Embed Size (px)
Citation preview

Determination of Exogenous Galactose
in Blood using “Specially Purifled”
Glucose Oxidase
Sheldon S. Waldstein, Alvin Dubin, Albert Newcomer,
and Charles H. McKenna
A method is described for the determination of exogenous galactose in blood by amodification of the Nelson-Somogyi hexose procedure in which glucose oxidase is
used to eliminate interfering glucose. The use of “specially purified” glucoseoxidase (SPGO) achieves low, regular blanks and satisfactory replicates, and
enhances accuracy, in contrast to the results obtained with crude enzyme prepara-tions. The ready availability of SPGO makes the method satisfactory and practical
for the clinical laboratory.
rFBE ACC�TRATE DETERMINATION of exogenous galactose in blood by a
method which is rapid and easily adapted to routine blood sugar ineth-
ods is desirable, particularly for the study of liver function and dis-
ease. (ilucose oxidase has been useful for elimination of glucose from
biologic fluids ill order to measure other sugars by methods in which
glucose would be an interfering substance (1). For the past 5 years,
we have used glucose oxidase for this purpose in a colorimetric method
for the determination of exogenous galactose in blood. Until recently,
most enzyme preparations that have been available commercially have
been crude, and when used in this manner have had the important dis-
advantage of producing high, irregular, and unpredictable blanks (1).
In the past 2 years, we have tested and used a preparation, designated
“speciallY purified” glucose oxidase (SPGO) by the manufacturer,
that has proved to he satisfactory for colorimetry.
From the Department of Medicine, Northwestern University Medical School, and the
Hektoen Institute for Medical Research of the Cook County Hospital, Chicago, Ill.
This study was supported by Research Grant H-2187 from the National Heart Tiistitute,
U.S. Public Health Service.
Received for publication Apr. 6, 1962.
381

382 WALDSTEIN fT AL. Clinical Chemistry
It is the purpose of this report to present a simple modification of
the Nelson-Somogyi hexose method suitable for the measurement of
exogenous galactose in blood, and in which the use of SPGO has elim-
inated the irregular color development which occurs when crude glu-
cose oxidase is used.
Materials and MethodsReagents
SPGO* Stock enzyme SPGO is supplied as a lyophilized powder
in vials containing 4400 U. of glucose oxidase.t In addition, catalase
is present in an unquantitated amount. When protracted storage is
anticipated, store the vials in the refrigerator in the dark. To prepare
a working enzyme solution containing 400 U./ml. dissolve the lyophi-
]ized powder in distilled water. Ordinarily, no more than a weeks’
supply of working enzyme solution should be kept on hand.
Zinc sulfate (ZnSO4 . 7H,O), reagent-grade, w/v 5%
Barium hydroxide, reagent-grade, O.3N Upon titration, 5 ml. of
zinc sulfate solution diluted to 25 ml. with distilled water should re-
quire 4.7-4.8 ml. of the barium hydroxide solution to produce a pink
color with phenolphthalein. Adjust the solutions as needed.
Caprylic alcohol, reagent-grade
Somogyi copper reagent (2) Before use, add 1 ml. 15% copper
sulfate (3) to each 24 ml. of this reagent.
Nelson arsenomolybd ate solution (2)
Galactose standard solutions Prepare solutions of galactose in
varying concentrations between 10 and 300 mg./100 ml. with saturated
benzoic acid solution. Galactose, C.P.,1 or 50% galactose injectable
solutionsl are satisfactory. Store in the refrigerator for no more than
2 weeks.
Apparatus
Oxygen train The reaction is carried out in 175 X 22-mm. bac-
teriologic culture tubes. The reaction tubes are closed with rubber
stoppers through which pass two hollow glass tubes, one of which ex-
*Fer,neozynle Type 785, Fermco Laboratories, Inc., Chicago, Ill., distributed by Pfanstiehl
Laboratories, Inc., Waukegan, Ill.
tA unit of glucose oxidase is defined by the manufacturer of SPGO as that amount ofenzyme which will cause up take of 10 ml. oxygen per minute, using 3.3% glucose substrate,
at 300 and pH 5.9.
Available from Pfanstiehl Laboratories, Inc.

Vol. 10, No. 5, 1964 DETERMINATION OF GALACTOSE 383
tends to within 5 mm. of the bottom of the reaction tube and the other
of which extends to a distance well above the surface of the fluid. Con-
nect these to a source of oxygen, whose flow should be adjusted care-
fully to provide equal oxygenation by noting the bubbling in all tubes.
A suitable rack to support the tubes may be constructed from a 2 X 4-
iii. board.
Heparinized containers Add 3 drops of heparin solution, 1000 TJ./
ml., to a number of 25-ml. containers and dry at 60#{176}.These contain
sufficient heparin to provide adequate anticoagulation for as much as
25 ml. of blood.
Preparation of Samples
Reagent blank without enzyme (RB) Add 16 ml. of distilled water
to a reaction tube.
Reagent blank with enzyme (EB) Add 15 ml. of distilled water
and 1 ml. of working enzyme solution to a reaction tube.
Control blood sample Before the introduction of galactose into
the subject, collect (without stasis) 10-25 ml. whole blood for the
measurement of fasting blood sugar and residual reducing substances,
and for the construction of the standard curve for galactose. When
desired, plasma may be substituted for whole blood.
Galact’ose standard curve For each standard, add 13 ml. of dis-
tilled water, 1 ml. of control blood, 1 ml. of the appropriate galactose
standard solution, and 1 ml. of working enzyme solution to a reaction
tube. The galactose standard curve should be constructed to include
the anticipated range of galactose concentration in the unknown
samples.
Residual reducing substances (RB) Add 14 ml. of distilled water,
1 ml. of control blood, and 1 ml. of working enzyme solution to a
reaction tube.
Blood glucose Add 15 ml. of distilled water and 1 ml. of control
blood to a reaction tube.
Unknown samples Collect, without stasis, 3-4 ml. whole blood per
sample at various times after the introduction of galactose into the
subject. Immediately upon collection, add each sample to a heparinized
container, mix, and pipet 1 ml. into a reaction tube containing 14 ml.
of distilled water. Add 1 ml. of working enzyme solution.
Where applicable in all samples described above, add the blood as
soon after shedding as practical, but add the working enzyme solution
just before reaction.

384 WALDSTEIN fT AL. Clinical Chemistry
Procedure
Reaction of Blood with Glucose Oxidase
Add a fine drop of caprylic alcohol to each reaction tube. Connect
the oxygen train and oxygenate the mixtures at room temperature for
30 mm. (Care should be taken that each oxygen delivery tube is placed
to the same depth in each reaction tube.) After 30 mm., disconnect the
oxygen train carefully to avoid aspiration of the sample into the
delivery tubes. From this point on, follow the usual hexose method.
Precipitation
Add 2 ml. of barium hydroxide to each reaction tube, mix, and allow
to stand until all color change has been completed. Add 2 ml. of the
zinc sulfate solution, shake thoroughly, and filter through medium
filter paper. The mixture is now a 1 :20 dilution of the original sample,
and should yield at least 14 ml. of a clear and colorless filtrate which
contains no enzyme activity. Should the filtrate be distinctly colored
or turl)id, discard the sample.
Color Development
Add 1 ml. of the filtrate to a Folin tube. Add I ml. of the modified
copper reagent. Place the Folin tubes in a boiling-water bath for 20
mm. Although glucose develops maximum color in 10-iS mm., galac-
tose does not reach its maximum in less than 20 mm. (3). Cool the
tubes rapidly in an ice bath. Add 1 ml. of the arsenomolybdate reagent
rapidly. Mix thoroughly, and after all foaming has ceased, dilute with
distilled water to 25 ml.
Colorimetry
Galactose develops approximately 83% of the absorbance as glucose
under identical conditions (3). Readings may be made in an Evelyn
colorimeter, using the macrocuvet (10-mm. slit) at 515 mp�, or in other
appropriate instruments. Set the colorimeter to 100% transmittance
using the reagent blank without enzyme (RB). Read the reagent
blank containing enzyme (EB) against the RB. Satisfactory enzyme
solutions produce an EB which gives no more color than 95% T
against the RB. Reset the colorimeter to 100% T, using the EB. Read
all unknown samples and galactose standards against the EB. Con-
struct the standard curve. It sho#{252}ldcross the ordinate at the RS value.
Read the values for the unknown samples from the standard curve for
galactose constructed for each run.

Vol. 10, No. 5, 1964 DETERMINATION OF GALACTOSE 385
ResultsShelf Life
Under usual laboratory conditions, SPGO appeared stable for pro-
tracted periods. Seven lots of SPGO solution were frozen for up to
9 months and tested at regular monthly intervals for activity against
glucose. No loss of activity was noted. Lyophilized preparations of
SPGO stored in the refrigerator or at room temperature in the dark
were likewise found to maintain full activity for several months. When
such precautions were not taken, partial loss of activity occurred.
Recalibratiori of the unitage of a batch of lyophilized SPGO after 10
months of storage at room temperatures under ambient 1i�-ht showed
that 10% of the original glucose oxidase activity and 30% of the
original catalase activity was lost (4). Thus it can be expected that no
appreciable loss of glucose oxidase activity will occur for protracted
periods if lyophilized SPGO is stored in darkness in the refrigerator.
For the present method, the enzyme solution must produce a low
and regular blank. Storage of the working solution was found to re-
sult in an increasing blank with time, so that eventually the color of
the blank exceeded the color produced by low concentrations of galac-
tose. This occurred at varying intervals, depending upon the mode
of storage. Working enzyme solutions stored in the refrigerator ex-
cept when used remained satisfactory for 10-14 days. Solutions
frozen immediately after preparation were stored for periods up to
1 year, and were as satisfactory when thawed as were freshly prepared
solutions. Daily thawing and freezing, however, appeared to cause
more rapid deterioration than storage at refrigerator temperatures.
Therefore it has been our practice to divide large volumes of working
enzyme solution into aliquots (each of a volume sufficient for the num-
ber of weekly determinations) and to freeze them. At the beginning
of each week, a new aliquot is thawed, stored in the refrigerator, and
discarded at the end of the week. A careful record of each EB is kept,
and the remaining frozen aliquots of the original batch discarded if
the EB begins to show consistently excessive color. Six- to 12-month
supplies of working enzyme solution have been stored and used satis-
factorily in this way. With the present availability of lvophilized
SPGO in convenient quantities, however, such extremes of storage
time for working solutions of the enzyme need not be encountered.
Enzyme Characteristics which Affect the Method
Glucose oxidase exhibits significant activity over a pH range of 4-8(s). Gluconic acid is generated rapidly, and in unbuffered aqueous

386 WALDSTEIN FT AL. Clinical Chemistry
solutions, a prompt drop of pH below 4.0 occurs, which limits the re-
action. Buffering is therefore necessary in aqueous media. A phos-
phate buffer at pH 7.0 has proved satisfactory for the present method,
since it is near the optimum pH of the enzyme and does not interfere
with the protein precipitants used. In blood, the buffers contained
within a 1-ml. sample have been found to be sufficient to maintain the
pH of the reaction mixture well within the range of optimum activity.
The enzyme is active over a wide range of temperature. Impairment
in activity is noted at 0#{176},but at room temperatures and at 37#{176},destruc-
tion of glucose in the medium is virtually complete within the specified
reaction time. Accordingly, the method is carried out at room tem-
perature.
Oxygen, required for the oxidation of glucose, must be supplied to
the reaction mixture. Although it has been possible to secure oxygena-
tion by placing the 16-mi. reaction mixture in a large, flat-bottomed
flask, thereby creating a thin layer of fluid into which oxygen can pass
by diffusion, optimum oxygenation has been achieved by bubbling
oxygen through the mixture.
Oxidation of glucose in the presence of glucose oxidase is a first-
order reaction. Under the conditions of the method, the reaction ap-
proaches completion within 15 mm. but does not reach equilibrium for
60-120 mm. Since it has been shown previously that glucose oxidase
attacks galactose to a limited extent (3), a variety of reaction times
were studied to select one resulting in the most glucose removal with
the least galactose loss. A minimum of 15 mm. and a maximum of 60
mm. met these objectives when the recommended concentration of
enzyme was used. Therefore, 30 mm. of reaction time was selected
as most convenient.
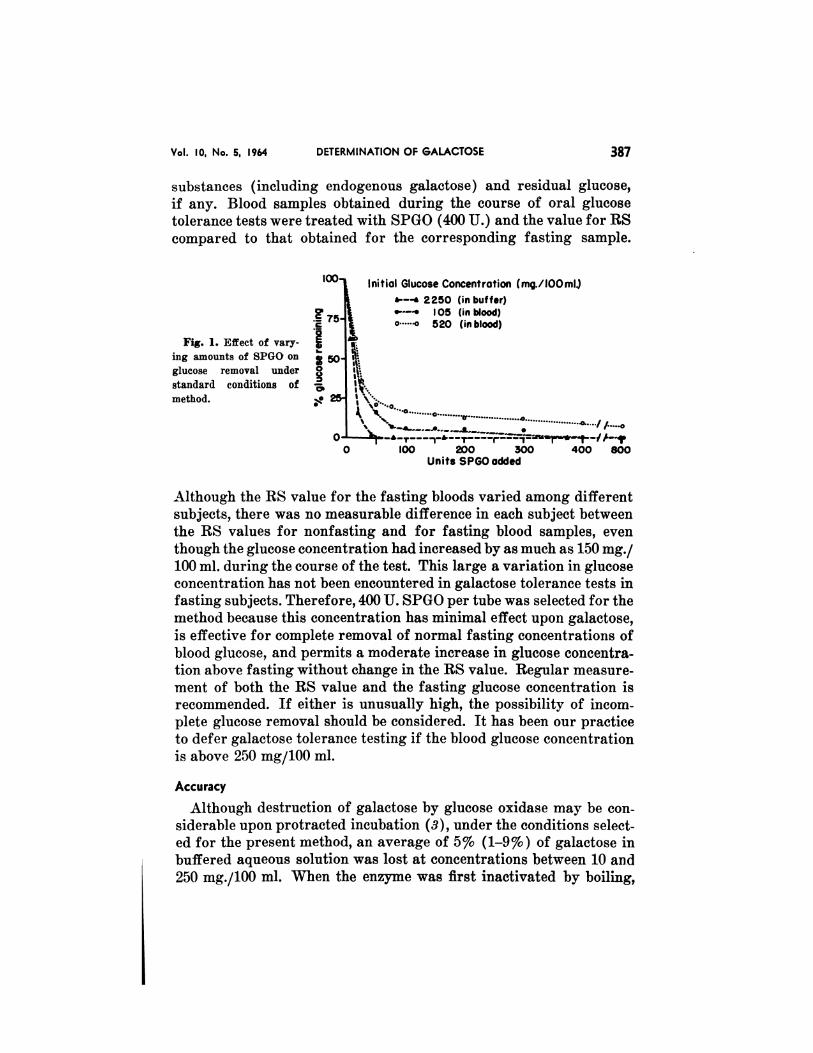
The effect of enzyme concentration was studied in buffered aqueous
solution and in blood (Fig. 1). In buffer, approximately 40 U. SPGO
pci’ tube was sufficient to eliminate glucose at an initial concentrationgreater than 2000 mg./i00 ml. In blood, however, 400 U. of SPGO per
tube was necessary to effect elimination of glucose at a typical fasting
concentration, and 800 U. SPGO per tube did not effect complete re-
moval of glucose when an initial concentration over 500 mg./100 ml.
was present. The reason for inhibition of enzyme activity in blood is
unknown. Because blood may contain peroxidase inhibitors, an at-
tempt was made to overcome the inhibition by addition of catalase in
excess of that already contained in SPGO. No effect upon the inhibi-
tion was found. When blood is reacted with the enzyme, the value for
residual reducing substances (RS) represents nonglucose-reducing
Initial Glucose Concentration (mgI 100 nil.)
�--‘ 2250 (in buffer)I 05 (in blood)
#{176} 520 (inblood)
0 100 200 300Units SPGO added
400 800
Vol. 10, No. 5, 1964 DETERMINATION OF GALACTOSE 387
substances (including endogenous galactose) and residual glucose,
if any. Blood samples obtained during the course of oral glucose
tolerance tests were treated with SPGO (400 U.) and the value for RS
compared to that obtained for the corresponding fasting sample.
Fig. 1. Effect of vary-
jug amounts of SPGO on
glucose removal under
standard conditions of
method.
Although the RS value for the fasting bloods varied among different
subjects, there was no measurable difference in each subject between
the RS values for nonfasting and for fasting blood samples, even
though the glucose concentration had increased by as much as 150 mg./
100 ml. during the course of the test. This large a variation in glucose
concentration has not been encountered in galactose tolerance tests in
fasting subjects. Therefore, 400 U. SPGO per tube was selected for the
method because this concentration has minimal effect upon galactose,
is effective for complete removal of normal fasting concentrations of
blood glucose, and permits a moderate increase in glucose concentra-
tion above fasting without change in the RS value. Regular measure-
ment of both the RS value and the fasting glucose concentration is
recommended. If either is unusually high, the possibility of incom-
plete glucose removal should be considered. It has been our practice
to defer galactose tolerance testing if the blood glucose concentration
is above 250 mg/100 ml.
Accuracy
Although destruction of galactose by glucose oxidase may be con-
siderable upon protracted incubation (3), under the conditions select-
ed for the present method, an average of 5% (1-9%) of galactose in
buffered aqueous solution was lost at concentrations between 10 and
250 mg./100 ml. When the enzyme was first inactivated by boiling,

N represents Number of pairs; D., means difference between pairs; S.D., Standard Devia-
tion of difference between pairs.
388 WALDSTEIN FT AL. Clinical Chemistry
complete recovery of all concentrations of galactose was obtained. In
blood, less galactose is probably lost because of the inhibition of
enzyme activity. Nevertheless, it is recommended that, in each blood
studied, a standard curve for galactose be constructed encompassing
the anticipated range of blood concentrations.
An estimate of the accuracy of the present method was obtained by
analysis of the difference between duplicate measurements of known
concentrations of galactose between 10 and 250 mg./100 ml., both in
buffered aqueous solution and in whole blood (Table 1). These differ-
ences include procedural error as well as error due to variation iii
enzyme activity. Concentration had no apparent effect upon accuracy.
In aqueous solution, the difference between duplicates was well within
1% T. This is of the same order of accuracy obtained for the Nelson-
Somogyi hexose method alone using the same instrumentation, and
suggests that the enzyme, SPGO, contributes little sampling error
per se to the method. In blood, greater sampling variation was found,
although the mean difference between duplicates was 1% T or less.
The reason for the variation being greater in blood than in aqueous
solution is not known, but may be related to the partial inhibition of
enzyme activity in blood, noted above. Because of the shallow slope
of the galactose standard curve, the mean difference between dupli-
cates is equivalent to approximately 2 mg./100 ml. galactose in aqueous
solution, and to 4 mg./100 ml. galactose in blood; differences as great
as 10 mg./100 ml. may be encountered in blood.
On the other hand, when ordinary commercial crude glucose oxidase
was employed under the conditions of the present method, duplication
was unsatisfactory (Fig. 2). The differences between duplicates using
a crude preparation were equivalent to 30-40 mg./100 ml. galactose.
Table 1. DIFFERENCES BErWEEN DUPLICATES OF GALACPOSE STANDARDS WITH USE OF SPGO
Concentration gaiaetose
added (mg/joe mi)
Galactos e standards in buffer G&actose standards in blood
N D.±S.D. (% T) N D.±S.D. (% T)
10 41 0.22 ± 0.28 43 0.71 ± 0.72
25 72 0.39 ± 0.50 66 0.84 ± 0.90
40 30 0.20 ± 0.23 27 0.93 ± 0.72
50 76 0.28 ± 0.30 70 0.78 ± 0.83
100 64 0.37 ± 0.30 60 0.82 ± 0.78
150 49 0.50 ± 0.55 46 1.17 ± 0.89
200 55 0.37 ± 0.30 49 1.09 ± 0.97
250 22 0.44 ± 0.44 19 1.05 ± 0.84
All concentrations (ToTAL) 409 0.41 ± 0.32 380 0.90 ± 0.82

#{149}
#{149}
S
S
S
Vol. 10, No. 5, 1964 DETERMINATION OF GALACTOSE 389
Variation of such magnitude is largely inherent in crude preparations
of the enzyme and does not appear only in the presence of glucose or
galactose. The color produced by duplicate blanks containing no en-
zyme or of duplicate blanks containing SPGO has been consistently
#{149}Crude glucose o.s’idose.#{149} I O�pe�ilol/ypurif�jd
Fig. 2. Variation of final color developed glucose oxidose
under conditions of method from replicate - 1 :“
samples of blood (with and without added
galactose) reacted with crude glucose oxidase
or SPGO. Dotted lines indicate 1% trans. -
mittance difference. -
GLUCOSE GALACTOSE GALACTOSE9Omg./IOOml. 25 mg/lOOmI. l5OmG/I00 ml.
within 1% T, but the color produced by duplicate blanks containing
crude glucose oxidase varies irregularly and has been as great as
6% T. For these reasons, crude glucose oxidase is unsatisfactory for
the present method and the use of SPGO is recommended.
Discussion
In 1928, Muller (6) obtained an enzyme from fungi of Aspergillus
and Penicillium capable of catalyzing the oxidation of glucose. He
named this enzyme “glucose oxidase,” and although it has since been
shown to function as a dehydrogenase (7), the original and familiar
designation has been used in this report.
In this country, glucose oxidase is commonly derived from the mold
mycelia of Aspergillus niger grown on liquid media containing glucose.
Most preparations available here are crude, if highly concentrated.
However, preparations of greater purity have been made and used in
reported methods (1,8-10). One such preparation, now available corn-
mercially, contains “specially purified” glucose oxidase (SPGO);its coenzyme, FAD; and amounts of catalase adequate for the removal
of 11202 as it is formed.
Glucose oxidase catalyzes the following reaction:
glucoSeC,;Hi.,0,i + 1120 + Oi . � C611,207 + H20�
oxidase
Alternative approaches for the use of the enzyme in the determina-
tion of glucose have been developed. In one, the hydrogen peroxide
produced is reacted in the presence of peroxidase with a chromogenic
oxygen acceptor such as o-tolidine or o-dianisidine. The intensity of
the resulting color is proportional to the amount of glucose (11). This

390 WALDSTEIN ET AL. Clinical Chemistry
approach is the basis for the commonly used test strips for estimating
urine glucose (12). In another method, oxygen uptake is measured
manometrically, there being a stoichiometric relation between glucose
oxidized and oxygen consumption (13). In a third approach, total
hexose is measured before and after incubation of the sample with the
enzyme, the difference representing glucose (14).
Of more than 50 sugars (other than glucose) tested, glucose oxidase
has been found to catalyze the oxidation of galactose (3), mannose,
xylose, 6-methylglucose, 4-6-dimethylglucose (5), and 2-deoxy-d-glu-
cose (is). Since it attacks these sugars to a very limited extent it is
possible to use the enzyme to eliminate glucose from biologic fluids
and to determine these “foreign” sugars by appropriate means with-
out interferelice. In this manner, Tygstrup et al. (9) used “notatin”
to measure galactose, and Froesch et al. (1) used a commercially un-
available, highly purified glucose oxidase preparation to measure
inulin. Because the Nelson-Somogyi method is used widely for the
measurement of blood glucose, it seemed to us appropriate to adapt it
in like manner for the measurement of galactose. The procedure, as
developed in this laboratory, adds but 35-40 mm. to the usual glucose
method and requires little additional equipment. The use of SPGO,
a potent, stable, and uniform preparation of glucose oxidase, has
eliminated the irregularities produced by crude glucose oxidase and
has greatly enhanced the accuracy of galactose determination. The
commercial availability of SPGO should facilitate measurement of
galactose and permit greater use of galactose tolerance tests in clin-
ical medicine.
References
1. Froesch, E. H., Reardon, J. B., and Renold, A. E., J. Lab. 4� Gun. Med. 50, 918 (1957).
2. Nelson, N., J. Biol. Chem. 153, 375 (1944).
3. w�ld�t�th, S. S., Areilla, H. A., and Dubin, A., Quart. Bull. I’Torthwe,itern Univ. Mcd.
School 31, 323 (1957).
4. Se�tt, D., Personal communication, April, 1961.
5. Keilin, D., and Hartree, E. F., Bioche’m. J. 42, 221 (1948).
6. Miller, D., Bioehem. Ztschr. 199, 136 (1928).
7. Theorell, H., in The Enz2,mes (vol. 2). J. Summer, and K. Myrback, Eds. Academic,
New York, 1951, p. 348.
8. Coulthard, C. E., Miehaelis, R., Short, W. F., Sykes, G., Skrimshire, 0., Standfast, A.,
Birkinshaw, J., and Raistriek, H., Biochem. J. 39, 24 (1945).
9. Tygstrup, N., Winkler, K., Lund, E., and Engell, H. C., Scand. J. Clin. 4� Lab. Invest.
6,43 (1954).
10. Marks, \T�, Clin. Chim. Acta 4, 395 (1959).11. Saifer, A., and Gerstenfeld, S., J. Lab. 4�Ctin. Med. 51,448 (1958).
12. Comer, J. P., Anal. (Them. 28, 1748 (1956).
13. Keilin, D., and Hartree, E. F., Biochein. J. 42, 230 (1948).
14. Froeseh, E. B., and Renold, A. E., Diabetes 5, 1 (1956).
15. MeComb, B. B., and Yushok, W. D., J. Franklin Inst. 265, 417 (1958).










![Index [application.wiley-vch.de]€¦ · 1,4-linked3,6-anhydro-𝛼-L-galactose 256 1,3-linked𝛽-D-galactose 256 liverbiology celltypes 302 extracellularmatrix 303 histologicalstructure](https://img.pdfslide.us/doc/110x75/5f8b7e1f5ead5860b4325134/index-14-linked36-anhydro-l-galactose-256-13-linked-d-galactose.jpg)


















