
(Horowitz et al., 1986b). The corresponding increase in efficiency reduces metabolic heat
production. Finally, since there is an increase in myocardial contractile performance following
endurancetraining(SchaibleandScheuer,1979)andHAtypicallyinvolvesenduranceexercise,then
overallcardiacfunctionappearstobeenhanced.
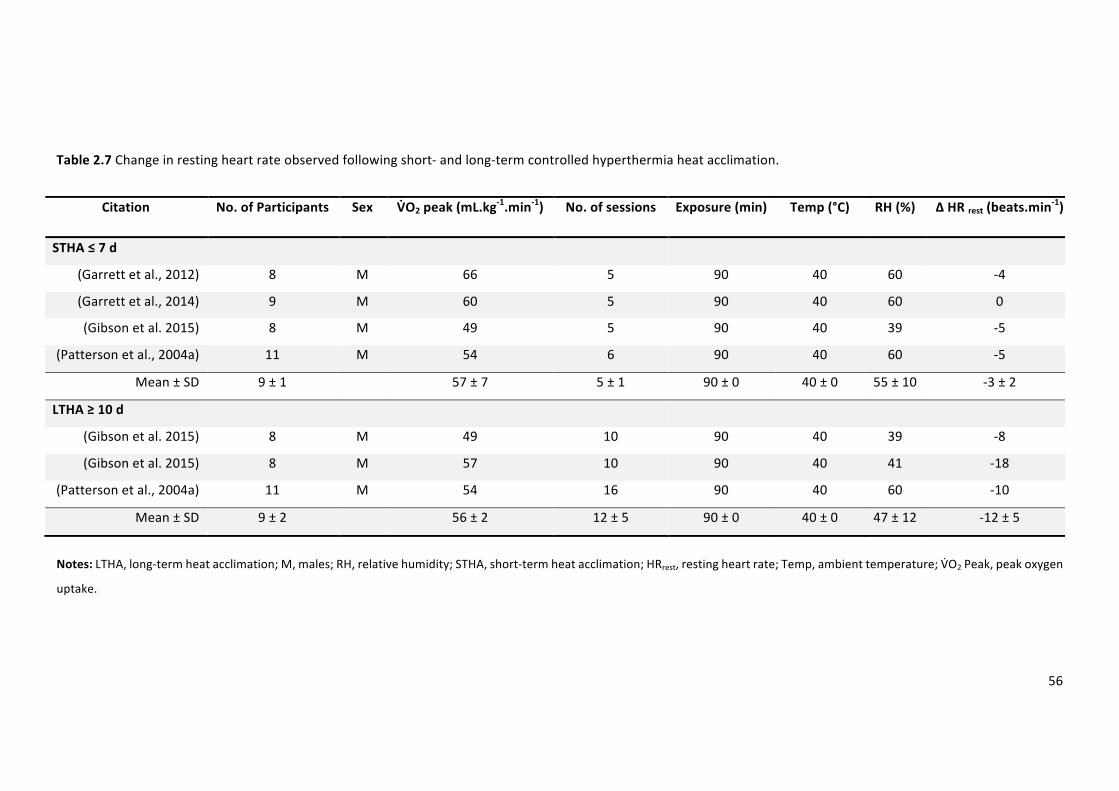
56
Table2.7Changeinrestingheartrateobservedfollowingshort-andlong-termcontrolledhyperthermiaheatacclimation.
Citation No.ofParticipants Sex V9O2peak(mL.kg-1.min-1) No.ofsessions Exposure(min) Temp(°C) RH(%) ∆HRrest(beats.min-1)
STHA≤7d
(Garrettetal.,2012) 8 M 66 5 90 40 60 -4
(Garrettetal.,2014) 9 M 60 5 90 40 60 0
(Gibsonetal.2015) 8 M 49 5 90 40 39 -5
(Pattersonetal.,2004a) 11 M 54 6 90 40 60 -5
Mean±SD 9±1 57±7 5±1 90±0 40±0 55±10 -3±2
LTHA≥10d
(Gibsonetal.2015) 8 M 49 10 90 40 39 -8
(Gibsonetal.2015) 8 M 57 10 90 40 41 -18
(Pattersonetal.,2004a) 11 M 54 16 90 40 60 -10
Mean±SD 9±2 56±2 12±5 90±0 40±0 47±12 -12±5
Notes:LTHA,long-termheatacclimation;M,males;RH,relativehumidity;STHA,short-termheatacclimation;HRrest,restingheartrate;Temp,ambienttemperature;VTO2Peak,peakoxygen
uptake.
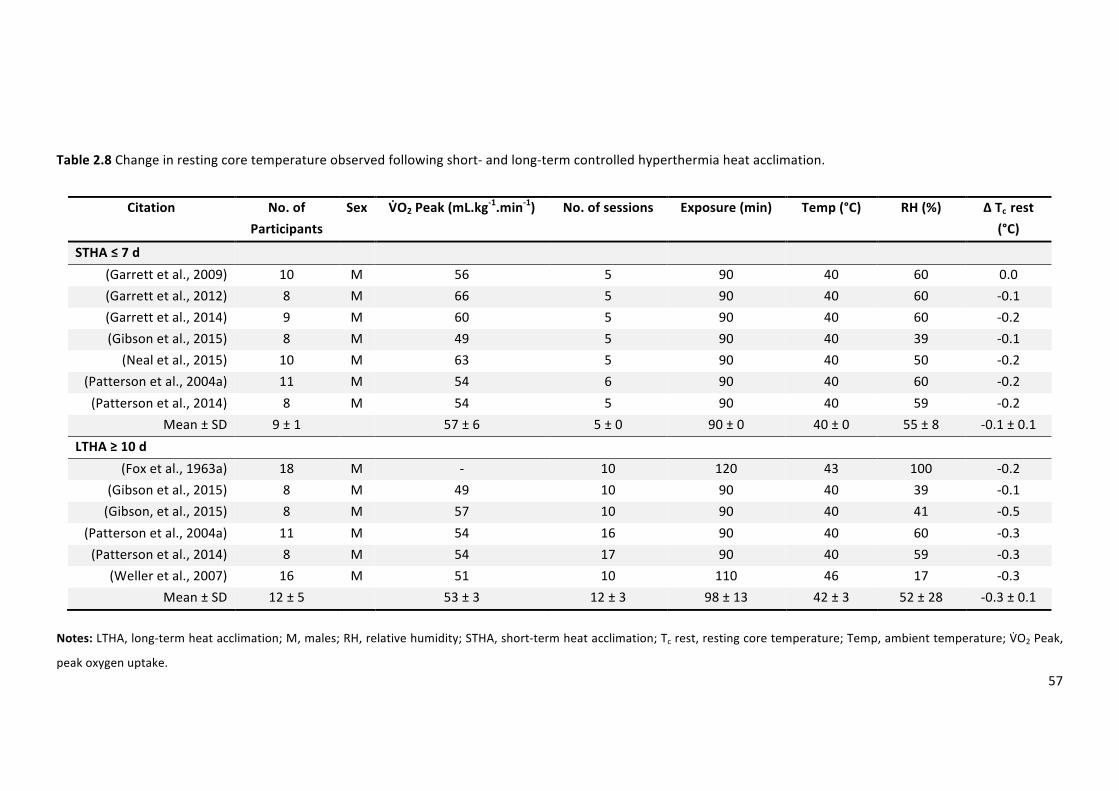
57
Table2.8Changeinrestingcoretemperatureobservedfollowingshort-andlong-termcontrolledhyperthermiaheatacclimation.
Citation No.of
Participants
Sex V9O2Peak(mL.kg-1.min-1) No.ofsessions Exposure(min) Temp(°C) RH(%) ∆Tcrest
(°C)
STHA≤7d
(Garrettetal.,2009) 10 M 56 5 90 40 60 0.0(Garrettetal.,2012) 8 M 66 5 90 40 60 -0.1(Garrettetal.,2014) 9 M 60 5 90 40 60 -0.2(Gibsonetal.,2015) 8 M 49 5 90 40 39 -0.1(Nealetal.,2015) 10 M 63 5 90 40 50 -0.2
(Pattersonetal.,2004a) 11 M 54 6 90 40 60 -0.2(Pattersonetal.,2014) 8 M 54 5 90 40 59 -0.2
Mean±SD 9±1 57±6 5±0 90±0 40±0 55±8 -0.1±0.1
LTHA≥10d
(Foxetal.,1963a) 18 M - 10 120 43 100 -0.2(Gibsonetal.,2015) 8 M 49 10 90 40 39 -0.1(Gibson,etal.,2015) 8 M 57 10 90 40 41 -0.5
(Pattersonetal.,2004a) 11 M 54 16 90 40 60 -0.3(Pattersonetal.,2014) 8 M 54 17 90 40 59 -0.3
(Welleretal.,2007) 16 M 51 10 110 46 17 -0.3Mean±SD 12±5 53±3 12±3 98±13 42±3 52±28 -0.3±0.1
Notes:LTHA,long-termheatacclimation;M,males;RH,relativehumidity;STHA,short-termheatacclimation;Tcrest,restingcoretemperature;Temp,ambienttemperature;VTO2Peak,
peakoxygenuptake.

58
2.15.4. Thermotolerance
Thermotoleranceoracquiredcellularthermotoleranceisthenomenclatureusedtodescribethe
cellular adaptation caused by a single, or repeated severe, but non-lethal heat exposure that
preconditionsanorganismtosurviveasubsequentandotherwiselethalheatstress(Kuennenet
al.,2011;Moseley,1997).IthasbeensuggestedthatthermaltoleranceandHAshareacommon
basis,asbothmaypotentiallybegovernedbytheheatshockresponse(HSR)(Kuennenetal.,2011).
Assuch, thermal toleranceandHAarecomplimentary,asHAreducesheatstrainandtolerance
increases survival toagivenheat strain. Forexample,Maloyanetal. (1999)demonstrated that
rodents,withfullydevelopedthermotolerance,survive60%moreheatstrainthanwhatwouldhave
been initially lethal. Thermal tolerance is associatedwithheat shockproteins (HSP)2 binding to
denaturedornascentcellularpolypeptidesandprovidingprotectionandacceleratingrepairfrom
heat stress, as well as fever, hypoxia, ischemia, viral infection, energy depletion, and acidosis
(Kregel,2002).HSParegroupedintofamiliesbasedupontheirmolecularmass(8to110kDa),with
HSP72beingparticularlyresponsivetoheatstressandexercise(Locke,1997).Heatshockprotein
families have different cellular locations and functions. At the intracellular (iHSP) level, HSP’s
process stress-denaturedproteins,manageprotein fragments,maintain structuralproteins,and
chaperoneotherproteinsacrosscellmembranes.Intheextracellular(eHSP)milieu,itissuggested
thatHSPactasasignal,triggeringanimmuno-stimulatoryresponse(Pockley,2003).
Heatshockproteinexpressionincreasesduringandfollowingexposuretoheatstresswithdiffering
responses across various tissues (e.g., brain and liver exhibit a greater response than skeletal
muscle).Aftertheinitialexposure,HspmRNAlevelspeakwithinanhourandsubsequentprotein
synthesisdependsuponboththeseverityandcumulativeheatstress(Maloyanetal.,1999).Passive
heat exposure and physical exercise both elicit HSP synthesis (Febbraio and Koukoulas, 2000);
however, the combination of exercise and heat exposure elicits a greater response than either
stressorindependently(Skidmoreetal.,1995).Duringexerciseintheheat,theexpressionofeHsp
hasbeenshowntobebothdurationandintensitydependent,relatingtothelevelofhyperthermia
attainedandrateofriseinTc(Périardetal.,2012).
2The termheat shock protein (HSP)will be used throughout this thesis to describe the proteinexpression whereas heat shock protein mRNA (Hsp mRNA) will be used to describe the geneexpression.

59
Horowitz&Robinson(2007)reportedthatHAincreasesHSP70reservesandacceleratestheHSRin
animals.Inhumans,theHSRduringHAandtheconcomitantexpressionofiHspandeHspremain
somewhatunclear.Following2-dexerciseheatexposureareducedbasallevelofeHsp72andan
increaseexpressionimmediatelypost-exercisehasbeenreported(Marshalletal.,2006).However,
ithasalsobeenshownthatiHSP72isunaffectedoverthesametimecourse(Marshalletal.,2007).
Yamadaetal.(2007)demonstratedthatwhenHAwasextendedto10d,basaliHSP72expression
increased, which blunted the post-exercise induction response. In contrast, eHSP72 remained
unchanged.During a 15-dHA regimen, basal eHSP72 progressively increased and post-exercise
expressiondecreasedina29-yr.oldmaleultra-marathonrunnerpreparingfortheMarathondes
Sables,suggestingthatalongerHAperiodmayinducegreatercellularadaptations(Sandströmet
al.,2008).McClungetal.(2008)conductedafurtherstudyinwhichiHSP72andiHSP90responses
to10-dofexerciseHAwerecorrelatedwithphysiologicaladaptations.SuchthatHAincreasedbasal
levels of both iHSP72 and iHSP90 and individuals who demonstrate the greatest physiological
adaptation exhibit a reduced post-exercise expression (measured ex vivo via water bath
incubation).11-dofcontrolledhyperthermia(1.0°CTcelevation)HAresultedinanincreaseinbasal
iHSP72levels,whileeHSP72expressionremainedunchanged(Magalhãesetal.,2010).Similarto
the findings of McClung et al. (2008), Magalhães et al. (2010) reported a blunted expression
followingexerciseheatexposure inboth iHSP72andeHSP72.Thus, itappearsthat iHSPmaybe
moresensitivetoheatstressthaneHSPandanincreaseinbasallevelduringHAresultsinablunting
of theacute response toexerciseasHAdevelops.ThemeasurementofHsp72mRNA,offersan
alternative marker of the magnitude of the cellular stress response required for increased
thermotolerance,allowingforthedeterminationofwheretheinhibitionhasoccurred.Gibsonet
al.(2015)reportedequalsessionalHsp72mRNAtranscriptionacrossa10-dHAusingbothfixedand
controlledhyperthermia (figure2.9)protocols.TheequalHsp72mRNAincreasesoccurringafter
equal,reducedorincreasedTcfollowingboth5and10-dHA,suggestthataslongasaminimum
endogenouscriterionissurpassed,additionalendogenousthermoregulatorystrainisnotoffurther
benefit,noriscontinualexerciseloadcrucialsolongashyperthermiaispresent.
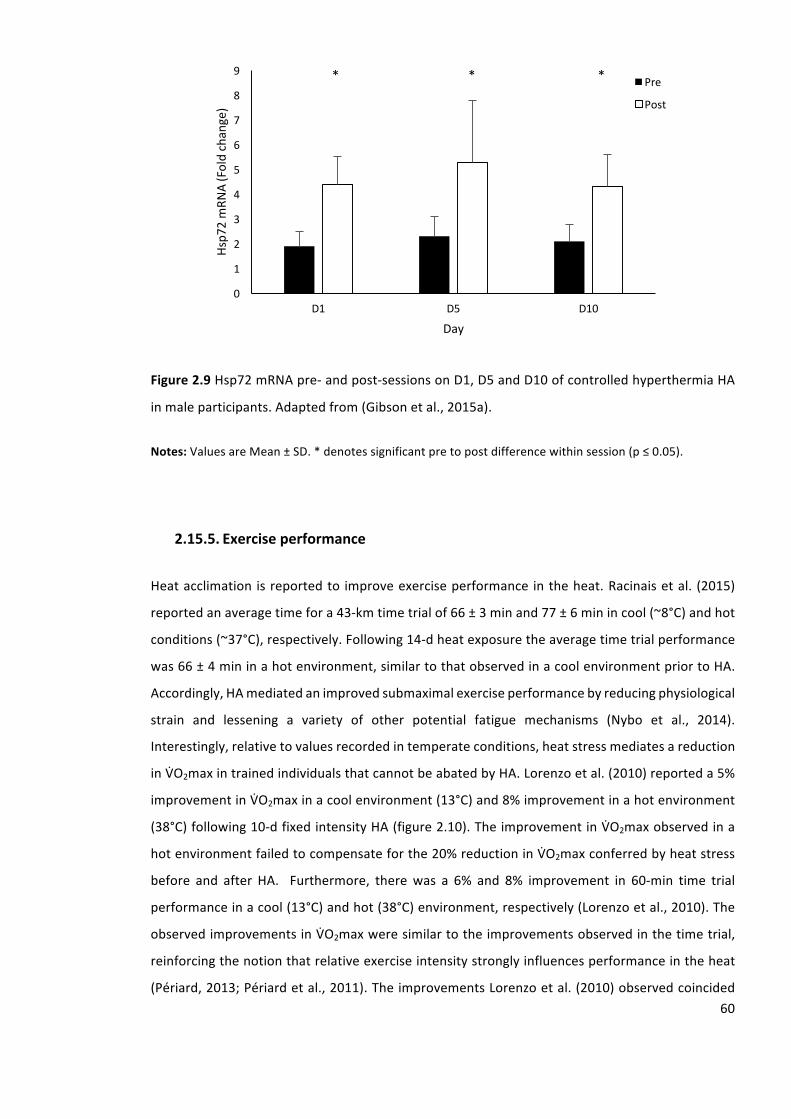
60
Figure2.9Hsp72mRNApre-andpost-sessionsonD1,D5andD10ofcontrolledhyperthermiaHA
inmaleparticipants.Adaptedfrom(Gibsonetal.,2015a).
Notes:ValuesareMean±SD.*denotessignificantpretopostdifferencewithinsession(p≤0.05).
2.15.5. Exerciseperformance
Heatacclimation isreportedto improveexerciseperformance intheheat.Racinaisetal. (2015)
reportedanaveragetimefora43-kmtimetrialof66±3minand77±6minincool(~8°C)andhot
conditions(~37°C),respectively.Following14-dheatexposuretheaveragetimetrialperformance
was66±4mininahotenvironment,similartothatobservedinacoolenvironmentpriortoHA.
Accordingly,HAmediatedanimprovedsubmaximalexerciseperformancebyreducingphysiological
strain and lessening a variety of other potential fatigue mechanisms (Nybo et al., 2014).
Interestingly,relativetovaluesrecordedintemperateconditions,heatstressmediatesareduction
inVgO2maxintrainedindividualsthatcannotbeabatedbyHA.Lorenzoetal.(2010)reporteda5%
improvementinVgO2maxinacoolenvironment(13°C)and8%improvementinahotenvironment
(38°C)following10-dfixedintensityHA(figure2.10).TheimprovementinVgO2maxobservedina
hotenvironmentfailedtocompensateforthe20%reductioninVgO2maxconferredbyheatstress
before and after HA. Furthermore, therewas a 6% and 8% improvement in 60-min time trial
performanceinacool(13°C)andhot(38°C)environment,respectively(Lorenzoetal.,2010).The
observedimprovementsinVgO2maxweresimilartotheimprovementsobservedinthetimetrial,
reinforcingthenotionthatrelativeexerciseintensitystronglyinfluencesperformanceintheheat
(Périard,2013;Périardetal.,2011).TheimprovementsLorenzoetal.(2010)observedcoincided
0
1
2
3
4
5
6
7
8
9
D1 D5 D10
Hsp7
2mRN
A(Foldchange)
Day
Pre
Post
***
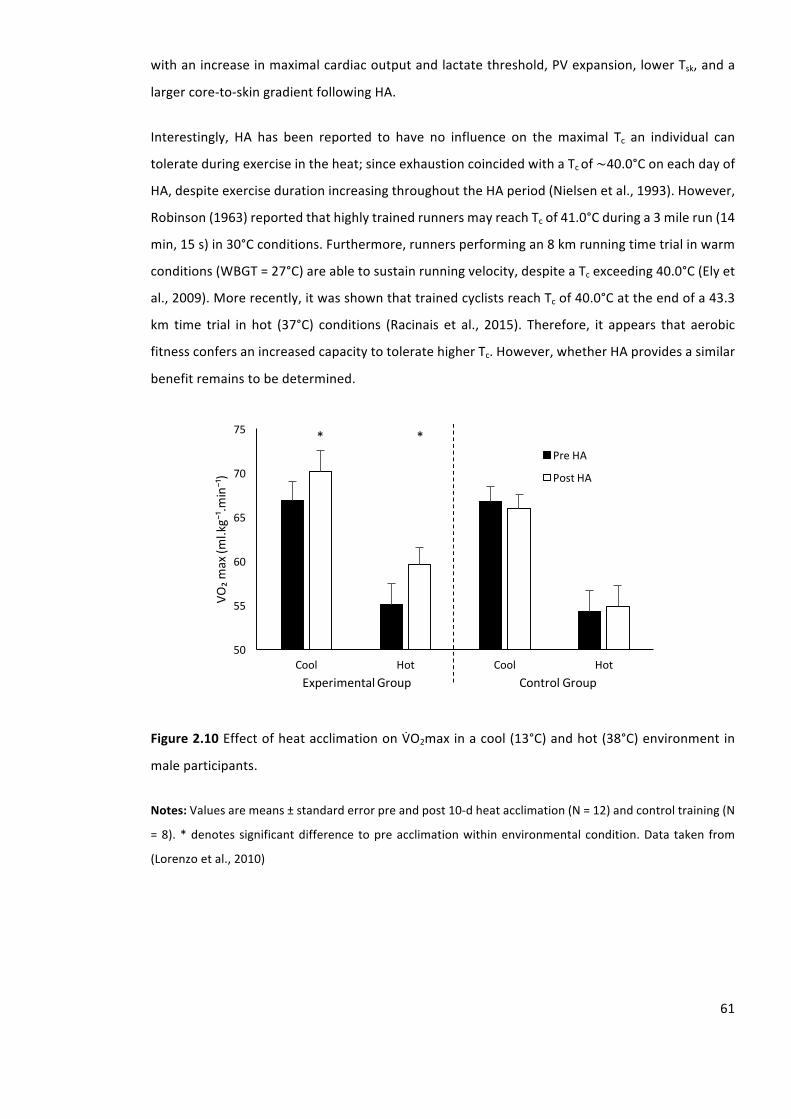
61
withanincreaseinmaximalcardiacoutputandlactatethreshold,PVexpansion,lowerTsk,anda
largercore-to-skingradientfollowingHA.
Interestingly, HA has been reported to have no influence on themaximal Tc an individual can
tolerateduringexerciseintheheat;sinceexhaustioncoincidedwithaTcof∼40.0°ConeachdayofHA,despiteexercisedurationincreasingthroughouttheHAperiod(Nielsenetal.,1993).However,
Robinson(1963)reportedthathighlytrainedrunnersmayreachTcof41.0°Cduringa3milerun(14
min,15s)in30°Cconditions.Furthermore,runnersperformingan8kmrunningtimetrialinwarm
conditions(WBGT=27°C)areabletosustainrunningvelocity,despiteaTcexceeding40.0°C(Elyet
al.,2009).Morerecently,itwasshownthattrainedcyclistsreachTcof40.0°Cattheendofa43.3
km time trial in hot (37°C) conditions (Racinais et al., 2015). Therefore, it appears that aerobic
fitnessconfersanincreasedcapacitytotoleratehigherTc.However,whetherHAprovidesasimilar
benefitremainstobedetermined.
Figure2.10EffectofheatacclimationonVgO2maxinacool(13°C)andhot(38°C)environmentin
maleparticipants.
Notes:Valuesaremeans±standarderrorpreandpost10-dheatacclimation(N=12)andcontroltraining(N
=8).*denotessignificantdifference topreacclimationwithinenvironmentalcondition.Data taken from
(Lorenzoetal.,2010)
50
55
60
65
70
75
Cool Hot Cool Hot
VO₂m
ax(m
l.kg⁻¹.m
in⁻¹)
ExperimentalGroupControlGroup
PreHA
PostHA
**

62
2.15.6. Perceptualresponses
Areduction in thermal sensation (TS) ispotentiallybeneficial forperformance in theheat since
thermal discomfort drives true behavioural thermoregulation (Flouris, 2011). Initiation of these
responsepathwaysduringexerciseelicitsbehaviouralresponseswhichreduceworkrate(Tucker
etal.,2006,2004).Thisisundesirablewithregardstooptimalperformanceintheheat.However,
HAisreportedtoreduceTSduringexercise,performedatasetintensityintheheat.Gibsonetal.,
(2015b)reportedasignificantreductioninbothmeanandpeakTSduringa30-minfixedintensity
heatstresstestfollowing10-d,butnot5-d,ofcontrolledhyperthermiaHA.Theseobservationsare
supportedbyGarrettetal.,(2012)whoobservednochangesinthermalcomfortfollowing5-dof
controlledhyperthermiaHA.Incontrast,Nealetal.,(2015)reportedparticipantshadanimproved
thermalcomfortfollowing5-dcontrolledhyperthermiaHAduringa20-minfixedintensitycycling
intemperateconditions(22°C,60%).TheroleofTSorthermalcomfortareyettobefullyelucidated
withregardstoHA.
2.16. Timecourseofheatacclimation
Heat acclimation is a relatively rapid process that begins on the first day of exposure to a hot
climate.Duringthefirst4-7dofexposure75-80%oftheadaptationsoccur(ArmstrongandMaresh,
1991;Pandolf,1998).TheArmstrongandMareshtableforplateaudaysofphysiologicaladaptation
duringheatacclimatisation(ArmstrongandMaresh,1991)hasevolvedtobeatemporalpatterning
modelofHAacquisition(table2.9).Duringtheinitialheatexposure,physiologicalstrainishigh,as
demonstratedbyanelevatedTcandHR.However,thephysiologicalstraininducedbyheatstress
progressively decreases each day of exposure (Buono et al., 2009a; Houmard et al., 1990).
Cardiovascularfunctionadaptsmostrapidlyin4–5dandvirtuallycompletefollowingsevendaily
exposures(Gibsonetal.,2015;Fujiietal.,2012;Garrettetal.,2009).Inaddition,majorityofthe
thermoregulatoryresponsesincludingreductionsinTskandTctypicallyoccurduringSTHA(Gibson
etal.,2015;Fujiietal.,2012;Garrettetal.,2009).TheexpansionofPV,alongwiththermoregulatory
cardiovascular and metabolic adaptations, improves the perception of effort in athletes.
Traditionally, STHA has failed to provide a strong-enough stimulus for sudomotor adaptation
(Cotteretal.,1997;Sunderlandetal.,2008).However,Buonoetal.(2009)reportedasignificant
increaseinSWRfollowing4-dofHAtohumidconditions.Thus,STHAappearstobesufficientat
inducing performance-enhancing adaptations (Lorenzo et al., 2010), which may be more
pronouncedafterfluidregulatorystrainfromapermissivedehydrationHAregimen(Garrettetal.,
2014,2011;TaylorandCotter,2006).

63
Table2.9Plateaudaysofphysiologicaladaptations(thepointatwhichapproximately85%ofthe
adaptationoccurs)duringheatacclimatisation.
AdaptationDaysofheatacclimatisation
1 2 3 4 5 6 7 8 9 10 11 12 13 14
HRdecrease ______________
PVexpansion ______________
Tredecrease ______________
RPEdecrease ______________
SweatNaCldecrease __________________
SWRincrease ___________________________
RenalNaCldecrease __________________
Notes:HR,heartrate;NaCl,SodiumChloride;PV,plasmavolume;Tre,rectaltemperature;SWR,sweatrate.
AdaptedfromArmstrongandMaresh,(1991).
Theplateauinresponsesfollowing3-6dHA(table2.9)haspreviouslybeenviewedasfavourable.
However,traditionalmethodsofHAmayconstraintheadaptationandthustheplateaumaysimply
reflectincompleteadaptation.Thereisanabsenceofinformationonthetemporalpatterningof
controlledhyperthermiaHA,thisisproblematicsinceitisessentialthatwherepossibletheduration
ofHAadministeredelicitsoptimaladaptation.
2.17. Heatacclimationdecay
Heatacclimationisatransientprocessandwillgraduallydisappearifnotmaintainedbycontinued
repeated exercise-heat exposure. Literature is consistent in reporting that the physiological
adaptationsassociatedwithHAarerelativelyshort-termandmaydecayonlyafewdaysorweeks
afterremovalfromheatexposure(ArmstrongandMaresh,1991).Thefirstadaptationstooccur
aretypicallythefirsttodecay,withthephysiologicaladaptationsthattakelongertodevelop,such
assudomotorfunction,willhaveaslowerrateofdecay(Taylor,2000).Flourisetal.(2014)assessed
thedecayof14-doffixedintensityHA.TheobservedchangesinTcpersistforatleast2-wksafter
removalfromheatexposure;whilethechangesinHRandHRvariabilitydecayfasterandareonly

64
partlyevidentafter2-wksofnon-exposuretoheat.Furthermore,Garrettetal.(2009)assessedthe
decayof5-dcontrolledhyperthermiaHA following2,9,16and23-dwithoutexposure toheat.
STHAinducedadaptationspermittingincreasedheatloss;theseadaptationspersistedforone,but
not 2-wks following HA. Thus, when using a cross over design to assess two HA methods,
considerationneedstobetakentoensureanadequatewashoutperiodisused.
Furthermore, the re-inductionofHAafter its lossoccursmarkedly faster thanduring the initial
induction period (Tetievsky et al., 2008).Weller et al. (2007) assessed the re-induction of 10-d
controlled hyperthermia HA following 12 and 26-d without exposure to heat. It took 2-4 d
respectivelytorestorethepreviousobservedphysiologicalbenefits.Thus,onceadaptationtothe
heathadbeenattained,thetimethatindividuals’mayspendincoolerconditionsbeforereturning
toahotenvironment,couldbeaslongasonemonthwithouttheneedforextensivere-adaptation
toheat.Thisobservation isconsistentwithresearchconcerningadaptationmemory,withsome
acquiredfunctionalchangesbeingrapidlyrestored(Tetievskyetal.,2008).Thus,whenusingacross
over design to assess two HA methods, consideration needs to be taken to ensure trials are
successfullyrandomisedtoreducetheeffectofHAmemory.
2.18. Females’responsestoheatacclimation
There is a dearthof literature assessing females’ responses toHA. Previously, thephysiological
responses ofmales and females to 10-d fixed intensityHAwere assessedwith females initially
exhibiting lower Tre and HR, despite a lower SWR compared withmales (Avellini et al., 1980).
Following HA, the physiological strainwas similar betweenmales and females, althoughmales
maintained a greater SWR. This study adopted a traditional HA protocol which results in a
progressivedeclineintheadaptationstimulusoverthedurationofHA.Controlledhyperthermia
ensures consistent potentiating stimuli for adaptation throughout the HA period, eliciting
reductionsinthermalstrainandincreasesinworkcapacityduringbothSTHA(Garrettetal.,2012,
2009)andLTHA(Pattersonetal.,2004a),potentiallypromotingmorecompleteadaptation(Taylor
andCotter,2006).Furthermore,Sunderlandetal.(2008)assessedtheresponsesoffemalegames
players responses to STHA. Results suggest, that females fail to establish typical phenotypic
adaptations, namely reduction in Tre and HR despite a 33% increase in intermittent sprint
performance in the heat. These typical adaptive responses have been previously observed in
trainedmalesfollowingSTHA(Buonoetal.,1998;Fujiietal.,2012;Garrettetal.,2011;Poirieret
al., 2015; Racinais et al., 2012); suggesting females may require LTHA to achieve adaptation.
However,duetotheself-pacedexerciseadministeredpreandpostHAbySunderlandetal.(2008),
females were exercising at a higher absolute intensity following HA, suggesting an increase in

65
metabolicheatproduction;potentiallynegatinganyimprovementsinthermoregulationachieved
throughHA.Itremainsunknowntheextenttowhichfemales’adapttothecontrolledhyperthermia
model of HA. The absence of evidence concerning effective STHA protocols in females is
problematicforathletes,sinceSTHAisapreferredregimeasitprovideslessdisruptionfromquality
trainingpriortocompetitioncomparedwithtraditionalHAprotocols.
2.19. Aimofthesis
AHTTusinga runningmodeofexercise,dataon females’phenotypicandcellular responses to
controlledhyperthermiaHAandestablishingoptimisedHAstrategiesforfemalesarerequired.This
thesiswillthereforeconsistofaseriesofexperimentswhichwillfirstlyassesstherepeatabilityand
thesensitivityofaRHTT.Oncethishasbeenestablished,theRHTTwillbeusedtocomparethe
phenotypicandcellularadaptationresponsestocontrolledhyperthermiainmalesandfemales.A
furtherexperimentwill thenbeperformed toevaluateanovelHA strategy in females across a
short-termtimescale.
2.19.1. Researchquestionsarisingfromtheliteraturereview
The literature review has identified a number of areas for investigation still outstanding. The
incidenceofheat-related illnesses in running isapparent,yet there iscurrentlynostandardised
methodtoevaluatearunner’sheattolerance.ThedevelopmentofaRHTTiswarranted.Itiswell
understoodthatcontrolledhyperthermiaHAelicitsmoreoptimaladaptationcomparedwithfixed
intensityprotocols.Aseriesofintegratedphysiologicalandcellularadaptationselicitanenhanced
exerciseheattolerance.Whilstthetemporalpatterningandlengthofexposurerequiredtoelicit
phenotypic and cellular adaptations following controlled hyperthermia HA are becoming well
understoodinmales,littleprogressionhasbeenmadewithregardstofemales’responses.Thus,a
comparisonofmales’andfemales’phenotypicandcellularresponsestocontrolledhyperthermia
HAiswarranted.Oncefemales’temporalpatterningassociatedwithcontrolledhyperthermiaHA
hasbeenidentified,itremainstobedeterminedwhethertherearemoreoptimisedHAmethods
toestablishanacceleratedadaptioninfemales.Passiveheatexposurestosaunasandhotwater
have successfully elicited phenotypic adaptation in males; consideration of these protocols is
requiredinfemales.

66
2.19.2. Proposedresearchstudiesandhypotheses
Thefollowingresearchquestionsandhypothesesareproposedforthisthesis.
Repeatabilityofarunningheattolerancetest
Aim:Toestablishtherepeatabilityofapracticalrunningtesttoevaluateanindividual’sabilityto
tolerateexerciseheatstress.
Hypotheses: Itwasprimarilyhypothesised thatphysiological responsesduring the runningheat
tolerancetestwillhaveastrongpositivecorrelationonrepeatedtrials.Secondly,therewillbeno
differences inphysiological responsesduring the runningheat tolerance testbetween repeated
trials.
Sensitivityofarunningheattolerancetest
Aim: To identify the sensitivity of the runningheat tolerance test to changes in heat tolerance
followingshort-termheatacclimation.
Hypotheses:Itwasprimarilyhypothesisedthattherewillbedifferencesinphysiologicalresponses
duringtherunningheattolerancetestfollowingshort-termheatacclimation.Secondly,therunning
heattolerancetestwillbesensitivetoindividualdifferencesinthemagnitudeofadaptation.
Acomparisonofmaleandfemaletemporalpatterningtoshort-andlong-termheatacclimation
Aim: To determine the sex differences in thermoregulatory, cardiovascular, and sudomotor
adaptationtoshort-termandlong-termheatacclimation.
Hypotheses:ItwasprimarilyhypothesisedthatmaleswillachievegreaterreductionsinTreandHR
followingSTHAcomparedwithfemales.Secondly,thattherewillbenodifferencesinthereduction
inTreandHRbetweenmalesandfemalesfollowingLTHA.Finally,thatfemaleswillachieveagreater
increaseinSWRfollowingSTHAandLTHAcomparedwithmales.
SexcomparisonofleukocyteHsp72mRNAtranscriptionduringheatacclimation
Aim:Toexaminethesexdifferencesinheatshockprotein72mRNAtranscriptionduringcontrolled
hyperthermiaheatacclimationovershort-andlong-termtimescales.

67
Hypothesis:Itwashypothesisedthatheatshockprotein72mRNAresponsewillbelowerinfemales
comparedtomalesacrossthecourseofcontrolledhyperthermiaheatacclimation.
Passiveheatexposureprecedingshort-termheatacclimationacceleratesphenotypicadaptation
infemales.
Aim:Todeterminewhethershort-termheatacclimationprecededbyapassiveheatexposureto
sauna-likeconditionsacceleratedheatadaptationinfemales.
Hypotheses:Itwasprimarilyhypothesisedthatshort-termHAprecededbyapassiveheatexposure
willresultinagreaterplasmavolumeexpansioninfemalescomparedwithheatacclimationalone.
Secondly,short-termheatacclimationprecededbyapassiveheatexposurewillproduceagreater
sudomotoradaptationinfemalescomparedwithheatacclimationalone.

68
3. Generalmethods
________________________________________________________________________________
Thischapterdescribesthegeneralmethodsandmaterialsusedwithintheexperimentalchapters
of this thesis.When additional ormodifiedmeasureswere used, full descriptions are included
withinthemethodssectionoftherelevantexperimentalchapters.
3.1. Ethics,healthandsafety
AllexperimentsreportedwithinthisthesiswereapprovedbytheUniversityofBrightonResearch
Ethics&GovernanceCommitteeandconductedinaccordancewiththeguidelinesoftherevised
DeclarationofHelsinki, 2013.All experimentationwas carriedout in linewith theUniversityof
Brighton’sstandardoperatingproceduresandriskassessmentlaboratoryguidelines.
Experimenters
Therewere at least two experimenters present throughout each experimental trial within this
thesis.Oneexperimenterremainedwithintheenvironmentalchamberattendingtotheparticipant
and one experimenter remained outside the environmental chamber, ensuring the safety of
individualswithin the chamber for the duration of each trial. Furthermore, at least one of the
experimenterspresentduringtheexperimentaltrialswasaqualifiedfirstaider.
Equipmentcleaningprocedure
To avoid contamination, all equipment used was cleaned before and after use. Respiratory
apparatuswassoakedin1%Virkondisinfectant(AntecInternational,UK)foraminimumof10-min
aspermanufacturerinstructions.Apparatuswasthenthoroughlyrinsedanddriedpriortouse.HR
monitorstrapsweresoakedfor10-minin1%Virkondisinfectant,rinsedanddriedfollowinguse.
Allskinthermistorswerecleanedusingalcoholwipes(MölnlyckeHealthcare,Sweden)beforeand
afteruse.All otherequipment including the cycleergometerand themotorised treadmillwere
wipeddownusingdisinfectantsurfacespray(Bioguard,UK)followinguse.
Controlofsubstanceshazardoustohealth
Control of substances hazardous to health formswere completed and passed onto theWelkin
LaboratoriesBiologicalSafetyofficer forapproval foreverypowderandsolutionusedwithinall

69
experimentspresentedwithinthisthesis.Theseformsoutlinedanyknownrisksassociatedwiththe
powderorsolutionandthecorrectstoring,handling,anddisposalprocedures.
Wastedisposal
Rectalthermistorsandallothernon-reusablewasteweredisposedofbyincineration.Allsharps,
suchasneedlesandlancetsforthemeasurementofbloodweredisposedofinadesignatedsharps
bin.Biologicalmaterialandwastewerehandledanddisposedofinlinewithrelevantguidelines.
Criteriaforterminationofexperiments
Writteninformedconsent(Appendix1)andamedicalhistory(Appendix2)wasobtainedfromeach
participantpriortoanytestingandallparticipantswereinformedoftheirrighttowithdrawfrom
thestudyatanytimewithoutobligationtogivereason.ExercisewasterminatedifTre≥39.7°Cin
linewiththeuniversitiesethicallimits,theparticipantwithdrewduetovolitionalexhaustion,the
participantscouldnolongermaintaintheexerciseintensitydespitestrongverbalencouragement,
or the participant demonstrated signs of heat illness including heat syncope, exhaustion,
disorientation, nausea or vomiting. Upon removal from the chamber, appropriate cooling
techniqueswereadopted.Coolingtechniquesinvolvedcoldwateringestion,coldwaterimmersion
of the hands and feet and sitting in front of a fan. Participants were permitted to leave the
laboratorywhenTrereturnedtowithin0.5°Cofbaselinemeasuresobtainedpriortotesting.
3.2. Participants
Recruitment
Participants were recruited via an email to University of Brighton students and through
advertisementonsocialmedia,includingFacebookandTwitter.Thisprovidedbasicdetailsabout
thestudyandcontactdetails fortheprincipalexperimenter.Prospectiveparticipantswerethen
providedwithaverbalexplanationofthestudy,inadditionanexplanationoftherisksandbenefits.
Adetailedparticipantinformationpackwasthenprovided.
Medicalcriteria
Athletes,aged18–35yrs.oldwhoincludedrunningandcyclingwithintheirweeklytrainingand
hadarecenthistoryofcompetinginanenduranceevent,whohadnoknowninjury,respiratory
problems,diabetes,orcardiacproblemswererecruited.Participantswerenotselectedtotakepart
inanyofthestudiesifanycontraindicationswereidentified.Participantsweredeemedsuitableif

70
theyhadbeenabsentfromrepeatedexternalheatexposureforthepreviousthreemonthsandhad
noknownpreviousincidentofheatillness.Participantswereexcludedfromtakingpartiftheywere
takinganydietarysupplementsormedicationotherthanthecontraceptivepill,hadtakenpartin
otherlaboratoryexperimentsinthethreemonthspreceding,hadexperiencedanaphylacticshock
symptomstoneedles,probesorothermedicalequipmentthatmightprecludethemfrominserting
arectalprobe,orhadaknownhistoryofrectalbleeding,analfissures,haemorrhoids,oranyother
conditionoftherectum.Femaleswhohad irregularmenstrualcycleorwhowereusinganother
formofhormonalcontraceptionotherthanthecombinationpillwereexcludedfromtakingpart.
3.3. Facilities
AlltestingtookplaceinBritishAssociationofSportandExerciseSciencesaccreditedlaboratories,
attheWelkinsiteoftheUniversityofBrighton,EastbourneCampus.Allpreliminarytestingtook
placeintheresearchlaboratory.Allexperimentalheattrialswereconductedintheenvironmental
physiology laboratory which housed an environmental chamber 4.5 x 3.5 x 3m high (WatFlow
controlsystem;TISS,Hampshire,UK).
3.4. Pre-trialdietandexercisestandardisation
Experimentaltrialswereconductedatthesametimeofdayforallparticipantstoavoidaneffect
of circadian variation on physiological variables (Atkinson and Reilly, 1996). Experimentation
occurredduring theUKwinter (meanambient temperatureof5°C); therefore,participantshad
beenabsentfromrepeatedexternalheatexposurefortheprevious3months.Participantswere
instructedtorefrainfromhotbaths,saunarooms,steamrooms,andsunbedsduringthethree
months prior to the experimental testing and throughout the testing period. Furthermore,
participantswere instructed to refrain fromexposure tohypobaric or hyperbaric environments
duringthethreemonthspriortotheexperimentaltestingandthroughoutthetestingperiod.
48-hrspriortoconductingthetrials,participantswereinstructedtomaintainnormalhydrationand
diet,andrefrainfromtheconsumptionofalcohol(Yodaetal.,2005),caffeine(DelCosoetal.,2009),
glutamine, generic supplementation and exhaustive exercise. 2-hrs prior to arrival participants
wereinstructedtoslowlyconsume3-5mL.kg-1(250–400mL)ofwatertoensureadequatehydration
(Sawkaetal.,2007).Participantswererecommendedtoconsumeasmallbreakfastconsistingof
30-50 g ofmultigrain cereal with semi-skimmedmilk, or two slices of wholemeal toast with a
toppingofchoice,2-hrspriortoarrival.Participantswereinstructedtomaintainidenticaldietin
theimmediate48-hrspriortoeachexperimentalsession.

71
Hydrationassessment
Toensureequalandadequatehydrationbetweentrialstheassessmentofhydrationstatuswas
performed.When twooutof the following threecriteriawereachieved,adequatehydration to
performthe trialwasassumed;anosmolalityvalueof≤700mOsm.kg-1,aurinespecificgravity
(USG)valueof≤1.020,oraBMwithin1%ofdailyaverage(Sawkaetal.,2007).Theseexperimental
controls were not violated for any participant for any of the preliminary or experimental
procedures.
Urineosmolality
Urineosmolalitywasmeasuredusingahandheldmicroosmometer(AdvancedInstruments Inc.,
Massachusetts,USA).Themicroosmometerwascalibratedprior toeverysampleusingdistilled
water(Osmolality0mOsm.kg-1).Approximately1mLofurinewasplacedintotheosmometerlens,
wherebythesamplewasmeasuredusingwaterfreezingpointdepression.
Urinespecificgravity
Urinespecificgravitywasassessedusingavisualhandheldrefractometer(IndexInstrumentsLtd.,
Cambridge,UK).Therefractometerwascalibratedpriortoeverysampleusingdistilledwater(USG
1.000). Approximately 2 mL of urine was placed onto a glass lens of the refractometer. The
refractometerwasthenhelduptothelight,whiletheexperimenterlookedthroughtheeyelens
andrecordedthevaluesfromthescalewithin.
Repeatability3ofurinaryanalysis
A urine sample was collected and the same sample analysed for osmolality and USG on ten
occasions.Betweenanalysestheosmometerandrefractometerwascalibratedwithdistilledwater.
Technicalerrorasacoefficientofvariation(TE(CV%)),andintra-classcorrelationcoefficientwith
95%confidenceintervals(ICC95%CI)werecalculatedfromduplicatemeasuresof10samples(table
3.1).
3Reliability,repeatability,reproducibilityandagreementaretermsoftenusedinterchangeably.Thetermrepeatabilitywillbeusedconsistentlythroughoutthisthesissinceitspecificallyreferstothevariationinrepeatedmeasurementsmadeonthesameparticipantunderidenticalconditions.
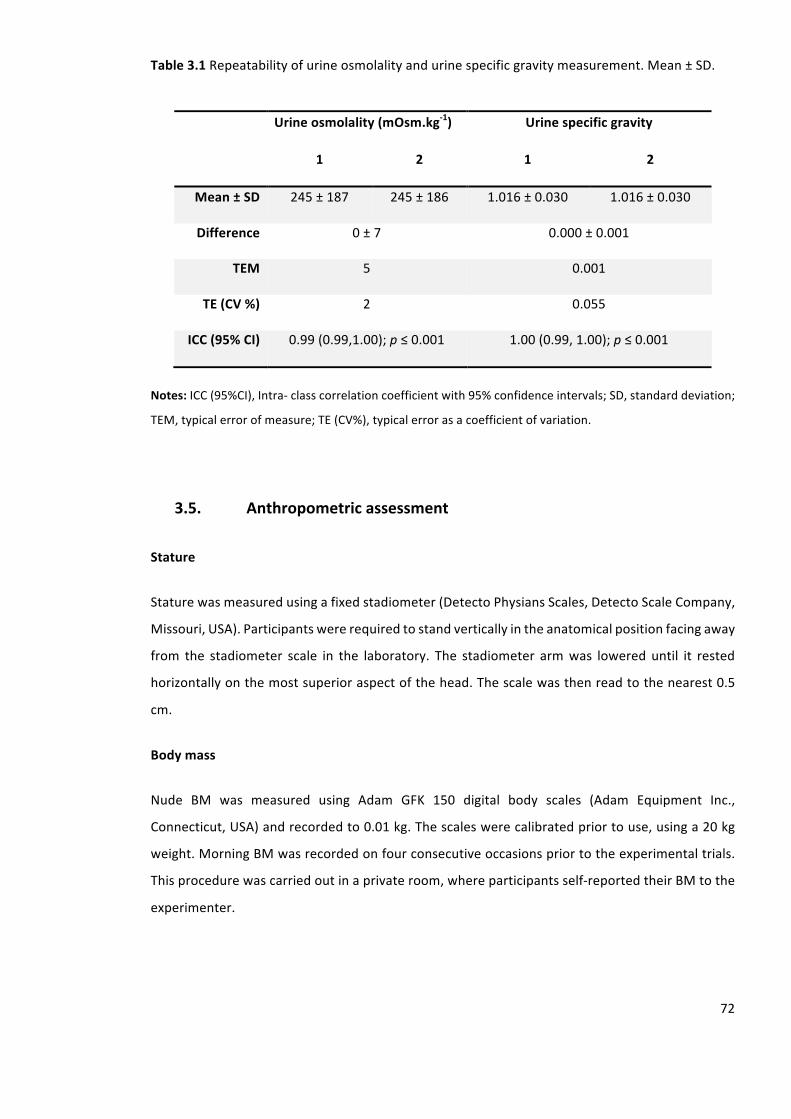
72
Table3.1Repeatabilityofurineosmolalityandurinespecificgravitymeasurement.Mean±SD.
Urineosmolality(mOsm.kg-1) Urinespecificgravity
1 2 1 2
Mean±SD 245±187 245±186 1.016±0.030 1.016±0.030
Difference 0±7 0.000±0.001
TEM 5 0.001
TE(CV%) 2 0.055
ICC(95%CI) 0.99(0.99,1.00);p≤0.001 1.00(0.99,1.00);p≤0.001
Notes:ICC(95%CI),Intra-classcorrelationcoefficientwith95%confidenceintervals;SD,standarddeviation;
TEM,typicalerrorofmeasure;TE(CV%),typicalerrorasacoefficientofvariation.
3.5. Anthropometricassessment
Stature
Staturewasmeasuredusingafixedstadiometer(DetectoPhysiansScales,DetectoScaleCompany,
Missouri,USA).Participantswererequiredtostandverticallyintheanatomicalpositionfacingaway
from the stadiometer scale in the laboratory. The stadiometer armwas lowered until it rested
horizontallyonthemostsuperioraspectofthehead.Thescalewasthenreadtothenearest0.5
cm.
Bodymass
Nude BM was measured using Adam GFK 150 digital body scales (Adam Equipment Inc.,
Connecticut,USA)andrecordedto0.01kg.Thescaleswerecalibratedpriortouse,usinga20kg
weight.MorningBMwasrecordedonfourconsecutiveoccasionspriortotheexperimentaltrials.
Thisprocedurewascarriedoutinaprivateroom,whereparticipantsself-reportedtheirBMtothe
experimenter.

73
Bodysurfacearea
Following measurement of both stature and BM, body surface area (BSA) was subsequently
calculatedusingEquation3.1(DuBoisandDuBois,1916).
Equation3.1Calculationofbodysurfacearea(BSA)
BSA=0.007184xBM0.425xH0.725
Where:BM=bodymassinkg,H=statureinmeters
Skinfolds
Skin fold thickness was determined from the right side of each participant whilst stood in the
anatomicalpositioninambientlaboratoryconditionsof20±1°Cand40±5%RH.Sumofskinfolds
wasdeterminedfromfoursites(DurninandWomersley,1974);thebicep,triceps,subscapular,and
supra-iliacareausingHarpendenskinfoldcallipers(BatyInternational,WestSussex,UK).Skinfold
thicknesswasmeasuredtothenearest2mm.Measuresweretakenintriplicateandtheaverage
recorded.Ifvalueswerenotwithin0.4mmadditionalmeasuresweretaken.
Thebicepskinfoldwaslocatedatthepointontheanteriorsurfaceofthearminthemidlineatthe
levelof themid-acromial-radiale landmark.The tricepskin foldwas locatedat thepointon the
posteriorsurfaceofthearm,inthemidline,atthelevelofthemid-acromial-radialelandmark.The
subscapularskinfoldwaslocated2cmlaterallyandobliquelydownwardsfromtheundermosttip
oftheinferiorangleofthescapula.Thesupra-iliacskinfoldwaslocatedatthepointontheiliac
crest where the midaxilla on the longitudinal axis of the body meets the ilium (Durnin and
Womersley,1974).
Whentheselocationshadbeenmarked,afirmgraspwastakenoftheskinandsubcutaneousfat
ensuringnomuscleorfasciawastakenusingthethumbandindexfinger.Thenthecontactsurface
of thecalliperswasplacedata90-degreeangle to the skin foldapproximately1 cmbelow the
fingers. The pressure was then slightly released between the fingers, ensuring that a greater
pressure was applied by the callipers. The handle of the callipers was then released and the
measurementwasrecordedapproximately4safterthepressurewasreleased.
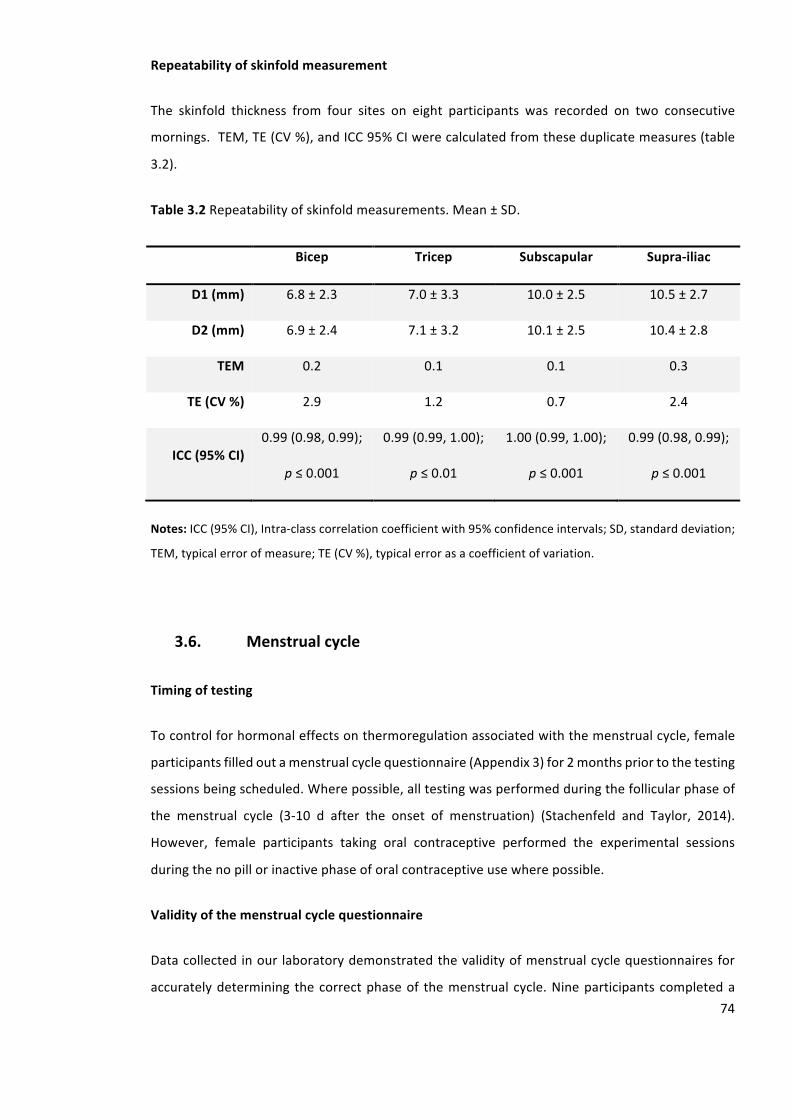
74
Repeatabilityofskinfoldmeasurement
The skinfold thickness from four sites on eight participants was recorded on two consecutive
mornings.TEM,TE(CV%),andICC95%CIwerecalculatedfromtheseduplicatemeasures(table
3.2).
Table3.2Repeatabilityofskinfoldmeasurements.Mean±SD.
Bicep Tricep Subscapular Supra-iliac
D1(mm) 6.8±2.3 7.0±3.3 10.0±2.5 10.5±2.7
D2(mm) 6.9±2.4 7.1±3.2 10.1±2.5 10.4±2.8
TEM 0.2 0.1 0.1 0.3
TE(CV%) 2.9 1.2 0.7 2.4
ICC(95%CI)0.99(0.98,0.99);
p≤0.001
0.99(0.99,1.00);
p≤0.01
1.00(0.99,1.00);
p≤0.001
0.99(0.98,0.99);
p≤0.001
Notes:ICC(95%CI),Intra-classcorrelationcoefficientwith95%confidenceintervals;SD,standarddeviation;
TEM,typicalerrorofmeasure;TE(CV%),typicalerrorasacoefficientofvariation.
3.6. Menstrualcycle
Timingoftesting
Tocontrolforhormonaleffectsonthermoregulationassociatedwiththemenstrualcycle,female
participantsfilledoutamenstrualcyclequestionnaire(Appendix3)for2monthspriortothetesting
sessionsbeingscheduled.Wherepossible,alltestingwasperformedduringthefollicularphaseof
the menstrual cycle (3-10 d after the onset of menstruation) (Stachenfeld and Taylor, 2014).
However, female participants taking oral contraceptive performed the experimental sessions
duringthenopillorinactivephaseoforalcontraceptiveusewherepossible.
Validityofthemenstrualcyclequestionnaire
Datacollectedinour laboratorydemonstratedthevalidityofmenstrualcyclequestionnairesfor
accuratelydetermining thecorrectphaseof themenstrual cycle.Nineparticipantscompleteda
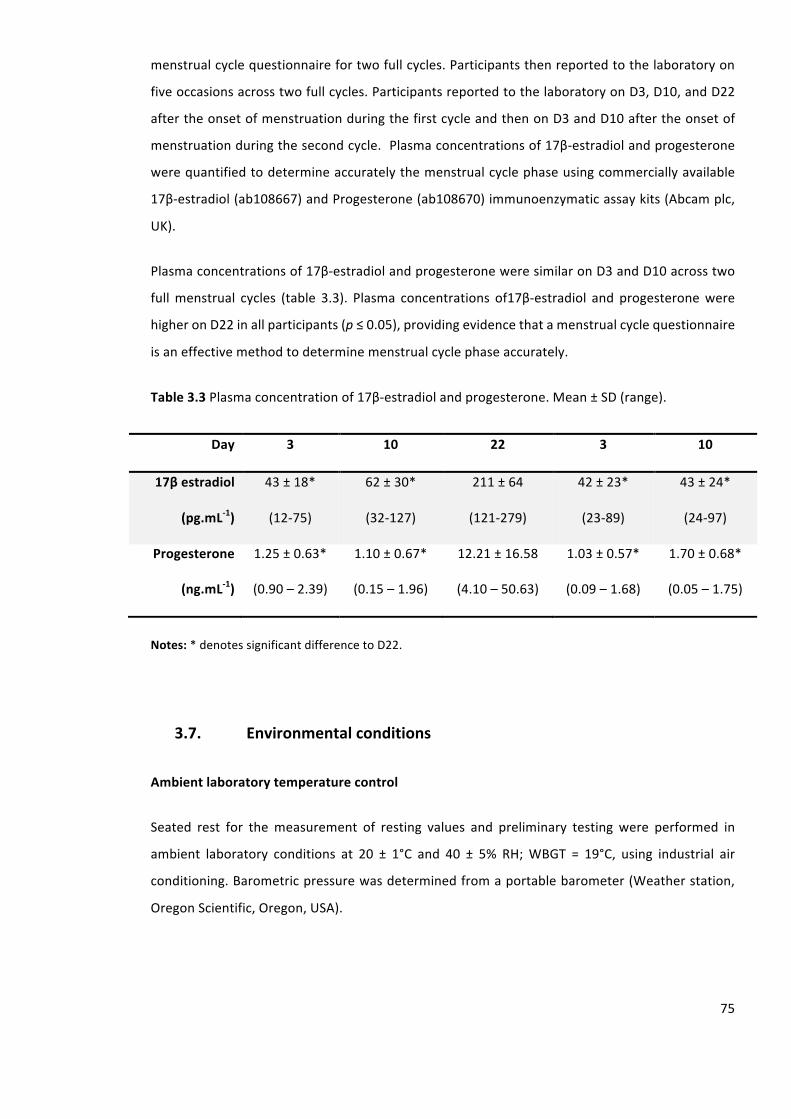
75
menstrualcyclequestionnairefortwofullcycles.Participantsthenreportedtothelaboratoryon
fiveoccasionsacrosstwofullcycles.ParticipantsreportedtothelaboratoryonD3,D10,andD22
aftertheonsetofmenstruationduringthefirstcycleandthenonD3andD10aftertheonsetof
menstruationduringthesecondcycle.Plasmaconcentrationsof17β-estradiolandprogesterone
werequantifiedtodetermineaccuratelythemenstrualcyclephaseusingcommerciallyavailable
17β-estradiol(ab108667)andProgesterone(ab108670)immunoenzymaticassaykits(Abcamplc,
UK).
Plasmaconcentrationsof17β-estradiolandprogesteroneweresimilaronD3andD10acrosstwo
fullmenstrual cycles (table 3.3). Plasma concentrations of17β-estradiol and progesteronewere
higheronD22inallparticipants(p≤0.05),providingevidencethatamenstrualcyclequestionnaire
isaneffectivemethodtodeterminemenstrualcyclephaseaccurately.
Table3.3Plasmaconcentrationof17β-estradiolandprogesterone.Mean±SD(range).
Day 3 10 22 3 10
17βestradiol
(pg.mL-1)
43±18*
(12-75)
62±30*
(32-127)
211±64
(121-279)
42±23*
(23-89)
43±24*
(24-97)
Progesterone
(ng.mL-1)
1.25±0.63*
(0.90–2.39)
1.10±0.67*
(0.15–1.96)
12.21±16.58
(4.10–50.63)
1.03±0.57*
(0.09–1.68)
1.70±0.68*
(0.05–1.75)
Notes:*denotessignificantdifferencetoD22.
3.7. Environmentalconditions
Ambientlaboratorytemperaturecontrol
Seated rest for themeasurement of resting values and preliminary testing were performed in
ambient laboratory conditions at 20 ± 1°C and 40 ± 5% RH;WBGT = 19°C, using industrial air
conditioning.Barometricpressurewasdeterminedfromaportablebarometer(Weatherstation,
OregonScientific,Oregon,USA).

76
Experimentaltemperatureandhumidity
Allexerciseheatsessionswereperformedinalarge,purpose-builtenvironmentalchamberwith
computer controlledmonitoring andmaintenance of desired environmental conditionswith an
availablerangeof-20°Cto+50°Cand20to95%RH.AsanindexofheatstressWBGTwascalculated.
Unlessotherwisestated,experimentaltrialswereconductedin40°Cand40%RH(WBGT=33°C).
Duringexerciseheat stressexperimental trials,manual recordingof thechamberconditionwas
performedevery5-mintodescribeaccuratelytheenvironmentexperiencedbyparticipants.
3.8. Thermoregulatorymeasures
Rectaltemperature
RectaltemperaturewasmeasuredtoindicateTc,usingageneralpurposedisposableprobe(Henley,
Reading,UK)attachedtoadatameter(MeasSpec4600,measurementSpecialities,VirginiaUSA)
measuredtotheaccuracyof0.01°C.Therectalprobewasmeasuredatadepthof10cmpastthe
analsphincterwithazincoxidetapebungappliedtotheprobepriorto insertiontoensurethe
correctdepthwasattainedandmaintainedthroughouttesting.
Repeatabilityofrectalthermistor
Therepeatabilityoftherectal thermistorwasdeterminedpriortoexercise inarestedstate.An
individualwasexposedtotemperatelaboratoryconditionsinarestedstatefor20-mintostabilise
Tre.MeasurementsofTrewerethentakeneveryminfortwo10-minperiods.TEM,TE(CV%),and
ICC95%CIwerecalculatedfromduplicatemeasures(table3.4).

77
Table3.4Repeatabilityofrectalthermistors.Mean±SD.
RestingTre(°C)
Measures 1 2
Mean±SD 37.4±0.0 37.4±0.02
Difference 0.02±0.02
TEM 0.02
TE(CV%) 0.04
ICC(95%CI) 0.99(0.98,1.00);p≤0.001
Notes: ICC, intra-classcorrelationcoefficient;TEM, typicalerrorofmeasure;TE (CV%), typicalerrorasa
coefficientofvariation;Tre,rectaltemperature.
Skintemperature
Tskwasrecordedusingskinthermistors(EltekLtd,Cambridge,UK)attachedtofoursitesontheright
sideofthebodyconnectedtoaSquirreltemperaturelogger(Squirrel1000series,EltekLtd.,UK).
Thefoursiteswerethechest(midpointofthepectoralismajo),thearm(midpointofthetriceps
brachii lateral head), the upper leg (midpoint of the rectus femoris), and the lower leg
(gastrocnemiuslateralhead)MeanTskwascalculatedusingEquation2.4(Ramanathan,1964).
Repeatabilityoftheskinthermistors
Therepeatabilityoftheskinthermistorwasdeterminedbyrecordingthemeantemperatureoffour
skinthermistorsexposedtoambientconditions.Theambientconditionswerestableat20°Cand
40%RHthroughoutthedatacollection.Theskinthermistorswerelefttostabilisefor20-minand
thenmeasurementswererecordedeveryminfortwo10-minperiods.Datawasthenaveragedover
each10-minperiod.TEM,TE(CV%),andICC95%CIwerecalculatedacrossthetwo10-minperiods
(table3.5).

78
Table3.5Repeatabilityofskinthermistors.Mean±SD.
Meanskinthermistor(°C)
Measures 1 2
Mean±SD 20.3±0.1 20.2±0.1
TEM 0.1
TE(CV%) 0.3
ICC(95%CI) 0.91(0.70,0.98);p≤0.001
Notes:ICC(95%CI),intra-classcorrelationcoefficientwith95%confidenceintervals;SD,standarddeviation;
TEM,typicalerrorofmeasure;TE(CV%)technicalerrorasacoefficientofvariation.
3.9. Cardiopulmonarymeasures
Metabolicgasanalysis
Expired metabolic gas was measured using a breath-by-breath online gas analysis system
(MetalyzerSport,Cortex,Germany).Calibrationwasperformedaspermanufacturer’sinstructions.
Calibration was performed for pressure, volume turbine and gas sensors prior to each use.
Barometric pressure was determined and calibrated to the device from a portable barometer
(GA690,CastleGroupLtd,UK).Volumecalibrationrequiredsimulatedinspirationandexpirationvia
amanualsyringe(3L,HansRudolph,Germany)forfiveacceptablecycleselicitingaflowrateof2
to4L.s-1.According to themanufacturer’s instructions, the flowsensormeasuredvalues to the
accuracyof±2%witharangeof0.05-20L.s-1.Gasconcentrationcalibrationrequiredtwoknown
O2andtwoknownCO2concentrations,knownasgas1andgas2,tobesampleduntilstabilisation
ofsensorshadbeenachieved.Accordingtothemanufacturer’sinstructions,theO2sensorandCO2
sensorsmeasured values to the accuracy of 0.1 Vol%with a range of 0 to 60% and 0 to 13%
respectively.
Inaccordancewithmanufacturer’sguidelines,gas1wasrequiredtorepresentclosetotheinspired
fractionandgas2closetotheexpiredfraction.Gas1wastakendirectlyfromambientconditions
(inspiredfractionO2=0.2093,inspiredfractionCO2=0.0005),Gas2wasdrawnfromacontained

79
gascylinder(expiredfractionO2=0.17,expiredfractionCO2=0.05)intoaclamp-sealedcollection
bag,whichwas immediatelyaffixed to thesample line. All respiratorygasexchangedatawere
averagedovera10speriod.
Heartrate
HeartratewasrecordedusingaPolarHRmonitor(PolarElectroOyo,Kempele,Finland).Achest
strap transmitter was securely affixed to the participant and the sensors dampened to aid
conductivity.ThewristwatchreceiverwasalsoworntoenableliveHRdatarecording.According
tothemanufacturer’sinstructions,theHRmonitorisaccurateduringsteadystateconditionsto±
1%or±1beat.min-1,whicheverislarger.
3.10. Physiologicalstrainindex
Thephysiologicalstrainindex(PSI)wascalculatedfromexercisingTreandHRusingEquation3.2.
Equation3.2Calculationofphysiologicalstrainindex(PSI)
PSI=5(Tret-Treo)*(39.5-Treo)-1+5(HRt-HRo)*(180-HRo)-1
Where:TreoandHRoarethe initialandTretandHRtaresimultaneousmeasurements takenatanytime
(Moranetal.,1998).
3.11. Haematologicalmeasures
Capillarylactatesample
Forcapillarylactatesamplingthefingertipwasfirstcleanedusinganalcoholwipeandlefttoairfor
approximately15s.Theskinwaspuncturedusingasingleuselancet(Acc-ChekSafe-T-Pro,Roche
Diagnosticsltd.,WestSussex,UK).Applyinglightpressuretothesitea~200µlofcapillaryblood
wascollectedfromthefingertipintoaheparin-fluoridecoatedmicrovettetubeandsubsequently
analysed for lactate concentration using an automated analyser (YSI 2300 Stat Plus, YSI UK,
Hampshire, UK). The analyser was calibrated immediately before each session using the
manufacturer’s 5 mmol.L-1 standard, set to self-calibrate every 25-min and verified after each
sessionusingthesamemanufacturerstandard.

80
Repeatabilityofcapillarybloodlactateanalysis
Arestingcapillarybloodsample,combinedwithsamplesatapproximately1mmol.L-1,2mmol.L-1,
3mmol.L-1,4mmol.L-1,and8mmol.L-1wereanalysedinduplicate.TEM,TE(CV%),andICC95%CI
werecalculatedfromduplicatemeasures(table3.6).
Table3.6Repeatabilityofcapillarybloodlactateanalysis.Mean±SD.
Lactateconcentration(mmol.L-1)
1 2
Mean±SD 3.30±2.66 3.32±2.69
TEM 0.03
TE(CV%) 1.02
ICC(95%CI) 1.00(0.99,1.00);p<0.001
Notes:ICC(95%CI),Intra-classcorrelationcoefficientwith95%confidenceintervals;SDstandarddeviation;
TEM,typicalerrorofmeasure;TE(CV%),technicalerrorasacoefficientofvariation.
3.12. Sweatrate
Towel-driednudeBMwasmeasuredandrecordedtothenearestgrammebeforeandimmediately
afteralltrialsasameasureofwholebodySWR.BetweenthesetwomeasuresofnudeBM,fluid
intakewas restricted and valueswere corrected for urine output. Valueswere uncorrected for
respiratoryandmetabolicweightlossesastheywereassumedassimilarbetweentrialsduetothe
matchedexerciseintensityandenvironmentalconditions.SWRwascalculatedandreporteding.hr-
1usingEquation3.3.SWRwasalsocalculatedandreportedrelativetoBSA(SWRBSA) ing.hr-1.m2
usingEquation3.4.
Equation3.3Calculationofwholebodysweatrate(SWR)
SWR(g.hr-1)=[BMpre(g)–(BMpost(g)+urineoutput(g)]/exerciseduration(min)*60
Where:BMpre=bodymasspriortobeginningthetrial,BMpost=bodymassfollowingthetrial.

81
Equation3.4Calculationofwholebodysweatraterelativetobodysurfacearea(SWRBSA)
SWRBSA(g.hr-1.m2)=SWR(g.hr-1)/BSA(m2)
Where:BSA,bodysurfacearea;SWR,sweatrate.
3.13. Perceptualscales
Ratingofperceivedexertion
The rating of perceived exertion scale (RPE), also known as the Borg scale (Borg, 1962), is a
subjectivetooltoassesssubjectiveperceptionofeffortduringexercise.RPErangedfrom6(very,
verylight),through13(somewhathard),to20(very,veryhard)alonga15-pointscale(figure3.1).
RPEhasbeenreportedtobecloselyrelatedtobothmetabolicandcardiac intensityparameters
(Scherr et al., 2013). Participants were provided with standardised instructions that provided
specificcommandsonhowtoreportoverallfeelingsofexertionthroughouttheexperimentaltrials.
Thestandardisedinstructionsincludedclearunderstandingofanchoringthetopandbottomratings
topreviousexperiencesofnoexertionatall(RPE=6)andmaximalexertion(RPE=20)(Maugeret
al.,2013).Withinthisthesis,RPEwasrecordedat10-minintervalsthroughouttheRHTTandHA
sessions.DuringthepreliminarylactatethresholdandVgO2peaktest,RPEwasrecordedinthefinal
15sofeachstage.
Thermalsensation
The thermal sensation (TS) scale (Toner et al., 1986) is a subjective tool to assess thermal
perception.TSranges from0.0 (unbearablycold), through4.0 (comfortable), to8.0 (unbearably
hot)alonga17-pointscale(figure3.1).TShasbeenreportedtobecloselyrelatedtomeasuresof
ambient air temperature,metabolic rate, Tre, and Tsk during exercise (Gagge et al., 1969). The
standardisedinstructionsprovidedtotheparticipantsincludedclearunderstandingofanchoring
the top and bottom ratings to previous experiences of being unbearably cold (TS = 0) and
unbearablyhot(TS=8).WithinthisthesisTSwasrecordedat10-minintervalsthroughouttheRHTT
andHAsessions.

82
Figure3.1Ratingofperceivedexertion(RPE)andthermalsensation(TS)scales.
3.14. Cyclingexercisetrials
Cycleergometry
All cycle ergometry was performed on a cycle ergometer (Monark 874E, Monark Exercise AB,
Vansbro,Sweden).Foralltrialstheseatheightwassettoensurea25°angleinthekneeflexion
whenat6o’clockpedalcrankpositionwasattained.Powerwascontrolledbymanipulatingthe
resistanceonthesuspendedweightpanusinga fixedcadenceof80revolutionspermin (rpm).
Calculationsofpoweroutputsinwatts(W)duringcyclingergometryweremadeasfollows.
Equation3.5Calculationofworkduringcycleergometry
Work(J)=Force(N)xDistance(m)
=Massontheweightpan(kg)xAccelerationduetogravity(m.s-1)xPedalrevolutions(r.s-
1)xFlywheeldistanceperrevolution(m)

83
Equation3.6Calculationofpowerfromworkduringcycleergometry
Power(W)=Work(J)/Time(s)
Cyclingpeakoxygenuptake
Agradedexercisetestwasperformedusingacycleergometerintemperatelaboratoryconditions
(WBGT=19°C)todetermineparticipants’VgO2peak.Participantsperformeda5-minwarmupat60
Wandwereinformedtomaintainaconstantcadenceof80rpm.Thecyclingintensitywassetto
80Wand resistancewasapplied to the flywheel toelicita16 to24W.min-1 increase (selected
depending on the BM of the participant). These intensities were selected to ensure volitional
exhaustionwas reached in8-15min. In the final10 sofeachstageHRwas recorded.VgO2,and
respiratoryexchangeratio(RER)weremeasuredcontinuouslyandaveragedforeach10speriod
throughouteachstage.Thetestwasterminatedwhenparticipantsreachedvolitionalexhaustion
and/or the cadence could no longer be maintained at 80 ± 5 rpm despite strong verbal
encouragement.3-minfollowingthecompletionofthetestacapillarybloodsamplewascollected
fortheanalysisoflactateconcentration.VgO2peakwasidentifiedatthepowerassociatedwiththe
highestVgO2averagedover10sec.VgO2peakwasdeterminedwhenthreeoutofthefollowingfour
criteriawhereachieved;aplateauinVgO2(increasebylessthan2mL.kg-1.min-1),anRER≥1.15,HR
reached 10 beats.min-1 from age predicted max and blood lactate ≥ 8 mmol.L-1. The protocol
adoptedwasaccordingtotheBritishAssociationofSportandExerciseScienceGuidelines(Davison
andWooles,2007).
Heatacclimationprotocol
ThedailyHAsessionsconsistedofa90-minexposure to40°C,40%RH (WBGT=33°C).Exercise
intensitywassetatcycling65%VgO2peak(Appendix4)fromtheoutsetandadjustedwithwork:
restintervalstoachieveandmaintainaTre~38.5°C(Garrettetal.,2012;Pattersonetal.,2004a),or
if participants were unable to maintain a cadence of 80 ± 5 rpm despite strong verbal
encouragementAn example of this protocol for one participant is presented in Figure 3.2. The
controlledhyperthermiamodelofHAwasselectedsinceitensuresaconstantadaptationstimulus
(Foxetal., 1963a), and is reported to result inperformanceandphysiological improvements in
highlytrainedathletes(Garrettetal.,2012).Acyclingmodeofexercisewasselectedsincethereis
lessvariabilityinefficiencycomparedwithrunningbetweenparticipantsthus,ensuringasimilar
stress.Furthermore,duetotheconsecutivenatureoftheHAsessions;cyclingincurslessmuscle
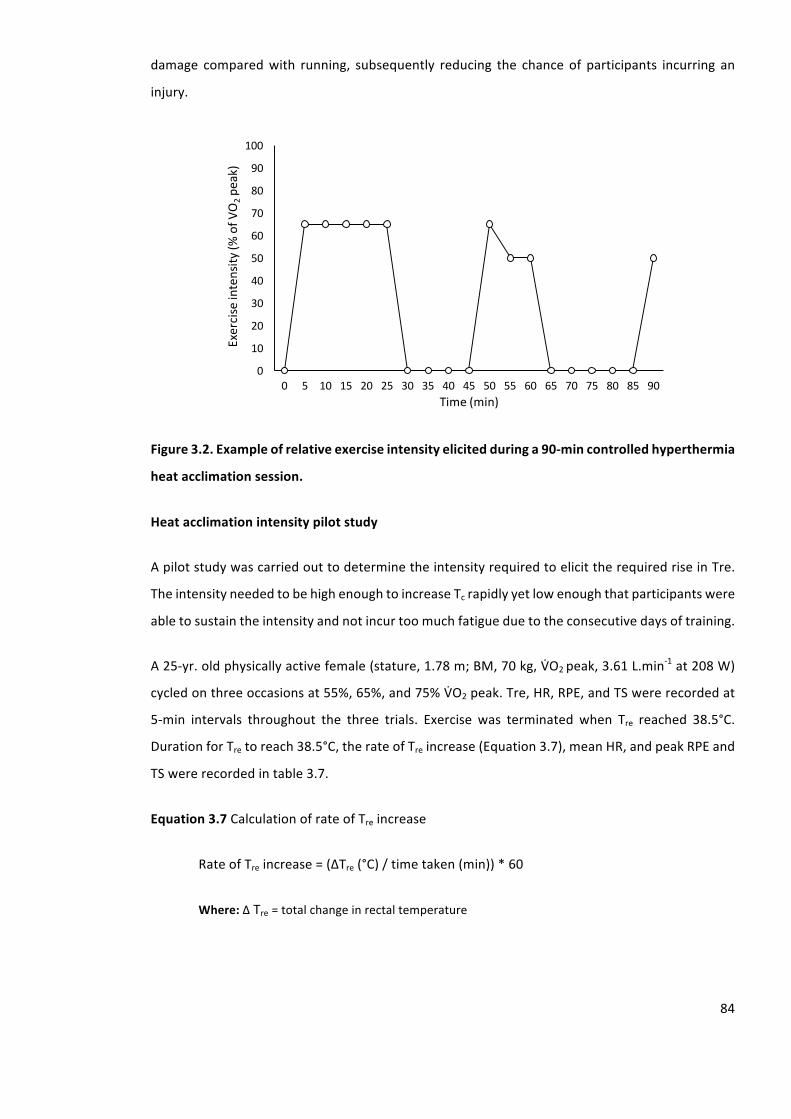
84
damage comparedwith running, subsequently reducing the chance of participants incurring an
injury.
Figure3.2.Exampleofrelativeexerciseintensityelicitedduringa90-mincontrolledhyperthermia
heatacclimationsession.
Heatacclimationintensitypilotstudy
ApilotstudywascarriedouttodeterminetheintensityrequiredtoelicittherequiredriseinTre.
TheintensityneededtobehighenoughtoincreaseTcrapidlyyetlowenoughthatparticipantswere
abletosustaintheintensityandnotincurtoomuchfatigueduetotheconsecutivedaysoftraining.
A25-yr.oldphysicallyactivefemale(stature,1.78m;BM,70kg,VgO2peak,3.61L.min-1at208W)
cycledonthreeoccasionsat55%,65%,and75%VgO2peak.Tre,HR,RPE,andTSwererecordedat
5-min intervals throughout the three trials. Exercise was terminated when Tre reached 38.5°C.
DurationforTretoreach38.5°C,therateofTreincrease(Equation3.7),meanHR,andpeakRPEand
TSwererecordedintable3.7.
Equation3.7CalculationofrateofTreincrease
RateofTreincrease=(∆Tre(°C)/timetaken(min))*60
Where:∆Tre=totalchangeinrectaltemperature
0
10
20
30
40
50
60
70
80
90
100
0 5 10 15 20 25 30 35 40 45 50 55 60 65 70 75 80 85 90
Exerciseintensity
(%ofV
O2pe
ak)
Time(min)
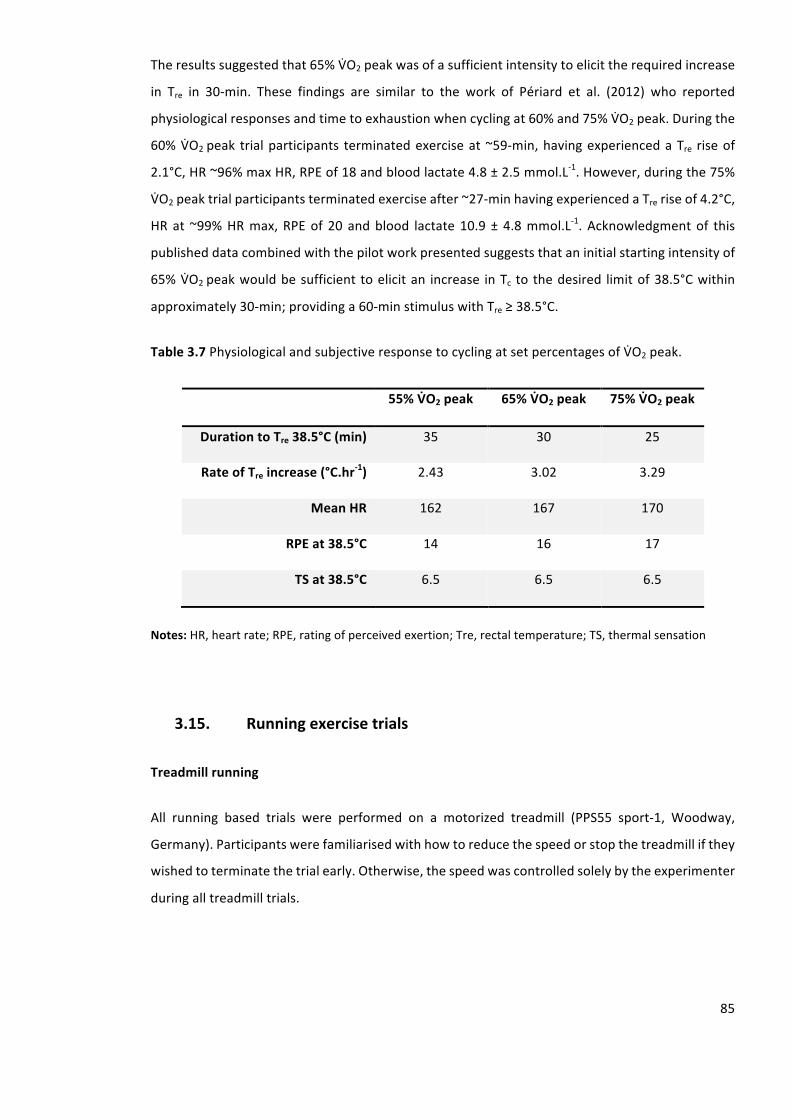
85
Theresultssuggestedthat65%VgO2peakwasofasufficientintensitytoelicittherequiredincrease
in Tre in 30-min. These findings are similar to the work of Périard et al. (2012) who reported
physiologicalresponsesandtimetoexhaustionwhencyclingat60%and75%VgO2peak.Duringthe
60%VgO2peak trialparticipants terminatedexerciseat~59-min,havingexperiencedaTre riseof
2.1°C,HR~96%maxHR,RPEof18andbloodlactate4.8±2.5mmol.L-1.However,duringthe75%
VgO2peaktrialparticipantsterminatedexerciseafter~27-minhavingexperiencedaTreriseof4.2°C,
HRat ~99%HRmax,RPEof 20 andblood lactate10.9±4.8mmol.L-1.Acknowledgmentof this
publisheddatacombinedwiththepilotworkpresentedsuggeststhataninitialstartingintensityof
65%VgO2peakwouldbesufficient toelicitan increase inTc tothedesired limitof38.5°Cwithin
approximately30-min;providinga60-minstimuluswithTre≥38.5°C.
Table3.7PhysiologicalandsubjectiveresponsetocyclingatsetpercentagesofVgO2peak.
55%VcO2peak 65%VcO2peak 75%VcO2peak
DurationtoTre38.5°C(min) 35 30 25
RateofTreincrease(°C.hr-1) 2.43 3.02 3.29
MeanHR 162 167 170
RPEat38.5°C 14 16 17
TSat38.5°C 6.5 6.5 6.5
Notes:HR,heartrate;RPE,ratingofperceivedexertion;Tre,rectaltemperature;TS,thermalsensation
3.15. Runningexercisetrials
Treadmillrunning
All running based trials were performed on a motorized treadmill (PPS55 sport-1, Woodway,
Germany).Participantswerefamiliarisedwithhowtoreducethespeedorstopthetreadmillifthey
wishedtoterminatethetrialearly.Otherwise,thespeedwascontrolledsolelybytheexperimenter
duringalltreadmilltrials.

86
Runninglactatethreshold
A graded exercise test was performed using a motorized treadmill in temperate laboratory
conditions(WBGT=19°C)todetermineparticipants’lactatethreshold.Participantsperformeda5-
minwarmupataspeed1km.hr-1belowthestartingspeed;thiswastypicallyaverylightjog(7
km.hr-1to9km.hr-1).Theinitialrunningspeedforthetestwassetbetween8km.hr-1and10km.hr-
1with a 1% gradient (Jones andDoust, 1996). Participants then performed five to nine, 3-min,
incremental (0.8 km.hr-1) stages on a treadmill. VgO2 and RER were recorded continuously
throughoutthestageandaveragedforeach10speriod.Inthefinal10sofeachstageHRandRPE
wererecorded.Oncompletionofeachstage,exercisewasmomentarilypausedandlactatesample
wastaken.Thetestwasterminatedwhenlactatereached4.0±0.5mmol.L-1.Lactatethresholdwas
determinedusingthepointatwhichbloodlactateincreased1mmol.L-1aboverestingvalue(Jones
and Doust, 1998). The protocol adoptedwas according to the British Association of Sport and
ExerciseScienceGuidelines(Jones,2007).
Runningpeakoxygenuptake
Followinga15-minrecoveryfromdeterminationofthelactatethreshold,participantsperformed
1-minincremental(1%gradient)stages.OnthecompletionofeachstageVgO2,RERandHRwere
recorded. The testwas terminatedwhen participants reached volitional exhaustion and/or the
speed of the treadmill could no longer be maintained despite strong verbal encouragement.
Immediatelypost and3-min following the completionof the test, a capillaryblood samplewas
collected for the analysis of lactate concentration. VgO2 peak was identified at the intensity
associatedwiththehighestVgO2averagedover10s.VgO2peakwasdeterminedwhenthreeoutof
thefollowingfourcriteriawhereachieved;aplateauinVgO2(increaseof≤2mL.kg-1.min-1),anRER
≥1.15,HRreached10beats.min-1fromagepredictedmax,andbloodlactate≥8mmol.L-1.
3.16. Heattolerancetests
Runningheattolerancetest
TheRHTTwasperformedonamotorizedtreadmillandconsistedof30-minrunningat9km.hr-1
and2%gradient inambientconditionsof40°Cand40%RH(WBGT=33°C).Following20-minof
seatedrest intemperatureconditions,Tre,HR,Tsk,RPE,andTSwererecorded.Participantsthen
enteredtheenvironmentalchambertoperformtheRHTT;whereTre,HR,andTskwererecordedat
5-min intervals and RPE and TS at 10-min intervals. Towel-dried nude BM was measured and
recordedtothenearestgrammebeforeandafteralltrialsasameasureofSWR.

87
Runningheattolerancetestpilotwork
Introduction
TheIsraeliDefenceForce(IDF)developedawalkingHTTtoevaluateifmilitarypersonnel,whohad
experiencedexertionalheatillness,weresafetoreturntoduty.Theprotocolconsistsof120-min
walkingonatreadmillatapaceof5km.hr-1anda2%gradientinambientconditionsof40°Cand
40%RH(WBGT=33°C).At theendof theexposure,heattolerancewasdefinedasapeakTre≤
38.0°C,peakHR≤120beats.min-1,andSWR≥780g.h-1.Heatintolerancewasdefinedas,peakTre
>38.5°C,peakHR>145beats.min-1,(Moranetal.,2007)andapeakPSI≥6(Moranetal.,2004).
There is an instant elevation in the rate of thermogenesis at the onset of physical activity. As
exerciseintensityincreases,especiallyunderhotenvironmentalconditions,athermalimbalance
persistsresultinginacontinuallypositiverateofchangeinbodyheatstorage,increasingbodyheat
contentandasustainedriseinTcgivingagradedincreaseofheatstrain(Jay&Kenny,2007).There
areseverallimitationsassociatedwiththeIDFHTT,whicharemoreapparentwhenconductingthe
testonenduranceathletes(Johnsonetal.,2013).Endurancerunnerstypicallyrunathighintensities
for shorter durations than military personnel. The intensity of the protocol of the IDF HTT is
considerablybelowwhatanathletewouldtrainandcompete.Thus,theIDFHTTmisrepresentsthe
metabolicheatproductionenduranceathletesmayexperienceandpotentiallymaymisdiagnose
theirsusceptibilitytoahyperthermicstate.
Theaimofthispilotstudywastodeterminewhethertherewasasimilarpatternofheattolerance
andheatintolerancebetweentheIDFHTTandtheRHTT.ItwashypothesizedthattheRHTTwould
have a strong positive relationship for the participants’ thermoregulatory and cardiovascular
measuresduringtheIDFHTTandtheRHTT.
Materialsandmethods
Sixteen (8 males; 8 females) healthy individuals volunteered and provided written informed
consenttoparticipateinthepilotstudy(mean±standarddeviation(SD),age,23±5yrs.;BM,67.07
±10.96kg;stature,1.76±0.01m).
Participantsperformedtwotrialsinarandomisedorderseparatedby5-7d.TheIDFHTTinvolved
120-minwalkingat5km.hr-1and2%gradientinhotambientconditionsof40°Cand40%RH(WBGT
= 33°C). The RHTT was conducted using the previously described procedure. Nude BM was
88
measuredpre-andpost-trialasameasureofSWR.At5-minintervalsthroughoutthetrials,HRand
Trewererecorded.
AlldatawerefirstcheckedfornormalityusingtheShapiro-Wilkmethod.Asameasureofretest
correlation,ICCwith95%CIwerecalculatedforeachphysiologicalmeasureduringIDFHTTandthe
RHTT.Pairedsampledttestswerecalculatedtoidentifyanydifferencesinphysiologicalmeasures
betweentheIDFHTTandtheRHTT.Alldatawasanalysedusingastandardstatisticalpackage(SPSS
version20.0),andreportedasmean±SD.Statisticalsignificancewasacceptedatthelevelofp≤
0.05.
Results
Table3.8presentsthemean±SDdataforthephysiologicalvariablesmeasuredduringtheIDFHTT
andtheRHTT.TherewerenoobserveddifferencesbetweentheIDFHTTandtheRHTTinmeasures
ofTre rest (t (15)= -0.597,p=0.559)andHRrest (t (15)=1.370,p=0.191).However, therewere
significantdifferencesbetweentheIDFHTTandtheRHTTinmeasuresofpeakTre(t(15)=-5.567,p
≤0.001),peakHR,(t(15)=-22.753,p≤0.001)andSWR(t(15)=-8.180,p≤0.001).
Table3.8PhysiologyvariablesmeasuredduringtheIDFHTTandtheRHTT.Mean±SD.
PhysiologicalVariables IDFHTT RHTT
Trerest(°C) 37.17±0.26 37.18±0.26
Trepeak(°C) 38.30±0.28 38.82±0.47*
HRrest(beats.min-1) 65±8 64±8
HRpeak(beats.min-1) 135±18 182±15*
SWR(g.hr-1) 637±188 1721±675*
Notes:HR,heartrate;IDFHTT,IsraeliDefenceForceheattolerancetest;RHTT,runningheattolerancetest;
Tre,rectaltemperature;andSWR,sweatrate.*denotessignificantlydifferencetoIDFHTT(p≤0.05).
Figure3.3presentstherawdataforTrerestandHRrest.Strongcorrelations(ICC(95%CI))were
observedbetweentheIDFHTTandtheRHTTforTrerest(r=0.938(0.822-0.978),p≤0.001)andHRrest
(r)=0.959(0.881-0.986),p≤0.001).Figure3.4presentstherawdataforTrepeakandHRpeak.Strong
correlations(ICC(95%CI))wereobservedbetweentheIDFHTTandtheRHTTforTrepeak(r=0.702
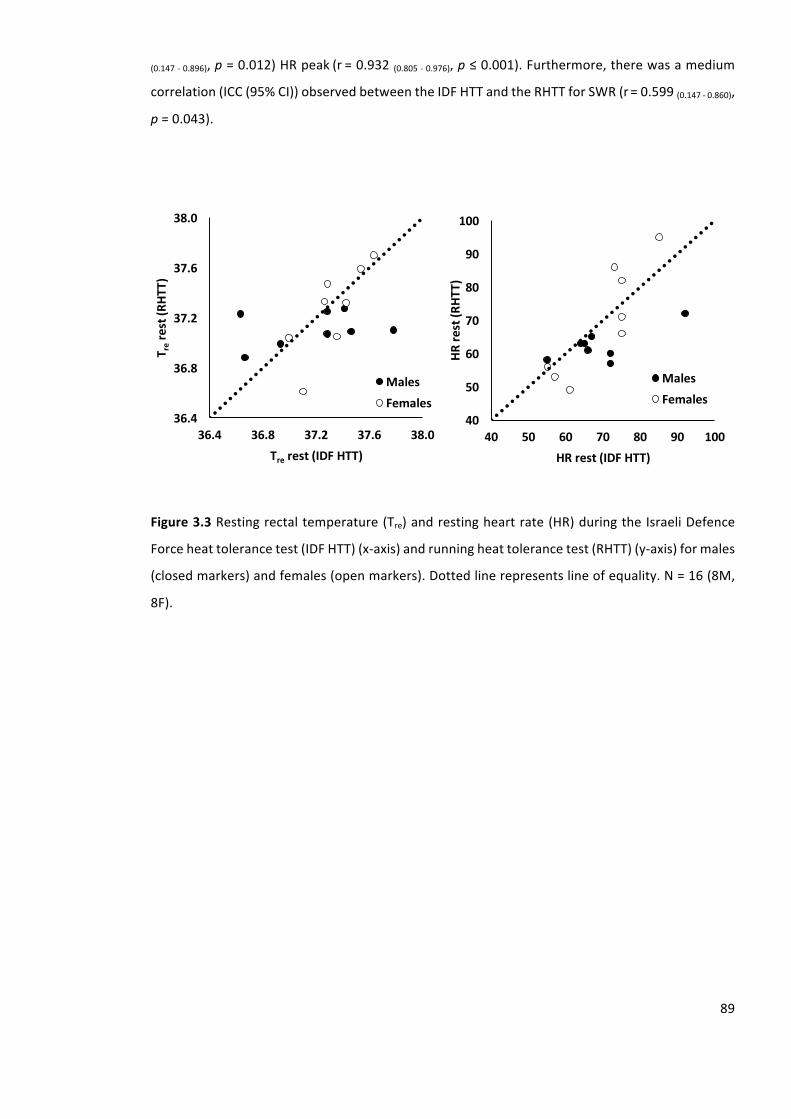
89
(0.147-0.896),p=0.012)HRpeak(r=0.932(0.805-0.976),p≤0.001).Furthermore,therewasamedium
correlation(ICC(95%CI))observedbetweentheIDFHTTandtheRHTTforSWR(r=0.599(0.147-0.860),
p=0.043).
Figure3.3Restingrectaltemperature(Tre)andrestingheartrate(HR)duringtheIsraeliDefence
Forceheattolerancetest(IDFHTT)(x-axis)andrunningheattolerancetest(RHTT)(y-axis)formales
(closedmarkers)andfemales(openmarkers).Dottedlinerepresentslineofequality.N=16(8M,
8F).
40
50
60
70
80
90
100
40 50 60 70 80 90 100HR
rest(R
HTT)
HRrest(IDFHTT)
MalesFemales
36.4
36.8
37.2
37.6
38.0
36.4 36.8 37.2 37.6 38.0
T rerest(R
HTT)
Tre rest(IDFHTT)
MalesFemales
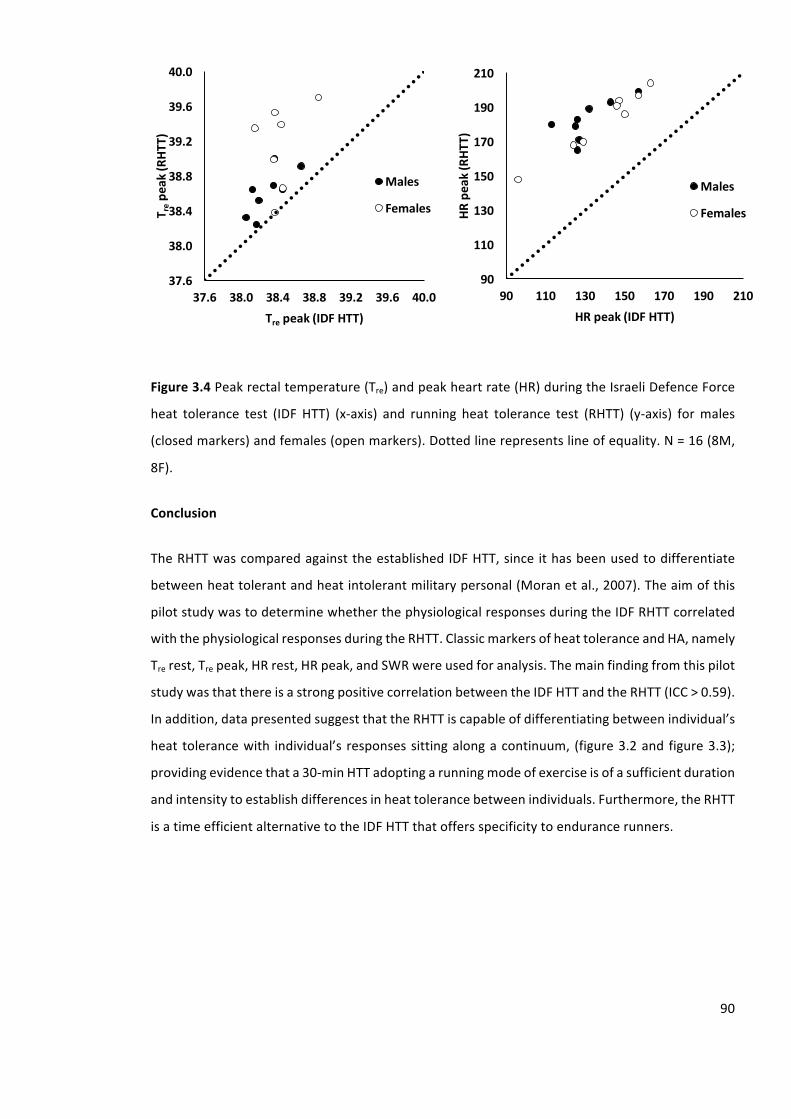
90
Figure3.4Peakrectaltemperature(Tre)andpeakheartrate(HR)duringtheIsraeliDefenceForce
heat tolerance test (IDF HTT) (x-axis) and running heat tolerance test (RHTT) (y-axis) formales
(closedmarkers)andfemales(openmarkers).Dottedlinerepresentslineofequality.N=16(8M,
8F).
Conclusion
TheRHTTwascomparedagainsttheestablishedIDFHTT,sinceithasbeenusedtodifferentiate
betweenheattolerantandheatintolerantmilitarypersonal(Moranetal.,2007).Theaimofthis
pilotstudywastodeterminewhetherthephysiologicalresponsesduringtheIDFRHTTcorrelated
withthephysiologicalresponsesduringtheRHTT.ClassicmarkersofheattoleranceandHA,namely
Trerest,Trepeak,HRrest,HRpeak,andSWRwereusedforanalysis.Themainfindingfromthispilot
studywasthatthereisastrongpositivecorrelationbetweentheIDFHTTandtheRHTT(ICC>0.59).
Inaddition,datapresentedsuggestthattheRHTTiscapableofdifferentiatingbetweenindividual’s
heat tolerancewith individual’s responsessittingalongacontinuum, (figure3.2and figure3.3);
providingevidencethata30-minHTTadoptingarunningmodeofexerciseisofasufficientduration
andintensitytoestablishdifferencesinheattolerancebetweenindividuals.Furthermore,theRHTT
isatimeefficientalternativetotheIDFHTTthatoffersspecificitytoendurancerunners.
37.6
38.0
38.4
38.8
39.2
39.6
40.0
37.6 38.0 38.4 38.8 39.2 39.6 40.0
T repe
ak(R
HTT)
Tre peak(IDFHTT)
Males
Females
90
110
130
150
170
190
210
90 110 130 150 170 190 210
HRpeak(RHT
T)
HRpeak(IDFHTT)
Males
Females

91
3.17. Statisticalanalysis
3.17.1. Poweranalysis
ThenumberofparticipantsrequiredforeachexperimentalchapterwasdeterminedusingG*Power
version3.1inaccordancewithestablishedguidelinesforaprioridetermination(Farrokhyaretal.,
2013)Thevalueofαandβweresetas0.05and0.8,respectively.
3.17.2. Normalityandsphericityofdata
Normaldistribution
Datawascheckedforskewnessandkurtosistodeterminewhetherthedistributionofscoreswas
approximatelynormal.Ifvaluesofskewnessandkurtosiswerebetween-1.96and1.96datawas
assumednormallydistributed(Field,2013).
Furthermore, the Shapiro-Wilk method was used to compare the scores in the data set to a
normallydistributedsetofdatawithanidenticalmeanandSD.Ifthetestwasnon-significant(i.e.
p≥0.05),thenthedistributionwasdeemednotsignificantlydifferentfromthenormaldistribution
anddatawasassumednormallydistributed(Field,2013).
Sphericity
SphericitywasassessedusingtheMauchly’stest,whichteststhehypothesisthatthevarianceof
thedifferencesbetweenconditionsisequal.IfMauchly’steststatisticwassignificant(p≤0.05),it
wasconcludedthat thereweresignificantdifferencesbetweenthevariancesofdifferencesand
therefore,theconditionofsphericitywasnotmet.Iftheassumptionofsphericitywasnotmetthe
degreesoffreedomwereadjustedusingtheGreenhouse-Geissermethod.IfMauchly’steststatistic
wasnon-significant(p≥0.05)thenitwasconcludedthatthevarianceofdifferenceswereabout
equal(Field,2013).
3.17.3. Repeatabilitystatistics
Typicalerrorofmeasure(TEM)
WithinthisthesistheTEMwascalculatedtorepresenttheabsoluterepeatabilityofmeasurements
used.TEMwascalculatedfromtheSDofthedifferencescorebythesquarerootoftwo(Hopkins,
2000a).

92
Equation3.8Calculationoftypicalerrorofthemeasure(TEM)
TEM=SDdiff/√2.
Where:SDdiff=standarddeviationofthedifferencebetweentherepeatedmeasures;√2=square
rootof2.
Coefficientofvariation
WithinthisthesistheTEMhasbeenexpressedasapercentageofitsrespectivemeantoformthe
technicalerrorasacoefficientofvariation(TE(CV%))(Hopkins,2000a).
Equation3.9Calculationoftechnicalerrorasacoefficientofvariation(TE(CV%))
TE(CV%)=(TEM/Mean)*100
Where:TEM=typicalerrorofmeasure,Mean=themeanofthemeanofthepairedsamples
Intraclasscorrelationcoefficient
WithinthisthesisICCwith95%CIwerecalculatedusingSPSSasameasureofretestcorrelation.
Priortoconductingthistestdatawasfirstcheckedfornormalityaspreviouslydescribed.
Equation3.10Calculationofintraclasscorrelation
ICC=(SD2–sd2)/SD2
Where:SD=betweensubjectstandarddeviation,sd=withinsubjectstandarddeviation
3.17.4. Statisticaltestsofdifference
Statisticalsignificancelevel
Withinthisthesisastatisticalsignificancelevelwassettodescribetheprobability(p)ofseeinga
differenceasbig(orbigger)thantheoneyousaw,giventhatthereisnodifference.Thethreshold

93
pvalueforallexperimentswithinthisthesiswassetat5%thus,giving95%confidencethatthere
wasadifferenceorassociation.Withinthisthesis,apvalue≤0.05indicatesthattheobservedeffect
isunlikelytohavearisenpurelybychance,providingevidencetorejectthenullhypothesis.
TypeIandtypeIIerror
Astatisticalsignificancelevelof0.05wasselectedwithinthisthesistoavoidthechancesoftypeI
and type IIerror.Type Ierroroccurswhenyou incorrectly reject thenullhypothesis.Thus,you
wouldsuggestarelationshipordifference,wheninrealitynoneexists.Thistypicallyoccurswhen
thestatisticalsignificancelevelistoohigh(e.g.p≤0.1).Incontrast,typeIIerroroccurswhenyou
accept the null hypothesis when it is false. Thus, you would suggest no relationship, when a
relationshipexists.Thistypicallyoccurswhenthestatisticalsignificancelevel istoolow(e.g.p≤
0.01).
Pairedsamplesttest
Withinthisthesispairedsamplettestswereusedtodeterminestatisticalsignificancebetweentwo
experimentalconditions,whenthesameparticipantswereusedduringbothconditions.Theratio
ofthesevariancesisknownasthet-ratio.Priortoconductingthettest,datawerefirstcheckedfor
skewnessandkurtosistodeterminewhether
Analysisofvariance(ANOVA)
Analysisofvariance(ANOVA)isawayofcomparingtheratioofsystematicvariancetounsystematic
varianceinanexperimentalstudy.TheratioofthesevariancesisknownastheF-ratio.ANOVAisa
collection of statistical models used to analyse the difference among group means and their
associatedprocedures.PriortoconductinganANOVAwithintherespectivestudiesdatawasfirst
checkedfortheassumptionofnormaldistributionandsphericityaspreviouslydescribed(3.17.2.).
3.17.5. Effectsizes
Withinthisthesis,effectsizeshavebeenusedtopresentthemagnitudeofthereportedeffectsin
a standardised metric, providing information on the practical significance of the findings. The
selectedeffect size reported for each statistical test,wasbasedon the recommendations from
Lakens'(2013)reviewoncalculatingandreportingeffectsizes.

94
Cohen’sd
Inthisthesis,effectsizesforttestshavebeenreportedasCohen’sd.Cohen’sdhasbeenusedto
describethestandardisedmeandifferenceofaneffect.ACohen’sdvalueof0.2referstoasmall
effect,0.5asamediumeffectand0.8asalargeeffectbasedonbenchmarkssuggestedby(Cohen,
1988).Cohen’sdinbetweensubjects’designcanbereadilyinterpretedasapercentageoftheSD
suchthataCohen’sdof0.5meansthedifferenceequalstohalfaSD.
Equation3.11CalculationofCohen’sd(d)
d=(X1–X2)/SD
Where:X1=meanfirstsetofdata,X2=meansecondsetofdata,SD=standarddeviation
Partialetasquared
In this thesis, effect sizes forANOVAanalyses, havebeen reported as partial eta squared (np2)
(LevineandHullett,2002).ThiswascalculatedbySPSS.Partialetasquaredexpressesthesumof
squaresoftheeffectinrelationtothesumofsquaresoftheeffectandthesumofsquaresofthe
errorassociatedwiththeeffect.
3.17.6. Posthocanalysis
Tocombatthefamilywiseerrorrateassociatedwithmultiplecomparisons,thelevelofsignificance
forindividualtestswasalteredusingBonferronicorrections.TheBonferronimethodswasselected
withinthisthesisasitisextremelyrobust(althoughslightlyconservative)andcontrolsthealpha
levelregardlessofthemanipulation.IndoingthisitensuresthatthecumulativetypeIerrorremains
below0.05(Field,2013).
Anyadditionalstatisticalanalysesusedwiththisthesisareoutlineintherespectivechapter.

95
4. Repeatabilityofarunningheattolerancetest
________________________________________________________________________________
4.1. Abstract
AtpresentthereisnostandardisedHTTprocedurethatusesrunningasamodeofexercise.The
current study aimed to establish the repeatability of a practical running test to evaluate an
individual’s ability to tolerate exercise heat stress. Sixteen (8M, 8F) participants performed the
RHTT(30-min,9km.hr-1,2%elevation)ontwoseparateoccasionsinahotenvironment(40°Cand
40%RH).TherewerenodifferencesinpeakTre(RHTT1:38.82±0.47°C,RHTT2:38.86±0.49°C,r=
0.93,TEM=0.13°C),peakTsk(RHTT1:38.12±0.45°C,RHTT2:38.11±0.45°C,r=0.79,TEM=0.30°C),
peakHR(RHTT1:182±15beats.min-1,RHTT2:183±15beats.min-1,r=0.99,TEM=2beats.min-1),
orSWR(RHTT1:1,721±675g.hr-1,RHTT2:1,716±745g.hr-1,r=0.95,TEM=162g.hr-1)between
RHTT1andRHTT2(p≥0.05).Resultsdemonstrategoodagreement,strongcorrelationsandsmall
differencesbetweenrepeatedtrials,andtheTEMvaluessuggestlowwithin-participantvariability.
TheRHTTwaseffectiveindifferentiatingbetweenindividuals’physiologicalresponses;supporting
aheattolerancecontinuum.ThefindingssuggesttheRHTTisarepeatablemeasureofphysiological
strainintheheatandmaybeusedtoassesstheeffectivenessofacuteandchronicheat-alleviating
procedures,whenusingarepeatedmeasuresdesign.
4.2. Introduction
Duringexercise inahotenvironment,activemusclesperformworkcausingan increase inbody
heatcontent.Thesechangesaremodulatedbytherateofrelativeheatproduction(Cramerand
Jay,2014),andrepresenttherateofchangeinbodyheatstorage,whichinturnreflectsthebalance
betweenmetabolicheatproduction,heatabsorbedfromtheenvironmentandtotalbodyheatloss
(Jay and Kenny, 2007). Individuals vary in their ability to withstand heat stress, with some
demonstratingadecreasedcapabilitytodissipateheatandgreaterriseinbodyheatcontentunder
the same exercise heat stress (Epstein, 1990). These individuals have been described as heat
intolerant and are often characterized by an earlier and greater rise in Tc, a greater storage of
metabolicheat,ahigherphysiologicalstraintomoderateintensityexerciseintheheat,andreduced
sweatingsensitivity(Epsteinetal.,1983;Moranetal.,2004).
Anindividual’sheatintolerantstatemaybetemporaryorpermanent(Epstein,1990;Moranetal.,
2007;Ruelletal.,2014),stemmingfromtransientpredisposingfactors,suchasanacuteinjuryto

96
thethermoregulatorycentre,insufficientHA,dehydrationorinfectiousdisease(Epstein,1990).In
addition,alastingthermoregulatorydysfunctionmaystemfromconditionssuchascardiacdisease,
impairmenttosweatglands(Epstein,1990),ordifferencesingeneexpression(Moranetal.,2006).
Congenital factors such as ectodermal dysplasiamay also compromise heat tolerance in some
individuals(Epstein,1990).Asidefromthesepredisposingfactors,thehighexerciseintensitythat
endurancerunnersexperienceduringcompetitionscombinedwithextremeambientconditions,
mayelicitunavoidableuncompensableheatproduction.Theevaporativeheatlossrequirementto
maintainathermalsteadystateexceedsthemaximalevaporativecapacityoftheindividualinthe
given environment causing a continual rise in Tc. Thework by Nielsen (1996) provides data to
suggestamarathonrunnermayexperienceuptoa1.0°CriseinTcevery~9-minwhenracinginhigh
ambientconditions(≥35°C,≥60%RH),whenradiantandconvectiveheatlossisnegligible.This
rate of rise in Tcwould result in the runner reaching a Tc of 40.0°Cwithin 25-30min,with the
immediatedangersofheatexhaustion.Highincidenceofexertionalheatillnesshasbeenreported
inlongdistancerunners,with31%and53%ofthetotalnumberofcasesofexertionalheatillness
during the 1992 New Orleans U.S. Olympic Trials and the 1996 Atlanta Olympics, respectively
(Martin,1997).Whetherheatintoleranceispermanentoracquired,theconsequencesofexertional
heatillnessamongenduranceathletesemphasisesthevalueofatesttoevaluateanindividual’s
abilitytowithstandexerciseheatstress.
ExperimentalprocedureshavebeenusedtocauseariseinTcunderrestingandexerciseconditions
tochallengethethermoregulatoryresponses(Inoueetal.,2005;Johnsonetal.,2013;Kenneyand
Hodgson,1987;Montainetal.,1994)andthusassesstheabilityofanindividualtowithstandheat
stressandtoevaluateheatdissipatingmechanisms.TheIsraeliDefenceForce(IDF)developeda
HTTtoevaluatewhetherexertionalheatillnessinmilitarypersonnelwastemporaryorpermanent,
supportingasafereturntoduty(Moranetal.,2004).Theprotocolinvolves120-minwalkingona
treadmillatapaceof5km.hr-1anda2%gradientinhotambientconditionsof40°Cand40%RH
(WBGT=33°C).Heattoleranceisdeterminedattheendoftheexposure,usingcriteriaofpeakTre
≤38.0°C,peakHR≤120beats.min-1,andSWR≥780g.h-1.Deviationsfromthespecifiedcriteria
indicateagreaterstateofheatintolerance,whereasapronouncedplateauinbothTreandHRisa
definitivesignofheattolerance.

97
There is an instant elevation in the rate of thermogenesis at the onset of physical activity. As
exercise intensity increases,especially inanuncompensable4environment,a thermal imbalance
persists.Thisresultsinacontinuallypositiverateofchangeinbodyheatstorage,increasingbody
heatcontentandasustainedriseinTc,givingagradedincreaseofheatstrain(JayandKenny,2007).
TheIDFHTTmaybeappropriateforspecificoccupationalsituationsduetothelow-to-moderate
intensitycoupledwiththelongexposuretimethatislikelytobeexperiencedinmilitaryscenarios.
AcknowledgingtheworkcarriedoutbytheIDF,limitationsassociatedwiththeHTTremainwhen
examiningendurancerunners.Exertionalheatillnessiscompoundedbyuncompensableheatstress
whichisinturn,influencedbythedurationandintensityofexercise.Therelativeworkintensityof
an endurance runner training and competing in the heat is markedly higher compared with
occupationalactivities.Therefore,theIDFHTTmaynotbeapplicabletoanendurancepopulation
duetothedurationandintensityoftheprotocol.ThelowintensitynatureofthecurrentHTTwill
not reflect themetabolic heat production endurance runners experience andmaymisdiagnose
theirsusceptibilitytoahyperthermicstate,pointingtothebenefitofaRHTT.Atpresentthereisno
standardised HTT procedure adopting a runningmode of exercise and such a test would offer
greaterecologicalvalidityforendurancerunners.
Moran and colleagues (2004) assessed the heat tolerance of nineteen male participants and
concludedthatthedurationofaHTTcannotbeshorterthan120-min,sincetoleranceat60-min
wasunabletopredicttoleranceat120-min.TheworkbyEpsteinandcolleagues(1983)andmore
recentlybyMoranandcolleagues(2007)contradictsthesefindings,astherateofincreaseinTre
andHRduringthefirst20-30minwasconsiderablydifferentbetweenthoseindividualsdeemed
heatintolerantandthoseheattolerant.Thisevidencesuggeststhatitmaybeplausibletoassess
anindividual’sabilitytowithstandexerciseheatstressin30-min.AshorterRHTTrequiringnoprior
testingwouldprovideamoretimeefficientscreeningprocedureforrunners.
To assess and monitor changes in heat tolerance a protocol needs to be reliable to minimise
measurementerror,duetobiologicalvariationandequipmentnoise(AtkinsonandNevill,1998).
Typically,theassessmentofrepeatabilityusesperformancemarkersmoreoftenthanphysiological
markers,andespeciallythermoregulatorymarkers.Whentherepeatabilityofphysiologicalmarkers
has been assessed, it has often been between two pieces of equipment measuring the same
4 Compensableheat stress refers to a steady state Tc,whereby theheat lossmatches theheatproduction. Uncompensable heat stress refers to an imbalance between the evaporativerequirementandtheevaporativecapacityofanindividual.

98
physiological variable. Consequently, there is limited evidence comparing physiologicalmarkers
during repeated trials using a set intensity exercise protocol. The repeatability of mean aural
temperatureandmeanHRduringafixedintensitycyclingheatstresstest,whichinvolvedthree20-
mincycleboutsseparatedby8-minrestwasassessedinadolescents(Brokenshireetal.,2009).ICC
of0.58and0.95formeanauraltemperatureandmeanHR,respectivelyandaTE(CV%)of0.1%
and3%formeanauraltemperatureandmeanHR,respectivelywerereportedandareindicativeof
strongmeasurementrepeatability.Determiningtherepeatabilityofphysiologicalmeasuresduring
aHTTwouldprovideconfidenceintervalsandallowthetesttobeusedtodetermineachangein
heattolerancefollowingacuteorchronicheat-alleviatinginterventions.Thus,itmayenablespecific
guidanceonpreparationrequiredpriortotrainingorcompetinginhighambientconditions.
WhileacknowledgingthelimitationsoftheIDFHTT,pilotdata(3.16.)establishedsimilarpatterns
ofheattoleranceandheatintoleranceprovidingsupporttoexaminetherepeatabilityoftheRHTT.
Thisstudyaimedtoestablishtherepeatabilityofapracticalrunningtesttoevaluateindividuals’
heattolerance.TheprimarilyhypothesiswasthatphysiologicalresponsesduringtheRHTTwillhave
astrongpositivecorrelationonrepeatedtrials.Thesecondaryhypothesiswasthattherewillbeno
differences inphysiological responsesduring the runningheat tolerance testbetween repeated
trials.
4.3. Materialsandmethods
Participants
Sixteen(8males;8females)athletes(3.2.)volunteeredandprovidedwritteninformedconsentto
participateinthestudy(mean±SD;age,23±5yrs.;BM,67.07±10.96kg;stature,1.76±0.10m;
BSA,1.82±0.19m2;sumoffourskinfolds,43±15mm;speedatlactatethreshold,11.7±1.8km.hr-
1;andrunning,VgO2peak49±7mL.kg-1.min-1).ThestudywasapprovedbytheUniversityofBrighton
Research Ethics & Governance Committee and conducted in line with the health and safety
guidelines(3.1.)andinaccordancewiththeguidelinesoftherevisedDeclarationofHelsinki,2013.
Preliminarytesting
During the first visit to the laboratory standardised anthropometric assessmentwas conducted
(3.5.)followedbyagradedexercisetesttodetermineparticipants’lactatethresholdandVgO2peak
(3.15.).Breath-by-breathexpiredairwasmeasuredusingonlinegasanalysisthroughouttheRHTT
(3.9.).Acapillary lactatesample(3.11.)wastakeoncompletionofeachstageduringthelactate
thresholdtestandoncompletionoftheVgO2peaktest.

99
Runningheattolerancetest
All trials were conducted with pre-trial diet and exercise standardisation (3.4.). Participants
providedaurinesampleonarrivaltoensureadequatehydration.Thetimingoftestingforfemale
participants was conducted in accordance with the menstrual cycle standardisation guidelines
(3.6.). Fiveparticipantswereusingoral contraceptive. twousedYasmin (30µgethinylestradiol/
3mgdrospirenone)andthreeusedMicrogynon(30µgethinylestradiol/150µglevonorgestrel).All
testingwasperformedonamotorisedtreadmill(3.15.).
TheRHTT (3.16.)wasperformedon twooccasions. Following a 20-min stabilisationperiod in a
temperateenvironment(3.7.),measuresofTre(3.8.),HR(3.9.),Tsk(3.8.),RPE,andTS(3.13.)were
recorded,andparticipantsenteredtheenvironmentalchamber(3.7.).ToweldriednudeBMwas
alsorecordedtocalculateSWR(3.12.).ThroughouttheRHTTTre,HR,andTskwererecordedat5-
minintervals.RPEandTSwererecordedat10-minintervals.Exercisewasterminatedinlinewith
universityguidelines(3.1.).Participantsrepeatedtheprocessbetween5-7dlatertoassessthe
repeatabilityoftheRHTT.MeanTsk(Equation2.4)andPSIweresubsequentlycalculated(Equation
3.2).
Statisticalanalyses
AlldatawerefirstcheckedfornormalityusingtheShapiro-Wilkmethod.Physiologicalmeasures
duringRHTT1andRHTT2wereexaminedusingabatteryofrepeatabilitystatistics (3.17.3).Asa
measureofretestcorrelation,ICCwith95%CIwascalculatedforeachvariable.TEMwascalculated
andexpressedasapercentageofitsrespectivemeantoformtheTE(CV%).Bland-Altmanlimitsof
agreement(LOA)plotsshowingthemeanbiasand95%CIwereproduced.Theindividualparticipant
differences,betweenthetwotrialsforeachvariablewereplottedagainsttherespectiveindividual
means.Pairedsampledttestswerecalculatedtoidentifyanydifferencesinphysiologicalmeasures
between RHTT1 and RHTT2 (3.17.4). Effect sizes (Cohens’ d) were calculated to analyse the
magnitude of the interaction (Lakens, 2013) (3.17.5). All data were analysed using a standard
statistical package (SPSS version 20.0), and reported asmean ± SD. Statistical significancewas
acceptedatthelevelofp≤0.05.
4.4. Results
MeasuresofICC,TEM,theTE(CV%)andmeanbiaswithLOAforkeymarkersofheattoleranceare
presentedintable4.1.StrongcorrelationspresentedasICCwereobservedinpeakTre(r=0.93),
peakTsk(r=0.95),peakHR(r=0.99),peakPSI(r=0.98),andSWR(r=0.95)betweenRHTT1and

100
RHTT2(figure4.1).TheseobservedsimilaritiesarefurthersupportedbyalowTEMandTE(CV%)
forthekeymarkersofheattolerancebetweenRHTT1andRHTT2.
Figure 4.2 demonstrates small mean bias and LOA between trials for physiological measures
betweenthetwotrials.Inaddition,therewerenodifferencesobservedinpeakTre(t(15)=-0.785,p
=0.445,d=0.20),peakTsk(t(15)=0.223,p=0.827,d=0.06),HRpeak(t(15)=-1.005,p=0.331,d=
0.25),peakPSI (t (15)= -0.969,p=0.348,d=0.24),andSWR(t (15)=0.087,p=0.931,d=0.02)
betweenRHTT1andRHTT2.
StrongcorrelationspresentedasICC(95%CI)wereobservedinTrerest(r=0.88(0.38,0.90),p≤
0.001),Tremean(r=0.92(0.77,0.97),p≤0.001),HRrest(r=0.91(0.76,0.97),p≤0.001),andHR
mean(r=0.97(0.98,0.99),p≤0.001)betweenRHTT1andRHTT2.Theseobservedsimilaritiesare
furthersupportedbyalowTEMandTE(CV%)forTrerest(0.17°C,0.45%),Tremean(0.15°C,0.39%),
HRrest(3beats.min-1,5%),andHRmean(3beats.min-1,2%).ThereweresmallmeanbiasandLOA
forTrerest[-0.04,(-0.50,0.42)],Tremean[-0.04,(-0.45,0.37)],HRrest[0,(-8,8)],andHRmean[1,
(-8,10)].Furthermore,therewerenodifferencesinTrerest,(t(15)=-0.722,p=0.481,d=0.18)Tre
mean,(t(15)=-0.770,p=0.453,d=0.19)HRrest(t(15)=-0.301,p=0.768,d=0.08),andHRmean
(t(15)=-1.003,p=0.332,d=0.25)betweenRHTT1andRHHT2.
Strongcorrelations(ICC)wereobservedforpeakRPE(r=0.96,(0.88,0.99),p≤0.001)andpeakTS
(r=0.92,(0.76,0.97),p≤0.001)betweenRHTT1andRHTT2.Theseobservedsimilaritiesarefurther
supportedbyalowTEMandTE(CV%)forpeakRPE(0.6,4%),andpeakTS(0.2,3.2%).Therewere
smallmeanbiasandLOAforpeakRPE[-0.1,(-1.7,1.6)],peakTS[0.1,(-0.6,0.7)]betweenRHTT1
andRHTT2.Furthermore,therewerenodifferencesinpeakRPE(t(15)=-0.293,p=0.774,d=0.08)
andpeakTS(t(15)=0.145,p=0.0.27,d=0.28)betweenRHTT1andRHHT2.
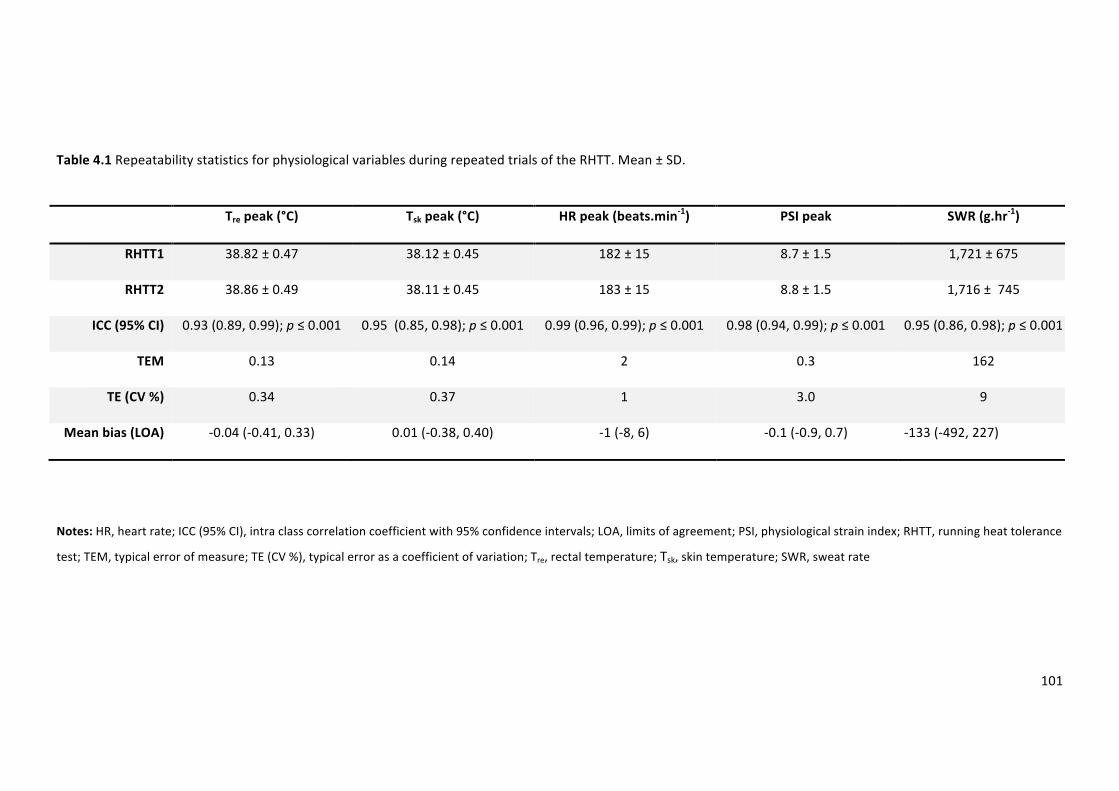
101
Table4.1RepeatabilitystatisticsforphysiologicalvariablesduringrepeatedtrialsoftheRHTT.Mean±SD.
Trepeak(°C) Tskpeak(°C) HRpeak(beats.min-1) PSIpeak SWR(g.hr-1)
RHTT1 38.82±0.47 38.12±0.45 182±15 8.7±1.5 1,721±675
RHTT2 38.86±0.49 38.11±0.45 183±15 8.8±1.5 1,716±745
ICC(95%CI) 0.93(0.89,0.99);p≤0.001 0.95(0.85,0.98);p≤0.001 0.99(0.96,0.99);p≤0.001 0.98(0.94,0.99);p≤0.001 0.95(0.86,0.98);p≤0.001
TEM 0.13 0.14 2 0.3 162
TE(CV%) 0.34 0.37 1 3.0 9
Meanbias(LOA) -0.04(-0.41,0.33) 0.01(-0.38,0.40) -1(-8,6) -0.1(-0.9,0.7) -133(-492,227)
Notes:HR,heartrate;ICC(95%CI),intraclasscorrelationcoefficientwith95%confidenceintervals;LOA,limitsofagreement;PSI,physiologicalstrainindex;RHTT,runningheattolerance
test;TEM,typicalerrorofmeasure;TE(CV%),typicalerrorasacoefficientofvariation;Tre,rectaltemperature;Tsk,skintemperature;SWR,sweatrate
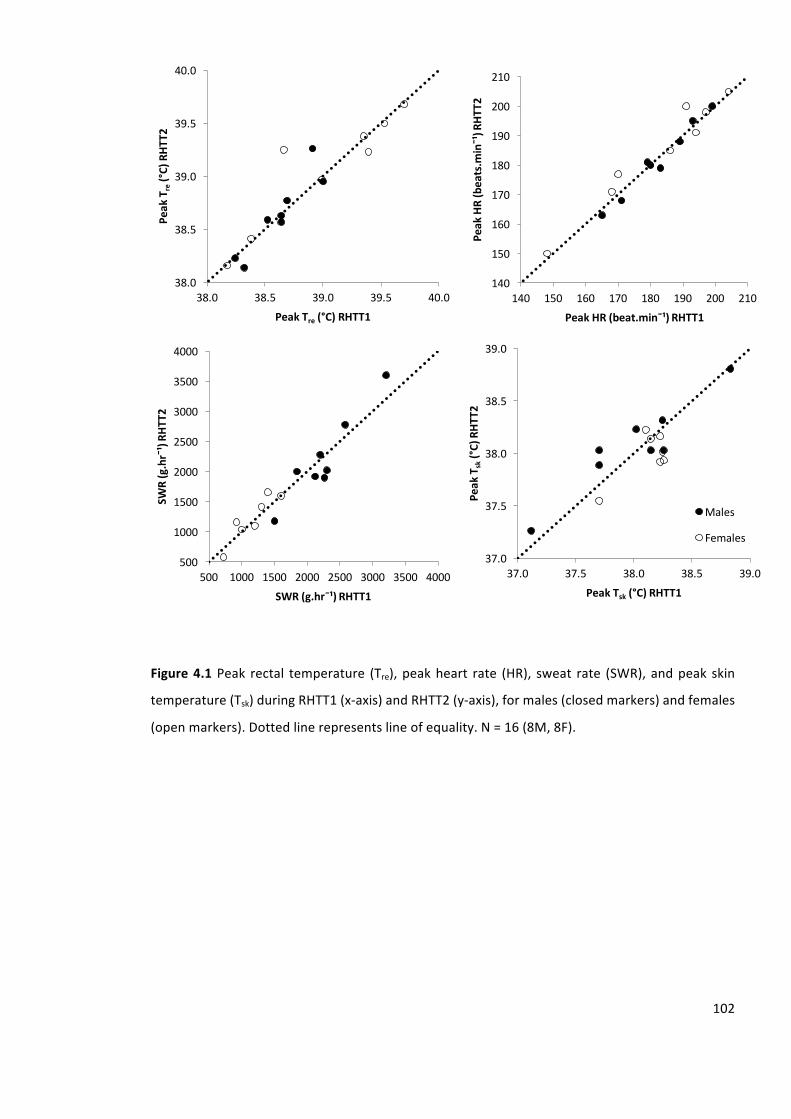
102
Figure4.1 Peak rectal temperature (Tre),peakheart rate (HR), sweat rate (SWR),andpeak skin
temperature(Tsk)duringRHTT1(x-axis)andRHTT2(y-axis),formales(closedmarkers)andfemales
(openmarkers).Dottedlinerepresentslineofequality.N=16(8M,8F).
38.0
38.5
39.0
39.5
40.0
38.0 38.5 39.0 39.5 40.0
PeakT
re(°C)RHT
T2
PeakTre (°C)RHTT1
140
150
160
170
180
190
200
210
140 150 160 170 180 190 200 210
PeakHR(beats.m
in¯¹)R
HTT2
PeakHR(beat.min¯¹)RHTT1
500
1000
1500
2000
2500
3000
3500
4000
500 1000 1500 2000 2500 3000 3500 4000
SWR(g.hr¯¹)RH
TT2
SWR(g.hr¯¹)RHTT1
37.0
37.5
38.0
38.5
39.0
37.0 37.5 38.0 38.5 39.0
PeakT
sk(°C)RHT
T2
PeakTsk (°C)RHTT1
Males
Females
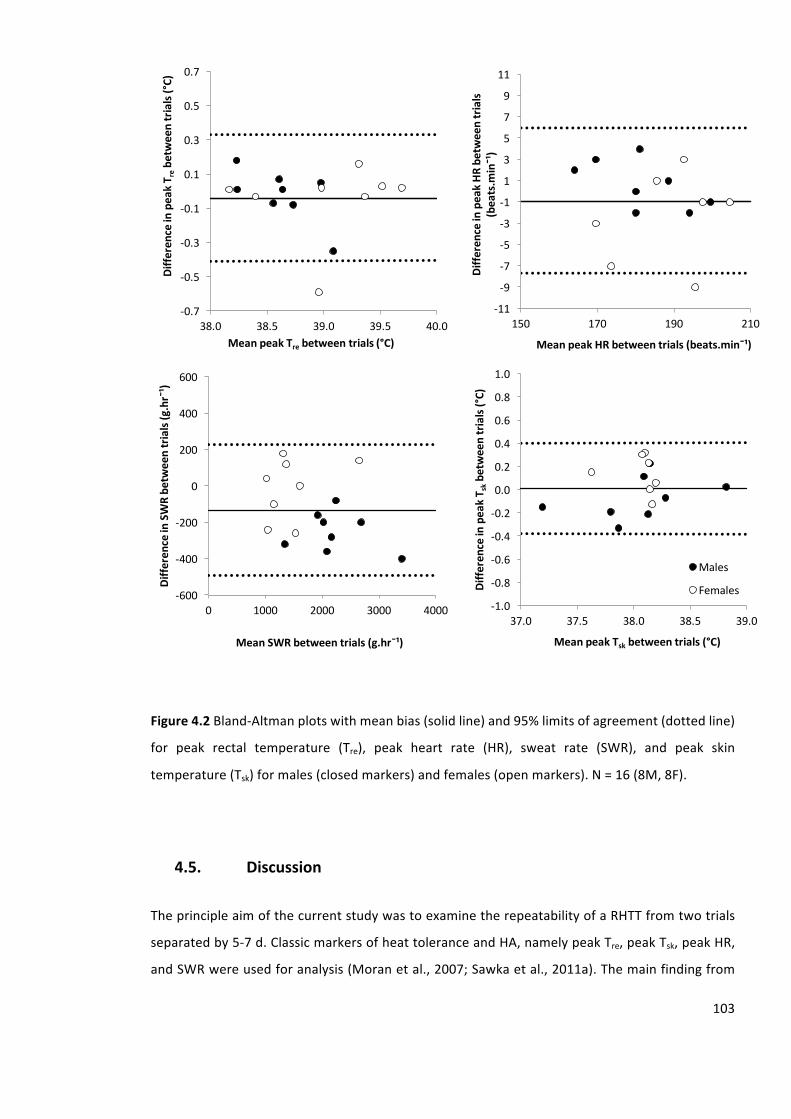
103
Figure4.2Bland-Altmanplotswithmeanbias(solidline)and95%limitsofagreement(dottedline)
for peak rectal temperature (Tre), peak heart rate (HR), sweat rate (SWR), and peak skin
temperature(Tsk)formales(closedmarkers)andfemales(openmarkers).N=16(8M,8F).
4.5. Discussion
TheprincipleaimofthecurrentstudywastoexaminetherepeatabilityofaRHTTfromtwotrials
separatedby5-7d.ClassicmarkersofheattoleranceandHA,namelypeakTre,peakTsk,peakHR,
andSWRwereusedforanalysis(Moranetal.,2007;Sawkaetal.,2011a).Themainfindingfrom
-0.7
-0.5
-0.3
-0.1
0.1
0.3
0.5
0.7
38.0 38.5 39.0 39.5 40.0
Diffe
renceinpeakT r
ebe
tweentrials(°C)
MeanpeakTre betweentrials(°C)
-11
-9
-7
-5
-3
-1
1
3
5
7
9
11
150 170 190 210
Diffe
renceinpeakHR
betweentrials
(beats.m
in¯¹)
MeanpeakHRbetweentrials(beats.min¯¹)
-600
-400
-200
0
200
400
600
0 1000 2000 3000 4000
Diffe
renceinSWRbe
tweentrials(g.hr¯¹)
MeanSWRbetweentrials(g.hr¯¹)
-1.0
-0.8
-0.6
-0.4
-0.2
0.0
0.2
0.4
0.6
0.8
1.0
37.0 37.5 38.0 38.5 39.0
Diffe
renceinpeakT s
kbe
tweentrials(°C)
MeanpeakTsk betweentrials(°C)
Males
Females

104
thecurrentstudyisthattheRHTThadgoodagreement,strongcorrelationsandsmalldifferences
between repeated trials and the TEM values for these classic markers suggested low within-
participantvariability.Thesefindingsareinsupportoftheaimofthestudy.Specifically,theprimary
andsecondaryhypotheseswerebothsupported.ValuesofpeakTre,peakTsk,peakHR,andSWR
spreadalongacontinuum,withawiderangeof responses,challengingthepreviousperception
thatindividualsareeitherheatheattolerantorheatintolerant(Kresfelderetal.,2006;Moranet
al., 2007). The data presented in the current study, demonstrates that the RHTT has strong
repeatabilityandisabletodifferentiatebetweenindividualresponsestotheRHTT.Thesefindings
supporttheuseoftheRHTTinfutureinvestigationstogaugeindividual’srunningheattolerance,
andtomonitortheextentofacuteandchronicheat-alleviatingprotocols.
To the author’s knowledge, there are no studies assessing the repeatability of physiological
measures during a fixed intensity running protocol in the heat, making acceptable levels of
repeatability a priori difficult to determine. Furthermore, there is no literature assessing the
repeatabilityoftheclassicalIDFHTTwhichwouldoffercomparison.Asageneralrule,acorrelation
coefficientover0.90isconsideredtobehigh,between0.70–0.80moderate,andbelow0.70tobe
lowforphysiologicaltests(Vincent,1995). Inthecurrentstudy,ICCvaluesforpeakTre,peakTsk,
peakHR,peakPSI,andSWRwereallequalorgreaterthan0.93.Basedonthesepredetermined
criteria, it is reasonable to suggest that the physiological measures during the RHTT have an
acceptablelevelofrepeatabilitybetweenrepeatedtrialsbasedonICC.
Thedata inthecurrentstudydemonstrates lessvariabilitythanavailabledataonfixedintensity
cyclingprotocolsintheheatusingsimilarphysiologicalvariables.TheTE(CV%)of0.3%and1%for
peakTreandpeakHRrespectively,comparefavourablytotheTE(CV%)of0.3%and3.9%formean
TreandmeanHRreportedduringa60-min,fixedintensitycyclingprotocolinhothumidconditions
(Haydenetal.,2004).Furthermore,Brokenshireandcollegues(2009)investigatedtherepeatability
ofHR during a fixed intensity cycling heat stress test,which involved three 20-min cycle bouts
separatedby8-min rest. The ICCof0.95andTE (CV%)of3% formeanHRwere reportedand
assumed indicative of strongmeasurement repeatability. The ICC of 0.99 and TE (CV%) of 1%
reportedinthecurrentstudy,againcomparefavourablytothesefindingshighlightingthestrength
oftheRHTTintermsoflowmeasurementerror.
To aid interpretation and presentation of results when assessing individuals’ heat tolerance,
researchers havedichotomized the population and categorized individuals as either tolerant or
intolerant(Kresfelderetal.,2006;Moranetal.,2007).AltmanandRoyston(2006)reportedseveral
problemswithdichotomizingdata.Statisticalpowertodetectarelationshipmaybereduced,there

105
isanincreasedriskofreportingafalsepositive,apossibilityofunderestimatingtheextentofthe
variationintheoutcomebetweengroups,andusingtwogroupsconcealsanynon-linearityinthe
relationship.AltmanandRoyston(2006)stateusingmultiplecategories isgenerally favouredto
dichotomizingdata;withfourorfivegroupsthelossofinformationcanbequitesmallbutthere
arecomplexitiestoanalysis.Indeed,Moranandcolleagues(2007)acknowledgethatthelargerthe
deviationsfromnormalvaluesthemorepronouncedthestateofheatintolerance,thusimplyinga
continuum of heat tolerance. Similarly work from, Taylor and Cotter (2006) propose that heat
adaptation isa continuum,with thepositionofan individualalong thecontinuumrepresenting
progressiveincreasesinheattolerance.Thefindingsinthepresentstudy,demonstrateclearlythat
anindividual’sheattolerance,representedbythepeakTre,peakTsk,peakHR,peakPSI,andSWRis
acontinuousvariable;demonstratingthatheattolerancemaybemoreaccuratelycategorisedon
a continuum. Consequently, these findings support the use of the RHTT to track changes in
individuals’ ability to withstand exercise heat stress from acute and chronic heat-alleviating
interventions.
Casesofexertionalheat illness canoccuramongenduranceathletes, inextremecircumstances
leading to death; however, the epidemiology is not well documented in the literature.Martin
(1997)reportedthehighestincidenceofexertionalheatillnessoccurredinlongdistancerunners
duringthe1992NewOrleansU.S.OlympicTrialsandthe1996AtlantaOlympics;withlongdistance
runners accounting for 31% and 53% respectively, of the total cases of exertional heat illness.
Furthermore,Nielsen(1996)providesdatatosuggesttheincidenceofheatillnessisunavoidable
forendurance runnerswhencompeting inahighambient temperaturecombinedwithhighRH
withoutasevere reduction inenduranceperformance;highlighting the importanceof thorough
preparationtopreventtheincidenceofaheatillness.Thefindingsinthecurrentstudycouldbe
applied in a manner that would serve to minimise the number of athletes that suffer from
hyperthermia, by supporting amore complete evaluation and subsequent preparation prior to
trainingandcompetingintheheat(Johnsonetal.,2013).
TheTEMexpressedas a coefficientof variation canbeused toestimate sample size for future
studiesusingtheRHTTwhenasmallestworthwhilechangeisknown;usingtheformulaN=8s2/d
2
(where‘s’=typicalerrorexpressedasacoefficientofvariationand‘d’=thesmallestworthwhile
change (Hopkins, 2000b). A change of 0.4°C typically represents a significant reduction in Tre
followingacuteandchronicheat-alleviatingprocedures(Buonoetal.1998;Pattersonetal.2004;
Castleetal.2006;Garrettetal.2009).Accordingly,inthecurrentstudytodetecta0.4°Cchange,a
sample sizeof sixparticipantswouldbesufficientwhenpeakTre is thekeyvariableof concern,
assumingsimilarvariabilityamongtheparticipantsrecruited.

106
4.6. Limitations
The RHTT adopts a fixed absoluteworkload,which is an appropriate procedure to assess heat
tolerance when the experimental design is a repeated measure. However, the fixed absolute
workloadmaylimitapplicabilityofthetestwhencomparingbetweengroups,especiallywhenthey
are not matched. The recent work by Cramer & Jay (2014) reports when comparing between
individuals or groups unmatched from a biophysical perspective, exercise intensity should be
administeredusingfixedheatproductionrelativetoBMtopreventtheintroductionofsystematic
bias. Future research is warranted to quantify the potential differences in metabolic heat
production between participants of differing BM, BSA and fitness during the RHTT to ensure
unbiasedcomparisonofthermoregulatoryresponsesbetweenindividuals(CramerandJay,2014).
Furthermore,theroleoftheRHTTinidentifyingindividuals’susceptibilitytoexertionalheatillness
hasnotbeen identifiedwithin the current study.CollectingRHTTdataon individualswhohave
previously incurred exertional heat illness would test the value of the RHTT in identifying an
individual’ssusceptibilitytoexertionalheatillness.
4.7. Conclusion
ThemainfindingfromthecurrentstudywasthattheRHTTdemonstratedgoodagreement,strong
correlationsandsmalldifferencesbetweenrepeatedtrials.TheTEMvaluesshowed lowwithin-
participantvariability. Inaddition,data ispresentedtodemonstratethattheRHTT iscapableof
differentiating between individuals’ responses to the RHTT when adopting a running mode of
exercise;providingevidencethata30-minHTTprotocolissufficientinduration.Furthermore,the
findingsfromthepresentstudychallengethepreviousreportsthatheattoleranceisdichotomous,
sinceindividualresponseswerelinearlyspreadoveracontinuum.Thesefindings,couldbeusedto
observeindividuals’thermoregulatoryresponsesovertime,providinginformationtophysiologist,
coaches,andmedicalstafftoassistdecisionsaboutathletes’safetypriortotrainingandcompeting
intheheat.Inaddition,theRHTTcouldbeusedasatoolforinvestigatingtheeffectofacuteand
chronicheat-alleviatingprocedures.
ThischapterexploredtherepeatabilityoftheRHTT.Thisestablishedituseasameasurementtool.
Chapter5(Study2)willassessthesensitivityoftheRHTTtochangesinheattolerance.

107
5. Sensitivityofarunningheattolerancetest
________________________________________________________________________________
Study1(Chapter4)ofthisthesishasestablishedtheRHTTasreliable,modespecifictest.Toassess
thesensitivityoftheRHTTtochangesinheattolerance,datawaspooledfromtwofurtherstudies
ofwhicharenotpresentedelsewhereinthethesis.
5.1. Abstract
EstablishingwhethertheRHTTissensitivetochangesinheattoleranceisessentialtosupportfuture
use of the RHTT in providing a more complete evaluation of athletes’ heat tolerance, and to
quantifying the effectiveness of heat-alleviating procedures. This study aimed to identify the
sensitivity of the RHTT to changes in heat tolerance following STHA. Twenty-five (12M, 13F)
participantsperformedtwoRHTT,24-hrprecedingHA(RHTT1)and24hrsfollowing5-dcontrolled
hyperthermia(Tre=38.5°C)HA(RHTT2).TheRHTTinvolved30-minrunning(9km.hr-1,2%gradient)
in40°C,40%RH(WGBT=33°C).FollowingHA,valuesofpeakTre(RHTT1:38.94±0.48°C,RHTT2:
38.56±0.55°C,p≤0.001),peakTsk(RHTT1:37.47±0.49°C,RHTT2:37.06±0.70°C,p=0.003),and
peakHR(RHTT1:185±14beats.min-1,RHTT2:175±14beats.min
-1,p≤0.001)reduced,andSWR
increased(RHTT1:1,070±541g.hr-1,RHTT2:1,481±457g.hr
-1,p=0.003).Therangeofreductions
inmeasuresofpeakTre(-0.48°C,-0.03°C),peakTsk(-1.74°C,0.13°C),peakHR,(-24beats.min-1,-2
beats.min-1)andSWR(-460g.hr
-1,1640g.hr
-1)demonstratemarkedindividualdifferences.Thedata
suggeststheRHTTissensitivetochangesinheattoleranceandthatchangesinheattoleranceare
highlyindividual.
5.2. Introduction
Duringprolongedsubmaximalexerciseinahotenvironmentthereisariseinbodyheatstorage,
which in turn reflects thebalancebetweenmetabolicheatproduction,heatabsorbed fromthe
environmentandtotalbodyheatloss(JayandKenny,2007).Thereisawidevariationinindividual
abilitytowithstandexerciseheatstress(Epstein,1990).Underextremeconditionsofexertionin
theheatevenhealthy,heatacclimated,andendurancetrainedindividualswillincreasetheirbody
heat content. Individualswhoexperienceagreater storageofmetabolicheatdue toa reduced
sweatingsensitivityanda lowadaptivecapacitytoexercise intheheat,havebeendescribedas
heatintolerant(Epsteinetal.,1983;Moranetal.,2004).Theabilitytoquantifyanindividual’slevel

108
ofheattoleranceoffersvaluableinformationtosupportthepreventionoffutureoccurrenceofa
heat-relatedillness.
A RHTT was developed in Study 1 (Chapter 4). The protocol involves 30-min of running on a
treadmill(9km.hr-1anda2%gradient)inambientconditionsof40°Cand40%RH(WBGT=33°C).
Classicmarkersofheattoleranceandadaptationnamely,Tre,Tsk,HR,andSWRdemonstratedgood
agreement, strong correlations, (ICC>0.93) and small differences (p ≥ 0.05)between repeated
trials. Furthermore, the TEM values for these classicmarkers suggested lowwithin participant
variability.TheRHTTdemonstrateddifferentdegreesofheattolerancebetweenindividualswhich
reinforcesthattheRHTTisanappropriatetooltoevaluateheattolerance.Thereiscurrentlyno
evidence supporting the sensitivity of the RHTT to changes in heat tolerance. Establishing the
sensitivityoftheRHTTtochangesinheattolerancemaysupporttheuseoftheRHTTtomonitor
changesinheattolerancefollowingacuteandchronicheat-alleviatingprocedures.Inaddition,it
maysupporttheuseoftheRHTTtomonitorchangesinheattolerancethatoccurnaturallytohelp
identifyperiodsoftimewhenathletesareexperiencingperiodsoflowheattolerancetosupport
thepreventionofdevelopingaheat-relatedillness.
An individual’sheat tolerancehasbeendefinedas theability towithstandexerciseheat stress,
combinedwiththeirabilitytoadapttoexercisingintheheat(Shapiroetal.,1979).Routinely,HTTs
are used in isolation to assess heat tolerance following a heat illness incident to support the
individual’sreturntotraining.Recently,Johnsonandcolleagues(2013)providedcasestudydatato
supporttheuseofacombinedheattolerancescreeningprocedure,usingbothacuteandchronic
heatexposuretoassessatriathlete’sexerciseheattolerance.Theseexploratoryfindingssupporta
combined method to assess athletes’ heat tolerance to facilitate optimal preparation prior to
trainingandcompetingintheheat.
ThisstudyaimedtoestablishthesensitivityoftheRHTTtochangesinheattolerance.Furthermore,
this studyaimed toexplore the individual changes inheat tolerance followingHA. Theprimary
hypothesiswasthattherewillbedifferencesinphysiologicalresponsesduringtheRHTTfollowing
STHA.ThesecondaryhypothesiswasthattheRHTTwillbesensitivetoindividualdifferencesinthe
magnitudeofadaptation.
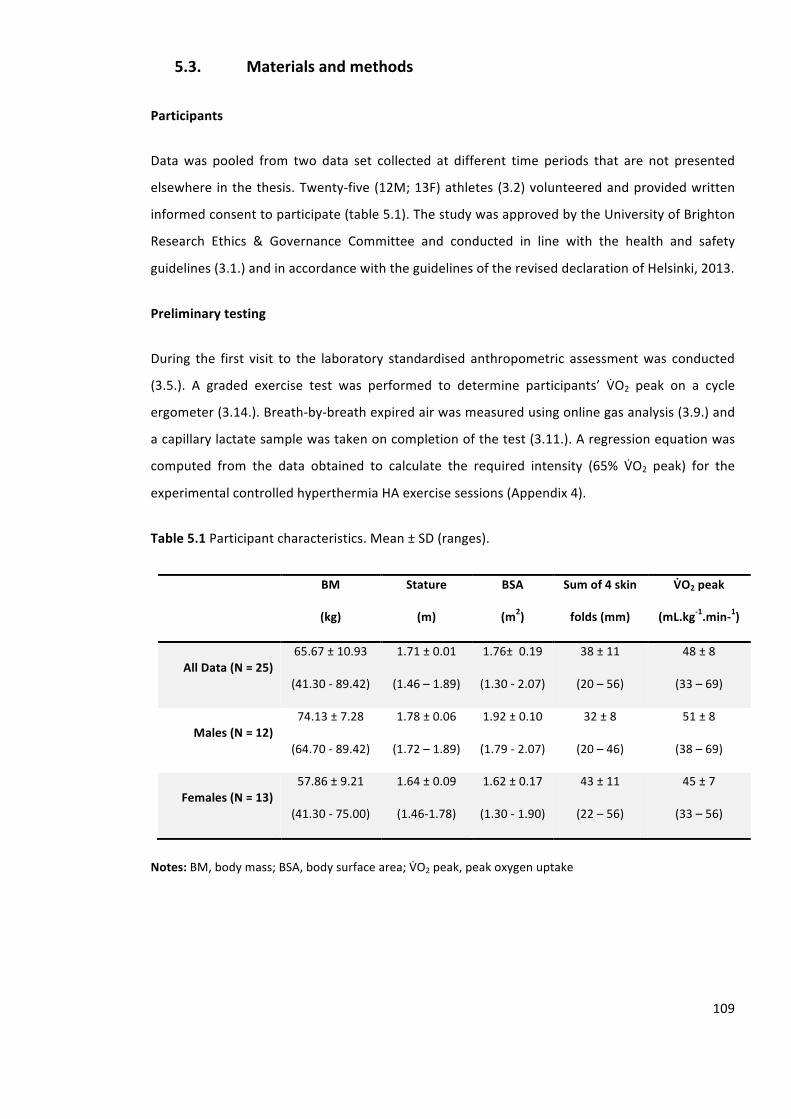
109
5.3. Materialsandmethods
Participants
Datawas pooled from two data set collected at different time periods that are not presented
elsewhere inthethesis.Twenty-five(12M;13F)athletes(3.2)volunteeredandprovidedwritten
informedconsenttoparticipate(table5.1).ThestudywasapprovedbytheUniversityofBrighton
Research Ethics & Governance Committee and conducted in line with the health and safety
guidelines(3.1.)andinaccordancewiththeguidelinesofthereviseddeclarationofHelsinki,2013.
Preliminarytesting
During the first visit to the laboratory standardised anthropometric assessmentwas conducted
(3.5.). A graded exercise test was performed to determine participants’ VnO2 peak on a cycle
ergometer(3.14.).Breath-by-breathexpiredairwasmeasuredusingonlinegasanalysis(3.9.)and
acapillarylactatesamplewastakenoncompletionofthetest(3.11.).Aregressionequationwas
computed from the data obtained to calculate the required intensity (65% VnO2 peak) for the
experimentalcontrolledhyperthermiaHAexercisesessions(Appendix4).
Table5.1Participantcharacteristics.Mean±SD(ranges).
BM
(kg)
Stature
(m)
BSA
(m2)
Sumof4skin
folds(mm)
VSO2peak
(mL.kg-1.min-1)
AllData(N=25)65.67±10.93
(41.30-89.42)
1.71±0.01
(1.46–1.89)
1.76±0.19
(1.30-2.07)
38±11
(20–56)
48±8
(33–69)
Males(N=12)74.13±7.28
(64.70-89.42)
1.78±0.06
(1.72–1.89)
1.92±0.10
(1.79-2.07)
32±8
(20–46)
51±8
(38–69)
Females(N=13)57.86±9.21
(41.30-75.00)
1.64±0.09
(1.46-1.78)
1.62±0.17
(1.30-1.90)
43±11
(22–56)
45±7
(33–56)
Notes:BM,bodymass;BSA,bodysurfacearea;VnO2peak,peakoxygenuptake

110
Experimentaldesign
Testing was completed over a 7-d period. The timing of testing for female participants was
conducted in line with the menstrual cycle standardisation guidelines (3.6.). Five female
participantswere using oral contraceptive: three usedMicrogynon, one used Rigevidon (30 µg
ethinylestradiol/ 150 µg levonorgestrel) and one used Yasmin (30 µg ethinylestradiol/ 3mg
drospirenone). All trialswere conducted in linewith pre-trial diet and exercise standardisation
(3.4.). Participants provided a urine sample upon arrival to ensure adequate hydration (3.4.).
VolunteersperformedfivecontrolledhyperthermiaHAsessionsonacycleergometer(3.14.)and
twoRHTTs(3.16.).ThefirstRHTTwasperformed24-hrpriortobeginningHA(RHTT1),thesecond
RHTTwasperformed24-hrfollowing5-dHA(RHTT2).Allexperimentaltrialswereperformedina
hotenvironment(3.7.).Exercisewasterminatedinlinewithuniversityguidelines(3.1.).Therewere
noincidencesofearlyterminationofexerciseduringtheRHTTwithinthisstudy.
Runningheattolerancetest
TheRHTT(3.16.)wasperformedontwooccasionsonamotorisedtreadmill(3.15.).Followinga20-
minstabilisationperiodinatemperateenvironment(3.7.),measuresofTre,Tsk(3.8.),HR(3.9.),RPE,
andTS(3.13.)wererecorded.Theparticipantsthenenteredtheenvironmentalchamber.Towel
driednudeBMwasalsorecordedtocalculateSWR(3.12.).ThroughoutthetestTre,Tsk,andHRwere
recordedat5-minintervals.RPEandTSwererecordedat10-minintervals.MeanTsk(Equation2.4)
andPSI(Equation3.2)weresubsequentlycalculated.
Statisticalanalyses
DatawereassessedfornormalityusingtheShapiro-Wilkmethod.Apairedsamplettest(3.17.4)
wascalculatedtoidentifyanydifferencesinphysiologicalmeasuresbetweenD1andD5ofHAand
betweenRHTT1andRHTT2.Effectsizes(Cohen’sd)werecalculatedtoanalysethemagnitudeof
theinteraction(Lakens,2013)(3.17.5.).Alldatawereanalysedusingastandardstatisticalpackage
(SPSSversion20.0),andreportedasmean±SD.Statisticalsignificancewasacceptedatthelevelof
p≤0.05.

111
5.4. Results
Heatacclimation
Allparticipantscompletedfiveconsecutivedaily90-minHAsessions.Participantsmaintainedthe
targetTreof38.5°Conaveragefor48±9minduringeachsession.Themeandurationofexercise
was64±12minpersession.Themeansessionintensity(includingrestperiods)was41±4%VnO2
peak;however, themeanexercise intensity (excludingrestperiods)was58±7%VnO2peak.The
meanTreandHRduringeachHAsessionwas38.27±0.13°Cand147±9beats.min-1,respectively.
Furthermore,themeanSWRduringeachHAsessionwas556±153g.hr-1.
TherewasareductioninTrerest(t(24)=2.192,p=0.037,d=0.408)andHRrest(t(24)=3.758,p≤
0.001,d=0.609)fromD1toD5ofHA.TherewasanincreaseinSWRduringtheHAsessionfrom
D1toD5(t(24)=-3.030,p=0.006,d=0.526).Furthermore,therewasanincreaseinSWRBSA(t(24)=
-2.917,p=0.008,d=0.512).
Runningheattolerancetest
Table5.2presentsthemean±SDdataforTrerest,peakTre,peakTsk,HRrest,peakHR,andSWR
duringtheRHTT.Therewasa0.16±0.29°Cand0.38±0.38°CreductioninTrerest(t(24)=2.517,p=
0.019,d=0.46)andpeakTre(t(24)=4.957,p≤0.001,d=0.71)respectively,fromRHTT1toRHTT2.
Adaptive responsevariedbetweenparticipantswith changes inTre rest ranging from -0.62°C to
0.41°CandpeakTrerangingfrom-1.43°Cto-0.03°C.Therewasa0.41±0.51°CreductioninpeakTsk
fromRHTT1andRHTT2(t(24)=3.348,p=0.003,d=0.56);changesrangedfrom-1.74°Cto0.93°C
(figure5.1).
Therewasa4±9beats.min-1and10±9beats.min
-1reductionsinHRrest(t(24)=2.195,p=0.038,
d=0.41)andpeakHR(t(24)=5.788,p≤0.001,d=0.76)respectively,fromRHTT1toRHTT2.Adaptive
responsesvariedbetweenparticipantswithchangesinHRrestrangingfrom-19beats.min-1to8
beats.min-1andpeakHRrangingfrom-24beats.min
-1to0beats.min
-1.
ThesechangesresultedinanaveragereductioninpeakPSIfromaveryhighstraintoahighstrain
(Moran et al., 1998) (t (24) = 5.268, p < 0.001, d = 0.73). Adaptive responses varied between
participantswithchangesinPSIpeakrangingfrom-0.06to4.30.
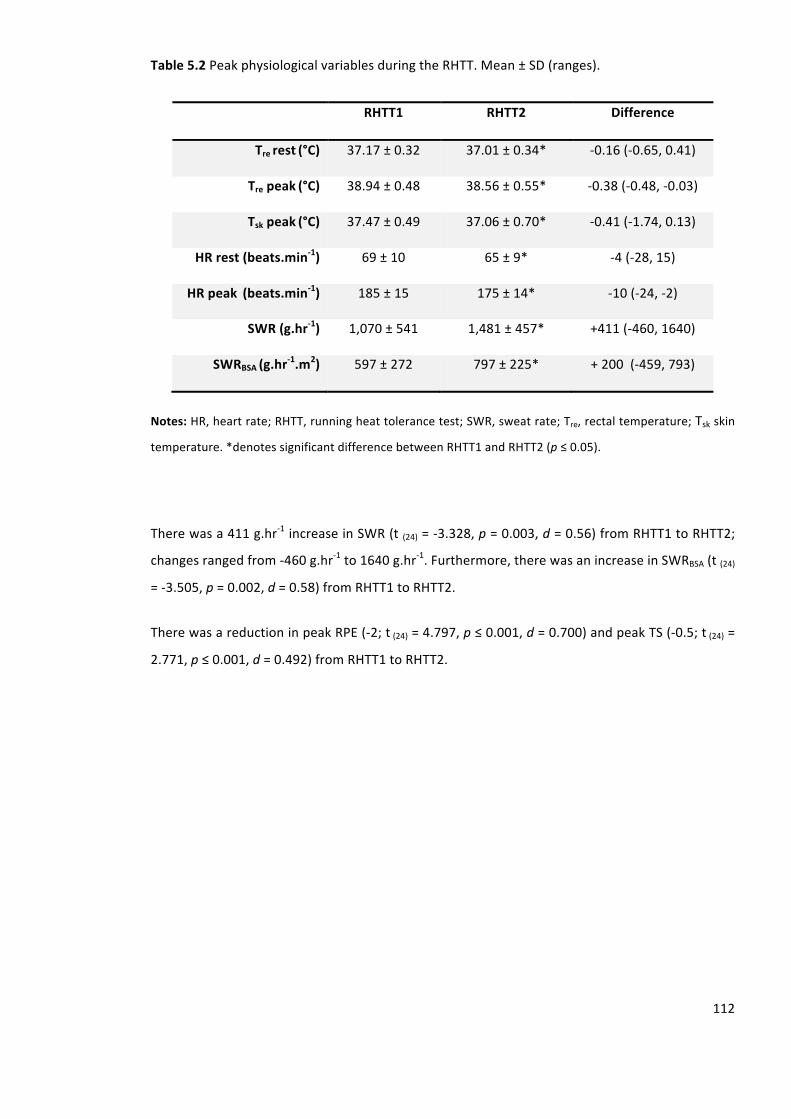
112
Table5.2PeakphysiologicalvariablesduringtheRHTT.Mean±SD(ranges).
RHTT1 RHTT2 Difference
Trerest(°C) 37.17±0.32 37.01±0.34* -0.16(-0.65,0.41)
Trepeak(°C) 38.94±0.48 38.56±0.55* -0.38(-0.48,-0.03)
Tskpeak(°C) 37.47±0.49 37.06±0.70* -0.41(-1.74,0.13)
HRrest(beats.min-1) 69±10 65±9* -4(-28,15)
HRpeak(beats.min-1) 185±15 175±14* -10(-24,-2)
SWR(g.hr-1) 1,070±541 1,481±457* +411(-460,1640)
SWRBSA(g.hr-1.m2) 597±272 797±225* +200(-459,793)
Notes:HR,heartrate;RHTT,runningheattolerancetest;SWR,sweatrate;Tre,rectaltemperature;Tskskin
temperature.*denotessignificantdifferencebetweenRHTT1andRHTT2(p≤0.05).
Therewasa411g.hr-1increaseinSWR(t(24)=-3.328,p=0.003,d=0.56)fromRHTT1toRHTT2;
changesrangedfrom-460g.hr-1to1640g.hr
-1.Furthermore,therewasanincreaseinSWRBSA(t(24)
=-3.505,p=0.002,d=0.58)fromRHTT1toRHTT2.
TherewasareductioninpeakRPE(-2;t(24)=4.797,p≤0.001,d=0.700)andpeakTS(-0.5;t(24)=
2.771,p≤0.001,d=0.492)fromRHTT1toRHTT2.
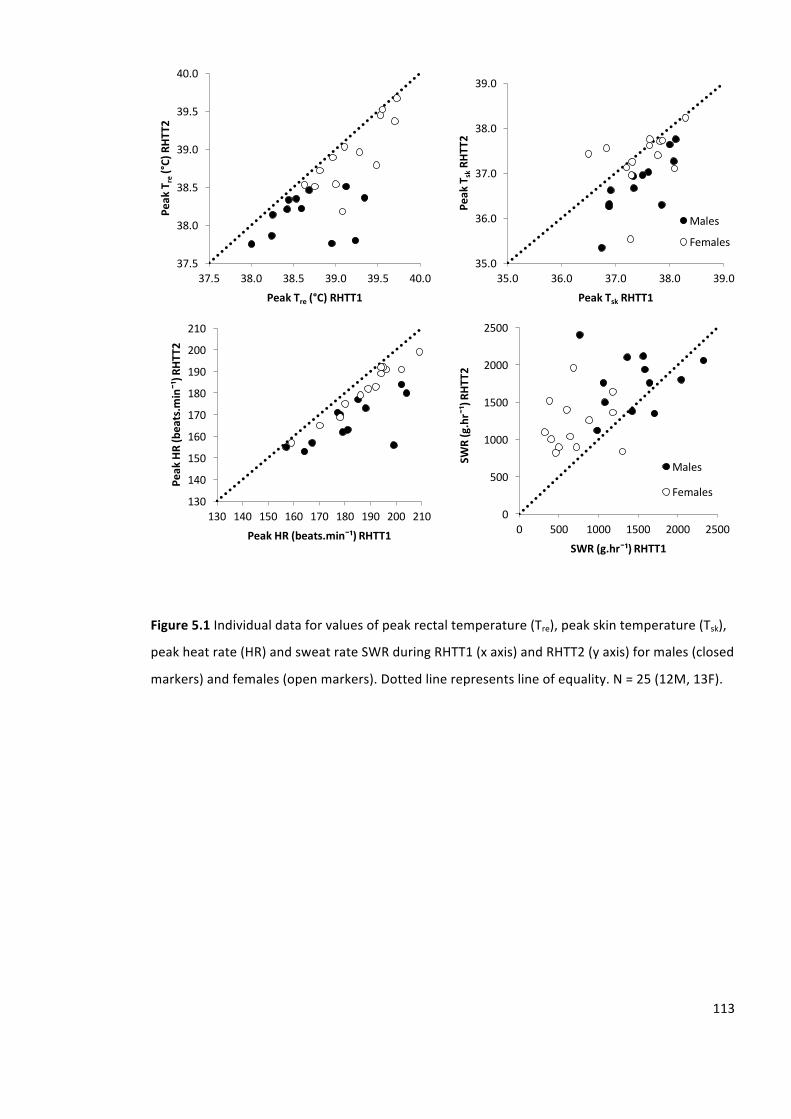
113
Figure5.1Individualdataforvaluesofpeakrectaltemperature(Tre),peakskintemperature(Tsk),
peakheatrate(HR)andsweatrateSWRduringRHTT1(xaxis)andRHTT2(yaxis)formales(closed
markers)andfemales(openmarkers).Dottedlinerepresentslineofequality.N=25(12M,13F).
37.5
38.0
38.5
39.0
39.5
40.0
37.5 38.0 38.5 39.0 39.5 40.0
PeakT
re(°C)RHT
T2
PeakTre (°C)RHTT1
35.0
36.0
37.0
38.0
39.0
35.0 36.0 37.0 38.0 39.0
PeakT
skRH
TT2
PeakTsk RHTT1
Males
Females
0
500
1000
1500
2000
2500
0 500 1000 1500 2000 2500
SWR(g.hr¯¹)RH
TT2
SWR(g.hr¯¹)RHTT1
Males
Females130
140
150
160
170
180
190
200
210
130 140 150 160 170 180 190 200 210
PeakHR(beats.m
in¯¹)R
HTT2
PeakHR(beats.min¯¹)RHTT1

114
5.5. Discussion
ThemainfindinginthecurrentstudywasthattheRHTTissensitivetothechangesinheattolerance
thatareassociatedwithSTHA,evidencedbyareductioninTre,Tsk,andHR,andanincreaseinSWR.
Thesefindingssupporttheaimofthestudy.Specifically,theprimaryandsecondaryhypotheses
were both supported. These results support the use of the RHTT to monitor changes in heat
tolerancewhenusingarepeatedmeasuresdesign.Furthermore,thedatademonstratesmarked
differencesbetweenparticipants’changesinheattolerance,suggestingheatadaptationishighly
individual.
Heatadaptationresponses
Therewasa0.38°CmeanreductioninpeakTrefollowingSTHAinthecurrentstudy.Study1(Chapter
4)identifiedtheTEMforpeakTreduringtheRHTTtobe0.13°C,suggestingatruereductionof>
0.25°C. These findings, are comparable to the~0.24°C reduction, previouslyobserved following
STHA using a controlled hyperthermia model (Garrett et al., 2009; Patterson et al., 2004a).
Furthermore,Study1(Chapter4)identifiedaTEMof0.14°CforpeakTskduringtheRHTT,the0.41°C
reductionobservedinthecurrentstudyalsosuggestsatruereductionof>0.27°C.Theobserved
reductions inthermoregulatorystrainarepotentiallyduetoanenhancedevaporativeheat loss,
thus reducing heat storage. These results demonstrate the sensitivity of the RHTT to monitor
changesinthermoregulatorystrainassociatedwithSTHA.
Therewas a 10 beats.min-1mean reduction in peakHRduring the RHTT following STHA in the
currentstudy.Study1(Chapter4)identifiedtheTEMforpeakHRduringtheRHTTtobe2beats.min-
1, suggesting a true reduction of > 8 beats.min
-1. These findings are comparable to the ~12
beats.min-1reductionpreviouslyobservedfollowingSTHAusingacontrolledhyperthermiamodel
(Garrettetal.,2009;Pattersonetal.,2004a).Thereductionincardiovascularstrainrepresentan
improved myocardial efficiency and ventricular compliance (Lorenzo et al., 2010). Due to the
matched exercise intensity of the RHTT, the observed reduction in cardiovascular strain
demonstrates the sensitivity of the RHTT to monitor adaptation to the cardiovascular system
associatedwithSTHA.
Therewasa411g.hr-1meanincreaseobservedinSWRfollowingSTHAinthecurrentstudy.Study
1(Chapter4)identifiedtheTEMforSWRduringtheRHTTtobe162g.hr-1,suggestingatrueincrease
of≥249g.hr-1;thesefindingsarecomparabletothe220g.hr
-1previouslyobservedfollowingSTHA
usingacontrolledhyperthermiamodel(Pattersonetal.,2004a).AnincreaseinSWRcouldbeeither

115
analteredafferentneuralactivityfromtheperipheralorcentralthermo-receptorscausingdifferent
integrationofthermalinformation,analteredefferentneuralactivityforagivenlevelofafferent
inputoranalteredeffectorresponse.Thepotentialmechanismsforthisincludealterationsinthe
Tcthresholdforsweatingonset(Shvartzetal.,1979),anincreasedsweatingsensitivity(Shvartzet
al.,1979)and/oreccrinesweatglandhypertrophy(Satoetal.,1990).Duetothematchedexercise
intensityduringtheRHTT,therepeatedtrialswereassumedtobematchedfortherequirementfor
evaporative heat loss thus, these results demonstrate the sensitivity of the RHTT to monitor
changesinsudomotorfunctionfollowingHA.
Individualresponses
Observing individual heat tolerance using both an acute and chronic heat exposure has been
suggestedtoofferamorecomprehensiveassessmentofindividualheattolerance(Johnsonetal.,
2013).However,themajorityofHAliteraturereportsaverageresponsesandfailstorecognisethe
differences inadaptiveresponses.Thedatademonstrated largedifferences inthemagnitudeof
adaptationbetweenindividuals.Forexample,oneparticipantinthecurrentstudydemonstrateda
largeadaptiveresponsefollowingSTHAwithpeakPSIreducingfromhigh(8.65)tomoderate(6.98)
strain, combined with a 150% increase in SWR. However, another participant experienced no
alterations in peak PSI and had only a 23% increase in SWR, demonstrating a small adaptive
response.
There are several factorswhich havebeen suggested to alter themagnitudeof the adaptation
response which may explain the variance observed in the current study. Firstly, the closer an
individualistotheirgeneticallydeterminedmaxima,thegreatertheadaptationresistanceasthe
adaptionreservenarrowswithprogressivelysmallgainsoccurringovertime(Prud’Hommeetal.,
1984).Furthermore,withinanygivenpopulationtherearehighrespondersandlowrespondersto
thermal stimulation (Taylor, 2014). The majority of young sedentary individuals who live in a
predominantlytemperateclimatewillbehighthermalresponders.Whereas,individualswholive,
workandtraininhighambientconditionswillshowadaptationresistance(Taylor,2014).
TheexerciseelicitedduringtheHAsessionswasperformedat65%ofVnO2peak.Thereareseveral
limitationsobservedwithsettingexercisepercentagebasedonVnO2peakwhichmayexplainthe
varianceobservedinthemagnitudeofadaptation incurrentstudy.TheVnO2peakvaluesforthe
participants’ in the current study ranged from33mL.kg-1.min
-1 to 69mL.kg
-1.min
-1. Thus, those
individualswithalowerVnO2peakworkedatalowermetabolicheatproductionprovidingalower
stimulusforsweatproduction(CramerandJay,2014;Gagnonetal.,2013,2008)andpotentially

116
constrainingadaptation.TheBMoftheparticipants’inthecurrentstudyrangedfrom41.3kgto
89.4kg.AlowerBMentailslessheatstorageandtherefore,alowerexerciseintensityisrequired
to increaseTc to38.5°C (Gagnonet al., 2009). Consequently, the stress imposedduring theHA
sessionswaslikelylowerinparticipantswithalowerBMcomparedwiththosewithahigherBM,
potentially constraining adaptation and influencing the time course of adaptation due to
inadequate endogenous heat strain. Furthermore, changes in Tc are not only a function of the
changeinbodyheatcontent,butalsoofitsmassandtissuecompositionasdefinedbythespecific
heat of the tissue of the human body. The participants’ sum of skin fold and thus, body fat
percentagerangedfrom20mmto56mm.Anindividualwithalowerbodyfatpercentagewillhave
ahigherspecificheat.Thus,atagivenchangeinbodyheatcontentparticipantswithalowbodyfat
percentage will have a lower change in Tc (Jay and Kenny, 2007), potentially constraining
adaptation.
Johnsonetal.,(2013)observedanimprovedheattolerancefollowing9-dHAinatriathletewho
hadpreviousincidencesofheatillness.Followingthisassessment,theathletewasdeemedsafeto
race,duetoasubstantialimprovementinheattolerance,andwentontocompletetheracewith
nosignsofexertionalheatillness.Thecurrentstudysupportsthismodeltoevaluateindividuals’
heat tolerance since therewere a variety of adaptive responses between individuals. Although
thresholds have not been established, those individuals who demonstrate small adaptive
responses,especiallywhencombinedwithahighphysiologicalstrainduringtheRHTT,will likely
experiencesubstantial strainduringcompetitionandpotentiallyatahigher riskofexperiencing
exertionalheatillness.Thus,whenassessingheattolerance,practitionersshouldconsiderusingthe
RHTTpreandpostaheat-alleviatingproceduresuchasHA.Thismethodisconsideredtoprovidea
more in-depth assessment of heat tolerance with information being provided on the initial
exposure and the adaptive capacity of an individual; to support the modification of future
preparationstrategiespriortotrainingandcompetingintheheat.
5.6. Limitations
TheRHTTmaybe limitedby its applicabilitywhen comparingbetweengroups, especiallywhen
participantsarenotmatchedforbiophysicalcharacteristics.Administeringafixedintensityprotocol
whencomparingbetweengroupswillelicitexerciseatavarietyofrequirementsforevaporative
heatloss,resultinginaninaccurateassessmentofthermoregulatorystrainbetweenparticipants
(CramerandJay,2014;Gagnonetal.,2013,2008).However,theRHTTisanappropriateprocedure
toassessheattolerancewhentheexperimentaldesignisarepeatedmeasure,ortotrackanathlete
overtimepriortotrainingandcompetinginhighambientconditions.

117
Furthermore, the roleof theRHTT toaid inclinicaldecisions followingexertionalheat illness in
athletes,andpredictingindividuals’susceptibilitytoexertionalheatillnessisnotknown.Collecting
dataonindividualswhohavepreviouslyincurredexertionalheatillnesswouldreinforcethevalue
oftheRHTT.Providingdiagnosticandprognosticinformationwouldhelpidentifyindividualswho
are at a greater risk of developing exertional heat illness. Future investigations are required to
gatherthisinformationtoimprovethepreparationofindividualspriortotrainingorcompetingin
theheat;ensuringinterventionsareimplementedtobestpreparetheindividual.
5.7. Conclusion
Inconclusion, this studydemonstrates that theRHTT is sensitive tomonitoradjustments in the
classicmarkersofheattoleranceandadaptationwithreductionsinmeasuresofpeakTre,peakTsk,
peakHR,andSWR.ThesefindingssupporttheuseoftheRHTTinfutureinvestigationstoassess
chronic heat-alleviating interventions. Furthermore, these findings could be used to support
optimalpreparation,withindividualisedstrategiesdevelopedbaseduponanindividual’sinitialheat
toleranceandtheiradaptivecapacity,priortotrainingandcompetingintheheat.TheRHTThas
the potential to be used to track changes in heat tolerance assisting decisionmaking about an
athlete’sabilitytotrainandcompeteinhighambientconditions,whilstadoptingamorefavourable
protocolforendurancerunners,duetotherunningmodeofexerciseadministered.

118
6. Acomparisonofmaleandfemaletemporalpatterningtoshort-andlong-termheatacclimation
________________________________________________________________________________
Study1(Chapter4)andStudy2(Chapter5)haveestablishedtheRHTTasareliable,modespecific
(Study1)andsensitivetest(Study2).Thereisapaucityofdataonfemales’responsestoHA.This
chapterwill provide original data onmales and females’ responses to HA using the RHTT as a
methodtoevaluatechangesinheattolerance.
6.1. Abstract
Thecurrentstudyassessedthesexdifferencesinthermoregulatoryandphysiologicaladaptationto
STHA (5d)andLTHA (10d).Sixteen (8M;8F)participantsperformedthreeRHTT,precedingHA
(RHTT1),followingSTHA(RHTT2)andLTHA(RHTT3).TheRHTTinvolved30-minrunning(9km.hr-1,
2%gradient)in40°C,40%RH(WBGT=33°C).FollowingSTHA,Trerest(Males:-0.24±0.16°C,p≤
0.001;Females:-0.02±0.08°C,p=0.597),peakTre(Males:-0.39±0.36°C,p≤0.001;Females-0.07
±0.18°C,p=0.504),andpeakHR(Males:-14±12beats.min-1,p≤0.001;Females:-5±3beats.min
-
1,p=0.164)reducedinmales,butnotfemales.FollowingSTHA,SWRBSAincreased(428±269g.hr
-
1.m
2,p=0.029)infemales,butnotmales(-11±286g.hr
-1.m
2,p=0.029).FollowingLTHA,Trerest
(Males:-0.04±0.15°C,p=0.459;Females:-0.22±0.12°C,p≤0.01)andpeakTre(Males:-0.05±
0.26°C,p=0.590;Females:-0.41±0.24°C,p≤0.01)reducedinfemales,butnotmales.Following
LTHA,SWRBSAincreasedinmales(308±346g.hr-1.m
2,p=0.029),butnotfemales(44±373g.hr
-
1.m
2, p = 0.733). Males and females responded to STHA; however, females required LTHA to
establishreductionsinthermoregulatoryandcardiovascularstrain.ThesefindingssuggestthatHA
protocolsshouldbedesignedtotargetsexdifferencesinthermoregulationforoptimaladaptation.
6.2. Introduction
Increasingambienttemperatureisknowntohaveadetrimentaleffectonenduranceperformance
(GallowayandMaughan,1997).Duringprolongedsubmaximalexerciseinhighambientconditions,
thereisagreaterrequirementforheatlossduetoeitherarateofheatgainfromtheenvironment,
oralowergradientfordryheatloss,typicallyresultinginagreaterchangeinbodyheatcontent
comparedtotemperateconditions.Manyathletes,soldiers,andmanualoperativesexposedtohigh
ambient conditionsare susceptible toheat illnesses; includingheat cramps,heat syncope,heat
exhaustion, and heat stroke. Prior to a heat illness, individuals vary in their ability to tolerate

119
exerciseheatstress,somedemonstratingadecreasedcapabilitytodissipateheatunderthesame
exerciseheatstress(Epstein,1990).Theseindividualsarecharacterizedbyanearlierandgreater
rise in Tc, greater storageofmetabolicheat, greaterphysiological strain, and reduced sweating
sensitivitywhenexercisingintheheat(Epsteinetal.,1983;Moranetal.,2004).
Malesandfemalesdifferintheirthermoregulatoryresponsestoexercise-heatstress,largelydue
to females having a reduced sudomotor function (Gagnon and Kenny, 2011) thus decreasing
evaporative heat loss capacity,with the resultant increase in physiological strain (Moran et al.,
1999). Ithasbeenshownthatmalesandfemalesdisplaysimilarratesofheatdissipationat low
requirements for heat loss. However, sex differences in sudomotor function have been
demonstratedbeyondacertainrequirementforheatloss(GagnonandKenny,2012).Incontrast,
whenmalesandfemalesdisplaysimilarheatlossforagivenheatproduction,femalesmaydisplay
ahigherchangeinTcduetophysicalcharacteristics(Gagnonetal.,2009;Havenith,2001).These
resultssuggestthatfemalesmayreachhyperthermiclevels inashortertimeperiodthanmales,
consequently females have beenmore frequently diagnosed as heat intolerant comparedwith
males (Druyanet al., 2012), potentiallyputting themat greater riskofobtainingaheat-related
illness.Theobservedsexdifferencesinthermoregulationarenotalwaysevident,butthedifference
maybecomemoreevidentastheheatstressincreases(GagnonandKenny,2012).Furthermore,
hormonal fluctuations associated with the menstrual cycle are suggested to modify central
regulatory mechanisms for thermoregulation (Inoue et al., 2005). Elevated progesterone
concentrations during the luteal phase of themenstrual cycle, have been reported to increase
resting Tc by ~0.34°C, the Tc onset threshold for sweating by 0.29°C, and the Tc threshold for
cutaneousvasodilationby0.23-0.30°C(Inoueetal.,2005).
Heatacclimation improvesheat transfer from thebody’s core to the skinandultimately to the
external environment, serving to attenuate physiological strain and improve exercise capacity
(Lorenzoetal.,2010;Sunderlandetal.,2008). HAreducesheatstorage,partiallyasa resultof
adaptationstothesudomotorfunctioncausingan increase inwholebodyevaporativeheat loss
(Poirier et al., 2015). Additionally, HA increases blood volume preserving stroke volume and
reducingHRatagivenworkload(Franketal.,2001;LorenzoandMinson,2010).
There is a dearth of literature assessing female responses to HA. Previously, the physiological
responses ofmales and females to 10-d fixed intensityHAwere assessedwith females initially
exhibiting lower Tre and HR, despite a lower SWR compared withmales (Avellini et al., 1980).
Following HA, the physiological strainwas similar betweenmales and females, althoughmales
maintained a greater SWR. This study adopted a traditional HA protocol which resulted in a

120
progressivedeclineintheadaptationstimulusoverthedurationofHA.Controlledhyperthermia
ensures consistent potentiating stimuli for adaptation throughout the HA period, eliciting
reductionsinthermalstrainandincreasesinworkcapacityduringbothSTHA(Garrettetal.,2012,
2009)andLTHA(Pattersonetal.,2004a),potentiallypromotingmorecompleteadaptation(Taylor
and Cotter, 2006). It remains unknown the extent to which females adapt to the controlled
hyperthermiamodelofHAandwhethertheirresponsesdiffertothatofmales.
HeatacclimationisoftenseparatedintoSTHA(≤8d)andLTHA(≥10d)(Garrettetal.,2011).STHA
is a preferred regime, as it provides less disruption of quality training prior to competition.
Approximately70%ofadaptationshavebeendemonstratedtooccurfollowingSTHA,evidencedby
reductionsinthermoregulatoryandcardiovascularstrain,combinedwithanimprovedsudomotor
function (Poirier et al., 2015).Acknowledgingpreviousobservations thatmales typically have a
superiorsudomotorfunctioncomparedwithfemales(GagnonandKenny,2011;Inoueetal.,2005),
itmaybeexpectedthatfemalesachievesuperiorsudomotoradaptationfollowingSTHAcompared
withmales.However,Sunderlandandcolleagues(2008)onlyachievedpartialHAintrainedfemales
witha33%increaseinintermittentsprintperformanceintheheat,despitenoalterationsinclassic
indicatorsofHAincludingHR,Tre,andSWR,followingSTHA.Thesetypicaladaptiveresponseshave
beenpreviouslyobserved intrainedmalesfollowingSTHA(Buonoetal.,1998;Fujiietal.,2012;
Garrettetal.,2011;Poirieretal.,2015;Racinaisetal.,2012);suggestingfemalesmayrequireLTHA
toachieveadaptation.Duetotheself-pacedexerciseadministeredpreandpostHA,participants
wereexercisingatahigherabsolute intensity followingHA,suggestingan increase inmetabolic
heatproduction;potentiallynegatinganyimprovementsinthermoregulationachievedthroughHA.
ResearchisrequiredtodeterminetheextenttowhichfemalesadapttoSTHAwhenusingafixed
intensityHTTtomonitoradaptations.
The primary aim of this study was to compare males and females’ thermoregulatory,
cardiovascular,andsudomotoradaptationtoSTHAandLTHAusingthecontrolledhyperthermia
modelofHA.Theprimaryhypothesis is thatmaleswillachievegreaterreductions inTreandHR
following STHA compared with females. The secondary hypothesis is that there will be no
differencesinthereductioninTreandHRbetweenmalesandfemalesfollowingLTHA.Thetertiary
hypothesis is that females will achieve a greater increase in SWR following STHA and LTHA
comparedwithmales.
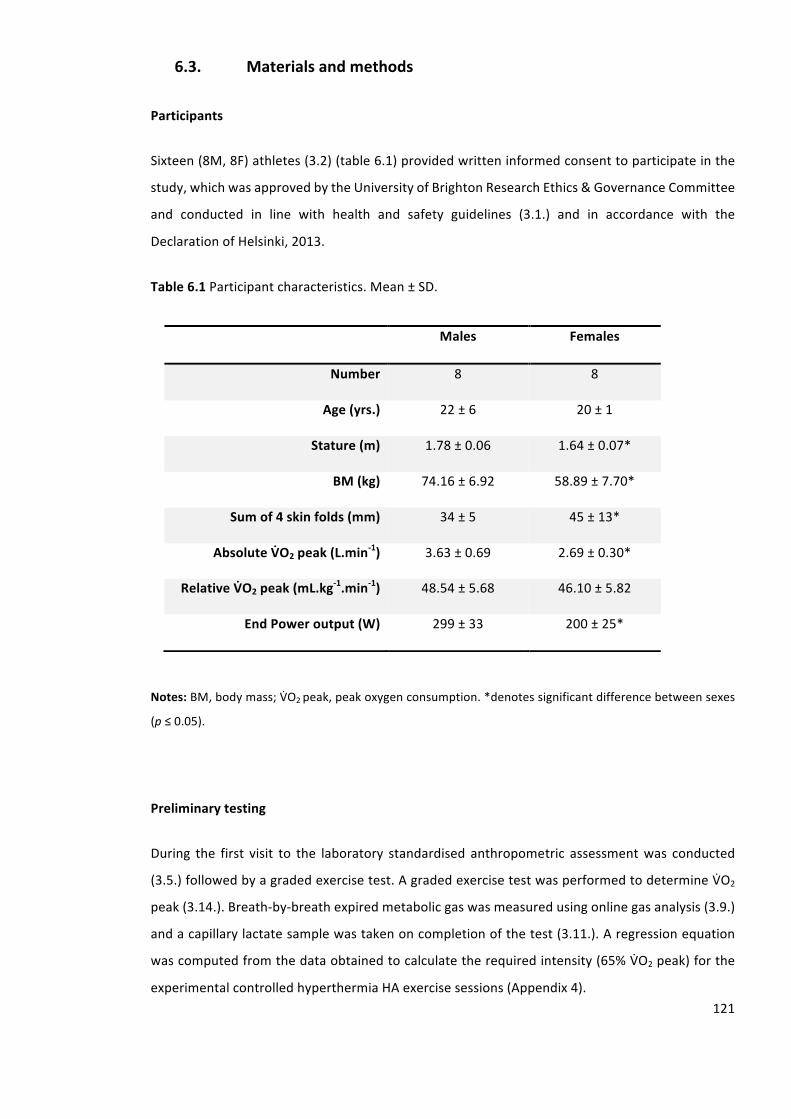
121
6.3. Materialsandmethods
Participants
Sixteen(8M,8F)athletes(3.2)(table6.1)providedwritteninformedconsenttoparticipateinthe
study,whichwasapprovedbytheUniversityofBrightonResearchEthics&GovernanceCommittee
and conducted in line with health and safety guidelines (3.1.) and in accordance with the
DeclarationofHelsinki,2013.
Table6.1Participantcharacteristics.Mean±SD.
Notes:BM,bodymass;VnO2peak,peakoxygenconsumption.*denotessignificantdifferencebetweensexes
(p≤0.05).
Preliminarytesting
During the first visit to the laboratory standardised anthropometric assessmentwas conducted
(3.5.)followedbyagradedexercisetest.AgradedexercisetestwasperformedtodetermineVnO2
peak(3.14.).Breath-by-breathexpiredmetabolicgaswasmeasuredusingonlinegasanalysis(3.9.)
andacapillarylactatesamplewastakenoncompletionofthetest(3.11.).Aregressionequation
wascomputedfromthedataobtainedtocalculatetherequiredintensity(65%VnO2peak)forthe
experimentalcontrolledhyperthermiaHAexercisesessions(Appendix4).
Males Females
Number 8 8
Age(yrs.) 22±6 20±1
Stature(m) 1.78±0.06 1.64±0.07*
BM(kg) 74.16±6.92 58.89±7.70*
Sumof4skinfolds(mm) 34±5 45±13*
AbsoluteVSO2peak(L.min-1) 3.63±0.69 2.69±0.30*
RelativeVSO2peak(mL.kg-1.min-1) 48.54±5.68 46.10±5.82
EndPoweroutput(W) 299±33 200±25*

122
Experimentaldesign
Testing was completed over a 17-d period. The timing of testing for female participants was
conducted in linewith themenstrual cycle standardisation guidelines (3.6.). Three participants
wereusingoralcontraceptive(Yasmin(30µgethinylestradiol/3mgdrospirenone).Alltrialswere
conducted in linewith pre-trial diet and exercise standardisation (3.4.). Participants provided a
urinesampleonarrivaltoensureadequatehydration(3.4.).Participantsperformed10HAsessions
(3.14.)separatedbythreeRHTT(3.16.).ThefirstRHTTwasperformed48-hrspriortobeginningHA
(RHTT1),thesecond48-hrsfollowing5-dHA(RHTT2)andthethird48-hrsfollowingtenHAsessions
(RHTT3). All experimental trials were performed in a hot environment (3.7.). Exercise was
terminatedinlinewithUniversityguidelines(3.1.).Therewerenoincidencesofearlytermination
ofexerciseduringtheRHTTwithinthisstudy.
Runningheattolerancetest
TheRHTT(3.16.)wasperformedonthreeoccasions.Followinga20-minstabilisationperiodina
temperate environment (3.7.), measures of Tre, Tsk, (3.8.) and HR (3.9.) were recorded, and
participantsenteredtheenvironmentalchamber.ToweldriednudeBMwasrecordedtocalculate
SWR (3.12.). Throughout the test Tre, Tsk, and HR were recorded at 5-min intervals. Mean Tsk
(Equation2.4)wassubsequentlycalculated.
Heatacclimation
HAinvolvedtwoblocksoffiveconsecutivecontrolledhyperthermiasessionsseparatedby48-hrs
(3.14.).
Statisticalanalyses
AlldatawerefirstcheckedfornormalityusingShapiro-Wilkandcorrectedforsphericityusingthe
Greenhouse Geisser method. An independent samples t-test was used to identify differences
betweenmaleandfemalecharacteristics.Atwo-way(3x2)mixeddesignANOVAwasperformed
toidentifydifferencesbetweentheperformanceandphysiologicalcharacteristicsduringSTHAand
LTHA,thephysiologicalresponsesonD1,D5andD10ofHAandthephysiologicalresponsesduring
RHTT1,RHTT2andRHTT3 (3.17.4.).Whenamaineffector interactioneffectwas found, results
werefollowedupusingaBonferronicorrectedposthoccomparison(3.17.6.).Effectsizes(partial
Eta squared (np2)) were calculated to analyse the magnitude and trends of the interventions

123
(3.17.5.). All data was analysed using a standard statistical package (SPSS version 20.0), and
reportedasmean±SD.Statisticalsignificancewasacceptedatthelevelofp≤0.05.
6.4. Results
Performanceresponsesduringheatacclimation(D1-D5andD5-D10)
Table6.2presents themean±SDdata for theperformanceandphysiological responsesduring
STHAandLTHA.Allparticipantscompletedten,90-minHAsessions.ANOVArevealedamaineffect
ofHAphaseonexerciseduration(F (1,14)=7.728,p=0.015,np2=0.356).Exercisedurationwas
lower in STHA (70 ± 8min) comparedwith LTHA (75 ± 7min). Therewasno interaction effect
betweenHAphaseandsexforexerciseduration(F(1,14)=0.340,p=0.569,np2=0.024).
TherewasamaineffectofHAphaseonexerciseintensity(F(1,14)=4.710,p=0.048,np2=0.252).
ExerciseintensitywaslowerinSTHA(57±6%VnO2peak)comparedwithLTHA(59±5%VnO2peak).
TherewasnointeractioneffectbetweenHAphaseandsexforexerciseintensity(F(1,14)=0.587,p
=0.456,np2=0.04).
TherewasamaineffectofHAphaseontotalwork(F(1,14)=16.272,p<0.001,np2=0.538).Total
workwaslowerinSTHA(484±105kJ)comparedwithLTHA(570±124kJ).Therewasnointeraction
effectbetweenHAphaseandsexfortotalwork(F(1,14)=0.186,p=0.673,np2=0.013).
ANOVArevealedamaineffectofHAphaseondurationofTre≥38.5°C(F(1,14)=4.982,p=0.042,np2
=0.262).ThedurationofTre≥38.5°CwashigherinSTHA(49±8min)comparedwithLTHA(46±8
min).TherewasnointeractioneffectbetweenHAphaseandsexfordurationofTre≥38.5°C(F(1,14)
=0.513,p=0.486,np2=0.035).
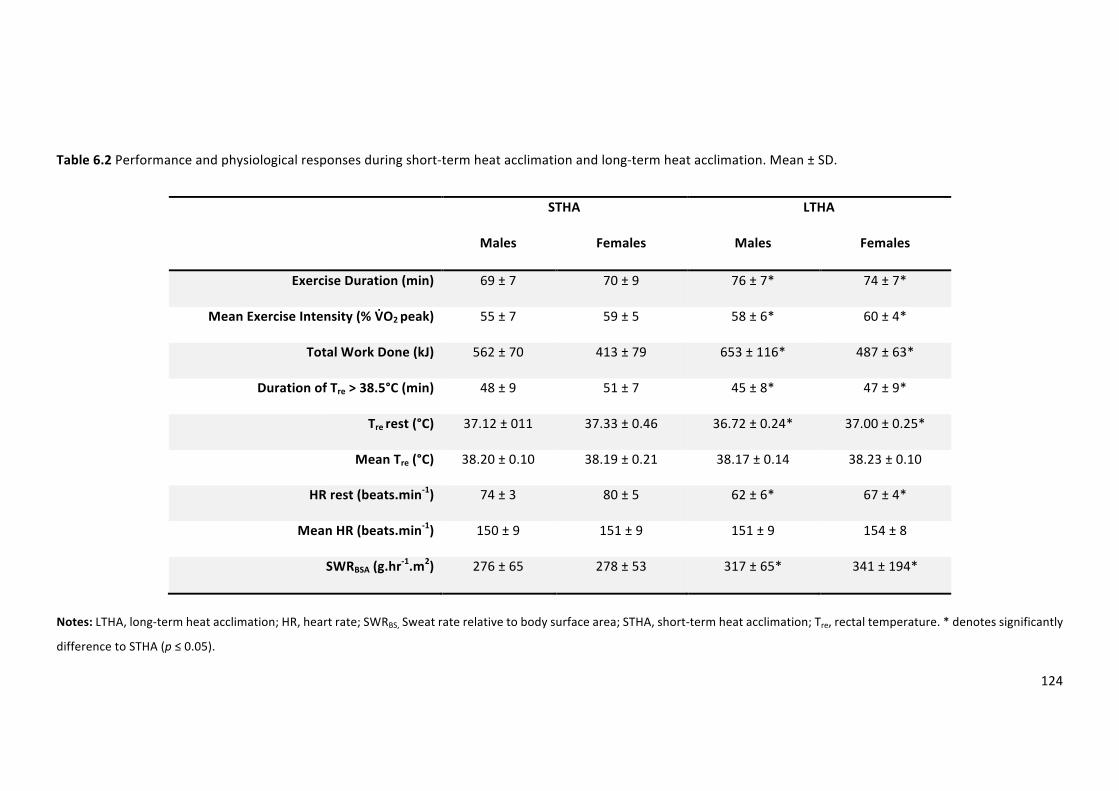
124
Table6.2Performanceandphysiologicalresponsesduringshort-termheatacclimationandlong-termheatacclimation.Mean±SD.
STHA LTHA
Males Females Males Females
ExerciseDuration(min) 69±7 70±9 76±7* 74±7*
MeanExerciseIntensity(%VBO2peak) 55±7 59±5 58±6* 60±4*
TotalWorkDone(kJ) 562±70 413±79 653±116* 487±63*
DurationofTre>38.5°C(min) 48±9 51±7 45±8* 47±9*
Trerest(°C) 37.12±011 37.33±0.46 36.72±0.24* 37.00±0.25*
MeanTre(°C) 38.20±0.10 38.19±0.21 38.17±0.14 38.23±0.10
HRrest(beats.min-1) 74±3 80±5 62±6* 67±4*
MeanHR(beats.min-1) 150±9 151±9 151±9 154±8
SWRBSA(g.hr-1.m2) 276±65 278±53 317±65* 341±194*
Notes:LTHA,long-termheatacclimation;HR,heartrate;SWRBS,Sweatraterelativetobodysurfacearea;STHA,short-termheatacclimation;Tre,rectaltemperature.*denotessignificantly
differencetoSTHA(p≤0.05).

125
Physiologicalresponsesduringheatacclimation(D1-D5andD5-D10)
Thermoregulatoryresponses
TherewasamaineffectofHAdayonTrerest(F(2,28)=37.281,p≤0.001,np2=0.727).Therewasa
reductioninTrerestfromD1toD5(-0.26±0.19°C,p≤0.001),fromD5toD10(-0.21±0.28,p=
0.002),andfromD1toD10(-0.47±0.20,p≤0.001).TherewasnointeractioneffectbetweenHA
dayandsexonTrerest(F(2,28)=1.732,p=0.195,np2=0.110).
There was no main effect of HA phase on mean Tre (F (1, 14) = 0.000, p = 0.988, np2 = 0.000).
Furthermore,therewasnointeractioneffectbetweenHAphaseandsexonmeanTre(F(1,14)=0.872,
p=0.366,np2=0.059).
Cardiovascularresponses
TherewasamaineffectofHAdayonHRrest(F(2,28)=24.137,p≤0.001,np2=0.633).Therewere
nochangesinHRrestfromD1toD5(-4±6beats.min-1,p=0.070).TherewasareductioninHRrest
fromD5toD10(-6±4beats.min-1p≤0.001)andfromD1toD10(-10±7beats.min-1,p≤0.001).
TherewasnointeractioneffectbetweenHAdayandsexonHRrest(F(2,28)=2.117,p=0.139,np2
=0.131).
There was no main effect of HA phase on mean HR (F (1, 14) = 3.059, p = 0.102, np2 = 0.179).
Furthermore, therewasno interactioneffectbetweenHAphaseand sexonmeanHR (F (1,14) =
0.716,p=0.412,np2=0.049).
Sudomotorresponses
TherewasamaineffectofHAdayonSWRBSA(F(2,28)=16.266,p≤0.001,np2=0.537).Therewasan
increasefromD1toD5(89±144g.hr-1.m2,p=0.043),fromD5toD10(87±114g.hr-1.m2,p=0.014)
andfromD1toD10(177±134g.hr-1.m2,p≤0.001).TherewasnointeractioneffectbetweenHA
dayandsexonSWRBSA(F(2,28)=2.806,p=0.077,np2=0.167).
TherewasamaineffectofHAphaseonSWRBSA(F(1,14)=21.737,p≤0.001,np2=0.608).SWRBSAwas
lower in STHA (277 ± 58 g.hr-1.m2) compared with LTHA (329 ± 79 g.hr-1.m2). There was no
interactioneffectbetweenHAphaseandsexonSWRBSA(F(1,14)=0.987,p=0.337,np2=0.066).

126
Thermoregulatoryresponsetoshort-termandlong-termheatacclimation
Table6.3presentsthemean±SDdataformalesandfemalesphysiologicalresponsesduringRHTT1,
RHTT2,andRHTT3.TherewasamaineffectofRHTTforTrerest(F(2,28)=26.084,p≤0.001,np2=
0.651).TrerestreducedfollowingSTHA(RHTT1toRHTT2)(-0.13±0.16°C,p=0.002),LTHA(RHTT2toRHTT3)(-
0.13±0.16°C,p=0.006)andLTHA(RHTT1toRHTT3)(-0.26±0.16°C,p≤0.001).Therewasaninteraction
effectbetweenRHTTandsexforTrerest(F(2,28)=5.282,p=0.011,np2=0.274).Trerestreduced
followingSTHA(RHTT1toRHTT2)inmales(-0.24±0.16°C,p≤0.001),butnodifferenceswereobserved
infemales(-0.02±0.08°C,p=0.597).TrerestreducedfollowingLTHA(RHTT2toRHTT3)forfemales(-0.22
±0.12°C,p≤0.001),butnodifferenceswereobservedinmales(-0.04±0.15°C,p=0.459).Trerest
reducedfollowingLTHA(RHTT1toRHTT3)forbothmales(-0.28±0.17°C,p≤0.001)andfemales(-0.24±
0.17°C,p≤0.001).
TherewasamaineffectofRHTT forpeakTre (F (2,28)=17.972,p≤0.001,np2=0.532).PeakTre
reducedfollowingSTHA(RHTT1toRHTT2)(-0.23±0.32°C,p=0.018),LTHA(RHTT2toRHTT3)(-0.26±0.30°C,p
=0.008)andLTHA(RHTT1toRHTT3)(-0.46±0.36°C,p≤0.001).Therewasaninteractioneffectbetween
RHTTandsexforpeakTre(F(2,28)=3.339,p=0.050,np2=0.193).PeakTrereducedfollowingSTHA
(RHTT1toRHTT2)formales(-0.39±0.36°C,p≤0.001),butnodifferenceswereobservedinfemales(-0.07
±0.18,p=0.504).PeakTre reduced followingLTHA (RHTT2toRHTT3) for females (-0.41±0.24°C,p≤
0.001),butnodifferenceswereobserved inmales (-0.05±0.26°C,p=0.590).PeakTre reduced
followingLTHA(RHTT1toRHTT3)forbothmales(-0.44±0.45°C,p=0.005)andfemales(-0.48±0.27°C,p
=0.003).
Figure6.1presentsTredataat5-minintervalsduringtheRHTTforbothmalesandfemales.There
wasamaineffectofRHTTandtimeonTre(F(12,168)=2.343,p=0.008,np20.143).FollowingSTHA
(RHTT1toRHTT2)therewasareductioninTreat5(p≤0.001),10(p=0.008),15(p=0.027),20(p=0.007)
and25(p=0.018)min.FollowingLTHA(RHTT2toRHTT3)therewerenodifferencesinTreat5(p=0.265),
10(p=0.347),15(p=0.138),20(p=0.346)and25(p=113)min.FollowingLTHA(RHTT1toRHTT3)there
wasareductioninTreat5(p=0.002),10(p=0.005),15(p=0.006),20(p=0.002)and25(p≤0.001)
min.TherewasnointeractioneffectbetweenRHTTandtimeandsexforTre(F(12,168)=1.055p=
0.402,np2=0.070).
TherewasnomaineffectofRHTTforTrechange(F(2,28)=2.502,p=0.100,np2=0.152).Therewas
nointeractioneffectbetweenRHTTandsexobservedforTrechange(F(2,28)=0.513,p=0.604,np2
=0.035).
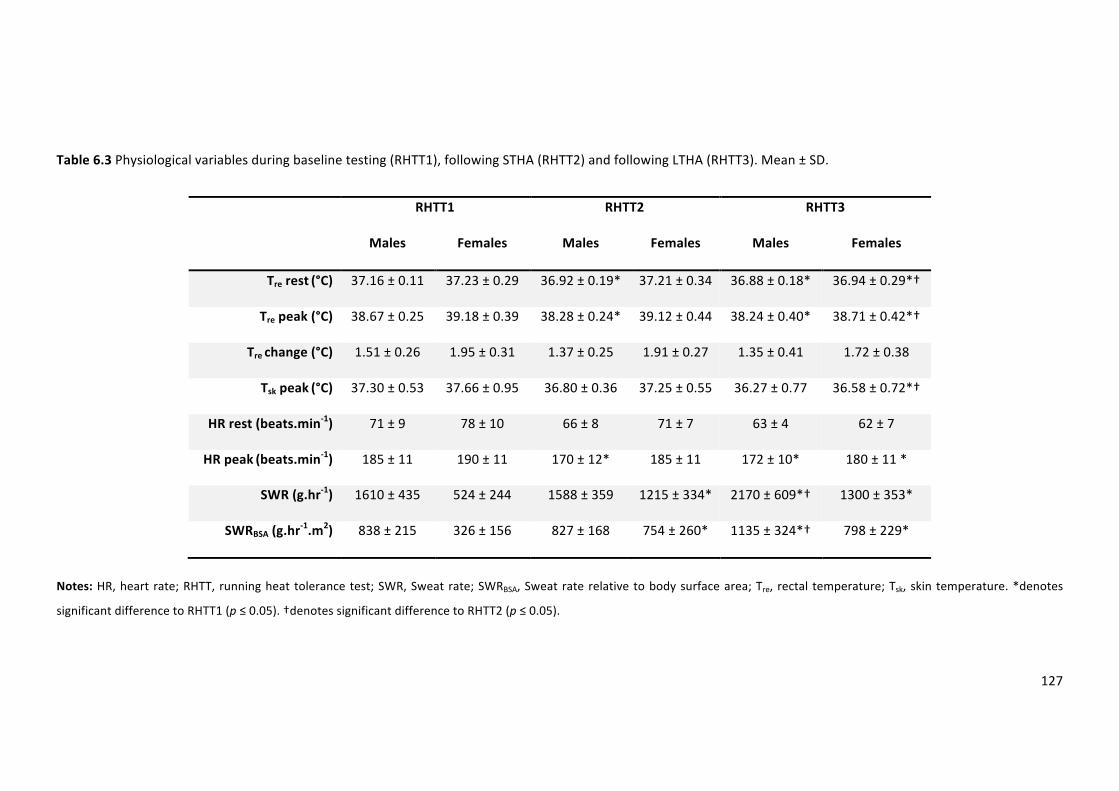
127
Table6.3Physiologicalvariablesduringbaselinetesting(RHTT1),followingSTHA(RHTT2)andfollowingLTHA(RHTT3).Mean±SD.
RHTT1 RHTT2 RHTT3
Males Females Males Females Males Females
Trerest(°C) 37.16±0.11 37.23±0.29 36.92±0.19* 37.21±0.34 36.88±0.18* 36.94±0.29*†
Trepeak(°C) 38.67±0.25 39.18±0.39 38.28±0.24* 39.12±0.44 38.24±0.40* 38.71±0.42*†
Trechange(°C) 1.51±0.26 1.95±0.31 1.37±0.25 1.91±0.27 1.35±0.41 1.72±0.38
Tskpeak(°C)37.30±0.53 37.66±0.95 36.80±0.36 37.25±0.55 36.27±0.77 36.58±0.72*†
HRrest(beats.min-1) 71±9 78±10 66±8 71±7 63±4 62±7
HRpeak(beats.min-1) 185±11 190±11 170±12* 185±11 172±10* 180±11*
SWR(g.hr-1) 1610±435 524±244 1588±359 1215±334* 2170±609*† 1300±353*
SWRBSA(g.hr-1.m2) 838±215 326±156 827±168 754±260* 1135±324*† 798±229*
Notes:HR,heart rate;RHTT, runningheat tolerancetest;SWR,Sweatrate;SWRBSA,Sweatraterelativetobodysurfacearea;Tre, rectal temperature;Tsk, skin temperature.*denotes
significantdifferencetoRHTT1(p≤0.05).†denotessignificantdifferencetoRHTT2(p≤0.05).
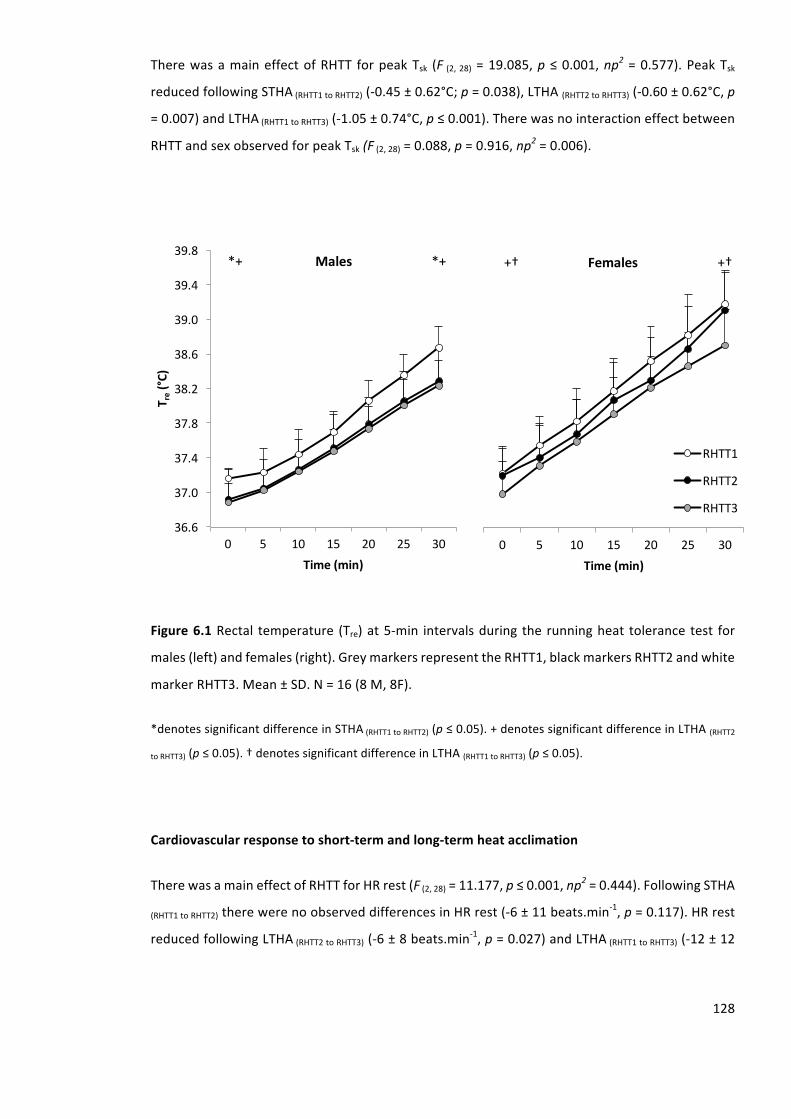
128
TherewasamaineffectofRHTT forpeakTsk (F (2,28)=19.085,p≤0.001,np2=0.577).PeakTsk
reducedfollowingSTHA(RHTT1toRHTT2)(-0.45±0.62°C;p=0.038),LTHA(RHTT2toRHTT3)(-0.60±0.62°C,p
=0.007)andLTHA(RHTT1toRHTT3)(-1.05±0.74°C,p≤0.001).Therewasnointeractioneffectbetween
RHTTandsexobservedforpeakTsk(F(2,28)=0.088,p=0.916,np2=0.006).
Figure6.1Rectal temperature (Tre)at5-min intervalsduring therunningheat tolerance test for
males(left)andfemales(right).GreymarkersrepresenttheRHTT1,blackmarkersRHTT2andwhite
markerRHTT3.Mean±SD.N=16(8M,8F).
*denotessignificantdifferenceinSTHA(RHTT1toRHTT2)(p≤0.05).+denotessignificantdifferenceinLTHA(RHTT2
toRHTT3)(p≤0.05).†denotessignificantdifferenceinLTHA(RHTT1toRHTT3)(p≤0.05).
Cardiovascularresponsetoshort-termandlong-termheatacclimation
TherewasamaineffectofRHTTforHRrest(F(2,28)=11.177,p≤0.001,np2=0.444).FollowingSTHA
(RHTT1toRHTT2)therewerenoobserveddifferencesinHRrest(-6±11beats.min-1,p=0.117).HRrest
reducedfollowingLTHA(RHTT2toRHTT3)(-6±8beats.min-1,p=0.027)andLTHA(RHTT1toRHTT3)(-12±12
36.6
37.0
37.4
37.8
38.2
38.6
39.0
39.4
39.8
0 5 10 15 20 25 30
T re(°C
)
Time(min)
*+Males *+
0 5 10 15 20 25 30
Time(min)
RHTT1
RHTT2
RHTT3
+† Females +†

129
beats.min-1,p=0.003).TherewasnointeractioneffectbetweenRHTTandsexobservedforHRrest
(F(2,28)=0.942,p=0.402,np2=0.063).
TherewasamaineffectofRHTTforpeakHR(F (2,28)=19.916,p≤0.001,np2=0.587).PeakHR
reducedfollowingSTHA(RHTT1toRHTT2)(-9±10beats.min-1,p=0.002),showednodifferencesfollowing
LTHA(RHTT2toRHTT3)(-2±8beats.min-1,p=1.000),butreducedfollowingLTHA(RHTT1toRHTT3)(-11±6
beats.min-1,p≤0.001).TherewasaninteractioneffectbetweenRHTTandsexforpeakHR(F(2,28)
=3.598,p=0.041,np2=0.204).PeakHRreducedfollowingSTHA(RHTT1toRHTT2) inmales(-14±12
beats.min-1,p≤0.001),nodifferenceswereobserved in females (-5±3beats.min
-1,p=0.164).
TherewerenodifferencesobservedforpeakHRfollowingLTHA(RHTT2toRHTT3)inbothmales(2±10
beats.min-1,p=0.505)andfemales(-5±5beats.min
-1,p=0.076).PeakHRreducedfollowingLTHA
(RHTT1toRHTT3) forbothmales(-13±7beats.min-1,p≤0.001)andfemales(-10±6beats.min
-1,p≤
0.001).
Figure6.2presentstheHR5-minintervaldataforbothmalesandfemales.Therewasnomaineffect
ofRHTTandtimeonHR(F(12,168)=0.845,p=0.604,np2=0.057).Therewasnointeractioneffect
betweenRHTTandtimeandsexforHR(F(12,168)=1.055p=0.401,np2=0.070).
Sudomotorresponsetoshort-termandlong-termheatacclimation
TherewasamaineffectofRHTTforSWR(F(2,28)=12.207,p≤0.001,np2=0.466).SWRincreased
followingSTHA(RHTT1toRHTT2)(334±590g.hr-1,p=0.042).Therewerenodifferencesobservedfor
LTHA(RHTT2toRHTT3)forSWR(334±636g.hr-1,p=0.131)however,anincreasewasobservedfollowing
LTHA(RHTT1toRHTT3)(668±529g.hr-1,p≤0.001).TherewasaninteractioneffectbetweenRHTTand
sexforSWR(F(2,28)=3.661,p=0.039,np2=0.270).SWRincreasedfollowingSTHA(RHTT1toRHTT2)in
females(691±412g.hr-1,p≤0.001),butnodifferenceswereobservedinmales(-22±533g.hr,p
=0.896).SWRincreasedfollowingLTHA(RHTT2toRHTT3)formales(583±638g.hr-1,p=0.016),butno
differenceswereobservedinfemales(85±564g.hr-1,p=0.696).SWRincreasedfollowingLTHA
(RHTT1toRHTT3)forbothmales(560±594g.hr-1,p≤0.001)andfemales(776±470g.hr
-1,p≤0.001).
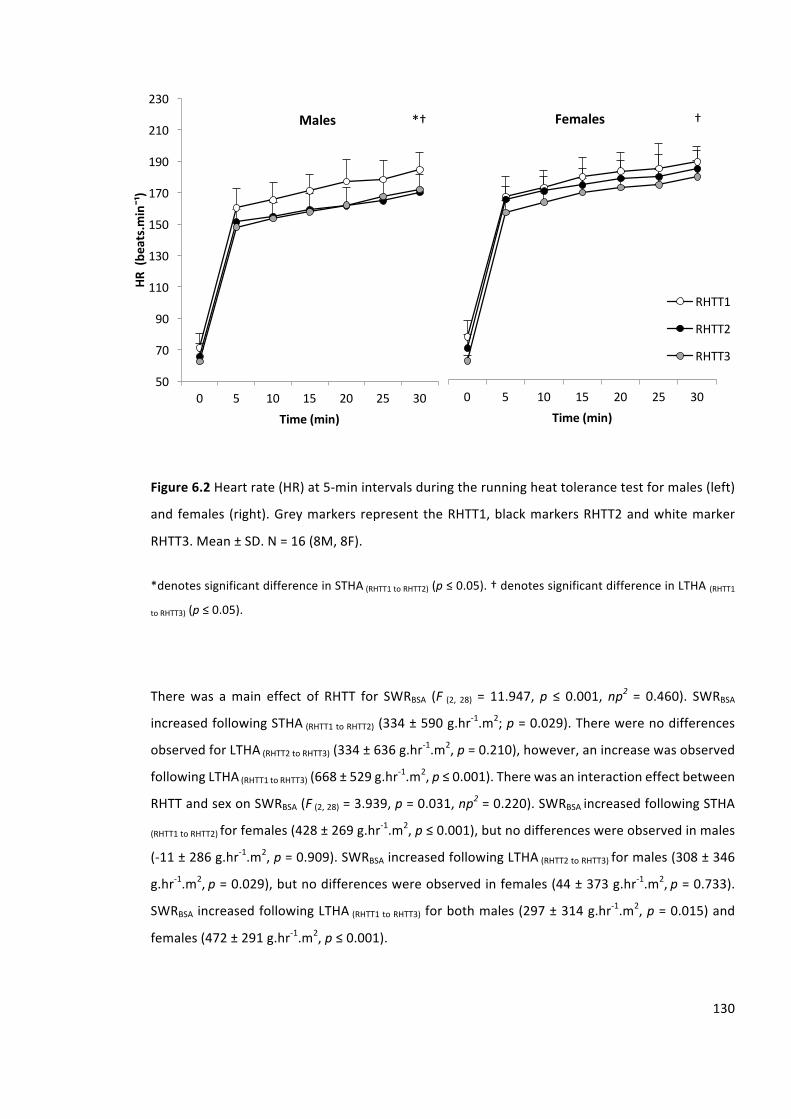
130
Figure6.2Heartrate(HR)at5-minintervalsduringtherunningheattolerancetestformales(left)
andfemales (right).Greymarkersrepresent theRHTT1,blackmarkersRHTT2andwhitemarker
RHTT3.Mean±SD.N=16(8M,8F).
*denotessignificantdifferenceinSTHA(RHTT1toRHTT2)(p≤0.05).†denotessignificantdifferenceinLTHA(RHTT1
toRHTT3)(p≤0.05).
Therewas amain effect of RHTT for SWRBSA (F (2, 28) = 11.947,p ≤ 0.001,np2 = 0.460). SWRBSA
increasedfollowingSTHA(RHTT1toRHTT2)(334±590g.hr-1.m
2;p=0.029).Therewerenodifferences
observedforLTHA(RHTT2toRHTT3)(334±636g.hr-1.m
2,p=0.210),however,anincreasewasobserved
followingLTHA(RHTT1toRHTT3)(668±529g.hr-1.m
2,p≤0.001).Therewasaninteractioneffectbetween
RHTTandsexonSWRBSA(F(2,28)=3.939,p=0.031,np2=0.220).SWRBSAincreasedfollowingSTHA
(RHTT1toRHTT2)forfemales(428±269g.hr-1.m
2,p≤0.001),butnodifferenceswereobservedinmales
(-11±286g.hr-1.m
2,p=0.909).SWRBSAincreasedfollowingLTHA(RHTT2toRHTT3)formales(308±346
g.hr-1.m
2,p=0.029),butnodifferenceswereobservedinfemales(44±373g.hr
-1.m
2,p=0.733).
SWRBSAincreasedfollowingLTHA(RHTT1toRHTT3)forbothmales(297±314g.hr-1.m
2,p=0.015)and
females(472±291g.hr-1.m
2,p≤0.001).
50
70
90
110
130
150
170
190
210
230
0 5 10 15 20 25 30
HR(be
ats.min¯¹)
Time(min)
Males *†
0 5 10 15 20 25 30
Time(min)
RHTT1
RHTT2
RHTT3
Females †

131
6.5. Discussion
The understanding of HA on experimentation using humans is primarily based upon male
participants. The application of HA to femalesmay be inappropriate due to sex differences in
thermoregulation.ThisstudyexaminedthesexdifferencesinthetemporalpatterningtoSTHAand
LTHA.Thedatademonstrates thatbothmalesand femalesachievepartialadaptation following
STHA;withmalesdemonstratinga reduction in thermoregulatoryandcardiovascular strainand
femalesdemonstratinganincreasedsudomotorfunction.FollowingLTHA,bothmalesandfemales
achievedadditionaladaptation;withfemalesdemonstratingareductioninthermoregulatorystrain
andmalesan increasedsudomotorfunction.Theseresultssuggestthatbothmalesandfemales
respondtoSTHAhowever,femalesrequireLTHAtoestablishreductionsinthermoregulatoryand
cardiovascularstrain.Thesefindingssupporttheaimofthestudywiththeprimary,secondaryand
tertiaryhypothesisalsobeingaccepted.
Short-termheatacclimation
Following STHA, approximately 70% ofmaximal adaptations has been reported to be achieved
(Poirieretal.,2015). Inthecurrentstudy,malesdemonstratedmoreadaptationfollowingSTHA
comparedwithfemales,withareductioninTrerest(-0.24±0.16°C)andpeakTre(-0.32±0.36°C);
thesechangesdidnotalterTrechange.ThemagnitudeofreductioninTreisverysimilartothe0.3°C
observedbyGarrettandcolleagues(2012)following5-dcontrolledhyperthermiaHA.Endurance
performanceismarkedlyimpairedinhotcomparedtotemperateenvironmentduetoanincrease
inTccausingadecreaseincentralactivation(NyboandNielsen,2001).AttenuationofthepeakTre
may lessen or delay the likelihood of individuals obtaining or expressing signs of heat-related
illnesseswhentraining,workingorcompetingintheheat,demonstratingtheeffectivenessofSTHA
inmales.
In thecurrentstudy,exercise intensity,exerciseduration,andtotalworkperformedwashigher
duringLTHAcomparedwithSTHA.Thisincreasedexerciseintensitywasadministeredtoelicitand
maintainthetargetTcof38.5°C.ThehigherexerciseintensityduringLTHAwouldresultinahigher
metabolic heat production compared with STHA. Since total heat loss during exercise is
predominantlyafunctionofevaporativeheatloss,agreaterrateofmetabolicheatproductionin
LTHA,withcomparableTrevaluesachieved,suggestsanincreaseinevaporativeheatlossandthus
reduced heat storage. These findings support that partial heat adaptationwas achieved during
STHA.

132
AreductionincardiovascularstrainwasachievedfollowingSTHAinmaleparticipants,evidenced
bya reduction inpeakHR. The14±12beats.min-1reduction inpeakHR is in accordancewith
previousobservationsfollowingSTHAusingcontrolledhyperthermia(Garrettetal.,2012;Patterson
et al., 2004a). The reduction in cardiovascular strain is potentially due to an increase in blood
volume,preservingstrokevolumeandreducingheattransferfromthebody’scoretotheskinand
ultimately,totheexternalenvironment.
Sunderland and colleagues (2008) assessed the effect of 4-d HA on female games players’
intermittent sprintperformance in theheat. Intermittent sprintperformance increasedby33%,
following 4-d HA, however there were no differences in peak Tre, peak HRand SWR. The self-
regulatednatureof the intermittent sprintprotocolusedprovidesnostandardisedendogenous
thermal load, potentially constraining adaptation in some individuals (Taylor andCotter, 2006).
Furthermore,anyreductioninthermoregulatoryandcardiovascularstrainmayhavebeennegated
duetoparticipantsperformingmoreworkfollowingHA.Thefindings inthecurrentstudyarein
agreementwiththesepreviousreports,withnoreductionsincardiovascularandthermoregulatory
strainduringSTHAinfemaleparticipants.
IncreasedSWRhavebeenreportedtooccurfollowingSTHA(Buonoetal.,2009;Pattersonetal.,
2004;Poirieretal.,2015).Pattersonandcolleagues(2004a)reportedthatonlysixHAsessionswere
requiredtoelicitpartialsudomotoradaptation,evidencedbyanelevatedlocalSWR.Thefindings
inthecurrentstudysupportpreviousfindingswitha428±269g.hr-1.m
2increaseinSWRinfemale
participantsfollowingSTHA.Although,femalesinthecurrentstudyhadalowerstimulusforsweat
production,duetoexercisingatalowermetabolicheatproductionpotentially,theynonetheless
improvedsweatproductiontoagreaterextentcomparedtomales,particularly followingSTHA.
Thisissurprising,basedonpreviousfindingswhichsuggestthattheproductionofsweatisthemain
driverforimprovementsinsweatingduringHA(Buonoetal.,2009).Assuch,itcouldbearguedthat
femalesadaptduringSTHAtoagreaterextentthanmales.However,theobservedincreaseinSWR
didnotresult inareducedthermalstraininfemaleparticipants.NoplateauwasobservedinTre
duringtheRHTTinfemaleparticipants,thustheenhancedSWRdidnotoffsettheuncompensatable
rateofmetabolicheatproductionobserved.
AnincreaseinwholebodySWRobservedinfemalesfollowingSTHAinthecurrentstudy,suggests
eitheranalteredafferentneuralactivityfromtheperipheralorcentralthermo-receptorscausing
differentintegrationofthermalinformation,analteredefferentneuralactivityforagivenlevelof
afferentinput,oranalteredeffectorresponse.Sexmodulatesperipheralcontrolofthesudomotor
function,thisisevidencedbyareducedthermosensitivity;resultinginfemaleshavingareduced

133
SWRcomparedwithmales(GagnonandKenny,2011).Consequently,itmaybehypothesisedthat
theenhancedsudomotorfunctioninthefemaleparticipantsfollowingSTHAinthecurrentstudyis
asaresultofperipheralchangestothethermosensitivityoftheeccrinesweatglands.Thepotential
mechanisms for this include an increased cholinergic sensitivity of the eccrine sweat gland and
increaseglandularhypertrophy(Buonoetal.,2009;LorenzoandMinson,2010).
In the current study, therewerenoobservedchanges inSWRduringSTHA (RHTT1 toRHTT2) inmale
participants. These findings may be due to a lower peak Tre in RHTT2 compared with RHTT1.
Specifically,inmaleparticipantsthepeakTreinRHTT1was38.67±0.25°CwhichproducedaSWRBSA
of838±215g.hr-1.m
2.ThepeakTreinRHTT2was38.28±0.24°CwhichproducedaSWRBSAof827±
168g.hr-1.m
2.ItiswellreportedthatanincreaseinTcstimulatessudomotorfunctionduringexercise
intheheat(Sawkaetal.,1989).Recentfindings,suggestthatwholebodySWRmayunderestimate
thetrueadaptationthatoccurredtothesweatglandfunctionfollowingHA(Buonoetal.,2009a;
Inoueetal.,1999).Buonoetal.(2009)reported20%increaseinwholebodySWR,whilepilocarpine
induced SWR increases by 63% following 8-dHA. Furthermore, Inoue et al. (1999) reportedno
changes in whole body SWR, while a significant improvement was observed for methycholine
inducedSWRfollowing8-dHA.
Long-termheatacclimation
The adaptive effects of LTHA arewell established, such that the extent towhich an individual
physiologicallyadaptstoHAisdependentuponthelengthofexposuretoheatstressconditions.In
the current study, therewasa reduction in the combined thermoregulatory and cardiovascular
strainandanenhancedsudomotorfunctionfollowingLTHAinallparticipants.Thesefindingsarein
accordance with the results reported by Avellini and colleagues (1980) when assessing sex
differencesinadaptationto10-dfixedintensityHA.Malesandfemaleswerereportedtoexpressa
similar adaptive response, evidencedby a reduction in Tre, an increased SWRand an improved
exercise capacity. Participants worked at the same absolute exercise intensity during the HA
session,therefore,theremayhavebeenvarietyinthephysiologicalstrainplaceduponparticipants.
When work rate remains constant, thermal strain during sequential exposures progressively
declines,constrainingadaptation(TaylorandCotter,2006).
Since controlled hyperthermia HA ensures equal thermal strain during every session, it can be
assumed that adaptation was not constrained during sequential sessions in the current study,
establishingmore complete adaptation (Taylor and Cotter, 2006). Additional adaptations were
observed during LTHA for the female participants, evidenced by reductions in measures of

134
thermoregulatory and cardiovascular strain from RHTT2 to RHTT3. These observed differences
werenotpresent in themaleparticipants, thus females requireLTHAtoestablish reductions in
thermoregulatoryandcardiovascularstrain.
LTHA established an improved sudomotor response in the current study. Increased SWR is not
uniquetothecurrentstudywithHAknowntoimproveperipheralandcentralmechanismsinvolved
insudomotorfunction,viaenhancedsweatglandsensitivity(Buonoetal.,2009a)andreductions
in the onset threshold for sweating, enhancing evaporative heat loss (Yamazaki andHamasaki,
2003).Completesudomotoradaptationhasbeensuggestedtotakebetween10-14dtoestablish
(ArmstrongandMaresh,1991),butthisbiphasicadaptationmaybemoreprotocoldependent.In
thecurrentstudyfemalesobtainednoadditionalbenefittothesudomotorfunctionasaresultof
LTHA,however,anincreaseinSWRwasobservedfromRHTT2toRHTT3inmaleparticipants.
6.6. Limitations
TheexerciseelicitedduringtheHAsessionswasperformedat65%ofVhO2peak.Females inthe
currentstudyhadalowerabsoluteVhO2peakcomparedwiththemaleparticipants,consequently
theyworkedatalowermetabolicheatproductionprovidingalowerstimulusforsweatproduction
(CramerandJay,2014;Gagnonetal.,2013,2008).Furthermore,femalesinthecurrentstudyhad
alowerBMcomparedwiththemaleparticipantswhichentailslessheatstorage,andthereforea
lower exercise intensity is required to increase their Tc to 38.5°C (Gagnon et al., 2009).
Consequently,thestressimposedduringtheHAsessionswaslikelylowerinfemaleparticipants,
potentially constraining adaptation and influencing the time course of adaptation, due to
inadequate endogenous heat strain. Future research is warranted to quantify these potential
differences between males and females. Future work should involve the implementation of a
controlled hyperthermia HA protocol where workload is administered using relative heat
production.ThismayfurtheroptimiseadaptationtoHAbyreducingindividualvariabilityassociated
withmetabolicheatproduction(Cramer&Jay2014)
Future work should implement greater control over hormonal alterations which alter
thermoregulatory responses associated with the menstrual cycle between repeated trials. An
elevationinprogesteroneconcentration,associatedwiththelutealphaseofthemenstrualcycle,
alters resting Tc, the threshold for sweating, and cutaneous vasodilation and consequently,
tolerancetoexerciseheatstress(Inoueetal.,2005).Inthecurrentstudy,theminimumnumberof
daystheprotocolrequiredforcompletionwas16,thuscrossingovertwomenstrualcyclephases.
Forthoseparticipantsnotusingoralcontraception,RHTT1andRHTT2wereperformedduringthe

135
follicularphaseoftheirself-reportedmenses,whenrestingTcandthethresholdfortheonsetof
sweatingandcutaneousvasodilation is lowercomparedwith the lutealphase.However,RHTT3
wasperformedduringthelutealphaseoftheirself-reportedmenses.Consequently,theextentof
the adaptation reported in females may have been smaller due to alterations in hormone
concentrationsofprogesteroneassociatedwiththemenstrualcycle(Inoueetal.,2005).Controlling
forthehormonalalterationassociatedwiththemenstrualcyclethroughoutHAwouldprovidea
greaterunderstandingintothetrueadaptationpresent.
Furthermore,inthecurrentstudy,changesinPVandfluidregulationwerenotmeasured,bothof
whichassistinthemaintenanceofanelevatedSWRandareducedcardiovascularstrainassociated
withHA(Taylor,2014).Consequently,theeffectoftheseadaptationsontheimprovementinSWR
andreductionincardiovascularstraininthecurrentstudyisnotknown.Futureresearchshouldbe
conductedobservingthesevariableswhilstaccountingforhormonalfluctuationsassociatedwith
themenstrualcycle,duetotheirknowneffectonfluidbalanceandPV.
6.7. Conclusion
Inthecurrentstudy,HAwaseffectiveinattenuatingphysiologicalstrainandimprovingexercise-
heattoleranceinbothmalesandfemalesandthusmayreducethelikelihoodofobtainingaheat-
relatedillnessduringtrainingorcompetitionintheheat.STHAisapreferredregimeforathletes
sinceitiseasiertoadoptwhensustainingqualitytrainingandtaperingperformanceintheweeks
beforecompetition.ThesefindingssuggestthatwhilstSTHAmaybeeffectiveinachievingpartial
adaptation inmalesandfemales,femalesrequireLTHAtoestablishreductions incardiovascular
andthermoregulatorystrain.
ThesefindingsprovideoriginaldatawhichwillsupporttheimplementationofHAprotocolsthat
are tailored to target sex differences in the temporal patterning of adaptation. An additional
elementofHAisanacquiredcellularthermotolerance.Study4(Chapter7)willcomparemalesand
females’cellularadaptation,specificallyHsp72mRNAacrossthecourseofSTHAandLTHA.

136
7. SexcomparisonofleukocyteHsp72mRNAtranscriptionduringheatacclimation
________________________________________________________________________________
InStudy3(Chapter6)itbecameapparentthatfemales’phenotypicresponsestoHAdifferedfrom
thoseofmales.AnadditionalelementtoHAinanacquiredcellularthermotolerance.Thisstudywill
explorethedifferencesbetweenmalesandfemales’cellularadaptationtoSTHAandLTHAusing10
outofthe16participantspresentedinStudy3(Chapter6).
7.1. Abstract
Thermotolerance is an acquired state of increased cytoprotection achieved following single or
repeatedexposurestoheatstress,inpartcharacterisedbychangesintheintracellular72kdaheat
shockprotein(HSP72;HSPA1A).FemaleshavedemonstratedreducedexerciseinducedHSP72in
comparisontomales.ThisstudyexaminedsexdifferencesinHsp72mRNAtranscriptionduringheat
acclimation(HA)toidentifywhethersexdifferenceswerearesultofdifferentialgenetranscription.
Tenparticipants(5M,5F)performed10,90-mincontrolledhyperthermiaHAsessionsover12-d.
Leukocyte Hsp72 mRNA was measured pre and post D1, D5, and D10, via RT-QPCR. HA was
evidenced by a reduction in resting Tre (-0.36 ± 0.53°C) and resting heart rate [(HR); -13 ± 7
beats.min-1] following HA (p ≤ 0.05). During HA no difference (p > 0.05) was observed in ΔTre
betweenmales(D1=1.52±0.19°C;D5=1.56±0.38°C;D10=1.80±0.28°C)andfemales(D1=1.52
±0.47°C;D5=1.40±0.22°C;D10=1.80±0.28°C).ThiswasalsotrueofmeanTredemonstrating
equalityofthermalstimuliformRNAtranscriptionandHA.Therewerenodifferences(p>0.05)in
Hsp72mRNAexpressionbetweenHAsessionsorbetweenmales(D1=+1.8±1.5fold;D5=+2.0±
1.0fold;D10=+1.1±0.4fold)andfemales(D1=+2.6±1.8fold;D5=+1.8±1.4fold;D10=+0.9±
1.9fold).ThisexperimentdemonstratesthatthereisnodifferenceinHsp72mRNAincreasesduring
HA between sexes when controlled hyperthermia HA is utilised. Gender specific differences in
exercise-inducedHSP72reportedelsewherelikelyresultfrompost-transcriptionalevents.
7.2. Introduction
Repeatedexposuretostressfulthermalenvironmentsinitiatesheatadaptationinhumans(Taylor,
2014).Heatadaptationincorporatestheinterrelatedacclimationandthermotolerance(Horowitz,
2014;Sawkaetal.,2011a).Aheatacclimatedphenotypedescribesenhancedheat losseffector
responsesandhypervolemiawhichmitigatephysiological,perceptual,andfunctionaldetriments

137
to heat exposure (Périard et al., 2015; Taylor, 2014). Thermotolerance or acquired cellular
thermotoleranceisthenomenclatureusedtodescribecellularadaptationscausedbyasingle,or
repeatedsevere,butnon-lethalheatexposure[e.g.heatacclimation(HA)](Moseley,1997).
HArepeatedlyinitiatestheheatshockresponse(HSR),typicallyincreasingvariousbasalheatshock
proteins(HSP),includingHSP72(HSP72).Inresponseto10-dHA,baselineintracellularHSP72has
beenshowntoincreaseby18%(McClungetal.,2008)whilstHsp72mRNAdemonstratesapattern
wherebytranscriptionoccurswithineachHAsession(+195%)beforereturningtobaseline24-hrs
later(Gibsonetal.,2015a,2015c).ThesetransientHAmediatedcellularadaptationstoiHSP72can
confercytoprotectiontosubsequentthermal(McClungetal.,2008)andnon-thermal(Gibsonetal.,
2015c)stressorsinvitro(McClungetal.,2008)andinvivo(Leeetal.,2016).Eloquentin-vitrodata
demonstrates that cytoprotection to stress (thermal or otherwise) is abolished when HSP72 is
knockedout(Drewetal.,2014;Leeetal.,2004;Senfetal.,2013)orblocked(Kuennenetal.,2011).
HA mediated in-vivo cytoprotection is dependent upon sufficient Hsp72 mRNA transcription
(Moranetal.,2006)andsubsequentHSP72proteintranslation(SilverandNoble,2012).
ControlledhyperthermiaHAresultsinagreaterHsp72mRNAcomparedwithmatchedtrainingin
coolconditionsasaresultofgreaterendogenousstimulifortranscription(Gibsonetal.,2015c).
Due to the consistent endogenous thermal stimulus there is an equality of Hsp72 mRNA
transcription during 10-d controlled hyperthermia HA (Gibson et al., 2015a). Thus, controlled
hyperthermiaisapreferredHAmethodcomparedwithtraditionalexogenouslyprescribedHAsince
it induces robust Hsp72 mRNA responses, ensuring sufficient and consistent increases in
endogenous stimuli throughout an in vivo chronic intervention, particularly when comparing
independentgroups.
Mortonandcolleagues (2009) reporteda sex specificHSPadaptation inhumanskeletalmuscle
following6-wks.ofcontinuousandintervaltraining.Specifically,HSP70increasedby38±41%and
23±36%followingcontinuousandintervaltrainingrespectivelyinmales(n=5);howeverfemales
(n=5)hadnochanges(3±37%and4±14%increaserespectively),despitesimilartrainingstatus,
training prescription and training adaptations (VhO2max) (Morton et al., 2009). Differential sex
responses reported by Morton et al. (2009) may be attributed to cytoprotective effects of
oestrogen.Elevatedoestrogenhasbeenshowntoaffordcellularprotection(Shinoharaetal.,2004),
accordinglyincreasedoestrogeninfemalesversusmalesmayprovideamechanismforinhibited
changesinHSP72expression(Bombardieretal.,2009).Oestrogenbindstotheoestrogenreceptor,
whichisamemberofthesteroidfamilyofnuclearreceptorsandistheoestrogenresponseelement
intargetgenes,leadingtothetranscriptionalregulationofmanygenes(Ogitaetal.,2003).Gillum

138
etal.(2013)reportedhigherintracellularHSP72concentrationsfollowingasingleboutofexercise
intheheatinmalescomparedwithfemales(inboththefollicularandlutealphaseofthemenstrual
cycle), despite similar baseline values and identical endogenous stimuli for Hsp72 mRNA
transcription(Gillumetal.,2013).Differentialsexresponseswerealsosuggestedtobearesultof
oestrogenprovidingcellularprotectionandthus,decreasingthenecessityfortranslationofHSP72
infemales.Although,stress-mediatedsexspecificdifferencesintheHSP72havebeenseen(Gillum
etal.,2013;Mortonetal.,2009),theyhavenotbeenexaminedatanmRNAlevelacrossthecourse
ofcontrolledhyperthermiaHA.
DeterminationofHsp72mRNAtranscriptioninfemaleswouldfacilitateidentificationofwhether
theinhibitedHSP72responseresultedfromabsentgenesignalling,ormitigatedproteintranslation,
potentiallyduetoelevatedoestrogen(Gillumetal.,2013;Mortonetal.,2009).Absenceofdatain
femalepopulationscouldbeproblematicforpractitionerswhomayadoptHAprotocolsthatare
informedbymechanisticcellularadaptationsfrommaleonlycohorts(Gibsonetal.,2015a;Meeet
al., 2015). Thismay reduce themagnitude to which females are protected against heat injury
(Moranetal.,2006).
The aim of the current study was to determine whether the Hsp72 mRNA response during
controlledhyperthermiaHA,differedbetweenmalesand females. Itwashypothesised that the
Hsp72mRNAresponsewillbelowerinfemalescomparedtomalesacrossthecourseofcontrolled
hyperthermiaHA.
7.3. Materialsandmethods
Participants
Based ona priori power analyses (3.17.1), four participants in each groupwould result in 95%
probabilityofdetectingadifferenceinHsp72mRNAacrossthecourseofcontrolledhyperthermia
HA(Gibsonetal.,2015a).Inlinewithpoweranalysis,andpreviousworkinthearea(Mortonetal.,
2009),ten(5M,5F)athletes(3.2)(table7.1)providedwritteninformedconsenttoparticipate.This
datawascollectedalongsideStudy3(Chapter6),howeverparticipantswerepre-determinedprior
todatacollectionfortheirinvolvementinbothstudies.ThestudywasapprovedbytheUniversity
ofBrightonResearchEthics&GovernanceCommitteeandconductedinlinewithhealthandsafety
guidelines(3.1)andinaccordancewiththeDeclarationofHelsinki,2013.
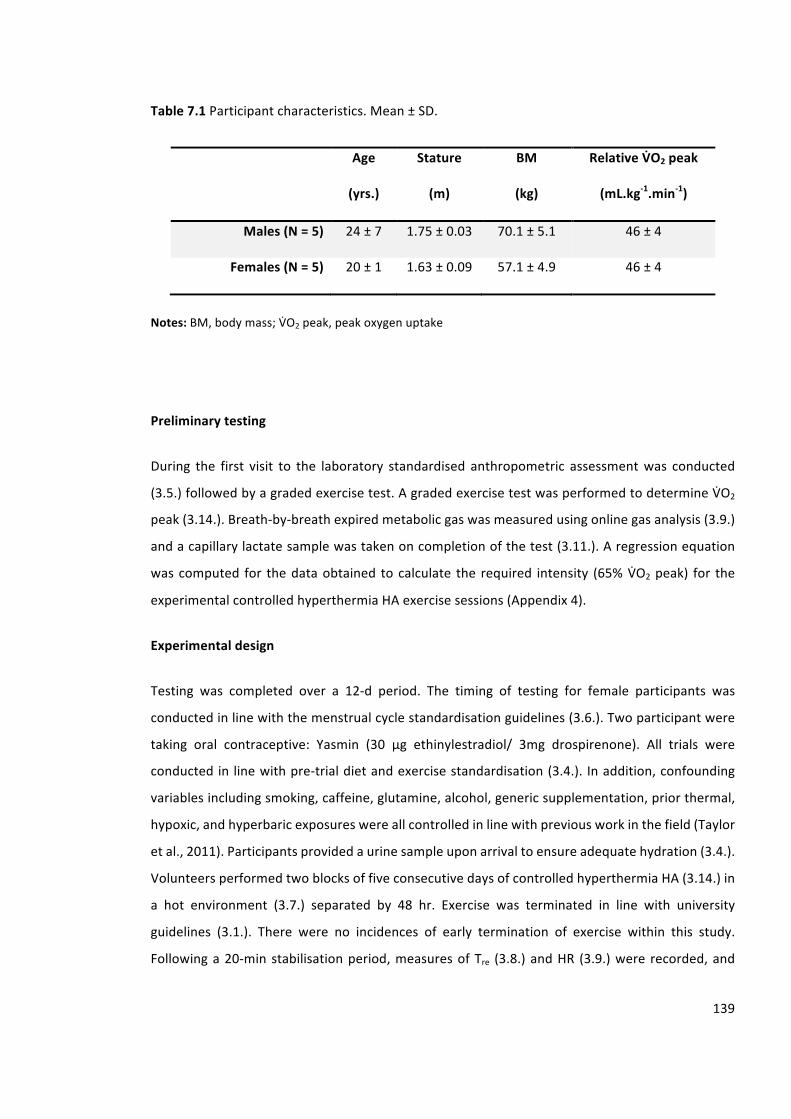
139
Table7.1Participantcharacteristics.Mean±SD.
Age
(yrs.)
Stature
(m)
BM
(kg)
RelativeVQO2peak
(mL.kg-1.min-1)
Males(N=5) 24±7 1.75±0.03 70.1±5.1 46±4
Females(N=5) 20±1 1.63±0.09 57.1±4.9 46±4
Notes:BM,bodymass;VhO2peak,peakoxygenuptake
Preliminarytesting
During the first visit to the laboratory standardised anthropometric assessmentwas conducted
(3.5.)followedbyagradedexercisetest.AgradedexercisetestwasperformedtodetermineVhO2
peak(3.14.).Breath-by-breathexpiredmetabolicgaswasmeasuredusingonlinegasanalysis(3.9.)
andacapillarylactatesamplewastakenoncompletionofthetest(3.11.).Aregressionequation
wascomputed for thedataobtained tocalculate the required intensity (65%VhO2peak) for the
experimentalcontrolledhyperthermiaHAexercisesessions(Appendix4).
Experimentaldesign
Testing was completed over a 12-d period. The timing of testing for female participants was
conductedinlinewiththemenstrualcyclestandardisationguidelines(3.6.).Twoparticipantwere
taking oral contraceptive: Yasmin (30 µg ethinylestradiol/ 3mg drospirenone). All trials were
conductedin linewithpre-trialdietandexercisestandardisation(3.4.). Inaddition,confounding
variablesincludingsmoking,caffeine,glutamine,alcohol,genericsupplementation,priorthermal,
hypoxic,andhyperbaricexposureswereallcontrolledinlinewithpreviousworkinthefield(Taylor
etal.,2011).Participantsprovidedaurinesampleuponarrivaltoensureadequatehydration(3.4.).
VolunteersperformedtwoblocksoffiveconsecutivedaysofcontrolledhyperthermiaHA(3.14.)in
a hot environment (3.7.) separated by 48 hr. Exercise was terminated in line with university
guidelines (3.1.). There were no incidences of early termination of exercise within this study.
Followinga20-minstabilisationperiod,measuresofTre (3.8.)andHR (3.9.)were recorded,and

140
participants entered the environmental chamber. Towel dried nude BM was also recorded to
calculateSWR(3.12.).ThroughouttheHAsessionsTre,HR,wererecordedat5-minintervals.
Blood sampling, RNA extraction, and one-step reverse transcription quantitative polymerase
chainreaction(RT-qPCR)
Inlinewithpreviousworkinthefield(Gibsonetal.,2015;Tuttleetal.,2015),venousbloodsamples
weretakenimmediatelybeforeandimmediatelypostexerciseheatexposureonD1,D5andD10
ofcontrolledhyperthermiaHA.Allbloodsamplesweredrawnfromtheantecubitalveininto6mL
EDTAVacuettetubes(GrenierBIO-One,Stonehouse,UK).1mLofvenousbloodwaspipettedinto
10mLof1in10redbloodcell lysissolution(10XredbloodCellLysisSolution;MiltenyiBiotech,
Bisley, UK). Samples were incubated for 15-min at room temperature then isolated via
centrifugationat400gfor5-minandwashedtwicein2mLphosphate-bufferedsalineat400gfor
5-min to isolateall leukocytes. Sampleswere then stored ina -83°C freezerand transported to
Bedfordshire University for the remainder of the analysis which was performed by a trained
laboratorytechnician.RNAwasextractedviathepreviouslyvalidatedacidguanidiumthiocyanate–
phenol–chloroform extraction method (Chomczynski and Sacchi, 1987). A standard RNA
concentrationwasproducedprior toPCrbyaddinga fixedamountofRNAtocellculturegrade
water togiveanequaldilution.Quantitywasdeterminedatanopticaldensityof260nmwhile
qualitywasdeterminedviathe260/280and260/230ratiosusingananodropspectrophotometer
(NanoDrop2000c;ThermoScientific,Waltham,MA,USA).
Hsp72mRNAwasquantifiedusingRT-QPCR.Primers(table7.2)weredesignedusingprimerdesign
software(PrimerQuestandOligoanalyzer;IntegratedDNATechnologies,Coralville,IA,USA).20μL
reactions containing 10 μL SYBR-Green RT-PCR Mastermix (Quantifast SYBRgreen Kit; Qiagen,
Manchester,UK),0.15μLforwardprimer,0.15μLreverseprimer,0.2μLreversetranscriptionmix
(QuantifastRTMix;Qiagen)and9.5μLsample(70ngRNA/μL)werepreparedinseparatetubes.
EachPCRreaction(RotorgeneQ;Qiagen)wasthenperformedas follows:10-min,50°C(reverse
transcription),5-min95°C(transcriptase inactivationandinitialdenaturation); followedby:10-s,
95°C(denaturation),30-s,60°C(annealingandextension)for40cycles.Fluorescencewasmeasured
following each cycle as a result of the incorporation of SYBR green dye into the amplified PCR
product.Melt curves (50 to95°C; rampprotocol5-s stages)wereanalysed foreach reaction to
ensure only the single gene of interest was amplified. The relative quantification of mRNA
expression foreachsamplewasassessedbydetermining the ratiobetween thecycle threshold
valueofthetargetmRNAandthecyclethresholdvaluesforβ2-microglobuinfoldchangeinrelative
mRNAexpressionwascalculatedusingthe2-ΔΔCTmethod(SchmittgenandLivak,2008).

141
Table7.2Hsp72mRNAprimersequences.
Gene NCBIAccession# Primer Sequence(5’→3’)Amplitude
length
Β2-Microglobulin
(β2-M)NM_004048
Forward
Reverse
CCGTGTGAACCATGTGACT
TGCGGCATCTTCAAACCT
91
Hsp72 NM_005345
Forward
Reverse
CGCAACGTGCTCATCTTTGA
TCGCTTGTTCTGGCTGATGT
198
Notes:NCBINationalCentreforBiotechnologyInformation
Statisticalanalysis
AlldatawerefirstcheckedfornormalityusingShapiro-Wilkandcorrectedforsphericityusingthe
GreenhouseGeissermethod.A twoway (3x2)mixeddesignanalysisofvariance (ANOVA)was
performed todeterminedifferencesbetween thephysiological andperformance characteristics
betweenD1,D5andD10inmalesandfemales(3.17.4.).Athree-way(3x2x2)mixeddesignANOVA
wasperformedtoidentifydifferencesbetweentheHsp72mRNA,preandpost,onD1,D5,andD10
of controlled hyperthermiaHAbetweenmales and females.When amain effect or interaction
effect was found, results were followed up using a Bonferroni corrected post hoc comparison
(3.17.6).Effectsizes[partialetasquared(np2)]werecalculatedtoanalysethemagnitudeandtrends
of the interventions (3.17.5.). All datawere analysed using a standard statistical package (SPSS
version20.0,IBM,Armonk,NewYork,USA)andreportedasmean±SD.Statisticalsignificancewas
acceptedatthelevelofp≤0.05.
7.4. Results
Evidenceofheatacclimation
Figure 7.1 presents the resting Tre and resting HR data for D1, D5 and D10 of controlled
hyperthermiaHA.TherewasamaineffectofdayonTrerest(F(2,16)=11.219,p≤0.001,np2=0.584).
NodifferenceswereobservedfromD1toD5(p=0.563).However,TrerestreducedfromD1toD10
(p=0.027)andfromD5toD10(p=0.003).Therewasnointeractioneffectbetweendayandsex
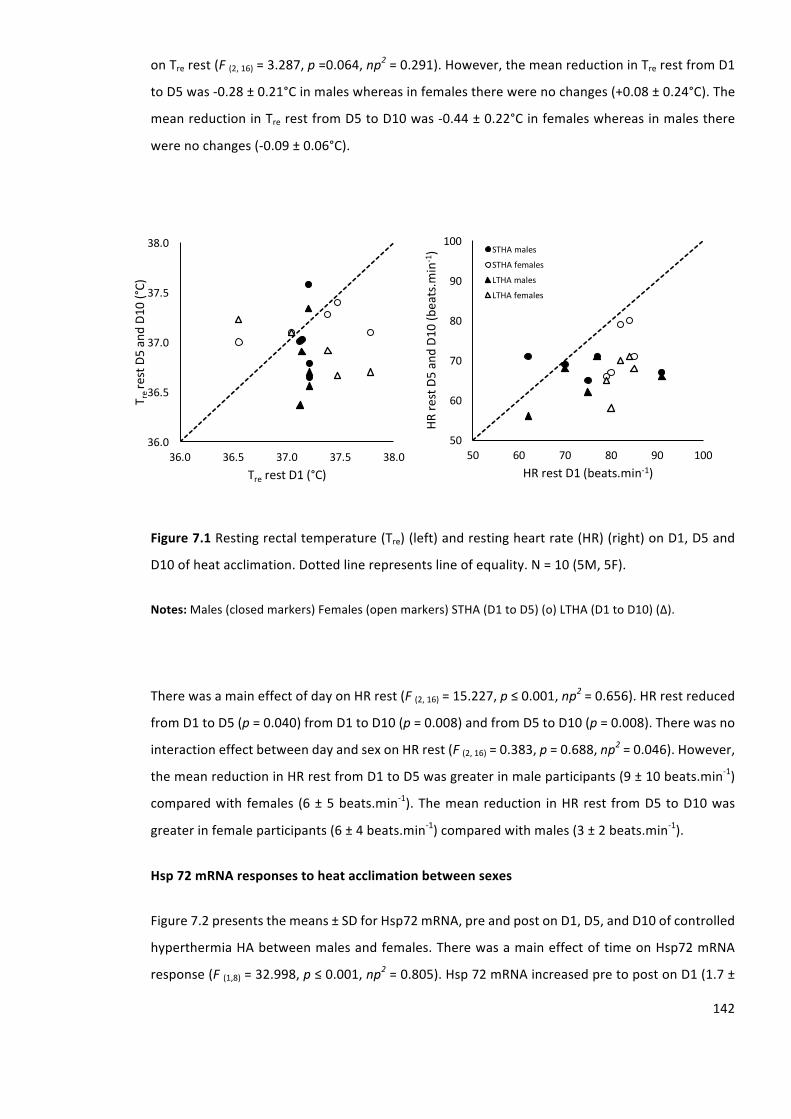
142
onTrerest(F(2,16)=3.287,p=0.064,np2=0.291).However,themeanreductioninTrerestfromD1
toD5was-0.28±0.21°Cinmaleswhereasinfemalestherewerenochanges(+0.08±0.24°C).The
meanreductioninTrerestfromD5toD10was-0.44±0.22°Cinfemaleswhereasinmalesthere
werenochanges(-0.09±0.06°C).
Figure7.1Restingrectaltemperature(Tre)(left)andrestingheartrate(HR)(right)onD1,D5and
D10ofheatacclimation.Dottedlinerepresentslineofequality.N=10(5M,5F).
Notes:Males(closedmarkers)Females(openmarkers)STHA(D1toD5)(o)LTHA(D1toD10)(∆).
TherewasamaineffectofdayonHRrest(F(2,16)=15.227,p≤0.001,np2=0.656).HRrestreduced
fromD1toD5(p=0.040)fromD1toD10(p=0.008)andfromD5toD10(p=0.008).Therewasno
interactioneffectbetweendayandsexonHRrest(F(2,16)=0.383,p=0.688,np2=0.046).However,
themeanreductioninHRrestfromD1toD5wasgreaterinmaleparticipants(9±10beats.min-1)
comparedwith females (6±5beats.min-1).Themeanreduction inHRrest fromD5toD10was
greaterinfemaleparticipants(6±4beats.min-1)comparedwithmales(3±2beats.min
-1).
Hsp72mRNAresponsestoheatacclimationbetweensexes
Figure7.2presentsthemeans±SDforHsp72mRNA,preandpostonD1,D5,andD10ofcontrolled
hyperthermiaHAbetweenmalesandfemales.TherewasamaineffectoftimeonHsp72mRNA
response(F(1,8)=32.998,p≤0.001,np2=0.805).Hsp72mRNAincreasedpretopostonD1(1.7±
36.0
36.5
37.0
37.5
38.0
36.0 36.5 37.0 37.5 38.0
TrerestD5andD10(°C)
Tre restD1(°C)
50
60
70
80
90
100
50 60 70 80 90 100
HRrestD5andD10(beats.m
in-1)
HRrestD1(beats.min-1)
STHAmales
STHAfemales
LTHAmales
LTHAfemales
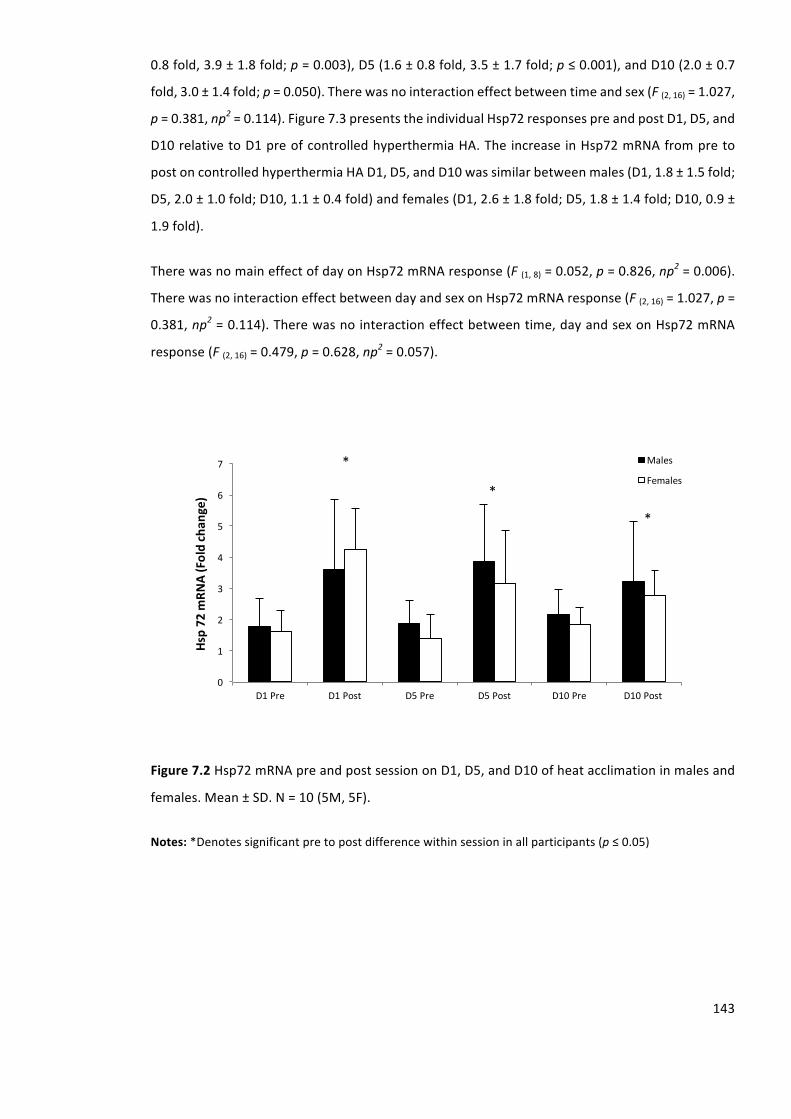
143
0.8fold,3.9±1.8fold;p=0.003),D5(1.6±0.8fold,3.5±1.7fold;p≤0.001),andD10(2.0±0.7
fold,3.0±1.4fold;p=0.050).Therewasnointeractioneffectbetweentimeandsex(F(2,16)=1.027,
p=0.381,np2=0.114).Figure7.3presentstheindividualHsp72responsespreandpostD1,D5,and
D10relativetoD1preofcontrolledhyperthermiaHA.TheincreaseinHsp72mRNAfrompreto
postoncontrolledhyperthermiaHAD1,D5,andD10wassimilarbetweenmales(D1,1.8±1.5fold;
D5,2.0±1.0fold;D10,1.1±0.4fold)andfemales(D1,2.6±1.8fold;D5,1.8±1.4fold;D10,0.9±
1.9fold).
TherewasnomaineffectofdayonHsp72mRNAresponse(F(1,8)=0.052,p=0.826,np2=0.006).
TherewasnointeractioneffectbetweendayandsexonHsp72mRNAresponse(F(2,16)=1.027,p=
0.381,np2=0.114).Therewasnointeractioneffectbetweentime,dayandsexonHsp72mRNA
response(F(2,16)=0.479,p=0.628,np2=0.057).
Figure7.2Hsp72mRNApreandpostsessiononD1,D5,andD10ofheatacclimationinmalesand
females.Mean±SD.N=10(5M,5F).
Notes:*Denotessignificantpretopostdifferencewithinsessioninallparticipants(p≤0.05)
0
1
2
3
4
5
6
7
D1Pre D1Post D5Pre D5Post D10Pre D10Post
Hsp72m
RNA(Foldchan
ge)
Males
Females
*
*
*
144
Figure7.3Hsp72mRNApretopostsessiononD1,D5andD10relativetoD1preofheatacclimation
inmales(opencircles)andfemales(closedcircles).N=10(5M,5F).
Comparableheatacclimationsessions
Table7.3presentsthemean±SDforperformanceandphysiologicalvariablesduringD1,D5,and
D10ofcontrolledhyperthermiaHA formalesand females.Therewasnomaineffectofdayon
meanTre (F (2,16) =2.536,p =0.143,np2=0.241). Furthermore, therewasno interactioneffect
betweendayandsexonmeanTre(F(2,16)=1.880,p=0.185,np2=0.190).Therewasnomaineffect
ofdayonTrechange(F(2,16)=3.042,p=0.076,np2=0.275).Furthermore,therewasnointeraction
effectbetweendayandsexonTrechange(F(2,16)=0.234,p=0.794,np2=0.028).Therewasnomain
effect of day onmeanHR (F (2, 16) = 0.488,p = 0.623,np2= 0.057). Furthermore, therewas no
interactioneffectbetweendayandsexonmeanHR(F(2,16)=0.242,p=0.788,np2=0.029).
Therewasamaineffectofdayonmeanpower(F (2,16)=11.608,p≤0.001,np2=0.592).Mean
powerwashigheronD5(p≤0.001)andD10(p=0.015)comparedtoD1.Therewerenodifferences
betweenD5andD10(p=1.000).Therewasnointeractioneffectbetweendayandsexonmean
power(F(2,16)=0.663,p=0.529,np2=0.076).
-2
-1
0
1
2
3
4
5
6
D1Pre D1Post D5Pre D5Post D10Pre D10Post
Hsp72m
RNA(fo
ldch
ange)
Males
Females
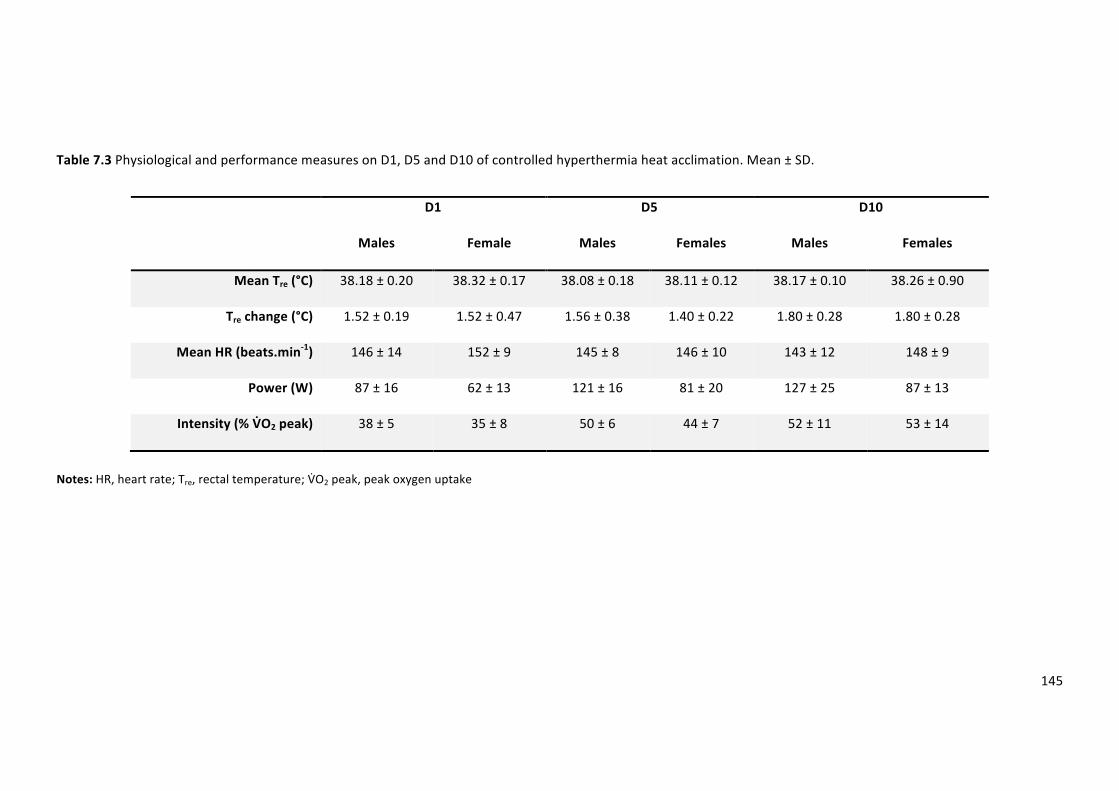
145
Table7.3PhysiologicalandperformancemeasuresonD1,D5andD10ofcontrolledhyperthermiaheatacclimation.Mean±SD.
D1 D5 D10
Males Female Males Females Males Females
MeanTre(°C) 38.18±0.20 38.32±0.17 38.08±0.18 38.11±0.12 38.17±0.10 38.26±0.90
Trechange(°C) 1.52±0.19 1.52±0.47 1.56±0.38 1.40±0.22 1.80±0.28 1.80±0.28
MeanHR(beats.min-1) 146±14 152±9 145±8 146±10 143±12 148±9
Power(W) 87±16 62±13 121±16 81±20 127±25 87±13
Intensity(%VHO2peak) 38±5 35±8 50±6 44±7 52±11 53±14
Notes:HR,heartrate;Tre,rectaltemperature;VJO2peak,peakoxygenuptake

146
Therewasamaineffectofdayonrelativeexerciseintensity(F(2,16)=21.593,p≤0.001,np2=0.730).
RelativeexerciseintensitywashigheronD5(p≤0.001)andD10(p≤0.001)comparedtoD1.There
werenodifferencesbetweenD5andD10(p=0.221).Therewasnointeractioneffectbetweenday
andsexonrelativeexerciseintensity(F(2,16)=0.1.034,p=0.378,np2=0.114).
7.5. Discussion
ThisisthefirststudytocompareHsp72mRNAexpressioninmalesandfemalesacrossthecourse
ofcontrolledhyperthermiaHA.Incontrasttoourhypothesis,thisexperimentdemonstratesthat
theHsp72mRNAresponseissimilarbetweenmalesandfemalesonD1,D5,andD10ofacontrolled
hyperthermiaHA.ThissuggeststhatsexdifferencesinHSPfollowingacute(Gillumetal.,2013)and
chronic (Morton et al., 2009) in vivo exercise bouts are due to post transcriptional events.
ControlledhyperthermiaHAresultedintypicalphenotypicadaptations,evidencedbyreductionin
resting Tre and HR across the course of controlled hyperthermia HA. Males and females
demonstrated equal physiological responses (∆Tre,mean Tre andHR) to eachHA sessionwhere
Hsp72mRNAwasmeasured. Accordingly, equality of these endogenous stimuli, both between
groups, and throughout HA elicited equal increases in Hsp72mRNA transcription. Comparable
transcription of Hsp72mRNA betweenmales and females, suggests endogenous stimuli which
inducetheHSRarethemostimportantcriteriaforincreasingHsp72mRNA(Gibsonetal.,2015a),
withnosexdependentinhibitionoramplificationintranscription.
Inthecurrentstudy,HAproducedasignificantincreaseinHsp72mRNAprovidingfurtherevidence
thatthecontrolledhyperthermiaHAmethodpresentsasufficientendogenousstresstosurpass
theminimum requirement to elicit increased transcription of Hsp72mRNA, in bothmales and
females, at the onset and culmination of discrete and repeated bouts of exercise-heat stress.
Gibsonetal.(Gibsonetal.,2015a)reportedanincreaseinHsp72mRNApretopostonD1(1.9±
0.6fold,4.9±1.1fold),D5(2.3±0.8fold,5.3±2.5fold)andD10(2.1±0.7fold,4.3±1.3fold)
duringa10-dcontrolledhyperthermiaHAprotocol inmaleparticipants.Thedata inthecurrent
studydemonstratesfemaleshaveacomparablemagnitudeofresponsetomales.Accordingly,this
data providesmechanistic support for practitioners prescribing controlled hyperthermiaHA for
femaleathletes.SustainedincreasesinHsp72mRNAthroughouttheHA,furtherdemonstratesthe
continuedstimulationofthepathwaysresponsibleforthermotolerance,i.e.theequalityofHSR,in
bothmalesandfemales.
Previously,agreaterHSP72increasehasbeenreportedinmalescomparedtofemales(Gillumet
al.,2013;Mortonetal.,2009);however,thesestudiesmeasuredtheprotein(HSP)whereasthe

147
currentstudymeasuredthegene(HspmRNA).Interestingly,thecurrentdatacontradictsParooet
al.(Parooetal.,2002)findings,whoreportedasexspecificHSRatthelevelofproteinandmRNA;
withmaleratshavingasignificantlyhigherHSP70(200%ofcontrol)andHsp70mRNA(+900%of
control) response following 60-min of exercise at 70% V\O2max when compared with females
(HSP70=100%ofcontrol;Hsp70mRNA=450%ofcontrol).Parooetal. (Parooetal.,2002)did
however provide mechanistic evidence for an interaction between HSP70, Hsp70 mRNA and
oestrogen. Ovariectomized female animals treated with a placebo demonstrated equivalent
increases in HSP72 (+150% of control) and Hsp72mRNA (+1,200% of control) tomales, whilst
endogenousoestrogenreturnedthetypicalinhibitedfemaleHSRresponse(HSP70100%ofcontrol;
Hsp70mRNA300%ofcontrol).Methodologically,Parooetal.(Parooetal.,2002)implementedthe
northernblottingtechniquewhichislesssensitivethantheRT-QPCRtechniqueusedinthecurrent
study,potentiallyexplainingthenon-significantlyincreasedHsp72mRNA.
Elevatedoestrogenaffordscellularprotection(Shinoharaetal.,2004)andthus,thiscytoprotective
pathway may inhibit changes in HSP72 translation (Bombardier et al., 2009). It is likely, that
oestrogen most greatly mediates post Hsp72 mRNA transcriptional changes which inhibit the
translationofHSP72.ThemechanismbywhichoestrogenattenuatesHSRmaybemediatedthrough
its indirect antioxidant properties by stabilising cellular membranes and attenuating oxidative
stress;suchaneffectcouldprotectthermalsensitivecellsagainstexercise-induceddamage,and
therebyresult inabluntedHSP72response(Parooetal.,2002).Thelackofobserveddifference
betweenmalesandfemalesinthecurrentstudy,maybearesultoflowoestrogenconcentrations,
whichmaynothavebeensufficienttoexertanantioxidanteffect.
7.6. Limitations
FutureworkshouldinvolvethemeasurementofHSP72proteinalongsideHsp72mRNAacrossthe
courseofcontrolledhyperthermiaHAinmalesandfemales,tohelpunderpinthetrueeffectofsex
ontheHSR;theabsenceofwhichisalimitationofthepresentstudy.Furthermore,oestrogenis
reported to have a dose dependent inhibition of HSP72 expression at the transcription level
(Shinoharaetal.,2004).FutureworkshouldinvestigatetheHSR,andsubsequentHSP72andHsp72
mRNAresponsetodiscreteandrepeatedboutsofexercise-heatstressinhighandlowoestrogen
conditionsandinpost-menopausalwomen,whonaturallyhaveloweroestrogenconcentrations.
ThisinformationwouldhelppractitionersimplementcontrolledhyperthermiaHAstrategiesthat
ensureanoptimalstimulusforcellularadaptation.

148
7.7. Conclusion
Males and females have equal Hsp72 mRNA expression throughout 10-d of controlled
hyperthermia HA. This suggests that there are no differences in the endogenous criteria to
transcribe Hsp72 mRNA via the HSR between males and females. Differences in basal HSP72
observedelsewherearethereforelikelytoresultfrominhibitedproteintranslation,potentiallydue
totheinfluenceofoestrogen.
ThisstudyexploredthecellularadaptationsinmalesandfemalesacrossthecourseofHA.Sinceno
sex differences in the cellular adaptation were observed, Chapter 8 will take a more applied
approachtoinvestigatewhetherthephenotypicadaptationscanbeacceleratedduringSTHA.

149
8. Passiveheatexposureprecedingshort-termheatacclimationacceleratesadaptationinfemales
________________________________________________________________________________
The initial experiments have established the RHTT is repeatable (Study 1), mode specific and
sensitive to changes in thermoregulatory responses (Study2). In Study3 (Chapter6) it became
apparent that females’ responses toHA differed from those ofmales, but thiswas not due to
differencesinthermotolerance(Study4).Thisstudywillexploreapotentialmethodtoaccelerate
females’responsetoSTHAusingtheRHTTtomonitorchanges.
8.1. Abstract
ThecurrentstudyassessedwhetherSTHAprecededbyapassiveheatacceleratedheatadaptation
infemales.NinefemalesperformedtwoHAinterventionsinarandomisedorderseparatedby7-
wks. A RHTTwas performed 24-hr pre (RHTT1) and 24-hr post (RHTT2) STHA. During the HACH
intervention participants preceded the HA session by 20-min seated rest in a temperate
environmentwearing shorts anda sportsbra (20°C, 40%RH;WBGT=19°C).During theHAsauna
intervention participants preceded theHA session by 20-min seated rest in a hot environment
(50°C,30%RH;WBGT>40°C),wearingshortsandsportsbraanda100%vinylsaunasuit.Trerest(-
0.28±0.16°C),peakTre(-0.42±0.22°C),HRrest(-10±4beats.min-1),peakHR(-12±7beats.min-1),
Treattheonsetofsweating(-0.29±0.17°C)reduced(p≤0.001)fromRHTT1toRHTT2inHAsauna;
butnotHACH.TS(p=0.002)andRPE(p≤0.001)reducedfromRHTT1toRHTT2inHAsauna;butnot
HACH.PlasmavolumeexpansionwasgreaterfollowingHAsauna(9.3±7.6%)comparedtoHACH(1.3±
5.0%)(p=0.013).Sweatrateincreased(p≤0.001)andsweatsodiumchloride(NaCl)concentration
reduced (p = 0.006) in both conditions. HAsauna was effective in attenuating thermoregulatory,
cardiovascularandperceptualstrain;thiswasnotachievedfollowingHACH.ThisnovelHAstrategy
providesatime-efficientmethodtostimulateHAinfemales.
8.2. Introduction
Heat acclimation improves thermal comfort (Gonzalez and Gagge, 1976), submaximal exercise
performance, and increases maximal aerobic capacity in the heat (Lorenzo et al., 2010). The
benefits arise from enhanced sudomotor and skin blood flow responses, PV expansion,
cardiovascularstabilityandanimprovedfluidbalance(Poirieretal.,2015).Asaresult,HAisthe
consensusrecommendationstrategytoattenuatethephysiologicalstrainassociatedwithtraining

150
and competing in the heat (Racinais et al., 2015). However, there remains a paucity of data
concerningbestpractice forHA in females;consequently, femaleathletesare implementingHA
strategiesbasedondatacollectedfrommaleparticipants.Theimplicationsofthisarethatfemales
maybeadoptingsub-optimalstrategiesthatprovidelittlebenefittoalleviatingphysiologicalstrain.
Heatacclimation isoftenseparated intoSTHA (≤8-d)andLTHA (≥18-d).Approximately70%of
maximal adaptations occur following STHA evidenced by reductions in thermoregulatory and
cardiovascularstrain(Poirieretal.,2015),combinedwithanimprovedsudomotorfunction(Buono
etal.,2009b).Study3(Chapter6)presentedevidencetosuggestthatfemalesachieveonlypartial
adaptationfollowingSTHA,evidencedwithanincreaseinwholebodySWRdespitenochangesin
other classic markers of HA. Moreover, females required LTHA to establish reductions in
cardiovascularandthermoregulatorystrain.Thesefindingsarefurthersupportedbytheworkof
Sunderlandetal.(2008)whoreportednodifferenceinTreandHRfollowing4-dHAinfemales.The
absenceofevidenceconcerningeffectiveSTHAprotocols in females isproblematic forathletes,
since STHA is a preferred regime as it provides less disruption from quality training prior to
competitionthanmoretraditionalHAprotocols(Gibsonetal.,2015b).
Recently,theconceptofcombiningpassiveheatexposureswithtemperatetraininghasreceived
considerableinterest.Scoonetal.(2007)andStanleyetal.(2014)reporteda7.1%and17.8%PV
expansion,respectively,whentemperatetrainingwasfollowedbya30-minsauna(87°C)exposure.
Furthermore,Stanleyetal.(2014)reportedthatPVreachedpeakexpansion(+17.8%)followingfour
exposures;highlightingtheeffectivenessofshort-termsaunaexposure.Multiplesaunaexposures
appear to specifically augment PV expansion, which may contribute to improved myocardial
efficiency(Horowitzetal.,1986b),increasedventricularcompliance(Horowitzetal.,1986a)and
improvedmaximalcardiacoutput(Lorenzoetal.,2010);allofwhichmaytranslateintoimproved
physicalperformance.Thus,itseemsplausiblethatcombiningpassiveheatexposurewithHAmay
resultinanacceleratedadaptationinfemalesthroughtargetingPVexpansion.
Despite observing an increase in SWR in females following STHA in Study 3 (Chapter 6), SWR
remained10% lower thanmales’baselineSWR.These findingswereevidentevenwith females
potentiallyworkingatahighermetabolicheatproductionandthusevaporativerequirement,due
tothefixedintensityprotocol.Duringuncompensableheatstress,theevaporativerequirementfor
heatbalanceexceedsthemaximumpossibleheatloss(Montainetal.,1994).Protectiveclothing
exacerbates the challenge of thermoregulation because of limited water vapour permeability
across the clothing layers, decreasing the rate of heat exchange (Cheung et al., 2000). When
wearingclothing that restricts sweatevaporation,muchof thesweatmaybecomeabsorbedor

151
trapped within the clothing, altering the rate of heat transfer. Thus, it seems plausible that
stimulatingsudomotorfunctionwhilstrestrictingsweatevaporationpriortoaHAsessionthrough
theuseofprotectiveclothingmayprovidemorecompletesudomotoradaptationduetogreater
thermalburden,withoutadditionaltrainingload.
STHAoffersamoreecologicallyvalidmethodforathletescomparedwithLTHA,thusitisessential
toestablisheffectiveSTHAmethodsforafemalepopulationwhilstlimitingtheexercisestimulus.
ThisstudyaimedtoinvestigatewhetherSTHA,precededbyapassiveheatexposure,designedto
target PV expansion and sudomotor adaptation, accelerated heat adaptation in females. The
primaryhypothesiswas thatSTHAprecededbyapassiveheatexposure (HAsauna)will result ina
greaterplasmavolumeexpansioninfemalescomparedwithHAalone.Thesecondaryhypothesis
isthatHAsaunawillproduceagreatersudomotoradaptationinfemalescomparedwithHAalone.
8.3. Materialsandmethods
Participants
Ninefemaleathletes(3.2)(Mean±SD;age,22±4yrs.;BM,59.33±11.55kg;stature,1.64±0.01
m;BSA,1.6±0.2m2;sumof4skinfolds,49±11mm;V\O2peak,50±4mL.kg-1.min-1)provided
writteninformedconsenttoparticipateinthecurrentstudywhichwasapprovedbytheUniversity
ofBrightonResearchEthics&GovernanceCommitteeandconductedinlinewithhealthandsafety
guidelines(3.1)andinaccordancewiththeDeclarationofHelsinki,2013.
Preliminarytesting
During the first visit to the laboratory standardised anthropometric assessmentwas conducted
(3.5.). A graded exercise test using amotorised treadmill was performed to determine lactate
threshold andV\O2 peak (3.15.). In addition,V\O2 peakwasdeterminedusing a cycle ergometer
(3.14.). A regression equation was computed for the data obtained to calculate the required
intensity (65% V\O2 peak) for the experimental controlled hyperthermia HA exercise sessions
(Appendix4).Breath-by-breathexpiredmetabolicgaswasmeasuredusingonlinegasanalysis(3.9.).
Acapillarylactatesamplewascollectedandanalysedattheendofeachstageduringthelactate
thresholdtestandoncompletionoftheV\O2peaktest(3.11.).

152
Experimentaldesign
The timing of testing for female participants was conducted in line with the menstrual cycle
standardisationguidelines(3.6.).Fourparticipantsweretakingoralcontraceptive:twousedYasmin
(30µgethinylestradiol/3mgdrospirenone)andtwousedMicrogynon(30µgethinylestradiol/150
µglevonorgestrel).Hormonalstatuswasconfirmedbytakingavenousbloodsampleonthefirst
andfinaldayofeachexperimentalintervention.Alltrialswereconductedinlinewithpre-trialdiet
and exercise standardisation (3.4.). Participants provided a urine sample upon arrive to ensure
adequate hydration (3.4.). All experimental trials were performed in a hot environment (3.7.).
Exercisewasterminatedinlinewithuniversityguidelines(3.1).Therewerenoincidencesofearly
terminationofexerciseduringtheRHTTwithinthisstudy.
Runningheattolerancetest
Participants performed twoRHTT (3.16.) on amotorised treadmill (3.15.) separated by fiveHA
sessions.ThefirstRHTTwasperformed24-hrspriortobeginningHA(RHTT1)andthesecond24-
hrsfollowing5-dHA(RHTT2).Followinga20-minstabilisationperiodinatemperateenvironment
(3.7.),measures of Tre, Tsk (3.8.), HR (3.9.), RPE, and TS (3.13.)were recorded, and participants
enteredtheenvironmentalchamber.ToweldriednudeBMwasalso recordedtocalculateSWR
(3.12.).ThroughoutthetestTre,Tsk,andHRwererecordedat5-minintervalsandRPEandTSat10-
min intervals. Mean Tsk (Equation 2.4) was subsequently calculated. Breath-by-breath expired
metabolic gas was measured using online gas analysis throughout the RHTT (3.9.). Relative
metabolicheatproduction(W.kg-1)wascalculatedinaccordancewiththeguidelinesofCramerand
Jay(2014).
ForearmsweatsampleswerecollectedduringeachoftheRHTTusingaMacroductsweatcollector
(Wescor,Logan,UT).Immediatelypriortoenteringthechambertheparticipants’skinwascleaned
withdeionizedwateranddriedimmediatelybeforesecuringtheMacroductsweatcollector.The
collectorswereheld in placeon themidpoint of the anterior forearmby aVelcro strap,which
preventedleakageandsamplecontamination.TheMacroductcollectorhasasurfaceareaof5.2
cm2.Sweatsecretedbythesweatglands is forcedfromtheductsunderhydraulicpressureand
flowsbetween theskinand theconcaveunder surfaceof theMacroductcollectorand into the
microbore tubing spiral.A small amountofbluedye ismixedwith the secretedsweatprior to
enteringthespiraltubingtoallowforvisualidentificationoftheonsetofsweating.Theforearm
sweat samples were analysed in duplicate for sweat NaCl using a sweat conductivity analyser
(SweatChek3120,Wescor,UK).

153
Heatacclimation
Inarandomised,cross-overdesign,participantscompletedtwo5-dHAinterventions(figure8.1).
Thetwointerventionswereseparatedby7-wks.toensurethedecayofadaptation(Pandolf,1998;
Poirieretal.,2015).Followinga20-minstabilisationperiod,measuresofTre(3.8.),HR(3.9.),RPE
andTS(3.13.)wererecorded,andparticipantsenteredtheenvironmentalchamber.Forthefirst
intervention(HACH)participantsenteredintoatemperateenvironment(3.7.)whilstwearingshorts
andasportsbra,forafurther20-minseatedrest.Forthesecondintervention(HAsauna)participants
enteredintoahotenvironment(50°C,30%RH;WBGT=40°C)whilstwearinga100%vinylsauna
suittoperformafurther20-minseatedrest.Thevinylsaunasuitcoveredthemajorityofthebody
with only the hands, feet, and head exposed. Following this, participants in both interventions
completedacontrolledhyperthermiaHAsession(3.14.)wearingshortsandasportsbrainahot
environment(3.7.).AllHAsessionswerecompletedonacycleergometer(3.14.).
Figure8.1Schematicoftheexperimentaldesign.
Notes:BM,bodymass;HA,heatacclimation;HR,heatrate;RH,relativehumidity;Tre,rectaltemperature.
Bloodsamplingandanalysis
Following20-minofseatedrestacapillarybloodsamplewastakenfromthefingertippriortoeach
oftheRHTTstoestimatePVexpansionusingEquation8.1(DillandCostill,1974).Capillaryblood
sampleswerecollectedinamicrocuvetteinduplicatefortheanalysisofhaemoglobinandanalysed
usingB-HaemoglobinPhotometer(HemocueLtd,Sweden).Capillarybloodsampleswerecollected
inglasscapillarytubesinduplicatefortheanalysisofhaematocritafterbeingcentrifugedat11,800
rpmfor3-min(Haemotospin1300Centrifuge,Hawksley&SonsLtd,UK).

154
Equation8.1CalculationforchangeinPlasmaVolume(∆PV)
∆PV,%=100(PVA–PVB)/PVB
Where:∆PV,changeinplasmavolume;PVA,preHAplasmavolume;PVB,postHAplasmavolume
VenousbloodsamplesweretakeninarestedstatepriortotheRHTTsandonD22ofthemenstrual
cycle.A6mLwholebloodsamplewasdrawnfromtheantecubitalfossa.Eachsamplewasdivided
into two EDTA tubes (Starstedt, Germany).Whole blood samples were centrifuged (Eppendorf
5702RCentrifuge,EppendorfUKLtd,UK)at4400rpmforaperiodof10-min.Plasmawaspipetted
(Eppendorfresearchpipettes,EppendorfUKLtd,UK)into1.5mLmicrotubes(Westernlaboratory
science,UK)andstoredat-86°C(SanyoVIPseries,SanyoElectricBiomedicalCoLtd,Japan)until
analysis. To perform the analysis, commercially available 17β-estradiol (ab108667) and
Progesterone (ab108670) immunoenzymaticassaykits (Abcamplc,UK)wereused.Quantitative
determination of plasma concentrations of 17β-estradiol and Progesterone were performed
accordingtothemanufacturer’sguidelines. Incubationof the96wellkit, includingtherequired
qualitycontrolstandardswasperformedonanorbitalplatformshaker(Titramax1000,HeidolpUK,
UK)at1.5mmorbitalvibrationandreadbyamicroplatereaderusingabsorptionat450nm(elx800,
BioTekUK,UK).Theintra-assayandinter-assayvariability,asdescribedbythemanufacturerwas
9% and 10% for 17β-estradiol and 4% and 9.3% for progesterone respectively. The lowest
detectableconcentrationof17β-estradiolandprogesterone,asdescribedbythemanufacturerwas
8.68pg.mL-1and0.05ng.mL-1,respectively.
Statisticalanalyses
AlldatawerefirstcheckedfornormalityusingtheShapiroWilktestandcorrectedforsphericity
usingtheGreenhouseGeissermethod.Atwoway(2x2)repeatedmeasuresANOVAwasperformed
to identify differences between the physiological and perceptual responses during RHTT1 and
RHTT2 between the HACH and HAsauna conditions and the physiological and HA method
characteristicsduringD1andD5ofHACHandHAsauna(3.17.4.).Whenamaineffectorinteraction
effect was found, results were followed up using a Bonferroni corrected post hoc comparison
(3.17.6.).Partialetasquared(np2)wasusedasameasureofeffectsize(3.17.5.).Apairedsamples
t-testwasperformedonthePVdata(3.17.4.).Alldatawereanalysedusingastandardstatistical
package(SPSSversion20.0,IBM,Armonk,NewYork,USA)andreportedasmean±SD.Statistical
significancewasacceptedatthelevelofp≤0.05.

155
8.4. Results
HAprotocolduringHACHandHAsauna(D1-D5)
Table8.1presentsthemean±SDdatafortheHAprotocolandphysiologicalresponsesforboth
conditions. All participants completed fiveHA sessions on consecutive days in both conditions.
DurationspentwithaTre≥38.5°Cwasonaverage52±11minacrossallsessionsinbothconditions.
DurationwithaTre≥38.5°CreducedinbothconditionsfromD1toD5(F(1,8)=8.147,p=0.021,np2
=0.505),however,nointeractioneffectbetweendayandconditionondurationwithaTre≥38.5°C
wasobserved(F(1,8)=1.488,p=0.257,np2=0.157).Nomaineffectforday(F(1,8)=5.139,p=0.053,
np2=0.391),orinteractioneffectbetweendayandconditiononexercisedurationwasobserved(F
(1,8)=2.250,p=0.172,np2=0.220).
RelativeexerciseintensityincreasedinbothconditionsfromD1toD5(F(1,8)=5.586,p=0.046,np2
=0.411),however,nointeractioneffectbetweendayandconditionwasobserved(F(1,8)=2.967,p
=0.123,np2=0.271).TotalworkincreasedinbothconditionsfromD1toD5(F (1,8)=5.593,p=
0.046,np2=0.411),however,nointeractioneffectbetweendayandconditionwasobserved(F(1,8)
=3.215,p=0.111,np2=0.287).
PhysiologicalresponsesduringHACHandHAsauna(D1–D5)
TherewasaninteractioneffectbetweendayandconditiononTrerest(F(1,8)=5.395,p=0.049,np2
=0.403).NodifferenceswereobservedbetweenconditionsonTrerestonD1(p=0.57).Trerest
reducedfromD1toD5(p=0.006)inHAsauna,butnotinHACH(p=0.268).Therewasaninteraction
effectbetweendayandconditiononmeanTre(F(1,8)=6.380,p=0.035,np2=0.444).Nodifferences
wereobservedbetweenconditionsformeanTreonD1(p=0.167).MeanTrereducedfromD1toD5
(p=0.016) in theHACHcondition,butnot inHAsauna (p=0.132).Therewasan interactioneffect
betweendayandconditiononHRrest(F(1,8)=5.924,p=0.041,np2=0.425).Nodifferenceswere
observedbetweenconditionsforHRrestonD1(p=0.251).HRrestreducedtoagreaterextent
fromD1toD5inHAsauna(p≤0.001)comparedtoHACH(p=0.009).Nomaineffectwasobservedfor
dayonmeanHR(F(1,8)=2.837,p=0.131,np2=0.262),furthermore,nointeractioneffectbetween
dayandconditionwasobserved(F(1,8)=0.059,p=0.814,np2=0.007).
SWRwashigher intheHAsaunaconditiononD1andD5comparedwithHACH(F (1,8)=21.719,p=
0.002,np2=0.731).Similarly,SWRBSAwashigherintheHAsaunaconditiononD1andD5compared
withHACH(F(1,8)=23.710,p≤0.001,np2=0.748)(table8.1).
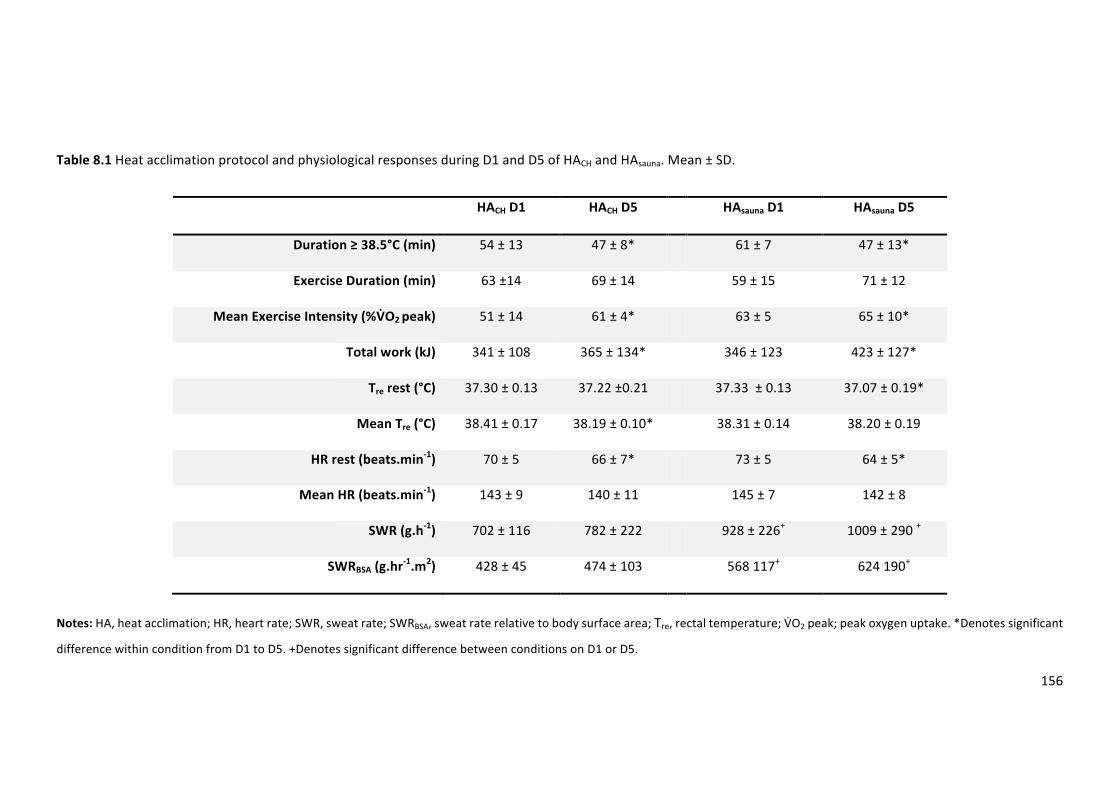
156
Table8.1HeatacclimationprotocolandphysiologicalresponsesduringD1andD5ofHACHandHAsauna.Mean±SD.
HACHD1 HACHD5 HAsaunaD1 HAsaunaD5
Duration≥38.5°C(min) 54±13 47±8* 61±7 47±13*
ExerciseDuration(min) 63±14 69±14 59±15 71±12
MeanExerciseIntensity(%VDO2peak) 51±14 61±4* 63±5 65±10*
Totalwork(kJ) 341±108 365±134* 346±123 423±127*
Trerest(°C) 37.30±0.13 37.22±0.21 37.33±0.13 37.07±0.19*
MeanTre(°C) 38.41±0.17 38.19±0.10* 38.31±0.14 38.20±0.19
HRrest(beats.min-1) 70±5 66±7* 73±5 64±5*
MeanHR(beats.min-1) 143±9 140±11 145±7 142±8
SWR(g.h-1) 702±116 782±222 928±226+ 1009±290+
SWRBSA(g.hr-1.m2) 428±45 474±103 568117+ 624190+
Notes:HA,heatacclimation;HR,heartrate;SWR,sweatrate;SWRBSA,sweatraterelativetobodysurfacearea;Tre,rectaltemperature;VRO2peak;peakoxygenuptake.*Denotessignificant
differencewithinconditionfromD1toD5.+DenotessignificantdifferencebetweenconditionsonD1orD5.
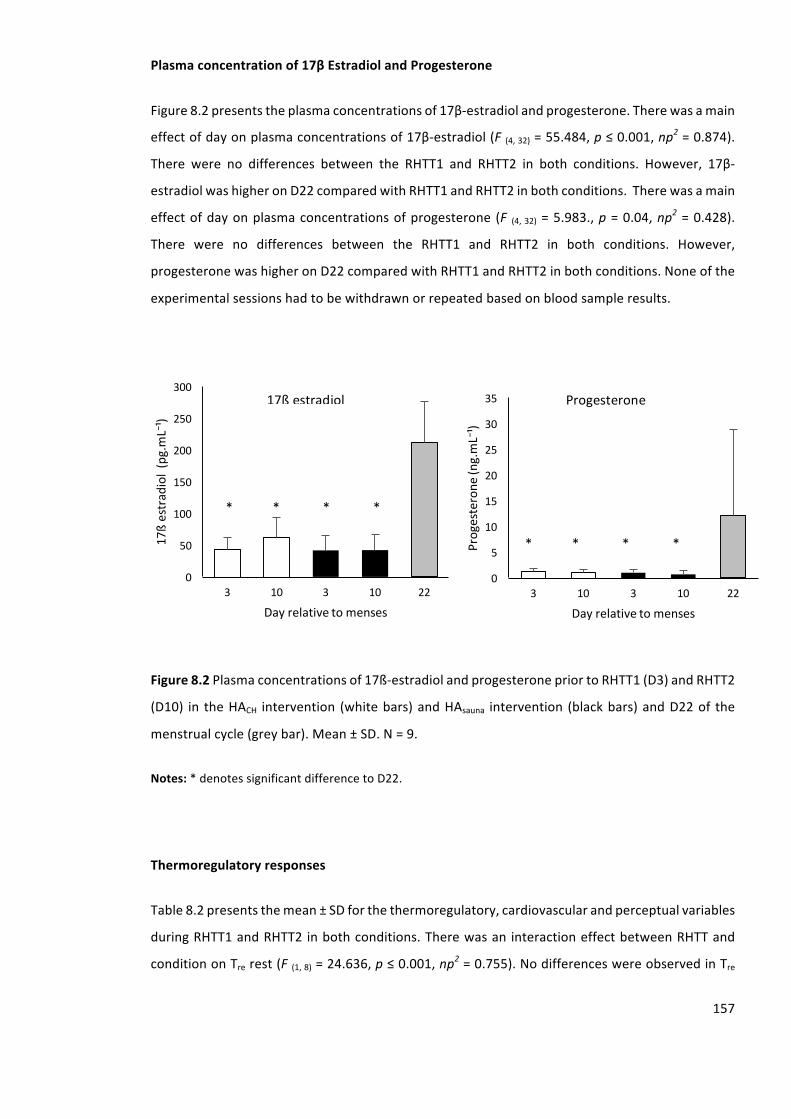
157
Plasmaconcentrationof17βEstradiolandProgesterone
Figure8.2presentstheplasmaconcentrationsof17β-estradiolandprogesterone.Therewasamain
effectofdayonplasmaconcentrationsof17β-estradiol(F(4,32)=55.484,p≤0.001,np2=0.874).
There were no differences between the RHTT1 and RHTT2 in both conditions. However, 17β-
estradiolwashigheronD22comparedwithRHTT1andRHTT2inbothconditions.Therewasamain
effectofdayonplasmaconcentrationsofprogesterone (F (4,32)=5.983.,p=0.04,np2=0.428).
There were no differences between the RHTT1 and RHTT2 in both conditions. However,
progesteronewashigheronD22comparedwithRHTT1andRHTT2inbothconditions.Noneofthe
experimentalsessionshadtobewithdrawnorrepeatedbasedonbloodsampleresults.
Figure8.2Plasmaconcentrationsof17ß-estradiolandprogesteronepriortoRHTT1(D3)andRHTT2
(D10) in theHACH intervention (whitebars)andHAsauna intervention (blackbars)andD22of the
menstrualcycle(greybar).Mean±SD.N=9.
Notes:*denotessignificantdifferencetoD22.
Thermoregulatoryresponses
Table8.2presentsthemean±SDforthethermoregulatory,cardiovascularandperceptualvariables
duringRHTT1andRHTT2inbothconditions.Therewasan interactioneffectbetweenRHTTand
conditiononTrerest(F(1,8)=24.636,p≤0.001,np2=0.755).NodifferenceswereobservedinTre
0
5
10
15
20
25
30
35
3 10 3 10 22
Progesterone
(ng.mL⁻¹)
Dayrelativetomenses
0
50
100
150
200
250
300
3 10 3 10 22
17ßestradiol(pg.mL⁻¹)
Dayrelativetomenses
****
****
17ßestradiol Progesterone
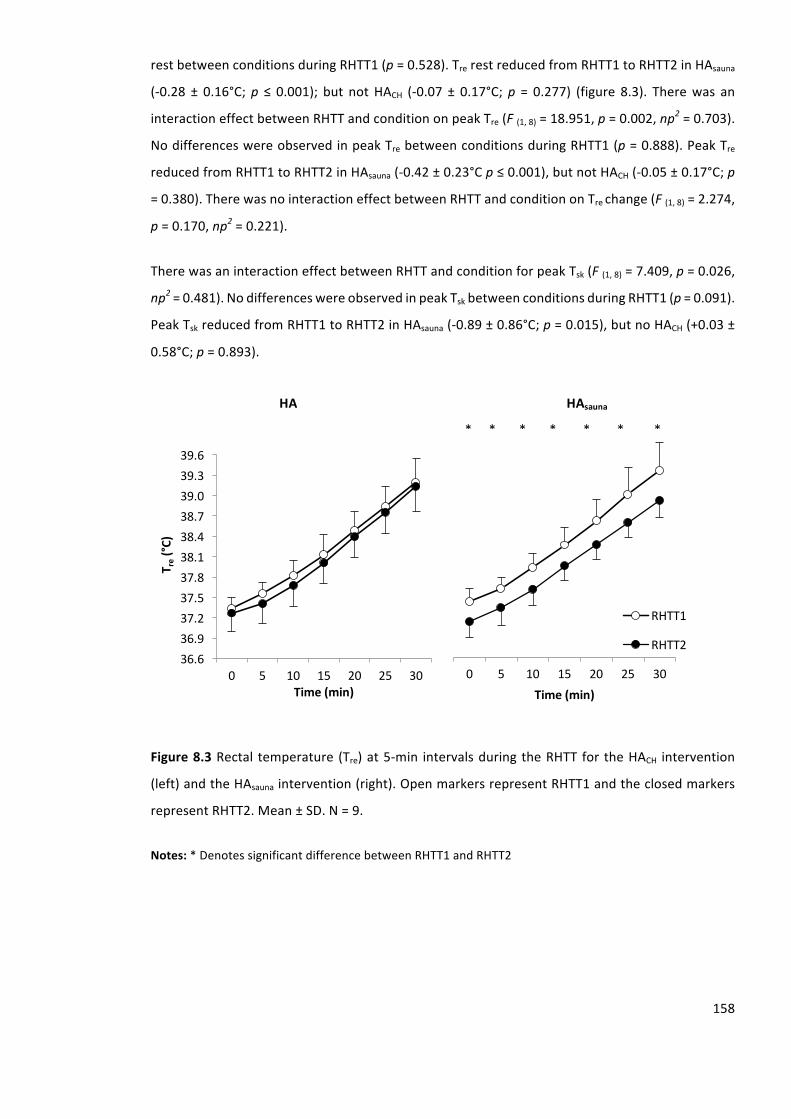
158
restbetweenconditionsduringRHTT1(p=0.528).TrerestreducedfromRHTT1toRHTT2inHAsauna
(-0.28 ± 0.16°C;p ≤ 0.001); but notHACH (-0.07 ± 0.17°C;p = 0.277) (figure 8.3). Therewas an
interactioneffectbetweenRHTTandconditiononpeakTre(F(1,8)=18.951,p=0.002,np2=0.703).
NodifferenceswereobservedinpeakTrebetweenconditionsduringRHTT1(p=0.888).PeakTre
reducedfromRHTT1toRHTT2inHAsauna(-0.42±0.23°Cp≤0.001),butnotHACH(-0.05±0.17°C;p
=0.380).TherewasnointeractioneffectbetweenRHTTandconditiononTrechange(F(1,8)=2.274,
p=0.170,np2=0.221).
TherewasaninteractioneffectbetweenRHTTandconditionforpeakTsk(F(1,8)=7.409,p=0.026,
np2=0.481).NodifferenceswereobservedinpeakTskbetweenconditionsduringRHTT1(p=0.091).
PeakTskreducedfromRHTT1toRHTT2inHAsauna(-0.89±0.86°C;p=0.015),butnoHACH(+0.03±
0.58°C;p=0.893).
Figure8.3Rectal temperature (Tre)at5-min intervalsduringtheRHTTfor theHACH intervention
(left)andtheHAsaunaintervention(right).OpenmarkersrepresentRHTT1andtheclosedmarkers
representRHTT2.Mean±SD.N=9.
Notes:*DenotessignificantdifferencebetweenRHTT1andRHTT2
0 5 10 15 20 25 30Time(min)
RHTT1
RHTT236.636.937.237.537.838.138.438.739.039.339.6
0 5 10 15 20 25 30
T re(°C
)
Time(min)
*******
HA HAsauna
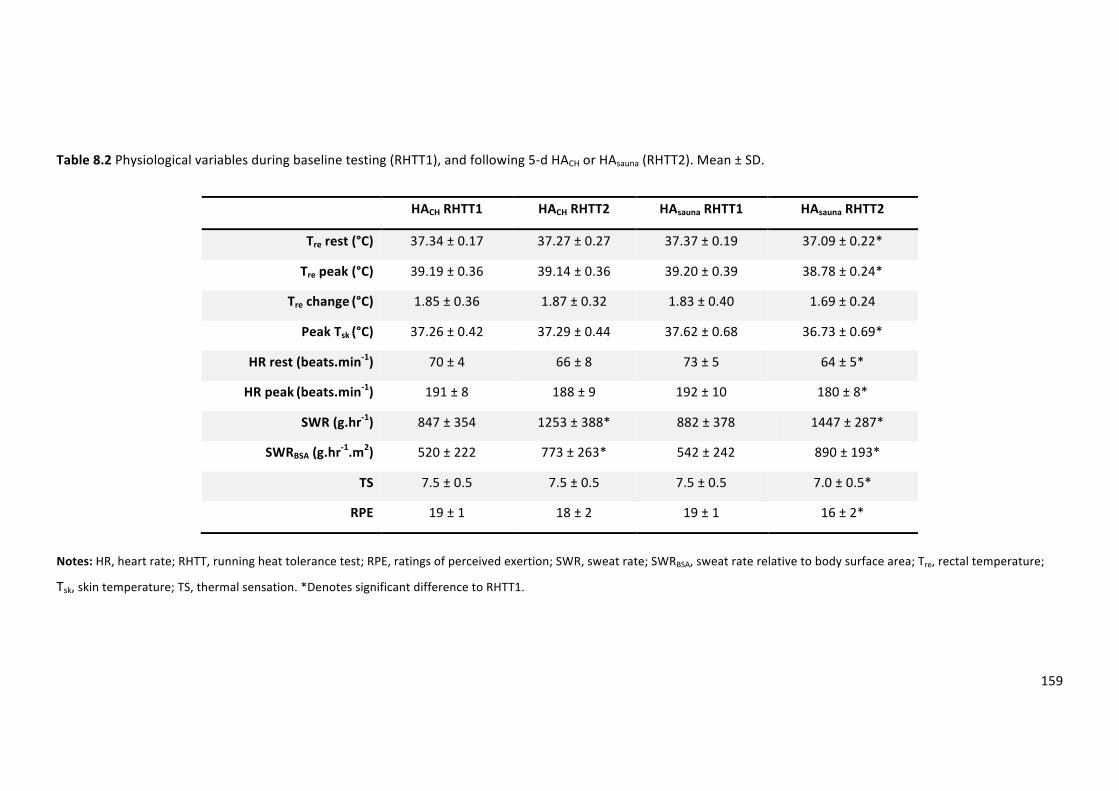
159
Table8.2Physiologicalvariablesduringbaselinetesting(RHTT1),andfollowing5-dHACHorHAsauna(RHTT2).Mean±SD.
HACHRHTT1 HACHRHTT2 HAsaunaRHTT1 HAsaunaRHTT2
Trerest(°C) 37.34±0.17 37.27±0.27 37.37±0.19 37.09±0.22*
Trepeak(°C) 39.19±0.36 39.14±0.36 39.20±0.39 38.78±0.24*
Trechange(°C) 1.85±0.36 1.87±0.32 1.83±0.40 1.69±0.24
PeakTsk(°C) 37.26±0.42 37.29±0.44 37.62±0.68 36.73±0.69*
HRrest(beats.min-1) 70±4 66±8 73±5 64±5*
HRpeak(beats.min-1) 191±8 188±9 192±10 180±8*
SWR(g.hr-1) 847±354 1253±388* 882±378 1447±287*
SWRBSA(g.hr-1.m2) 520±222 773±263* 542±242 890±193*
TS 7.5±0.5 7.5±0.5 7.5±0.5 7.0±0.5*
RPE 19±1 18±2 19±1 16±2*
Notes:HR,heartrate;RHTT,runningheattolerancetest;RPE,ratingsofperceivedexertion;SWR,sweatrate;SWRBSA,sweatraterelativetobodysurfacearea;Tre,rectaltemperature;
Tsk,skintemperature;TS,thermalsensation.*DenotessignificantdifferencetoRHTT1.
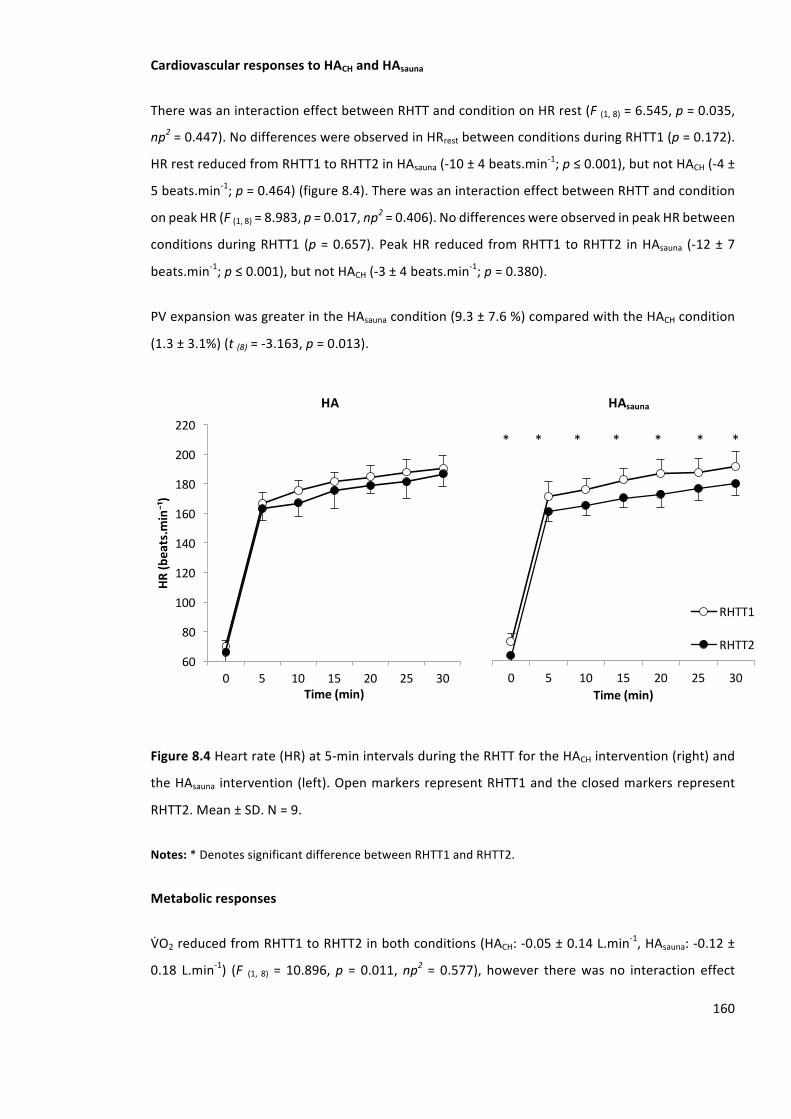
160
CardiovascularresponsestoHACHandHAsauna
TherewasaninteractioneffectbetweenRHTTandconditiononHRrest(F(1,8)=6.545,p=0.035,
np2=0.447).NodifferenceswereobservedinHRrestbetweenconditionsduringRHTT1(p=0.172).
HRrestreducedfromRHTT1toRHTT2inHAsauna(-10±4beats.min-1;p≤0.001),butnotHACH(-4±
5beats.min-1;p=0.464)(figure8.4).TherewasaninteractioneffectbetweenRHTTandcondition
onpeakHR(F(1,8)=8.983,p=0.017,np2=0.406).NodifferenceswereobservedinpeakHRbetween
conditionsduringRHTT1 (p=0.657).PeakHR reduced fromRHTT1 toRHTT2 inHAsauna (-12±7
beats.min-1;p≤0.001),butnotHACH(-3±4beats.min-1;p=0.380).
PVexpansionwasgreaterintheHAsaunacondition(9.3±7.6%)comparedwiththeHACHcondition
(1.3±3.1%)(t(8)=-3.163,p=0.013).
Figure8.4Heartrate(HR)at5-minintervalsduringtheRHTTfortheHACHintervention(right)and
theHAsauna intervention(left).OpenmarkersrepresentRHTT1andtheclosedmarkersrepresent
RHTT2.Mean±SD.N=9.
Notes:*DenotessignificantdifferencebetweenRHTT1andRHTT2.
Metabolicresponses
VYO2reducedfromRHTT1toRHTT2inbothconditions(HACH:-0.05±0.14L.min-1,HAsauna:-0.12±
0.18 L.min-1) (F (1, 8) = 10.896,p = 0.011,np2 = 0.577), however therewas no interaction effect
0 5 10 15 20 25 30Time(min)
RHTT1
RHTT260
80
100
120
140
160
180
200
220
0 5 10 15 20 25 30
HR(b
eats.m
in⁻¹)
Time(min)
*******
HA HAsauna

161
observedbetweenRHTTandcondition(F(1,8)=0.634,p=0.449,np2=0.073).Therewasnomain
effectofRHTTonRER(F(1,8)=0.139,p=0.719,np2=0.017)orinteractioneffectbetweenRHTTand
conditiononRER(F(1,8)=0.177,p=0.685,np2=0.022).Metabolicheatproductionreducedinboth
conditionsfromRHTT1toRHTT2(HACH:-0.3±0.7W.kg-1,HAsauna:-0.8±1.0W.kg-1;F(1,8)=10.896,
p = 0.011, np2 = 0.577). No interaction effect between RHTT and condition onmetabolic heat
productionwasobserved(F(1,8)=0.901,p=0.370,np2=0.101).
Sudomotorresponses
Sweatrate(F (1,8)=49.982,p≤0.001,np2=0.862)andSWRBSA(F (1,8)=52.796,p≤0.001,np2=
0.868),increasedinbothconditionsfromRHTT1toRHTT2.NointeractioneffectbetweenRHTTand
conditionwasobservedforSWR(F(1,8)=3.893,p=0.084,np2=0.327)andSWRBSA(F(1,8)=3.605,p
=0.094,np2=0.311)(figure8.5).
TherewasaninteractioneffectbetweenRHTTandconditionforsweatNaCl(F (1,8)=13.932,p=
0.006,np2=0.635).SweatNaClreducedfromRHTT1toRHTT2toagreaterextentinHAsauna(-16±
10mmol.L-1)comparedwithHACH(-5±2mmol.L-1).TherewasaninteractioneffectbetweenRHTT
andconditionforTreatonsetofsweating(F(1,8)=12.386,p=0.008,np2=0.608).Nodifferences
wereobservedinTreatonsetofsweatingbetweenconditionsduringRHTT1(HACH37.55±0.20°C,
HAsauna37.54±0.16°C;p=0.989).TheTreatonsetofsweatingreducedfromRHTT1toRHTT2in
HAsauna(-0.29±0.15°C;p≤0.001),butnotHACH(-0.08±0.17°C;p=0.137)(figure8.5).
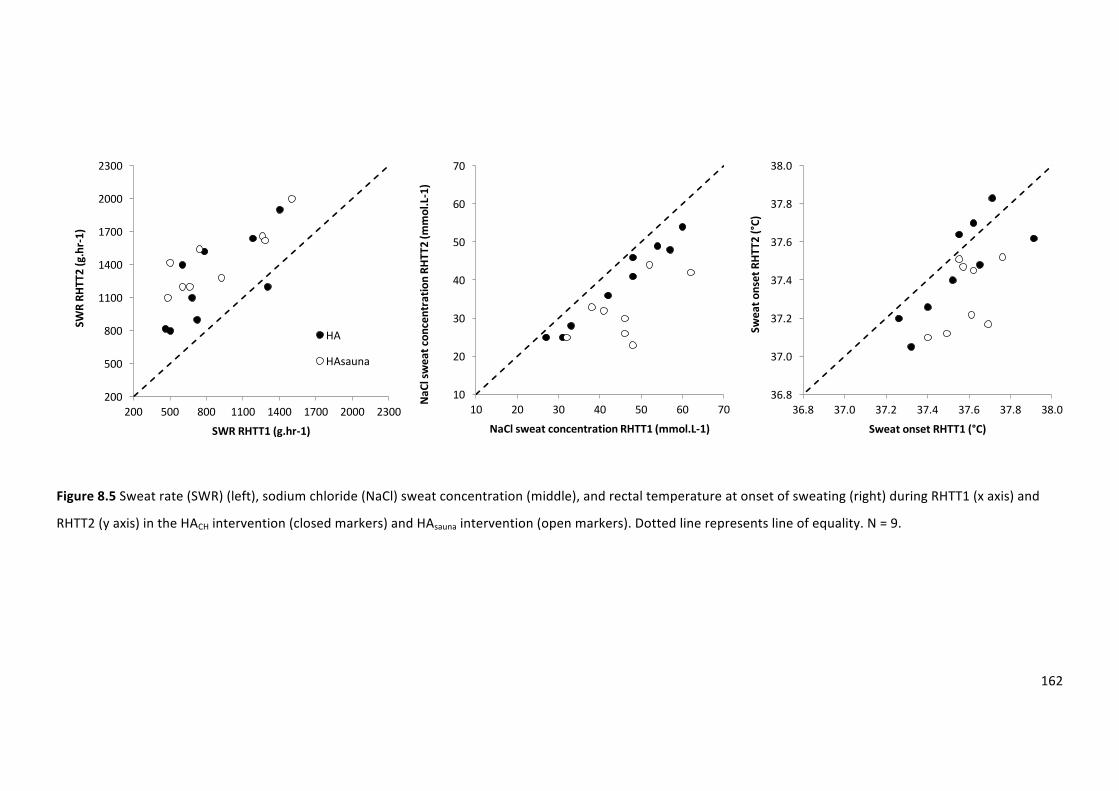
162
Figure8.5Sweatrate(SWR)(left),sodiumchloride(NaCl)sweatconcentration(middle),andrectaltemperatureatonsetofsweating(right)duringRHTT1(xaxis)and
RHTT2(yaxis)intheHACHintervention(closedmarkers)andHAsaunaintervention(openmarkers).Dottedlinerepresentslineofequality.N=9.
36.8
37.0
37.2
37.4
37.6
37.8
38.0
36.8 37.0 37.2 37.4 37.6 37.8 38.0
Sweato
nsetRHT
T2(°C)
SweatonsetRHTT1(°C)
10
20
30
40
50
60
70
10 20 30 40 50 60 70Na
Clsw
eatcon
centratio
nRH
TT2(m
mol.L-1)
NaClsweatconcentrationRHTT1(mmol.L-1)
200
500
800
1100
1400
1700
2000
2300
200 500 800 1100 1400 1700 2000 2300
SWRRH
TT2(g.hr-1)
SWRRHTT1(g.hr-1)
HA
HAsauna

163
Perceptualresponses
TherewasaninteractioneffectbetweenRHTTandconditiononTS(F(1,8)=67.600,p≤0.001,np2=
0.894). No differences were observed in TS between conditions during RHTT1 (p = 1.000). TS
reduced from RHTT1 to RHTT2 in HAsauna (p = 0.002), but not HACH (p = 0.729). There was an
interactioneffectbetweenRHTTandconditiononRPE(F(1,8)=24.143,p=0.001,np2=0.751).No
differenceswereobservedinRPEbetweenconditionduringRHTT1(p=0.347).RPEreducedfrom
RHTT1toRHTT2inHAsauna(p≤0.001),butnotHACH(p=0.081).
8.5. Discussion
ThecurrentstudydemonstratedthatprecedingSTHAwithapassiveheatexposureacceleratesthe
adaptation in females, with HAsauna resulting in PV expansion and superior adaptation to the
sudomotorfunctioncomparedwithHAalone.Thesefindingsareinsupportoftheaimofthestudy.
Specifically,theprimaryandsecondaryhypothesiswerebothsupported. Inaddition,thesedata
confirmedthefindingsfromStudy3(Chapter6)withfemalesfailingtoachievesomeoftheclassic
markers of controlled hyperthermia STHA, including a reduced thermoregulatory and
cardiovascular strain.However, followingHAsauna reductions in thermoregulatory, cardiovascular
andperceptualstrainwereobserved.TheadaptationpathwaywaslikelymediatedinpartbyPV
expansion and an improved thermoeffector and thermosensitivity response of the sudomotor
function.TheimportanceofthesefindingsisthatHAsaunamayaccelerateHAwithoutincreasingthe
exercise stimulus.However, it remainsunknown if thereareadditionalmodifications to theHA
protocol that may offer the potential for further accelerating the adaptations within a female
population.
HACHandHAsaunaweresuccessfullymatchedfortheprescribedtrainingparametersofdurationTre
≥38.5°C,exerciseduration,relativeexerciseintensityandtotalworkcompleted(table8.1).This
providesconfidencethattheadditionaladaptationsinducedfollowingHAsaunacomparedwithHACH
werearesultofthe20-minpreHAsessionexposuretoahotwhilstwearingasaunasuitcompared
withtemperateconditions.ThedataindicatethattheHAsaunaisaneffectiveHAstrategy,evidenced
byareductioninTrerest,peakTre,HRrest,peakHR,TSandRPEcombinedwithPVexpansionand
an enhanced thermoeffector (Tre at onset of sweating) and thermosensitivity (SWR) of the
sudomotorfunction.Thesefindingsare largely inkeepingwithotherresearchemployingsimilar
STHAregimesinmales(Garrettetal.,2012;Gibsonetal.,2015b;Nealetal.,2015).

164
PlasmavolumewasunchangedintheHACHcondition.Thesefindingsareinaccordancewithother
controlledhyperthermiaSTHAinvestigationsusingmaleparticipants(Garrettetal.,2009;Nealet
al.,2015).IthasbeensuggestedthatsaunaexposureactsasanindependentstimulusforHAby
augmentingPVexpansion(Scoonetal.,2007;Stanleyetal.,2015).Inthecurrentstudy,therewas
a9.3±7.6%PVexpansionfollowingtheHAsaunacondition.ThemagnitudeofPVexpansionisvery
similartothatobservedbyothersfollowingLTHAandlong-termsaunaexposure(Burketal.,2012;
Gibsonetal.,2015c;Stanleyetal.,2015);butexceededthe6.5%expansionreportedbyLorenzo&
Minson(2010)followingLTHAand7.1%expansionreportedbyScoonetal.(2007)followinglong-
termsaunaexposure.PVexpansionresultsinanincreasedvascularfillingtosupportcardiovascular
stability, increased specific heat capacity of blood and an improved skin blood flow responses
(Sawkaetal.,2011a).Thus,thereducedthermoregulatoryandcardiovascularresponsesobserved
inthecurrentstudywerelikelyinpartmediatedbythePVexpansion.
Enhancedthermosensitivityofthesudomotorfunctionwasobserved,evidencedbya407g.hr-1and
564g.hr-1increaseinSWRduringtheRHTTfollowingtheHACHandHAsaunacondition,respectively.
These findings are in accordance with the 691 g.hr-1 increases in SWR previously observed in
females following STHA (Study 3, Chapter 6). Furthermore, in the current study, there was an
enhancedthermoeffectorresponse,evidencedbyareductionintheTreattheonsetofsweatingin
theHAsaunacondition.Thus,theobservedincreaseinSWRmayinpartbemediatedbyanincreased
cholinergicsensitivityoftheeccrinesweatglandcausinganalteredafferentneuralactivityfrom
the central thermoreceptors that altered the integration of thermal information (Charkoudian,
2010). Despite an enhanced thermosensitivity following the HACH condition there were no
alterations in the thermoeffector response, thus, these findingsmay inpartbeexplainedbyan
increaseinglandularhypertrophy(Buonoetal.,2009b;LorenzoandMinson,2010).Thereduced
thermoregulatorystrainobservedinthecurrentstudywaslikelyinpartmediatedbyanenhanced
evaporativeheatlosscombinedwithareductioninmetabolicheatproduction.
Thisstudyisthefirststudytoprovidedataonsweatmineralconcentrationsandlossesfollowing
controlledhyperthermiaSTHAinafemalepopulation.BoththeHACHandHAsaunaconditionresulted
inareductioninsweatNaCl.ThemagnitudeofreductionfortheHAsaunaconditionexceededthat
reportedpreviouslyinmalesfollowingSTHA(Nealetal.,2015).Themechanismofsweatmineral
conservationisunclear,althoughpreviousdatasuggestssweatsodiumconservationfollowingHA
involvesincreasedsodiumionreuptakewithinthere-absorptiveductofthesweatgland(Satoand
Dobson,1970; Satoet al., 1971). This is anovel findingpresentedwithin the current studyhas
practicalrelevancetofemaleathleteswhomayexperiencechronicperfusesweatingduringtraining
whichmayraisethepotentialformineraldeficiencies(Chinevereetal.,2008).

165
HAsaunaresultedinareductionpretopostinTSandRPE.Thismayhaveimplicationsforself-paced
exerciseintheheat,asaconsequenceofTSbeingamaindriverofbehaviouralthermoregulation
(FlourisandSchlader,2015).Theobservedreductions inTSandRPE followingHAsauna isanovel
finding with practical relevance for athletes preparing to compete under hot environmental
conditions.However,furtherinvestigationsarerequiredtodeterminetheeffectofSTHA,withor
withoutadditionalsaunastressonpacingandtimetrialperformanceinfemales.
8.6. Limitations
Typically, it issuggestedthatphenotypicadaptationsdecayatarateofa1-dlossofacclimation
statusforevery2-dspentwithoutheatexposure(GivoniandGoldman,1973),supportingthe7-
wks between the twoHA conditions adopted in the current study. However, evenwith 7-wks.
betweenthetwoHAconditionsitispossiblethatthemolecularmemoryfromthepriorcondition
wasdifferentbetweenparticipants(Horowitz,2014).Thecurrentstudycontrolledforthisbyusing
arandomiseddesignhowever,furtherinvestigationisrequiredtoquantifythemolecularmemory
establishedduringthetwoconditions.
ChangesinPVwereestimatedviachangesinhaematocritandhaemoglobinconcentrations.This
methodhasbeenreportedtoreflectsmall-to-moderatechangesinPVaccurately,however,itmay
underestimatelargechangesinPV(LundvallandLindgren,1998).Nevertheless,atypicalerrorasa
coefficientofvariationof4.0%and3.9%forthemeasurementofhaematocritandhaemoglobin
respectively was observed between duplicate samples providing confidence that the change
observedisameaningfulchange.
Thecurrentstudyrecruitedphysicallyactivefemaleswhoperformedbothrunningandcyclingin
theirweeklytrainingregimenandhadanaverageVdO2peakof50mL.kg-1.min-1.Thus,theseresults
areconstrainedtoaphysicallyactive,femalepopulation.HAisastrategytypicallyadoptedbyhighly
trainedathletespriortotrainingandcompetingintheheat.Furtherinvestigationsarerequiredto
establishthemagnitudeofadaptation inahighlytrainedgroupoffemaleswhomayexperience
smallermarginalgainfromHA.
8.7. Conclusion
ThisisthefirststudytoinvestigatemethodstooptimiseSTHAspecificallyforfemales.TheHAsauna
conditionestablishedreductionsinthermoregulatory,cardiovascularandperceptualstrain,andan
improved thermosensitivity and thermoeffector sudomotor function. These findings provide
evidencethatprecedingaHAsessionwithapassiveheatexposuremayofferatime-efficientmeans

166
bywhichtofurtherenhanceacclimationinafemalepopulation.Furtherinvestigationsarerequired
to establish the optimal exposure to establish the greatest magnitude of adaptation whilst
minimisingtheexercisestimulusandexposureduration.

167
9. Generaldiscussion
________________________________________________________________________________
Thischapterwillbepresentedinfourmainsections.Firstly,theprinciplefindingsforeachofthe
studiespresentedwithinthisthesiswillbediscussedwithatableofhypothesesfromeachstudy
andwhethertheywereacceptedorrejected(table9.1).Secondly,amechanisticoverviewwillbe
provided,forthemainphysiologicalissuesarisingfromtheexperimentalstudies,usingpooleddata
fromacrossallthestudiespresentedwithinthisthesis.Possibledirectionsforfutureresearchwill
thenbeconsideredpriortodiscussingthepracticalapplicationandimpactofthedatapresented
withinthisthesis.
9.1. PrincipleFindings
9.1.1. Heattolerancetest
Thefirstexperimentalstudy(Chapter4)withinthisthesisintroducedthefirstHTTwhichadoptsa
runningmodeofexerciseandassesseditsrepeatability.Thedatafromthisstudysupportedthe
primaryandsecondaryhypothesis(table9.1).PhysiologicalresponsesduringtheRHTThadstrong,
positivecorrelationsandnosignificantdifferences inphysiological responsesbetweenrepeated
trials. FollowingChapter4, the sensitivityof theRHTT to changes inheat tolerance followinga
chronic,heat-alleviatingstrategywasassessed.Thedatafromthisstudy(Study2,Chapter5)also
supported the primary and secondary hypothesis (table 9.1). Thus, the RHTT was sensitive to
changesinheattoleranceandtoindividualdifferencesinthemagnitudeofadaptation.Theresults
fromthesetwostudiesindicatethattheRHTTisasensitivetoolwhichcanbeusedtoquantifythe
changes in tolerance following chronic heat-alleviating procedures when using a repeated
measuresdesign. Inaddition, the results suggest that theRHTTmaybeanappropriate tool for
physiologists,coachesandmedicalstafftousetohelpinformdecisionsregardingathletes’safety,
and to inform the severity of the heat-alleviating procedures required prior to training and
competingintheheat.
9.1.2. Sexcomparisonstoheatacclimation
Thedatafromthethird(Chapter6)andfourth(Chapter7)studiespresentedwithinthisthesiswere
collectedtogether,butreportedastwoseparatechapterssincetheywereaddressingverydistinct
questions.Theaimofthethirdstudywastoprovidethefirstdatacomparingmales’andfemales’
phenotypicadaptationstoSTHAandLTHAusingtheRHTTtoquantifythemagnitudeofadaptation.

168
HAwaseffectiveinattenuatingphysiologicalstrainandimprovingexerciseheattolerance.STHA
waseffectiveinachievingpartialadaptationinbothmalesandfemales.However,femalesrequired
LTHA to establish reductions in cardiovascular and thermoregulatory strain. The data from this
study supports the primary, secondary and tertiary hypothesis (table 9.1) and provides novel
informationonfemales’phenotypicresponsestocontrolledhyperthermiaHA.
Theaimofthefourthstudy(Chapter7)wastoprovidethefirstdatacomparingmales’andfemales’
cellularadaptionstoSTHAandLTHAviathequantificationofHsp72mRNA.Malesandfemaleshad
similarHsp72mRNAexpressionacrossthecourseofcontrolledhyperthermiaHA.Therewereno
differences in the endogenous criteria to elicit the HSR and the adaptation towards
thermotolerancebetweenmalesandfemalesoccursindiscriminately.Thisdataisnotinsupportof
thehypothesisthatHsp72mRNAresponsewillbeattenuatedinfemalescomparedtomalesacross
thecourseofcontrolledhyperthermiaHA(table9.1).Accordingly,thisdataprovidesmechanistic
supportforpractitionersprescribingcontrolledhyperthermiaHAforfemaleathletes.
9.1.3. Acceleratedheatacclimationresponsesinfemales
Theaimofthefinalstudypresentedwithinthisthesiswastoinvestigatewhetheritispossibleto
acceleratethephenotypicadaptionstoSTHAinfemalesduringthefollicularphaseofthemenstrual
cycle,confirmedbyplasmaconcentrationsof17ßestradiolandprogesterone.Todothis,anovel
HAmethodcombiningapassivesaunalikeexposurewithcontrolledhyperthermiawasused,which
combinedcontrolledhyperthermiaHAwithapassiveexposurein50°Cwhilstwearinga100%vinyl
saunasuit.ThisinterventionresultedinanexpansionofPVandanimprovedsudomotorfunction,
resultinginareducedrestingandpeakthermoregulatoryandcardiovascularstrain,inadditionto
areductioninperceptualresponses;allofwhichwerenotobservedfollowingHAalone.Thisnovel
dataisinsupportoftheprimaryandsecondaryhypotheses(table9.1).Thesefindingsprovidenew
evidencethatfemalescanacceleratethephenotypicadaptationsduringcontrolledhyperthermia
HA.
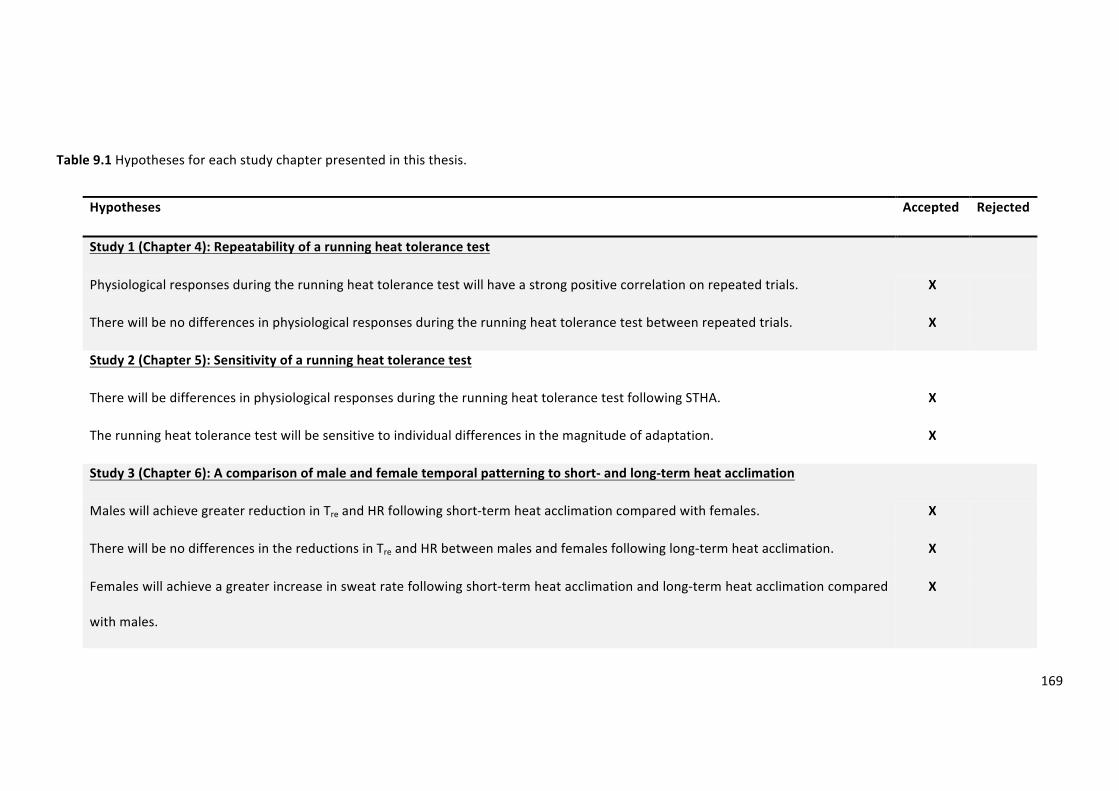
169
Table9.1Hypothesesforeachstudychapterpresentedinthisthesis.
Hypotheses Accepted Rejected
Study1(Chapter4):Repeatabilityofarunningheattolerancetest
Physiologicalresponsesduringtherunningheattolerancetestwillhaveastrongpositivecorrelationonrepeatedtrials. X
Therewillbenodifferencesinphysiologicalresponsesduringtherunningheattolerancetestbetweenrepeatedtrials. X
Study2(Chapter5):Sensitivityofarunningheattolerancetest
TherewillbedifferencesinphysiologicalresponsesduringtherunningheattolerancetestfollowingSTHA. X
Therunningheattolerancetestwillbesensitivetoindividualdifferencesinthemagnitudeofadaptation. X
Study3(Chapter6):Acomparisonofmaleandfemaletemporalpatterningtoshort-andlong-termheatacclimation
MaleswillachievegreaterreductioninTreandHRfollowingshort-termheatacclimationcomparedwithfemales. X
TherewillbenodifferencesinthereductionsinTreandHRbetweenmalesandfemalesfollowinglong-termheatacclimation. X
Femaleswillachieveagreaterincreaseinsweatratefollowingshort-termheatacclimationandlong-termheatacclimationcompared
withmales.
X

170
Study4(Chapter7):SexcomparisonofleukocyteHsp72mRNAtranscriptionduringheatacclimation
Hsp72mRNAresponsewillbelowerinfemalescomparedtomalesacrossthecourseofcontrolledhyperthermiaheatacclimation. X
Study5(Chapter8):Passiveheatexposureprecedingshort-termheatacclimationacceleratesphenotypicadaptationinfemales
Short-termheatacclimationprecededbyapassiveheatexposurewillresultingreaterplasmavolumeexpansioninfemalescompared
withheatacclimationalone.
X
Short-termheatacclimationprecededbyapassiveheatexposurewillproduceagreatersudomotoradaptationinfemalescompared
withheatacclimationalone.
X

171
9.2. MechanisticOverview
9.2.1. HeatTolerance
ThisthesishasintroducedthefirstHTTwhichadoptsarunningmodeofexercise.Datahasbeen
presentedtodemonstratethatthemethod isrepeatable,supportingtheuseoftheRHTTwhen
usingarepeatedmeasuresdesign.Interestingly,datahasalsobeenpresentedtodemonstratethat
individuals’ thermoregulatory responses to the RHTT are highly variable with participants’
responses along a heat tolerance continuum (figure 4.1). When exercising at a fixed absolute
workload as in the RHTT, typically this results in a similar absolute heat production between
individuals of a variety of biophysical factors. However, thiswill typically result in greater heat
productionrelativetoBMinfitterindividualswithalowpercentagebodyfat,sincetheyoftenhave
asmallerbodysize.Evidencehasbeenprovidedtosupportthatwhenmatchingexerciseintensity
atafixedheatproductionrelativetototalBMthesystematicdifferencesinTcbetweengroupsof
differentbodysize(CramerandJay,2014)andfitness(Jayetal.,2011)areeliminated.Thus,the
fixedworkloadoftheRHTTmaylimitapplicabilityofthetestwhencomparingbetweenindividuals,
especiallywhentheyarenotmatchedforbiophysicalfactors.Thismaypotentiallychallengethe
RHTTbeingable todemonstrate individualdifferences inheat tolerancewith theuseofaheat
tolerancecontinuum.
To identify the predominant factors responsible for the observed differences in changes in Tre
betweenindividualsduringtheRHTTandtodeterminewhethertheprotocolhasthepotentialto
beusedtocomparebetweenindependentgroups,allfemaledatapresentedwithinthisthesis(N
=30)waspooledtoenableretrospectiveanalysis.Part1oftheanalysesinvolvedallpooleddata(N
=30).Correlationcoefficients(r)wereestablishedbetweenchanges inTreduringtheRHTTwith
BSA,sumofskinfolds,VVO2peak,relativeexerciseintensity,andheatproductionexpressedinits
absoluteform(W),relativetoBSA(W.m2),andBM(W.kg-1).
Part2of thisanalyses, involved14 femaleswhowereselected fromthepooleddata toenable
comparisonsofTrechangebetweentwoindependentgroupsoflowBM(N=7;49.5±4.5kg)and
highBM(N=7;71.7±2.2kg).ThecriteriaforselectionoftheseparticipantsforthelowBMgroup
wasaBMof≤55kgand≥65kgforthehighBMgroup.Part3oftheanalysesinvolvedaseparate
14 females, whowere selected from this pooled data to enable the comparison of Tre change
betweentwoindependentgroupsoflowVVO2peak(N=7;41±2mL.kg-1.min-1)andhighVVO2peak
(N=7;57±5mL.kg-1.min-1).ThecriteriaforselectionoftheseparticipantsforthelowVVO2peak
was≤48mL.kg-1.min-1(performancelevel1and2)andforthehighVVO2peakwas≥52mL.kg-1.min-
1 (performance level 4 and 5) (Decroix et al., 2016). Independent sample t tests were used to

172
established differences between the independent groups’ data. Participant characteristics are
displayedintable9.2andpresentedasmean,SD,minimum,andmaximumvalues.
Figure 9.1 presents the correlation coefficients for the association between changes in Tre and
biophysicalandexerciseintensityvariables.Absoluteheatproduction(r=0.603,p≤0.001;figure
9.2),heatproductionrelativetoBSA(r=0.592,p≤0.001),andheatproductionrelativetoBM(r=
0.549,p=0.002)hadmediumpositivecorrelationswithchangeinTre.BSA(r=0.349,p=0.058),
sumofskinfolds(r=0.248,p=0.186),relativeVVO2peak(r=-0.204,p=0.278),andrelativeexercise
intensity(r=0.362p=0.051)didnothaveasignificantcorrelationwithchangeinTre(figure9.1).
Trechangesweresimilarbetweenthe lowBMgroup(1.7±0.2°C)andthehighBMgroup(2.0±
0.4°C)(t(12)=1.781,p=0.100)despiteahigherheatproductioninthehighBMgroup(lowBM:515
±78W;highBM:782±176W;t(12)=3.645,p=0.003).Interestingly,whenheatproductionwas
expressedrelativetoBMthesedifferenceswereeliminated(lowBM:10.5±1.7W.kg-1;highBM:
10.9±2.6W.kg-1;t(12)=0.403,p=0.694).
TrechangesweresimilarbetweenthelowVVO2peakgroup(2.0±0.4°C)andthehighVVO2peakgroup
(1.8 ± 0.4°C) (t (12) = 0.952, p = 0.360) despite a slightly, although not significantly higher heat
productioninthelowVVO2peakgroup(lowVVO2peak:735±189W;highVVO2peak:547±143W;t
(12) = 1.617, p = 0.132). Although, when heat production was expressed relative to BM these
differenceswereeliminated(lowVVO2peak:10.7±2.4W.kg-1:highVVO2peak:10.6±2.2W.kg-1;t(12)
=0.117,p=0.909).
OncompletionoftheRHTTabsoluteheatproduction(figure9.2),heatproductionrelativetoBSA,
and heat production relative to BM were found to be the best predictors of Tre change.
Furthermore, similar changes in Tre were observed in two independent groups with large
differencesinBMandfitness.Thesefindingsareexplainedbytheexerciseintensityadministered
during the RHTT equating to similar heat production relative to BM, permitting unbiased
independentgroupcomparisonsofTc.ThesefindingsaresupportedbytheworkofCramerandJay
(2014)whoreportedsimilarchangesinTrebetweentwoindependentgroupswithlargedifferences
in BM when exercise was administered at a fixed rate of heat production relative to BM.
Furthermore,Jayetal.(2011)reportedsimilarchangesinTrewhenexercisewasadministeredat
fixedratesofheatproductionrelativetoBMbetweenindependentgroupswithlargedifferences
in fitness. These findings provide initial evidence to support the use of the RHTT between
independentgroupssinceitappearsthatdespitetheexerciseintensitybeingfixed,therelativeheat
production is similar between independent groups of different BM and fitness. These

173
interpretationsare limited toa femalepopulation, further consideration shouldbemadewhen
usingmaleparticipants.
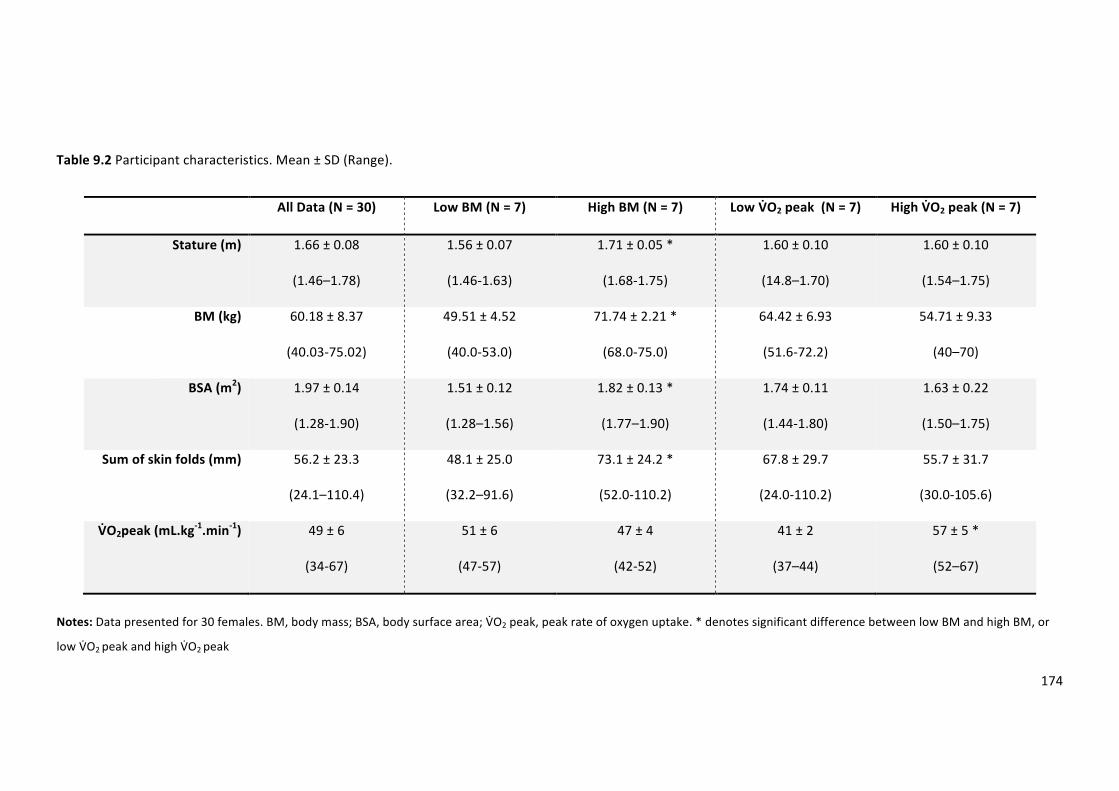
174
Table9.2Participantcharacteristics.Mean±SD(Range).
AllData(N=30) LowBM(N=7) HighBM(N=7) LowV>O2peak(N=7) HighV>O2peak(N=7)
Stature(m) 1.66±0.08
(1.46–1.78)
1.56±0.07
(1.46-1.63)
1.71±0.05*
(1.68-1.75)
1.60±0.10
(14.8–1.70)
1.60±0.10
(1.54–1.75)
BM(kg) 60.18±8.37
(40.03-75.02)
49.51±4.52
(40.0-53.0)
71.74±2.21*
(68.0-75.0)
64.42±6.93
(51.6-72.2)
54.71±9.33
(40–70)
BSA(m2) 1.97±0.14
(1.28-1.90)
1.51±0.12
(1.28–1.56)
1.82±0.13*
(1.77–1.90)
1.74±0.11
(1.44-1.80)
1.63±0.22
(1.50–1.75)
Sumofskinfolds(mm) 56.2±23.3
(24.1–110.4)
48.1±25.0
(32.2–91.6)
73.1±24.2*
(52.0-110.2)
67.8±29.7
(24.0-110.2)
55.7±31.7
(30.0-105.6)
V>O2peak(mL.kg-1.min-1) 49±6
(34-67)
51±6
(47-57)
47±4
(42-52)
41±2
(37–44)
57±5*
(52–67)
Notes:Datapresentedfor30females.BM,bodymass;BSA,bodysurfacearea;VPO2peak,peakrateofoxygenuptake.*denotessignificantdifferencebetweenlowBMandhighBM,or
lowVPO2peakandhighVPO2peak
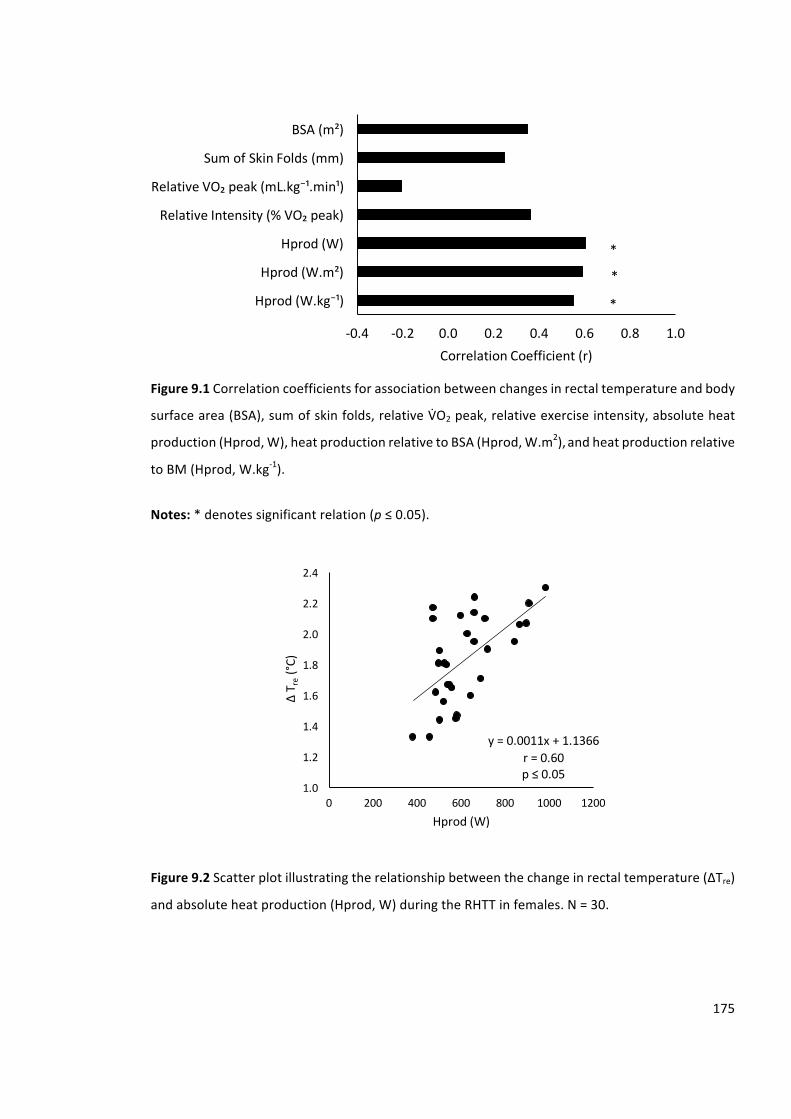
175
Figure9.1Correlationcoefficientsforassociationbetweenchangesinrectaltemperatureandbody
surfacearea(BSA),sumofskinfolds,relativeVCO2peak,relativeexerciseintensity,absoluteheat
production(Hprod,W),heatproductionrelativetoBSA(Hprod,W.m2),andheatproductionrelative
toBM(Hprod,W.kg-1).
Notes:*denotessignificantrelation(p≤0.05).
Figure9.2Scatterplotillustratingtherelationshipbetweenthechangeinrectaltemperature(∆Tre)
andabsoluteheatproduction(Hprod,W)duringtheRHTTinfemales.N=30.
-0.4 -0.2 0.0 0.2 0.4 0.6 0.8 1.0
Hprod(W.kg⁻¹)
Hprod(W.m²)
Hprod(W)
RelativeIntensity(%VO₂peak)
RelativeVO₂peak(mL.kg⁻¹.min¹)
SumofSkinFolds(mm)
BSA(m²)
CorrelationCoefficient(r)
*
*
*
y=0.0011x+1.1366r=0.60p≤ 0.05
1.0
1.2
1.4
1.6
1.8
2.0
2.2
2.4
0 200 400 600 800 1000 1200
∆T r
e(°C)
Hprod(W)

176
9.2.2. HeatAcclimation
Aprimaryobjectiveofthisthesiswastoexplainfemales’physiologicalandcellularresponsesto
controlled hyperthermia HA while also considering the effect of combining HA with passive
exposurestoacceleratethephysiologicalandperceptualresponsestoafixedexerciseheatstress.
Priortotheproductionofthisthesisalargebodyofresearchhadinvestigatedmales’responsesto
controlledhyperthermiaHA,however, therewasno literaturepublishedonfemales’ responses.
The limited data published on females, has used fixed (Avellini et al., 1980) or self-paced
(Sunderland et al., 2008) HA strategies which potentially constrained adaptation. This thesis
presents the first data set on females’ phenotypic and cellular adaptations to controlled
hyperthermiaHA.
Temporalpatterningofcontrolledhyperthermiaheatacclimationinfemales
Functionally,HAenhancesthecapacityofanorganismtotolerateheatstressbyaugmentingheat
dissipationandelevatingthebodytemperaturethatcanbesustainedintheheat(Horowitzand
Kodesh, 2010). Such resistance is a consequence of systemic adaptation, a chronic process
dependentuponheat-inducedreprogrammingofgeneexpression(HorowitzandKodesh,2010).
Traditionally,measuresofTre,HR,andPVwerereportedtoplateaufollowing3–8dofHA,with
SWRtaking8-14dtoreachaplateau(ArmstrongandMaresh,1991).Thisdataislimitedtomales’
responses.Therepeatedexposuretoaconstantoverloadproducesphysiologicalhabituation.Thus,
theobservedplateausinkeymarkersofHAmaybeanartefactofconstantworkratemethodsand
theinherentdecreaseinphysiologicalstrainobservedoverthedurationoftheprotocol.Toavoid
this inherent limitation and optimise adaptation, controlled hyperthermia is used. Controlled
hyperthermiaHAinvolvesrapidlyheatingdeeptissuestoatargetTc,whichisthensustainedfora
fixedduration.ControlledhyperthermiaHAensuresaprogressiveoverloadapproachwhichinduces
morecompleteadaptationwhilerevealingthemechanismsthatincreaseheattolerance(Taylor,
2014). In contrast to traditional protocols, data presented within this thesis using controlled
hyperthermiaHAdidnotresult intheclassicplateauinphysiologicalresponses.Consideringthe
responsestocontrolledhyperthermiaHAinthecurrentthesis,itislikelythatthismethodmayalter
thetemporalpatterningofadaptationwiththepotentialforsustainedadaptationastheprotocol
progresses.

177
Females’cardiovascularresponsestocontrolledhyperthermiaheatacclimation
Following controlled hyperthermia HA, reductions in resting HR have been observed in males
followingLTHA(-12beats.min-1)comparedwithSTHA(-4beats.min-1)(Garrettetal.2012;Gibson;
Gibson,Turner,etal.2015;Pattersonetal.2004a).Thiswasalsoobservedinthisthesisinfemales.
DatapresentedinStudy3(Chapter6)demonstratesanenhancedreductioninrestingHRfollowing
10-dofcontrolledhyperthermiaHA(-16beats.min-1)comparedwith5-d(-7beats.min-1)infemales.
ThesefindingsarefurthersupportedbythedatapresentedinStudy5(Chapter8)withonlysmall
reductionsinrestingHR(-4beats.min-1)observedfollowing5-dofcontrolledhyperthermiaHA.In
additiontodecreasedrestingHR,controlledhyperthermiaalsoreducedpeakHRduringtheRHTT.
Following STHA, peakHRwas reduced in both Study 3 (Chapter 6; -5 beats.min-1) and Study 5
(Chapter8;-3beats.min-1)infemaleparticipants.Anenhancedreductionwasobservedfollowing
LTHA in Study3 (Chapter 6; -10beats.min-1) and following STHA combinedwith a passiveheat
exposureinStudy5(Chapter8;-12beats.min-1).Thesefindingschallengeoriginalreportsthata
reduction in resting HR plateau following 6-d HA (Armstrong and Maresh, 1991). This further
demonstrates that the controlled hyperthermia method of HA is capable of augmenting the
physiologicaladaptationstoagreaterextentcomparedwithmoretraditionalmethods.
AreducedcardiovascularstrainfollowingHAhasbeenattributedtoanexpansionofPV.Following
controlledhyperthermiaHAanenhancedexpansionofPVisobservedinmalesfollowingLTHA(14
±1%)comparedwithSTHA(7±5%)(Garrettetal.,2014;Gibsonetal.,2015c;Pattersonetal.,2014,
2004b).InStudy5(Chapter8),PVremainedunchangedfollowingSTHA,howeverwhenSTHAwas
combined with a passive heat exposure PV increased by 9.3%, mirroring the changes in HR
responses. These findings suggest that following STHA PV has not reached full expansion,
challenging the original reports that expansion in PV plateau following 6-d HA (Armstrong and
Maresh,1991).
Females’thermoregulatoryresponsestocontrolledhyperthermiaheatacclimation
FollowingcontrolledhyperthermiaHA,anenhancedreductioninrestingTrehasbeenobservedin
males following LTHA (-0.3°C) comparedwith STHA (-0.2°C) (Garrett et al., 2014;Gibson et al.,
2015b;Neal et al., 2015; Patterson et al., 2004a).However,Gibson et al., (2015b) reported no
further reductions in restingTre following10-d (-0.09°C) comparedwith5-d (-0.10°C) controlled
hyperthermia HA. This was not observed for females in this thesis. Data presented in Study 3
(Chapter6)demonstratesnochanges in restingTre following5-d controlledhyperthermiaHA (-
0.02°C)infemales.ThesefindingsarefurthersupportedbythedatapresentedinStudy5(Chapter

178
8) with only a very small reduction in resting Tre (-0.07 °C) observed following 5-d controlled
hyperthermiaHA.However,inlinewithpreviousdataonmales,areductionwasobservedfollowing
LTHAinStudy5(Chapter8;-0.29°C)andfollowingSTHAcombinedwithpassiveheatexposurein
Study5(Chapter8;-0.28°C).AnotherclassicmarkerofHAisareducedexerciseTreowinginpart,
to a reduced resting Tre and improved heat dissipation. Following STHA, peak Tre remained
unchanged in both Study 3 (Chapter 6; -0.06°C) and Study 5 (Chapter 8; -0.05°C). However, a
reduction was observed following LTHA in Study 3 (Chapter 6; -0.47°C) and following STHA
combinedwithapassiveheatexposureinStudy5(Chapter8;-0.42°C).Thesefindingschallenge
original reports that reductions in resting Treplateau following 8-dHA (Armstrong andMaresh,
1991),withfemalesrequiringatleast10-dtoachievedreductionsinrestingTre.
Females’sudomotorresponsestocontrolledhyperthermiaheatacclimation
FollowingcontrolledhyperthermiaHAanenhancedincreaseinSWRhasbeenobservedinmales
followingLTHA(+0.46L.hr-1)comparedwithSTHA(+0.23L.hr-1)(Gibsonetal.,2015b,2015c;Neal
et al., 2015; Patterson et al., 2004a). This was also observed for females in this thesis. Data
presentedinStudy3(Chapter6)demonstratesamarginallyenhancedSWR(+0.78L.hr-1)following
10-dcontrolledhyperthermiaHAcomparedwith5-d(+0.69L.hr-1)infemales.Thesefindingsare
further supported by the data presented in Study 5 (Chapter 8) with large increases in SWR
occurringinthefirst5-dofcontrolledhyperthermiaHA(+0.41L.hr-1).Inthesefemaleparticipants,
approximately90%oftheSWRadaptationoccurredinthefirst5-dofcontrolledhyperthermiaHA.
ThesefindingschallengeoriginalreportsthatanincreaseinSWRplateausbetween8and14-dof
HA(ArmstrongandMaresh,1991)withaplateaupotentiallyoccurringmuchearlier.
The Armstrong and Maresh table for plateau days of physiological adaptation during heat
acclimatisation(ArmstrongandMaresh,1991)hasevolvedtobeatemporalpatterningmodelof
HAacquisition(table9.3).Toachieveaplateauinresponsesfollowing3-6dofHAwaspreviously
viewed favourable. However, taking into account the limitations of fixed intensity, traditional
methodsofHAandtheadvancedunderstandingofthecontrolledhyperthermiamethodofHA,the
plateaumaysimply reflect incompleteadaptation.Recent literature investigating thecontrolled
hyperthermia method of HA combined with data from this thesis on females’ responses to
controlledhyperthermiaHAaddstothetemporalpatterningmodelofHAacquisition.Controlled
hyperthermia HA ensures a progressive overload approach which induces more complete
adaptation thus, often a plateau in responses is not observed. Table 9.4 presents the days of
physiologicaladaptationduringcontrolledhyperthermiaHAinbothmalesandfemalesandusesan
adaptedformattothatofArmstrongandMaresh(1991).Thedatausedtoproducethistableisa

179
combinationofrecentcontrolledhyperthermiaHAstudies(Garrettetal.,2014,2012;Gibsonetal.,
2015b,2015c;Nealetal.,2015;Pattersonetal.,2014,2004a,2004b)anddatapresentedinthis
thesis.
Table9.3Plateaudaysofphysiologicaladaptations(thepointatwhichapproximately85%ofthe
adaptationoccurs)duringheatacclimatisation.
AdaptationDaysofheatacclimatisation
1 2 3 4 5 6 7 8 9 10 11 12 13 14
HRdecrease ______________
PVexpansion ______________
Tredecrease ______________
RPEdecrease ______________
SweatNaCldecrease __________________
SWRincrease ___________________________
RenalNaCldecrease __________________
Notes:HR,heartrate;NaCl,SodiumChloride;PV,plasmavolume;Tre,rectaltemperature;SWR,sweatrate.
AdaptedfromArmstrongandMaresh,(1991).
Inmales,theearlyadaptationsprimarily involveanimprovedcontrolofcardiovascularfunction,
includingexpandedPVanda reducedHR. In females theearlyadaptationsprimarily involvean
increased SWR with an improved control of cardiovascular function requiring ≥10 d. Taken
together, these data would appear to suggest that when using a controlled hyperthermia HA
method the temporal patterning is different to that reported when using constant workload
protocols. The key observation is that following 10-d controlled hyperthermia HA, there is a
sustainedadaptationastheprotocolprogresses.Inaddition,thesedatawouldappeartosuggest
that females adopt a slightly different temporal patterning to males when using a controlled
hyperthermiamethodofHA.

180
Table9.4Daysofphysiologicaladaptation inmalesandfemalesduringcontrolledhyperthermia
heatacclimation.
AdaptationSex
(M/F)
Daysofheatacclimation
1 2 3 4 5 6 7 8 9 10
ReducedHRrestM _________________________________________
F ______________________
PVexpansionM _________________________________________
F ________________
ReducedTrerestM _________________________________________
F ________________
SWRIncreaseM ____________________________
F _________________________
ReducedSweatNaClM __________________________________________
F _____________________________
Notes: Solid line representsmales and thebroken line represents females.HR, heart rate;NaCl, Sodium
Chloride;PV,plasmavolume;Tre,rectaltemperature;SWR,sweatrate.
Females’acquiredthermotolerancefollowingcontrolledhyperthermiaheatacclimation
Inaddition to thephenotypicadaptations tocontrolledhyperthermiadiscussed, this thesisalso
presented the first data set on females acquired cellular thermotolerance associated with
controlled hyperthermia STHA and LTHA. The data provides confidence that when using a
controlledhyperthermiamethodofHAequalsignals for theattainmentof thermotoleranceare
presentinbothmalesandfemales.TheequalHsp72mRNAincreasesoccurringinbothmalesand
females,whowererequiredtocompletearangeofexerciseintensitiestoachievethetargetTc,
suggeststhataslongasaminimumendogenouscriterion(38.5°C)(Gibsonetal.,2014)issurpassed,
additionalexerciseorthermalstimulusisnotoffurtherbenefit.Thesefindingsareinsupportof
recent observations that controlled hyperthermia (38.5°C and 39.0°C) and fixed intensity HA
methodsallelicitsimilarHsp72mRNAresponsesacrossthecourseofLTHAassumingaminimum

181
endogenouscriteriaisachieved(Gibsonetal.,2015a).Takentogether,thesedatawouldappearto
suggestthatexerciseperseisnotasignificantindicatorofHsp72mRNAresponseandthelevelof
hyperthermiaachievedisamoreimportantdriverfortheacquiredthermotolerance.
9.2.3. Acceleratedheatacclimationinfemales
TherewasclearevidenceofHA inStudy5(Chapter8)whencontrolledhyperthermiaSTHAwas
preceded by a sauna-like exposure to 50°C whilst wearing a 100% vinyl sauna suit. This was
demonstratedbyalowerrestingTre(-0.25±0.16°C)andalowerpeakTre(-0.42±0.22°C)following
the RHTT. AdditionalHA responses included, PV expansion (+9.3 ± 7.6%), a lower set point for
sweatingonset(-0.28±0.16°C),areductioninsweatNaCl(-16±10mmol.L-1)andanincreasein
SWR(+0.57±0.20L.hr-1).Furthermore,restingHR(-10±4beats.min-1),peakHR(-12±7beats.min-1), RPE (-3 ± 1), and TS (-0.5 ± 0.5) reduced. The mechanism responsible for establishing this
acceleratedadaptationexposurerequireselucidation.
Thetypeofthethermalloadinginducedduringheatexposure,toelicitadaptation,willinfluence
thevascularadaptationthatoccurs.TheoriginalworkbyFoxetal.(1964)andlaterReganetal.
(1994)demonstratedthatwhileHAisdependentuponthedegreeofTcelevation,theelevationof
Tskisimportantforcompleteadaptation.Duringpassiveheatingcutaneousbloodflowismaximised
sincethereis lesscompetitionfortheavailablecardiacoutput.However,whenexercisinginthe
heatthereisahigherdemandfromactivemuscleforbloodsupply,thuscutaneousbloodflowis
compromised(Kenneyetal.,2014).Thisdifferenceinbloodflowdistributionmayhelpexplainthe
importanceofelevatingTskforbothitsdirectimpactonthebloodvolume(Mawetal.,2000)and
for its role within the heat adaptation process (Taylor, 2014). HA modifies both the central
thresholds and the sensitivity of cutaneous blood flow, with the former reflecting altered
hypothalamicprocessing(Bruck,1986;Horowitz,2014).Thismechanismmayinpartexplainthe
observedadaptationswhencontrolledhyperthermiaHAwasprecededbyasauna-likeexposurein
Study5(Chapter8),withalikelyincreaseinTskoccurringduringthisexposure.
A mechanism which may explain some of the observed reductions in thermoregulatory strain
followingcontrolledhyperthermiaHAcombinedwithasauna-likeexposure isPVexpansion.An
increaseinvenousreturntotheheartenhancesthesensitivityofskinvasodilationinresponseto
increasedTcduringexercise.Noseetal.(1990)reporteda35%increaseinskinbloodflowwitha
150 mL increase in PV following intravenous saline infusion when exercising in a warm
environment.However,thesefindingsarechallengedbyTakenoetal.(2001)whoexaminedthe
effectsofPVexpansiononthesensitivityoftheskinvasodilatoryresponsetoanincreaseinTcina

182
coolandwarmenvironment.Theyfoundthattheincreaseinsensitivitywaslessinacoolthanwarm
environments despite a similar PV expansion. These results suggest that improvements of the
thermoregulatoryresponseafteraerobictrainingareprimarilycausedbyneuraladaptationofthe
thermoregulatorycentreinthehypothalamustorepeatedheatexposureduringtrainingandthat
increasedPVisnotacauseofimprovement,butaresultofadaptation.
Anelevatedsensitivityofthesweatingresponsealsoaccompaniesheatadaptation,andthisitis
apparentwhenheatadaptedindividualsareeitherheatexposed(Foxetal.,1964,1963a;Gotoet
al., 2010; Lorenzo andMinson, 2010; Patterson et al., 2004a), or stimulated pharmacologically
(Collinsetal.,1966).Foxetal.(1967)suggestedthisphenomenonwasduetosweatglandtraining
since inhibition of local sweating during HA resulted in less pronounced sweat flow during
subsequentheat stimulus. Conversely,whenskin regionswere repeatedly stimulatedeitherby
localheating (Foxet al., 1964),orpharmacologically (Collinset al., 1966)without simultaneous
wholebodyheating,thenthoseregionsrespondedtoauniformthermalimpulsewithgreatersweat
flows.Lorenzo&Minson(2010)recentlyverifiedtheselocaltrainingresponseswithanincreased
sensitivityofsweatglandswhenlocallyinfusedwithacetylcholinefollowingHA.ThehigherSWR
duringthesauna-likeexposurescomparedwiththetemperateexposurepriortotheHAsessionsin
chapter 8 may in part, explain the observed adaptations. Sensitivity changes are principally a
peripheralphenomenonhowever,itseemspossiblethatanenhancedsweatingresponsefollowing
HAcouldbeattributedtoacombinationofcentralandperipheralmechanisms.
The responsible mechanism for establishing accelerated adaptation following controlled
hyperthermiaHAcombinedwithapassiveexposuretosauna-likeconditionsrequireselucidation,
but likely includes increased central thresholds and sensitivity of cutaneous blood flow, PV
expansion and an increase in sweat sensitivity. Additional data is required to establish these
mechanismsfullyandverifythesefindingstosupportthedevelopmentofoptimisedHAprotocols.
TogetherthisknowledgecanhelpsupportthedevelopmentofHAforfemalesbyensuringprotocols
targettheforcingfunctionsofHA.
9.3. Directionsforfutureresearch
ThisthesishasintroducedthefirstHTTthatadoptsarunningmodeofexercise.Therepeatability
(Study1, Chapter 4), and the sensitivity (Study2, Chapter 5) of theRHTThasbeenestablished
supporting the use of the RHTT when using a repeated measures design. Furthermore,
retrospectiveanalysiswithinthegeneraldiscussion(9.2.1.)providedevidencetosupporttheuse
oftheRHTTbetweenindependentgroupssinceitappearsthatdespitetheexerciseintensitybeing

183
fixed, the relative heat production is similar between independent groups of different BM and
fitness.FutureresearchisrequiredtoestablishtherepeatabilityandsensitivityofaRHTTthatelicits
matchedheatproductionbetweenindividuals.Thiswouldprovideconfidenceintheabilityofthe
proceduretobeusedwhenassessingthermoregulatoryresponsesbetweenindependentgroups.
Thisthesishasprovidedthefundamentalstudiestosupportfutureresearchindevelopingamore
comprehensive understanding into the mechanisms responsible for the thermoregulatory
adaptations to controlled hyperthermia HA in females. This thesis has taken a very practical
approach with the application to endurance athletes being the predominant focus. Additional
researchisrequiredtodevelopmoreoptimisedHAstrategiesforfemales,tosupporttrainingand
competitionsinhotclimates.
Inthepresentthesis,controlledhyperthermiaHAhasbeenusedtoinvestigatefemales’phenotypic
adaptations.FromthedatapresentedinStudy3(Chapter6),thismethodwasconsideredeffective
when conductedover 10-d.However, 5-dwas not sufficient to establish thermoregulatory and
cardiovascularadjustments,unlesscombinedwithanadditionalpassivethermalstimulus(Study5,
Chapter8).TheseadaptationsareinpartexplainedbyPVexpansion,thussupportingcardiovascular
adjustments, in addition to an increase in evaporative cooling. Lorenzo and Minson, (2010)
investigatedthemechanismsresponsibleforphenotypicadaptationbylocallystimulatingtheskin
withspecificconcentrationsoftheendothelium-dependentvasodilatoryacetylcholine,infusedvia
micro-dialysisandbyperformingastandardisedlocalheatingprotocol,inrestingmales.Theresults
fromthisstudyprovideconfidencethatadaptationsoccurwithintheskins’microcirculationand
sweatglandapparatusfollowingHAinmales,promotingasuperiorthermoregulatoryresponse.In
addition,nochangeswereobservedinmaximalskinbloodflowfollowingHA,demonstratingthat
theobservedchangesareattributable to improvements incutaneousvascular functionandnot
structuralchanges.Developingagreaterlevelofsophisticationwiththeproceduresandmeasures
used toassessHA responses in females,would furtherourunderstanding into themechanisms
responsiblefortheadaptationsobservedinfemales.
Within this thesis, where possible females have been tested during the follicular phase of the
menstrual cycle. Despite the convenience of studying females in the follicular phase of the
menstrualcycle,thebroaderclinicalrelevanceofthefindingsmaybelimitedsincefemalesareonly
inthispartoftheircycleforapproximately50%oftheirreproductivelives.Furthermore,intense
physical exercise has been associatedwith variousmenstrual irregularities including shortened
luteal phase, anovulation and amenorrhea (Loucks and Horvath, 1985). In addition, post-
menopausal women display substantially reduced concentration of these female hormones. It

184
remainsunclearwhetherfemales’heatadaptationiscompromisedasaresultofovarianhormonal
fluctuationsassociatedwithdifferentphasesofthemenstrualcycle,menstrualirregularitiesandin
post-menopausalwomen.Futureresearchisrequiredtoestablishthelevelofacclimationachieved
duringthedifferentphasesofthemenstrualcycleandinfemaleswithmenstrualcycleirregularities.
This informationmaysupport theoptimisationofprotocolsand the timingofadministeringHA
strategies.
Todate,thereisnoliteratureonthetimecourseofthedecayandmaintenanceofheatadaptation
in females.This informationhasahigh levelofpractical importance, toensure theadaptations
achievedremainpresentduringcompetition.Theobservationmadewithinthecurrentthesis,that
thetemporalpatterningoftheinductionisdifferentbetweenmalesandfemales,mayleadoneto
speculate that the temporalpatterningof thedecaymayalsodiffer.Traditionally, forevery2-d
withoutheatexposure,1-dofHAislost,(GivoniandGoldman,1973)withthetemporalpatterning
mirroringtheinduction(ArmstrongandMaresh,1991).Furthermore,thelengthofdecayhasalso
beensuggestedtodependuponphysicalfitnessofparticipantsandthemagnitudeofadaptation
achieved (Pandolf, 1998). Future research is required to determine whether the type of HA
determines the rate of decay since these previous observations were based on traditional HA
methods. Furthermore, research is required to explore the decay rate of adaptations achieved
followingeffectiveSTHAprotocolsinfemales.Inaddition,methodstomaintaintheadaptationsor
reducethespeedofthedecaywouldhaveahighlevelofpracticalimportanceforathletes.
ControlledhyperthermiaHAusedwithinthisthesis, isanexerciseintensivemodelwherebyTcis
rapidlyincreasedbyanexercisestimulusandmaintainedbyacombinationofexerciseandseated
rest.TherearepracticaldisadvantagesofusingexerciseHAprotocols.Theseprotocolscanbecostly
andimpracticalfornon-acclimatedindividualslivingincoolerclimates,astheircompletionrequires
accesstoanenvironmentalchamberortemporaryrelocationtoahotclimate.Furthermore,the
exercise required to ensure an adequate stimulus for adaptation may interfere with athletes
taperingpriortocompetition.Usinganinterventioninvolvingpassiveexposures,forexamplesauna
exposuresorhotwaterimmersionmayovercomeanumberofthepracticallimitationswithcurrent
exerciseHAprotocols.Theuseofasaunaexposureandhotwaterimmersionfollowingtemperate
traininghavebeenreportedtoexpandPV,resultinginareducedthermoregulatoryandperceptual
straininmales(Stanleyetal.,2015;Zurawlewetal.,2015).Researchisrequiredtoestablishthe
effectiveness of these passiveHA strategies in a female population. This body of researchmay
supportafuturelongitudinalstudywhereathletesincorporatepassiveheatexposuresduringtheir
typical temperate training. This will establish whether it is possible to maintain complete

185
acclimationthroughouttraining,avoidingtherequirementforintensiveheat-alleviatingprocedures
andtoofferprotectionduringunexpectedheatstressduringtraining.
9.4. Practicalapplicationoffindings
The RHTT presented within this thesis offers a practical method to assess thermoregulatory
responsestoagivenheatstress,specificallyinrunners,butwithapplicationtootheroccupations
suchasthemilitary.ThepracticalapplicationoftheRHTTistwofold.Firstly,datapresentedwithin
thisthesissupportstheuseoftheRHTTtoassessthermoregulatorychangesthatoccurduetoacute
andchronicheatalleviatinginterventions.Duetothelowtypicalerrorandcoefficientofvariation
associationwith the key thermoregulatory variables, researchers can have confidence that the
effect observed is a true effect and not due to error within the measure. Secondly, the data
presentedwithinthisthesissupportstheuseoftheRHTTtoobserveindividualsthermoregulatory
responsesover time,providing information tophysiologists, coachesandmedical staff tomake
decisions regarding athlete’s safety prior to training and competing in the heat. The practical
importanceoftheRHTTmaystretchasfarassupportingmedicaldecisionsaboutathletes’safety
toreturntotrainingandcompetitionfollowingexertionalheatillnessand/orassociatedsymptoms
(Johnsonetal.,2013).
Thepracticalimportanceofthedatapresentedwithinthethirdexperimentalchapteristhatthe
temporalpatterningtoHAisinpartmediatedbysexdifferences.STHAisapreferredregimefor
athletessinceitiseasiertoadoptwhensustainingqualitytrainingandtaperingperformanceinthe
weeks prior to competition. However, data presented within this thesis provides compelling
evidencethatconsiderationsneedtobemadewhenusingcontrolledhyperthermiaSTHAstrategies
withfemales.STHAmaybeeffectiveinachievingpartialadaptation,howeverfemalesrequireLTHA
to establish reductions in cardiovascular and thermoregulatory strain. Based on this evidence,
physiologists need to ensure that when supporting female athletes with heat-alleviating
interventions, they adopt an adequate number of HA sessions to ensure athletes are fully
acclimated,evidencedbyaplateau(≤5%change)inresponses.Ensuringthatanadequatethermal
stimulus is achievedwill likely support a reduction in thepossibilityofobtainingaheat-related
illnessduringtrainingorcompetitionintheheat.
The practical importance of the data presented in Study 5 (Chapter 8) is that controlled
hyperthermia STHA used in conjunction with a passive heat exposure to sauna-like conditions
provides a practical alternative to LTHA in females. This procedure resulted in reduced
cardiovascular and thermoregulatory strain similar to that observed following LTHA in Study 3

186
(Chapter 6). The use of this combined thermal stimulus provides new knowledge regarding
techniqueswhich can accelerate STHA. Themeasurement of central and peripheral sudomotor
adaptationinStudy5(Chapter8)providesusefulpractical insight intothesudomotoradaptions
that occurs following controlled hyperthermia HA. The use of these techniques over STHA in
females provides new knowledge that may permit better understanding of the possible
mechanismstoexplaintheadaptationsobserved,supportingfurtheroptimisationofHAstrategies.
Inaddition,thisdatasupportsfutureinvestigationsintothebenefitsofpassiveversusexerciseheat
exposuresinfemales.Passiveprocedureswouldbepreferabletoathletessincetherewouldbeless
interferencewithqualitytrainingandtheycouldbeincorporatedmoreeasilyintoataperingperiod
priortocompetition.
Thisdatahaspracticalimportancebeyondyoung,healthy,femaleathletes.Thefindingsfromthese
studiesmay be applied in amanner to support a range of females from both an occupational
population who are exposed to high ambient conditions combined with a high work load, in
addition to the elderly, and those with comorbid medical conditions that may inhibit their
thermoregulatoryability.Withatotalof389exertionalheatillnesscasesreportedinUKmilitary
personnelduringan88monthperiod(September2007-December2014),with236occurringinthe
UK,ofthese125innon-summermonths,heatillnesshasbecomeamajorpriorityofthemilitary
(Staceyetal.,2015).Ensuringourmilitarypersonalareadequatelypreparedfortraining inhigh
ambientconditionsisofupmostimportance.Withthecurrentconsiderationstoallowfemalesinto
groundclosecombatroles,strategieswillneedtobeimplementedtoensuretheirsafetyfroma
heat stress perspective. The information provided within this thesis could support the
implementationofbeneficialstrategiestoreducethelikelihoodoffemalesexperiencingexertional
heatillness.Furthermore,theelderlycomparedwithyoungerindividuals,storemoreheatduring
shortexposurestodryandhumidheat(Stapletonetal.,2014),consequentlythedeathratesduring
heatwavesareconsiderablyhigherintheelderly.Thedatapresentedwithinthisthesiscouldalso
beusedinamannertoinformfutureinvestigationsandheat-alleviatinginterventionsforelderly
females.
9.5. Conclusion
The RHTT introduced within this thesis, offers a simple practical tool to evaluate changes in
thermoregulatoryresponses.DatapresentedwithinthisthesissupportsathletesusingtheRHTTto
monitor changes in thermoregulation over time. To validate the use of the RHTT further, a
longitudinal study is required to assess the effectiveness of the protocol in predicting the
occurrence of exertional heat illness and supporting medical decisions regarding returning to

187
training following an exertional heat illness. Few scientific studies have examined methods to
optimise females’ adaptation during STHA, despite a number of female endurance athletes
suffering from exertional heat illness and/ or experiencing decrements in performance due to
additionalheatstrain.Thevolumeofdatapresentedwithinthisthesisobservingfemales’responses
to HA, collectively provide an original contribution to the literature. The thesis provides novel
information on the temporal patterning of HA; with females’ only achieving partial adaptation
followingSTHAusingacontrolledhyperthermiaprotocol.Inaddition,thisthesisoffersthefirstdata
set toobserve reductions in cardiovascular and thermoregulatory strain in females, following a
novelSTHAprotocol.Theinclusionofapassiveheatexposuretosauna-likeconditions,priortoa
typicalHAprotocol is an excitingnewmethodwhichhasbeen shown to accelerate adaptation
responses infemales.Further investigationsarerequiredtoexamineotherpossiblemethodsby
whichtooptimiseacclimationprotocolsforfemaleathletesanddevelopagreaterunderstanding
intothemechanismsanddifferencesthatmayoccuracrossthemenstrualcycle.Whilstexploring
these methods, it is imperative that the HA procedure remains practical and accessible to all
individuals,whether athleteswhen tapering for competition in a hot climate, ormilitarywhen
preparingfordeployment.

188
References
Altman,D.G.,Royston,P.,2006.Thecostofdichotomisingcontinuousvariables.Br.Med.J.332,1080.Armstrong,L.,Maresh,C.,1991.Theinductionanddecayofheatacclimatisationintrainedathletes.Sport.
Med.12,302–312.Armstrong, L.E., Casa, D.J.,Millard-Stafford,M.,Moran, D.S., Pyne, S.W., Roberts,W.O., 2007. American
CollegeofSportsMedicinepositionstand.Exertionalheatillnessduringtrainingandcompetition.Med.Sci.SportsExerc.39,556–572.
Armstrong,L.E.,Hubbard,R.W.,Deluca,J.P.,Christensen,E.L.,1986.Procedures,selfpacedheatacclimation,in:USArmyResearchInstituteofEnvironmentalMedicine.NatickMassachusetts.
Astrand,P.O.,Saltin,B.,1961.Oxygenuptakeduringthefirstminutesofheavymuscularexercise.J.Appl.Physiol.16,971–976.
Atkinson,G.,Nevill,A.M.,1998.Statisticalmethodsforassessingmeasurementerror(reliability)invariablesrelevanttosportsmedicine.SportMed.26,217–238.
Atkinson,G.,Reilly,T.,1996.Circadianvariationinsportsperformance.Sport.Med.21,292–312.Avellini,B.A.,Kamon,E.,Krajewski,J.T.,1980.Physiologicalresponsesofphysicallyfitmenandwomento
acclimationtohumidheat.J.Appl.Physiol.49,254–261.Bass,D.B.,Kleeman,C.R.,Quinn,M.,Henschel,A.,Hegnauer,A.H.,1955.Mechanismsofacclimatizationto
heatinman.Medicine(Baltimore).34,323–380.Bauman,J.E.,1981.Basalbodytemperature:unreliablemethodofovualtiondetection.Fertil.Steril.36,729–
733.Beaudin, A.E., Clegg,M.E.,Walsh,M.L.,White,M.D., 2009. Adaptation of exercise ventilation during an
actively-inducedhyperthermiafollowingpassiveheatacclimation.Am.J.Physiol.Regul.Integr.Comp.Physiol.297,R605–R614.
Bligh, J., 2006. A theoretical consideration of the means whereby the mammalian core temperature isdefendedatanullzone.J.Appl.Physiol.100,1332–7.
Bligh,J.,Johnson,K.G.,1973.Glossaryoftermsforthermalphysiology.J.Appl.Physiol.35,941–961.Bombardier,E.,Vigna,C., Iqbal, S., Tiidus,P.M.,Tupling,A.R.,2009.Effectsofovarian sexhormonesand
downhillrunningonfiber-type-specificHSP70expressioninratsoleus.J.Appl.Physiol.106,2009–15.Borg,G.A.V.,1962.Psychophysicalbasesofpercievedexertion.Med.Sci.SportExerc.14,337–381.Boulant,J.a,2006.NeuronalbasisofHammel’smodelforset-pointthermoregulation.J.Appl.Physiol.100,
1347–54.Brodeur,V.B.,Dennett,S.R.,Griffin,L.S.,1989.Exertionalhyperthermia,icebaths,andemergencycareatthe
FalmouthRoadRace.J.Emerg.Nurs.15,304–312.Brokenshire, C.S., Armstrong, N., Williams, C.A., 2009. The reliability of adolescent thermoregulatory
responsesduringaheatacclimationprotocol.J.Sport.Sci.Med.8,689–695.Brothers,R.M.,Bhella,P.S.,Shibata,S.,Wingo,J.E.,Levine,B.D.,Crandall,C.G.,2009a.Cardiacsystolicand
diastolicfunctionduringwholebodyheatstress.Am.J.Physiol.Hear.Circ.Physiol.296,1150–1156.Brothers,R.M.,Wingo,J.E.,Hubing,K.a,DelCoso,J.,Crandall,C.G.,2009b.Effectofwholebodyheatstress
onperipheralvasoconstrictionduringlegdependency.J.Appl.Physiol.107,1704–9.Bruck,K.,1986.Basicmechanismsinthermallong-termandshort-termadaptation.J.Therm.Biol.II,73–77.Brunt,V.E.,Miner,J.A.,Meendering,J.R.,Kaplan,P.F.,Minson,C.T.,2012.17-Bestradiolandprogesterone
independentlyaugmentcutaneousthermalhyperemiabutnotreactivehyperemia.Micocirculation18,347–355.
Budd,G.M.,2008.Wet-bulbglobetemperature(WBGT)-itshistoryanditslimitations.J.Sci.Med.Sport11,20–32.
Buono,M.,Martha, S., Heaney, J., 2009a. Peripheral sweat gland function is improvedwith humid heatacclimation.J.Therm.Biol.34,127–130.
Buono,M.,Numan,T.,Claros,R.,Brodine,S.,Kolkhorst,F.,2009b.Isactivesweatingduringheatacclimationrequired for improvements inperipheral sweatgland function?Am. J.Physiol.Regul. Integr.Comp.Physiol.297,R1082–5.
Buono,M.J.,Heaney,J.H.,Canine,K.M.,1998.Acclimationtohumidheatlowersrestingcoretemperature.Am.J.Physiol.274,R1295–R1299.
Burk,A., Timpmann,S.,Kreegipuu,K., Tamm,M.,Unt,E.,Oöpik,V.,2012.Effectsofheatacclimationonendurancecapacityandprolactin response toexercise in theheat.Eur. J.Appl.Physiol.112,4091–4101.

189
Byrne,C.,Lim,C.L.,2007.Theingestibletelemetricbodycoretemperaturesensor:areviewofvalidityandexerciseapplications.Br.J.SportsMed.41,126–133.
Cabanac,M.,2006.Adjustablesetpoint:tohonorHaroldT.Hammel.J.Appl.Physiol.100,1338–46.Calzone,W.L.,Silva,C.,Keefe,D.L.,Stachenfeld,N.S.,2001.Progesteronedoesnotalterosmoticregulation
ofAVP.Am.J.Physiol.Regul.Integr.Comp.Physiol.281,2011–2020.Carpenter,A.J.,Nunneley,S.A.,1988.Endogenoushormonessubtlyalterwomens’responsetoheatstress.J.
Appl.Physiol.65,2313–2317.Casa,D.J.,Guskiewicz,K.M.,Anderson,S.A.,Heck,J.F.,Jimenez,C.C.,McDermott,B.P.,Miller,M.G.,Stearns,
R.L., Swartz, E.E., Walsh, K.M., 2012. National athletic trainers’ association position statement:Preventingsuddendeathinsports.J.Athl.Train.47,96–118.
Castle, P.,Mackenzie, R.W.,Maxwell, N.,Webborn, A.D.J.,Watt, P.W., 2011. Heat acclimation improvesintermittentsprintingintheheatbutadditionalpre-coolingoffersnofurtherergogeniceffect.J.SportsSci.29,1125–34.
Castle,P.C.,Macdonald,A.L.,Philp,A.,Webborn,A.,Watt,P.W.,Maxwell,N.S.,2006.Precoolinglegmuscleimprovesintermittentsprintexerciseperformanceinhot,humidconditions.J.Appl.Physiol.100,1377–1384.
Chapman, A.B., Zamudio, S., Woodmansee, W., Merouani, A., Osorio, F., Johnson, A.N.N., Moore, L.G.,Dahms,T.,Coffin,C.,Abraham,W.T.,Schrier,R.W.,1997.Systemicandrenalhemodynamicchangesinthelutealphaseofthemenstrualcyclemimicearlypregnancy.Am.J.Physiol.273,777–782.
Charkoudian, N., 2010. Mechanisms and modifiers of reflex induced cutaneous vasodilation andvasoconstrictioninhumans.J.Appl.Physiol.109,1221–8.
Charkoudian,N.,2001.Influencesoffemalereproductivehormonesonsympatheticcontrolofthecirculationinhumans.Clin.Auton.Res.11,295–301.
Charkoudian,N.,Halliwill, J.R.,Morgan,B.J.,Eisenach, J.H., Joyner,M.J.,2003. Influencesofhydrationonpost-exercisecardiovascularcontrolinhumans.J.Physiol.552,635–44.
Charkoudian,N.,Johnson,J.M.,2000.Femalereproductivehormonesandthermoregulatorycontrolofskinbloodflow.Exerc.SportSci.Rev.28,108–112.
Cheung, S.S., McLellan, T.M., Tenaglia, S., 2000. The thermophysiology of uncompensable heat stress:Physiologicalmanipulationsandindividualcharacteristics.Sport.Med.29,329–59.
Cheuvront,S.N.,Ely,B.R.,Kenefick,R.W.,Sawka,M.N.,2010.Biologicalvariationanddiagnosticaccuracyofdehydrationassessmentmarkers.Am.J.Clin.Nutr.92,565–573.
Cheuvront, S.N., Kolka,M. a, Cadarette, B.S., Montain, S.J., Sawka,M.N., 2003. Efficacy of intermittent,regionalmicroclimatecooling.J.Appl.Physiol.94,1841–8.
Chinevere,T.D.,Kenefick,R.W.,Cheuvront,S.N.,Lukaski,H.C.,Sawka,M.N.,2008.Effectofheatacclimationonsweatminerals.Med.Sci.SportExerc.40,886–91.
Chomczynski, P., Sacchi, N., 1987. Single-stepmethod of RNA isolation by acid guanidinium thiocyanate-phenol-chloroformextraction.Anal.Biochem.162,156–159.
Chung,S.C.,Goldfarb,A.H.,Jamurtas,A.Z.,Hegde,S.S.,Lee,J.,1999.Effectofexerciseduringthefollicularandlutealphasesonindicesofoxidativestressinhealthywomen.Med.Sci.SportExerc.31,409–413.
Cohen,J.,1988.Astatisticalpoweranalysisforthebehaviouralsciences.RoutledgeAcademic,NewYork.Collins,K.J.,Crockford,G.W.,Weiner,J.S.,1966.Thelocaltrainingeffectofsecretoryactivityontheresponse
ofeccrinesweatglands.J.Physiol.184,203–214.Cooper,K.E.,Cranston,W.I.,Snell,E.S.,1964.Temperatureintheexternalauditorymeatusasanindexof
centraltemperaturechanges.J.Appl.Physiol.19,1032–1034.Cotter,J.D.,Patterson,M.J.,Taylor,N.A.S.,1997.Sweatdistributionbeforeandafterrepeatedheatexposure.
Eur.J.Appl.Physiol.76,181–6.Coyne,M.D.,Kesick,C.M.,Doherty,T.J.,Kolka,M.a,Stephenson,L.a,2000.Circadianrhythmchangesincore
temperatureoverthemenstrualcycle:methodfornoninvasivemonitoring.Am.J.Physiol.Regul.Integr.Comp.Physiol.279,R1316–20.
Cramer,M.N.,Bain,A.R., Jay,O.,2012.Localsweatingontheforehead,butnotforearm, is influencedbyaerobicfitnessindependentlyofheatbalancerequirementsduringexercise.Exp.Physiol.97,572–82.
Cramer, M.N., Jay, O., 2014. Selecting the correct exercise intensity for unbiased comparisons ofthermoregulatoryresponsesbetweengroupsofdifferentmassandsurfacearea.J.Appl.Physiol.116,1123–32.
Crandall,C.G.,Shibasaki,M.,Wilson,T.E.,2010. Insufficientcutaneousvasoconstriction leadinguptoandduringsyncopalsymptomsintheheatstressedhuman.Am.J.Physiol.HeartCirc.Physiol.299,H1168–73.
DeSouze,M.J.,Miller,B.E.,Loucks,A.B.,Luciano,A.A.,Pescatello,L.S.,Campbell,C.G.,Lasley,B.L.,1998.High

190
frequencyoflutealphasedeficiencyandanovulationinrecreationalwomenrunners:Bluntedelevationinfollicle-stimulatinghormoneobservedduringlutealfolliculartransition.J.Clin.Endocrinol.Metab.83,4220–4232.
Decroix,L.,DePauw,K.,Foster,C.,Meeusen,R.,2016.GuidelinestoClassifyFemaleSubjectGroupsinSport-ScienceResearch.Int.J.SportsPhysiol.Perform.11,204–213.
DelCoso,J.,Estevez,E.,Mora-Rodriguez,R.,2009.Caffeineduringexerciseintheheat:Thermoregulationandfluid-electrolytebalance.Med.Sci.Sport.Exerc.41,164–173.
DeMartini,J.K.,Casa,D.J.,Belval,L.N.,Crago,A.,Davis,R.J.,Jardine,J.J.,Stearns,R.L.,2014.Environmentalconditionsandtheoccurrenceofexertionalheatillnessesandexertionalheatstrokeatthefalmouthroadrace.J.Athl.Train.49,478–485.
Denis,P.,Cazor,J.L.,Feret,J.,Weisang,E.,Lefrancois,1976.2,3Diphosphoglycerateredcellconcentrationchangesduringthemenstrualcycleinwomen.Biomedicine25,144.
Dill,D.B.,Costill,D.L.,1974.Calculationofpercentagechangesinvolumesofblood,plasma,andredcellsindehydration.J.Appl.Physiol.37,247–8.
Dill,D.B.,Jones,B.F.,Edwards,H.T.,Oberg,S.A.,1933.Salteconomyinextremedryheat.J.Biol.Chem.100,755–767.
Drew,B.G.,Ribas,V.,Le,J.A.,Henstridge,D.C.,Phun,J.,Zhou,Z.,Soleymani,T.,Daraei,P.,Sitz,D.,Vergnes,L.,Wanagat,J.,Reue,K.,Febbraio,M.A.,Hevener,A.L.,2014.HSP72isamitochondrialstresssensorcriticalforParkinaction,oxidativemetabolism,andinsulinsensitivityinskeletalmuscle.Diabetes63,1488–1505.doi:10.2337/db13-0665
Druyan,A.,Makranz,C.,Moran,D.,2012.Heattolerance inwomen-reconsideringthecriteria.Aviat.Sp.Environ.Med.83,53–60.
DuBois,D.,DuBois,E.F.,1916.Aformulatoestimatetheapproximatesurfaceareaifheightandweightbeknown.Ann.Intern.Med.17,863–871.
Durnin,J.V.G.,Womersley,J.,1974.Bodyfatassessedfortotalbodydensityanditsestimationfromskinfoldthickness:Measurementson481menandwomenaged16-72years.Br.J.Nutr.32,77.
Edwards,R.J.,Belyavin,A.J.,Harrison,M.H.,1978.Coretemperaturemeasurementinman.Aviat.Sp.Environ.Med.49,1289–1294.
Eichna,L.W.,Parl,C.R.,Nelson,N.,Horvath,S.M.,Palmes,E.D.,1950.Threeaspectsofthermalbalancewerestudied:Am.J.Physiol.163,585–597.
ElHelou,N.,Tafflet,M.,Berthelot,G.,Tolaini, J.,Marc,A.,Guillaume,M.,Hausswirth,C.,Toussaint, J.-F.,2012.Impactofenvironmentalparametersonmarathonrunningperformance.PLoSOne7,e37407.
Ely,B.R.,Ely,M.R.,Cheuvront,S.N.,Kenefick,R.W.,Degroot,D.W.,Montain,S.J.,2009.Evidenceagainsta40degreesCcoretemperaturethresholdforfatigueinhumans.J.Appl.Physiol.107,1519–25.
Epstein,Y.,1990.Heatintolerance:predisposingfactororresidualinjury?Med.Sci.SportExerc.22,29–35.Epstein,Y.,Moran,D.S.,Shapiro,Y.,Sohar,E.,Shemer,J.,1999.Exertionalheatstroke:acaseseries.Med.
Sci.SportExerc.31,224–228.Epstein, Y., Shapiro, Y., Shai, B., 1983. Role of surface area-to-mass ratio and work efficiency in heat
intolerance.J.Appl.Physiol.54,831–836.Farrokhyar,F.,Reddy,D.,Poolman,R.W.,Bhandari,M.,2013.Whyperformapriorisamplesizecalculation?
Can.J.Surg.56,207–213.doi:10.1503/cjs.018012Febbraio,M.a,Koukoulas,I.,2000.HSP72geneexpressionprogressivelyincreasesinhumanskeletalmuscle
duringprolonged,exhaustiveexercise.J.Appl.Physiol.89,1055–1060.Fiala,D.,Havenith,G.,Bröde,P.,Kampmann,B.,Jendritzky,G.,2012.UTCI-Fialamulti-nodemodelofhuman
heattransferandtemperatureregulation.Int.J.Biometeorol.56,429–441.Field,A.,2013.DiscoveringstatisticsusingIBMSPSSstatistics,4thed.Sage,London.Flouris,a.D.,Schlader,Z.J.,2015.Humanbehavioralthermoregulationduringexerciseintheheat.Scand.J.
Med.Sci.Sports25,52–64.Flouris,A.D.,2011.Functionalarchitectureofbehaviouralthermoregulation.Eur.J.Appl.Physiol.111,1–8.Flouris,A.D.,Poirier,M.P.,Bravi,A.,Wright-Beatty,H.E.,Herry,C.,Seely,A.J.,Kenny,G.P.,2014.Changesin
heart rate variability during the induction anddecayof heat acclimation. Eur. J. Appl. Physiol. 114,2119–28.
Forman,R.G.,Chapman,M.C.,Steptoe,P.C.,1987.Theeffectofendogenousprogesteroneonbasalbodytemepratureinstimulatedovariancycles.Hum.Reprod.2,631–634.
Fox,R.H.,Goldsmith,R.,Hampton,I.F.,Hunt,T.J.,1967.Heatacclimatizationbycontrolledhyperthermiainhot-dryandhot-wetclimates.J.Appl.Physiol.22,39–46.
Fox,R.H.,Goldsmith,R.,Hampton,I.F.G.,Lewis,H.E.,1964.Thenatureoftheincreaseinsweatingcapacityproducedbyheatacclimatization.J.Physiol.171,368–376.

191
Fox,R.H.,Goldsmith,R.,Kidd,D.J.,Lewis,H.E.,1963a.Acclimatizationtoheatinmanbycontrolledelevationofbodytemperature.J.Physiol.166,530–547.
Fox,R.H.,Goldsmith,R.,Kidd,D.J.,Lewis,H.E.,1963b.Bloodflowandotherthermoregulatorychangeswithacclimatizationtoheat.J.Physiol.548–562.
Frank,A.,Belokopytov,M.,Moran,D.,Shapiro,Y.,Epstein,Y.,2001.Changesinheartratevariabilityfollowingacclimationtoheat.J.BasicClin.Physiol.Pharmacol.12,19–32.
Fritzsche, R.G., Coyle, E.F., 2000. Cutaneous blood flow during exercise is higher in endurance-trainedhumans.J.Appl.Physiol.88,738–744.
Fritzsche,R.G.,Switzer,T.W.,Hodgkinson,B.J.,Coyle,E.F.,1999.Strokevolumedeclineduringprolongedexerciseisinfluencedbytheincreaseinheartrate.J.Appl.Physiol.86,799–805.
Fujii,N.,Honda,Y.,Ogawa,T.,Tsuji,B.,Kondo,N.,Koga,S.,Nishiyasu,T.,2012.Short-termexercise-heatacclimationenhancesskinvasodilationbutnothyperthermichyperpneainhumansexercisinginahotenvironment.Eur.J.Appl.Physiol.112,295–307.
Gagge,A.P.,Stolwijk, J.A.J., Saltin,H.,1969.Comfortand thermal sensationsandassociatedphysiologicalresponsesduringexerciseatvariousambienttemperatures.Environ.Res.2,209–229.
Gagnon,D.,Dorman,L.E.,Jay,O.,Hardcastle,S.,Kenny,G.P.,2009.Coretemperaturedifferencesbetweenmalesandfemalesduringintermittentexercise:physicalconsiderations.Eur.J.Appl.Physiol.105,453–61.
Gagnon,D.,Jay,O.,Kenny,G.P.,2013.Theevaporativerequirementforheatbalancedetermineswhole-bodysweatrateduringexerciseunderconditionspermittingfullevaporation.J.Physiol.591,2925–35.
Gagnon, D., Jay, O., Lemire, B., Kenny, G.P., 2008. Sex-related differences in evaporative heat loss: theimportanceofmetabolicheatproduction.Eur.J.Appl.Physiol.104,821–9.
Gagnon, D., Kenny, G.P., 2012. Sex differences in thermoeffector responses during exercise at fixedrequirementsforheatloss.J.Appl.Physiol.113,746–57.
Gagnon,D.,Kenny,G.P.,2011.Sexmodulateswhole-bodysudomotorthermosensitivityduringexercise.J.Physiol.589,6205–17.
Galloway,S.D.R.,Maughan,R.J.,1997.Effectsofambienttemperatureonthecapacitytoperformprolongedcycleexerciseinman.Med.Sci.SportExerc.29,1240–1249.
Garrett,aT.,Goosens,N.G.,Rehrer,N.J.,Patterson,M.J.,Harrison,J.,Sammut,I.,Cotter,J.D.,2014.Short-termheatacclimationiseffectiveandmaybeenhancedratherthanimpairedbydehydration.Am.J.Hum.Biol.26,311–20.
Garrett,A.T., Creasy,R., Rehrer,N.J., Patterson,M.J., Cotter, J.D., 2012. Effectivenessof short-termheatacclimationforhighlytrainedathletes.Eur.J.Appl.Physiol.112,1827–37.
Garrett,A.T.,Goosens,N.G.,Rehrer,N.J.,Rehrer,N.G.,Patterson,M.J.,Cotter,J.D.,2009.Inductionanddecayofshort-termheatacclimation.Eur.J.Appl.Physiol.107,659–670.
Garrett, A.T., Rehrer, N.J., Patterson,M.J., 2011. Induction and decay of short-term heat acclimation inmoderatelyandhighlytrainedathletes.Sport.Med.41,757–771.
Geddes,L.A.,Baker,L.E.,1967.Thespecificresistanceofbiologicalmaterial-acompendiumofdataforthebiomedicalengineerandphysiologist.Med.Biol.Eng.5,271–293.
Gibson, O.R., Dennis, A., Parfitt, T., Taylor, L., Watt, P.W., Maxwell, N.S., 2014. Extracellular Hsp72concentration relates to aminimum endogenous criteria during acute exercise-heat exposure. CellStressChaperones19,389–400.
Gibson,O.R.,Mee,J.A.,Taylor,L.,Tuttle,J.A.,Watt,P.W.,Maxwell,N.S.,2015a.Isothermicandfixed-intensityheatacclimationmethodselicitequalincreasesinHsp72mRNA.Scand.J.Med.Sci.Sports25,259–268.
Gibson,O.R.,Mee,J.A.,Tuttle,J.A.,Taylor,L.,Watt,P.W.,Maxwell,N.S.,2015b.Isothermicandfixedintensityheatacclimationmethodsinducesimilarheatadaptationfollowingshortandlong-termtimescales.J.Therm.Biol.49-50,55–65.
Gibson,O.R.,Turner,G.,Tuttle,J.A.,Taylor,L.,Watt,P.W.,Maxwell,N.S.,2015c.HeatAcclimationattenuatesphysiologicalstrainandtheHsp72,butnotHsp90αmRNAresponsetoacutenormobarichypoxia.J.Appl.Physiol.119,889–899.
Gillum,T.,Kuennen,M.,Gourley,C.,Dokladny,K.,Schneider,S.,Moseley,P.,2013.Sexdifferencesinheatshockprotein72expressioninperipheralbloodmononuclearcellstoacuteexerciseintheheat.Int.J.Endocrinol.Metab.11,8739.
Gisolfi,C.V.,Wenger,C.B.,1984.Temperature regulationduringexercise:oldconcepts,new ideas.Exerc.SportSci.Rev.12,339–372.
Givoni, B., Goldman, R.F., 1973. Predicting effects of heat acclimatization on heart rate and rectaltemperature.J.Appl.Physiol.35,875–879.
Gledhill,N.,Jamnik,R.,1994.Enduranceathletes’stokevolumedoesnotplateau:Majoradvantageisdiastolic

192
function.Med.Sci.SportExerc.26,1116–1121.Gonzalez-Alonso,J.,2003.Reductionsinsystemicandskeletalmusclebloodflowandoxygendeliverylimit
maximalaerobiccapacityinhumans.Circulation107,824–830.González-Alonso,J.,Crandall,C.G.,Johnson,J.M.,2008.Thecardiovascularchallengeofexercisingintheheat.
J.Physiol.586,45–53.Gonzalez,R.R.,Gagge,A.P.,1976.Warmdiscomfortandassociatedthermoregulatorychangesduringdryand
humidheatacclimatization.Isr.J.Med.Sci.12,804–807.Goto, M., Okazaki, K., Kamijo, Y., Ikegawa, S., Masuki, S., Miyagawa, K., Nose, H., 2010. Protein and
carbohydratesupplementationduring5-dayaerobictrainingenhancedplasmavolumeexpansionandthermoregulatoryadaptationinyoungmen.J.Appl.Physiol.109,1247–55.
Grucza,R.,Pekkarinen,H.,Titov,E.K.,Kononoff,A.,Hanninen,O.,1993.Influenceofthemenstrualcycleandoralcontraceptivesonthermoregulatoryresponsestoexerciseinyoungwomen.Eur.J.Appl.Physiol.67,279–285.
Guy, J.H., Deakin, G.B., Edwards, A.M., Miller, C.M., Pyne, D.B., 2015. Adaptation to hot environmentalconditions: an exploration of the performance basis, procedures and future directions to optimiseopportunitiesforeliteathletes.Sport.Med.45,303–11.
Hammel, H.T., Jackson, D.C., Stolwijk, J.A.J., Hardy, D., Stromme, S.B., 1963. Temperature regulation byhypothalamicproportionalcontrolwithanadjustablesetpoint.J.Appl.Physiol.18,1146–1154.
Harper-Smith, A.D., Crabtree, D.R., Bilzon, J.L.J.,Walsh, N.P., 2010. The validity of wireless iButtons andthermistorsforhumanskintemperaturemeasurement.Physiol.Meas.31,95–114.
Harrison,M.H.,1985.Effectsonthermalstressandexerciseonbloodvolumeinhumans.Physiol.Rev.65,149–209.
Havenith, G., 2001. Human surface tomass ratio and body core temperature in exercise heat stress—aconceptrevisited.J.Therm.Biol.26,387–393.
Havenith, G.,Middendorp, H. van, 1990. The relative influence of physical fitness, acclimatization state,anthropometricmeasuresandgenderonindividualreactionstoheatstress.Eur.J.Appl.Physiol.Occup.Physiol.61,419–427.
Hawes,R.,McMorran,J.,Vallis,C.,2010.Exertionalheatillnessinhalfmarathonrunners:experiencesoftheGreatNorthRun.Emerg.Med.J.27,866–867.
Hayden, G.,Milne, H.C., Patterson,M.J., Nimmo,M. a, 2004. The reproducibility of closed-pouch sweatcollectionandthermoregulatoryresponsestoexercise-heatstress.Eur.J.Appl.Physiol.91,748–51.
Hayward,J.S.,Eckerson,J.D.,Kemna,D.,1984.Thermalandcardiovascularchangesduringthreemethodsofresuscitationfrommildhypothermia.Resuscitation11,21–33.
Hessemer,V.,Bruck,K.,1985.Influenceofmenstrualcycleonthermoregulatory,metabolic,andheartrateresponsestoexerciseatnight.J.Appl.Physiol.59,1911.
Hopkins,W.G.,2000a.Reliabilityfromconsecutivepairsoftrials.Sport.Sci.4.Hopkins,W.G.,2000b.Measuresofreliabilityinsportsmedicineandscience.Sport.Med.30,1–15.Horowitz,M.,2014.Heatacclimation,epigenetics,andcytoprotectionmemory.Compr.Physiol.4,199–230.Horowitz,M.,2003.Matchingthehearttoheat-inducedcirculatoryload:heat-acclimatoryresponses.News
Physiol.Sci.18,215–21.Horowitz,M.,Kodesh,E.,2010.Molecularsignalsthatshapetheintegrativeresponsesoftheheat-acclimated
phenotype.Med.Sci.Sport.Exerc.42,2164–2172.Horowitz,M.,Peyser,Y.M.,Muhlrad,A.,1986a.Alterationsincardiacmyosinisoenxymesdistributionasan
adaptationtochronicenvironmentalheatstressintherat.J.Mol.Cell.Cardiol.18,511–515.Horowitz,M.,Robinson,S.D.M.,2007.Heatshockproteinsandtheheatshockresponseduringhyperthermia
anditsmodulationbyalteredphysiologicalconditions.Prog.BrainRes.162,433–446.Horowitz,M.,Shimoni,Y.,Parnes,S.,Gotsman,M.S.,Hasin,Y.,1986b.Heatacclimation:cardiacperformance
ofisolatedratheart.J.Appl.Physiol.60,9–13.Horvath,S.M.,Drinkwater,B.L.,1982.Thermoregulationandthemenstrualcycle.Aviat.Sp.Environ.Med.
53,790–794.Houmard, J.A., Costill, D.L., Davis, J.A.,Mitchell, J.B., Pascoe, D.D., Robergs, R.A., 1990. The Influence of
exerciseintensityonheataccliamtionintrainedsubjects.Med.Sci.SportExerc.22,615–620.Howe,A.S.,Boden,B.P.,2007.Heat-relatedillnessinathletes.Am.J.SportsMed.35,1384–95.Ilsley,A.H.,Rutten,A.J.,Runciman,W.B.,1983.Anevaluationofbodytemperaturemeasurement.Anaestheic
IntensiveCare11,31–39.Inoue,Y.,Havenith,G.,Kenney,W.L.,Loomis,J.L.,Buskirk,E.R.,1999.Exercise-andmethylcholine-induced
sweatingresponses inolderandyoungermen:effectofheatacclimationandaerobic fitness. Int. J.Biometeorol.42,210–6.

193
Inoue,Y.,Tanaka,Y.,Omori,K.,Kuwahara,T.,Ogura,Y.,Ueda,H.,2005.Sex-andmenstrualcycle-relateddifferencesinsweatingandcutaneousbloodflowinresponsetopassiveheatexposure.Eur.J.Appl.Physiol.94,323–32.
James,C.A.,Richardson,A.J.,Watt,P.W.,Maxwell,N.S.,2014.Reliabilityandvalidityof skin temperaturemeasurementbytelemetrythermistorsandathermalcameraduringexercise intheheat.J.Therm.Biol.45,141–149.
JansedeJonge,X.A.K.,2003.Effectsofthemenstrualcycleonexerciseperformance.Sport.Med.33,833–51.
Jay,O.,Bain,A.R.,Deren, T.M., Sacheli,M.,Cramer,M.N.,Hi,V., 2011. Largedifferences inpeakoxygenuptakedonotindependentlyalterchangesincoretemperatureandsweatingduringexercise.Am.J.Physiol.Regul.Integr.Comp.Physiol.301,832–841.
Jay,O.,Kenny,G.,2007.Thedeterminationofchangesinbodyheatcontentduringexerciseusingcalorimetryandthermometry.J.Human-EnvironmentalSyst.10,19–29.
Johnson,E.C.,Kolkhorst,F.W.,Richburg,A.,Schmitz,A.,Martinez,J.,Armstrong,L.E.,2013.Specificexerciseheatstressprotocol foratriathlete’sreturnfromexertionalheatstroke.Curr.SportsMed.Rep.12,106–9.
Johnson,J.M.,Kellogg,D.L.,2010.Localthermalcontrolofthehumancutaneouscirculation.J.Appl.Physiol.109,1229–38.
Jones,A.,Doust,J.,1998.Thevalidityofthelactateminimumtestfordeterminationofthemaximallactatesteadystate.Med.Sci.SportExerc.30,1304–1313.
Jones,A.M.,2007.Middleand longdistancerunning, in:Winter,E.M., Jones,A.M.,RichardDavison,R.C.,Bromley,P.D.,Mercer,T.H.(Eds.),SportandExercisePhysiologyTestingGuidelines.RoutledgeTaylorandFrancisGroup,LondonandNewYork,pp.147–154.
Jones,A.M.,Doust,J.H.,1996.A1%treadmillgrademostaccuratelyreflectstheenergeticcostofoutdoorrunning.J.SportsSci.14,321–327.
Jurkowski, J.E., Jones,N.L.,Toews,C.J.,Sutton, J.R.,1981.Effectsofmenstrualcycleonblood lactate,O2delivery,andperformanceduringexercise.J.Appl.Physiol.51,1493–1499.
Kaciuba-Uscilko,H.,Grucza,R.,2001.Genderdifferencesinthermoregulation.Curr.Opin.Clin.Nutr.Metab.Care4,533–6.
Keatisuwan,W.,Ohnaka,T.,Tochihara,Y.,1996.PhysiologicalresponsesofmenandwomenduringexerciseinhotenvironemtnswithequivalentWBGT.Appl.Hum.Sci.15,249–258.
Kenefick,R.W.,Cheuvront,S.N.,Sawka,M.N.,2007.Thermoregulatoryfunctionduringthemarathon.Sport.Med.37,312–315.
Kenefick,R.W.,Sawka,M.N.,2007.Heatexhaustionanddehydrationascausesofmarathoncollapse.Sport.Med.37,378–381.
Kenefick,R.W.,Sollanek,K.J.,Charkoudian,N.,Sawka,M.N.,2014.Impactofskintemperatureandhydrationonplasmavolumeresponsesduringexercise.J.Appl.Physiol.117,413–20.
Kenney,W.L.,1985.Areviewofcomparativeresponsesofmenandwomentoheatstress.Environ.Res.37,1–11.
Kenney,W.L.,Hodgson,J.L.,1987.Heattolerance,thermoregulationandageing.Sport.Med.4,446–456.Kenney,W.L., Stanhewicz, A.E., Bruning, R.S., Alexander, L.M., 2014. Blood pressure regualtion III:What
happenswhenonesystemmustservetwomasters:Temperatureandpressureregulation?Eur.J.Appl.Physiol.114,467–479.
Kenny,G.P.,Jay,O.,2013.Thermometry,calorimetry,andmeanbodytemperatureduringheatstress.Compr.Physiol.3,1689–719.
Kenny, G.P., Webb, P., Ducharme, M.B., Reardon, F.D., Jay, O., 2008. Calorimetric measurement ofpostexercisenetheatlossandresidualbodyheatstorage.Med.Sci.SportsExerc.40,1629–36.
Kolka,M.A.,Stephenson,L.A.,1997.Effectoflutealphaseelevationincoretemperatureonforearmbloodflowduringexerciseheat.J.Appl.Physiol.82,1079–1083.
Kolka,M.A., Stephenson, L.A., 1989.Control of sweatingduring thehumanmenstrual cycle. Eur. J.Appl.Physiol.Occup.Physiol.58,890–895.
Kondo,N.,Shibasaki,M.,Aoki,K.,Koga,S.,Inoue,Y.,Crandall,C.G.,2011.Functionofhumaneccrinesweatglandsduringdynamicexerciseandpassiveheatstress.J.Appl.Physiol.90,1877–1881.
Kregel,K.C.,2002.Molecularbiologyof thermoregulation InvitedReview:Heat shockproteins:modifyingfactorsinphysiologicalstressresponsesandacquiredthermotolerance.J.Appl.Physiol.92,2177–2186.
Kresfelder,T.,Claassen,N.,Cronje,M.,2006.Hsp70inductionandhsp70genepolymorphismsasindicatorsofacclimatizationunderhyperthermicconditions.J.Therm.Biol.31,406–415.
Kuennen,M.,Gillum,T.,Dokladny,K.,Bedrick,E.,Schneider,S.,2011.Thermotoleranceandheatacclimation

194
mayshareacommonmechanisminhumans.Am.J.Physiol.Regul.Integr.Comp.Physiol.1,524–533.Kuwahara,T.,Inoue,Y.,Abe,M.,Sato,Y.,Kondo,N.,2005.Effectsofmenstrualcycleandphysicaltrainingon
heatlossresponsesduringdynamicexerciseatmoderateintensityinatemperateenvironment.Am.J.Physiol.Regul.Integr.Comp.Physiol.228,R1347–R1353.
Lakens,D.,2013.Calculatingandreportingeffectsizestofacilitatecumulativescience:apracticalprimerfort-testsandANOVAs.Front.Psychol.4,1–12.
Lebrun,C.M.,McKenzie,D.C.,Prior, J.C., Taunton, J.E., 1995.Effectsofmenstrual cyclephaseonathleticperformance.Med.Sci.SportExerc.27,437–444.
Lee, B.J., Miller, A., James, R.S., Thake, C.D., 2016. Cross acclimation between heat and hypoxia: Heatacclimationimprovescellulartoleranceandexerciseperformanceinacutenormobarichypoxia.Front.Physiol.7,1–15.
Lee,S.H.,Kwon,H.M.,Kim,Y.J.,Lee,K.M.,Kim,M.,Yoon,B.W.,2004.EffectsofHsp70.1geneknockoutonthe mitochondrial apoptotic pathway after focal cerebral ischemia. Stroke 35, 2195–2199.doi:10.1161/01.STR.0000136150.73891.14
Lee, S.M.C., Williams, W.J., Schneider, S.M., 2000. Core temperature measurement during submaximalexercise:Esophageal,rectal,andintestinaltemperatures.Aviat.Sp.Environ.Med.71,939–945.
Lefrant,J.Y.,Muller,L.,deLaCoussaye,J.E.,Benbabaali,M.,Lebris,C.,Zeitoun,N.,Mari,C.,Saïssi,G.,Ripart,J., Eledjam, J.J., 2003. Temperaturemeasurement in intensive care patients: comparisonof urinarybladder, oesophageal, rectal, axillary, and inguinalmethods versus pulmonary artery coremethod.IntensiveCareMed.29,414–8.
Levine, T.R., Hullett, C.R., 2002. Eta Squared, Partial Eta Squared, and Misreporting of Effect Size inCommunicationResearch.Hum.Commun.Res.28,612–625.doi:10.1111/j.1468-2958.2002.tb00828.x
Libert, J.P., Candas, V., Sagot, J.C., Meyer, J.P., Vogt, J.J., Ogawa, T., 1984. Contribution of skin thermalsensitivitiesoflargebodyareastosweatingresponse.Jpn.J.Physiol.34,75–88.
Locke,M.,1997.Thecellularstressresponsetoexercise:roleofstressproteins.Exerc.SportSci.Rev.25,105–136.
Lorenzo,S.,Halliwill,J.R.,Sawka,M.N.,Minson,C.T.,2010.Heatacclimationimprovesexerciseperformance.J.Appl.Physiol.109,1140–1147.
Lorenzo, S.,Minson, C.T., 2010. Heat acclimation improves cutaneous vascular function and sweating intrainedcyclists.J.Appl.Physiol.109,1736–1743.
Loucks,A.B.,Horvath,S.M.,1985.Athleticamenorrhea:areview.Med.Sci.SportExerc.17,56–72.Low, D.A., Vu, A., Brown, M., Davis, S.L., Keller, D.M., Levine, B.D., Crandall, C.G., 2007. Temporal
thermometryfailstotrackbodycoretemperatureduringheatstress.Med.Sci.SportExerc.39,1029–1035.
Low,P..,2004.Evaluationofsudomotorfunction.Clin.Neurophysiol.115,1506–13.Lundvall,J.,Lindgren,P.,1998.F-cellshiftandproteinlossstronglyaffectvalidityofPVreductionsindicated
byHb/Hctandplasmaproteins.J.Appl.Physiol.84,822–829.Magalhães,F.D.C.,Amorim,F.T.,Passos,R.L.F.,Fonseca,M.A.,Oliveira,K.P.M.,Lima,M.R.M.,Guimarães,J.B.,
Ferreira-Júnior,J.B.,Martini,A.R.P.,Lima,N.R.V,Soares,D.D.,Oliveira,E.M.,Rodrigues,L.O.C.,2010.Heat and exercise acclimation increases intracellular levels of Hsp72 and inhibits exercise-inducedincreaseinintracellularandplasmaHsp72inhumans.CellStressChaperones15,885–95.
Maloyan,A.,Palmon,A.,Horowitz,M.,1999.HeatacclimationincreasesthebasalHSP72levelandaltersitsproductiondynamicsduringheatstress.Am.J.Physiol.276,R1506–15.
Marsh,S.A.,Jenkins,D.G.,2002.Physiologicalresponsestothemenstrualcycle.SportMed.32,601–614.Marshall, H.C., Campbell, S. a., Roberts, C.W.,Nimmo,M. a., 2007. Human physiological and heat shock
protein72adaptationsduringtheinitialphaseofhumid-heatacclimation.J.Therm.Biol.32,341–348.Marshall, H.C., Ferguson, R.A., Nimmo, M.A., 2006. Human resting extracellular heat shock protein 72
concentrationdecreasesduring the initial adaptation toexercise in ahot, humidenvironment. CellStressChaperones11,129–134.
Martin,D.E.,1997.Climaticheatstressandathleticperformance.TrackCoach139.Mauger, A.R.,Metcalfe, A.J., Taylor, L., Castle, P.C., 2013. The efficacy of the self pacedVO2max test to
measuremaximaloxygenuptakeintreadmillrunning.Appl.Physiol.Nutr.Metab.38,1211–1216.Maughan,R.J.,Mcarthur,M.,Shirreffs,S.M.,1996.Influenceofmenstrualstatusonfluidreplacementafter
exerciseinduceddehydrationinhealthyyoungwomen.Br.J.SportMed.30,41–47.Maw,G.J.,Mackenzie,I.L.,Comer,D.A.,Taylor,N.A.,1996.Wholebodyhyperhydrationinendurance-trained
malesdeterminedusingradionuclidedilution.Med.Sci.SportExerc.28,1038–1044.Maw, G.J., Mackenzie, I.L., Taylor, N.., 2000. Can skin temperature manipulation, with minimal core
temperaturechange,influenceplasmavolumeinrestinghumans?Eur.J.Appl.Physiol.81,159–162.

195
McClung,J.P.,Hasday,J.D.,He,J.R.,Montain,S.J.,Cheuvront,S.N.,Sawka,M.N.,Singh,I.S.,2008.Exercise-heatacclimationinhumansaltersbaselinelevelsandexvivoheatinducibilityofHSP72andHSP90inperipheralbloodmononuclearcells.Am.J.Physiol.Regul.Integr.Comp.Physiol.294,R185–191.
Mee, J.A., Gibson, O.R., Doust, J., Maxwell, N.S., 2015. A comparison of males and females’ temporalpatterningtoshort-andlong-termheatacclimation.Scand.J.Med.Sci.Sports25,250–258.
Mekjavic,I.B.,2006.Contributionofthermalandnonthermalfactorstotheregulationofbodytemperatureinhumans.J.Appl.Physiol.100,2065–2072.
Miller,P.B.,Soules,M.R.,1996.TheusefulnessofaurinaryLHkitforovulationpredictionduringmenstrualcyclesofnormalwomen.Am.J.Obstet.Gynecol.87,13–17.
Mitchell, D., Wyndham, C.H., 1969. Comparision of weighting formulas for calculating mean skintemperature.J.Appl.Physiol.26,616.
Montain, S.J., Sawka,M.N., Cadarette, B.S., Quigley,M.D.,McKay, J.M., 1994. Physiological tolerance touncompensable heat stress: effects of exercise intensity, protective clothing, and climate. J. Appl.Physiol.77,216–222.
Mora-rodriguez,R.,Coso,J.D.,Hamouti,N.,Estevez,E.,Ortega,J.F.,2010.Aerobicallytrainedindividualshavegreater increases in rectal temperature than untrained ones during exercise in the heat at similarrelativeintensities.Eur.J.Appl.Physiol.109,973–981.
Moran,D.S.,Eli-Berchoer,L.,Heled,Y.,Mendel,L.,Schocina,M.,Horowitz,M.,2006.Heatintolerance:doesgenetranscriptioncontribute?J.Appl.Physiol.100,1370–1376.
Moran,D.S.,Erlich,T.,Epstein,Y.,2007.Theheattolerancetest:anefficientscreeningtoolforevaluatingsusceptibilitytoheat.J.SportRehabil.16,215–21.
Moran,D.S.,Heled,Y.,Still,L.,Laor,A.,Shapiro,Y.,2004.Assessmentofheattoleranceforpostexertionalheatstrokeindividuals.Med.Sci.Monit.10,CR252–CR257.
Moran,D.S.,Mendal,L.,2002.Coretemperaturemeasurement:methodsandcurrentinsights.Sport.Med.32,879–85.
Moran,D.S.,Shapiro,Y.,Laor,A.,Izraeli,S.,Pandolf,K.B.,1999.Cangenderdifferencesduringexercise-heatstressbeassessedbythephysiologicalstrainindex?Am.J.Physiol.Regul.Integr.Comp.Physiol.276,R1798–R1804.
Moran,D.S.,Shitzer,A.,Pandolf,K.B.,1998.Aphysiologicalstrainindextoevaluateheatstress.Am.J.Physiol.Regul.Integr.Comp.Physiol.275,R129–R134.
Morton,J.P.,Holloway,K.,Woods,P.,Cable,N.T.,Burniston,J.,Evans,L.,Kayani,A.C.,McArdle,A.,2009.Exercise training-induced gender-specific heat shock protein adaptations in human skeletalmuscle.MuscleNerve39,230–233.
Moseley,P.L.,1997.Heatshockproteinsandheatadaptationofthewholeorganism.J.Appl.Physiol.83,1413–1417.
Nadal,E.R.,Bullard,R.W.,Stolwijk,J.A.,1971.Importanceofskintemperatureintheregulationofsweating.J.Appl.Physiol.31,80–87.
Nadal,E.R.,Cafarelli,E.,Roberts,M.F.,Wenger,C.B.,1979.Circulatoryregulationduringexerciseindifferentambianttemperatures.J.Appl.Physiol.46,430–437.
Nakamura,K.,Matsumura,K.,Hübschle,T.,Nakamura,Y.,Hioki,H.,Fujiyama,F.,Boldogköi,Z.,König,M.,Thiel,H.-J.,Gerstberger,R., Kobayashi, S., Kaneko, T., 2004. Identificationof sympatheticpremotorneuronsinmedullaryrapheregionsmediatingfeverandotherthermoregulatoryfunctions.J.Neurosci.24,5370–80.
Nakayama,T.,Suzuki,M., Ishizuka,N.,1975.Actionofprogesteroneonpreopticthermosensitiveneurons[abstract].Nature258.
Neal,R.A.,Corbett,J.,Massey,H.C.,Tipton,M.J.,2015.Effectofshort-termheatacclimationwithpermissivedehydration on thermoregulation and temperate exercise performance. Scand. J.Med. Sci. Sports.doi:10.1111/sms.12526
Nelson,M.D.,Haykowsky,M.J.,Petersen,S.R.,Delorey,D.S.,Stickland,M.K.,Cheng-baron,J.,Thompson,R.B.,2010.Aerobicfitnessdoesnotinfluencethebiventricularresponsetowholebodypassiveheatstress.J.Appl.Physiol.109,1545–1551.
Nielsen,B.,1996.OlympicsinAtlanta:afightagainstphysics.Med.Sci.SportExerc.28,665–668.Nielsen,B.Y.B.,Hales,J.R.S.,Strange,S.,Juel,N.,1993.Humancirculatoryandthermoregulatoryadaptations
withheatacclimationandexerciseinahot,dryenvironment.J.Physiol.460,467–485.Nose,H.,Mack,G..,Shi,X.R.,Nadel,E.R.,1988.Shiftinbodyfluidcompartmentsafterdehydrationinhumans.
J.Appl.Physiol.65,318–324.Nose,H.,Mack,G.W.,Shi,X.R.,Morimoto,K.,Nadel,E.R.,1990.Effectofsalineinfusionduringexerciseon
thermalandcirculatoryregulations.J.Appl.Physiol.69,609–616.

196
Nunneley,S.,1978.Physiologicalresponsesofwomentothermalstress:Areview.Med.Sci.Sports10,250–255.
Nybo,L.,Nielsen,B.,2001.Hyperthermiaandcentralfatigueduringprolongedexerciseinhumans.J.Appl.Physiol.91,1055–60.
Nybo,L.,Rasmussen,P.,Sawka,M.N.,2014.Performanceintheheat-physiologicalfactorsofimportanceforhyperthermia-inducedfatigue.Compr.Physiol.4,657–89.
Nybo,L.,Secher,N.H.,Nielsen,B.,2002.Inadequateheatreleasefromthehumanbrainduringprolongedexercisewithhyperthermia.J.Physiol.545,697–704.
Ogita,H.,Node,K.,Kitakaze,M.,2003.Theroleofestrogenandestrogen-relateddrugs incardiovasculardiseases.Curr.DrugMetab.4,497–504.
Pandolf,K.B.,1998.Timecourseofheatacclimationanditsdecay.Int.J.SportsMed.19,S157–S160.Paroo,Z.,Dipchand,E.S.,Noble,E.G.,2002.EstrogenattenuatespostexerciseHSP70expressioninskeletal
muscle.Am.J.Physiol.CellPhysiol.282,C245–51.Patterson, M.J., Stocks, J.M., Taylor, N. a S., 2014. Whole-body fluid distribution in humans during
dehydration and recovery, before and after humid-heat acclimation induced using controlledhyperthermia.ActaPhysiol.210,899–912.
Patterson,M.J.,Stocks,J.M.,Taylor,N.aS.,2004a.Humidheatacclimationdoesnotelicitapreferentialsweatredistributiontowardthelimbs.Am.J.Physiol.Regul.Integr.Comp.Physiol.286,R512–R518.
Patterson,M.J.,Stocks,J.M.,Taylor,N.aS.,2004b.Sustainedandgeneralizedextracellularfluidexpansionfollowingheatacclimation.J.Physiol.559,327–34.
Périard,J.D.,2013.Cardiovasculardeterminantsinvolvedinpacingunderheatstress.Sport.Med.43,643–5.Périard,J.D.,Cramer,M.N.,Chapman,P.G.,Caillaud,C.,Thompson,M.W.,2011.Cardiovascularstrainimpairs
prolongedself-pacedexerciseintheheat.Exp.Physiol.96,134–44.Périard,J.D.,Jay,O.,Alonso,J.M.,Coutts,A.J.,Flouris,A.D.,González-Alonso,J.,Hausswirth,C.,Lee,J.K.W.,
Nassis,G.P.,Nybo,L.,Pluim,B.M.,Roelands,B.,Sawka,M.N.,Wingo, J.,Racinais,S.,2016.Author’sReplytoBrocherieandMillet:‘IstheWet-BulbGlobeTemperature(WGBT)Index.Sport.Med.46,139–141.
Périard, J.D., Racinais, S., Sawka, M.N., 2015. Adaptations and mechanisms of human heat acclimation:Applicationsforcompetitiveathletesandsports.Scand.J.Med.Sci.Sports25,20–38.
Périard, J.D., Ruell, P., Caillaud, C., Thompson,M.W., 2012. Plasma Hsp72 (HSPA1A) and Hsp27 (HSPB1)expressionunderheatstress:influenceofexerciseintensity.CellStressChaperones17,375–83.
Pockley,G.,2003.Heatshockproteinsasregulatorsoftheimmuneresponse.Lancet362,469–76.Poirier,M.P.,Gagnon,D.,Friesen,B.J.,Hardcastle,S.G.,Kenny,G.P.,2015.Whole-bodyheatexchangeduring
heatacclimationanditsdecay.Med.Sci.SportExerc.47,390–400.Prud’Homme, D., Bouchard, C., Leblanc, C., Landry, F., Fontaine, E., 1984. Sensitivity ofmaximal aerobic
powertotrainingisgenotypedependent.Med.Sci.SportExerc.18,489–493.Racinais,S.,Alonso,J.M.,Coutts,a.J.,Flouris,a.D.,Girard,O.,González-Alonso,J.,Hausswirth,C.,Jay,O.,
Lee, J.K.W.,Mitchell,N.,Nassis,G.P.,Nybo, L., Pluim,B.M.,Roelands,B., Sawka,M.N.,Wingo, J.E.,Périard,J.D.,2015.Consensusrecommendationsontrainingandcompetingintheheat.Scand.J.Med.Sci.Sports25,6–19.
Racinais,S.,Mohr,M.,Buchheit,M.,Voss,S.C.,Gaoua,N.,Grantham,J.,Nybo,L.,2012.Individualresponsestoshort-termheatacclimatisationaspredictorsoffootballperformanceinahot,dryenvironment.Br.J.SportsMed.46,810–5.
Racinais,S.,Périard,J.D.,Karlsen,A.,Nybo,L.,2015.Effectofheatandheatacclimatizationoncyclingtimetrialperformanceandpacing.Med.Sci.SportsExerc.47,601–6.
Ramanathan,N.L.,1964.Anewweightingsystemformeansurfacetemperatureofthehumanbody.J.Appl.Physiol.19,531–533.
Randall,W.C.,1946.Quantificationandregionaldistributionofsweatglandsinman.J.Clin.Invest.25,761–767.
Rasmussen, P.,Nybo, L., Volianitis, S.,Møller, K., Secher,N.H.,Gjedde,A., 2010. Cerebral oxygenation isreducedduringhyperthermicexerciseinhumans.ActaPhysiol.199,63–70.
Regan,J.M.,Macfarlane,D.J.,Taylor,N.A.S.,1996.Anevaluationoftheroleofskintemperatureduringheatadaptation.ActaPhysiol.Scand.158,365–375.
Regan, J.M.,Macfarlane,D.J.,Taylor,N.A.S.,1994.Adaptation toheat:differences inducedby isothermalstrain following heat acclimation and thermoneutral exercise, in: International Conference onEnvironmentalErgonomics.pp.45–46.
RichardDavison,R.C.,Wooles,A.L.,2007.Cycling,in:Winter,E.,Jones,A.,RichardDavison,R.C.,Bromley,P.D.,Mercer,T.H.(Eds.),SportandExercisePhysiologyTestingGuidlines.RoutledgeTaylorandFrancis

197
Group,LondonandNewYork,pp.160–164.Richards,R.,Richards,D.,1984.ExertioninducedheatexhaustionandothermedicalaspectsoftheCity-to-
Surffunruns,1978-1984.Med.J.Aust.141,799–805.Roberts,M.F.,Wenger,C.B.,Stolwijk,J.A.,Nadal,E.R.,1977.Skinbloodflowandsweatingchangesfollowing
exercisetrainingandheatacclimation.J.Appl.Physiol.43,133–137.Roberts,W.O., 2010. Determining a “do not start” temperature for amarathon on the basis of adverse
outcomes.Med.Sci.SportExerc.42,226–232.Roberts,W.O.,2006.exertionalheatstrokeduringacoolweathermarathon:acasestudy.Med.Sci.Sport
Exerc.38,1197–1203.Roberts,W.O.,2000.A12-yrprofileofmedicalinjuryandillnessfortheTwinCitiesMarathon.Med.Sci.Sport
Exerc.32,1549–1555.Robertson,B.,Walter,E.,2010.“Coolrunnings”:heatstrokeincoolconditions.Emerg.Med.J.27,387–388.Robinson,J.,Charlton,J.,Seal,R.,Spady,D.,Joffres,M.R.,1998.Oesophageal,rectal,axillary,tympanic,and
pulmonaryarterytemperaturesduringcardiacsurgery.Can.J.Anaesth.45,317–323.Robinson,S.,1963.Temperatureregulationinexercise.Pediatrics32,691–702.Rowell,L.B.,Kraning,K.K.,Kennedy,J.W.,Evans,T.O.,1967.Centralcirculatoryresponsestoworkindryheat
beforeandafteracclimatization.J.Appl.Physiol.22,509–518.Ruell, P.A., Simar,D., Périard, J.D., Best, S., Caillaud, C., Thompson,M.W., 2014. Plasmaand lymphocyte
Hsp72responsestoexerciseinathleteswithpriorexertionalheatillness.AminoAcids46,1491–9.Saat, M., Sirisinghe, R.G., Singh, R., Tochihara, Y., 2005. Effects of short-term exercise in the heat on
thermoregulation, blood parameters, sweat secretion and sweat composition of tropic-dwellingsubjects.J.Physiol.Anthropol.Appl.HumanSci.24,541–549.
Saltin,B.,Hermansen,L.,1966.Esophageal,rectalandmuscletemperatureduringexercise.J.Appl.Physiol.21,1757–1762.
Sandström,M.E.,Siegler,J.C.,Lovell,R.J.,Madden,L.a,McNaughton,L.,2008.Theeffectof15consecutivedaysofheat-exerciseacclimationonheatshockprotein70.CellStressChaperones13,169–75.
Sato,F.,Owen,M.,Matthes,R.,Sato,K.,Gisolfi,C.V,1990.Functionalandmorphologicalglandwithheatacclimationchangesintheeccrinesweatglandwithheatacclimation.J.Appl.Physiol.69,232–236.
Sato,K.,Dobson,R.L.,1970.Regionalandindividualvariationsinthefunctionofthehumaneccrinesweatgland.J.Investig.Dematology54,443–449.
Sato,K.,Dobson,R.L.,Mali,J.W.H.,1971.Enxymaticbasisfortheactivetransportofsodiumintheeccrinesweatgland.J.Invest.Dermatol.57,10–16.
Sato,K.,Kang,W.H.,Saga,K.,Sata,K.T.,1989.Biologyofsweatglandsandtheirdisorders.I.Normalsweatglandfunction.J.Am.Acad.Dermotology20,537–563.
Sato,K.,Sato,F.,1983.Individualvariationsinstructureandfunctionofhumaneccrinesweatgland.Am.J.Physiol.Regul.Integr.Comp.Physiol.245,R203–R208.
Sawka,M.N., Burke, L.M., Eichner, E.R., Maughan, R.J., Montain, S.J., Stachenfeld, N.S., 2007. AmericanCollegeofSportsMedicinepositionstand.Exerciseandfluidreplacement.Med.Sci.SportExerc.39,377–390.
Sawka,M.N.,Coyle,E.F.,1999.Influenceofbodywaterandbloodvolumeonthermoregulatoryandexerciseperformanceintheheat.Exerc.SportSci.Rev.27,167–218.
Sawka, M.N., Gonzalez, R.R., Young, A.J., Dennis, R.C., Valarie, R.C., Pandolf, K.B., 1989. Control ofthermoregulatoryduringexerciseintheheat.Am.J.Physiol.Regul.Integr.Comp.Physiol.26,311–316.
Sawka,M.N.,Leon,L.R.,Montain,S.J.,Sonna,L.A.,2011a.Integratedphysiologicalmechanismsofexerciseperformance,adaptation,andmaladaptationtoheatstress.Compr.Physiol.1,1883–1928.
Sawka,M.N.,Toner,M.M.,Francesconi,R.P.,Pandolf,K.B.,1983.Hypohydrationandexercise:effectsofheatacclimation,genderandenvironment.J.Appl.Physiol.55,1147–1153.
Sawka,M.N.,Wenger,C.B.,Pandolf,K.B.,2011b.Thermoregulatoryresponsestoacuteexerciseheatstressandheatacclimation.Compr.Physiol.157–185.
Sawka,M.N.,Wenger,C.B.,Pandolf,K.B.,1993.Humanresponsestoexerciseheatstress.Schaible, T.F., Scheuer, J., 1979. Effects of physical training by running or swimming on ventricular
performanceofrathearts.J.Appl.Physiol.Repiratory,Environ.Exerc.Physiol.46,854–60.Scherr, J.,Wolfarth, B., Christle, J.W., Pressler, A.,Wagenpfeil, S., Halle,M., 2013. Associations between
Borg’s rating of perceived exertion and physiological measures of exercise intensity. Eur. J. Appl.Physiol.113,147–55.
Schlader,Z.J.,Raman,A.,Morton,R.H.,Stannard,S.R.,Mündel,T.,2011a.Exercisemodalitymodulatesbodytemperatureregulationduringexerciseinuncompensableheatstress.Eur.J.Appl.Physiol.111,757–66.

198
Schlader,Z.J.,Simmons,S.E.,Stannard,S.R.,Mündel,T.,2011b.Physiology&Behavior:Theindependentrolesoftemperatureandthermalperceptioninthecontrolofhumanthermoregulatorybehavior.Physiol.Behav.103,217–224.
Schmittgen,T.D.,Livak,K.J.,2008.Analyzingreal-timePCRdatabythecomparativeCTmethod.Nat.Protoc.3,1101–1108.
Schutte,P.,2010.Heatstressmanagementinhotmines,in:Brune,J.P.(Ed.),ExtractingtheScience:ACenturyofMiningResearch.USA,pp.30–34.
Scoon,G.S.M.,Hopkins,W.G.,Mayhew,S.,Cotter,J.D.,2007.Effectofpost-exercisesaunabathingontheenduranceperformanceofcompetitivemalerunners.J.Sci.Med.Sport10,259–262.
Senay,L.C.,1973.Bodyfluidandtemperatureresponsesofheatexposedwomenbeforeandafterovulationwithandwithoutrehydration.J.Physiol.232,209–219.
Senay, L.C.,Mitchell, D.,Wyndham, C.H., 1976. Acclimatization in a hot, humid environment: body fluidadjustments.J.Appl.Physiol.40,786–796.
Senf,S.M.,Howard,T.M.,Ahn,B.,Ferreira,L.F., Judge,A.R.,2013.Lossofthe InducibleHsp70DelaystheInflammatoryResponsetoSkeletalMuscleInjuryandSeverelyImpairsMuscleRegeneration.PLoSOne8.
Shapiro,Y.,Hubbard,R.W.,Kimbrough,C.M.,Pandolf,K.B.,1981.Physiologicalandhematologicalresponsestosummerandwinterdryheatacclimation.J.Appl.Physiol.50,792–798.
Shapiro,Y.,Magazanik,A.,Udassin,R.,Ben-Baruch,G.,Shvartz,E.,Shoenfeld,Y.,1979.Heatintoleranceinformerheatstrokepatients.Ann.Intern.Med.90,913–916.
Shibasaki,M.,Wilson,T.E.,Crandall,C.G.,2006.Neuralcontrolandmechanismsofeccrinesweatingduringheatstressandexercise.J.Appl.Physiol.100,1692–1701.
Shinohara,T.,Takahashi,N.,Ooie,T., Ichinose,M.,Hara,M.,Yonemochi,H., Saikawa,T.,Yoshimatsu,H.,2004. Estrogen inhibits hyperthermia-induced expression of heat-shock protein 72 andcardioprotectionagainstischemia/reperfusioninjuryinfemaleratheart.J.Mol.Cell.Cardiol.37,1053–1061.
Shvartz, E., Bhattacharya, A., Sperinde, S.J., Brock, P.J., Sciaraffa, D., Van Beaumont,W., 1979. Sweatingresponsesduringandmoderateconditioning.J.Appl.Physiol.46,675–680.
Silva,N.L.,Boulant, J.a,1986.Effectsof testosterone,estradiol,and temperatureonneurons inpreoptictissueslices.Am.J.Physiol.250,R625–R632.
Silver,J.T.,Noble,E.G.,2012.Regulationofsurvivalgenehsp70.CellStressChaperones17,1–9.Skidmore,R.,Gutierrez,J.a,GuerrieroJr,V.,Kregel,K.C.,1995.HSP70inductionduringexerciseandheat
stressinrats:roleofinternaltemperature.Am.J.Physiol.268,R92–R97.Smith,A.,Crabtree,D.,Bilzon,J.,Walsh,N.,2010.ThevalidityofwirelessiButtonsandthermistorsforhuman
skintemperaturemeasurement.Physiol.Meas.31,95–114.Stacey,M.J.,Parsons,I.T.,Woods,D.R.,Taylor,P.N.,Ross,D.,JBrett,S.,2015.Susceptibilitytoexertionalheat
illnessandhospitalisationriskinUKmilitarypersonnel.Br.Med.J.OpenSportExerc.Med.1,e000055.Stachenfeld, N.S., Pietro, L.D.I., Palter, S.F., Nadel, E.R., Nina, S., Dipietro, L., Steven, F., 1998. Estrogen
influencesosmoticsecretionofAVPandbodywaterbalanceinpostmenopausalwomen.Am.J.Physiol.Regul.Integr.Comp.Physiol.274,187–195.
Stachenfeld,N.S.,Silva,C.,Keefe,D.L.,2000.Estrogenmodifiesthetemperatureeffectsofprogesterone.J.Appl.Physiol.88,1643–1649.
Stachenfeld,N.S.,Silva,C.,Keefe,D.L.,Kokoszka,C.A.,Nadel,E.R.,1999.Effectsoforalcontraceptivesonbodyfluidregulation.J.Appl.Physiol.87,1016–1025.
Stachenfeld, N.S., Taylor, H.S., 2014. Challenges and methodology for testing young healthy women inphysiologicalstudies.Am.J.Endocrinol.Metab.06519,849–853.
Stanley,J.,Halliday,A.,D’Auria,S.,Buchheit,M.,Leicht,A.S.,2015.Effectofsauna-basedheatacclimationonplasmavolumeandheartratevariability.Eur.J.Appl.Physiol.115,785–94.
Stapleton,J.M.,Larose,J.,Simpson,C.,Flouris,A.D.,Sigal,R.J.,Kenny,G.P.,2014.Doolderadultsexperiencegreaterthermalstrainduringheatwaves?Appl.Physiol.Nutr.Metab.39,292–8.
Stephenson, L.A., Kolka, M.A., 1999. Esophageal temperature threshold for sweating decreases beforeovulationinpremenopausalwomen.J.Appl.Physiol.86,22–28.
Stephenson,L.A.,Kolka,M.A.,1993.ThermoregulationinWomen.Exerc.SportSci.Rev.21,231–262.Stephenson,L.A.,Kolka,M.A.,1985.Menstrualcyclephaseandtimeofdayalterreferencesignalcontrolling
armbloodflowandsweating.Am.J.Physiol.Regul.Integr.Comp.Physiol.249,R186–R191.Sunderland,C.,Morris,J.G.,Nevill,M.E.,2008.Aheatacclimationprotocolforteamsports.Br.J.SportsMed.
42,327–333.Syrop,C.H.,Hammond,M.G.,1987.Diurnalvariationsinmidlutealserumprogesteronemeasurements.Fertil.

199
Steril.47,67–70.Takamata, A., Yoshida, T., Nishida, N., Morimoto, T., 2001. Relationship of osmotic inhibition in
thermoregulatoryresponsesandsweatsodiumconcentrationinhumans.Am.J.Physiol.Regul.Integr.Comp.Physiol.280,623–629.
Takeno,Y.,Kamijo,Y.I.,Nose,H.,2001.Thermoregulatoryandaerobicchangesafterendurancetraininginahypobarichypoxicandwarmenvironment.J.Appl.Physiol.91,1520–8.
Tankersley, C.G., Nicholas, W.C., Deaver, D.R., Mikita, D., Kenney, W.L., 1992. Estrogen replacement inmiddle-agedwomen :thermoregulatoryresponsestoexerciseintheheat.J.Appl.Physiol.73,1238–1245.
Taylor,L.,Midgley,A.W.,Chrismas,B.,Hilman,A.R.,Madden,L.a,Vince,R.V,McNaughton,L.R.,2011.DailyhypoxiaincreasesbasalmonocyteHSP72expressioninhealthyhumansubjects.AminoAcids40,393–401.
Taylor,N.A.S.,2000.Principlesandpracticesofheatadaptation.J.Hum.EnvionmentalSyst.4,11–22.Taylor,N.A.S.,Cotter,J.D.,2006.Heatadaptation:Guidlinesfortheoptimisationofhumanperformance.Int.
J.SportsMed.7,33–57.Taylor,N.A.S.,Tipton,M.J.,Kenny,G.P.,2014.Considerationsforthemeasurementofcore,skinandmean
bodytemperatures.J.Therm.Biol.46,72–101.doi:10.1016/j.jtherbio.2014.10.006Taylor,N.S.,2014.Humanheatadaptation.Compr.Physiol.4,325–65.Taylor,W.F.,Johnson,J.M.,O’Leary,D.,Park,M.K.,1984.Effectofhighlocaltemperatureonreflexcutaneous
vasodilation.J.Appl.Physiol.57,191–196.Tenaglia,S.a,McLellan,T.M.,Klentrou,P.P.,1999.Influenceofmenstrualcycleandoralcontraceptiveson
tolerancetouncompensableheatstress.Eur.J.Appl.Physiol.Occup.Physiol.80,76–83.Tetievsky,A.,Cohen,O.,Eli-Berchoer,L.,Gerstenblith,G.,Stern,M.D.,Wapinski,I.,Friedman,N.,Horowitz,
M.,2008.Physiologicalandmolecularevidenceofheatacclimationmemory:a lesson fromthermalresponsesandischemiccross-toleranceintheheart.Physiol.Genomics34,78–87.
Toner,M.M.,Drolet,L.L.,Pandolf,K.B.,1986.Perceptualandphysiologicalresponsesduringexerciseincoolandcoldwater.Percept.Mot.Skills62,211–220.
Tucker,R.,Marle,T.,Lambert,E.V,Noakes,T.D.,2006.Therateofheatstoragemediatesananticipatoryreductioninexerciseintensityduringcyclingatafixedratingofperceivedexertion.J.Physiol.574,905–915.
Tucker, R., Rauch, L., Harley, Y.X.R., Noakes, T.D., 2004. Impaired exercise performance in the heat isassociatedwithananticipatoryreductioninskeletalmusclerecruitment.Eur.J.Physiol.448,422–30.
Turk,J.,Thomas,I.R.,1975.Artificialacclimatizationtoheat.Ann.Occup.Hygeine17,271–278.Tuttle, J. a,Castle,P.C.,Metcalfe,A.J.,Midgley,A.W.,Taylor, L., Lewis,M.P.,2015.Downhill runningand
exerciseinhotenvironmentsincreaseleukocyteHsp72(HSPA1A)andHsp90α(HSPC1)genetranscripts.J.Appl.Physiol.118,996–1005.
Vincent,W.J.,1995.StatisticsinKinesiology.HumanKinetics.Webb,P.,1995.ThePhysiologyofheatregualtion.Am.J.Physiol.Regul.Integr.Comp.Physiol.268,R838–
R850.Weller,A.S.,Linnane,D.M.,Jonkman,A..,Daanen,H.A.M.,2007.Quantificationofthedecayandre-induction
ofheatacclimationindry-heatfollowing12and26dayswithoutexposuretoheatstress.Eur.J.Appl.Physiol.102,57–66.
Wells,C.L.,Horvath,S.M.,1974.Responsestoexerciseinahotenvironmentasrelatedtothemenstrualcycle.J.Appl.Physiol.36,299–302.
Whipp,B.J.,Wasserman,K.,1972.Oxygenuptakekinetics forvarious intensitiesofconstant-loadwork. J.Appl.Physiol.33,351–356.
Williams,T.J.,Krahenbuhl,G.S.,1997.Menstrualcyclephaseandrunningeconomy.Med.Sci.SportExerc.29,1609–1618.
Wilson,T.E.,Brothers,R.M.,Tollund,C.,Dawson,E.a,Nissen,P.,Yoshiga,C.C.,Jons,C.,Secher,N.H.,Crandall,C.G.,2009.EffectofthermalstressonFrank-Starlingrelationsinhumans.J.Physiol.587,3383–92.
Wilson,T.E.,Cui,J.,Zhang,R.,Crandall,C.G.,2006.Heatstressreducescerebralbloodvelocityandmarkedlyimpairsorthostatictoleranceinhumans.Am.J.Physiol.Regul.Integr.Comp.Physiol.291,R1443–8.
Wingo,J.E.,Low,D.a,Keller,D.M.,Brothers,R.M.,Shibasaki,M.,Crandall,C.G.,2010.Skinbloodflowandlocal temperature independentlymodify sweat rate during passive heat stress in humans. J. Appl.Physiol.109,1301–6.
Wyndham,C.H.,1973.Thephysiologyofexerciseunderheatstress.Annu.Rev.Physiol.35,193–220.Wyndham,C.H.,1951.Effectofacclimatizationoncirculatoryresponsestohighenvironmentaltemperatures.
J.Appl.Physiol.4,383–395.

200
Wyndham,C.H.,Benade,aJ.,Williams,C.G.,Strydom,N.B.,Goldin,A.,Heyns,aJ.,1968.Changesincentralcirculationandbodyfluidspacesduringacclimatizationtoheat.J.Appl.Physiol.25,586–593.
Wyndham,C.H., Rogers,G.G., Senay, L.C.,Mitchell,D., 1976.Acclimization in ahot, humidenvironment:cardiovascularadjustments.J.Appl.Physiol.40,779–785.
Yaglou,C.P.,Minard,D.,1957.Controlofheatcasualtiesatmilitarytrainingcenters.AMAArch.Ind.Heal.16,302–316.
Yamada,P.M.,Amorim,F.T.,Moseley,P.,Robergs,R.,Schneider,S.M.,2007.Effectofheatacclimationonheatshockprotein72andinterleukin-10inhumans.J.Appl.Physiol.103,1196–204.
Yamazaki,F.,Hamasaki,K.,2003.Heatacclimationincreasesskinvasodilationandsweatingbutnotcardiacbaroreflexresponsesinheat-stressedhumans.J.Appl.Physiol.95,1567–1574.
Yoda,T.,Crawshaw,L.I.,Nakamura,M.,Saito,K.,Konishi,A.,Nagashima,K.,Uchida,S.,Kanosue,K.,2005.Effectsofalcoholonthermoregulationduringmildheatexposureinhumans.Alcohol36,195–200.
Zurawlew,M.J.,Walsh,N.P.,Fortes,M.B.,Potter,C.,2015.Post-exercisehotwaterimmersioninducesheatacclimation and improves endurance exercise performance in the heat. Scand. J. Med. Sci. Sport.doi:10.1111/sms.12638

201
Appendices
Informedconsent
INFORMEDCONSENTFORM
Declaration
I herebyvolunteertotakepartinthisresearchprojectwhichinvestigates
Theprincipalinvestigatorhasexplainedtomysatisfactionthepurposeoftheexperimentandthepossiblerisksinvolved.IhavehadtheprinciplesandtheprocedureexplainedtomeandIhavealsoreadtheparticipantinformationsheet.Iunderstandtheprinciplesandproceduresfully.
IamawarethatIwillberequiredto:
Havemyaerobiccapacityandbodycompositionmeasured
Performcyclingexerciseforupto90-mininhotconditionsonoccasions
Havecoretemperaturemonitoredviarectalthermometryonmultipleoccasions.
Iunderstandhowthedatacollectedwillbeused,andthatanyconfidentialinformationwillnormallybeseenonlybytheresearchersandwillnotberevealedtoanyoneelse.
Iunderstandthat Iamfreetowithdrawfromthe investigationatanytimeandthat Iamundernoobligationtogivereasonsforwithdrawalortoattendagainforexperimentation.
IagreethatshouldIwithdrawfromthestudy,thedatacollecteduptothatpointmaybeusedbytheresearcherforthepurposesdescribedintheinformationsheet.
Iunderstandthattheresultsofthestudycanbemadeknowntome.
Furthermore, if I am a student, I am aware that taking part, or not taking part in this experiment, will neither bedetrimentalto,norfurthermypositionasastudent.
Iunderstandtoobeythelaboratory/studyregulationsandtheinstructionsoftheinvestigatorsregardingsafety,subjectonlytomyrighttowithdrawdeclaredabove.
SignatureoftheSubject Date
SignatureoftheInvestigator Date

202
Medicalquestionnaire
MEDICALQUESTIONNAIRE
Name Age
Areyouingoodhealth? Yes No
IfNo,pleaseprovidegivedetails
Howoftendoyoucurrentlyparticipateinvigorousphysicalactivity?
<Oncepermonth
Oncepermonth
2-3timesperweek
4-5timesperweek
5+timesperweek
Haveyousufferedfromaseriousillnessoraccident? Yes No
Ifyes,pleasegiveparticulars:
Doyousuffer,orhaveyoueversufferedfrom:
RespiratoryProblems(e.g.Asthma,Bronchitis?) Yes No
Diabetes? Yes No
Epilepsy? Yes No
High/LowBloodPressure Yes No
Cardiovascularproblems Yes No
Areyoucurrentlytakingmedicationordietarysupplements? Yes No
Ifyes,pleasegivedetails:
Inthelast3months,haveyouconsultedyourGPforanycondition? Yes No
Ifyes,pleasegiveparticulars
Areyoucurrently/recentlytakingpartinanyotherlaboratoryexperiments? Yes No

203
PLEASEREADTHEFOLLOWINGCAREFULLY
Personswillbeconsideredunfittoparticipateinthestudyifthey:
Areunsureofthetestprotocolandthepossiblerisksanddiscomfortsdesignatedonthesubjectinformationsheet;
Theanswersgivenonthemedicalquestionnaireorinformedconsentformdonotmeettherequiredcriteria;
Havesuspendedtrainingduetoaninjury
Haveaknownhistoryofmedicaldisordersincluding,highbloodpressureandheartorlungdisease
Havebeenverified,ordocumentedashavinganybloodcarriedinfections(Hepatitis,HIV),
Havehadapreviousincidenceofhyperthermia,heatstrokeorotherheat-relatedillness
Havesymptomsofnausea,lightheadednessoranaphylacticshocktoneedles,probesorothermedicaltypeequipment
Havechronicoracutesymptomsofgastrointestinalbacterialinfections
Haveahistoryofinfectiousdiseases
DECLARATION
I herebyvolunteertobeaparticipantinexperimentsandinvestigationsconductedduringtheperiodcommencing
MyrepliestotheabovequestionsarecorrecttothebestofmybeliefandIunderstandthattheywillbetreatedwiththestrictestconfidence.Theexperimenterhasfullyinformedmeof,andIhaveunderstood,thepurposeoftheexperimentandpossiblerisksinvolved.
IunderstandthatImaywithdrawfromtheexperimentatanytimeandthatIamundernoobligationtogivereasonsforwithdrawalortoattendagainforexperimentation.
Iundertaketoobeythelaboratory/studyregulationsandtheinstructionsoftheexperimenterregardingsafety,subjectonlytomyrighttowithdrawdeclaredabove.
SignatureoftheSubject Date
SignatureoftheInvestigator Date

204
Menstrualcyclequestionnaire
Werequireinformationregardingyourmenstrualcycletiming.Wherepossibletestingwillbescheduledinthefollicularphaseofthemenstrualcycle(Day3–10).
Wemayrequireavenousbloodsampletoquantifyconcentrationsof17-ßestradiolandprogesterone.Thiswillenableustoconfirmwhetherthetestingwasconductedduringthecorrectphaseofthemenstrualcycle.
Itisessentialthatyoutakeyourtimetocompletethisquestionnairetoensureaccuracyoftheinformationprovided.
Whatistheaveragelengthofyourcycle?
Areyourmenstrualcyclesregular?
Pleaseprovidethestartdateofyouprevioustwomenstrualperiods?
Werethesetwocyclestypicalofyouraveragecycle?
Doyoutakeoralcontraceptiveorusehormonereplacement?
Ifsopleaseprovidedetails:
Pleaseprovideanyadditionalnoteswhichmayberelevant:
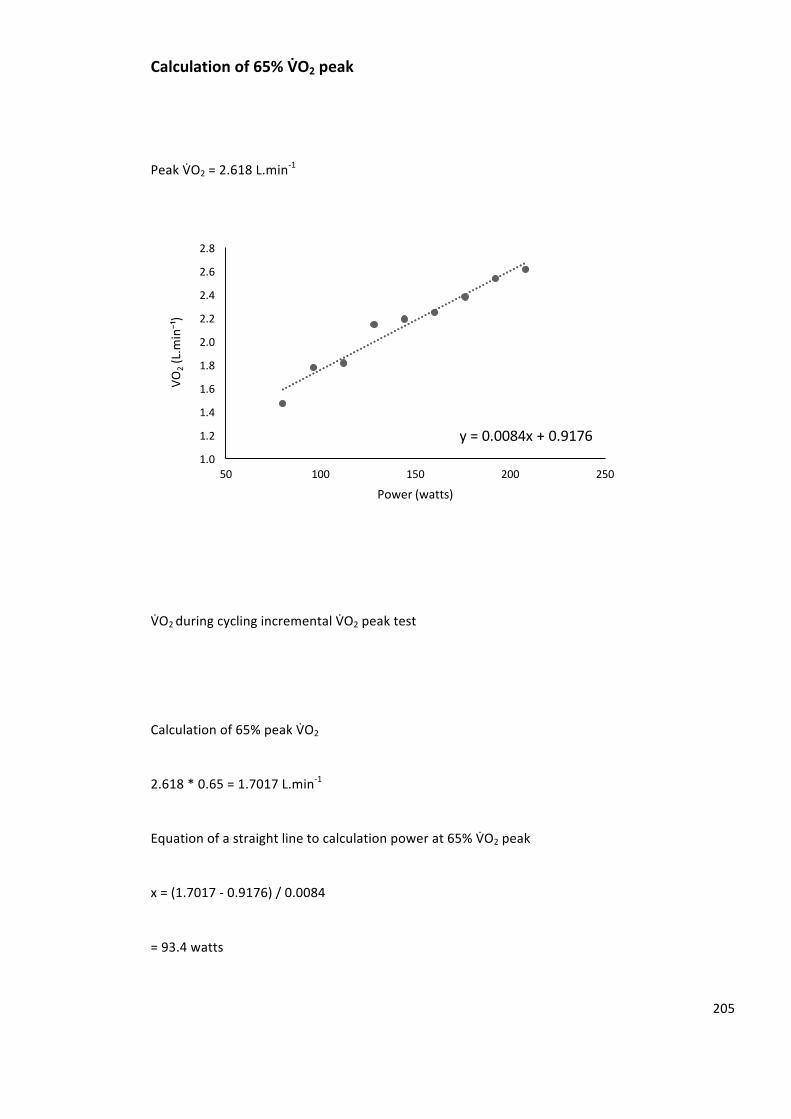
205
Calculationof65%VbO2peak
PeakVCO2=2.618L.min-1
VCO2duringcyclingincrementalVCO2peaktest
Calculationof65%peakVCO2
2.618*0.65=1.7017L.min-1
Equationofastraightlinetocalculationpowerat65%VCO2peak
x=(1.7017-0.9176)/0.0084
=93.4watts
y=0.0084x+0.91761.0
1.2
1.4
1.6
1.8
2.0
2.2
2.4
2.6
2.8
50 100 150 200 250
VO2(L.m
in⁻¹)
Power(watts)





























