Embed Size (px)
Citation preview

of November 27, 2018.This information is current as
Invasive Pulmonary AspergillosisNeutrophil-Mediated Host Defense inNecessary Components of CXC Chemokine Receptor-2 Ligands Are
C. Tsai, Sergio A. Lira and Theodore J. StandifordBorna Mehrad, Robert M. Strieter, Thomas A. Moore, Wan
http://www.jimmunol.org/content/163/11/60861999; 163:6086-6094; ;J Immunol
Referenceshttp://www.jimmunol.org/content/163/11/6086.full#ref-list-1
, 21 of which you can access for free at: cites 54 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 1999 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on N
ovember 27, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

CXC Chemokine Receptor-2 Ligands Are NecessaryComponents of Neutrophil-Mediated Host Defense in InvasivePulmonary Aspergillosis1
Borna Mehrad*, Robert M. Strieter,* Thomas A. Moore,* Wan C. Tsai,* Sergio A. Lira †, andTheodore J. Standiford2*
Invasive pulmonary aspergillosis is a devastating complication of immunosuppression, which occurs in association with neutrophildysfunction or deficiency. ELR1 CXC chemokines are a subfamily of chemokines that play a critical role in neutrophil chemotaxisand activation both in vitro and in vivo. We hypothesized that interaction of these ligands with CXC chemokine receptor-2(CXCR2), their sole murine receptor, is a major component of neutrophil-dependent pulmonary host defense againstAspergillusfumigatus. In immunocompetent animals, neutrophils were recruited to the lung in response to intratracheally administeredA.fumigatus conidia. In a model of transient in vivo depletion of neutrophils, animals developed invasive pulmonary aspergillosis,associated with delayed influx of neutrophils into the lung. In both normal and neutrophil-depleted animals, the ELR1 CXCchemokines MIP-2 and KC were induced in response to intratracheal administration of conidia. Ab-mediated neutralization of thecommon ELR1 CXC chemokine receptor, CXCR2, resulted in development of invasive disease indistinguishable from the diseasein neutrophil-depleted animals, while control animals were highly resistant to the development of infection. CXCR2 neutralizationwas associated with reduced lung neutrophil influx and resulted in a marked increase in mortality compared with controls. Incontrast, animals with constitutive lung-specific transgenic expression of KC were resistant to the organism, with reduced mor-tality and lower lung burden of fungus. We conclude that CXCR2 ligands are essential mediators of host defense againstA.fumigatus, and may be important targets in devising future therapeutic strategies in this disease.The Journal of Immunology,1999, 163: 6086–6094.
I nvasive pulmonary aspergillosis is a common and devastat-ing complication of immunosuppression. Evidence suggestsan alarming recent increase in the incidence of this disease,
which has been attributed to the increasing prevalence of immu-nocompromised patients (1–3). Current therapy for this infection islimited by a poor rate of response (4, 5), such that the mortality ofinvasive pulmonary aspergillosis is.85% despite best availabletreatment (6).
Invasive aspergillosis has been noted to occur in the setting ofagranulocytosis since its original description (7), and deficienciesin neutrophil number or function have since been recognized to bestrongly associated with the development of invasive aspergillosisin diverse populations of patients (8–11). In vitro studies haveshown that normal neutrophils efficiently damageAspergillushy-phae (12, 13). In contrast, dysfunctional neutrophils, such as thosetreated with corticosteroids or those isolated from patients with
myeloperoxidase (MPO)3 deficiency or chronic granulomatousdisease are inefficient at killingAspergillushyphae (14, 15). Invivo animal studies have established a causative relationship be-tween neutrophil dysfunction or deficiency and the development ofinvasive aspergillosis (16, 17). However, factors that regulate neu-trophil recruitment and activation in invasive aspergillosis havenot been characterized.
CXC chemokines are a family of related 8- to 10-kDa polypep-tide molecules that are potent chemotactic factors for numerouspopulations of immune cells and are characterized by four con-served cysteine residues at the N terminus, the first two of whichare separated by a nonconserved amino acid. The CXC chemo-kines are further subdivided into those that contain the amino acidsequence glutamic acid-leucine-arginine (the ELR motif) immedi-ately preceding the CXC sequence, and those that do not. Thehuman ELR1 CXC chemokines include IL-8, the growth-relatedoncogene (GRO) chemokines, epithelial neutrophil-activating pro-tein-78 (ENA-78), neutrophil-activating peptide-2 (NAP-2), andgranulocyte chemotactic peptide-2 (GCP-2). Murine ELR1 CXCchemokines include macrophage inflammatory protein-2 (MIP-2),KC, GCP-2, LPS-induced CXC chemokine (LIX), and Lungkine(18, 19). ELR1 CXC chemokines have been shown to mediateneutrophil chemotaxis and activation in vitro (20–24) and in di-verse in vivo settings (25–27). Several ELR1 CXC chemokineshave been shown to be expressed in murine models of bacterialpneumonia (28), but the neutralization of specific chemokines hasnot altered survival dramatically (29).
Two human ELR1 CXC chemokine receptors have been iden-tified, and are designated CXC chemokine receptor-1 (CXCR1)and CXC chemokine receptor-2 (CXCR2). These receptors are ex-clusively expressed on the surface of human granulocytes, and are
*Department of Medicine, Division of Pulmonary and Critical Care Medicine, Uni-versity of Michigan Medical School, Ann Arbor, MI 48109; and†Department ofImmunology, Schering-Plough Research Institute, Kenilworth, NJ 07033
Received for publication July 20, 1999. Accepted for publication September 15, 1999.1 This work was supported in part by National Institutes of Health Grants1K08HL04220-01, HL57243, HL58200, and P50HL60289.2 Address correspondence and reprint requests to Dr. Theodore J. Standiford, Uni-versity of Michigan Medical Center, 6301 MSRB III, 1150 West Medical CenterDrive, Ann Arbor, MI 48109-0360. E-mail address: [email protected] Abbreviations used in this paper: MPO, myeloperoxidase; CCR1, CC chemokinereceptor-1; CXCR2, CXC chemokine receptor-2; GMS, Gomori methanamine silver;ELR, glutamic acid-leucine-arginine; ENA-78, epithelial neutrophil activating pro-tein-78; GCP-2, granulocyte chemotactic protein-2; GRO, growth-related oncogene;H&E, hematoxylin and eosin; i.t., intratracheal; LIX, LPS-induced CXC chemokine;MIP-1a; macrophage inflammatory protein-1a; MIP-2, macrophage inflammatoryprotein-2; NAP-2, neutrophil activating peptide-2; BAL, bronchoalveolar lavage;GMS, Gomori methanamine silver.
Copyright © 1999 by The American Association of Immunologists 0022-1767/99/$02.00
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

involved in neutrophil chemotactic and degranulation responses(30–32). CXCR1 binds IL-8 and GCP-2, while CXCR2 binds allELR1 CXC chemokines with equal avidity (33, 34). The CXCR1is not expressed in the mouse, but the murine CXCR2 is highlyhomologous to human CXCR2. Like its human counterpart, mu-rine CXCR2 is promiscuous and has been shown to bind to severalmurine ELR1 CXC chemokines. Neutrophils from mice with tar-geted deletion of CXCR2 fail to respond to macrophage inflam-matory protein-2 (MIP-2) or KC in vitro, but exhibit normal che-motaxis to C5a (35). In in vivo studies, CXCR2 knockout mice failto develop tissue neutrophilic infiltrates in response to s.c. uratecrystals (36), suggesting that ELR1 CXC chemokines are the ma-jor mediator of neutrophil influx in this model, and that CXCR2 isthe sole receptor for these ligands. The role of CXCR2 in thesetting of infections has not been examined. The presence of mul-tiple ligands and the promiscuity of the receptors have made thestudy of human ELR1 CXC chemokines difficult. In this context,the mouse system, which lacks CXCR1, provides a simplified sys-tem for studying the ligand-receptor interaction of ELR1 CXCchemokines.
In this study, we hypothesized that the interaction of ligandswith CXCR2 is a major component of neutrophil-dependent pul-monary host defense againstA. fumigatusby mediating neutrophildeployment in response to the organism. To test this hypothesis,we evaluated the outcome and severity of infection in the setting ofCXCR2 neutralization, and in the setting of compartmentalizedoverexpression of KC in the lung.
Materials and MethodsReagents
Polyclonal antimurine MIP-2 and KC Abs used in the ELISAs were pro-duced by immunization of rabbits with carrier-free murine rt MIP-2 andKC (R&D Systems, Minneapolis, MN) in multiple intradermal sites withCFA, as previously described (37, 38). Polyclonal goat antimurine CXCR2Abs used in in vivo neutralization studies were produced by immunizationof a goat with murine rt CXCR2 peptide in multiple intradermal sites withCFA. The peptide sequence, Met-Gly-Glu-Phe-Lys-Val-Asp-Lys-Phe-Asn-Ile-Glu-Asp-Phe-Phe-Ser-Gly, is a portion of the seven transmem-brane receptor that resides on the cell surface and has previously beenshown to be the binding site for ligands (39). Abs were purified over aprotein A column and endotoxin contamination excluded byLimuluslysateassay (ICN Biomedical, Costa Mesa, CA). In CXCR2 neutralization ex-periments, 0.5 ml goat antimurine CXCR2 serum, or control goat serum,was administered i.p. 2 h prior toA. fumigatusadministration. In prelim-inary studies, administration of this Ab did not affect the number of cir-culating neutrophils over the subsequent 4 days (data not shown). In sep-arate experiments, administration of anti-CXCR2 abrogated the influx ofneutrophils into the peritoneum of normal mice in response to exogenousKC (data not shown).
Animals
Specific pathogen-free C57BL/6 mice (6 to 8-wk-old females, The JacksonLaboratories, Bar Harbor, ME) were used in all experiments except thoseinvolving KC transgenic animals. Specific pathogen-free transgenic KCmice CC511/1 (6 to 8-wk-old females) were generated on a B6D2 back-ground by microinjection of fertilized eggs carrying the KC transgene intheir genome. The expression of the KC transgene was localized to the lungby incorporating regulating elements of the CC10 gene, which encodes a10-kDa protein produced by nonciliated bronchial epithelial cells (Claracells) (40–42). Age-matched wild-type B6D2 mice served as controls. Allmice were housed in specific pathogen-free conditions within the animalcare facility at the University of Michigan until the day of sacrifice.
Preparation and use of RB6-8C5 mAb for in vivo neutrophildepletion
RB6-8C5 is a rat anti-mouse monoclonal IgG2b directed against Ly-6G,previously known as Gr-1, an Ag on the surface of murine granulocytes.The Ag expression increases with cell maturity, and is absent from pre-cursor cells. The Ab, originally used for flow cytometry, is a complement-fixing isotype, well suited for in vivo cell depletion (43–47). The Ab pro-
duced by TSD BioServices (Germantown, NY) by i.p. injection of RB6-8C5 hybridoma into nude mice and ascites collection. One hundredmicrograms of RB6-8C5 was administered i.p. 1 day prior to challengewith intratracheal (i.t.)A. fumigatusor vehicle. This resulted in peripheralblood neutropenia (absolute circulating neutrophil count,50 cells/ml) bydays 1 and 3 after Ab administration in both infected and control animals,with a return of peripheral counts to pretreatment levels by day 5.
Preparation and administration ofA. fumigatusconidia
We chose to useA. fumigatusstrain 13073 (American Type Culture Col-lection, Manassas, VA) in our studies, as this strain has previously beenshown to induce invasive aspergillosis in immunocompromised mice (48).The organism was grown on Sabouraud dextrose agar plates (Becton Dick-inson, Cockeyville, MD) for 7 to 10 days at 37°C. The surface of each platewas then washed with 100 ml of sterile 0.1% Tween 80 (SigmaUltra, St.Louis, MO) in normal saline. The resulting suspension of conidia wasfiltered through sterile gauze to remove clumps and hyphal debris, and thenwashed once and resuspended in 4 ml of 0.1% Tween-80. The concentra-tion of Aspergillusconidia in the suspension was determined by a particlecounter (Z2 particle analyzer, Coulter, Hialeah, FL). The suspension wasthen diluted to the desired concentration, and the concentration was againmeasured prior to administration. In preliminary experiments, the numberof particles determined by the particle counter was in close agreement withthe number of viable CFUs found by serial dilution and plating of thesuspension. On the day of inoculation, each animal was anesthetized with1.8–2 mg pentobarbital i.p. Using standard aseptic technique, the tracheawas exposed and a 30-ml inoculum (A. fumigatussuspension or 0.1%Tween-80) was administered via a sterile 26-gauge needle. The skin inci-sion was closed with surgical staples. Animals were challenged with in-ocula ranging from 1–23 106 to 1–2 3 107 conidia in variousexperiments.
Lung harvest
At designated time points, the mice were sacrificed by CO2 asphyxiation.The chest cavity was opened aseptically, and the pulmonary vasculaturewas perfused with PBS via the right ventricle. For histologic examination,lungs were perfused with 1 ml 4% paraformaldehyde in PBS, inflated with1 ml 4% paraformaldehyde in PBS via the trachea, and then excised enbloc. Lungs for various assays were perfused with 1 ml of PBS containing5 mM EDTA, removed, frozen in liquid nitrogen, and stored at220°Cuntil the day of the assay. Lungs for cytokine and MPO assays were ho-mogenized in 1 ml of 23 complete protease inhibitor cocktail buffer(Boehringer Mannheim, Mannheim, Germany) in PBS, using a tissue ho-mogenizer (Biospec Products, Bartlesville, OK). A 900-ml aliquot of PBSwas added to 900ml from each sample, sonicated for 10 s, and centrifugedat 5003 g for 10 min. Supernatants were passed through a 0.45-micronfilter (Gelman Sciences, Ann Arbor, MI), and stored at 4°C for cytokineELISA.
Lung chitin assay
Given that molds (includingAspergillusspecies) do not reliably form re-productive units in tissue, we employed an assay for chitin to measure theburden of organisms in lungs. Chitin is a component of the hyphal wall thatis absent from mammalian cells and conidia. The assay was adapted froma previously described method, which demonstrated a direct correlationbetween the weight of hyphae and level of chitin, as detected by assay (49).Lungs were homogenized in 5 ml distilled water and centrifuged (15003g, 5 min, 20°C). The supernatants were discarded, pellets resuspended insodium lauryl sulfate (3% w/v), and heated at 100°C for 15 min. Sampleswere then centrifuged (15003 g, 5 min, 20°C), pellets washed with dis-tilled water, and resuspended in 3 ml KOH (120% w/v). Samples were thenheated at 130°C for 60 min. After cooling, 8 ml of ice-cold ethanol (75%v/v) was added to each sample, and tubes were shaken until ethanol andKOH made one phase. Samples were incubated on ice for 15 min, and 0.3ml of Celite suspension (supernatant of 1 g of Celite 545 (Fisher Scientific,Pittsburgh, PA) added to 75% ethanol and allowed to stand for 2 min) wasadded to each. Samples were centrifuged (15003 g, 5 min, 4°C), andsupernatants were discarded. Pellets were washed once with ethanol (40%v/v) and twice with distilled water, and resuspended in 0.5 ml distilledwater. Standards, consisting of 0.2 ml distilled water and 0.2 ml glu-cosamine (10mg/ml), were made up. A total of 0.2 ml NaNO2 (5% w/v)and 0.2 ml KHS04 (5% w/v) was added to each standard, and 0.5 mlNaNO2 (5% w/v) and 0.5 ml KHS04 (5% w/v) was added to each tissuepreparation; all samples were mixed gently for 15 min, and then centri-fuged (15003 g, 2 min, 4°C). Two 0.6-ml aliquots of supernatant fromeach tissue preparation were transferred to separate tubes. A total of 0.2 mlammonium sulfamate was added to each tube, and all tubes were shaken
6087The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

vigorously for 5 min. A fresh solution of 3-methyl-2-thiazolone hydrazoneHCl monohydrate (50 mg in 10 ml distilled water) was made, and 0.2 mlwas added to each tube. Samples were then heated to 100°C for 3 min andcooled. A total of 0.2 ml of FeCl3 . 6H2O (0.83% w/v) was added to each,and OD was measured at 650 nm after 25 min. Chitin content, measured inglucosamine equivalents, was measured by the following formula: chitincontent5 {[5 3 (OD of organ2 OD of control organ)]/(OD of glu-cosamine2 OD of water)}.
Lung MPO activity
Lung MPO activity was measured as a marker of neutrophil sequestration,as described previously (50). Briefly, a 100-ml aliquot of each lung ho-mogenate was added to 100ml of a buffer containing 50 mM potassiumphosphate (pH 6.0), 5% hexadecyltrimethylammonium bromide, and 5mM EDTA. Samples were sonicated for 10 s and centrifuged at 30003 gfor 15 min. The supernatant was mixed 1:15 with a buffer containing 1 Mmonobasic potassium phosphate, 1 M dibasic potassium phosphate, 3%H202, and o-dianisidine hydrochloride, and read at 490 nm. MPO unitswere calculated as the change in absorbency over time.
Chemokine ELISA
Murine MIP-2 and KC levels were quantified using a modification of adouble-ligand method, as described previously (51). Briefly, flat-bottom96-well microtiter plates (Immuno-Plate I 96-F, Nunc Roskilde, Denmark)were coated with 50ml/well of rabbit anti-cytokine Ab (1mg/ml in 0.6 MNaCl, 0.26 M H3BO4, and 0.08 N NaOH, pH 9.6) for 16 h at 4°C and thenwashed with PBS, pH 7.5, 0.05% Tween-20 (wash buffer). Microtiter platenonspecific-binding sites were blocked with 2% BSA in PBS and incubatedfor 90 min at 37°C. Plates were rinsed four times with wash buffer, anddiluted (neat and 1:10) cellfree supernatants (50ml) in duplicate wereadded, followed by incubation for 1 h at 37°C. Plates were washed fourtimes, followed by the addition of 50ml/well biotinylated rabbit amti-cytokine Abs (3.5mg/ml in PBS, pH 7.5, 0.05% Tween-20, and 2% FCS),and plates incubated for 30 min at 37°C. Plates were washed four times,Streptavidin-peroxidase conjugate (Bio-Rad, Richmond, CA) was added,and the plates were incubated for 30 min at 37°C. Plates were washed againfour times and chromogen substrate (Bio-Rad) was added. The plates wereincubated at room temperature to the desired extinction, and the reactionwas terminated with 50ml/well of 3M H2SO4 solution. Plates were read at490 nm in an ELISA reader. Standards were1⁄2 log dilutions of recombi-nant murine MIP-2 or KC, from 1 pg/ml to 100 ng/ml. This ELISA methodconsistently detected the relevant cytokine at concentrations above 25 pg/ml. The ELISAs did not cross-react with IL-1, IL-2, IL-4, IL-6, IL-10 orIFN-g. In addition, each ELISA did not cross-react with other members ofthe murine CXC or CC chemokine families.
Statistical analysis
Data were analyzed by a Power Macintosh 8600/300 computer using InStatversion 2.01 statistical package (GraphPad Software, San Digeo, CA). Sur-vival data were compared using the Fisher’s exact test. All other data wereexpressed as mean6 SEM and compared using an unpaired two-tailMann-Whitney (nonparametric) test. Probability values were consideredstatistically significant if they were less than 0.05.
ResultsInflux of neutrophils into the lungs in response toA. fumigatus
We first determined the time course of neutrophil influx into thelungs of normal C57BL/6 mice in response to challenge with1–23 107 A. fumigatusconidia. Lungs were harvested at varioustime points after challenge for histology and measurement of MPOactivity, a surrogate measure of neutrophil presence. Lung MPOlevels were elevated over baseline at 8 h after inoculation, andreached a plateau at 24 h (Fig. 1). Histology showed evidence ofa patchy peribronchial infiltration of inflammatory cells withinboth the interstitial and alveolar compartments at 24 h, which pre-dominantly consisted of neutrophils, and to a lesser extent, ofmononuclear cells (data not shown). The cellular infiltrate wasresolving by day 4 after inoculation, and predominantly consistedof mononuclear cells (data not shown).
Effect of neutrophil depletion on host response toA. fumigatus
In order to assess the contribution of neutrophils to host defenseagainstA. fumigatus, we next challenged neutrophil-depleted micewith i.t. conidia. Neutrophil depletion was achieved by i.p. admin-istration of 100mg of RB6-8C5 mAb i.p. 1 day before i.t.A.fumigatus. In preliminary studies, we found that administered ofthis dose of RB6-8C5 resulted in peripheral blood neutropenia (ab-solute circulating neutrophil count,50 cells/ml) by days 1 and 3after Ab administration in both infected and control animals, witha restoration of peripheral counts to pretreatment levels by day 5;administration of control serum did not affect the peripheral countsor lung histology (data not shown). We found a dose-dependentmortality in mice pretreated with RB6-8C5, depending on the sizeof A. fumigatusinoculum (Fig. 2). Importantly, pretreatment withRB6-8C5 leads to greater than 90% mortality in mice challenged
FIGURE 1. Lung MPO activity afterA. fumigatuschallenge in normalmice. Lung MPO activity was measured at various times afterA. fumigatuschallenge in normal mice (1–23 107 conidia). Af, A. fumigatus; MPO,myeloperoxidase. Experimentaln 5 6 at each time point.p, p , 0.05 ascompared with animals receiving vehicle. Data shown are representative ofthree separate experiments.
FIGURE 2. Effect of transient neutrophil depletion on survival afterA.fumigatuschallenge in normal mice. Mice were treated with i.p. saline orRB6-8C5 mAb 1 day prior to i.t. challenge withA. fumigatus(1.653 106
or 1.893 106 conidia). E6,3106; RB6-8C5, rat anti-mouse Ly-6G IgG2bmAb. Experimentaln 5 15 per group.p, p , 0.05 as compared withanimals receiving i.p. saline. Data pooled from two separate experiments.
6088 CXCR2 IN INVASIVE ASPERGILLOSIS
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
with 1.893 106 conidia, while no deaths occurred in control an-imals challenged with the same inoculum.
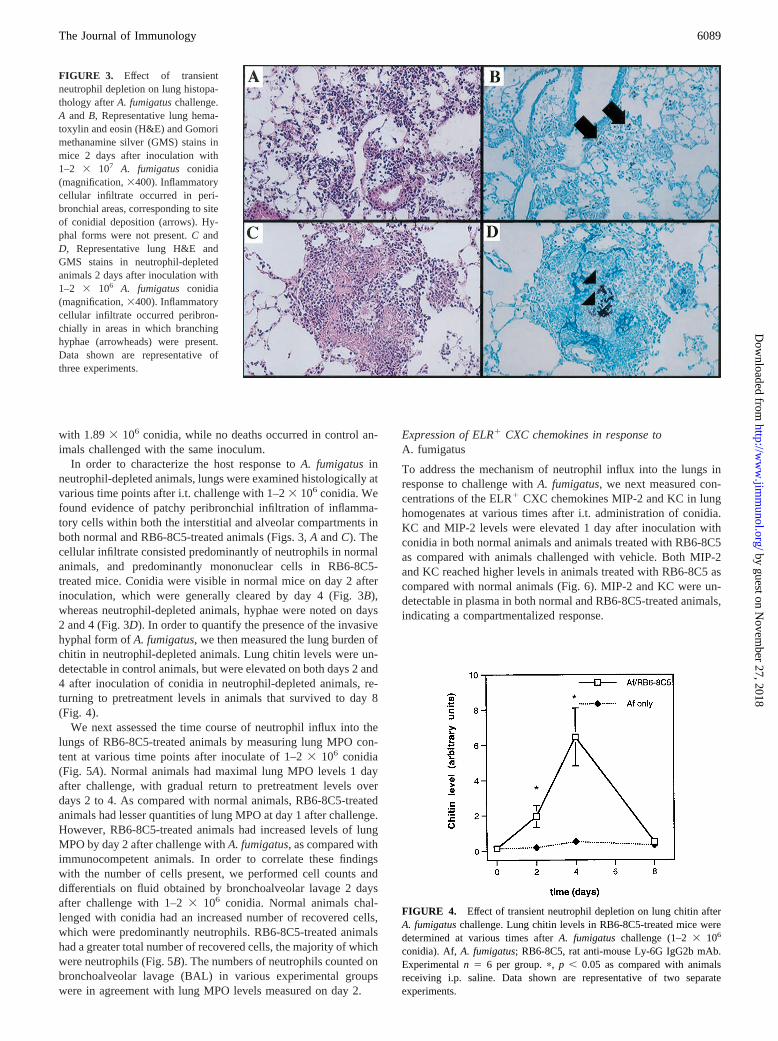
In order to characterize the host response toA. fumigatusinneutrophil-depleted animals, lungs were examined histologically atvarious time points after i.t. challenge with 1–23 106 conidia. Wefound evidence of patchy peribronchial infiltration of inflamma-tory cells within both the interstitial and alveolar compartments inboth normal and RB6-8C5-treated animals (Figs. 3,A andC). Thecellular infiltrate consisted predominantly of neutrophils in normalanimals, and predominantly mononuclear cells in RB6-8C5-treated mice. Conidia were visible in normal mice on day 2 afterinoculation, which were generally cleared by day 4 (Fig. 3B),whereas neutrophil-depleted animals, hyphae were noted on days2 and 4 (Fig. 3D). In order to quantify the presence of the invasivehyphal form ofA. fumigatus, we then measured the lung burden ofchitin in neutrophil-depleted animals. Lung chitin levels were un-detectable in control animals, but were elevated on both days 2 and4 after inoculation of conidia in neutrophil-depleted animals, re-turning to pretreatment levels in animals that survived to day 8(Fig. 4).
We next assessed the time course of neutrophil influx into thelungs of RB6-8C5-treated animals by measuring lung MPO con-tent at various time points after inoculate of 1–23 106 conidia(Fig. 5A). Normal animals had maximal lung MPO levels 1 dayafter challenge, with gradual return to pretreatment levels overdays 2 to 4. As compared with normal animals, RB6-8C5-treatedanimals had lesser quantities of lung MPO at day 1 after challenge.However, RB6-8C5-treated animals had increased levels of lungMPO by day 2 after challenge withA. fumigatus, as compared withimmunocompetent animals. In order to correlate these findingswith the number of cells present, we performed cell counts anddifferentials on fluid obtained by bronchoalveolar lavage 2 daysafter challenge with 1–23 106 conidia. Normal animals chal-lenged with conidia had an increased number of recovered cells,which were predominantly neutrophils. RB6-8C5-treated animalshad a greater total number of recovered cells, the majority of whichwere neutrophils (Fig. 5B). The numbers of neutrophils counted onbronchoalveolar lavage (BAL) in various experimental groupswere in agreement with lung MPO levels measured on day 2.
Expression of ELR1 CXC chemokines in response toA. fumigatus
To address the mechanism of neutrophil influx into the lungs inresponse to challenge withA. fumigatus, we next measured con-centrations of the ELR1 CXC chemokines MIP-2 and KC in lunghomogenates at various times after i.t. administration of conidia.KC and MIP-2 levels were elevated 1 day after inoculation withconidia in both normal animals and animals treated with RB6-8C5as compared with animals challenged with vehicle. Both MIP-2and KC reached higher levels in animals treated with RB6-8C5 ascompared with normal animals (Fig. 6). MIP-2 and KC were un-detectable in plasma in both normal and RB6-8C5-treated animals,indicating a compartmentalized response.
FIGURE 3. Effect of transientneutrophil depletion on lung histopa-thology afterA. fumigatuschallenge.A and B, Representative lung hema-toxylin and eosin (H&E) and Gomorimethanamine silver (GMS) stains inmice 2 days after inoculation with1–2 3 107 A. fumigatus conidia(magnification,3400). Inflammatorycellular infiltrate occurred in peri-bronchial areas, corresponding to siteof conidial deposition (arrows). Hy-phal forms were not present.C andD, Representative lung H&E andGMS stains in neutrophil-depletedanimals 2 days after inoculation with1–2 3 106 A. fumigatus conidia(magnification,3400). Inflammatorycellular infiltrate occurred peribron-chially in areas in which branchinghyphae (arrowheads) were present.Data shown are representative ofthree experiments.
FIGURE 4. Effect of transient neutrophil depletion on lung chitin afterA. fumigatuschallenge. Lung chitin levels in RB6-8C5-treated mice weredetermined at various times afterA. fumigatuschallenge (1–23 106
conidia). Af, A. fumigatus; RB6-8C5, rat anti-mouse Ly-6G IgG2b mAb.Experimentaln 5 6 per group.p, p , 0.05 as compared with animalsreceiving i.p. saline. Data shown are representative of two separateexperiments.
6089The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Effect of CXCR2 blockade on host defense againstA. fumigatus
To assess the contribution of the ELR1 CXC chemokines to hostresponse againstA. fumigatus, we next assessed survival of ani-mals pretreated with anti-CXCR2 Abs. In preliminary studies, ad-ministration of this Ab to normal animals did not affect the numberof circulating neutrophils over the subsequent 4 days (data notshown). Administration of anti-CXCR2 serum to normal animals2 h before challenge with 1–23 107 conidia resulted in a markedincrease in mortality, with.90% mortality in animals treated withanti-CXCR2, as compared with minimal mortality in immunocom-petent animals treated with a similar inoculum ofA. fumigatus(Fig. 7).
In order to determine the cause of increased lethality in anti-CXCR2-treated animals, mice were sacrificed on day 2 after chal-lenge with conidia, and lungs were harvested for histology, chitincontent, and MPO activity. Lungs of animals treated with controlserum contained a dense peribronchial, predominantly neutrophiliccellular infiltration, corresponding to areas of conidia deposition(Fig. 8, A and B). Hyphae were not present. However, lungs of
animals treated with anti-CXCR2 serum contained patchy peri-bronchial mononuclear cellular infiltration and fibrin deposition,with a paucity of neutrophils as compared with anti-CXCR2-treated animals (Fig. 8C). Importantly, fungal stainsshowed few conidia and large numbers of hyphae, indicating thedevelopment of invasive disease (Fig. 8D).
To quantify the amount of hyphae present, lung chitin levelswere measured 2 days after challenge with conidia, in animalsreceiving anti-CXCR2 or control serum. A 30-fold increase in lungchitin content was noted in animals treated with anti-CXCR2 wildcompared with control animals (Fig. 9A). Furthermore, lungs ofanimals treated with anti-CXCR2 contained 63% less myeloper-oxidase at 2 days after challenge withA. fumigatus, as comparedwith animals treated with control serum (Fig. 9B), corroboratingthe histologic observation of paucity of neutrophils.
Effect of compartmentalized overexpression of KC on hostdefense againstA. fumigatus
Having demonstrated the neutralization of the common receptorfor ELR1 CXC chemokines results in marked impairment of host
FIGURE 5. Lung MPO activity and BAL cells afterA. fumigatuschal-lenge in transiently neutrophil-depleted mice.A, Whole lung MPO activitymeasured at various times afterA. fumigatuschallenge in normal mice andthose treated with RB6-8C5 (1–23 106 conidia)B, Cell numbers in bron-cho-alveolar lavage fluid on day 2 afterA. fumigatuschallenge in normalmice and those treated with RB6-8C5 (1–23 106 conidia). Af,A. fumiga-tus; MPO, myeloperoxidase; RB6-8C5, rat anti-mouse Ly-6G IgG2b mAb.Experimentaln 5 6 for each group.p, p , 0.05 as compared with animalsreceiving i.t. vehicle.
FIGURE 6. Time-dependent production of KC and MIP-2 protein inlungs from normal and neutrophil-depleted mice after the i.t. administra-tion of vehicle orA. fumigatusconidia.A, Lung KC levels 1–23 106 A.fumigatusconidia or vehicle challenge.B, Lung MIP-2 levels 1–23106 A.fumigatusconidia or vehicle challenge. Af,A. fumigatus; RB6-8C5, ratanti-mouse Ly-6G IgG2b mAb. Experimentaln 5 6 for each group.p, p ,0.05 as compared with animals receiving i.t. vehicle. Data shown are rep-resentative of three separate experiments.
6090 CXCR2 IN INVASIVE ASPERGILLOSIS
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

defense againstA. fumigatus, we sought to determine if the over-expression of the ligand KC results in enhanced clearance of thepathogen. Since immunocompetent animals are naturally resistantto infection withAspergillusspecies, and patients with defects inneutrophil numbers or function are at highest risk of developingAspergillusinfections, we utilized transiently neutrophil-depletedmice to assess the effect of KC overexpression in this infection.Transient neutrophil depletion was achieved in wild-type andtransgenic animals with lung-specific overexpression of KC by i.p.injection of 100mg of RB6-8C5 mAb. This resulted in peripheralblood neutropenia (absolute neutrophil count,50 cells/ml) ondays 1 and 3 after injection in all animals, with restoration ofperipheral counts to pretreatment levels by day 5 (data not shown).Animals were challenged with 1–23 106 i.t. A. fumigatusconidiaafter 24 h. Lung histology of uninfected wild-type and transgenicanimals was normal and was not affected by administration ofRB6-8C5 mAb.
In survival studies, KC transgenic animals had a 46% reductionin mortality compared with wild-type animals (Fig. 10A). Lungchitin levels measured 2 days after challenge with conidia showedKC transgenic animals to have a 67% decrease in lung chitin con-tent as compared with wild-type controls (Fig. 10B). Lung histol-ogy obtained on day 2 showed a greater accumulation of neutro-phils associated with areas of fungal hyphae in KC transgenicmice, as compared with wild-type animals (data not shown).
DiscussionInvasive pulmonary aspergillosis is a devastating complication ofimmunosuppression, which is associated with abnormal numbersor function of neutrophils. ELR1 CXC chemokines have beenshown to have a critical role in neutrophil-mediated events in otherdiseases. We therefore tested the hypothesis that the interaction ofthese ligands with CXCR2 is a major component of neutrophil-dependent pulmonary host defense againstA. fumigatus, mediatingneutrophils deployment in response to the organism.
In order to evaluate the role of neutrophils in host defenseagainstA. fumigatusin an animal model, we developed a model oftransient in vivo neutrophil depletion. The Ab-mediated depletionof the available pool of mature neutrophils allowed the organism toestablish tissue invasion, which was not observed in animals withintact neutrophil number and function. The dynamics of reductionand early recovery of neutrophil numbers in this model, induced bya single dose of RB6-8C5 1 mAb on day21, are analogous to theclinical recovery of neutrophils in patients after chemotherapy orbone marrow transplantation. Two days after i.t. challenge withAspergillus, the number of neutrophils in the lungs of RB6-8C5-treated animals was larger than that in normal controls challengedwith the same inoculum. This greater influx of neutrophils in RB6-8C5-treated animals was in response to fungal tissue invasion andoccurred in the context of peripheral blood neutropenia, represent-ing maximal deployment of the limited pool of newly producedneutrophils to the site of infection. An advantage of this model isthat it is unaffected by the pleotropic influence of chemotherapeu-tic agents on various other immunologically active cells. Further-more, clinical and histological features of invasive aspergillosis inthis model resembled those seen in human disease.
FIGURE 7. Effect of anti-CXCR2 serum on survival in normal mice.Mice were treated with either control or anti-CXCR2 serum i.p. 2 h priorto inoculation withA. fumigatus(2 3 107 conidia).p, p , 0.05 as com-pared with animals receiving control serum. Af,A. fumigatus; anti-CXCR2, anti-CXCR2 serum. Experimentaln 5 15/group. Data shown arerepresentative of two experiments. Pooled data from two separateexperiments.
FIGURE 8. Effect of CXCR2 neutraliza-tion on lung histopathology afterA. fumiga-tus challenge.A andB, Representative lungH&E and GMS stains in mice treated withcontrol serum, 2 days after inoculation with1–2 3 107 A. fumigatusconidia (magnifica-tion, 3400). Inflammatory cellular infiltrateoccurred in peribronchial areas, correspond-ing to site of conidial deposition. Hyphalforms were not present.C andD, Represen-tative lung H&E and GMS stains in micetreated with anti-CXCR2 serum, 2 days afterinoculation with 1–23 106 A. fumigatusconidia (magnification,3400). Predomi-nantly mononuclear inflammatory cellularinfiltrate occurred peribronchially, in areas inwhich branching hyphae were present.
6091The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Given that the ELR1 CXC chemokines have been shown tomediate neutrophil recruitment and activation in diverse condi-tions, we elected to study their contribution to neutrophil-mediatedhost defense againstA. fumigatus. Other studies have suggested arole for chemokines in host defense againstA. fumigatus. For ex-ample, isolated rat alveolar macrophages have been shown to pro-duce MIP-1a, MIP-2, and KC, as well as TNF-a in response toA.fumigatusconidia in vitro (52), and knockout mice lacking CCchemokine receptor-1 (CCR1), a receptor for MIP-1a and RAN-TES, developed disseminated infection when administeredA. fu-migatusi.v. (53). In a previous study, we showed that neutraliza-tion of TNF resulted in reduced lung neutrophil influx associatedwith attenuated levels of lung CXC and CC chemokines, includingMIP-2, MIP-la, and JE (54). In this context, we examined theimportance of ELR1 CXC chemokines as a group againstA. fu-migatus, by blocking their common receptor in the mouse. Thisapproach allowed us to overcome the difficulties in simultaneousneutralization of the biologic effects of multiple ligands, whichmay have redundant and overlapping functions. The effects ofELR1 CXC chemokines in humans are mediated via bothCXCR-1 and CXCR-2. While the relative simplicity of the murine
model allowed us to address the role of these mediators in hostdefense againstA. fumigatus, the dissimilarities between the mu-rine and human systems represent a limitation of the present study.We employed a goat anti-murine serum, which did not depletecirculating neutrophils but prevented in vivo influx of neutrophilsinto the peritoneum in response to exogenous KC. The pneumoniathat resulted fromAspergillusinoculation in the setting of CXCR2neutralization was indistinguishable from that caused by neutro-phil depletion. The residual lung accumulation of neutrophils inthe setting of CXCR2 neutralization indicates that other mecha-nisms, such as other chemotactic factors or differential expressionof adhesion molecules, may play a role in neutrophil influx inresponse toA. fumigatus. However, the magnitude of effect ofCXCR2 neutralization highlights the essential role of CXCR2 li-gands in host defense againstA. fumigatus, nearly to the exclusionof other chemotactic mediators.
The two best-studied murine ELR1 CXC chemokines, MIP-2and KC, are functional homologues of the human ELR1 CXCchemokines, IL-8 and Gro- chemokines (55, 56). MIP-2 and KC
FIGURE 9. Effect of anti-CXCR2 serum on lung chitin content andMPO activity afterA. fumigatuschallenge.A, Lung chitin levels in micetreated with anti-CXCR2 serum or control serum 2 days afterA. fumigatuschallenge (1–23 107 conidia).B, Lung MPO activity in mice treated withanti-CXCR2 serum or control serum 2 days afterA. fumigatuschallenge(1–2 3 107 conidia). AF, A. fumigatus; aCXCR2, anti-CXCR2 serum;MPO, myeloperoxidase. Experimentaln 5 6 for each group.p, p , 0.05as compared with animals receiving i.t. vehicle. Data shown are represen-tative of two separate experiments.
FIGURE 10. Effect of lung-specific overexpression of KC on the out-come ofA. fumigatuspneumonia in neutrophil-depleted mice. All micewere treated with i.p. RB6-8C5 mAb 1 day prior to challenge withA.fumigatus(1–23 106 conidia).A, Survival after challenge with conidia inwild-type and transgenic animals. Experimentaln 5 13 or 14 animals/group.p, p , 0.05 compared with wild-type animals. Pooled data from twoseparate experiments.B, Lung chitin levels 2 days after challenge withconidia in wild-type and transgenic animals. Experimentaln 5 6 per group.p, p , 0.05 as compared with animals receiving i.t. vehicle.
6092 CXCR2 IN INVASIVE ASPERGILLOSIS
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

have been shown to mediate neutrophil-dependent host defense inother models of pneumonia. Specifically, in a murine model ofPseudomonaspneumonia, MIP-2 and KC levels were associatedwith presence of neutrophils in lung (57), and in murineKlebsiellapneumonia, MIP-2 was shown to be involved in recruitment ofneutrophils, but had only modest effects on clearance of the patho-gen and survival of animals (29). In preliminary studies, we foundthat Ab-mediated depletion of MIP-2 resulted in minimal changein survival in animals challenged withAspergillus (data notshown).
We found markedly greater levels of KC and MIP-2 in the lungsof transiently neutrophil-depleted animals, as compared with im-munocompetent animals, challenged withA. fumigatus. This dif-ference may be due to the greater burden of organisms in animalswith transient neutrophil depletion. Other mechanisms may in-clude local negative feedback on the production of the chemo-kines, or their receptor-mediated uptake by neutrophils. Thepresent study showed that lung-specific overexpression of KC re-sulted in augmented host defense againstAspergillus, as mani-fested by reduced mortality and lung fungal burden. Given thatimmunocompetent mice are intrinsically resistant toAspergillusinfection, and that neutropenic patients are at greatest risk of de-veloping invasive aspergillosis, we used a mAb to transiently de-plete neutrophils to assess the effect of KC overexpression in thelung. The limited pool of available neutrophils induced in theseanimals is analogous to the clinical setting in which invasive as-pergillosis occurs. The mechanism of the protective effect of KCagainstA. fumigatusis likely via enhanced recruitment of this lim-ited pool of available neutrophils to the lung, as suggested byhistology, and may also involve greater neutrophil antifungal ac-tivity. These findings are consistent with the prior observation of aprotective role for KC in murine bacterial pneumonia, which wasassociated with an augmented neutrophil influx (58).
The substantial effect of CXCR2 neutralization suggests thatmultiple ligands may be involved. Other candidate murine ELR1
CXC chemokines are less well characterized. A newly describedmurine chemokine, LIX, has been shown to share structural ho-mology with the human chemokines ENA-78 and GCP-2 (18), andis chemotactic for neutrophils in vitro (59). In a murine model ofsystemic endotoxemia, the expression of the ELR1 CXC chemo-kines was found to differ markedly in various organs (19), withprominent expression of MIP-2 in the lung, LIX in the heart, andequal expression of KC in lung, heart, and liver. Another recentlydescribed murine ELR1 CXC chemokine, Lungkine, has also beenshown to be chemotactic for neutrophils in vitro, and is constitu-tively expressed by lung epithelial cells, but not expressed in otherorgans. The preferential expression of chemokines in specific or-gans is of interest, and suggests that the various ELR1 CXC che-mokines may have distinct biological roles.
This study has identified CXCR2 ligands as required compo-nents of host defense againstA. fumigatus. This family of medi-ators may be important targets in devising future therapeutic strat-egies against invasive aspergillosis. We have shown thattransgenic overexpression of KC in the lungs is feasible and is notcomplicated by detrimental effects (58). Future studies will eval-uate the effect of overexpression or augmentation of other ELR1
CXC chemokines on the outcome of invasive aspergillosis.
References1. Fraser, D. W., J. I. Ward, L. Ajello, and B. D. Plikaytis. 1979. Aspergillosis and
other systemic mycoses: the growing problemJ. Am. Med. Assoc. 242:1631.2. Groll, A. H., P. M. Shah, C. Mentzel, M. Schneider, G. Just-Nuebling, and
K. Huebner. 1996. Trends in the postmortem epidemiology of invasive fungalinfections at a university hospital.J. Infect. 33:23.
3. Denning, D. W. 1998. Invasive aspergillosis.Clin. Infect. Dis. 26:781.
4. Denning, D. W., and D. A. Stevens. 1990. Antifungal and surgical treatment ofinvasive aspergillosis: review of 2,121 published cases.Rev. Infect. Dis. 12:1147.
5. Walsh, T. J., J. W. Heimenz, N. L. Seibel, J. R. Perfect, G. Horwith, L. Lee,J. L. Sibler, M. J. DiNubile, A. Reboli, E. Bow, J. Lister, and E. J. Anaissie. 1998.Amphotericin B lipid complex for invasive fungal infections: analysis of safetyand efficacy in 556 cases.Clin. Infect. Dis. 26:1383.
6. Denning, D. W. 1996. Therapeutic outcome in invasive aspergillosis.Clin. Infect.Dis. 23:608.
7. Rankin, N. E. 1953. Disseminated aspergillosis and moniliasis associated withagranulocytosis and antibiotic therapy.Br. Med. J. 183:918.
8. Khoo, S. H., and D. W. Denning. 1994. Invasive aspergillosis in patients withAIDS. Clin. Infect. Dis. 19(Suppl 1):S41.
9. Wald, A., W. Leisenring, J. A. van Burik, and R. A. Bowden. 1997. Epidemiol-ogy of Aspergillusinfections in a large cohort of patients undergoing bone mar-row transplantation.J. Infect. Dis. 175:1459.
10. Cohen, M. S., R. E. Isturiz, H. L. Malech, R. K. Root, C. M. Wilfert, L. Gutman,and R. H. Buckley. 1981. Fungal infection in chronic granulomatous disease: theimportance of the phagocyte in defense against fungi.Am. J. Med. 71:59.
11. Gerson, S. L., G. H. Talbot, S. Hurwitz, B. L. Strom, E. J. Lusk, andP. A. Cassileth. 1984. Prolonged granulocytopenia: the major risk factor for in-vasive pulmonary aspergillosis in patients with acute leukemia.Ann. Int. Med.100:345.
12. Schaffner, A., H. Douglas, and A. Braude. 1982. Selective protection againstconidia by mononuclear and against mycelia by polymorphonuclear phagocytesin resistance toAspergillus: observations on these two lines of defense in vivoand in vitro with human and mouse phagocytes.J. Clin. Invest. 69:617.
13. Levitz, S. M., M. E. Selsted, T. Ganz, R. I. Lehrer, and R. D. Diamond. 1986. Invitro killing of spores and hyphae ofAspergillus fumigatusandRhizopus oryzaeby rabbit neutrophil cationic peptides and bronchoalveolar macrophages.J. In-fect. Dis. 154:483.
14. Rex, J. H., J. E. Bennett, J. I. Gallin, H. L. Malech, and D. A. Melnick. 1990.Normal and deficient neutrophils can cooperate to damageAspergillus fumigatushyphae.J. Infect Dis. 162:523.
15. Roilides, E., K. Uhlig, D. Venzon, P. A. Pizzo, and T. J. Walsh. 1993. Preventionof corticosteroid-induced suppression of human polymorphonuclear leukocyte-induced damage ofAspergillus fumigatushyphae by granulocyte colony-stimu-lating factor andg interferon.Infect Immun. 6l:4870.
16. Morgenstern, D. E., M. A. Gifford, L. L. Li, C. M. Doerschuk, andM. C. Dinauer. 1997. Absence of respiratory burst in X-linked chronic granulo-matous disease mice leads to abnormalities in both host defense and inflammatoryresponse toAspergillus fumigatus. J. Exp. Med. 185:207.
17. Cenci, E., A. Mencacci, C. Fe d’Ostiani, G. Del Sero, P. Mosci, C. Montagnoli,A. Bacci, and L. Romani. 1998. Cytokine- and T helper-dependent lung mucosalimmunity in mice with invasive pulmonary aspergillosis.J. Infect. Dis. 178:1750.
18. Smith, J. B., L. E. Rovai, and H. R. Herschman. 1997. Sequence similarities of asubgroup of CXC chemokines related to murine LIX: implications for the inter-pretation of evolutionary relationships among chemokines.J. Leukocyte Biol.62:598.
19. Rovai, L. E., H. R. Herschman, and J. B. Smith. 1998. The murine neutrophil-chemoattractant chemokines LIX, KC, and MIP-2 have distinct induction kinet-ics, tissue distributions, and tissue-specific sensitivities to glucocorticoid regula-tion in endotoxemia.J. Leukocyte Biol. 64:494.
20. Baggiolini, M., A. Walz, and S. L. Kunkel. 1989. Neutrophil-activating peptide-1/interleukin 8, a novel cytokine that activates neutrophils.J. Clin. Invest. 84:1045.
21. Richardson, M. D., and M. Patel. 1995. Stimulation of neutrophil phagocytosis ofAspergillus fumigatusconidia by interleukin-8 andN-formylmethionyl-leucyl-phenylalanine.J. Med. Vet. Mycol. 33:99.
22. Thelen, M., P. Peveri, P. Kernen, V. von Tscharner, A. Walz, and M. Baggiolini.1988. Mechanism of neutrophil activation by NAF, a novel monocyte-derivedpeptide agonist.FASEB J. 2:2702.
23. Wolpe, S. D., G. Davatelis, B. Sherry, B. Beutler, D. G. Hesse, H. T. Nguyen,L. L. Moldawer, C. F. Nathan, S. F. Lowry, and A. Cerami. 1988. Macrophagessecrete a novel heparin-binding protein with inflammatory and neutrophil che-mokinetic properties.J. Exp. Med. 167:570.
24. Wolpe, S. D., and A. Cerami. 1989. Macrophage inflammatory proteins 1 and 2:members of a novel superfamily of cytokines.FASEB J. 3:2565.
25. Broaddus, V. C., A. M. Boylan, J. M. Hoeffel, K. J. Kim, M. Sadick,A. Chuntharapai, and C. A. Hebert. 1994. Neutralization of IL-8 inhibits neutro-phil influx in a rabbit model of endotoxin-induced pleurisy.J. Immunol. l52:2960.
26. Sekido, N., N. Mukaida, A. Harada, I. Nakanishi, Y. Watanabe, andK. Matsushima. 1993. Prevention of lung reperfusion injury in rabbits by a mono-clonal antibody against interleukin-8.Nature 365:654.
27. Xing, Z., M. Jordana, H. Kirpalani, K. E. Driscoll, T. J. Schall, and J. Gauldie.1994. Cytokine expression by neutrophils and macrophages in vivo: endotoxininduces tumor necrosis factor-a, macrophage inflammatory protein-2, interleu-kin-1b, and interleukin-6 but not RANTES or transforming growth factor-b1mRNA expression in acute lung inflammation. [Published erratum appears in1994, Am. J. Respir. Cell. Mol. Biol. 10:346]. Am. J. Respir. Cell. Mol. Biol.10:148.
28. Huang, S., J. D. Paulauskis, J. J. Godleski, and L. Kobzik. 1992. Expression ofmacrophage inflammatory protein-2 and KC mRNA in pulmonary inflammation.Am. J. Pathol. 141:981.
29. Greenberger, M. J., R. M. Strieter, S. L. Kunkel, J. M. Danforth, L. L. Laichalk,D. C. McGillicuddy, and T. J. Standiford. 1996. Neutralization of macrophageinflammatory protein-2 attenuates neutrophil recruitment and bacterial clearancein murineKlebsiella pneumomiae. J. Infect. Dis. 173:159.
6093The Journal of Immunology
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

30. Loetscher, P., M. Seitz, I. Clark-Lewis, M. Baggiolini, and B. Moser. 1994. Bothinterleukin-8 receptors independently mediate chemotaxis: Jurkat cells trans-fected with IL-8R1 or IL-8R2 migrate in response to IL-8, GROa and NAP-2.FEBS Lett. 341:187.
31. Morohashi, H., T. Miyawaki, H. Nomura, K. Kuno, S. Murakami,K. Matsushima, and N. Mukaida. 1995. Expression of both types of human in-terleukin-8 receptors on mature neutrophils, monocytes, and natural killer cells.J. Leukocyte Biol. 57:180.
32. Thomas, K. M., L. Taylor, and J. Navarro. 1991. The interleukin-8 receptor isencoded by a neutrophil-specific CDNA clone, F3R.J. Biol. Chem. 266:14839.
33. Moser, B., C. Schumacher, V. von Tscharner, I. Clark-Lewis, and M. Baggiolini.1991. Neutrophil-activating peptide 2 and gro/melanoma growth-stimulatory ac-tivity interact with neutrophil-activating peptide 1/interleukin 8 receptors on hu-man neutrophils.J. Biol. Chem. 266:10666.
34. Ahuja, S. K., and P. M. Murphy. 1996. The CXC chemokines growth-regulatedoncogene (GRO) alphaa, GROb GROg, neutrophil-activating peptide-2, andepithelial cell-derived neutrophil-activating peptide-78 are potent agonists for thetype B, but not the type A, human interleukin-8 receptor.J. Biol. Chem. 271:20545.
35. Lee, J., G. Cacalano, T. Camerato, K. Toy, M. W. Moore, and W. I. Wood. 1995.Chemokine binding and activities mediated by the mouse IL-8 receptor.J. Im-munol. 155:2158.
36. Terkeltaub, R., S. Baird, P. Sears, R. Santiago, and W. Boisvert. 1998. Themurine homolog of the interleukin-8 receptor CXCR-2 is essential for the occur-rence of neutrophilic inflammation in the air pouch model of acute urate crystal-induced gouty synovitis.Arthritis Rheum. 41:900.
37. Evanoff, H. L., M. D. Burdick, S. A. Moore, S. L. Kunkel, and R. M. Strieter.1992. A sensitive ELISA for the detection of human monocyte chemoattractantprotein-1 (MCP-l).Immunol. Invest. 21:39.
38. Strieter, R. M., S. L. Kunkel, M. D. Burdick, P. M. Lincoln, and A. Walz. 1992.The detection of a novel neutrophil-activating peptide (ENA-78) using a sensitiveELISA. Immunol. Invest. 21:589.
39. Hebert, C. A., A. Chuntharapai, M. Smith, T. Colby, J. Kim, and R. Horuk. 1993.Partial functional mapping of the human interleukin-8 type A receptor: identifi-cation of a major ligand binding domain.J. Biol. Chem. 268:18549.
40. Lira, S. A., P. Zalamea, J. N. Heinrich, M. E. Fuentes, D. Carrasco, A. C. Lewin,D. S. Barton, S. Durham, and R. Bravo. 1994. Expression of the chemokineN51/KC in the thymus and epidermis of transgenic mice results in marked in-filtration of a single class of inflammatory cells.J. Exp. Med. 180:2039.
41. Lira, S. A., M. E. Fuentes, R. M. Strieter, and S. K. Durham. 1997. Transgenicmethods to study chemokine function in lung and central nervous system.Meth-ods Enzymol. 287:304.
42. Lira, S. A. 1996. Genetic approaches to study chemokine function.J. LeukocyteBiol. 59:45.
43. Fleming, T. J., M. L. Fleming, and T. R. Malek. 1993. Selective expression ofLy-6G on myeloid lineage cells in mouse bone marrow: RB6–8C5 mAb to gran-ulocyte-differentiation antigen (Gr-1) detects members of the Ly-6 family.J. Im-munol. 151:2399.
44. Lai, L., N. Alveradi, L. Maltais, and H. C. Morse III. 1998. Mouse cell surfaceantigens: nomenclature and immunophenotyping.J. Immunol. 160:3861.
45. Lewinsohn, D. M., R. F. Bargatze, and E. C. Butcher. 1987. Leukocyte-endothe-lial cell recognition: evidence of a common molecular mechanism shared byneutrophils, lymphocytes, and other leukocytes.J. Immunol. 138:4313.
46. Pennline, K. J., F. Pellerito, M. DaFonseca, P. Monahan, M. I. Siegel, andS. R. Smith. 1990. Flow cytometric analysis of recombinant murine GM-CSF(rmuGM-CSF) induced changes in the distribution of specific cell populations invivo. Cytometry 11:283.
47. Tepper, R. I., R. L. Coffman, and P. Leder. 1992. An eosinophil-dependent mech-anism for the antitumor effect of interleukin-4.Science 257:548.
48. Allen, S. D., K. N. Sorensen, M. J. Nejdl, C. Durrant, and R. T. Proffit. 1994.Prophylactic efficacy of aerosolized liposomal (AmBisome) and non-liposomal(Fungizone) amphotericin B in murine pulmonary aspergillosis.J. Antimicrob.Chemother. 34:1001.
49. Lehmann, P. F., and L. 0. White. 1975. Chitin assay used to demonstrate renallocalization and cortisone-enhanced growth ofAspergillus fumigatusmyceliumin mice. Infect. Immun. 12:987.
50. Goldblum, S. E., K. M. Wu, and M. Jay. 1985. Lung myeloperoxidase as a mea-sure of pulmonary leukostasis in rabbits.J. Appl. Physiol. 59:1978.
51. Greenberger, M. J., R. M. Strieter, S. L. Kunkel, J. M. Danforth, R. E. Goodman,and T. J. Standiford. 1995. Neutralization of IL-10 increases survival in a murinemodel ofKlebsiella pneumoniae. J. Immunol. 155:722.
52. Shahan, T. A., W. G. Sorenson, J. D. Paulauskis, R. Morey, and D. M. Lewis.1998. Concentration- and time-dependent upregulation and release of the cyto-kines MIP-2, KC, TNF, and MIP-1a in rat alveolar macrophages by fungal sporesimplicated in airway inflammation.Am. J. Respir. Cell. Mol. Biol. 18:435.
53. Gao, J. L., T. A. Wynn, Y. Chang, E. J. Lee, H. E. Broxmeyer, S. Cooper,H. L. Tiffany, H. Westphal, J. Kwon-Chung, and P. M. Murphy. 1997. Impairedhost defense, hematopoiesis, granulomatous inflammation and type 1-type 2 cy-tokine balance in mice lacking CC chemokine receptor 1.J. Exp. Med. 185:1959.
54. Mehrad, B., R. M. Strieter, and T. J. Standiford. 1999. Role of TNF-a in pulmo-nary host defense in murine invasive aspergillosis.J. Immunol. 162:1633.
55. Yoshimura, T., and D. G. Johnson. 1993. cDNA cloning and expression of guineapig neutrophil attractant protein-1 (NAP-1): NAP-1 is highly conserved in guineapig. J. Immunol. 151:6225.
56. Tekamp-Olson, P., C. Gallegos, D. Bauer, J. McClain, B. Sherry, M. Fabre,S. van Deventer, and A. Cerami. 1990. Cloning and characterization of cDNAsfor murine macrophage inflammatory protein 2 and its human homologues.J. Exp. Med. 172:911.
57. Kooguchi, K., S. Hashimoto, A. Kobayashi, Y. Kitamura, I. Kudoh,J. Wiener-Kronish, and T. Sawa. 1998. Role of alveolar macrophages in initiationand regulation of inflammation inPseudomonas aeruginosapneumonia.Infect.Immun.66:3164.
58. Tsai, W. C., R. M. Strieter, J. M. Wilkowski, K. A. Bucknell, M. D. Burdick,S. A. Lira, and T. J. Standiford. 1998. Lung-specific transgenic expression of KCenhances resistance toKlebsiella pneumoniaein mice.J. Immunol. 161:2435.
59. Wuyts, A., A. Haelens, P. Proost, J. P. Lenaerts, R. Conings, G. Opdenakker, andJ. Van Danune. 1996. Identification of mouse granulocyte chemotactic protein-2from fibroblasts and epithelial cells: functional comparison with natural KC andmacrophage SA inflammatory protein-2.J. Immunol. 157:1736.
6094 CXCR2 IN INVASIVE ASPERGILLOSIS
by guest on Novem
ber 27, 2018http://w
ww
.jimm
unol.org/D
ownloaded from





























