Embed Size (px)
Citation preview

Journal of the American Society of Nephrology 75
Characterization of Cimetidine Transport in LLCPKI Cells”2
R. Bendayan,3 B. Lo, and M. Silverman
P. Bendayan, Faculty of Pharmacy, University of To-
ronto, Toronto, Ontario, Canada
B. Lo, M. Silverman, Department of Medicine, Universityof Toronto, Toronto, Ontario, Canada
(J, Am. Soc. Nephrol. 1994; 5:75-84)
ABSTRACTIn this study, cimetidine uptake and its regulation byLLCPK1 monolayers were investigated. Uptake wastemperature dependent with kinetic and specificitycharacteristics typical of a carrier-mediated mech-anism. With cimetidine uptake in the presence of anexcess concentration of the potent inhibitor quini-dine as a measure of nonspecific transport, the esti-mated kinetic parameters for cimetidine uptake at37#{176}Cunder steady-state conditions are Km 32.3 ±
6.4 �M and Vmax 20.2 ± 2. 1 pmol/mg per minute.Amiloride, quinidine, and quinine inhibited cimeti-dine uptake, whereas N�-methylnicotinamide, tetra-ethylammonium, and guanidine did not. The uptakeof cimetidine was increased in the presence of a cell
-� lumen W gradient, consistent with the behavior ofa cimetidine-W antiport system. Furthermore, the ac-tivity of both the Na�-H� exchanger and H�-ATPase
acted to dissipate the cell -� lumen W gradient,thereby decreasing net cimetidine transport. Theseresults suggest that there is a cimetidine-W ex-change system in LLCPKI cells and thatthe net secre-tion of organic base in vivo may be regulated by
luminal acidification mechanisms.
Key Words: Cimetidine, kidney, transport. LLCPKI cells
N umerous drugs of therapeutic usefulness areeither weak acids or weak bases and are elimi-
nated primarily in urine. Renal excretion Is deter-mined by: ( 1 ) glomerubar filtration of free drug; (2)solubibity in plasma versus urine on the basis of drug
pKa and liposobubility, and (3) specialized tubular
I Received November 22, 1993. Accepted March 15, 1994.2 Presented in preliminary form at the Annual Meeting of the American society
for Pharmacology and Experimental Therapeutics, August 1991. Published inabstract form in the Phormocologist, 1991 (#372).3 Correspondence to Dr. Reina Bendayan, Faculty of Pharmacy, University ofToronto, 19 Russell 5treet, Toronto, Ontario, Canada M5S 252.
1046-6673/050 1-0075$03.00/0Journal of the American Society of NephrologyCopyright C 1994 by the American society of Nephrology
secretory systems for organic anions and cations.Tubular secretion is not only a very efficient mech-anism of drug elimination (1 ) but is also the basis fordrug-drug Interactions, i.e. , cimetidine-procainamide
or cimetidine-nicotine (2,3), and drug nephrotoxicity,I.e. , acute tubular necrosis Induced by cephaboridine
(4).The transport characteristics of various endoge-
nous and exogenous catlons have been determinedin different animal species with brush bonder mem-
brane vesicles (BBMV) and basolateral membranevesicles. Studies with Ni�methylnicotmnamide (NMN),
tetraethylammonium (TEA), and cimetidmne in BBMVhave shown that their transport is saturable (Imply-ing the presence of a carrier), is specific, and is
stimulated by the countertransport of protons (5-12).
The accumulation of TEA has been studied in
monolayers of LLCPK1 and oppossum kidney (OK)cells and has been shown to be a carrier-mediatedprocess. However, contrary to previous observations
with brush border membrane vesicles, cation/proton
exchange at the luminal surface of LLCPK1 and OKcells did not appear to represent the major mecha-
nism for organic cation uptake (1 3, 1 4). Furthermore,it has recently been reported that TEA and NMN are
transported by LLCPKI cells grown on permeablesupports, such as collagen-coated filters, from thebasolateral to the apical side by a mechanism that istemperature dependent, saturable, and inhibitableby other organic cations, but transport from the api-cal to the basobateral side was found to be paracel-lulan (1 5, 1 6). Thus, there appears to be disagreement
about the nature of organic base transport at theluminal surface as defined experimentally in BBMV
prepared from whole-kidney cortex compared withtransport at the luminal surface of epithebial cell
lines in culture.Although the luminal transport of organic bases
involves net secretion (cell -� lumen), it is likely thatthe putative exchange system for organic bases bo-cated at the apical surface is operationally symmet-rical. In this study, therefore, we chose to exploit
LLCPK1 monolayens grown on impermeable supportsas an experimental model system to help define the
kinetics and specificity of uptake of a weak organicbase, cimetidine. Our results suggest the presence of
a specific cimetidine uptake at the apical surface ofLLCPKI cells that is energy dependent and function-ally regulated by the Na�-H� antiporten and H�-ATP-ase.

Cimetidine Transport by LLCPKI Cells
76 Volume 5 ‘ Number I ‘ 1994
METHODS
Culture of LLCPK1 Cells
LLCPK1 cells (Passage’s 198 to 240) were main-tamed by serial passage in 25-cm2 and 75 cm2 Falconplastic tissue culture flasks (Becton Dickinson, Lin-
coln Park, NJ). The cells were fed with Dulbecco’smodified Eagle medium containing L-glutammne andD-glucose (Gibco Laboratories, Grand Island, NY),1 0% fetal bovine serum (Gibco), and 0.5% penicillinand streptomycmn suspension (Gibco). The cells wereincubated at 37#{176}Cin an atmosphere of 95% ain-5%CO2. When confluent monolayens were formed, theywere subcultuned by trypsinization with 0.05% tryp-sin-EDTA (Gibco). On a regular basis, cells were keptfrozen at -70#{176}Cin a solution containing fetal bovineserum and 1 0% dimethylsulfoxide.
Confluent LLCPK1 cells appeared as a monolayeron the surface of the plastic dish. The presence ofmultiple domes was characteristic of confluent epi-thelial cells grown on a flat surface. Electron micros-copy confirmed that the monolayer was mainlyformed of epithebial cells with the microvillar apicalsurface extending upward into the medium.
To verify that these LLCPK1 cells retained physio-logic membrane transport properties, we measureda-methyl-o-glucopynanoside uptake in the presence
and absence of a sodium gradient using methodspreviously developed in this laboratory (17). It wasconfirmed that a-methyl-D-glucopynanoside uptake
was significantly increased in the presence of a so-
dium gradient and inhibited (90%) in the presence of
1 0 �zM phbonizmn (data not shown).
Uptake Experiments by LLCPK1 Monolayer Cells
For these experiments, cells were grown on plasticFalcon 24-well plates with growth areas of 2 cm2.
Wells were seeded at a density of 2 x 1 O� cells/well.
Cell confluency with dome formation was reachedafter 3 to 4 days. and uptake measurements weregenerally performed after 4 days.
The uptake of [3Hjcimetidmne ( 1 6.5 Ci/mmol; Amen-
sham) by LLCPK1 cells was determined by the methoddescribed by McKmnney et at. ( 1 3). Cells were firstwashed three times with 0.5 mL of incubation mediacontaining (in millimolan concentrations): sodiumchloride, 1 39; dibasic potassium phosphate, 2.5;magnesium sulfate, 1 .2; calcium chloride, 2.0; glu-
cose, 5.5; L-alanine, 6.0; sodium citrate, 1 .0; andsodium lactate, 4.0 (pH 7.4). The cells were then
preincubated in the same solution for 30 mm at 37#{176}C.After preincubation, 0.5 mL of the standard incuba-
tion media containing 1 �Ci/mL of [3H]cimetidmne andcold cimetidine at the desired concentration wasadded to the cells. Incubation was terminated with
an excess of ice-cold 0. 1 6 M sodium chloride (stopsolution). After the removal of the stop solution, the
cells were solubilized in 1 mL of 1 N sodium hydrox-
ide, and after 30 mm, they were transferred to a
scintillation vial containing 500 �L of SN HC1. Radio-activity was determined by standard liquid scintilla-tion techniques (Beckman scintillation counter).
“Zero time” uptake was subtracted from the nadioac-tivity measured at other incubation times. Cimetidine
uptake was standardized to the quantity of cell pro-tein in each culture plate. Protein was determined bythe Bradford method (1 8) with bovine serum albuminas a standard. Extracellular space as measured by
I3Hlmnulmn was small (less than 5%); therefore, nocorrection was applied. [3HINMN (66 Ci/mmol) uptake
by LLCPKI was determined by applying a methodsimilar to that described for cimetidine. Two methodshave been applied for the determination of cimetidine
specific uptake, i.e. , uptake measured at 4#{176}Cor up-
take measured in the presence of an excess concen-tration of the organic base inhibitor qumnidmne. Up-take studies performed under voltage-clamped con-ditions used the potassium ionophore valmnomycmn ata concentration of S ,zg/mg of protein per milliliter of
LLCPK1 cells. Because valinomycmn stock solution isprepared in ethanol, we performed control expeni-ments where cimetidine uptake was determined in
the presence of ethanol alone at a concentrationsimilar to the one in the valmnomycmn experiments,
i.e. , 0. 1 5%. Our results indicate that cimetidine up-take by LLCPK1 cells is not altered by this concentra-
tion of ethanol. Part of this work has been presented
in a preliminary form (19).
Fluorescence Microscopy Experiments
Fluorescence microscopy was performed with aNikon Diaphot-TMD Inverted microscope attached toa spectrofluorometer (Spex DM-3000). IntracellularpH was measured with the pH-sensitive dye biscar-
boxyethylcanboxyfluonescein (BCECF). Monolayenswere grown on glass coversbips. Cells were preincu-bated at 37#{176}Cin standard incubation media on Na�-free media for 30 mm and were then loaded with 3�M dye by preincubatlon for 5 to 7 mm. Cells wereexcited with wavelengths of 436 and 500 nm. Fluo-
nescence was recorded at 530 nm. By the calculationof the emission ratios after 436- and 500-nm exci-tation, a pH-dependent, cellular dye content-inde-
pendent value is generated. Calibration was pen-formed in each experiment by the use of nigericin(25 �tM) in potassium chloride buffers of varying ex-
tracellular pH (pH 6.8 to 7.8) with the following com-position in millimolan concentrations: KC1, 120;NaCl, 20; N-hydnoxyethylpiperazmne-N’-2-ethanesul-fonic acid (HEPES), 20 (20).
C�37 Celsius
�‘C
C.
,�
:�
Bendayan et al
Journal of the American Society of Nephrology 77
Statistical Analysis
Each experimental point is the mean of quadrupli-cate determinations, and each protocol was per-formed at least two times with two different sets ofcultured cells. Results are presented as the averageof two different experiments on as a typical expeni-ment. Statistical analysis was determined by use ofthe t test for unpaired data. Correlation betweenvalues was studied by least-squares regression analy-sis. A P value of <0.05 was considered statisticallysignificant.
Materials
[3Hjcimetidmne (16.5 Ci/mmol; Amersham), [3HJNMN (66 Ci/mmol; Amensham), [i4C1methyl�a�n�glu�
copyranoside (275.0 mCi/mmob; Dupont, NEN Re-
search Products, Boston, MA), and [‘4C]mnulin (1 to 3mCi/g; Dupont, NEN Research Products) were pun-
chased. All other chemicals were purchased fromSigma Chemical Co. (St. Louis, MO) and are of thehighest purity available. Bafibomycin A was pun-chased from Dr. Abtendorf’s laboratory (Univensit#{228}tOnasbruck, Onasbr#{252}ck, Germany) (2 1 ). BCECF was
purchased from Molecular Probes Inc.
RESULTS
Uptake Properties of Cimetidine by LLCPK1Cells
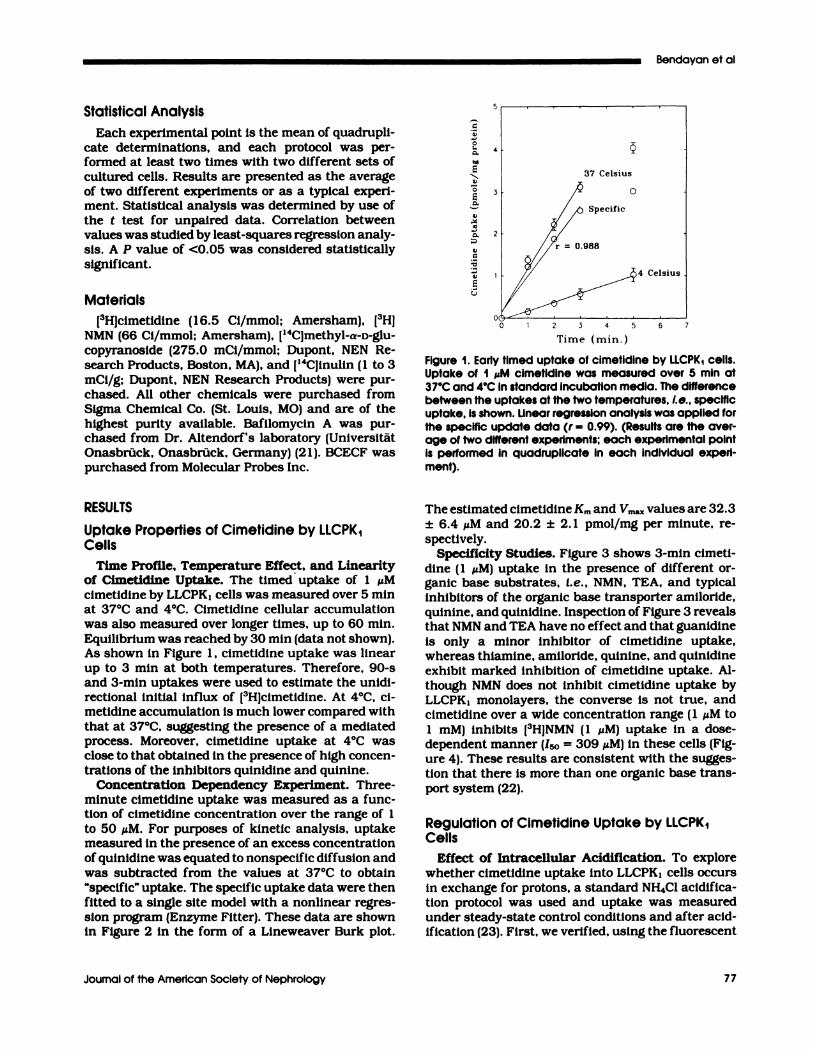
Time Profile, Temperature Effect, and Linearityof Cimetidine Uptake. The timed uptake of 1 �Mcimetidine by LLCPKI cells was measured oven 5 mmat 37#{176}Cand 4#{176}C.Cimetidine cellular accumulation
was also measured over longer times, up to 60 mm.
Equilibrium was reached by 30 mm (data not shown).As shown in Figure 1 , cimetidine uptake was linear
up to 3 mm at both temperatures. Therefore, 90-sand 3-mm uptakes were used to estimate the unidi-rectional initial influx of [3H]cimetidmne. At 4#{176}C,ci-metidine accumulation is much lower compared withthat at 37#{176}C,suggesting the presence of a mediatedprocess. Moreover, cimetidine uptake at 4#{176}Cwasclose to that obtained in the presence of high concen-tnations of the inhibitors qumnidmne and quinine.
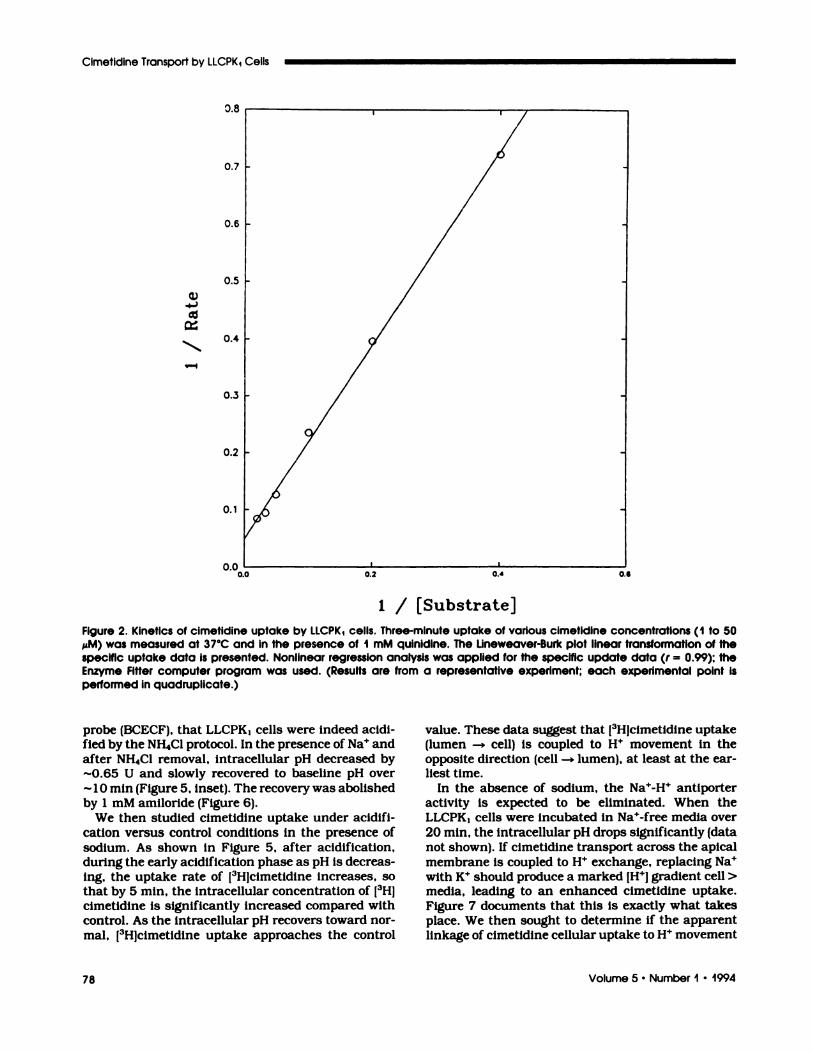
Concentration Dependency Experiment. Three-minute cimetidine uptake was measured as a func-tion of cimetidine concentration over the range of 1
to 50 �M. For purposes of kinetic analysis, uptakemeasured in the presence of an excess concentrationof qumnidmne was equated to nonspecific diffusion and
was subtracted from the values at 37#{176}Cto obtain“specific” uptake. The specific uptake data were then
fitted to a single site model with a nonlinear regnes-sion program (Enzyme Fitter). These data are shownin Figure 2 in the form of a Lineweaven Burk plot.
// ,/� Specific
� � - _�4 Celsius
/ :� � �(�‘ . .T � 4 � � -.
Tinic ( ii�in.
Figure 1. Early timed uptake of cimetidine by iLCPK1 cells.Uptake of I ,�M cimetidine was measured over 5 mm at
37#{176}Cand 4#{176}CIn standard Incubation media. The differencebetween the uptakes at the two temperatures, i.e. , specificuptake, is shown. iinear regression analysis was applied forthe specific update data (r = 0.99). (Results are the aver-age of two different experiments; each experimental pointis performed in quadruplicate in each individual experi-ment).
The estimated cimetidine Km and Vm� values are 32.3
± 6.4 �M and 20.2 ± 2. 1 pmol/mg per minute, ne-
spectiveby.
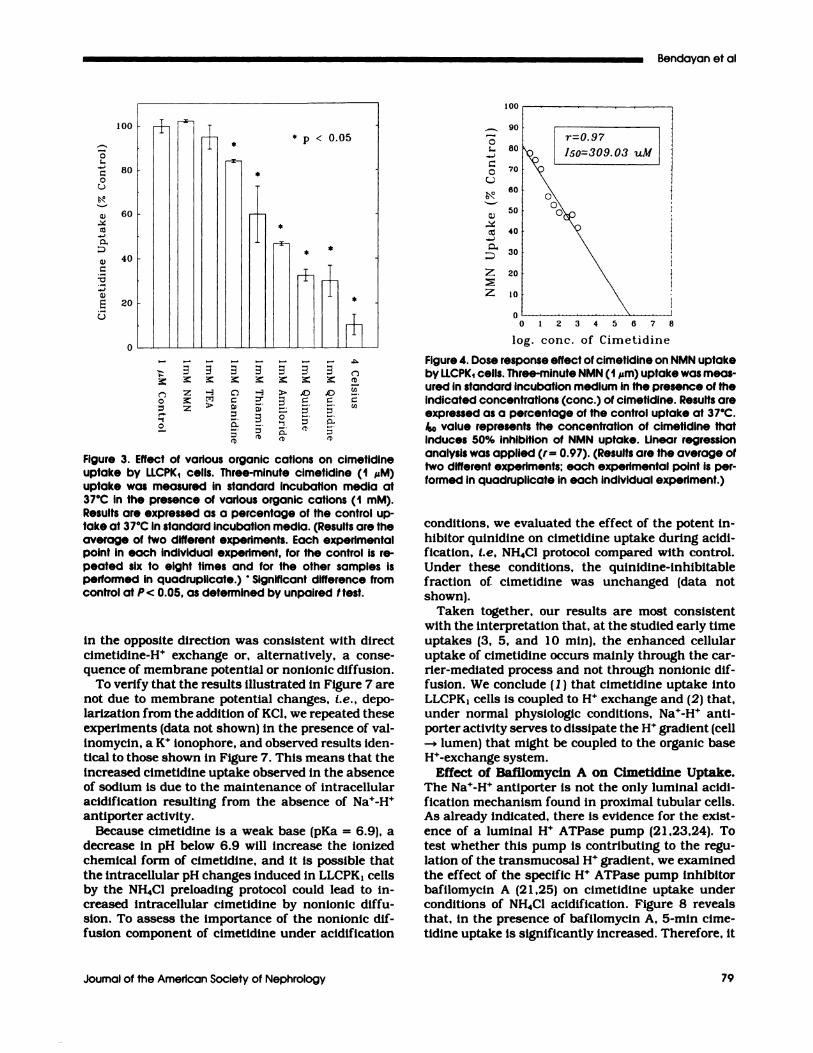
Specificity Studies. Figure 3 shows 3-mm cimeti-dine (1 �M) uptake in the presence of different or-ganic base substrates, I.e. , NMN, TEA, and typicalinhibitors of the organic base transporter amiboride,quinine, and qumnidmne. Inspection of Figure 3 revealsthat NMN and TEA have no effect and that guanidmneis only a minor inhibitor of cimetidine uptake,whereas thiamine, amibonide, quinine, and qumnidmneexhibit marked inhibition of cimetidine uptake. Al-
though NMN does not inhibit cimetidine uptake byLLCPKI monolayens, the converse is not true, andcimetidine over a wide concentration range ( 1 �tM to1 mM) inhibits [3HJNMN (1 �tM) uptake in a dose-dependent manner (150 = 309 �zM) in these cells (Fig-ure 4). These results are consistent with the sugges-tion that there is more than one organic base trans-port system (22).
Regulation of Cimetidine Uptake by LLCPK1Cells
Effect of Intracellular Acidification. To explorewhether cimetidine uptake into LLCPK1 cells occurs
in exchange for protons, a standard NH4C1 acidifica-
tion protocol was used and uptake was measuredunder steady-state control conditions and after acid-ification (23). FIrst, we verified, using the fluorescent
1 / [Substrate]
Cimetidine Transport by LLCPKI Cells
78 Volume 5 . Number I - 1994
Figure 2. Kinetics of cimetidine uptake by iiCPK1 cells. Three-minute uptake of various cimetidine concentrations (1 to 50
MM) was measured at 37#{176}Cand in the presence of I mM quinidine. The Lineweaver-Burk plot linear transformation of thespecific uptake data is presented. Nonlinear regression analysis was applied for the specific update data (r = 0.99); theEnzyme Fitter computer program was used. (Results are from a representative experiment; each experimental point isperformed in quadruplicate.)
probe (BCECF), that LLCPK� cells were indeed acidi-fied by the NH4C1 protocol. In the presence of Na� and
after NH4C1 removal, intracellular pH decreased by-0.65 U and slowly recovered to baseline pH over
1 0 mm (Figure 5, inset). The recovery was abolishedby 1 mM amibonide (Figure 6).
We then studied cimetidine uptake under acidifi-cation versus control conditions in the presence of
sodium. As shown in Figure 5, after acidification,during the early acidification phase as pH is decreas-ing, the uptake rate of [3H]cimetidmne increases, sothat by 5 mm, the intracellular concentration of [3H1cimetidine is significantly increased compared withcontrol. As the intracellular pH recovers toward nor-
mab, I3Hlcimetidine uptake approaches the control
value. These data suggest that [3Hjcimetidine uptake(lumen -+ cell) is coupled to H� movement in theopposite direction (cell -� lumen), at least at the ear-liest time.
In the absence of sodium, the Na�-H� antiporten
activity is expected to be eliminated. When the
LLCPK� cells were incubated in Na�-fnee media oven20 mm, the intracellular pH drops significantly (datanot shown). If cimetidine transport across the apical
membrane is coupled to H� exchange, replacing Na�with K� should produce a marked [W] gradient cell>media, leading to an enhanced cimetidine uptake.Figure 7 documents that this Is exactly what takesplace. We then sought to determine if the apparent
linkage of cimetidine cellular uptake to H� movement
fl�n
C
C
C
C
CC
C,
8)
8)
ioo� �
80
60 L40
20
* p < 0.05
100
90
80
70
60
50
40
30
20
10
C
z
z
C
(�)
C/C
CC/C
Bendayan et al
Journal of the American Society of Nephrology 79
�z � �c �z;� � � -� > .0 �;� � C �‘ � �
�> � � a�: � 0 � -#{149}
�- �. : �� �
Figure 3. Effect of various organic cations on cimetidineuptake by LLCPK1 cells. Three-minute cimetidine (1 �M)uptake was measured in standard incubation media at37#{176}Cin the presence of various organic cations (1 mM).Results are expressed as a percentage of the control up-take at 37#{176}Cin standard incubation media. (Results are theaverage of two different experiments. Each experimentalpoint in each individual experiment, for the control is re-peated six to eight times and for the other samples isperformed in quadruplicate.) ‘ Significant difference fromcontrol at P< 0.05, as determined by unpaired I test.
In the opposite direction was consistent with direct
cimetidine-H� exchange or, alternatively, a conse-quence of membrane potential or nonionic diffusion.
To verify that the results illustrated in Figure 7 arenot due to membrane potential changes, i.e. , depo-larization from the addition of KC1, we repeated theseexperiments (data not shown) in the presence of val-
Inomycin, a K� ionophone, and observed results iden-tical to those shown in Figure 7. This means that the
increased cimetidine uptake observed in the absenceof sodium is due to the maintenance of intracellularacidification resulting from the absence of Na�-H�antiporter activity.
Because cimetidine is a weak base (pKa = 6.9), a
decrease in pH below 6.9 will increase the ionizedchemical form of cimetidine, and it is possible that
the intracellular pH changes induced in LLCPK1 cellsby the NH4C1 preboading protocol could lead to in-creased intracellular cimetidine by nonionic diffu-sion. To assess the importance of the nonionic dif-fusion component of cimetidine under acidification
r0.97 �kc#{231}� Iso=309.03 uM
0 1 2 3 4 5 6 7 8
log. cone. of Cimetidine
Figure 4. Dose response effect of cimetidine on NMN uptakeby LLCPK1 cells. Three-minute NMN (1 ,�m) uptake was meas-ured in standard incubation medium in the presence of theindicated concentrations (conc.) of cimetidine. Results areexpressed as a percentage of the control uptake at 37#{176}C./SO value represents the concentration of cimetidine thatinduces 50% inhibition of NMN uptake. Linear regressionanalysis was applied (r = 0.97). (Results are the average oftwo different experiments; each experimental point is per-formed in quadruplicate in each individual experiment.)
conditions, we evaluated the effect of the potent in-hibitor quinidmne on cimetidine uptake during acidi-fication, I.e. NH4C1 protocol compared with control.Under these conditions, the qumnidine-mnhibitabbefraction of cimetidine was unchanged (data not
shown).Taken together, our results are most consistent
with the interpretation that, at the studied early time
uptakes (3, 5, and 10 mm), the enhanced cellularuptake of cimetidine occurs mainly through the can-nier-mediated process and not through nonionic dif-fusion. We conclude ( 1 ) that cimetidine uptake intoLLCPK1 cells is coupled to H� exchange and (2) that,under normal physiologic conditions, Na�-H� anti-porter activity serves to dissipate the H� gradient (cell
-* lumen) that might be coupled to the organic baseH�-exchange system.
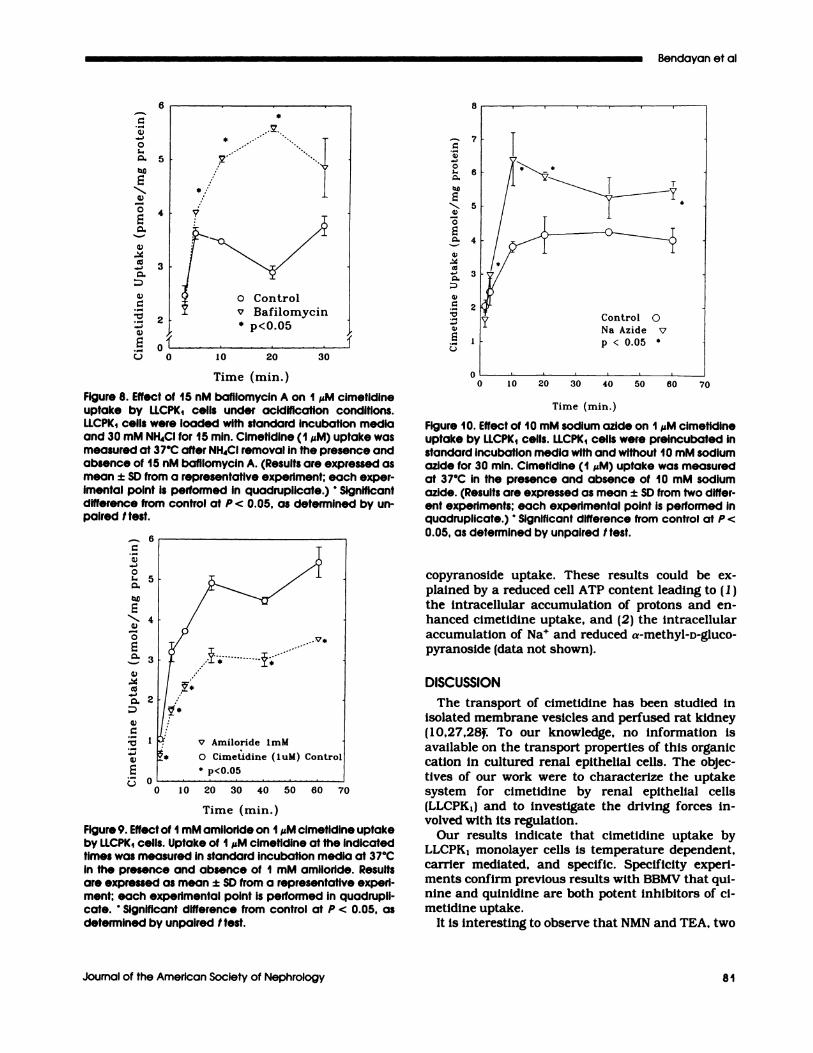
Effect of Bafilomycin A on Cimetidine Uptake.The Na�-H� antiporter is not the only luminal acidi-fication mechanism found in proximal tubular cells.As already indicated, there is evidence for the exist-
ence of a luminal H� ATPase pump (21 ,23,24). Totest whether this pump is contributing to the regu-lation of the tnansmucosal W gradient, we examinedthe effect of the specific H� ATPase pump inhibitorbafibomycmn A (2 1 ,25) on cimetidine uptake underconditions of NH4C1 acidification. Figure 8 revealsthat, in the presence of bafibomycmn A, 5-mm cime-
tidine uptake is significantly increased. Therefore, it

4.5
4.0
3.5
3.0
2.5
2.0
1.5
1.0
0.00
0 0 20 30 40 50 60 70
10 20 30 40
CI-,0�
EC)
C
E
C)
CC
0.
C)C
C)
E
Time (mm.)
Figure 5. Time profile of cimetidine uptake by LiCPK1 cellsunder acidification conditions. iLCPK1 cells were loadedwith 30 mM NH4CI for 15 mm. Cimetidine (1 �M) uptake wasmeasured in standard incubation media at 37#{176}Cafter NH4CIremoval. (Results are expressed as mean ± SD of two differ-ent experiments; each experimental point is performed inquadruplicate.) Significant difference from control at P <
0.05, as determined by unpaired Itest. Inset shows the timecourse of �pH of LiCPK1 cells after 15 mm of exposure andthe subsequent removal of 30 mM NH4CI. (Intracellular pHmeasured by quantitative fluorescence microscopy.)
is likely that, under normal physiologic conditions,
H� ATPase activity in addition to the Na�-H� anti-porter contributes to the dissipation of the apicaltransmembrane H� gradient and thus potentiallyplays a role In regulating the uptake of organic bases.
Effect of Amiloride on Cimetidine Uptake. Amil-oride is an organic base and a well-known inhibitorof the Na�-H’� antiporten system. If amiloride inter-
acts only with the Na�-H� antiporter, and not withthe organic base transporter, then the addition ofamibonide to the incubation medium should increase
cimetidine uptake. In fact, as shown in Figure 9,amiboride inhibits cimetidine uptake in the presence
of sodium. This result is best explained by postubat-ing that amilonide competes with cimetidine for a
common carrier system.
Sodium Azide Experiments. Sodium azide, an in-hibitor of oxidative phosphorylation (26), signifl-cantly enhances cimetidine uptake in the presenceof sodium (Figure 1 0) but decreases a-methyl-D-gbu-
�1
pH
Time (mm.)
Figure 6. Effect of amiloride on iLCPK1 cells, intracellular pHrecovery from an NH4CI acid load. Time course of iiCPK1intracellular pH at baseline and after 15 mm of exposureand the subsequent removal of 30 mM NH4CI (acidification).followed by pH recovery and the inhibition of this recoveryby the addition of I mM amiloride.
-9C
C1�C-
7
E-.-.... 6
C)
C
E5C-
8)4
CC
C-3
C
‘C
C)
0 50
Time (mm.)
Figure 7. Time profile of cimetidine uptake in the presenceand absence of sodium by iLCPK1 cells. Uptake of I �Mcimetidine was measured in standard incubation media atthe Indicated times at 37#{176}Cin the presence and absenceof sodium. In the absence of sodium, the standard incuba-tion media contained 139 mM KCI. (Results are expressedas mean ± SD from two different experiments; each exper-imental point Is performed in quadruplicate.) Significantdifference from control at P < 0.05, as determined by un-paired Itest.
10 20 30 40
Cimetidine Transport by LLCPKI Cells
80 Volume 5 . Number I #{149}1994
S
7
6
5
4
:110 20 30
Control C
Na Azide �
p < 0.05 #{149}
10 20 30 40 50 60 70
6
V Amiloride 1mM
0 Cimetidine (luM) ControL
S p<0,05
Bendayan et al
Journal of the American Society of Nephrology 81
C
C)
CC.,C,
bIC
C)
0
C-
C)
CC
C,
C)
C
,�
C)
6
5
4
3
2
01�
0
5
Time (mm.)
Figure 8. Effect of 15 nM bafilomycin A on I �M cimetidineuptake by LLCPK1 cells under acidification conditions.LLCPK1 cells were loaded with standard incubation mediaand 30 mM NH4CI for 15 mm. Cimetidine (1 �M) uptake wasmeasured at 37#{176}Cafter NH4CI removal in the presence andabsence of 15 nM bafilomycin A. (Results are expressed asmean ± SD from a representative experiment; each exper-imental point is performed in quadruplicate.) Significantdifference from control at P < 0.05, as determined by un-paired I test.
CC)
CC..
C-
E.�.,, 4
C)
C
E
C)‘�
CC
C,
a)C
C)
2
0 iO 20 30 40 50 60 70
Time (mm.)
Figure 9. Effectof I mM amiloride on I � cimetidine uptakeby LLCPK1 cells. Uptake of I �M cimetidine at the indicatedtimes was measured in standard incubation media at 37#{176}Cin the presence and absence of I mM amiloride. Resultsare expressed as mean ± SD from a representative experi-ment; each experimental point is performed in quadrupli-cate. ‘ Significant difference from control at P < 0.05, asdetermined by unpaired I test.
C8)
C
-8)
C
C,
C)
CC
C,
8)C
‘C
3
0
Time (mm.)
Figure 10. Effect of 10 mM sodium azide on I zM cimetidineuptake by LLCPK1 cells. LLCPK1 cells were preincubated instandard incubation media with and without 10 mM sodiumazide for 30 mm. Cimetidine (1 �M) uptake was measuredat 37#{176}Cin the presence and absence of 10 mM sodiumazide. (Results are expressed as mean ± SD from two differ-ent experiments; each experimental point is performed inquadruplicate.) ‘ Significant difference from control at P <
0.05, as determined by unpaired I test.
copyranoside uptake. These results could be ex-
plalned by a reduced cell ATP content beading to (1)
the intracellular accumulation of protons and en-
hanced cimetidine uptake, and (2) the intracellularaccumulation of Na� and reduced a-methyb-D-gluco-pynanoside (data not shown).
DISCUSSION
The transport of cimetidine has been studied inisolated membrane vesicles and perfused rat kidney
(10,27,28� To our knowledge, no information is
available on the transport properties of this organiccation in cultured renal epithelial cells. The objec-
tives of our work were to characterize the uptakesystem for cimetidine by renal epithelial cells(LLCPK1) and to investigate the driving forces in-volved with its regulation.
Our results indicate that cimetidine uptake byLLCPK1 monolayer cells is temperature dependent,carrier mediated, and specific. Specificity expeni-ments confirm previous results with BBMV that qui-
nine and qumnidine are both potent inhibitors of ci-metidine uptake.
It is interesting to observe that NMN and TEA, two

Cimetidine Transport by LLCPKI Cells
82 Volume 5 . Number I . 1994
standard substrates of the organic base transporter,
did not significantly affect cimetidine uptake. Cime-tidine, however, was found to be a potent inhibitorof NMN uptake. These results corroborate data pro-
vided by Miyamoto et at. on guanidmne transport and
suggest that multiple carrier systems exist for on-
ganic cations at the renal brush bonder membrane(I.e. , guanidine uptake was only minimally inhibitedby organic catlons such as TEA, NMN, and choline,
but was significantly inhibited by other organic cat-ions, i.e. , amiloride, cbonidine, imiprammne, and han-
maline) (22).
Little Information is presently available on the reg-ulatory processes involved with organic base trans-porters in LLCPKJ cells. Studies with NMN and ci-metidine in purified renal BBMV have suggested thattheir transport is driven by an outwardly directedproton gradient, probably generated by the Na�-H�
antiporter system in vivo (7,9, 1 0, 1 1 ,27). Although aNa�-H� antiporten system has been identified inLLCPK1 cells (29), no information is available on the
coupling of this exchanger with the luminal transportof organic cations. It has been shown, however, thatapical membranes Isolated from the LLCPK1 cell lineretain a carrier system for the organic cation TEA.This carrier system presents characteristics similarto those observed In renal BBMV isolated from ratkidney cortex, including an organic cation/protonexchange (30).
The acidification experiments performed with the
standard NH4C1 protocol led to a small but consistentincrease of initial cimetidine uptake in the presenceof sodium. This increase at early times is consistentwith the enhancement of cimetidine uptake in ex-change for protons moving from cell -+ lumen. Suchresults would corroborate previous findings in pun-fied membrane vesicles from various animal species’kidneys (7,10). On the other hand, because cimeti-dine is a weak base (pKa = 6.9), it is likely thatintracellular acidification could play a robe in en-hancing cimetidine intracellular accumulation due tothe effects of nonlonic diffusion. To assess this pos-
sibibity more directly, we evaluated the effect of thepotent inhibitor quinidine on cimetidine uptake un-den acidification conditions and observed that thequmnidmne-mnhibitabbe fraction under control and
acidification conditions remains the same. This sug-gests that the enhanced cimetidine uptake underacidification conditions occurs mainly through a can-rien-mediated process, i.e. , H�-onganic base ex-
change.We also Investigated the effect of replacing Na� by
K� and found that cimetidine uptake is significantly
enhanced in the absence of sodium. Intracellular pHmeasurements obtained by fluorescence microscopyconsistently showed a reduced pH when Na� wasreplaced by K�. In principle, replacing Na� by K�
would cause membrane depolarization and shouldlead to decreased organic cation uptake, rather than
the increase that was observed. Thus, changes inmembrane potential cannot account for the increaseIn cimetidine uptake when K� replaces Na�. Mc-
Kinney et at. also studied the effect of intracellular
and extnacellular pH alterations on TEA Influx andefflux In both LLCPK1 and OK cells but did not ob-
serve any significant changes. In these studies,McKinney et at. did not perform intracellular pHmeasurements using fluorescence microscopy expen-iments. Therefore, their results are difficult to inter-pret because the cells may have compensated for themore acidic and/or alkaline external pH oven the 30-
to 60-mm incubation time used for influx/effluxmeasurements. Our results on cimetidine uptake alsodiffer from studies with TEA (1 3, 1 4) in which intra-
cellular acidification by NH4C1 had little effect onTEA uptake by LLCPK1 and OK epithelial cell lines.Perhaps, the observed variance between cimetidineand TEA is because their cellular uptake might be
mediated by a different organic cation transporter.Amiboride, an organic base and an inhibitor of the
Na�-H� antiporter system reduced cimetidine uptakein the presence of sodium. This result is consistedwith the interpretation that amiloride competes withcimetidine for the organic base carrier system. Along
these lines, amilonide has previously been shown toshare the organic base transporter in purified renal
membrane vesicles from dog kidney (12) and in OK
cells (14).Although the Na�-W antiporten is the main cell
acidification mechanism, some active proton trans-port is primarily driven by proton pumps energized
by ATP hydrolysis (3 1 -34). Recently Jehmlich et at.
have shown that the carboxyl group reagent N,N’-
dlcycbohexylcarbodiimide, the sulfhydryb group re-agent N-ethybmaleimide, and bafilomycin inhibit therenal H�-ATPase pump. Bafibomycin was also shownto be a potent Inhibitor of brush bonder membranevacuolar ATPases (1 5 nM abolished ATP-driven H�
uptake) (24). More recently, Manger and Koeppen (25)Identified a Na�-Independent, K�-independent N-
ethylmabeimide- and bafilomycmn A-sensitive H�-
ATPase in outer medullary collecting duct cells grownin primary culture. Although there is no direct prooffor the existence of W-ATPase pumps in LLCPKI
cells, evidence strongly suggests their existence (23).We have tested the effect of bafibomycin A on the
uptake of cimetidine and observed an enhancementof the uptake in the presence of sodium, suggesting
that, by preventing proton efflux via H�-ATPase,more protons are available mntracellularly and forexchange with luminal organic bases, i.e. , cimeti-
dine.The enhanced effect of the sodium azide pretreat-
ment of LLCPK1 cells on cimetidine uptake Is ex-

Bendayan et al
Journal of the American Society of Nephrology 83
plained by sodium azide-induced ATP depletion ub-
timately compromising the primary active transportsystems such as Na�-K�-ATPase and H�-ATPase, aswell as the Na�-coupled secondary transport system
i.e. , Na�-H� exchange. The net result is gradual intra-cellular acidification, which will favor increased ci-metidine uptake through H� exchange.
In summary, we have obtained new informationabout a unique cimetldine-H� antiport system inLLCPK1 cells. The cell uptake assay used in this
system involved LLCPK1 monolayens grown in plasticdishes. We cannot, however, exclude the possibility
that a component of the measured cimetidine uptaketook place across the basobateral membrane. Never-
thebess, we have demonstrated the functional linkageof cimetidine uptake to the Na�-H� exchanger and
H�-ATPase. The nature of this linkage may providevaluable insights into factors that regulate net tu-
bular organic base secretion. Both H4-ATPase andthe Na�-H� antiporten act to dissipate cell -� lumen(H�) gradients. Consequently, under steady-state con-ditions, they would act to reduce cell uptake reab-sorption and instead promote net secretion (all otherfactors being equal). In other words, renal cell acidi-fication mechanisms may act to regulate transepi-thelial organic base transport. This hypothesis, aswell as the membrane localization of the cimetidine-H’� exchanger, will require further investigation.
ACKNOWLEDGMENTSThis study was supported by an operating grant awarded by the
Kidney Foundation of Canada. R. Bendayan Is a Career Scientist,
Ministry of Health of Ontario. B. Lo is a recipient of a Medical
Research Council of Canada Studentship. The authors thank Ms.
Wanda Georgis for her excellent technical assistance and Ms. Jen-
nifer Brown for her excellent secretarial assistance.
REFERENCES
1 . Werner IM, Mudge GH: Renal tubular mecha-nisms of excretion of organic acids and bases.Am J Med 1964;36:743-�62.
2. Somogyi AA, McLean A, Heinzow B: Cimeti-dine-procainamide, pharmacokinetic interac-tion in man: Evidence of competition for tubularsecretion of basic drugs. Eur J Clin Pharmacol1 983;25:339-345.
3. Bendayan R, Sullivan JT, Hamilton C, FreckerRC, Sellers EM: Effect of cimetidine and raniti-dine on the hepatic and renal elimination ofnicotine In humans. Eur J Clin Pharmacob 1990;38: 165-169.
4. Tune BM, Wu KY, Kempson RI: Inhibition oftransport and prevention of toxicity of cephab-oridine in the kidney. Dose responsiveness ofthe rabbit and guinea pig to probenecid. J Phan-macob Exp Then 1 977;202:466.
S. Kinsella JL, Holohan PD, Pessah NI, Ross CR:The transport of organic ions in renal corticalluminal and antiluminal membrane vesicles. J
Pharmacol Exp Then 1979;209:443-50.6. Holohan PD, Ross CR: Mechanisms of organic
cation transport in kidney plasma membranevesicles: � Ph studies. J Pharmacol Exp Then1981 ;216:294-298.
7. Sokol PP, Holohan PD, Ross CR: Electroneutnaltransport of organic cations in canine renalbrush border membrane vesicles. J PharmacobExp Then 1985;233:694-699.
8. Takano M, Inul K!, Okano T, Saito H, Hon R:Carrier mediated transport systems of tetnaeth-ylammonium in rat renal brush bonder and baso-lateral membrane vesicbes. Biochim BiophysActa 1984;773:1 13-24.
9. Wright SH: Transport of N’-methylnicotlnamldeacross brush border membrane vesicles fromrabbit kidney. Am J Physiob 1985;249:F903-F91 1.
1 0. Gisclon L, Wong FM, Giacomini MK: Cimetidinetransport in isolated luminal membrane vesiclesfrom rabbit kidney. Am J Physiob 1987;253:F141-F150.
1 1 . Hsyu PH, Giacomini KM: The pH gradient-de-pendent transport of organic cations in the renalbrush border membrane, studies with acridineorange. J Biol Chem 1 987;262:3964-3968.
1 2. Bendayan R, Sellers EM, Silverman M: Relativeinhibition kinetics of various cationic drugs onN’-methylnicotlnamide uptake by brush bordermembrane vesicles from the dog kidney cortex.Can J Physiol Pharmacob 1990;68:467-475.
13. McKinney TD, DeLeon C, Speeg KV: Organiccation uptake by a cultured renal epithelium. JCell Physiol 1988;137:513-520.
14. McKmney TD, Scheller MB, Hosford M, Mc-Ateer JA: Tetraethylammonium transport byOK cells. J Am Soc Nephrob 1 990; 1:902-909.
1 5. Fauth C, Rossier B, Roch-Ramel F: Transportof tetraethybammonium by a kidney epithellalcell line (LLCPK1). Am J Physiol 1988;2S4:F35 1 -F357.
16. Fouda AK, Fauth C, Roch-Ramel F: Transportof organic cations by kidney epithellal cell line(LLCPK1). J Pharmacol Exp Then 1990;252:286-292.
1 7. Yau C, Rao L, Silverman M: Sugar uptake intoa primary culture of dog kidney proximal tubularcells. Can J Physiol Pharmacol 1985;63:417-426.
1 8. Bradford MM: A rapid and sensitive method forthe quantitation of microgram quantities of pro-tein utilizing the principle of protein dye binding.Anal Blochem 1976;72:248-254.
19. Bendayan R, Silverman M: Comparison of N’-Methylnicotmnamide and cimetidine uptake byLLCPK, monolayer cells [Abstract 372[. ThePharmacologist 1 99 1 ;3:2 1 1.
20. Larsson SI:I, Fukuda Y, KOlare 5, Aperia A:Proliferation and intracellular pH in culturedproximal tubular cells. Am J Physiol 1990;258:F697-F704.
2 1 . Bowman EJ, Siebers A, Altendorl K: Bafibo-mycins A class of Inhibitors of membrane AT-Pases from microorganisms, animal cells andplant cells. Proc Natl Acad Sci USA 1 988;85:7972-7976.
22. Miyamoto Y, Tiruppathi C, Ganapathy V, Lei-bach FH: Multiple transport systems for organiccatlons In renal brush border membrane vesi-

Cimetidine Transport by LLCPKI Cells
84 Volume 5 . Number I - 1994
des. Am J Physiol 1989;256:F540-F548.23. Jans AWl!, Amsier K, Griewel B, Kinne RKH:
Regulation of intracellular pH in LLCPK, cellsstudied by 31P-NMR spectnoscopy. Biochim Blo-phys Acta 1987;927:203-212.
24. Jehmlich K, Sablotini J, Simon B, BruckhardtG: Biochemical aspects of H�-ATPase in renalproximal tubules: Inhibition of N, N’-dicyclo-hexyl carbodlimide, N-ethylmaleimide and bafi-lomycin. Kidney Int 1 99 1 ;40:564-70.
25. Manger TM, Koeppen BM: Characterization ofacid-base transporters in cultured outer medul-lary collecting duct cells. Am J Physiol 1992;263:F996-F 1003.
26. Frederick KA, Babish JG: Evaluation of muta-genicity and other adverse effects of occupa-tional exposure to sodium azide. Regub ToxicobPharmacol 1 982;2:308-322.
27. Takano M, hail K, Okano T, Hon R: Cimetidinetransport in rat renal brush bonder and basobat-eral membrane vesicles. Life Sci 1985;37:1579-1585.
28. Somogyl AA, Horens CM, Muirhead MR. Boch-nen F: Renal tubular secretion of amibonide andits inhibition by cimetidine in humans and inanimal model. Drug Metab Dispos 1 989; 17:
190-196.29. Cantiello HF, Scott JA, Rabito CA: Polarized
distribution of the Na�/W exchange system in arenal cell line (LLCPK,) with characteristics ofproximal tubular cells. J Blob Chem 1986;261:3252-3258.
30. Inui K, Saito H, Hon R: W-gradient-dependentactive transport of tetnaethylammonium cationin apical-membrane vesicles isolated from kid-ney epithelial cell line LLCPK1. Biochem J 1985;227:199-203.
3 1 . Kinne-Saffran E, Beauwens R, Kinne R: AnATP-dnlven proton pump in brush border mem-branes from rat renal cortex. J Membr Biol 1982;64:67-76.
32. Brown D, Hirsch S, Gluck S: Localization of aproton-pumping ATPase in rat kidney. J ClinInvest 1988;82:21 14-2126.
33. Burckhardt G, Warnock DG: Mechanism of H�secretion In the proximal convoluted tubule.Semin Nephrol 1990;10:93-103.
34. Khadouri C, Cheval L, Messy S, Barlet-Bas C,Doucet A: Characterization and control of pro-ton-ATPase along the nephron. Kidney mt i � i;40:�71-S78.


















![GARBAGE COLLECTION - Toronto · 2002-03-14 · GARBAGE COLLECTION City of T ronto Code, said Chapter 844 shall prevail.)o Amendments noted where applicable.] GENERAL REFERENCES Nuisances](https://img.pdfslide.us/doc/110x75/5f8cfdf45152966a4516e733/garbage-collection-toronto-2002-03-14-garbage-collection-city-of-t-ronto-code.jpg)









