Embed Size (px)
Citation preview

JPET #123356
1
Reciprocal regulation of eNOS and NADPH oxidase by betulinic acid in
human endothelial cells
Katja Steinkamp-Fenske, Larissa Bollinger, Hui Xu, Ying Yao,
Sven Horke, Ulrich Förstermann and Huige Li
From the
Department of Pharmacology,
Johannes Gutenberg University, Mainz, Germany
JPET Fast Forward. Published on May 11, 2007 as DOI:10.1124/jpet.107.123356
Copyright 2007 by the American Society for Pharmacology and Experimental Therapeutics.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
2
Running title: Regulation of eNOS and NADPH oxidase by betulinic acid
Correspondence to: Huige Li, MD, PhD
Department of Pharmacology
Johannes Gutenberg University
Obere Zahlbacher Strasse 67
D-55131 Mainz
Germany
Telephone: + 49 (6131) 39-36929
Fax: + 49 (6131) 39-36611
E-mail: [email protected]
The manuscript contains
• 23 text pages;
• 8 figures;
• 0 Tables;
• 40 references;
• 224 words in Abstract;
• 486 words in Introduction and
• 1240 words in Discussion.
Abbreviations:
BH4, (6R)-5,6,7,8-tetrahydro-L-biopterin; HUVEC, human umbilical vein endothelial cells;
L-NAME, NG-nitro-L-arginine methyl ester; NO, nitric oxide, eNOS, endothelial NO
synthase; ZSS, Zizyphi Spinosi Semen
Section: Cardiovascular
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
3
ABSTRACT
Nitric oxide (NO) produced by endothelial NO synthase (eNOS) is a protective
principle in the vasculature. Many cardiovascular diseases are associated with reduced NO
bioactivity and eNOS uncoupling due to oxidative stress. Compounds that reverse eNOS
uncoupling and increase eNOS expression are of therapeutic interest. Zizyphi spinosi semen
(ZSS) is one of the most widely used traditional Chinese herbs with protective effects on the
cardiovascular system. In human umbilical vein endothelial cells (HUVEC) and HUVEC-
derived EA.hy 926 cells, an extract of ZSS increased eNOS promoter activity, eNOS mRNA
and protein expression, as well as NO production in a concentration- and time-dependent
manner. Major ZSS constituents include saponins such as jujuboside A and B and
pentacyclic triterpenes such as betulin and betulinic acid. Jujuboside A, jujuboside B, or
betulin had no significant effect on eNOS expression whereas betulinic acid increased eNOS
mRNA and protein expression in HUVEC and EA.hy 926 cells. Interestingly, betulinic acid
also attenuated the expression of NADPH oxidase subunits Nox4 and p22phox, thereby
reducing oxidative stress and improving eNOS function. Consequently, betulinic acid-treated
endothelial cells showed an increased production of bioactive NO (as indicated by a higher
efficacy in stimulating cGMP generation in RFL-6 reporter cells). Thus betulinic acid
possesses combined properties of eNOS upregulation and NADPH oxidase downregulation.
Compounds such as betulinic acid may have a therapeutic potential in cardiovascular disease.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
4
Introductions
Nitric oxide (NO) produced by endothelial NO synthase (eNOS) is an important
vasodilator and plays a protective physiological role in the vasculature (Li and Forstermann,
2000a). Blockade of NO synthesis with pharmacological NOS inhibitors causes significant
peripheral vasoconstriction and elevation of blood pressure. Mice with a disrupted eNOS
gene are hypertensive and lack endothelium-dependent, NO-mediated vasodilation (Huang et
al., 1995). Besides its role in controlling blood pressure, NO protects the blood vessels from
thrombosis by inhibiting platelet aggregation and adhesion. In addition, endothelial NO
possesses multiple anti-atherosclerotic properties (Li and Forstermann, 2000a). In agreement
with this concept, pharmacological inhibition of eNOS causes accelerated atherosclerosis in
rabbits (Cayatte et al., 1994) and in mice (Kauser et al., 2000), and deficiency in eNOS
accelerates the development of atherosclerosis in apolipoprotein E-knockout (apoE-KO) mice
(Kuhlencordt et al., 2001).
Due to the antithrombotic, anti-atherosclerotic and antihypertensive properties of
endothelial NO, the eNOS enzyme could be an interesting target for the prevention or therapy
of cardiovascular disease. NO synthesis can be modulated by eNOS activity and/or eNOS
gene expression (Li et al., 2002b; Li et al., 2002c). In vivo upregulation of eNOS gene
expression had been considered to be a reasonable strategy to prevent cardiovascular disease.
However, results from such experiments were disappointing. For example, eNOS-
overexpressing transgenic mice on an apoE-KO background (eNOS-TG/apoE-KO) developed
larger atherosclerotic lesions than apoE-KO mice (Ozaki et al., 2002). Also in animal models
of hypertension or diabetes, eNOS has been found upregulated, however, the upregulated
enzyme turned out to be dysfunctional (Hink et al., 2001; Mollnau et al., 2002). A
dysfunctional eNOS produces reactive oxygen species (ROS) at the expense of NO (Xia et al.,
1998). This has been referred to as eNOS “uncoupling” (Forstermann and Munzel, 2006).
The main reason for eNOS uncoupling is a deficiency of the essential eNOS cofactor (6R)-
5,6,7,8-tetrahydro-L-biopterin (BH4) (Channon, 2004). Oxidation of BH4 due to NADPH
oxidase-mediated vascular oxidative stress may represent a major cause of BH4 deficiency in
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
5
many cases (Forstermann and Munzel, 2006). Suppression of oxidative stress by
downregulating the expression or activity of vascular NADPH oxidase has been shown to
restore eNOS functionality (Hink et al., 2001; Mollnau et al., 2002).
Therefore, a correction of eNOS uncoupling combined with upregulation of eNOS
expression may be a promising therapeutic strategy for cardiovascular disease.
Zizyphi Spinosi Semen (ZSS, Suanzaoren in Chinese) is the dried seeds of Zizyphus
jujuba Mill. var. spinosa (Bunge). ZSS has been widely used since antiquity in Traditional
Chinese Medicine for the treatment of neurasthenia, anxiety and insomnia. However, modern
pharmacological studies revealed that this herb also has potent actions on the cardiovascular
system, such as anti-arrhythmic and blood pressure-lowering effects (Huang, 1999).
Here we report that betulinic acid from ZSS upregulates eNOS and, at the same time,
reduces the expression of NADPH oxidase in human endothelial cells. This results in
enhanced bioactive NO. Therefore, compounds like betulinic acid may have therapeutic
potential for cardiovascular diseases.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
6
Methods
Materials.
Aqueous extract of ZSS: 10g ZSS were crushed, incubated with 100 ml of water for
30 min and then boiled for another 30 min. The solution was collected using paper filter.
After adding 100 ml water, the solid phase was boiled again for 30 min, and the mixture
filtered with a paper filter. Filtrates from the both steps were pooled, mixed with an equal
volume of ethanol and placed in darkness at 4°C for 24h. The precipitate was removed by
centrifugation at 1000 g for 10 min. The supernatant was filtered using a 0.45 µm membrane.
The solution was concentrated using a rotary evaporator at 40°C to reduce the volume down
to 2 ml. This was considered a stock solution and used at different dilutions in the
experiments. A control stock solution was generated by evaporating a mixture of 100 ml
water and 100 ml ethanol down to 2 ml.
Jujuboside A (CAS number 55466-04-1) and B (CAS number 55466-05-2) were
purchased from ChromaDex (Santa Ana, CA, USA). Betulin [lup-20(29)-ene-3β,28-diol;
CAS number 473-98-3] and betulinic acid [3β-3-Hydroxy-lup-20(29)-en-28-oic acid, CAS
number 472-15-1] are from Sigma (Taufkirchen, Germany).
Cell culture.
Human umbilical vein endothelial cells (HUVEC) were isolated by collagenase
digestion as described (Li et al., 1998). HUVEC-derived EA.hy 926 endothelial cells were
kindly provided by Dr. Cora-Jean Edgell (Chapel Hill, NC). EA.hy 926 endothelial cells
were grown under 10% CO2 in Dulbecco’s modified Eagle’s medium (DMEM, Sigma)
supplemented with 10% fetal bovine serum, 2 mM L-glutamine, 1 mM sodium pyruvate, 100
U/ml penicillin, 100 µg/ml streptomycin, and 1x HAT (hypoxanthine, aminopterin and
thymidine) (Invitrogen, Karlsruhe, Germany) (Li and Forstermann, 2000b).
Analysis of eNOS promoter activity in stably transfected EA.hy 926 cells.
A stable EA.hy 926 cell line was generated by transfection of EA.hy 926 cells with
pGL3-eNOS-Hu-3500-neo, which contains a neomycin-resistance gene and a 3.5 kb promoter
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
7
fragment of human eNOS driving the luciferase reporter gene (Li et al., 1998). These EA.hy
926 cells were cultured in medium containing 1 mg/ml compound G418. For analysis of
eNOS promoter activity, the stably transfected cells were incubated with ZSS extract for 18
hours. Then, cells lysates were prepared and luciferase activities were determined as
described (Li et al., 1998). The luciferase activity, normalized for protein concentration of
cell lysates, was used as a determinant of eNOS promoter activity.
RNase protection assay for eNOS mRNA analyses.
EA.hy 926 cells were incubated with ZSS extract for 18 hours and total RNA was
isolated. The expression of eNOS mRNA was analyzed by RNase protection assay as
described previously (Li et al., 2002a; Li et al., 1998).
Real-Time RT-PCR for mRNA expression analyses.
In some experiments, eNOS mRNA expression was analyzed with quantitative Real-
Time RT-PCR using an iCyclerTM iQ System (Bio-Rad Laboratories, Munich, Germany). 500
ng of total RNA was used for Real-Time RT-PCR analysis with the QuantiTect™ Probe RT-
PCR kit (Qiagen, Hilden, Germany). Sequences of used eNOS primers were
GTGGCTGTCTGCATGGACCT (forward) and CCACGATGGTGACTTTGGCT (reverse).
The sequence of the dual-labeled TaqMan probe was
AGTGGAAATCAACGTGGCCGTGCTGC (Li et al., 2004). TaqMan Gene Expression
Assays (pre-designed probe and primer sets) were obtained from Applied Biosystems (Foster
City, CA, USA) for analyzing the mRNA expression of Nox4 and p22phox (assay ID
Hs00276431_m1 and Hs00609145_m1, respectively). mRNA expression levels of target
genes were normalized to TATA box binding protein mRNA (TBP, Applied Biosystems,
assay ID Hs00427620_m1).
Western blot for eNOS protein analyses.
Confluent EA.hy 926 cells were incubated with ZSS extract for 18 hours and total
protein was isolated. Western blotting was performed using 50 µg of protein and a
monoclonal anti-eNOS antibody (BD Biosciences Pharmingen, San Diego, CA), as described
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
8
previously (Li and Forstermann, 2000b). Immunocomplexes were developed using an
enhanced horseradish peroxidase/luminol chemiluminescence reagent (PerkinElmer Life
Sciences, Boston, MA) according to the manufacturer’s instructions.
Determination of NO synthesis using an NO Analyzer.
EA.hy 926 cells were treated with ZSS extract for 18 hours and the oxidation products
of NO, nitrite and nitrate, were assayed as a measure of NO synthesis in the supernatants.
After reduction of nitrate with nitrate reductase, total nitrite was determined by NO-ozone
chemiluminescence using a NOATM 280 NO Analyzer (Sievers, Boulder, CO). Total protein
content of the cells was determined (Bradford), and nitrite levels were normalized for protein
(Li et al., 2002a).
Reporter cell assay for determination of NO production.
EA.hy 926 cells were treated with 10 µM betulinic acid for 18 hours and NO
production bioassayed with RFL-6 rat lung fibroblasts as reporter cells (Li and Forstermann,
2000b). After the 18 hour-treatment, EA.hy cells and RFL-6 cells were washed twice with
Locke’s solution. Then, EA.hy 926 cells were incubated with Locke’s solution (154.0 mM
NaCl, 5.6 mM KCl, 2.0 mM CaCl2, 1 mM MgCl2, 3.6 mM NaHCO3, 5.6 mM glucose, 10.0
mM HEPES, pH 7.4).) containing 200 U/ml SOD, 100 µM L-arginine and 10µM betulinic
acid, and RFL-6 cells with Locke’s solution containing 600 µM IBMX for 30 min. After the
preincubation, EA.hy cells were incubated for 3 min at 37°C in Locke’s solution containing
200 U/ml SOD, 300 µM IBMX, 100 µM L-arginine, and 10µM betulinic acid. Then, the
conditioned media containing the NO released from the EA.hy cells were transferred onto the
RFL-6 cells which contain guanylyl cyclase, and another incubation of 3 min at 37°C was
performed. The reaction was stopped by aspiration of the solution, adding 1 ml of ice-cold 50
mM sodium acetate, pH 4.0, and rapidly freezing the cells with liquid nitrogen. The cGMP
content of the RFL-6 samples was determined by radioimmunoassay as described. The basal
cGMP content of the RFL-6 cells (receiving no conditioned media) was subtracted from all
samples (Li and Forstermann, 2000b).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
9
Measurement of ROS production by L-012 chemiluminescence.
Production of ROS was determined using the luminol derivative L-012 [8-amino-5-
chloro-7-phenylpyridol[3,4-d]pyridazine-1,4(2H,3H)dione] (Li et al., 2006). Briefly, EA.hy
926 cells were treated with 10 µM betulinic acid for 18 hours in 96-well plates and L-012-
derived chemiluminescence was measured using a Microplate Luminometer (Berthold) in
Hanks ́buffered salt solution (PAA Laboratories) containing 500 µM L-012. In additional
experiments, cells were pre-treated with 300 µM apocynin for 48h prior to betulinic acid
treatment. The photon counts were normalized for protein content of the cell lysates.
Statistics
Statistical differences between mean values were determined by analysis of variance
(ANOVA) followed by Fisher’s protected least-significant-difference test for comparison of
different means.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
10
Results
Increased eNOS promoter activity, eNOS expression and NO production in response to
a ZSS extract.
EA.hy 926 cells stably transfected with a 3.5 kb human eNOS promoter fragment were
treated with an aqueous extract of ZSS. This treatment resulted in a concentration-dependent
increase in eNOS promoter activity (Fig. 1A). The maximal increase was seen with a 1:100
dilution of the aqueous ZSS extract (5g raw material per ml as stock solution). Treatment
with the control solutions at any dilution did not have any effect on eNOS promoter activity.
For further experiments, control solutions at a 1:100 dilution was used as control.
Treatment of naïve EA.hy 926 cells with ZSS resulted in a concentration-dependent
increase in eNOS mRNA expression, as analyzed with RNase protection assay (Figs. 1B and
1C). Fig. 1D demonstrates the time-dependence of eNOS upregulation in response to
aqueous ZSS extract. A maximum effect was reached after 18 hours. Therefore, this time
frame was chosen for all other experiments. An 18-hour treatment of EA.hy 926 cells with
aqueous ZSS extract also increased eNOS protein (as analyzed with Western blot, Figs. 2A
and 2B).
Finally, ZSS treatment for 18 hours increased NO synthesis in EA.hy 926 cells in a
concentration-dependent manner, as determined by the NO-ozone chemiluminescence assay
(Fig. 2C).
Effects of ZSS constituents on eNOS expression in human endothelial cells.
Jujuboside A and B had no effect or on eNOS mRNA expression in human EA.hy 926
endothelial cells (Fig. 3). These compounds did not change eNOS promoter activity, either
(up to 30 µM and an 18 hour incubation, data not shown). Also betulin had no significant
effect on eNOS mRNA expression (Fig. 4A). In contrast, an 18 hour-treatment with betulinic
acid significantly increased eNOS mRNA expression in both EA.hy 926 cells (Fig. 4A) and in
HUVEC (Fig. 4B).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
11
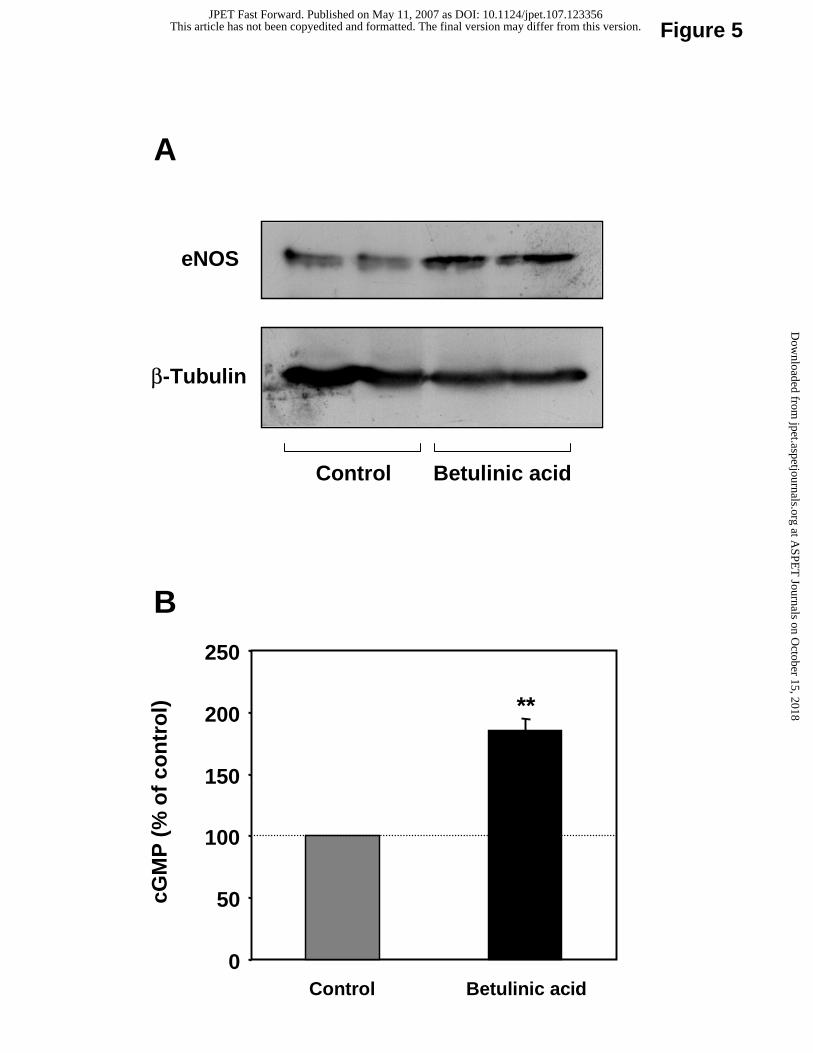
Increased eNOS protein expression and NO production in response to betulinic acid.
Treatment of EA.hy 926 cells with 10 µM betulinic acid for 18 hours resulted in a
significant induction of eNOS protein, as determined by Western blot (Fig. 5A). The
conditioned media from these cells showed higher potency in stimulating cGMP generation in
RFL-6 reporter cells, indicating an increased NO production by EA.hy 926 cells (Fig. 5B).
Downregulation of Nox4 and p22phox expression and reduced ROS production in
response to betulinic acid.
Treatment with 10 µM betulinic acid for 18 hours resulted in marked downregulation
of Nox4 and p22phox mRNA expression in EA.hy 926 cells (Fig. 6A) as well as in HUVEC
(Fig. 6B). This effect seems to be independent of the eNOS-upregulating effect of betulinic
acid, because it was not prevented by the NOS inhibitor NG-nitro-L-arginine methyl ester (L-
NAME, Fig. 7). In addition, treatment of EA.hy 926 cells with the NO donor spermine
NONOate did not reduce Nox4 expression (Fig. 7). The reduced expression of NADPH
oxidase subunits by betulinic acid was associated with a reduction of ROS production (Fig.
8). The effect of betulinic acid on ROS production was likely to be NADPH oxidase-
dependent, because it was absent in the presence of apocynin, a NADPH oxidase inhibitor
(Fig. 8).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
12
Discussion
ZSS is one of the most commonly used Chinese herbs. It is a sedative and hypnotic
drug with additional effect on the cardiovascular system (Huang, 1999).
ZSS protects cardiomyocytes from ischemic injury. Damage of cultured neonatal rat
myocardial cells by deprivation of oxygen and glucose was markedly reduced by ZSS total
saponins (Chen et al., 1990). Anoxia-reoxygenation of cultured neonatal rat myocardial cells
results in increased intracellular malondialdehyde and lipid peroxides, increased intercellular
calcium concentration, and decreased SOD activity. All of these parameters were reversed by
ZSS total saponins (Chen et al., 1990; Wan et al., 1997). In an animal model of endotoxin-
fever, the reduced SOD protein levels were prevented by treatment with ZSS (Peng et al.,
1995).
ZSS has also blood pressure-lowering effects. Intravenous injection of an aqueous
solution of ZSS extract markedly decreased blood pressure in anesthetized rats, dogs and cats
without any significant effect on the coronary blood flow, heart rates or myocardial
contractility (Gu et al., 1987). Oral treatment of spontaneously hypertensive rats with ZSS
jujubosides resulted in reduction of blood pressure (Zhang et al., 2003). Blood pressure
reduction was observed as early as 30 min, lasted at least 3.5 hours and the effect declined
after 7.5 hours (Zhang et al., 2003). Moreover, treatment of hypercholesterolemic rabbits
with ZSS for three months resulted in a reduction of total cholesterol, triglyceride and LDL,
an increase in HDL, and reduced atherosclerosis (Wu et al., 1989).
Molecular mechanisms underlying these cardiovascular effects are poorly understood.
Interestingly, treatment of rats with ZSS results in increased plasma levels of NO by unknown
mechanisms (Wang and Xie, 2004). Expression or activity of NOS isoforms was not analyzed
in that study.
The present study is the first that systematically investigates the effect of ZSS and its
constituents on the gene expression of eNOS. We found that ZSS increases eNOS promoter
activity (Fig. 1A), eNOS mRNA (Fig. 1B-D) and protein expression (Figs. 2A and 2B), as
well as NO production (Fig. 2C) in human endothelial cells.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
13
The active constituents of ZSS include saponins, triterpenoids, flavonoids, alkaloids
and fatty acids (Huang, 1999; Li et al., 2005b; Zhao et al., 2006). The most important ZSS
saponins are triterpenoid oligoglycosides such as jujubosides A and B (Bai et al., 2003;
Huang, 1999; Wang et al., 2005). The content of jujubosides A and B in ZSS is about 400
mg/kg (Wang et al., 2005). Triterpenoids found in ZSS include betulin, betulinic acid (Bai et
al., 2003; Li et al., 2005b).
Jujuboside A and B showed no effect on eNOS promoter activity or eNOS mRNA
expression (Fig. 3). Also betulin had no effect on eNOS expression (Fig. 4A). Interestingly,
treatment of human endothelial cells with betulinic acid resulted in a significant upregulation
of eNOS mRNA (Fig. 4) and protein (Fig. 5A) expression. The contents of betulinic acid and
betulin in ZSS are about 7 mg/kg and 3 mg/kg, respectively (Li et al., 2005b). In the present
study, we used ZSS extract at a concentration of 5g ZSS per ml. When cells were treated with
this extract at a 1:100 dilution, the estimated final concentration of betulinic acid is in the low
micromolar range. As shown in Fig. 4B, betulinic acid increased eNOS mRNA expression
even at 1 µM. Thus, betulinic acid is likely to be one of the compounds responsible for the
eNOS upregulation produced by ZSS.
Upregulation of eNOS does not necessarily result in an increase in bioactive NO.
Under pathological conditions of oxidative stress, eNOS is often uncoupled and dysfunctional
(Forstermann and Munzel, 2006). In the vasculature, ROS can be produced by several
enzyme systems, including enzymes of the respiratory chain, cytochrome P450
monoxygenases, uncoupled eNOS, xanthine oxidase, and NADPH oxidases. Although all of
these enzymes contribute to the oxidative burden, evidence is accumulating that an initial
generation of ROS by NADPH oxidases triggers the release of ROS by the other enzymes
(Griendling, 2004). Moreover, NADPH oxidase-mediated oxidative stress is involved in the
development and progression of atherosclerosis (Lassegue and Clempus, 2003). In diseased
human coronary arteries, about 60% of vascular superoxide is derived from NADPH oxidases
(Guzik et al., 2006).
The NADPH oxidase enzyme complex consists of 2 essential membrane-bound
catalytic subunits, gp91phox/Nox2 and p22phox, which compose flavocytochrome b558, and
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
14
several cytosolic regulatory components (Lassegue and Clempus, 2003). In the vasculature,
the catalytic subunit gp91phox/Nox2 is a member of a family of homologous proteins termed
Nox. In addition to gp91phox/Nox2, Nox1, and Nox4 are also expressed in blood vessels
(Griendling, 2004; Lassegue and Clempus, 2003). Both Nox2 and Nox4 are found in
endothelial cells (Griendling, 2004). Nox4 is likely the major Nox isoform and the
predominant ROS source in endothelial cells (Ago et al., 2004; Hu et al., 2005).
Interestingly, betulinic acid significantly reduced the expression of NADPH oxidase
catalytic subunits Nox4 and p22phox (Fig. 6), which was associated with a reduction in ROS
production (Fig. 8).
Therefore, betulinic acid possesses a dual protective action on the vasculature: (i) it
upregulates eNOS expression, thereby producing more eNOS protein, and (ii) it
downregulates NADPH oxidase, thereby maintaining the upregulated eNOS enzyme in a
functional state.
In the past, we have found analogous combined effects of some other compounds. The
PKC inhibitor midostaurin, for example, upregulates eNOS expression by a PKC-independent
mechanism (Li and Forstermann, 2000b), and reduces vascular Nox1 expression by PKC
inhibition (Li et al., 2006). By reducing NADPH oxidase-mediated oxidative stress,
midostaurin reverses eNOS uncoupling in spontaneously hypertensive rats and in
atherosclerosis-prone apoE-KO mice, which was associated with NO-mediated vasodilation
and blood pressure reduction (Li et al., 2005a; Li et al., 2006). Also red wine and its
constituents (such as resveratrol, cinnamic acid and hydroxycinnamic acid) upregulate eNOS
while preserving eNOS functionality (Wallerath et al., 2002; Wallerath et al., 2005; Wallerath
et al., 2003). Direct antioxidant properties of these compounds as well as the demonstrated
downregulation of Nox4 by resveratrol (Li et al. unpublished data) may contribute to this
effect.
Thus, the combination of eNOS upregulation and NADPH oxidase downregulation
may result in enhanced levels of bioactive NO and thus vascular protection. Indeed, treatment
of endothelial cells with betulinic acid increased bioactive NO (that stimulated cGMP
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
15
generation in RFL-6 reporter cells; Fig. 5B), indicating that the upregulated eNOS by
betulinic acid is maintained under a functional state.
The current study has certain limitations. (i) The measurements of NO were indirect
as nitrite/nitrate or stimulation of soluble guanylyl cyclase. At least the latter indicates,
however, that bioactive NO was increased by betulinic acid. (ii) The main effects are shown
on healthy cells that are not stressed. Further experiments are needed to find out whether
betulinic acid can reverse eNOS uncoupling under conditions of oxidative stress. The
suppression of NADPH oxidase expression by betulinic acid (Figs. 6A and B) would favor
such an action. (iii) Betulinic acid has been shown to have pro-apoptotic effects in cancer
cells (Rzeski et al., 2006). We have confirmed this effect in the alveolar epithelial carcinoma
cell line A549/8 (data not shown). In HUVEC, however, betulinic acid did not induce
apoptosis. At concentrations of 1 and 10 µM, betulinic acid even reduced caspase 3/7 activity
in HUVEC (data not shown).
In conclusion, the present study demonstrates that betulinic acid from the Chinese herb
ZSS upregulates eNOS expression and downregulates NADPH oxidase expression in human
endothelial cells. This results in an increased amount of functional eNOS protein, enhanced
production of bioactive NO and thus vasoprotection (even in disease conditions with
increased oxidative stress).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
16
Acknowledgments
We thank Dr. Andrew Marston, Laboratoire de Pharmacognosie et Phytochimie, Ecole
de Pharmacie Genève-Lausanne, Université de Genève, Switzerland, for methodological
suggestions and helpful discussions. The manuscript contains parts of the doctoral theses of
K.S.-F. and of L.B.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
17
References
Ago T, Kitazono T, Ooboshi H, Iyama T, Han YH, Takada J, Wakisaka M, Ibayashi S,
Utsumi H and Iida M (2004) Nox4 as the major catalytic component of an endothelial
NAD(P)H oxidase. Circulation 109:227-233.
Bai YJ, Cheng G, Tao J, Wang B, Zhao YY, Liu Y, Ma LB and Tu GZ (2003) [Structure
identification of jujuboside E]. Yao Xue Xue Bao 38:934-937.
Cayatte AJ, Palacino JJ, Horten K and Cohen RA (1994) Chronic inhibition of nitric oxide
production accelerates neointima formation and impairs endothelial function in
hypercholesterolemic rabbits. Arterioscler Thromb 14:753-759.
Channon KM (2004) Tetrahydrobiopterin: regulator of endothelial nitric oxide synthase in
vascular disease. Trends Cardiovasc Med 14:323-327.
Chen XJ, Yu CL and Liu JF (1990) [Protective effects of total saponins of semen Ziziphi
spinosae on cultured rat myocardial cells]. Zhongguo Yao Li Xue Bao 11:153-155.
Forstermann U and Munzel T (2006) Endothelial nitric oxide synthase in vascular disease:
from marvel to menace. Circulation 113:1708-1714.
Griendling KK (2004) Novel NAD(P)H oxidases in the cardiovascular system. Heart 90:491-
493.
Gu WX, Liu JF, Zhang JX, Liu XM, Liu JS and Chen YR (1987) [Blood pressure-lowering
effect and mechanism of total saponins from Ziziphus Mill seeds]. Di Yi Jun Yi Da
Xue Xue Bao 7:8-10.
Guzik TJ, Sadowski J, Guzik B, Jopek A, Kapelak B, Przybylowski P, Wierzbicki K, Korbut
R, Harrison DG and Channon KM (2006) Coronary artery superoxide production and
nox isoform expression in human coronary artery disease. Arterioscler Thromb Vasc
Biol 26:333-339.
Hink U, Li H, Mollnau H, Oelze M, Matheis E, Hartmann M, Skatchkov M, Thaiss F, Stahl
RA, Warnholtz A, Meinertz T, Griendling K, Harrison DG, Forstermann U and
Munzel T (2001) Mechanisms underlying endothelial dysfunction in diabetes mellitus.
Circ Res 88:E14-E22.
Hu T, Ramachandrarao SP, Siva S, Valancius C, Zhu Y, Mahadev K, Toh I, Goldstein BJ,
Woolkalis M and Sharma K (2005) Reactive oxygen species production via NADPH
oxidase mediates TGF-beta-induced cytoskeletal alterations in endothelial cells. Am J
Physiol Renal Physiol 289:F816-F825.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
18
Huang KC (1999) The pharmacology of Chinese herbs. CRC Press, Boca Raton.
Huang PL, Huang Z, Mashimo H, Bloch KD, Moskowitz MA, Bevan JA and Fishman MC
(1995) Hypertension in mice lacking the gene for endothelial nitric oxide synthase.
Nature 377:239-242.
Kauser K, da Cunha V, Fitch R, Mallari C and Rubanyi GM (2000) Role of endogenous nitric
oxide in progression of atherosclerosis in apolipoprotein E-deficient mice. Am J
Physiol Heart Circ Physiol 278:H1679-H1685.
Kuhlencordt PJ, Gyurko R, Han F, Scherrer-Crosbie M, Aretz TH, Hajjar R, Picard MH and
Huang PL (2001) Accelerated atherosclerosis, aortic aneurysm formation, and
ischemic heart disease in apolipoprotein E/endothelial nitric oxide synthase double-
knockout mice. Circulation 104:448-454.
Lassegue B and Clempus RE (2003) Vascular NAD(P)H oxidases: specific features,
expression, and regulation. Am J Physiol Regul Integr Comp Physiol 285:R277-297.
Li H and Forstermann U (2000a) Nitric oxide in the pathogenesis of vascular disease. J
Pathol 190:244-254.
Li H and Forstermann U (2000b) Structure-activity relationship of staurosporine analogs in
regulating expression of endothelial nitric-oxide synthase gene. Mol Pharmacol
57:427-435.
Li H, Hergert SM, Schafer SC, Brausch I, Yao Y, Huang Q, Mang C, Lehr HA and
Forstermann U (2005a) Midostaurin upregulates eNOS gene expression and preserves
eNOS function in the microcirculation of the mouse. Nitric Oxide 12:231-236.
Li H, Junk P, Huwiler A, Burkhardt C, Wallerath T, Pfeilschifter J and Forstermann U
(2002a) Dual effect of ceramide on human endothelial cells: induction of oxidative
stress and transcriptional upregulation of endothelial nitric oxide synthase. Circulation
106:2250-2256.
Li H, Oehrlein SA, Wallerath T, Ihrig-Biedert I, Wohlfart P, Ulshöfer T, Jessen T, Herget T,
Forstermann U and Kleinert H (1998) Activation of protein kinase C alpha and/or
epsilon enhances transcription of the human endothelial nitric oxide synthase gene.
Mol Pharmacol 53:630-637.
Li H, Wallerath T and Forstermann U (2002b) Physiological mechanisms regulating the
expression of endothelial-type NO synthase. Nitric Oxide 7:132-147.
Li H, Wallerath T, Munzel T and Forstermann U (2002c) Regulation of endothelial-type NO
synthase expression in pathophysiology and in response to drugs. Nitric Oxide 7:149-
164.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
19
Li H, Witte K, August M, Brausch I, Godtel-Armbrust U, Habermeier A, Closs EI, Oelze M,
Munzel T and Forstermann U (2006) Reversal of endothelial nitric oxide synthase
uncoupling and up-regulation of endothelial nitric oxide synthase expression lowers
blood pressure in hypertensive rats. J Am Coll Cardiol 47:2536-2544.
Li H, Xia N, Brausch I, Yao Y and Forstermann U (2004) Flavonoids from artichoke (Cynara
scolymus L.) up-regulate endothelial-type nitric-oxide synthase gene expression in
human endothelial cells. J Pharmacol Exp Ther 310:926-932.
Li L-M, Liao X, Peng S-L and Ding L-S (2005b) Chemical constituents from the seeds of
Ziziphus jujuba var. spinosa (Bunge) Hu. Journal of Integrative Plant Biology 47:494-
498.
Mollnau H, Wendt M, Szocs K, Lassegue B, Schulz E, Oelze M, Li H, Bodenschatz M,
August M, Kleschyov AL, Tsilimingas N, Walter U, Forstermann U, Meinertz T,
Griendling K and Munzel T (2002) Effects of angiotensin II infusion on the
expression and function of NAD(P)H oxidase and components of nitric oxide/cGMP
signaling. Circ Res 90:e58-e65.
Ozaki M, Kawashima S, Yamashita T, Hirase T, Namiki M, Inoue N, Hirata K, Yasui H,
Sakurai H, Yoshida Y, Masada M and Yokoyama M (2002) Overexpression of
endothelial nitric oxide synthase accelerates atherosclerotic lesion formation in apoE-
deficient mice. J Clin Invest 110:331-340.
Peng Z, Zhang H, Cheng S and Guo W (1995) [Protective effect of semen Ziziphi spinosae on
superoxide dismutase reduction in mice with endotoxin fever]. Zhongguo Zhong Yao
Za Zhi 20:369-370.
Rzeski W, Stepulak A, Szymanski M, Sifringer M, Kaczor J, Wejksza K, Zdzisinska B and
Kandefer-Szerszen M (2006) Betulinic acid decreases expression of bcl-2 and cyclin
D1, inhibits proliferation, migration and induces apoptosis in cancer cells. Naunyn
Schmiedebergs Arch Pharmacol 374:11-20.
Wallerath T, Deckert G, Ternes T, Anderson H, Li H, Witte K and Forstermann U (2002)
Resveratrol, a polyphenolic phytoalexin present in red wine, enhances expression and
activity of endothelial nitric oxide synthase. Circulation 106:1652-1658.
Wallerath T, Li H, Godtel-Ambrust U, Schwarz PM and Forstermann U (2005) A blend of
polyphenolic compounds explains the stimulatory effect of red wine on human
endothelial NO synthase. Nitric Oxide 12:97-104.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
20
Wallerath T, Poleo D, Li H and Forstermann U (2003) Red wine increases the expression of
human endothelial nitric oxide synthase: a mechanism that may contribute to its
beneficial cardiovascular effects. J Am Coll Cardiol 41:471-478.
Wan HY, Ding L, Kong XP, Liu SJ and Chen XJ (1997) [Effect of total saponins of semen
Ziziphi spinosae on hypoxia-reoxygenation injury in myocardial cells]. Zhongguo
Bingli Shengli Za Zhi 13:522-526.
Wang X and Xie M (2004) [Effects of suanzaoren decoction on serum NO and the Level of
cytokines in rats]. Beijing Zhong Yi Yao Da Xue Xue Bao 11:16-18.
Wang XL, Zhang YJ, Chen MX and Wang ZF (2005) [Optimization study on extraction
technology of the seed of Ziziphus jujuba var. spinosa by orthogonal design with
multi-targets]. Zhongguo Zhong Yao Za Zhi 30:753-755.
Wu SX, Lang XC, Jia BY, Zhao SX, Li MX and Lan MY (1989) [Effects of Ziziphus spinosa
Hu on serum lipoprotein and experimental atherosclerosis]. Zhongguo Zhong Yao Za
Zhi 14:434-451, 448.
Xia Y, Tsai AL, Berka V and Zweier JL (1998) Superoxide generation from endothelial
nitric-oxide synthase. A Ca2+/calmodulin-dependent and tetrahydrobiopterin
regulatory process. J Biol Chem 273:25804-25808.
Zhang D, Yuang BX and Sun H (2003) [The effect of jujuboside on rats with spontaneous
hypertension]. Xi'an Jiao Tong Da Xue Xue Bao 24:59-60.
Zhao J, Li SP, Yang FQ, Li P and Wang YT (2006) Simultaneous determination of saponins
and fatty acids in Ziziphus jujuba (Suanzaoren) by high performance liquid
chromatography-evaporative light scattering detection and pressurized liquid
extraction. J Chromatogr A 1108:188-194.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
21
Footnotes
This work was supported by the Collaborative Research Center SFB 553 (project A1 to H.L.
and U.F.) from the DFG (Deutsche Forschungsgemeinschaft), Bonn, Germany.
Address correspondence to: Dr. Huige Li, Department of Pharmacology, Johannes Gutenberg
University, Obere Zahlbacher Strasse 67, D-55131 Mainz, Germany. E-mail:
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
22
Legends for Figures
Fig. 1. Extract of Zizyphi Spinosi Semen (ZSS) enhances eNOS promoter activity
and eNOS mRNA expression. Panel A: Human EA.hy 926 endothelial cells were stably
transfected with a 3.5-kb human eNOS promoter fragment driving a luciferase reporter gene.
Cells were treated with the ZSS extract at the indicated dilutions (stock solution
corresponding to 5g raw ZSS per ml). Luciferase activity was analyzed as a determinant of
eNOS promoter activity. Panels B-D: Human EA.hy 926 endothelial cells were treated for 18
hours with ZSS extract and eNOS mRNA expression analyzed with RNase protection assays.
Panel B demonstrates an original gel (performed in triplicate). The gel shows the protected
bands for eNOS and for β-actin (used for normalization). Panels C and D demonstrate the
concentration- and time-dependency of ZSS’s effect on eNOS mRNA expression,
respectively. Symbols represent mean ± SEM, n = 9-15 (*p< 0.05; **p<0.01, ***p<0.001,
compared with control).
Fig. 2. ZSS extract increases eNOS protein expression and NO production. EA.hy
926 cells were treated for 18 hours with ZSS extract and eNOS protein expression analyzed
with Western blot using a monoclonal anti-eNOS antibody. Panel A shows a representative
blot of three independent experiments. β-tubulin was used for normalization. Panel B
illustrates results of densitometric analyses. Panel C: Oxidation products of NO, nitrite and
nitrate, in the cell culture supernatant were assayed by NO-ozone chemiluminescence using a
NO Analyzer. Symbols represent mean ± SEM, n = 9-18 (*p< 0.05, **p< 0.01, compared
with control).
Fig. 3. The saponins jujuboside A and B have no effect on eNOS expression.
Human EA.hy 926 cells were treated with jujuboside A or B for 18 hours (1-10 µM) and
eNOS mRNA expression was analyzed with quantitative Real-Time RT-PCR. Columns
represent mean ± SEM, n = 9-12.
Fig. 4. The triterpenoid betulinic acid increases eNOS mRNA expression. Human
EA.hy 926 cells (panel A) or HUVEC (panel B) were treated with betulin or betulinic acid (1-
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

JPET #123356
23
10 µM) for 18 hours and eNOS mRNA expression was analyzed with quantitative Real-Time
RT-PCR. Columns represent mean ± SEM, n = 9-15 (*p< 0.05; **p<0.01, compared with
control).
Fig. 5. Betulinic acid enhances eNOS protein expression and NO production.
EA.hy 926 cells were treated for 18 hours with betulinic acid (10 µM) and eNOS protein
expression was analyzed with Western blot using a monoclonal anti-eNOS antibody. Panel A
shows a representative blot of three independent experiments. β-tubulin was used for
normalization. Panel B: Human EA.hy 926 cells were treated with betulinic acid (10 µM) for
18 hours. Bioactive NO from EA.hy 926 cells was bioassayed using guanylyl cyclase-
containing RFL-6 reporter cells. cGMP content in RFL-6 cells was determined with
radioimmunoassay. Columns represent mean ± SEM, n = 8-18 (**p< 0.01 compared with
control).
Fig. 6. Betulinic acid downregulates the mRNA expression of Nox4 and p22phox.
Human EA.hy 926 cells (panel A) or HUVEC (panel B) were treated with 10 µM betulinic
acid (BA) for 18 hours. Co: control. mRNA expression of NADPH oxidase catalytic subunits
Nox4 and p22phox was analyzed with quantitative Real-Time RT-PCR. Columns represent
mean ± SEM, n = 9-15 (**p< 0.01).
Fig. 7. Nox4 downregulation by betulinic acid is NO-independent. Human EA.hy
926 cells were treated with betulinic acid (BA, 10 µM), the NOS inhibitor L-NAME (1 mM)
or the NO donor spermine NONOate (NONOate, 10 µM) for 18 hours. Co: control. mRNA
expression of Nox4 was analyzed with quantitative Real-Time RT-PCR. Columns represent
mean ± SEM, n = 9-12 (**p< 0.01, compared with control).
Fig. 8. Betulinic acid reduces ROS production in EA.hy 926 endothelial cells.
Human EA.hy 926 cells, with or without apocynin pre-treatment (300 µM, 48 hours), were
treated with 10 µM betulinic acid (BA) for 18 hours. Co: control. ROS production was
measured with L-012-derived chemiluminescence. Columns represent mean ± SEM, n = 9-12
(**p < 0.01).
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

A
D
Figure 1
eNO
S m
RN
A(%
)
ZSS 18 hours
C
100
140
180
220
Control 1:1000 1:300 1:100
**
*
ZSS 1:100 (hours)
0 6 12 18 24
100
150
200
**
** **
100
150
200
250
300
1:1001:3001:1000
eNO
S p
rom
ote
r ac
tivi
ty(%
)
ZSS***
*
Vehicle
Control
B
eNOS
β-actin
Control 1:1000 1:300 1:100
ZSS 18 hoursZSS 18 hours
eNO
S m
RN
A(%
)
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

eNOS
β-Tubulin
ControlZSS
1:1000
100
200
300
400
500
eNO
Sp
rote
in(%
)
Control ZSS1:100
**
Figure 2
0
100
200
300
400
500
Control 1:1000 1:300 1:100
ZSS
Nit
rite
/Nit
rate
(%
)
*
**
A B
C
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from

0
50
100
150
Jujuboside A (µM) Jujuboside B (µM)
eNO
Sm
RN
A (
%)
Figure 3
0 1 3 10 0 1 3 10
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from
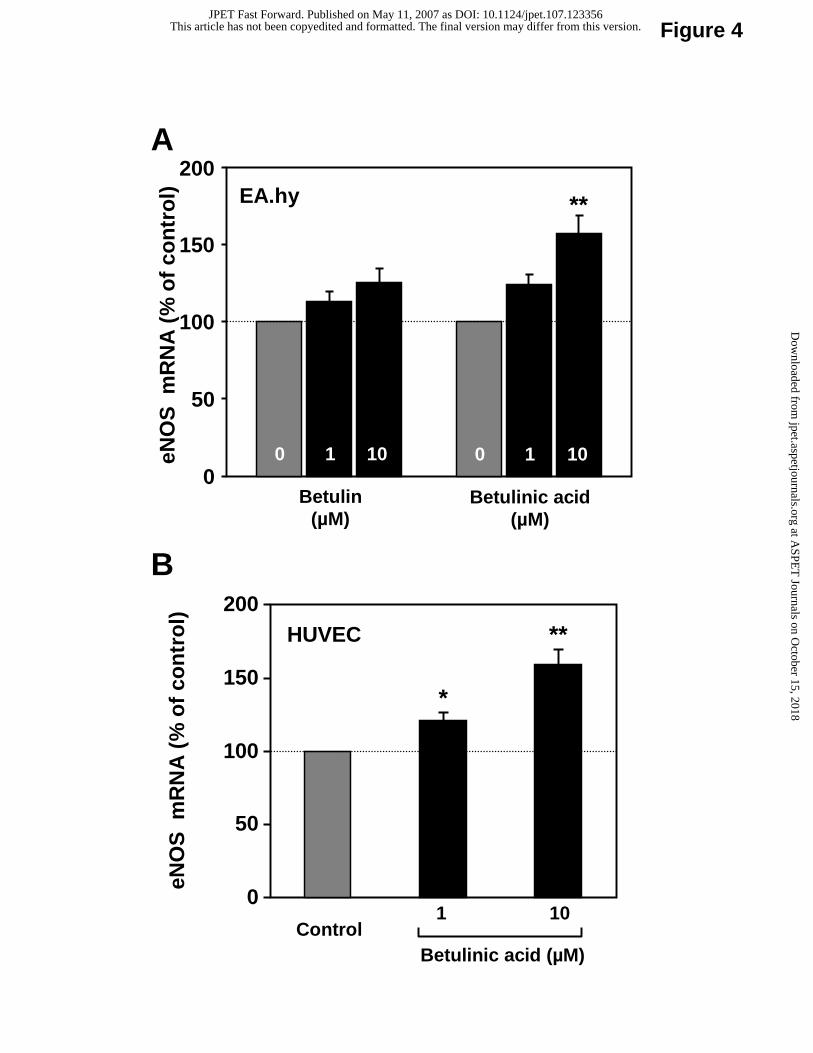
Figure 4
HUVEC
0
50
100
150
200
eNO
Sm
RN
A (
% o
f co
ntr
ol)
ControlBetulinic acid (µM)
1 10
EA.hy
0
50
100
150
eNO
Sm
RN
A (
% o
f co
ntr
ol)
200
Betulin(µM)
10
**
**
*
10
Betulinic acid(µM)
1010
A
B
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from
0
50
100
150
200
250
cGM
P(%
of
con
tro
l)
Control Betulinic acid
**
A
B
Betulinic acidControl
eNOS
β-Tubulin
Figure 5This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from
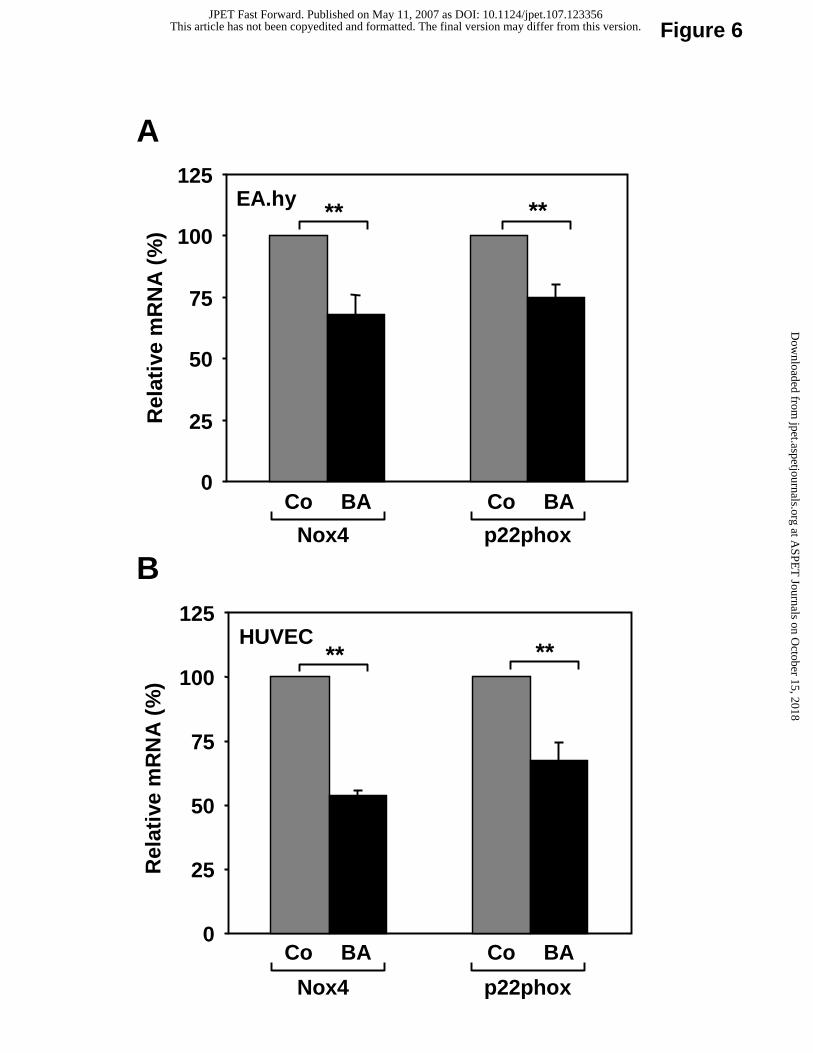
Figure 6
HUVEC
0
25
50
75
100
125
Rel
ativ
e m
RN
A(%
)
Nox4 p22phox
Co BA Co BA
** **
Nox4 p22phox
EA.hy
0
25
50
75
100
125R
elat
ive
mR
NA
(%)
Co BA Co BA
****
A
B
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from
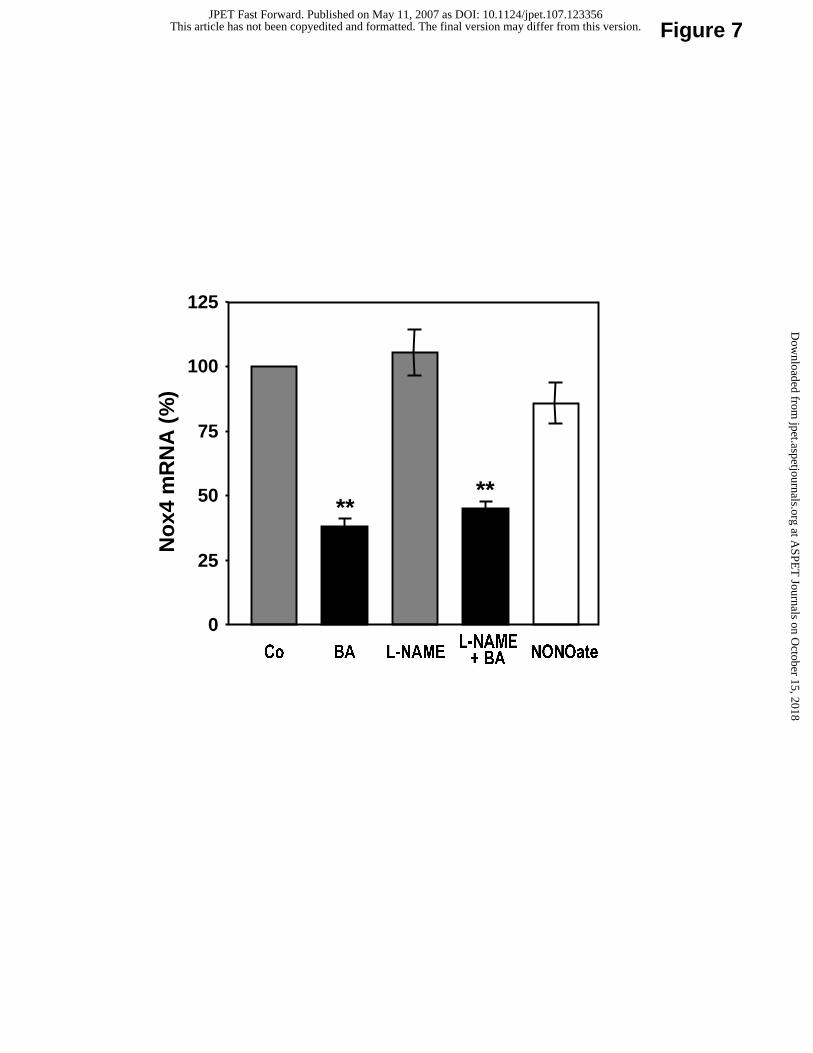
Figure 7
0
25
50
75
100
125
Co BA L-NAME NONOate
No
x4 m
RN
A (
%)
L-NAME+ BA
****
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from
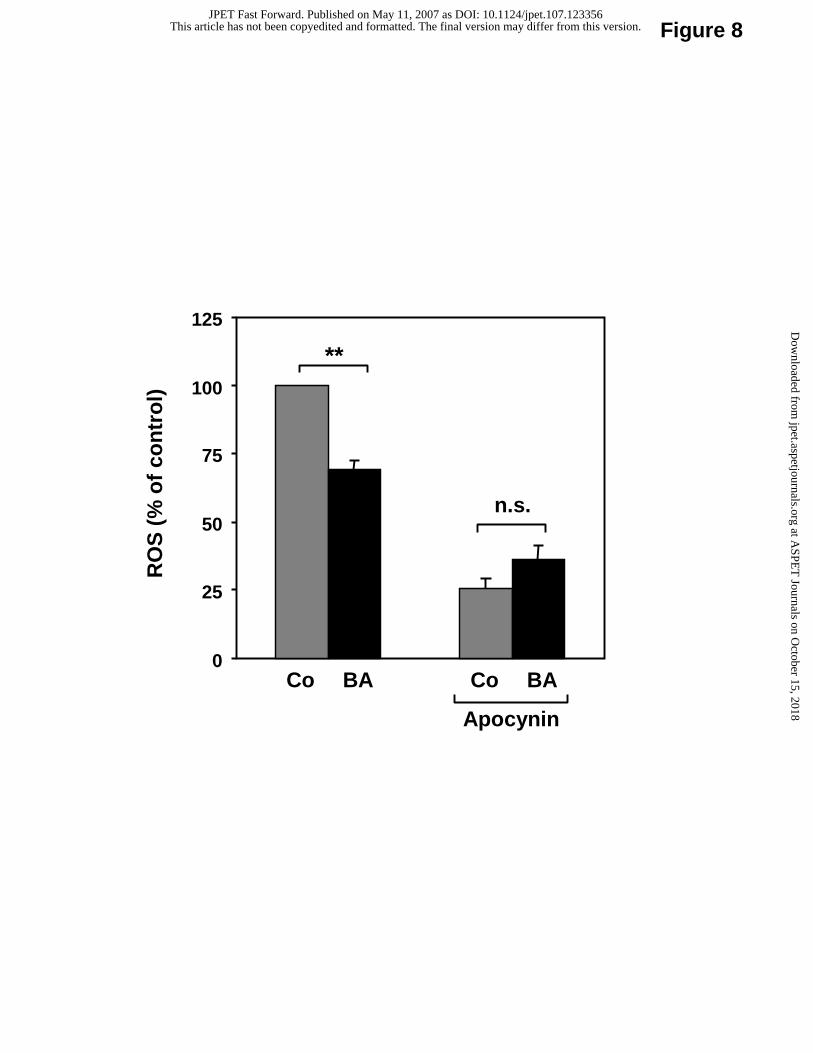
Figure 8
0
25
50
75
100
125
RO
S (
% o
f co
ntr
ol)
Apocynin
Co BACo BA
**
n.s.
This article has not been copyedited and formatted. The final version may differ from this version.JPET Fast Forward. Published on May 11, 2007 as DOI: 10.1124/jpet.107.123356
at ASPE
T Journals on O
ctober 15, 2018jpet.aspetjournals.org
Dow
nloaded from





























