Embed Size (px)
Citation preview

of June 22, 2018.This information is current as
LungkineLacking Expression of the CXC Chemokine Impaired Pulmonary Host Defense in Mice
Sergio A. LiraWiekowski, Albert Zlotnik, Theodore J. Standiford andVassileva, Denise J. Manfra, Donald N. Cook, Maria T. Shu-Cheng Chen, Borna Mehrad, Jane C. Deng, Galya
http://www.jimmunol.org/content/166/5/3362doi: 10.4049/jimmunol.166.5.3362
2001; 166:3362-3368; ;J Immunol
Referenceshttp://www.jimmunol.org/content/166/5/3362.full#ref-list-1
, 10 of which you can access for free at: cites 22 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2001 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on June 22, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Impaired Pulmonary Host Defense in Mice Lacking Expressionof the CXC Chemokine Lungkine1
Shu-Cheng Chen,* Borna Mehrad,† Jane C. Deng,‡ Galya Vassileva,* Denise J. Manfra,*Donald N. Cook,* Maria T. Wiekowski,* Albert Zlotnik, § Theodore J. Standiford,‡ andSergio A. Lira2*
Lungkine (CXCL15) is a novel CXC chemokine that is highly expressed in the adult mouse lung. To determine the biologicfunction of Lungkine, we generated Lungkine null mice by targeted gene disruption. These mice did not differ from wild-type micein their hematocrits or in the relative number of cells in leukocyte populations of peripheral blood or other tissues, including lungand bone marrow. However, Lungkine null mice were more susceptible toKlebsiella pneumoniainfection, with a decreasedsurvival and increased lung bacterial burden compared with infected wild-type mice. Histologic analysis of the lung and assess-ment of leukocytes in the bronchioalveolar lavage revealed that neutrophil numbers were normal in the lung parenchyma, butreduced in the airspace. The production of other neutrophil chemoattractants in the Lungkine null mice did not differ from thatin wild-type mice, and neutrophil migration into other tissues was normal. Taken together, these findings demonstrate thatLungkine is an important mediator of neutrophil migration from the lung parenchyma into the airspace. The Journal of Im-munology,2001, 166: 3362–3368.
Chemokines are a large family of 8- to 10-kDa polypeptidemolecules that regulate the immune system in many as-pects, including chemotaxis of leukocytes, hemopoiesis,
and angiogenesis (1–4). Based on the composition of a cysteinemotif located near the N terminus of these molecules, they areclassified into four subgroups: C, CC, CXC, and CX3C. The CXCchemokines are characterized by four cysteine residues at the Nterminus, the first two of which are separated by a nonconservedamino acid. They are further classified by the presence or the ab-sence of the amino acid sequence glutamic acid-leucine-arginine(the ELR motif) immediately preceding the CXC sequence. Nearlyall of the known neutrophil-targeted chemokines belong to theELR1CXC chemokine family.
Lungkine is an ELR1CXC chemokine that is highly expressedby lung airway epithelial cells (5). It has recently been designatedCXCL15, according to the new nomenclature system (6). The ex-pression of Lungkine is up-regulated in response to a variety ofinflammatory stimuli, including intratracheal challenge with LPS,Nippostronglyus, andAsperigillus(5). Because Lungkine inducesneutrophil recruitment in vivo and in vitro (5) and because it ishighly expressed in the lung in response to pathogens, it has been
hypothesized that this chemokine may have a distinct biologicalrole in pulmonary physiology, including host response to infection.
Recently, a novel hemopoietic regulatory factor, namedWECHE, was isolated from an endothelial cell line using a dif-ferential gene expression method and was shown to be identicalwith Lungkine (7). During development, the expression of eitherthe transcript or the protein of Lungkine/WECHE was found insites of hemopoietic stem and progenitor cell production and main-tenance, including day 9.5 dorsal aorta, day 10.5 yolk sac, aorta-gonad-mesonephrons, and fetal liver. In vitro, Lungkine inhibitsthe development of erythroid (BFU-E) progenitor cells obtainedfrom either mouse fetal liver or bone marrow and is chemotacticfor bone marrow cells in Transwell assays (7).
In an attempt to determine the biological roles of Lungkine invivo, we have generated Lungkine null (2/2) mice. In this study,we report that Lungkine2/2 mice have defects in pulmonary hostdefense.
Materials and MethodsGene targeting of Lungkine
Two BAC clones containing genomic copies of the mouse Lungkine genewere identified in a mouse 129/Sv-derived embryonic stem (ES)3 cellgenomic library (Incyte Genomics, St. Louis, MO) using PCR primerscorresponding to the mouse Lungkine cDNA: GV15 (59-TTAGGCATCACTGCCTGTCA-39) and GV16 (59-AATGAAGCTTCTGTATAGT39). Several fragments containing the Lungkine gene were identified bySouthern blot analysis of BAC plasmid DNA using a 500-bp cDNA probelabeled with [g-32P]dCTP (3000 Ci/mmol; Amersham, Arlington Heights,IL) by random priming (Megaprime DNA Labeling System; AmershamPharmacia Biotech, Piscataway, NJ). TwoBamHI fragments (7.6 kb each)that contained either the 59half or the 39half of the Lungkine cDNA weresubcloned into pBluescript (Stratagene, La Jolla, CA) and mapped by re-striction digest. The complete intron and exon sequences of these twosubclones were further analyzed using an Applied Biosystems 377 DNAsequencer (Applied Biosystems, Foster City, CA). DNA fragments corre-sponding to the 59and 39 regions of the Lungkine locus were, in turn,subcloned from this plasmid into the pOSDupDel vector (a gift from Oliver
*Department of Immunology, Schering-Plough Research Institute, Kenilworth, NJ07033; †Division of Pulmonary and Critical Care Medicine, University of TexasSouthwestern Medical Center, Dallas, TX 75390;‡Department of Medicine, Divisionof Pulmonary and Critical Care Medicine, University of Michigan Medical School,Ann Arbor, MI 48109; and§DNAX Research Institute, Palo Alto, CA 94304
Received for publication October 12, 2000. Accepted for publication December14, 2000.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby markedadvertisementin accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported in part by National Institutes of Health GrantsRO1HL56402 (to T.J.S.), RO1HL57243 (to T.J.S.), P50HL60289 (to T.J.S.), andK08HL04220 (to B.M.) and American Lung Association Grant RG-005-N (to B.M.).2 Address correspondence and reprint requests to Dr. Sergio A. Lira, Department ofImmunology, Schering-Plough Research Institute, 2015 Galloping Hill Road, Ken-ilworth, NJ 07033. E-mail address: [email protected]
3 Abbreviations used in this paper: ES, embryonic stem; p.i., postinoculation; BAL,bronchoalveolar lavage; MIP-2, macrophage-inflammatory protein-2.
Copyright © 2001 by The American Association of Immunologists 0022-1767/01/$02.00
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Smithies, University of North Carolina, Chapel Hill, NC) at either end ofthe neomycin resistance gene (neo). This targeting vector DNA was lin-earized using the restriction enzymeNotI before its introduction into 129/Sv-derived ES cells by electroporation. Lungkine-targeted ES clones wereidentified by Southern blotting. DNA was digested withBamHI, trans-ferred to nylon membranes, and hybridized to a radiolabeled DNA probederived from a region upstream of the 59region of homology. Cells whoseDNAs were of the structure predicted for the targeted locus were used togenerate chimeric mice using standard blastocyst injection procedures.Several correctly targeted ES cells were injected into blastocysts to gen-erate chimeric mice. Lungkine heterozygous (1/2) offspring were identi-fied using a PCR-based screening strategy using three oligonucleotideprimers corresponding to the region of homology, the neo gene, and thedeleted region of the Lungkine gene. Lungkine2/2 mice were generated byinterbreeding these heterozygous mice.
Mice
For the experiments described here we used Lungkine2/2 mice generatedin the 129 3 BL6 background. Age- and sex-matched (129 SvEv3C57BL/6)F2 control mice (Lungkine1/1) were obtained from The JacksonLaboratory (Bar Harbor, ME). All mice were housed in a specific patho-gen-free environment. All experiments in which animals were used wereconducted in accordance with the institutional guidelines of Schering-Plough and the University of Michigan.
RNA analysis
Total RNA was extracted from lung tissues of wild-type and Lungkine2/2
mice using an ULTRASPEC RNA isolation reagent (Biotecx, Houston,TX). Twenty micrograms of RNA was electrophoresed in a 1% agarose geland blotted onto a Duralone membrane (Stratagene). A Lungkine cDNAprobe was32P-labeled using the Megaprime DNA labeling random primerkit (Amersham Pharmacia Biotech) according to the manufacturer’s in-structions. Hybridization was conducted at 68°C in QuickHyb hybridiza-tion solution (Stratagene), and the blots were washed according to themanufacturer’s protocol. The membranes were then exposed overnight tox-ray film at 270°C with an intensifying screen.
Histology and immunohistochemistry
Tissues were either fresh-frozen for cryosection or perfused, inflated (forlung only), fixed in 4% paraformaldehyde, and processed for paraffin sec-tions. Routinely, 5-mm paraffin sections were cut and stained with hema-toxylin and eosin. For immunostaining, fresh-frozen sections were fixedwith cold acetone and stained with anti-CD11b/Mac-1 or anti-Gr-1 Abs(PharMingen, San Diego, CA). Binding of the Ab was amplified with aVectastain Elite ABC kit (Vector Laboratories, Burlingame, CA) and de-tected with a diaminobenzidine substrate kit from Vector Laboratories.Slides were counterstained with hematoxylin.
Preparation of lung single-cell suspension
Single-cell suspensions of the lungs were prepared as previously described(8). Briefly, freshly resected lungs were minced with scissors to a fineslurry and incubated at 37°C for 30 min in an RPMI 1640 solution (Sigma,St. Louis, MO) containing 1 mg/ml type A collagenase (Roche, Indiana-polis, IN) and 30 U/ml DNase (Sigma). The solutions were then drawn upand down 20 times in 10-ml syringes (Becton Dickinson, Franklin Lakes,NJ) to disperse the cells mechanically. The resulting cell suspensions werepelleted, resuspended, passed through Nitex mesh filters (Tetko, KansasCity, MO), and passed through a 20% Percoll gradient (Sigma) before cellcounting under a hemocytometer. Cytospin slides of this suspension werethen prepared and stained (Diff-Quik Stain set; Dade Behring, Newark,DE), and differential cell counts were determined using a high-power mi-croscope. The absolute number of a leukocyte subtype was determined bymultiplication of the percentage of that cell type by the total number oflung leukocytes in that sample.
Flow cytometry
Total lung leukocytes were isolated from animals as described above. Foridentification of lung leukocyte subsets, FITC- or PE-conjugated CD4,CD8a, CD3, CD11c, B220, pan NK, Mac-1, F4/80, and Gr-1 were used (allreagents from PharMingen). In addition, cells were stained with anti-CD45(PharMingen), allowing discrimination of leukocytes from nonleukocytesand thus eliminating any nonspecific binding of cell surface markers onnonleukocytes. Stained samples were kept in the dark at 4°C until analyzedon a FACScan cytometer (Becton Dickinson, San Jose, CA) usingCellQuest software (Becton Dickinson). Leukocyte populations were ana-lyzed by gating on CD45-positive cells of the appropriate light scatter
characteristics and then were examined for FL1 and FL2 fluorescence ex-pression. The absolute number of a leukocyte type was determined bymultiplication of the percentage of that cell type by the total number oflung leukocytes in that sample. For analyses of bone marrow and blood,single-cell suspensions were prepared from individual tissues by passagethrough a 100-mm pore size nylon cell strainer (BD Biosciences Labware,Bedford, MA) in RPMI 1640 medium containing 10% FCS.
Hematology
Blood samples were collected from the infraorbital sinus into sterile evac-uated tubes with added EDTA (Vacutainer Systems; Becton Dickinson,Rutherford, NJ). Hematologic values were determined with an automatedsystem (Cell-Dyn 3500; Abbott, Chicago, IL). Platelet counts were per-formed manually when the instrument was unable to provide accurateplatelet counts due to excessive clumping or excessively large platelets.
Klebsiella pneumoniaeinoculation
We chose to useK. pneumoniaestrain 43816, serotype 2 (American TypeCulture Collection, Manassas, VA) in our studies because a murine modelof pneumonia has been well characterized using this strain (9, 10).K.pneumoniaewas grown in tryptic soy broth (Difco, Detroit, MI) for 18 hat 37°C. The concentration of bacteria in broth was determined by mea-suring the amount of absorbance at 600 nm. A standard of absorbenciesbased on known CFU was used to calculate the inoculum concentration. Aninoculum of 13 102 organisms was chosen, as this dose allowed for thedevelopment of substantial inflammation by 36–48 h without excessivemortality in wild-type animals. Mice were anesthetized with;1.8–2 mg ofpentobarbital/animal i.p. The trachea was exposed, and 30ml of inoculumor saline was administered via a sterile 26-gauge needle. The skin incisionwas closed with surgical staples.
Determination of lungK. pneumoniaeCFU
At the time of sacrifice, the right ventricle was perfused with 1 ml of PBS,the lungs were removed aseptically and placed in 3 ml of sterile saline, andthe tissues were homogenized with a tissue homogenizer under a ventedhood. The lung homogenates were placed on ice, and serial 1/5 dilutionswere made. Ten microliters of each dilution was plated on soy-base bloodagar plates (Difco) and incubated for 18 h at 37°C, and then the colonieswere counted.
Bronchoalveolar lavage (BAL)
BAL was performed to obtain airspace cells as previously described (11).The trachea was exposed and intubated using a 0.97-mm outside diameterpolyethylene catheter. BAL was performed by instilling PBS containing5 mM EDTA in 1-ml aliquots. Approximately 5 ml of lavage fluid wasretrieved per mouse. Cytospins were then prepared from BAL cells andstained with Diff-Quik (Dade Behring, Newark, DE), and differentialcounts were determined.
Lung harvesting for cytokine analysis
At designated time points, mice were sacrificed by carbon dioxide inhala-tion, and blood was collected by direct cardiac puncture. The pulmonaryvasculature was perfused with 1 ml of PBS containing 5 mM EDTA via theright ventricle, and whole lungs were harvested for assessment of the var-ious cytokine protein levels. Lungs were homogenized in 1.5 ml of com-plete protease inhibitor lysis buffer (Roche). Homogenates were incubatedon ice for 30 min, then centrifuged at 2500 rpm for 10 min. Supernatantswere collected, passed through a 0.45-mm pore size filter (Gelman Sci-ences, Ann Arbor, MI), then stored at220°C for assessment of cytokinelevels.
Murine cytokine ELISA
Murine TNF-a, macrophage-inflammatory protein-2 (MIP-2), and KCwere measured using a modification of a double-ligand method as de-scribed previously (12). Briefly, flat-bottom 96-well microtiter plates (Im-muno-Plate I 96-F; Nunc, Copenhagen, Denmark) were coated with 50ml/well rabbit Ab against the various cytokines (1mg/ml in 0.6 M NaCl,0.26 M H3BO4, and 0.08 M NaOH, pH 9.6) for 16 h at 4°C and thenwashed with PBS (pH 7.5) and 0.05% Tween 20 (wash buffer). Nonspecificbinding sites on the microtiter plates were blocked with 2% BSA in PBSand incubated for 90 min at 37°C. After rinsing four times with washbuffer, the plates were added with diluted (undiluted and 1/10) cell-freesupernatants (50ml) in duplicate, followed by incubation for 1 h at 37°C.Plates were washed four times, followed by the addition of 50ml/wellbiotinylated rabbit Abs against the specific cytokines (3.5mg/ml in PBS
3363The Journal of Immunology
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

(pH 7.5), 0.05% Tween 20, and 2% FCS), and incubated for 30 min at37°C. Streptavidin-peroxidase conjugate (Bio-Rad, Richmond, CA) wasthen added after washing the plates, and incubation proceeded for 30 minat 37°C. Plates were washed again four times, and chromogen substrate(Bio-Rad) was added. The plates were incubated at room temperature to thedesired extinction, and the reaction was terminated with 50ml/well 3 MH2SO4 solution. Plates were read at 490 nm in an ELISA plate reader.Standards were half-log dilutions of recombinant murine cytokines from 1pg/ml to 100 ng/ml. This ELISA method consistently detected murine cy-tokine concentrations.25 pg/ml. The ELISA did not cross-react withIL-1, IL-2, IL-4, or IL-6. In addition, the ELISA did not cross-react withother members of the murine chemokine family, including murine MIP-1,JE, RANTES, or ENA-78.
In vivo neutrophil chemotaxis
The mouse air-pouch model for in vivo chemotaxis has been described indetail previously (13). Briefly, sex- and age-matched Lungkine2/2 andwild-type mice were anesthetized with ether. On experimental day 0, 5 ml
of sterile air was injected s.c. under the dorsal skin; the resultant space wasreinjected with 3 ml of sterile air on day 3. On day 5, 0.2mg of Escherichiacoli LPS (Sigma) in 1 ml of carboxymethylcellulose (0.5% in saline; Fluka,Buchs, Switzerland) was injected into the pouches. The animals were sac-rificed 4 h later, and the air-pouchs were lavaged with 2 ml of sterile PBS.The resulting cell suspensions were pelleted, resuspended, and countedunder a hemocytometer. Cytospin slides were prepared and stained (Diff-Quik Stain set; Dade Behring), and differential cell counts were determinedusing a high-power microscope. The absolute number of a leukocyte typewas determined by multiplication of the percentage of that cell type by thetotal number of lung leukocytes in that sample.
Statistical analysis
Data were analyzed using the InStat version 2.01 statistical package(GraphPad software; GraphPad, San Diego, CA). Survival data were com-pared using Fisher’s exact test. All other data were expressed as themean6 SEM and compared using an unpaired two-tailed Mann-Whitney
FIGURE 1. Generation of Lungkine2/2 mice.A, Targeting strategy. The DNA structures of the targeting vector, the Lungkine locus, and the targetedlocus are shown. Arrows indicate the transcriptional direction of each gene. Open rectangles, exons. A DNA probe (filled rectangle) from a 59 upstreamhomologous region of Lungkine gene was used to screen the ES cells. A 7.6- and a 5-kbBamHI fragment were expected for the wild-type Lungkine locusand for the targeted locus, respectively. Restriction enzymes shown are: H,HindIII; H/P, HindIII/PlmI; B, BamHI; and E,EcoRI.B, An analysis of mousegenomic DNA by PCR. An example of PCR products of wild-type (1/1), Lungkine heterozygous (1/2), and Lungkine null (2/2) mice is shown.C,Northern blot analysis of Lungkine2/2 mice. Total RNA was isolated from lungs of two separate Lungkine2/2 lines (L #25 and L #69) and from wild-typemice. A total of 20mg of total RNA was loaded on each lane. Lungkine cDNA was used as a probe for the hybridization. No Lungkine mRNA was detectedin lungs from either line of Lungkine2/2 mice.
3364 LUNGKINE IN HOST DEFENSE
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
U (nonparametric) test. Ap , 0.05 was considered to be statisticallysignificant.
ResultsGeneration of Lungkine2/2 mice
DNA sequence analysis of the Lungkine genomic locus revealedthat it contains three introns and four exons. The translation startsite is located in exon 1, and the open reading frame spans all fourexons (Fig. 1A). A targeting strategy was designed to delete a 3-kbsequence of Lungkine genomic DNA that includes the entire firstand second exons and part of the third exon (Fig. 1A). The target-ing vector shown in Fig. 1 was used to transfect ES cells. Twoindependent ES cell clones containing the targeted loci were iden-tified by Southern blot analysis and injected into mouse blasto-cysts. Chimeras obtained from both clones were bred withC57BL/6J mice to generate heterozygotes (Lungkine1/2). PCRanalysis of tail DNA from these founders confirmed germlinetransmission of the targeted allele (Fig. 1B). Further intercrossingof these Lungkine1/2 mice yielded Lungkine2/2 mice within theexpected Mendelian ratios, demonstrating that Lungkine expres-sion is not essential for embryonic development.
Northern blot analysis was used next to examine LungkinemRNA expression in the lung tissues of Lungkine2/2 mice. Asshown in Fig. 1C, a Lungkine mRNA of the predicted size wasdetected in the wild-type mouse, but not in either line ofLungkine2/2 mice.
Lungkine2/2 mice develop normally and have a normalcomplement of leukocytes in the lung
The Lungkine2/2 mice developed normally and were fertile. Rou-tine histologic analysis of all organs was unremarkable. GivenLungkine’s expression pattern and its chemotactic properties, wefocused our initial analysis on the lung using immunohistochem-istry and flow cytometry. Anti-Mac-1 and anti-Gr-1 Abs were usedin immunohistochemical staining to examine the resident macro-phage and neutrophil populations in the lung tissues ofLungkine2/2 animals. No significant differences from wild-typemice were seen in the number or localization of these cell types inthe lungs of Lungkine2/2 mice (data not shown). The residentleukocyte population in lung tissue was further analyzed by flowcytometry. We did not detect significant differences in the absoluteor relative numbers of distinct leukocytes (CD3-, CD4-, CD8a-,CD11c-, B220-, pan NK-, Mac-1-, F4/80-, and Gr-1-expressingsubsets) between Lungkine2/2 and wild-type animals (n5 10;data not shown).
Lungkine2/2 mice have normal leukocyte subpopulations
To evaluate a possible role for Lungkine in hemopoiesis, leukocytesubpopulations in peripheral blood and bone marrow were ana-lyzed by flow cytometry. Compared with wild-type mice,Lungkine2/2 mice had no significant changes in the absolute orrelative numbers of distinct cell types (CD11c-, B220-, pan NK-,Mac-1-, F4/80-, Gr-1-, CD4-, CD8a-, and CD3-expressing subsets;n 5 14 for bone marrow andn 5 10 for blood; data not shown).Similarly, hematocrits and total white blood cell counts ofLungkine2/2 mice did not differ significantly from those of wild-type mice (n5 15; data not shown).
Lungkine2/2 mice are more susceptible to pneumonia inducedby Klebsiella
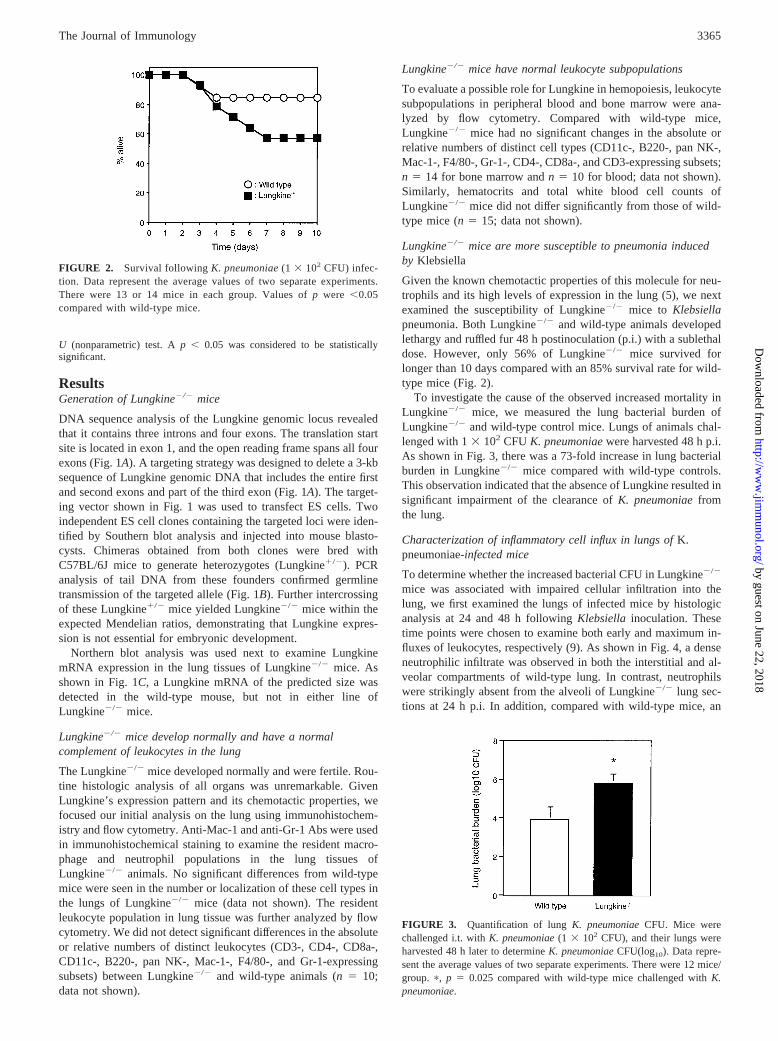
Given the known chemotactic properties of this molecule for neu-trophils and its high levels of expression in the lung (5), we nextexamined the susceptibility of Lungkine2/2 mice to Klebsiellapneumonia. Both Lungkine2/2 and wild-type animals developedlethargy and ruffled fur 48 h postinoculation (p.i.) with a sublethaldose. However, only 56% of Lungkine2/2 mice survived forlonger than 10 days compared with an 85% survival rate for wild-type mice (Fig. 2).
To investigate the cause of the observed increased mortality inLungkine2/2 mice, we measured the lung bacterial burden ofLungkine2/2 and wild-type control mice. Lungs of animals chal-lenged with 13 102 CFU K. pneumoniaewere harvested 48 h p.i.As shown in Fig. 3, there was a 73-fold increase in lung bacterialburden in Lungkine2/2 mice compared with wild-type controls.This observation indicated that the absence of Lungkine resulted insignificant impairment of the clearance ofK. pneumoniaefromthe lung.
Characterization of inflammatory cell influx in lungs ofK.pneumoniae-infected mice
To determine whether the increased bacterial CFU in Lungkine2/2
mice was associated with impaired cellular infiltration into thelung, we first examined the lungs of infected mice by histologicanalysis at 24 and 48 h followingKlebsiella inoculation. Thesetime points were chosen to examine both early and maximum in-fluxes of leukocytes, respectively (9). As shown in Fig. 4, a denseneutrophilic infiltrate was observed in both the interstitial and al-veolar compartments of wild-type lung. In contrast, neutrophilswere strikingly absent from the alveoli of Lungkine2/2 lung sec-tions at 24 h p.i. In addition, compared with wild-type mice, an
FIGURE 2. Survival followingK. pneumoniae(1 3 102 CFU) infec-tion. Data represent the average values of two separate experiments.There were 13 or 14 mice in each group. Values ofp were ,0.05compared with wild-type mice.
FIGURE 3. Quantification of lungK. pneumoniaeCFU. Mice werechallenged i.t. withK. pneumoniae(1 3 102 CFU), and their lungs wereharvested 48 h later to determineK. pneumoniaeCFU(log10). Data repre-sent the average values of two separate experiments. There were 12 mice/group. p, p 5 0.025 compared with wild-type mice challenged withK.pneumoniae.
3365The Journal of Immunology
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
increase in basophilically stained bacterial particles was seen inlungs of Lungkine2/2 mice at 48 h p.i. (Fig. 4,C and D). Tofurther characterize the apparent absence of airspace neutrophils inLungkine2/2 lung at 24 h p.i., we quantified the cell numbers inthe BAL. In uninfected mice no differences in the relative or ab-solute numbers of total BAL cells, neutrophils, or mononuclearcells were noted between wild-type and Lungkine2/2 mice. How-ever, at 24 h p.i., an average 10-fold increase in the numbers ofBAL neutrophils was observed in wild-type mice compared withbaseline, whereas there was no appreciable increase in the numbersof airspace neutrophils in Lungkine2/2 mice (Fig. 5A). In contrast,at 48 h the numbers of BAL neutrophils in Lungkine2/2 mice waseven greater than that in wild-type animals, although this differ-ence was not significant (p 5 0.14). No significant differences innumbers of BAL mononuclear cells were observed at either 24 or48 h following Klebsiellachallenge (Fig. 5B).
Lungkine is produced primarily by airway epithelial cells and issecreted into the airway (5). To determine whether Lungkine pref-erentially affects infiltration of neutrophils into the airway or into
the parenchyma, we infected mice withK. pneumoniaand pre-pared cells from both the lung airspace (BAL) and the parenchyma24 h after bacterial challenge. As shown above, a reduction in thenumbers of infiltrating neutrophils was observed in the airspacecompartment of Lungkine2/2 mice compared with that in wild-type mice. In contrast, higher numbers of neutrophils were ob-served in the parenchyma of Lungkine2/2 mice compared withthat of wild-type mice, although no statistically significant differ-ence was seen between the two groups (Fig. 6). This result sug-gests that a major function of Lungkine is to facilitate migration ofneutrophils from the parenchyma into the airway.
Analysis of lung neutrophil-chemotactic mediators andextrapulmonary neutrophil chemotaxis in Lungkine2/2 mice
We next addressed whether a reduction in airway neutrophils ofLungkine-deficient mice is secondary to impaired production ofother neutrophil chemotactic mediators. Levels of TNF-a, MIP-2,and KC in the lung were measured 24 h p.i. withK. pneumoniae.No significant differences in the levels of these cytokines were
FIGURE 4. Lung histopathology followingK.pneumoniaeinoculation. Hematoxylin- and eosin-stained sections of lung harvested 24 h (AandB)and 48 h (CandD) afterK. pneumoniaeinocula-tion. A and C, Wild-type mice; B and D,Lungkine2/2 mice. In Lungkine2/2 mice, bacteriaparticles are visible under higher magnification at48 h p.i. (arrows ininset of D; magnification,3300). One of two representative experiments isshown.
FIGURE 5. Cell numbers in BAL of wild-type and Lungkine2/2 mice 24 and 48 h follow-ing K. pneumoniaeinoculation. Panels depictBAL neutrophil and mononuclear cell numbersat baseline and 24 and 48 h after i.t.K. pneu-moniaeadministration, respectively. Data repre-sent the average values of four separate experi-ments for neutrophils and two separateexperiments for mononuclear cells. There were 4mice/group for uninfected animals and 10–22/group forKlebsiella-challenged animals.p, p 50.0012 compared with wild-type mice chal-lenged withK. pneumoniae.
3366 LUNGKINE IN HOST DEFENSE
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

seen between lungs of wild-type and Lungkine-deficient animals(data not shown), indicating that the reduced neutrophil influx intothe airway of Lungkine2/2 mice is independent of other neutrophilchemoattractants.
To assess whether the Lungkine deficiency results in a generaldefect in neutrophil mobilization, we measured neutrophil chemo-taxis at an extrapulmonary site. We used a well-characterizedmodel in which LPS is injected into an s.c. air-pouch, (13, 14). Asshown in Fig. 7, at 4 h postinjection, an early time point at whichthe accumulation of neutrophils can be reliably assessed (14), therewas no significant difference in the number of neutrophils betweenwild-type and Lungkine2/2 animals (1.033 107 6 2.93 106 and8.6 3 106 6 2.7 3 106 neutrophils, respectively). This indicatesthat Lungkine is not generally required for neutrophil traffickingand suggests that it is specific to the airspace of the lung.
DiscussionIn this report, we describe the generation and preliminary charac-terization of mice lacking the novel CXC chemokine Lungkine. Inthe adult mouse, Lungkine is produced at appreciable levels onlyby lung epithelial cells and collects in the lung airspace, suggestingthat it might function in pulmonary host defense. We now showthat deletion of the Lungkine gene is associated with diminishedhost defense against the pulmonary pathogenK. pneumoniae.
The rapid clearance of bacterial pathogens from the respiratorytract is mediated by resident alveolar macrophages and neutrophilsthat are recruited from the blood into the airspace (15–17). Thisneutrophil recruitment is mediated by the production in the lung ofchemotactic cytokines (16). ELR1 CXC chemokines, includingMIP-2 and KC, contribute to antibacterial host defense by affectingneutrophil trafficking and activation (9, 11, 18).
The increased mortality in Lungkine2/2 mice following infec-tion with K. pneumoniademonstrates that Lungkine is a chemo-kine that has a central role in pulmonary host defense. Our findingthat the numbers of neutrophils are decreased in the BAL of in-fected Lungkine2/2 mice at 24 h p.i. (but not in the parenchyma)suggests that one function of Lungkine might be to direct neutro-phils from the parenchyma into the airspace. This reduction maybe of notable functional importance, given the key role of neutro-phils in innate defense against bacteria in the respiratory tract thathave not been killed by resident alveolar macrophages (15). Infact, at 48 h p.i., the burden of bacteria was considerably greater inLungkine2/2 mice than in wild-type control animals. The in-creased bacterial load may lead to tissue invasion and dissemina-tion that is not adequately controlled despite the impressive influxof neutrophils seen in Lungkine2/2 mice at this time. A similarincrease in both neutrophils and bacteria was seen in mice lackingthe receptor for neutrophil chemoattractant C5a following infec-tion with Pseudomonas aeruginosa, although the C5a-deficientmice did not display the early deficit in airway neutrophils seen inthe Lungkine2/2 mice (19). This increased neutrophilic contentfound in the airways of both these mouse strains is probably sec-ondary to an increased bacterial load. However, as in all geneti-cally engineered, loss-of-function mice, we cannot exclude thepossibility that other unidentified, compensatory neutrophil attract-ants are up-regulated in Lungkine2/2 mice. Lungkine protein hasbeen shown by others to be secreted into the alveolar space inresponse to inflammatory stimuli (5). This pattern of expressionand secretion may favor directed migration of leukocytes from thevascular and/or interstitial spaces into the airway lumen and alve-olar spaces. If that is the case, it is unclear why so few neutrophilsare normally present within the lung airspace and airway despitethe fact that Lungkine is expressed at high levels in these struc-tures. Apparent paradoxes of this sort are not unique to Lungkine.Other chemokines, such as eotaxin (20, 21), have potent chemoat-tractant roles in vitro and are expressed at high levels in tissues thatare not normally infiltrated by eosinophils, the target cells ofeotaxin. We propose that Lungkine may act as a permissive factor,facilitating the transmigration of neutrophils into the airways dur-ing the early phases of inflammation. For example, Lungkine mayact specifically on activated neutrophils or may work in concertwith undefined molecules that are present only on the surface ofinflamed endothelium. It is also possible that Lungkine, like otherELR1 CXC chemokines, may affect polymorphonuclear leukocyteactivities, including respiratory burst and antimicrobial activities(1, 2, 22). Finally, the site of production (distal airway epithelialcells) and the magnitude of expression show similarities to theexpression of defensin proteins (23), and this raises the possibilitythat Lungkine may have direct antimicrobial properties that are
FIGURE 6. Parenchymal neutrophil cell numbers in the lungs of wild-type and Lungkine2/2 mice 24 h afterK. pneumoniaeinoculation. Datarepresent the average values of two separate experiments. There were 12mice/group.p 5 0.29 compared with wild-type mice challenged withK.pneumoniae.
FIGURE 7. Number of neutrophils in the s.c. air-pouch in wild-type andLungkine2/2 mice after LPS instillation. LPS was injected into the air-pouches in wild-type and Lungkine2/2 mice, and the infiltrating cells werelavaged at 4 h after injection as described inMaterials and Methods. Therewere six mice per group.p 5 0.65 compared with wild-type mice chal-lenged with LPS.
3367The Journal of Immunology
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

independent of its effects on neutrophils. Exploring these mecha-nistic possibilities will probably contribute to a better understand-ing of the role of Lungkine in host defenses and to insights into thebiology of a subset of chemokines expressed constitutively in spe-cific organs.
AcknowledgmentsWe thank Channa Young, Petronio Zalamea, Linda Hamilton, Susan Ab-bondanzo, and Peggy Monahan for excellent technical assistance. Wethank Dr. Lee Sullivan for informative discussion and support throughoutthe course of this work. We also thank the scientists at Schering-PloughClinical Pathology Laboratory for their excellent technical assistance inhematology.
References1. Luster, A. D. 1998. Chemokines: chemotactic cytokines that mediate inflamma-
tion. N. Engl. J. Med. 338:436.2. Rollins, B. J. 1997. Chemokines.Blood 90:909.3. Broxmeyer, H. E., and C. H. Kim. 1999. Chemokines and hematopoiesis. In
Chemokines and Cancer. B. J. Rollins, ed. Humana Press, Totowa, p. 263.4. Broxmeyer, H. E., and C. H. Kim. 1999. Regulation of hematopoiesis in a sea of
chemokine family members with a plethora of redundant activities.Exp. Hema-tol. 27:1113.
5. Rossi, D. L., S. D. Hurst, Y. Xu, W. Wang, S. Menon, R. L. Coffman, andA. Zlotnik. 1999. Lungkine, a novel CXC chemokine, specifically expressed bylung bronchoepithelial cells.J. Immunol. 162:5490.
6. Zlotnik, A., and O. Yoshie. 2000. Chemokines: a new classification system andtheir role in immunity.Immunity 12:121.
7. Ohneda, O., K. Ohneda, H. Nomiyama, Z. Zheng, S. A. Gold, F. Arai,T. Miyamoto, B. E. Taillon, R. A. McIndoe, R. A. Shimkets, et al. 2000.WECHE: a novel hematopoietic regulatory factor.Immunity 12:141.
8. Huffnagle, G. B., M. F. Lipscomb, J. A. Lovchik, K. A. Hoag, and N. E. Street.1994. The role of CD41 and CD81 T cells in the protective inflammatory re-sponse to a pulmonary cryptococcal infection.J. Leukocyte Biol. 55:35.
9. Greenberger, M. J., R. M. Strieter, S. L. Kunkel, J. M. Danforth, L. L. Laichalk,D. C. McGillicuddy, and T. J. Standiford. 1996. Neutralization of macrophageinflammatory protein-2 attenuates neutrophil recruitment and bacterial clearancein murineKlebsiellapneumonia.J. Infect. Dis. 173:159.
10. Greenberger, M. J., R. M. Strieter, S. L. Kunkel, J. M. Danforth, R. E. Goodman,and T. J. Standiford. 1995. Neutralization of IL-10 increases survival in a murinemodel ofKlebsiellapneumonia.J. Immunol. 155:722.
11. Tsai, W. C., R. M. Strieter, J. M. Wilkowski, K. A. Bucknell, M. D. Burdick,S. A. Lira, and T. J. Standiford. 1998. Lung-specific transgenic expression of KCenhances resistance toKlebsiella pneumoniaein mice.J. Immunol. 161:2435.
12. Rovai, L. E., H. R. Herschman, and J. B. Smith. 1998. The murine neutrophil-chemoattractant chemokines LIX, KC, and MIP-2 have distinct induction kinet-ics, tissue distributions, and tissue-specific sensitivities to glucocorticoid regula-tion in endotoxemia.J. Leukocyte Biol. 64:494.
13. Edwards, J. C., A. D. Sedgwick, and D. A. Willoughby. 1981. The formation ofa structure with the features of synovial lining by subcutaneous injection of air:an in vivo tissue culture system.J. Pathol. 134:147.
14. Perretti, M., J. G. Harris, and R. J. Flower. 1994. A role for endogenous histaminein interleukin-8-induced neutrophil infiltration into mouse air-pouch: investiga-tion of the modulatory action of systemic and local dexamethasone.Br. J. Pharmacol. 112:801.
15. Toews, G. B., G. N. Gross, and A. K. Pierce. 1979. The relationship of inoculumsize to lung bacterial clearance and phagocytic cell response in mice.Am. Rev.Respir. Dis. 120:559.
16. Standiford, T. J. 1997. Cytokines and pulmonary host defenses.Curr. Opin.Pulm. Med. 3:81.
17. Broug-Holub, E., G. B. Toews, J. F. van Iwaarden, R. M. Strieter, S. L. Kunkel,R. Paine III, and T. J. Standiford. 1997. Alveolar macrophages are required forprotective pulmonary defenses in murineKlebsiella pneumonia: elimination ofalveolar macrophages increases neutrophil recruitment but decreases bacterialclearance and survival.Infect. Immun. 65:1139.
18. Huang, S., J. D. Paulauskis, J. J. Godleski, and L. Kobzik. 1992. Expression ofmacrophage inflammatory protein-2 and KC mRNA in pulmonary inflammation.Am. J. Pathol. 141:981.
19. Hopken, U. E., B. Lu, N. P. Gerard, and C. Gerard. 1996. The C5a chemoattrac-tant receptor mediates mucosal defence to infection.Nature 383:86.
20. Rothenberg, M. E., A. D. Luster, and P. Leder. 1995. Murine eotaxin: an eosin-ophil chemoattractant inducible in endothelial cells and in interleukin 4-inducedtumor suppression.Proc. Natl. Acad. Sci. USA 92:8960.
21. Ganzalo, J. A., G. Q. Jia, V. Aguirre, D. Friend, A. J. Coyle, N. A. Jenkins,G. S. Lin, H. Katz, A. Lichtman, N. Copeland, et al. 1996. Mouse eotaxin ex-pression parallels eosinophil accumulation during lung allergic inflammation butit is not restricted to a Th2-type response.Immunity 4:1.
22. Frevert, C. W., S. Huang, H. Danaee, J. D. Paulauskis, and L. Kobzik. 1995.Functional characterization of the rat chemokine KC and its importance in neu-trophil recruitment in a rat model of pulmonary inflammation.J. Immunol. 154:335.
23. Ganz, T. 1999. Defensins and host defense.Science 286:420.
3368 LUNGKINE IN HOST DEFENSE
by guest on June 22, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
















![Pulmonary Abnormalities in Mice with ... · We have developed a model of pulmonary PCM in male BALB/c mice induced by the intranasal inoculation of P. brasiliensis conidia [3]. This](https://img.pdfslide.us/doc/110x75/5f84c4aac1fff6621111927e/pulmonary-abnormalities-in-mice-with-we-have-developed-a-model-of-pulmonary.jpg)












