Embed Size (px)
Citation preview

Exp. Eye Res. (1982) 35, 69-75
Compar i son of Po lyunsaturated Fatty Acid Leve l s in N o r m a l and Mature Cataractous H u m a n Lenses*
L o u i s I~OSENFELD AND ABRAHAM SPECTOR
Biochemistry and Molecular Biology Laboratory, Department of Ophthalmology, College of Physicians and Surgeons, Columbia University, New York,
New York 10032, U.S.A.
(Received 2 December 1981, New York)
Polyunsaturated fatty acids (PUFAs) isolated from lipids of age matched senile normal and totally opaque cataractous human lenses were identified and quantitated by thin-layer and gas chromatography. The PUFAs found in human lenses, as confirmed by mass spectrometry, are 18:2, 20:2, 20:3, 20:4, 20:5, 22:3, and 22:4. The total PUFA content found in human lenses (2 % of the total fatty acid population) is low compared to other human tissue. No significant differences were observed in either the PUFA composition or total amount between normal and cataractous lenses, except 20:4 and 20:5 which are decreased in cataracts.
Key words: oxidation ; malondialdehyde ; thin-layer chromatography; gas chromatography; gas chromatography-mass spectroscopy.
1. In troduct ion
The discovery of protein oxidat ion in the h u m a n lens upon ca ta rac t format ion (Dische and Zil, 1950; Garner and Spector, 1980a, b; Spector et al. 1980) has p rompted an invest igation into the oxidat ion of other lens components , par t icular ly lipids. Suggestions of fa t ty acid oxidat ion during cataractogenesis come from the observat ions of extensive membrane polypept ide oxidat ion (Garner and Spector, 1980b), and from a report of a small increase of thiobarbi tur ic acid react ive species in ca ta rac t (Bhuyan, B h u y a n and Podos, 1981), indicating the presence of malondia ldehyde (MDA), an oxidat ion produc t of po lyunsa tura ted f a t ty acids (PUFAs) (Dahle, Hill and Holman,
1962). Lipid peroxidat ion products have been implicated in causing cell damage in
atherosclerosis, in liver injury, in the ageing process, and in other pathological conditions (Bidlack and Tappel, 1973 ; Tappel, 1973). Lipid peroxidat ion occurs more readily in tissues with a high abundance of subcellular organelles rich in P U F A s (Bidlack and Tappel, 1973). I n addit ion to the high concentra t ion of P U F A s in organelle membranes, the presence of peroxide-generat ing systems contr ibutes to lipid oxidat ion (Tappel, 1973). H u m a n lens fiber cells are essentially devoid of subcellular organelles, bu t the lens is exposed to peroxide from the aqueous, and significantly elevated levels of hydrogen peroxide have been detected in the aqueous of certain ca tarac t pat ients (Spector and Garner, 1981). Since low levels of MDA have been detected in the h u m a n lens (Bhuyan et al., 1981) it was of interest to determine the abundance of unsa tu ra ted f a t ty acids in this tissue and whether there is a significant decrease in cataract .
* A preliminary report of this material was presented at the 1981 Association for Research in Vision and Ophthalmology meeting in Sarasota, Florida.
0014-4835/82/070069+07 $02.00/0 �9 1982 Academic Press Inc. (London) Limited 69

70 L. ROSENFELD AND A. SPECTOR
2. Mater ia l s and M e t h o d s
Normal human lenses were obtained from the Eye Bank of New York, usually within 24-36 hr of death. Senile cataract lenses were obtained from the Columbia-Presbyterian Medical Center, usually within 2 hr of extraction. All lenses were stored at -70~ until used. Pairs of normal or mature, totally opaque cataractous human lenses (classification: M, CXA +4, CXP +4, SCA 1~176 N +4, NS pale yellow to yellow by the method of Chylack, 1978) were used. The average age of the donors was 65 or 68 years (S.D.--+9) for normal lenses or cataracts, respectively.
Lenses were directly homogenized in methanol, and enough chloroform was added to make a chloroform-methanol 2 : 1 (v/v) solution. All solvents contained 0"01 ~o BHT as antioxidant. Total lipid extraction of the lenses was performed as previously described (Rosenfeld and Spector, 1981), using the modified Folch procedure according to Cotlier, Obara and Toftness (1978). This procedure has been found to extract the lipid quantitatively from both cataracts and normal lenses (Rosenfeld and Spector, 1981). Attempts to detect residual lipid in the extracted insoluble residue indicate only negligible amounts of material. Furthermore, recoveries of lipid in the normal or cataract populations varied no more than -+ 7 ~o. After evaporation of the solvents with nitrogen, fatty acid methyl esters (FAMEs) were methanolyzed from the residual lipids by treatment with 4 % sulfuric acid in methanol for 5 hr at 80~ (Rosenfeld and Spector, 1981). After extraction with hexane, the solvents were evaporated with nitrogen and the residues were dissolved in chloroform. Methanolysis and hexane extraction yield quantitative recoveries of FAMEs, as determined by treating standards in the same manner.
The FAMEs were separated on the basis of the number of double bonds in their carbon chain by silver ion thin-layer chromatography (TLC) as described by Morris (1964, 1966). Silica gel G plates (20 cm x 20 cm, Redi/Plate, Fisher Scientific) were uniformly sprayed to saturation with an aqueous 10 % silver nitrate solution and reactivated by heating for 30 min at 110~ before use. The concentrated FAME preparation from two lenses was applied as one spot on the TLC plate. A standard PUFA methyl ester mixture (PUFA-2, Supelco) and a blank were run in parallel with each group of samples to check the quanti tat ion of the methods. For the standard mixture, overall reproducibility from run to run was found to be within ___ 2'2 ~o. The relative amounts of standard PUFAs after TLC treatment were found to be within -+1"5~o of the untreated mixture. The plates were developed with hexane--ethyl ether-methanol, 85:15:1'5 (v/v). Spots were visualized under UV light after spraying with an alcoholic solution of 2',7'-dichlorofluorescein (0'2 %) which did not interfere with the assay (Stahl, 1969). One such TLC separation is illustrated in Fig. 1. Regions A, B, C, and D, corresponding to zero or one, two, three and four or more double bonds, respectively, for the standard, were scraped from the plate for each sample. Cholesterol in the samples did not interfere with the subsequent anal.yses. Although fatty acids with two and three unsaturated bonds were not visualized by this procedure, it will be shown that such species were indeed present in these preparations. The silica from each region was transferred to a Pasteur pipet fitted with glass wool, and the material was eluted with at least four volumes of chloroform.
After concentration, the samples were analysed by gas chromatography (GC) as described (Rosenfeld and Spector, 1981). A known amount of the FAME 23:0 was added to all samples as a quantitative internal standard. Aliquots were injected onto a Hewlett-Packard 5880A gas chromatograph with a 6 ft (1"83 m) by 2 mm column of 10 % Silar-10C on 100/120 Gas Chrom Q (Applied Sciences Laboratories). A temperature gradient of l~ from 155 to 210~ was employed, using nitrogen (30 ml/min) as the carrier gas. The fatty acids were quantitated using the internal standard method and were identified on the basis of their retention times compared to standard FAME mixtures RM-3 and PUFA-2 (Supelco). The identities were confirmed by gas chromatographic-mass spectrometric (GC-MS) analysis utilizing a Finnegan Model 9500 GC and Model 3300 MS, using the Finnegan Model 6000 data system, as previously described (Rosenfeld and Spector, 1981), except that the technique of chemical ionization-mass spectrometry (CI-MS) with methane gas was used (Vine, 1980).
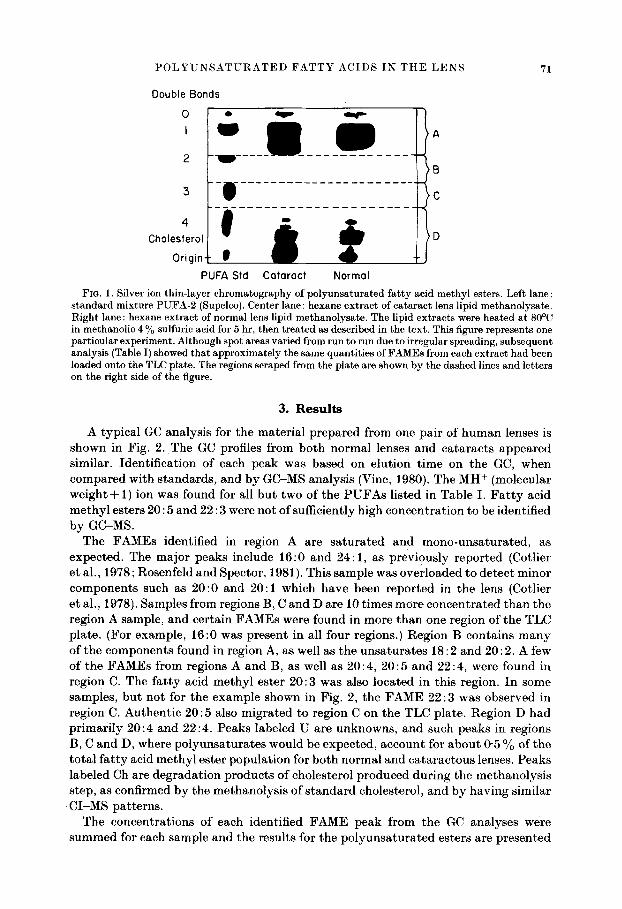
POLYUNSATURATED FATTY ACIDS IN THE LENS 71
Double Bonds
0 o ~ ~ }
- -
2 -"qP" 'B
Cholesterol ~ , D i
Origin-- 0 . J
PUFA Std Cotoroct Normel
FIG. 1. Silver ion thin-layer chromatography of polyunsaturated fatty acid methyl esters. Left lane : standard mixture PUFA-2 (Supelco). Center lane: hexane extract of cataract len s lipid methanolysate. Right lane : hexane extract of normal lens lipid methanolysate. The lipid extracts were heated at 80~ in methanolic 4 % sulfuric acid for 5 hr, then treated as described in the text. This figure represents one particular experiment. Although spot areas varied from run to run due to irregular spreading, subsequent analysis (Table I) showed that approximately the same quantities of FAMEs from each extract had been loaded onto the TLC plate. The regions scraped from the plate are shown by the dashed lines and letters on the right side of the figure.
3. R e s u l t s
A typ ica l GC ana lys i s for the ma te r i a l p r e p a r e d f rom one pa i r of h u m a n lenses is shown in Fig. 2. The GC profiles f rom bo th no rma l lenses a n d ca t a r ac t s a p p e a r e d s imilar . Iden t i f i ca t ion of each peak was based on e lu t ion t ime on the GC, when c o m p a r e d wi th s t anda rds , and b y GC-MS ana lys i s (Vine, 1980). The M H + (molecular w e i g h t + 1) ion was found for all b u t two of the P U F A s l i s ted in Table I. F a t t y acid m e t h y l esters 20:5 and 22 : 3 were no t of suff iciently high concen t r a t i on to be ident i f ied b y GC-MS.
The F A M E s ident i f ied in region A are s a t u r a t e d and m o n o - u n s a t u r a t e d , as expec ted . The ma jo r peaks include 16:0 and 24:1 , as p rev ious ly r epo r t ed (Cotlier e t al. , 1978; Rosenfe ld and Spector , 1981). This sample was ove r loaded to de t ec t minor componen t s such as 20 :0 and 20:1 which have been r epo r t ed in the lens (Cotlier e t al. , 1978). Samples f rom regions B, C and D are 10 t imes more c onc e n t r a t e d t h a n the region A sample, and cer ta in F A M E s were found in more t h a n one region of the TLC pla te . (For example , 16:0 was p resen t in all four regions.) Reg ion B conta ins m a n y of the componen t s found in region A, as well as the u n s a t u r a t e s 18:2 and 20:2. A few of the F A M E s from regions A and B, as well as 20 : 4, 20 : 5 and 22 : 4, were found in region C. The f a t t y acid m e t h y l ester 20 :3 was also loca ted in this region. I n some samples , b u t no t for the example shown in Fig. 2, the F A M E 22:3 was observed in region C. Au then t i c 20 :5 also mig ra t ed to region C on the TLC pla te . Reg ion D had p r i m a r i l y 20 :4 and 22:4 . Peaks labe led U are unknowns , and such peaks in regions B, C and D, where p o l y u n s a t u r a t e s would be expec ted , account for a b o u t 0"5 % of t he t o t a l f a t t y acid m e t h y l es ter popu l a t i on for bo th no rma l and c a t a r a c t o u s lenses. Peaks labe led Ch are d e g r a d a t i o n p roduc t s of cholesterol p roduced dur ing the me tha no ly s i s s tep, as confirmed b y the me thano lys i s of s t a n d a r d cholesterol , and b y hav ing s imi lar C I - M S pa t t e rns .
The concen t ra t ions of each ident i f ied F A M E peak from the GC ana lyses were s u m m e d for each sample and the resul ts for the p o l y u n s a t u r a t e d esters are p resen ted

72 L. R O S E N F E L D A N D A. S P E C T O R
W 0') Z 0
(D b..I r r
23:0 I
B
20:4 Ch ~' 2~5 22:4 ~'
' ~ ' ~ C
22:4
0 5 0 60 R E T E N T I O N T I M E ( m i n u t e s )
FIG. 2. Gas chromatographic profiles of the mater ia l scraped and eluted from regions A, B, C and D from one lane of a TLC plate similar to the one i l lustrated in Fig. 1. A Hewle t t -Packa rd 5880A Gas Chromatograph with a 6 ft (I'83 m ) x 2 m m column of l0 % Silar-10C on 100/120 Gas Chrom Q was employed, using a t empera tu re gradient of l ~ from 155 to 210~ Corrections were made for solvent contaminants .
POLYUNSATURATED FATTY ACIDS IN THE LENS
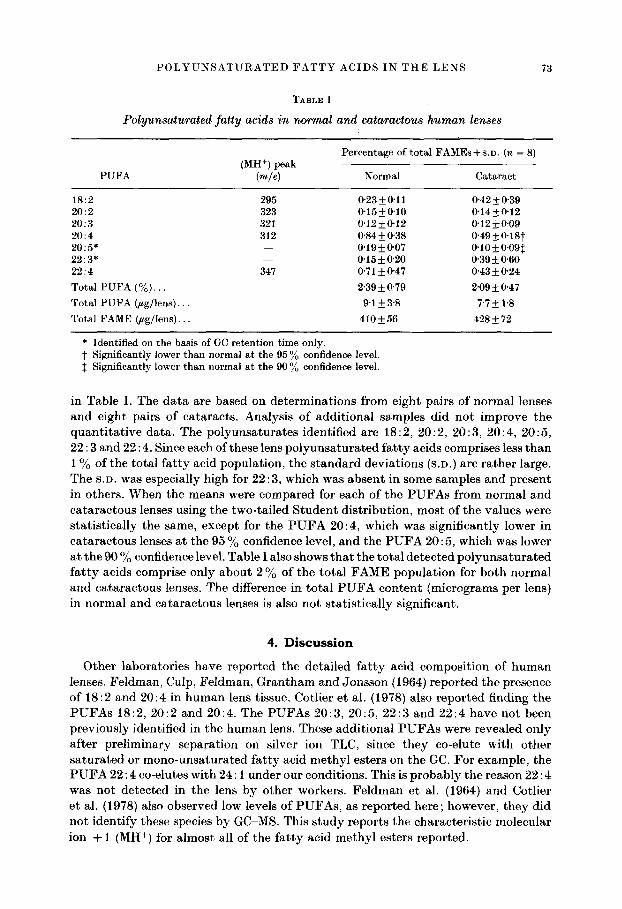
TABLE I
Polyunsaturated fatty acids in normal and cataraetous human lenses
73
PUFA
Percentage of total FAMEs + S.D. (n = 8) (MH +) peak
(re~e) Normal Cataract
18:2 20:2 20:3 20:4 20:5" 22:3* 22:4 Total PUFA (%)... Total PUFA (~g/lens)... Total FAME (gg/lens)...
295 323 321 312
347
0"23_+0'11 0"42_+0'39 0-15_+0'10 0-14__.0"12 0'12_+0-12 0"12• 0"84-+0'38 0-49-+ 0.18t 0'19-4-0"07 0'10• 0'15+0"20 0'39___0'60 0"71 • 0"47 0"43 • 0"24 2'39_+0'79 2'09_+0'47 9-1 -+ 3'8 7"7 -+ 1"8
410• 428•
* Identified on the basis of GC retention time only. t Significantly lower than normal at the 95 ~o confidence level. :~ Significantly lower than normal at the 90 ~o confidence level.
in Table I. The da t a are based on determinat ions from eight pairs of normal lenses and eight pairs of cataracts . Analysis of addit ional samples did not improve the quant i ta t ive data. The polyunsa tura tes identified are 18:2, 20:2, 20 : 3, 20:4, 20:5, 22 : 3 and 22:4. Since each of these lens po lyunsa tu ra ted f a t ty acids comprises less than 1% of the total f a t ty acid populat ion, the s tandard deviat ions (S.D.) are ra ther large. The S.D. was especially high for 22 : 3, which was absent in some samples and present in others. When the means were compared for each of the P U F A s from normal and ca taraetous lenses using the two-tailed S tuden t distr ibution, most of the values were stat ist ically the same, except for the P U F A 20:4, which was significantly lower in ca tarac tous lenses at the 95 % confidence level, and the P U F A 20 : 5, which was lower at the 90 % confidence level. Table I also shows tha t the tota l detected po lyunsa tu ra ted f a t t y acids comprise only about 2 % of the total F A M E popula t ion for both normal and ca taractous lenses. The difference in tota l P U F A conten t (micrograms per lens) in normal and ca tarac tous lenses is also not stat ist ically significant.
4. D i s c u s s i o n
Other laboratories have reported the detailed f a t ty acid composi t ion of h u m a n lenses. Feldman, Culp, Feldman, Gran tham and Jonsson (1964) reported the presence of 18:2 and 20:4 in h u m a n lens tissue. Cotlier et al. (1978} also reported finding the P U F A s 18:2, 20:2 and 20:4. The P U F A s 20:3, 20:5, 22 :3 and 22:4 have not been previously identified in the h u m a n lens. These addit ional P U F A s were revealed only after prel iminary separat ion on silver ion TLC, since they co-elute with other sa tura ted or mono-unsa tura ted fa t ty acid methyl esters on the GC. For example, the P U F A 22: 4 eo-elutes with 24:1 under our conditions. This is p robab ly the reason 22: 4 was not detected in the lens by other workers. Fe ldman et al. (1964) and Cotlier et al. (1978} also observed low levels of PUFAs , as repor ted here; however, they did not identify these species by GC-MS. This s tudy reports the characterist ic molecular ion + 1 (MH +) for a lmost all of the fa t ty acid methy l esters reported.

74 L. ROSENFELD AND A. SPECTOR
Assuming 1 mol MDA (malondialdehyde) is formed per mole of polyunsaturated fa t ty acid, and assuming an average chain length of 20 carbons, the amount of MDA reported in the literature (Bhuyan et al., 1981) for cataractous lenses would represent the oxidation of approximately 3 ~o of the PUFA population. A greater loss of lens PUFA would be expected, since such MDA values do not accurately indicate the actual quant i ty of MDA produced in vivo. The MDA may be involved in cross-linking, or it could be degraded by aldehyde dehydrogenase, which has been found in the human lens (Jedziniak and Rokita, 1981). Also MDA represents only one of several possible oxidation products (Dahle et al., 1962). Nevertheless, the MDA level reported does not represent the loss of a large fraction of the fa t ty acid pool, al though it may have impor tant biological significance. As a comparison, over 50 % of sulfhydryl groups and 20 ~o ofmethionine residues have been found to be oxidized in cataractous lens proteins (Garner and Spector, 1980a, b). This does not mean tha t protein oxidation is the only significant event in cataractogenesis. A small amount of oxidized lipid intermediate, such as lipid peroxide, may be necessary to initiate oxidation of proteins (Dahl et al., 1962}; however, the unusually low concentration of polyunsatura ted fa t ty acids in the human lens makes this an unlikely possibility.
The present study has shown tha t there is no significant difference i n the overall PUFA levels in normal and cataractous lenses. Quanti ta t ive values presented in Table I were difficult to obtain, owing to the small amount of PUFA present in the lens and to the limitations of the technique. Large s tandard deviations were obtained for many of the values. However, the basic conclusions are unaltered : the human lens has very little PUFA, compared to other tissue (Kuksis, 1978), and there is no apparent difference in total P U F A content when comparing normal and cataractous lenses. The total PUFA level in the human lens was found to be about 2 % of the total FAME content. The PUFA level for other human tissues has been reported to range from 2"5 to 67 % of the total FAME population (Kuksis, 1978). The unusually low PUFA content, as well as the high cholesterol content (Broekhuyse, 1973 ; Rosenfeld and Spector, 1981), in the human lens may contribute to the remarkable rigidity of the fiber cell membrane (Li, Spector, Cogan and Schacter, 1981}.
A C K N O W L E D G M E N T S
The authors thank Mr Vincent Saltemach for operating the GC-MS. Louis Rosenfeld was supported by National Eye Institute Training Program Grant No. EY07006-06.
R E F E R E N C E S
Bhuyan, K.C., Bhuyan, D.K. and Podos, S .M. (1981). Evidence of increased lipid peroxidation in cataracts. IRCS Med. Sci. 9, 126-7.
Bidlack, W.R. and Tappel, A.L. (1973). Damage to microsomal membrane by lipid peroxidation. Lipids 8, 177-82.
Broekhuyse, R. M., (1973). Membrane lipids and proteins in ageing lens and cataract. Ciba Found. Symp. 19, 135-49.
Chylack, L. T. (1978). Classification of human cataracts. Arch. Ophthalmol. 90, 888-92. Cotlier, E., Obara, Y. and Toftness, B. (1978). Cholesterol and phospholipids in protein
fractions of human lens and senile cataract. Biochim. Biophys. Acta 530, 267-78. Dahle, L. K., Hill, E. G. and Holman, R. T. (1962). The thiobarbituric acid reaction and the
autoxidations of polyunsaturated fatty acid methyl esters. Arch. Biochem. Biophys. 98, 253-61.
Dische, Z., and Zil, H. (1950). Studies on the oxidation of cysteine to cystine in lens proteins during cataract formation. Proc. A. Res. Ophthalmol. 19th Meeting, p. 105.

POLYUNSATURATED FATTY ACIDS IN THE LENS 75
Feldman, G. L., Culp, T. W., Feldman, L. S., Grantham, C. K. and Jonsson, H. T. (1964). Phospholipids of the bovine, rabbit, and human lens. Invest. Ophthalmol. 3, 194-7.
Garner, M. H. and Spector, A. (1980a). Selective oxidation of cysteine and methionine in normal and senile cataractous lenses. Proc. Natl. Acad. Asci. U.S.A. 77, 1274-7.
Garner, M. H. and Spector, A. (1980b). Sulfur oxidation in selected human cortical cataracts and nuclear cataracts. Exp. Eye Res. 31,361-9.
Jedziniak, J. and Rokita, J. (1981). Aldehyde metabolism in the human lens. Invest. Ophthalmol. 20, Suppl., 90.
Kuksis, A. (1978), Fat ty acid composition of glycerolipids of animal tissues. In Handbook of Lipid Research. Fatty Acids and Glycerides. Vol. 1, pp. 381-442. Plenum Press, New York.
Li, L. K., Speetor, A., Cogan, U. and Schachter, D. (1981). Lipid dynamics and lipid-protein interaction in calf lens fiber membranes. Invest. Ophthalmol. 20, Suppl., 36.
Morris, L. J. (1964). Specific separations by chromatography on impregnated thin layers. Lab. Pract. 13, 284-98.
Morris, L. J. (1966). Separations oflipids by silver ion chromatography. J. Lipid Res. 7, 717-32. Rosenfeld, L. and Speetor, A. (1981). Changes in lipid distribution in the: human lens with
the development of cataract. Exp. Eye Res. 33, 641-50. Speetor, A., Garner, M. H., Roy, D., Garner, W. H., Farnsworth, P. N. and Shyne, S. (1980).
Oxidation of lens proteins. In Red Blood Cell L~ns Metabolism (Ed. Srivastava, S. K.). Vol. 9, pp. 81-93. Elsevier/North Holland, Amsterdam.
Spector, A. and Garner, W. H. (1981). Hydrogen peroxide and human cataract. Exp. Eye Res. 33, 673-81.
Stahl, E. (1969}. Thin Layer Chromatoqraphy : A Laboratory Handbook. Springer Verlag, Berlin. See pp. 866-7.
Tappel, A. L. (1973). Lipid peroxidation damage to cell components. Fed. Proc., 32, 1870-4. Vine, J. (1980). Analysis of fatty acid methyl esters by high-resolution gas chromatography-
chemical ionisation mass spectrometry. J. Chromatogr. 196, 415-24.





























