Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 83, pp. 7775-7779, October 1986Cell Biology
Enhancement of galactose/N-acetylgalactosamine receptor activityon the surface of freshly isolated rat hepatocytes: Evidence formasking of receptor sites by inhibitors derived fromcollagenase preparations
(endocytosis/hepatic parenchymal ceils/asialoglycoproteins/perfused liver)
NANCY LACLAIR STULTS AND YUAN CHUAN LEE*Department of Biology and The McCollum Pratt Institute, The Johns Hopkins University, Baltimore, MD 21218
Communicated by Saul Roseman, June 24, 1986
ABSTRACT Rat hepatocytes prepared by collagenaseperfusion of the liver are known to exhibit increased binding ofasialoorosomucoid (ASOR) after prior treatment with EDTAor after warming at 37°C. The cause of the apparent increasein the surface binding activity of the galactose/N-acetylgalac-tosamine (Gal/GalNAc) receptor on freshly isolated rathepatocytes was investigated. Binding experiments using threedifferent galactose-terminated ligands revealed up to a 2- to6-fold increase in the level of surface receptor sites on rathepatocytes upon prior incubation at 4°C with 10 mM GalNAcor 10 mM EDTA or at 37°C compared to untreated cells. Withdigitonin-permeabilized cells, it was shown that the newlyexposed receptor sites most likely originated from maskedsurface receptor sites, as no alteration in the size of the internalpool of receptor was observed. Collagenase preparations werefound to inhibit the binding of 121I-labeled ASOR to theGal/GalNAc receptor. Exposure of hepatocytes to collagenaseresulted in a significant decrease in 12.5-labeled ASOR binding,which was reversible upon treatment with GalNAc or EDTA at4°C or upon warming at 37°C. Perfusion ofEDTA through theisolated whole liver at 0-20C to remove any possible boundendogenous ligands did not result in a significant increase in thelevel of 1251-labeled ASOR binding, while perfusion of colla-genase caused a marked decrease in the binding activity of theliver. We conclude that the enhancement of Gal/GalNAcreceptor activity on the surface of freshly isolated hepatocytesby temperature and EDTA is potentially an artifact of thecollagenase perfusion method.
The galactose/N-acetylgalactosamine (Gal/GalNAc) recep-tor of the parenchymal cells of mammalian liver functions inthe binding and internalization of galactose- and GalNAc-terminated oligosaccharides and glycoproteins (for review,see refs. 1-4). This receptor system has been extensivelystudied in both the isolated perfused liver (5) and in isolatedhepatocytes prepared by a two-step collagenase-perfusiontechnique (6) with the goal of elucidating the pathway andmolecular mechanisms of receptor-mediated endocytosis.The first step in the pathway of Gal/GalNAc receptor-mediated endocytosis is the specific binding of ligand toreceptors on the plasma membrane of hepatocytes. In thecase of freshly isolated rat hepatocytes, there is considerablevariability in the literature regarding the surface bindingbehavior of asialoorosomucoid (ASOR), particularly in themagnitude of the receptor site number. The sources ofvariability in site numbers for this receptor are potentiallynumerous with regard to the conditions of the collagenaseperfusion protocol and the binding assay and have been
acknowledged (7). In addition, several treatments have beenreported to affect the expression of surface receptor activityin this system and include temperature (8-12) and EDTA (9,13).
In this paper we present evidence that collagenase prepa-rations contain a carbohydrate-containing inhibitor(s) that isspecifically bound by rat hepatocytes during the collagenase-perfusion step. This inhibitor(s) was readily dissociated fromthe receptor by displacing agents at 40C and apparently wascleared from the hepatocyte surface upon warming at 370C.These findings not only further explain the large reportedrange of binding parameters for this receptor system butsuggest that receptor enhancement (that is, the increase insurface receptor site number observed after treatment offreshly isolated hepatocytes with EDTA or after warming)may represent the removal of collagenase inhibitor(s) fromsurface receptor sites. In addition, these results indicate thatreceptor enhancement observed when freshly isolatedhepatocytes are warmed cannot be explained entirely on thebasis of receptor repair or on a temperature-sensitive mod-ulation of surface receptor activity, which has recently beenproposed (11).
MATERIALS AND METHODSReagents. The vitamins and amino acids used in the
preparation of medium were from Sigma or Calbiochem.Reagent-grade chemicals, including disodium EDTA, werepurchased from Baker. Hepes was obtained from ResearchOrganics (Cleveland, OH). Trypan blue, crystal violet,digitonin, monosaccharides, EGTA, polyvinylpyrrolidone(PVP-40), and DNase I were from Sigma. Bovine serumalbumin (BSA) (fatty acid-free) was obtained from Armour(Kankakee, IL). The 4-methylumbelliferyl a-keto glycosideof N-acetylneuraminic acid was prepared in this laboratory(14). The collagenase preparations from Clostridium histolyt-icum were purchased from Cooper Biomedical (Malvern, PA)and Sigma. Arthrobacter ureafaciens neuraminidase was agift from Y. Uchida. Carrier-free Na125I (13-17 mCi/pg; 1 Ci= 37 GBq) was obtained from Amersham. Orosomucoid wasdonated by M. Wickerhauser of the American Red CrossFractionation Center and was desialylated by using immobi-lized neuraminidase (15). BSA containing 44 mol of 1-thio-D-galactopyranoside per mol of protein, linked via an amidinogroup (Gal44-AI-BSA) and a,3-diaspartamide of tris[(,B-lac-
Abbreviations: ASOR, asialoorosomucoid; BSA, bovine serum albu-min; di-tris-lac, a,,-diaspartamide of tris[(P-lactosyloxy)methyl](6-aminohexanamido)methane; Ga144-AI-BSA, bovine serum albumincontaining 44 mol of 1-thio-D-galactopyranoside per mol of proteinlinked via an amidino group; Gal/GalNAc, galactose/N-acetylga-lactosamine.*To whom reprint requests should be addressed.
7775
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement"in accordance with 18 U.S.C. §1734 solely to indicate this fact.
Dow
nloa
ded
by g
uest
on
Nov
embe
r 4,
202
1
7776 Cell Biology: Stults and Lee
tosyloxy)methyl](6-aminohexanamido)methane (di-tris-lac)modified with p-nitrophenyl-tert-butyloxy-carbonyl-L-tyrosine were prepared as described (16, 17). The cellmedium consisted of a modified Eagle's medium (pH 7.4)supplemented with amino acids (18). The medium contained10 mM Hepes for use at 0-40C (medium I) or 50 mM Hepesfor use at 370C (medium II).Methods. The microbiuret assay (19) was used to measure
protein concentrations. Cell concentration was determinedby counting either the cells directly or crystal violet-stainednuclei (6, 20). Collagenase was heat-denatured by boiling for%15 min in water at a concentration of 5-10 mg/ml. Underthese conditions, there were no remaining detectable colla-genase, clostripain, caseinase, or trypsin activites (21). Sev-eral lots of collagenase were screened for neuraminidaseactivity by measuring the hydrolysis of sialic acid fromorosomucoid (22) or 4-methylumbelliferyl a-keto glycoside ofN-acetyl neuraminic acid (14).
Preparation of Isolated Rat Hepatocytes. Hepatic parenchy-mal cells were prepared by modification of the two-stepcollagenase perfusion technique (6) from either maleSprague-Dawley rats obtained from Harlan (Walkersville,MD) or from male F334 weanling rats purchased from theNational Cancer Institute (Frederick, MD) (23). All cellpurification steps and subsequent manipulations were carriedout at 40C unless otherwise indicated. The resulting hepato-cyte preparations were routinely >90% viable as judged bymorphology and exclusion of trypan blue.Whole Liver Perfusion. Liver perfusion experiments were
carried out as described at 0-2°C (5). The following stepswere carried out in the whole liver perfusion experiment: (i)125I-labeled ASOR (125I-ASOR) was added to the recirculat-ing perfusion system to a final concentration of 65 nM andallowed to bind to the liver for 30 min, (it) unbound ligand wasremoved from the liver by perfusion with buffer for 10 min,(iii) buffer containing 10 mM EDTA was perfused through theliver for 5 min to remove specifically bound ligand, and (iv)calcium was restored to the liver by perfusion with buffer for10 min. The amount of EDTA-dissociable radioactivityobtained from the liver in the third step was used to calculatethe level of specifically bound 1251-ASOR. This binding cycle(ligand, wash, EDTA, wash) was repeated several times todetermine the effect of passing collagenase through the liver.
Treatments of Rat Hepatocytes. Rat hepatocytes at 4°C(2.5-5 x 106 per ml) were treated with 10 mM EDTA,GalNAc, GlcNAc, or mannose or with 100 nM di-tris-lac inmedium I at 4°C. After the incubation period, the cellsuspensions were washed 2-3 times by resuspension andcentrifugation at 50 x g with medium I. Hepatocytes werewarmed rapidly by diluting a concentrated cell stock 1:5 to1:10 (usually to 2.5 x 106 cells per ml) into warmed mediumII containing 0.1% BSA (medium II/BSA). Incubations werecarried out in a waterbath shaker at 80-100 rpm inpolycarbonate flasks. After the cells were warmed, they wererapidly cooled by adding a 5- to 10-fold volume of ice-coldmedium I, followed by centrifugation and resuspension.Hepatocytes (2.5 x 106 per ml) were treated with 0.5 mg ofcollagenase per ml, and after the incubation period, the cellswere washed with medium I. To reverse the observedaggregation of hepatocytes when incubated with collagenaseat 37°C, the cells were treated with DNase (6.7 ,g/ml) at 4°Cfor 30 min.
Ligand Binding. Ligands were radioiodinated by a modi-fication of the chloramine-T method (24). Direct bindingassays were carried out as previously outlined, and nonspe-cific binding was routinely assessed by using a nondilutingEDTA release protocol (25). The data were analyzed by usinga modified version of the nonlinear multiparameter regres-sion program LIGAND (26) as described (25). Nonspecificbinding was calculated based on the LIGAND-determined
value of the N parameter, which is the limiting [bound]/[free]ligand ratio. The graphical display of the data represents totalbinding from which nonspecific binding obtained from the Nparameter has been subtracted. The binding affinity ofdifferent lots of collagenase for the Gal/GalNAc receptor onrat hepatocytes was estimated by using an inhibition assaywith 115I-ASOR as the labeled ligand (27).Measurement of Internal Receptor Sites. The digitonin-
permeabilization technique (28) was used to measure internalreceptor binding after masking surface receptor sites byincubation with 2 AM Gal4-AI-BSA for 1 hr at 40C. After freeGal44-AI-BSA was washed away, the cells were treated with0.055% digitonin at 40C for 15-20 min. Digitonin was addedto the cell suspension in absolute ethanol. After centrifuga-tion and removal of the medium, the cells were resuspendedin medium I/BSA and subjected to the direct binding assaywith 125I-ASOR.
RESULTS
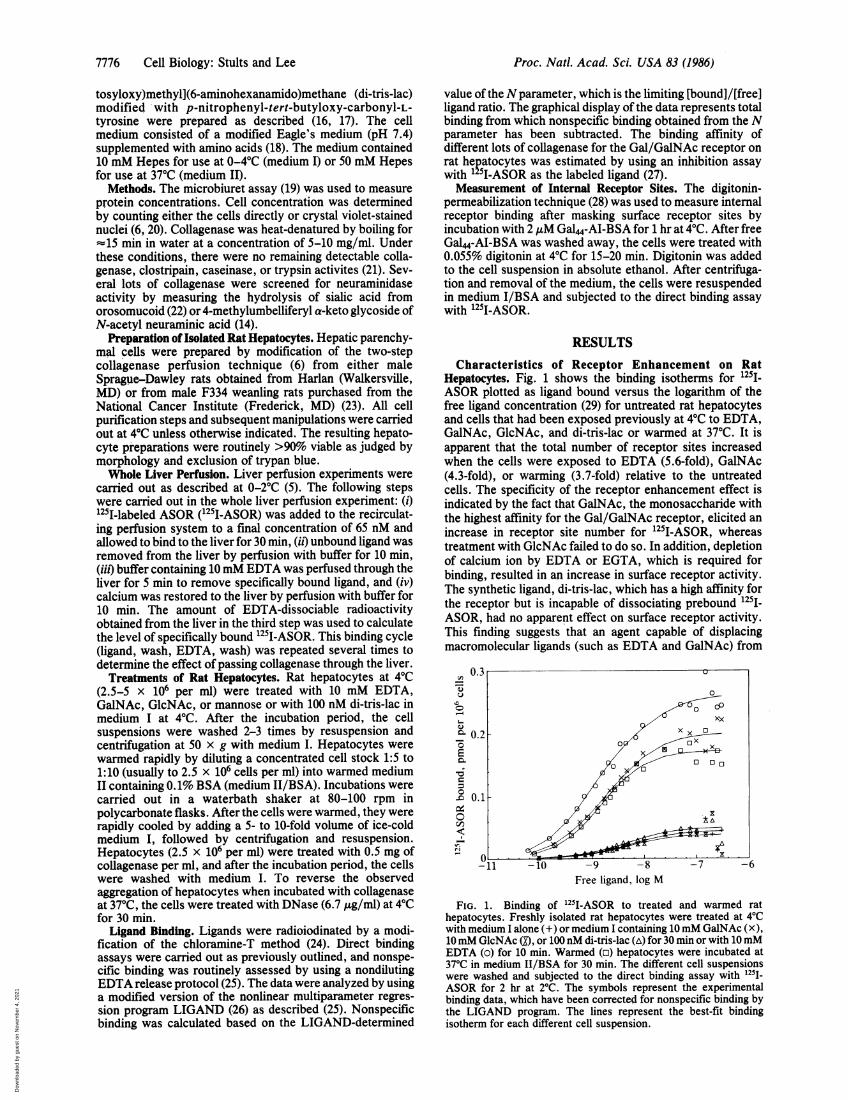
Characteristics of Receptor Enhancement on RatHepatocytes. Fig. 1 shows the binding isotherms for 125I1ASOR plotted as ligand bound versus the logarithm of thefree ligand concentration (29) for untreated rat hepatocytesand cells that had been exposed previously at 40C to EDTA,GalNAc, GlcNAc, and di-tris-lac or warmed at 370C. It isapparent that the total number of receptor sites increasedwhen the cells were exposed to EDTA (5.6-fold), GalNAc(4.3-fold), or warming (3.7-fold) relative to the untreatedcells. The specificity of the receptor enhancement effect isindicated by the fact that GalNAc, the monosaccharide withthe highest affinity for the Gal/GalNAc receptor, elicited anincrease in receptor site number for 125I-ASOR, whereastreatment with GlcNAc failed to do so. In addition, depletionof calcium ion by EDTA or EGTA, which is required forbinding, resulted in an increase in surface receptor activity.The synthetic ligand, di-tris-lac, which has a high affinity forthe receptor but is incapable of dissociating prebound 125i-ASOR, had no apparent effect on surface receptor activity.This finding suggests that an agent capable of displacingmacromolecular ligands (such as EDTA and GalNAc) from
0.3 U
0
0 P- X XX
0.2-~0
-11 -10 - -
.n0.1X
o0')
-11 -10 -9 -8 -7 -6Free ligand, log M
FIG. 1. Binding of 125I-ASOR to treated and warmed rathepatocytes. Freshly isolated rat hepatocytes were treated at 40Cwith medium I alone (+) or medium I containing 10mM GalNAc (X),10 mM GlcNAc (O, or 100 nM di-tris-lac (A) for 30 min or with 10mMEDTA (o) for 10 min. Warmed (o) hepatocytes were incubated at370C in medium II/BSA for 30 min. The different cell suspensionswere washed and subjected to the direct binding assay with l25I-ASOR for 2 hr at 20C. The symbols represent the experimentalbinding data, which have been corrected for nonspecific binding bythe LIGAND program. The lines represent the best-fit bindingisotherm for each different cell suspension.
Proc. Natl. Acad. Sci. USA 83 (1986)
Dow
nloa
ded
by g
uest
on
Nov
embe
r 4,
202
1
Proc. Natl. Acad. Sci. USA 83 (1986) 7777
the Gal/GalNAc receptor is necessary for the enhancementeffect. In view of these results, we reasoned that warmingwas also effecting the removal of a carbohydrate-containingmolecule(s) from surface receptor sites; however, the mech-anism of displacement from the surface receptor was pre-sumably internalization. Freshly isolated hepatocytes alsoexhibited a similar increase in surface receptor activity forthe small synthetic cluster ligand 125I-labeled di-tris-lac andthe neoglycoprotein "251-labeled Gat4-AI-BSA after EDTAand GalNAc treatment or after incubation at 37°C.
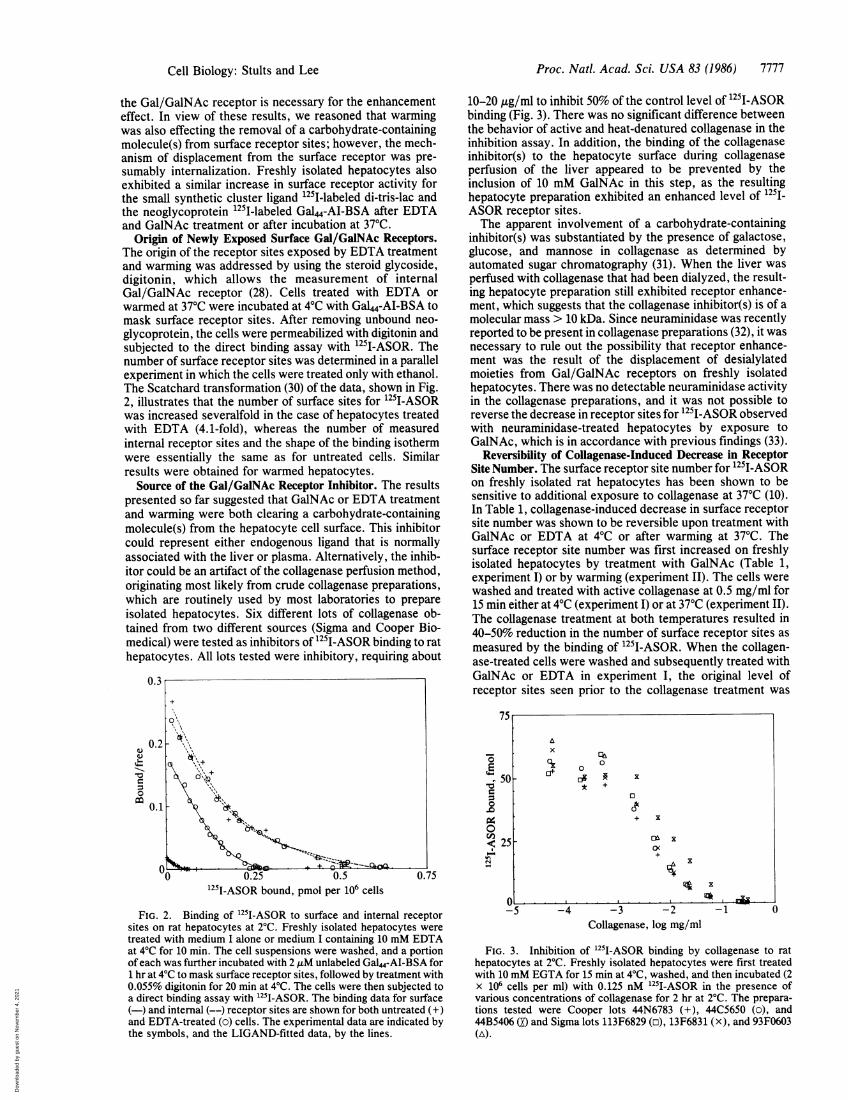
Origin of Newly Exposed Surface Gal/GalNAc Receptors.The origin of the receptor sites exposed by EDTA treatmentand warming was addressed by using the steroid glycoside,digitonin, which allows the measurement of internalGal/GalNAc receptor (28). Cells treated with EDTA orwarmed at 37°C were incubated at 4°C with GaL4-AI-BSA tomask surface receptor sites. After removing unbound neo-glycoprotein, the cells were permeabilized with digitonin andsubjected to the direct binding assay with 1251I-ASOR. Thenumber of surface receptor sites was determined in a parallelexperiment in which the cells were treated only with ethanol.The Scatchard transformation (30) of the data, shown in Fig.2, illustrates that the number of surface sites for 1251I-ASORwas increased severalfold in the case of hepatocytes treatedwith EDTA (4.1-fold), whereas the number of measuredinternal receptor sites and the shape of the binding isothermwere essentially the same as for untreated cells. Similarresults were obtained for warmed hepatocytes.
Source of the Gal/GalNAc Receptor Inhibitor. The resultspresented so far suggested that GalNAc or EDTA treatmentand warming were both clearing a carbohydrate-containingmolecule(s) from the hepatocyte cell surface. This inhibitorcould represent either endogenous ligand that is normallyassociated with the liver or plasma. Alternatively, the inhib-itor could be an artifact of the collagenase perfusion method,originating most likely from crude collagenase preparations,which are routinely used by most laboratories to prepareisolated hepatocytes. Six different lots of collagenase ob-tained from two different sources (Sigma and Cooper Bio-medical) were tested as inhibitors of 1251I-ASOR binding to rathepatocytes. All lots tested were inhibitory, requiring about
0.3
0.2
0.1X
0 0.25 0.5 0.751251I-ASOR bound, pmol per 106 cells
FIG. 2. Binding of 251I-ASOR to surface and internal receptorsites on rat hepatocytes at 2°C. Freshly isolated hepatocytes weretreated with medium I alone or medium I containing 10 mM EDTAat 4°C for 10 min. The cell suspensions were washed, and a portionof each was further incubated with 2 ,uM unlabeled Gal"-Al-BSA for1 hr at 4°C to mask surface receptor sites, followed by treatment with0.055% digitonin for 20 min at 4°C. The cells were then subjected toa direct binding assay with 1251-ASOR. The binding data for surface(-) and internal (--) receptor sites are shown for both untreated (+)and EDTA-treated (o) cells. The experimental data are indicated bythe symbols, and the LIGAND-fitted data, by the lines.
10-20 pug/ml to inhibit 50% of the control level of 125I-ASORbinding (Fig. 3). There was no significant difference betweenthe behavior of active and heat-denatured collagenase in theinhibition assay. In addition, the binding of the collagenaseinhibitor(s) to the hepatocyte surface during collagenaseperfusion of the liver appeared to be prevented by theinclusion of 10 mM GalNAc in this step, as the resultinghepatocyte preparation exhibited an enhanced level of 1251_ASOR receptor sites.The apparent involvement of a carbohydrate-containing
inhibitor(s) was substantiated by the presence of galactose,glucose, and mannose in collagenase as determined byautomated sugar chromatography (31). When the liver wasperfused with collagenase that had been dialyzed, the result-ing hepatocyte preparation still exhibited receptor enhance-ment, which suggests that the collagenase inhibitor(s) is of amolecular mass > 10 kDa. Since neuraminidase was recentlyreported to be present in collagenase preparations (32), it wasnecessary to rule out the possibility that receptor enhance-ment was the result of the displacement of desialylatedmoieties from Gal/GalNAc receptors on freshly isolatedhepatocytes. There was no detectable neuraminidase activityin the collagenase preparations, and it was not possible toreverse the decrease in receptor sites for 125I-ASOR observedwith neuraminidase-treated hepatocytes by exposure toGalNAc, which is in accordance with previous findings (33).
Reversibility of Collagenase-Induced Decrease in ReceptorSite Number. The surface receptor site number for 1251I-ASORon freshly isolated rat hepatocytes has been shown to besensitive to additional exposure to collagenase at 370C (10).In Table 1, collagenase-induced decrease in surface receptorsite number was shown to be reversible upon treatment withGalNAc or EDTA at 4°C or after warming at 370C. Thesurface receptor site number was first increased on freshlyisolated hepatocytes by treatment with GalNAc (Table 1,experiment I) or by warming (experiment II). The cells werewashed and treated with active collagenase at 0.5 mg/ml for15 min either at 4°C (experiment I) or at 37°C (experiment II).The collagenase treatment at both temperatures resulted in40-50% reduction in the number of surface receptor sites asmeasured by the binding of 1251I-ASOR. When the collagen-ase-treated cells were washed and subsequently treated withGalNAc or EDTA in experiment I, the original level ofreceptor sites seen prior to the collagenase treatment was
-
00
$
-4e
75
Ax ciiOX 0
* +0
+ Kan z
25 OX
4K
0 , I &-5 -4 -3 -2
Collagenase, log mg/ml-1 U
FIG. 3. Inhibition of 125I-ASOR binding by collagenase to rathepatocytes at 2°C. Freshly isolated hepatocytes were first treatedwith 10 mM EGTA for 15 min at 4°C, washed, and then incubated (2x 106 cells per ml) with 0.125 nM 1251-ASOR in the presence ofvarious concentrations of collagenase for 2 hr at 2°C. The prepara-tions tested were Cooper lots 44N6783 (+), 44C5650 (o), and44B5406 (O) and Sigma lots 113F6829 (o), 13F6831 (x), and 93F0603(A).
Cell Biology: Stults and Lee
A
Dow
nloa
ded
by g
uest
on
Nov
embe
r 4,
202
1

Proc. Natl. Acad. Sci. USA 83 (1986)
Table 1. Reversibility of collagenase-induced decrease in"'I-ASOR binding to rat hepatocytes
Temperature, % of controlTreatment °C binding
Experiment IGalNAc-treated hepatocytes 4 100+ Collagenase 4 51+ GalNAc 4 116+ EDTA 4 119+ Mannose 4 64
Experiment IIWarmed hepatocytes 37 100+ Collagenase 37 40+ GalNAc 4 90+ EDTA 4 84+ Warming 37 77
Freshly isolated rat hepatocytes were either incubated with 10 mMGalNAc (I) for 30 min at 40C or warmed (II) for 45 min at 370C toincrease the number of surface receptor sites. After the cells werewashed, they were exposed to active collagenase at 0.5 mg/ml for 15min either at 40C (experiment I) or at 370C (experiment II). The cellswere then washed free of collagenase and subsequently treated with10mM GalNAc, 10mM EDTA, or 10 mM mannose at 40C or warmedfor 45 min at 37TC. After each treatment, an aliquot of cells wasincubated with 100 nM 125I-ASOR for 2 hr at 20C.
restored, whereas treatment with mannose was not effective.In experiment II, in addition to GalNAc and EDTA treat-ment, warming recovered most of the original receptoractivity. Exposure of freshly isolated rat hepatocytes toheat-denatured collagenase yielded similar results.
Receptor Enhancement Does Not Occur in the IsolatedPerfused Liver. By using the protocol outlined in Table 2, theisolated perfused liver was used to address the question ofwhether receptor enhancement is an artifact of the collagen-ase perfusion method for the preparation of isolatedhepatocytes. 125I-ASOR was bound by and removed from theliver in four consecutive cycles at 0-2°C. Perfusion ofEDTAthrough the liver after the first binding cycle with 65 nM1251-ASOR (Table 2, step A) did not cause a significantincrease in the amount of ligand bound during the secondbinding cycle (step B). However, perfusion of -40 mg ofactive collagenase through the liver prior to the third bindingcycle reduced the amount of 125I-ASOR bound by about 40%(step C). After subsequent EDTA perfusion, there wasrecovery ofthe original level ofligand bound (step D). Similarresults were obtained using 1251-labeled Gal44-AI-BSA orheat-denatured collagenase, or both.
Table 2. Effect of EDTA and collagenase on 251I-ASOR bindingin the isolated perfused liver at 0-20C
Binding 125I-ASOR bound,Step cycle Treatment pmol/g of liver
A 1 64B 2 EDTA 70C 3 Collagenase 41D 4 EDTA 73
A 6.1 g rat liver was prepared for perfusion at 0-20C. In the firstbinding cycle (step A), 65 nM 1"I-ASOR was perfused through theliver for 30 min. After unbound ligand was washed out, the amountof specifically bound '25I-ASOR was determined by perfusion of 10mM EDTA through the liver for 5 min as described. After replace-ment of calcium to the liver by perfusion with buffer, a secondbinding cycle (step B) was carried out as described for step A.Approximately 40 mg of active collagenase was then perfusedthrough the liver. After washout of the collagenase by perfusion withbuffer, two additional binding cycles (C and D) were performed. Thelevel of nonspecific binding in this experiment was estimated to be-15%.
DISCUSSION
Our results indicate that collagenase preparations contain ahigh-affinity ligand(s) that binds to the Gal/GalNAc receptorduring the hepatocyte-isolation procedure and that it may belargely responsible for the modulation of receptor activityobserved when freshly isolated hepatocytes are warmed orexposed to GalNAc or EDTA. This inhibitor was readilydissociated from the receptor, resulting in a 2- to 6-foldincrease in receptor sites for the neoglycoprotein Ga44-AI-BSA, the synthetic cluster ligand di-tris-lac, and the glyco-protein ASOR. It also appeared to be cleared from surfacereceptor sites upon incubation at 370C, which resulted in asimilar increase in receptor number for the same ligands. Itwas also possible to demonstrate the release of inhibitorymaterial from freshly isolated hepatocytes by treatment withEDTA or GalNAc at 40C (data not shown). The size of theinternal pool of receptors as assessed by digitonin perme-abilization of hepatocytes did not change upon EDTA treat-ment or warming, which suggests that only the surface poolof receptor participates in the phenomenon described here.
Perfusion of EDTA through the isolated whole liver wasnot accompanied by a severalfold increase in receptor bind-ing activity, which suggests that endogenous ligand-that is,ligand normally associated with the receptor in vivo-if itindeed exists, does not appear to play a major role in theenhancement of surface receptor activity seen with isolatedhepatocytes. As a potential source of exogenous ligand,collagenase preparations were shown to interact directly withthe Gal/GalNAc receptor by their inhibitory effect on thebinding of 125I-ASOR to hepatocytes. In addition, exposureof isolated hepatocytes and the whole liver to collagenaseresulted in a significant reduction in surface receptor activity,which was reversible by the same treatments that causereceptor enhancement. The effect of collagenase on theGal/GalNAc receptor was shown to be primarily nonenzy-matic, since both active and heat-denatured collagenaseyielded similar results.EDTA was generally more effective than GalNAc at the
same concentration in the removal of the collagenase inhib-itor(s) as evidenced by the exposure of slightly more sites (seeFig. 1). This result was not unreasonable, given our repeatedobservation that the release of prebound macromolecularligands from the surface of hepatocytes by 10 mM GalNAc isonly 80-90% complete in 30 min, whereas 10 mM EDTAeffects their release very rapidly (within 15 s). Warming alsoappeared to reveal fewer receptor sites compared to EDTAtreatment, which is probably related to the report that only70-90% of a prebound ligand (ASOR) is internalized by rathepatocytes when incubated at 37°C (13). With regard to theefficacy of EDTA in the exposure of more binding sites, thisagent has been proposed to expose unoccupied yet crypticsurface receptors on freshly isolated rat hepatocytes thatwere previously in a nonfunctional state at some stage in therecycling process (11, 13). Removal of the collagenaseinhibitor from the hepatocyte surface by these varioustreatments appeared not only to affect the receptor sitenumber but also to alter the affinity constants (within a rangeof about 5-fold) for the different ligands. This is not surprisingbecause partial saturation ofthe Gal/GalNAc receptor on thesurface of rabbit hepatocytes with a macromolecular ligand(ASOR) affects the binding ofgalactose-terminated ligands tothe remaining accessible receptors in a specific manner (25).
It has been shown that the Gal/GalNAc surface receptoractivity on freshly isolated hepatocytes can be reversiblymodulated by changes in temperature, which has beenproposed to represent an energy-dependent externalizationand/or activation of receptor that was previously unable tobind ligand (3, 11). For this reason and the potential for theendocytosis of the bound collagenase inhibitor, the previous
7778 Cell Biology: Stults and Lee
Dow
nloa
ded
by g
uest
on
Nov
embe
r 4,
202
1

Proc. Natl. Acad. Sci. USA 83 (1986) 7779
thermal history of freshly isolated hepatocytes would beexpected to affect variously the magnitude of the increase insurface receptor activity associated with warming at 370C ortreating with GalNAc or EDTA. Seglen's procedure for thepreparation of isolated hepatocytes recommends that, afterthe collagenase perfusion step at 37TC, the isolation andpurification of the cells be carried out at 0-40C (6), which wehave adhered to strictly. However, if the liver cell prepara-tion is handled at temperatures above 10'C but below 370C,endocytosis of the bound collagenase inhibitor as well asdown-modulation of surface receptor activity (11) may takeplace. In addition, the amount of collagenase inhibitor asso-ciated with freshly isolated hepatocytes would be dependenton the time, dose, and preparation of collagenase used for theperfusion. These factors would contribute also to variabilityin the fold activation of surface receptor sites by the differenttreatments.Our data indicate that the size of the pools of surface
receptor that are exposed by warming and by treatment withGalNAc or EDTA are comparable. In contrast, Weigel andcoworkers reported that the number of new receptor sitesexposed upon warming at 370C was 3-fold greater than thatobtained after EDTA treatment at 40C (11). They believe thisdiscrepancy to be attributable to the fact that they carry outthe isolation and purification of their cells at room temper-ature or above and that any collagenase inhibitor bound onthe surface of their cells would be internalized during thistime (P. H. Weigel, personal communication). Thus, treat-ment of their hepatocyte preparation with EDTA wouldexpose the EDTA cryptic pool of receptor, while warming at370C would increase the number of surface receptor sites thathad undergone down-modulation during the period when thecells were at room temperature (11). In fact, we were able todemonstrate a decrease in surface receptor activity whenhepatocytes whose surface had been freed of the collagenaseinhibitor were incubated at room temperature.
Nevertheless, based on the results presented here, thepossibility must be considered that the increase in surfacereceptor number observed when freshly isolated hepatocytesare warmed or treated with EDTA or GalNAc represents theclearance of collagenase inhibitor. The potential presence ofan exogenous ligand on surface and internal Gal/GalNAcreceptor sites also complicates the unresolved issue ofwhether or not receptor recycling in this system is constitu-tive. It is possible that the collagenase inhibitor(s) may haveprovided the signal for internalization and recycling of theGal/GalNAc receptor. In summary, experiments that dealwith the dynamics of the Gal/GalNAc receptor on freshlyisolated hepatocytes prepared by the collagenase perfusionmethod must be reevaluated in view of the evidence that avariable and significant proportion of their receptor sites maybe occupied by an exogenously derived ligand. Moreover,these findings are probably not unique to the Gal/GalNAcreceptor and may be extended to other receptor systemsstudied using cells prepared with crude collagenase prepara-tions.
The authors thank Dr. R. Reid Townsend for helpful suggestionsthroughout the course of this work. We are grateful to Dennis Redderfor preparation of the hepatocytes and perfused liver and to Dr.Reiko T. Lee for providing the di-tris-lac. We also appreciate theexpert technical assistance of T. Chai, G. Lee, and M. Hardy in someof the binding experiments. This work was supported by research
grants from the National Institutes of Health (AM9970 and CA21901)and the National Science Foundation (DCB-8509638). This is con-tribution 1331 from The McCollum Pratt Institute, The JohnsHopkins University, Baltimore, MD 21218.
1. Stockert, R. J. (1983) in Receptor-Mediated Endocytosis, eds.Cuatrecasas, P. & Roth, T. (Chapman & Hall, New York), pp.167-185.
2. Harford, J. & Ashwell, G. (1985) in Endocytosis, eds. Pastan,I. & Willingham, M. C. (Plenum, New York), pp. 69-81.
3. Weigel, P. H. (1985) in Vertebrate Lectins, eds. Parent, J. B.& Olden, K. (Van Nostrand Reinhold, New York), in press.
4. Breitfeld, P. P., Simmons, C. F., Jr., Strous, G., Geuze, H. J.& Schwartz, A. L. (1985) Int. Rev. Cytol. 97, 47-95.
5. Dunn, W. A., Wall, D. A. & Hubbard, A. L. (1980) MethodsEnzymol. 98, 225-240.
6. Seglen, P. 0. (1976) Methods Cell Biol. 13, 29-83.7. Schwartz, A. L., Rup, D. & Lodish, H. F. (1980) J. Biol.
Chem. 225, 9033-9036.8. Steer, C. J. & Ashwell, G. (1980) J. Biol. Chem. 255,
3008-3013.9. Weigel, P. H. (1980) J. Biol. Chem. 255, 6111-6120.
10. Zeitlen, P. L. & Hubbard, A. L. (1982) J. Cell. Biol. 92,634-647.
11. Weigel, P. H. & Oka, J. A. (1983) J. Biol. Chem. 258,5089-5094.
12. Fiete, D., Brownell, M. D. & Baenziger, J. U. (1983) J. Biol.Chem. 258, 817-823.
13. Weigel, P. H. & Oka, J. A. (1982) J. Biol. Chem. 257,1201-1207.
14. Myers, R. W., Lee, R. T., Lee, Y. C., Thomas, G. H.,Reynolds, L. W. & Uchida, Y. (1980) Anal. Biochem. 101,166-174.
15. Stults, N. L., Lin, P., Hardy, M., Lee, Y. C., Uchida, Y.,Tsukada, Y. & Sugimori, T. (1983) Anal. Biochem. 135,392-400.
16. Lee, Y. C., Stowell, C. P. & Krantz, M. J. (1976) Biochemis-try 15, 3956-3963.
17. Lee, R. T., Lin, P. & Lee, Y. C. (1984) Biochemistry 23,4255-4261.
18. Schnaar, R. L., Weigel, P. H., Kuhlenschmidt, M. S., Lee,Y. C. & Roseman, S. (1978) J. Biol. Chem. 253, 7940-7951.
19. Zamenhof, S. (1957) Methods Enzymol. 3, 696-704.20. Patterson, M. K., Jr. (1979) Methods Enzymol. 58, 141-152.21. Millipore (1979) Worthington Enzymes Manual (Millipore,
Bedford, MA).22. Uchida, Y., Tsukada, Y. & Sugimori, T. (1977) J. Biochem.
(Tokyo) 82, 1425-1433.23. Townsend, R. R., Hardy, M. R., Wong, T. C. & Lee, Y. C.
(1986) Biochemistry, in press.24. Greenwood, F. C., Hunter, W. M. & Glover, J. S. (1%3)
Biochem. J. 89, 114-123.25. Hardy, M. R., Townsend, R. R., Parkhurst, S. M. & Lee,
Y. C. (1985) Biochemistry 24, 22-28.26. Munson, P. J. & Rodbard, D. (1980) Anal. Biochem. 107,
220-239.27. Lee, Y. C., Townsend, R. R., Hardy, M. R., Lonngren, J.,
Arnaap, J., Haraldsson, M. & Lonn, H. (1983) J. Biol. Chem.258, 199-202.
28. Weigel, P. H., Ray, D. A. & Oka, J. A. (1983) Anal. Biochem.113, 437-449.
29. Klotz, I. (1982) Science 217, 1247-1249.30. Scatchard, G. (1949) Ann. N. Y. Acad. Sci. 51, 660-672.31. Lee, Y. C. (1972) Methods Enzymol. 28, 68-73.32. Hatton, M. W. C., Berry, L. R., Krestynski, F., Sweeney,
G. D. & Regoeczi, E. (1983) Eur. J. Biochem. 137, 311-318.33. Stockert, R. J., Howard, D. J., Morell, A. G. & Scheinberg,
I. H. (1980) J. Biol. Chem. 255, 9028-9029.
Cell Biology: Stults and Lee
Dow
nloa
ded
by g
uest
on
Nov
embe
r 4,
202
1





























