Embed Size (px)
Citation preview

Journal ofNeurochemistryRaven Press, Ltd., New York© 1994 International Society for Neurochemistry
Characterization of Lysophospholipid MetabolizingEnzymes in Human Brain
Human Neurochemical Pathology Laboratory Clarke Institute of Psychiatry, Toronto, Ontario, Canada
Abstract : Lysophospholipids are generated during theturnover and breakdown of membrane phospholipids . Wehave identified and partially characterized three enzymesinvolved in the metabolism of lysophospholipids in humanbrain, namely, lysophospholipase, lysophcspholipid :acyl-CoA acyltransferase (acyltransferase), and lysophospho-lipid:lysophospholipid transacylase (transacylase) . Eachenzyme displayed comparable levels of activity in biop-sied and autopsied human brain, although in all casesthe activity was somewhat lower in human than that inrat brain . All three enzymes were localized predominantlyin the particulate fraction, with lysophospholipase pos-sessing the greatest activity followed by acyltransferaseand transacylase . Lysophosphatidylcholine possessed aKm in the micromolar range for lysophospholipase andtransacylase, and in the millimolar range for acyltransfer-ase, whereas arachidonyl-CoA displayed a Km in the mi-cromolar range for acyltransferase . The three enzymesdiffered in their pH optima, with lysophospholipase beingmost active at pH 8.0, transacylase at pH 7.5, and acyl-transferase at pH 6.0 . Both bromophenacyl bromide andN-ethylmaleimide inhibited lysophospholipase activityand, to a lesser extent, that of acyltransferase and trans-acylase . None of the enzyme activities were affected bythe presence of dithiothreitol or EDTA, although particu-late lysophospholipase was activated approximately two-fold by the addition of 5 MM M9CI2 or CaC12 but not KCI .Transacylating activity was stimulated by CoA, the EC5oof activation being 6.8 AM. Acyltransferase displayed anapproximately threefold preference for arachidonyl-CoAover palmitoyl-CoA, whereas the acylation rate of differ-ent lysophospholipids was in the order lysophosphatidyli-nositol > 1-palmitoyl lysophosphatidylcholine > 1-oleoyllysophosphatidylcholine > lysophosphatidylserine > ly-sophosphatidylethanolamine . This, and the preference ofhuman brain phospholipase A2 for phosphatidylinositol,suggests that this phospholipid may possess a higherturnover rate than the other phospholipid classes exam-ined . Human brain homogenates also possessed the abil-ity to transfer fatty acid from lysophosphatidylcholine tolysophosphatidylethanolamine . In addition, we also pres-ent evidence that diacylglycerophospholipids can act asacyl donors for the transacylation of lysophospholipids .We have therefore demonstrated the presence of, andpartially characterized, three enzymes that are involvedin the metabolism of lysophospholipids in human brain .Our results suggest that lysophospholipase may be the
Brian M. Ross and Stephen J . Kish
1839
major route by which lysophospholipids are removedfrom the cell membrane in human brain . However, allthree enzymes likely play an important role in the remod-eling of membrane composition and thereby contributeto the overall functioning of membrane-associated pro-cesses . Key Words: Lysophospholipase-Acyl-CoAaysophospholipid acyltransferase-Lysophospho-lipid:lysophospholipid transacylase-Human brain-Ly-sophospholipids-Rat brain .J. Neurochem . 63,1839-1848 (1994) .
Lysophospholipids are generated along with fattyacids during the hydrolysis of membrane phospholip-ids by phospholipases A, (EC 3.1 .1 .32) and A2 (EC3.1 .1 .4) (PLAT and PLA2 , which hydrolyze the sn-1and sn-2 acyl groups, respectively, and are collectivelyreferred to as PLA) . The monoacyl lysophospholipidsso formed are potentially toxic in nature due to theirdetergent-like properties that, in sufficient quantities,cause membrane disruption and ultimately cell lysis(Weltzien, 1979) . Thus, levels of lysophospholipidsare strictly controlled and are normally present at lowconcentrations in the membrane compared with theirdiacyl counterparts (Koski et al ., 1978 ; Wiley et al .,1992) . Nevertheless, changes in lysophospholipid lev-els within the subtoxic range may serve to alter thefunctioning of membrane-associated processes such ashormonal secretion (Metz, 1986), calcium ion trans-port (Rustenbeck and Lenzen, 1992), the activity ofmembrane-associated enzymes such as protein kinase
Received January 27, 1994 ; revised manuscript received March22, 1994 ; accepted March 31, 1994 .
Address correspondence and reprint requests to Dr . B. M. Rossat Human Neurochemical Pathology Laboratory, Clarke Institute ofPsychiatry, 250 College Street, Toronto, Ontario, Canada M5T l R8 .Abbreviations used: acyltransferase, acyl-CoA :lysophospholipid
acyltransferase; DMSO, dimethyl sulfoxide; DTT, dithiothreitol ;lysoPC, lysophosphatidylcholine ; lysoPE, lysophosphatidylethanol-amine; lysoPI, lysophosphatidylinositol ; lysoPS, lysophosphatidyl-serine ; PC, phosphatidylcholine ; PE, phosphatidylethanolamine ; PI,phosphatidylinositol ; PLA, phospholipase A; PS, phosphatidylser-ine; transacylase, lysophospholipid :lysophospholipid transacylase .

1840
C and adenylate cyclase (Shier et al ., 1976 ; Oishi etal ., 1988), and the modulation of G protein-dependentsignal transduction (Flavahan, 1993) . Lysophospho-lipids are also formed in the maintenance and remodel-ing of the membrane's acyl group composition (Sunand MacQuarrie, 1989) .
Lysophospholipids are metabolized by a number ofdifferent enzymes, resulting in either reacylation toform phospholipids or in their further breakdown .Thus, lysophospholipase (EC 3.1 .1 .5) hydrolyzes theremaining fatty acid to produce free fatty acid andglycerophosphodiesters such as glycerophosphocho-line . Alternatively, reesterification can take place cata-lyzed by either acyl-CoA:lysophospholipid acyltrans-ferase (EC 2.3.1 .23 ; abbreviated to acyltransferase)and lysophospholipid :lysophospholipid transacylase(abbreviated to transacylase) . Acyltransferase reversesthe action of PLA by reesterifying a fatty acid onto amolecule of lysophospholipid . The fatty acid used inthis reaction is in the form of a CoA ester producedin an ATP-driven catalysis by acyl-CoA synthase .Transacylase works in a similar manner, but in thiscase the acyl donor is a second molecule of lysophos-pholipid .
Recently, several studies have implicated alteredphospholipid metabolism in a variety of human neuro-degenerative diseases including Alzheimer's disease(Pettegrew et al ., 1988 ; Blusztajn et al ., 1990 ; Nitsch etal., 1992 ; Smith et al ., 1993) and olivopontocerebellaratrophy (Kish et al., 1990, Nitsch et al ., 1993), as wellas psychiatric disorders such as schizophrenia (Pette-grew et al ., 1991 ; Fujimoto et al ., 1992) . In principle,many of the observations made in these studies couldbe explained by alterations in the functioning of thelysophospholipid metabolizing enzymes . However, todate, the reacylation enzymes have never been charac-terized in human brain, and lysophospholipase has onlybeen the subject of limited investigation (Farooqui etal ., 1988) . This study, therefore, aims to provide base-line information regarding the human enzymes,thereby allowing their role in disorders of the humanCNS to be established .
MATERIALS AND METHODS
Brain materialTemporal cortices (Brodmann area 21) from the brains
of four neurologically normal subjects of age 53 { 13 years(mean ± SEM) and postmortem interval 8 ± 1 h were usedthroughout this study . The brains had been frozen at autopsyand stored at -80°C . Biopsied temporal cortex was obtainedfrom three individuals (age, 25 ± 3 yr) who had undergonetemporal lobotomy for epileptic seizures . The biopsied tissuewas from nonepileptogenic regions as determined by depth-recording electrography . All tissue was free of gross infarctand tumors, and autopsied tissue did not show any indicationof neurodegenerative disease as assessed by neuropathologi-cal examination .
J. Neurochem., Vol. 63, No. 5, 1994
B. M. ROSS AND S. J. KISH
MaterialsRadiochemicals were purchased from New England Nu-
clear . Unlabeled phospholipids were obtained from Sigma.The lysophosphatidylinositol (lysoPI) used contained a mix-ture of esterified palmitic (58%) and stearic acids (42%),and lysophosphatidylserine (lysoPS) contained primarily es-terified stearic acid (90%) . All other chemicals were ob-tained from either BDH or Sigma and were of the highestgrade commercially available. TLC plates were 20-cm What-man silica gel 150A plates with linear-K loading strips(LK5D) .
Preparation of substrateChloroformic solutions of phospholipids, or chloroform/
methanol solutions of lysophospholipids, were transferred toa glass tube, the solvent removed under a gentle stream ofnitrogen, and the residue resuspended in water by sonicationon ice for four 10-s periods separated by 30 s, using a Bon-will Biosonics sonicator set at 50% power equipped witha 3-mm-diameter probe . Radiolabeled phosphatidylcholine(PC) and phosphatidylethanolamine (PE) were used at thesupplied specific activity, whereas lysophosphatidylcholine(lysoPC) and arachidonyl-CoA were diluted with unlabeledcompound as indicated .
Tissue preparationBrain tissue was dissected free of small blood vessels
and white matter, disrupted on ice by sonication (BonwillBiosonics sonicator set at 70% power) in 5 volumes of ice-cold homogenization buffer (1 mM EDTA, 1 mM EGTA,50 mM HEPES, pH 7 .4), and centrifuged at 100,000 ga, for30 min at 4°C to separate particulate from cytosolic material .The particulate fraction was washed once by resonicationand centrifugation, and the cytosolic fraction was respun toremove any remaining particulate material .
Standard assay proceduresAll assays were performed in duplicate with homogeniza-
tion buffer being used in blank reactions . In all cases bufferblanks gave the same values as boiled tissue preparations .
LysophospholipaseAliquots of tissue preparations were diluted using 10 mM
HEPES buffer, pH 7.4, and kept on ice before use . Lysophos-pholipase activity was assayed by incubating 0.25-0.5 legof protein, depending on which cellular fraction was used,in 25 MI of incubation medium containing 50 yM 1- [`C ] -palmitoyl lysoPC (28 .5 mCi/mmol) and 100 mM glycine,pH 8.0, at 37°C for 30 min . Reactions were terminated bythe addition of 25 ul of ethanol containing 1 % acetic acidand 100 Mg/ml of unlabeled fatty acid. The reaction productswere separated by TLC on silica gel plates developed withthe organic phase of isooctane/ethyl acetate/acetic acid/wa-ter (75 :55 :4:100 by vol) (Rordorf et al ., 1991) . The bandcorresponding to fatty acid (rf = 0.84) was scraped intoscintillation fluid and radioactivity quantitated using liquidscintillation spectroscopy .
AcyltransferaseAcyltransferase was assayed by incubating 25 ug of pro-
tein in 25 p1 of incubation medium containing 10 mM 1-palmitoyl lysoPC, 100 pM [1- ` 4C ] arachidonyl-CoA (13 .9mmol/mCi), 8.5% ethanol (from the 50% alcohol used asthe solvent for arachidonyl-CoA), and 100 mM sodium ace-tate, pH 6.0, for 5 min at 37°C . The reaction was terminated

LYSOPHOSPHOLIPID METABOLISM IN HUMAN BRAIN
TABLE 1. Activities of lysophospholipid metabolizing enzymesin rat and human brain
Each preparation was assayed under standard conditions (see Materials and Methods) .Values shown are mean nanomoles of fatty acid produced (lysophospholipase) or incorporated(acyltransferase or transacylase) per hour per milligram of protein ± SEM of data obtainedfrom three temporal cortices (autopsied or biopsied human tissue) or three whole rat brains .Note that assuming the phospholipid synthesized in the transacylase reaction is all dueto transacylation, the rate of phospholipid formation will be one-half that of fatty acidincorporation .
by the addition of 125 MI of 0.5 M HCl and extracted twicewith 300 pl of chloroform/methanol (2 :1, vol/vol) . Thepooled organic phases were then evaporated to dryness byvacuum centrifugation and resuspended in 40 pl of chloro-form/methanol (2:1, vol/vol) containing 100 pg/ml of theappropriate unlabeled standards. The resuspended lipidswere then analyzed by TLC on silica gel plates developedwith chloroform/methanol/water (65:35:4 by vol) (van denBosch et al ., 1973), which permitted the rapid separation ofPC (rf = 0.40) from lysoPC (rf = 0.13) and fatty acid (rf= 1 .00) . In some experiments, the plate was first developedfor 10 cm with chloroform/methanol/acetic acid/water(75:45:8 :2 by vol) (Kramer et al ., 1984), which permittedthe consistent separation of lysoPC (rf = 0.07), PC (rf= 0.27), phosphatidylserine/phosphatidylinositol (PS/PI)(rf = 0.63), and PE (rf = 0.90) . To better separate PEfrom the solvent front, the plate was dried and redevelopedwith the organic phase of isooctane/ethyl acetate/aceticacid/water (75:55:4:100 by vol) . Routinely, the band corre-sponding to PC was scraped into scintillation fluid and radio-activity quantitated using liquid scintillation spectroscopy .
TransacylaseTransacylase was assayed by incubating 50 pg of protein
in 25 MI of incubation medium containing 200 AM [ 1-'°C ] -palmitoyl lysoPC (28.5 mCi/mmol) and 100 mM HEPES,pH 7.5, for 30 min at 37°C . The reaction was terminatedand the PC formed quantitated as described for the assay ofacyltransferase.
PLAZ digestion of reaction productsTo determine the proportion of radioactively labeled fatty
acids at the sn-1 and sn-2 positions, the PC formed in theacyltransferase and transacylase reactions was isolated byTLC as described above. The area of silica gel containingthe PC band was then scraped into 1 ml of chloroform/methanol (2 :1, vol/vol) and incubated with agitation at roomtemperature for 10 min. The solvent was aspirated and thesilica gel extracted with a further 0.5 ml of chloroform/methanol . The combined solvent extracts were then dried byvacuum centrifugation and the residue resuspended in 50 MIof dimethyl sulfoxide (DMSO) . This was then incubated inthe presence or absence of 80 units of PLAZ from Crotalusadamanteus venom (Sigma) in 50 mM glycylglycine, pH8.9 (total volume, 150 pl) for 1 h at 37°C . The incubation
184 1
mixture was then extracted with chloroform/methanol fol-lowed by TLC to separate and quantitate PC, lysoPC, andfatty acid as described above.
MiscellaneousProtein concentration was assayed using the Bio-Rad pro-
tein assay according to the manufacturer's instructions, usingbovine serum albumin as a standard .
RESULTS
GeneralThe activities of lysophospholipase, acyltransferase,
and transacylase were determined in homogenates andin particulate and cytosolic fractions prepared fromhuman temporal cortex . In all cases the enzymes werefound to be localized mainly in the particulate fraction,with the cytosol containing relatively low levels ofactivity (Table 1) . Each enzyme was somewhat moreactive in rat than in humanbrain (Table 1 ; on average,rat brain homogenates contained 55% more lysophos-pholipase activity than human homogenates, 230%more acyltransferase activity, and 43% more transacy-lase activity) . However, biopsied human temporal cor-tex gave activities comparable with those determinedin autopsied tissue .
Under optimized conditions the enzyme assays werefound to be linear with respect to protein and time overthe following ranges (data not shown) : lysophospholi-pase, up to 1 yg of protein over 45 min ; acyltransfer-ase, up to 100 yg of protein over 6 min; transacylase,up to 75 hg of protein over 30 min. The rate ofincorpo-ration of fatty acids into PC by acyltransferase wassimilar irrespective of whether the radiolabel was pres-ent in the lysoPC or arachidonyl-CoA (see Table 5) .In the absence of exogenous lysoPC, some incorpora-tion of radioactivity into PC was still observed, indicat-ing the presence of low levels of endogenous lysoPC(see Table 5) . However, under the reaction conditionsnormally used, the rate of incorporation of labeled ara-chidonic acid into endogenous lysoPC was < 1% oftotal activity .
J. Neurochem., Vol. 63, No . 5, 1994
Fraction Lysophospholipase Acyltransferase Transacylase
Homogenate 275 ± 8 87 ± 6 0.28 ± 0.03Particulate 145 ± 7 147 ± 2 0.32 ± 0.06Cytosol 15 ± 5 Not detectable 0.04 ± 0.02Biopsy (homogenate) 325 ± 29 96 ± 9 0.35 ± 0.10Rat (homogenate) 427 ± 26 287 ± 6 0.40 ± 0.10
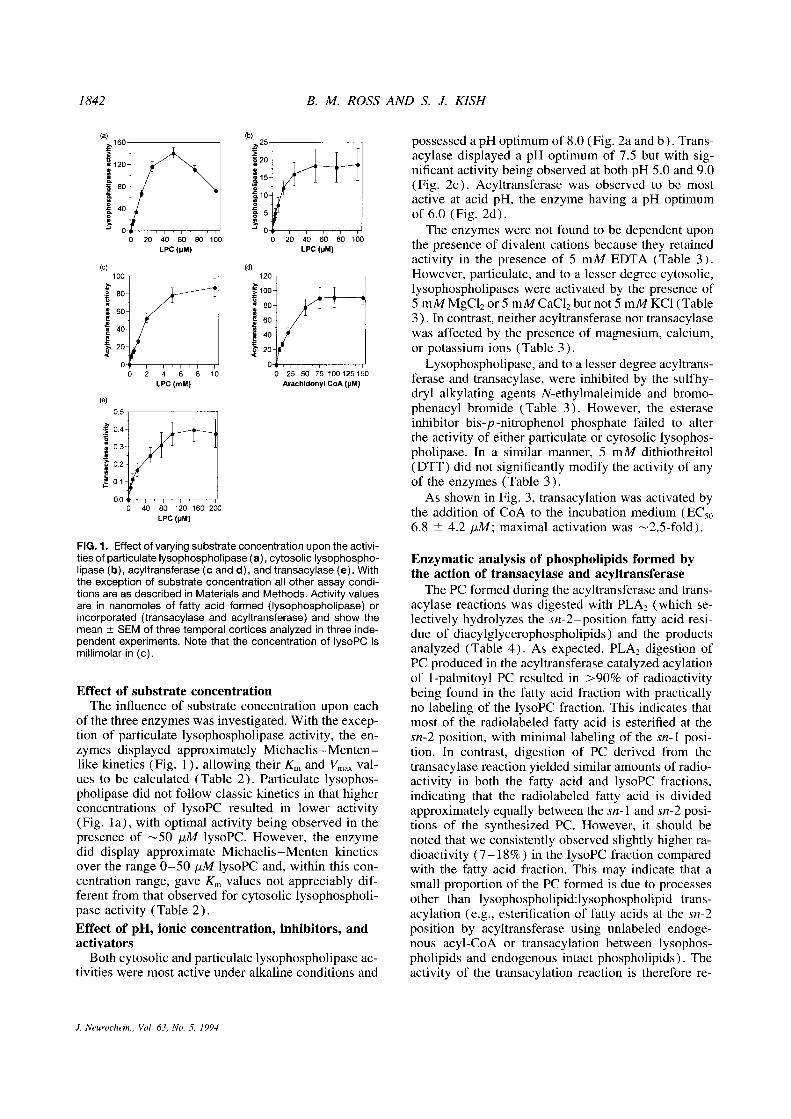
1842
FIG. 1. Effect of varying substrate concentration upon the activi-ties of particulate lysophospholipase (a), cytosolic lysophospho-lipase (b), acyltransferase (c and d), and transacylase (e) . Withthe exception of substrate concentration all other assay condi-tions are as described in Materials and Methods. Activity valuesare in nanomoles of fatty acid formed (lysophospholipase) orincorporated (transacylase and acyltransferase) and show themean ± SEM of three temporal cortices analyzed in three inde-pendent experiments. Note that the concentration of lysoPC ismillimolar in (c) .
Effect of substrate concentrationThe influence of substrate concentration upon each
of the three enzymes was investigated . With the excep-tion of particulate lysophospholipase activity, the en-zymes displayed approximately Michaelis-Menten-like kinetics (Fig . 1), allowing their Km and Vmax val-ues to be calculated (Table 2) . Particulate lysophos-pholipase did not follow classic kinetics in that higherconcentrations of lysoPC resulted in lower activity(Fig . la), with optimal activity being observed in thepresence of -50 jtM lysoPC . However, the enzymedid display approximate Michaelis-Menten kineticsover the range 0-50 yM lysoPC and, within this con-centration range, gave Km values not appreciably dif-ferent from that observed for cytosolic lysophospholi-pase activity (Table 2) .Effect of pH, ionic concentration, inhibitors, andactivatorsBoth cytosolic and particulate lysophospholipase ac-
tivities were most active under alkaline conditions and
J. Neurochem., Vol . 63, No . 5, 1994
B. M. ROSS AND S. J. KISH
possessed a pH optimum of 8.0 (Fig . 2a and b) . Trans-acylase displayed a pH optimum of 7.5 but with sig-nificant activity being observed at both pH 5 .0 and 9.0(Fig . 2c) . Acyltransferase was observed to be mostactive at acid pH, the enzyme having a pH optimumof 6.0 (Fig . 2d) .The enzymes were not found to be dependent upon
the presence of divalent cations because they retainedactivity in the presence of 5 mM EDTA (Table 3) .However, particulate, and to a lesser degree cytosolic,lysophospholipases were activated by the presence of5 MMMgC12 or 5 mM CaC12 but not 5 mM KCl (Table3) . In contrast, neither acyltransferase nor transacylasewas affected by the presence of magnesium, calcium,or potassium ions (Table 3) .
Lysophospholipase, and to a lesser degree acyltrans-ferase and transacylase, were inhibited by the sulfhy-dryl alkylating agents N-ethylmaleimide and bromo-phenacyl bromide (Table 3) . However, the esteraseinhibitor bis-p-nitrophenol phosphate failed to alterthe activity of either particulate or cytosolic lysophos-pholipase . In a similar manner, 5 mM dithiothreitol(DTT) did not significantly modify the activity of anyof the enzymes (Table 3) .As shown in Fig . 3, transacylation was activated by
the addition of CoA to the incubation medium (ECso6.8 ± 4.2 uM; maximal activation was ^-2.5-fold) .
Enzymatic analysis of phospholipids formed bythe action of transacylase and acyltransferaseThe PC formed during the acyltransferase and trans-
acylase reactions was digested with PLAZ (which se-lectively hydrolyzes the sn-2-position fatty acid resi-due of diacylglycerophospholipids) and the productsanalyzed (Table 4) . As expected, PLAZ digestion ofPC produced in the acyltransferase catalyzed acylationof 1-palmitoyl PC resulted in >90% of radioactivitybeing found in the fatty acid fraction with practicallyno labeling of the lysoPC fraction . This indicates thatmost of the radiolabeled fatty acid is esterified at thesn-2 position, with minimal labeling of the sn-1 posi-tion . In contrast, digestion of PC derived from thetransacylase reaction yielded similar amounts of radio-activity in both the fatty acid and lysoPC fractions,indicating that the radiolabeled fatty acid is dividedapproximately equally between the sn-1 and sn-2 posi-tions of the synthesized PC . However, it should benoted that we consistently observed slightly higher ra-dioactivity (7-18%) in the lysoPC fraction comparedwith the fatty acid fraction . This may indicate that asmall proportion of the PC formed is due to processesother than lysophospholipid:lysophospholipid trans-acylation (e.g ., esterification of fatty acids at the sn-2position by acyltransferase using unlabeled endoge-nous acyl-CoA or transacylation between lysophos-pholipids and endogenous intact phospholipids) . Theactivity of the transacylation reaction is therefore re-

LYSOPHOSPHOLIPID METABOLISM IN HUMAN BRAIN
TABLE 2 . Km and V�,ax values for lysophospholipid metabolizing enzymes inautopsied human brain
All enzymes were assayed as described in Materials and Methods except for varying thesubstrate concentration . Values are given as mean -_ SEM of data obtained from threetemporal cortices analyzed in independent experiments . K�, values are micromolar and V,nllxvalues as nanomoles of fatty acid produced (lysophospholipase) or incorporated (acyltransfer-ase and transacylase) per hour per milligram of protein. K,� values of lysoPC for acyltransferasewere calculated using an arachidonyl-CoA concentration of 100 PM, and for arachidonyl-CoA using a lysoPC concentration of 10 mM.Km and V�,ax values were calculated from substrate concentrations in the range 2-50 pM.
ported in terms of fatty acid incorporated rather thanamount of phospholipid formed .
Substrate preference of acyltransferaseAcyltransferase displayed an approximately three-
fold preference for arachidonyl-CoA (C20:4) over pal-mitoyl-CoA (C16:0) (Table 5) . There were also dif-ferences in the rate of acylation of the different lyso-phospholipid classes ; lysoPI was acylated most rapidly
FIG. 2. Effect of varying pH upon the activities of particulatelysophospholipase (a), cytosolic lysophospholipase (b), trans-acylase (c), and acyltransferase (d) . The buffers used for eachpH range are as follows: pH 5-6.5, 100 mM sodium acetate;pH 7-8, 100 mM HEPES; and pH 8.5-9, 100 mM glycine. Allother assay conditions are as described in Materials and Meth-ods. Activity values are in nanomoles of fatty acid formed (lyso-phospholipase) or incorporated (transacylase and acyltransfer-ase) and show the mean ± SEM of three temporal cortices ana-lyzed in three independent experiments.
1843
followed by lysoPC and lysoPS, whereas acylation oflysophosphatidylethanolamine (lysoPE) was barelydetectable under the conditions used (Table 5) . Thefatty acid esterified to the lysophospholipid also af-fected the rate of incorporation as acylation of 1-palmi-toyl (C16 :0) lysoPC proceeded approximately twiceas rapidly as that of I -oleoyl (C18:1) lysoPC (Ta-ble 5) .
Transacylation between phospholipid classesHuman brain homogenates possessed the ability to
transfer fatty acid from lysoPC to lysoPE but not tolysoPS or lysoPI (Table 6) . In addition, incubation ofsn-2-labeled PE with lysoPC resulted in the formationof labeled PC, indicating that the preparations con-tained enzyme(s) capable of catalyzing phospho-lipid:lysophospholipid transacylation (Table 6) . How-ever, incubation of either dilabeled or sn-2-labeledPC with lysoPE did not result in the formation of la-beled PE (Table 6) .
DISCUSSION
The timely removal of lysophospholipids producedby the action of PLA is an important process in themaintenance and functioning of brain membranes . Sev-eral enzymes have been suggested to play a role in thisprocess including lysophospholipase, acyltransferase,and lysophospholipid transacylase (Sun and MacQuar-rie, 1989) . We have now demonstrated the presenceof, and partially characterized, these enzymes in humanbrain . All three enzymes possessed similar activitiesin biopsied versus autopsied human tissue, suggestingthat they are relatively stable postmortem . Moreover,the activity of lysophospholipase and transacylase inrat brain was only 50% higher than that in human,although acyltransferase was several times more activein rat compared with human brain .
J. Neurochem, Vol . 63, No . 5, 1994
Enzyme Substrate Km
Lysophospholipase(particulate) LysoPC 19 .3 i 2 .4° 183 ± 9°
Lysophospholipase(cytosol) LysoPC 8 .1 ± 2 .3 19 .7 ± 4.8
Acyltransferase LysoPC 2,600!- 500 114 ± 20Arachidonyl-CoA 21 .6 ± 4 .0 102 i 13
Transacylase LysoPC 29 .5 ± 0 .4 0 .44 ± 0 .08
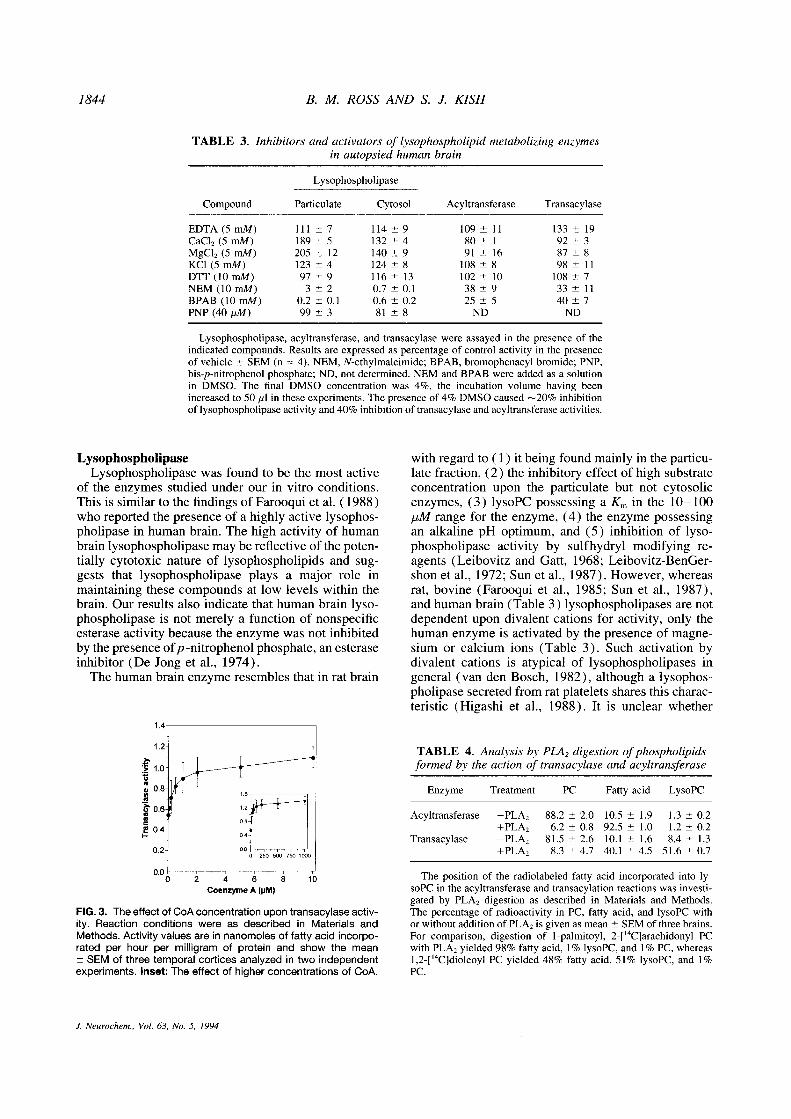
1844
TABLE 3 . Inhibitors and activators of lysophospholipid metabolizing enzymesin autopsied human brain
Lysophospholipase, acyltransferase, and transacylase were assayed in the presence of theindicated compounds. Results are expressed as percentage of control activity in the presenceof vehicle ± SEM (n = 4) . NEM, N-ethylmaleimide ; BPAB, bromophenacyl bromide; PNP,bis-p-nitrophenol phosphate; ND, not determined . NEM and BPAB were added as a solutionin DMSO . The final DMSO concentration was 4%, the incubation volume having beenincreased to 50 Ml in these experiments. The presence of 4% DMSO caused ^20% inhibitionof lysophospholipase activity and40% inhibition of transacylase and acyltransferase activities.
LysophospholipaseLysophospholipase was found to be the most active
of the enzymes studied under our in vitro conditions .This is similar to the findings of Farooqui et al . (1988 )who reported the presence of a highly active lysophos-pholipase in human brain. The high activity of humanbrain lysophospholipase may be reflective of the poten-tially cytotoxic nature of lysophospholipids and sug-gests that lysophospholipase plays a major role inmaintaining these compounds at low levels within thebrain . Our results also indicate that human brain lyso-phospholipase is not merely a function of nonspecificesterase activity because the enzyme was not inhibitedby the presence ofp-nitrophenol phosphate, an esteraseinhibitor (De Jong et al ., 1974) .The human brain enzyme resembles that in rat brain
FIG. 3. The effect of CoAconcentration upon transacylase activ-ity . Reaction conditions were as described in Materials andMethods. Activity values are in nanomoles of fatty acid incorpo-rated per hour per milligram of protein and show the meanSEM of three temporal cortices analyzed in two independent
experiments. Inset: The effect of higher concentrations of CoA.
J. Neurochem ., Vol. 63, No. 5, 1994
B. M. ROSS AND S. J. KISH
with regard to (1) it being found mainly in the particu-late fraction, (2) the inhibitory effect of high substrateconcentration upon the particulate but not cytosolicenzymes, (3) lysoPC possessing a K�, in the 10-100pM range for the enzyme, (4) the enzyme possessingan alkaline pH optimum, and (5) inhibition of lyso-phospholipase activity by sulfhydryl modifying re-agents (Leibovitz and Gatt, 1968 ; Leibovitz-BenGer-shon et al ., 1972; Sun et al ., 1987) . However, whereasrat, bovine (Farooqui et al ., 1985 ; Sun et al., 1987),and human brain (Table 3) lysophospholipases are notdependent upon divalent cations for activity, only thehuman enzyme is activated by the presence of magne-sium or calcium ions (Table 3) . Such activation bydivalent cations is atypical of lysophospholipases ingeneral (van den Bosch, 1982), although a lysophos-pholipase secreted from rat platelets shares this charac-teristic (Higashi et al ., 1988) . It is unclear whether
TABLE 4 . Analysis by PLA2 digestion ofphospholipidsformed by the action of transacylase and acyltransferase
Enzyme
Acyltransferase
Transacylase
The position of the radiolabeled fatty acid incorporated into ly-soPC in the acyltransferase and transacylation reactions was investi-gated by PLAZ digestion as described in Materials and Methods.The percentage of radioactivity in PC, fatty acid, and lysoPC withor without addition of PLAZ is given as mean - SEM of three brains.For comparison, digestion of 1-palmitoyl, 2-[' °C]arachidonyl PCwith PLAZ yielded 98% fatty acid, 1% lysoPC, and 1 % PC, whereas1,2-[' °C]dioleoyl PC yielded 48% fatty acid, 51% lysoPC, and 1%PC .
Compound
Lysophospholipase
Particulate Cytosol Acyltransferase Transacylase
EDTA (5 mM) III ± 7 114 - 9 109 ± 11 133 ± 19CaC1 2 (5 m1d) 189 ± 5 132 - 4 80 !- 1 92 ± 3MgC1 2 (5 mM) 205 ± 12 140 ± 9 91 ± 16 87 ± 8KCl(5mM) 123-4 124-8 108-8 98-11DTT(10mM) 97-9 116-13 102-10 108-7NEM (10mM) 3-2 0.7-0.1 38-9 33-11BPAB (10mM) 0.2-0.1 0.6-0.2 25-5 40-7PNP (40 pM) 99 - 3 81 - 8 ND ND
Treatment PC Fatty acid LysoPC
-PLAZ 88.2-2.0 10.5-1 .9 1 .3-0.2+PLAZ 6.2-0.8 92.5-1.0 1 .2-0.2-PLAZ 81 .5 ± 2.6 10 .1 ± 1.6 8.4 ± 1.3+PLAZ 8.3 ± 4.7 40 .1 ± 4.5 51 .6 ± 0.7

LYSOPHOSPHOLIPID METABOLISM IN HUMAN BRAIN
TABLE 5 . Substrate preference ofautopsied human brain acyltransferase
The indicated concentrations of lysophospholipid and acyl-CoA were incubated along with25 pg of homogenate protein in 100 mM sodium acetate, pH 6 .0, for 5 min at 37 °C . C16 :0,palmitoyl ; C18 :0, stearoyl ; C18 :1, oleoyl ; C20:4, arachidonyl. LysoPI contained a mixtureof C16 :0 and C18:0 fatty acids (see Materials and Methods) . Results are expressed as meannanomoles of fatty acid incorporated into PC, PE, PI, or PS as appropriate per hour permilligram of protein ± SEM of three temporal cortices and are representative of threeindependent experiments .
the observed stimulation by magnesium or calciumions is due to a direct activatory effect upon the enzymeor by altering the physical state of the substrate .
AcyltransferaseAcyltransferase was observed to be substantially
more active, under our in vitro conditions, than trans-acylase in both autopsied and biopsied human brain .Acyltransferase displayed an acid pH optimum com-pared with the neutral pH optimum of rat brain micro-somal acyltransferase (Vaswani and Ledeen, 1989) butwas inhibited by sulfhydryl group modifiers in a waysimilar to the rat islet acyltransferase (Metz, 1987) .
TABLE 6 . Transacylation activity between different lysophospholipid andphospholipid classes in autopsied human brain
We also observed that lysoPC displayed a K�, in themillimolar range for human brain acyltransferase com-pared with the micromolar range reported for the en-zyme prepared from other sources (e.g ., van Heusdenet al ., 1980; Gross and Sobel, 1982; Vaswani andLeeden, 1989) . If this represents the actual affinity ofthe enzyme for lysophospholipids in vivo, then onewould expect this enzyme to be of relatively minorimportance compared with lysophospholipase . How-ever, because we measured the activity of the enzymein whole tissue homogenates as opposed to the micro-somal preparations used by others, we cannot rule outthat the higher levels of endogenous diacylphospholip-
The indicated "C-labeled phospholipid (100 yM) was incubated along with 50 Pg of brainhomogenate protein in a 100 mM HEPES, pH 7.5 buffer for 45 min at 37°C, either in thepresence or absence of 100 pM of the indicated unlabeled compound. The table shows the rateof incorporation of labeled fatty acid into each phospholipid class expressed as a percentage ofthat observed in the absence of any additions . C16 :0, palmitoyl ; C18 :1, oleoyl ; C20 :4, arachi-donyl ; NA, not applicable . Values shown are the mean ± SEM of that observed in threetemporal cortices and are representative of two independent experiments . The absolute rates(i .e., rate above blank reactions) or incorporation in the absence of additions for ['"C]lysoPC,C16 :0, were 249 _ 71, 12 ± 2, and 47 ± 21 pmol/h/mg of protein of PC, PS/Pl, and PE,respectively, for PE, 1-C16 :0, 2-['"C]C20:4, 11 ± 1 and 7 ± 1 pmol/h/mg of protein of PCand PS/Pl, respectively, for PC, 1-C16:0, 2-['"C]C20 :4, 34 _ 3 and 8 ± 1 pmol/h/mg ofprotein of PE and PS/PI, respectively, and for PC, 1,2-['"CIC18 :1, 28 - 3 and 9 ± 7 pmol/h/mg of protein of PE and PS/PI, respectively .
1845
J. Neurochem., Vol . 63, No . 5, 1994
Labeled lipid Addition PC PS/PI PE
['"C]LysoPC, C16 :0 LysoPI 98 ± 28 90 ± 5 101 - 9LysoPS 103 ± 21 86 ± 5 100 ± 11LysoPE 81 ± 17 92 ± 14 548 ± 117
PE, 1-C16:0,2-['"C]C20 :4 LysoPC 3,470 ± 587 118 ± 41 NAPC, 1-C16 :0, 2-['"C]C20 :4 LysoPE NA 92 i 17 107 ± 25PC, 1,2-['"C]C18 :1 LysoPE NA 82 ± 33 84 ± 9
Lysophospholipid Acyl-CoAFatty acid incorporation
into phospholipids
None too 11M [ 14C]C20:4 CoA 0.8 ± 0 .110 mM ['"C]lysoPC, C16:0 None Not detectable10 mM lysoPC, C16:0 100 pM ['"C]C20 :4 CoA 87.0 ± 14 .210 mM ['"C]lysoPC, C16:0 100 pM C20:4 CoA 78.0 ± 2 .710 mM ['"C]lysoPC, C16:0 100 pM C16:0 CoA 25.7 ± 2 .81 mM lysoPC, C16:0 100 pM ['"C]C20:4 CoA 49.3 8 .61 trim lysoPC, C18 :1 100 pM ['"C]C20:4 CoA 25 .7 ± 5 .81 mM lysoPE, C16 :0 100 pM ['"C]C20:4 CoA 1 .6 ± 0 .61 mM IysoPI, C16:0/C18 :0 100 1.1M [' 4C]C20:4 CoA 111 ± 15 .71 rum lysops, C18:0 100 AM [ 14C]C20:4 CoA 5.0 1 .4

1846
ids present in our assay decreases the availability ofthe lysophospholipid substrate, thereby resulting in ahigher apparent Km value. It is interesting that humanbrain acyltransferase displayed a preference for bothacyl donors and acceptors . Thus, palmitoyl-CoA actedas a poorer acyl donor than arachidonyl-CoA, as hasbeen reported for the rat brain enzyme (Masuzawa etal ., 1989 ; Sun and MacQuarrie, 1989) . Furthermore,the preference exhibited by acyltransferase for lysoPIand lysoPC is in agreement with that reported by mostinvestigators for the rat brain enzyme both in vitro(Sun and MacQuarrie, 1989 ; Cassagne et al ., 1991 ;but see Masuzawa et al ., 1989) and in vivo (De Georgeet al ., 1989) . Such differences in the reacylation ratesof lysophospholipid types may have important conse-quences for the metabolic routes by which each speciesis eliminated from the membrane. For example, giventhat brain lysophospholipase appears to hydrolyze bothlysoPC and lysoPE at approximately the same rates(Leibovitz and Gatt, 1968), our results suggest thatlysophospholipase may play a greater role in the re-moval of lysoPE (a poor acyltransferase substrate)than it does for lysoPC (a good acyltransferase sub-strate) . This, of course, assumes that lysophospholi-pase and acyltransferase are present in same subcellu-lar compartments . Furthermore, the preference of acyl-transferase for lysoPI is mirrored by the substratepreference of human brain PLAZ for PI over PC andPE (unpublished data) . These observations suggestthat the turnover of PI is more rapid than for otherclasses of phospholipids, possibly reflecting the roleof PI in intracellular signaling .
TransacylationTwo types of putative transacylation activity were
detected in human brain, namely, transfer offatty acidsbetween lysophospholipids (lysophospholipid :lysophos-pholipid transacylase) and from phospholipids to lyso-phospholipids (phospholipid:lysophospholipid trans-acylase) . Due to the crude nature of the preparationsused, we cannot rule out that these reactions are cata-lyzed by multiple enzymes, perhaps involving the se-quential hydrolysis and reesterification of fatty acids .However, the likely requirement by such a pathwayfor ATP (to allow the formation of acyl-CoA interme-diates) makes such a possibility somewhat unlikely .Lysophospholipid:lysophospholipid transacylase ac-tivity has previously been identified in several tissuesincluding heart, gastric mucosa, and lung in which thetransacylation activity copurifies with a more activelysophospholipase (Vianen and van den Bosch, 1978 ;Lin et al ., 1979 ; van Heusden et al ., 1980 ; Gross andSobel, 1982 ; Gross, 1991 ; but see Sanjanwala et al .,1989) . Although it is unclear whether the lysophos-pholipase and transacylase activities of human brainare localized on the same polypeptide chain, the differ-ent pH optima and inhibitor sensitivities of each en-zyme make it unlikely that the two reactions are cata-
J. Neurochem., Vol. 63, No. 5, 1994
B. M. ROSS AND S. J. KISH
lyzed by the same active site . Nevertheless, myocardiallysophospholipid :lysophospholipid transacylase dis-plays similarities with that in human brain includingits pH optimum (Gross and Sobel, 1982) and stimula-tion by CoA (Sanjanwala et al ., 1989) . Moreover, thelysophospholipid :lysophospholipid transacylase activi-ties of rat and human brain possess a similar affinity forlysoPC and are both stimulated by CoA (Premkumar etal ., 1993), although the EC50 of CoA in rat brain (80W) is somewhat higher than that in human brain (71W) . The functional significance of CoA stimulationof lysophospholipid :lysophospholipid transacylase isunclear but may serve to couple acyltransferase activity(which releases CoA) to transacylation (Premkumaret al ., 1993) . Thus, the acyltransferase catalyzed esteri-fication of fatty acids to, for example, lysoPC mayactivate the transacylation of other lysophospholipidsnot readily acylated by this route (such as lysoPE) . Inthis regard we observed that human brain homogenatespossessed the ability to catalyze the transfer of fattyacids between lysoPC and lysoPE (Table 6) . Althoughthe rate of these reactions was much lower than thatobserved for acyltransferase, we must stress that theenzymes were not being assayed under identical reac-tion conditions . Indeed, using assay conditions differ-ent from those described herein, Premkumar et al .(1993) observed that the activity ofrat brain lysophos-pholipid:lysophospholipid transacylase was higherthan that of acyltransferase .
The transfer of fatty acids between phospholipidsand lysophospholipids has been the subject of limitedinvestigation in brain, although it has been studied inother systems (MacDonald and Sprecher, 1991) . Ma-suzawa et al . (1989) reported that transacylation oc-curred between the sn-2 position of PC and that oflysoPE in microsomes prepared from rat brain . How-ever, under the conditions used in the present investiga-tion, we did not observe any significant transfer ofradioactivity between either sn-1 (arachidonyl andoleoyl)- or sn-2-labeled (oleoyl) PC and lysoPE(Table 6) . In addition, we did not detect any transacy-lation between PC and lysoPS, as occurs in dog heart(Reddy and Schmid, 1987) . Nevertheless, humanbrain homogenates did possess the ability to catalyzethe transfer of radiolabeled arachidonic acid from thesn-2 position of PE to lysoPC . Such a mechanism,along with the other deacylation/reacylation enzymes,may be of significant importance in the generation ofphospholipid species that are not formed in the initialincorporation of fatty acids into membranes . For exam-ple, the administration of radiolabeled arachidonic acidto rats, although initially resulting in the assimilationof radioactivity into PI and PC, is followed by a slowrise in the amount of radiolabeled PE, suggesting aredistribution of arachidonic acid between phospho-lipid classes (De George et al., 1989) .
In summary, we have identified three enzymes inhuman brain capable of detoxifying the lysophospho-

LYSOPHOSPHOLIPID METABOLISM IN HUMAN BRAIN
lipids produced by the action of PLA . Our results sug-gest that lysophospholipase may be the major route bywhich lysophospholipids are removed from the cellmembrane in human brain. However, all three enzymesmay play an important role in the remodeling of mem-brane composition and thereby contribute to the overallfunctioning of membrane-associated processes . Our in-vestigation of lysophospholipid metabolism shouldalso prove useful for subsequent studies of the enzymesin pathological conditions of the human brain.
Acknowledgment : This study was supported by U.S . NIHgrant NS26043-06.
REFERENCESBlusztajn J . K., Gonzalez-Coviella I . L ., Logue M., Growdon J. H .,
and Wurtman R . J . (1990) Levels of phospholipid catabolicintermediates, glycerophosphocholine and glycerophosphoetha-nolamine, are elevated in brains of Alzheimer's disease but notof Down's syndrome patients . Brain Res. 536, 240-244 .
Cassagne C ., Juguelin H ., and Boiron F . (1991) Phospholipid acyla-tion by mouse sciatic nerve microsomes . Biochim. Biophys .Acta 1070, 119-126 .
De George J . J ., Noronha J. G ., Bell J ., Robinson P ., and RapoportS . I . (1989) Intravenous injection of [ 1-' °C] arachidonate toexamine regional brain lipid metabolism in anaesthetised rats .J . Neurosci. Res. 24, 413-423 .
De Jong J. G . N ., van den Bosch H ., Rijken D., and van DeenenL. L . M . (1974) Studies on lysophospholipases. III . The com-plete purification of two proteins with lysophospholipase activ-ity from beef liver . Biochim . Biophys. Acta 369, 50-63 .
Farooqui A. A ., Pendley C. E ., Taylor W. A ., and Horrocks L . A .(1985) Studies on diacylglycerol lipases and lysophospholi-pases of bovine brain, in Phospholipids in the Nervous System,Volume 2: Physiological Roles (Horrocks L . A � Kanfer J . N.,and Porcellati G ., eds), pp . 179-192 . Raven Press, New York .
Farooqui A . A., Liss L ., and Horrocks L . A . (1988) Stimulation oflipolytic enzymes in Alzheimer's disease . Ann. Neurol . 23,306-308 .
Flavahan N . A . (1993) Lysophosphatidylcholine modifies G protein-dependent signaling in porcine endothelial cells. Am. J. Physiol.264, H722-H727 .
Fujimoto T ., Nakano T ., Takano T., Hokazono Y ., Asakura T ., andTsuji T . (1992) Study of chronic schizophrenics using 31p mag-netic resonance chemical shift imaging. Acta Psychiatr. Scand.86, 455-462.
Gross R . W. (1991) Purification of lysophospholipase and lysophos-pholipase-transacylase from rabbit myocardium. Methods Enzy-mol. 197, 475-482 .
Gross R . W . and Sobel B . E . (1982) Lysophosphatidylcholine me-tabolism in the rabbit heart. Characterization of metabolic path-ways and partial purification of myocardial lysophospholipase-transacylase . J. Biol. Chem. 257, 6702-6708.
Higashi S ., Kobayashi T ., Kudo I., and Inoue K. (1988) Purificationand characterisation of lysophospholipase released from ratplatelets. J. Biochem. (Tokyo) 103, 442-447 .
Kish S . J ., Robitaille Y., Ball M ., Gilbert J ., Deck J . H. N ., ChangL . J ., and Schut L . (1990) Glycerophosphoethanolamine con-centration is elevated in brain of patients with dominantly-inher-ited olivopontocerebellar atrophy . Neurosci. Lett. 120, 209-211 .
Koski Y ., Jungalwala F . B ., and Kolodny E . H . (1978) Normalityof erythrocyte phospholipid in Duchenne muscular dystrophy .Clin . Chim. Acta 85, 295-298 .
Kramer R . M ., Pritzker C. R ., and Deykin D. (1984) Coenzyme A-mediated arachidonic acid transacylation in human platelets . J.Biol. Chem . 259, 2403-2406 .
1847
Leibovitz Z . and Gatt S . (1968) Isolation of lysophospholipase, freeof phospholipase activity, from rat brain . Biochim. Biophys.Acta 164, 439-441 .
Leibovitz-BenGershon Z., Kobiler I., and Gatt S . (1972) Lysophos-pholipases of rat brain . J. Biol. Chem. 247, 6840-6847 .
Lin Y . N ., Wassef M . K., and Horowitz M . I. (1979) Lysophospholi-pase and transacylase activities of rat gastric mucosa . Arch .Biochem. Biophys. 193, 213-220.
MacDonald J . 1. and Sprecher H . (1991) Phospholipid fatty acidremodeling in mammalian cells . Biochim . Biophys. Acta 1084,105-121 .
Masuzawa Y., Sugiura T., Sprecher H., and Waku K . (1989) Selec-tive acyl transfer in the reacylation of brain glycerophospholip-ids . Comparison of three acylation systems for 1-alk-1 '-enylglycero-3-phosphoethanolamine, 1-acylglycero-3-phosphoethano-lamine and 1-acylglycero-3-phosphocholine in rat brainmicrosomes. Biochim . Biophys. Acta 1005, 1-12 .
Metz S . A . (1986) Lysophosphatidylinositol, but not lysophospha-tidic acid, stimulates insulin release. Biochem . Biophys. Res.Commun. 138, 720-727 .
Metz S . A . (1987) Metabolism of lysophospholipids in intact ratislets . The insulin secretagogue p-hydroxymercuribenzoic acidimpairs lysophosphatidylcholine catabolism and permits its ac-cumulation . Biochem. J. 241, 863-869 .
Nitsch R . M ., Blusztajn J . K ., Pittas A . G ., Slack B . E., GrowdonJ. H ., and Wurtman R. J . (1992) Evidence for a membranedefect in Alzheimer's disease/ senile dementia of the Alzheimertype. Proc. Nall. Acad. Sci. USA 89, 1671-1675 .
Nitsch R . M ., Blusztajn J. K., Doyle F. M ., Robitaille Y ., WurtmanR . J ., Growdon J . H ., and Kish S . J . (1993) Phospholipid metab-olite levels are altered in cerebral cortex of patients with dominantly inherited olivopontocerebellar atrophy . Neurosci. Lett.161, 191-194 .
Oishi K ., Raynor R . L, Charp P . A ., and Kuo J . F . (1988) Regulationof protein kinase C by lysophospholipids . J. Biol . Chem . 260,7889-7895 .
Pettegrew J . W., Panchalingam K ., Moossy J ., Martinez J ., Rao G .,and Boller F. (1988) Correlation of phosphorus-31 magneticresonance spectroscopy and morphological findings in Alzhei-mer's disease. Arch. Neurol. 45, 1093-1096 .
Pettegrew J . W., Keshavan M . S ., Panchalingam K ., Strychor S .,Kaplan D . B ., Tretta M . G ., and Allen M . (1991) Alterationsin brain high-energy phosphate and membrane phospholipidmetabolism in first episode, drug-naive schizophrenics . A pilotstudy of the dorsal prefrontal cortex by in vivo phosphorus 31nuclear magenetic resonance spectroscopy . Arch. Gen . Psychia-try 48, 563-568 .
Premkumar N ., Sun G. Y ., and MacQuarrie R . A . (1993) Acylationof lysophosphatidylcholine by brain membranes . J. Neurosci.Res. 35, 321-326 .
Reddy P . V . and Schmid H . H . (1987) Acylation of dog heartlysophosphatidylserine by transacylase activity . Biochim . Bio-phys. Acta 922, 379-385 .
Rordorf G., Uemura Y ., and Bonventre J . V . (1991) Characterizationof phospholipase A Z (PLAZ ) activity in gerbil brain: enhancedactivities of cytosolic, mitochondrial, and microsomel formsafter ischemia and reperfusion . J. Neurosci. 11, 1829-1836 .
Rustenbeck I . and Lenzen S. (1992) Effect of lysophospholipids,arachidonic acid and other fatty acids on regulation of Ca"transport in permeabilised pancreatic islets . Cell Calcium 13,193-202 .
Sanjanwala M., Sun G. Y ., Cutrera M., and MacQuarrie R . A. (1989)Purification of lysophosphatidylcholine transacylase from bo-vine heart muscle microsomes and regulation of activity bylipids and coenzyme A . Biochim. Biophys . Acta 1006, 203-208 .
Shier W . T., Baldwin J . H ., Nilsen-Hamilton M., Hamilton R . T ., andThanassi N. M . (1976) Regulation of guanylate and adenylatecyclase activities by lysolecithin . Proc . Natl . Acad. Sci. USA73, 1586-1590.
J. Neurochem., Vol. 63, No. 5, 1994

1848
Smith C . D ., Gallenstein L. G . I., Layton W. J., Kryscios R . J ., andMarkesbery W . R . (1993) 3'P magnetic resonance spectroscopyin Alzheimer's and Pick's disease . Neurobiol. Aging 14, 85-92 .
Sun G . Y . and MacQuarrie R . A . (1989) Deacylation-reacylation ofarachidonyl groups in cerebral phospholipids . Ann . NY Acad.Sci. 559, 37-55 .
Sun G . Y ., Tang W., Huang S . F ., and MacQuarrie R. (1987) Lyso-phospholipase activity in rat brain subcellular fractions . Neuro-chem . Res. 12, 451-458 .
van den Bosch H . (1982) Phospholipases, in Phospholipids (Haw-thorne J . N . and Ansell G . B ., eds), pp . 313-358 . Elsevier,Amsterdam .
van den Bosch H ., Aarsman A . J ., De Jong J . G . N ., and van DeenenL . L . M . (1973) Studies on lysophospholipases . 1 . Purificationand some properties of a lysophospholipase from beef pancreas .Biochim . Biophys. Acta 296, 94-104.
van Heusden G . P. H., Vianen G . M ., and van den Bosch H . (1980)
J. Neurochem., Vol . 63, No . 5, 1994
B. M. ROSS AND S. J. KISH
Differentiation between acyl coenzyme A:lysophosphatidylcho-line acyltransferase and lysophosphatidylcholine :lysophosphati-dylcholine transacylase in the synthesis of dipalmitoylphosphati-dylcholine in rat lung . J. Biol. Chem . 255, 9312-9318 .
Vaswani K . K . and Ledeen R . W . (1989) Purified rat brain myelincontains measurable acyl-CoA:lysopholipid acyltransferase(s)but little, if any, glycerol-3-phosphate acyltransferase . J. Neuro-chem . 52, 69-74 .
Vianen G . M . and van den Bosch H . (1978) Lysophospholipase andlysophosphatidylcholine:lysophosphatidylcholine transacylasefrom rat lung : evidence for a single enzyme and some aspectsof its properties . Arch . Biochem. Biophys . 190, 373-384 .
Weltzien H . U. (1979) Cytolytic and membrane-perturbing proper-ties of lysophosphatidylcholine . Biochim. Biophys . Acta 559,259-287 .
Wiley M., Przetakiewicz M., Takahashi M., and Lowenstein J . M .(1992) An extended method for separating and quantitatingmolecular species of phospholipids . Lipids 27, 295-301 .





























