Embed Size (px)
Citation preview

THE PRODUCTION OF CHAINS BY DIPLOCOCCUS PNEUMONIAE INMAGNESIUM DEFICIENT MEDIA
EMERIC J. ROCHFORD AND R. J. MANDLEDepartment of Bacteriology, The Jefferson Medical College of Philadelphia, Philadelphia, Pennsylvania
Received for publication May 27, 1953
The classical morphological appearance ofDiplococcus pneumonute in artificial culturemedia is that of pairs of cocci. Short chains con-sting of 2 to 4 pairs of organisms, aranged
linearly, are seen in young cultures. The lengthmay vary with the serological type. Chains arerelatively uncommon with Type I but are en-countered frequently with Type III. It is notknown if chaining in this genus is an expressionof incomplete cell division or is related to thecharacteristics of the capsular integument.
It was observed in this laboratory that theaddition of sodium citrate to a mediumotherwisesuited for the growth of the pneumococcus re-sulted in the organisms being found in chainsrather than as isolated diplococci. Okamoto (1940)using quatemary ammonium compounds andBadger (1944) using ethanolamine observedlike rsults.
Similar effects upon other organisms have beenrecorded. Filamentous forms of Cloidum per-fringens were produced by Webb (1948) by grow-ing the organisms in a medium deficient in Mg++.Subculture of these organisms in a Mg++ richmedium caused a reversion to the normal form.This reversion could not be accomplished by thesubstitution of metallic ions other than Mg++.Studies on the morphology of organisms grownin a Mg++ deficient medium were extended byWebb (1949) to include other groups of organ-isms such as clostridia and gram positive aerobicbacilli. Shankar and Bard (1952) investigatedC. perfringenm with respect to the production offilamentous cells in Mg++ deficient media andfound that Ca++ played a role in the productionof filaments even at optimal levels of Mg++and other ions.The relation of trace element concentration to
the change in morphology of D. pneumoniae wasinvestigated to ascertain which ions were in-volved in this alteration and to relate morphol-ogy and growth to metallic ion concentration.
MATERIAL AND METHODS
Diplococus pneumoniae, Types I, II, and arough strain of Type II (D39R), were used inthis study. The strains were maintained by serialtransfer in brain heart infusion broth, the cul-tures being checked at frequent intervals fortheir identity and dissociation status. The inoculawere prepared from 18 to 24 hour cultures inbrain heart infusion broth, grown at 37 C, washedtwice, and resuspended in one-half of the originalvolume of sterile 0.067 m phosphate buffer, pH7.4. Of this suspension, 0.15 ml was used asinoculum for each 5 ml t medium.
All glassware was washed thoroughly with dis-tilled water following the usual chemical treat-ment with chromic acid, NaOH, and dilute HCLTwo methods were employed routinely in
this study for removal of the ions:(1) The medium was treated with salts which
had the property of complexing or precipitatingthe positive divalent ions. These salts were:Sodium citrate, sodium potassium tartrate,potassium oxalate, sodium hexametaphosphate("calgon"), and the sodium salt of ethylene di-amine tetraacetic acid ("versene")).1
(2) A cationic exchange column was used. Acolumn, 24 inches long by 1 inch in diameter,was packed firmly with the resin material,"amberlite IR-120".2 A volume of 200 ml ofthe medium (made up in a concentration 2.5times that normaly used) was passed throughthe column. The first effluent, 50 ml, was dis-carded. The remainder was collected and dis-pensed in chemically clean test tubes in 2 mlportions. The tubes containing the medium werebrought to a final volume of 5 ml with the addi-tion of distilled water or solutions of the salts
I Supplied through the courtesy of the Bers-worth Chemical Company, Framingham, Massa-chusetts.
'Supplied generously by the Rohm and HaasCompany, Philadelphia, Pennsylvania.
554
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
PRODUCTION OF CHAINS BY D. PNEUMONIAE
of Mg++, Ca++, Mn++, and Fe++ in the concen-trations desired.The resin treated medium and the distilled
water were tested for the presence of free Ca++and Mg++ ions by the method of Buckley et al.(1951). This method is sensitive to free Ca++and Mg++ in amounts as small as one ug perml. Within the limits of the method, no detec-table amounts of either ion could be determinedin the distilled water or the resin treated medium.
organic ions in the usual medium were varied inorder to ascertain the optimum levels necessaryfor the production of chains. This was accom-plished by adding various complexing agents tothe medium or by removing the metallic ionsfrom the medium by passing it through a cationicexchange column. Various metallic ions wereresupplied in turn to the resin treated mediumand the effect upon the growth and chain forma-tion of D. pneumoniae observed.
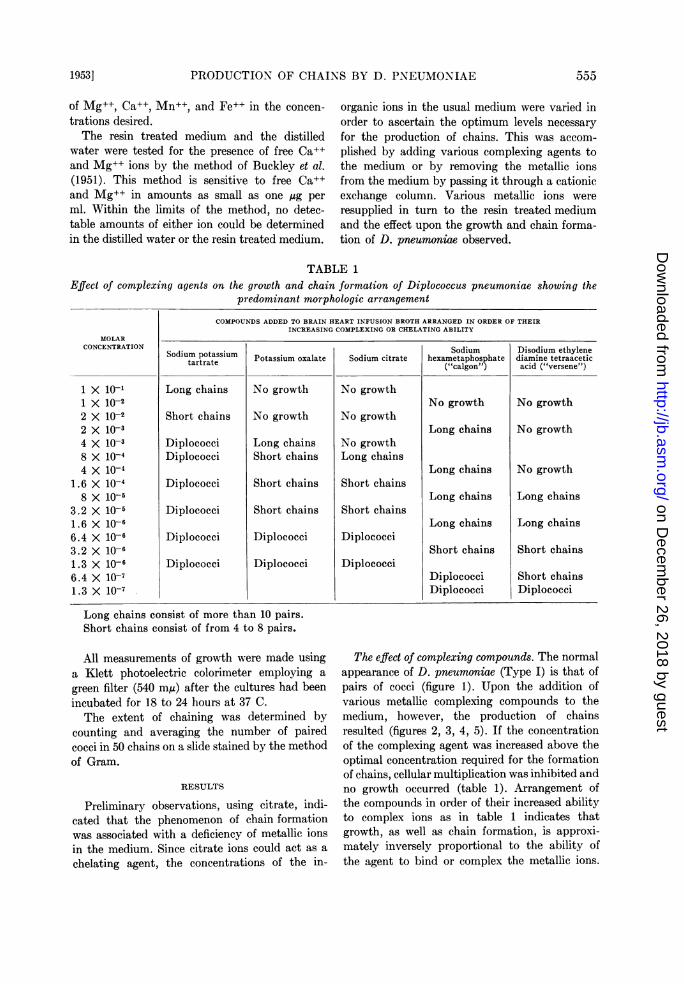
TABLE 1Effect of complexing agents on the growth and chain formation of Diplococcus pneumoniae showing the
predominant morphologic arrangementCOMPOUNDS ADDED TO BRAIN HEART INFUSION BROTH ARRANGED IN ORDER OF THEIR
INCREASING COMPLEXING OR CHELATING ABILITYMOLAR
CONCENTRATION Sodium Disodium ethyleneSodium potassium Potassium oxalate Sodium citrate hexametaphosphate diamine tetraacetictartrate ("calgon") acid ("versene")
1 X 10-1 Long chains No growth No growth1 X 10-I No growth No growth2 X 10-2 Short chains No growth No growth2 X 10-3 Long chains No growth4 X 10-3 Diplococci Long chains No growth8 X 10-4 Diplococci Short chains Long chains4 X 10-4 Long chains No growth
1.6 X 10-4 Diplococci Short chains Short chains8 X 10-5 Long chains Long chains
3.2 X 10-5 Diplococci Short chains Short chains1.6 X 10-6 Long chains Long chains6.4 X 10-6 Diplococci Diplococci Diplococci3.2 X 10-6 Short chains Short chains1.3 X 10-6 Diplococci Diplococci Diplococci6.4 X 10-7 Diplococci Short chains1.3 X 10-7 Diplococci Diplococci
Long chains consist of more than 10 pairs.Short chains consist of from 4 to 8 pairs.
All measurements of growth were made usinga Klett photoelectric colorimeter employing agreen filter (540 m,u) after the cultures had beenincubated for 18 to 24 hours at 37 C.The extent of chaining was determined by
counting and averaging the number of pairedcocci in 50 chains on a slide stained by the methodof Gram.
RESULTS
Preliminary observations, using citrate, indi-cated that the phenomenon of chain formationwas associated with a deficiency of metallic ionsin the medium. Since citrate ions could act as a
chelating agent, the concentrations of the in-
The effect of complexing compounds. The normalappearance of D. pneumoniae (Type I) is that ofpairs of cocci (figure 1). Upon the addition ofvarious metallic complexing compounds to themedium, however, the production of chainsresulted (figures 2, 3, 4, 5). If the concentrationof the complexing agent was increased above theoptimal concentration required for the formationof chains, cellular multiplication was inhibited andno growth occurred (table 1). Arrangement ofthe compounds in order of their increased abilityto complex ions as in table 1 indicates thatgrowth, as well as chain formation, is approxi-mately inversely proportional to the ability ofthe agent to bind or complex the metallic ions.
1953] 555
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

EMERIC J. ROCHFORD AND R. J. MIANDLE[
Figuire 1. Typical morphology of Diplococcuispneunmoniae, Type I, when grown in brain heartinfusion broth.
Figuire 2. Morphological appearance of Diplo-coccuis pne?imoniae, Type I, when grown in brainheart infusion broth containing "calgon" (4 X10-4 M).
Figutre 3. Morphological appearance of Diplo-cocculs pneumoniae, Type I, when grown in brainheart infusion broth containinig sodium potassiumtartrate (0.1 iu).
Figuire 4. Morphological appearance of Diplo-coccius pneumnioniae, Type I, when grown in brainheart infusion broth containiing "versene" (8 X10-5 M).
Figure 5. Morphological appearance of Diplo-coccIIs pnemnoniae, Type I, when grown in brainheart infusion broth containing potassium oxalate(4 X 10O3 M).
Figure 6. Morphological appearance of Diplo-coccus pneumoniae, Type I, when grown in resintreated broth containing 0.4 ,ug MIg++ and 1 ,ugCa++ per ml.
It was noted also that the length of the chainsappeared to incr-ease with the propoitionalincrease of the comp)lexing ability of the agentuisedI.
\'arious complexing agents were able to pro-(luce the same result. This suggested that theactioIn in eachi case was similar, namely that theavailability of the critical inorganic ion or ionsbecamle a limiting factor and thereby resultedlin the piroduction of long chains.
The effect of the addition of salts to resin treatedmtiedia. In anI atteml)t to (leimonstiate which of themetallic ions was responsible for the growth an(dclhain foimation, vaiious concentrations of saltswere a(lded to the resin treated medium. Theme(dium, after passage thiough the column, wasunable to support the girowth of D. pneurmoniacunless both Mg++ an(d Ca++ were ieturnedl tothe resin tieatedl broth. In the absence of oneoI both of these, growth (lid not occur even whenFe++ or Mn++ was uise(d singly or- in combination.Growth in this me(lium was (lelendent theiefoieipon the concentrations of Ca++ an(d Mg++.When the Mg++ concentration was inerease(l
in the presence of a constant concentrationi ofCa++ (1 ,ug per ml), which was in excess of thegrowth requirement, growth would inerease in a
mannei directly l)roportional to the Mg++ levelpiesenit in the medlium as shown in figure 15.At a concentration of 0.7 to 0.8 jig per ml ofMg++ or greater, the amount of growth wouldapproach the maximum for this system. Growthwould occuIr, but no chains woul(d be formed whenthe concentration of Mg++ was below 0.4 jigper ml. A higher eoncentration, 0.4 to 0.6 ,jg perml, would result in the plroduction of chains(figure 6). Above this level of Mg++ the cellswould revert to their normal appearance ofdiplococci and the chaining phenomenon wrouldbe lost (figure 10).
Cellular growth was dependent also upon theCa++ concentiationi present in the culture mediumeven in the presence of an excess of Mg++. Al-though the growth of the organism was relatedto the amount of Ca++ present in the medium,the variation of the Ca++ concentration did noteffect an alteration of the morphology of theorganisms from that of diplococci to that ofchains (figure 16). When Ca++ was added to asystem which contained a level of Mg++ (0.4 to0.6 jig per ml) which is in the range of the forma-tion of chains, the cellular morphology would
[VO L. 66556
. *. t
.,*
I
II
* . 11-I
:1 I
0 1
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

PRODUCTION OF CHAIN
(
8
f>\t
9 ' i0-V,
0
Figure 7. Conditions the same as in figure 6but with the Ca++ raised to 1.4 jig per ml.
Figure 8. Conditions the same as in figure 6but with the Ca++ raised to 2.0,gg per ml.
Figure 9. Conditions the same as in figure 6 butwith the Ca++ raised to 5,ug per ml.
Figure 10. Morphological appearance of Diplo-coccus pneumoniae, Type I, when grown in resintreated broth containing 2,g Mg++ and 5 ,g Ca++per ml.
Figure 11. Normal appearance of a rough strainof Diplococcus pneumoniae, Type II, when grownin brain heart infusion broth.
Figure 12. Morphological appearance of a roughstrain of Diplococcus pneumoniae, Type II, whengrown in brain heart infusion broth containing0.2 per cent choline.
Figure 13. Morphological appearance of Diplo-coccus pneumoniae, Type I, when grown in brainheart infusion broth containing 0.2 per centcholine.
Figutre 14. Conditions the same as figure 13
!;S BY D. PNEUMONIAE 557
remain that of chains regardless of the amountof Ca++ present (figure 16) (figures 6, 7, 8, 9).The Mg++ requirement for the production of
diplococcal foims was not met by the substitutionof Ca++, Fe++, or Mn++ either singly or in com-bination. However, it was observed that the num-ber of cells which contributed to making up thechain length appeared to increase when Mn++was added to the medium which contained alevel of Mg++ optimal for chain formation (0.4to 0.6 ,ug per ml) (similar to figure 2). The Mn++did not shift the range of the Mg++ action for.chain formation, but rather its effect seemed tobe upon the length of individual chains.These results were obtained from observations
upon the growth and morphology of D. pneu-moniae, Type I. However, identical results wereobserved with Type II. In order to determinewhether this chaining phenomenon was the resultof some property of the capsular mateiial of theorganism, the experiments were conducted usinga rough strain Type II (D39R) pneumococcus.It was demonstrated that again chaining occurredwhen the Mg++ level was suboptimal (figure 11,12). Since the chaining effect could be producedwith this organism, it was felt that the chainingphenomenon was associated with the cell and itsmechanism for disengaging the paired cocci ratherthan with the capsular substance.
The effect of the addition of choline to normaland resin treated media. In light of the above, itwas desired to confirm the observation of Oka-moto (1940) that the presence of an excess ofcholine promoted long chain formation of D.pneumoniae. The results presented in figure 17demonstrate that in a concentration of 0.20 percent choline chloride, in the brain heart infusionmedium, extremely long chain formations of thepneumococci occurred (figure 13). The concen-tration of the choline was increased up to oneper cent and long chain formation persisted(figure 14). Also, the addition of choline to themedium, up to the amounts used above, had astimulatory effect upon the growth of D. pneu-moniae (figure 17). The same results were obtainedwhen a rough strain derived from D. pneumoniae,Type II, was used (figure 12).
but with the choline concentration raised to oneper cent.
Figures 1 to 14 represent a X 1,200 magnifica-tion of gram stained preparations.
1953]
f .
A
-1
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

EMRIIIC J. ROCHFORD AND 1H. ,J. MANDLE
AWhen varying concentiationis of choline chlo-iide were added to resin tr-eate(d me(lium forti-fied with 0.4 to 0.6 Mg Mg++ and(1 1 Mg Ca++ per
5orNORMAL GROWTH = 70
40-
CI
Hr
30FZONE OF CHAINFORMATION
Co CONSTANT 1.0 ug/ML
20p
lo1
0
0.2 0.4 0.6 0.8 1.0
,ug/ ML MAGNESIUM
Fiqgure 15. The effect of ]Mg-+ on the growthan(l chain formation of Diplococc s pneiumoniaein resin treated brain heart infusion broth.
50
NORMAL GROWTH -70
40
.*---- CHAINS
I,ug/ML
00.49g/ML
H20rc,+
02 0.4 0.6 08 1.0
uq/ ML CALCIUM
Fiqure 16. The effect of Ca++ on the growth andchain formation of Diplococcuts pneunhsoniae in thepresence of a limited magniesium concentration.
ml, the optimal amount of Mg++ for chain forma-tion, long chains of the cocci weire observed.However, in contrast to untreated medium, con-
centrations of one per cent choline chloride tendedto inhibit the growth of the organism. When
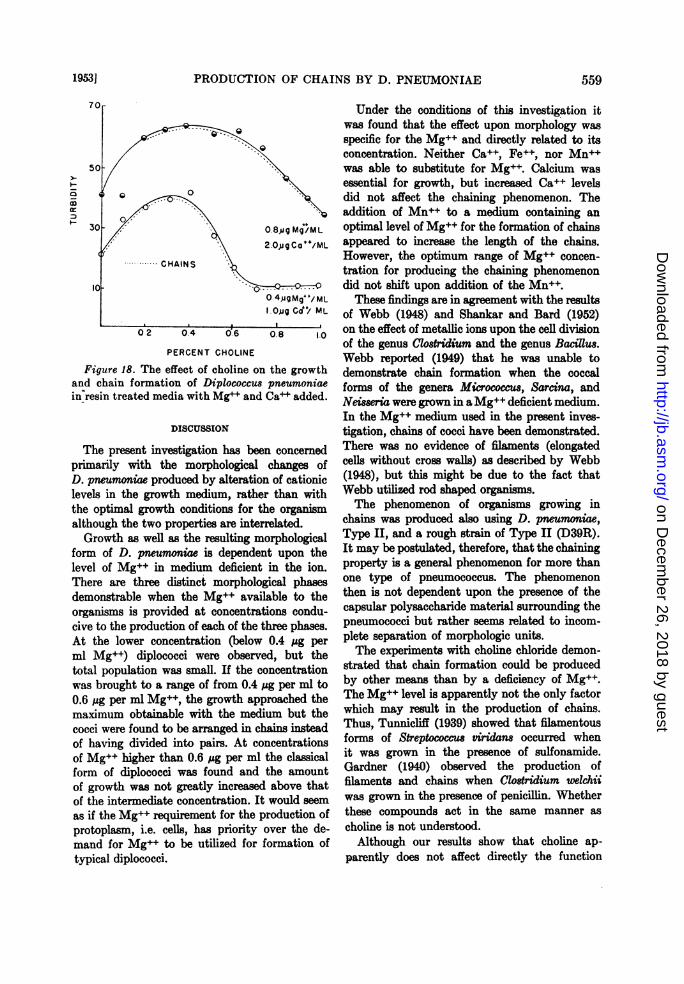
the salt level was (louble(l, the amount of growthincreased, lbut in this system one per cent cholinestill exhibited an inhibitory effect upon the growthof the organism (figure 18).
In or(ler to (leteimine whether choline had theproperty of complexing metallic ions, excesscholine (one per cent) was added to a watersolution which contained 1 ,g Mg++ and 1 MgCa++ per ml. A test for free Mg++ indlicatedl thatthie salts remlined ionized in the presence of thecholine. This riesult was in contrast to similarexperiments using oxalate or versene in which it
80k
70
olf<--~~~
CHAINS
02 0.4 0.6 0.8 1.0
PERCENT CHOLINF
Figurse 17. The effect of choline on the growthand chain formation of Diplococcuis pneum1Roniaein brain heart medium.
was apparent that the ions wrere free in the solu-tion. These findings do not exclude the possibilitythat choline is a weak complexing agent, for itis likely that the reagents used in the test couldcompete successfully for the Mg++ in such amanner as to remove it from the choline and thusgive the appearanice that the Mg++ had not beenbound. However, the extreme length of the chainsobtaine(d when choline was used (figure 14) ascompared to the relativel short ones receivedwNhen a complexing agent such as potassium oxa-late was use(l (figure 5) would make it seem thatthe organism was less able to remove Mg++from choline (if it were bound) than from oxa-late which formns a sufficiently stable complex withMg++ when tested by the colorimetric method.Therefore, the choliine does not appear to bindor complex the metallic ions when it promotesthe formation of long chains in a growth system.
[VOL. 66558
90r
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from
PRODUCTION OF CHAINS BY D. PNEUJMONIAE
I-
E!4 Q 02.O,uqCot/ML
- CHAINS '
cr.~~~~~~~~:30.
0.4pgMg'/MLI 0ug CcP/ ML
0 2 0.4 0 6 0.8 1.0
PERCENT CHOLINE
Figure 18. The effect of choline on the growthand chain formation of Diplococcus pneumoniaein'resin treated media with Mg++ and Ca++ added.
DISCUSSION
The present investigation has been concemedprimarily with the morphological changes ofD. pneumoniae produced by alteration of cationiclevels in the growth medium, rather than withthe optimal growth conditions for the organismalthough the two properties are interrelated.Growth as well as the resulting morphological
form of D. pneumonwae is dependent upon thelevel of Mg++ in medium deficient in the ion.There are three distinct morphological phasesdemonstrable when the Mg++ available to theorganisms is provided at concentrations condu-cive to the production of each of the three phases.At the lower concentration (below 0.4 ug perml Mg++) diplococci were observed, but thetotal population was small. If the concentrationwas brought to a range of from 0.4 jig per ml to0.6 ,ug per ml Mg++, the growth approached themaximum obtainable with the medium but thecocci were found to be arranged in chains insteadof having divided into pairs. At concentrationsof Mg++ higher than 0.6 ,g per ml the classicalform of diplococci was found and the amountof growth was not greatly increased above thatof the intermediate concentration. It would seem
as if the Mg++ requirement for the production ofprotoplasm, i.e. cells, has priority over the de-mand for Mg++ to be utilized for formation oftypical diplococci.
Under the conditions of this investigation itwas found that the effect upon morphology wasspecific for the Mg++ and directly related to itsconcentration. Neither Ca++, Fe++, nor Mn++was able to substitute for Mg++. Calcium wasessential for growth, but increased Ca++ levelsdid not affect the chaining phenomenon. Theaddition of Mn++ to a medium containing anoptimal level of Mg++ for the formation of chainsappeared to increase the length of the chains.However, the optimum range of Mg++ concen-tration for producing the chaining phenomenondid not shift upon addition of the Mn++.These findings are in agreement with the results
of Webb (1948) and Shankar and Bard (1952)on the effect of metallic ions upon the cell divisionof the genus Clostridium and the genus Bacillus.Webb reported (1949) that he was unable todemonstrate chain formation when the coccalforms of the genera Microcu, Sarcina, andNeisseria were grown in aMg++ deficient medium.In the Mg++ medium used in the present inves-tigation, chains of cocci have been demonstrated.There was no evidence of filaments (elongatedcells without cross walls) as described by Webb(1948), but this might be due to the fact thatWebb utilized rod shaped organisms.The phenomenon of organisms growing in
chains was produced also using D. pneu?nonuie,Type II, and a rough strain of Type II (D39R).It may be postulated, therefore, that the chainingproperty is a general phenomenon for more thanone type of pneumococcus. The phenomenonthen is not dependent upon the presence of thecapsular polysaccharide material surrounding thepneumococci but rather seems related to incom-plete separation of morphologic units.The experiments with choline chloride demon-
strated that chain formation could be producedby other means than by a deficiency of MgThe Mg++ level is apparently not the only factorwhich may result in the production of chains.Thus, Tunnicliff (1939) showed that filamentousforms of Streptococcus viridans occurred whenit was grown in the presence of sulfonamide.Gardner (1940) observed the production offilaments and chains when Clostridium welchiiwas grown in the presence of penicillin. Whetherthese compounds act in the same manner ascholine is not understood.Although our results show that choline ap-
parently does not affect directly the function
1953] 559
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from

560EMERIC J. ROCHFORD AND R. J. MANDLE
of the Mg++ in its relationship to the growth andcell division of the organism, it is interesting tonote that the presence of the choline in the growthsystem is capable of producing the same effectupon the morphology of D. pneumoniue as thatof a Mg++ deficiency. It might be that the cholineexerts its influence upon some metabolic processrelated to mechaniism for the disengagement ofthe diplococcal unit with which Mg++ is alsoclosely associated, that is, that the sites of actionof each lie along a common metabolic pathway.Such a relationship would help explain whycholine does not demonstrate any complexingability in the presence of Mg++ and yet has thesame result as that of a Mg++ deficiency.
SUMMARY
The controlled production of long chains ofDiploc pneumonuae was achieved by the useofMg++ deficient media. It was shown that chainformation is a function of the Mg++ concentra-tion and that the progressive changes in mor-phology are exhibited by the population as awhole, dependent upon critical gradients of theions available after the requirements for cellularsynthesis have been met.A comparison has been made with the effect
of suboptimal concentrations of Mg++ on themorphology of D. pneumoniae to the same mor-phologic effect produced by the addition of cholineto brain heart infusion. Although the morpho-logical appearance is identical, the mechanismresulting in chaining evidently is dissimilar.
REFERENCES8BADGER, ELIZABETH 1944 The nutritional re-
quirements of a strain of type III pneumo-coccus. J. Bact., 47, 509-518.
BUCKLEY, E. S., GIBSON, J. G., AND BORToLOTTI,THERESA R. 1951 Simplified titrimetrictechniques for the assay of calcium and mag-nesium in plasma. J. Lab. Clin. Med., 38,751-761.
GARDNER, A. D. 1940 Morphological effects ofpenicillin on bacteria. Nature, 146, 837.
OKAMOTO, H. 1940 Characteristic long chaingrowth of pneumococci produced by quater-nary ammonium compounds. Japan. J. Med.Sci. IV. Pharmacol., 12, 143-156.
SHANKAR, K., AND BARD, R. C. 1952 The effectof metallic ions on the growth and morphologyof Clostridium perfringens. J. Bact., 63, 279-290.
TUNNICLIFF, RUTH 1939 The action of prontosilsoluble and sulfanilamide on the phagocyticactivities of leucocytes and on the dissociationof streptococci. J. Infectious Diseases, 64,279-290.
WEBB, M. 1948 The influence of magnesium oncell division. I. The growth of Clostridiumwelchii in complex media deficient in mag-nesium. J. Gen. Microbiol., 2, 275-287.
WEBB, M. 1949 The influence of magnesium oncell division. II. The effect of magnesiumon the growth and cell division of variousbacterial species in complex media. III. Theeffect of magnesium on the growth of bacteriain simple chemically defined media. J. Gen.Microbiol., 5, 485-495.
560 [VOL. 66
on Decem
ber 26, 2018 by guesthttp://jb.asm
.org/D
ownloaded from





























