Embed Size (px)
Citation preview

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
1
TID
SK
RIF
T
Kungl. Skogs- ochLantbruksakademiensÅrg. 142 • Nr 13 • År 2003
Crop and Forest Biotechnologyfor the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
2
Ansvarig utgivare: Akademiens sekreterare och VD: Bruno NilssonRedaktör: Gunilla Agerlid

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
3
Crop and Forest Biotechnologyfor the Future
International Conference in FalkenbergSeptember 17–19, 2001
in cooperation withSwedish Foundation for Strategic Research
andBertebos Foundation
Maria Larsson
Redovisningen sammanställd under medverkan avakademiråd Agnetha Alriksson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
4

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
5
Table of Contents
Introduction ................................................................................................................................ 7
Genetically modified organisms in food and agriculture: Where are we? Where are we going?Lecturer: Louise Fresco, Ass. Director General, FAO, Rome, Italy .............................................. 8
Ripening ...................................................................................................................................... 9Control of fruit ripening and quality
Lecturer: Prof Donald Grierson, Nottingham University, UK ............................................ 9Better keeping qualities
Lecturer: Dr. Jean-Claude Pech, INRA-ENSAT, Toulouse, France ................................... 10
Fatty acid modification ............................................................................................................ 11Desaturation of fatty acids
Lecturer: Prof. Ernst Heinz, Institut für allgemeine Botanik, Hamburg, Germany ......... 11Three milestones in oil biosynthesis
Lecturer: Dr. Ulf Ståhl, Swedish University of Agricultural Sciences, Uppsala, Sweden 12
Starch ........................................................................................................................................ 13Designing starch
Lecturer: Dr Alison Smith, John Innes Centre, Norwich, UK .......................................... 14The starch synthesis in barley endosperm
Lecturer: Prof. Christer Jansson, Swedish University of Agricultural Sciences,Uppsala, Sweden ............................................................................................................ 15
Genomics, proteomics and metabolomics ............................................................................. 1625 500 genes in the Arabidopsis genome
Lecturer: Prof. Michael Bevan, John Innes Centre, Norwich, UK ................................... 16Brassica differs from Arabidopsis and Capsella
Lecturer: Dr Renate Schmidt, Max-Delbreuch-Laboratorium in der MPG; Germany ..... 17Analyse 14 000 clones in a microarray
Lecturer: Prof. Ellen Wisman, Michigan State University, USA ...................................... 18Study the change in protein quantity
Lecturer: Dr. Kathryn S. Lilley, University of Cambridge, UK ......................................... 19Metabolic profiling: A metabolic engineering perspective
Lecturer: Dr. Alisdair Fernie, Max-Planck-Institute, Potsdam, Germany ........................ 21
Plant stress and signalling ....................................................................................................... 22Hormonal regulation of oxidative cell death
Lecturer: Prof. Jaako Kangasjärvi, University of Turku, Finland ................................... 22Defence responses to Leptosphaeria maculans
Lecturer: Dr. Christina Dixelius, Swedish University of Agricultural Sciences,Uppsala, Sweden ............................................................................................................ 23
Wound responses and insect defenceLecturer: Dr. Johan Meijer, Swedish University of Agricultural Sciences,Uppsala, Sweden ............................................................................................................ 25
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
6
Regulation of the plasma membrane H+-ATPase by phosphorylation and 14-3-3Lecturer: Prof. Marianne Sommarin, Lund University, Sweden ..................................... 26
Forest genetics ......................................................................................................................... 27Genetic engineering improves poplars
Lecturer: Dr. Wout Borjean, University of Ghent, Belgium ............................................. 27Functional genomics of wood formation
Lecturer: Prof. Göran Sandberg, Swedish University of Agricultural Sciences,Umeå, Sweden ............................................................................................................... 28
Enzymes for making and breaking wood fibresLecturer: Prof. Tuula Teeri, Royal Institute of Technology, Stockholm, Sweden ........... 29
The longer night length influences the phytochromeLecturer: Dr. Stefan Jansson, Umeå University, Sweden ................................................ 30
Plant development ................................................................................................................... 31A conifer perspective on reproductive organ development in plants
Lecturer: Prof. Peter Engström, Uppsala, University ...................................................... 32Regulation of cambial development
Lecturer: Dr. Rishikesh Bhalerao, Swedish University of Agricultural Sciences,Umeå, Sweden ................................................................................................................ 32
Members of the SHI gene family are involved in the control of cell expansionLecturer: Dr. Eva Sundberg, Uppsala University, Sweden .............................................. 33
The hidden half: Dissecting the genetic and hormonal control of root developmentLecturer: Prof. Malcolm Bennett, Nottingham University, UK ........................................ 35
Transgenics: Not “if” but “why” ............................................................................................... 35Where are we heading?....................................................................................................... 36But what is a transgenic?.................................................................................................... 36Stable or unstable? ............................................................................................................. 36Antisense techniques ......................................................................................................... 36What can be learnt from Arabidopsis thaliana .................................................................. 37
Food or non food applications ................................................................................................ 37More and better food to feed the world ............................................................................ 37Replace mineral oils with vegetable oils ........................................................................... 38Energy in the field ............................................................................................................... 38Designing starch ................................................................................................................. 38Forest engineering makes wonder ..................................................................................... 38
Science, media and the consumers......................................................................................... 39Consumers are concerned ................................................................................................. 39Legislation is needed .......................................................................................................... 39
Conclusions .............................................................................................................................. 39
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
7
and the Swedish Foundation for StrategicResearch (SSF). This is the brief backgroundto the current International Conference onCrop and Forest Biotechnology for the Future.
The conference focused on what biotech-nology science can do for the long-term sup-ply of agricultural and forestry products. De-spite the absence of four speakers from theUS and Australia (who were unable to comedue to the events in New York on September11) the programme was broad, thorough andinspiring. It is an intriguing task to documentthis conference properly and completely. Themost important result of the scientific work,as Prof. Donald Grierson put it, is the increasedunderstanding of how it all works, but ofcourse it is also a challenge to be able to adaptthis increased knowledge in practical horti-culture, agriculture and forestry.
Introduction
Olof and Brita Stenström have establishedthe Bertebos Foundation in order to promoteresearch, development and education withinthe food industry. Through the auspices ofKSLA, the Foundation has established a pres-tigious prize, the Bertebos prize. In 2001 thisprize was awarded to Professor Donald Grier-son, Nottingham University, UK, for his pio-neering research on control of fruit ripeningand quality. The Bertebos Foundation alsoholds a seminar in Falkenberg, close to Slö-inge, where the Stenström family have theirproperty, Berte Mill, ”SIA Ice Cream” and afarm. In the seminar, the prize winner and oth-er scientists from around the world gather aslecturers and participants. The 2001 confer-ence was organised by the Bertebos Founda-tion in co-operation with the Royal SwedishAcademy of Agriculture and Forestry (KSLA)
Olof Stenström, Donald Grierson och Brita Stenström. Photo: Maria Larsson.
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
8
Genetically modifiedorganisms in food andagriculture: Where arewe? Where are we going?A transgenic plant has just one or a few genesaltered and this generates a new variety withdifferent properties. One gene might influ-ence the aroma and other characteristics thatseemed to be independent of the desired he-reditary character. One fact that makes thetransgenic plant different from a plant devel-oped with traditional plant breeding is thatthe transgenic plant can contain genes fromany living being, while the traditionally bredplant can receive hereditary characters onlyfrom plants from the same species.
It is easy to recognise the great potential aswell as the complications with geneticallymodified organisms, GMOs. Louise Fresco,Asst. Director General of the FAO, claimedthat the FAO needs to assess GMOs in termsof their impact on food security, poverty, bio-safety and the sustainability of agriculture.Will GMOs increase the amount of food in theworld and make more food accessible to thehungry? She wanted to discuss not the speci-fics of GMO technology, but the context inwhich they are developed and deployed andhow public opinion and government policyon GMOs are formed.
In most countries there is no consensus onhow biotechnology and GMOs in particularcan focus on the key challenges of the foodand agricultural sector. Scientists see geneticmodification as a major new set of tools, theindustry looks at GMOs as opportunities forcorporate profit (but with public acceptanceas a stumbling block) and governments oftenlack coherent policies in relation to GMOs.
The FAO has been conducting a world-wide inventory of agricultural biotechnologyapplications and products, with special refer-
ence to developing countries, to test the hy-pothesis that biotechnology applications incrop science – particularly GM crops – havenot addressed the needs of developing coun-tries.
The total area cultivated with GMO cropsin the world is rapidly growing and currentlystands at 44.2 million hectares. About 75% ofthe area planted with GM crops is in indust-rialised countries. Around 16% of the acreageof the four crops soybean, maize, cotton andcanola is under GM varieties. Two traits – in-sect resistance (mainly based on Bt) and her-bicide tolerance – dominate.
Only seven developing countries commer-cially cultivate GM crops. The dominant cropsare soybean and cotton and the traits are her-bicide tolerance and insect resistance. Onlyin China was one GM cotton locally devel-oped and commercialised. Other countriesobtained genetic constructs or varieties fromindustrialised countries.
However, to ensure that GM crops make anoptimal contribution to world food security,food safety and sustainability, three greatproblems must be solved. Firstly, GMOs mustbe directed to the right research prioritiessuch as drought and heat tolerance, im-proved nutrient uptake and rooting, biologi-cal nitrogen fixation, responses to carbon di-oxide and tolerance to key abiotic stresses,such as salinity and drought. Secondly, agreednational and international instruments of gov-ernance must be developed. Thirdly, the ac-cess to research and new technologies fordeveloping countries, poor producers andconsumers must be facilitated.
Louise Fresco’s speech formed a back-ground to the scientific achievements thatwere presented during the seminar. Little ofthe research has been done to lighten theburdens of the poor, but many achievementscan be utilised to develop a sustainable andsecure food chain in poor countries.
Read Louise Fresco’s full speech at http://www.fao.org/ag/magazine/GMOs/pdf
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
9
Ripening
Ripening and senescence go hand in hand butby delaying senescence in fruits and vegetab-les the sustainability is prolonged, which is anadvantage to producers, trade and consum-ers.
Control of fruit ripening andqualityTomato fruit development and ripening is in-fluenced by the growth hormone ethylene(ethene). Twenty five years ago, Prof. DonaldGrierson and his research team at Notting-ham University and other scientists began tostudy genes that regulate ripening. One ofthese genes encoded a receptor for ethylene,which detects the presence of the hormone infruit cells that switch on the ripening process.They later found that ethylene activates acascade of genes that coordinate many bio-logical changes and thereby affect fruit qual-ity: colour, flavour, texture and even the rateof ripening in the tomato. Similar changesoccur in many fruits.
Having identified some of the genes ex-pressed during ripening, Prof. Grierson andhis team wanted to find out whether theycould control their expression with geneticmodification techniques. They found that byusing the antisense technique or sense genesilencing (co-suppression), messenger RNA(mRNA) and enzyme activity for polygalactu-ronase, phytoene synthase and ACC oxidase,and other enzymes could be reduced. Thismodified texture, changed succulence andcolour, and demonstrated that senescencecould actually be delayed if expression of theright genes was modified.
This is a very precise method and only onegene out of tens of thousands is inactivated.The effect is related to the specific antisensegene that is used and it is inherited. From
tomatoes with the antisense gene, progenytomatoes can be obtained that have a de-layed ripening process or altered quality. Thismeans that a new variety is created. Around1990, Grierson found that if this was donewith a sense gene, a similar knockout effectwas obtained. The results of the two proce-dures are very similar, so there must be acommon mechanism between sense geneactivation and antisense gene activation.
The key point is that if ethylene productionor the response to ethylene can be control-led, the rate at which produce progresses tooverripeness and spoilage can be regulated.Between 20 and 50% of all fresh produce (in-cluding tomatoes) deteriorates before it reach-es the consumer. Preventing this deteriora-tion is an advantage to the producers, tradeand consumers. The synthesis of ethylenerequires a short biochemical pathway involv-ing a compound found in all living cells calledSAM (S-adenosyl methionine). To make ethyl-ene the plant has to control two steps, first toconvert SAM to ACC (1- amino-cyclo-propane-1 carboxylic acid); then to convert ACC toethylene. Ethylene causes ripening and se-nescence in leaves, flowers and fruit and oncethe process is started, it is very difficult tostop. A low ethylene tomato, however, has aslower deterioration process and is thereforeedible during a longer period.
A number of genes are needed to controldifferent aspects of the ripening process. Theexpression by GM procedures affecting onegene can be inhibited without interfering withanother and some interesting and potentiallyvery useful biotechnological applications canbe obtained.
To summarise, Prof. Grierson and his col-leagues have found more than 25 ripeningrelated genes and are beginning to under-stand the role of several of these genes incontrolling processes like texture and succu-lence, pro-vitamin A contents, storage andpost-harvest deterioration and ethylene.
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
10
Better keeping qualitiesThe methods used in Nottingham have beenfurther developed and adapted to producecantaloupe melons with better keeping qual-ities.
Cantaloupe Charentais melons are verytasty, sweet French melons with good flavourbut with poor keeping qualities. They over-ripen quickly and are chilling sensitive. Thecantaloupe Charentais melon therefore repre-sented a good target fruit for improving stor-ability through biotechnology.
Melons, bananas, pears and peaches and,of course, tomatoes belong to a climacterictype of fruit where ethylene plays a majorrole. They are characterised by a rise in res-piration during ripening associated with aburst of ethylene production. In addition, eth-ylene can stimulate its own biosynthesis (au-tocatalysis). There is another type of fruitsuch as grapes and pineapples in which eth-ylene does not seem to play a major role butscientists have not yet identified the otherfactor that plays that major role, according toDr. Jean-Claude Pech, INRA-ENSAT in Tou-louse.
In order to increase the storage life of mel-ons, he and his colleagues decided to try toslow down ripening by inhibiting ethyleneproduction. They isolated the melon ACCoxidation gene (that Donald Grierson had dis-covered in tomato) and put it in antisenseorientation. Ethylene was inhibited almost to100% in the antisense melon in a way thatgave a significant difference in storability be-tween the antisense (AS) and the wild type(WT) melon. After 15 days of storage at 25°C,the WT was not edible any longer while theAS type had retained firmness and some otherquality attributes. By comparing the compo-sition of the two types of fruit, Dr Pech andhis colleagues were able to show that sugar
accumulation and the colour of the flesh weresimilar and therefore corresponded to ethyl-ene independent processes. On the otherhand, the colour of the rind and the formationof the peduncular abscission zone wereethylene-dependent processes. Softening ofthe flesh has two components, dependentand independent of ethylene. These data sug-gest that climacteric fruit ripening involvesethylene-independent events and thereforecomprises some non-climacteric regulation.
By submitting AS fruit to ethylene, INRA-ENSAT scientists found that the various ripen-ing pathways were sensitive to levels of ethyl-ene (2.5 to 5 ppm) that are much lower thaninternal ethylene present in ripe fruit (80 to100 ppm). Attached fruit had a higher rate ofethylene production than detached fruit butrespiration was lower in attached fruit.
Wild type melons showed chilling injuryafter storage at low temperatures (2 weeks at2°C). The injury was particularly visible uponrewarming. In contrast, the AS melon showedvery little damage, demonstrating that ethyl-ene was a stimulator of chilling injury.
Besides providing a tool for understandingthe mechanisms of fruit ripening, these trans-genic melons could give rise to new post-harvest handling methods:
• Harvesting all fruit at the same time in thefield (possibly mechanically) when all ofthem have reached sufficient sensoryqualities without risk of over-ripening.
• Storing at low temperature without chill-ing damage.
• Shipping to distant markets, treating withadapted levels of ethylene.
• Sorting for quality.• Delivery to the consumer of fruit at an op-
timum sensory quality.
However, such a scheme needs to be valida-ted at the practical level.
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
11
Fatty Acid Modification
Vegetable oils are important commercial pro-ducts, of which about 90% are used for humanconsumption. Non-food applications includesoap and detergents, paints, lubricants andfuel (bio-diesel). The use is dependent on thequality, which in turn depends on the fattyacid composition. Although several hundredfatty acids of different structures exist, it islikely that only about 50 enzymes/genes areinvolved in the synthesis of the most impor-tant fatty acids. The annual oilseed crops yieldfrom 0.4–1.8 tons of oil per hectare, which isfar exceeded by oil palm with up to 7 tons/ha/year. The annual global production/consump-tion of plant oils adds up to about 100 miotons compared to about 20 mio tons of animalfats. All edible oils together would result inabout 40 g oil/person/day given a global po-pulation of 6 billion people.
Desaturation of fatty acidsIn industrialised countries, about three timesas much oil, 120 g, is consumed per personper day despite the fact that for years medicaland health authorities have been recommend-ing a reduction in this consumption to about60 g per person per day, which correspondsto about 30% of the recommended daily 2000kcalories. In addition, the oils prevailing inpresent-day food items of the Western worldare characterised by a predominance of sat-urated and at most diunsaturated linoleicacid. A healthy and balanced mixture of vari-ous fatty acids, particularly considering lino-leic (ω-6) and linolenic acid (ω-3) in a ratio ofabout 5:1, is not provided. These two polyun-saturated fatty acids cannot be synthesizedby man and, therefore, are essential fatty ac-ids required for the biosynthesis of variouseffectors controlling human physiology onvarious occasions and at different points. The
recommended daily 2 g of linolenic acid arecontained in about 500 g of green vegetablefresh weight, whereas this ω-3-fatty acid ishardly found in any of the other food itemsappreciated by all of us for their conveniencein storage and preparation.
From a biochemical point of view the dailyconsumed fatty acids should include about10 g of linoleic and 2 g of linolenic acid and asignificant reduction in all saturated fatty acidsand a corresponding increase in oleic acid.Therefore, a reduction in cholesterol-contain-ing animal (dairy) fats and the use of choles-terol-free plant oils whenever possible can berecommended. With respect to these criteria,in particular due to the presence of linoleicand linolenic acid in a ratio of about 3:1, rape-seed oil is superior to olive and sunfloweroils, since both do not contain the essentiallinolenic acid. Since we all can hardly avoidthe flooding of our daily diet by saturated andat most diunsaturated (ω-6, but not ω-3) lino-leic acid, Prof. Ernst Heinz, Institut für allge-meine Botanik in Hamburg, recommends twoservings of fat (not lean!) fish per week. Thisprovides the C20- and C22-ω-3- and ω-6-long-chain polyunsaturated fatty acids that are thedirect effector precursors made by the hu-man body from dietary linoleic and linolenicacid.
In view of this situation it is an obviousidea to produce plant seed oils containingthese very long-chain polyunsaturated fattyacids normally only found in fish (”oceanic“fatty acids). This goal may be realised by genetechnology provided that the biochemistry ofoil biosynthesis is known in detail. Also, allthe required genes and the transformationtechnique to introduce these genes into annu-al oilseed crops must be at hand. Severalgroups world-wide have concentrated on thistransgenic approach, because it would:• Provide a healthy oil beneficial for human
nutrition.• Represent a consumer-orientated trans-
genic product.
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
12
• Guarantee a sustainable production.• Protect the marine resources from over-
fishing.
At the end of the year 2002, most genes requi-red for this task have been cloned and onlyone is still missing. The oilseeds of potentialuse for this project include rapeseed, soybe-an, linseed, sunflower and peanut, which allcan be genetically modified. The most suc-cessful approach so far published has resul-ted in canola oil with 44% γ-linolenic acid,which is a good basis for the implementationof the final reactions leading to C20- and C22-polyunsaturated fatty acids.
Seed oils of higher plants contain threepredominant fatty acids: oleic, linoleic andlinolenic acid, which all have 18 carbon at-oms and from one to three double bonds.These double bonds are introduced by desat-urase enzymes, in which two large familiesexist in all plants: soluble enzymes for thefirst double bond and membrane-bound de-saturases for the following double bonds. Thegene-technologically most relevant enzymesbelong to the latter group. In addition to theprevailing cis-desaturation, members of thisgroup catalyse a wide variety of reactionssuch as trans-desaturation, hydroxylation,epoxidation, conjugation and decarbonyla-tion. The acyl groups used as substrates arethioesters of coenzyme A or oxygen esters inglyco- and phospholipids. These seeminglyunrelated reactions are due to the versatilityof a di-iron complex believed to be bound inthe active site of these enzymes. All the de-saturases required for the above-mentionedproject have been cloned from various organ-isms including algae, mosses and fungi. Butapart from the desaturases, two elongasesare required for converting polyunsaturatedC18- to C20- and finally to C22-fatty acids.From these, the final elongase is still not avail-able in cloned form, blocking the productionof polyunsaturated C22-fatty acids.
Three milestones in oilbiosynthesisThe fatty acids (FA) found in available vegeta-ble oil qualities are:• The five common fatty acids: 16:0 (palmi-
tic), 18:0 (stearic), 18:1 (oleic), 18:2 (lino-leic) and 18:3 (linolenic), found in oilpalm,canola, sunflower, soya bean and linseed.
• Exotic fatty acids: 10:0 (capric), 12:0 (lau-ric) and 14:0 (myristic), found in palm ker-nel and coconut, 22:1 (euric acid) found inhigh erucic rape.
There are more than 200 exotic fatty acidstructures identified in nature and many ofthese can be found in high levels, up to 90% ofFA in natural vegetable oils. The chain lengthsdiffer from C8 to C24 fatty acids. The desatura-tion can be at odd positions, trans, conjugatedor triple. The substitutions can be hydroxy,epoxy, furanoid or keto. Exotic fatty acids canhave branched, cyclic or fluoro modifications.They are used already today by the industrybut many more have potentially interestingindustrial applications. Most of the exotic fat-ty acids are naturally produced by plants thatare not suitable for modern agriculture sys-tems. However, biotechnology can in the futu-re be used to design a desired oil quality in adesired oil crop. A desired fatty acid can beobtained with the usage of the big gene libra-ry in nature or by protein engineering.
The first milestone in the scientific chal-lenge is to obtain a general understanding oflipid biosynthesis and metabolism in plantsand especially in developing oil seeds. Oilseeds are highly specialised plant organs asregards their glycerolipid composition. Oilseeds contain 1% plastidial galactolipids, 1%plastidial phospholipids, 3% cytosolic phos-pholipids and 95% tricyglycerols.
The second milestone in the scientific chal-lenge is to clone genes for the biosynthesis ofexotic fatty acids. There is already a long listof cloned genes and it is rapidly growing.
M. Larsson
K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
13
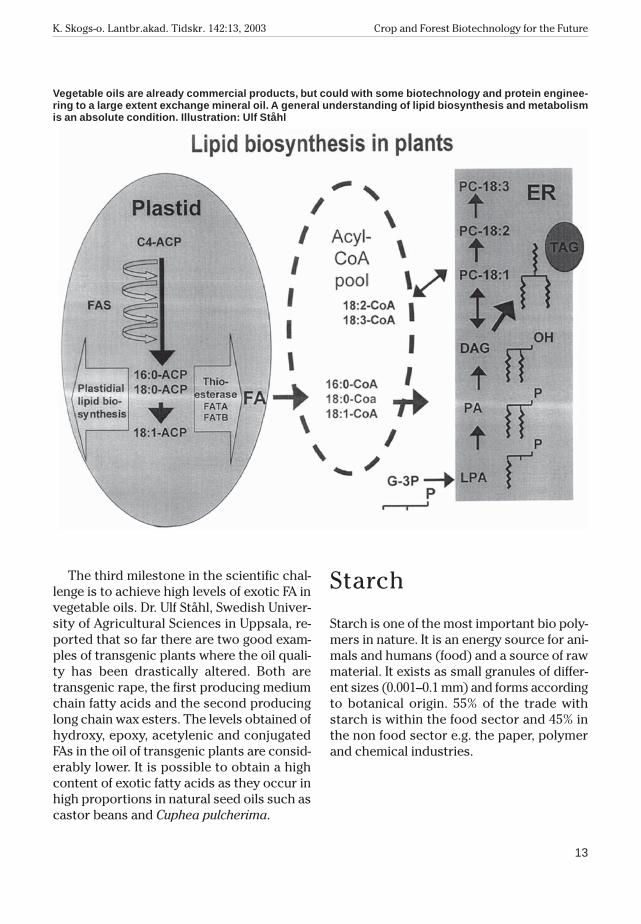
The third milestone in the scientific chal-lenge is to achieve high levels of exotic FA invegetable oils. Dr. Ulf Ståhl, Swedish Univer-sity of Agricultural Sciences in Uppsala, re-ported that so far there are two good exam-ples of transgenic plants where the oil quali-ty has been drastically altered. Both aretransgenic rape, the first producing mediumchain fatty acids and the second producinglong chain wax esters. The levels obtained ofhydroxy, epoxy, acetylenic and conjugatedFAs in the oil of transgenic plants are consid-erably lower. It is possible to obtain a highcontent of exotic fatty acids as they occur inhigh proportions in natural seed oils such ascastor beans and Cuphea pulcherima.
Starch
Starch is one of the most important bio poly-mers in nature. It is an energy source for ani-mals and humans (food) and a source of rawmaterial. It exists as small granules of differ-ent sizes (0.001–0.1 mm) and forms accordingto botanical origin. 55% of the trade withstarch is within the food sector and 45% inthe non food sector e.g. the paper, polymerand chemical industries.
Vegetable oils are already commercial products, but could with some biotechnology and protein enginee-ring to a large extent exchange mineral oil. A general understanding of lipid biosynthesis and metabolismis an absolute condition. Illustration: Ulf Ståhl
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
14
Designing starchStarch is the major form in which plants storetheir carbon – around 80% of the dry weightin potato tubers and the endosperm of cere-als and up to 50% for peas and beans. Starchis the main source of carbohydrates in ourdiet but it also serves as a thickener, texturiz-er or stabilizer in both the food and non foodindustries.
Starch extracted from the crop (maize,wheat, potato, cassava etc.) undergoes post-extraction processing consisting of chemical,enzymatic or physical modifications. Thisgenerates starches with many different prop-erties (different pastes and gels with varyingcharacteristics) suitable for a wide range ofdifferent industrial uses.
But the post-extraction processing hassome major disadvantages, it is energy con-suming and expensive and can generate pol-luting waste. It would be better if we coulddesign the right type of starch in the crop sothat post-extraction modification would notbe needed.
Starch consists entirely of glucose poly-mers. There are two types of glucose polymersin the granule: 20–30% long linear amylose and70–80% highly branched amylopectin.
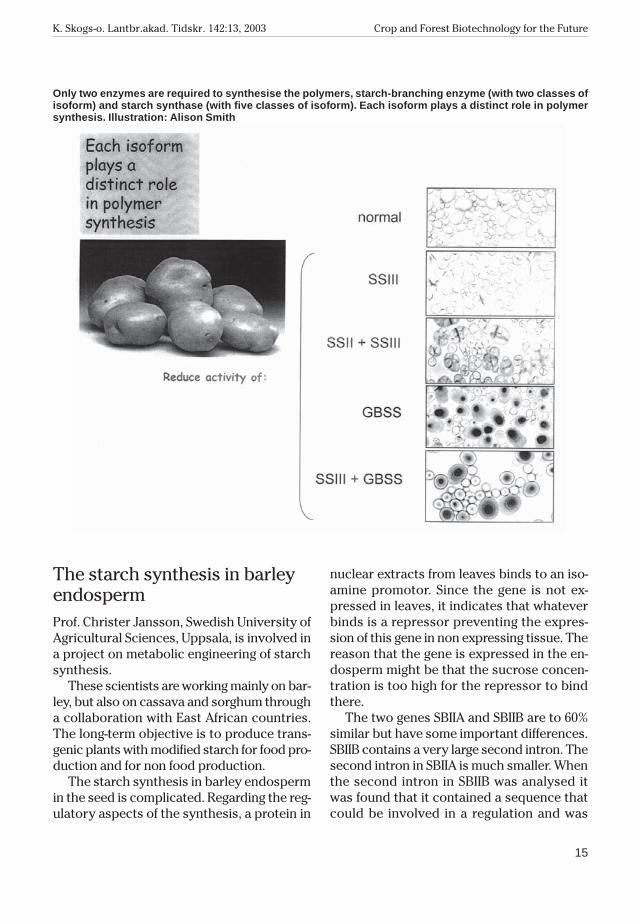
Only two enzymes are required to synthe-size the polymers. They are starch synthaseand starch-branching enzyme. The two en-zymes exist as multiple, conserved isoforms:there are five different isoforms of starch syn-thase and two of starch-branching enzyme.Each isoform makes a different contributionto the synthesis of the glucose polymers, andwithout these different classes of isoformsthe plant would not be able to make some-thing so complex as a starch granule. For ex-ample, in a potato tuber three classes ofstarch synthase are found. SSII and SSIII areresponsible for different aspects of the syn-thesis of amylopectin and GBSS is responsi-ble for the synthesis of amylose. The functionof each isoform cannot be replaced by other
isoforms, and the contribution of each is de-pendent on which other isoforms are present.
Debranching enzyme (isoamylase) is alsonecessary for normal starch synthesis. Whenit is reduced or eliminated, the plant synthe-sises a highly-branched glucose polymercalled phytoglycogen that is not normallypresent, and also synthesises many morestarch granules than normal. The precise roleof isoamylase is not known, but two possibleexplanations have been put forward.1. The direct model proposes that isoamyla-
se is directly necessary for the synthesisof amylopectin, because it removes someof the branches from a precursor calledpreamylopectin, allowing it to become or-ganised to form the granule. If isoamylaseis reduced or eliminated, less of the pre-amylopectin will become incorporatedinto starch granules. Instead it will becomephytoglycogen.
2. The indirect model proposes that isoamy-lase is only indirectly involved in thestarch synthesis. The starch synthase andstarch-branching enzyme are allowed tobe entirely responsible for the starch syn-thesis. The role of isoamylase is to de-grade any soluble products of the action ofthese enzymes on malto-oligosaccharidespresent in the stroma surrounding thegranule. If isoamylase is reduced or elimi-nated these malto-oligosaccharides arebuilt up by starch synthase and starch-branching enzyme and become phytogly-cogen.
Dr Alison Smith, John Innes Centre in Nor-wich, knows quite a lot about the enzymesthat are necessary for starch synthesis but asyet has not successfully put them into a con-vincing pathway that tells us how starch ismade. Nonetheless she and her colleaguesare beginning to gain sufficient informationfor “designing” starch properties in a rationalway.
M. Larsson
K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
15
The starch synthesis in barleyendospermProf. Christer Jansson, Swedish University ofAgricultural Sciences, Uppsala, is involved ina project on metabolic engineering of starchsynthesis.
These scientists are working mainly on bar-ley, but also on cassava and sorghum througha collaboration with East African countries.The long-term objective is to produce trans-genic plants with modified starch for food pro-duction and for non food production.
The starch synthesis in barley endospermin the seed is complicated. Regarding the reg-ulatory aspects of the synthesis, a protein in
nuclear extracts from leaves binds to an iso-amine promotor. Since the gene is not ex-pressed in leaves, it indicates that whateverbinds is a repressor preventing the expres-sion of this gene in non expressing tissue. Thereason that the gene is expressed in the en-dosperm might be that the sucrose concen-tration is too high for the repressor to bindthere.
The two genes SBIIA and SBIIB are to 60%similar but have some important differences.SBIIB contains a very large second intron. Thesecond intron in SBIIA is much smaller. Whenthe second intron in SBIIB was analysed itwas found that it contained a sequence thatcould be involved in a regulation and was
Only two enzymes are required to synthesise the polymers, starch-branching enzyme (with two classes ofisoform) and starch synthase (with five classes of isoform). Each isoform plays a distinct role in polymersynthesis. Illustration: Alison Smith
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
16
similar to the bbox from patatine promotor (astorage protein in potato). This bbox con-firms to specificity to the patatine gene. Theproject drew the conclusion that the genewas involved in regulation in barley. There isa protein or a protein complex binding to thiselement in the second intron. There is also,apparently, a protein binding somewhere out-side. So again, a repressor binds to this ele-ment. The susiba 1-gene and susiba 2-genehave regulatory roles.
The hypothesis is that the susiba 2-gene(the longer of the two susiba genes that arecracked) is expressed only in endosperm.The encoding repressor binds to a sugar re-sponsible element. In the endosperm thesusiba 2-gene encoding transcription factor isactive, while the transcription factor for susi-ba 1 is not active. The susiba 1-gene is insteadactive in the source of the leaves. When thesusiba 1-gene is active it encodes a repressorwhich binds to the bbox-like element in thesecond intron.
Genomics, Proteomicsand MetabolomicsGenomics is the analysis of genomes i.e. thesum of the genetic material in a cell, individ-ual or species.
Proteomics is the systematic analysis ofqualitative and quantitative changes of pro-teins expressed by a cell or tissue in responseto a specific biological perturbation. Globally,it will increase understanding of biologicalmechanisms. It will also give an identification/screening of diagnostic markers.
Metabolomics is the analysis of the metab-olism in the organism.
The sequencing of Arabidopsis thaliana wasa great breakthrough that can be used to im-prove the genetic knowledge of other plantsand to test different analysing instruments.
25 500 genes in the ArabidopsisgenomeArabidopsis thaliana has been the most wide-ly studied plant the last ten years. It was thefirst plant to have its genome sequenced, in2000, and by analysing epigenetics and chro-mosome structure it is possible to under-stand the role of methylation in regulatingchromosome structure and gene expression.
The work with analysing every single genewill continue to 2005.
Arabidopsis has approximately 25 500genes that are very closely packed in the ge-nome. About 60% of the genes match ESTs(Expressed Sequence Tags) which is verygood experimental evidence that most of thegenes are functional and means that they ex-press RNA.
Drosophila has round about 14 000 genesand the nematode Caenorhabditis elegans justunder 20 000 genes (while the human genomehas up to 40 000 genes). Arabidopsis, Dro-sophila and C. elegans make roughly about11 000–14 000 different types of proteins.Yeast does not make more than about 5 000proteins. Arabidopsis has a larger number ofgene families than the other organisms men-tioned. Around 1200 genes are present as tan-dem duplications.
There is evidence of duplication but not oftriplication or greater repetitions. The pro-cess identifies duplicated regions of the ge-nome. 60% are duplicated, which is the sameproportion as is identified by the direct DNAcomparison.
Once a sequence of genes is obtained, itcan be compared with all other genes. Aunique pattern is obtained. Arabidopsis de-votes the largest proportion of its genes tometabolism, like any other organism, for ex-ample a bacteria. However, because it is mul-ticellular it devotes a large amount of itsgenes to transcriptional control, cellular com-munication, signal transduction, control ofcell division and cell growth etc.
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
17
30% of the genes are presently unclassifed.They are either false gene predictions, maybe5–10%, or plant specific, which provides aninteresting subject for future systematic ana-lysis.
There are 29 gene families of 1700 knowntranscription factors in Arabidopsis and 16 ofthose families are unique to the plant. Prof.Michael Bevan, John Innes Centre in Norwich,finds the question of why plants should haveevolved such a diverse range of DNA-boundproteins very interesting.
The genome sequence provides a com-plete record of all of the genes and of all theprotein sequence because of the exquisiteprecision of nucleotide sequence. From thatit is possible to study expression patterns ofgenes and to build up knowledge of co-regu-lated genes, the so-called regulon.
Brassica differs fromArabidopsis and CapsellaIn the plant kingdom there exist a lot of differ-ences in genome size. Whereas in other or-ganisms there is quite a small range of ge-nome size, in angiosperms there is really atremendous variation in genome size inplants. Lilies have a much bigger genome butnot so many more genes than Arabidopsis.Most likely it is repetitive elements spreadbetween genes that make up the plant differ-ences in genome size. Arabidopsis has 130mbp/1C, rice has 450, tomato 950, maize 2 500and wheat 16 000 mbp/1C.
Plants probably differ significantly in thenumber of repetitive elements. In spite of thebig difference in genome size, the generalstructure of the chromosomes is often verysimilar. That means that the order of markersor genes along the chromosomes is oftenvery similar, if not identical.
Comparative genetic mapping experimentsreveal co-linearity of genomes. Tomato andpotato genomes are almost identical while
rice and maize genomes are very different. Inmaize there is some ancient duplication, sofor every rice gene there are two genes inmaize.
Dr. Renate Schmidt has worked in Max-Delbreuch-Laboratorium with comparativegenome analysis in crucifers. She and her col-leagues have chosen to compare Arabidopsisthaliana 2n=10, Capsella rubella 2n=16 andBrassica oleracea 2n=18. Arabidopsis andCapsella are much more closely related thaneither of them is to Brassica.
When maps of Arabidopsis and Capsellaare compared, some growth chromosomechanges would be expected. Despite thegrowth translocation events, the rest of themap looks largely co-linear. There are somesmall differences that could not have beenseen if the Arabidopsis genome had not beensequenced, because all the markers in Capsel-la could actually be linked back to the Arabi-dopsis genome sequence.
The microsynteny analysis of Arabidopsisthaliana and Capsella rubella shows not onlythat most of the genes are similar but alsothat they are in the same order and the spac-ing between the genes is roughly the same,although not identical. The transposable ele-ments are different in both species. A genethat is destroyed in Capsella seems to befunctioning in Arabidopsis. Another differ-ence between the genomes is that tandemduplications that are present in Capsella arethe equivalent of a single gene in Arabidopsis.We know that a lot of tandem duplicationsoccur in the Arabidopsis genome and thecomparison between the Arabidopsis andCapsella shows that this is a general featureof plant genomes. Tandem duplication seemto be very important and very frequent.
More than 80% of the Capsella segmentscan be placed back on the Arabidopsis se-quence. At least 50% of the repetitive DNAsequences go back to the low-copy nuclearDNA sequences. The sequences that matchto the Arabidopsis sequences are not only
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
18
gene coding sequences but aligned sequenc-es such as exon, intron and intergenic se-quences.
When Arabidopsis and Brassica are com-pared, it is found that exon lengths andsequences are highly conserved, whereasintron and intergenic sequences are muchmore varied.
Two highly homologous partial copies ofgene F are found on Brassica leracea Chromo-some 3.
Dr Schmidt found large differences intransgene expression in independent trans-formants. The expression level itself, when itgets over a certain threshold, is responsiblefor the gene silencing.
Analyse 14 000 clones in amicroarrayIt is possible to identify in/dels between eco-types of Arabidopsis thaliana using EST-micro-arrays.
With the microarray, all the genes can beviewed at the same time. Prof. Ellen Wismanfrom Michigan State University is studyinggene expressions in different ArabidopsisESTs to find out how many genes are regulat-ed by daylight. With spotted microarrays,there is a glass slide which is the microscopeslide and then inside this one can spot highdensity genome-DNA, EST-clones, CD-clonesor any DNA source that is amplified by PCR.They are spotted in high density.
Arabidopsis and Capsella are much more closely related than either of them is to Brassica. All the markersin Capsella could be linked back to the Arabidopsis genome sequence. Illustration: Renate Schmidt
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
19
The AFGC array analyses about 14 000clones: 9200 EST clones from all tissues, 2 000EST clones from developing seeds and 3 000GSTs (gene specific tags). The array controlsare:• Negative/spiking controls.• Transgenes (BAR, BT, BASTA, Luci-
ferase…).• Positive controls.• Dilutions of chromosomal DNA.
These are later needed to analyse the data.Former arrays could analyse more than
11 000 clones and 99% of the re-sequencedclones in the old collection were correct. El-len Wisman expects to have the same degreeof resolution in the new collection.
The AFGC performs experiments for cus-tomers and 45% of these customer experi-ments are genotype comparisons of mutants,usually on Arabidopsis. The scientists taketwo biological repetitions, each with onetechnical repetition. That is a minimum. Ifsmall differences are expected, more repeti-tions are needed to provide higher confi-dence in the data; for example if comparisonsbetween mutants and wild types are beingmade.
Gene expression studies have been usedfor gene discoveries. For the first time it ispossible to have a global view of processesthat were impossible before. This is a kind ofexperience where only the number of re-sponding genes gives an answer.
Future perspective for global gene expres-sion studies:• Gene discovery.• Global view (circadian rhythm, epigenic
changes).• Approach old problems (hybrid vigour).• Diagnostic tool (transgenes).• New hypothesis (looking for patterns over
many experiments).• Gene discovery in poorly characterized
species.
Future perspectives: as Columbia ESTs wereused, spots with higher intensities in otherecotypes are expected to result from an in-crease in gene copy number. In/dels can bemapped with QTLs to identify potential genefunctions and interesting agronomic traits.Combining array expression in SMD may giveclues to phenotypic differences between eco-types. In/dels common between ecotypesmight suggest a close relationship of groupsof ecotypes. Phylogenetic relationships ba-sed on in/dels can be compared to more tra-ditional methods.
Study the change in proteinquantityDescriptive proteomics analyse the proteomecomponents under a set of defined circum-stances. Quantitative proteomics analyse therelative amounts of proteins expressed undertwo or more sets of circumstances. To studythe change in protein quantity scientists con-sider several factors such as tissue or celltype, development stage, infection, disease,mutation, environmental effects and pesti-cides and herbicides. In the quantitative ana-lysis of the proteome, currently techniquesare used that measure quantitative changes.They are 2-dimensional polyacrylamide gelelectrophoresis (2D PAGE) and stable isotopelabelling/multidimensional liquid chromato-graphy.
The advantages of 2D PAGE are:• 2D PAGE visualises many proteins at once
(whole organisms, tissue types, sub-cellu-lar fractions and multi protein complexes).
• It is a relatively quick technique• The ease of detection (silver stain in the
interval 1–100 ng, collodial coomassie at50 ng–50 µg and SYPRO dyes in the inter-val 1 ng–100 µg.The method also has some disadvantages:
Only proportions of proteomes are visualised(membrane proteins, proteins with extremes
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
20
of pI and proteins with extremes of molecularweight (MW) are poorly represented), theresolution is co-migrated and there are lowabundance species, (1 000–5 000 copies percell), 90% of the protein is the product of 10%of the genome. Other disadvantages are poorgel to gel reproducibility and that only onesample is taken at a time.
The Difference Gel Electrophoresis (DiGE)advantages are that the method gives:• Comparison of two samples simultaneous-
ly (it is the most sensitive method withbroad dynamic range, it gives reproduci-bility – no gel to gel variation – and it hasa multiplexing possibility).
• No staining (it requires fluorescent imagerwith integral spot cutter and it is compat-ible with mass spectrometry.
• Quantitation (image analysis performedby ImageMaster2D – output excel spread-sheet of Cy3/Cy5 ratios.
Dr. Kathryn S. Lilley and her group at theUniversity of Cambridge were one of the firstsites in the UK to work with the DiGE system.They are content with the method after analy-sing the data achieved with the DiGE system.
There are some points to be considered inplant proteomics: Phenols can cause proteincross-linking. The genetic background and
Proteomics is the systematic analysis of qualitative and quantitative changes of proteins expressed by acell or tissue in response to a specific biological perturbation. Illustration: Kathryn Lilley
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
21
the standardised growth conditions must beconsistent. The development stage must bematched. Contamination must be controlled.RUBISCO dominates 2D gel if photosyntheticmaterial is used.
In the future there will be new techniquessuch as pre/sub fractionation techniques, iso-tope affinity tags, capillary electrophoretictechniques, multidimensional liquid chroma-tography and protein chips.
Metabolic profiling: A metabolicengineering perspectiveThe approach for manipulation of starch andamino acid metabolism in the potato tuber isto move away from naive early approachesand towards predictable strategies. A frame-work of MCA is used for future strategies.
Detailed characterisation based on metabol-ic profiling for understanding past “failures”.
Dr. Alisdair Fernie, Max-Planck-Institute inPotsdam, describes four strategies:1. Targeting AGPase: Overexpression of an
unregulated bacterial AGPase resulted inincreased starch yields. (Stark et al., 1991) However, in a separate experiment theincreased flux to starch was coupled withan increased starch degradation (Sweet-love et al., 1996).
2. Targeting the plastid ATP:ADP transporter:Overexpression of an AtAATP has a con-trol coefficient of 0.78 for starch synthesis.
3. Targeting the plastidial PGM. Alisdair Fer-nie’s group generated transgenic plants inwhich the plastidial isoform of phospho-glucomutase was inhibited. These plantswere characterised by severe reductionsin starch contents, proving the involve-
The four strategies for manipulation of starch and amino acid metabolism in the potato tuber are 1) targe-ting AGPase, 2) targeting the plastidial ATP, 3) targeting the plastidial PGM and 4) targeting sucrose break-down. Illustration: Alisdair Fernie
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
22
ment of pPGM in the starch synthesispathway. Antisense plants exhibiting inhibition inpPGM are a phenocopy of those exhibitingAGPase. pPGM has a relatively high con-trol coefficient for starch synthesis. There-fore ectopic expressions of a non-plantPGM both individually and in combinationwith other proteins represent a novelstrategy that may enhance starch yield.
4. Targeting sucrose breakdown. Primarycharacteristics of plants engineered seemto have more efficient sucrose metabo-lism.
The observed increase in amino acids wassurprising given the use of a tissue-specificpromoter. It had previously been thought thatamino acids were synthesised in the leavesprior to transport to the tuber. This demon-strates that the tuber has the necessarymachinery for the de novo biosynthesis of allamino acids and illustrates the importance ofmetabolic profiling in genomics approaches.
Metabolic profiling allows comprehensivephenotyping. When carried out on diversesystems this also allows the identification ofphenocopies. The measurement of many dif-ferent metabolites within a single extract al-lows evaluation of “dependency” relation-ships between metabolites.
Overall conclusions: A greater understand-ing of the control processes regulating metab-olism is required before metabolism engineer-ing becomes routine. Alisdair Fernie hopes toachieve this by assessing the steady-stateconcentrations of a wide range of metabolitesand by the development of sensitive and accu-rate methods of determining cellular fluxes.
Future perspectives: Use of acquired know-ledge for rational design of genetic manipula-tion strategies. To look deeper at the metabol-ic networks involved by a combinatorial ap-proach using both radiolabelled and stableisotopes. To look at disparities in metabolismduring different developmental stages.
Plant stress andsignalling
Plants in all cultivation systems are more orless sensitive to stress which limits horticul-ture, agriculture and forestry. Stress can befrost, drought, heat, salinity and air pollution.All kinds of stress decrease the production.With more stress tolerant varieties, yieldswould be improved and new crops could begrown.
Hormonal regulation of oxidativecell deathTo get a better understanding of stress, stressrecognition and what kind of signalling theplant is using to resist the stress, Prof. JaakoKangasjärvi, University of Turku, started bystudying the effects of ozone (O3). Ozone is aconvenient tool for studying the role of oxy-gen radicals in signalling.
Ozone is an air pollutant and reacts withdifferent components of the cell wall, formingreactive oxygen species there. It can neverenter the cell.
Traditionally, people who have been stud-ying ozone from an environmental point ofview, as a pollutant, have believed that theseradicals physically destroy the cells, like awound response. But actually, there is plentyof evidence that reactive oxygen species arecentrally involved in cellular signalling.
The classical example is the oxidativeburst in plant-pathogen interaction, but it canalso be found in the regulation of senescenceand so on. Looking at recent papers, reactiveoxygen species have been observed in manydifferent stresses and also in some of thedevelopmental aspects.
Arabidopsis columbia ecotype is tolerantbut certain Arabidopsis mutants are sensitiveto ozone, which leads to cell death. When
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
23
Prof. Kangasjärvi finds a mutant with onemutation in one gene that leads to acquiredsensitivity he can later on identify that geneand say that this pathway is involved in theozone lesion formation. The scientific grouphave named their mutants rcd (radically in-duced cell death). In the rcd1 mutant, the celldeath is spread from cell to cell in the leaffrom the initiation sites. The cell death contin-ues even after exposure to ozone in clean air,until almost the whole leaf is dead.
Rcd1 is a single co-dominant trait in chro-mosome one. It is hypersensitive to ozoneand superoxide but not to hydrogen perox-ide. Rcd1 has been shown to increase initialcell death rates and prolong spreading celldeaths and superoxide is sufficient to triggerthe lesion formation.
Instead of ozone damage, scientists arenowadays speaking more of the role of reac-tive oxygen species and how they interactwith different signalling pathways, how theseregulate expression of target genes and howthey regulate cell death. Cell death in thiscase is an active genetically programmedprocess that requires metabolism, transcrip-tion and translation. Since there is a propaga-tion of cell death, there must also be signalsthat go from the dying cells to the next ones– cell to cell communication.
Ethylene has been shown to be involved inseveral abiotic stresses, including oxidativestresses. Ethylene signalling is required in celldeath and superoxide accumulation.
In the oxidative cell death cycle, salicylicacid enhances the toxicity, ethylene amplifiessuperoxide accumulation and jasmonates in-hibit salicylic acid.
Conclusions: Rcd1, the radical hypersensi-tive mutant, exhibits a high ethylene peakwhich contributes to the spread of lesions.The ethylene insensitive ein2 is highly tole-rant to radicals because of the ethylene sensi-tivity and it has increased ethylene evolutionbecause it has lost the feedback regulation ofethylene biosynthesis.
• ROS (reactive oxygene species) are impor-tant biological regulators of cell death dur-ing oxidative stress.
• ROS regulation of cell death is tightly cou-pled with hormonal regulation.
• ROS and hormones work really as an inter-connective network where everything af-fects everything else and not as separatelinear pathways. There is cross talk thataffects the sensitivity of other pathwaysand so on, which is something that Prof.Kangasjärvi and his group are going toaddress in the future.
• Ozone is a convenient tool in probing therole of reactive oxygene species in the reg-ulation of these things.
Defence responses to Leptosphaeria maculansThe fungal pathogen Leptosphaeria maculans(anamorph: Phoma lingam) causes blacklegor stem canker on several species withinBrassicaceae. This plant pathogen is the ma-jor threat to crops like oilseed rape (Brassicanapus) and turnip rape (B. rapa) and causesconsiderable crop losses in Canada, Australiaand Central Europe.
The rapid evolution of new and more ag-gressive (virulent) isolates demands a moreefficient use of resistance sources within theplant population. Resistance to L. maculans ismainly found in Brassica species containingthe B-genome, i.e. Brassica nigra, B. junceaand B. carinata. Traditional breeding pro-grammes have utilised this germplasm as asource of resistance, but transferring genesfrom interspecific hybrids demands mucheffort.
Dr. Christina Dixelius and her colleagues atthe Swedish University of Agricultural Scien-ces in Uppsala have cloned the first gene, Lm1,from B. nigra, that confers resistance to L.maculans. Homologous sequences have alsobeen found in Arabidopsis thaliana, rice and
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
24
partly in the nin gene from Lotus japonicus.The latter is interesting since some signalpathways in the symbiotic interactions aresimilar to those found in pathogenic systems.The Lm1 protein has two transmembrane re-gions and it is tempting to speculate that thisunique part senses pathogenic attack, direct-ly or as a part of a signal transduction pathway,while the part containing conserved motifsacts as a cytoplasmic effector domain.
Arabidopsis thaliana and Brassica speciesare closely related even if they phylogeneti-cally belong to different genera within theBrassicaceae family. The similarity is furtherevident on nucleotide level where compari-sons between B. napus genes and their A. thal-iana orthologues show more than 85% iden-tity even though the Brassica and Arabidopsislineages diverged 12.2–19.2 million years ago.A large number of pathogen resistance genes
are characterised to a more or lesser extent inA. thaliana. In addition to this, mutant studieshave provided insight into A. thaliana basaldefence responses against various patho-gens. Interestingly, it has been demonstratedthat the A. thaliana genome can be a source ofresistance to L. maculans.
A number of A. thaliana mutants impairedin defined parts of various signalling path-ways related to plant stress induced by otherpathogens are known. Mutants such as pad(phytoalexin accumulation deficient), npr(non-expressor of PR genes), eds (enhancesdisease susceptibility), ndr (non-specific dis-ease resistance), lsd (lesions simulating dis-ease), and cpr (constitutive expresser of patho-genesis related (PR) proteins) have been eval-uated in the L. maculans system. However,only pad3 has given a susceptible phenotype.This finding is in contrast to e.g. the Alternaria
The fungal pathogen Leptosphaeria maculans causes blackleg or stem canker on several species withinBrassicaceae. The disease is spread with the rain and the wind. Source: B. Howlett
M. Larsson
K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
25
and Perenospora systems and suggests thatthere is another type of signalling in the L.maculans system. The pad3 mutant is nowbeing crossed with lms1, lms2 and the RacGapmutant to reveal their reciprocal relation-ships. Such clarification is of high importancesince lms1 and lms2 give a susceptible pheno-type but show wild type level of camalexin.
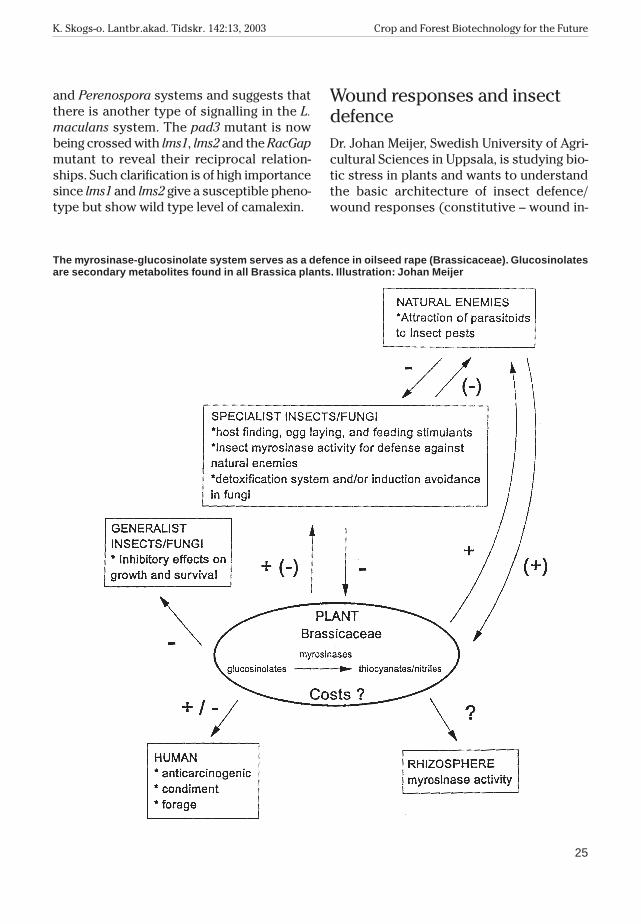
Wound responses and insectdefenceDr. Johan Meijer, Swedish University of Agri-cultural Sciences in Uppsala, is studying bio-tic stress in plants and wants to understandthe basic architecture of insect defence/wound responses (constitutive – wound in-
The myrosinase-glucosinolate system serves as a defence in oilseed rape (Brassicaceae). Glucosinolatesare secondary metabolites found in all Brassica plants. Illustration: Johan Meijer
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
26
ducible), the regulation of wound responses(signalling, cross-talk and gene activation)and the possibility to improve the insect pestresistance.
Model organisms are Brassica napus (oil-seed rape), Arabidopsis thaliana and Pichiapastoris (for overexpression).
The myrosinase-glucosinolate systemserves as a defence in oilseed rape. Glucosi-nolates are secondary metabolites found inall Brassica plants. These plants also housethe enzyme myrosinase. Myrosinase de-grades glucosinolates into toxic products e.g.thiocyanates/nitriles, which are harmful topests. A number of other proteins are associ-ated with myrosinase in seed extracts, suchas myrosinase binding protein (MBP) whichis a lectin and myrosinase associated protein(MyAP), and which houses a lipase motif. Therole of complex formation is not clear.
Myrosinase cellular location: The distribu-tion of myrosinase containing idioblasts isdifferent in Arabidopsis thaliana and Brassicanapus. In Arabidopsis, myrosinase is localisedin idioblasts present in the phloem. In Brassi-ca napus, myrosinase is localised in idioblas-tic myrosine cells in the ground tissue and inidioblasts in the phloem.
Myrosin “grains” form a continuous reticu-lum in Brassica napus myrosine cells as visu-alised by confocal laser scanning microscopy.
The third myrosinase gene (TGG3) in Ara-bidopsis is a pseudogene but is specificallyexpressed in stamen and petal.
Conclusions for oilseed rape seed analysis:• SeedMBP ubiquitous expression except
myrosin cells• Myrosinase-MBP complexes do not exist
in vivo• SeedMBP necessary for complex forma-
tion• Free MB myrosinase is soluble and active• No effect of as seedMBP on pests tested.
Down-regulation of vegetative and wound-inducible myrosinase binding proteins in
Brassica napus affect susceptibility differenti-ally in different Brassica specialist herbivo-res.
Regulation of the plasmamembrane H+-ATPase byphosphorylation and 14-3-3The plant plasma membrane H+-ATPase is anabundant enzyme that builds up an electro-chemical gradient across the plasma mem-brane by pumping protons out of the cell.This gradient provides the energy that drivessecondary active transport of nutrients andsolutes across the plasma membrane. Conse-quently, the H+-ATPase is thought to play amajor role in many cell processes and its ac-tivity is regulated by a number of physiologi-cal factors, including hormones, blue lightand fungal toxins.
Prof. Marianne Sommarin and her col-leagues at Lund University have earliershown that treating intact leaves with the fun-gal toxin fusicoccin results in activation of theH+-ATPase and that this activation is achievedby binding of 14-3-3 protein to the C terminusof the H+-ATPase.
14-3-3 proteins constitute a family of eukary-otic proteins that are key regulators of a largenumber of processes ranging from mitosis toapoptosis. In plants, the most well studied ex-amples are the plasma membrane H+-ATPaseand two enzymes of the primary metabolism,nitrate reductase and sucrose phosphate syn-thase. Nitrate reductase and sucrose phos-phate synthase are both inhibited by 14-3-3binding, in contrast to the activation seen withthe plasma membrane H+-ATPase. 14-3-3 pro-teins usually bind to phosphorylated motifs intheir target proteins. However, there is no ami-no acid sequence in the C terminus of the H+-ATPase with any obvious similarity to the es-tablished binding motifs for 14-3-3.
Following in vivo phosphorylation usingradiolabelled 32Pi and fusicoccin, Prof. Som-
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
27
marin and her group identified a phosphory-lated amino acid in the C terminus of the H+-ATPase, namely Thr-948. Thr-948 is the penul-timate amino acid in the C terminus of mostplasma membrane H+-ATPase isoforms. Theycould show that phosphorylated Thr-948 ispart of a binding site for 14-3-3 in vitro, a bind-ing site that differs strongly from the estab-lished binding motifs. The physiological im-portance of this novel motif was shown byheterologous expression of a plant H+-ATPasein yeast. A mutation in the motif (exchange ofThr-948 for alanine) abolished 14-3-3 bindingand activation of the plant H+-ATPase, andprohibited yeast growth, which in this yeaststrain was dependent on H+ pumping by theplant H+-ATPase (the gene for the yeasts ownH+-ATPase was inactivated).
In all plants analysed so far, the plasmamembrane H+-ATPase and 14-3-3 proteins areencoded by multigene families. The genomeof the model plant Arabidopsis harbourstwelve H+-ATPase genes and fifteen 14-3-3genes, and most of them are expressed. Anobvious question is whether the many iso-forms in multicellular organisms reflect differ-ent functional roles, i.e. does isoform specifi-city towards target proteins exist? Is thisachieved by differences in tissue/subcellulardistribution and/or sequence differences inthe interaction sites between the two compo-nents? The scientists are presently investigat-ing the expression pattern and tissue/subcel-lular localization of the different isoforms inplants exposed to various forms of stress toaddress this issue. In addition, they are study-ing the interaction between the different 14-3-3 isoforms and phosphorylated peptidesmimicking the C-terminal binding motifs ofthe different H+-ATPase isoforms.
Forest genetics
According to the FAO report “The State of theWorld’s Forests”, there is a yearly decline of12 million hectares due to an increasing con-sumption of wood and all kinds of wood prod-ucts and due to an increased urbanisation. Itis mainly in developing countries that forest-ry land is being converted to agriculturalland. This decline is expected to continue. Inthe same time there is a growing environmen-tal awareness of the need to protect the re-maining forests. The simple conclusion is thatthere is a need to increase the yield and thequality of wood of tree species that can begrown in agro forestry systems. Forest gene-tics can be a valuable tool for this, especiallyconsidering the long rotation of forest treesand the natural slow improvement with tradi-tional tree breeding.
Genetic engineering improvepoplarsBreeding is the main approach to improve-ment of quality and yield. Genetic engineeringcan also be used. Disease resistance is themajor selection criterion for breeding in pop-lar trees and an important factor determiningyield.
When breeders started the resistancework against the major fungus
Melampsora larici – populina, they usedinterspecific crosses between the resistantAmerican Populus deltoides and the nativePopulus nigra. The result was highly produc-tive interspecific hybrids that were complete-ly resistant to this pathogen. These cloneshave been spread over Europe.
Over the years, new varieties of the patho-gen have occurred and broken down the re-sistance. As a consequence, there is current-ly not a single resistant clone that is commer-cially available.
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
28
Together with the Poplar Breeding Insti-tute, Dr. Wout Borjean and his team at theUniversity of Ghent have initiated a pro-gramme to understand the genetics of inher-itance of the disease resistance in order toprogress slowly to a marker resistant selec-tion.
The first step was to create genetic maps ofthe three most important poplar species forbreeding world-wide. The maps were used todissect resistance to Melampsora. The scien-tists grew the poplars in greenhouses to makesure that they were not infected by other dis-eases. Approximately 50% are completely re-sistant while the other 50% are spread over allthe other classes. This is an agreement with amodel where the female parent, which is re-sistant, is heterozygous for a dominant genewhich confers resistance and then segregatesinto a Mendelian ratio in the population.
Resistance to the three races completelyco-segregates so there is a major locus.
Prof. Borjean was interested in cloning theresistance gene by positional cloning. He thenhad to saturate the locus with genetic mar-kers. His team looked for primer combina-tions – “fingerprints” – that gave them mark-ers that were present in the resistant parent,absent in the susceptible parent, will bepresent in the bulk of resistant individualsand will be absent in the bulk of susceptibleindividuals. Using 540 primer combinationsthey have identified 11 markers, and could,with the markers, identify 16 recombinants intheir 502 progeny plants from two mappingpedigrees. The scientific team ended up witha locus of ten markers in coupling and one inreversion.
Interestingly enough, it looks as thoughthis is a large resistance gene cluster.
The future plans for this project are to mapthese R-gen clusters and see whether theycoincide with qtls. It is also important to inte-grate all the poplar maps world-wide.
Genetic engineering – in this case engineer-ing lignin biosynthesis – is also a tool to im-
prove wood quality for the pulp and paperindustry. Roughly 50% of wood is made up ofcellulose, 25% hemicellulose and 25% lignin.For making high quality paper this lignin frac-tion needs to be degraded by chemicals andneeds to be extracted and separated from thecellulose. This is a very energy requiring andtoxic process. The question is thereforewhether it is possible to reduce the amount oflignin in trees or to change the composition oflignin in order to make it easier to extract.
A lignin molecule is a polymer with threeunits and with the creation of different typesof interunit linkages. If the ratio of these unitsin lignin can be changed, this will also changethe ratio of the linkages and as a consequencewill interfere with the extractability of lignin.
The intentions of Prof. Borjean were tomodify the composition of lignin. He clonedthe different enzymes of this pathway, makingantisense constructs and putting them intransgenic poplars. The first enzyme ana-lysed was AS Comt. It did not change theamount of lignin but it drastically changedthe composition. Prof. Borjean also found anovel unit incorporated into the transgenicplant.
The results so far indicate that it is pos-sible to reduce the lignin in the tree as well aschange the composition of the lignin and thatless chemicals are needed to process thetransgenic poplars.
Functional genomics of woodformationA tree has round about 40 000 genes. Prof.Göran Sandberg, Swedish University of Agri-cultural Sciences in Umeå, and other Swedishscientists are working in a national EST-pro-gramme – The Swedish Tree Functional Ge-nomics – on the first large scale sequence ana-lysis of genes in a tree. They are especially in-terested in identifying all the genes that directthe formation and structure of the fibre.
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
29
The project concentrates on two differenttypes of genes that are crucial to the practicalimplementation in forestry. One type affectsformation of wood and stem growth. Theother type of gene is important for stress tol-erance. Wood formation and the ability to sur-vive under harsh climates are unique charac-ters for trees.
The initial stemcell can develop into differ-ent types of cells. Wood properties are deter-mined in a developmental gradient, with thedevelopmental decision being taken in thecambium.
In early spring, the number of xylem moth-er cells determines the amount of wood pro-duced together with seasonal and other ef-fects on the gradient. A bit later in the season,the cell has gone through the elongation zoneand starts to form the secondary wall. Whenthis cell enters the expansion zone, it has onlythe primary cell wall. In late season, the cellgets the secondary wall much earlier. There isalso an effect of the climate on this gradient.
The dormant cambium provides a way fora conifer to survive for 700 years.
The point with the technology is that withthe wide range of data and all pathways foramino acid synthesis etc, experiments can beinitiated. The advantage is that it is possibleto go in and look at all the pathways at thesame time.
The most important thing for wood devel-opment is gene function. Functional genom-ics is just a way to get to gene function. Theproject has a database with real tissue specif-ic expression of genes. If there is a gene thatis expressed in the cell division zone, it mightbe a candidate for regulation of this process.This is a way to find key regulatory factors.
How can gene function be analysed? Ifthere is high homology to an Arabidopsisorthologue, it can actually be done in Arabi-dopsis because secondary growth can be in-duced in Arabidopsis. If it cannot be done inArabidopsis, it has to be done in poplars.Poplars are much closer to Arabidopsis than
tomatoes or tobacco are to Arabidopsis. Pop-lar ESTs have a high similarity to Arabidopsis,but Arabidopsis has big families, which is adisadvantage if one wants to knock out genesand be sure they are compatible.
Basically, when going from the expressionpattern in poplars it can be predicted thatthis has something to do with vascular forma-tion and then it can easily be checked thatthe mutant is found in Arabidopsis.
If the process has to be carried out in pop-lar, there is a method to select good candi-date genes and the project is now evaluatingstrategies to do this. The goal is to knock outat least 2 000 poplar genes at the Centre forForest Biotechnology and Chemistry. Theproject will select interesting candidates, notknock out in random.
Metabolic profiling is another tool that isbeing developed.
Enzymes for making andbreaking wood fibresEnzymes can be used to change the structure,surface, composition and function of the fibresfrom the cell walls of plants/trees. By identify-ing, examining and modifying enzymes, scien-tists can make better and more exact changes.
Prof. Tuula Teeri, Royal Institute of Tech-nology in Stockholm, is an expert on the en-zymes that are involved in both the biosyn-thesis and biodegradation of wood fibres.The three major components of wood arecellulose, lignin and hemicellulose. Celluloseis the load-bearing structure that one usuallywants to maintain in all kinds of application:papers, tables, houses etc. Chemically it is avery simple polymer, stacked together withother molecules in sheets and the sheetsstack on each other forming crystals. Theselong fibrils pack further to long fibres, usedby the pulp and paper industry.
The cell wall also consists of other compo-nents like hemicellulose and with a backbone
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
30
of one or different sugars which can be xy-lose, glucose etc. It is linked to different sidegroups which have the effect that these poly-mers usually are totally or partly soluble,amorphous or only very partially insoluble.The cell wall holes between cellulose andhemicellulose are filled in with lignin.
Sometimes it is desirable to get rid of orreduce the lignin when making paper. For ex-ample, residual lignin causes yellowing inpaper. There are many mechanical and chem-ical processes involved in pulp and paperproduction but a possibility is that some ofthem could be displaced or facilitated withenzymatic processes.
In nature, there are many microorganisms,fungi and bacteria that, with the help of extra-cellular enzymes, break down and utilise plantmaterial in a controllable way. Nature hasprobably developed thousands of enzymes todeal with the different aspects of wood degra-dation or biosynthesis. To achieve total hy-drolysis, it is necessary to use mixtures ofenzymes or even a whole organism, since nosingle enzyme is able to achieve completedegradation of the complex raw material.
In contrast, for pulp bleaching, paper mak-ing or the textile industry, one selects one ortwo enzymes that can remove selected fibrecomponents but that do not harm the fibresfurther. The extracellular enzymes are spe-cialised in different components in the cellwall and affect the composition as well as thestructure and other characters.
With enzymes, it is possible to reduce theamount of lignin for bleaching or do somesurface modification of the cell walls to per-form better in paper making. Enzymes canmake them react better with other agents.
In addition, enzymes could improve thereactivity of the main component, cellulose,because there is very little chemistry on thecellulose surface.
Prof. Teeri observes that efficient largescale production of enzymes is not yet pos-sible, but she is sure that the pathways will be
very different from now on and that there willfind new enzymes to utilise.
The longer night lengthinfluences the phytochromeTwo major questions concern how the leafadapts to its environment and what is thegenetic basis of autumn senescence.
32% of ESTs (Expressed Sequence Tags)encode photosynthetic proteins and 37%encode chloroplast proteins in leaf proteinsynthesis. All plastid-encoded proteins arepart of multi-protein complexes together withnuclear-encoded subunits for which synthe-sis can be estimated. The plastid protein syn-thesis in young Populus leaves correspondsto approximately 25% of nuclear protein syn-thesis. 50% of all leaf proteins are located inthe chloroplast.
The previous day’s weather is most impor-tant in determining PsbS mRNA levels, sothere must be a signal that is stable over-night.
RNA accumulation precedes yellowing inautumn senescence. Different sets of genesare expressed and 2 700 different genes werefound among 5 200 ESTs sequenced. Many ofthem have ”unknown” functions. There aremany proteases, stress-induced genes etc.During the senescence, there is a change fromphotosynthesis to respiration. RNA disap-pears two weeks after yellowing and the generegulation patterns change. The genetic pro-cess is developmentally controlled.
The hypothesis for the genetic regulationof autumn senescence is that the longer nightlength influences the phytochrome. With sig-nal transduction the tree obtains cold hardi-ness and is subject to senescence. The long-term goal is to analyse every step and learn tomanipulate the senescence for different needs.Earlier senescence might increase the hardi-ness and later senescence might increase theyield.
M. Larsson
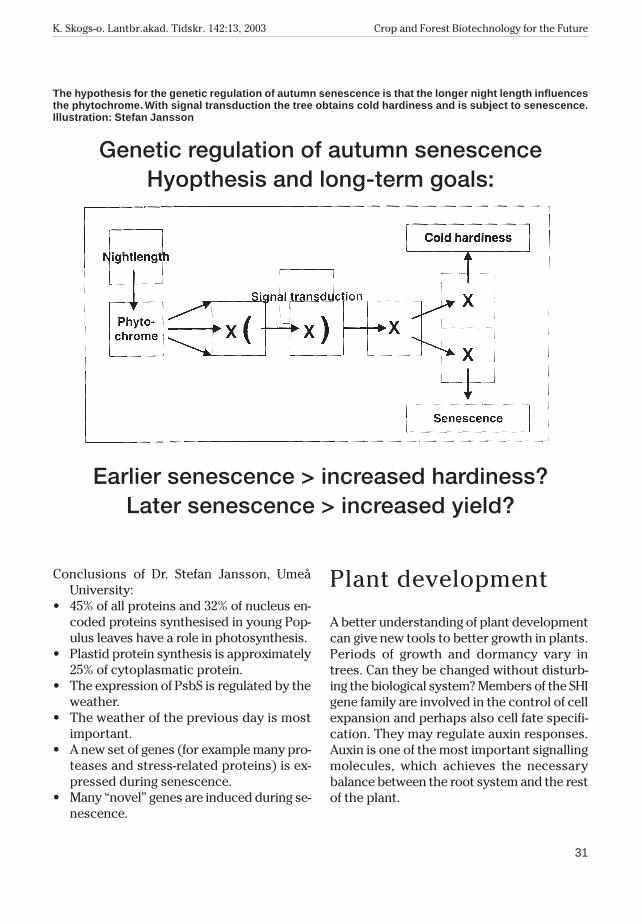
K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
31
Conclusions of Dr. Stefan Jansson, UmeåUniversity:
• 45% of all proteins and 32% of nucleus en-coded proteins synthesised in young Pop-ulus leaves have a role in photosynthesis.
• Plastid protein synthesis is approximately25% of cytoplasmatic protein.
• The expression of PsbS is regulated by theweather.
• The weather of the previous day is mostimportant.
• A new set of genes (for example many pro-teases and stress-related proteins) is ex-pressed during senescence.
• Many “novel” genes are induced during se-nescence.
Plant development
A better understanding of plant developmentcan give new tools to better growth in plants.Periods of growth and dormancy vary intrees. Can they be changed without disturb-ing the biological system? Members of the SHIgene family are involved in the control of cellexpansion and perhaps also cell fate specifi-cation. They may regulate auxin responses.Auxin is one of the most important signallingmolecules, which achieves the necessarybalance between the root system and the restof the plant.
The hypothesis for the genetic regulation of autumn senescence is that the longer night length influencesthe phytochrome. With signal transduction the tree obtains cold hardiness and is subject to senescence.Illustration: Stefan Jansson
Genetic regulation of autumn senescenceHyopthesis and long-term goals:
Earlier senescence > increased hardiness?Later senescence > increased yield?
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
32
A conifer perspective onreproductive organ developmentin plantsAll the major angiosperm families were estab-lished 80–90 million years ago, which meansthat they are relatively young compared toother land plants. They are also closely rela-ted.
Most of the work on conifer trees has beenfocused on reproductive organs, mainly forpractical reasons. Male and female reproduc-tive organs are born on separate axes. Theseed cone is surrounded by the remnants ofthe pollen cones. The angiosperm flower, onthe other hand, is a truly unisexual organism.
Are there genetic similarities between theregulating mechanisms that control seedcone in conifers and the mechanisms thatcontrol the angiosperm flower (Arabidopsis)?Pollen cone is the male organ and seed conethe female.
The starting point of the work was datafrom Arabidopsis and other angiosperms.The basis for the ABC-model is two Arabidop-sis mutants. In total, the Arabidopsis has ahundred MADS-box genes and 4 of these arepart of A, B, and C functions. The genes presentin Arabidopsis have evolved by a series ofgene duplications indicated by branches inthe “evolutionary tree” and followed by se-quence divergence and functional diver-genes. The increasing complexity of the genefamily is perhaps related to the increasingcomplexity of the structure.
There are MADS-box genes that are activeduring the reproductive development in con-ifers as well. Prof. Peter Engström and hiscollegues at the Uppsala University isolated afew clones.
In angiosperms, a gene that is expressed inone type of organ has a role related to thatspecific organ. There are functions related toaugiar development and augiar developmentis something different from genosperms andangiosperms.
The scientists found three genes that arephylogenetically related to AP3 and three pes-tilata genes that have the B-genes in the an-giosperms. The DAL 11, 12 and 13 genes are allrelated to B but they have different functions.
Angiosperm genes can convert females tomales.
B-genes and C-genes are present but no A-genes have been found. The B- and C-typesystem for regulating the reproduction devel-opment is an ancient feature common togenosperms and angiosperms.
By studying the reproductive developmentof Arabidopsis, the scientists have at leastcome up with a possible model for how repro-ductive development is controlled in its cen-tral aspects in conifers.
Regulation of cambialdevelopmentDr. Rishikesh Bhalerao and his collegues atthe Swedish University of Agricultural Scien-ces in Umeå are working with two model sys-tems, Arabidopsis and poplars. The chosenprocess can be studied much better in pop-lars and the knowledge can be utilised in thefurther Arabidopsis research.
These scientists are studying the seasonalactivity of the vascular cambium.
The tree shifts between active growth anddormancy. Some trees stop growing ratherearly in the north, which is because theyneed a longer preparation period for the win-ter. But different trees need various amountsof time for the preparation depending on howfast the tree gains cold hardiness. From a pro-duction point of view, a longer period ofgrowth and consequently a short winterpreparation period would be beneficial. Theactual winter is hard to influence.
Dr Bhalerao wants to find the answers tothree questions:• What is the role of auxin transport in dor-
mancy transition?
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
33
• What are the targets of dormancy inducingsignals in the cell cycle machinery?
• How does auxin sensitivity regulate cell di-vision?
Auxin transport has been shown to play a keyrole in cell division in the vascular cambiumand in maintaining its identity as a merisma-tic tissue.
Poplars grow rather fast (compared withother trees), they have a limited genome size,some genetics work is possible on them andthere exists a very excellent EST sequencingprogramme. Transcriptional profiling andgenomics etc. are more reasons for usingpoplars.
The vascular cambium makes a transitionfrom active to passive when the tree ceasesto grow. The auxin transport is reduced as thetree stops growing, but even after the cell di-visions have stopped there is considerablecapacity in the cambium to transport auxinfor a long time. That means there are twomechanisms operating. One which reducesthe capacity first and the cells later. Thosecells lose their capacity to transport auxin.The other mechanism operates when thepoplars break. There is a tremendous in-crease in the transport capacity in the vascu-lar cambium – prepared to handle all the aux-in that comes down.
The auxin transport is reduced becausethe auxin handling genes are turned off.
Cell cycle machinery is very conservedfrom animals to plants and basically consistsof four phases. Six enzymes (two in poplar)interact with a group of proteins, called cyc-lins. The cyclins are specific for the differentphases and help the transitions. The dormantcambium is supposed to be “arrested” at oneof the phases. The protein level is cut downduring dormancy. This is the first level of reg-ulation, which is post transitional. At the sec-ond level, the activity of protein goes downeven when the level is constant. These activ-ities are all driven by a pathway called cell
division auxin transporters. There are veryfew genes in common between the activecambium and the dormant cambium. Thereare fewer expressed genes but they are ex-pressed more often. Dr Bhalerao and his teamcannot yet answer the question of how activ-ity goes up and down when the switch fromactive to dormant cambium occurs.
In another project, the development ofxylem is being studied. Xylem is arrangedinto distinct developmental zones. How doesthis kind of pattern get specified? There areseveral genes involved in different kinds ofregulation. Chilling alters the sensitivity of theresting cambium to the applied 1AA gene.Dormancy is a complex but excellent system.
Members of the SHI gene familyare involved in the control ofcell expansionDr. Eva Sundberg, Uppsala University, is inter-ested in genetic regulation of plant develop-ment and has focused on Arabidopsis and theSHI gene family, as it appears that they areinvolved in the regulation of development.
The SHI gene family consists of at least tenmembers and they interfere with GA- and aux-in signalling when constitutively expressed.The SHI gene family encodes proteins thathave a RING domain, which is characterisedby a number of conserved cysteine and histi-dine residues. It has been suggested that RINGdomains are exclusively involved in protein-protein interactions. A large number of theRING finger proteins has been shown to beinvolved in ubiquitination for targeted de-gradation. In addition, some proteins includ-ing RING 1 itself appear to be involved in tran-scriptional repression via interaction withchromatin remodelling proteins.
The SHI gene was identified by activationtagging. The shi mutant, over-expressing SHIfrom a strong and constitutive promoter, re-sembles GA deficient dwarfs in several as-
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
34
pects; reduced internode elongation, darkergreen rosette leaves and delayed flowering inshort days. However, exogenous applicationof GA does not restore the dwarf phenotype ofshi. A recent hypothesis predicts that GA sig-nalling is regulated by a de-repressible sys-tem. Dr Sundberg is therefore testing wheth-er SHI is acting as a repressor of GA respon-ses. GA responses and signalling are wellcharacterised in barley aleurones. At the timeof germination, GA is transported from the em-bryo into the starchy endosperm of the barleygrain. GA signalling in the aleurone tissueresults in the activation of genes encoding hy-drolytic enzymes. These hydrolytic enzymesare secreted into the starchy endospermwhere the starch is degraded into solutes thatwill be used by the germinating embryo.
Dr. Sundberg could show that SHI, whenexpressed in barley aleurones, can repress
GA induction of the genes encoding hydrolyt-ic enzymes. This suggests that SHI at leastwhen ectopically expressed can act as a re-pressor of GA induced gene expression.
Constitutive SHI-expression also appearsto repress auxin responses, such as lateralroot initiation. Dr. Sundberg has screened forinsertion mutations in SHI and the other SHI-family members to get a better understandingof the gene function. Knock-out mutants ofboth SHI and another member of the genefamily, STY2, show no phenotypic deviationsfrom wild type suggesting that members ofthe family may have redundant functions.This is supported by the fact that constitutiveexpression of other SHI-family members con-fers similar changes as 35S::SHI.
The only member of the gene family whosefunction cannot be compensated for by otherfamily members is STY1. Sty1 shows a subtle
A recent hypothesis predicts that GA signalling is regulated by a de-repressible system. SHI, when expres-sed in barley aleurones, can repress GA induction of the genes encoding hydrolytic enzymes.Illustration: Eva Sundberg
GA receptornGA receptorn
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
35
style and stigma phenotype, which is en-hanced in sty1 sty2 double mutants. The stylecells are irregularly shaped and reduced innumber in single and double mutants. In ad-dition, a novel trait, leaf serration, appears inthe sty1 sty2 double mutant.
Conclusions: Members of the SHI gene fam-ily are involved in the control of cell expan-sion and perhaps also cell fate specification.They may regulate both gibberellin and aux-in responses. They exhibit partly redundantfunctions.
The hidden half: Dissecting thegenetic and hormonal controlof root developmentThere must be a balance between the rootsystem and the rest of the plant. The balanceis achieved through root shoot signallingmolecules. One of the most important signal-ling molecules is auxin, which is transportedto the root where it regulates root architec-ture. Classically, for some seventy years or so,it has been imagined that auxin is transport-ed to the root through polar auxin transport.Prof. Malcolm Bennett, Nottingham Universi-ty, challenges that concept.
The group at Nottingham University hasrecently demonstrated that auxin producedin aerial tissues has a profound effect on rootarchitecture. Following seedling germination,aerial derived auxin is transported to the roottip where it promotes lateral root initiation.Following seven days of growth, auxins pro-duced in the very first leaf buds promote theemergence of lateral roots. Hence, the signalsthat go between the shoot and the root are ofgreat importance to root architecture.
There are several different auxin carriers inplant cells. Auxin is transported into the cellthrough an influx carrier. Auxin then moves tothe base of the cell where it exits through anefflux carrier. Mutations in both of these car-riers have been identified in Arabidopsis.
Mutations in two auxin influx carrier genes,AUX1 and LAX3, both affect lateral root devel-opment. The aux1 lax3 double mutant exhib-its an additive lateral root phenotype. Thegenetic additivity is likely to result from thenon-overlapping expression patterns of AUX1and LAX3. The function of the AUX1 protein isto provide the cells of the developing lateralroot primordia with the ability to accumulateauxin and therefore promote cell division. Inthe case of LAX3, Prof. Bennett believes thatits role is to facilitate the radial redistributionof auxin from a vascular source to the devel-oping lateral root primordia. If both genes areknocked out it will be much more difficult forthe root cells to obtain auxin and divide.
Transgenics: Not “if”but “why”Let us now change perspectives and discussthe outlines and the results of the conference.In common opinion transgenics in agriculture– but not in medical use! – are controversial.GMOs are a fact of public preoccupation andopinion, which politicians must take into ac-count. There is further discussion of this issuelater in this report.
However, when scientists meet to reporton and exchange scientific achievementsthere are very few objections against thetechnology itself. Genetically modified orga-nisms (GMOs) are a fact of modern agricul-ture and are here to stay. The advantages aregreater than the disadvantages.
The starting point for Louise Fresco, FAO,was that genetic modification has increasedproduction in some crops but that the tech-nology has so far addressed too few challeng-es, in few crops of relevance to productionsystems in many developing countries. Ge-netic modification is not a good in itself, buta tool integrated into a wider research agen-
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
36
da, where public and private science can ba-lance each other. Scientists must not be blind-ed by the glamour of cutting-edge molecularscience for its own sake. Louise Fresco isworried about the disconnection between thelaboratory and the field and the decline ofagronomy as an integrative science. She wantsto see a scientific approach that in a globalisedworld serves local agriculture and local foodsecurity and helps keeping the rural areasliveable and attractive to young people insearch of employment.
Where are we heading?Gene technology can be utilised to increasethe quality of the plant material. So far, themost widely used transgenics are resistant topesticides rather than to pests. However, sci-ence seems – at least in the near future – to beable to meet the demand of consumers andproducers in developing healthier, strongerand better producing plants. The New Scien-tist has just recently reported that biotechresearchers in the US have knocked out agene in soya that causes allergic reactions.They used a “gene silencing” technique calledsense suppression, a technique that is alsoused by Donald Grierson and other scientistswho participated in the seminar.
Louise Fresco’s demands could have beenchallenging for the scientists present. Maybethey were – at least they agreed with her. How-ever, none of the scientists at the seminardevoted themselves to the poor people in thethird world – those for whom Louise Frescohad acted as a spokeswoman. Rather, the sci-entists devoted themselves to their science,albeit a science with many promising applica-tions: non rotting tomatoes, healthier oils andcrops that are resistant towards frost, droughtand diseases.
Genetic engineering uses expensive tech-niques – how are we to share the money?How can we be most efficient? How can scien-
tists, universities and research institutes re-tain their independence in scientific outlineand control of the results? Many importantquestions – but beyond the aim of this semi-nar.
But what is a transgenic?A transgenic plant is a plant with one genealtered – and all of a suddenit obtains com-pletely new properties. One gene might influ-ence the aroma and other characteristics thatin the mother organism seemed to be inde-pendent of the desired hereditary character.
A genetically modified organism – GMO – isdifferent from an organism developed withtraditional animal or plant breeding, as theGMO can contain genes from any living being,while the traditionally bred animal/plant canreceive hereditary characters only from indi-viduals from the same species.
It is still unclear what consequences theorigin of the gene might cause to the GMO.
Stable or unstable?A critical point with GMOs is the stability.When a GM experiment is carried out, therecan be a range of expression levels whichvary from one plant to another. Some of thoseare unstable and unwanted for further use.However, according to Donald Grierson’sexperience of tomatoes, it is quite easy toidentify, measure and select tens or maybehundreds of lines that are stable. Some of hisGM tomato lines are twelve years old.
Antisense techniqueAntisense technique means that isolatedgenes are replaced one by one in a “back-ward” order. Donald Grierson first introducedthe technique to study the ripening and
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
37
senescence process in tomatoes. However,the use is much wider than that. Silencing agene allows one to study the reactions of theorganism, to get a proper understanding ofthe biological function and role of the actualgene. It provides an exquisite and very pre-cise view of a gene’s function. The sense genemakes sense-RNA and the antisense genemakes antisense-RNA. Changing the structureof the sense gene affects the way in whichsilencing occurs.
Using anti-sense or knock-out techniques,beta-carotene genes in tomatoes and goldenrice have been identified. Beta-carotene is apro vitamin A and gives the tomato/rice ayellow colour.
What can be learnt fromArabidopsis thalianaArabidopsis thaliana is a tiny, plain plant – aweed to many people. Despite its humble na-ture, it has become famous because in theyear 2000, it became the first plant to be total-ly genetically sequenced. By analysing epi-genetics and chromosome structure it is pos-sible to understand the role of methylation inregulating chromosome structure and geneexpression. And now it seems that all scien-tists are working on Arabidopsis.
There are several reasons for working onArabidopsis thaliana. Arabidopsis has a veryshort growth cycle – results can be achievedwithin five weeks – and the plant has natural-ly a lot of varieties, which makes it suitable touse for finding tools for expressing naturalvariation. For instance, there exists a Stock-holm variety with a perennial behaviour, anadaptation to the short growth period to thelatitude of the Stockholm area.
In addition, with very little experimenta-tion plant genomes can be looked at in a veryefficient way because Arabidopsis can beused as a template to learn more about other
genomes. If there is high homology to anArabidopsis orthologue, gene function in Ara-bidopsis can be analysed because it is possi-ble to induce secondary growth in Arabidop-sis.
Surprisingly enough, poplar trees are clo-ser genetically to Arabidopsis than tomatoesor tobacco.
Food or non-foodapplications
Agriculture is in the first instance about foodproduction. With a scarcity of food in vastareas of the world, the food problem and sci-entific work on food has a priority in agricul-ture and horticulture.
However, starvation and food shortagescannot be sustainably solved with distribu-tion of food from the rich to the poor world.That fact opens up for non-food applications,at least in the rich world.
More and better food to feedthe worldThe biggest advantage with gene technologyis that once all the difficulties with the tech-nique have been solved, improvements willcome much quicker than with conventionalplant breeding.
Drought-stricken Southern African farmerswould benefit from a drought resistant maizevariety. A-vitaminised golden rice is already afact.
Fruit and vegetables with a prolongedgrowing period (with later senescence) aresuitable for farmers, trade and consumers.New unsaturated oils can reduce heart dis-ease.
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
38
Replace mineral oils withvegetable oilsThe annual consumption of mineral oil is3200 million tonnes, of which 220 milliontonnes are used in the chemical industry. Dr.Ulf Ståhl pointed out that after some morescientific work it would be possible to replacethese finite, non renewable and chemicallycomplex mineral oils, which require exten-sive chemical processing, with renewablevegetable oils with defined and desired qual-ities that do not require a lot of chemicalprocessing.
Of course, that is a complicated task and itis a scientific challenge restricted by publicopinion. The industrial capacity for industri-al crops is uncertain, since the existing indu-stry effectively forms a barrier against build-ing a new industry. Mineral oil is a difficultcompetitor as it is relatively cheap. New oilsmight be toxic, which means that they couldby no means be mixed with edible oils andthat measures would have to be taken to pro-tect the environment, birds and animal feeds.
Plastics are the most important potentialof oilseed plants.
Energy in the fieldSalix is a common energy crop in Sweden. Ithas been genetically improved and yieldsmuch more than the wild type. However, en-ergy could also be produced in other forms.Enzymes could make methane productionfrom manure more efficient, genetically mod-ified grasses could yield more energy andvegetable oil could replace mineral oil.
Designing starchStarch can be used both as a food and as anon food crop. “Designing” starch propertiescan be done in different ways.
• By modifying the amount of amylose it ispossible to modify the properties in a rad-ical way. A potato without amylose can forinstance be used for textile fibres.
• Amylopectin can change the starch syn-thesis and the branching enzymes. Thestarch from an amylopectin free potato ishardly swollen at all.
• Little is known about controlling granulesize, shape and number but they are be-lieved to be important. When isoamylase ispresent, glucan polymers/precursors in thestroma are degraded by isoamylase. Whenisoamylase is absent, the glucan polymers/precursors build up in the stroma. Elabora-tion by synthases and branching enzymesare interesting possibilities of the technique.Accumulation of the starch granules and/orthe phytoglycogen molecules are depend-ent on the synthetic enzymes available.
Forest engineering makeswonderTrees have an extremely long growth rotationand therefore traditional breeding is a slowimprovement process. Not surprisingly, sci-entists have turned to genetic engineering forsolutions to quality problems, slow growthand different kinds of stress sensitivity.
Just as interesting are the attempts to re-duce/replace chemicals in wood fibre process-ing with transgenics. The challenge is to de-sign wood quality for different purposes, forinstance produce a tree with less lignin to suitthe pulp and paper industry. The new tech-nique would save money and the environ-ment. But while creating a new tree, it is im-portant that laboratory studies, greenhouseexperiments and field trials go hand in hand.So far, the few harvested trees with reducedlignin have proved to be safe and can with-stand stormy weather, as well as other types ofstress. The growth and yield are unaffectedand it is important that they continue to be so.
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
39
Science, media andthe consumersLouise Fresco pointed out that as sharehold-ers in the GMO debate, scientists must recog-nise that there is also a substantial public dis-trust of science. Maybe this distrust opensthe way for non scientists and non scientificarguments in the debate. Donald Griersonhas participated in many television debatesand has been interviewed by the media manytimes. He was surprised and annoyed by theignorance and sometimes even the lack ofinterest in finding out the truth about GMfoods. Prof. Kristina Glimelius disagreed withthis. Often in ongoing research, it is hard tofind true facts and a number of facts must bepresented – some for and some against.
However, scientists have an interest and amoral obligation in participating in the publicdebate. To reach the public, they have to ex-plain their work in a simpler vocabulary thanthey use to address each other. Not every-body has that talent, but the talented scien-tists can get good publicity and outnumberignorant media giants.
Consumers are concernedPublic perception is necessary for the accept-ance of GM products.
Sometimes it is tempting to say that con-sumers are ignorant and overreact. There aremany examples of strange headlines such as“GM food stunts your growth”. But in factthere are also many sound and clever argu-ments against GM foods.
There are still unsolved problems in thetechnique and the applicability. And, proba-bly more important, people want to knowwhether they are buying GM products. Suchfoods must be labelled so that if consumersdo not approve of them, they have the choiceof not buying them.
Monsanto reduced the British GM marketto nil by marketing unlabelled GM soybeans,a practice that enraged the consumers. Themarket still has not recovered, nor has con-sumer confidence. There is a lesson to belearnt.
Legislation is neededMany countries still lack a thorough legisla-tion on GMOs. National and internationalagreements are important to set a standardbut also to decide what is legal and what isillegal.
A functioning legislation is necessary forfuture applications.
ConclusionsWhat Science Can Do in Crop and Forest Bio-technology of the Future was the theme of anInternational Conference on Crop and ForestBiotechnology for the Future in Falkenberg. Itwas held on September 17–19, 2001 and or-ganised by the Swedish Foundation for Stra-tegic Research (SSF), The Royal SwedishAcademy of Agriculture and Forestry (KSLA)and Bertebos Foundation.
That Genetically Modified Organisms(GMO) are a fact of modern agriculture andare here to stay was the starting point forLouise Fresco, Assistant Director General ofthe FAO. To ensure that GM crops make anoptimal contribution to world food security,food safety and sustainability, three greatproblems must be solved, according to Lou-ise Fresco. Firstly, GMOs must be directed tothe right research priorities such as droughtand heat tolerance, improved nutrient uptakeand rooting, biological nitrogen fixation, re-sponses to carbon dioxide and tolerance tokey abiotic stresses, such as salinity anddrought. Secondly, agreed national and inter-
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
40
national instruments of governance must bedeveloped. Thirdly, the access to researchand new technologies for developing coun-tries, poor producers and consumers mustbe facilitated.
The speech of Louise Fresco formed a back-ground to a wide range of scientific achieve-ments in the genetics of horticultural andagricultural plants and forest trees.
Ripening and senescence: Ripening and se-nescence go hand in hand, but by delayingsenescence in fruits and vegetables the sus-tainability is prolonged, which is an advanta-ge to producers, trade and consumers. TheBertebos prize winner of 2001, Prof. DonaldGrierson, showed from his own scientificwork on tomatoes that this is possible withan antisense technique. This is a very precisemethod and only one gene out of tens of thou-sands is inactivated. The effect is related tothe specific antisense gene that is used and itis inherited. From tomatoes with the antisen-se gene, one can produce progeny tomatoesthat have a delayed ripening process or alte-red quality. The key point is that if ethyleneproduction or the response to ethylene canbe controlled, the rate at which produce pro-gresses to overripeness and spoilage can beregulated. (Antisense technique means thatthe isolated genes are put back one by one ina “backward” order. By silencing a gene onecan study the reactions of the organism, toget a proper understanding of the biologicalfunction and role of the actual gene.)
Fatty acid modification: From a biochemicalpoint of view the daily consumption of fattyacids should include about 10 g of linoleicand 2 g of linolenic acid and a significant re-duction in all saturated fatty acids and a cor-responding increase in oleic acid. In view ofthis situation, it is an obvious idea to produ-ce plant seed oils containing very long-chainpolyunsaturated fatty acids normally onlyfound in fish (“oceanic“ fatty acids). This goal
may be realised by gene technology, providedthat the biochemistry of oil biosynthesis isknown in detail. At the end of the year 2002,most genes required for this task have beencloned, and only one is still missing.
Biotechnology can in the future be used todesign a desired oil quality in a desired oilcrop. A desired fatty acid can be obtainedwith the usage of the vast gene library in na-ture or by protein engineering.
Starch: Starch is the major form in whichplants store their carbon – around 80% of thedry weight in potato tubers and the endos-perm of cereals and up to 50% for peas andbeans. Starch is the main source of carbohy-drates in our diet but it also serves as a thick-ener, texturizer or stabilizer in both food andnon food industries.
Starch extracted from the crop (maize,wheat, potato, cassava etc) undergoes post-extraction processing consisting of chemical,enzymatic or physical modifications. Thisgenerates starches with many different prop-erties (different pastes and gels with varyingcharacteristics) suitable for a wide range ofdifferent industrial uses.
But post-extraction processing has somemajor disadvantages. It would be better if theright type of starch could be designed in thecrop so that post-extraction modificationwould not be needed. Scientists are beginningto gain sufficient information for “designing”starch properties in a rational way.
Genomics, proteomics and metabolomics:Arabidopsis thaliana was the first plant tohave its genome sequenced and by analysingepigenetics and chromosome structure, it ispossible to understand the role of methyla-tion in regulating chromosome structure andgene expression. Once a sequence of genes isobtained, this can be compared with all othergenes. The genome sequence provides a com-plete record of all of the genes and of all theprotein sequence because of the exquisite
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
41
precision of nucleotide sequence. From thatit is possible to study expression patterns ofgenes and to build up knowledge of co-regu-lated genes, the so-called regulon.
In the plant kingdom there exist a lot of dif-ferences in genome size. In spite of the largedifference in genome size, the general struc-ture of the chromosomes is often very similar.That means that the order of markers orgenes along the chromosomes is often verysimilar, if not identical.
Proteomics analyse the proteome compo-nents under a set of defined circumstances.To study the change in protein quantity, sci-entists consider several factors such as tissueor cell type, development stage, infection,disease, mutation, environmental effects andpesticides and herbicides.
A greater understanding of the controlprocesses regulating metabolism is requiredbefore metabolic engineering becomes rou-tine. This can be achieved by assessing thesteady-state concentrations of a wide rangeof metabolites and by the development ofsensitive and accurate methods of determin-ing cellular fluxes.
Plant stress and signalling: Ozone is a con-venient tool for studying the role of oxygenradicals in signalling (to get rid of the stress).ROS (reactive oxygen species) are importantbiological regulators of cell death during ox-idative stress. ROS regulation of cell death istightly coupled with hormonal regulation.ROS and hormones work as an interconnecti-ve network.
The fungal pathogen Leptosphaeria macu-lans causes blackleg or stem canker and is themajor threat to crops like oilseed rape andturnip rape. It has been demonstrated thatthe Arabidopsis thaliana genome can be asource of resistance to L. maculans. The re-sults of Arabidopsis work can be applied toBrassica.
The myrosinase-glucosinolate systemserves as a defence in oilseed rape. Glucos-
inolates are secondary metabolites found inall Brassica plants. These plants also housethe enzyme myrosinase, which degrades glu-cosinolates into toxic products, harmful topests. Down-regulation of vegetative andwound-inducible myrosinase binding pro-teins in Brassica napus affects susceptibilitydifferentially in different Brassica specialistherbivores.
The H+-ATPase is thought to play a majorrole in many cell processes and its activity isregulated by a number of physiological fac-tors, including hormones, blue light and fun-gal toxins. In all plants analysed so far, theplasma membrane H+-ATPase and 14-3-3 pro-teins are encoded by multigene families.
Forest genetics: Genetics can be used to im-prove quality and yield in forest trees. A Swe-dish project concentrates on two differenttypes of genes that are crucial to the practicalimplementation in forestry. One type affectsformation of wood and stem growth. Theother type is important for stress tolerance.Wood formation and ability to survive underharsh climates are unique characters fortrees.
Lignin biosynthesis can be used to im-prove wood quality for the pulp industry.With enzymes it is possible to reduce theamount of lignin for bleaching or to do somesurface modification of the cell walls to per-form better in paper making. Enzymes canmake cells react better with other agents. Inaddition, enzymes could improve the reactiv-ity of the main component, cellulose.
Plant development: Periods of growth anddormancy vary in trees The vascular cambi-um makes a transition from active to passivewhen the tree stops growing. The auxin trans-port is reduced as the tree stops growing, buteven after the cell divisions have stoppedthere is considerable capacity in the cambi-um to transport auxin for a long time. Auxin isone of the most important signalling mole-
Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
42
cules, which achieves the necessary balancebetween the root system and the rest of theplant.
Members of the SHI gene family are in-volved in the control of cell expansion and
perhaps also cell fate specification. They mayregulate auxin responses and the study ofgene control of these processes could be ofgreat benefit to forestry.
M. Larsson

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
43
Förteckning över tidigare utgivna nummer
År 2001; Årgång 140Nr 1 Sälen – resurs eller problemNr 2 Skogliga konsekvensanalyser 1999Nr 3 Framtida möjligheter till ökat utnyttjande av naturresurserNr 4 Verksamhetsberättelse 2000 Kungl. Skogs- och LantbruksakademienNr 5 Landskapet: restprodukt eller medvetet skapat?Nr 6 Ecologically Improved Agriculture – Strategy for SustainabilityNr 7 Sverige i det europeiska samarbetetNr 8 Bekämpningsmedel i vatten – vad vet vi om förekomst och effekter?Nr 9 Odlingssystem, växtnäring och markbördighet – 18 års resultat från en tidigare ej uppgödslad jordNr 10 Svenska fiskets framtid och samhällsnyttaNr 11 Vem sätter värde på den svenska skogen? – Trämekanisk framsynNr 12 Nutrition och folkhälsa i EU-perspektiv OCH Miljöpåverkan och framtidens matvanor OCH
Framtidens mat i framtidens kökNr 13 Konkurrenskraftig virkesförsörjning– ett kraftprov för skogsteknologin!Nr 14 Fritidsodlingens och stadsgrönskans samhällsnyttaNr 15 Debatt – Hur kan marknad och miljö förenas? Exemplet spannmålsproduktion
År 2002; Årgång 141Nr 1 Genteknik – en skymf mot Gud eller nya möjligheter för mänskligheten?Nr 2 Genmodifierade grödor. Varför? Varför inte? – Genetically Modified Crops. Why? Why Not?Nr 3 Skogsfrågor i ”Konventionen om biologisk mångfald”Nr 4 Mindre kväveförluster i foderodling, foderomvandling och gödselhantering!Nr 5 Bondens nya uppdrag OCH Shaping U.S. Agricultural PolicyNr 6 Verksamhetsberättelse 2001Nr 7 Sustainable forestry to protect water quality and aquatic biodiversityNr 8 Foder – en viktig länk i livsmedelskedjan!Nr 9 Fortbildning för landsbygdsutvecklareNr 10 Hållbart jordbruk – kunskapssammanställning och försök till syntesNr 11 Närproducerad mat. Miljövänlig? Affärsmässig? Djurvänlig?Nr 12 Nya kunskaper inom bioteknik och genetik för nya tillämpningar på husdjur SAMT
Fria Varuströmmar – konflikt med djur- och folkhälsa? Vilka möjliga utvägar finns? SAMT
Exempel på verksamhet inom JordbruksverketNr 13 Bland skärgårdsgubbar och abborrar på MöjaNr 14 EU och EMU – broms eller draghjälp för skogen?Nr 15 Hur kan skogsbruk och kulturmiljövård förenas?Nr 16 Vilket kött äter vi om 10 år? Rött, vitt, svenskt, importerat? Vi får det samhälle vi äter oss till!Nr 17 Avsättning av skogsmark
År 2003; Årgång 142Nr 1 Det sydsvenska landskapet, framtidsvisioner och framtidssatsningar SAMT
Idéer för framtidens skogslandskapNr 2 Viltets positiva värdenNr 3 Inför toppmötet i JohannesburgNr 4 Kapital för landsbygdsföretagareNr 5 Kompetensförsörjningen i svenskt jordbrukNr 6 Fiskets miljöeffekter – kan vi nå miljömålen?Nr 7 Verksamhetsberättelse 2002Nr 8 De glesa strukturerna i den globala ekonomin – kunskapsläge och forskningsbehovNr 9 Tro och vetande om husdjurens välfärd (Enbart publicerad på www.ksla.se)Nr 10 Svenska satsningar på ökad träanvändning (Enbart publicerad på www.ksla.se)Nr 11 Kapital för landsbygdsföretagare (Enbart publicerad på www.ksla.se)Nr 12 Feminisering av Moder natur? Östrogener i naturen och i livsmedelNr 13 Crop and Forest Biotechnology for the Future

K. Skogs-o. Lantbr.akad. Tidskr. 142:13, 2003
44
Kungl. Skogs- och Lantbruksakademiens Tidskrift (KSLAT) har, under olika namn, utkommit sedan1813, då akademien grundades. Från och med 1994 utges KSLAT som en numrerad serie av skrifter
(15–20 häften/år) med egna titlar. Innehållet består huvudsakligen av dokumentering frånakademiens sammankomster och seminarier – även debattnummer förekommer –
och speglar akademiens verksamhetsområde; de areella näringarnaoch till dessa knutna verksamheter.
Prenumerationspris 350 kr/år.
Kungl. Skogs- och LantbruksakademienDrottninggatan 95 B, Box 6806, 113 86 Stockholm
Tel 08-54 54 77 00, Fax 08-54 54 77 10, Postgiro 18 32 80 - 7
ISSN 0023–5350 ISBN 91–89379-55-1 Tex
t & T
ryck
Tot
ab A
B, E
skils
tuna
200
3
Pris 75 kr