Embed Size (px)
Citation preview

378
Conservation Biology, Pages 378–388Volume 15, No. 2, April 2001
Countryside Biogeography of Moths in a Fragmented Landscape: Biodiversity in Native and Agricultural Habitats
TAYLOR H. RICKETTS,* GRETCHEN C. DAILY, PAUL R. EHRLICH, ANDJOHN P. FAY
Center for Conservation Biology, Department of Biological Sciences, Gilbert Hall, 371 Serra Mall,Stanford University, Stanford, CA 94305–5020, U.S.A.
Abstract:
Studies of fragmented landscapes, especially in the tropics, have traditionally focused on the nativefragments themselves, ignoring species distributions in surrounding agricultural or other human-dominatedareas. We sampled moth species richness within a 227-ha forest fragment and in four surrounding agricul-tural habitats (coffee, shade coffee, pasture, and mixed farms) in southern Costa Rica. We found no signifi-cant difference in moth species richness or abundance among agricultural habitats, but agricultural siteswithin 1 km of the forest fragment had significantly higher richness and abundance than sites farther than
3.5 km from the fragment. In addition, species composition differed significantly between distance classes(but not among agricultural habitats), with near sites more similar to forest than far sites. These results sug-gest that (1) different agricultural production regimes in this region may offer similar habitat elements andthus may not differ substantially in their capacities to support native moth populations and (2) that the ma-jority of moths may utilize both native and agricultural habitats and move frequently between them, form-ing “halos” of relatively high species richness and abundance around forest fragments. Correlations betweenspecies richness and the amount of nearby forest cover, measured over circles of various radii around thesites, suggest that halos extend approximately 1.0–1.4 km from the forest edge. The extent of these halos likelydiffers among taxa and may influence their ability to survive in fragmented landscapes.
Biogeografía Rural de Polillas en un Paisaje Fragmentado: Biodiversidad en Hábitats Nativos y Agrícolas
Resumen:
Los estudios de paisajes fragmentados, especialmente en los trópicos, tradicionalmente se han en-focado en los fragmentos nativos per se, ignorando las distribuciones de especies en áreas agrícolas circun-dantes o en otras áreas dominadas por humanos. Muestreamos la riqueza de polillas dentro de un frag-mento de bosque de 227 hectáreas y en cuatro hábitats agrícolas (café, café con sombra, pastizal y camposmixtos) en el Sur de Costa Rica. Encontramos que no hubo diferencias significativas en la riqueza de especieso en la abundancia de polillas entre los hábitats agrícolas, sin embargo, los sitios agrícolas cercanos (
,
1 km)al fragmento de bosque tuvieron una riqueza de especies y abundancia significativamente mayor que las delos sitios lejanos (
.
3.5 km) al fragmento. Además, la composición de especies fue significativamente difer-ente entre las clases de distancia (pero no entre los hábitats agrícolas), siendo los sitios cercanos más simi-lares al bosque que los sitios retirados. Estos resultados sugieren que (1) los diferentes regímenes de produc-ción agrícola en esta región pueden ofrecer elementos de hábitat similares y por lo tanto pueden no diferirsubstancialmente en lo que se refiere a su capacidad para sostener poblaciones de polillas nativas y (2) quela mayoría de las polillas pueden utilizar tanto hábitatsnativos como agrícolas y mover frecuentemente en-tre ellos, formando “halos” con una riqueza de especies y una abundancia relativamente altas alrededor delos fragmentos del bosque. Las correlaciones entre la riqueza de especies y la cantidad de cobertura forestal
*
email [email protected] submitted August 4, 1999; manuscript accepted June 21, 2000.

Conservation BiologyVolume 15, No. 2, April 2001
Ricketts et al. Countryside Biogeography of Moths
379
circundante, medida en círculos de diferente radio alrededor de los sitios de estudio, sugiere que los halos seextienden aproximadamente 1.0–1.4 km del borde del bosque. La extension de estos halos posiblemente difi-
ere entre taxones y puede influenciar sus habilidades para sobrevivir en paisajes fragmentados.
Introduction
Although preservation of large areas of continuous, orig-inal habitat is crucial to the conservation of many spe-cies, human-dominated landscapes make up the majorityof land area in many parts of the world (Pimentel et al.1992; Vitousek et al. 1997). Fragmented landscapes thusare becoming increasingly important to the conserva-tion of biodiversity, and understanding the ecologicalprocesses at work in these landscapes has become oneof the central goals of conservation biology (Saunders etal. 1991; Robinson et al. 1992).
Studies of fragmented landscapes traditionally have fo-cused on the fragments themselves and ignored the sur-rounding agricultural or other human-dominated habitats,collectively dubbed the “matrix” (Laurance & Bierregaard1997). This patch-centric view has been driven in part bytwo bodies of theory commonly applied to fragmentationstudies: island biogeography and metapopulation dynam-ics, which consider fragments to be islands in an ocean-like matrix (MacArthur & Wilson 1967; Levins 1969;Soulé et al. 1992; Hanski 1998). Typically, however, thearea between patches of native habitat is not a uniformwasteland but comprises a variety of land-use types ofwidely differing—and sometimes considerable—capac-ity to support native biotas. Thus, an exclusive focus onnative fragments may miss important opportunities tounderstand biogeographic patterns and conserve biodi-versity in the overall landscape.
More recently, an increasing number of studies hastaken a more inclusive approach, considering fragmentedlandscapes as complex mosaics of habitats that vary inquality with respect to different groups of organisms (e.g.,Hansson & Angelstam 1991; Hobbs et al. 1993; Wiens1995; Gascon et al. 1999). Biodiversity studies in Europein particular have long included human-dominated habi-tats, in part because agricultural plots (e.g., unimprovedpasture) are now the primary habitat for many species(Thomas 1984).
The majority of studies that incorporate the matrix,however, remain focused on the remnants of native hab-itat and examine how the type of surrounding matrix af-fects species living in the fragments themselves (Aberget al. 1995; Sisk et al. 1997; Moilanen & Hanski 1998).Especially in the tropics, few studies directly examinebiogeographic patterns across the entire countryside byincluding both forest remnants and human-dominatedhabitats such as crops, pasture, fallow fields, and gar-
dens (Hutto 1989; Holloway et al. 1992; Perfecto & Snel-ling 1995; Power 1996; Greenberg et al. 1997; Daily etal. 2001). Such studies can provide insight into (1) thecapacities of different countryside habitats to supportnative biotas, (2) species characteristics that confer asurvival advantage in human-dominated areas, and (3)the movement patterns of organisms in complex land-scapes (Daily 1999).
Moths are a useful group for this kind of biogeographi-cal and conservation research. The Lepidoptera com-pose a diverse and abundant taxon in many forest sys-tems and play important roles as herbivores, pollinators,and prey ( Janzen 1987; Barlow & Woiwod 1989). In ad-dition, members of the Lepidoptera are often host-spe-cific ( Janzen 1988) and thus may serve as indicators ofnative plant diversity and local land management ( Er-hardt & Thomas 1991; Luff & Woiwod 1995). More than90% of known lepidopteran species are moths, themajority of which are nocturnal ( Janzen 1988; Young1997). These species can be surveyed readily in largenumbers with light traps, allowing relatively efficientestimation of geographic patterns of diversity and abun-dance.
We explored the biogeography of moths in a range ofcountryside habitats in southern Costa Rica. We sampledmoths in five habitat types—native forest, coffee, shadecoffee, pasture, and mixed farms—to address threequestions: (1) Do moth species richness, abundance,and composition differ among habitats? (2) Are mothspecies richness, abundance, and composition in agri-cultural sites related to proximity to native forest? (3) Dospecies with differing habitat affiliations show corre-sponding differences in discernible traits?
Methods
Study Sites
Prior to 1960, the Las Cruces area of Costa Rica was mostlycontinuous tropical moist forest. Now it is largely defor-ested, densely populated, and converted to a complexmosaic of agricultural habitats and forest fragments (Fig.1). Within this landscape lies the Las Cruces station ofthe Organization for Tropical Studies (OTS), with its227-ha forest reserve, the largest remaining forest frag-ment at this elevation within 15 km.
We sampled moth species at 16 sites (Table 1, Fig. 1),

380
Countryside Biogeography of Moths Ricketts et al.
Conservation BiologyVolume 15, No. 2, April 2001
representing the five major habitats in the region: (1)coffee grown in full sun in plots of approximately 1–30ha, often with bananas planted sparsely between rows;(2) shade coffee, also typically grown in plots of 1–30ha, planted among monospecific stands of shade trees(often legumes, such as
Erythrina
sp. and
Inga
sp.); (3)cattle pastures of approximately 10–30 ha planted in ex-otic grasses and typically containing scattered nativetrees to provide shade; (4) mixed farms, typically 1–3ha, containing small plots (
,
0.5 ha) of pasture, coffee,sugar cane, yucca, vegetable gardens, and abandoned orfallow areas; and (5) the Las Cruces forest reserve.Smaller forest fragments, typically 1–30 ha, were scat-tered throughout the landscape, but we did not samplein these areas (Fig. 1).
For coffee, pasture, and mixed sites, we selected twosites
,
1 km from the Las Cruces forest reserve (hereaf-
ter “near sites”) and two sites
.
3.5 km from the reserve(“far sites”) (Table 1; Fig. 1). We sampled only oneshade coffee site in each distance class because we wereunable to locate others. The two distance classes werechosen arbitrarily, with the intent to capture the ex-tremes of a possible distance gradient in moth richnessand community composition.
The forest map (Fig. 1) was classified from a 1994Landsat TM image. Details of the image classification aredescribed elsewhere (Daily et al. 2000). Although sitesP-F2 and M-F2 appear to be near large forest patches, thetopography at the southern edge of the map (Fig. 1)dropped steeply away from the area containing ourstudy sites (approximately 1100 m above sea level) tothe coastal plain (approximately 30 m above level). Con-sequently, the forests depicted at the southern edge ofthe map occurred at much lower elevations and were
Figure 1. Map of Las Cruces region of southern Costa Rica, showing the distribution of forest remnants (shaded areas) and locations of moth sampling sites (site codes as in Table 1). Large forest areas visible on the southern edge of the map oc-cur at much lower elevations and were not included in the study.

Conservation BiologyVolume 15, No. 2, April 2001
Ricketts et al. Countryside Biogeography of Moths
381
unlikely to influence the moth diversity in these agricul-tural sites. We therefore considered sites P-F2 and M-F2far sites because they were located
.
3.5 km from anylarge patches at similar elevations.
Moth Sampling
To sample moths we used bucket traps with 12-watt fluo-rescent black lights powered by 12-volt batteries. Mothsare attracted to the light, strike transparent baffles sur-rounding the lamp, and fall through a funnel into abucket below. We placed strips of solid insecticide (2.2dichlorovinyl phosphate) inside each bucket to kill thetrapped moths. Traps either were placed on the groundor were hung from trees or fence posts 1–2 m from theground so that the light would not be quickly attenuatedby dense understory vegetation or coffee shrubs. Trapheights were kept constant within each habitat.
There is considerable uncertainty about the distanceover which moths are effectively attracted to traps ofthis kind. Although most evidence indicates a “radius ofattraction” of 50–200 m, published estimates vary from
,
5 m to
.
700 m (Dufay 1964; Hartstack et al. 1971;Baker & Sadovy 1978; McGeachie 1997). We thus tookgreat care when selecting sites to ensure that thelight traps illuminated only the target habitat. First, wemasked each trap with black sheet plastic for two-thirdsof its circumference, so we could “aim” traps to illumi-nate only the target habitat and minimize draw fromother habitats. Second, we placed each trap at the bot-tom of a small valley or hillside consisting of only the tar-get habitat so that, no matter how large the radius of at-traction was, the trap was effectively shielded fromilluminating other habitats. No forest patches were di-rectly visible from any of the agricultural sites. The twoforest traps, although located over 500 m from the near-
est forest edge, were also masked for two-thirds of theircircumferences to keep sampling effort equal.
We sampled sites in rotation between 26 January and1 March 1998, with four to eight traps in operation eachnight. Traps were operated for 2 hours, between 1.5 and3.5 hours after sunset; thus, we could guard themagainst vandalism and limit the number of individualssampled. In all, eight samples were collected per site.We collected over a full lunar cycle to control for the ef-fects of the moon on trap effectiveness (Nag & Nath1991), and we arbitrarily reassigned traps and batteriesto sites each night to remove any bias of lamp brightnessor battery strength.
We sorted the moth samples to morphospecies, basedon external morphology and color patterns of wings, an-tennae, and bodies. Morphospecies richness has beenshown to correlate strongly with actual species richnessin samples of several taxa (e.g., Oliver & Beattie 1996).Although restricting analyses to morphospecies sacri-fices potentially useful autecological information (Gold-stein 1997), tropical insect faunas often contain manyundescribed species, and ecological data are typicallysparse and unreliable ( Janzen 1988). Therefore, sortingspecimens to morphospecies, although not ideal, offersan efficient method of estimating relative species rich-ness in highly diverse and poorly documented taxa. Indi-viduals with wing lengths of
,
12 mm were excluded toavoid the high rate of sorting errors among microlepi-doptera. We retained up to four specimens of each mor-phospecies. For simplicity, we hereafter refer to mor-phospecies as species.
Analyses
We analyzed the effects of habitat and distance class onmoth species richness with a two-way analysis of vari-
Table 1. Moth sampling results for the 16 sampling sites in the Las Cruces area of Costa Rica.
Site code Habitat Distance class
a
Richness Richness (
.
1)
b
Abundance
Forest 1 forest — 129 34 200Forest 2 forest — 174 55 307M-N1 mixed near 84 19 172M-N2 mixed near 140 40 440M-F1 mixed far 79 21 175M-F2 mixed far 47 15 97P-N1 pasture near 128 33 270P-N2 pasture near 139 57 482P-F1 pasture far 33 9 58P-F2 pasture far 38 8 54C-N1 coffee near 93 18 139C-N2 coffee near 115 27 172C-F1 coffee far 34 5 43C-F2 coffee far 27 5 43SC-N shade coffee near 74 23 132SC-F shade coffee far 49 16 82
a
Near,
,
1 km from Las Cruces forest reserve; far,
.
3.5 km from Las Cruces forest reserve.
b
Nonsingleton species richness, the number of species represented in a site by more than one individual.

382
Countryside Biogeography of Moths Ricketts et al.
Conservation BiologyVolume 15, No. 2, April 2001
ance (ANOVA). No estimate of variance was possible forshade coffee sites, so we excluded shade coffee fromthe ANOVA. To examine the possibility of an errant re-sult due to the high number of rare or vagrant species,we repeated the ANOVA and excluded species collectedonly once in a given site, termed singletons. We also re-peated the ANOVA using total abundance instead of spe-cies richness.
To examine the similarity of species compositionamong sites, we calculated pairwise Jaccard similaritycoefficients (Colwell & Coddington 1994). The Jaccardindex is a ratio of the number of species shared by twosamples to the number of total species in the twopooled samples. We used a multidimensional scaling(MDS) algorithm (SYSTAT 7.0) to reveal any patterns incommunity similarity among sites. In addition, we testedwhether sites were clustered significantly by agriculturalhabitat or by distance class in the MDS plot, using thePRIMER software package (Carr 1997). PRIMER usesrandomization techniques to estimate the probability ofobtaining the observed degree of clustering by chance.
For each treatment of habitat and distance class (Ta-ble 1), we calculated the proportion of unique species,(i.e., those found in only that treatment). To reduce thepossible influence of rare or vagrant species on these re-sults, we repeated this analysis using only species foundin a given treatment more than once.
Jerry Powell at the University of California at Berkeleyhelped us assign 643 of the 705 morphospecies (91%) tofamilies. Using these 643 species, we compared mothfamily composition among habitats and between dis-tance classes using
R
3
C
tests of independence.Wing length generally correlates with overall body
size in the Lepidoptera and is often assumed to indicatepotential flight strength, although this relationship hasrarely been tested ( Van Dyke & Matthysen 1999). Wemeasured the forewing length of all retained specimensto estimate the mean wing length of each species. Wethen computed the mean wing length of the samplesfrom each distance class and compared the means usinga one-way ANOVA.
To further examine the scale of the response of mothdiversity to local forest cover, we tested the correlationbetween species richness and amount of nearby forestmeasured at a variety of geographic scales. Using a geo-graphic information system (GIS), we calculated the areaof forest inside circles of 400-m radius around each siteand correlated these measures of forest cover with mothspecies richness across the 14 agricultural sites. We thenrepeated the process, increasing the radii of the circlesby 200 m each time, up to 2400 m (Fig. 2). By graphingthe strength of these correlations against the radii of thecircles over which forest cover is measured, we canidentify the neighborhood scale(s) at which moth spe-cies richness is most strongly correlated with the densityof nearby forest cover (including both the Las Cruces re-
serve and smaller forest fragments). In this analysis weignored forests to the south and southwest of our studyarea (Fig. 2) that occurred at substantially lower eleva-tions. We set an elevation filter in the GIS to exclude allforests below 700 m above sea level (400 m below thegeneral elevation of the study area).
The measures of local forest cover for the 14 siteswere not independent because circles from differentsites overlapped substantially (Fig. 2). This violates a ba-sic assumption of correlation analyses (Hargrove & Pick-ering 1992). The problem is especially severe among thenear sites, which are clustered around the Las Crucesforest reserve. In an effort to reduce this problem, weexcluded four of the near sites (one near site of each ag-ricultural habitat) and repeated the scale analysis. Tocompletely remove the effect of the Las Cruces reserve
Figure 2. Concentric circles used to measure nearby forest cover at different spatial scales. For clarity, only circles of radius 400 m and 800 m are shown.
Conservation BiologyVolume 15, No. 2, April 2001
Ricketts et al. Countryside Biogeography of Moths
383
on the measurements of nearby forest cover, we re-peated the analysis excluding all near sites.
Results
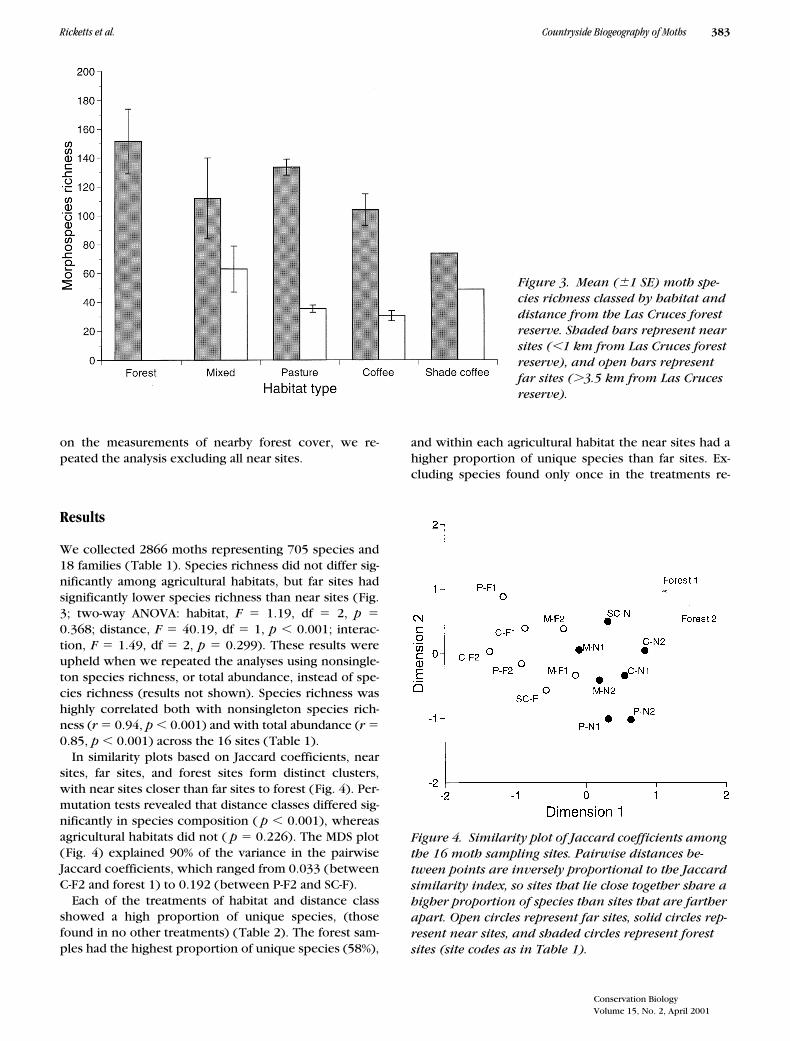
We collected 2866 moths representing 705 species and18 families (Table 1). Species richness did not differ sig-nificantly among agricultural habitats, but far sites hadsignificantly lower species richness than near sites (Fig.3; two-way ANOVA: habitat,
F
5
1.19, df
5
2,
p
5
0.368; distance,
F
5
40.19, df
5
1,
p
,
0.001; interac-tion,
F
5
1.49, df
5
2,
p
5
0.299). These results wereupheld when we repeated the analyses using nonsingle-ton species richness, or total abundance, instead of spe-cies richness (results not shown). Species richness washighly correlated both with nonsingleton species rich-ness (
r
5
0.94,
p
,
0.001) and with total abundance (
r
5
0.85,
p
,
0.001) across the 16 sites (Table 1).In similarity plots based on Jaccard coefficients, near
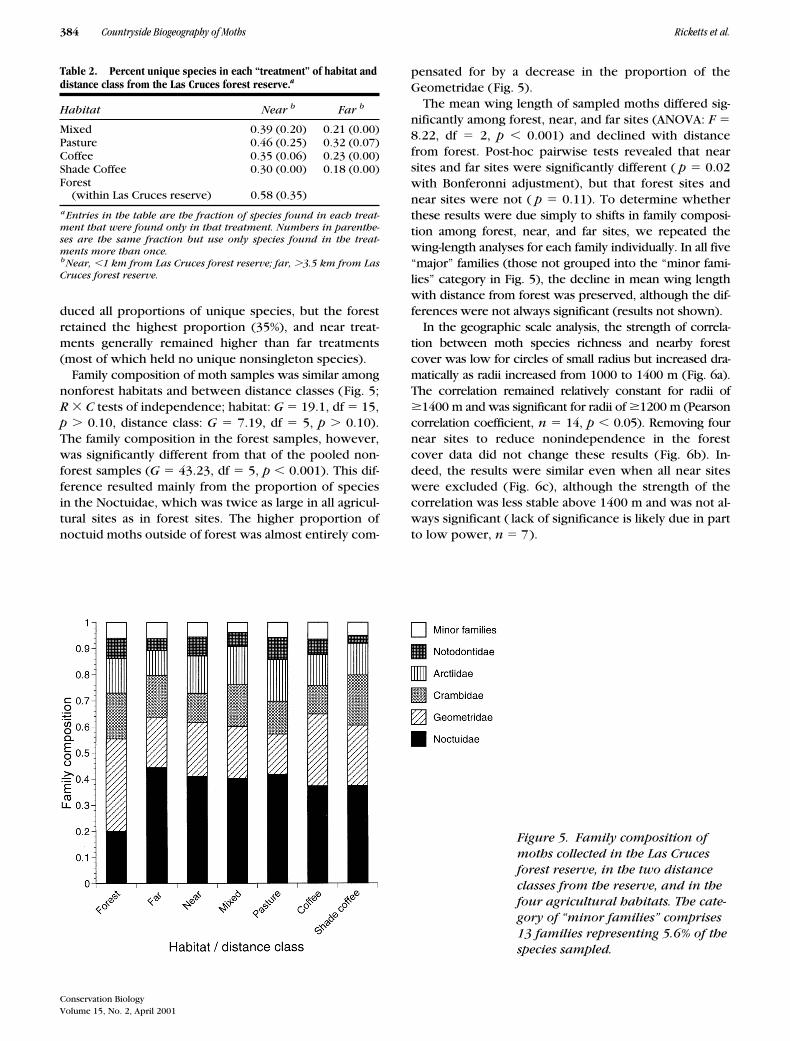
sites, far sites, and forest sites form distinct clusters,with near sites closer than far sites to forest (Fig. 4). Per-mutation tests revealed that distance classes differed sig-nificantly in species composition (
p
,
0.001), whereasagricultural habitats did not (
p
5
0.226). The MDS plot(Fig. 4) explained 90% of the variance in the pairwiseJaccard coefficients, which ranged from 0.033 (betweenC-F2 and forest 1) to 0.192 (between P-F2 and SC-F).
Each of the treatments of habitat and distance classshowed a high proportion of unique species, (thosefound in no other treatments) (Table 2). The forest sam-ples had the highest proportion of unique species (58%),
and within each agricultural habitat the near sites had ahigher proportion of unique species than far sites. Ex-cluding species found only once in the treatments re-
Figure 3. Mean (61 SE) moth spe-cies richness classed by habitat and distance from the Las Cruces forest reserve. Shaded bars represent near sites (,1 km from Las Cruces forest reserve), and open bars represent far sites (.3.5 km from Las Cruces reserve).
Figure 4. Similarity plot of Jaccard coefficients among the 16 moth sampling sites. Pairwise distances be-tween points are inversely proportional to the Jaccard similarity index, so sites that lie close together share a higher proportion of species than sites that are farther apart. Open circles represent far sites, solid circles rep-resent near sites, and shaded circles represent forest sites (site codes as in Table 1).
384
Countryside Biogeography of Moths Ricketts et al.
Conservation BiologyVolume 15, No. 2, April 2001
duced all proportions of unique species, but the forestretained the highest proportion (35%), and near treat-ments generally remained higher than far treatments(most of which held no unique nonsingleton species).
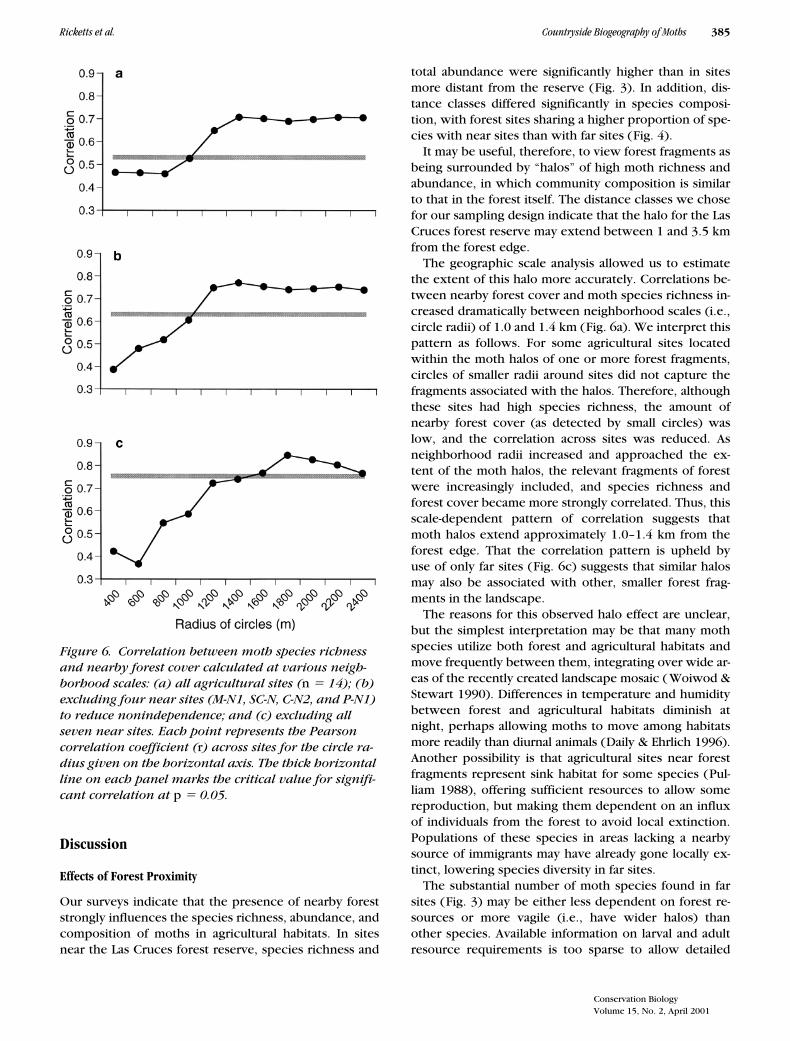
Family composition of moth samples was similar amongnonforest habitats and between distance classes (Fig. 5;
R
3
C
tests of independence; habitat:
G
5
19.1, df
5
15,
p
.
0.10, distance class:
G
5
7.19, df
5
5,
p
.
0.10).The family composition in the forest samples, however,was significantly different from that of the pooled non-forest samples (
G
5
43.23, df
5
5,
p
,
0.001). This dif-ference resulted mainly from the proportion of speciesin the Noctuidae, which was twice as large in all agricul-tural sites as in forest sites. The higher proportion ofnoctuid moths outside of forest was almost entirely com-
pensated for by a decrease in the proportion of theGeometridae (Fig. 5).
The mean wing length of sampled moths differed sig-nificantly among forest, near, and far sites (ANOVA:
F
5
8.22, df
5
2,
p
,
0.001) and declined with distancefrom forest. Post-hoc pairwise tests revealed that nearsites and far sites were significantly different (
p
5
0.02with Bonferonni adjustment), but that forest sites andnear sites were not (
p
5
0.11). To determine whetherthese results were due simply to shifts in family composi-tion among forest, near, and far sites, we repeated thewing-length analyses for each family individually. In all five“major” families (those not grouped into the “minor fami-lies” category in Fig. 5), the decline in mean wing lengthwith distance from forest was preserved, although the dif-ferences were not always significant (results not shown).
In the geographic scale analysis, the strength of correla-tion between moth species richness and nearby forestcover was low for circles of small radius but increased dra-matically as radii increased from 1000 to 1400 m (Fig. 6a).The correlation remained relatively constant for radii of
$
1400 m and was significant for radii of
$
1200 m (Pearsoncorrelation coefficient,
n
5
14,
p
,
0.05). Removing fournear sites to reduce nonindependence in the forestcover data did not change these results (Fig. 6b). In-deed, the results were similar even when all near siteswere excluded (Fig. 6c), although the strength of thecorrelation was less stable above 1400 m and was not al-ways significant ( lack of significance is likely due in partto low power,
n
5
7).
Table 2. Percent unique species in each “treatment” of habitat and distance class from the Las Cruces forest reserve.
a
Habitat Near
b Far b
Mixed 0.39 (0.20) 0.21 (0.00)Pasture 0.46 (0.25) 0.32 (0.07)Coffee 0.35 (0.06) 0.23 (0.00)Shade Coffee 0.30 (0.00) 0.18 (0.00)Forest
(within Las Cruces reserve) 0.58 (0.35)aEntries in the table are the fraction of species found in each treat-ment that were found only in that treatment. Numbers in parenthe-ses are the same fraction but use only species found in the treat-ments more than once.bNear, ,1 km from Las Cruces forest reserve; far, .3.5 km from LasCruces forest reserve.
Figure 5. Family composition of moths collected in the Las Cruces forest reserve, in the two distance classes from the reserve, and in the four agricultural habitats. The cate-gory of “minor families” comprises 13 families representing 5.6% of the species sampled.
Conservation BiologyVolume 15, No. 2, April 2001
Ricketts et al. Countryside Biogeography of Moths 385
Discussion
Effects of Forest Proximity
Our surveys indicate that the presence of nearby foreststrongly influences the species richness, abundance, andcomposition of moths in agricultural habitats. In sitesnear the Las Cruces forest reserve, species richness and
total abundance were significantly higher than in sitesmore distant from the reserve (Fig. 3). In addition, dis-tance classes differed significantly in species composi-tion, with forest sites sharing a higher proportion of spe-cies with near sites than with far sites (Fig. 4).
It may be useful, therefore, to view forest fragments asbeing surrounded by “halos” of high moth richness andabundance, in which community composition is similarto that in the forest itself. The distance classes we chosefor our sampling design indicate that the halo for the LasCruces forest reserve may extend between 1 and 3.5 kmfrom the forest edge.
The geographic scale analysis allowed us to estimatethe extent of this halo more accurately. Correlations be-tween nearby forest cover and moth species richness in-creased dramatically between neighborhood scales (i.e.,circle radii) of 1.0 and 1.4 km (Fig. 6a). We interpret thispattern as follows. For some agricultural sites locatedwithin the moth halos of one or more forest fragments,circles of smaller radii around sites did not capture thefragments associated with the halos. Therefore, althoughthese sites had high species richness, the amount ofnearby forest cover (as detected by small circles) waslow, and the correlation across sites was reduced. Asneighborhood radii increased and approached the ex-tent of the moth halos, the relevant fragments of forestwere increasingly included, and species richness andforest cover became more strongly correlated. Thus, thisscale-dependent pattern of correlation suggests thatmoth halos extend approximately 1.0–1.4 km from theforest edge. That the correlation pattern is upheld byuse of only far sites (Fig. 6c) suggests that similar halosmay also be associated with other, smaller forest frag-ments in the landscape.
The reasons for this observed halo effect are unclear,but the simplest interpretation may be that many mothspecies utilize both forest and agricultural habitats andmove frequently between them, integrating over wide ar-eas of the recently created landscape mosaic ( Woiwod &Stewart 1990). Differences in temperature and humiditybetween forest and agricultural habitats diminish atnight, perhaps allowing moths to move among habitatsmore readily than diurnal animals (Daily & Ehrlich 1996).Another possibility is that agricultural sites near forestfragments represent sink habitat for some species (Pul-liam 1988), offering sufficient resources to allow somereproduction, but making them dependent on an influxof individuals from the forest to avoid local extinction.Populations of these species in areas lacking a nearbysource of immigrants may have already gone locally ex-tinct, lowering species diversity in far sites.
The substantial number of moth species found in farsites (Fig. 3) may be either less dependent on forest re-sources or more vagile (i.e., have wider halos) thanother species. Available information on larval and adultresource requirements is too sparse to allow detailed
Figure 6. Correlation between moth species richness and nearby forest cover calculated at various neigh-borhood scales: (a) all agricultural sites (n 5 14); (b) excluding four near sites (M-N1, SC-N, C-N2, and P-N1) to reduce nonindependence; and (c) excluding all seven near sites. Each point represents the Pearson correlation coefficient (r) across sites for the circle ra-dius given on the horizontal axis. The thick horizontal line on each panel marks the critical value for signifi-cant correlation at p 5 0.05.

386 Countryside Biogeography of Moths Ricketts et al.
Conservation BiologyVolume 15, No. 2, April 2001
analysis of any shared characteristics among speciesfound in far sites ( Janzen 1988). At a coarser taxonomiclevel, however, we found no differences in family com-position between distance classes, suggesting that dis-tance effects do not differ markedly among moth families.If vagility were an important factor, we would expectstrong-flying species to be overrepresented in far sites.The trend in wing length from different distance classes,however, was the opposite of what we expected: meanwing length decreased with distance from forest. Winglength is an untested indicator of flight strength in moths,however, making interpretation of this result tenuous.
The extent of any forest-related halos probably variesamong taxonomic groups. In the same area of CostaRica, Daily et al. (2001) found no decline in species rich-ness or abundance of birds, even at distances of 6 kmfrom large forest patches. In a German agricultural land-scape, Tscharntke et al. (1998) found that diversity andabundance of natural enemies of trap-nesting hymenopteradeclined abruptly with isolation of over 600 m from pri-mary habitat. Taxa with larger halos might be expected tohave a survival advantage in fragmented landscapesbecause populations isolated in a single fragment arewidely thought to face increased extinction risk from de-mographic, environmental, and genetic factors (e.g.,Meffe & Carroll 1997). Identifying traits such as vagilityand resource generality that accompany large-diversityhalos may therefore help predict the consequences ofhabitat fragmentation for native biotas.
Effects of Habitat
Three lines of evidence suggest that local crop types andproduction regimes do not strongly influence patternsof moth diversity in this landscape. First, species rich-ness and total abundance did not differ significantlyamong agricultural habitats (Fig. 3). Second, the speciescomposition analyses revealed that sites did not clustersignificantly by agricultural habitat (Fig. 4). Third, wefound no significant differences in moth family composi-tion among nonforest habitats (although Noctuidae werestrongly overrepresented in all agricultural habitats rela-tive to forest; Fig. 5).
At least three factors may be responsible for this unex-pected result. First, moths may not be utilizing resourcesin agricultural habitats at all, and they may have beensampled only while moving among forest patches. Sec-ond, our crop-based habitat classification may be irrele-vant to moths. Any resources in agricultural sites arelikely to be found not in the crops themselves (e.g., cof-fee shrubs, exotic pasture grasses), but in small areas offallow or abandoned land, crop verges, seminatural ri-parian strips, or small pockets of native vegetation.These resources may be relatively homogeneously dis-tributed across the countryside, regardless of the type ofagricultural land use. Third, some agricultural habitats
may contain more resources than others, but the vagilityof many moths obscures these differences in our sam-ples. For example, mixed agricultural plots typicallycontained more diverse vegetation and seminatural areasthan other agricultural habitats and thus may have con-tributed disproportionately to the resources available inthe agricultural landscape. If these resources are impor-tant to moths, then replacing small, diverse farms withmore extensive and homogeneous production regimesmay reduce species richness and abundance in the land-scape in general. Comparing species diversity in landscapeswith different proportions of crop types and managementregimes may help to illuminate these issues further.
In our surveys, 58% of the species collected in the LasCruces reserve were not found anywhere outside theforest (Table 2). The proportion of species unique toeach of the treatments of habitat and distance class wasalso considerable, however, making it difficult to esti-mate what fraction of the moth fauna truly was re-stricted to forest. One relatively clear pattern, however,is that far treatments consistently had lower proportionsof unique species than did near treatments. This resultsuggests that species found in far sites tend to be rela-tively widespread or common, a finding also reported byHolloway et al. (1992).
Several authors have compared species diversity amongnatural and disturbed habitats in the tropics, but togetherthe studies yield few generalities. Janzen (1973), in sur-veys of Costa Rican beetles and bugs, reported sharp dropsin diversity from primary forest to disturbed habitats butonly subtle differences among disturbance types, a resultsimilar to ours. Holloway et al. (1992) found similar pat-terns in Indonesian moths, but samples of carrion beetlesin the same sites showed no differences in diversity be-tween native and disturbed habitats, perhaps because ofthe generality of carrion beetle diets. Holloway also re-ported a large increase in the proportion of noctuid speciesoutside intact forest, a result similar result to ours (Fig. 5).
Roth et al. (1994) found that the species richness ofground-foraging ants in Costa Rica was significantlylower in all agricultural habitats than in forest. In addi-tion, species richness declined significantly with increasedagricultural intensity among crop types. Differences in in-dividual vagility may help explain why agricultural habitatsappear to influence diversity patterns in ants more than inmoths. Roth et al. (1994) point out, however, that intheir study agricultural intensity and proximity to forestare confounded; the most intensely managed cropswere also farthest from native forest. Therefore, theymay have measured in part the effects of distance as wellas management practice.
Caveats
Several caveats deserve mention. First, our data incorpo-rate some degree of pseudoreplication (Hargrove &

Conservation BiologyVolume 15, No. 2, April 2001
Ricketts et al. Countryside Biogeography of Moths 387
Pickering 1992). Most important, our near sites all sur-rounded a single fragment, the Las Cruces forest reserve(Fig. 1). We see no reason to suspect that the reservewas not representative of local forest, and because itwas the only remnant of any size remaining in the area,suitable replicates were not available. Second, althoughwe found no significant differences among agriculturalhabitats in moth species richness, the low statisticalpower of these tests (only four sites of each habitat) mayrender any real differences difficult to detect. Whateverdifferences actually existed among agricultural habitats,however, they were probably small compared to the in-fluence of nearby forest (Fig. 3).
Third, tropical insect faunas are notoriously diverseand difficult to sample adequately, and species accumu-lation curves for our sites indicate little reduction inslope toward an asymptote. Other studies of tropical Lep-idoptera have reported nonasymptotic species accumula-tion curves, even after a full year of sampling (Price et al.1995; Devries et al. 1997). Because species richness andtotal abundance were correlated across our sites, onemay suspect that we sampled a gradient in abundancebut not species richness (Gotelli & Graves 1996). The sim-ilarity plot (Fig. 4), however, provides strong evidence ofan actual faunal shift between distance classes, not simplythe effect of different-sized samples of the same speciespool.
Fourth, because our analyses were based on identicalsampling effort (trap hours), our results rest on the as-sumption that sampling efficiency was equivalent amongsites. Differences in sampling efficiency would influenceour results if, for example, denser vegetation reducedthe attraction radius of traps in some sites, resulting in asmaller sample of individuals and lower apparent spe-cies richness. Although this problem may affect compar-isons among habitats, which clearly vary in vegetationdensity (e.g., pasture vs. forest), there is no reason to ex-pect that trap efficiencies will differ systematically be-tween near and far sites of the same habitat. Thus, thedistance results we report are probably relatively robustwith respect to this potential problem, but habitat com-parisons should be regarded with greater caution.
Finally, although our results raise several points ofgeneral interest, our findings are limited by sampling de-sign. First, we sampled only nocturnal moths larger than12 mm in wing length that are attracted to light traps.Diurnal species and the diverse microlepidopteran faunamay exhibit different patterns than those we found. Sec-ond, we sampled moths for only a portion of a singleyear. The diversity, abundance, and composition of lepi-dopteran communities have been shown to change sea-sonally (Devries et al. 1997) and between years ( Wolda1978). Additional studies with moths and other taxa indifferent seasons will help determine the generality ofthe patterns we documented here.
Acknowledgments
We thank the farmers surrounding Las Cruces who withgood humor permitted us to sample from their proper-ties at night. G. Burtness, E. Bush, D. Campos, T. Davis,A. Ehrlich, J. Hughes, and R. Murillo provided valuableassistance with trapping and sorting. A. Sánchez-Azofeifadeveloped the forest map of our study area, J. Powellidentified our morphospecies to family, and C. Boggssupplied the inspiration for the neighborhood scale anal-yses. We thank the staff of the Las Cruces station of theOrganization for Tropical Studies for general field assis-tance and support. S. Bailey, C. Boggs, K. Carney, R. Car-roll, C. Devine, J. Hellmann, J. Hughes, K. Holl, J. Pow-ell, N. Sanders, and two anonymous reviewers improvedthe manuscript with their thoughtful comments. Sup-port was provided by the Winslow Foundation, the Ko-ret Foundation, the National Aeronautics and Space Ad-ministration, P. and H. Bing, and a National ScienceFoundation graduate fellowship to T.H.R.
Literature Cited
Aberg, J., G. Jansson, J. E. Swenson, and P. Angelstam. 1995. The effectof matrix on the occurrence of hazel grouse (Bonasa bonasia) inisolated habitat fragments. Oecologia 103:265–269.
Baker, R. R., and Y. Sadovy. 1978. The distance and nature of the lighttrap response of moths. Nature 276:818–821.
Barlow, H. S., and I. P. Woiwod. 1989. Moth diversity of a tropical for-est in Peninsular Malaysia. Journal of Tropical Ecology 5:37–50.
Carr, M. R. 1997. PRIMER user manual (Plymouth Routines in Multi-variate Ecological Research). Plymouth Marine Laboratory, Ply-mouth, United Kingdom.
Colwell, R. K., and J. A. Coddington. 1994. Estimating terrestrial biodi-versity through extrapolation. Philosophical Transactions of theRoyal Society of London, Series B 345:101–118.
Daily, G. C. 1999. Developing a scientific basis for managing Earth’slife-support systems. Conservation Ecology 3: http://www.consecl.org/vol3/iss?/art14.
Daily, G. C., and P. R. Ehrlich. 1996. Nocturnality and species survival.Proceedings of the National Academy of Sciences of the UnitedStates of America 93:11709–11712.
Daily, G. C., P. R. Ehrlich, and G. A. Sánchez-Azofeifa. 2000. Countrysidebiogeography: utilization of human dominated habitats by the avi-fauna of southern Costa Rica. Ecological Applications 11:1–13.
Devries, P. J., D. Murray, and R. Lande. 1997. Species diversity in verti-cal, horizontal, and temporal dimensions of a fruit-feeding butterflycommunity in an Ecuadorian rainforest. Biological Journal of theLinnean Society 62:343–364.
Dufay, C. 1964. Contribution a l’etude du phototropism des Lepidop-tres noctuids. Annales des Sciences Naturelles Zoologie et BiologieAnimale 6:281–406.
Erhardt, A., and J. A. Thomas. 1991. Lepidoptera as indicators ofchange in the semi-natural grasslands of lowland and upland Eu-rope. Pages 213–236 in N. M. Collins and J. A. Thomas, editors. Theconservation of insects and their habitats. Academic Press, London.
Gascon, C., T. E. Lovejoy, R. O. Bierregaard, J. R. Malcolm, P. C.Stouffer, H. L. Vasconcelos, W. F. Laurance, B. Zimmerman, M.Tocher, and S. Borges. 1999. Matrix habitat and species richness intropical forest remnants. Biological Conservation 91:223–229.
Goldstein, P. Z. 1997. How many things are there? A reply to Oliver

388 Countryside Biogeography of Moths Ricketts et al.
Conservation BiologyVolume 15, No. 2, April 2001
and Beattie, Beattie and Oliver, Oliver and Beattie, and Oliver andBeattie. Conservation Biology 11:571–574.
Gotelli, N. J., and G. R. Graves. 1996. Null models in ecology. Smithso-nian Institution Press, Washington, D.C.
Greenberg, R., P. Bichier, A. C. Angon, and R. Reitsma. 1997. Bird pop-ulations in shade and sun coffee plantations in central Guatemala.Conservation Biology 11:448–459.
Hanski, I. 1998. Metapopulation dynamics. Nature 396:41–49.Hansson, L., and P. Angelstam. 1991. Landscape ecology as a theoreti-
cal basis for nature conservation. Landscape Ecology 5:191–202.Hargrove, W. W., and J. Pickering. 1992. Pseudoreplication: a sine qua
non for regional ecology. Landscape Ecology 6:251–258.Hartstack, A. W., Jr., J. P. Hollingsworth, R. L. Ridgway, and H. H. Hunt.
1971. Determination of trap spacings required to control an insectpopulation. Journal of Economic Entomology 64:1090–1100.
Hobbs, R. J., D. A. Saunders, and G. W. Arnold. 1993. Integrated land-scape ecology: a Western-Australian perspective. Biological Con-servation 64:231–238.
Holloway, J. D., A. H. Kirk-Spriggs, and C. Vun Khen. 1992. The re-sponse of some rain forest insect groups to logging and conversionto plantation. Philosophical Transactions of the Royal Society ofLondon Series B 335:425–436.
Hutto, R. L. 1989. The effect of habitat alteration on migratory landbirds in a west Mexican tropical deciduous forest: a conservationperspective. Conservation Biology 3:138–148.
Janzen, D. H. 1973. Sweep samples of tropical foliage insects: effectsof seasons, vegetation types, elevation, time of day, and insularity.Ecology 54:687–709.
Janzen, D. H. 1987. Insect diversity of a Costa Rican dry forest: whykeep it, and how? Biological Journal of the Linnean Society 30:343–356.
Janzen, D. H. 1988. Ecological characterization of a Costa Rican dryforest caterpillar fauna. Biotropica 20:120–135.
Laurance, W. F., and R. O. Bierregaard, editors 1997. Tropical forestfragments. University of Chicago Press, Chicago.
Levins, R. 1969. Some demographic and genetic consequences of envi-ronmental heterogeneity for biological control. Bulletin of the En-tomological Society of America 15:237–240.
Luff, M. L., and I. P. Woiwod. 1995. Insects as indicators of land-usechange: a European perspective, focusing on moths and groundbeetles. Pages 399–422 in R. Harrington and N. E. Stork, editors. In-sects in a changing environment. Academic Press, London.
MacArthur, R. H., and E. O. Wilson. 1967. The theory of island bioge-ography. Princeton University Press, Princeton, New Jersey.
McGeachie, W. J. 1997. The effects of air temperature, wind vectorsand nocturnal illumination on the behaviour of moths at mercury-vapour light-traps. Ph.D. thesis. Ecological Physics Research Group,Cranfield Institute of Technology, Cranfield, United Kingdom.
Meffe, G. K., C. R. Carroll, and Contributors. 1997. Principles of con-servation biology. 2nd edition. Sinauer Associates, Sunderland,Massachusetts.
Moilanen, A., and I. Hanski. 1998. Metapopulation dynamics: effects ofhabitat quality and landscape structure. Ecology 79:2503–2515.
Nag, A., and P. Nath. 1991. Effect of moon light and lunar periodicityon the light trap catches of cutworm Agrotis ipsilon (Hufn.)moths. Journal of Applied Entomology 111:358–360.
Oliver, I., and A. J. Beattie. 1996. Invertebrate morphospecies as surro-gates for species: a case study. Conservation Biology 10:99–109.
Perfecto, I., and R. Snelling. 1995. Biodiversity and the transformationof a tropical agroecosystem: ants in coffee plantations. EcologicalApplications 5:1084–1097.
Pimentel, D., U. Stachow, D. A. Takacs, H. W. Brubaker, A. R. Dumas,J. J. Meaney, J. A. S. Oneil, D. E. Onsi, and D. B. Corzilius. 1992.Conserving biological diversity in agricultural forestry systems:most biological diversity exists in human-managed ecosystems. Bio-science 42:354–362.
Power, A. G. 1996. Arthropod diversity in forest patches and agroeco-systems of tropical landscapes. Pages 91–110 in J. Schelhas and R.Greenberg, editors. Forest patches in tropical landscapes. IslandPress, Washington, D.C.
Price, P. W., I. R. Diniz, H. C. Morais, and E. S. A. Marques. 1995. Theabundance of insect herbivore species in the tropics: the high localrichness of rare species. Biotropica 27:468–478.
Pulliam, H. R. 1988. Sources, sinks, and population regulation. TheAmerican Naturalist 132:652–661.
Robinson, G. R., R. D. Holt, M. S. Gaines, S. P. Hamburg, M. L. Johnson,H. S. Fitch, and E. A. Martinko. 1992. Diverse and contrasting ef-fects of habitat fragmentation. Science 257:524–526.
Roth, D. S., I. Perfecto, and B. Rathcke. 1994. The effects of manage-ment systems on ground-foraging ant diversity in Costa Rica. Eco-logical Applications 4:423–436.
Saunders, D. A., R. J. Hobbs, and C. R. Margules. 1991. Biological con-sequences of ecosystem fragmentation: a review. Conservation Bi-ology 5:18–32.
Sisk, T. D., N. M. Haddad, and P. R. Ehrlich. 1997. Bird assemblages inpatchy woodlands: modeling the effects of edge and matrix habi-tats. Ecological Applications 7:1170–1180.
Soulé, M. E., A. C. Alberts, and D. T. Bolger. 1992. The effects of habi-tat fragmentation on chaparral plants and vertebrates. Oikos 63:39–47.
Thomas, J. A. 1984. The conservation of butterflies in temperate coun-tries: past efforts and lessons for the future. Pages 333–353 in R. I.Vane-Wright and P. R. Ackery, editors. The biology of butterflies.Academic Press, London.
Tscharntke, T., A. Gathmann, and I. Steffan Dewenter. 1998. Bioindica-tion using trap-nesting bees and wasps and their natural enemies:community structure and interactions. Journal of Applied Ecology35:708–719.
Van Dyke, H., and E. Matthysen. 1999. Habitat fragmentation and in-sect flight: a changing ‘design’ in a changing landscape? Trends inEcology and Evolution 14:172–174.
Vitousek, P. M., H. A. Mooney, J. Lubchenco, and J. M. Melillo. 1997.Human domination of Earth’s ecosystems. Science 277:494–499.
Wiens, J. A. 1995. Habitat fragmentation: island v. landscape perspec-tives on bird conservation. Ibis 137:S97–S104.
Woiwod, I. P., and J. A. Stewart. 1990. Butterflies and moths: migrationin the agricultural environment. Pages 189–202 in R. G. H. Bunceand D. C. Howard, editors. Species dispersal in agricultural habi-tats. Bellhaven Press, London.
Wolda, H. 1978. Fluctuations in abundance of tropical insects. TheAmerican Naturalist 112:1017–1046.
Young, M. 1997. The natural history of moths. T & A Poyser, London.





























