Embed Size (px)
Citation preview

Cell-free fusion of bacteria-containing phagosomeswith endocytic compartmentsUlrike Becken, Andreas Jeschke, Katharina Veltman1, and Albert Haas2
Cell Biology Institute, University of Bonn, Ulrich-Haberland-Strasse 61a, 53121 Bonn, Germany
Edited by Reinhard Jahn, Max Planck Institute for Biophysical Chemistry, Goettingen, Germany, and accepted by the Editorial Board October 4, 2010 (receivedfor review June 2, 2010)
Uptake of microorganisms by professional phagocytic cells leads toformation of a new subcellular compartment, the phagosome,which matures by sequential fusion with early and late endocyticcompartments, resulting in oxidative and nonoxidative killing ofthe enclosed microbe. Few tools are available to study membranefusion between phagocytic and late endocytic compartments ingeneral and with pathogen-containing phagosomes in particular.We have developed and applied a fluorescence microscopy assayto study fusion of microbe-containing phagosomes with different-aged endocytic compartments in vitro. This revealed that fusionof phagosomes containing nonpathogenic Escherichia coli withlysosomes requires Rab7 and SNARE proteins but not organelleacidification. In vitro fusion experiments with phagosomes con-taining pathogenic Salmonella enterica serovar Typhimuriumindicated that reduced fusion of these phagosomes with earlyand late endocytic compartments was independent of endosomeand cytosol sources and, hence, a consequence of altered phago-some quality.
latex beads ∣ phagocytosis ∣ macrophage ∣ phagolysosome ∣ vacuole
Phagocytosis is the receptor-mediated ingestion of largeparticles, usually by professional phagocytes, i.e., neutrophils,
dendritic cells, monocytes, or macrophages, resulting in forma-tion of a new cytoplasmic compartment, the phagosome (1).
The development of a phagosome into a fully degradativephagolysosome is temporally and spatially ordered in that thenewly formed “early” phagosomematures vectorially into a “late”phagosome after fusion with late endosomes and finally into aphagolysosome by fusion with lysosomes (1). This process is ac-companied by loss of early endocytic markers [e.g., transferrinreceptor (TfR), Rab5] and acquisition of late endocytic markers[e.g., lysosome associated membrane protein 1 (LAMP1), cathe-psin D, Rab7] (1, 2). Fusion yield (fusion events per time periodper organelle) generally decreases with endosome or phagosomeage (2, 3). Rab GTPases and SNARE proteins are key regulatorsof all vesicle fusion events in the endocytic pathway. Rab proteins,in concert with tethering factors, specifically bridge membranesto be fused (4) with Rab5 supporting early (3, 5) and Rab7supporting late (6) fusion events, but additional Rabs may beinvolved (7). SNARE proteins on both partner membranesare downstream effectors of Rab proteins and drive membranefusion by formation of intermembrane quarternary SNARE com-plexes (8) composed of one R-SNARE helix from one partnermembrane and Qa-, Qb-, and Qc-SNARE helices from the other.Following membrane fusion, quarternary SNARE complexesare disassembled under ATP hydrolysis by the N-ethylmaleimidesensitive factor (NSF) reforming activated SNAREs (8).
Several intracellular pathogens interfere with maturation oftheir phagosomes into fully degradative compartments, e.g., bydisconnection of the phagosome from the endocytic pathway(e.g., Legionella pneumophilae) or by the arrest of phagosomema-turation at a prelysosome stage (e.g.,Mycobacterium tuberculosis)(1). Macrophage phagosomes containing Salmonella enterica(SCP) acidify (9) and acquire LAMP1 but exclude mannose6-phosphate receptor (10). Whereas several groups observed re-
duced fusion of SCPs with lysosomes compared to phagosomescontaining inert particles (10–12), others did not (13). Fusogeni-city of SCPs with other endocytic organelles in macrophages hasbeen investigated little (10).
To truly understand the (dys)function of phagosome matura-tion at a molecular level, the fusion of endocytic with microbe-containing phagocytic organelles ought to be investigated inreconstituted systems, yet most published assays used bead-containing but not microbe-containing phagosomes (3, 5) andwere limited to fusion of early phagosomes with early endocyticorganelles (14, 15). Fusion of lysosomes with microbe-containingphagosomes has been reconstituted only in permeabilized macro-phages (16). Here, we present a generally applicable in vitroassay to quantify fusion of phagosomes containing pathogenicor harmless microorganisms or inert particles with endocyticorganelles of different maturation stages.
ResultsMaturation of Phagosomes Containing Heat-Killed Escherichia coliDH5α or Latex Beads in Vivo. Kinetic analysis of phagolysosomeformation in J774E macrophages using fluorescently labeled,heat-killed, IgG-opsonized E. coli showed that colocalization ofbacteria with preloaded lysosomal dye was almost quantitativeafter 30 min at 37 °C (Fig. S1 and SI Text). Therefore “30-minphagosomes” (phagolysosomes) were used in fusion experiments.Phagosomes containing IgG-coupled latex beads prepared after30-min infection and 60-min chase were enriched in LAMP1 butnot TfR (Fig. S2A). At this time point 84% (SD� 4, n ¼ 3) ofbeads colocalized with preloaded lysosomal dye.
Preparation of Endocytic Vesicles. Using pulse/chase protocols forferrofluid (10-nm paramagnetic particles) (17), different-agedendocytic compartments were purified from postnuclear superna-tants (PNSs) and analyzed biochemically (Fig. 1 and SI Text).Compared with PNSs, 100∕00 (pulse/chase) endosomes werestrongly enriched in early endocytic TfR and Syntaxin 13 (Stx13),yet contained little late endocytic LAMP1 or mature cathepsin D.Both Rab5 and Rab7 were enriched and, on a single-organellelevel, 50% of endosomes were positive for both Rabs (Fig. S3and SI Text). 100∕150 endosomes were also enriched in TfRand Stx13, but showed more late endosome characteristics, suchas loss of Rab5, with only modest enrichment of lysosomal BSArhodamine and lysosomal acid β-galactosidase activity. Lyso-somes (50∕1200) were strongly enriched in LAMP1, cathepsinD, acid β-galactosidase activity, and BSA rhodamine (Fig. 1 Aand B) and lacked early endocytic markers.
Author contributions: U.B. and A.H. designed research; U.B., A.J., and K.V. performedresearch; U.B. and A.H. analyzed data; and U.B. and A.H. wrote the paper.
The authors declare no conflict of interest.
This article is a PNAS Direct Submission. R.J. is a guest editor invited by the Editorial Board.1Present address: Institute for Infectiology, ZMBE, University of Münster,Von-Esmarch-Strasse 56, 48149 Muenster, Germany.
2To whom correspondence should be addressed. E-mail: [email protected].
This article contains supporting information online at www.pnas.org/lookup/suppl/doi:10.1073/pnas.1007295107/-/DCSupplemental.
20726–20731 ∣ PNAS ∣ November 30, 2010 ∣ vol. 107 ∣ no. 48 www.pnas.org/cgi/doi/10.1073/pnas.1007295107

Characterization of Phagosome–Lysosome Fusion in Vitro. Cell-freefusion of E. coli-containing phagolysosomes (ECPs) with lyso-somes (Fig. 2) progressed quickly and reached a plateau afterapproximately 40 min (Fig. 3A). In vitro fusion required physio-logical temperature and ATP, it was partly dependent on cytosol,was inhibited by the fast Ca2þ chelator 1,2-bis(2-aminophenoxy)ethane-N′,N′,N′,N′-tetraacetate (BAPTA), the NSF inhibitorN-ethylmaleimide (NEM), and by recombinant Rab guanosinenucleotide dissociation inhibitor (RabGDI), which can extractRab(GDP) proteins from membranes (Fig. 3B). Fusion wassensitive to the calmodulin inhibitors trifluoperazine and W7,but not to W5, which has lower affinity for calmodulin (Fig. S4).These data agree with results from other in vitro fusion reactions
(18–23). Neither bafilomycin A1, a specific inhibitor of thevATPase, nor nigericin, a Kþ∕Hþ antiporter and proton gradientuncoupler, inhibited ECP-lysosome fusion in vitro, althougheither drug effectively collapsed lysosome pH gradients in vitro(SI Text and Fig. S5). Nocodazole inhibited ECP-lysosome fusionin vitro to a minor degree (Fig. 3B).
In vitro fusion of pure phagosomes containing inert latex beads(LBPs) with lysosomes (Fig. 3 and Fig. S2) proceeded with thesame kinetics as ECP-lysosome fusion (Fig. 3A), was also depen-dent on physiological temperature and ATP, and was inhibited byNEM and RabGDI (Fig. 3C).
To directly test whether membrane fusion and not possiblymembrane attachment was quantified, we performed a micro-scopic Foerster resonance energy transfer (FRET) analysis,which yields a signal only after vesicle content mixing but notduring vesicle attachment (24, 25). In samples containing anATP-depleting system or an inhibitory soluble Q-SNAREmixture (see below), percentages of bacteria with meanFRET intensities > 0 and percentages of colocalization were re-duced by the same degree (Fig. 3D), although membrane attach-ment should have occurred normally (26).
Fig. 1. Preparation of endocytic vesicles. Macrophages were incubatedwith ferrofluid and magnetic compartments were isolated from a PNS.(A) Western blot analysis of PNS and purified endocytic compartments. Fiftyor 10 μg protein of PNS, 10 μg protein for each purified fraction were added.Pulse/chase times for ferrofluid are indicated. Arrowheads indicate the posi-tions of cathepsin D (Cat D) precursor (p), intermediate (i), and mature (m)forms. (B) Quantification of specific lysosomal acid β-galactosidase activities(U∕mg) and of fluorescence (FU) of lysosomal BSA rhodamine. Shown aremeans and standard deviations of three independent experiments.
Fig. 2. Assay for cell-free phagosome–endosome fusion. (A) Schematic re-presentation of the cell-free phagosome–endosome fusion assay. See textfor details. (B) Micrograph showing a representative image section of anin vitro ECP-lysosome fusion reaction prepared for fluorescence microscopy.In the overlay, phagosomes containing E. coli are shown in green, lysosomesin red. Enlargements show bacteria not colocalizing with lysosomal fluor(Upper) or colocalizing with lysosomal fluor (Lower). (Right) False color-coded (open arrowhead) images of FRET intensity for each pixel (black:low intensity; white: high intensity). Bar: 2 μM.
Fig. 3. In vitro fusion of ECPs or LBPs with lysosomes. Fusion rates understandard conditions (control) were set as 100%. This translates to 3–12%of the bacteria or latex beads being positive for the lysosomal fluor. Meansand standard deviations of ≥3 independent experiment are shown. �p < 0.05,��p < 0.01 compared to control. (A) Time course of phagosome–lysosome fu-sion. Fusion by 75 min (ECPs) or by 60 min (LBPs) was set as 100%. Fusionreactions contained ECPs (open circles, error bars downward) or LBPs (closedsquares, error bars upward). (B and C) Compounds added at the beginning ofthe fusion reaction: 4 mM NEM, 10 mM BAPTA, 0.5 mg∕mL RabGDI, 40 nMbafilomycin A1, 1 μM nigericin, 20 μM nocodazole. Reactions contained ECPs(B) or LBPs (C). (D–F) Reactions contained ECPs. (D) Colocalization of bacteriawith lysosomal BSA rhodamine and percentage of bacteria with meanFRET intensity > 0were calculated and expressed in percent of the respectivecontrol. One hundred percent (SD� 73, n ¼ 7) of colocalizing bacteriashowed mean FRET intensity > 0. (E) Affinity purified anti-Rab7 or (F) GST-Rab7 WTor GST-Rab7T22N were added at the indicated final concentrations.
Becken et al. PNAS ∣ November 30, 2010 ∣ vol. 107 ∣ no. 48 ∣ 20727
BIOCH
EMISTR
Y
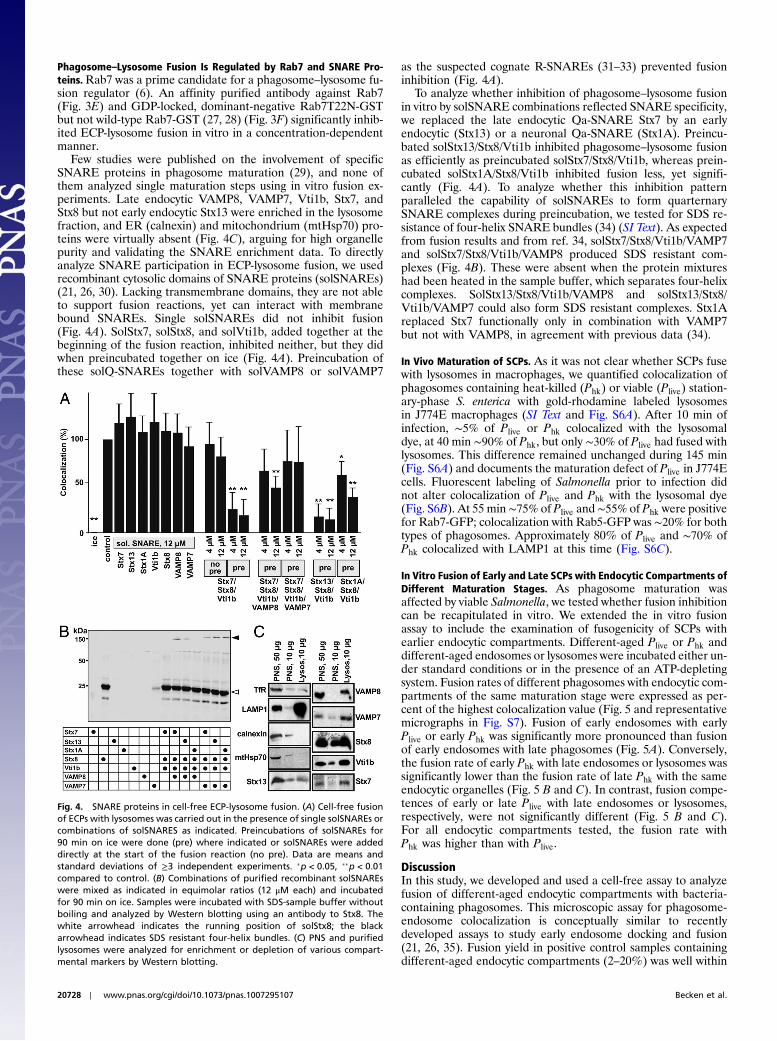
Phagosome–Lysosome Fusion Is Regulated by Rab7 and SNARE Pro-teins.Rab7 was a prime candidate for a phagosome–lysosome fu-sion regulator (6). An affinity purified antibody against Rab7(Fig. 3E) and GDP-locked, dominant-negative Rab7T22N-GSTbut not wild-type Rab7-GST (27, 28) (Fig. 3F) significantly inhib-ited ECP-lysosome fusion in vitro in a concentration-dependentmanner.
Few studies were published on the involvement of specificSNARE proteins in phagosome maturation (29), and none ofthem analyzed single maturation steps using in vitro fusion ex-periments. Late endocytic VAMP8, VAMP7, Vti1b, Stx7, andStx8 but not early endocytic Stx13 were enriched in the lysosomefraction, and ER (calnexin) and mitochondrium (mtHsp70) pro-teins were virtually absent (Fig. 4C), arguing for high organellepurity and validating the SNARE enrichment data. To directlyanalyze SNARE participation in ECP-lysosome fusion, we usedrecombinant cytosolic domains of SNARE proteins (solSNAREs)(21, 26, 30). Lacking transmembrane domains, they are not ableto support fusion reactions, yet can interact with membranebound SNAREs. Single solSNAREs did not inhibit fusion(Fig. 4A). SolStx7, solStx8, and solVti1b, added together at thebeginning of the fusion reaction, inhibited neither, but they didwhen preincubated together on ice (Fig. 4A). Preincubation ofthese solQ-SNAREs together with solVAMP8 or solVAMP7
as the suspected cognate R-SNAREs (31–33) prevented fusioninhibition (Fig. 4A).
To analyze whether inhibition of phagosome–lysosome fusionin vitro by solSNARE combinations reflected SNARE specificity,we replaced the late endocytic Qa-SNARE Stx7 by an earlyendocytic (Stx13) or a neuronal Qa-SNARE (Stx1A). Preincu-bated solStx13/Stx8/Vti1b inhibited phagosome–lysosome fusionas efficiently as preincubated solStx7/Stx8/Vti1b, whereas prein-cubated solStx1A/Stx8/Vti1b inhibited fusion less, yet signifi-cantly (Fig. 4A). To analyze whether this inhibition patternparalleled the capability of solSNAREs to form quarternarySNARE complexes during preincubation, we tested for SDS re-sistance of four-helix SNARE bundles (34) (SI Text). As expectedfrom fusion results and from ref. 34, solStx7/Stx8/Vti1b/VAMP7and solStx7/Stx8/Vti1b/VAMP8 produced SDS resistant com-plexes (Fig. 4B). These were absent when the protein mixtureshad been heated in the sample buffer, which separates four-helixcomplexes. SolStx13/Stx8/Vti1b/VAMP8 and solStx13/Stx8/Vti1b/VAMP7 could also form SDS resistant complexes. Stx1Areplaced Stx7 functionally only in combination with VAMP7but not with VAMP8, in agreement with previous data (34).
In Vivo Maturation of SCPs. As it was not clear whether SCPs fusewith lysosomes in macrophages, we quantified colocalization ofphagosomes containing heat-killed (Phk) or viable (Plive) station-ary-phase S. enterica with gold-rhodamine labeled lysosomesin J774E macrophages (SI Text and Fig. S6A). After 10 min ofinfection, ∼5% of Plive or Phk colocalized with the lysosomaldye, at 40 min ∼90% of Phk, but only ∼30% of Plive had fused withlysosomes. This difference remained unchanged during 145 min(Fig. S6A) and documents the maturation defect of Plive in J774Ecells. Fluorescent labeling of Salmonella prior to infection didnot alter colocalization of Plive and Phk with the lysosomal dye(Fig. S6B). At 55 min ∼75% of Plive and ∼55% of Phk were positivefor Rab7-GFP; colocalization with Rab5-GFP was ∼20% for bothtypes of phagosomes. Approximately 80% of Plive and ∼70% ofPhk colocalized with LAMP1 at this time (Fig. S6C).
In Vitro Fusion of Early and Late SCPs with Endocytic Compartments ofDifferent Maturation Stages. As phagosome maturation wasaffected by viable Salmonella, we tested whether fusion inhibitioncan be recapitulated in vitro. We extended the in vitro fusionassay to include the examination of fusogenicity of SCPs withearlier endocytic compartments. Different-aged Plive or Phk anddifferent-aged endosomes or lysosomes were incubated either un-der standard conditions or in the presence of an ATP-depletingsystem. Fusion rates of different phagosomes with endocytic com-partments of the same maturation stage were expressed as per-cent of the highest colocalization value (Fig. 5 and representativemicrographs in Fig. S7). Fusion of early endosomes with earlyPlive or early Phk was significantly more pronounced than fusionof early endosomes with late phagosomes (Fig. 5A). Conversely,the fusion rate of early Phk with late endosomes or lysosomes wassignificantly lower than the fusion rate of late Phk with the sameendocytic organelles (Fig. 5 B and C). In contrast, fusion compe-tences of early or late Plive with late endosomes or lysosomes,respectively, were not significantly different (Fig. 5 B and C).For all endocytic compartments tested, the fusion rate withPhk was higher than with Plive.
DiscussionIn this study, we developed and used a cell-free assay to analyzefusion of different-aged endocytic compartments with bacteria-containing phagosomes. This microscopic assay for phagosome-endosome colocalization is conceptually similar to recentlydeveloped assays to study early endosome docking and fusion(21, 26, 35). Fusion yield in positive control samples containingdifferent-aged endocytic compartments (2–20%) was well within
Fig. 4. SNARE proteins in cell-free ECP-lysosome fusion. (A) Cell-free fusionof ECPs with lysosomes was carried out in the presence of single solSNAREs orcombinations of solSNARES as indicated. Preincubations of solSNAREs for90 min on ice were done (pre) where indicated or solSNAREs were addeddirectly at the start of the fusion reaction (no pre). Data are means andstandard deviations of ≥3 independent experiments. �p < 0.05, ��p < 0.01compared to control. (B) Combinations of purified recombinant solSNAREswere mixed as indicated in equimolar ratios (12 μM each) and incubatedfor 90 min on ice. Samples were incubated with SDS-sample buffer withoutboiling and analyzed by Western blotting using an antibody to Stx8. Thewhite arrowhead indicates the running position of solStx8; the blackarrowhead indicates SDS resistant four-helix bundles. (C) PNS and purifiedlysosomes were analyzed for enrichment or depletion of various compart-mental markers by Western blotting.
20728 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1007295107 Becken et al.

the range of other systems reconstituting fusion of endosomes(21, 35, 36) or phagosomes (3, 37).
Authentic phagosome-lysosome fusion was observed in vitro,as reaction kinetics matched those of related cell-free fusionreactions (5, 21). Further, colocalization was strictly dependenton a physiological temperature and presence of ATP. Additionof cytosol increased fusion, but was not absolutely required ashas been seen with other in vitro fusion reactions (21, 22), likelyreflecting that fusion-enhancing peripheral membrane proteinssuch as NSF or Rab-GTPases are present on isolated membranes.Like late endosome–lysosome fusion (23), phagosome–lysosomefusion was reduced by calmodulin inhibitors, supporting previousreports on a role for calmodulin in phagosome–lysosometethering (18). Fusion was also strongly inhibited by NEM andRabGDI, reflecting the requirements for NSF and Rab proteins(19, 20). An important Rab-GTPase in phagosome–lysosomefusion was Rab7. Purified Rab proteins were not geranylgerany-lated and, hence, wild-type Rab7 did not promote fusion. Domi-nant-negative Rab7 inhibited phagosome–lysosome fusion, as itsfunction is not fully dependent on the lipid modification (38, 39).
Reconstitution of not only ECP- but also LBP-lysosome fusiondemonstrates the versatility of the presented assay. IsolatedLBPs are almost pure allowing in vitro fusion experiments in abiochemically defined system, albeit with a nonphysiologicalphagosome load. The presented reconstitution system can helpto molecularly investigate many of the observations made overthe years with latex bead-fed phagocytes.
SolSNAREs have previously been used to test SNARErequirements in fusion of endocytic organelles in vitro or in semi-intact cells (21, 26, 30). In our study, preincubated combinationsof soluble Qa/b/c-SNAREs but not of Qa/b/c/R-SNAREs inhib-ited ECP-lysosome fusion. Because formation of SNAREfour-helix bundles may proceed through Qa/b/c intermediates(40) and because our solSNAREs form SDS resistant quaternarycomplexes in vitro, we interpret this as follows: Ternary, three-helix solQa/b/c-SNARE complexes assemble during preincuba-tion on ice at a low rate (34), similar to binary three-helixSNAP-25/Stx1 complexes (41). The preassembled Q-SNAREsengage organelle embedded cognate R-SNARE(s) in futile bind-ing reactions and so efficiently block membrane fusion. Fusion
inhibition is prevented, when solQa/b/c- and solR-SNAREs forma quaternary complex already during preincubation.
In vitro formation of SDS resistant complexes (as in refs. 34,42, 43) and fusion inhibition by solQa/b/c-SNARE complexeswere promiscuous in that they also occurred when late endocyticStx7 (Qa) was replaced by early endosomal Stx13 (Qa) or neu-ronal Stx1A (Qa). There is precedence for SNARE promiscuityin the endocytic system, e.g., the functional replacement of Vti1bby other SNAREs in Vti1b knockout mice (44). In contrast,homotypic early endosome fusion in vitro was inhibited onlyby early endosomal (solStx13/Vti1a/Stx6) but not neuronal(solStx1/SNAP-25) SNARE domains (21). Yet, combinationsof solStx7/Vti1b/Stx8 or solStx13/Vti1b/Stx8 inhibited phago-some–lysosome fusion more efficiently compared to solStx1A/Vti1b/Stx8. As the first two SNARE groups form SDS resistantfour-helix bundles with either solVAMP7 or solVAMP8, whichare both present on lysosomes, but the third one only withsolVAMP7, this could indicate that both of these R-SNAREsare able to mediate phagosome–lysosome fusion. Inhibition bysolStx1A/Vti1b/Stx8 is then less efficient, because only one ofthese R–SNAREs is inactivated by this complex.
Acidification of newly formed phagosomes supports phagoly-sosome formation (24) and is important for autophagosome–lysosome fusion (45). It may be important for vesicle buddingin endosome and phagosome maturation (46, 47), yet, its rolein fusion of late phagosomes or endosomes with lysosomes hasnot been analyzed directly. As neither bafilomycin nor nigericininhibited phagosome–lysosome fusion, organelle acidificationseems not to be required for this specific subreaction of phago-lysosome formation.
To further understand diversion of phagosome maturation byintracellular pathogens, we compared fusion competence of earlyand late phagosomes containing live or heat-killed S. enterica withdifferent-aged endocytic vesicles. We have shown that in vitro,early endosomes fuse more avidly with early than with late Phkand late endosomes and lysosomes are more fusogenic with latethan with early Phk as expected. However, some fusion occurredbetween early Phk and late endocytic organelles and, conversely,late Phk and early endosomes. One explanation for this unex-pected finding is heterogenity in the maturation stages of the pha-gosomes in that 10–20% of late Phk were still positive for earlyendocytic Rab5, yet negative for lysosomal gold rhodamine. Rab7is present on the majority of endosomes that are only 0–10 minold, and this may explain fusion competence between early endo-somes and late Phk in vitro. In vivo, this fusion may not readilyoccur due to spatial segregation from late phagosomes andphagolysosomes. Whether the presence of both Rabs on mostearly endosomes is a general macrophage characteristic remainsto be investigated.
Fusion competence of both Plive and Phk with early endosomesdecreased as phagosomes aged. But whereas Phk acquired fusioncompetence with late endocytic organelles, this was much less thecase with Plive. It has been suggested (48) that late SCPs in macro-phages retain Rab5 and Rab5 effectors, stay fusogenic with earlyendosomes, and that this sustained fusion occurs via an uncon-ventional mechanism independent of ATP hydrolysis. We, too,observed ATP-independent fusion of late Plive with early endo-somes, yet the fusion extent was much lower than fusion of earlyPlive with early endosomes. And in our study, Rab7-GFP but notRab5-GFP was associated with the majority of late Plive. Hence,the strong fusion defect between late Plive and lysosomes was notcaused by an early arrest as proposed in ref. 48 but rather by anarrest at a late maturation step.
Our data show that the low fusion competence of Plive withlysosomes in vivo was reconstituted in vitro and that reducedfusion competence of Plive was not restricted to lysosomes. Suchgenerally reduced fusion competence of Plive is in line with thereported reduced accessibility of SCPs to incoming transferrin
Fig. 5. In vitro fusion of Salmonella-containing phagosomes with different-aged endocytic compartments. In vitro fusion experiments were carriedout with early (10 min) or late (55 min) phagosomes containing live (Plive)or heat-killed (Phk) S. enterica. Fusion rates of phagosomes with endosomesof different age cannot be directly compared, because fluor contents andvolume of endocytic organelles likely change with maturation. Therefore,only fusion reactions with same-stage endocytic compartments were com-pared. Only phagosomes containing an intact membrane (bacteria werenot antibody accessible) were considered. (A) Fusion reactions contained100∕00 endosomes (early endosomes, EE), (B) 100∕150 endosomes (late endo-somes, LE), or (C) 100∕1200 endosomes (lysosomes, Lys). Data are the meansand standard deviations of ≥3 independent experiments. Asterisks or valuesabove open bars without brackets mark p values comparing adjacent openand filled bars. ��p < 0.01; *: p < 0, 05.
Becken et al. PNAS ∣ November 30, 2010 ∣ vol. 107 ∣ no. 48 ∣ 20729
BIOCH
EMISTR
Y

and dextran (10). Salmonella effector proteins such as spiC (49),which are secreted from the phagosome into the cytosol of in-fected cells via a type III secretion system, were discussed tobe prime candidates for altering maturation of the SCP. However,phagosomes that contained Salmonella generally defective ineffector protein secretion showed no enhanced fusion withlysosomes (11); hence, the role of effector proteins remainselusive. Against this background it was revealing that fusion ofPlive with lysosomes in vitro was diminished with lysosomes fromuninfected cells and did not require cytosol prepared from in-fected macrophages, which would be a prime source of secretedeffectors. This suggests that the intraphagosomal bacteria hadturned phagosomes fusion-incompetent before they were iso-lated. Whether reprogramming requires prior bacterial secretionor the presence of certain surface molecules, such as lipopolysac-charides, remains to be investigated. It has been proposed thatdiminished phagosome–lysosome fusion in vivo may be the resultof an impaired capability of SCPs to attach to microtubules (MTs)and their reduced trafficking (50, 51). Disruption of MTs bynocodazole had only a minor effect on standard phagosome–lysosome fusion in our study, demonstrating that a possible needfor MTs in phagosome maturation in vivo would be bypassed invitro. Hence, inhibition of phagosome movement is not the (only)decisive factor in trafficking diversion of SCPs.
Material and MethodsChemicals. Antibodies, plasmids and recombinant proteins, buffers, and pre-paration of cytosol are described in SI Text. Cultivation of mammalian andbacterial cells and their use in infection experiments is described in SI Text.
Fluorescent Labeling of Endocytic Compartments. To label lysosomes for invitro fusion experiments with ECPs or LBPs, macrophages were incubatedovernight (oN) in complete medium containing 50 μg∕mL BSA rhodamine(5 μg∕mL for FRETanalysis). Cells were rinsed with PBS, incubated in completemedium containing ferrofluid (SI Text) for 5 min, rinsed twice with PBS, andchased at 37 °C for 2 h. For in vitro fusion experiments with SCPs, ferrofluidwas incubated oN in BSA rhodamine in PBS and diluted in complete mediumresulting in 100 μg∕mL BSA rhodamine and 6 μL∕mL ferrofluid. Macrophageswere incubated for 10 min at 37 °C with BSA rhodamine/ferrofluid, rinsedtwice with PBS, and chased as indicated. For biochemical analysis of purifiedendocytic compartments (SI Text), lysosomes were labeled with 50 μg∕mLBSA rhodamine oN, chased for 2 h, and ferrofluid was added. To quantifyphagosome–lysosome colocalization in vivo, macrophages were incubatedin complete medium containing gold rhodamine for 1 h (infection withE. coli: OD520 ¼ 1; infection with S. enterica: OD520 ¼ 0, 4) or containing50 μg∕mL BSA rhodamine oN, rinsed thrice with PBS and chased for 3 h(infection with E. coli) or 2 h (infection with S. enterica or latex beads).
Preparation of Phagosomes and Endocytic Compartments. Infection ofmacrophages is described in SI Text. Bacteria-containing phagosomes wereprepared as in ref. 5 with modifications: Cells were sequentially washedwith PBS∕5 mM EDTA and homogenization buffer (HB), resuspended inHB∕10 mg∕mL BSA∕1 × protease inhibitors, and homogenized in a Dounce
homogenizer. A PNS was prepared at 450 × g for 3 min and centrifugedin a fixed angle rotor: When the rotor reached 12;000 × g, the centrifugewas turned off, the supernatant was centrifuged at 12;000 × g for 6 min,and the pellet was resuspended in HB∕10 mg∕mL BSA. Preparation of LBPsis described in SI Text.
Using pulse/chase protocols, endocytic compartments were loaded withferrofluid (EMG 508, Ferrotec Europe) and purified from a PNS as in ref. 17with modifications (SI Text).
In Vitro Fusion Assay. Standard fusion reactions contained 1.5 μL of 10 × saltsolution (15 mM MgCl2, 1 M KCl), 1.5 μL of 10 × ATP regenerating system(SI Text), 0.15 μL 100 mM DTT, 2 mg∕mL cytosol, 2 μL of the endosome orlysosome preparation and 2 μL of the phagosome preparation. Volumes wereadjusted to 15 μL with HB. Each reaction contained lysosomes from 7.5 × 106
or late or early endosomes from 1.3 × 107 macrophages. Per reaction, bacter-ia-containing phagosomes from 7.5 × 105 macrophages or LBPs (final concen-tration: A600 ¼ 0.3) were added. In control experiments, cytosol was replacedby HB. Recombinant proteins or pharmacologicals were either from a stock inHB or carrier-containing control samples were included. Fusion experimentswere at 37 °C for 60 min and stopped by placing the tubes on ice. To identifybacteria not surrounded by an intact phagosome membrane, samples withSCPs were incubated with 5 μg∕mL rabbit anti-Salmonella antibody at 4 °Cfor 30 min. This labeling was omitted in samples containing ECPs or LBPs.Samples were diluted with 1 mL cold HB. Poly-L-lysine coated coverslips wereplaced in a 24-well plate, 500 μL of the diluted fusion sample were added perwell, and plates were centrifuged at 1;800 × g for 15 min at 4 °C. Sampleswere fixed with 2.5% glutardialdehyde and 2% formaldehyde (FA) in HB(samples with ECPs or LBPs) or 3% FA in HB (samples with SCPs and samplesfor FRETanalysis). After rinsing three times with PBS, samples were incubatedin 1 mg∕mLNaBH4 in PBS (ECPs or LBPs) or 50 mMNH4Cl in HB (SCPs and FRETanalysis) for 30 min. Samples with SCPs were stained with a Cy5-conjugatedsecondary antibody. Coverslips were mounted in Mowiol. Samples wereexamined using a Zeiss Axioplan (ECPs or LBPs) or a Zeiss Axio Observer.Z1microscope (SCPs and FRET analysis) equipped with 100×/1.4 oil immersionobjectives and appropriate filter sets. Percentages of beads or green bacteriacolocalizing with lysosomal BSA rhodamine were determined from 300particles each in duplicates. In samples with SCPs, Cy5-labeled bacteria wereexcluded from analysis. All coverslips were anonymized. FRET analysis wasperformed according to the principle described by Youvan et al. (52) usingZeiss Axio Vision 4.7.1 software. ATTO488-labeled bacteria were definedas regions of interest (R) and mean FRET intensity was calculated for eachR. Per sample, 200 bacteria were analyzed, and percentages of Rs with meanFRET intensities > 0 were expressed as percent of the positive control. FRETintensity for every pixel was displayed in a false color-coded image.
Statistical Analysis. Means and standard deviations were calculated fromindependent experiments. Data were analyzed by the two-tailed unpairedStudent’s t test with significance assumed at p < 0.05 and high significanceat p < 0.01.
ACKNOWLEDGMENTS. We thank all colleagues who generously contributedplasmids and antibodies. This study was supported by the DeutscheForschungsgemeinschaft (SFB 645) with initial support by the VolkswagenFoundation.
1. Haas A (2007) The phagosome: Compartment with a license to kill. Traffic 8:311–330.2. Desjardins M, Huber LA, Parton RG, Griffiths G (1994) Biogenesis of phagolysosomes
proceeds through a sequential series of interactions with the endocytic apparatus.J Cell Biol 124:677–688.
3. Jahraus A, et al. (1998) In vitro fusion of phagosomes with different endocyticorganelles from J774 macrophages. J Biol Chem 273:30379–30390.
4. Zerial M,McBride H (2001) Rab proteins as membrane organizers.Nat RevMol Cell Biol2:107–117.
5. Alvarez-Dominguez C, et al. (1996) Phagocytosed live Listeria monocytogenesinfluences Rab5-regulated in vitro phagosome-endosome fusion. J Biol Chem271:13834–13843.
6. Harrison RE, et al. (2003) Phagosomes fuse with late endosomes and/or lysosomesby extension of membrane protrusions along microtubules: Role of Rab7 and RILP.Mol Cell Biol 23:6494–6506.
7. Smith AC, et al. (2007) A network of Rab GTPases controls phagosomematuration andis modulated by Salmonella enterica serovar Typhimurium. J Cell Biol 176:263–268.
8. Jahn R, Scheller RH (2006) SNAREs: Engines for membrane fusion.Nat RevMol Cell Biol7:631–643.
9. Drecktrah D, Knodler LA, Ireland R, Steele-Mortimer O (2006) The mechanism ofSalmonella entry determines the vacuolar environment and intracellular gene expres-sion. Traffic 7:39–51.
10. Rathman M, Barker LP, Falkow S (1997) The unique trafficking pattern of Salmonellatyphimurium-containing phagosomes in murine macrophages is independent of themechanism of bacterial entry. Infect Immun 65:1475–1485.
11. Garvis SG, Beuzon CR, Holden DW (2001) A role for the PhoP/Q regulon in inhibitionof fusion between lysosomes and Salmonella-containing vacuoles in macrophages.Cell Microbiol 3:731–744.
12. Buchmeier NA, Heffron F (1991) Inhibition of macrophage phagosome-lysosomefusion by Salmonella typhimurium. Infect Immun 59:2232–2238.
13. Oh YK, et al. (1996) Rapid and complete fusion of macrophage lysosomes with phago-somes containing Salmonella typhimurium. Infect Immun 64:3877–3883.
14. Mukherjee K, et al. (2000) Live Salmonella recruits N-ethylmaleimide-sensitive fusionprotein on phagosomal membrane and promotes fusion with early endosome. J CellBiol 148:741–753.
15. Vergne I, et al. (2004) Mycobacterium tuberculosis phagosome maturation arrest:Mycobacterial phosphatidylinositol analog phosphatidylinositol mannoside stimu-lates early endosomal fusion. Mol Biol Cell 15:751–760.
16. Mukherjee K, et al. (2002) Diverting intracellular trafficking of Salmonella to thelysosome through activation of the late endocytic Rab7 by intracellular delivery ofmuramyl dipeptide. J Cell Sci 115:3693–3701.
17. Li HS, Stolz DB, Romero G (2005) Characterization of endocytic vesicles using magneticmicrobeads coated with signalling ligands. Traffic 6:324–334.
20730 ∣ www.pnas.org/cgi/doi/10.1073/pnas.1007295107 Becken et al.

18. Stockinger W, et al. (2006) Differential requirements for actin polymerization, calmo-dulin, and Ca2þ define distinct stages of lysosome/phagosome targeting.Mol Biol Cell17:1697–1710.
19. Ullrich O, et al. (1993) Rab GDP dissociation inhibitor as a general regulator for themembrane association of Rab proteins. J Biol Chem 268:18143–18150.
20. Malhotra V, et al. (1988) Role of an N-ethylmaleimide-sensitive transport componentin promoting fusion of transport vesicles with cisternae of the Golgi stack. Cell54:221–227.
21. Brandhorst D, et al. (2006) Homotypic fusion of early endosomes: SNAREs do notdetermine fusion specificity. Proc Natl Acad Sci USA 103:2701–2706.
22. Haas A, Conradt B, Wickner W (1994) G-protein ligands inhibit in vitro reactions ofvacuole inheritance. J Cell Biol 126:87–97.
23. Pryor PR, et al. (2000) The role of intraorganellar Ca2þ in late endosome-lysosome het-erotypic fusion and in the reformation of lysosomes from hybrid organelles. J Cell Biol149:1053–1062.
24. Yates RM, Hermetter A, Russell DG (2005) The kinetics of phagosome maturation as afunction of phagosome/lysosome fusion and acquisition of hydrolytic activity. Traffic6:413–420.
25. Wu P, Brand L (1994) Resonance energy transfer: Methods and applications. Anal Bio-chem 218:1–13.
26. Geumann U, et al. (2008) SNARE function is not involved in early endosome docking.Mol Biol Cell 19:5327–5337.
27. Cantalupo G, et al. (2001) Rab-interacting lysosomal protein (RILP): The Rab7 effectorrequired for transport to lysosomes. EMBO J 20:683–693.
28. Bucci C, et al. (2000) Rab7: A key to lysosome biogenesis. Mol Biol Cell 11:467–480.29. Collins RF, Schreiber AD, Grinstein S, Trimble WS (2002) Syntaxins 13 and 7 function at
distinct steps during phagocytosis. J Immunol 169:3250–3256.30. Hua Y, Scheller RH (2001) Three SNARE complexes cooperate to mediate membrane
fusion. Proc Natl Acad Sci USA 98:8065–8070.31. Mullock BM, et al. (2000) Syntaxin 7 is localized to late endosome compartments,
associates with VAMP 8, and is required for late endosome-lysosome fusion. Mol BiolCell 11:3137–3153.
32. Pryor PR, et al. (2004) Combinatorial SNARE complexes with VAMP7 or VAMP8 definedifferent late endocytic fusion events. EMBO Rep 5:590–595.
33. Antonin W, et al. (2000) The R-SNARE endobrevin/VAMP-8 mediates homotypic fusionof early endosomes and late endosomes. Mol Biol Cell 11:3289–3298.
34. AntoninW, et al. (2000) A SNARE complexmediating fusion of late endosomes definesconserved properties of SNARE structure and function. EMBO J 19:6453–6464.
35. Rizzoli SO, et al. (2006) Evidence for early endosome-like fusion of recently endocy-tosed synaptic vesicles. Traffic 7:1163–1176.
36. Robinson LJ, Aniento F, Gruenberg J (1997) NSF is required for transport from early tolate endosomes. J Cell Sci 110(Pt 17):2079–2087.
37. Mayorga LS, Bertini F, Stahl PD (1991) Fusion of newly formed phagosomes withendosomes in intact cells and in a cell-free system. J Biol Chem 266:6511–6517.
38. Li G, Barbieri MA, Colombo MI, Stahl PD (1994) Structural features of the GTP-bindingdefective Rab5 mutants required for their inhibitory activity on endocytosis. J BiolChem 269:14631–14635.
39. Stenmark H, et al. (1994) Inhibition of Rab5 GTPase activity stimulates membranefusion in endocytosis. EMBO J 13:1287–1296.
40. Fiebig KM, Rice LM, Pollock E, Brunger AT (1999) Folding intermediates of SNAREcomplex assembly. Nat Struct Biol 6:117–123.
41. Fasshauer D, et al. (1997) Structural changes are associated with soluble N-ethylma-leimide-sensitive fusion protein attachment protein receptor complex formation.J Biol Chem 272:28036–28041.
42. Fasshauer D, et al. (1999) Mixed and non-cognate SNARE complexes. Characterizationof assembly and biophysical properties. J Biol Chem 274:15440–15446.
43. Yang B, et al. (1999) SNARE interactions are not selective. Implications for membranefusion specificity. J Biol Chem 274:5649–5653.
44. Atlashkin V, et al. (2003) Deletion of the SNARE vti1b in mice results in the loss of asingle SNARE partner, Syntaxin 8. Mol Cell Biol 23:5198–5207.
45. Kawai A, et al. (2007) Autophagosome-lysosome fusion depends on the pH in acidiccompartments in CHO cells. Autophagy 3:154–157.
46. Gu F, Gruenberg J (2000) ARF1 regulates pH-dependent COP functions in the earlyendocytic pathway. J Biol Chem 275:8154–8160.
47. Beron W, Mayorga LS, Colombo MI, Stahl PD (2001) Recruitment of coat-protein-com-plex proteins on to phagosomal membranes is regulated by a brefeldin A-sensitiveADP-ribosylation factor. Biochem J 355:409–415.
48. Parashuraman S, Madan R, Mukhopadhyay A (2010) NSF independent fusion of Sal-monella-containing late phagosomes with early endosomes. FEBS Lett 584:1251–1256.
49. Uchiya K, et al. (1999) A Salmonella virulence protein that inhibits cellular trafficking.EMBO J 18:3924–3933.
50. Shotland Y, Kramer H, Groisman EA (2003) The Salmonella SpiC protein targets themammalian Hook3 protein function to alter cellular trafficking. Mol Microbiol49:1565–1576.
51. Marsman M, et al. (2004) Dynein-mediated vesicle transport controls intracellularSalmonella replication. Mol Biol Cell 15:2954–2964.
52. Youvan D, et al. (1997) Calibration of fluorescence resonance energy transfer inmicroscopy using genetically engineered GFP derivates on nickel chelating beads.Biotechnology 3:1–18.
Becken et al. PNAS ∣ November 30, 2010 ∣ vol. 107 ∣ no. 48 ∣ 20731
BIOCH
EMISTR
Y