Embed Size (px)
Citation preview

Relation to the book themes?
System development:
Basic concepts – no specific links
Scale:
Clarification of some scale issues:
differences among species, habitat
structure
Chapter 1
Understanding species and habitat
Major points to remember
Species present in an
ecological system
(community or
ecosystem) are usually
local populations that
may differ from other
populations of the same
taxonomic species living
somewhere else.
Species membership in an ecological system ranges from almost permanent to most
accidental. Their roles in and effects on the system may correlate with the nature of
membership but they also may be disproportionately different.
Habitat is difficult to define as it changes with the perception of species occupying
it. Its relative nature is best managed by invoking a hierarchy of structure and
features.
Habitat structure and attributes of its component patches change over time as a
result of various factors.

We made some comments about the habitat in the introduction. One was that the habitat
and the niche (species characteristics) will be used in different ways and that they do
complement but do not overlap. In this chapter we will continue developing a perspective
on what habitat is and also we will attempt to get a more refined view on what a species in
a community really is, if there is a simple answer.
Species
As populations
Communities form from interacting networks of species. But what do we mean by species?
The membership of a community is not just a list of species. A community is likely
composed of species fragments or local populations. In fact, most species observed in a
community live also somewhere else. It is a rare situation that a whole species lives within
one community unless we consider a very large community. Now, you can see how the
change of extent (Introduction) affects the interpretation of community membership. This
will become clearer soon, especially when you review Figure 1.1. Let’s consider various
possibilities.
A community may comprise species entirely living in it, present because they are
present in the surrounding habitats, present only in portions of the community habitat, or
present in far away habitats that are also suitable but not connected to the habitat of interest
at the ecological time scale (although they may have been connected historically, Fig. 1.1).
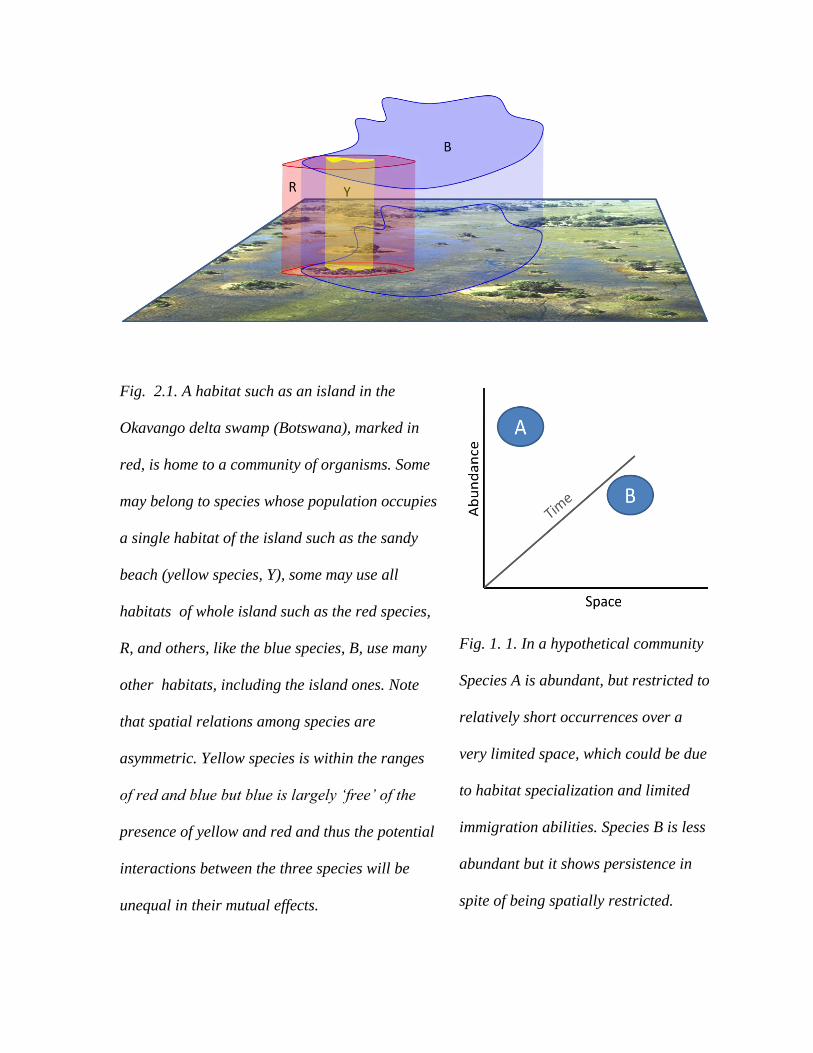
Fig. 2.1. A habitat such as an island in the
Okavango delta swamp (Botswana), marked in
red, is home to a community of organisms. Some
may belong to species whose population occupies
a single habitat of the island such as the sandy
beach (yellow species, Y), some may use all
habitats of whole island such as the red species,
R, and others, like the blue species, B, use many
other habitats, including the island ones. Note
that spatial relations among species are
asymmetric. Yellow species is within the ranges
of red and blue but blue is largely ‘free’ of the
presence of yellow and red and thus the potential
interactions between the three species will be
unequal in their mutual effects.
Fig. 1. 1. In a hypothetical community
Species A is abundant, but restricted to
relatively short occurrences over a
very limited space, which could be due
to habitat specialization and limited
immigration abilities. Species B is less
abundant but it shows persistence in
spite of being spatially restricted.

Thus, a local community is often a mixture of species to which the
local habitat is very important, species to which this habitat is optional or
occasional, and species that have any kind of intermediate relationship with
it. This applies to time dimension, too. Some species persist in a habitat
almost permanently, while others are mere transients. Also, some species are
represented by many while others by few individuals. The phase space
describing all the combinations along these dimensions can be filled in a variety of ways.
Figure 1.2 illustrates a couple of combinations as examples. Indeed, any single community
or ecosystem will have only some of this space filled and the way it is filled may depend on
history, kind of physical environment, and the scale of habitat of interest (particularly, its
extent).
Furthermore, the species observed in a community may show varying degrees of
integration with their own broader populations. How distinct locally they are depends on
whether individuals migrate into community or are born in it. The latter depends on habitat
isolation relative to species dispersal; poorly dispersed species are likely to rely more on
local reproduction than on immigration of individuals from outside. The
level of integration a local population has with the remaining populations of
the species matters because it determines to what degree the local population
depends on what happens in the local community. The less connected it is to
the outside, the more likely its interactions with other species in the local
community will determine its growth or demise. Those species whose local individuals are
most linked to the outside populations are transients: Individuals of some species, whether
How a local
population is
integrated with
‘sister’ populations
outside affects its
responses to other
community
members
‘Species’ in a
community or
ecosystem are
usually small local
populations of
species that have
much broader
geographical and
ecological
distribution
plant or animals, may show up only occasionally because they either pass by and interact
opportunistically or because their seeds arrived by chance, germinated and grew into a
plant specimen that is unlikely to continue to reproduce within this community. Transient
species may have a significant effect on
the local community but they do not
depend on it – their dynamics depends on
many other factors well outside of the
local community. For example, the
Serengeti population of wildebeest does
not depend on the local grassland, even if
one considers large patches of, say, 100
square kilometers. Rather, they follow
the pattern of rains and grass growth over
hundreds of kilometers and can easily
move on if the local habitat is inadequate
due to poor grass growth, grass lost to
other grazers, or due to concentration of
resident predators.
This example brings another
point worth considering. Broadly
ranging species may have only a brief
interest in the local community. At the
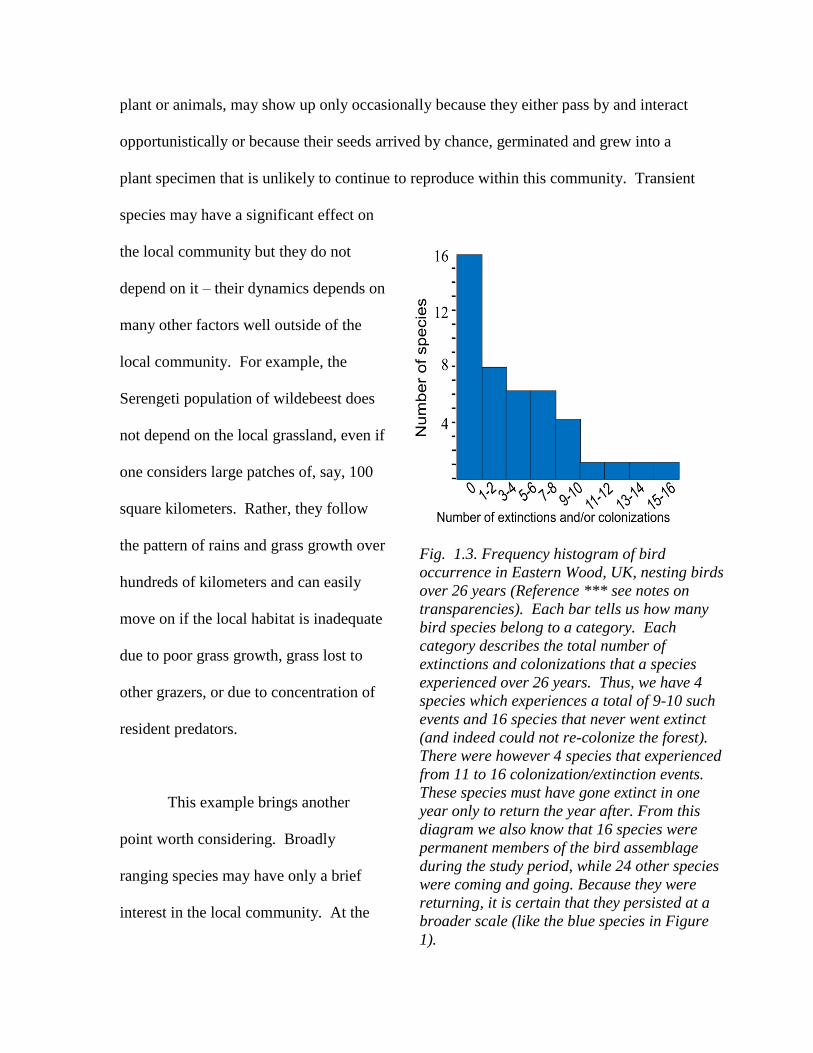
Fig. 1.3. Frequency histogram of bird
occurrence in Eastern Wood, UK, nesting birds
over 26 years (Reference *** see notes on
transparencies). Each bar tells us how many
bird species belong to a category. Each
category describes the total number of
extinctions and colonizations that a species
experienced over 26 years. Thus, we have 4
species which experiences a total of 9-10 such
events and 16 species that never went extinct
(and indeed could not re-colonize the forest).
There were however 4 species that experienced
from 11 to 16 colonization/extinction events.
These species must have gone extinct in one
year only to return the year after. From this
diagram we also know that 16 species were
permanent members of the bird assemblage
during the study period, while 24 other species
were coming and going. Because they were
returning, it is certain that they persisted at a
broader scale (like the blue species in Figure
1).

same time they are members of a much larger ecological system and thus their dynamics is
best understood at the scale of a larger system. A study of birds in a small forest patch in
Great Britain reveals some composition complexity typical of practically every ecological
system, whether small or large (Fig. 1.3).
Thus, we may think that each community is made up of species whose membership
ranges from obligatory (always present in the area delineating the community) to optional.
Species participate in community processes unequally with respect to their numbers, time
of ‘involvement’, and activities they engage in. The balance between the various classes of
members may change depending on the habitat/community type, its stage of development
(succession), and external influences such as pollution, destruction of surrounding habitat,
and many others.
Why is it important to remember that an ecological system is made up of
populations while we use species names to identify their general characteristics? An
example helps make the point. Troost et al. (2005) investigated adaptation of protists
living in under a gradient of light. In a homogeneous environment these
theoretical organisms are generalists because they use mixotrophy: they use
both photosynthesis and consumption of organic materials to satisfy their
energetic and nutritional needs. However, when presented with a gradient of
light availability, they specialize. Populations living where light is scarce
give up photosynthesis and switch to heterotrophy alone. Populations living in well lit
conditions specialize in autotrophic feeding via photosynthesis. The implication for
Populations of the
same species may
have widely
different
characteristics
depending on the
host habitat

community ecologists is that the same taxonomic species may in fact act as two or three
distinctly different ecological entities depending on the characteristics of habitat. What is
more, introducing spatial heterogeneity also makes evolution of these populations sensitive
to other environmental conditions, such as total nitrogen content or light intensity. As such,
it provides an explanation of why mixotrophs are often more dominant in nutrient-poor
systems while specialist strategies are associated with nutrient-rich systems (Troost et al.
2005).
This example and preceding considerations tell us that using the species name alone
does not give us a good sense of what traits this species has in ecological terms. More
information is required and remembering that actually it is the population that is a member
of community helps to keep this in better perspective.
Species traits
Not only species differ in when and for how long they are members of an ecological system
but they also do so in virtually any other characteristic. If the list of species is sufficiently
long, we will find clear gradients of other features. Species that differ in the number of
offspring they produce may form a reproductive strategy gradient (recall r- and K-
strategists). Species that differ in kinds of food, shelter, or microhabitat needed may form a
gradient of resource utilization. Autotrophic species that differ in biomass or rate of
growth may form a gradient of contributions to the energy budget of the local ecosystem.
Species that employ chemical defenses may form still another gradient of traits (by
chemical effectiveness or concentration). Or species restricted by various adverse or
unsuitable features of the habitat may form a gradient of habitat specialization. The
universality of this gradient is well illustrated by Dyer et al. (2007) who studied host
specificity of butterflies. Host specificity measures one of the dimensions that one could
include in species niche. They found that in every of eight locations they investigated from
Canada to Ecuador there were species that used five or less plant host genera as well as
some species that used more plant genera. Thus, they found that each butterfly assemblage
is a collection of species of different levels of specialization. In addition they found that
butterflies (their caterpillars) are progressively more specialized in the tropics.
Yet, while the above picture indicates some fluidity to which species make up a
community, some constraints apply. The community membership may appear haphazard
but local habitat attributes impose soft rules on which species can participate in communal
living and how they do so.
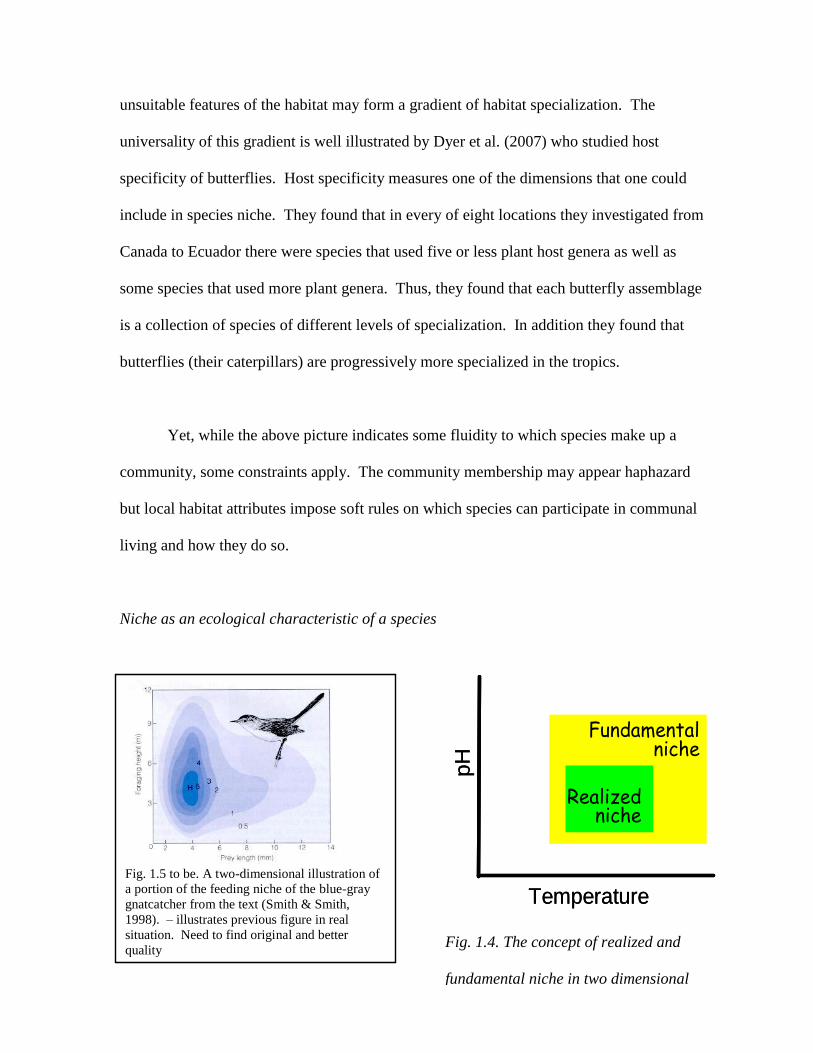
Niche as an ecological characteristic of a species
Fig. 1.4. The concept of realized and
fundamental niche in two dimensional
space.
Fig. 1.5 to be. A two-dimensional illustration of
a portion of the feeding niche of the blue-gray
gnatcatcher from the text (Smith & Smith,
1998). – illustrates previous figure in real
situation. Need to find original and better
quality
Temperature
pH
Temperature
pH
Fundamental niche
Realized niche

So far we recognized that species differ
in their traits how this is important to
understanding makeup of communities. These
differences and some of their consequences
have been examined by ecologists by
employing a concept of ecological niche.
Several approaches exist to characterization
and quantification of species traits but among
those the niche breadth is the most common
and well developed. Other concepts, related
but also slightly different in their emphasis or
scope, are tolerance and ecological range.
Niche does not have a good definition but it
can be loosely summarized as a set of biotic
and abiotic dimensions that describes
conditions a species can live under (or lives
under). Because it describes a species, niche is
analogous to some extent to morphology. This
view of the niche is due to E.G. Hutchinson
who first defined the niche as a species
characteristic. Usually, niche is portrayed using
one, two, or three dimensions, where values
suitable for a species become sides of a box.
Fig. 1.6. Species 1 can exist in a narrow range
of pH but large range of temperatures (a).
Species 2 can tolerate fairly broad ranges of
both pH and temperature. The two species
overlap over a small range of temperature
values. The two species can overlap in both or
many dimensions (b). If we added a third
dimension such as the size of food items each
species can consume, we could calculate the
volume of each box. This led ecologists (E.G.
Hutchinson) to talk of niche as a
multidimensional volume.
Temperature
pH
Temperature
pH
Temperature
pH
Temperature
pH
Niche #2
Niche #1
Indeed, many dimensions are required for a more complete description of a species’ niche.
In two dimensions the box becomes a picture of the species’ niche.
Is mentioned earlier, ecologists used the niche concept to ask important questions
about species abundance and distribution, diversity-productivity relationship, community
stability, ecosystem functioning , and biological invasions . Current views highlight the
importance of the niche concept in ecology, and understanding different properties of
niches (e.g. use of environmental space, resource use) is also a key for understanding
community organisation (Chase and
Leibold, 2003).
Schematic on the right (Fig.
1.5) uses two variables only and
depicts two versions of a
hypothetical species’ niche. The
yellow areas describe the
combinations of temperature and
moisture that the species can live in
and reproduce. This resource space is
known as the fundamental niche.
By contrast, the green area describes
the actual combinations of these two
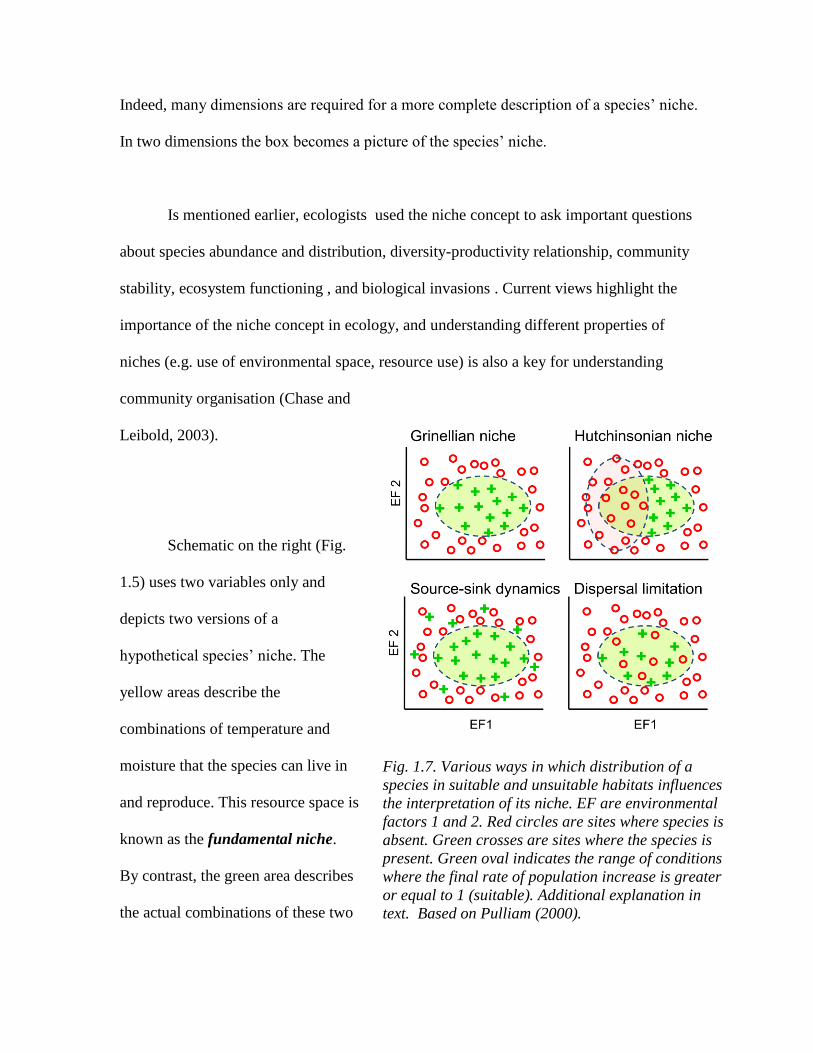
Fig. 1.7. Various ways in which distribution of a
species in suitable and unsuitable habitats influences
the interpretation of its niche. EF are environmental
factors 1 and 2. Red circles are sites where species is
absent. Green crosses are sites where the species is
present. Green oval indicates the range of conditions
where the final rate of population increase is greater
or equal to 1 (suitable). Additional explanation in
text. Based on Pulliam (2000).

variables that the species utilizes in its habitat. This subset of the fundamental niche is
known as the realized niche. Fundamental niche represents species potential or capability
while the realized niche is often seen as being reduced by biotic factors such as competition
or predation. For example, leopards can and often do hunt during both the day and night.
However, in areas of high lion density, a species that strongly competes with leopards and
kills them whenever possible, leopards limit their hunting to the night. Thus their temporal
dimension for hunting is much smaller than their behavioral abilities permit. Furthermore,
a species may easily disperse to habitats that are suboptimal and that would not maintain
the population through the natural growth. Such habitats may appear as suitable if one
relied on the distribution of species alone. Some of the possible relations between species
niche, its distribution, and its habitat were reviewed by Pulliam (2000) and appear in Figure
1.7. Grinellian niche views species as being present in suitable habitat (final rate of
population increase equal or greater than 1). Hutchinsonian niche implies exclusion of
species from some sites due to biotic interactions – actual distribution is thus less than the
potential one. Source-sink dynamics permits species existence outside suitable patches
(final rate of increase less than 1). Dispersal limitation emphasizes the possibility that
species is absent from suitable sites because it is unable to disperse to them after they
formed or after species disappeared from them. The different processes captured by each
cartoon together underscore the difficulty of deducing niche properties from distribution
and habitat properties. They further underscore the need for clear separation of the three
concepts.

Using the box convention to depicting niches permits a quick analysis whether the
two species would compete or enter in any other interaction. Consider two niches that may
overlap (Fig. 1.6a). Here the overlap is over the section of temperature values but no
overlap exists along pH values. Thus, the two species are unlikely to compete because
different pH requirements prevent
them from occurring together. When
niches overlap such that the two boxes
intersect to some degree (Fig.1.6b),
competition, predation, or symbiotic
relationships may take place. The
more they overlap, the stronger the
competition among species if the
overlap is over a resource both use.
Two species may overlap in different
ways. The various permutations (Fig.
1.8) suggest different ways species are
likely to interact. For example, under
the coextensive scenario, it is likely
that one species will be excluded
because it has no range of
environmental conditions where it an
avoid competition. The reciprocal
scenario is likely to lead to a
Fig. 1.8. Niches can overlap (or not) in
different ways. Here a niche of a species is
reduced to one dimension only, that of a
resource gradient. Frequency of occurrence
indicates how often (or how many individuals)
a species is present at a particular value of
resource (e.g., size of food particles). Each
curve represents a niche of one species.
Adopted from
http://www.montana.edu/~wwwbi/staff/creel/bi
o405/405lec19.pdf
Overla
ps
Non-o
ve
rlaps
Fre
quency o
f occurr
ence
Resource gradient
Included
Reciprocal
Coextensive
Asymmetric
Abutting Disjunct
substantial reduction of realized niches but both species should be able to coexist by
selecting patches of habitat where they do not have to compete directly with the other
species. The asymmetric scenario gives more habitat to one of the species but the other can
still find a decent range of condition where to retreat. The case of lions and leopards would
fit here nicely. The non-overlapping situations are also of some interest because,
depending on whether the species abut or are disjunct (distinctly separated) on the resource
gradient, their potential for interaction is large or small if conditions slightly change. You
can try to think what outcomes would be likely under the remaining scenarios in Figure 1.8.
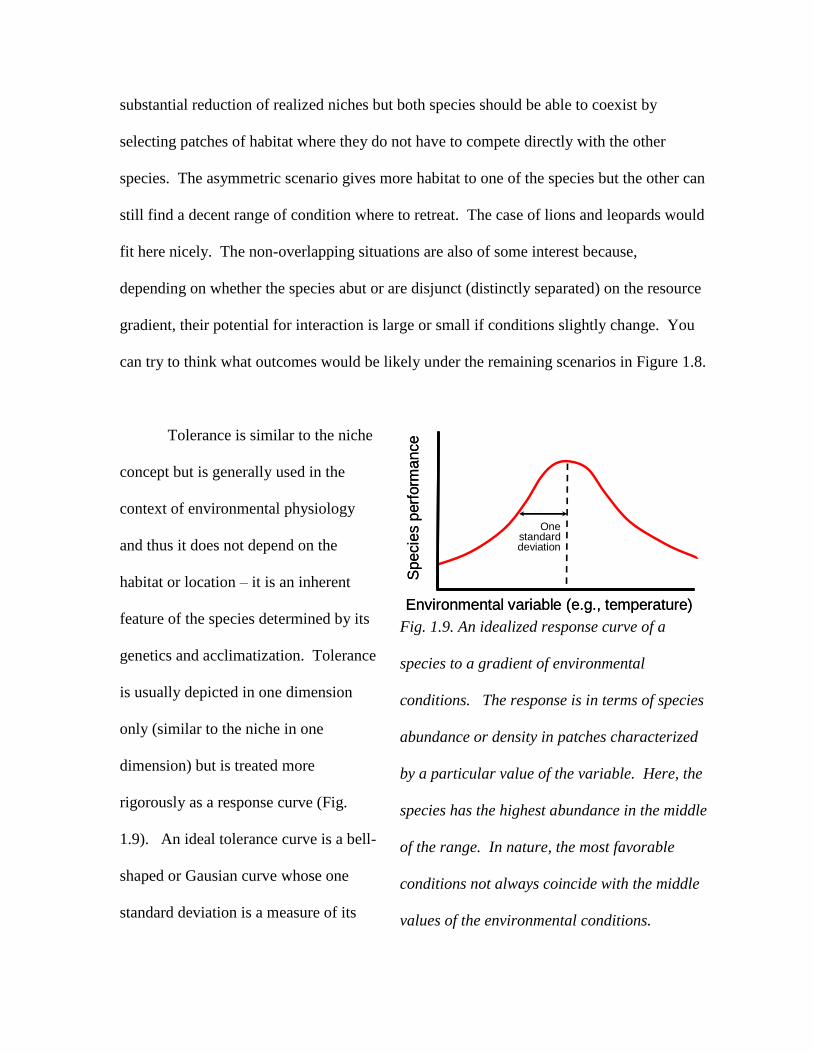
Tolerance is similar to the niche
concept but is generally used in the
context of environmental physiology
and thus it does not depend on the
habitat or location – it is an inherent
feature of the species determined by its
genetics and acclimatization. Tolerance
is usually depicted in one dimension
only (similar to the niche in one
dimension) but is treated more
rigorously as a response curve (Fig.
1.9). An ideal tolerance curve is a bell-
shaped or Gausian curve whose one
standard deviation is a measure of its
Fig. 1.9. An idealized response curve of a
species to a gradient of environmental
conditions. The response is in terms of species
abundance or density in patches characterized
by a particular value of the variable. Here, the
species has the highest abundance in the middle
of the range. In nature, the most favorable
conditions not always coincide with the middle
values of the environmental conditions.
Environmental variable (e.g., temperature)
Specie
s p
erf
orm
ance
Environmental variable (e.g., temperature)
Specie
s p
erf
orm
ance
One standard deviation

spread. The concept of the niche discussed above has been inspired by the tolerance curve
and it indistinguishable from it when a single variable/dimension is considered only. When
it comes to variables that do not affect physiology, the curve has a much lesser probability
of having bell shape. A response curve to predator is more likely to be monotonically
declining – it is not logical to expect that the species will improve its performance when its
predator density increases.
Ecological range is a related concept to that of the realized niche but it has some
additional flexibility. Which of the descriptors of species’ ability is used depends on the
question. For example, in a system of rock pools with differing environmental conditions
species can occur in some pools but cannot occur in others. However, species are often
absent from some of the pools whose conditions they can tolerate. The reason for such
absences is not therefore their tolerance (or niche parameters) but something else. A
species may fail to be present by chance when dispersing individuals simply missed the
opportunity; it may be absent due to history if it was pushed to extinction by past
perturbations and failed to return; and it can be absent because it is systematically or
occasionally reduced to zero numbers by competition, predation, or diseases. Neither
chance events nor history can be part of the niche volume because they would make a niche
volume not a characteristic of a species but a random value. However, the ecological range
is an effective measure of a species’ performance in a specified landscape – landscape of
rock pools in our example. Ecological range of a species may be different in a different
landscape. However, the effective (actual) ecological range in any specified landscape tell

us a lot about the species success and its ability to meet successfully other
species occupying the same space.
Habitat
What do we mean by habitat?
Habitat is rather difficult to define. A useful approach would be to ask what is habitat from
the user (a local population of a species) point of view. When you think of any two
species, even closely related species that occupy the same area, you will realize that they
use that area differently. Wolves and coyotes may share Yellowstone National Park but
hunt different prey, and coyotes keep their distance from wolves while wolves do no limit
their movement and hunting on account of coyotes. In Algonquin Provincial Park in
Ontario moose and white-tail deer live in the same area but use open spaces, lake shores
and swamps differently as well as show different levels of vulnerability to predators and
harsh winters. Thus, wolves and coyotes or moose and white-tail deer experience their
habitat differently. For all ecological purposes they live in somewhat different habitats
even though they occupy the same ecological system. Such differences in how coexisting
species see their habitat depend on the evolutionary distance between them as well as on
another, very important aspect of habitat structure (see further).
Ecological range
and the niche,
realized niche in
particular, are
related but not the
same

Note also that the niche determines what habitat a species perceives. However,
even though the niche and habitat of a species may be described using the same
dimensions, their meaning is different.
Niche is a suit of species attributes,
which is relatively constant. Habitat may
have many other attributes or values
along a particular axis that species does
not. Species attributes, or the niche, can
be seen as an interpreter of habitat
attributes.
One can illustrate differences in
habitat perceptions by species by
graphing relevant dimensions and giving
them numerical values to reflect their importance for each species (Fig. 1.10). Indeed, the
axes selected for the figure are only to give the sense of diversity of factors involved. Their
full enumeration and explanation how they can be made quantitative lies outside this
chapter scope. Note that the polygons produced for each species describe the relationship
between the species attributes and the attributes of local environment. Thus, they are not
strictly habitat attributes or niches of the two species (see Introduction for the term
definitions). Nevertheless, such descriptions may be very useful in thinking about how
each species is likely to perform in the local habitat. An example from the Amazon area
demonstrates the difference between the two concepts quite well.
Figure 1.10. Habitat view from perspective of
two different species. Each axis describes a
factor or group of factors that affect
significantly species performance. For
instance, blue species is under strong
pressure from competitors while the yellow
species in not. Green area shows overlap of
the two species.

Fine at al. (2004) wondered why they
observed distinct plant communities on two
different types of soil, sand and clay. They
transplanted seedlings of 20 species from six
genera of phylogenetically unrelated pairs of
edaphic (=soil) specialist trees. Trees growing
normally on clay soils were transplanted into
sandy soils and vice versa. They also
manipulated the presence of herbivores. They
found that clay specialist species grew
significantly faster than white-sand specialists
in both soil types when protected from
herbivores. However, when unprotected, white-sand specialists dominated in white-sand
forests and clay specialists dominated in clay forests. Fine and colleagues concluded that
habitat specialization in that system results from an interaction of herbivore pressure with
soil type. What is of interest to us here is that species abilities resulting from their
evolutionarily acquired traits (the niche) were obscured by contingent attributes of the
environment – not even one factor but an combination of two environmental axes (soil and
herbivores).
Box. 1.1. To consider… Where is the Figure 1.10 coming from? First, recall that the niche is an ecological characterization of species. It is usually thought of in terms of a variety of dimensions and species performance along those dimensions (Introduction). These dimensions may be exactly as those shown in Fig. 1.9 but may also include others that do not apply in the habitat we consider. For example, a species may have anti-predator defenses but when predators are absent, a comparison would make no sense. Thus, a value on an axis is produced by relating species traits to the state of the environment. If both species need shelter equally but the shelter is too small for the yellow, it will have a lower value on its axis.

Habitat structure
The differences in how species perceive
habitat and how they respond to it have
major systematic consequences. Species
occupying a local habitat differ not only in
response to specific habitat properties but
also in resolution of habitat grain (see
Introduction). However, the habitat grain
differences are not simply due to the fact
that some habitat components are large
while others are small. Rather they are due
to how those components are arranged
with respect to each other. But first …
Consider again the Okavango Delta
swamp. The picture (Fig. 1.11) shows a
mosaic of habitats: treed areas, sands, patches of open water, floating vegetation, animal
paths, and so on. What we see in the picture is also habitat but not in the way we tried to
define it earlier. Let us thing of this kind of habitat as a landscape feature – something that
is not very precise but good enough as a starting point for further considerations.
Organisms related to this habitat in different ways. Fish eagles use most of the habitat.
They perch on trees but hunt in the open water. Buffalo, lechwe antelopes, hippos, and
elephants move easily through as well as graze or rest on the land, open water, and shallow
Fig. 1.11. Any habitat, like this Okavango
Delta swamp, can be seen at several levels of
resolution. At the lowest resolution, one can
see it as a habitat type, or as a mixed habitat of
water and land. At some finer scale (smaller
grain) one can distinguish minor habitats
composing aquatic and terrestrial habitats
(e.g., trees, grasses, reeds). Further increases
in resolution would reveal even smaller habitat
types.
reed and floating vegetation patches. However, monkeys are restricted to
dry areas while tiger fish is at home in open or quasi open water. From the
perspective of these organisms, habitat is actually a collection of more or
less suitable patches. Some organisms distinguish smaller, better defined kinds of habitat
than others. Thus, organisms living there see the swamp habitat as a mosaic of different
habitat types, which in turn are composed of mosaics of smaller habitat types, and so on.
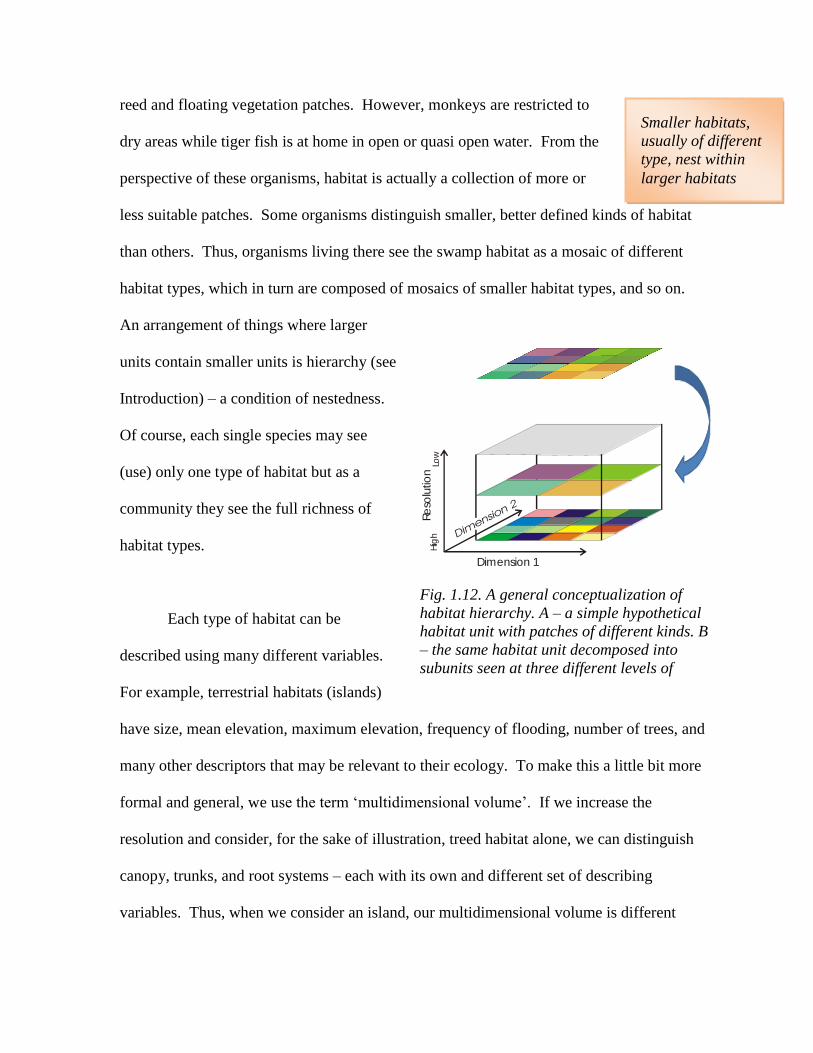
An arrangement of things where larger
units contain smaller units is hierarchy (see
Introduction) – a condition of nestedness.
Of course, each single species may see
(use) only one type of habitat but as a
community they see the full richness of
habitat types.
Each type of habitat can be
described using many different variables.
For example, terrestrial habitats (islands)
have size, mean elevation, maximum elevation, frequency of flooding, number of trees, and
many other descriptors that may be relevant to their ecology. To make this a little bit more
formal and general, we use the term ‘multidimensional volume’. If we increase the
resolution and consider, for the sake of illustration, treed habitat alone, we can distinguish
canopy, trunks, and root systems – each with its own and different set of describing
variables. Thus, when we consider an island, our multidimensional volume is different
Smaller habitats,
usually of different
type, nest within
larger habitats
Fig. 1.12. A general conceptualization of
habitat hierarchy. A – a simple hypothetical
habitat unit with patches of different kinds. B
– the same habitat unit decomposed into
subunits seen at three different levels of
resolution
Dimension 1
Re
solu
tion
Low
Hig
h

from that of a treed habitat, and so on. To capture this trend, we can start with a working
assumption that habitat is a nested hierarchy of multidimensional volumes (Fig. 1.12).
Because any dimension except time can be mapped onto space, we interpret the
habitat as a hierarchical mosaic of patches arranged such that lower level patches nest
within higher level patches, with this being true for each successive level of resolution.
Furthermore, each level and its patches may involve a distinct set of attributes. For
example, the top level in our Okavango swamp will be characterized by a ratio of water to
land. But each island (or water habitat) can be described using different variables (size,
elevation, plant cover, etc., but no longer by the ratio of water to land surface!) and each of
these habitats has its own descriptors that are appropriate. For the sake of graphical
illustration, we will use two dimensions only instead of the more realistic multidimensional
volume. This simplification gives the representation of habitat (or its model) a spatial
appearance even though this appearance represents a simplification. It does not purport to
convey any specific configuration of actual habitats in space. The model only identifies the
total amount of space a particular habitat unit occupies on average relative to a higher level
unit. Each unit can take various configurations in space as a patch that is either contiguous
or fragmented to varying degrees. Regardless of spatial configuration, each two distinct
and different habitat types are represented in the model as two subunits. Extending this
approach permits representation of the whole habitat as a nested structure of units emerging
at increasing levels of resolution (higher resolution reveals more detail – it reveals finer
habitat units).

Changing resolution, however,
has other consequences that go beyond a
mere observation of smaller fragments.
Smaller fragments of larger units have
different qualities. A familiar analogy
helps to explain this regularity: a picture
or a wall painted in yellow and blue
squares will look exactly like that, from
a short distance: a mosaic of blue and
yellow squares. When one moves far
enough, the squares are no longer visible
and the picture or wall will become
green. This effect of scale is used in
photography, digital imaging and many
other fields to produce millions of color of just 3 or 4 basic colors. In ecology examples
are less familiar and the effects of scale not always intuitively obvious but this is why we
need to be aware of them. Consider a simple two-species system: a symbiotic entity made
of a fungus and an alga (a organism called lichen, Fig. 1.13). At a very small scale, one
observes either algal cells or fungal hyphae. As one expands the scale (and reduces
resolution), a lichen appears. Properties of lichen are radically different from properties of
either of its constituents. Symbiosis (relation among components) and size of the new
system changed everything.
Fig. 1.13. Lichen –a symbiotic system
composed of two species, an alga and a fugus.
The red portions are fungal fruiting bodies.

In more general terms, it is
useful to think of habitat as a nested
mosaic of patches and represent it, as
we did in the Figure 1.14. This figure
merely emphasizes that any ecological
pattern or process adds to, or is a
constituent part of a process at a larger
scale. Furthermore, it implies that an
answer to any ecological question has
only limited validity and may change at
a scale above or below of the one at
which the examination has been conducted. For example, we cannot ask if a lichen is auto-
or heterotrophic but we can at the level of its components. Lichen itself is both auto- and
heterotrophic. We will discuss this in greater detail in Chapter 2 that concerns research
methodology in ecology.
As lower level habitat units become smaller with each level of
subdivision, they also become more fragmented (Fig. 1.14; lower panel). At
the same time, each habitat unit may occur in nature as a single block or it
may split into a number of fragments. Sometimes one type of habitat may be
embedded within another. In this case one habitat shows connection while
the other appears as isolated patches. In other situations habitats may occur
Fig. 1.14. From multitude to simplicity:
Different configurations of two habitat subunit
types (continuous, embedded, fragmented) in
upper panel are represented as a simple
hierarchical habitat model. Other
configurations may exist without, however,
changing the general way of capturing them by
the model.
Lower levels of
hierarchy consist of
many small but
different habitat
fragments:
diversification of
habitat is most
intensive at fine
scale of resolution

as isolated patches neighboring similar or different patch types (Fig. 1.14; two left panels).
Furthermore, patches may be of different sizes and, indeed, the sizes of individual patches
rarely resemble one another in shape and size. Irrespective of the number of fragments into
each conceptual unit is split on the ground, the model remains the same (as shown in the
figure on the Figure 1.14; lower panel) where all four configurations of patches lead to the
same representation of the habitat structure.
Because habitat fragmentation within one habitat type (say yellow) has various
consequences for the movement and performance of species, the model shown in the lower
panel cannot be directly used to deal with such ecological processes. Rather it aims at
organizing our thinking about the fundamental nature of the habitat. Additional features of
habitat can be added onto this model as required by a particular research question. The
number of different features that may be of interest to an ecologist is high. A few examples
of habitat fragmentation (Fig. 1.15) give a good idea of the range of combinations that
occur in nature or have been imposed on nature by human activity. One needs also to
remember that the habitats shown below are perceived by organisms differently than by the
camera that took pictures. Again, what features are important to organisms depends on the
factors discussed above. For example, if one is interested in the rodent communities
assembling in cleared forest patches, one might ask questions about the immigration of
individuals, width of forest stands that separate the cleared areas, number predators capable
of hunting in open areas and the answers to these questions might be relatively different if
we considered a forest rodent that ventures into open areas versus an obligatory user of
such areas.

Inadvertently Figure 1.15 illustrates another important fact about habitat and its
structure. Two left panels show pictures of habitats that once looked differently. Habitats
change. Their structure (types of subunits and their mutual arrangement) changes, too.
Some of these changes are slow and reflect long-term interactions between biota and
physical environment, usually soils or water chemistry, but some
of them are rapid and disruptive. Changes can affect habitat at
any and all resolutions. A tree may die and create a gap in the
canopy – an event of great importance in the dynamics of tropical
forests where seedlings wait for a death of an adult tree to gain
Habitat structure
may change at
many levels due to
internal or external
causes; changes at
higher level of
resolution are fast
Fig. 1.15. Examples of habitat patch configurations (clockwise): Embedded patches of
clear-cut forest habitat in Oregon; fragmented patches of moss and rock in Ontario; mixed
multiscale forest and clear –cut habitat in Poland; mixed embedded and fragmented coral
reefs of the Great Barrier Reef in Australia.

light. At a forests stand level, changes are slow but at the individual herb or tree level
changes are much more frequent due to grazing, trampling, or seasonal life cycle. Structure
of the forest stand however may persist for longer periods of time. The causes of change
may both be internal (a tree fell as a result of its size and trunk or root system weakened by
parasites or weight added by vines and epiphytes) or it may be external (a strong wind or
lightning or soil erosion by the nearby stream). Internal causes of change as well as
external are very likely to differ depending on the scale of habitat under consideration. We
will look at some of these aspects, particularly at the links between spatial scale and
temporal scale of processes in the next chapter.
Self-test questions
1. Which portions of the phase space in Figure 1 would be likely to fill with
species if the figure pertained to a highly variable, unpredictable habitat?
How it would differ if the habitat was very stable and highly diversified into
smaller patches of distinct qualities to which species could specialize?
2. A sandy beach of an island is at a higher or lower level of habitat hierarchy
with respect to the island itself? Is it at a higher or lower level of resolution
with respect to the same island?
3. How many different habitat types can you think of when looking at each
picture in Figure 1.8? Try to write down all the possibilities …
Suggested readings
Wu 1999. ***





























