Embed Size (px)
Citation preview

Ecotoxicology 5, 9-21 (1996)
Changes in uptake phytotoxicity of cadmium in J A S W A N T S I N G H l, P.N. V I S W A N A T H A N l, P O O N A M G U P T A 2. and S A N T H A DEVI 2
JEcotoxicology Section, Industrial Toxicology Research Centre, P Box-80, M. G. Marg, Lucknow-226001, India :Electron Microscopy Section, National Botanical Research Institute, Lucknow, India
of calcium caused by Salvinia molesta
The effect of Ca on the uptake of Cd by root and leaves of Salvinia molesta was investigated at different time intervals and under different photoperiods. For detailed study on uptake and interaction, 45Ca and 1°9Cd were used and it was found that there was a higher uptake of 45Ca in the root and leaves at 48 h and a concurrent reduction in l°°Cd content at 48 h suggesting alterations in Ca functions due to the phytotoxicity of Cd. Ultrastructural changes due to cadmium toxicity included swirling of thylakoid membranes of the chloroplasts as well as detachment of the tips of trichomes from the leaf.
Keywords: cadmium; calcium; phyotoxicity; Salvinia molesta; uptake
Introduction
The phytotoxic effects of environmental pollution by cadmium have been well documented (Barcelo and Poschenrieder 1990, Chawla et al., 1991, Rasico et al., 1993). Van Balen et al. (1980) and Sela et al. (1990) reported uptake and interactions of essential trace metals with toxic metals. Rauser and Ackerley (1987) reported electron- dense granules containing Cd and variable proportions of Ca, Fe, Ni, Pb and Zn. Calcium fluxes and functions could be a major site of toxic mechanisms especially in oxidative stress and cytotoxicity (Orrenius et al. 1989). Cadmium is known to interfere with Ca channel dynamics and reserves (Viarengo and Nicotera 1991). Khan et al. (1984) suggested that calcium provides protective mechanisms against Cd toxicity and X-ray microanalytical studies reveal the formation of co-precipitates with Cd and phosphate and immobilization of soluble Cd in the cells. In view of the above, it was studied whether the uptake and translocation of Cd are influenced by Ca at different time intervals and photoperiods in aquatic plant Salvinia molesta, along with ultrastructural changes caused by Cd exposure.
Materials and methods
Plants of S. molesta (Mitchell) were collected from stagnant ponds of Quilon (Kerala, India) and maintained in artificial ponds at the National Botanical Research Institute, Lucknow. Before the start of the experiment plants were moved in an incubator and
*To whom correspondence should be addressed.
0963 9292 ~'~ L, 1996 Chapman & Hall

10 Singh, Viswanathan, Gupta, and Devi
maintained in 3% Hoagland's solution (EPA 1975) under standard physiological con- ditions at 25 2 0.5 BE, with a 16 h light and 8 h dark photoperiod of 200 ~molm -2 s -1 fluorescent light (Philips). The culture medium was supplemented with CdC12.H20 (Loba- Chemie Indoaustranal Co.) to obtain a 0.1 mg1-1 Cd concentration (Gupta and Devi 1992). The cultures were run in triplicates and the plants were harvested after 16 h (light period), 24 h (dark period), 40 h (light period) and 48 h (dark period), washed with double-distilled water, the root and leaf portions were separated, blotted dry and fresh weights were recorded. Cadmium-free controls were also studied for comparisons with the treated set of experiments. For the whole experiment plants were grown and maintained as described above unless otherwise specified.
Estimation of metals
Weighed amounts of dried material were digested using sulphuric and nitric acid mixture and the solution was brought to a known volume with deionized water. The Cd content was determined on a Perkin-Elmer atomic absorption spectrophotometer model AA 303, using a cadmium lamp at a wavelength of 228.8 nm. For calcium estimation, lanthanum nitrate was added to the samples and the Ca content was measured at 422.7 nm wavelength using a calcium lamp.
Radioactive studies
45Ca uptake was done in Cd-treated and control plants by incorporating 15 ~Ci of 45Ca (specific activity 1 mCi, BARC, Bombay, India).
The incorporation of 1°9Cd was studied by using 0.1 ~Ci 1°9Cd (specific activity 1 mCiug -1 Cd, Radiochemical center, Amersham, UK). The radioactive Ca and Cd were mixed in 750 ml of 3% Hoagland's medium containing 0.1 mg 1 -a Cd only. Plants were harvested after 16, 24, 40 and 48 h intervals and the root and leaf portions were separated and washed with double-distilled water, using 10% homogenate and after standard processing, 45Ca radioactivity was measured on a LKB Rack Beta counter Model 1215/1216. Gamma radiations of 1°9Cd, in each sample were recorded on an LKB ultrogamma counter for 1 min. Corrections were made for background and natural decay of isotopes during the experiment. The student 't'-test described by Fisher (1950) was employed to calculate the statistical significance between the control and experimental values.
Electron microscopic studies
For transmission of electron microscopy, the leaf and root parts of treated (0.1 mg 1-1 Cd) and control plants were pre-fixed in 2.5% glutaraldehyde, buffered in sodium cacodylate for 24 h, post-fixed in 1% osmium tetraoxide (24 h) and embedded in eponaraldite (Hayat 1981). Sections were cut on an LKB ultramicrotome using glass knives, stained with uranyl acetate and lead citrate and examined under a Philips-400 TEM.
For scanning electron microscopy (SEM) samples were fixed in 2.5% glutaraldehyde, washed with 0.1 M cacodylate buffer (pH 7.2) dehydrated in ethanol series, critical point dried through CO2 mounted on stubs and coated with gold, before observation on a JEOL 35C scanning electron microscope.
Cadmium calcium interaction in phytotoxicity 11
Results
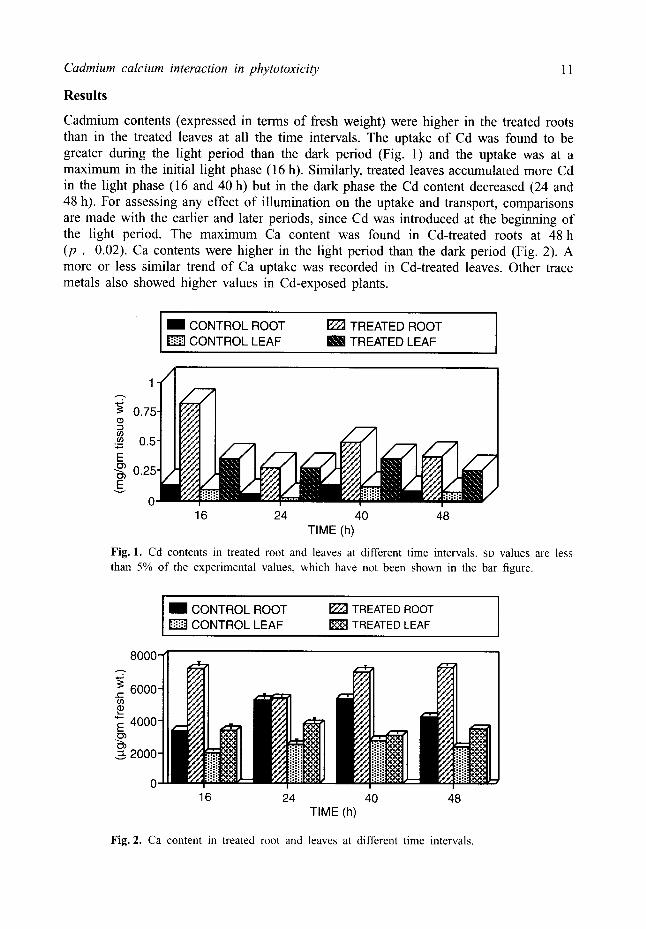
Cadmium contents (expressed in terms of fresh weight) were higher in the treated roots than in the treated leaves at all the time intervals. The uptake of Cd was found to be greater during the light period than the dark period (Fig. 1) and the uptake was at a maximum in the initial light phase (16 h). Similarly, treated leaves accumulated more Cd in the light phase (16 and 40 h) but in the dark phase the Cd content decreased (24 and 48 h). For assessing any effect of illumination on the uptake and transport, comparisons are made with the earlier and later periods, since Cd was introduced at the beginning of the light period. The maximum Ca content was found in Cd-treated roots at 48 h (p , 0.02). Ca contents were higher in the light period than the dark period (Fig. 2). A more or less similar trend of Ca uptake was recorded in Cd-treated leaves. Other trace metals also showed higher values in Cd-exposed plants.
I I CONTROL ROOT CONTROL LEAF
TREATED ROOT mini TREATED LEAF
/ 1
,--.?.
o.5.
0.25" i /
0- 16 24 40 48
TIME (h)
Fig. 1. Cd contents in treated root and leaves at different time intervals. SD values are less than 5% of the experimental values, which have not been shown in the bar figure.
l 1 CONTROL ROOT F22J TREATED ROOT ] D CONTROL LEAF ~ TREATED LEAF
8000-
6000-
4000-
~2000-
i I i
16 24 40 TIME (h)
I
48
Fig. 2. Ca content in treated root and leaves at different time intervals.
12 Singh, Viswanathan, Gupta, and Devi
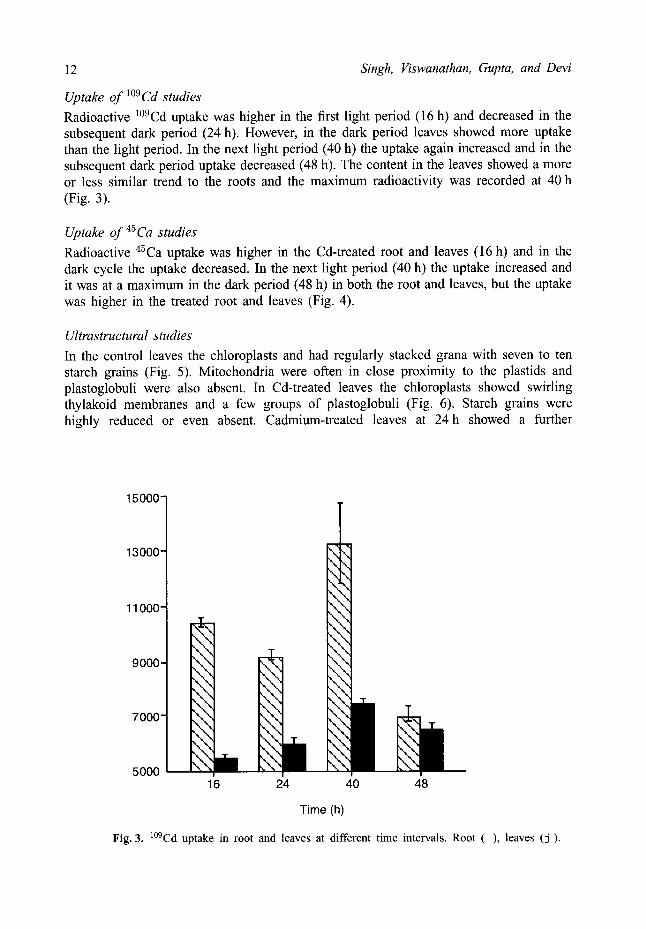
Uptake of 1°9Cd studies Radioactive l°9Cd uptake was higher in the first light period (16 h) and decreased in the subsequent dark period (24 h). However, in the dark period leaves showed more uptake than the light period. In the next light period (40 h) the uptake again increased and in the subsequent dark period uptake decreased (48 h). The content in the leaves showed a more or less similar trend to the roots and the maximum radioactivity was recorded at 40 h (Fig. 3).
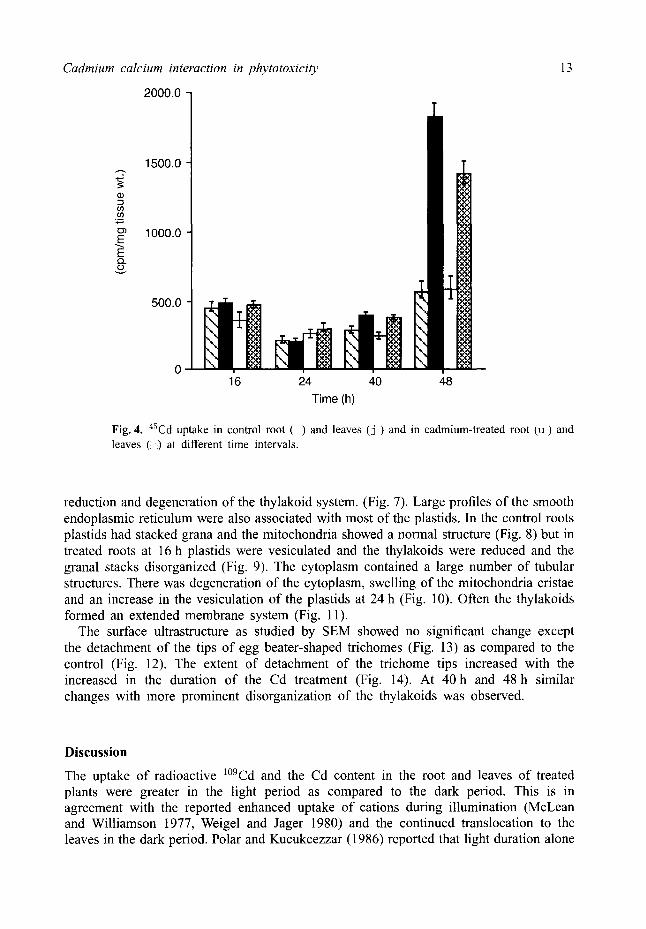
Uptake of 45Ca studies Radioactive 45Ca uptake was higher in the Cd-treated root and leaves (16 h) and in the dark cycle the uptake decreased. In the next light period (40 h) the uptake increased and it was at a maximum in the dark period (48 h) in both the root and leaves, but the uptake was higher in the treated root and leaves (Fig. 4).
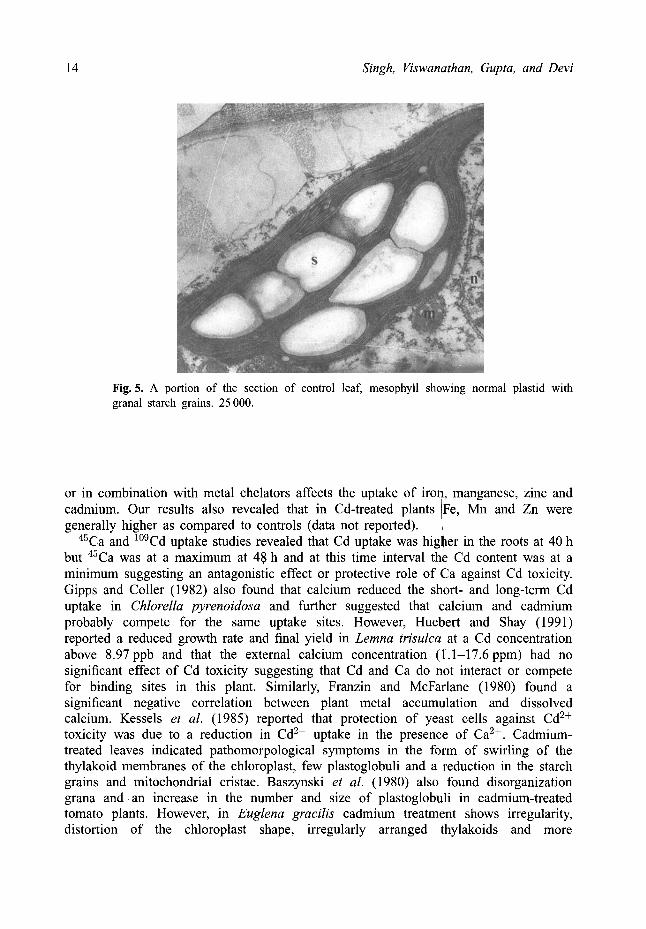
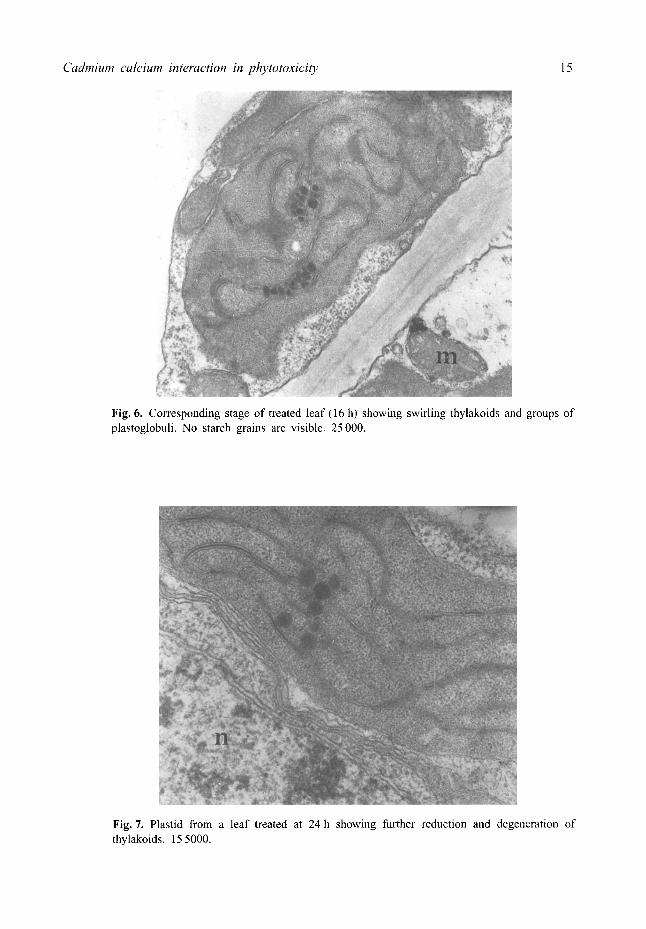
Ultrastructural studies In the control leaves the chloroplasts and had regularly stacked grana with seven to ten starch grains (Fig. 5). Mitochondria were often in close proximity to the plastids and plastoglobuli were also absent. In Cd-treated leaves the chloroplasts showed swirling thylakoid membranes and a few groups of plastoglobuli (Fig. 6). Starch grains were highly reduced or even absent. Cadmium-treated leaves at 24 h showed a further
15000-
13000-
11000-
9000-
7000-
5000
%_%_ %_%_ ...........
%_'..._ . . % \
"....%- "..._ \
\ \ \ \ ".... \ '.... \
16 24 40 48
Time (h)
Fig. 3. l°9Cd uptake in root and leaves at different time intervals. Root ( ) , leaves (j).
Cadmium calcium interaction in phytotoxicity
2000.0 7
13
1500.0
if) .,..,
1000.0 E
Q .
500.0
16 24 40 48
Time (h)
Fig. 4. 45Cd uptake in control root ( ) and leaves ( j ) and in cadmium-treated root (u) and leaves ( ) at different time intervals.
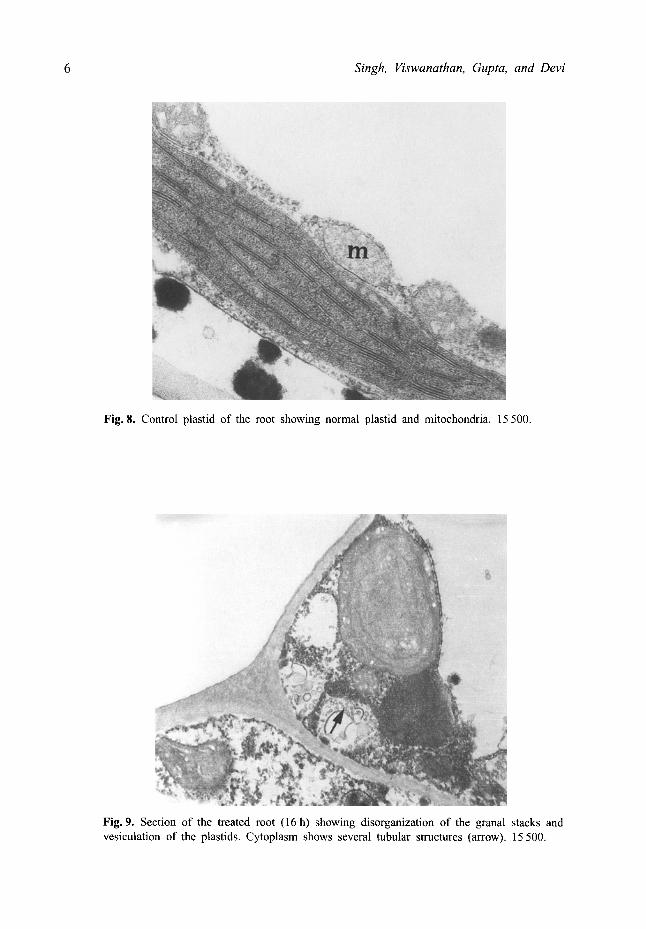
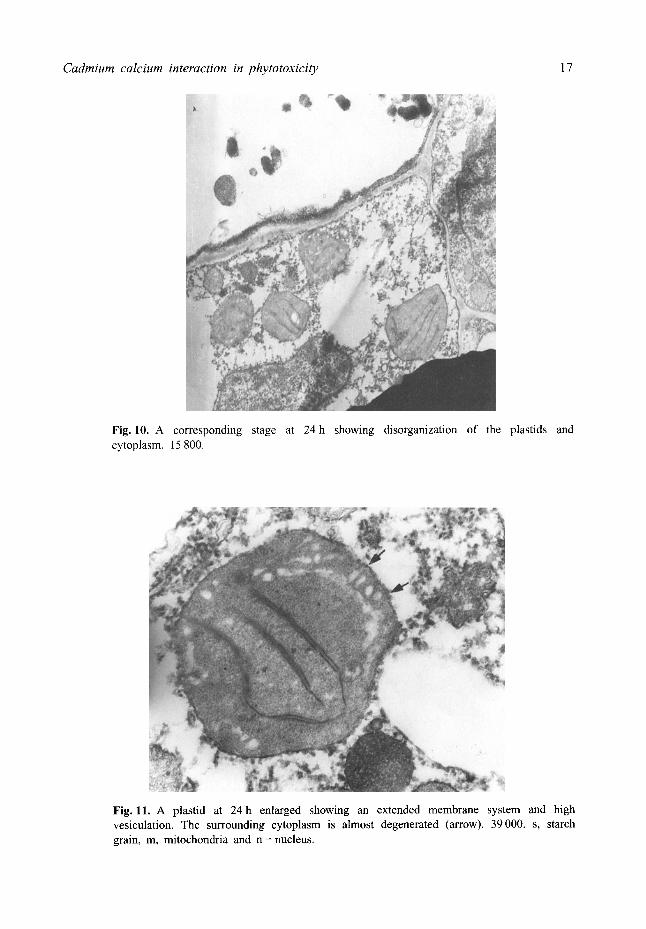
reduction and degeneration of the thylakoid system. (Fig. 7). Large profiles of the smooth endoplasmic reticulum were also associated with most of the plastids. In the control roots plastids had stacked grana and the mitochondria showed a normal structure (Fig. 8) but in treated roots at 16 h plastids were vesiculated and the thylakoids were reduced and the granal stacks disorganized (Fig. 9). The cytoplasm contained a large number of tubular structures. There was degeneration of the cytoplasm, swelling of the mitochondria cristae and an increase in the vesiculation of the plastids at 24 h (Fig. 10). Often the thylakoids formed an extended membrane system (Fig. 11).
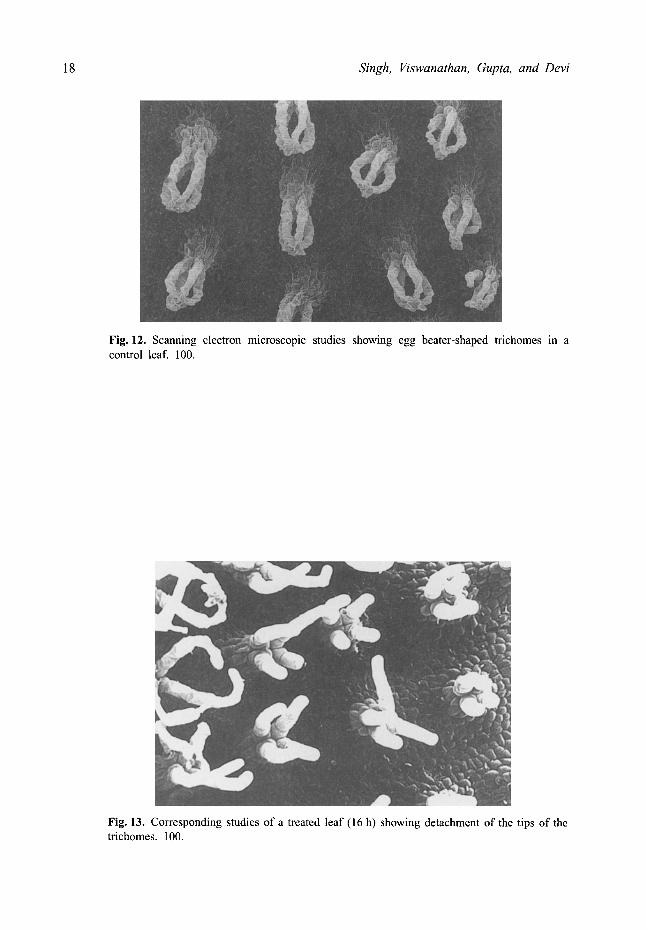
The surface ultrastructure as studied by SEM showed no significant change except the detachment of the tips of egg beater-shaped trichomes (Fig. 13) as compared to the control (Fig. 12). The extent of detachment of the trichome tips increased with the increased in the duration of the Cd treatment (Fig. 14). At 40 h and 48 h similar changes with more prominent disorganization of the thylakoids was observed.
Discussion
The uptake of radioactive 1°9Cd and the Cd content in the root and leaves of treated plants were greater in the light period as compared to the dark period. This is in agreement with the reported enhanced uptake of cations during illumination (McLean and Williamson 1977, Weigel and Jager 1980) and the continued translocation to the leaves in the dark period. Polar and Kucukcezzar (1986) reported that light duration alone
14 Singh, Viswanathan, Gupta, and Devi
Fig. 5. A portion of the section of control leaf, mesophyll showing normal plastid with granal starch grains. 25 000.
or in combination with metal chelators affects the uptake of iroq, manganese, zinc and cadmium. Our results also revealed that in Cd-treated plants I Fe, Mn and Zn were generally higher as compared to controls (data not reported).
45Ca and l°9Cd uptake studies revealed that Cd uptake was higher in the roots at 40 h but 45Ca was at a maximum at 48 h and at this time interval the Cd content was at a minimum suggesting an antagonistic effect or protective role of Ca against Cd toxicity. Gipps and Coller (1982) also found that calcium reduced the short- and long-term Cd uptake in Chlorella pyrenoidosa and further suggested that Calcium and cadmium probably compete for the same uptake sites. However, Huebert and Shay (1991) reported a reduced growth rate and final yield in Lemna trisulca at a Cd concentration above 8.97ppb and that the external calcium concentration (1.1-17.6ppm) had no significant effect of Cd toxicity suggesting that Cd and Ca do not interact or compete for binding sites in this plant. Similarly, Franzin and McFarlane (1980) found a significant negative correlation between plant metal accumulation and dissolved calcium. Kessels et al. (1985) reported that protection of yeast cells against Cd 2+ toxicity was due to a reduction in Cd 2+ uptake in the presence of Ca 2+. Cadmium- treated leaves indicated pathomorpological symptoms in the form of swirling of the thylakoid membranes of the chloroplast, few plastoglobuli and a reduction in the starch grains and mitochondrial cristae. Baszynski et al. (1980) also found disorganization grana and an increase in the number and size of plastoglobuli in cadmium-treated tomato plants. However, in Euglena gracilis cadmium treatment shows irregularity, distortion of the chloroplast shape, irregularly arranged thylakoids and more
Cadmium calcium interaction in phytotoxicity 15
Fig. 6. Corresponding stage of treated leaf (16 h) showing swirling thylakoids and groups of plastoglobuli. No starch grains are visible. 25 000.
Fig. 7. Plastid from a leaf treated at 24 h showing further reduction and degeneration of thylakoids. 15 5000.
6 Singh, Viswanathan, Gupta, and Devi
Fig. 8. Control plastid of the root showing normal plastid and mitochondria, 15 500.
Fig. 9. Section of the treated root (16 h) showing disorganization of the granal stacks and vesiculation of the plastids, Cytoplasm shows several tubular structures (arrow). 15 500.
Cadmium calcium interaction in phytotoxicity 17
Fig. 10. A corresponding stage at 24h showing disorganization of the plastids and cytoplasm. 15 800.
Fig. 11. A plastid at 24 h enlarged showing an extended membrane system and high vesiculation. The surrounding cytoplasm is almost degenerated (arrow). 39000. s, starch grain, m, mitochondria and n = nucleus.
18 Singh, Viswanathan, Gupta, and Devi
Fig. 12. Scanning electron microscopic studies showing egg beater-shaped trichomes in a control leaf. 100.
Fig. 13. Corresponding studies of a treated leaf (16 h) showing detachment of the tips of the trichomes. 100.

Cadmium calcium interaction in phytotoxicity 19
Fig. 14. Further detachment of the tips of the trichomes of a treated leaf (24 h), leaving only the stubs. 100.
plastoglobuli as compared to the control (Duret et al. 1986). Since many aspects of ultrastructural and functional organization are regulated by Ca, interference by Cd (Kauss 1987) in Ca functions could lead to such effects. Ghoshroy and Nadaka vukaren (1990) revealed a time-dose effect of Cd on chloroplast development in soyabean. Inhibition by Cd (Wu and Lorenzen 1984), chloroplast development and photosynthesis (Stobart et al. 1985) support this. Vazquez et al. (1992), observed that many plastids from Cd-treated bean plants are enlarged, indicating an inhibition of plastid divisions and exhibit abundant electron-dense deposits resembling phytoferritin. Sela et al. (1990), reported the formation of precipitates of Cd with phosphate and calcium in the xylem cells of the shoot bundle of Cd-treated Azolla roots along with an increase in the phosphate content. The present results indicate that interaction with the uptake and functions of trace elements especially Ca could be a major locus of phytotoxicity of cadmium. Whether the higher uptake is due to Cd-induced damage to membranes and whether higher intracellular Ca levels could lead to cytotoxicity is not clear.
Acknowledgements
Thanks are due to Dr R.C. Srimal, Director of the Industrial Toxicology Research Centre and Dr P.V. Sane, Director of the National Botanical Research Institute, Lucknow for their interest in this work. Financial support to J.S. by the Council of Scientific and Industrial Research (CSIR) and to RG. by the Department of Science and Technology, N. Delhi, is gratefully acknowledged.

20 Singh, Viswanathan, Gupta, and Devi
References
Barcelo, J. and Poschenrieder, C.H. (1990) Plant water relations as affected by heavy metal stress: a review. J Plant Nutr. 13, 1-37.
Baszynski, T., Wajda, L., Krol, M., Wolinska, D., Krupa, Z. and Tukendrof, A. (1980) Photosynthesis activities of cadmium treated tomato plants. Physiol. Plant. 48, 365-70.
Chawla, G., Singh, J. and Viswanathan, EN. (1991) Effect of pH and temperature on the uptake of cadmium by L. minor. Bull. Environ. Contain. Toxicol. 47, 84-90.
Duret, S., Bonaly, J., Bariaud, A., Vannereau, A. and Mestre, J.C. (1986) Cadmium induced ultrastructural changes in Euglena cells. Environ. Res. 39, 96-193.
EPA (1975) Recommended test protocols: approach for evaluation of toxicity to aquatic vascular plants. In: Rubinstein, R. et al. (eds) Test methods for assessing the effects of chemicals" on plants. Washington DC: EPA-560-5-75-008, final report, US Environmental Protection Agency, pp. 3-117, 3-120.
Fisher, R.A. (1950) Statistical Methods for Research Work, 1 lth edition. Edinburgh: 0liver and Boyd. Franzin, W.G. and McFarlane, G.A. (1980) An analysis of the aquatic macrophyte Myriophyllum
exalbescens as an indicator of the metal contamination of aquatic ecosystem near a base metal smelter. Bull. Environ. Contam. Toxieol. 24, 597-605.
Ghoshroy, S. and Nadakavukaren, M.J. (1990) Influence of cadmium on the ultrastructure of developing chloroplasts in soyabean and corn. Exp. Environ. Bot. 30, 187 92.
Gipps, J.E and Coller, B.A.W. (1982) Effect of some nutrient cations on uptake of cadmium by Chlorella pyrenoidosa. Aust. J. Mar. Freshwater Res. 33, 979-87.
Gupta, M. and Devi, S. (1992) Cadmium sensitivity inducing structural responses in Salvinia molesta Mitchell Bull. Environ. Contam. Toxicol. 49, 43643.
Hayat, M.A. (1981) Principles and Techniques of Electron Microscopy, Vol. I. New York: Van Nostrand-Reinhold.
Huebert, D.B. and Shay J.M. (1991) The effect of cadmium and its interaction with external calcium in submerged aquatic macrophyte Lemna trisulca L. Aquat. Toxicol. 20, 57-71.
Kauss, H. (1987) Some aspects of calcium dependent regulation in plant metabolism. Ann. Rev. Plant Physiol. 38, 47-72.
Kessels, B.G.E, Belde, P.J.M. and Borst-Pauwells, G.W.EH. (1985) Protection of Saceharomyces cerevisiae against Cd 2+ toxicity by Ca 2+. J. Gen. Microbiol. 131, 2533-7.
Khan, D.H., Duckett, J.G., Fankland, B. and Kirkhan, J.B. (1984) An X-ray microanalytical study of the distribution of cadmium in roots of Zea mays L. J. Plant Physiol 115, 19-28.
McLean, M.W. and Williamson, EB. (1977) Cadmium accumulation by the marine red alga Porphyra umbilicalis. Physiol. Plant. 41, 268-72.
Orrenius, S., McConkey, D.J., Bellomo, G. and Nicotera, P. (1989) Role of the Ca 2+ in toxic cell killing. Trends" Pharmacol. Sci. 10, 281-5.
Polar, E. and Kucukcezzar, R. (1986) Influence of some metal chelators and light regimes on bioaccumulation and toxicity of Cd in duckweed (Lemna gibba). Physiol. Plant. 66, 87-93.
Rasico, N., Veccha, ED., Ferretti, M., Merlo, L. and Ghisi, R. (1993) Some effects of cadmium on maize plants. Arch. Environ. Contam. Toxicol 25, 244-9.
Rauser, W.E. and Ackerley, C.A. (1987) Localization of cadmium in granules within differentiating and mature root cells. Can J Bot. 65, 643 6.
Sela, M., Fritz, E., Huttermann, A. and Tel-Or, E. (1990) Studies on cadmium localization in the water fern Azolla. Physiol. Plant. 79, 547-53.
Stobart, A.R., Griffiths, WT., Bukhari, J.A. and Shewood, R.P. (1985) The effect of the Cd 2+ on the biosynthesis of chlorophyll in leaves of barley. Physiol. Plant. 63, 293-8.
Van Balen, E., Van DeGeijn, S.C. and Desmet, G.M. (1980) Autoradiography evidence for the incorporation of cadmium into calcium oxalate crystals. Zeitschriftfur Pflanzen Physiol. 97, 123- 33.

Cadmium calcium interaction in phytotoxicity 21
Vazquez, M.D., Poschenrieder, C.H. and Barcelo, J. (1992) Ultrastructural effects and localization of low cadmium concentration in bean roots. New Phytol. 120, 215 26.
Viarengo, A. and Nicotera, 13. (1991) Possible role of Ca 2+ in heavy metal cytotoxicity. Comp. Biochem. Physiol. 1000, 81-4.
Weigel, H.J. and Jager, H.J. (1980) Subcellular distribution and chemical form of cadmium in bean plants. Plant Physiol. Lancaster 65, 480 2.
Wu, J.T. and Lorenzen, H. (1984) Effect of copper on photosynthesis in synchronous Chlorella cells. Bot. Bull. Academia Sinica 25, 125-32.





























