Embed Size (px)
Citation preview

JOURNAL OF BACTERIOLOGY, May 1991, p. 2962-2968 Vol. 173, No. 90021-9193/91/092962-07$02.00/0Copyright © 1991, American Society for Microbiology
Characterization of a Neopullulanase and an cx-Glucosidase fromBacteroides thetaiotaomicron 95-1
KAREN A. SMITH* AND ABIGAIL A. SALYERS
Department of Microbiology, University of Illinois, Urbana, Illinois 61801
Received 20 August 1990/Accepted 19 February 1991
Previously, we constructed a gene disruption in the pullulanase I gene of Bacteroides thetaiotaomicron 5482A.This mutant, designated B. thetaiotaomicron 95-1, had a lower level of pullulanase specific activity than didwild-type B. thetaiotaomicron but still exhibited a substantial amount of pullulanase activity. Characterizationof the remaining pullulanase activity present in B. thetaiotaomicron 95-1 has identified an a(1->4)-D-glucosidicbond cleaving pullulanase which has been tentatively designated a neopullulanase. The neopullulanase(pullulanase II) is a 70-kDa soluble protein which cleaves a(1-4)-D-glucosidic bonds in pullulan to producepanose. The neopullulanase also cleaved a(1-4) bonds in amylose and in oligosaccharides of maltotriosethrough maltoheptaose in chain length. An a-glucosidase from B. thetaiotaomicron 95-1 was characterized. Thea-glucosidase was partially purified to a preparation containing three proteins of 80, 57, and 50 kDa. Puliulanand amylose were not hydrolyzed by the a-glucosidase. a(1->4)-D-Glucosidic oligosaccharides from maltose tomaltoheptaose were hydrolyzed to glucose by the a-glucosidase. The a-glucosidase also hydrolyzed a(1-6)-linked oligosaccharides such as panose (the product of the pullulanase II action on pullulan) and isomaltotriose.
Bacteroides thetaiotaomicron, a gram-negative obligateanaerobe that is found in the human colon in high numbers(6), can ferment a wide variety of polysaccharides (16).Among these polysaccharides are the starch amylose and thestarchlike polysaccharide pullulan (Fig. 1). Previously, wereported the cloning and characterization of pullulanasefrom B. thetaiotaomicron. The cloned pullulanase (puilula-nase I) was active as a monomer of approximately 77 kDaand cleaved ac(1-6)-D-glucosidic linkages of pullulan toproduce maltotriose (18). Directed insertional mutagenesiswas used to construct a pullulanase I-minus mutant of B.thetaiotaomicron so that the physiological significance of thecloned pullulanase could be investigated. Subsequent exam-ination of B. thetaiotaomicron 95-1 (the pullulanase I-minusmutant) revealed that although the cloned pullulanase com-prised approximately 30 to 50% of the total pullulanaseactivity in B. thetaiotaomicron, this enzyme was not essen-tial for growth of B. thetaiotaomicron on pullulan. In fact,the 95-1 mutant was able to grow on pullulan at a rate similarto that of wild-type B. thetaiotaomicron. Therefore, theremust be a second pullulan-degrading enzyme in B. thetaio-taomicron.Four types of pullulan-hydrolyzing enzymes have been
described: (i) a glucoamylase (EC 3.2.1.3) (11, 19) whichhydrolyzes pullulan from the nonreducing ends to produceglucose, (ii) a pullulanase (EC 3.2.1.41) (1) from Klebsiellapneumoniae which hydrolyzes a(1-*6)-D-glucosidic linkagesof puilulan to produce maltotriose, (iii) an isopullulanase (EC3.2.1.57) (14) from Aspergillus niger which hydrolyzesa(1-*4)-D-glucosidic linkages of pullulan to produce isopa-nose, and (iv) a neopullulanase from Bacillus stearothermo-philus which hydrolyzes a(1-*4)-D-glucosidic linkages inpullulan to produce panose (7). The most common pullula-nase is the Klebsiella pullulanase type, which hydrolyzes thea(1-*6)-glucosidic linkages of pullulan to produce maltotri-ose. The B. thetaiotaomicron pullulanase I was this type ofpullulanase. In this report we describe the purification and
* Corresponding author.
characterization of a second pullulanase (II), tentativelydesignated a neopullulanase, from B. thetaiotaomicron. Inaddition, we describe another enzyme, an a-glucosidase,which may be involved in pullulan and starch breakdown byB. thetaiotaomicron.
MATERIALS AND METHODS
Bacterial strains and media. B. thetaiotaomicron 5482A(ATCC 29148) was obtained from the culture collection ofVirginia Polytechnic Institute Anaerobe Laboratory, Blacks-burg. B. thetaiotaomicron 95-1, a derivative of B. thetaio-taomicron 5482A, has been described previously (18). Itcontains an insertion in the a(1--6)-glucosidic linkage-cleav-ing pullulanase I gene. B. thetaiotaomicron 5482A and 95-1were grown in defined medium similar to the basal mediumdescribed by Varel and Bryant (20) except that 0.1 Mpotassium phosphate buffer (pH 7.2) was used in place ofcarbonate buffer. The final concentration of carbohydrate inthe medium was 0.5%. The atmosphere was 80% nitrogen-20% carbon dioxide. To B. thetaiotaomicron 95-1 minimalmedium broth, erythromycin at 10 ,ug/ml was added. Pullu-lan was obtained from Sigma Chemical Co., St. Louis, Mo.Enzyme assays. (i) Pullulanase and amylase assays. Pullu-
lanase and amylase activities were measured by determiningthe rate of increase in reducing sugar concentration whenpullulan or amylose was incubated with enzyme at 37°C. Apullulan concentration of 4 mg/ml was found to be saturat-ing. However, amylose is fairly insoluble, and a saturatedsolution contains approximately 2 mg of amylose per ml. Thereaction mixture (2.0 ml) contained 1.8 ml of pullulan (4mg/ml) or amylose (2 mg/ml) in 20 mM potassium phosphatebuffer (pH 6.5) and 0.2 ml of the appropriately dilutedenzyme. Samples of 0.25 ml were removed at intervalsduring the 30-min incubation and heated for 5 min at 100°C.The increase in reducing sugar was measured by the methodof Dygert et al. (2). One unit of enzyme was defined as 1 ,ugof reducing sugar equivalent (glucose) produced per min in20 mM potassium phosphate buffer (pH 6.5) at 37°C. Forcolumn fractions, the reaction mixture (0.7 ml) contained
2962
on January 10, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

ENZYMES OF B. THETAIOTAOMICRON 95-1 2%3
n i
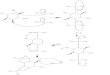
Pullulan JG G-G-J Amylose [G-G-G-G-G-G]
Maltotnose G-G-G Panose G Isomaltotriose G
G-G G
Maltose G-G Isomaltose G
G
FIG. 1. Structures of carbohydrates used in this study. G, Glu-cose; n, the a(1-4) bond cleaved by the Bacillus stearothermophi-lus neopullulanase and the B. thetaiotaomicron pullulanase II; i, thea(l1- 4) bond cleaved by the A. niger isopullulanase; p, the a(1- 6)bond cleaved by the K. pneumoniae pullulanase and the B. theta-iotaomicron pullulanase I. Horizontal lines indicate at(1- 4)-D-glucosidic bonds; vertical lines indicate a(1- 6)-D-glucosidic bonds.
0.35 ml of pullulan (4 mg/ml) or amylose (2 mg/ml), 0.3 ml of20 mM potassium phosphate buffer (pH 6.5), and 0.05 ml ofeach column fraction. Samples of 0.25 ml were drawn at 0and 30 min and heated at 1000C. The reducing sugar concen-
tration was measured as described above.(ii) a-Glucosidase assay. a-Glucosidase activity was mea-
sured by determining the rate of hydrolysis ofp-nitrophenyl-a-D-glucopyranoside at 37°C. The reaction mixture (0.5 ml)contained 0.4 ml of 20 mM potassium phosphate buffer (pH6.5), 0.05 ml of 20 mMp-nitrophenyl-a-D-glucoside, and 0.05ml of the appropriately diluted enzyme. The increase in A405was measured with a Gilford 250 recording spectrophotom-eter. A unit of enzyme activity was defined as 1 ,umol ofp-nitrophenol liberated per min in 20 mM potassium phos-phate buffer (pH 6.5) at 370C. The extinction coefficient forp-nitrophenol under these conditions was 1.3 x 104 M-1cm-l. Column fractions were assayed for a-glucosidaseactivity by mixing in the wells of a microtiter plate 75 ptl of20 mM p-nitrophenyl-a-D-glucopyranoside in potassiumphosphate buffer (pH 6.5) and 75 RId of the column fraction.The microtiter plate was incubated at 37°C for 1 to 4 h. Wellswere inspected visually for yellow color formation.
Purification of a soluble pullulanase from B. thetaiotaomi-cron 95-1. B. thetaiotaomicron 95-1 was grown in 2 liters ofdefined medium which contained pullulan as the sole carbonsource and erythromycin (10,ug/ml) to maintain the chromo-somal insertion. The cells were harvested at an opticaldensity (650 nm) of 0.7 to 1.0 by centrifugation (10,000 x g,20 min, 4C), washed twice, and resuspended in 20 to 40 mlof potassium phosphate buffer (pH 7.5). Resuspended cellswere disrupted by sonication four times for 3 to 4 min at 50to 60% maximum tip output. Disrupted cells were centri-fuged (15,000 x g, 20 min, 4°C) twice to remove cell debris,and the supernatant fluid (cell extract) was used in thepurification scheme described below.
(i) Step 1: ultracentrifugation. Cell extract was centrifugedat 200,000 x g for 12 h at 4°C, and the supernatant wasdecanted. The pellet from this centrifugation was resus-
pended in 20 ml of 20 mM potassium phosphate buffer (pH7.5) and centrifuged again (200,000 x g, 2.5 h, 4°C). The firstultracentrifugation step pelleted most of the pullulanaseactivity with the membranes. The subsequent membranewash contained the largest portion of pullulanase activity.This wash was applied to the DEAE-Sephacel column.
(ii) Step 2: DEAE-Sephacel chromatography. The secondsoluble protein fraction (membrane wash) was applied to aDEAE-Sephacel column (1.5 by 12 cm) which had beenequilibrated with 20 mM potassium phosphate buffer (pH7.5). The column was washed with buffer until no elutingprotein was detected by A280. The column was eluted with alinear 0 to 0.15 M NaCl gradient, and 1.9-ml fractions werecollected. The fractions were assayed for puilulanase, amy-lase, and a-glucosidase activity as well as NaCl concentra-tion and protein (A280). Fractions that contained pullulanaseactivity (fractions 28 through 40) but no a-glucosidase activ-ity were pooled.
(iii) a-Cyclodextrin-Sepharose 6B affinity column chroma-tography. The pooled pullulanase activity from the previousstep was concentrated in Amicon Centriprep 30 concentra-tors (Amicon, Beverly, Mass.) in a clinical centrifuge at 4°C.The Amicon Centriprep 30 was also used to desalt andequilibrate the pooled fractions to 20 mM potassium phos-phate (pH 6.5).The concentrated activity was applied to an a-cyclodex-
trin-Sepharose 6B column (1 by 7 cm) which had beenequilibrated with 20 mM potassium phosphate buffer (pH6.5). The a-cyclodextrin column was prepared according tothe manufacturer's directions, using 3 ,umol of a-cyclodex-trin (Sigma) per g of epoxy-activated Sepharose 6B (Phar-macia). The column was washed extensively with bufferuntil no eluting protein was detected (A280). The column waseluted with a linear gradient of 0 to 0.15 M NaCl, and 1.7-mlfractions were collected. The fractions were assayed forpullulanase, amylase, and a-glucosidase activity as well asNaCl concentration and protein (A280). Fractions 26 through36, which contained the highest levels of pullulanase activ-ity, were pooled, concentrated, and desalted in AmiconCentriprep 30 concentrators. The enzyme preparation wasstored in 20% glycerol at -20°C. A portion of the prepara-tion was used to determine the enzyme's substrate speci-ficity and end products.
Partial purification of the B. thetaiotaomicron 95-1 a-glu-cosidase. (i) Steps 1 and 2. Ultracentrifugation and DEAE-Sephacel anion-exchange chromatography were performedas described above for the pullulanase. The DEAE-Sephacelfractions (fractions 21 through 27) containing oa-glucosidaseactivity but no pullulanase activity were pooled and concen-trated in Amicon Centriprep 30 concentrators.
(ii) Step 3: Mono Q ion-exchange chromatography. Thea-glucosidase concentrate was dialyzed against 4 liters ofTris-HCl (pH 7.0) overnight at 4°C and then applied to anFPLC Mono Q column (Pharmacia, Uppsala, Sweden)which had been equilibrated with 20 mM Tris-HCl (pH 7.0).Under these conditions, the a-glucosidase activity remainedon the column but could be eluted with a gradient of 2 to 7%20 mM Tris-HCl-0.2 M NaCl. The flow rate was 1 ml/min,and 0.5-ml fractions were collected. Fractions were assayedfor enzyme activity and protein concentration. Fractionscontaining high levels ofa-glucosidase activity were pooled,concentrated in Amicon Centricon 30 microconcentrators,and stored in 20% glycerol at -20°C. A portion of the pooledfractions was incubated with the various carbohydrates todetermine the enzyme's breakdown products and substratespecificity.
Estimation of B. thetaiotaomicron 95-1 pullulanase nativemolecular weight. The native molecular weight of the solublepullulanase of B. thetaiotaomicron 95-1 was estimated by gelfiltration chromatography on a Bio-Rad AO.5M column. Thecolumn was equilibrated and eluted with 0.25 M NaCl-20mM potassium phosphate buffer (pH 6.5). Molecular mass
VOL. 173, 1991
on January 10, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2964 SMITH AND SALYERS
standards were alcohol dehydrogenase (150 kDa), phosphor-ylase b (97 kDa), bovine serum albumin (68 kDa), carbonicanhydrase (29 kDa), and lysozyme (14 kDa).
Gel electrophoresis. Proteins were separated by sodiumdodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) on 10% polyacrylamide gels as described by Laemm-li (8). Proteins were visualized by staining with Coomassiebrilliant blue of FASTSTAIN (Zoion Research Allston,Mass.). Molecular mass standards were myosin (H chain;200 kDa), phosphorylase b (97 kDa), bovine serum albumin(68 kDa), ovalbumin (43 kDa), carbonic anhydrase (29 kDa),and lysozyme (14 kDa).For isoelectric focusing (IEF), tube gels (9 mm by 13 cm)
were used. The IEF gels contained 5% acrylamide, 12%glycerol, 0.5% Triton X-100, and 2% ampholytes (Serva,Garden City Park, N.Y.) (for the pH range 4 to 9, the ratio ofpH 3 to 10/pH 5 to 8/pH 7 to 9 was 3:1:1 [vol/vol/vol]; for thepH range 4 to 7, the ratio of pH 4 to 6/pH 3 to 7/pH 3 to 10was 2:1:1 [vol/vol/vol]). Electrophoresis was done as de-scribed by Righetti and Drysdale (12). After electrophoresis,gels were cut into 0.5-cm slices. Gel slices to be assayed forenzyme activity were eluted in 0.5 ml of 20 mM potassiumphosphate buffer (pH 6.5). For determination of isoelectricpoint, a pH gradient gel was prepared in an identical mannerexcept that slices were eluted with 0.5 ml of deionized waterand then the pH of each slice was determined. Triton X-100(0.5%) was included in the IEF tube gels used for electro-phoresis of the Triton X-100 membrane extract (see below).
Protein determination. In most cases, protein concentra-tion was measured by the method of Lowry et al. (10). Forthe final step in enzyme purification, protein concentrationswere measured by the method of Whitaker and Granum (21).
Solubilizatlon of membrane-bound enzyme activities fromB. thetaiotaomicron membranes. The membrane pellet, whichhas been washed twice with 20 mM potassium phosphatebuffer (pH 7.5) and twice with 20 mM potassium phosphatecontaining 0.25 M NaCl (pH 7.5), was suspended in 3 ml of20 mM potassium phosphate buffer (pH 7.5). Then 1 M KCl,Triton X-100, and 1 M potassium phosphate buffer (pH 7.5)were added to make a final concentration of 0.15 M KCl-0.1M KPO4-1.5% Triton X-100 (5). This solution was agitatedgently for 3 h at 4°C and centrifuged at 200,000 x g for 2.5 hat 4°C. The resulting Triton X-100 extract was collected, andany remaining membranes were suspended in 20 mM potas-sium phosphate buffer (pH 7.5). The enzyme activitiescontained in the membrane extract were then separated onan IEF tube gel (pH range 4 to 7), and gel slice eluants wereused to digest the various carbohydrates.
Analysis of carbohydrate breakdown products. The par-tially purified pullulanase, a-glucosidase, and fractions fromthe IEF gel of the Triton X-100 membrane extract were usedto digest various carbohydrates (20 mg/ml in 20 mM KPO4[pH 6.5]) at 37°C. Carbohydrates tested were pullulan,maltoheptaose, maltohexaose, maltopentaose, maltotet-raose, maltotriose, panose, isomaltotriose, maltose, andmaltitol (Sigma) (see Fig. 1 for structures). A portion (20 ,ul)of each carbohydrate was taken prior to incubation at 37°C,and a second portion (20 to 25 R.l) was taken after digestionfor approximately 12 to 18 h. Each portion was spotted on
Whatman 3MM chromatography paper and resolved bydescending paper chromatography. Chromatograms were
developed for 20 to 30 h in a solvent system of ethylacetate-glacial acetic acid-H20 (3:1:1 [vol/vol/vol]), and thecarbohydrates were visualized by spraying the chromato-gram with a p-anisidine-phthalic acid mixture (15).
TABLE 1. Pullulanase and amylase activities and specificactivities during the stages of purification
Sp act Total activity (U)Fraction (U/mg of protein)a
Puilulanase Amylase Pullulanase Amylase
Cell extract 42 68 8,285 13,300Soluble protein 103 144 2,958 4,150DEAE-Sephacel 838 1,389 1,890 3,130a-Cyclodextrin column 996 832 741 620
a One unit of activity is defined as 1 ,ug of reducing sugar equivalent(glucose) produced per min at 37°C.
RESULTS
Characteristics of the B. thetaiotaomicron 95-1 pullulanase.As reported previously, the pullulanase I insertional mutantB. thetaiotaomicron 95-1 was able to grow on pullulan andretained 50 to 70%o of the wild-type level of pullulanaseactivity. The remaining activity was presumably due to oneor more pullulanases distinct from the enzyme characterizedpreviously. To facilitate characterization and purification ofthe remaining enzymes, we used extracts from B. thetaio-taomicron 95-1 to eliminate interference from the previouslycharacterized pullulanase I.Approximately 80% of the total pullulanase activity was
either soluble or eluted from the membranes with 20 mMpotassium phosphate buffer (pH 7.5). The other 20%o ofpullulanase activity remained associated with the mem-branes. The soluble pullulanase activity was loaded onto aDEAE-Sephacel column and, after washing, eluted with alinear gradient of 0.0 to 0.15 M NaCl in potassium phosphatebuffer (pH 7.5). A single peak of pullulanase activity elutedat approximately 0.15 M NaCl. Also, a single peak ofa-glucosidase activity eluted at approximately 0.12 M NaCl.The nonoverlapping portions of the pullulanase (fractions 28to 40) and a-glucosidase (fractions 21 to 27) peaks werepooled. Approximately 64% of the pullulanase activity in thesoluble protein fraction was recovered from the DEAE-Sephacel column (Table 1).The next step in the purification utilized an a-cyclodex-
trin-Sepharose 6B column. a-Cyclodextrin is an inhibitor ofsome types of pullulan-cleaving enzymes, and the use ofo-cyclodextrin affinity column chromatography in purifica-tion of a(1-4)-glucosidic linkage-hydrolyzing pullulanaseshas been reported previously (3). The B. thetaiotaomicron95-1 pullulanase activity was retained by the column. Sincethe a-glucosidase was not retained by the column, this stepremoved any remaining a-glucosidase activity from thepullulanase preparation. Elution of the a-cyclodextrin-Seph-arose column with a linear gradient of 0.0 to 0.15 M NaClyielded a single peak of pullulanase activity which waspooled in fractions 26 through 36. About 11 to 13% of thetotal pullulanase activity contained in the soluble ceil proteinwas recovered after the affinity chromatography step. Thepooled and concentrated fractions from the oa-cyclodextrin-Sepharose 6B column were analyzed by SDS-PAGE, and themost purified preparation (Fig. 2, lane b) contained only oneprotein band. This preparation of the 95-1 pullulanase wasused to digest various carbohydrates to determine its sub-strate specificity and end products (Table 2).The previously characterized B. thetaiotaomicron pullu-
lanase I produced maltotriose from pullulan but did nothydrolyze amylose. Thus, pullulanase I appears to be spe-cific for a(1-6)-glucosidic linkages. However, the pullula-
J. BACTERIOL.
on January 10, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

ENZYMES OF B. THETAIOTAOMICRON 95-1 2965
a b c d e
i
FIG. 2. Ten percent SDS-PAGE gel of the most purified prepa-rations of the a(1-+6)-pullulanase (I) and the a(1- 4)-pullulanase (IIor neopullulanase). Lanes: a, pullulanase I (30 jig) purified fromEscherichia coli containing pKS3-17, the pullulanase I clone; b,pullulanase II (27 ,ug) purified from B. thetaiotaomicron 95-1.Arrowheads indicate positions of protein standards.
nase examined in this study (pullulanase II) degraded amy-lose as well as pullulan (Table 1). Also, the product ofpullulan digestion was panose rather than maltotriose (Fig.3). Although maltotriose and panose had similar migrationdistances on paper chromatograms, maltotriose yielded ayellow-brown product while panose produced a green-brownproduct when the chromatogram was developed with thep-anisidine-phthalic acid reagent. Furthermore, to confirmthat the pullulanase II end product was panose rather thanisopanose, the pullulanase II pullulan hydrolysate was di-gested with A. niger amyloglucosidase and then analyzed bypaper chromatography. When amyloglucosidase hydrolyzespanose, glucose is the only product, but isomaltose andglucose would be the products produced from isopanose.The only product detected was glucose. Thus, the pullula-nase II end product was panose rather than isopanose. Inaddition, pullulanase II hydrolyzed maltotriose [a(1-*4)] tomaltose and glucose, whereas it did not hydrolyze isomal-totriose [ot(1-*6)]. These characteristics identified the puri-fied pullulanase II as an enzyme which cleaved the a(1-*4)-rather than the a(1->6)-D-glucosidic bonds. Pullulanase IIwas able to cleave ,B-cyclodextrin (a cyclic molecule of
TABLE 2. Comparison of end products of carbohydrate digestionfor pullulanase I, pullulanase II, and the a-glucosidase
End products of digestion with:aSubstrate
Pullulanase I Pullulanase II a-Glucosidase
Pullulan G3 G'Maltoheptaose - G, G2 (G-G5) GMaltohexaose - G, G2 (G-G5) GMaltopentaose - G, G2 (G-G4) GMaltotetraose - G, G2 (G-G3) GMaltotriose - G, G2 GPanose - - GIsomaltotriose - - GMaltose - - GMaltitol - -
Degrades amylose No Yes No
a G, Glucose; G2, maltose; G3, maltotriose; G3', panose; G4, maltotetraose;G5, maltopentaose; -, the carbohydrate was not hydrolyzed by the enzyme.Minor products of digestion are given in parentheses.
AI.
FIG. 3. Paper chromatogram of pullulan digested with pullula-nase II. Lanes: a, pullulan digested with a most purified preparationof pullulanase II; b, undigested pullulan; c, pullulan (undigested),panose, and glucose; d, pullulan (undigested), maltotriose, andglucose; e, glucose.
maltoheptaose), suggesting that the enzyme cleaves thecarbohydrate molecule endolytically.IEF gels of the purified preparation of the pullulanase II
indicated that the enzyme's pI was approximately 5.4. Inaddition, gel filtration on a Bio-Rad AO.5M column estimatedits native molecular mass at approximately 70 kDa. Thedenatured molecular mass from SDS-PAGE is approxi-mately 70 kDa (Fig. 2, lane b). Pullulanase I had beendetermined previously (18) to have a pl of 5.6, a nativemolecular mass of approximately 77 kDa, and a denaturedmolecular mass of approximately 80 kDa (Fig. 2, lane a).
Characterization of the B. thetaiotaomicron 95-1 a-glucosi-dase. The a-glucosidase activity was separated from thepullulanase activity at the DEAE-Sephacel anion-exchangechromatography step. The B. thetaiotaomicron 95-1 solubleprotein fraction which was loaded on the DEAE-Sephacelcolumn contained approximately 24 to 27% of the totala-glucosidase activity contained in the crude cell extract.Subsequent membrane washes continued to wash ot-glucosi-dase activity off the membranes. Approximately 70% of thetotal a-glucosidase activity in the crude cell extract wasrecovered when the activities of the soluble fractions weresummed. Approximately 14 to 19% of the ca-glucosidaseactivity contained in the crude cell extract was recoveredafter the DEAE-Sephacel anion-exchange chromatographystep. The a-glucosidase activity fraction contained many
a b
- 200kDa
- 97kDa
- 68kDa
- 43kDa
VOL. 173, 1991
on January 10, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2966 SMITH AND SALYERS
TABLE 3. End products of digestion with Triton X-100-solubilized membrane fractions
End products of digestion with:aSubstrate
Peak pI 4.4 Peak pI 5.4 Peak pl 5.8
Pullulan G3' G3 Trace GMaltohexaose G, G2 G, G2 GMaltopentaose G, G2, G3 G, G2 GMaltotetraose G, G2, G3 G, G2 GMaltotriose G, G2 G, G2 GPanose - - NDIsomaltotriose - Trace G2' GMaltose - - ND
Degrades amylose Yes Yes No
a G, Glucose; G2, maltose; G2', isomaltose; G3, maltotriose; G3, panose;-, the enzyme did not degrade the substrate; ND, not determined.
proteins after the DEAE-Sephacel step. Chromatography ona Mono Q column produced an a-glucosidase preparationcontaining only three proteins, as determined by SDS-PAGE: 80, 57, and 50 kDa. The pl of the ot-glucosidase was5.7.
This preparation of the a-glucosidase was used to deter-mine the enzyme's substrate specificity and end products(Table 3). The enzyme had little or no activity againstpuilulan, amylose, and ,-cyclodextrin but hydrolyzed G2through G7 to glucose. Also the enzyme degraded bothpanose and isomaltotriose to glucose. Thus, it appeared thatthe ca-glucosidase is able to hydrolyze both a(1-*4) anda(l-+6) linkages.
Characterization of enzyme activities from B. thetaiotaomi-cron 95-1 membranes. As reported previously (18), 16 to 18%
1.0
0.8
° 0.6
00.4
0
c 1.0IV 0.8Cu
0)
0 0.6
0.4
0.2
0
0ELI.0
00
of the pullulanase activity of B. thetaiotaomicron 5482Aremained associated with the membranes even after 0.25 MNaCl washes. This was also true for the 95-1 mutant.Approximately 50% of the activity which remained with themembranes was released by Triton X-100. Electrophoresisof this membrane extract on IEF tube gels (pH range 4 to 7)revealed three activity peaks (Fig. 4). Two activity peaks (pl4.4 and 5.4) hydrolyzed both pullulan and amylose but didnot hydrolyze p-nitrophenyl-ca-D-glucopyranoside. The thirdpeak (pl 5.8) had activity against the p-nitrophenyl-a-D-glucopyranoside but no activity against pullulan and littleactivity against amylose.
DISCUSSIONOur results demonstrate that B. thetaiotaomicron has at
least two soluble pullulanases. Pullulanase I cleaves oa(1-36)-D-glucosidic bonds and produces maltotriose from pullulan.In contrast, pullulanase II cleaves ot(1-->4)-D-glucosidic link-ages and produces panose from pullulan. Furthermore,pullulanase II, unlike pullulanase I, hydrolyzed amylose aswell as pullulan.There was pullulanase activity which remained associated
with the B. thetaiotaomicron 95-1 membranes after repeatedsalt washes and had to be solubilized with Triton X-100. Thismembrane activity appeared to be due to two differentenzymes. One had a pI value and substrate specificityvirtually identical to those of pullulanase II and could be amembrane form of pullulanase II. The other enzyme differedfrom pullulanases I and II with respect to pI but had asubstrate specificity spectrum similar to that of pullulanaseII. There have been several reported cases in Bacteroidesspecies of enzymes which cleave the same substrate or bondin a substrate but are genetically distinct from each other (4,
pH0 X N CD I_ 0oOa N _ )X X 0X No N- It CD 0 cN )M (D N a 0 _ cm c C N a) cU co It 0NU CU C') C'i o st t 't It I) e t U) It) It) It ) ItO ED 6161I I I I IIII i , , i i
I * ~ Amylase
Pullulanase
i[i11, Iaa-Glucosidase
4 6 8 10 12 14 16
GEL Slice18 20 22 24
FIG. 4. Pullulanase, amylase, and a-glucosidase activities in a Triton X-100 extract of B. thetaiotaomicron 95-1 membranes that had beenanalyzed on an IEF tube gel (pH range 4 to 7).
J. BACTERIOL.
3
5
on January 10, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

ENZYMES OF B. THETAIOTAOMICRON 95-1 2967
9). Thus, it is possible that the pI 4.4 enzyme peak is a thirdpullulanase.
Since both of the Bacteroides pullulanases degraded pul-lulan to trisaccharides but did not degrade the trisaccharides,B. thetaiotaomicron presumably needs an enzyme whichhydrolyzes these products to glucose. The a-glucosidaseappears to be such an enzyme. The most purified preparationcleaved small a(1-*4) oligosaccharides (G7 through G2) toglucose but did not hydrolyze amylose (<5%). Thus, thisenzyme appeared to hydrolyze only short-chain oligosaccha-rides. The a-glucosidase also cleaved panose and isomalto-triose to glucose, indicating that the enzyme was able tohydrolyze both a(1-*4)- and a(1->6)-D-glucosidic bonds.The ability of the a-glucosidase to hydrolyze a(1-*6)-D-glucosidic bonds may be important in the breakdown ofpullulan, since pullulanase I does not cleave a(1-*6) linkagesin small oligosaccharides such as panose. Moreover, the factthat the a-glucosidase can hydrolyze maltoheptaose butcannot hydrolyze 3-cyclodextrin suggests that the enzymeacts in an exo rather than endo manner to degrade oligosac-charides. Although most of the a-glucosidase activity issoluble, some activity remained with the membranes. The pl5.8 peak of the Triton X-100 membrane extract had the samesubstrate specificity spectrum as the soluble a-glucosidaseand a similar pl. Again, this may be a membrane-bound formof the soluble enzyme.
B. thetaiotaomicron grows on amylose as well as onpullulan. Thus far, we have not found an amylase thathydrolyzes amylose but not pullulan. However, pullulanaseII degrades both pullulan and amylose. Thus, despite theabsence of a specific amylase, pullulanase I, pullulanase II,and the a-glucosidase acting in concert should be able tohydrolyze either pullulan or amylose to glucose.The pullulanase II examined in this study had properties
similar to those of three previously characterized types ofpuilulan-degrading enzymes: an isopullulanase from A. ni-ger, an a-amylase of Thermoactinomyces vulgaris (13, 17),and a neopullulanase of Bacillus stearothermophilus (7). TheBacteroides pullulanase II, Thermoactinomyces a-amylase,and Bacillus neopullulanase produce panose from pullulanhydrolysis. The Bacillus neopullulanase also produces malt-ose and glucose in addition to panose (panose/maltose/glucose ratio of 3:1:1). Furthermore, both the Bacteroidespullulanase II and the Bacillus neopullulanase appeared toact endolytically, since they degrade ,B-cyclodextrin. TheThermoactinomyces a-amylase was not tested for the abilityto hydrolyze P-cyclodextrin. Both the Thermoactinomycesoa-amylase and the Bacteroides pullulanase II hydrolyzedamylose. The fact that the Thermoactinomyces oa-amylasecan hydrolyze pullulan differentiates it from most a-amy-lases found in nature, since most ax-amylases are unable tohydrolyze pullulan. However, the activity of the Thermo-actinomyces a-amylase against pullulan was approximatelyone-fourth of its activity against amylose, whereas theactivity of the Bacteroides pullulanase II against amylosewas approximately equal to its activity against pullulan. TheBacillus neopullulanase had very little activity against starch(i.e., only small amounts of glucose and maltose weredetected on paper chromatograms after starch was digestedfor 24 h with the neopullulanase).The Bacteroides pullulanase II resembles the A. niger
isopullulanase in that it hydrolyzed only a(1-4)-D-gluco-sidic linkages but differs in that isopullulanase producedisopanose from pullulan rather than panose. The preferencefor only a(1-*4) linkages differentiates the B. thetaiotaomi-cron pullulanase II from the Bacillus neopullulanase and the
Thermoactinomyces a-amylase, which hydrolyzed botha(1->4)- and a(1-+6)-D-glucosidic bonds. Thus, the Bacteroi-des pullulanase II appears not to be identical to any of thefour types of previously reported pullulan-degrading en-zymes, although it shares some properties with two types.Since the Bacteroides pullulanase II produces panose frompullulan, we have tentatively designated it a neopullulanase.
ACKNOWLEDGMENTS
This work was supported by Public Health Service grant RO1 Al17876 from the National Institute of Allergy and Infectious Diseases.Karen Smith was supported by Cell and Molecular Biology traininggrant GM 07283 from the National Institutes of Health.
REFERENCES1. Bender, H., and K. Wallenfels. 1966. Pullulanase (an amylopec-
tin and glycogen debranching enzyme) from Aerobacter aero-genes. Methods Enzymol. 8:555-559.
2. Dygert, S., L. H. Li, D. Florida, and J. A. Thoma. 1965.Determination of reducing sugar with improved precision. Anal.Biochem. 13:367-374.
3. Enevoldsen, B. S., L. Reimann, and N. L. Hansen. 1977. Bio-specific affinity chromatography of pullulanase. FEBS Lett.79:121-124.
4. Gherardini, F. C., M. Babcock, and A. A. Salyers. 1985.Purification and characterization of two a-galactosidases asso-ciated with catabolism of guar gum and other a-galactosides byBacteroides ovatus. J. Bacteriol. 161:500-506.
5. Gherardini, F. C., and A. A. Salyers. 1987. Characterization ofan outer membrane mannanase from Bacteroides ovatus. J.Bacteriol. 169:2031-2037.
6. Holdemann, L. V., I. J. Good, and W. E. Moore. 1976. Humanfecal flora variation in bacterial composition within individualsand possible side effects of emotional stress. Appl. Environ.Microbiol. 31:359-375.
7. Kuriki, T., S. Okada, and T. Imanaka. 1988. New type ofpullulanase from Bacillus stearothermophilus and molecularcloning and expression of the gene in Bacillus subtilis. J.Bacteriol. 170:1554-1559.
8. Laemmli, U. K. 1970. Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature (London)227:680-685.
9. Linn, S., T. Chan, L. Lipeski, and A. A. Salyers. 1983. Isolationand characterization of two chondroitin lyases from Bacteroidesthetaiotaomicron. J. Bacteriol. 156:859-866.
10. Lowry, 0. H., N. J. Rosebrough, A. L. Farr, and R. J. Randall.1951. Protein measurement with the Folin phenol reagent. J.Biol. Chem. 193:265-275.
11. Pazur, J. H., and T. Ando. 1960. The hydrolysis of glycosyloligosaccharides with a(1-+4) and a(1->6) bonds by fungalamyloglucosidase. J. Biol. Chem. 235:297-302.
12. Righetti, P. G., and J. W. Drysdale. Isoelectric focusing, p.333-587. In T. S. Work and E. Work (ed.), Laboratory tech-niques in biochemistry and molecular biology, vol. 5. Elsevier/North-Holland Publishing Co., New York.
13. Sakano, Y., S. Hiraiwa, J. Fukushima, and T. Kobayashi. 1982.Enzymatic properties and action of Thermoactinomyces vul-garis a-amylase. Agric. Biol. Chem. 46:1121-1129.
14. Sakano, Y., N. Masuda, and T. Kobayashi. 1971. Hydrolysis ofpullulan by a novel enzyme from Aspergillus niger. Agric. Biol.Chem. 35:971-973.
15. Salyers, A. A., and M. O'Brien. 1980. Cellular location ofenzymes involved in chondroitin sulfate breakdown by Bac-teroides thetaiotaomicron. J. Bacteriol. 143:772-780.
16. Salyers, A. A., J. R. Verceeloti, S. E. West, and T. D. Wilkins.1977. Fermentation of mucin and plant polysaccharides bystrains of Bacteroides from the human colon. Appl. Environ.Microbiol. 33:319-322.
17. Shimizu, M., M. Kanno, M. Tamura, and M. Suekane. 1978.Purification and some properties of a novel a-amylase producedby a strain of Thermoactinomyces vulgaris. Agric. Biol. Chem.
VOL. 173, 1991
on January 10, 2020 by guesthttp://jb.asm
.org/D
ownloaded from

2968 SMITH AND SALYERS
42:1681-1688.
18. Smith, K. A., and A. A. Salyers. 1989. Cell-associated pullula-nase from Bacteroides thetaiotaomicron: cloning, characteriza-tion, and insertional mutagenesis to determine role in pullulanutilization. J. Bacteriol. 171:2116-2123.
19. Ueda, S., K. Fujita, K. Komatsu, and N. Nakashima. 1963.Polysaccharide produced by the genus Pullularia. I. Production
of polysaccharide by growing cells. Appl. Microbiol. 11:211-215.20. Varel, V. H., and M. P. Bryant. 1974. Nutritional features of
Bacteroides fragilis subsp. fragilis. Appl. Microbiol. 28:251-257.
21. Whitaker, J. R., and P. E. Granum. 1980. An absolute methodfor protein determination based on difference in absorbance at235 and 280 nm. Anal. Biochem. 109:156-159.
J. BACTERIOL.
on January 10, 2020 by guesthttp://jb.asm
.org/D
ownloaded from