Embed Size (px)
Citation preview

Experimental Parasitology 119 (2008) 336–342
Contents lists available at ScienceDirect
Experimental Parasitology
journal homepage: www.elsevier .com/ locate /yexpr
Cultivation of Trypanosoma cruzi epimastigotes in low glucose axenic mediashifts its competence to differentiate at metacyclic trypomastigotes
Ana R. De Lima a,b, María C. Navarro a, Rosa Y. Arteaga a, Víctor T. Contreras a,b,*
a Laboratorio de Protozoología, Centro BioMolP, Universidad de Carabobo, Bárbula, Valencia, Estado Carabobo, Venezuelab Departamento de Morfofisiopatología, Escuela de Bioanálisis, Facultad de Ciencias de la Salud, Universidad de Carabobo, Bárbula, Valencia, Estado Carabobo, Venezuela
a r t i c l e i n f o
Article history:Received 23 August 2007Received in revised form 3 March 2008Accepted 10 March 2008Available online 15 March 2008
Index Descriptors and Abbreviations:Trypanosoma cruziCompetenceMetacyclogenesisEpimastigotesMetacyclic trypomastigotes
0014-4894/$ - see front matter � 2008 Elsevier Inc. Adoi:10.1016/j.exppara.2008.03.003
* Corresponding author. Address: VLN 1500 P.O. Bo5685, USA. Fax: +58 241 8673342.
E-mail address: [email protected] (V.T. Contre
a b s t r a c t
This study offers an insight into why Trypanosoma cruzi epimastigotes lose their capacity to differentiateinto metacyclic forms, if maintained in culture media long-term through serial passages. The biologicaland metabolic behaviour of two T. cruzi strains isolated from various origins (human, opossum), andmaintained under two schedules (alternate triatomine/mouse passages and serial culture media) werecompared. To determine the effect of the environment on the parasites, the epimastigotes were grownunder extreme conditions (high and low glucose concentrations), and the glucose consumption, ammoniaproduction and changes in pH, either in one compartment (along the growth curve) or two compartments(induced metacyclogenesis) were compared. The glucose effect on the stages involved in metacyclogen-esis at antigenic level was also evaluated. The results indicate that T. cruzi adapts to various environmen-tal conditions and also that the ability of epimastigotes to undergo metacyclogenesis are influenced bythe maintenance schedule. Antigenic profile analysis supports the idea that epimastigotes adapted to cul-ture media do not complete their molecular differentiation into the trypomastigote metacyclic stage.These transition forms conserve some degree of gene expression of the epimastigote stage.
� 2008 Elsevier Inc. All rights reserved.
1. Introduction clogenesis. Moreover, the reservosomes, organelles specific to this
Trypanosoma cruzi, the parasitic protozoan responsible for Cha-gas disease, exhibits at least four stages in its complex life cycle.Amastigotes and blood-stream trypomastigotes are present in themammalian host, whereas epimastigotes and metacyclic trypom-astigotes are in the insect vector (De Souza, 2002). T. cruzi multipli-cation and differentiation in the invertebrate host occurs in distinctcompartments. In the host midgut, the parasite grows and dividesextracellularly, mainly in the epimastigote form in the nutritional-rich environment. Similar forms have been grown readily inculture. The differentiation of epimastigotes into metacyclic try-pomastigotes (metacyclogenesis) occurs at the end of the intestinaltract in which nutrition conditions are decreased (Schaub, 1989;Kollien and Schaub, 2000). Multiplication and differentiation canbe simulated in chemically complex media containing glucose,amino acids, lipids and proteins, in which spontaneous metacyclictrypomastigotes appear as the growth curve plateaus (Camargo,1964). During the exponential phase of the growth curve, epim-astigotes prefer to consume glucose over amino acids if both arepresent (Cannata and Cazzulo, 1984). Conversely, proteins and lip-ids appear to be used as fuel by the epimastigotes during metacy-
ll rights reserved.
x 025685, Miami, FL 33102-
ras).
stage and which contain proteins and lipids that had previouslybeen accumulated during exponential growth, disappear duringepimastigote differentiation into metacyclics (Soares et al., 1989).
In axenic cultures, several factors have been implicated in themetacyclogenesis process; among these are the strain or clone uti-lized (Crane and Dvorak, 1982; Garcia and Dvorak, 1982), the dura-tion of in vitro cultures (Chiari, 1974), addition of insecthaemolymph (Wood and Pipkin, 1969; Wood and Souza, 1976),osmolarity (Adroher et al., 1988a; Osuna et al., 1990), the initialpH of the media (Ucros et al., 1983), the use of L-proline as the solesource of carbon (Contreras et al., 1985; Krassner et al., 1990) andnutritional stress. Metacyclic production in the absence of multi-plication can be induced in two steps: first, growing the epimastig-otes in a nutrition-rich medium; second, stressing the parasites for2 h in a chemically defined medium that simulates the urine of tri-atomine, known as TAU (triatomine artificial urine), and then incu-bating the epimastigotes in TAU supplement containing proline,glutamate, aspartate and glucose (TAU3AAG) (Contreras et al.,1985). These results could only be obtained repetitively with epim-astigotes from the end of exponential growth, suggesting that theparasite requires a competence state to respond to TAU3AAG med-ium (Goldenberg et al., 1984). This competence state has been de-fined as the ability of a heteroxenous parasite to respond tostimulus induced by the environment with morphological trans-formation and appears to be achieved usually after vigorousgrowth (Deane et al., 1984; Vickerman, 1985).

A.R. De Lima et al. / Experimental Parasitology 119 (2008) 336–342 337
There is some evidence of T. cruzi carbohydrate metabolismchanging during the differentiation of the epimastigote into ametacyclic form. Thus, the activity of the key glycosomal enzymesresponsible for glycolysis decreases at the same time as the activityof key enzymes in the tricarboxylic acid cycle increases (Adroheret al., 1988b, 1990). Moreover, the differential expression of prote-ases occurs along the growth curve (Pance and Henriquez, 1992) asmuch as that in the model of two compartments (Bonaldo et al.,1991). Thus, the ability for epimastigotes to respond to nutritionalstress may be related to protein metabolic activation, such thatthey may be able to use their own lipids, amino acids and proteinsas fuel.
In this study, we demonstrated that T. cruzi epimastigotes cul-tured in low glucose axenic media shift its ability to differentiate,thus leading to greater yields of metacyclic trypomastigotes.
2. Materials and methods
2.1. Parasite maintenance and culture conditions
Two isolates of T. cruzi were used throughout this study: a non-cloned (EP) strain, isolated from a fatal human case, and a clone(Dm30L) isolated from opossum. Both parasites were maintainedat least 15 years by alternate triatomine/mouse passages (triato-mine condition) at 27 �C in axenic medium (culture condition), aspreviously described (Contreras et al., 1994). Parasites were cul-tured at 27 �C in LITB medium [5.4 mM KCl, 150 mM NaCl,24 mM glucose, 5% (v/v) liver extract, 0.02% (w/v) hemin, 2% (w/v) yeast extract, 1.5% (w/v) tryptose, 10% calf bovine serum] (Goi-tia-Aular and Boiso, 1982).
Primary cultures contained parasites at a final concentration of2 � 106 cells/ml in 125 ml Erlenmeyer flasks containing 20 ml ofLITB medium in the absence of or supplemented with various glu-cose concentrations (10, 24, 48 mM). Cultures were initiated intriplicate. Growth was monitored by counting in a Neubauerhaemocymetric chamber, and the percentage of metacyclic formswas monitored with Giemsa-stained smears. At least 300 parasiteswere counted per sample.
2.2. Glucose, ammonia and pH estimation
On various days of the growth curve, culture aliquots were con-centrated by centrifuging at 14,000g for 5 min (Eppendorf micro-centrifuge) and the supernatants were used to measure theglucose and ammonia concentration, and pH. We measured glu-cose and ammonia concentrations by glucose oxidase and theBerthelot reaction (Gordon et al., 1978) assays, respectively.
2.3. In vitro differentiation
Metacyclic trypomastigotes were obtained in vitro usingTAU3AAG medium, as previously described (Contreras et al.,1985; Bonaldo et al., 1988). Briefly, epimastigotes, at the endof the exponential growth phase in LITB medium with variousglucose concentrations, were harvested by centrifugation at8500g and incubated for 2 h in artificial triatomine urine(TAU, 190 mM NaCl, 17 mM KCL, 2 mM CaCl2, 2 mM MgCl2,8 mM phosphate buffer pH 6.0), at a density of 5 � 107 cells/ml. The parasites were then incubated in TAU3AA medium(TAU supplemented with 10 mM L-proline, 50 mM L-glutamateand 2 mM L-aspartate) or TAU3AAG (TAU3AA supplementedwith 10 mM glucose) up to a final volume of 70 ml in Rouxflasks with a concentration of 5 � 106 cells/ml. Each inductionexperiment was carried out in triplicate. All parasite sampleswere then incubated at 27 �C for 96 h. The percentage of meta-
cyclic forms was monitored by counting in a Neubauer haem-ocymetric chamber, and by Giemsa-stained smears. Metacyclictrypomastigotes in TAU3AA or TAU3AAG medium were purifiedat the end of the fourth day. The parasites were centrifuged at10,000g, resuspended in TAU medium, treated for 30 min at37 �C with fresh guinea pig serum, and separated on DEAE cel-lulose (Souza, 1983), as previously described (Contreras et al.,1994).
2.4. In vivo glucose consumption in T. cruzi vector intestinal tract
Starving nymphs of 3rd-instars of Rhodnius prolixus were fed torepletion with mice heparinised blood (6.7 mM glucose), using thefeeding apparatus described by Garcia et al. (1984). The glucoseconcentration was determined using the glucose oxidase enzy-matic assay at various times (0, 10, 30 min, 1, 3, 6 and to 12 h) afterfeeding.
2.5. Lysis of parasites
Epimastigotes grown in LITB medium (10 and 48 mM glucose)and purified metacyclic trypomastigotes, obtained from epimastig-otes grown in LITB medium supplemented with 10 and 48 mM glu-cose in TAU3AAG medium, were re-suspended (25 mg/ 100 ll) indeionised water supplemented with a protease inhibitor cocktail:0.1 mM E-64 [trans-epoxysuccinyl-L-leucilamido(4 guanidino)-bu-tane], 0.5 mM TPCK (N-tosil-L-phenylalanine chloromethylketone), 0.5 mM TLCK (Na-p-tosil-L-lysine chloromethyl ketone),25 lM leupeptin, 1 mM 1,10-phenanthroline. The homogenateswere frozen quickly in dry ice-ethanol, thawed in a 37 �C waterbath and vigorously vortexed for 3 min at room temperature. Afterthree cycles of freezing and thawing, the broken parasites werecentrifuged at 14,000g for 10 min at 4 �C. The supernatants werecollected, and were equilibrated to a final concentration of150 mM NaCl. The supernatants were then frozen at �70 �C forstorage until use. The protein content of the supernatants werequantified using the Coomassie Plus assay (Pierce, Rockford,Illinois).
2.6. Western blot analysis
Four microgram of protein from whole cell lysates of T. cruziepimastigotes and metacyclic forms were run on a 10% SDS–poly-acrylamide gel (Laemmli, 1970). Pre-stained, wide range (6.5–205 kDa, Sigma) molecular weight standards were included in eachgel as markers. After the electrophoresis, the gels were equilibratedin transfer buffer [0.25 M Trizma base (Sigma), 192 mM glycine(Sigma) and 20% v/v methanol (Fisher, Pittsburg, P.A)] and trans-ferred to nitrocellulose membrane (Pierce, Rockford, Illinois) at100 V for an hour in a BioRad Trans-Blot� SD unit, according tothe method described by Towbin et al. (1979). The membraneswere blocked in TBS (10 mM Tris/HCl, 150 mM NaCl, pH 7.2) con-taining 5% non-fat milk powder. The membranes were incubatedovernight at 4 �C with a 1:1000 dilution of a hyperimmune rabbitantiserum against the four stages of T. cruzi (anti-EMTA) (Contreraset al., 2002). After eight washes with TBS, membranes were incu-bated (2 h at 37 �C) with 1:2500 dilution of peroxidase-conjugategoat anti-rabbit total immunoglobulins and developed using theLuminol chemiluminiscense system, according to the protocol sup-plied by the manufacturer (Super-Signal, Pierce, Rockford). Finally,the blots were wrapped in plastic wrap and viewed using AGFAfilm and DuPont Cronex-Plus intensifying screens. The immuno-blots were then scanned using a BioRad Imaging Densitometermodel GS-690, and the profiles were analyzed using the BioRadMolecular Analyzer Software Ver. 1.2.
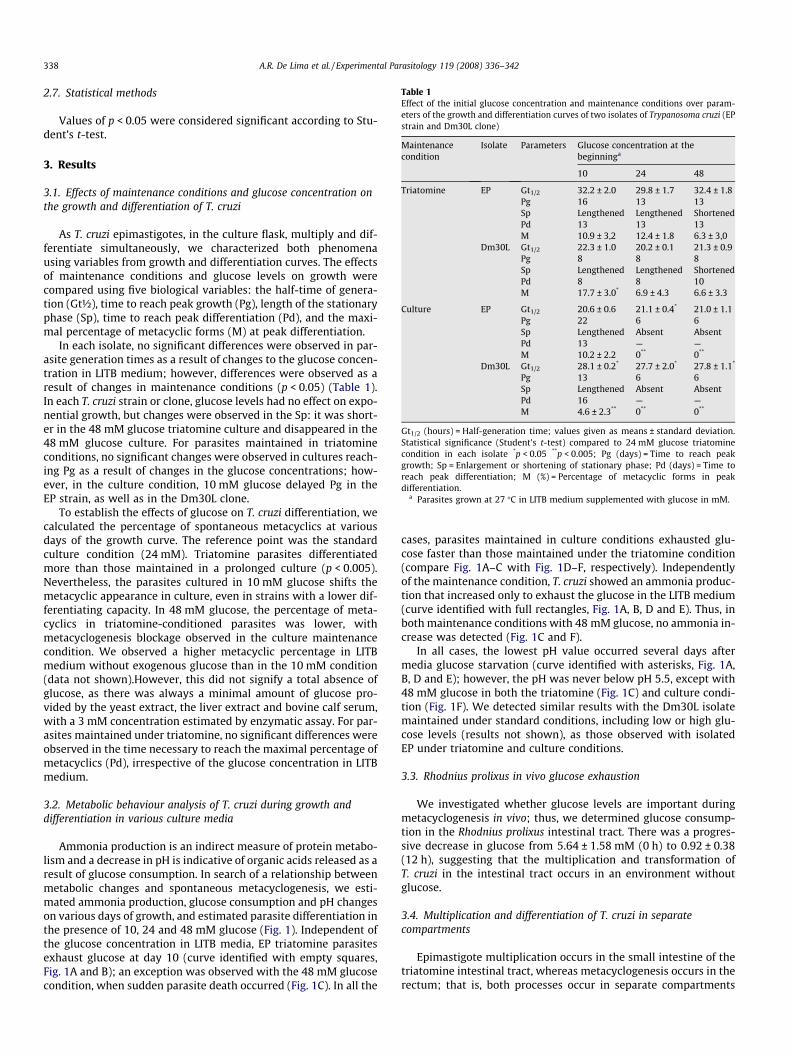
Table 1Effect of the initial glucose concentration and maintenance conditions over param-eters of the growth and differentiation curves of two isolates of Trypanosoma cruzi (EPstrain and Dm30L clone)
Maintenancecondition
Isolate Parameters Glucose concentration at thebeginninga
10 24 48
Triatomine EP Gt1/2 32.2 ± 2.0 29.8 ± 1.7 32.4 ± 1.8Pg 16 13 13Sp Lengthened Lengthened ShortenedPd 13 13 13M 10.9 ± 3,2 12.4 ± 1.8 6.3 ± 3,0
Dm30L Gt1/2 22.3 ± 1.0 20.2 ± 0.1 21.3 ± 0.9Pg 8 8 8Sp Lengthened Lengthened ShortenedPd 8 8 10M 17.7 ± 3.0* 6.9 ± 4.3 6.6 ± 3.3
Culture EP Gt1/2 20.6 ± 0.6 21.1 ± 0.4* 21.0 ± 1.1Pg 22 6 6Sp Lengthened Absent AbsentPd 13 — —M 10.2 ± 2.2 0** 0**
Dm30L Gt1/2 28.1 ± 0.2* 27.7 ± 2.0* 27.8 ± 1.1*
Pg 13 6 6Sp Lengthened Absent AbsentPd 16 — —M 4.6 ± 2.3** 0** 0**
Gt1/2 (hours) = Half-generation time; values given as means ± standard deviation.Statistical significance (Student’s t-test) compared to 24 mM glucose triatominecondition in each isolate *p < 0.05 **p < 0.005; Pg (days) = Time to reach peakgrowth; Sp = Enlargement or shortening of stationary phase; Pd (days) = Time toreach peak differentiation; M (%) = Percentage of metacyclic forms in peakdifferentiation.
a Parasites grown at 27 �C in LITB medium supplemented with glucose in mM.
338 A.R. De Lima et al. / Experimental Parasitology 119 (2008) 336–342
2.7. Statistical methods
Values of p < 0.05 were considered significant according to Stu-dent’s t-test.
3. Results
3.1. Effects of maintenance conditions and glucose concentration onthe growth and differentiation of T. cruzi
As T. cruzi epimastigotes, in the culture flask, multiply and dif-ferentiate simultaneously, we characterized both phenomenausing variables from growth and differentiation curves. The effectsof maintenance conditions and glucose levels on growth werecompared using five biological variables: the half-time of genera-tion (Gt½), time to reach peak growth (Pg), length of the stationaryphase (Sp), time to reach peak differentiation (Pd), and the maxi-mal percentage of metacyclic forms (M) at peak differentiation.
In each isolate, no significant differences were observed in par-asite generation times as a result of changes to the glucose concen-tration in LITB medium; however, differences were observed as aresult of changes in maintenance conditions (p < 0.05) (Table 1).In each T. cruzi strain or clone, glucose levels had no effect on expo-nential growth, but changes were observed in the Sp: it was short-er in the 48 mM glucose triatomine culture and disappeared in the48 mM glucose culture. For parasites maintained in triatomineconditions, no significant changes were observed in cultures reach-ing Pg as a result of changes in the glucose concentrations; how-ever, in the culture condition, 10 mM glucose delayed Pg in theEP strain, as well as in the Dm30L clone.
To establish the effects of glucose on T. cruzi differentiation, wecalculated the percentage of spontaneous metacyclics at variousdays of the growth curve. The reference point was the standardculture condition (24 mM). Triatomine parasites differentiatedmore than those maintained in a prolonged culture (p < 0.005).Nevertheless, the parasites cultured in 10 mM glucose shifts themetacyclic appearance in culture, even in strains with a lower dif-ferentiating capacity. In 48 mM glucose, the percentage of meta-cyclics in triatomine-conditioned parasites was lower, withmetacyclogenesis blockage observed in the culture maintenancecondition. We observed a higher metacyclic percentage in LITBmedium without exogenous glucose than in the 10 mM condition(data not shown).However, this did not signify a total absence ofglucose, as there was always a minimal amount of glucose pro-vided by the yeast extract, the liver extract and bovine calf serum,with a 3 mM concentration estimated by enzymatic assay. For par-asites maintained under triatomine, no significant differences wereobserved in the time necessary to reach the maximal percentage ofmetacyclics (Pd), irrespective of the glucose concentration in LITBmedium.
3.2. Metabolic behaviour analysis of T. cruzi during growth anddifferentiation in various culture media
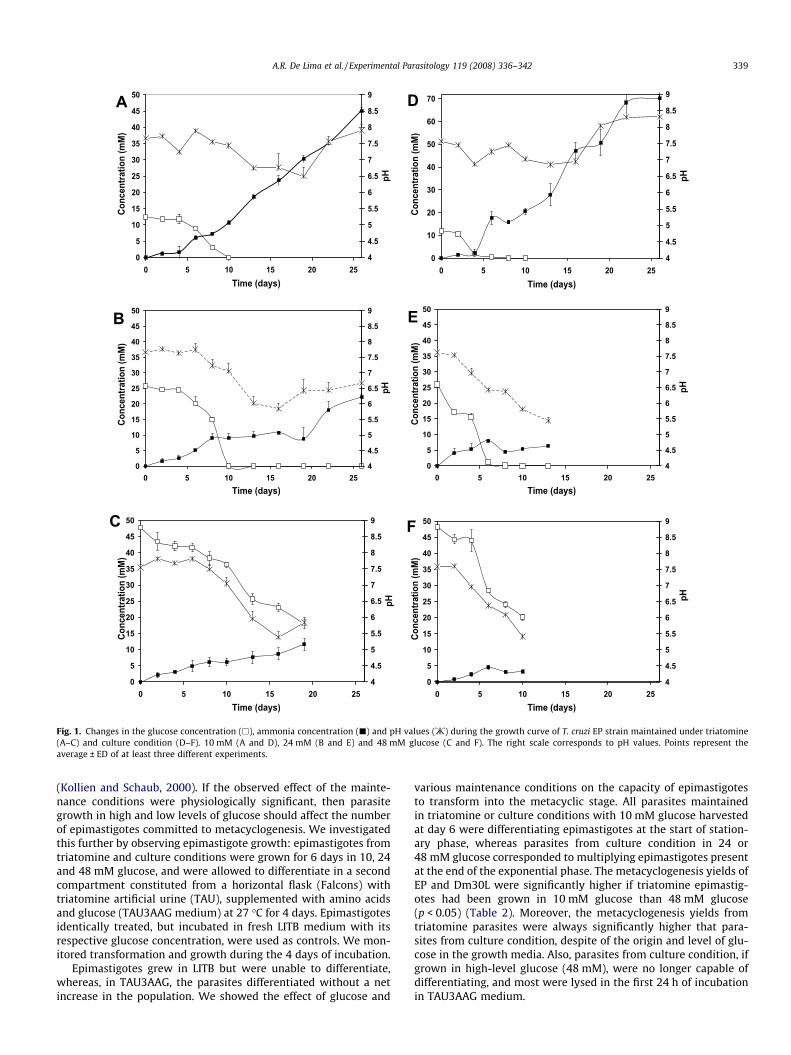
Ammonia production is an indirect measure of protein metabo-lism and a decrease in pH is indicative of organic acids released as aresult of glucose consumption. In search of a relationship betweenmetabolic changes and spontaneous metacyclogenesis, we esti-mated ammonia production, glucose consumption and pH changeson various days of growth, and estimated parasite differentiation inthe presence of 10, 24 and 48 mM glucose (Fig. 1). Independent ofthe glucose concentration in LITB media, EP triatomine parasitesexhaust glucose at day 10 (curve identified with empty squares,Fig. 1A and B); an exception was observed with the 48 mM glucosecondition, when sudden parasite death occurred (Fig. 1C). In all the
cases, parasites maintained in culture conditions exhausted glu-cose faster than those maintained under the triatomine condition(compare Fig. 1A–C with Fig. 1D–F, respectively). Independentlyof the maintenance condition, T. cruzi showed an ammonia produc-tion that increased only to exhaust the glucose in the LITB medium(curve identified with full rectangles, Fig. 1A, B, D and E). Thus, inboth maintenance conditions with 48 mM glucose, no ammonia in-crease was detected (Fig. 1C and F).
In all cases, the lowest pH value occurred several days aftermedia glucose starvation (curve identified with asterisks, Fig. 1A,B, D and E); however, the pH was never below pH 5.5, except with48 mM glucose in both the triatomine (Fig. 1C) and culture condi-tion (Fig. 1F). We detected similar results with the Dm30L isolatemaintained under standard conditions, including low or high glu-cose levels (results not shown), as those observed with isolatedEP under triatomine and culture conditions.
3.3. Rhodnius prolixus in vivo glucose exhaustion
We investigated whether glucose levels are important duringmetacyclogenesis in vivo; thus, we determined glucose consump-tion in the Rhodnius prolixus intestinal tract. There was a progres-sive decrease in glucose from 5.64 ± 1.58 mM (0 h) to 0.92 ± 0.38(12 h), suggesting that the multiplication and transformation ofT. cruzi in the intestinal tract occurs in an environment withoutglucose.
3.4. Multiplication and differentiation of T. cruzi in separatecompartments
Epimastigote multiplication occurs in the small intestine of thetriatomine intestinal tract, whereas metacyclogenesis occurs in therectum; that is, both processes occur in separate compartments
0
10
20
30
40
50
60
70
0 5 10 15 20 25Time (days)
Conc
entra
tion
(mM
)
4
4.5
5
5.5
6
6.5
7
7.5
8
8.5
9
pH
0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20 25Time (days)
Conc
entra
tion
(mM
)
4
4.5
5
5.5
6
6.5
7
7.5
8
8.5
9
pH
0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20 25Time (days)
Conc
entra
tion
(mM
)
4
4.5
5
5.5
6
6.5
7
7.5
8
8.5
9
pH
0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20 25Time (days)
Conc
entra
tion
(mM
)
4
4.5
5
5.5
6
6.5
7
7.5
8
8.5
9
pH0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20 25Time (days)
Conc
entra
tion
(mM
)
4
4.5
5
5.5
6
6.5
7
7.5
8
8.5
9
pH0
5
10
15
20
25
30
35
40
45
50
0 5 10 15 20 25Time (days)
Conc
entra
tion
(mM
)
4
4.5
5
5.5
6
6.5
7
7.5
8
8.5
9
pH
A
B
C
D
E
F
Fig. 1. Changes in the glucose concentration (h), ammonia concentration (j) and pH values (:) during the growth curve of T. cruzi EP strain maintained under triatomine(A–C) and culture condition (D–F). 10 mM (A and D), 24 mM (B and E) and 48 mM glucose (C and F). The right scale corresponds to pH values. Points represent theaverage ± ED of at least three different experiments.
A.R. De Lima et al. / Experimental Parasitology 119 (2008) 336–342 339
(Kollien and Schaub, 2000). If the observed effect of the mainte-nance conditions were physiologically significant, then parasitegrowth in high and low levels of glucose should affect the numberof epimastigotes committed to metacyclogenesis. We investigatedthis further by observing epimastigote growth: epimastigotes fromtriatomine and culture conditions were grown for 6 days in 10, 24and 48 mM glucose, and were allowed to differentiate in a secondcompartment constituted from a horizontal flask (Falcons) withtriatomine artificial urine (TAU), supplemented with amino acidsand glucose (TAU3AAG medium) at 27 �C for 4 days. Epimastigotesidentically treated, but incubated in fresh LITB medium with itsrespective glucose concentration, were used as controls. We mon-itored transformation and growth during the 4 days of incubation.
Epimastigotes grew in LITB but were unable to differentiate,whereas, in TAU3AAG, the parasites differentiated without a netincrease in the population. We showed the effect of glucose and
various maintenance conditions on the capacity of epimastigotesto transform into the metacyclic stage. All parasites maintainedin triatomine or culture conditions with 10 mM glucose harvestedat day 6 were differentiating epimastigotes at the start of station-ary phase, whereas parasites from culture condition in 24 or48 mM glucose corresponded to multiplying epimastigotes presentat the end of the exponential phase. The metacyclogenesis yields ofEP and Dm30L were significantly higher if triatomine epimastig-otes had been grown in 10 mM glucose than 48 mM glucose(p < 0.05) (Table 2). Moreover, the metacyclogenesis yields fromtriatomine parasites were always significantly higher that para-sites from culture condition, despite of the origin and level of glu-cose in the growth media. Also, parasites from culture condition, ifgrown in high-level glucose (48 mM), were no longer capable ofdifferentiating, and most were lysed in the first 24 h of incubationin TAU3AAG medium.

Table 2Metacyclogenesis yield of induced epimastigotes forms of Trypanosoma cruzi (EP,Dm30L) in TAU3AAG medium maintained in triatomine or culture conditions andcultured in LITB media with various glucose concentrations
Glucose concentration inculture media (mM)
Percentage of metacyclic forms (X ± SD)
EP strain maintained Dm30L maintained
Triatomine Culture Triatomine Culture
10 63 ± 3.1* 39 ± 11.0 59 ± 8.1* 32 ± 8.124 42 ± 2.2 18 ± 4.1* 39 ± 9.2 27 ± 6.4*
48 35 ± 1.4 0** 13 ± 3.2* 0**
Epimastigotes were grown for 6 days and harvested by centrifugation, and wereincubated for 4 days in TAU3AAG at 27 �C using equal number of parasites/ml. Thein vitro differentiation was monitored daily by counting cells in a haemocymetricchamber and by differential counting in Giemsa-stained smears. Values representonly metacyclic forms at the end of day 4 and correspond to means ± standarddeviation of at least three experiments. Statistical significance (Student’s t-test)compared to 24 mM glucose triatomine condition in each isolate *p < 0.05,** p < 0.005.
205
116
66
45
29
14.2
6.5
Mr(kDa)
Epi Meta
10 48 10 48
*
**
*
*
**
Fig. 2. Western blot analysis of T. cruzi (EP strain, triatomine condition) using anti-EMTA serum diluted 1/1000. Epi refers to epimastigotes grown in LITB mediumwith 10 and 48 mM glucose (lanes 10 and 48, respectively). Meta indicates purifiedmetacyclic trypomastigotes differentiated in TAU3AAG medium, derived from ep-imastigotes already described (lanes 10 and 48, respectively). Mr corresponds torelative mobility of the molecular weight markers, in kilodaltons (kDa).
340 A.R. De Lima et al. / Experimental Parasitology 119 (2008) 336–342
3.5. Metabolic behavioural analysis of T. cruzi under nutritional stress
We investigated the metabolic performance during differentia-tion in TAU3AAG medium (second compartment). Thus, we deter-mined the glucose consumption, pH change and ammoniaproduction in both isolates maintained under triatomine and cul-ture conditions and grown in LITB media with 10, 24 and 48 mMglucose. Contrary to expectation, no glucose consumption, andnet increase in ammonia or pH decrease was observed. Epimastig-otes incubated in fresh LITB medium were used as a control; theyproduced ammonia and consumed glucose without significantchanges in pH, as previously observed in the culture media. Similarresults to that obtained in TAU3AAG medium were observed in theassays using TAU3AA medium (without glucose) and TAUP med-ium (only 10 mM L-Proline as a carbon source) with the EP strainand Dm30L clone, if grown under standard conditions and condi-tions of low or high glucose levels (results not shown).
3.6. Western blot analysis of metacyclic trypomastigotes derived fromepimastigotes grown in conditions of high and low glucose levels
The glucose concentration in axenic medium indicated largemetabolic changes; thus, we studied changes in expression ofany stage-specific antigens. Total proteins from epimastigotesgrown for 6 days in LITB media with 10 mM and 48 mM glucose,and their derived metacyclics differentiated in TAU3AAG mediumand purified through DE-52, were analysed by Western blot, usinga serum against the four stages of T. cruzi (Anti-EMTA) (Fig. 2).Epimastigotes of the 10 mM condition (Epi-10) showed a patterncharacterised by five faint bands of Mr 150, 125, 116, 41, 29 and27 kDa (dark triangles at left of lane 10, Epi, Fig. 2), three intenseclusters of antigens that were not well-resolved (66/64/62, 55/50and 46/45/43 kDa, asterisk at left of lane10, Epi) and three sharpbands (60, 57 and 31 kDa, arrowheads at left of lane 10, Epi). Wecompared this profile with that displayed by epimastigotes of the48 mM glucose condition (Epi-48); there are significant changesrepresented by the appearance of at least six antigenic bands (42,40, 38, 36, 35 and 33 kDa, arrowheads at left of lane 48, Epi), whichare absent in Epi-10, indicating that this antiserum reveals changesin antigenic expression, despite morphological and polypeptidedifferences between these epimastigotes (results not shown). Inderived-metacyclic trypomastigotes from Epi-10, anti-EMTA ser-um revealed at least five antigens (180, 110, doublet 90/84, 25and 20 kDa, arrowheads at left of lane 10, Meta), which are absentin Epi-10 and Epi-48, suggesting that these antigens are metacy-clic-specific. More significant antigenic differences between the
derived-metacyclic trypomastigotes from Epi-10 and Epi-48(Fig. 2 lane 10 vs. lane 48, Meta) are represented by the absenceof any metacyclic-specific (180, 110, 80, 34 and 25 kDa arrowheadsat left of lane 10, Meta) and common antigens (doublet 64/62, 50,43 and 39/38 kDa, asterisks at left of lane 10, Meta), and also by thepresence of two specific antigens of epimastigote (57 and 55 kDa,arrowheads on the right side of lane 48, Meta). Finally, the anti-genic profiles of derived-metacyclic trypomastigotes from Epi-48,revealed by anti-EMTA serum, resemble the antigenic profiles ofEpi-10 more than the derived-metacyclic trypomastigotes fromthe Epi-10 profile.
4. Discussion
One of the aims of our study was to demonstrate that parasitesmaintained long term in cultures exhibit a lower capacity totransform into metacyclic trypomastigotes, in relation to thosemaintained by alternate invertebrate/vertebrate passages. Never-theless, if parasites maintained in standard culture conditions weregrown in low glucose axenic media, we observed a shift in the per-centage yields of metacyclics, similar to those detected in epim-astigotes recently isolated from vertebrate host. Thus, it ispossible to maintain T. cruzi in a laboratory setting, without alter-nating between invertebrate/vertebrate passages to conservemetacyclogenesis capacity.
Morphologically alternating along of the evolutionary cycle is acharacteristic feature of some important members of the Trypano-somatidae family. Competence has been defined, in parasites that

A.R. De Lima et al. / Experimental Parasitology 119 (2008) 336–342 341
alternate between various host species, as the capacity to respondto stimulus induced by the environment with morphological trans-formation (Deane et al., 1984). Thus, the T. cruzi competence totransform may be influenced by the maintenance schedule in thelaboratory (Table 1). Thus, T. cruzi from a same isolate (cloned ornon-cloned), maintained by alternate triatomine/mouse passagesor by serial passages in axenic medium, demonstrated differentbiological behaviour, in terms of growth and differentiating capac-ity, in response to high and low glucose levels. However, parasitesfrom separate isolates (EP and Dm30L), if maintained under thesame schedule, demonstrated similar biological behaviour.
We suggest that there are two epimastigote populations, phys-iologically different if observing the growth curve: one of multiply-ing parasites adapted to consume glucose; and the other ofdifferentiating parasites, with replicative capacity, and committedto transform spontaneously into the metacyclic form, in responseto nutritional stress. If as observed in this report, the glucose con-centration in the triatomine intestinal tract is very low, then theepimastigotes probably survive using a proteolytic metabolism inthe invertebrate host. By contrast, the glycolytic metabolism al-lows parasites maintained in media supplement with glucose, asthat used routinely in most laboratories, to endure cultureconditions.
Our analysis of glucose consumption, ammonia production andchanges to pH, obtained using one compartment, provide an inter-esting insight into the effect of metabolites produced duringgrowth in metacyclogenesis. Increased acidification (pH 5.5) lysedthe epimastigotes, but metacyclogenesis occurred without acidify-ing the culture medium, suggesting that pH is not a determinantfactor in differentiation, as previously demonstrated (Lanar,1979; Lopetegui and Sosa Miatello, 1982; Ucros et al., 1983). Theaddition of 48 mM glucose to LITB medium eliminated stationaryphase, and caused parasite sudden death. This could be due tothe excessive accumulation of organic acids (Fernandes and Castel-lani, 1966; Caceres and Fernandes, 1976; Cazzulo et al., 1985; Sán-chez-Moreno et al., 1995), causing pH values to decrease below 5.5(Fig. 1).
The fact that parasites maintained in LITB medium with 10 mMglucose exhibit increasing ammonia production simultaneouslywith a shift in metacyclic percentages suggest that parasites grownin media with little glucose increase their commitment to differen-tiate. This might be related to an increased protein metabolism.Accordingly, epimastigotes release large quantities of ammoniato the growth medium after depletion of the available carbohy-drates (Cazzulo, 1994; Urbina, 1994). In addition, the reservosomecontent is massively diminished during metacyclogenesis (Figuei-redo et al., 2004; Cunha-e-Silva et al., 2006) and specific inhibitorsof proteases (Franke de Cazzulo et al., 1994) block the metacyclo-genesis process (Bonaldo et al., 1991).
It became clear from these experiments using two compart-ments (Table 2) that the response of the epimastigotes kept in var-ious maintenance conditions and grown in various levels ofglucose is a measurement of their competence for differentiation.Thus, epimastigotes without the capacity to produce ammoniacannot survive in a medium with similar conditions to triatomineurine. Moreover, epimastigotes that were able to produce high lev-els of ammonia yielded the highest percentages of metacyclics,indicating that these epimastigotes are able to endure an environ-ment suitable for metacyclogenesis. It is worth mentioning thatparasites in differentiation showed no glucose consumption, noammonia production or pH change in TAU3AAG medium, i.e. sup-plemented with amino acids, and glucose at low pH. As observedduring the metacyclogenesis, specific proteases are activated andlipid, amino acid and protein reserves are used as fuel (Bonaldoet al., 1991; Soares et al., 1989); thus, competent epimastigotes
may manage the metabolites accumulated in the reservosomesduring differentiation very efficiently.
We demonstrated the existence of antigenic differences be-tween multiplying and differentiating epimastigotes (Fig. 2, Epi).Metacyclic trypomastigotes obtained from multiplying and differ-entiating epimastigotes differentiated in identical conditions, anddemonstrated significant differences at the antigen level (Fig. 2,Meta), confirming that they were derived of physiologically differ-ent epimastigotes. By contrast, metacyclics obtained from multi-plying epimastigotes exhibited an antigenic profile similar toepimastigotes, indicating that they correspond to transition formsthat have not completed their molecular differentiation processyet. It is worth emphasising that the Western blot analysis wasperformed with metacyclics obtained from TAU3AAG medium,treated with fresh guinea pig serum and purified by chromatogra-phy. Thus, the exhibited profile was not the product of a mixture ofepimastigotes and metacyclics.
In summary, our study shows that T. cruzi epimastigotes main-tained in axenic culture are capable of adjusting their physiologicalstatus depending on the environmental conditions, and are able totransform into metacyclic trypomastigotes. It would still be pre-mature to assume that the events studied here exactly illustratemetacyclogenesis within the invertebrate host, but they may beuseful for generating a new testable hypothesis.
Acknowledgments
This research was supported by Grants from FONACIT S1-2001000683 (MCN, VTC, ARDL), CDCH-FCS-2006006 (MCN, VTC)and CDCH-FCS-2003005 (VTC, MCN, ARDL). We thank Dr. StenioFragoso for his critical suggestions and helpful discussion of themanuscript. We thank Diana Graterol, Wilmer Pineda and JhonnyAlbanesse for their excellent technical assistance.
References
Adroher, F.J., Lupiañez, J.A., Osuna, A., 1988a. Influence of saccharides and sodiumchloride on growth and differentiation of Trypanosoma cruzi. Cell Differentation22, 165–170.
Adroher, F.J., Osuna, A., Lupiañez, J.A., 1988b. Differential energetic metabolismduring Trypanosoma cruzi differentiation. I. Citrate synthase, NADP-Isocitratedehydrogenase and succinate dehydrogenase. Archives of Biochemistry andBiophysics 267, 252–261.
Adroher, F.J., Osuna, A., Lupiañez, J.A., 1990. Differential energetic metabolismduring Trypanosoma cruzi differentiation. II: Hexokinase, phosphofructokinase,and pyruvate kinase. Molecular and Cellular Biochemistry 94, 71–82.
Bonaldo, M.C., Souto-Padrón, T., de Souza, W., Goldenberg, S., 1988. Cell-substrateadhesion during Trypanosoma cruzi differentiation. Journal of Cell Biology 106,1349–1358.
Bonaldo, M.C., D́Escoffier, L.N., Salles, J.M., Goldenberg, S., 1991. Characterizationand expression of proteases during Trypanosoma cruzi metacyclogenesis.Experimental Parasitology 73, 44–51.
Caceres, O., Fernandes, J., 1976. Glucose metabolism, growth and differentiation ofTrypanosoma cruzi. Revista Brasileira de Biologia 36, 397–410.
Camargo, E.P., 1964. Growth and differentiation in Trypanosoma cruzi I. Origin ofmetacyclic trypanosomes in liquid media. Revista do Instituto de MedicinaTropical 6, 93–100.
Cannata, J.J.B., Cazzulo, J.J., 1984. The aerobic fermentation of glucose byTrypanosoma cruzi. Comparative Biochemistry and Physiology 79B, 297–308.
Cazzulo, J.J., 1994. Intermediate metabolism in Trypanosoma cruzi. Journal ofBioenergetics and Biomembranes. 26, 157–165.
Chiari, E., 1974. Growth and differentiation of Trypanosoma cruzi culture forms keptin laboratory for different periods of time. Revista do Instituto de MedicinaTropical de Sao Paulo 16, 61–67.
Contreras, V., Salles, J., Thomas, N., Morel, C., Goldenberg, S., 1985. In vitrodifferentiation of Trypanosoma cruzi under chemically defined conditions.Molecular and Biochemical Parasitology 16, 315–327.
Contreras, V.T., Araque, W., Delgado, V., 1994. Trypanosoma cruzi: metacyclogenesisin vitro. I. Changes in the properties of metacyclics trypomastigotes maintainedin the laboratory by different methods. Memorias do Instituto Oswaldo Cruz 89,253–259.
Contreras, V.T., Navarro, M.C., De Lima, A.R., Arteaga, R., Duran, F., Azkue, J., Franco,Y., 2002. Production of amastigotes from metacyclic trypomastigotes ofTrypanosoma cruzi. Memorias do Instituto Oswaldo Cruz 97, 1213–1220.

342 A.R. De Lima et al. / Experimental Parasitology 119 (2008) 336–342
Crane, M., Dvorak, J., 1982. Trypanosoma cruzi: spontaneous transformation by a Ystrain variant in liquid medium. Experimental Parasitology 54, 87–92.
Cunha-e-Silva, N., Sant́Anna, C., Pereira, M.G., Porto-Carreiro, I., Jeovanio, A.L., deSouza, W., 2006. Reservosomes: multipurpose organelles? ParasitologyResearch 99, 325–327.
De Souza, W., 2002. From the cell biology to the development of newchemotherapeutic approaches against trypanosomatids: dreams and reality.Kinetoplastid Biology and Disease 1, 1–21.
Deane, M.P., Moriearty, P.L., Thomaz, N. 1984. Cell differentiation intrypanosomatids and other parasitic protozoa. In: C.M. Morel (Ed.), Genes andAntigens of Parasites. A Laboratory Manual. UNDP/WORLD BANK/WHO Brasil,pp. 11–21.
Fernandes, J.F., Castellani, O., 1966. Growth characteristics and chemicalcomposition of Trypanosoma cruzi. Experimental Parasitology 18, 195–202.
Figueiredo, R.C., Rosa, D.S., Gomes, Y.M., Nakasawa, M., Soares, M.J., 2004.Reservosome: an endocytic compartment in epimastigote forms of theprotozoan Trypanosoma cruzi (Kinetoplastida, Trypanosomatidae). Correlationbetween endocytosis of nutrients and cell differentiation. Parasitology 129,431–438.
Franke de Cazzulo, B.M., Martínez, J., North, M.J., Coombs, G.H., Cazzulo, J.J., 1994.Effects of proteinase inhibitors on the growth and differentiation ofTrypanosoma cruzi. FEMS Microbiology Letters 124, 81–86.
Garcia, E., Azambuja, P., Contreras, V.T., 1984. Large-scale rearing of Rhodniusprolixus and preparation of metacyclic trypomastigotes of Trypanosoma cruzi. In:Morel, C.M., (Ed.), Genes and Antigens of Parasites. A Laboratory Manual. UNDP/WORLD BANK/WHO, Brasil, pp. 43–46.
Garcia, E.S., Dvorak, J.A., 1982. Growth and development of two Trypanosoma cruziclones in the arthropod Dipetalogaster maximus. American of Journal TropicalMedicine and Hygiene 31, 259–262.
Goitia-Aular, M., Boiso, J.F., 1982. Cultivo de Trypanosoma cruzi (cepa ElpidioPadrón) en un medio semidefinido Efecto de algunas variaciones en lacomposición y condiciones del mismo. Acta Cientifíca Venezolana 33, 488–496.
Goldenberg, S., Contreras, V.T., Salles, J.M., Bonaldo, M.C., De Lima, F., Linss, J.,Lafaille, J., Valle, D., Morel, C., 1984. Facts and hypothesis on Trypanosoma cruzidifferentiation. Memorias do Instituto Oswaldo Cruz 79, 39–44.
Gordon, S.A., Fleck, A., Bell, J., 1978. Optimal conditions for the estimationammonium by the Berthelot reaction. Annals of Clinical Biochemistry 15,270–275.
Kollien, A.H., Schaub, G.A., 2000. The development of Trypanosoma cruzi inTriatominae. Parasitology Today 16, 381–387.
Krassner, S., Granger, B., Phermsangngnam, P., Le, T., Linden, V., 1990. Furtherstudies on substrates inducing metacyclogenesis in Trypanosoma cruzi. Journalof Protozoology 37, 128–132.
Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of thehead of bacteriophage T4. Nature 227, 680–685.
Lanar, D.E., 1979. Growth and differentiation of Trypanosoma cruzi cultivated with aTriatoma infestans embryo cell line. Journal of Protozoology 26, 457–462.
Lopetegui, R., Sosa Miatello, C., 1982. Development and differentiation ofTrypanosoma cruzi in liquid cell-free media. Revista Latinoamericana deMicrobiologia 24, 125–132.
Osuna, A., Adroher, F.J., Lupiañez, J.A., 1990. Influence of electrolytes and non-electrolytes on growth and differentiation of Trypanosoma cruzi. CellDifferentiation and Development 30, 89–95.
Pance, A., Henriquez, D., 1992. Changes in proteolytic activity during the growth ofTrypanosoma cruzi epimastigotes. Biochemistry International 27, 613–623.
Sánchez-Moreno, M., Fernández-Becerra, M.C., Castilla-Calvente, J.J., Osuna, A.,1995. Metabolic studies by1 H NMR of different forms of Trypanosoma cruzi asobtained by ‘‘in vitro” culture. FEMS Microbiology Letters 133, 119–125.
Schaub, G.A., 1989. Trypanosoma cruzi: quantitative studies of development of twostrains in small intestine and rectum of the vector Triatoma infestans.Experimental Parasitology 68, 260–273.
Soares, M.J., Souto-Padrón, T., Bonaldo, M.C., Goldenberg, S., de Souza, W., 1989. Astereological study of the differentiation process in Trypanosoma cruzi.Parasitology Research 75, 522–527.
Souza, M.A., 1983. Surface electrical charge of bloodstream trypomastigotes ofTrypanosoma cruzi strains. Memorias do Instituto Oswaldo Cruz 78, 497–500.
Towbin, H., Stachelin, T., Gordon, J., 1979. Electrophoretic transfer of proteins frompolyacrylamide gels to nitrocellulose sheets: procedure and some applications.Proceedings of the National Academic of Sciences USA 79, 4350–4354.
Ucros, H., Granger, B., Krassner, S.M., 1983. Trypanosoma cruzi: effect of pH on‘‘in vitro” formation of metacyclic trypomastigotes. Acta Tropica 40, 105–112.
Urbina, J.A., 1994. Intermediary metabolism of Trypanosoma cruzi. ParasitologyToday 10, 107–110.
Vickerman, K., 1985. Developmental cycles and biology of pathogenictrypanosomes. Nature 208, 762–766.
Wood, D.E., Pipkin, C.A., 1969. Multiplication and differentiation of Trypanosomacruzi in an insect cell culture system. Experimental Parasitology 24, 176–183.
Wood, D.E., Souza, O.E., 1976. Trypanosoma cruzi: effects of Rhodnius prolixusextracts on in vitro development. Revista do Instituto de Medicina Tropical deSao Paulo 18, 93–96.





























