Embed Size (px)
Citation preview

ASSESSMENT OF HERBIVORE INDUCED PLANT VOLATILES IN
JUVENILE HOPS BY EXPOSURE TO METHYL SALICYLATE
By
RITA M. ABDELLA
A thesis submitted in partial fulfillment of
the requirements for the degree of
MASTER OF SCIENCE IN CHEMISTRY
WASHINGTON STATE UNIVERSITY
Department of Chemistry
DECEMBER 2010

ii
To the Faculty of Washington State University:
The members of the Committee appointed to examine the thesis of RITA MARIE ABDELLA
find it satisfactory and recommend that it be accepted.
__________________________
Vincent R. Hebert, PhD., Chair
___________________________
Herbert H. Hill, PhD.
____________________________
Steven G. Metcalf, PhD.
_____________________________
William D. Samuels, PhD.

iii
ACKNOWLEDGEMENT
First, I would like to thank Dr. Vincent Hebert for acting as my advisor throughout the
completion of this thesis. Dr. Hebert‟s keen eye and attention to detail were most appreciated
during the writing of the manuscript and thesis. I would also like to thank Jane Le Page for all
the analytical work that was performed during this study. A big thank you goes out to Dr.
William Samuels, who has provided me with the encouragement to keep going and for the
instruction over the course of this degree. Many thanks also go out to the remaining members of
my committee; Dr Herbert Hill and Dr. Steven Metcalf.
Lastly, I would like to thank my grandchildren: Linden, Loghan and Makena Kastl, for
being patient with Grammy for not making a lot of their sports events and to my husband Charlie
McDonald and my daughter, Astrid Kastl for seeing me through this. And finally I want to thank
my father, Richard Abdella, for instilling the importance of education in me and in memoriam to
my mother Mary Abdella, who wasn‟t here to see me finish. Mom, you would have been proud.

iv
ASSESSMENT OF HERBIVORE INDUCED PLANT VOLATILES IN
JUVENILE HOPS BY EXPOSURE TO METHYL SALICYLATE
Abstract
by Rita Marie Abdella
Washington State University
December 2010
Chair: Vincent R. Hebert
The volatile secondary plant product methyl salicylate (MeSA) exhibits numerous properties,
including the attraction of beneficial insects to plants undergoing attack from herbivores. This
capability to attract beneficial species is being explored as a “green” alternative to pesticides in
hop fields and grape vineyards. While some field studies have been performed using MeSA as
an attractant for predatory species or herbivore repellant, its capability to act as a priming
mechanism to incite the production of herbivore induced plant volatiles (HIPVs) has not been
examined. To examine if MeSA can intra-specifically induce production of volatiles beneficial
to insect attraction, juvenile hops plants were placed in environmentally controlled enclosures in
close proximity to MeSA controlled release dispensers over a 72-hour exposure period.
Immediately after exposure, replicate exposed plants (and respective control plants) were placed
into individual glass chambers. The chamber atmosphere from both MeSA exposed and control
hop plants were sampled by solid phase microextraction (SPME) to chemically profile volatile
emissions after the 72-hour MeSA exposure period. SPME collections were analyzed by gas
chromatography-mass spectroscopy (GC-MS) to identify volatile compound emissions. After

v
completion of the above examinations, the control hop and the treated hop plan were combined
within in a common enclosure to assay if MeSA primed hop plants could chemically “cross-talk”
with untreated plants to warn of impending herbivory. After this combined period, plant
chamber emissions were chemically profiled by SPME-GC/MS. Spectral profiling assessment
suggests MeSA exposed hops may increase production of the sesquiterpene, (E, E) α farnesene,
an identified HIPV. These year-to-year emission observations lend support to previous field
investigations that indicate airborne MeSA emissions may elicit production of HIPVs that can
serve as beneficial insect attractants. In the second combined control-MeSA hop treated
chamber evaluation series, the control plant head space volatiles showed a relative increase in
production of the sesquiterpene, α –caryophyllene, a known HIPV. The observed trend in
increased production of α–caryophyllene suggests a volatile signal(s) between plants in close
proximity may induce the production of chemicals needed in plant defense from insect
herbivory. Although the head space emission information collected from chamber evaluations
indicate relatively greater production of certain HIPVs after MeSA hop exposure, differences in
plant rearing (rhizomes versus soft wood cuttings), other environmental conditions (greenhouse
versus growth chamber plant rearing) likely resulted in the appreciable variation in emission
profiles among the replicated MeSA exposure trials conducted from 2007 through 2009. The
emission data herein should be viewed as qualitative. Certain evident trends in HIPV production,
however, bears further investigation. The experimental procedures developed herein should serve
to better design future evaluations for understanding and characterizing chemically induced plant
defense from herbivory.

vi
Table of Contents
Page
Acknowledgements…………………………………………………………………………iii
Abstract……………………………………………………………………………………..iv
Table of Contents…………………………………………………………………………...vi
List of Tables……………………………………………………………………………….vii
List of Figures……………………………………………………………………………...viii
Acronyms and Abbreviations………………………………………………………………ix
Preface………………………………………………………………………………………1
CHAPTER 1 Literature Review………………………………………………….4
Introduction…………………………………………………………………………5
Section I: Plant Response to Damage………………………………………………5
.
Section II: Recent developments and future applications for herbivore induced plant
volatile…………………………………………………………………22
Section III: Most recent controversies and trends in HIPV research……………...24
Section IV: Conclusions……………………………………………………….…..27
CHAPTER 2 Assessment of the Production of Herbivore Induced Plant Volatiles in
Juvenile Hops Using Methyl Salicylate…………………………….….40
1. Introduction…………………………………………….………41
CHAPTER 3 Final Conclusions and Future Work……………………….………66
APPENDIX A Analytical Summary Report……………………………….………71

vii
Tables
Chapter 2 Tables
Table 1 Summary of Compounds Collected in Experiment 1from unexposed (Control) and
Methyl Salicylate (MeSA) exposed juvenile hop H. humulus by treatment year….. 57
Table 2 Summary of Compounds Collected in Experiment 2 from unexposed (Control)
and Methyl Salicylate (MeSA) exposed juvenile hop H. humulus following 48 hours
combined exposure by treatment year…………………………………………….…58
Appendix Tables
Table 1 Sample Inventory and History………………………………………….……76
Table 2 Interval Data……………………………………………………………….…83
Table 3 Treated and Control Hop Replicate Data……………………………….……93
Table 4 Greenhouse Air Sample Data…………………………………………….…..97
Table 5 Treated and Control Plant Before and After Combined Exposure………….98

viii
Figures
Chapter 1 Figures
Figure 1 History of research on airborne plant-plant signaling…………………….…35
Figure 2 Impact of C6 Volatiles on Herbivore-Plant Interaction……………………..36
.
Figure 3 Volatile compounds from corn seedlings (Zea mays) undamaged (A),
first hour after feeding (B) and several hours after feeding (C)……………..37
Figure 4 Factors causing and traits affected by volatile-mediated signaling
among plants………………………………………………………………...38
Figure 5 Relative Distances for HIPV Emission ………………………………….…..39
Chapter 2 Figures
Figure 1 Chromatographic profiles of volatiles from unexposed (A) and MeSA
treated hops (B)……………………………………………………………….59
Figure 2 Structural identification of ten of sixteen compounds in head space……..….60
Figure 3 Structural identification of six of sixteen compounds in head space…….…..61
Figure 4 Volatiles from control plant by growth age……………………………..….…62
Appendix Figures
Figure 1 Comparative data from 3/22/07-Treated vs. Control………………………...103
Figure 2 Treated vs. Treated after 48 hrs combination…………………………….…..104

ix
Acronyms and Abbreviations
Compounds with six carbons C6
Green Leaf Volatiles GLV
Herbivore Induced Plant Volatiles HIPV
Irrigated Agricultural Experimental Research Center IAREC
Jasmonic Acid JA
Methyl Jasmonate MeJA
Methyl Salicylate MeSA
Salicylic Acid SA
Systemic Acquired Resistance SAR
(E,E)-4,8,12- trimethyl-1,3,7,11-tridecatraene TDTT
Volatile Organic Carbon VOC

1
Preface
The field of chemical ecology has been exploring the communication between plants
using chemical signaling since the 1980‟s. Early studies indicate that interspecies
communication existed as a function of perceived threat from insect damage. It was noted that in
interspecies communication certain chemical signals were emitted either via transpiration
through leaves or via a wound area caused by insect damage. This signal provided warning to
adjacent plants, which in turn continued the process and became part of a large network of
communication. These signals were relayed over a large area, providing plants of the same
species a means to develop defense mechanisms as well as provide a warning signal.
The concept of induced chemical plant protection as well as communication with
beneficial insects was considered controversial in the late 1970‟s. Studies in the early 1980‟s
indicated, however, that various secondary chemicals were emitted by plants that served as not
only alarm signals in the general vicinity, but were used by predatory insects to locate the
herbivorous insects causing injury to the plants. The concept of this plant-herbivore-predator
interaction provided the basis for an exciting research area in the study of plant-insect
interactions. Recent research in the field of chemical ecology has yielded a great deal of
information regarding the use of specific plant generated volatile chemicals as a green alternative
to pesticides.
The primary research addressed in this thesis is “can airborne plant exposure to methyl
salicylate (MeSA),”prime” inter and intra plant signaling by the production of herbivore

2
induced plant volatiles (HIPVs) in hop plants?” In conjunction with Dr. David James of the
Irrigated Agricultural Research Center (IARC) and his staff, particularly Larry Wright, juvenile
hop plants were exposed to MeSA air emissions from a commercially produced dispenser for a
period of 72 hours to assess the capability of this volatile compound to stimulate intra-plant
production of HIPVs. To assay if MeSA could trigger secondary metabolite production, the
treated plants as well as an unexposed control plant were placed in individual glass chambers
(developed by the James group) and transported to the Food and Environmental Quality (FEQL)
at Washington State University-Tri-Cities campus for analysis by solid phase microextraction
(SPME) and gas chromatography-mass spectroscopy (GC-MS). All lab analyses were performed
by FEQL Research Analyst, Jane Le Page. My primary role in the research was the collection
and interpretation of chemical emission data and reporting.
In this body of work Chapter 1 “Herbivore Induced Plant Volatiles and Chemical
Signaling- Effects on the Plant-Prey-Predator Interaction” discusses the relevant literature
regarding the plant synthesis of volatile organic compounds such as terpenes, fatty acid
derivatives and phenolics that provide “direct” defenses against attack from herbivorous pests, as
well as bacteria and fungi. These induced volatile emissions can also serve the plant in mounting
an “indirect” defense by attracting predatory insects while under herbivore attack. In turn, plants
in the vicinity of the stressed plant can “eavesdrop” on these SOS signals and use the signal to
begin mounting their own lines of defense. Chapter 2 “Assessment of Herbivore Induced Plant
Volatiles in Juvenile Hops by Exposure to Methyl Salicylate” is in the process of being

3
developed as a manuscript submission for the Journal of Chemical Ecology. Lastly, Chapter 3
discusses my final summary remarks and possibilities for future work. Appendix A is the
Analytical Summary Report. This regulatory science document provides a detailed overview of
the initial trials as performed in 2007.

4
Chapter 1
Herbivore Induced Plant Volatiles and Chemical Signaling
Effects on Plant-Prey-Predator Interaction

5
Introduction
Natural product chemical communication and the understanding of how this
communication affects plant-prey-predator interactions is an important aspect of chemical
ecology. While the above chemical communication is receiving increasing investigative
attention, considerable debate on the mechanisms of biosynthesis and function of these plant
volatile compound emissions still exists. This chapter review covers the plant‟s myriad of
responses to pathogen-insect injury, the production and biosynthesis of the herbivore induced
plant volatiles that include the C6 green leaf volatiles, the terpenes/terpenoids as well as other
plant secondary metabolites that can repel harmful but attract beneficial insects. The importance
of herbivore induced plant volatiles is highlighted; specifically their importance on plant-prey-
predator relationships and the impact of recent studies in the field of chemical ecology. My
intent is to provide a framework from the existing literature on the primary tenet of this thesis
“the ability of the secondary metabolite, methyl salicylate to “prime” intra and inter-plant
stimulus of the production of herbivore induced volatiles.”
Section 1: Plant response to damage
Plants emit volatile organic compounds in response to feeding by herbivorous insects.
These emissions vary from plant to plant, genotype and cultivar, species and age of herbivore
and time of the duress that the plant undergoes. Although mechanistically the purpose for these

6
secondary plant metabolites are not fully understood, assays have verified that emitted chemical
compounds act as toxins or repellents to drive away insect pest or provide antibiotic properties as
a defense against pathogens, may have developed a biosynthetic pathway to release volatile
chemical blend that were attractive to parasitic or predatory insects. (Turlings et al., 1995;
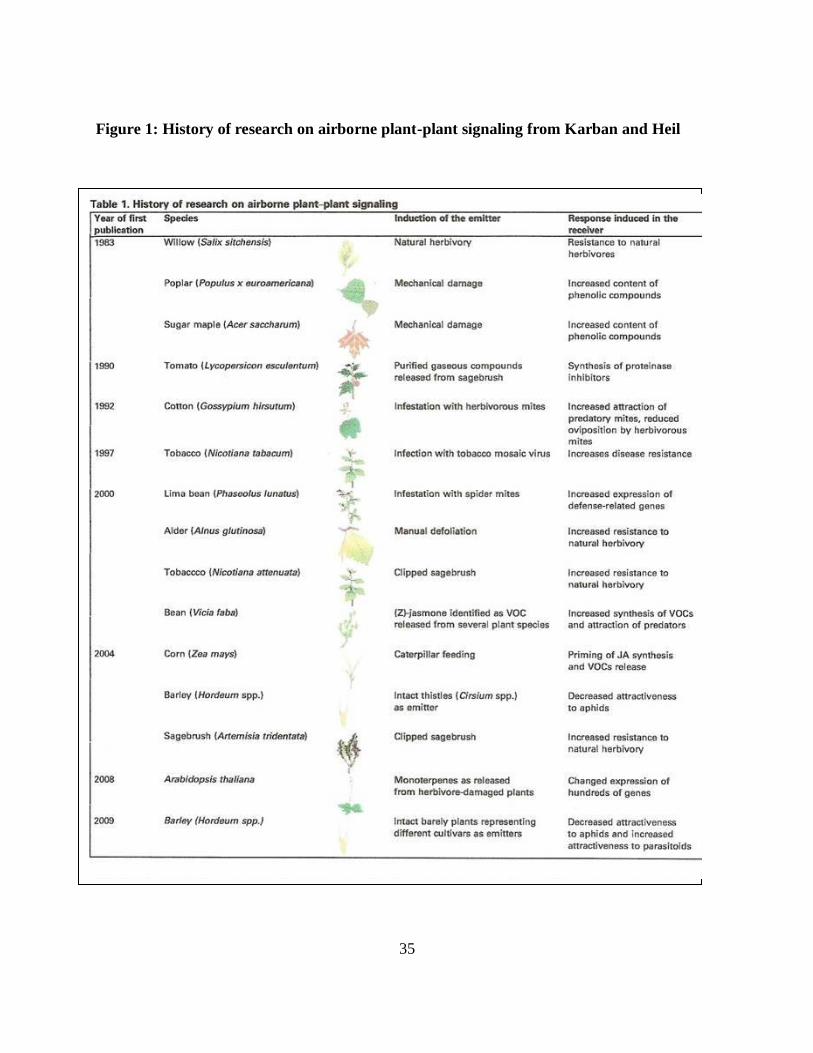
Walling, 2000). Research in this area of chemical ecology is rapidly growing. As an example,
Karban and Heil (2010) illustrates in Figure 1 the recent investigative history and acceleration of
plant-to-plant signaling research in forest, agricultural crop, and native plant communities.
Section 1.1 Direct and indirect response to herbivore damage
Section 1.1.1 Direct responses to herbivore damage
Plants have developed a wide variety of mechanisms to control injury from foraging
insects. Biosynthesized chemicals that provide an immediate negative impact against herbivore
damage are classified as direct defense compounds (Arimura et al., 2005). These defenses
include the production of terpenoids, alkaloids, and other biosynthesized secondary metabolites
which are toxic or repellent to the insect initiated through the production of defense genes
(Arimura et al., 2005). The plant phytohormone, ethylene, was suggested to be the chemical
messenger for the transport of volatile signals between adjoining plant systems, providing the
means to turn on defense genes in the plant as protection from herbivores (Farmer and Ryan,
1990). These investigators suggest that the defense genes initiate proteinase inhibitors to make
the plant less palatable to herbivores, and also acted as part of the plant‟s systemic alarm system
to carry the damage information throughout the entire plant. Although no biological evidence

7
existed at that time that any other compound than ethylene could provide atmospheric signaling,
exogenous application of the plant lipid compound, methyl jasmonate (MeJA) by these
investigators did elicit proteinase response in tomatoes (Lycopersicom esculentum) at a rate
much higher than exhibited by mechanical wounding. When treated plants were housed with
control plants, the controls developed low levels of proteinase inhibitor indicating that the
volatile methyl jasmine was affecting the control plants. Tobacco and alfalfa plants that were
also exposed to MeJA treated plants by these researchers expressed proteinase inhibition.
Section 1.1.2 Indirect responses to herbivore damage
Analogous to direct response, indirect responses are also induced by herbivore attack.
However, indirect responses initiate predator attraction to the herbivore through chemical
signaling. Herbivore injured leaves emit volatile compounds of varying types and amounts. The
release of plant volatile compounds is a cyclic process occurring primarily on a diurnal cycle.
(Turlings et al., 1995). Many plants release six carbon (C6) green leaf volatiles upon the first
indication of herbivore injury (Turlings et al.,1995). Other plants, such as cotton (Gossypium
hirsutum L.), store compounds in the leaves which are released from lysegenous glands upon
injury (Rose et al., 1996). The stored compounds largely exist as the C6 volatiles and mono and
sesquiterpenes. As the injury continues, secondary metabolites are biosynthesized for up to three
days following the injury. Identified metabolites include α and β farnesene, (E) -4, 8 dimethyl-1,
3, 7 nonatriene and the terpenoid, linalool (Rose et al, 1996). Cucumber plants undergoing
attack by spider mites show an increase in the terpenes, (E)-β-ocimene and (E)-1, 4-dimethyl-1,

8
3, 7 –nonatriene. Comparatively, the green leaf volatiles, (Z)-3-hexen-1-yl, and (Z)-3-hexen-1-ol
produced by non–infested plants comprise 35 to 50% of total volatiles. These researchers
observed that chemical concentrations were reduced to approximately 1% of total volatile
emissions in spider mite infested plants. An increase in the production of terpenes peaking
several hours following the initial damage and coordinated with the photocycle, indicate the
plant‟s ability to optimize production of volatile emissions at the time when the predator is most
likely in search of prey (Turlings et al.,1995). These herbivore induced chemical signals allow
predators to seek their prey by honing in on the volatile blend being emitted by the plant and
discriminately choose their prey.
Section 1.2 Providing a warning
Plants under attack can benefit from the presence of associated plants by emitting odors
that attracted predators, repel herbivores or mask the attractive odor of a host plant (Price et al.,
1980). Baldwin and Schultz (1983) observed that mechanically damaged poplar ramets and
sugar maple seedling emitted phenolic compounds. Wounded seedlings placed in proximity to
undamaged poplar and sugar maple seedlings emitted these compounds at a higher rate than the
true controls and continued to emit phenolic compounds for approximately 75 hours after
exposure. The authors suggested that ethylene may have played a part in the transmission of
these signals, since it is a product of wounding in many species of plant and has the capability to
influence the plant‟s ability to biosynthesize these phenolic compounds. This was the first
observation that plants may be employing volatile chemicals as a mode of communication.

9
However, the concept of plant derived compounds providing signals to chemically cross-talk to
adjoining plants was largely written off to “methodological grounds” by many (Dicke et al.,
1990). This was due in part to a lack of supporting quantifiable data to illustrate that the plants
were emitting chemical compounds and poor understanding of intra-interspecies chemical
signaling. However, later investigations performed on cotton plants indicated that this transfer of
chemical-based information between plants could be assayed. Cotton plants sharing air space in
close proximity to herbivore injured cotton plants emit similar amounts of the homoterpene, (E,
E)-4, 8, 12-trimethyl-1, 3,7,11 tridecatraene (Rőse et al., 1996). This data indicates a possible
induction factor from the injured plant to the control indicating impending damage. In 2006,
Karban et al., performed a series of experiments with sagebrush (Artemisia tridentata) to
determine the effectiveness of the transfer of volatiles from wounded plants to conspecifics in the
area. They concluded that while volatiles do in fact transfer information from a single plant to
adjacent plants, airflow is necessary to carry the volatile signal both to nearby plants as well
from one section of the plant to another. These airborne signals can affect plants up to 60 cm
(approximately 2 feet) away from the source. This is an increase of their initial estimate from
their first trials where they reported that volatiles were only carried less than an inch away
(Karban et al., 2003).
Section 1.3 Attraction of predatory insects
While the stimulation of the proteinase inhibiting genes was found to be directly
correlated to the plant chemical emission (Ryan and Green, 1990) the full breadth of the effect of

10
plant volatiles on herbivores and their natural enemies has yet to be established. It was proposed
that the relationship between the releases of volatile semiochemicals (A chemical emitted by a
plant or animal that evokes a behavioral or physiological response in another organism) were the
direct result of injury by herbivores (Pare and Tumlinson 1996; Dicke, 1999). The release of
semiochemicals triggered the movement of prey movement and provided the subsequent
attractants to predators. Herbivores use the presence of these airborne natural products to aid in
the decision to stay in a specific location and feed or take off and search for food. These plant
volatile emissions are important in the movement of the predator as well. Predatory mites will
remain in an area where prey related volatiles exist until they are no longer detected, and can
distinguish between volatile semiochemicals related to different species. In a 1994 study of the
bird-cherry-oat aphid, Rhopalosiphum padi, the compound methyl salicylate was found to inhibit
the settling of R. padi aphids on barley (Hordeum sativum) (Poacaea) and spring wheat, (Tricium
aestivum), when the plants were sprayed with methyl salicylate. Petterson and co-workers
(1994), hypothesized that the compound may have acted as a repellent to R. Padi, inhibiting
colonization of the aphid, thereby preventing overfeeding. This information coincides with a
study performed by Campbell et al. (1993), where they noted that while methyl salicylate as part
of a volatile blend containing hexenal and β caryophyllene emitted from hops was attractive to
the hop aphid, Phorodon humuli, increased levels of methyl salicylate eliminated positive
response. It was noted by these investigators that these volatiles increased as a function of aphid
population, and it is thought to have the effect of repellence to prevent over-colonization of the
aphids, therefore limiting food supplies. Price and others (1980) first recognized that plants

11
often communicate with beneficial insects via chemical emissions that are attractive to these
insects. These emissions may prevent herbivory to an adjacent species by providing a repellent
or masking odor that reduce attack from insect pests, while attracting the predatory insect to
them (Dicke et al., 1990). In order to initiate this relationship, plants provide predators with a
distinct blend of chemicals that provide an odor trail to lead them to the injured plant. This
release is a distinct response to herbivore injury releasing volatiles that incite a systemic response
in the plant (Turlings, 1995). However, in order for these chemical cues to be beneficial to the
predator, Turlings suggest that signals must be clear enough to clearly seek out their prey without
interference from background odors to in a reliable manner.
Herbivore induced volatile chemical blends must be produced in sufficient amounts to
attract the predator, specific to the plant species, and genotype. Dicke (1999) found that
volatiles in the headspace volume of Cox Orange Pippin apples infested with the two spotted
spider mite (Tetranychus urticae) contained the terpenes (E)-β ocimene and (E, E) –α farnesene
representing 5% of the total blend, while the headspace analysis of Summer Red apples infested
by the same species exhibited terpenoid levels that represented 25% and 55% of the total volatile
blend respectively. Specialist parasitoid wasps can distinguish between non-host and prey
infested corn, cotton and tobacco plants by responding to herbivore induced volatile emissions
(Arimura et al., 2005). This task becomes more difficult when the herbivore feeds on several
different plant species. Pare and Tumlinson (1999) have observed that specific chemical blends
are associated with a positive interaction with the host, increasing the wasp‟s attraction to the
specific volatile blend. It has been suggested that the volatile blend emitted by plants undergoing

12
herbivore injury may not be the only method beneficial insects use to identify herbivore
presence. Environmental conditions, competition from other insect predators-enemies,dietary
needs or behavioral factors can affect a predator‟s response to plant volatiles, therefore
questioning the reliability of the herbivore induced volatile blend as a valid indicator of
herbivore presence on predator attractions (Dicke, 1999).
Section 1.4 Herbivore Induced Plant Volatiles and Plant Signaling
Plants are capable of emitting complex volatile chemical blends upon damage from
wounding. Wounding can occur from mechanical damage from ripping or tearing, or from the
chewing action of herbivorous insects. In research conducted by Turlings and Tumlinson
(1992), corn seedlings (Zea mays) were mechanically damaged in manner that mimics feeding
injury from caterpillars of the beet armyworm (Spodotera exigua) by scratching the leaves of the
seedling with a razor blade, and placing the regurgitant of the caterpillar into the wounded area.
The damaged leaves were removed and placed into a volatile collection device and the headspace
volatiles collected. Similarly, undamaged leaves from the same plant as well as control plants
were sampled and analyzed. The leaves damaged by the caterpillar regurgitant responded as if
caterpillar damage had occurred producing the terpenoid: linalool, and the terpene (E)-1, 4-
dimethyl-1, 3, 7–nonatriene and β-farnesene. Undamaged leaves from the same plants exhibited
production of (Z)-3-hexen-1-yl acetate and indole. These investigators report that the production
of the volatiles, (Z)-3-hexen-1-yl acetate and indole indicate that plant defenses are systemic,
and are induced by the damaged leaves themselves. In a 2000 study by Ozawa et al., lima beans

13
Phaseolus lunatus were infested with beet army worm Spodoptera exigua, the common army
worm, Mythinma separata, and the phytophagous mite, Tetranychus urticae to induce herbivore
volatiles. A second series of experiments using un-infested lima bean plants were conducted to
evaluate if chemical induction can be associated with insect feeding injury. Aqueous solutions of
methyl jasmonate (MeJA), a product of the jasmonic acid (JA) pathway, together with aqueous
and gaseous methyl salicylate (MeSA) concentrations (salicylic acid (SA) pathway product)
were assayed. It was earlier thought that the JA and SA pathways were mutually exclusive.
However, application of aqueous MeJA to leaves produced volatiles that mimicked damage
invoked by S. exigua and M. separata, while leaves treated with aqueous MeSA emitted volatiles
similar to those from T. urticae damaged leaves. Surprisingly, the application of gaseous MeSA
followed by aqueous MeJA and followed with a second application of gaseous MeSA, produced
volatiles mimicking those elicited by T. urticae. This information was exciting in the
identification of the biosynthesis of volatiles based on the method of predator attack. This
research indicates that chewing herbivores elicit the JA pathway, while phloem feeding insects
elicit the SA pathway.
Other investigations show that the corn leaf aphid Rhopalosiphum maidis has a distinct
preference for the odor of healthy corn seedlings over those that have been treated with the
sesquiterpene, β-farnesene when offered both odors in a y-tube olfactometer. These choices
were elicited without the aphids having any physical or visual contact with the plants. β-
farnesene is a known aphid repellent and the clean air sample was overwhelmingly chosen by
aphids regardless of gender or age (Bernasconi et al., 1998).

14
Section 1.5 Biosynthesis and induction
Plant species differ in their chemical response to herbivore damage. The chemical
composition of volatiles vary among the type of plant tissue (cotton leaves, bolls), varieties and
cultivars (de Moraes et al., 1998), genotype (Loughrin et al., 1995), attacking herbivore
(Takabayashi and Dicke 1996; Dicke, 1999) and even the developmental stage of the herbivore
(Takabayashi et al., 1995). European corn borer larvae (2nd and 3rd
instar Ostrinia nubilalis)
were allowed to feed on corn seedlings. Another set of plants experienced the same feeding with
older more experienced larvae (6th
instar). Headspace analysis of the plants fed on by the young
caterpillars revealed a volatile blend that included green leaf volatiles, while the plants fed upon
by the older caterpillars released volatiles that were not significantly different than those seen in
mechanical wounding (Takabayashi et al., 1999). This data conflicts with information by
Turlings et al., 1995; Pare et al. 1998 who demonstrated that plant response to mechanical
wounding produced a much different volatile blend than insect feeding. (de Moraes et al., 1998)
It is apparent that the species of plant determines the type and timing of herbivore
induced plant volatiles. Corn (Z. mays) releases C6 green leaf volatiles initially when under
insect attack, as do tomato, potato and tobacco and others (Tumlinson and Waecker, 2004).
Cotton plants immediately release the terpenes, α-pinene and β-caryophyllene. This is likely due
to the terpenes acting as toxins or repellents to discourage further attack by herbivores. Higher
concentrations of (Z)-3-hexen-1-ol, α-pinene, (Z)-3-hexenyl acetate and β-caryophyllene were
exhibited by cotton plants after 24 hrs following damage. (de Moraes et al., 1998).

15
1.6 Biosynthesis of herbivore induced plant volatiles
The production of herbivore induced volatiles has been shown to serve a number of
functions, including the attraction of predators, the repellence of herbivores, a means of
communication among plants and the production of antimicrobial compounds to aid the plant in
wound damage. The production of these secondary plant metabolites cannot be fully understood
until we gain a better understanding of the biosynthetic pathways and the relationship of these
pathways in plant defense. While earlier research suggested that volatile organic compounds
were carried by ethylene gas (Ryan et al., 1980), research in the mid to late 1990‟s indicates
volatile plant emissions were produced by one of the three biosynthetic pathways- the
lipoxygenase (octadecanoid) pathway-producing the green leaf volatiles (which are not
systemically released, but are produced by plants when freshly cut or damaged), the jasmonic
acid pathway (known to initiate the plant defense system), the isoprenoid pathway (which
produces terpenoids), and the shikimic acid/tryptophanic pathway which produces indole and
methyl salicylate (Pare and Tumlinson,1995).
Section 1.7 Jasmonic acid, green leaf volatiles and the octadecanoic pathway
Jasmonic acid (JA) is biosynthesized by many plants by the octadecanoid pathway. This
pathway produces the green leaf volatiles produced by corn and other plants immediately
following herbivore injury (Turlings et al., 1995; Figure 2).

16
JA has a dual role in the production of herbivore induced plant volatiles. As the end
product of this biosynthetic pathway, JA appears to be the key information pathway in plant
discrimination between mechanical wounding and herbivore injury. Exogenous application of
JA were found to elicit a volatile blend that more closely resemble that from herbivore damage
and produced defense gene initiation that is similar to observed production following herbivore
damage (de Bruxelles et al., 2001). Wild type lima bean plants treated with exogenous JA
effectively reacted to treatment that mimicked herbivore feeding, and suffered less from the
consequences of herbivore attack than control plants (Heil, 2004). JA has been shown to attract
predatory mites and prohibit embryonic development and increase egg mortality in the two
spotted mite, Tetranychus urticae on tomato plants (Ament et al., 2004) and JA increased the
attraction of the parasitic wasp Anagrus nilaparvatae to rice plants and decreased the settling
behavior of the brown leaf hopper Nilaparavata lugens. Volatile production from infestation
from N. lugens and the application of JA was shown to produce different volatile blends and it is
thought that N. lugens feeding does not trigger the JA pathway. This information indicates that
different pathways may cause the production of different volatile blends, however the resultant
volatiles are attractive to the parasitic wasp (Lou et al., 2005).
Research performed with cis-jasmone (3-methyl-(cis-2-penten-1-yl)-2-cyclopentyl-1-one)
indicates the compound is beneficial as an aphid repellent, and as an attractant for aphid
predators such as the mite eating ladybeetle, Stethorus punctum picipes. Laboratory trials and
electroanntenaography studies indicate that cis-jasmone is far better as an attractant for predators
than JA and further increases plant defense gene production over JA (Birkett et al., 2000). To

17
assess the possibility of the cis-jasmone on the predatory wasp, Cotesia marginventris, a known
parasitoid of the beet armyworm, wind tunnel tests were used to facilitate the wasps‟ choice
between control plants, mechanically damaged plants and those mechanically damaged and
treated (Turlings and Tumlinson, 1992). Overall these researchers found that the wasps showed
a distinct preference for the treated plants indicating the plant emissions distinctly attracted the
wasp. C6 Green leaf volatiles are the precursors for JA production. The C6 volatiles are normally
emitted at low levels by healthy plants and are released rapidly following herbivore or
mechanical damage. These compounds stimulate wound response gene production, reduce
reproductive rates in aphids and spider mites and reduce caterpillar feeding, and can be
attractants for the Colorado potato beetle and specialist aphids. C6 volatiles also have
antimicrobial and antifungal properties (Walling, 2000; Figure 3).
Mechanical damage from chewing insects elicits different response from that of sucking
or piercing insects. Phloem feeding insects such as aphids and whiteflies do not mechanically
damage the plant tissue in the same manner as do the chewing caterpillars and beetles. These
insects as well as those classified as cell content feeders, puncture the plant cells and rupture the
membranes. Since this method of feeding does not impact the plant in the same manner as the
chewing damage of most herbivores, the plant does not often recognize this insect attack, instead
it is perceived as a pathogen (Walling, 2000). The pathogen response initiates the JA pathway
and the SA pathways which stimulate the production of defense genes and provide secondary
metabolites such as MeSA, a known aphid repellent (Shulaev et al., 1997, Bernasconi et al.,
1998). The production of the volatiles MeJA and MeSA is the result of these types of herbivore

18
piercing injury. Corn seedlings (Z. mays) exposed to caterpillar induced green leaf volatiles, (Z)-
3- hexenal, (Z)-3-hexenol and (Z)-3-hexenyl acetate induced increased production of JA
(Engelberth et al., 2004).
The C6 green leaf volatiles can be used to determine the presence of pathogenic
bacterium or molds affecting plant species. Lima beans release the C6 volatiles 3-hexenol and
(E)-2-hexenal 15-24 hours following inoculation with Pseudnomona syringae pv.phaseolicoa
and pepper leaves release similar C6 aldehydes and alcohols following inoculation with the
bacterial pathogen Xanthanmonas campestris pv. vesicatoria (De Moraes et al., 2004). The
cassava green mite (Munonychelhs tanajoa ) feeds on the cassava plant. Cassava is an important
food crop in the tropics and losses due to the herbivore damage are costly. The mite‟s natural
enemy is an entomopathogenic fungus, Neozygites tanajoae, which is found on the plant and
attacks by the emission of spores which are fatal to the mite. A number of green leaf volatiles
affect the rapid sporulation of the fungi. These included Z-(3) –hexen-1-ol; the terpenes E-
(trans)β- ocimene and (E)-4,8 dimethyl 1,3,7 nonatriene and, the terpenoid linalool as well as
methyl salicylate (Hountondjietal et al., 2005).
Section 1.8 The isoprenoid pathway- the terpenes and the terpenoids
The majority of all volatiles systemically released by herbivore feeding are terpenes (Pare
and Tumlinson, 1996), The most commonly identified terpenes from herbivore injured plants
include (E,E)-4,8,12-trimethyl-1,3,7,11 tridecatraene (TDTT) and (E,E)-1,4-dimethyl-1,3,7 –
nonatriene. These compounds appear in the herbivore induced volatile blends of many plant

19
species including corn, lima beans, tomatoes, tobacco, cotton and others. Cotton plants
damaged by herbivores release relatively high amounts of the terpenes: (E,E) –α-farnesene, (E)-
β-ocimene, (E,E)-4,8,12-trimethyl-1,3,7,11 tridecatraene (TDTT) and (E,E)-1,4-dimethyl-1,3,7 –
nonatriene and the terpenoid, linalool. While herbivore wounding initiates the release of the
terpenoids, β-caryophyllene and α-pinene, which are stored in the leaves (Rodriguez-Saona et al.,
2002) the sesquiterpenoids such as linalool are released hours following continual insect injury.
This indicates that the compounds are biosynthesized via a pathway that allows the plant to
produce these compounds de novo to purportedly enhance the attraction of predators (Rőse et al.,
1996). Corn seedlings (Z. mays) produce a volatile blend while undergoing herbivore attack
from the pea aphid, Acyrthsiphon pisum that attracts the twelve spot lady beetle, Coleomegilla
maculata and the green lacewing, Chyrsoperla carnea (Zhu et al., 1999). The components of the
blend include the sesquiterpenes, β-farnesene and β-caryophyllene and the C6 volatile 3-hexen-
1-ol (Z). The above predator species strongly responded via electroanntenography to β-
farnesene above all others. This is likely due to the fact that this terpene is a known aphid alarm
signal, allowing the predators to seek out their prey by using the volatile as a scent trail (Zhu et
al., 1999).
Section 1.9 Methyl salicylate as a volatile semiochemical
Methyl salicylate (MeSA), the methyl ester of salicylic acid, another common plant
volatile was the subject of further research for its role in chemical signaling. In a 1997 study by
Shulaev et al., tobacco plants inoculated with the tobacco mosaic virus exposed to airborne

20
methyl salicylate displayed increased disease resistance. When these treated plants were housed
with control plants, the exposed plants transmitted signals that increased the production of the
defense mechanism gene in controls (Shulaev et al., 1997). This would indicate that plant
volatile production is a systemic response and is not simply located within the individual plant‟s
boundaries, and that they are capable of influencing like species of plant that are in the vicinity
of the threat. While both JA and SA play important roles in the defense of plants against
herbivore attack both directly and indirectly, there is a differing opinion in the literature on the
benefits of each. The JA pathway may inhibit the SA pathway which provides a separate
secondary pathway to the induction of defense genes in response to wounding (Walling, 2000).
SA levels are not affected by mechanical wounding, indicating that salicylic acid is a secondary
elicitor of defense gene production (Walling, 2000). Lima beans undergoing attack by the two
spotted spider mite (T. urticae) exhibited a different volatile response than leaves treated with the
exogenous application of JA (de Bruxelles et al., 2001). This response could not be replicated
until the application of methyl salicylate was applied or application of jasmonic acid followed by
methyl salicylate was used. This response indicates that some species such as (T.urticae) elicit a
duel response through the JA-SA pathway. Other recent research indicates that there may be an
antagonistic relationship between the JA and SA pathways. Traditionally, the SA defense
pathways are activated in defense of pathogenic attack, while JA dependent defenses are initiated
by herbivore wounding. The SA and JA pathways are considered to be mutually exclusive, each
providing its own defense mechanism. Recent evidence has shown that cross talk between the
pathways does exist and can be detrimental to the plant. Tomato plants exposed to BTH

21
(benzothiadiazol), a defense mimicking chemical, exhibited increased resistance to the pathogen
P. syringae, but increased optimum feeding conditions for the corn ear worm (Heliocoverpa zea)
(Piertse et al., 2001). In contrast, increased SA levels have been shown to lower resistance from
attack by spotted cucumber beetles while undergoing pathogen induced systemic acquired
resistance from the fungus, Colltotrichum orbiculare (Piertse et al., 2001).
Many predatory insect species including the predatory mite, Phytoseiulus persimilis, the
green lacewing, Chrysopa nigricornis and the predatory bug Anthocoris nemoralis are attracted
to MeSA (James, 2003b). MeSA baited traps were placed in juice grape vineyards and hop yards
by James and Price (2004) to assess the attractiveness of predators of two spotted mite (T.
urticae) and hop aphid (Phorodon humuli). Collection of both prey and predators were
performed using sticky bait traps, canopy shake and leaf sampling and then assessed. Four
families of insects including hover flies, wasps, dance flies and flesh flies along with a
significant number of parasitic wasps were trapped on the sticky traps in the methyl salicylate
baited vineyard blocks. The hop yard baited traps also yielded high numbers of the mite eating
ladybird beetle, S. punctum picipes and the predatory bug, Orius tristicolor. These numbers
coincided with a decrease in aphid and mite numbers. It was noted by James that predator
numbers reached the highest concentration in late July. This timing may be a function of the
higher airborne MeSA concentrations that occurred in the vineyard during spring that may have
been a deterrent to some predators (James and Price, 2004).

22
Section 2.0 Recent developments and future applications for herbivore induced plant
volatiles
Plant specific volatile emission blends have been proven to attract predatory
insects to food sources without visual or vibrational cues that may otherwise be an attractant.
But is the primary function of chemical signaling the communication between plant and
predator? Or are there multifaceted uses for chemical signaling? Is it possible that plants may
use chemical signaling as a warning to potential threat to “keep off”? Often times, the chemical
signaling serves a third and very distinct purpose as a repellent, therefore keeping potential
attackers or females seeking a place for their eggs at a safe distance. Plant volatiles may provide
parasitic wasps cues on the location of their hosts. The parasitic wasp, Trissolcus basalis, uses
plant volatiles to locate plants damaged by and containing egg masses of their prey, the southern
green stink bug, Nezara viridula. Undamaged leaves or egg masses alone were unattractive to
the wasp, as were newly hatched larvae. This information clearly links the production of plant
volatiles as a consequence of the specialized situation of herbivore damage and oviposition
(Colazza et al., 2003).
The research of the past twenty years has brought a new wave of interest. Could
biosynthetic volatiles be manufactured and combined and used to draw beneficial insects? The
need to perform field trials was understood to be the best way to assess the effect of volatiles on
predators. In a follow up field study of his 2003 and 2004 experiments, James (2005) tested
fifteen synthetic HIPV‟s in an open field and a hop yard in Washington State. The experiment

23
consisted of a number of sticky cards baited with the synthetic volatiles. James noted that eleven
different species and families of insects were attracted to thirteen of the synthetic volatiles.
It is apparent that identifying the specific attractant(s) for predatory and parasitic insects
can serve as a tool in integrated pest management. The recruitment of specialized natural
enemies can affect pest populations and reduce agricultural losses. Augmenting the biosynthesis
of plant production of specific volatiles, priming plants with volatile blends that are attractive to
predators, and the production of synthetic herbivore induced volatile blends are all being
investigated. To date, the mouse ear cress (Arabidposis thaliana) has been transgenically
modified to change the C6 green leaf volatile production following herbivore attack by cabbage
butterfly larvae (Pieris rapae), as well as the response to grey mold Botyris cinerea (Halitscke et
al., 2008). The parasitic wasp, Cotesia glomerata, is known to be attracted to the green leaf
volatiles (Z)-3-hexenal and (E)-2-hexenal. Following wounding, (Arabidposis) demonstrated a
thirty fold increase in the production of (Z)-3-hexenal following wounding (Shiojiri et al., 2006).
Two strains of the plant were genetically modified. One strain was modified to enhance C6
volatile production to twice the level of that of the control plants and the other modified to
suppress C6 volatile levels.(≈ 25% of controls). A similar study performed with genetically
volatile inhibited wild tobacco plants (Nicotiana attenuata) dramatically decreased attraction of
the generalist predator Geocoris pallens (Halitschke et al., 2008).
The use of herbivore induced plant volatiles to prime plants is attracting a great deal of
interest. The knowledge of the plants use of these signals and their uses in the application of
agricultural pest control are key areas of research for improving crop management and

24
improvement of plant defense. This concept has garnered worldwide attention and interest,
particularly in countries such as Brazil that rely heavily on agricultural products (Arab and
Bento, 2006). Manipulation of the expression of volatiles from plants as a means to attract
predatory insects is a beneficial tool in the control of insect pests, but more research is necessary.
Further research in crop plants must be preformed and the volatile chemical signals used must be
carefully selected to target only those species that effectively control the desired pest population,
as well as consideration of the effects that these volatiles may have upon neighboring species
(Turlings and Ton, 2006). In order to successfully use volatiles as a recruitment tool for pest
control and crop management, the plant must be able to identify and prioritize these signals to
their advantage (Choudary et al., 2008). For example, (Z)-3-hexenol, was found to be the
primary attractant for the parasitic wasp Opius dissitus, in the control of the pea leafminer,
Liriomyza huidobrensis, a pest that has invaded all zoogeographic regions and has a broad
variety of hosts (Wei et al., 2007).
Section 3 Recent controversies and trends in HIPV research
Section 3.1 HIPV’s act as priming agents for intra-plant signaling
Heil and Silva Bueno (2007) specifically identify the so-called green leaf volatiles as one
of the most important series of compounds for plant signaling In their estimation, the fact that
these compounds are gaseous and easily transported through the air aids in the contact of more
plant area than any of the other lesser volatile herbivore induced secondary metabolites. While
the question of benefits to the signaling plants fitness remains an important topic, it does appear

25
that within plant as well as within a community of plants, all reap some benefit from the
herbivore induced chemical communication. They hypothesize that C6 volatiles may be the
quickest method to intra-specifically prime the plant‟s defense system-even quicker than
systemic defense hormones JA and SA thereby improving the timing of response. Plant internal
signals would be much slower traveling through the xylem and phloem. In this investigation, the
authors‟ state that what has previously identified as plant to plant signaling is perhaps “plants
„eavesdropping‟ on what is within plant signaling worn on the outside.” In their 2008 study,
Frost et al., (2008) further questioned the “communication” aspect of herbivore induced
volatiles. It was noted that plants that receive the message of impending herbivory, by the
presence of herbivore induced plant volatiles begin the defense mechanisms necessary to ward
off the threat. But unlike the plant under attack, the receptor plant pays fewer costs to overall
fitness than the plant sending the message out. Furthermore, this study calls out the differences
between “priming” and “induction” as two separate functions, citing the application of C6
volatiles to tomato plants, which in turn triggered the release of a number of volatile terpenes,
but direct defenses were not measured and priming effects could not be inferred. Frost and co-
workers further clarified the difference by stating that “if induction of volatile compounds in
response to herbivory is a cry for help, the induction of volatiles in response to a volatile signal
may be more of a whisper, which appears to be correspondingly less attractive to predators”. In
a 2008 addendum to the article, Frost et al., further clarification on the specialization of HIPVs
can have on plant behavior was discussed. In particular, the C6 volatiles were previously
believed to provide the signal from plants under going herbivory as a “cry for help”. While

26
capable of stimulating both priming and defenses in a number of plant species, they do not
appear to convey “any context–dependent information” (Frost et al., 2008). The problem with
these compounds, as was pointed out by Heil and Bueno is that unlike the sesquiterpenes, the C6
volatiles are rapidly dispersed and quickly dissipated thus making them excellent messengers for
the interplant priming capabilities. Frost et al. (2008) hypothesized that the reason for this is the
fact that the C6 volatiles primary purpose is to act as an intra plant signal for priming. This
function alleviates the problem of conflicting information from nearby plant communities who
have different enemies releasing the same or similar HIPV‟s. But as the authors point out, the
HIPV‟s including the C6 volatiles are capable of being detected by parasitoids and predators, up
to tens of meters away, indicating that volatile transport distances may not be the limiting factor.
It is more likely a function of the receiving plants capability to detect and employ the signal at
some threshold concentration. Frost et. al. (2008) proposed that this threshold concentration
may vary widely due to the physiological differences in the ability of insects and plants to detect
and use these signals. This purported ability would allow the receiving plant to ignore the signals
of a distant plant–which may not be providing accurate information on impending herbivore
injury (Figure 5). This information appears to fall in line with Dicke and Baldwin (2009) who
proposed that perhaps the absence rather than the presence of a volatile signal may in fact
provide relevant information.

27
Section 3.2 Production of Herbivore Induced Plant Volatiles and Fitness
Consequences
An interesting concept that requires a great deal of further study is the question of the
fitness of the emitting plant, a topic that was recently addressed by Dicke and Baldwin (2009). .
The fitness of the HIPV emitting plant to date has never been assessed, likely due to the inability
to accurately measure the exchange of volatiles between plants in the field. While evidence
exists that both lima bean tendrils (Heil and Silva Bueno, 2007) and sagebrush (Karban et al.,
2003), exhibited increased defenses following exposure to volatiles from herbivore and
mechanically injured plants, the effect on the emitting plant still remains unanswered. More
likely, the emitting plant is more subject to herbivory than its uninjured neighbor, and improving
its neighbors fitness may in fact make it even more attractive to attack. Heil and Karban (2009)
recently posed the question “should the phenomenon therefore be termed „eavesdropping‟ rather
than „communication‟? Likely, the answer may be yes, since it is hypothesized that plant
signaling evolved from intra plant signaling and over time, plants may have developed receptors
to use the signal to benefit themselves and other within a community. Dicke and Baldwin (2010)
agree with this concept stating that until a HIPV “deaf” and “mute” plant species is developed it
will be difficult to discern the effects of HIPV‟s as a part of a community, and the overall effects
of the compounds holistic effect on the plant-rather than the individual tri-trophic interactions.

28
Section 4 Conclusion
The production of herbivore induced plant volatiles is an important area for research.
The understanding of the effects of the production of these compounds as a function of overall
plant fitness as well as the effects of the production of these compounds is an area for more
research.
The study of the production of these compounds reaches across many fields of interest;
chemistry, biology, ecology, entomology and agriculture. The increasing concerns with the
effects of agricultural chemicals on human health and the environment make this an important
topic for further research.
Recent information in the literature indicates a trend into the investigation of the
determination of the mechanisms of interplant signaling and the concepts of eavesdropping by
nearby plants. This is an area that is rapidly expanding in both basic and applied integrated pest
management research for conservation biology.

29
References
Ament, K., Kant, M.R., Sabelis, M.W., Haring, M.A., and Schuurink, R.C. 2004 Jasmonic
Acid is a Key Regulator of Spider Mite Induced Volatile Terpenoids and Methyl Salicylate
Emission in Tomato. Plant Phys. 135: 2025-2037
Arab, A. and Bento, J.M.S. 2006 Plant Volatiles: New Perspectives for Research in Brazil
Neotropical Ent. 35(2): 151-158.
Arimura G., Kost, C. and Boland,W. 2005 Herbivore–induced, indirect plant defenses Bio.
Biophys. Acta 1734:91-111
Bernasconi, M.L., Turlings, T.C.J., Amnrosetti, L., Bassetti, P. and
Dorn, S. 1998 Herbivore- Induced Emissions of Maize Volatiles Repel the Corn Leaf
Aphid, Rhopalosiphum maidis. Ent. Exp. App. 87: 133-142
Birkett, M.A., Campbell, C.A.M., Chamberlain K., Guerri, E., Hick, A.J., Martin, J.L.,
Mattheis, M., Napier, J.A., Petterson, J., Pickett, J.A., Poppy, G.M., Pon, E.M., Pye, B.J.,
Smart, L.E., Wadhams, L.J., and Woodcock,C. M. 2000 New Roles for cis-jasmone as an
Insect Semiochemical and in Plant Defense. Proc. Natl. Acad. Sci. 97(16) :9329-9334
Campbell, C.A.M., Petterson, J., Pickett, J.A., Wadhams, L.J. and Woodcock, C.M. 1993
Spring Migration of Damson-Hop Aphid, Phorodon Humuli (Homoptera, Aphididae), and
Summer Host Plant Derived Semiochemicals Released on Feeding. J. Chem. Ecol.19: 7
1569-1576
Choudary, D.K., Johri, B.N. and Prakash, A. 2008 Volatiles As Priming Agents that Initiate
Plant Growth and Defense Responses Current Sci. 94(5):595-604.
Collazza, S., Fucarino, A., Peri, E., Salerno, G., Conti, E., and Bin, F. 2004 Insect
Oviposition Induces Volatile Emissions in Herbaceous Plants that Attract Egg Parasitoids J.
Exp. Biol 207: 47-53
D‟Alessandro, M. and Turlings, T.C.J., 2005 In Situ Modification of Herbivore –Induced
Plant Odors: A Novel Approach to Study the Attractiveness of Volatile Organic Compounds
to Parasitic Wasps. Chem. Senses 30 :739-753
Das, P.D., Raina, R., Prasad, A.R., and Sen, A. 2007 Electroanntenogram responses of the
Potato Tuber Moth, Phtorimaea operullela (Lepidoptera;Gelichiidae) to Plant Volatiles. J
Biosci. 32(2):339-349

30
DeBoer, J.G., Hardijik, C.A., Posthumus, M.A. and Dicke, M. 2008 Prey and Non-Prey
Arthropods Sharing a Host Plant: Effects on Induced Volatile Emission and Predator
Attraction. J. Chem. Ecol.34: 281-290
deBruxelles, G.L. and Roberts, M.R 2001 Signals Regulating Multiple Responses to
Wounding and Herbivores. Crit. Reviews in Plant Sci. 20(5):487-521.
DeMoreaes, C.M., Lewis,W.J., Pare, P.W. Alborn, H.T and Tumlinson, J.H. 2004 Induced
Plant Signaling and its Implication for Environmental Sensing. J. Tox. Env. Health, Part A
67: 819-834
Dicke, M., Sabelis, M.W., Takabayashi, J., Bruin, J. and Posthumus, M.A. 1990 Plant
Strategies of Manipulating Predator-Prey Interactions Through Allelochemicals: Prospects
for Application in Pest Control. J. Chemical Ecol 16: 3091-3110.
Dicke, M. 1999 Are Herbivore-Induced Plant Volatiles Reliable Indicators of Herbivore
Identity to Finding Carnivorous Arthropods? Ent. Exp. App.91: 131-142.
Dicke M. and Hiller, M. 2003 Induced Plant Defenses: From Molecular Biology to
Evolutionary Ecology. Basic Appl. Ecol. 4: 3-14.
Dicke, M and Baldwin, I.T. 2009 The evolutionary context for herbivore –induced plant
volatiles: beyond the cry for help. Trends in Plant Science 15:3 167-175
Dicke, M., van Loon, J.J.A. and Soler, R. 2009 Chemical complexity of volatiles from
plants induced by multiple attacks. Nature Chemical Bio. 5:5 317-324
Engelberth, J., Alborn, H.T., Schemelz, E.A. and Tumlinson, J.H. 2003 Airborne Signals
Prime Plants Against Insect Herbivore Attack. Proc. Natl. Acad. Sci. 101(6):1781-1785.
Farmer, E.E. and Ryan, C.A. 1990 Interplant Communications: Airborne Methyl Jasmonate
Induces Synthesis of Proteinase Inhibition in Plant Leaves. Proc. Natl. Acad. Sci.87:7713-
7716.
Frost, C.J., Mescher, M.C., Carlson, J.E. and De Moraes, C.M. 2008 Plant Defense
Priming against Herbivores: Getting Ready for a Different Battle. Plant Physiology
146: 818-824
Frost, C.J., Mescher, M.C., Carlson, J.E. and DeMoraes, C.M. 2008 Why do distance
limitations exist on plant-plant signaling via airborne volatiles? Plant Signaling and
Behavior 3:7 466-468

31
Gershenzon, J. 2007 Plant Volatiles Carry Both Public and Private Messages. Proc. Natl.
Acad. Sci .104(13): 5257-5258
Halitscke, R, Stenberg, J.A., Kessler, D., Kessler, A. and Baldwin, I.T. 2008 Shared
Signals- „Alarm Calls‟ from Plants Increase Appparency to Herbivores and their Enemies in
Nature. Ecol.Letters 11: 24-34.
Heil, M. 2004 Induction of Two Indirect Defenses Benefits Lima Bean (Phaseolus
lunatus, Fabacaea) in Nature. J. Chemical Ecol 92: 527-536.
Heil, M and Karban, R. 2009 Explaining evolution of plant communication by airborne
signals. Trends in Ecology and Evolution 25:3 137-144.
Heil, M. and Silva Bueno, J.C.S. 2007 Within-plant Signaling by Volatiles Leads to
Induction and Priming of an Indirect Plant Defense in Nature. Proc. Natl. Acad. Sci.
104(13)5467-5472.
Heil, M and Silva Bueno, J.C.S. 2007 Herbivore –Induced Volatiles as Rapid Signals in
Systemic Plant Response. Plant Signaling and Behavior 2: 3 191-193
Holopainen, J. 2004 Multiple Functions of Inducible Plant Volatiles. Trends in Plant Sci.
9(11): 529-533.
Hountoondji, F.C.C., Sabelis, M.W., Hanna, R. and Janssen, R 2005 Herbivore-Induced
Plant Volatiles Trigger Sporulation in Entomopathogenic Fungi: The Case of Neozygites
tanajoae Infecting the Cassava Green Mite. J. Chemical Ecol 31(5)1003-1021.
James, D.G. 2003 Synthetic Herbivore-Induced Plant Volatiles as Field Attractants for
Beneficial Insects. Environ. Entomol. 32: 977-982.
James, D.G. 2005 Further Field Evaluations of Synthetic Herbivore –Induced Plant
Volatiles as Attractants for Beneficial Insects. J. Chemical Ecol 31 (3): 481-495
James, D.G. and Price, T.S. 2004 Field Testing of Methyl Salicylate for Recruitment and
Retention of Beneficial Insects in Grapes and Hops. J. Chemical Ecol 30 (8): 1613-1628.
Karban, R., Maron, J., Felton, G.W., Ervin, G. and Eichenseer, H. 2003 Herbivore damage
to sagebrush induces resistance in wild tobacco, evidence for eavesdropping between plants.
Oikos 100:325-332.

32
Karban, R., Shiojiri, K., Huntzinger, M. and McCall, A.C. 2006 Damage Induced
Resistance in Sagebrush: Volatiles Are Key to Intra- and Interplant Communication
Ecology 87 (4) 922-930.
Kessler, A. and Baldwin, I.T. 2004 Herbivore-Induced Plant Vaccination, Part I.: The
Orchestration of Plant Defenses and their Fitness Consequences in the Wild Tobacco
Nicotiana attenuate. The Plant J. 38:639-649.
Korth, K.L. 2003 Profiling the Response of Plants to Herbivorous Insects. Genome Biol.
4(7) 221-225.
Lou,Y.G., Du, M.H., Turlings, T.C.J., Cheng, J.A. and Shan, W.F. 2005 Exogenous
Applications of Jasmonic Acid Induces Volatile Emissions in Rice and Enhances Parasitism
of Nilaparvata lugens Eggs by the Parasitoid Anagrus nilaparvatae. J. Chemical Ecol 31
(9): 1985-2002
Maeda, T. and Takabayashi, J. 2001 Production of Herbivore-Induced Plant Volatiles and
their Attractiveness to Phytoseius persimilis (Acari:Phytoseiidae) with Changes of
Tetranychus urticae(Acari: Tetranychidae) Density on a Plant. Appl. Entomol.Zool. 36(1)
47-52.
Margolies, D.C., Sabelis, M.W. and Boyer, Jr., J.E. 1997 Response of a Phytoseiid
Predator to Herbivore-Induced Plant Volatiles: Selection on Attraction and Effect on Prey
Exploitation. J. Insect Behavior 10(5): 695-709.
Ozawa, R., Arimura, G., Takabayashi, J.,Shimoda, T. and Takaaki, N. 2000 Involvement
of Jasmonate and Salicylate –Related Signaling Pathways for the Production of Specific
Herbivore Induced Volatiles in Plants. Plant Cell Phyisol. 41(4): 391-398.
Paré, P.W. and Tumlinson, J.H. 1996 Plant Volatile Signals in Response to Herbivore
Feeding. Florida Ent. 79(2) 93-103.
Paré, P.W. and Tumlinson, J.H. 1997 De Novo Biosynthesis of Volatiles Induced By
Insect Herbivory in Cotton Plants. Plant Physiol.114: 1161-1167.
Paré, P.W., Alborn, H.T. and Tumlinson, J.H. 1998 Concerted Biosynthesis of an Insect
Elicitor of Plant Volatiles. Proc. Natl. Acad. Sci. 95: 13971-13975.
Paré, P.W. and Tumlinson, J.H. 1999 Plant Volatiles as Defense Against Insect
Herbivores. Plant Physiol.121: 325-331.

33
Pettersson, J., Pickett, J.A., Pye, B.J., Quiroz, A., Smart, L.E., Wadhams, L.J., and
Woodcock, C.M. 1994 Winter Host Reduces Colonization by Bird-Oat-Cherry Aphid,
Rhopalosiphum padi (L.) (Homoptera, Aphididae) and Other Aphids in Cereal Fields J.
Chem. Ecol. 20(10): 2565-2574.
Pieterse, C.M.J., Ton, J and Van Loon, L.C. 2001 Cross-Talk Between Plant Defense
Signaling Pathways: Boost or Burden? AgBiotechNet 3:1-8.
Pickett, J.A. Rasmussen, H.B., Woodcock, C.M., Matthes, M. and Napier, J.A. 2003 Plant
Stress Signaling: Understanding and Exploiting Plant-Plant Interactions. Biochem. Soc.
Trans. 31: 123-127
Popof, B., Stange, G. and Abrell, L. 2005 Volatile Organic Compounds as Signals in a
Plant-Herbivore System: Electrophysiological Responses in Olfactory Sensilla of the Moth
Cactoblastis cactorum. Chem Senses 30: 51-68.
Price, P.W., Bouton, C.E., Gross, P., McPheron, B.A., Thompson, J.N. and Weis, A.E. 1980
Interaction Among Three Trophic Levels: Influence of Plants on Interactions Between Insect
Herbivores and Natural Enemies. Ann. Rev Ecol. Syst.11: 41-65.
Rodriguez-Saona, C.R, Crafts-Bradner, S.J., Williams III, L. and Paré, P.W. 2002 Lygus
hesperus Feeding and Salivary Gland Extracts Induce Volatile Emissions in Plants. J.
Chem. Ecol. 28(9): 1733-1747.
Rodriguez-Saona, and Frost, C.J. 2009 New evidence for a multi-functional role of
herbivore induced volatiles in defense against herbivores. Plant Signaling and Behavior 5:1
58-60
Röse, U.S.R., Manukian, A., Heath, R.R. and Tumlinson, J.H. 1996 Volatile
Semiochemicals Released from Undamaged Cotton Leaves: A Systemic Response of Living
Plants to Caterpillar Damage. Plant Physiol. 111:487-495.
Shijori, K., Kishimoto, K., Ozawa, R., Kugimiya, S., Urashimo, S., Arimura, G., Horiuchi, J.,
Nishioka, T., Matsui, K. and Takabayashi, J. 2006 Changing Green Leaf Volatile
Biosynthesis in Plants: An Approach for Improving Plant Resistance Against Both
Herbivores and Pathogens. Proc. Natl. Acad. Sci. 103(45): 16672-16676.
Shulaev, V., Silverman, P. and Rasskin, I. 2005 Airborne Signaling by Methyl Salicylate
in Plant Pathogen Resistance Nature 385: 718-721.

34
Takabayashi, J., Takahashi, M., Dicke, M., and Posthumus, M.A. 1995 Developmental
Stage of the Herbivore Pseudaletia separate Affects Production of Herbivore-Induced
Synomone by Corn Plants. J. Chem. Ecol. 21:273-278.
Takabayashi, J. and Dicke, M. 1996 Plant-Carnivore Mutualism Through Herbivore-
Induced Carnivore Attractant. Trends Plant Sci. 1: 109-113.
Turlings, T.C.J. and Tumlinson, J.H. 1992 Systemic Release of Chemical Signals by
Herbivore-Injured Corn. Proc. Natl. Acad. Sci. 89: 8399-8402.
Turlings, T.C.J., Loughrin, J.H., Mc Call, P.J., Röse, U.S.R, Lewis, W.J. and Tumlinson, J.H.
1995 How Caterpillar-damaged Plants Protect Themselves by Attracting Parasitic Wasps.
Proc. Natl. Acad. Sci. 92: 4169-4174.
Turlings, T.C.J., and Wäckers, F. 2004 Recruitment of Predators and Parasitoids by
Herbivore-injured Plants. In Advances in Insect Chemical Ecology (R.T. Cardế and J.G.
Miller eds.) Cambridge University Press pp.21-75.
Turlings, T.C.J. and Ton, J. 2006. Exploiting Scents of Distress: The Prospect of
Manipulating Herbivore-Induced Plant Odors to Enhance the Control of Agricultural Pests.
Current Opinion in Plant Biology 9: 421-427.
van Poecke, R.M.P. and Dicke, M. 2002. Induced Parasitoid Attraction by Arabidopsis
thaliana: Involvement of the Octadecanoid and Salicylic Acid Pathway. J. Exp. Botany
53(375): 1793-1799.
Walling, L.L. 2000 The Myriad Plant Responses to Herbivores. J. Plant Growth Regul.
19: 195-216.
Wei, J., Wang, L., Zhu, J., Zhang, S., Nandi, O.I. and Kang, L. 2007 Plants Attract
Parasitic Wasps to Defend Themselves Against Insect Pests by Releasing Hexenol.
PLoS ONE 2(9): 1-7.
Zhu, J., Cosse, A.A., Obryckii, J.J., Boo, K.S. and Baker, T.C. 1999 Olfactory Reactions
of the Twelve Spotted Lady Beetle, Coleomegilla maculata And the Green Lacewing,
Chrysoperla carnea to Semiochemicals Released From Their Prey and Host Plant:
Electroanntenogram and Behavioral Responses. J. Chem. Ecol. 25(5) :1163-1177
35
Figure 1: History of research on airborne plant-plant signaling from Karban and Heil

36
Figure 2 Impact of C6 Volatiles on Herbivore Plant Interaction from Walling, 2000

37
Figure 3:Volatile compounds from corn seedlings (Zea mays) undamaged (A), first hour after
feeding(B) and several hours after feeding (C) Turlings and Tumlinson, 1992

38
Figure 4: Factors causing and traits affected by volatile-mediated signaling among plants
from Karban and Heil, 2010

39
Figure 5: Relative distances for HIPV emissions from Frost et al., 2008
A schematic representing differences in the relative distances over
which parasitoids and plants can respond to herbivore-induced volatile
(HIPV) emissions. The herbivore-wounded plant (far left) is wounded by
herbivores and releases HIPVs (represented by gray arrow). Based on our
recent work and work from other systems, systemic regions of the wounded
plant can respond (☑) to these HIPVs. Undamaged conspecific and heterospecific
neighboring plants close to the wounded plant may also respond (?)
to the HIPVs in what has been termed „eavesdropping‟. However, parasitoids
evidently respond to HIPVs from a greater distance than do plants. A better
understanding of the mechanisms of HIPV reception in plants is required to
understand the basis for such apparent distance limitations on plant-to-plant signaling

40
Chapter 2
Assessment of Herbivore Induced Plant Volatiles in Juvenile Hops
by Exposure to Methyl Salicylate

41
INTRODUCTION
Methyl salicylate (MeSA), the methyl ester of salicylic acid, is a secondary plant
metabolite (Hardie et al.,1994, Pare and Tumlinson 1996 and Walling 2000) MeSA has been
found to be an initiator for the production of defense genes against pathogens (Shulaev et. al,
1997) as well as an airborne plant signaling compound (Dicke et al., 1990; Bernasconi et al.,
1998, Hardie et al., 1994, Pickett et al., 2003, Karban et al., 2003, Kessler and Baldwin, 2001,
Arimura et al., 2005, Ozawa et al., 2000). Along with MeSA, a number of other volatile
compounds are released when plants are damaged by feeding from herbivores. These compounds
commonly known as herbivore induced plant volatiles (HIPV)‟s are believed to provide
communication between plants giving warnings of possible insect infestation, allowing plants to
defend themselves (Choudhary et. al. 1998, Dicke, 1998, Dicke 1999, Karban and Baldwin
1997).
MeSA has also been identified as an attractant for beneficial insects (Turlings and Ton,
2006, Turlings and Waecker, 2004; Zhu et al., 2007; James 2003, 2003a; James and Price, 2004;
Heil and Bueno, 2007) including the predatory mite, Phytoselius persimilis, the green lacewing,
Chrysopa nigricornis, and the predatory bug, Anthocoris nemoralis (James 2003a, James and
Price 2004, Hardie et. al 1994). Olfactory studies performed by Hardie et al., 1994 and
Bernasconi et al., 1998 indicate that MeSA also repels several aphid species including the black
bean aphid, Aphis fabae.
The specific use of MeSA as a signaling attractant of beneficial insects has received
attention but has not been thoroughly investigated. To examine the use of MeSA as an attractant

42
to beneficial insects, James and Price, (2004) placed MeSA dispensers in a 10 ha juice grape
(var. Concord) vineyard and a ca. 1 ha hop yard during the growing season. Analysis of
populations of predatory and herbivorous arthropods demonstrated a sharp rise in beneficial
insects in MeSA treatment compared to untreated crop areas. As a follow-up to field
examinations of James, we focused on a controlled environment-chamber study design to
address the following questions: (1) does MeSA exposure initiate intra-plant production of
herbivore induced volatiles?, and (2) can exposure to MeSA stimulate inter-plant signaling? To
address the first question, juvenile hop plants were exposed for 72 hours to dispensers emitting
MeSA then evaluated to determine if production of HIPVs could be stimulated. To address our
second question, the MeSA exposed hop plants were combined with non-exposed control hops
plants to assay if volatile emissions from the treated plants could in turn stimulate HIPV
production in control plants. HIPV emission profiles were obtained from glass chambers by
sampling the air headspace volatiles of exposed, combined, and control hop plants using space
solid phase microextraction (SPME). Mass spectral assessment of chromatographically resolved
volatile emission peaks were performed using gas chromatography–mass spectroscopy (GC-
MS).
METHODS and MATERIALS
Growing conditions for MeSA exposed and non-exposed control hop plants: Juvenile hops
(Humulus lupulus L.) Var. Chinook were grown from rhizomes under greenhouse conditions
(16:8 day:night, average temperatures 18.9-36.7 ºC) for the 2007 and the first three experiments

43
of 2008 . Hop plants were exposed to MeSA when they reached a height of approximately 10-12
cm (ca. 30 day growth). In the later experiments of 2008 and in the 2009 trials, juvenile hops
grown from root cuttings were used. These root-cutting plants were ca. 60 days old before
exposure to MeSA. Root cuttings were grown in environmentally controlled growth chambers
at 25 ◦ C and an 8:16 L: D photoperiod.
Plant Treatment: For all evaluations, Hop plants were separated into control and treated
environmental growth systems. In 2007 greenhouse evaluations, single plants were exposed to
air concentrations of methyl salicylate from impregnated sachets for approximately 72 hrs.
Following exposure, both treated and control plants (from separate greenhouses) were enclosed
in separate 2L borosilicate glass chambers (Figure 1). Two slotted 10 cm x 10 cm aluminum
base plates were aligned to minimize ambient air leakage and possible cross-contamination. The
glass chambers were then strapped down to secure them to the base plates. The chambered
plants were transported to the Washington State University –Tri Cities Food and Environmental
Quality Lab (FEQL) for SPME GC/MS spectral determination of chamber head space volatiles.
The experiment was replicated 4 times in 2007, 6 times in 2008 and three times in 2009.

44
Figure 1: Juvenile Hops in head space chambers
Volatile Collection and Analysis In all trials, the 2L plant-chamber systems were allowed to
equilibrate to room temperature ca. 2 hrs before the first series of headspace evaluations
(Experiment 1). The 2L glass chambers were outfitted with septa to allow collection of volatiles
by solid phase micro extraction (SPME; Figure 2). After 48 hrs, the control and treated plants
were placed into a common 10 L glass container and the two plants remained side-by-side for 48
hrs, at which time they were separated and placed into individual chambers (Experiment 2).
These separated chambered plants were allowed to equilibrate ca. 4 hrs before headspace SPME
sampling to examine possible inter-plant chemical signaling.

45
Figure 2: SPME sampling septa
A Carbowax-DVB film SPME fiber (Supelco, PA, USA) was inserted into the septa and
exposed to the headspace air for 5 minutes. The SPME fiber was removed then immediately
thermally desorbed at 200 C for 30 seconds into the injection port of an Agilent 6890 Gas
Chromatograph connected to a 5973 Mass Spectrometer. Analyses were duplicated at each
timed interval and performed on both treated and control plants. All analyses were performed
using the Agilent 6890 Gas Chromatograph with the 5973 Mass Spectrometer. Compounds were
desorbed into the injection port using a pulse splitless mode at 20⁰ C into an Alltech EC-WAX
column (30m x 0.32 mm I.D. x 0.25 μM film thickness). After an initial column temperature of
50 ⁰C for one minute, the temperature was raised 5
⁰ C/minute to a final temperature of 260
⁰ C for

46
5 minutes. The column was directly coupled to the ion source of the Agilent 5973 Mass
Spectrometer which was operating in EI (electron ionization) mode scanning from mass 50 to
350 amu at 2.5 scans/ sec. Spectra for the resolved compounds were searched using the National
Institute of Standards Technology (NIST) 98 library where they were compared for best match.
The percentage of total area counts associated with each individual resolved peak was noted and
compared against total area count among all combined peak areas in an attempt to chemically
profile the relative contribution of each compound spectrally identified. It is important to note
that the area counts are not used for quantification of the identified compounds, but as a means to
characterize the instrument response at each interval and to compare the response between the
treated and control plants at each sampling interval.
Results
Table 1 and 2 summarize the proportional contributions of HIVPs emanating from
unexposed control and MeSA exposed juvenile hop plants. Table 1 provides profile data for
plants exposed or not exposed to MeSA (Experiment 1), while Table 2 provides profile data for
non-exposed plants from the first experiment after combination with the MeSA-treated plants
(Experiment 2). Figure 1 chromatographically displays separation of 6 of the 16 reported HIPVs
observed during the course of this study. Figure 2 shows the differences in identified HIPV
emissions from juvenile control plants raised as rhizomes (2007-2008) and from root cuttings
(2008-2009).

47
Experiment 1: 2007 evaluations of volatiles from MeSA exposed hops
During March-May 2007, a total of four sets of juvenile hops (treatments and controls)
raised under greenhouse conditions were sampled and the headspace volatiles analyzed after
exposure to MeSA. Each area count in Table 1 represents the average response from duplicate
evaluations conducted for each interval treated and control chamber sample.
Controls: A total of ten compounds were identified in the head space analysis. The
control plant volatile blend consisted largely of the green leaf volatile 3-hexen-1-ol, (Z) acetate,
(ca. 34% of total area counts), and the alkyl aldehydes, nonanal and decanal (ca. 3% and 4%,
respectively). Low to moderate levels of the sesquiterpenes, iso-caryophyllene, α–caryophyllene
and α – farnesene were also present (ca. 2%, 6% and 2%, respectively). Best match spectra
identified germacrene B at 3%. Methyl salicylate comprised ca. 4% of total area counts.
Treated: The sesquiterpene, (E, E) α–farnesene showed a three-fold higher level (2% -
6%) than in the control. Methyl salicylate comprised ca. 17% of total area counts, > 4 times
higher than the control. Total area counts were lower in 3-hexen-1-ol, (Z) acetate (ca. 25%),
while the nonanal area counts were slightly higher at ca. 7%. Decanal counts were slightly lower
at 2%. Overall levels of α–caryophyllene were the same as the control (6%), while iso-
caryophyllene levels decreased slightly (1%).
Experiment 1: 2008 evaluations of volatiles from MeSA exposed hops
A series of six sets of control and MeSA exposed treated chamber evaluations was
conducted in 2008. In the first three trials, the juvenile hops were grown under similar

48
greenhouse conditions as in 2007. However, in the latter trials of 2008, (9/20-10/2), hop plants
were grown in individual growth chambers to exclude thrips injury to foliage. All other
handling, including sampling and analysis were performed as before.
Controls: A total of ten volatile compounds were identified. Volatiles from control plants
showed lower amounts of 3-hexen-1-ol, (Z) acetate (21 % of total area counts), α-farnesene (2%)
and α- caryophyllene (4%) compared to 2007 control plants. Several long chain fatty acids:
nonanoic acid (ca 10%), tetradecanoic acid (ca.3%) and the fatty acid alcohol, dodecanol (ca.
4%), as well as the sesquiterpene germacrene B (ca.3%), were present. A C6 volatile, 3-hexen-
1-ol, contributed trace (ca.<1% ) amounts to total counts. MeSA contributions comprised ca. 3%
of total area counts.
Treated: Nine compounds were identified in the treated plant head space analysis. The
contribution of 3-hexen-1-ol, (Z), acetate (ca. 25 to 29%) was slightly higher than in the control
plant. The sesquiterpene, germacrene D (ca. 2%), MeSA (7%) and α-farnesene (ca. 1%) were
also higher than the control. Spectral contributions from α–caryophyllene (trace), tetradecanoic
acid (ca. 1%) and nonanoic acid (trace) were lower than obtained with the control plant.
Experiment 1: 2009 evaluations of volatiles from MeSA exposed hops
Control: A total of five compounds were identified. The C6 volatiles 3-hexen-1-ol, (Z),
acetate and 3-hexen-1-ol proportionally contributed ca. 36 % and 2% to the total area counts,
respectively. The sesquiterpene, α-caryophyllene contributed ca. 3 % and β-caryophylllene, was

49
present in trace amounts. α-farnesene, contributed ca. 4% and MeSA contributed ca. 3% to the
total area counts.
Treated: A total of six compounds were identified. The C6 volatiles 3-hexen-1-ol, (Z),
acetate and 3-hexen-1-ol contributed 41 and 2% to the area counts, respectively. α-
caryophyllene occurred at trace levels while MeSA was not detected. α-farnesene was present at
ca. 5 % of total area counts. Figure 3
Experiment 2: 2007-2009 evaluations of volatiles from unexposed hops after exposure to hops
previously treated with MeSA.
To determine if MeSA exposed hops use the elevated expressions of volatile signals to
provide a conspecific plant with information regarding potential herbivore attack, we combined
the control and treated hops from the first experiment commencing in the latter trials of 2007.
In 2007, volatiles from control plants (i.e. the control plants from the earlier experiment)
following exposure to the MeSA treated plants comprised five compounds: 3-hexen-1-ol, Z
acetate: nonanal, decanal, iso-caryophyllene and α-caryophyllene. These compounds yielded 2%,
9%, 21%, 4% and 6 % respectively to the total area counts, with all but 3-hexen-1-ol, Z acetate;
and α-caryophyllene showing elevated levels compared to the levels prior to exposure to the
treated plant. α-farnesene, MeSA and the fatty acids, nonanoic and tetradecanoic acid, as well as
dodecanol were non-detectable. Analysis of volatiles from the MeSA-treated plants showed
higher levels of 3-hexen-1-ol, (z) acetate (ca.12%) and nonanal (ca..10%) than the control plants
but lower levels of decanal (ca.19%) α –caryophyllene (ca. 3%) and iso caryophyllene (ca2%).

50
In 2008 higher levels of α –caryophyllene (ca. 23%), MeSA (ca. 4%) and nonanoic acid
(ca. 10%) occurred in volatiles from the control plants after exposure to the MeSA treated plants,
compared to previous levels. Levels of the C6 volatile 3-hexen-1-ol, (Z) acetate (ca. 20%) and
3-hexen-1-ol (ca. 5%) were similar to previous levels.
In 2009 levels of α-caryophyllene (ca. 25%) and β caryophyllene (9%) were higher in
the control plants after exposure to MeSA treated plants than before exposure (ca. 4% and not
detected, respectively). In the MeSA-treated plants the presence of β-caryophyllene (ca.10%), α-
farnesene (ca. 8%) and α-caryophyllene (ca 29%) were notable.
Discussion
The data provided here suggest that young hop plants exposed to airborne MeSA are
stimulated to produce certain volatiles at elevated levels compared to non-exposed plants. Some
of these volatiles may be attractants for predators and parasitoids of hop plant herbivores. In our
experiments hop plants were not damaged by herbivores, simply exposed to one of the major
volatiles emitted by plants (including hops) when attacked by herbivores. Thus, the volatile
responses we have demonstrated may be part of „defense priming‟ rather than full defense.
Defense priming is a process in which the responses to an anticipated challenge (mediated by
volatiles from neighboring plants suffering attack) from a herbivore or a pathogen, are initiated
(Engleberth, 2006). Intra plant defense priming has been observed in lima beans (Phaseolus
lunatus) (Heil and Silva Bueno, 2007; Ozawa et al., 2000). The process of priming allows the
receiving plant to begin the production of systemic defenses without seriously affecting plant

51
fitness. This allows the plant to keep the necessary resources in reserve for full defense
production in the event of actual herbivore damage (Karban and Heil, 2009). Plants are also
capable of priming conspecifics (Karban et al., 2003; Heil and Silva Bueno, 2007), thereby
establishing the interspecific communication network to allow the receiving plant to begin the
production of systemic defenses in anticipation of herbivore attack.
Volatile production in juvenile hop plants exposed to airborne MeSA
Spectral evaluation of the head space SPME of MeSA treated hop plants identified
sixteen compounds previously associated with herbivore damage. In 2007, analyses of head
space volatiles revealed an increase in average area counts of three HIPV‟s : nonanal, (E,E) α
farnesene and MeSA. Contributions to total area counts from 3-hexen-1-ol (Z) acetate, decreased
slightly in the treated plant but increased contributions from this compound occurred in 2008 and
2009. Another volatile associated with herbivore injury, 3-hexen-1-ol was detected in 2009. All
of these compounds have been identified as part of the initial “call to arms” signal of plants
undergoing herbivore damage (Engelberth, 2006)
While the increase in the aliphatic aldehyde, nonanal, showed a greater than two fold
increase over control plants and MeSA production was > four times that of control plants, the
increased production of the sesquiterpene (E,E) α-farnesene was particularly notable. In 2007
(E,E) α-farnesene levels in MeSA treated plants were three times higher than in control plants
(ca 2% to 6%), a trend that also occurred in 2008 and 2009 (levels of 1 and 5% in treated plants,
not detected in control plants).

52
(E,E) α-farnesene was identified in head space analysis of lima beans (P. lunatus)
damaged by the two-spotted spider mite (T. urticae) (Ozawa et al., 2000), as well as in the head
space of Psylla infested pears (Pyrus communis L) (Scutareanu et al., 1997). Farnesene was
shown to be an olfactory stimulant or attractant for at least two species of predatory ladybird
beetles (Zhu et al. 1999; Francis et al., 2004), a predatory bug (Scutareanu et al., 1997) and a
predatory mite (Kong et al., 2005), and it is likely that this compound plays a role in attracting
natural enemies of pest arthropods. Farnesene is also produced in aphids as an alarm pheromone
and repels conspecifics (Pickett et a;. 1992).
While values for all of the identified compounds were inconsistent across the three trials,
the data indicate notable differences in emission profiles of non exposed control plants and
MeSA treated plants. All of the compounds we have identified in hop plant emissions have also
been found in the volatile blends from corn (Turlings and Tumlinson, 1992); lima bean (Shimoda
et al. 2000, Dicke et al., 1990); and pear (Scutareaneu et al., 1996), following herbivore injury.
The use of MeSA in these experiments was to attempt to prime hop plants to begin the
production of HIPV‟s . A review of the existing literature (Ozawa et.al, 2000) indicates that
MeSA can prime plants to begin the process of stepping up their defense mechanisms, and as a
result of the initiation of this mechanism, begin the increased production of volatiles related to
herbivore injury. We believe that the data provided here illustrates the ability of MeSA to initiate
the production of HIPV‟s in juvenile hop plants.

53
Volatile production in hop plants confined with conspecifics previously exposed to MeSA
In Experiment 2, control hop plants from the first experiment were combined for 48 hrs
with MeSA treated hops from the same experiment. Our goal was to test the hypothesis that a
„naive‟ hop plant can „eavesdrop‟ on volatiles from a conspecific previously exposed to MeSA,
and produce its own blend of volatiles. Our data suggest that a hop plant can indeed respond to
the bouquet of volatiles produced by another hop plant that has been stimulated by exposure to
MeSA. This response is characterized by production of volatiles that may prime the plant for
defense against herbivores or provide chemical signals for attracting natural enemies of
herbivores. The volatiles produced by eavesdropping hop plants at elevated levels (4-25 X) were
decanal, iso-caryophyllene, α-caryophyllene and methyl salicylate. Methyl salicylate is well
established as an attractant for a number of natural enemy species (James, 2003; James and
Price, 2004) and caryophyllene has been reported as an attractant for the green lacewing,
Chrysoperla carnea (Flint et al., 1979).
Variability in Composition of Volatiles Produced
A number of environmental issues and problems as well as the type of SPME fiber we
used may have contributed to data inconsistencies seen during the three years of this study. For
example, possible airflow contamination between greenhouses, occasional pest incidence and
differences in plant ages may have contributed to poor reproducibility. Herbivore induced plant
volatiles differ greatly in type and amount as a function of the growth age of the plant. We used
hops grown from rhizomes with a leaf age of ca. thirty days in 2007 and the first three trials in

54
2008. In the later trials of 2008 and in 2009, hops grown from soft wood cuttings, with a leaf
age of ca. sixty days were used. In studies on soybeans (Glycine max) and pears (Pyrus
communis L.) younger plants produced higher levels of fatty esters, aldehydes and alcohols as
well as sesquiterpenes such as (E,E) α-farnesene and β-caryophyllene. (Zhu et al., 2005,
Scutareanu et al., 1997). Krofta and Nesvadba (2005) noted that the variety of hop, hop age and
type of matrix sampled can affect the volatile blend. Our data on volatiles released according to
plant age (Figure 2) support this assumption and this should be considered in future research.
The use of the growth chamber in the later trials of 2008 and in 2009 may also have had
some effect on volatile production. Treatments that limited or reduced air flow between plants
prevented induced resistance in sagebrush (Artemesia tridentata) (Karban et al., 2006).
Similarly, Heil and Silva Bueno (2007) noted that intra plant communication in lima beans ( P.
lunatus L.) was also affected by the lack of air flow. They hypothesized that lianas such as the
lima bean have large areas of separation between plant areas and that air flow was necessary to
provide the access to the volatiles to begin systemic defense. While not a liana, hops do have a
similar configuration, raising the question of the effects of the importance of air flow to provide
volatile signals capable of reaching areas a significant distance from the source of herbivore
injury.
Krofta and Nesvadba (2005) reviewed an analytical procedure to assess the use of SPME
to isolate hop oils from different matrices. β-pinene, myrcene, limonene, linalool, gerianol, β-
caryophyllene, α-humulene, β-farnesene, MeSA and γ-cadinene were identified in HS-SPME
analysis of hops. Several types of SPME fibers including PDMS (polydimethylsiloxane) of

55
various sizes were used to analyze hop essential oils. In the method development of head space
SPME analysis for this study, we investigated several types of SPME fiber for optimal adsorbent
capability including 7μm and 100 μm polydimethylsiloxane (PDMS) fibers, as well as a 75 μm
Carboxen/polydimethylsiloxane and a 75μm Carbowax/divinylbenzene (CW/DVB) bonded
fiber. After some experimentation, the 75μm Carbowax/divinylbenzene (CW/DVB) (Supleco,
USA) was chosen to perform the volatile analysis. Krofta and Nesvadba (2005), suggested a
PDMS 30μm fiber (Supelco, USA) was best suited for isolation of hop oil volatiles from female
plants, while a PDMS/DVB (Divinyl Benzene) SPME filter was better suited for head space
analysis of male plants. This is due to the higher concentration of lupulin glands present in the
male plants. Since we did not sex segregate the plants used in this study, this may have played a
factor in the adsorptive capability of the fiber.
The presence of elevated levels of α-caryophyllene and β-caryophyllene was notable.
These terpenes have been identified in HIPV blends released by lima beans and kidney beans
and are used by the two-spotted spider mite predator Phytoseiulus persimilis (Acarina:
Phytoseiidae) to locate prey (Maeda and Takabayashi 2001).
Although methyl salicylate has previously been identified as a compound deserving
investigation into its signaling capabilities (Pare and Tumlinson 1999, James and Grassowitz,
2005), its capability to function as a priming compound is less certain. Engelberth (2006)
suggested that salicylic MeSA along with MeJA were among the few volatiles with potential to
serve in inter plant communication”.

56
While the data are relatively inconsistent, our results indicate trends that may illustrate
that MeSA is capable of inducing the production of herbivore induced plant volatiles in exposed
plants. Furthermore, the data also indicated trends in the production of HIPV‟s in plants exposed
to MeSA may be capable of eliciting production of herbivore induced plant volatiles in nearby
plants. However, much more research is required to fully characterize the emission of volatiles
from hop plants exposed to MeSA or conspecifics exposed to MeSA. Primed or fully alerted hop
plants mediated by deployment of synthetic MeSA in the field, may have great potential in
improving plant defense and enhancing biological control, thus reducing crop damage and
pesticide use.

6057
57
57
a % GC area expresses the proportions of each compound in the total blend of volatiles, calculated from the mean of two repetitions b N= 4 Mean of four sets of juvenile hop plants for 2007 trial c N=6 Mean of six sets of juvenile hop plants for 2008 trial d N=3 Mean of three sets of juvenile hop plants for 2009 trial
trace = <1.0% of overall area counts
ND= Not detected
2007 2008 2009
%GC area
N=4b %GC area
N=6c %GC area
N=3d
Volatile Control
C1
MeSA exposed
T1
Control
C1
MeSA
Exposed
T1
Control
C1
MeSA
Exposed
T1
3-hexen-1-ol (Z ), acetate 34 25 21 29 36 41
3-hexen-1-ol ND ND trace trace 2 2
Nonanal 3 7 ND ND ND ND
Decanal 4 2 ND ND ND ND
Iso-caryophyllene 2 1 ND ND ND trace β-caryophyllene ND ND ND ND trace trace
α-caryophyllene 6 6 4 trace 3 4
germacrene D ND trace 3 2 ND ND
α-farnesene 2 6 ND 1 ND 5
Methyl Salicylate 4 17 3 7 4 ND
germacrene B trace trace 3 ND ND ND
muurolene ND ND ND ND ND ND
γ cadinene ND ND ND ND ND ND
dodecanol 1 trace 4 ND ND ND
nonanoic acid trace ND 10 trace ND ND
Tetradecanoic acid ND ND 3 1 ND ND
Table 1: Summary of compounds collected in Experiment 1 from unexposed (control) and methyl salicylate (MeSA) exposed
juvenile hops a

6058
58
58
Table 2: Summary of compounds collected in Experiment 2 from unexposed (control) and methyl salicylate (MeSA)
exposed juvenile hop plants following combination for 48 hrs a
2007 2008 2009
%GC area %GC area %GC area
Volatile Unexposed
Controlb
C2
Control
C1
MeSA
exposed
T1
Unexposed
Control
C2
Control
C1
MeSA
Exposed
T1
Unexposed
Control
C2
Control
C1
MeSA
Exposed
T1
3-hexen-1-ol (Z ), acetate 34 2 12 20 20 27 30 14 19
3-hexen-1-ol ND ND ND 7 5 6 2 2 ND
Nonanal 3 9 10 ND ND ND ND ND ND
Decanal 4 21 19 ND ND ND ND ND ND
Iso-caryophyllene 2 4 2 2 3 2 ND ND ND
β-caryophyllene ND ND ND 2 ND ND trace 9 10
α-caryophyllene 6 6 3 4 23 9 ND 25 29
germacrene D ND ND ND ND ND 1 ND ND ND
α-farnesene 2 ND ND ND ND trace 16 9 8
Methyl Salicylate 4 ND 2 trace 4 ND 1 2 1
germacrene B trace ND ND ND ND ND ND ND ND
murrolene ND ND ND ND trace trace trace ND ND
γ cadinene ND ND ND trace 1 1. ND ND ND
dodecanol 1 ND ND ND trace ND ND ND ND
nonanoic acid trace ND ND ND 10 trace ND ND ND
Tetradecanoic acid ND ND ND ND 2 1 ND ND ND
a % GC area expresses the proportions of each compound in the total blend of volatiles
b Unexposed control plant was not used in 2007 trials. Data is original control plant data from Table 1
trace = <1.0% of overall area counts
ND= Not detected

6059
59
59
B
Figure 1: Chromatographic profiles of 6 of the 16 retention times of volatiles by unexposed control hop plants (A) and MeSA
treated plants (B). Peaks: (1) 3-hexen-1-ol, (Z) acetate,(2) 3-hexen-ol, (3). β-caryophyllene) (4) α-caryophyllene (5) (E,E) α-
farnesene (6) Methyl salicylate
A
B

6060
60
60
Figure 2: Structural identification of ten of sixteen identified compounds

6061
61
61
Figure 3: Structural identification of remaining six of sixteen compounds

62
Figure 4: Volatiles from control plant by growth age
Volatiles from root cuttings plant ca. 60 days growth
38 1
5
2 3
3-hexen-1-ol Z acetate
3-hexen-1-ol
α-caryophyllene
MeSA
α farnesene
Volatiles from rhizome
ca. 30 days growth
20
2 1 1 2 3
2
4
6 8 2
3-hexen-1-ol Z acetate Nonanal
3-hexen-1-ol
Decanal
iso-caryophyllene
alpha caryophyllene Alpha farnesene
MeSA
Dodecanol
Nonanoic acid
tetradecanoic acid

63
REFERENCES
ARIMURA, G., KOST, C. and BOLAND, W. 2005 Herbivore–induced, indirect plant
defenses Bio. Biophys Acta. 1734:91-111
BERNASCONI, M.L., TURLINGS, T.C.J., AMBROSETTI, L., BASSETTI, P. and
DORN, S. 1998 Herbivore- induced emissions of maize volatiles repel the corn leaf
aphid, Rhopalosiphum maidis. Ent. Exp. App. 87: 133-142
CAMPBELL, C.A.M., PETTERSSON, J., PICKETT, J.A., WADHAMS, L.J. and
WOODCOCK, C.M. 1993 Spring migration of Damson Hop Aphid Phorodon humuli
(Homoptera, Aphididae) and summer host plant-derived semiochemicals released on
feeding. J. Chem. Ecol. 19: 1569-1576
CHOUDARY, D.K., JOHRI, B.N. and PRAKASH, A. 2008 Volatiles as priming
agents that initiate plant growth and defense responses. Current Sci. 94(5):595-604.
DeBOER, J.G., HARDIJIK, C.A., POSTHUMUS, M.A. and DICKE, M. 2008 Prey and
Non-Prey Arthropods Sharing a Host Plant: Effects on Induced Volatile Emission and
Predator Attraction. J. Chem. Ecol.34: 281-290
DICKE, M. 1999 Are Herbivore-Induced Plant Volatiles Reliable Indicators of
Herbivore Identity to Finding Carnivorous Arthropods? Ent. Exp. App.91: 131-142.
DICKE, M., SABELIS, M.W., TAKABAYASHI, J., BRUIN, J., POSTHUMUS, M.A.
1990 Plant strategies of manipulation of predator-prey interactions through
allelochemicals: prospects for application in pest control. J. of Chem. Ecol. 16:3091-
3118
DICKE, M., SCHÜTTE, C., and DIJKMAN, H. 2000 Change in behavioral response
to Herbivore Induced Plant Volatiles in a predatory mite population. J. of Chem. Ecol.
26: (6) 1497-1511
ENGEBERTH, J 2006 Smelling danger and getting prepared: Volatile signals as
priming agents in chemical defense response. Essay 13.8 Plant Physiology 4th ed.
FLINT, H. M., SALTER, S. S. and WALTERS, S. 1979 Caryophyllene: An attractant
for the green lacewing. Environ. Entomol. 8(6): 1123-1125.
FRANCIS, F., LOGNAY, G. and HAUBRUGE, E. 2004 Olfactory responses to aphid
and host plant volatile releases: (E)-x-Farnesene an effective kairomone for the predator,
Adalia bipunctata. J. Chem. Ecol. 30(4): 741-755.

64
HALITSCKE, R, STENBERG, J.A., KESSLER, D., KESSLER,A. and BALDWIN, I.T.
2008 Shared signals- „Alarm calls‟ from plants increase apparency to herbivores and
their enemies in nature. Ecol.Letters 11: 24-34.
HARDIE, J. ISSACS, R., PICKETT, J.A., WADHAM, L.J. and WOODCOCK, C.M.
1994 Methyl salicylate and (-)- (1R,5S) Myrtenal are plant derived repellents for black
bean aphid, Aphis fabae SCOP. (Homotera:aphididae. J. of Chem. Ecol. 20 (11) 2847-
2855.
HEIL, M. and SILVA BUENO, J.C.S. 2007 Within–plant signaling by volatiles leads to
induction and priming of an indirect plant defense in nature. Proc. Natl. Acad. Sci 104:
(13) 5467-5472.
KONG, C., HU, F., XIAOHUA, X., ZHANG, M. and LIANG W. 2005. Volatile
semiochemicals in the Aegeratum conyzoides intercropped citrus orchard and their effects
on mites Amblyseius newsami and Panonychus citri. J. Chem. Ecol. 31(9): 2193-2203.
JAMES, D.G. 2003 Synthetic Herbivore-Induced Plant Volatiles as Field Attractants for
Beneficial Insects. Environ. Entomol. 32: 977-982.
JAMES, D.G. and PRICE, T.S. 2004 Field Testing of Methyl Salicylate for
Recruitment and Retention of Beneficial Insects in Grapes and Hops. J. Chemical Ecol
30 (8): 1613-1628.
JAMES, D.G. and GRASSWITZ, T.R. 2005 Synthetic herbivore-induced plant
volatiles increase capture of parasitic wasps. Bio-Control 50: 871-880
KARBAN, R., MARON, J., FELTON, G.W., EICHENSEER, H. 2003
Herbivore damage to sagebrush induces resistance in wild tobacco: evidence for
eavesdropping between plants. Oikos 100: 325-332.
KESSLER, A. and BALDWIN, I.T. 2001 Defensive function of herbivore induced
volatile emissions in nature. Science 291: 2141-2144.
KESSLER, A. and BALDWIN, I.T. 2004 Herbivore–induced plant vaccination
Part 1: The orchestration of plant defenses in nature and their fitness consequences in the
wild tobacco Nicotiana attenuate. The Plant Journal 38: 639-649.
KOST, CHRISTIAN and HEIL, MARTIN 2008 The defensive role of volatile emission
and extrafloral nectar secretion for Lima Bean in nature. J. Chem. Ecol. 34:2-13.
KROFTA, KAREL and NESVADBA, VLADIMIR 2005 Proc. 1
st IS on Humulus
Acta Hort. ISHS 2005.
.

65
OZAWA, R., ARIMURA, G., TAKABAYASHI, J., SHIMODA, T and NISHIORA, T.
2000 Involvement of jasmonate and salicylate-related signaling pathways for the
production of special herbivore induced volatiles in plants. Plant Cell Phyisol.
41(4):391-398
PARẾ, P.W. and TUMLINSON, J.H. 1999 Plant volatiles as defense against insect
herbivores. Plant Physiol.121: 325-331.
PETTERSSON, J., PICKETT, J.A., PYE, B.J., QUIROZ, A., SMART,L. E. ,
WADHAM,L.J. and WOODCOCK,C.M. 1994 Winter host component reduces
colonization by bird cherry-oat aphid Rhopalosiphum padi L (Homoptera, Aphididae) and
other aphids in cereal fields. J. of Chem. Ecol. 20: (10) 2565-2574.
PICKETT, J. A., WADHAMS, L. J. and WOODCOCK, C. M. 1992. The chemical
ecology of aphids. Ann. Rev. Entomol. 37: 67-90.
SCUTAREANU, P. DRUKKER, B., BRUIN, J., POSTHUMUS, M.A., AND SABELIS,
M.W. 1997 Volatiles from Psylla-infested pear trees and their possible involvement in
attraction of anthrocorid predators. J. of Chem. Ecol. 23: (10) 2241-2260.
SHULAEV, V., SILVERMAN, P. and RASKIN, I. 2005 Airborne signaling by methyl
salicylate in plant pathogen resistance. Nature 385: 718-721.
TURLINGS, T.C.J. and TUMLINSON, J.H. 1992 Systemic release of chemical
signals by herbivore-injured corn. Proc. Natl. Acad. Sci. 89: 8399-8402.
TURLINGS, T.C.J., LOUGHRIN, J.H., Mc CALL, P.J., RÖSE, U.S.R, LEWIS, W.J. and
TUMLINSON, J.H. 1995 How caterpillar-damaged plants protect themselves by
attracting parasitic wasps. Proc. Natl. Acad. Sci. 92: 4169-4174.
TURLINGS, T.C.J., and WÄCKERS, F. 2004 Recruitment of predators and parasitoids
by herbivore-injured plants. In Advances in Insect Chemical Ecology (R.T. Cardế and J.G.
Miller eds.) Cambridge University Press pp.21-75.
.
TURLINGS, T.C.J. and TON, J. 2006. Exploiting scents of distress: The prospect of
manipulating herbivore-induced plant odors to enhance the control of agricultural pests.
Current Opinion in Plant Biology 9: 421-427.
WALLINGS, L.L 2000 The myriad plant response to herbivores. J. Plant Growth Reg.
19 195-216.
ZHU, J. and PARK, K-C. 2005 Methyl salicylate , a soybean aphid plant volatile
attractive to the predator Coccinella semtempunctata. J. of Chem. Ecol. 31(8): 1733-1746.

66
ZHU, J., COSSE, A. L., OBRYCKI, J. J., BOO, K. S. and T. C. BAKER, 1999. Olfactory
reactions of the twelve-spotted ladybeetle, Coleomegilla maculata and the green lacewing,
Chrysoperla carnea to semiochemicals released from their prey and host plant:
Electoantennogram and behavioral responses. J. Chem. Ecol. 25 (5): 1163-1177.

67
Chapter 3
Final Conclusions and Future Work

68
This 3-year plant chamber volatile head space emission evaluation showed trends that
imply MeSA exposed hop plants can elicit plant defense by chemically induced intra and inter
plant communication.
The chamber head space SPME emission information profiled the relative percent
contribution of sixteen herbivore induced plant volatiles to total volatile emission. Comparisons
of MeSA exposed to control plants suggest MeSA itself may elicit HIPV production aiding in
plant defense. When MeSA exposed plants were in close proximity to MeSA treated plants, a
relative rise in certain sesquiterpenes may indicate that HIPV chemical induction pathways
become operational. Previous studies with sagebrush and tobacco (Karban et al., 2004) and lima
beans (Heil and Silva Bueno, 2007) support the hypothesis that uninjured plants located near
plants undergoing herbivore injury will begin the production of volatile compounds. The
compounds emitted from the injured plant provide the uninjured plant with information to
increase its production of defense mechanisms. As the uninjured plant begins its own production
of volatiles, other nearby plants may use these volatiles to begin the process of increasing their
defense mechanisms in anticipation of possible herbivore injury.
While some field studies have been performed using methyl salicylate as an attractant for
predatory species (James, 2003, James and Price, 2004) or as a repellent for herbivory
(Pettersson et al., 2004), the use of methyl salicylate as a priming mechanism to elicit the
production of HIPVs in hops has not been performed. Although the head space emission
information collected from chamber evaluations indicate relatively greater production of certain
HIPVs after MeSA hop exposure, differences in plant rearing (rhizomes versus soft wood

69
cuttings) and other environmental conditions (greenhouse versus growth chamber plant rearing)
could account for appreciable variation in emission profiles among the replicated MeSA
exposure trials conducted from 2007 through 2009. In order to evaluate the capability of plants
to chemically communicate the need for defense, further field testing must be performed. The
data collected in these screening experiments will serve to better design future evaluations in
order to understand the function of intra-plant as well as inter-plant signaling .

70
Appendix A
Analytical Summary Report

71
ANALYTICAL SUMMARY REPORT
Characterization of Volatile Emissions from Juvenile Hop Plants after exposure to Methyl Salicylate by Gas Chromatograph/Mass
Spectroscopy
Author
Rita Abdella
Testing Facility Food and Environmental Quality Laboratory
Department of Entomology
Washington State University
2710 University Drive
Richland, WA 99354-1671
FEQL Study No.: 0507
Laboratory Research Manager
Dr. Vince Hebert
Research Conducted for:
Dr. David James
WSU-AERC, Prosser, WA
Study Timetable
Study Initiation Date: 3/2007
Experimental Termination Date: 9/2007
Report Date
7/2007

72
CERTIFICATION
The undersigned hereby declare that this study was performed under my supervision according
to the procedures described herein, and that this report provides a true and accurate record of the
results obtained.
Analytical Research Director: ___________________________ Date:
Vincent R. Hebert, Food and Environmental Quality Laboratory
Washington State University, Tri-Cities Campus, Richland WA
Analytical work performed by:
Rita Abdella, WSU-FEQL Graduate Researcher
Jane LePage, WSU-FEQL Research Chemist

73
TABLE OF CONTENTS
Page
Certification 2
Table of Contents 3
Archives (location of raw data) 4
Analytical Summary 5
I. Objective/Introduction 5
II. Sample Inventory/History 5
Table 1: Sample Inventory & History
III. Standard Preparation 7
IV. Analytical Procedure 8
A. Air Sampling 8
B. SPME Data Analyses 9
V. Results and Discussion 10
Table 2: Interval Data 11
Table 3 Treated and Control Hop Plant Replicate data 18
Table 4 Greenhouse Air Sample Data 21
Table 5 Treated and Control Hop Data before/after
Combination 26
Figure 2 28

74
ARCHIVES (LOCATION OF RAW DATA)
The original raw data, correspondence logs, and all relevant information for the study titled: “Characterization of Volatile Emissions from juvenile Hop Plants after Exposure to Methyl Salicylate by Gas Chromatography/Mass Spectroscopy” FEQL project number 0507, along with certified originals of the signed analytical summary report will be maintained by the testing facility. Exact copies of the analytical summary report and relevant information for the construction of this study will be transferred to WSU-
AERC, Prosser upon request.
Laboratory Research Director: Vincent Hebert
Testing Facility: Food and Environmental Quality Laboratory
Department of Entomology
Washington State University
Richland, WA 99354-1671

75
ANALYTICAL SUMMARY REPORT
I. Objectives/Introduction
The Food and Environmental Quality Laboratory (FEQL) in Richland, WA evaluated the volatile
emissions from juvenile hop plants following a timed exposure to methyl salicylate. Methyl
salicylate (MeSA) is a common volatile produced by plants when attacked by insect pests and can
aid natural enemies in locating their prey (Shulaev, et al., 1997; James, 2003, Kunart, et al., 2002).
Besides being a plant to insect signal, it has been suggested that MeSA elicits plant to plant
communication as well with the consequence that the hop plants themselves may start producing
their own predator-attracting volatiles (David James personal communication). To test this
hypothesis, a series of greenhouse/laboratory experiments were conducted from March 20 through
May 14th
2007 to evaluate volatile emission profiles from juvenile hop plants exposed to MeSA
when compared to control (non-exposed) plants. These studies were conducted at the WSU Prosser
Agricultural Research Center greenhouse facilities to evaluate chemical signaling following
exposure to methyl salicylate. The plants were contained in separate control and treated
greenhouses under comparable environmental conditions. The plants were of the same approximate
growth age and possessed an approximate total surface area of 30-50 cm2 when evaluated. For each
of the four trials, the treated plant was exposed to methyl salicylate from an impregnated sachet for
approximately 72 hrs. After the exposure interval, a control plant and a treated plant were enclosed
individually in 2.0 liters glass chambers that contained a sampling portal. The two chambered
plants were then transported to the Food and Environmental Quality Lab of Washington State
University –Tri-Cities. (WSU-FEQL). Upon arrival, the plants were allowed to equilibrate at room
temperature for ca. 2-hours before sampling the air surrounding each plant by Solid Phase Micro-
Extraction (SPME). The fiber of this diffusion-based sampling devise was exposed to the chamber
air for five minutes before thermal desorption into an Agilent 6890 Gas Chromatograph with a 5973
Mass Spectrometer. After 24 hrs a second set of SPME analyses were performed to evaluate
changes in chemical emission among the control and treatment chambered plants. After analyzing
the 24-hour SPME samples, the control and treatment plants were removed from their individual
containers, and placed together in a 10 liter glass chamber for 48 hours and then again separated to
their individual chambers. After allowing four hours for equilibration, a final set of SPME analyses
were drawn on the individual containers to measure the effects of exposure. The gas
chromatographic identity profiles from four replicate side-by side control-treatment evaluations are
reported herein.
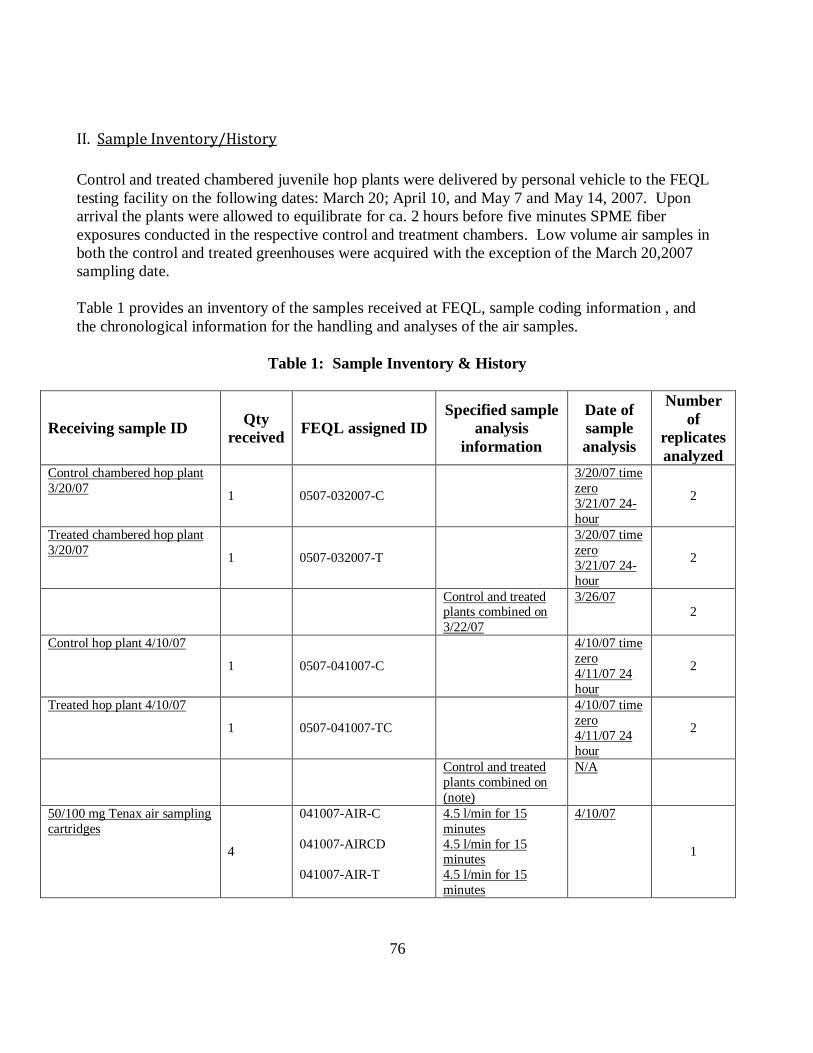
76
II. Sample Inventory/History
Control and treated chambered juvenile hop plants were delivered by personal vehicle to the FEQL
testing facility on the following dates: March 20; April 10, and May 7 and May 14, 2007. Upon
arrival the plants were allowed to equilibrate for ca. 2 hours before five minutes SPME fiber
exposures conducted in the respective control and treatment chambers. Low volume air samples in
both the control and treated greenhouses were acquired with the exception of the March 20,2007
sampling date.
Table 1 provides an inventory of the samples received at FEQL, sample coding information , and
the chronological information for the handling and analyses of the air samples.
Table 1: Sample Inventory & History
Receiving sample ID Qty
received FEQL assigned ID
Specified sample
analysis
information
Date of
sample
analysis
Number
of
replicates
analyzed Control chambered hop plant
3/20/07 1 0507-032007-C
3/20/07 time
zero
3/21/07 24-
hour
2
Treated chambered hop plant
3/20/07 1 0507-032007-T
3/20/07 time
zero
3/21/07 24-
hour
2
Control and treated plants combined on
3/22/07
3/26/07 2
Control hop plant 4/10/07
1 0507-041007-C
4/10/07 time
zero
4/11/07 24
hour
2
Treated hop plant 4/10/07
1 0507-041007-TC
4/10/07 time
zero
4/11/07 24
hour
2
Control and treated
plants combined on
(note)
N/A
50/100 mg Tenax air sampling
cartridges
4
041007-AIR-C
041007-AIRCD
041007-AIR-T
4.5 l/min for 15
minutes
4.5 l/min for 15 minutes
4.5 l/min for 15
minutes
4/10/07
1

77
Receiving sample ID Qty
received FEQL assigned ID
Specified sample
analysis
information
Date of
sample
analysis
Number
of
replicates
analyzed 041007-AIR-TD 4.5 l/min for 15
minutes
Control hop plant 5/7/2007
1 0507-050707-C
5/7/07 time
zero
5/8/07 24
hour
3
Treated hop plant 5/7/2007
1 0507-050707-T
5/7/07 time
zero
5/8/07 24
hour
3
Control and treated
plants combined on
5/9/07
5/11/07
2
50/100 mg Tenax air sampling
cartridges
4
050707-AIR-C
050707-AIRCD
050707-AIR-T
050707-AIR-TD
4.25 L/min for 15
minutes
4.25L/min for 15 minutes
4.5L/min for 15
minutes
4.5L/min for 15
minutes
5/7/07
1
Control hop plant 5/14/2007
1 0507-051407-C
5/14/07
Time Zero
5/15/07 24
hour
3
Treated hop plant 5/14/2007
1 0507-051407-T
5/14/07
Time Zero
5/15/07 24
hour
3
Control and treated plants combined on
5/15/07
5/17/07 2
50/100 mg Tenax air sampling
cartridges
4
051407-AIR-C
051407-AIRCD
051407-AIR-T
051407-AIR-TD
4.25 L/min for 15
minutes
4.25 L/min for 15
minutes
4.5L/min for 15
minutes
4.5L/min for 15
minutes
5/14/07
1

78
III. Standard Preparation
Standards were prepared to bracket the range of residues in the samples. The following test
substances, standards, and standard dilutions were used throughout this study:
Test substance
Compound Reference No. Source
Methyl Salicylate 13141 Sigma-Aldrich
Stock Solution
Compounds Solution Number Solvent Concentration
Methyl salicylate 13142 Hexane 1 mg/mL
Dilutions of Stock Solutions
Compounds Solution Number Solvent Concentration
Methyl Salicylate 13142-1 Ethyl Acetate 0.5 ug/mL
Methyl Salicylate 13142-2 Ethyl Acetate 1.0 ug/mL
Methyl Salicylate 13142-3 Ethyl Acetate 2.00.5 ug/mL
Methyl Salicylate 13142-4 Ethyl Acetate 5.0 ug/mL
Methyl Salicylate 13142-5 Ethyl Acetate 10.0 ug/mL
Methyl Salicylate 13142-6 Ethyl Acetate 0.2. ug/mL
Methyl Salicylate 13142-7 Ethyl Acetate 100 mg/mL
Methyl Salicylate 13142-8 Ethyl Acetate 0.005 u/mL
Methyl Salicylate 13142-9 Ethyl Acetate 0.1 ug/mL
All standard solutions were stored in the freezer at ca.-20˚ C (freezer I.D. Comet). The dilutions
made during the course of the study are recorded in the FEQL analytical laboratory standards book.
The expiration date of the linearity standards is 10/15/2007.

79
IV. Analytical Procedure
A. Air Samples
Air sampling was performed using 50/100 mg Tenax cartridges for both control and treated
plant greenhouses to assess ambient methyl salicylate concentrations on three of the four
interval sampling dates. Two 50/100 mg Tenax cartridges were collocated on a specially
designed air sample device that was attached to a portable low volume air sample pump
(Figure 1). Air samples were collected for 15 minutes and flow rates were verified using a
Gilmont flow meter following air sampling. Flow rates are presented in Table 1.
After the air samples were collected, the Tenax tubes were eluted with 3 mL of ethyl
acetate, into a 15 mL graduated centrifuge tube and the final volume adjusted to 3mL with
ethyl acetate for analysis by gas chromatograph/mass spectroscopy (GC/MS). Extracts were
analyzed by GC/MS Selective Single Ion Monitoring (SIM) mode to quantify methyl
salicylate (see chromatographic conditions below).
B. SPME Data Analysis
Each interval set of chambered plants (control and treatment) taken on a particular interval
date were analyzed in duplicate by SPME. Upon arrival, the plants were allowed to
equilibrate at room temperature for ca. 2-hours before sampling the air surrounding each
plant by Solid Phase Micro-Extraction (SPME). A 7 µm film thickness polysiloxane fiber
was inserted into the respective control/treatment chamber for 5 or 10 minutes. Afterwards,
the exposed fiber was thermal desorbed for 30 seconds into the hot (200oC) injection port of
a Agilent 6890 Gas Chromatograph.
Methyl salicylate and other compounds were determined using an Agilent 6890N Gas
Chromatograph with 5973N Mass Selective Detector (MSD) in total ion chromatography
(TIC) mode. Typical operating conditions are described below:
GC Conditions:
Columns: Alltech 19654 EC-WAX 30mx0.32mmI.D. x0.25 μm film thickness
(Alltech Inc.)
Carrier gas: Ultrapure helium

80
Oven Program: Initial temp 50˚C for 1 minute
Ramps:
5˚C/minute to 260˚C hold for 5 minutes
Injection Volume: 2 μL
Injector: Pulsed Splitless, 200˚C
Detector: Temperatures:
MS Quad 280˚C
Scan : 50-350 m/z or Single Ion Monitoring (SIM) at 152 m/z for MeSA in Tenax
The chromatograms generated in each analysis were integrated to indicate retention time and
a library search using the Data Analysis function of the ChemStation software was
performed to identify chromatographic peaks of interest. Following peak identification, the
peaks of interest were compared against the NIST library to determine the best match. The
relative proportion (percentage of area counts) associated with individual peaks per run was
also generated to compare chromatographic results of combined control and treatment
evaluations.
V. Study Findings
On March 20, 2007 results from SPME evaluations indicated that the control plant methyl
salicylate emissions made up approximately 4% to 8% of total peak area counts. A
moderate response (approximately 25% total area count) of 3-Hexen-1-ol was observed with
lesser air emissions of formate (10%), acetic acid (10%), and 1 to3% of other organic acids
being detected. Nonanol was present in very low concentrations (approximately 2% of total
area counts). The treated plant organic emissions indicated a higher percentage of methyl
salicylate (23%-25% of total area counts) present together with α-farnasene (18%-20%).
Detected air emissions of 3-hexen-1-ol were considerably lower in the treated samples (8-
9%). After approximately 24 hours, the second set of analysis on both the control and treated
plants indicated that in the control, methyl salicylate concentrations remained stable (5% of
total area counts), while nonanal concentration increased slightly (2 to 6.9% of total area
counts). The relative proportion of methyl salicylate, nonanal and α-farnasene in the treated
plant data increased (8.6%-13%; 25%-35% and 20%-44% respectively). Other trace level
organic acids such as propanoic, and 1-dodecanol concentrations were decreased overall.
On April 10, 2007 another set of control and similarly exposed MeSA treatment juvenile
hop plants were obtained and transported to the FEQL for analysis. SPME were performed
in duplicate for each sample in an identical manner as before. Analysis of the control plant
indicates the presence of methyl salicylate (2% of total area counts). 3-hexen-1-ol, formate
and hexen-1-ol, acetate were present (22 % and 40% of total area counts), respectively. The
treated plants were sampled for 5 minutes and analyzed. Total contribution due to methyl
salicylate in the treated sample was12% to 13% of the total area counts, while hexen-1-ol,

81
acetate and 3-hexen-1-ol, formate levels contributed 38% and 40% of the total area counts;
α-farnasene contributed approximately 2.4% to total area counts. The plants were placed in
a common container and after 24 hours, removed and isolated. After a short time for
equilibration, the plants were sampled, using a five minute sample time and the data
reviewed The control plant indicated an increase in contribution to area counts due to
methyl salicylate from 2.5% to 13% of total area counts. The contribution due to α-
farnasene increased from 2.4% to 24 % after the combined exposure. The treated plant
analysis suggests that there was no change in methyl salicylate. However, a decrease in the
contribution due to hexen-1-ol acetate from 22% to 7% and a reduction in α-farnasene to
17% was observed.
On May 7, 2007 the third set of juvenile hop plants were obtained and transported to WSU-
FEQL. The control plant was sampled using a 5 minute sample time and the data was
analyzed. Butanoic acid contributed approximately 15% of the total area counts, and 3-
hexen-1-ol contributed approximately 5% of the total area counts. No indication of α-
farnasene or methyl salicylate was present. The treated plant was sampled using a five
minute sample time. The data indicated that the majority of the contribution to total area
counts came from. α-farnasene (12% of total area counts), 2% from 3-hexen-1-ol, Z formate
and 2% was due to 3-hexen-1-ol, acetate and methyl salicylate. Following the resulting
exposure of the control plant to the treated, and the allowed time for equilibration after
separation, the plants were resampled for 5 minutes. The control plant indicated no presence
of methyl salicylate or α-farnasene. Several metabolites of methyl salicylate were indicated,
with benzoic acid, 2,4 bis-tri-methyl increasing from 4% in the original sample to 29% after
exposure. Benzoic acid was also present in the sample following exposure contributing
approximately 38% of the total area counts. The treated plant indicated a two fold increase
in levels of 3-hexen-1-ol acetate (5.5% to 12.6%) and α-farnasene (11% to 22%). Methyl
salicylate increased only slightly from 3% to 4 %
On May 14, 2007 the final set of plants was obtained and transported to WSU-FEQL for
sampling and analysis. The control plant was sampled for five minutes and the data
analyzed. Of the compounds present, 3-hexen-1-ol, acetate contributed 63% of the total area
counts, while 3-hexen-1-ol, formate contributed 6%of the total area counts, with 2% of the
area counts were attributable to α-farnasene and 3% to methyl salicylate. The treated plant
sample indicated 73% of the total area counts attributable to 1,4 hexadiene, 66% of the total
area counts attributable to 3-hexen-1-ol, acetate and 19-22% attributable to methyl
salicylate. After the exposure of the control plant to the treated plant, the control plant
exhibited a twofold decrease in the area counts of 3-hexen-1-ol (63%-32%) and an increase
in decanal area counts form 1% to 32% and an increase in nonanal area counts from 7% to
11%. The treated plant exhibited a reduction in 1,4 hexadiene area counts from 73% to 12%,
and a increase in methyl salicylate area counts from 19% to 22.5%. Decanal, not present in
the original sample, contributed 33% to the total area counts. The use of total area counts is
used as an indicator of instrument response to the compounds identified in this study. It is

82
important to note that the area counts are not used for quantification of the identified
compounds, but as a means to characterize the instrument response at each interval and to
compare the response between the treated and control plants at each sampling interval.
Table 2 provides the raw data for each sampling interval.

83
Table 2
Interval Data Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
0507-032007T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) 814065 8.605 9459860
Nonanal 326078 3.447
Decanal 458094 4.843
α- Farnesene 1521523 16.084
α- Farnesene 432908 4.576
2-(2-ethoxyethoxy)ethanol 469708 4.965
Methyl Salicylate 2397953 25.349
0507-032007T Treated Hop Plant #2 3-hexen-1-ol, acetate(Z) 768558 12.92 5947831
Nonanal 127756 2.15
Decanal 94991 1.6
α- Farnesene 1414151 23.8
α- Farnesene 397482 6.7
2-(2-ethoxyethoxy)ethanol 297726 5.006
Methyl Salicylate 2418796 40.667
0507-032007T Analyzed-3/21/07 3-hexen-1-ol, acetate(Z)
Nonanal a
Decanal a
α- Farnesene a
α- Farnesene a
2-(2-ethoxyethoxy)ethanol a
Methyl Salicylate a 0
0507-0320007-T+C Analyzed 3/26/07 3-hexen-1-ol, acetate(Z) 219837 3.8 7616994
Nonanal 431364 5.7

84
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
Decanal 1563544 20.57
α- Farnesene 625386 8.2
α- Farnesene 185276 2.4
2-(2-ethoxyethoxy)ethanol 288022 3.8
Methyl Salicylate 0 0
0507-032007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 2329355 30.583 5723682
Nonanal 220425 2.890
Decanal 451755 5.9
2-(2-ethoxyethoxy)ethanol 755645 9.92
α-caryophyllene 173026 2.9
α- Farnesene 89497 1.17
Methyl Salicylate 351522 4.6
0507-032007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1300934 33.959 46278881
Nonanal 220425 2.894 46278881
Decanal 451755 5.9 46278881
2-(2-ethoxyethoxy)ethanol 755645 9.92 46278881
α- Farnesene 130565 5.6 10351201
α- Farnesene 3.84
α-caryophyllene 203753 2.27 10351201
Methyl Salicylate
0507-032007-C Control Hop Plant-Analyzed
4/12/07
3-hexen-1-ol, acetate(Z) a
Nonanal a
Decanal
2-(2-ethoxyethoxy)ethanol a
α- Farnesene

85
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
α-caryophyllene
Methyl Salicylate
0507-032007-T+C Analyzed 3-hexen-1-ol, acetate(Z) 422220 3.1
Nonanal 518683 1.6
Decanal 1392646 10.27
2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene 640638 4.7
α- Farnesene 231907 1.7
α-caryophyllene 916545 6.76
Methyl Salicylate ND NA
0507-041007T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) ND NA
Nonanal ND ND
Decanal 1118089 4.9
α- Farnesene ND NA
α- Farnesene ND NA
2-(2-ethoxyethoxy)ethanol ND NA
Methyl Salicylate 239808 7.2
0507-041007T Treated Hop Plant #2 3-hexen-1-ol, acetate(Z) 912455 22.4 4081824
Nonanal ND NA
Decanal ND NA
α- Farnesene 827617 20.3
2-(2-ethoxyethoxy)ethanol ND NA
Methyl Salicylate 293808 7.2
0507-041007T Analyzed 4/11/07 3-hexen-1-ol, acetate(Z) ND NA 4894139

86
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
1,4 Hexadiene 74077 1.5
Decanal ND NA
α-caryophyllene 193386 3.95
α- Farnesene ND NA
2-(2-ethoxyethoxy)ethanol 76157 1.6
Methyl Salicylate 259129 5.3
0507-04100007-T+C Analyzed 4/1207 3-hexen-1-ol, acetate(Z) ND NA Nonanal 208334 5.2 Decanal 1563544 α- Farnesene ND NA 2-(2-ethoxyethoxy)ethanol 58403 1.5 Methyl salicylate 703476 17.7 0507-041007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1819153 5.2 35538598
Nonanal 2731535 7.7
Decanal 66156617 18.6
2-(2-ethoxyethoxy)ethanol 1558482 4.4
α-caryophyllene 158727 0.5
α- Farnesene 233958 .7
α- Farnesene 389787 1.1
Methyl Salicylate 316555 0.9
0507-041007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1193144 15.9 7494237
Nonanal ND NA Decanal ND NA 2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene 167961 2.2 α- Farnesene ND NA

87
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
α-caryophyllene ND NA Methyl Salicylate ND NA 0507-041007-C Control Hop Plant-Analyzed
4/11/07
3-hexen-1-ol, acetate(Z) 4452532 14.936 2295343
Nonanal 445232 14.936 Decanal 64963 2.18 2-(2-ethoxyethoxy)ethanol 542430 1.8 α- Farnesene 703697 23.6 α- Farnesene 386125 12.95 α-caryophyllene 047063 1.58 Methyl Salicylate ND 8.5 0507-041007-T+C Analyzed 4/12/07 3-hexen-1-ol, acetate(Z) 204035 8.9 2295343
Nonanal 161730 7.05 Decanal 335964 14.637
2-(2-ethoxyethoxy)ethanol ND
α- Farnesene 384013 16.7
α- Farnesene 103568 4.51
α-caryophyllene ND 0
Methyl Salicylate 195957 8.5 0507-050707T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) 9179031 20.064 Nonanal ND NA Decanal 129729 0.284 2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene 1927476 4.2 α- Farnesene 589647 1.3 α- -caryophyllene 9716891 21.2

88
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
Methyl Salicylate 4669829 10.2 0507-050707T Treated Hop Plant #2 3-hexen-1-ol, acetate(Z) 6968532 16.9 Nonanal ND NA Decanal ND NA 2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene 2147307 5.20 α- Farnesene 643020 1.6 α- -caryophyllene 1283423 3.11 Methyl Salicylate 4181594 10.134 0507-050707T Analyzed-5/8/07 3-hexen-1-ol, acetate(Z) 415025 5.51 Nonanal 198353 2.63 Decanal 400859 5.33 α-caryophyllene 214003 2.84 α- Farnesene 718421 9.53 α- Farnesene 183674 2.63 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate 198394 2.6 0507-050707-T+C Analyzed 5/11/07 3-hexen-1-ol, acetate(Z) 440886 12.6 Nonanal ND NA Decanal 80203 2.29 α-caryophyllene ND NA α- Farnesene 562628 16.11 α- Farnesene 229122 6.56 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate 143342 4.10 0507-0507007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 3207682 40.9 7830680

89
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
2,4 Hexadiene 157109 2.00 Decanal 124481 1.6 α-caryophyllene 743635 9.5 α- Farnesene ND NA Methyl Salicylate 238036 3.0 0507-050707-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1572577 45.8 3431347
2,4 Hexadiene 98949 2.9
Decanal ND NA
2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene ND NA
α-caryophyllene 507225 14.8
Methyl Salicylate 172376 5.0
0507-050707-C Control Hop Plant-Analyzed 5/8/07 3-hexen-1-ol, acetate(Z) 61513 5.3 1158917
Nonanal 172576 14.5
Decanal 314223 27.1 2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene ND NA α-caryophyllene 103434 8.9 Methyl Salicylate ND NA 0507-0050707-T+C Analyzed 5/11/07 3-hexen-1-ol, acetate(Z) 92578 3.4 2723343
Nonanal ND NA Decanal ND NA 2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene ND NA
α-caryophyllene ND NA
Methyl Salicylate ND NA

90
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
0507-051407T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) 665224 72.99 911396
Nonanal ND NA
Decanal ND NA
2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene ND NA
α- -caryophyllene 72414 7.9
Methyl Salicylate 173758 19.1
0507-051407T Treated Hop Plant #2 3-hexen-1-ol, acetate(Z) 406461 66.02 615634
Nonanal ND NA Decanal ND NA 2-(2-ethoxyethoxy)ethanol ND NA α- -caryophyllene 69893 11.4 α- Farnesene ND NA Methyl Salicylate 139280 22.62 0507-051407T Analyzed-5/8/07 3-hexen-1-ol, acetate(Z) 96874 11.7 825568 Nonanal 98288 11.9 Decanal 270914 32.82 α-caryophyllene ND NA α- Farnesene ND NA 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate 186533 22.6 0507-0541407-T+C Analyzed 5/17/07 3-hexen-1-ol, acetate(Z) 210563 11.9 1771614
Nonanal 291961 16.5
Decanal 632736 35.8
α-caryophyllene ND NA α- Farnesene ND NA

91
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate ND NA 0507-0514007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 4172270 63 6610360
2,4 Hexadiene 444197 6.7 Decanal 66558 1.0 α-caryophyllene 392699 5.94 α- Farnesene 87774 1.33 α- Farnesene 77023 1.2 Methyl Salicylate 227442 3.44 0507-0514007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 2117851 64.99 3258699
2,4 Hexadiene 193580 5.94
Decanal 68089 2.09
α- Farnesene ND NA α-caryophyllene 225693 6.926 Methyl Salicylate 155260 4.76 0507-051407-C Control Hop Plant-Analyzed
5/15/07
3-hexen-1-ol, acetate(Z) 487363 32.7 1491983
Nonanal 244290 16.4 Decanal 404379 27.1 2,4 bis[trimethyl],benzoic acid 39791 2.7 α- Farnesene ND NA α-caryophyllene ND0 NA Methyl Salicylate ND0 NA 0507-0051407-T+C Analyzed 5/17/07 3-hexen-1-ol, acetate(Z) ND NA 2088546
Nonanal 368825 17.7
Decanal 860842 41.22

92
Sample Identification Compound ID Area Counts % Total
Area
Total
Area
counts
2,4 bis[trimethyl],benzoic acid 76764 3.7
α- Farnesene ND NA
α-caryophyllene ND NA
Methyl Salicylate ND NA
ND = None detected
NA = Not applicable a Different SPME device used for comparison

93
Table 3
Treated and Control Hop Replicate Data
Sample Identification Compound ID Area Counts % Total
Area
Total Area
counts
0507-032007T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) 814065 8.605 9459860
Nonanal 326078 3.447
Decanal 458094 4.843
α- Farnesene 1521523 16.084
α- Farnesene 432908 4.576
2-(2-ethoxyethoxy)ethanol 469708 4.965
Methyl Salicylate 2397953 25.349
0507-032007T Treated Hop Plant #2 3-hexen-1-ol, acetate(Z) 768558 12.92 5947831
Nonanal 127756 2.15
Decanal 94991 1.6
α- Farnesene 1414151 23.8
α- Farnesene 397482 6.7
2-(2-ethoxyethoxy)ethanol 297726 5.006
Methyl Salicylate 2418796 40.667
0507-032007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 2329355 30.583 5723682
Nonanal ND ND
Decanal 451755 5.9
α-caryophyllene 173026 2.9
α- Farnesene 640638 NA
α- Farnesene 231907 NA
Methyl Salicylate ND NA
0507-032007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 2280630 48.895 4664357
Nonanal 108436 2.325

94
Sample Identification Compound ID Area Counts % Total
Area
Total Area
counts
Decanal 141391 3.031
2-(2-ethoxyethoxy)ethanol 615503 13.2
α-caryophyllene 222789 4.8
α- Farnesene 88849 1.91
α- Farnesene ND NA
Methyl Salicylate 141103 3.025
0507-041007T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) ND NA 10351201 Nonanal ND ND Decanal 1118089 4.9 α- Farnesene ND NA α- Farnesene ND NA 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate 239808 7.2 0507-041007T Treated Hop Plant #2 3-hexen-1-ol, acetate(Z) 912455 22.4 4081824
Nonanal ND NA
Decanal ND NA
α- Farnesene 827617 20.3
α- Farnesene ND NA
2-(2-ethoxyethoxy)ethanol ND NA
Methyl Salicylate 293808 7.2
0507-041007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1819153 5.2 35538598
Nonanal 2731535 7.7
Decanal 66156617 18.6
2-(2-ethoxyethoxy)ethanol 1558482 4.4
α-caryophyllene 158727 0.5
α- Farnesene 233958 .7
α- Farnesene 389787 1.1
Methyl Salicylate 316555 0.9
0507-041007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1193144 15.9 7494237

95
Sample Identification Compound ID Area Counts % Total
Area
Total Area
counts
Nonanal ND NA Decanal ND NA 2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene 167961 2.2 α-caryophyllene 507225 14.782 Methyl Salicylate ND NA 0507-050707T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) 9179031 20.064 Nonanal 1085129 2.37 Decanal 129729 0.284 2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene 1927476 4.2 α- Farnesene 589647 1.3 α- -caryophyllene 1421623 3.10 Methyl Salicylate 4669829 21.239 0507-050707T Treated Hop Plant #2 3-hexen-1-ol, acetate(Z) 6968532 16.9 Nonanal 729633 1.78 Decanal ND NA 0 2-(2-ethoxyethoxy)ethanol 729633 1.768 α- Farnesene 2147307 5.20 α- Farnesene 643020 1.6 α- -caryophyllene 1283423 3.11 Methyl Salicylate 4181594 10.134 0507-050707-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 4172270 63.1 6610360
2,4 Hexadiene 444197 6.72 Decanal 66558 1.0 α-caryophyllene 392699 5.94 α- Farnesene 87774 1.33

96
Sample Identification Compound ID Area Counts % Total
Area
Total Area
counts
α- Farnesene 77023 1.2 Methyl Salicylate 227442 3.44 0507-050707-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 2117851 64.99 3258699
2,4 Hexadiene 193580 5.94
Decanal 68089 2.09
2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene ND NA α-caryophyllene 225693 6.926 Methyl Salicylate 155260 4.76 0507-050707-T+C Analyzed 5/11/07 3-hexen-1-ol, acetate(Z) 440886 12.6 Nonanal ND NA Decanal 80203 2.29 α-caryophyllene 215205 6.1 α- Farnesene 562628 16.11 α- Farnesene 229122 6.56 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate 143342 4.10

97
Table 4
Greenhouse Air Sample Data
Receiving
sample ID
Fortification
(ug) Peak Count
Calculated
Concentration
(ug/mL)
Final
Volume
(mL)
Dilution
Factor
Total
MeSA
(ug)
Total
Air
Volume
(m3
)
MeSA
Air Conc.
(ug/m3
)
0507-050707-C1 195698 0.112 3 3 0.33 0.11 2.98
0507-050707-C2 1622658 0.094 3 3. 0.28 0.11 2.51
0507-050707-T1 277593 0.155 3 3 0.47 0.11 4.15
0507-050707-T2 521720 0.286 3 3 0.86 0.11 7.63
0507-FS-1 0.3 178157 0.102 3 3 0.31
0507-FS-2 0.3 123137 0.073 3 3 0.22
0507-051407-C1 ND 3 3 0.00 0.05 0.00
0507-051407-C2 ND 3 3 0.00 0.04 0.00
0507-051407-T1 219647 0.141 3 3 0.42 0.05 9.42
0507-051407-T2 163045 0.111 3 3 0.33 0.05 6.33

98
Table 5
Treated and Control Plant Before and After Combined Exposure
Sample Identification Compound ID
Average
Area Counts c
Average
% Total
Area
Average
Total Area
Counts
0507-032007T Treated Hop Plant 3-hexen-1-ol, acetate(Z) 791311.5 10.76 7703845.5
Nonanal 226917 2.8
Decanal 276542.5 3.27
α-caryophyllene ND NA
α- Farnesene 1467837 19.94
α- Farnesene 415195 5.6
2-(2-ethoxyethoxy)ethanol 383717 4.99
Methyl Salicylate 2408374.5 33.0
0507-0320007-Treated T+C Analyzed 3/26/07 3-hexen-1-ol, acetate(Z) 219837 3.8 7616994
Nonanal 431364 5.7
Decanal 1563544 20.57
α-caryophyllene ND NA
α- Farnesene 625386 8.2
α- Farnesene 185276 2.4
2-(2-ethoxyethoxy)ethanol 288022 3.8
Methyl Salicylate 2418796 40.667
0507-032007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 2304992.5 39.8 5194019.5
Nonanal 164430.5 2.325
Decanal 296573 2.8
2-(2-ethoxyethoxy)ethanol 685574 11.6
α-caryophyllene 97652.5 3.9
α- Farnesene 89173 1.5

99
Sample Identification Compound ID
Average
Area Counts c
Average
% Total
Area
Average
Total Area
Counts
α- Farnesene ND NA
Methyl Salicylate 246312.5 3.8
0507-032007-T+C Analyzed 3-hexen-1-ol, acetate(Z) 422220 3.1
Nonanal 518683 1.6
Decanal 1392646 10.27
2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene 640638 4.7
α- Farnesene 231907 1.7
α-caryophyllene 916545 6.76
Methyl Salicylate
0507-041007T Treated Hop Plant 3-hexen-1-ol, acetate(Z) 912455 22.4 Nonanal ND ND Decanal 1118089 4.9 α- Farnesene 827617 20.3 α- Farnesene ND NA α-caryophyllene 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate 249468.5 6.3 0507-041007-T+C Analyzed 4/12/07 3-hexen-1-ol, acetate(Z) 204035 8.9 2295343
Nonanal 161730 7.05 Decanal 335964 14.637
2-(2-ethoxyethoxy)ethanol ND
α- Farnesene 384013 16.7
α- Farnesene 103568 4.51
α-caryophyllene ND 0
Methyl Salicylate 195957 8.5

10
0
Sample Identification Compound ID
Average
Area Counts c
Average
% Total
Area
Average
Total Area
Counts
0507-041007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1506148.5 10.5 21516418
Nonanal 2731535 7.7
Decanal 66156617 18.6
2-(2-ethoxyethoxy)ethanol 1558482 4.4
α-caryophyllene 158727 0.5
α- Farnesene 200960 2.9
α- Farnesene 389787 1.1
Methyl Salicylate 316555 0.9
0507-041007-T+C Analyzed 4/12/07 3-hexen-1-ol, acetate(Z) 204035 8.9 2295343
Nonanal 161730 7.05 Decanal 335964 14.637
2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene 384013 16.7
α- Farnesene 103568 4.51
α-caryophyllene ND NA
Methyl Salicylate 195957 8.5 0507-050707T Treated Hop Plant #1 3-hexen-1-ol, acetate(Z) 8073782 18.5 43506992
Nonanal ND NA Decanal 129729 0.284 2-(2-ethoxyethoxy)ethanol ND NA α- Farnesene 2037392 4.7 α- Farnesene 616334 1.5 α- -caryophyllene 5569257 12.2 Methyl Salicylate 4425712 10.2 0507-050707-T+C Analyzed 5/11/07 3-hexen-1-ol, acetate(Z) 440886 12.6
Nonanal ND NA
Decanal 80203 2.29
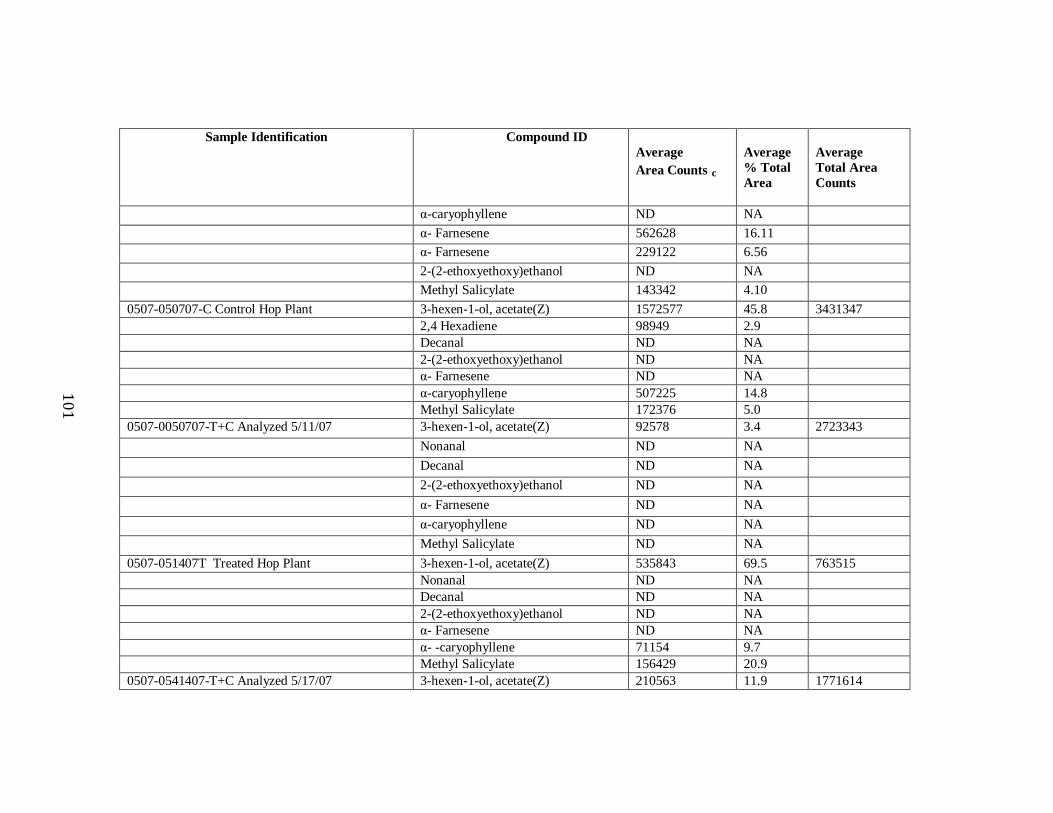
10
1
Sample Identification Compound ID
Average
Area Counts c
Average
% Total
Area
Average
Total Area
Counts
α-caryophyllene ND NA α- Farnesene 562628 16.11 α- Farnesene 229122 6.56 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate 143342 4.10 0507-050707-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 1572577 45.8 3431347
2,4 Hexadiene 98949 2.9
Decanal ND NA
2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene ND NA
α-caryophyllene 507225 14.8
Methyl Salicylate 172376 5.0
0507-0050707-T+C Analyzed 5/11/07 3-hexen-1-ol, acetate(Z) 92578 3.4 2723343
Nonanal ND NA Decanal ND NA 2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene ND NA
α-caryophyllene ND NA
Methyl Salicylate ND NA 0507-051407T Treated Hop Plant 3-hexen-1-ol, acetate(Z) 535843 69.5 763515
Nonanal ND NA
Decanal ND NA
2-(2-ethoxyethoxy)ethanol ND NA
α- Farnesene ND NA
α- -caryophyllene 71154 9.7
Methyl Salicylate 156429 20.9
0507-0541407-T+C Analyzed 5/17/07 3-hexen-1-ol, acetate(Z) 210563 11.9 1771614

10
2
Sample Identification Compound ID
Average
Area Counts c
Average
% Total
Area
Average
Total Area
Counts
Nonanal 291961 16.5
Decanal 632736 35.8
α-caryophyllene ND NA α- Farnesene ND NA 2-(2-ethoxyethoxy)ethanol ND NA Methyl Salicylate ND NA 0507-0514007-C Control Hop Plant 3-hexen-1-ol, acetate(Z) 3145062 64 4934530
2,4 Hexadiene 318889 6.3 Decanal 67324 1.6 α-caryophyllene 309196 6.4 α- Farnesene 87774 1.33 α- Farnesene 77023 1.2 Methyl Salicylate 191351 4.1 0507-0051407-T+C Analyzed 5/17/07 3-hexen-1-ol, acetate(Z) ND NA 2088546
Nonanal 368825 17.7
Decanal 860842 41.22
2,4 bis[trimethyl]benzoic acid 76764 3.7
α- Farnesene ND NA
α-caryophyllene ND NA
Methyl Salicylate ND NA

10
3
Figure 1
Comparative Data from 3/22/07 Sampling Data
Treated vs. Control 3/22/07
0 5000000 10000000 15000000 20000000 25000000 30000000
3-hexen-1-ol, acetate(Z)
Nonanal
Decanal
α-caryophyllene
α-Farnesene
2-(ethoxy)ethanol
Methyl Salicylate
Area Counts TIC
3/22 Treated
3/22 Control

10
4
Figure 2 : Treated vs. Treated after 48hrs combination 3/22/07
Treated vs Treated After Combination
0 1000000 2000000 3000000 4000000 5000000 6000000
3-hexen-1-ol, acetate(Z)
Nonanal
Decanal
α-caryophyllene
α- Farnesene
α- Farnesene
2-(2-ethoxyethoxy)ethanol
Methyl Salicylate
Treated Hop 3/22/07
Treated Hop After Comb





























