Embed Size (px)
Citation preview

Best Available Cpy
AFRRI REPORTS&~ ~ Jly *August 6 September 1988
A1
7VOLD
DTICnELECTE
DefenseN Nula Agency~Are ocs aiboog eerh nttt
Ar e Foce Apprbo eo r plReeare; dInstitumte

REPRODUCTION QUALITY NOTICE
This document is the best quality available. The copy furnishedto DTIC contained pages that may have the following qualityproblems:
" Pages smaller or larger than normal.
* Pages with background color or light colored printing.
* Pages with small type or poor printing; and or
* Pages with continuous tone material or colorphotographs.
Due to various output media available these conditions may ormay not cause poor legibility in the microfiche or hardcopy outputyou receive.
F7lf this block is checked, the copy furnished to DTICcontained pages with color printing, that when reproduced inBlack and White, may change detail of the original copy.
This document contain
blank pages that were
not filmed
UNCLASSIFIED
SECURITY CLASSIFICATION OF THIS PAGE
REPORT DOCUMENTATION PAGEI& REPORT SECURITY CLAS SIFICAIIO lb RESTRICTIVE MARKINGS
UNCLASSIFIED________________ _____
2. SECuRITY CLASSIFICATION AUTHORITY 3 OISTRIBUTIDNIAVAILABILITY OF REPORT
Approved for public rclease; distribution2b OE CLASSIF )CATION, DOWNGRADING SCHEOU LE unlimited.
4 PERIOCRMING ORGANIZATION REPORT NUMBERISI 5 MONITORING ORGANIZATION REPORT NUMBE RISI
SR88-20 - SR88-296.NA 4: PERFORMING ORGANIZATION 00.OFC YBL 7.NM F OIOIGOGNZTO
Ar med Forces Radiobiology II%111p~le, OL .. AM O
Research Institute IF~6c. ADDRESS ICily. Slate and ZIP Code) 7b. ADDRESS (City. Slate and ZIP Code)
Defense Nuclear AgencyBethesda, Maryland 20814-5145
So, NAME OF FUNOiNGSPONSORINO 186. OFFICE SYMBOL 9. PROCUREM6ENT INSTRUMENT IDENTIFICATION NUMBERORGANIZATtON W j appicable)
Defense Nuclear Agency JDNABc ADDRESS Cjty. Stale and ZIP CadeI TO SOURCE OF FUNDING NOS
IPR OGRAM PROJECT TASK WORK UNIT
Washington, DC 20305 ELEMENT NO. NO. NO, NO.
I I TLE Itc*iude Sec-Iy Ctaaf~caI~onI NWEDAFRRI Reports, Jul-Sep 1988 QAXM
12. PERSONAL AUTHORISI
13. TYPE OF REPORT 13b. TIME COVERED 14DATE OF REPORT iYr, Mo.. Day) 15. PAGE COUINt
Reprints/Technical FROM TO ____ 1988 November 85416 SUPPLEMAENTARY NOTATION
17 COSATI CODES 1B. SUBJECT TERMS ICOItiflW on rwlrfal Ifneccfuia and identif~y by block number)
FIELD GRO0UP SUB OR
19. ABSTRACT (Confin.ue on ntuerse it'neceuat-, and identify by block ,,u,,berI
This volume contains AFRRI Scientific Reports SR88-20 through SR88-29 for Jul-Sep 1988.
20. DISTRIBUTION/AVAILABILITY OF ABSTRACT 21 ABSTRACT SECURITY CLASSIFICATION
UNCLASSIFIED/UNLIMITEO 7 SAME AS RPT. -:3 OTIC USERS vj UNCLASSIFIED22s. NAME OF RESPONSIBLE iNDIVICU \L 226 TELEPHONE NUMBER 1 22c OFFICE SYMBOL
.M. E. Greenville Inl~ude Area. Cadet ISD(202)295-3536 ID
DO FORM 1473, 83 APR EDITION OF I JAN 73 IS OBSOLETE. UNCLASSIFIEDSECURITY CLASSIFICATION OP THIS PAGE

CONTENTS;
Scientific Reports
SR88-20: Dubois, A., Fiala, N., Boward, C. A., and Bogo, V. -Prevention andtreatment of the gastric symptoms of radiation sickness,
SR88-21: Dubois, A., and Walker, R. 1. "Prospects for management ofgastrointestinal injury associated with the acute radiation syndrome;
SR88-22: Holahan, P. K., Knizner, S. A., Gabriel, C. M., and Swenberg, C. E.Alterations in phosphate metabolism during cellular recovery of radiation damagein yaast;
SR88-23: Litten, R. Z., Carcillo, J. A., and Roth, B. L. Alterations inbidirectional transmembrane calcium flux occur without changes in protein kinaseC levels in rat aorta during sepsis;-
SR88-24: Miller, J. II., Wilson, W. E., Swenberg, C. E., Myers, L. S., Jr., andChariton, D. C. ,odeling radical yields in oriented DNA exposed to high-LETradiationj -
SR88-25: Miller, J. H., Wilson, W. E., Swenberg, C. E., Myers, L. S., Jr., andCharlton, D. E. 'Stochastic model of free radical yields in oriented DNA exposed todensely ionizing radiation at 77K;
SR88-26: Neta, R. In vivo effects and interactions of recombinant interleukin-1and tumor necrosis factor in radioprotection and in induction of fibrinogen)
SR88-27: Neta, R., and Oppenheim, J. J. Cytokines in therapy of radiationinjury;
SR88-28: Tolliver, J. M., and Pellmar, T. C. -Effects of dithiothreitol, asulfhydryl reducing agent, on CA,1 1 pyramidal cells of the guinea pig hippocampus in
SR88-29: Vogc,S. N., Kaufman, E. N., Tate, M. D., and Neta, R. -Recombinantinterleukin-l(r and recombinant tumor necrosis factor'iY synergize in vivo toinduce early endotoxin tolerance and associated hematopoietic changes. ( \ W)
Accession For
NTIS GRA&I
DTIC TAB 0UnarmouncedJust lfication,
'cc'4-' Distribution/
Ava i1abi1i ty CodesAvail and/or
ODit Special

AMUEO POO"$S PIAOgC4OLOGY0GSMARCH iNSrITUTS
IcMrU111c AKIAT
R Il INRI 1 RH115. 595-604 (1488 SR88-20
Prevention and Treatment of the Gastric Symptomsof Radiation Sic;kness
ANDRE DtuBolS AND NANCY, FIALA
lal rn ri'I( n sis '~i'q.u~i iid ~i c'rSo,.'. 1,esi~e D s(a~ Diisin.Depitient of ifeditine.
Iw inorreier% it. ,% I Pu ivrit o liu'lisa/i,Scu',es. sandtDeparipnent t!it u/ i ln' ' re
Rash. s/is I, i Rs's.ar I Imnife,. Iehewda. .tluriand 20814
AND
CHESTER A. BoWARD AND VICTOR BoGo
Dei).artmnt il BehaiA',jsral S(,.'ns,'. . tripled Forces Radtss'sudogc Research Itivittute&ificsda. .tarviand 208:4
Dt PAois. Ft"s. N.. Bow-ARD. C. .AND Bouo. V, Prevention and Treatment of theGastnc Si, mptoms of Radiation Sickness. Radiai. Rev. 115,595-604 (1988).
C'urrently, available treatments for radiation-induced nausea and vomiting either are ineffec-tise or reduce performance. The new antiemetic and gastrokinetic agent zacopnide was testedin rhesus monkevs to assess its behavioral toxiicity and its ability to inhibit radiation-inducedemesis. Zac~pride (intragastnic. 0.3 mg/kg) or a placebo was given blindly and randomly in thebasal state and 15 min before a whole-body 800 cGy '"Co -y-radiation dose lexcept for the legswhich wiere partially protected to permit survival of some bone marrow). We determine(; (1)gastnic emptying rates- (2) the presence and frequency of retching and vomiting. and (3) theeffect of zacopnide on the performance of a visual discrimination task in nonirradiated subjects.No vomiting. retching, or decreased performance was observed after either placebo o, zacopndein the control state. Following irradiation pius placebo, 70 emeses were observed in 5 of 6 mon-ke%,s. and 353 retches were observed in all 6 monkeys. In contrast. only I emesis was observedin 1 4f6 monke~s and 173 retches were seen in 4of6 monkeys after irradiation plus zacopridetP 0.01 I. Zacopnde also significantly inhibited radiation-induced suppression of gastnc emp- A
toing. When given after the first vomiting episode in a separate group of irradiated monkeys,zacopnde compietelk prevented any subsequent vomiting, The present results demonstrate thatintragastnic administration of Lacopride significantly inhibited radiation-induced retching, vom-iting, and suppression olgastric empty ing in rhesus monkeys and did not cause detectable behav-io~ral side effects w4hen giv~en to nonradiated monkeys. This observation has important implica-tions in the treatment at radiation sickness. C i98 Academic Priem t0C.
INTRODUCTION
Therapeutic and accidental exposure to doses of radiation greater than 150 c~ycauses nausea, vomiting. and suppression of gastric emptying in man, monkeys, anddogs (1-4). In addition, the normally low doses of radiation that may be encounteredin space could play a role in the man-space program, because some of the symptomsobserved in space sickness and radiation sickness are similar (5). Medications cur-
0033-7587/88 S3.00Copynjhi t. i988 by Acadecsl Prema. I nc.All nghiisofresroduction in any fornm reserved.

596 DUBOIS ET AL.
rently a'ailable to treat radiation- and space-induced nausea and vomiting either areinetflctive or reduce performance ability. For example. we previously reported (4, 6)
that mctodlopramide. but not domperidone, effectively prevents radiation-induced\omiting in rhesus monkeys. However. metoclopramide is known to cause involun-tarn mo\ements in treated patients (7).
Recently. several benzamide derivatives of the metoclopramide class have beenintroduced for the treatment of emesis and/or gastroparesis. and these derivativesmay not have behavioral side effects. Therefore, we evaluated the action of one ofthese agents. zacopride. on radiation-induced vomiting and gastric suppression inrhesus monkeys. Specifically. we studied the possibility of preventing and treating
radiation-induced vomiting and suppression ofgastric emptying using zacopride. Wealso determined the possible side effects of this therapeutic agent on gross behaviorand on pertormance ofa \ isual discrimination task.
MATERIAL AND METHODS
r, ent.-lour male domestic rhesus monke.s..tlaaa mdatta. mean weight 3.1 ± 0.2 kg. were used inthese experiments. Monkeys were quarantined on arrival and screened for evidence of disease before beingreleased from quarantine. They were maintained in an AALAC accredited facility and were held in individ-ual stainless steel cages in conventional holding rooms maintained at 21 ± I'C with 50 ± 10% relativehumidits. Animals were on a 12-h light/dark full-spectruri lighting scale with no twilight and were pro-%ided %, ith tap water ad lituni. commercial primate chow, and fruits.
After adaptation to a primate-restraining chair, six monkeys were trained to discriminate between acircle and a square (correct) randomly presented every 10 s on backlit press-plates. mounted on an eye-lesel response panel (8). An incorrect response or failure to respond within 0.8 s resulted in a 3-mA shock.Incoming efficiency on the visual discriminaion task was 9" ± 2% in the six monkeys. Percentage correctchoice was assessed during 18 min (100 trials) before, and fcr 180 min (1000 trials) after, oral administra-tion of a placebo or zacopride (AHR I 190B. A. H. Robins Co. Richmond. VA). A repeated measuredesign was used as follows: (a) baseline control without oral administration of fluid; and (b) blind adminis-tration of either 0.3 mg/kg zacopride in 5% glucose solution or 0.2 ml/kg of 5% glucose solution.
Twelve other chair-adapted monkeys were studied on 3 separate days after an overnight fast as follows:(1) and (2) on two control days after random and blind intragastric administration of either placebo (0.2 ml/kg) or 7acopride (0.3 mg/kg), and (3) on irradiation day after intragastric administration of either placebo orzacopnde given blindly and in random order 15 min before exposure. These doses of zacopride wereselected based on previous monkey experiments (unpublished observations) demonstrating that thesedoses did not produce noticeable side effects. Studies were performed in the morning and started 30 minafter drug administration and 15 min after either sham-radiation (on control days) or radiation exposure.
On control days, the animals were brought to the exposure room and the doors were closed for 3 min.but no radiation was deliveied. On irradiation day. monkeys were placed between two large. 10--Ci 6wCoirradiators. and the animals received nonuniform radiation exposure through, positioning of lead wallshields in front of and behind their legs (9). Phantom studies demonstrated that a I-min exposure resultedin midtissue doses of 8(X) c(;y for torso and abdomen. 896 cGy for head. 584 cGy for femurs, and 425 LGyfor tibiae )9i.
The six remaining monkeys were studied once in the basal state without treatment and again after irradi-ation as descnbed above but wihout drug administration prior to exposure. After one episode of vomitinghad occurred. 1.3 mg/kg zacopride was given intragastrically, and the study was performed as describedabove. If vomiting recurred again within 3 min of zacopride administration, a second dose of medicationwas administered.
Each monkey was monitored for 3 h on control days and 6 h on irradiation days using a videocameraand a sideo cassette recorder. The videotapes were blindly evaluated at a later time for vomiting, retching,and any other side effect. During this evaluation, vomiting was defined as a succession of strong and briefcontractions of thoracic and abdominal muscles leading to the expulsion of gastric contents through themouth; retching was defined as nonproductive vomiting (4).

RADIATION. VOMITING. AND GASTRIC FUNCTION 597
100 . .
Correct ----- Baseline
90 -- Glucose PlaceboZacopride
so I I I I
Pre 36 72 106 144 180Time Prriod (min)
Fit;. I. Effect of /acopride on visual discrimination performance. Six morkeys were tested repeatedly at2-week intersals without treatment (baseline). alter oral p'acebo, and after oral zacopride administration(6.3 mg/kg).
A pre% iousl. descnbed and validated marker dilution technique (10) was used to determine concurrentlygastinc secretion and gastric emptying during a 43-min fasting period and for 60 min after the injection ofan 80-ml water meal (postmeal period). In the present studies, as previously reported (4), this techniquewas slightly modified in that [' "TcJDTPA (diethylenetriamine pentacetic acid) was used as the markerinstead of phenol red. This intubation method requires only the sequential sampling of the gastric contents,and it permits the concurrent measurement of intragastric volume, gastric emptying, and gastric secretion.A 12-French double lumen nasoga.tric tube was placed in the stomach and its position was verified by thewater recovery test (II). Starting 45 min later, samples of the mixed gastric contents were aspirated justbefore and immediate]% after intragas-ric administration of 5 to 20 ml of a ( Tc]DTPA test solution (30mCi/ 100 ml HO: pH 7.4: 37"C). After centrifugation of the samples, the clear supernatants were assayedlor "Tc concentrations using an autogamma counter (1282 Compugamma LKB Instruments. Inc..(;aithersburg. MD). These determinations were repeated every 10 min during the basal period and afterintragastnc instillation of an 80-ml water meal containing J 'TcJDTPA (3 uCi/100 ml: pH 7.4; 37'C).
Intragastric solumes of fluid I,. I'2. land amounts of 'Tc (Tci. Tc.-. -) were determined at thetime of each sampling using the marker dilution principle (4. 10. 12. 13). Fractional emptying rate (g) wasthen determined for each 10-min interval (1) between two dilutions, assuming that emptying was a first-order process (exponential) dunng a given 1-min interval. However. since g was allowed to vary fromntersal to interval, no general assumption was required regarding emptying over the total duration of the
experiment. We used the following equation:
g = -[og,(Tc,Tc1)]/t.
Net fluid output ( R, in milliliters per minute was then determined for the corresponding interval, assum-ing that R, remained constant oer the given interval and using the equation:
R, = [V, - l' 1 .exp - J. , /[I - exp'f].
Intragastric solumes of fluid and masses of w"Tc were then recalculated, taking into account these firstestimates of fractional emptying and fluid output, which were in turn recalculated. This iterative processwAas repeated until the improement of the solution was less than IP per iteration.
Uhese calculations were performed using a locally developed program and a PDP- 10 computer (Divisionof('omputer Research and rechnology. National Institutes of Health, Bethesda. MD). The assumptionsinsolsed hase been described and discussed elsewhere (0) and are based on original coninbutions byIildes and Dunlop (12) and George (13). However, in contrast to their method, the present techniqueallows correction for emptying and secretion that occur during the I-min marker dilution interval, andthis technique can be applied during fasting. On irradiation day. intervals with occurrence of vomitingwere not taken into account for calculation of i and R.
Statistical esaluation of ,isual-discrimination performance data was assessed using a two-way analysisof %ariance. The statistical signficance of differences observed for fractional emptying rate and fluid outputwas esaiuated using a three-factor (treatment, time. and monkey) analysis of variance with repeated mea-
598 DUBOIS ET AL.
100.090.0
AC -- PlACE9O (mnvaesc: 0.2 m0 )$U 0.0
Lu 70.0 ZACOPME (infteaga*ic; 0.3 mglkg)U.0 60.0
Wj 50.0
40.0z_.j 30.04
20.0
100 I-I
0.070 ie"S 1 emeels
In Si6 monkeys In 1/6 monkey*
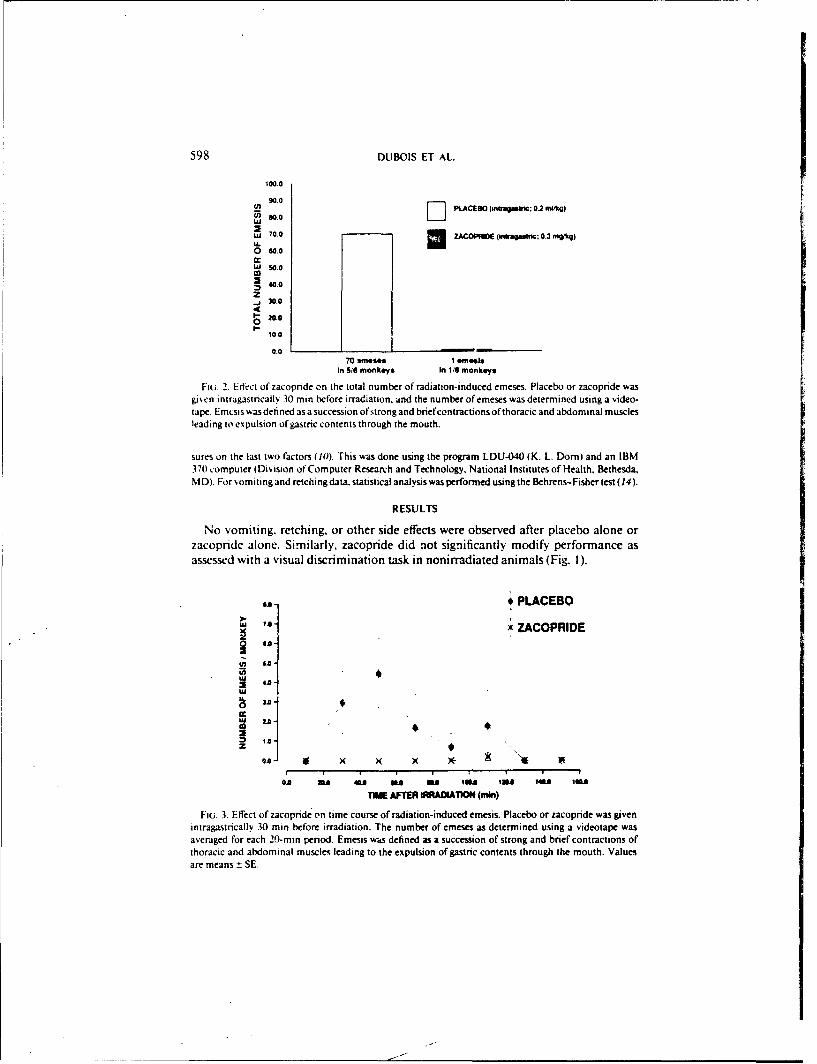
FIG. 2. Edect of zacopnde on the total number of radiation-induced emeses. Placebo or zacopride wasgiven intragastncally. 30 min before irradiation. and the number of emeses was determined using a video-tape. Emcsis was defined as a succession ofstrongand brief contractions of thoracic and abdominal musclesleading to expulsion of gastric contents through the mouth.
sures on the last two factors (I). This was done using the program LDU-040 (K. L. Dorn) and an IBM370 computer (Division of Computer Research and Technology. National Institutes of Health. Bethesda,MD). For ,.omiting and retching data. statistical analysis was performed using the Behrens-Fisher test (14).
RESULTS
No vomiting. retching. or other side effects were observed after placebo alone orzacopride alone. Similarly, zacopride did not significantly modify performance asassessed with a visual discrimination task in nonirradiated animals (Fig. 1).
u. * PLACEBO
7*- x ZACOPRIDE
-it 1.0-
LU
n. I
z O"0.0 2.6"IIII
0* 3 431 63 me is Ml 143 1
TiE AFTER MADWTOW (mi)
Fiu. 3. Effect of zacopride on time course of radiation-induced emesis. Placebo or zacopride was givenintragastncally 30 min before irradiation. The number of emeses as determined using a videotape wasaveraged for each 20-rn period. Emesis was defined as a succession of strong and brief contractions ofthoracic and abdominal muscles leading to the expulsion of gastric contents through the mouth. Valuesare means ± SE.
RADIATION. VOMITING. AND GASTRIC FUNCTION 599
400.0
U
i_300.0D PLco(ituamc0.
cr ~ZACOPF40DE (intragautnic; 0.mnkg)
w200.0
z~J100.0
0
3S3 retches 173 retche.In 6/8 monkeys. In 416 monkeys
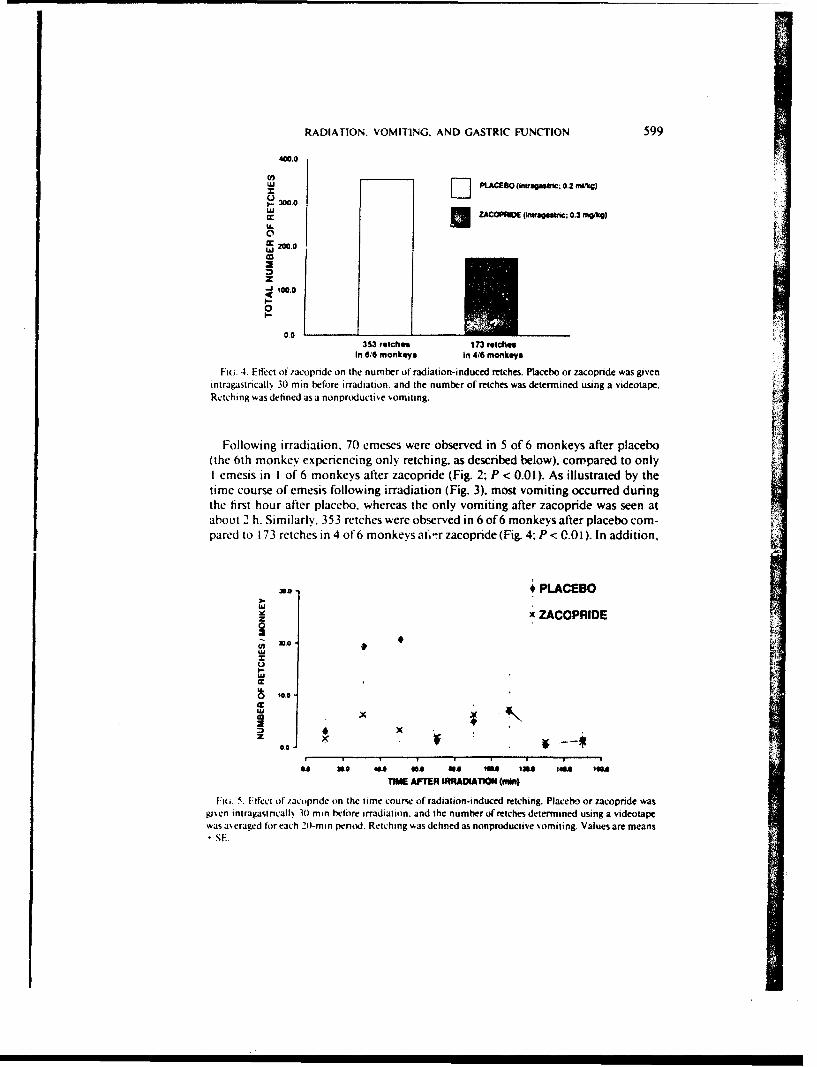
Fiaa. 4. Effect of .'acopnide on the number of radiation-induced retches. Placebo or zacopndce was givenintragastricallx 30 min before irradiation, and the number of retches was determined using a videotape.Retching was defined as a nonproductive vomiting.
Following irradiation. 70 emeses were observed in 5 of 6 monkeys after placeboF
(the 6th monkeN eprncgonly retching. as described below), compared to onlyI emesis in I of 6 monkeys after zacopride (Fig. 2; P < 0.01). As illustrated by thetime course of emesis following irradiation (Fig. 3). most vomiting occurred duringthe first hour after placebo, whereas the only vomiting after zacopride was seen atabout 2 h. Similarly. 353 retches were observed in 6 of 6 monkeys after placebo com-pared to 173 retches in 4 of' 6 monkeys ai.- r zacopride (Fig. 4. P < 0.0 1). In addition,
* PLACEBO
xZACOPRIDE
WAx
z.
TWM AFTER IRRAIATIO (min)
vi. 5. Effect of /acorinde on the time course of radiation-induced retching. Placebo or zacopride wasgisen intragastncalls 301 min before irradiation, and the number of retches determined using a videotapewas a'eraged for each 20-min penod. Retching was defined as nonproductive somiting. Values are means
-SE.
600) DUBOIS ET AL.
U,
iZACOPRIDEJL(mntrga~tric; 0.3 mgikg)
0WA 20A
z 081 ( )( )( )( X X
00 200 ft's 1,00 Iai. 1200 140. INA0
TIME AFTER IRRADIATMO (min)
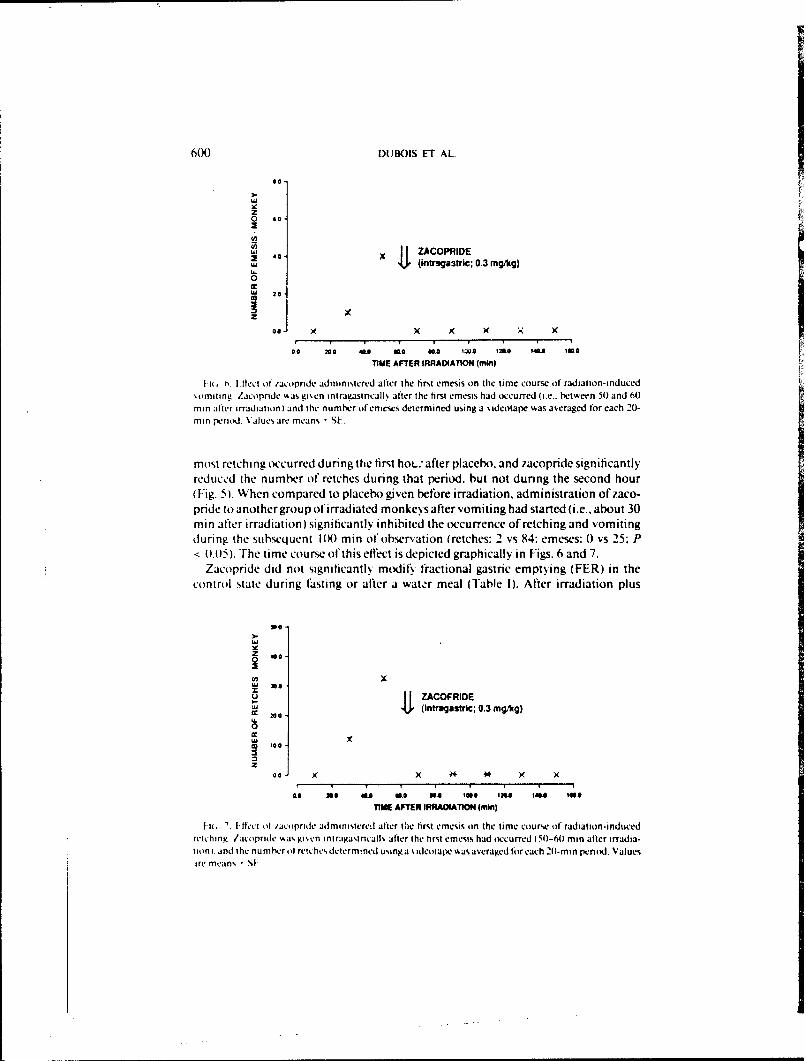
I it H ~iet of racornde administered after the first emesis on the time course of radiation-induced%omniting. Zacopride %a gi~cn intrau.astricall% after the first emesis had occurred (i.e.. between 50) and 60min After irradiation) and the numhc-r ofenieses determined using a %ideotaple %as averaged for each 20-min period. Values are means -SL.
most retching occurred during the first hoL.; after placebo. and zacopride significantlyreduce d the number ot retches during that period, but not during the second hour(JFig. 5). When compared to placebo given before irradiation, administration of zaco-pride to another group of irradiated monkeys after vomiting had started (i.e., about 30mi n after irradiation) significantly inhibited the occurrence of retching and vomitingduring the subsequent IN8) min of observation (retches: 2 vs 84: emeses: 0) vs 25: P
<(0.05). The time course of this etfct is depicted graphically in Figs. 6 and 7.Zacopride did not significantly modii, fractional gastric emptying (FER) in the
control state during ftasting or after a water meal (Table 1). After irradiation plus
OZ 00
(A xWj 300
U flZACOFRIDE200~ (Intuugastric: 0.3 mg/kg)
0
00 x x x4 x ( )
0-0 no "a* its0 "A loss is". iOU. 10
TIME AFTER IRRAOIATION (min)
16 ' If- I It /act pfride adm inistered after tCie first emesim on the time course of radiation-i nducedretching. /acopride ~as gi'.cn iniragastricallt after the first emesis had occurred (50-60min after irradia-tion). and the numher ol reiches determined usinga sideotape %asaveraged or each 2(1-min pento Valuesire means S1

RADIATION. vOMI'ING. AND GASTRIC FUNCTION 601
T-ABLE I
I Ilice ol laopride, Radiaion. and a Waler Meal on Fractional l-repluing Rate (in 1; /rmn)
( sarol hwilrraldi(tio
Placet') 3.41 0.92 421 '0.64 1.1 5 t 0.20 0)36 + O.12tacopide 4,M) 944 4.37 -0.09 2.72 ± 0.94*t 0.66 0 0.200t
\Vq' Valuesare -SF.*' - 0.05 compared to control.t 1' - 0.05 compared to placetxo using ,\NOV A with repeatcld measures.
placebo, gastric empt\ing was signifi.'antly slowed during fasting and after a meal(Table I: P < 0.05). effects which were significantly inhl;-ited by zacopride (P < 0.05).However, irradiation still produced significant suppression of both fiisting and post-mea! FER esen after zacopride (P < 0.05).
DISCUI'SION
In the present studies, we dete;mined the emetic and gastroplegic effects of nonuni-form I irradiation, studied the efficacy of zacopride in the prevention and treatmentof radiation-induced vomiting and gastric suppression. and evaluated the potentialside effects of this medication in the rhesus monkey.
In the basal nonirradiated state, intragastric administration of zacopride did notinduce vomiting, retching. or abnormal behavior. In addition, zacopride did not im-pair performance on a visual discrimination task, demonstrating that it was not be-haviorally toxic for this iask. This finding was recently confirmed using a murinemotor perlbrmance task ((15) and unpublished observations). Determining ifa newdrug has behavioral toxicity is important because this side effect may be as limiting asthe adverse effect (emesis) the drug is designed to treat. e.g.. in an emergency radiationsituation (6).
Following irradiation, we determined the precise time course of vomiting andretching as %ell as the alterations of gastric emptying. Retching and vomiting startedabout 30 min after irradiation, decreased markedly after 70 min. and disappearedafter 120 min. The present 30-min delay differs markedly from previous observationsofa delay of almost I h after irradiation with doses of 400-550 cGy (2) and ofa delayof less than 5 min following a dose of 1200 cGy (17). Thus. the interval betweenirradiation and vomiting appears to be inversely proportional to the dose received.In contrast, both the emetic resoonse and the gastnc suppression were similar to thoseobservcd after total-bod. irradiation in the same animal model with midtissue dosesto the torso and abdomen identical to those used in the present experiments (SOOcGy) (4). This dose of 800 cGy was selected because it was twice the ED 0 for vomitingas previously determined by others for monkeys (2. 17). However, due to the expo-sure system chosen for the present experiments, the head midtissue doses were 30%higher than in our previous studies (4). and the tibia and femur midtissue doses

602 DUBOIS ET AL.
x'ere. resnectixel%. 47"' and 27"; lower. Taken together, these data suggest that theabdomen and torso are the most important targeis for the initiation of the prodromals% ndrome.
In the nonke exposed to 8(0 cGy "Co. intragastric administration of zacopride15 min before irradiation prevented radiation-induced vomiting. hi addition. zaco-pride significantly inhibited retching. although ihe time course of retching appears tohaive been only minimally altered bN zacopride (Fig. 5). Due to the small number ofsubjects, the diffrence did not reach a level ofstatistical sigtficance but it is probable,h,, the effect shown on Fig. 5 was real. In addition. intragastric adminis,..ation of,acopride after the first episode completely suppressed retching and vomiting for thesubsequent I1 min. Since zacopride did not significantly modify radiation-inducedretching and vomiting ,ror, 60 to 120 min after exposure. the duration of the ai,!i-emetic efiect of zacopride in the present model appears to be about I h. Similarly.tacopride inhibited the suppression o,'gastric emptying indUced by irradiation duringfasting and after the water meal. Thus. after irradiation plus zacopride. gastric empty-ing was dec-eased by only 40"; during fasting and by 85% after the meal. Silice themeal Aas given about I h after iacopiide. it appears that the duration of the gastroki-netic eifect ofthe drug. like that of its antiemetic effect, was approximately I h. Takentogether. these observations suggest that the current formulation of zacopride shouldbe admii-istered twice: once before irradiation and once after exposure. Although theimprovement of gastric emptying induced by zacopride using the present frequencyand dose of administration is not complete, it markedly improves the possibility oforal rehydration after exposure to radiation. It is most remarkab!e that zacopride hasan antiemetic effect even when given intragastrically after irradiation and that it canactually interrupt iadiation-induced vomiting and retching.
The mechanism by which radiation causes emesis and gastr.,: inhibition. as well asthe mechamisnl by which zacopride prevents these effects. remains hypothetical. Thecentral nervous s.stem appears to play a pivotal role in radiation-induced prodromals. mptoms. as suggested by the observed rise of plasma 3-cndorphin following irradia-tion (4). This rise is similar to the one observed after exposure to stress (18, 19). whichis known to inhibit gastric function (0). Thus the rise of plasma 1-endorphin maybe responsible in part for radiation-induced vomiting and gastric inhibition, sincethis tpe I'feffect has been observed after exogenous administration of opioids (21).Irradiation could cause the release of 1-endorphin or of another humoral mediatorby initially actiating the peripheral end ofafferent nerves. A direct effect of irradia-tion on the brain app,.ars unlikely for at least three reasons. First. shielding of the are-postrema (chemoreceptor trigger zone) does not prevent ra'Jiation-indtced vomiting(22). Second. increasing the dose delivered to the head by 50", in the present studydoes not modif, signilicantl, the symptoms (4). Third. ablation ofthe area postremain cats does not present radiation-induced vomiting (23). Zacopride is a benzamidederivative with gastrokinetic properties which does not protect against emcsis causedb, apomorphine-induced activation of the dopa:nine receptors of the area postrema(24). 1 loweer. intravenous (iv) or intracerebroventricular zacopride prevents emesisinduced b. either i chcmotherapy agent (25). Since tacopride is not a doparnineantagonist. it is not neuroleptic and does not cause extrapyramidal. cardiovascular, orautonomic nervous s% stem side eltiects. Given these facts, it is probable that zacopride

RADIArION. VOMITING. AND G.-.STRIC FUNCTION 603
prevents radiation-ir.duced voniting and retching by acting at a site different thanthe area postrerna. either centrally or at the peripher-'. but the exact mechanism ofaction remains to tie defined.
In conclusion, we observed that radiation-induced emesis was accompanied bysuppression of gastric emptying in monkeys. In addition. intragastric administrationof zacopride significantly inhibited radiation-induced retching, vomiting, and sup-pres-ion of gastric empilving. Although zacopride does not appear to cause detectableadv erse behavioral side effects, further studies are needed to confirm this perception.1 he present observations have important Implications in the treatment of radiationsickness and it will be important to determine if they can be confirmed in clinicalstudies.
ACKNOWLEDGMENTS
%ke thank D~rs. R. S. *\lphin and W, Smith. A5. I11. Robins Co.. Richmond. Virginia. for generousl%suppl'.ng the iacopride and placebo. We also th:i'ik N. Fleming. M. Fl~nn. J. Stewsart. and J. Warrcnl~ltz10r their \aluablc support in animal handling aj '1 radiopharmaceutical preparation and administration.I his research %%a% supported h\, the Armed Foices Radiobiologv Research Institute. Defense Nuclear\gcnc\. unlder "4ork unit (X59. [ he opinion% and assertions contained herein are those of the authors
and should not he coinstrued as official or reflecting the siesss of the Department of Defense or the Uini-formed Scr\ ice, t'ni'ersits oft Flalth Scien':estor the l'efist Nuclear Agenes. I he esperiments reportedhcr -in Acre conducted acciirding ito the principles set forth in the Guidoikrlhe(ure'and t iiI .ahoraieirr'
Immals/. Institute of Anial Resources. National Research Council. OhFEWk PubI. No. NIFII78-23.
RI-cFivIL: February 8. 1988: REVISED: April 12. 1988
REFFE R FNCES
IR. A. (tN i\ s. Some etlects ofioniting radiation on the physiolog of the gastroiriestinal tract: AResiew, Radhia Re 5. 167-188(19561.
G C. R, Nitmi in'. nd R. W. Yoiit. Emesis in monke~s following exposure to ionizing radiation.hi at .%pai i. n ir, %nIled .46, 1 70-1-72(11975).
VA Ii PAiiHs. J, i'. J %(intt S. M. P. GRiSom. R. E-4(. andiJ. J. CON.KIN. Altered gastric emptying andpres ention of radiation induced %omiti ng in dogs. iaslro'werologi 86. 444-448 11984).P
4 V. 11)iios si . G, P.sf NI i Ii I . P.. R. EFsn. A. D)tRAKOVIC. J. J. CONKt-IN. and A. Dt!BoiU. Effect ofioni..ig radi..tiiir on gastic secretion and gasine motiity in monkeys. (;a.%roenw'r. lg 89. 374-
111 1405)
J I *' K,,[ R and A. (.s \N Bitt1. Etiological factors in space motion sickness.. Anat Spaice Environ.Wied 54, 67>-(,Xl I19 )
f) A INi Biiis. Sptsof radiation-induced gastrointestinal injurs -nd radioprotection. I'harmnato./iher 39. 6"-"2 (1 9SXt.
K S4i Ii I i/I -D~I Iif I i Metocliipramide. 6astro'ntertioi- 77, 768-779 (1979).. %% IA iioi f' and R. 'A. )(i N(. Ninke% performance after partial hods irradiation. .lerosp. Mted.
42. ;i);- SO' ( 11971 1R I .iKo R R Ski i t N . F'. I N~ I itH. v IN i ts. R. A. IDo'..sttt. and r. J. Mm. Vi Iitv.
, oser\ Irom scscre hentatoiiiei suppression using recomitiant human granuloc~te-macrO.phage colons stimulating lactor /-. \11 lloptol 16, 34.4-348 11989).
/0) A. )IN 84)1. Gp fI. NAl It ISN.. P. Ns -tRim wti . and J. 1). GAsRDNER. Gastricemptyingand secre-lion in the rhesus n'onkes. Oni~ J1 /'lii swi 232, !86s-!92 (1977).
IIJ. %I l-tst \N'i. R. J, PIm si ()it, and W. Sttw t s, .omparatise es6aluation ofwater recosery test andfluoroisopic screening in positioning a nasogastrtw- tube. duning gastric secretory studies. Br. I ld.

604 IMBOIS Ii I M
k.. H iti ii snd 1) 1 I~iNI ni'. ho V .Iti r t-,timating the ritesofpsatnrcw retion and cmptuing.kat?'I 1 11, V. 29.'X3-XSJ11 H
13 ) (ni.Ni.% cinicil method lor mraunrng the rate ofgitstnc emptstng: I hedoub'le samplingtc'.trimeal t,,., 9. 2v-242 I
14 (i %% SNI iti g~and %\ G (wiR .Sn',uia Vtlhii. 5th ed.. pp. 87-98. Iowa State Lin..Pres. Vines. I0%J. 981)
I5 B~x,4o. I \ lfi Ind R.\V.N~oi %(.ompnnsn oacclerod and rotard lsittit indetectingethanol- ind acrslimide-induced pe'r itmance decirer-cnt in rats: Res iew ofe~penmentaI consid.-r-atlitifls t rtating rod s%%tem'.Vu., ./e. 7115-787 t91).
I'. V ,4 VJ J\s tis,. andJ U1 N-:s. hastorat toxici :.ndetficacy ofWRY22 as aradtupriite.tint Radixl W, 104. 8S2-194)1 tJX).
I I I 1 Vi 5!I i nk! MI (i Nix~154wii Radition-induce-d emisis in monkeys. Radit' Re', 82.
Is R 4, i i isuN. I V JHi, Riisit p. S. SliNti I.. N. Lime,. C. Ri'. tR. W. Vsi. and F. Bi xst..1.. rid.rphin and adrentxortikcotrop~n are sereted concomitantlN b% the pituitar-, ;];nd .Swc
Po J Ri sii i. I lmNI i., C Ri\11ii . N. 1 1%(,. R. (it iti.%~t. and F. Hi (xHit. Foot-shock inuucedNtrevs inc:reases i-cnd()rphin in rat blood but not briin. \auiri' 270. 618-621) (1977).
.,I \) Dt ii' and H 11 N\it t .i IN). labituation ot'gastnic function suppression in monkeys after re-
rajted treec-aorant asoidance wession'.. Iiol Psi iho/ 6C 524-528 (1978).11 1 I1 iif \- I Iii t. Vo\1 \it. j *VHNot t). and A. IN Bois. Ftfects of met-erkephalin and nlk'-
one on gastric empis ig and secretion in rhesus monkes... On J I'hvksof 245. G I 964i2(X) (1983).N It I OntNN and S C A . %(. I txcusatemetic aitofl ltowitng irradiation. J'roj Saw. E.%p IiiI .Med
I I I HiPAWiN%. I . I . %1i ('.NRtI M. I .B. [NSI Pit . J. JiNsosi. and R BoRISO%. Acute radiation-inrduced % omiting in area rsstrema-iaatd cat%. Radial Res. 109, 430-4391(1997).
-' R S Vt vi w,. \k I Sw i i v.(C. H. J %i K'Ais. 1). V DRtOt'PI I %I A. and 1I .FS,%%( t i (. Zacopride1 1, 11 R - I IlI 'iiBi. a unique and potent gastrointestinal. prokinetic and anticmetic arent in laboratory
animals IbI)i,. i, i 311liii. 482Sti 1988i *bstracttV.I Sti iii., 1, M (sit siit s. and R. S. Ait ills 1he emetic actis6its of centrally administered,ispajtin and its antagonismn h% ,acopridc.J I'/,arm J'/iarnaiii/ 40. 142-1431 1988).

GASTROENTEROLOGY 1988;95:500-7
A I XO ONCS NAOIOSOLOGVINUACM iNSTITUTE
SCIENTIFIC REOiRT
SR88-21
Prospects for Management ofGastrointestinal Injury Associated Withthe Acute Radiation Syndrome
ANI)RE DJUIBOIS and RICHARD 1. WALKERI~itqii,. I)i i..'&s I)p, itn. )eprtmenl of Medicine. U niformed Servicvs U niversitv of the
di h Si wH i , arod Armed For( (- Raidibiolm:v Rewin h Instilutfe. Bethesd,. Mirvliid
The effect of total-body ionizing radiation on the heightened the general awareness to nuclear haz-digestive tract is dose-dependent and time-depe- ards. The medical experience in Russia should alertndent. At low doses 11.5 Gy). one observes only a physicians that high-dose total-body radiation injuryshort prodromal syndrome consisting of nausea, can occur and we must be prepared to treat suchvomiting, and gastric suppression. At doses -6 Gy, injuries.the prodromal syndrome is more marked, and it is The first comprehensive description of the acutefollowed after a 2-5-day remission period by a radiation syndrome in humans was provided bysubacute syndrome, characterized by diarrhea and Vlempelmann and colleagues (1) based on the expe-hematochezia. This gastrointestinal syndrome is rience they acquired while treating 10 patients in-superimposed onto a radiation-induced bone mar- volved in two radiation accidents at Los Alamosrow suppression. The combination of intestinal and National Laboratory on August 21. 1945. and on Mayhemopoietic syndromes results in dehydration. ane- 21. 1946. These cases pointed out the complexity ofmia, and infection, leading eventually to irrevers- the pathophysiology of radiation injury.ible shock and death. The treatment of prodromalsymptoms is based on the administration of anti- In his classic 1956 paper, Quastler {2) hypothe-emetics and gastrokinetics. although an effective sized that total-body irradiation arr,.sts the produc-
treatment devoid of side effects is not yet available tion of new epithelial cells from the crypts offor human therapy. The treatment of the gastroin- Lieberkuhn. The diminished replacement of epithe-testinal subacute syndrome remains difficult and lial cells combined with normal sloughing of differ-unsuccessful after exposure to total body doses entiated cells leads to the depletion of mature intes--8-10 Gy. Sapportive therapy to prevent infection tinal surface epithelial cells. This loss of epithelialand dehydration may be effective if restoration or cells causes a breakdown of the barrier between therepopulation of the intestinal and bone marrow intestinal luminal contents and permits entry ofstem cells does occur. In addition, bone marrow toxic substances into the systemic circulation, whichtransplantation may improve the prospect of treat- (:an be lethal. In addition, as discussed !y Moore (3).ing the hemopoietic syndrume. although the experi- changes of the metabolic balance observed afterence gained in Chernobyl suggests that this treat- total-body irradiation may bear similarities withment is difficult to apply in the case of nuclear those seen after surgical injury. In both situations.accidents. Administration of radioprotectants be- there is an increase in the extracellular component atfore irradiation decreases damage to healthy cells, the expense of intracellular metabolism, increasedwhile not protecting cancerous tissues. In the fu- urinary nitrogen excretion, loss of nitrogen and po-ture, stimulation of gastrointestinal and hemopoiet- tassium. and a tendency to retain sodium.ic progenitor cells may be possible using cell growth In 1965. Bond and his colleagues (4) reviewed theregulators, but much remains to be done to improve accumulated human and animal experience; theythe treatment of radiation damage to the gastroin- postulated a disturbance of cellular kinetics in mul-testinal tract. tiple-organ systems that manifested itself in distinct
tirinig the past .t) yr. much has been learned components: the hemopoietic syndrome, the gastro-
about the physiologic mechanisms causing radi-at ion injury. an( the receit events in Chernobvl have 0016-5085,88/$3.50

August 19O88 RAI)IATION INJURY 501
intestinal syndrome, and the cardiovascular or cen- as rats and mice (7). The median effective dose fortral nervous system sv'ndrone. The' also recognized voniting is -2 (v for total 1)o(id' exposure to ,-rays:that the i(tacute radiation sv'ndronie is characterize(d bY it is believed to be -2 (v for neutron irradiational acute )hase. also Callett prodronial synldrome. also. although some differen(ces mar exist in animalsand a sul)aCUte phase. also called bone marrow and (8). In humans. radiation-induced vomiting is pre-'astroi ntestinal svnil romes. 'hese two phases of c:eded andi accompanied bv nausea and anorexiaradiation sickness are separated by an apparent (1.4): in animals. hypersalivation, chewing, andremission durin g which the patient may be coin- yawning are observed, and these symptoms may bepletely symlptom-free. considered to be the physiologic equivalents of nau-
Although we recognize that bone marrow depres- sea (9). In addition, gastric emptying, gastric motil-sion with its hematologic and immunologic sequelae ity, and gastric secretion are temporarily suppressedis of extreme importance and represents a medical (10.11). For doses >9 Gy, diarrhea is often observed,problem frequent lv encountered in irradiated per- and the prognosis is particularly poor if diarrhea issons. we have elected to review specifically the less explosive and bloody. These symptoms may be ex-commonl (: onsi(ered( gastrointestinal component of plained by the gross alterations of the myoclectricthe Itacute radiation svndrome. ''lh large amount of activity of the small intestine that were observed ininformation on the pathophysioogy of radiation- dogs exposed to 9.4-C abdominal -),radiation. aninduced gut dvsfunction is summarized along with initial increase of intestinal motility immediatelyits implications for current and future therapeutic after exposure was followed by decreased motilityintervention. The long-tern effects of radiation (such 1-4 days later (12).as enteropathy. fibrosis, and carcinogenesis) will not These symptoms are potentially important from abe considered here. and the reader is referred to the diagnostic standpoint because they can be usedreviews or Mlorgenstern et al. (5) and Fry (6). within 1 or 2 h of e.posure to qualitatively estimate
It must be remembered that the degree of gastro- the dose of radiation received. Similar symptoms areintestinal injury following irradiation will depend observed after local irradiation, although to a lesserupont a variety of :ontditions. For example. many, of extent. In the case of local irradiation, the thresholdthe side etfects described in this review are not for vomiting and diarrhea is lowest for the abdomen,observed after exposure to similar doses during local the irradiation of which causes nausea and vomitingirradiation or in preparation for bone marrow trans- after doses greater than about 1.5 Gy (8).plantation. It is not known whether this is entirely The mediators involved in these early effects ofdue to the tow dose rate, to the fractionted irradia- radiation are unknown. Direct or indirect radiationtion. or to concurrent treatment with antibiotics or effects on the central nervous system probably play abone marrow infusion. Furthermore, neutron radia- pivotal role. although areas of the brain that aretion is much more destructive to intestinal crypt involved remain ill defined, as is the nature of thecells than are similar quantities of ,,-photons. Once neurotransmitters mediating these effects (13). Theinjury to the gastrointestinal tract has reached suffi- vomiting center and the vagal nuclei are thought tocient levels to produce symptoms associated with be necessary, but the precise role of the areathe acute radiation syndrome, mechanisms of organ postrema is still controversial (14-16). The stimula-failure and interventions woul be expected to be lion of the central nervous centers could result fromthe same regardless of the circumstances of their the radiation-induced release of free radicals. ororigin. Therefore. this review reports primarily data from other substances such as the endotoxins pro-obtained from experiences with photons delivered duced by intestinal microorganisms that have beenpromptly, as may occur in an accident. Htowever, shown to enter the bloodstream of animals (17) andthese findings should be pertinen't to any situation in humans (18) after irradiation. Alternatively, the pe-which the total dose received an( the tlose rate are ripheral afferent nerves could be directly stimulatedabove a given threshold. by these endogenous substances. Whatever initiates
the general response of the body, a release of variousPathophysiology of Gastrointestinal circulating chemicals (i.e., -endorphin. histamine,Injury prostaglandins, endotoxins) has been observed after
total-body irradiation, but their role in producing theAcute lodiotion Sickness or rodromol early effects of radiation remains to be defined.S '~tn ro me -Endorphin could play a role because endogenous
Immediately after total body irradiation with and exogenous opiates are known to cause vomitingdoses >1.5 Gy. vomiting is frequently observed in all (19), to slow gastric emptying, and to suppressthe mammals that can vomit, i.e., cats, dogs, mon- gastric acid output (20). The role of histamine in thekeys. and humans. but not in those that cannot, such pathogenesis of the symptoms of the prodromat

502 Il tiirs xN1) W\KR(;\S''ROENTE*-'ROI.O(;' Vol. 115. No, 2
syndrome remrai ns unrclear ad pjrobably involves ti ndings of Fine aiid c:oworkers (32-34), who de-histailine I 1 -receptors: histainine 11-receptor agon- tected bacterial enidotoxin of intestinal origin in theists (i riot caulse vomliting aInd they stimulate gastric plasiiia of animials after it variety of severe traunmasec:retionrin ,.ri~gstric eniltiii (21). whereats the( ep~isodles. Eidotoxin-coritaining pairticles in tile ini-opposite is observed Ater total-liocl irradliationi ( III. testine may penietrate the epithiehiii barrier via thelIn contrast. prostaglainliris coold be c.amuidaites ats intercellular route. Thel( inidrence )f dhisrurpted inter-ie(Iiators of tho~ observ-ed symiptoms hieratis (it) cellular tight junctions followed at biphiasic. pattern
the v are releatsec after irradiation (22)and (b) simrilar to that seen for the dletectioni of emidotoxin intheir effects are similar t,, those occurring after rriouse livers after irradiation (35). This increase intotafl-bodv irradiation (24.25). Finally, the fact tlhot intestinal permieability after irradiation could be due
coib nd agtonya nt I gh spinal cord sect ion to the action (of hurnoral mediators oii this organ. Aprevents radiation-induced vomniting (Ii suggests variety of vasoa .tive substances have been shown tothat an afferent or efferent nervous mechanism,. or inicrease intestinal piermneabil ity to endotoxini (:36).both, is involved, although the rreurot ransmnitter mne- For examinple. severe disrUptionI Of the tight junction(halting this effect has riot vet been dIetinedl (261). In comiplex was seen iii rabbits infused with histaminegenerail. there are rio morphologic chianiges of ga~stro- but riot in those animals giveni saline (37).intestinal smooth mnuscles or intestinal 1nuce'ia dur- Although not always associated directly with nior-ig the prodromal syridrorne (12). although some tality. endotoxin may havec profound effects on radi-
a) terat ions of parietal 1 cell uIt rastructo re have been at ion victims. F~or examinple. endotoxins may contrib-described ( 11). ute to immunoSUppression in the host. but thev' can
also produce subsequent beneficial effectF in com-
SubauteGasroitesina Stndrmt romiised subjects (:m8). Such as stimulation of boneSuboute ostoiotstirul nidrnremarrow repair after irradiation (39). Sublethal endo-
Radiation-induced vomniting usually' ceases toxemiia may be beneficial in other types of trauma aswithins 24 h of total-body irradiation. and gastric well. For exaniple. Spillert et al. (40) reported thatfunrction is normal 2 (lays after 8-Cy -y-exposure (11). endetoxin decreased burn severity when given toPatierits and ariimaIs then experience at rel~itively rnice i ntmediately after thermial inijury. On the othersvmiptoni-free period thadt may last 2-7 days. de- hand. endotoxins released shortly after irradiation ofpend inrg on the (lose received. If t Iiis (lose is >'5 Gv. animals (17) or humans ( 18) may also contribute toat second phase of radiation sickness appears within early performance dlecrements associated with radi-1 wk of irradliat ion. One observes stomiat it is. abdoni- at ion.irial bloating, gastrointestinal ileus. diarrhea, arid Endogenous enteric bacteria appear not to play agua iac-posi tive or bloody stools (4) as well as sepsis. major role in pure intestinal radiation death de-dlehydhrationi. and shock. IThis synd(romle is character- scribed after (loses >12 Gv as there was no sepsis orizeol by' ele(;troilvte imibalanrce (27.28) arid, as shown endotoxemia at the tine of dleath in rats with acuteby mnetabol ic balance studies (:3). bears siiilarit ies intestinal injury (41). Furthermore. preirrad iationwith thle Situat ion observed duonring the postoperative containination of the gastrointestinal tract withperiodl and after surgical injury. P'seudomonos aerugilioso (lid not modify survival
The cause of these symptoms is omleX, and their time of animals dying from pure intestinal syndromepathogeniesis is still not (:ompletelv understood. Al- within 3-4 dlavs of irradiation (42). In1 contrast,ter (loses of radiation -2 Cv. the turnover of intesti- (nostirradiatiomi infection from endogenous entericial IcrellIs is decreased. leachinrg to atrophl'~ of the vilIIi bacteria was aii important factor after exposure to the
(4). 1 n acddit ion. rad iat ion pirodluces alterat ions of lower doses of radiat ion that cause latter death by atransport in the raibbit i eor m as ev~rl atecl in vitroi Ombinatioi of intostinal and hemtopoietic injurieswith LUssing chambers. Short circ;uit current. trans- (42).epithelial potential. and resistanc(e were all iii- Intestinal microorganisms are a major source ofc~reased dose-dependently 1-4 days after tota I-body infection in irradiated ind(1ividuals. Changes in theexposure to 7.5-1 2-Cs' y-rad httion (29). These numbers of facultatively anaerobic bacteria, whichchanges are similar to those observed after adininis- could become opportunistic pathogens after irraolia-tration of bacterial toixinis or secretagogues, and they tion. have been monitored in experimental animalsmay be responsible for dlecreased intestinal absorp- (35). Ilea were removed from rats at intervals afterticin of electrolytes, fluids, and nutrients in vivo sublethal (5 Gv) or lethal (10 Gy) cobalt 6() irradia-(30,31). t ion and cultured quantitatively for microorgan isms.
An intestinal injury with immunologic arid phivs- The facultative flora were significantly reduced iniologic consequences is increased permeability of numbers 24 h after sublethal irradiation but reachedthe epithelial barrier. This concept is consistent with preirradiation levels 7-11 days later. Lethal (10 Gy)

August 1988 RADIATION INJURY 503
radiation also caused a reduction in numbers of alteration of normal intestinal barrier function couldfacultative flora at 24 11 aiter irradiation but, in enhance the likelihood of systemic infection as wellcontrast to sublethally irradiated rats, facultative as permit intestinal contents to damage the epithelialpopulations began to increase by 7 days postirradi- lining. Irradiation may cause a reduction of mucusation and were increased several times above normal secretion either through a decrease in the number oflevels by day 11, This period of excessive coloniza- goblet cells in the mucosa or through lymphocyte (Ttion of the ileui by facultativelv anaerobic flora cell) loss from radiation exposure (491. Bile secretioncoincided with the beginning of the time that deaths could also affect mucus integrity (50). Although theoccurred in rats. Disturbed intestinal microecology mechanism is unknown, it was re(ently shown thathas also been seen in other animal models of irradi- the continuity of the mucous blanket can be (e-atiom injury and has been associated with sepsis and graded after irradiation (511. Although other physi-death (35). Changes in the intestinal flora, coupled ologic and immunologic changes are probably alsowith impairment of the normal harrier function of involved in influencing postirradiation microbio-the gastrointestinal tract. allow the bowel to serve as logic events in the intestine, destruction of thea reservoir for pathogens that can enter the portal mucous barrier could alter colonization resistanceand systemic ci rcu lat ions and fuel the ongoing sep- and permit pathogen access to the cpithelium.tic process. Tbis process may become rapidly over-whelmning in a subject further compromised by mar- Alterations of Intestinal Blood Flow androw failure and profound inmmUnosuppressien. Microcirculation
In addition, loss of colonization resistance is asso-ciated with shifts in microbial populations in com- The role of alterations of intestinal blood flowpromised individuals. Van der Waaii (43) has shown in the pathophysiology of the acute, subacute, andthat opportunistic pathogens in the digestive tract chronic radiation syndromes remains unclear. Mea-are the major source of infection in animals with surements of total small intestinal blood flow in ratsdecreased defensive capacity. Colonization-resistant exposed to 5-Gy total-body -y-radiation failed toanaerobic flora contribute to the control of these demonstrate consistent changes. In contrast, intesti-facultativelv anaerobic pathogens. but when coloni- nal blood flow decreased during the first 2 h afterzation resistance is lost. the opportunistic flora are exposure to 10 Gy but increased significantly by 4-6able to multiply excessively on mucosal surfaces. h postirradiation (52-54). In rats exposed to whole-This event is associated with invasion of normally body ,-radiation of either 9 or 10 Gy. Suskevic andsterile tissues by endogenous flora. Selective decon- Uklonskava (55) observed marked fluctuations intamination of the digestive tract with antibiotics ihat blood flcw in the first hours after irradiation. Aneliminate pathogens but do not disturb anaerobic initial decrease was followed by a pronounced in-flora (which maintain colonization resistance) has (:rease at 6 h postirradiation and then by a sharpsuccssfully been used to prevent infection in pa- decrease from the second to the third day.tients with burns (44) or granulocytopenia (45). Extended observations of postirradiation blood
There is a relationship between numbers of intes- flow to the small intestine showed a continuedtinal microorganisms and their translocation to mes- decrease at 6 and 12 mo after abdominal x-irradia-enteri(; lymph nodes (461. Increased numbers of tion. with a fractionated (1.91 Gy/day) exposure ofbacteria in the lumen of irradiated subjects could 28.71 Gy. However, both the jejunum and ileumcause opportunistic infections through this process. showed a blood flow at control level when exposedRecent data may help identify the route by which to only a single dose of 5.74 Gy x-radiation (56). Intranslocation occurs, suggesting that M cells overly- contrast, blood flow to the large intestine was in-ing lymphoid follicles of the gastrointestinal tract are creased through the 6 mo of postirradiation observa-part of a major antigen-sampling system (47). Some tion, but began a decline to below control levels bybacteria can atta:h to and be transported through 12 mo postirradiation.these cells, where they should be processed by Thus, variations exist according to the species.macrophages and lymphocytes as an initial step in organs, source of radiation, method of exposurethe mucosal immune response. If the normal fune- (fractionated or single), technique of blood flowtion of this system is impaired by radiation or other measurement, and time of measurement after irradi-trauma, an easy route of ingress to the body would be ation. The response appears to be triphasic afterprovided, exposure to doses >6 Gy: an initial decrease in blood
Many organisms colonizing the intestine, includ- flow is followed in a few hours by an increase thating those conferring colonization resistance, are lo- lasts a few (lays and, in turn, gives way to a long-calized in the mucous barrier, a major structure lasting decrease in total blood flow.- 450 .m thick overlying the epithelium (48). An\ The microcirculation of the intestine also appears

-104 )1lIWIMIS ANtD WAIkKEK (;ASlR( rl:NrlRnw);Y Vol. 95. .Ni. 2
to be altered atter irradiation. Severity-two hours I Iowevt'r. thierapeu'itic dloses of rnetoiloirariniie doafter exposure to I i-(v nhixv(I neuitron -y-radiatin lnlfot seemn to be effective in humnans if given afterthe villmus I apillarv netwo(rk o the dlog sirmill intes- vo in it i it, has startedi (641. Ili addition. this mnedica-
i'slii lrulel( t'll il i'striictiori. but thn inrtestinal it cIrosses the hllloid-lrajin barrier .01(1 inlirb~ts striati..Iipillar v looiud tl(ii\ vher 1-rainr ot inuco,i \v's inl- uhcpiiie re~epitors 165S). Ili cointrast, thre l1wriliiralcrceased it t hia: tini (.17l. () i lusive ' erootloia'l dolpidiuile antagonist (holliilridlili doles 110t CaIuse(.hi1aiig'es w\ere toild illi the subinii. (silh arteriodes cenltral side effects biec~ause it iibits onily thleof rats 4 uhavs ifter exposuire to 1 4.6-GY. x-radiiatiin dubiianinie receptoirs loc;atedl outside fte blood-brainl58). Siouil.irlv. cliical evidence of iiicroivasi.1ilar barrier (661i and( it pirevents radliation-induiced vonil-
(:hianges inl lunis with radliatioin boiwel disease has itinig in thel (log (10). IHot ever. domiperidone doesbeen furnishedl by Carr et a)l. (591. These studlies also niot appear to be effective against either radliationi-
idio. atedl that alterations were oIbservedl in fte mul- iniducedl oastroplegia or radiation-induced v'omitinigcousal xvast latuirl in patienlts 1 28 nio after radio- in the niorukey' (10.11). Ini conitrast. clinical trialsthiertip\. Ili addiitioin. Silskovic midi 1'kloskava 155) seeml to indicate that dorujerion ina\' lie etfectiveiiservil t hat uerrnehmilitv i rrureiseh tirtiilh inl rats ill hum11anls (67). alt iougli double-hinhd placebo-3 (iavs afteor exposure to whuol-biuhdv y-rauhiationr ot conitroilledl studies will be necessary to) (onfirin this9l 1t) v. i uding.
Ini ats vxiuused to doses of upl to 15 Gy. the A lidimber of newer ahitienleti(: arid oastrokineticiiirlivasiulatulrl of the irite'Sti i'i s founld to be aglents are currently, being tested iii both aiinials anrdnriirl 4 thavs to 4 ino postirradiation (610. However, I atielits. Recently. one( of these mredlications (Zaco-Ater expoullre ti 15--t 30 v. decreased vascu Ian tv pride. A. H. Robins Co.. Richmiond. Va.) wais foundwas observ'ed in all lavers of the bowel. includoing to be effective in the prevention and treatment of thevariations inl 1inirl wvid~thI and obstru(;t ion Oif yes- prodroanal siidrome ) vonit inrg. retc;hing, and gastricsels. which occurred more frequently at thle highler emptying suppression) in mnonkeys while not caus-doses (60). These vascular changes mady be respon- ing undesirable side effects (68).sible for at decreased capillary filtrationi coefficiment.which has beven observedl after irradiation. Ini adi Subicuite WRoiat ionl Svildrorrieturn, later experimnnirts provided evidence of ultra-struc~turalI changes that correlated with these changes The troatment of t- sub~acuLte. gastrointestinalin ca pilIlary filItrat ion coefficient, suggesting that thel syndrome is based onl soupportiveC t herapy to p~reventearly decrease in thlis coefficien t s een in ft(i gr ops in fect ion and dehydration. alt ho ugh ulIt imate so r-exposed to 20) and 25 (;v may result fromt perica pi I- vival depends on bone marrow and( intest inal stemlarv fibrosis. cell[ restoration or repoptilat ion. This t herapy ill-
ncl des plasma volume expansion, platelets, and an-
Prospects for Management tibiotics. wvhich enhance survival after intestinalProdomo)injury caused by radiation. With these therapeutic
Svndonri rieasures, survival may he polssible upl to 15 Gy. butThe prevent ion and] treat merit 'radialition- toital-body irradiation above 20 (A' is riot manage-
ndhuced vomiting c:an be ac;hi evedl witi n ieuroleti cs able. Finally, the progno~sis becomnes much more(clilorproniazine. promlet haine) or even general anl- serious if irradiation injury is combined withI ther-est hesia (61). FHowever, this type of a pproach is not inal or mechanical injury, as may oiccur in an acci-desirable becauISe Of thel side effec~ts of these miedi- dent such as the Chernobvl disaster.C;at ions, wh(' I'icfrt her dlepress gastric em ptyinrg amir Current research is attemp)tinrg to rirevent the darn-appetite arid may increase the risk of pin orary age to thc intestine by using a variety of radioproted:-infection, ants. 'rhese compounds appear to reduce initial
A more prom isinrg approach has been t he use! of damage to stem cells in the cry pts arid therebyant idopami nergic agents. The oldest (h))am ine anl- dec;rease the effect of at given dlose of radhiation.tagonrist is rnetoclopram ide whichi. inr addit ion to its U nder experimental cond it ions a dose reductionantiemlet ic properties, has a potent gastroki net ic ef- factor can then be calculated to quantitatively eval-fect arid prevents rad iatiori-irlIIAX ucdVOrnit ing arid tiate the efficacy of radi oprotectants. For example,(last rojplegia in monkeys (62). This gastrokinetic ac(- the thiol derivative group (ethiofos or WR-2721)tion appears to be i ndeperndent of its ant idoparnin- improves survival of the stem cells of the intestinalergic properties and may be related to a mnetoclopra- crypts in addition to those of the bone marrow andmidle-induced release of acetylcholirne arid other has a dose reduction factor of 1.25-1.60 (31.69).neuropeptitles within the myenteric plexuts (63. Furthermore. ethiofos enemas in rats demonstrated a

August 1988 RADJIATIOJN INJURY 505
dose reduction factor of 1.6 compared with controls ants, possibly in combination with growth factors(70). As the compound was not absorbed into the that enhance stem cell recovery. may soon Le avail-circulation, it appears that ethiofos can exert its able to prevent or to rapidly repair gastrointestinalradioprotective action by a direct. nonsystemic effect damage. Selective decontamination with poorly ab-onl oastrointestinal mucosa. Recent evidence has also sorbed antibiotics can now offset some consequencesbeen presented suggesting that the use of prostaglan- of immune suppression in thle intestine and futuredims alone and especially in (cmbination with W11- studies may reveal means to nonspecifically enhance2721 could prevent damadge to the epithelial cells of mucosal immunitv as systemic immunity (;anl nowintestinal villi. being therefore truly radioprotective be stimulated.and cvtoprotective (69).
Stemn cell survival in the intestine is a probable Referencesevent, even if radioprotectants are not used. For this 1 eplanLLsnH ofa G h cl aito
reaontheindctin o adinitraionof ell svndrome: a study of nine cases and a review of the p~roblemn.growth regulators after radiation offers future possi- An Iner Med 1952:36:279-510.bilities for enhancement of intestinal recovery. 'Fbis5 2 Quasi ler H. The nature of intestinal radiation dleath. Radiatapproach is a lrddV beinrg applied to tho stem c:ell Rvs 156;4:303-201.co0m part men t oft the bon e mnarrow of irradiated sutb- :t. .5oo re FDI. Metabolic c:are of tile so rgitcal patient. Phi ladel -jects (71). phia: WB Saunders, 1959A921-4.
4. Bond VP. Fliedner TM, Archambeau [0. Mammalian radia-Other methods to promote intestinal recovery may tion 1:!thdlIitv: a dist urbanoce in celIlular kinetics. New York:be closer at hand. Post irrad iation enteropathy is Ac~ademriic .1965.exac~erbated by bile and pancreatic proteases (72,73). 5. Morgenstern L. Thompson R. Friedman NB. The modernEffec;ts of these substances mav be enhanced when enigma of radiation enteropathy: seqluelae and solutions. Amthle MUCOUS barrier is lost after injurv (51.74). These ISur 1977.134:166i-72.
6,. Fry RIMI. Experimental radiation carcinogenesis: W~hat haveproblems c:an be alleviated and cellular recovery we learned? Radial Res 1981:87:224-39.enhanced in experimental animals fed elemental 7. Conard RA. Effect of x-irradiation on intestinal motility in. thediets containing amino ac;ids before irradiation rat. Am I Phvsiol 1951;165:375-85.(74--76). Finally, therapeutic effectiveness of dlemon- 8. Young RW. Mechanisms and treatment of radiation-inducedtial diets hats also been shown in patients undergoing nausea and vomiting. In: Davis CI, Lake-Bakaar GV. Grahame-
0 Smith DG, eds. Nausea and vomiting: mechanisms and treat-radiation therapy (77). and numerous other com- inent. Berlin: Springer-Verlag 1986:94-109.pounds such as micronutrients (e.g., selenium, vita- 9. D~ubois A. Mueller C. O'Connell L, Durakovic A. Abnormalmnins A and E) are also currently uinder studs' as gastric emptying and prevention of radiation-induced vomit-p~otent ial rad ioprotectanlts. ig in primates labstrl. Gastroenterology 1985-88:1370.
111. Dubois A. Jacobus JP. Grissom MP. Eng Rk, Conklin 11.Altered gastric emptying and prevention of radiation-induced
Summary vomiting in dfogs. Gastroenterology 1984;8ii:444-8.11. Dorval ED), M~ueller GP, Eng RR. Durakovic A. Conklin 11,
Three types of injury occur in thel gastrointes- Dlubois A. Effect of ionizing radiation on gastric secretion andfnalI tract after radiation. Emesis and gastric su[p- gastric motility in monkeys. (;astroenterology 1985:89:374-
presitm ar casedby echniss sillunkown Summers RW. Flatt Al. Prihoda M. Mitros FA. Small intesti-and have effects that complic:ate radiotherapy and nal motility in dlogs after irradiation in lory. Dig Dis Scithe treat menit of peo[ple receiving exposures in acci- 1 987:3t2:14012-1t0.(lent or weapon detonation sc;enarios. Sufficient 1:t. Ilorisn HL. Borison R. McCarthy LE. Role of the areadamage to thle epithel ial barrier and the intestinal postremia in vomiting and related functions. Fed Prot: 1984;mnicroci rcu lation inmpa irs gastrointestinal function. 4:1:2!155-8.
14. C hinn [if, Wanig SC. locus of erneti c inin full owing i rradiwvhich c:an have let hal con~tsequenlces. The mucosal aton Pr Soc Exp Hint Med 1954:85:472-4.immlune system and the ecology of thle colonization Ii. Brizzee KR. Neat L.M. Williams PM. Tile chemoreceptorresistant flora art! also disrupted after radiation ex- trigger zone for emesis in the monkey. Am I Phvhiol 1955;posure. Thus, mortality and morbidity are inicreased 180:6i5!)-62.
binfectious comJpl icat ions, its well its physiologic 16. Itorison HL. Site of emetic action of x-radiation in the cat. Ifaly e 17. Walke RL ednevGD. 1id 3e CB. AXseptic: endotoxem ia inl
Prges s en md t ~oto tephsolgc radiation injurv ant] graft-vs-host disease. Radiat Res 1975:atnd immunologic consequences of radiat ion injury 62:242-9.t ll the astrointestinal tract. New-general ion anti- 18. Maxwell A. Caffin St.. wells MIT. Radiotherapy. endotoxemniaemnet ics may soon control sonme oebi litating effects of andI nausea. Lancet V18ti:i: 1148-9.
raditio. F rthrmor. N. L~efebvre RA. Willems iL Hogaert MG. Gastric relaxation andraiaio. urhemoeSupportive therapy with vomiting hy apomorphine. morphine andI fentanyl in thefluids aid platelets, as well as controlled (liets, can c onscious (log. Eur I Pharmacol 1981:69:1319-45.now minimize sonic radiation injury. Radioprotect- 20. Shea-Donnhue PT. Adams N, Arnold 1. Dubois A. Effects of

506 DUBOIS01 AND) WALKER CASTROENTEROLOCYVol. 95. No. 2
Moe-oikophl in d nna1( l'loxoint onl gastric enipt ilig andI secre- tract in experimental animals and its conse90uinces for ifec-ton in rhesus mionkeys. ;.\n I Phyvsiol It8:1:245:GC 196-2003. tion prevenitioni. ac~quisition (of lew btitii idi the prevoti-
2 1 Iuot s A Nomp leggi 1). MI vrs L, Castell DOI. H ista mine H, lion and spread of bacteria bietween cage mates. Ili: vanif derrieptor stimuolation iork asos gastit emtyigllsr. s Waaij D.. 'terhoef 1. eds. Now criterid for anltimicrobial therai-
troiltoriliig\ 1978; 74 10328. pv: mai ntenanc~e of digest ive trTact colonizatin rosistaIt.22 Steel 1K. Rallt.- MA.-. \Vnltv % I\V, v al, I Irinarv oei.rt-tiiin of Amisterdamn: Excerpta Meddca 1971:4:3-53.
%t i.nut It-Oi nes. tcr'iii ino, pr~istialatinin 1K and tliron- 44. ',ao Saenle IIKI. ijaseni 11, Sair EW. Selei 'tive dmontani-bo\,iii H3 fron nic(,e opisvd It thiile-liiil irraiation fromi iiatiiin in burn patients: effects oil wound liealiiig. Prilinii-ill -ili,iiiii-d nleutron l I-u it I Radial Bold 1 98W0:0:95-717 narv results. Ini: van der Waaiij 1). Vorhoef 1. eds. New criteriia
2.3 I)idinis A. Do rsal ED,) Stool L,. Fiala N. C onkliin 11. E~ffect of for antimicrobial therapiv: inaintenanice of itigestive tractinizing raidiation n lrostaglaiidiis and] gastriic secretioii inl tolonlization resistance. Amsterdam. Excerpta Nlodiia 3979:
rhesus iniiiikevs. Radialt Re-s 198':110:1289-113. 2 16- 22,
2-4. Nonlplggi 1). M LrI. Caistell 1DO. Duoios A. Effin.t iOt a 45. Older NH. Nieweg HO. Sleijfer DiT. Ot al. InfeCtiOnl prevvil-
pn istaglanin E1., aalog on gastric em ptvi ng anil set;rot ionI inl tio(1 ii granuocys~topen ic patien ts 1w selective decontam ina-rh-s us iiiikevs. I P harmon Il Es p Ther 1980:212:49 1-5. t ion of the digestive tract. In: van der Waai 1l). Verhoef 1, eds.
25. Shea-I )i 1i i PT. \Ihers L.. ( Xstel IDO). Dubho is A. Effei .1of Newv criteria for ant imicrobial t hera py: maintenance of d igos-I inst acvi: oin gastri ni: m3t viig a ml seicretion iii rheosus t ito tract colonizat ion resistance. Amasterda m: E xce~rptanionuies. (;astroni,iril yogv H8078476-9. NlMI-dia 1979:11 :1-6i.
0 C arpeniter DO( . Blriggs 1)1. Knox AP. Striiminger NI.. Railia- 46f. Steffen ER. Berg RD.. Relationship between icecnal 3p03pulatiii
iin-iiidoici-i iesis ili the doti. i-ff Is of le~sions aind dugs. levols of indigenous bacteria aind frailsloication to the meseii-Rid it Ris I 98ti: 18:3(37- Ill. teni: I vmph nodes. Inifect tinmuni 1983311:1252-9.
27 s 3 ter W(I . A\rmstrn \%il WI). Elec oriy It e niot alI is si aftIer totl 47. Siiellvr MIC. Stroher W. M icellIs anti1 host defense. I Infect IDisboilv \-irrailiatiuu. Ramliat Res 1456:5 1 89-204. 1986:1.14:7:17-41.
28. 1 its 1. Gerbier GBH. El es triil vt e liss, the ma in .a useo if ideathl 48. Rozee KR. Cooper 1). Lamn K. Costorton I 'V. M icrobhil fltora ofinomtheipCastroi ntestina I svld ri ne? Radiat Res I 73:55:33- tlit! mouse i leurm mucous Ia ' er and1( epi thtel iat sourface. A P P
28. Environ Microbiol 1982:431:1451-63.29 (iuiter-Sinlitl Pl. Gamlma radiat ion aiffoits active elecntriolvte 49. Lake AM. Btoch K1. Neutra MR. Walker WVA. Intestinal goblet
trans po rt h\ raibit i loom: biasalI Na and CI irauis port. Ail) I cell mucus release. It. In vivo stimu(1lat ion hv anutigen Ii thelPlisiol I!986:231:1 54i)-. immunized rat. I tmmonol 1979:122:834-7.
3.11shbaughl CC (.Su(t toii 1, Richmond CR. The quesioil of 50. Soulli van MF. HouIse 1V. M~ole RH. The moicus.(top ttingeleitri l vie loss in the inotestinial Id1eath svndronie of rad iaition act ion of bile in the small i ntest inie oif tho irradiated rat. Ihr Iilailatw. Raiit Res 19603;3:1.814-24. ExpJ Pathot 1965:46:25-44.
.31 Herrera 1. Gage T. \Inliupt T. 'tigneulle R. Diubois A. Effects 51. Walker RI. Brook 1. Costerton 1WV. MacVittie T. MOMa MI..Of raui iatioii nil iiitestinal absorption ill dogs: protecitiion Iv Possible association of mmucous blanket illtegritv with postir- AW R-2 72 1 Iaibt r . GastrioeinterolIogy 1987:A2.14:14. radiation colonization resistance. Rad iat Res 1985:1,346-
:Q2. Co a P1. Ishivama M. Roizurni S. Woodlruff P. Kaufman A. 57.E inv 1. Role of e ndi tiixoi i of initest inalI origin in earl% deiat h 52. Dielgado G. Butterfield A B. titschil Ii A. et il. \tiasur ociff rom lIarge lbuirins. S org Cvniecoa l Obstet 1974:1:38:725 -30t. blood flow 1wv the inu Itip le radiiadti ye*' rstirIoiliit
H3.. Raivin HA. ine 1. Bliologital imnpliications of intestiinal indo. in radiated gastrouintestinl ltiss'ie. Amt I Cliii Oiucol 1983:toxins. Foil Priii 1962:21 :65A 6:4631-7.
14 Woodtlrufft l'W 1. O'13 rri ll DEK. Rizii S. F-ino I. Role of the 5:3. lanossv C. Intestinal blood flow in gast roin testinal i rrai a tioninti-sti nl fliira in iipor trijil I liifeu(i Dis 197:3,128:29i3-4. svndrome in the rat. Acta Mted ,\cad Sici Huing 19t69:263:1:3-21.
.33. Walker RI. Pitrvazik MI. Assoiition of b~acteria aiiid iildo- 54. jailoss G. Cardiac output and its distribution in the terinalilixill withi piist trailola i-vvilits Ill: Ninnevnian 1, oil. Iiiiinuiio- stage iof the gastrointestinal irraidiationi sv-iniriinl (radiation
Igi. .ijise ique iiies if iertm l i(il rit ia m ati. in irv. H~i. s lIo k lJ. A t~ta M lin A cad Sci H un~g I % 9:2 6:23-9.Filtno: i3 T rs itv Park Press. 1983:1-1 5. 5 5. Suskevic L.N. I lklonskava I. Changes in ce~rta in fu nctionialI
I. W Uiv%,s P., Fiine 1. Priiiuionm of fatal onmlntixiu. sliom.k by% properties of the vslsof the intestines in whtle body%,isiiaitivi! silistuumcs. (;astriiiiieriliig 1117:6~4:285-t31I. gammak irradiation at superlethiat diises. Radiohiologiva 1975:
37 Pnrv.izinik M. Hiker WV. Walker RI. Disruption of the goblet 15:771-5.i'vll itte~rto-lular piiilitioii followiing histamine infusion of uthe 56.1 Volenec R. Votlitka t. Chmielar V. Brich P. Vavrova EK.
rihilit loomi. Expierienia I 983t:39:5 14-8. Changes in regional bloodt flow atl Late intervals after singleIli Walker RI. Ptntial uises tf iiiilotxiii in i.iifmpriiliisi-l ili.- aiii fractionated x-rat- irraidiatiiin iilrrlts. Sb) Ved Pr lek Fak
.3s.li: Noi v A.. oil. tBeiti.al effoits iif taniotoxizi. New I nit Rarlovv 197!1:22:459t-69.Yo'rk: lPleiiiii 198:3.19-212. 57. Ratial 1, Baun S1. Wvant DE1. Cainie intestinal yasoaitivit\
ti) \lim ittie T1. Walker RI. IEntotoxin-iiuiei alteratioins inl hiring the development of the gastrointestinal radiation stol-Sailil vriiiimliiesis: i.iilunv-stinnlatiulg faitoir. icilionv- lriime. Radiat Res 1972:5t0:5"8'18.
fourinig tolls in Cultuire. id growthl Of C~ells ill iliffilSin 58a. Eiliv HA. (:asaritt (M;. Initestinual vascular chanlges in thelirilrs tEx\i lenmatil I 978.:t 1t-8, acu~ti. radiation intestinal svidiroiiii. Ili: Sullivani ME. ed.
41). Spilolrt C R. G litiiiii S.S. MiCovt-rn III Jr. lazaro ElJ. E~fis(ts (if Casirointestinal raidiatiin iiury. Neow York: Exiorpita Medica(otitOtosinl in iinirin uoiiriis. Ailv Shoi.k Res 1981 :5:16t3-6i. 19t68::385-45.
41 f-;.rai P1. let.ksoii RI.. %lariatio NIS. The iuutest illa r,iiatiiii 5!V Carr NI). Pullen tBR. Hasliton [IS. Stcoliolil [IF, %tiirovasicilarsvnroiiii. sepsis unit voiiloxiii. Raidial Res 19(35:131442- studios in human radiain bowel disease. G;ut 1984:25:448-
.)4.42. 3 ;trai i 1I', Jackson RI.. Mariano %IS. Effei.ts of pseiuiomoiias til. Erikssun B. Microangiograpubic pattern in the small intestine
t iuitimiilatin or iititiioi. ideconitaminlationl iif the intestine iif the icat after irradtiatioin. Scnanct I Castroeteorl 1982;17:oil aizite raitiation lethality. Radial Res ',985 1034:1115-40t5, 887-4i.
4:3. 'an der Waaii 1). Thle colonnization resistanc~e ot Ile( digestive fl. Cmirdts RE. Animal-model stuidies of radiation-induced

,ugus. 1988 R1 DIATION INjURY 507
emesis and its control. Technical Report SAM-TR-86-26. San 71. Monroy RL, Skelly RR, Davis TT, et al. Enhanced hematopoi-Antonio, Tex.: U.S. Air Force School of Aerospace Medicine, etic recovery after bone marrow transplantation in a monkey1982. model using recombinant human granulocyte-macrophage
62. Dubois A. Danquechin Dorval E. O'Connell L. Durakovic A. colony stimulating factor (GM-CSF. Fed Proc 1987:46:1365.Conklin JJ. Treatment of vomiting and of gastric emptying 72. Geraci JP, Dunston SG. Jackson KL, Mariano MS, Holeski C,suppression in primates after gamma irradiation. I Nucl Med Eaton DL. A reexamination of the role of bile in radiation
1984;25:96A. induced intestinal death in rats, Radi3t Res 1987;109:47-57.63. Crosswell AR. Buvniski JP. Metoclopramide mechanism of 73. Morgenstern L. Hiatt N. Injurious effect of pancreatic secre-
action studies in the chemically or field stimulated guinea-pig tions on postradiation enteropathy. Gastroenterology 1967;ileum. Fed Proc 1984:43:3867A. 53:923-9.
64. Sokol GH. Greenberg HM, McCarthy S, Sledjeski L. Lyman G. 74. Langlois P, Williams HB, Gurd FN. Effect of an elemental dietRadiation induced nausea: the comparative efficacy of oral on mortality rates and gastrointestinal lesions in experimen-metoclopramide versus prochlorperazine and placebo. A tal burns. I Trauma 1972;12:771-6.double blind randomized study (abstrj. Proc Am Soc Clin 75. Hugon IS. Bounous G. Elemental diet in the management ofOncol Annu Meet 1986:5:970. the intestinal lesions produced by radiation in the mouse.
65. Schulze-Delrieu K. Metoclopramide. Gastroenterology 1979: Can I Surg 1972;15:18-26.77:768-79. 76. Mohiuddin M, Kramer S. Therapeutic effect of an elemental
66. Laduron PM. Levsen !E. Domperidone, a specific in vitro diet on proline absorption across the irradiated rat smalldopamine antagonist, devoid of in vivo central dopaminergic intestine. Radiat Res 1978:75:660-3.
activity. Biochem Pharmacol 1979:28:2161-5. 77. Bounous G. The use of elemental diets (luring cancer therapy.67. Bernier J, Huys J. Jomperidone in the symptomatic treatment Anticancer Res 1983;3:299-304.
of radiotherapy-induced nausea and vomiting. Postgrad MedI 1979;55(Suppl 1):50-4.
68. Dubois A. Fiala N, Bogo V. Prevention and treatment of thegastric symptoms of radiation sickness. Radiat Res (in press). Received November 12. 1987. Accepted March 7, 1988.
69. Hanson WR. Radiation protection of murine intestine by Address requests for reprints to: Andre Dubois, M.D., Ph.D..WR-2721, 16-16 dimethyl prostaglandin E2 and the combina- Department of Medicine, Room A3075, Uniformed Services Uni-tion of both agents. Radiat Res 1987;111:361-73. versity of the Health Sciences, Bethesda, Maryland 20814.
70. France HG Jr. Jirtle RL. Mansbich CM I1. Intracolonic WR The opinions and assertions contained herein are the private2721 protection of the rat colon from acute radiation injury, ones of the authors and are not to be construed as official policy orGastroenterology 1986;91:644-50. as reflecting the views of the Department of Defense.
__!

AOSS*AC" IMBYITUTIF
IN I. j. RA I AV. 14 10.1 1988, i_ 54, No. 4, 545 562 =11116TIFICREPORTSR88-22
Alterations in phosphate metabolism during cellularrecovery of radiation damage in yeast
CARO)LI NE M. GAB~RIEL. and CH ARLES E. SWF\BERI(;t
Radiation Biochemistry D~epartment,Armed Forces Radiob.ology Research Institute,Bethesda, Maryland 20814-5145, I'S.A.
Received 14 .4tigust 1987; revision received 13.April 1988.;MCOPied 14 AIpril 1988,
We ha\ c examiined alterations in phosphate pools during cellular recovery fromradiation damage in intact, ss ild-type diploid yeast cells using 3 IP nuclearmagnetic resonance (N\IR) spectroscopy. C oncurrent cell survival analysis wsasdetermined follow~ing exposure to "'Co -,-radiation. Cells h~ld in citrate-buffered saline (CBS) showsed increased survival witth increasing time afterirradiatnon (liquid hoilding recoivery, LIM) with no further recovery beyond49 h. Addition of l(R) mmol dm 'glucose tio the recoverv medium resulted ingreater recovery. In the presence of S mmoldm 2-deoxvglucose (2-l)G),LAIR swas completely inhibited. NNIR analyses w&ere done on cells perfused inagarose threads and maintained under conditions similar to those in the survivalstudies. P was observable by N MR only when glucose %%as present in therecioverv medium. In control cells, ATP concentrations increased and plattauedwith increasing recovsery time. With increasing radiation dose the increase inATlP was of lesser magnitude, and after 2X)yno increase ssas o~bserved.'Piese observatiiins suggest that either !he production of ATP' in irradiated cellsis suppressed or there is enhanced ATP utilization for repair of radiationdamage. In CBS with l1ttmmoldm 3glucose, a dose-dependent decrease inpolv phosphate (poly.P) s%as detectable w ith no concurrent increase in inorgsenicphosphate (P,). In the absence of an external energy source, such as glucose,there was a slight increase in P'. This suggests that polyP? may be used as analternative energy supply. When 2-DG was present in the recovery medium,polyP) decreased, but there was a simultaneous increase in P, with increasingradilation dose and recovery time. Tlhis su~ggests that the polyp are hydrolyzed asa source of phosphates for repair of radiation damage.
1. IntroductionP~hosphorus (31iP) nuclear magnetic resonance (N\IR) spectroscopy is currently
being used to monitor cellular metabolism and energetics both in vitro and in vivo(Navon et al. 1977, Jacobsen and Cohen 1981, Nicolay et al. 1983). In vitro NMRspectra can be generated from perchloric acid (PCA) exiracts obtained from cells(Navon et al. 1977). 1 lowever, this technique involves total disruption of the cellsand therefore does not allow dynamic studies of cell metabolism. In addition, PCAextracts permit the observation of only acid-soluble metabolites. In vivo NMRspectroscopy can bK used to observe major mobile (unbound) metabolites in cells ortissues. Large macromolecules or highly immobilized (bound) molecules will not
t Reprint requests and correspondence should be addressed to: C. E. Swenberg,D~epartment of Radiation Sciences, Al RI, Building 42, NMNCNCR, Bethesda, MD) 20814-5 145, V.S.A.

546 P. K. Ilolahan et ail.
generate anl observable NN\ R resonance. Biecause N \ JR spectroscoipy has aninherent lack if sensim t\' it irequires the use Of a large number att cells tfor Obtainingspectra \%ith it L ad signal -ta-noise ratio. (Cells are densely packed, \\li ch results inlat nah oliclState anld etuat~~ l Cell death due to the aIccumulation ot toxicInelta bol teS. A techn11] iqu th11at MoVrcomes these d Iiticulties and al Ilo\\s l arge n umb ersat CLlS til he iM1iiitaIiitdl 0 . \ able! Ph\SiolOgical state has been described b\ FOxalland (Ohlen (I 1983). 1In this me-thod, cells are embedded in an agarose mnatri\ anld canhe perfuscaU \\ th media IOr ha ffters containing nutrients. D iffiusion ot metabolitesthro ugh the agzarase reaches at vrv rapid equilibrium ( loxall elida. I 984). Ini thisStud\ \\e have utilized this technique to mionitor phosphate metabolism duringliq a id holding recoverv tldlow\Ing imnizing irradiation under a variety of conditions
tar wild-type: diploid yeast.Iiph Id evast (S.acharwinices ceri'7isiac) exhibits w\ell-dueumntted recoverv
frami pontially lethal dJinIaVe (iqi(Ld hloldinlg or dark recovery) (Patrick el oid.I 9041. Inl yeast this rci Ocr\ has,- been attributed to the repair at' dIouble-strandbreaks unIder ni ngri i\th ci1itos(laknegShae ia.I980). Since It isk iias\ i that enecrgy: is rVLlui red for recover\ tram radiation dlamage (Jam ieal. 19)77)nl add ition to cel-l grox% th anld maintenance (Jam ef al. 1 982), this investigation w\as
Undertaken to determine if Changes inl "T1 N\I R spectra \old correlate w\ith cellsensiti\ it\ to radiation killing. Repair requires the synthesis of fragments ot nucleicacids, production Of repair enzymnes. and encr. USually' in the form otf .ATP'.Pal\ phOsphates, fO iund in \yeast, are piolymers of Orthiiphiisphate (inlorganic phos-
phate) link-ed %\ith high-energy phiisphoanhyd ride bonds (Yoshida 19)55). Tlhe chainlength of these poly phosphates can vary triim several to several hundred phosphateresidues ( I lariOld 1 900. Tlhese pOly phosphates can be separated into tour tractiOns:PP 1. PP. lPP, and IT,~ (H arold 1906. Kulaev 1 979). PP and lIT, are the mosthighly pal~merized (nican chain length of 5i and 260) phosphate residues, respee-tivelv) and appear to he located neatr the cell plasma membrane. D~ue to their lengthanid associatior .\ith the plasma membrane, these phosphates are probably notdetected by NN\ I R spectrosciopy. Fractions IT,1 and IT, are much less polymerized,hit% .g chain lengths of 2 401 orthophosphate residues, and are located throughoutthe cell, Ii the c-tiiplasm. nucleus and vacuoiles ( Kulaev 1979). The physiologicalrile Of pi ii phosphates is not kno\\ n, but it has been proposed that they may serv eal its a stare of phosphate biind energy. (Ib, as at phosphate reserve, and iir (c) atsdrect phi sphilrvati IV agents In nmetabolic paths\ avs (N\I udd eli al. 958, N icOlay
it al. 1 983, Wood 1900)t. , 21 investigations (see Kulacy 19)79)) have shown thatI ractii n IlI), accunmulates du riiig nlucleic aid hiiisvnthesis w\hereas fraction Pl10 istisvd tu r the biiusvn thesis ut nuicleic acids and nucleotide triphOsphates. IHence. \\eha~ o measuiredl the variatiuons in phiosphate puOOIls during liquid holding fulloiwing
- irraldiatiOin In %ild -t vpe eASt using NN\ R spectroscoupy. Changes in the cellulariiitiblii si i t phi usphates m\crc correlated with cellular recOivery determined 1w cell-at r% i. Al experimnrts.
2. Materials and methods2. 1,. (e/l culture and svrrii'al assayv
Wild-t\1pe diploid \east cells tuObtained from Yeast ( ,enetic Stock ('enter,litrkeles, (A: .Saechro,Pnvccs reirevisiac, strain X2 I X4XS288C) were Cultured inY 1K 'I)tiied iunilI per cent \cast extract. 2 per cent peptone and 2 per cent dextrose;
ciw i LabOratiories, Inc., \ ladisOin, W~I). L ong-term Cultures \\ere: maintained at

Phosphates andi radiatinp damtage 547
4 C oin agar (2 per cent) platt-s. For dark recovery experiments, cells .vere grown to
plateauff Phase inl iquid YEITD broth at 30 C tor 24 48h before experiments. CellsI. r sjti ial analysis \\~ere dil uted inl phosphate-buffered s~'ine ( PBS), anidirradiated ill a cci11 densitv ofi I X nilcel l. Prior to irradiation, cells \%ere
soicareud fo r 31 S to cr50 re at sinlle cell suspension. Cell survival \%as determi ned bydili nig i he cell suispension and plating On nutrient Y EI)D avar. Cells were held oil
ice to enISUre no rcpar r of radiation diraage before plating. Agar plate.; were then
inciIC~bite'd at 304 (' for 3 4 days to allow% iolony torrnatior *DI )laUd platingv experiments \\erc conducted by holding the cells (I 0" cells fil)
in PBlS til t he dark it 204 C for the i ndicated times before planing oin nutrient agar.A-s before, thie cells %\ere incubated at 30) C to allow colony formation after plating.I n somne experiments, citrate-buffered saline (CBS) was substituted for PBS duringthe irradiation and recovery periodL.
2 2. Radiationt wd fir i hemiical treat ment
ilateral I rradiatio ns \%ecre pertormed using a "Co -,-i rradiator (dose rate
appi d\ imael v2 m iin). Cells \\eire Irradiated atl 4 C in at continuously shakenit t-\\ .iter bath.
For c\pvrinlcnts involving che-mical treatments the chemicals w\ere presentwdUrin-, i rrldilt)Ol tindai the recover:; period. 2-I )oxv-il-glucose wais obtained fromSignia (herolicall Co. (St L ouis, mo( ). Solutions were prepared in PBIS and filtered
through af sterile 442 pm Nalgene filter on the day of the experiment. Cells wvere
re sspended inl PBlS containing the chemicals i'mmediatelyV prior to irradiation.
2.3. Prepwat iof eeiis for VA.IR e.xperitpents
A\ssignment of resonances in the NM H spectrum \%as accomplished by use ofPCA .\\traicts (of kclls. Resoinances were aissigned b\ comparing chemical shifts to
litnerat ure \il des (Salhianv et al/. i 975. Navon e't a/. 1479) hy (1) ptertorroing plitit ratioins and obster\ ing Changes in chemical shifts, (2) adding certain enzymes (e.g.v]l ti w-ti- phuisphanase) anid noting resulting alterations in the spectrum, and (3)
piigthe (.\trict 171 akdditionl Of kiiom~n phosphorylated compounds and notingspectral chatngis. When obtaining IPCA extracts, cells weiv eIrradiated at a celltleiisitv (it ; x 10" cells nil anid theni pellered, %\~ashed, and resuspended in 049 per
ccent \A a(I.lho iphatvs \% crc extracteid \\ith cold 0i per cent ( w v ) perchioric acid,then t'rvezi-t hasmcd three. times in at d r\ ice cthanui! slurr\. The extract waistunitritugicl and. ile( t-iriatit, containing aIcid-soluble phosphates, was neu-traih/id to p11 7 0i h1% additionl of 2 nioldn t- .I 2co, Preci pitateid KC10 4 %%vas
reills ci b\ ccinrifuganilton aiid 1ISbsequent filtration of the extract with af 04? pm
htnr D,) IS sid&Ld ( Wi per cent lby iOu oli to the samrple! Solution.
/ot 711. dark red our cryeperiments "tcrc Co nducted using 3 x 104)' viable cells.Th iuf eelIs re cent rifutugei nd resuispentded in a nmixSture of 442 ni! 04'9 per ':cnt\A]( anmd I14nml li~ gii-elprtic(lgt 4 agarose 4 ~(Corp, Rockland,\l I to I finalt on m'Olcitratiion ot 4491 per -cnt lg"t agarose. Tihe iigarist cell mixture\i, thlen c rurit'ed, Uinder mild prussuri, thriih narrow (445 mm id.) Tetlon
tmibillo tit ii ice bath ito a 12-mm sirv\% -cap NIl tubhe (Wilniad, Buiena, NJ).eclI hiss diirin riv\trtisioin \\a s - 11) per cent. The 'spaulhetti-I ike' agarose threads
set? led ait the botutonm (It the tube inl Suith at \\!I\ that all cells wecre w\ithin theid toti-req ic t rarimit ncr rcet ~cr regLion ot tile N \11H probel(, A hi l lo%% Kel-FI
inset t \%is sitated Ill th' N \I tubei, illii4M irig1 t he c-ontinulous perfusion iif the cells
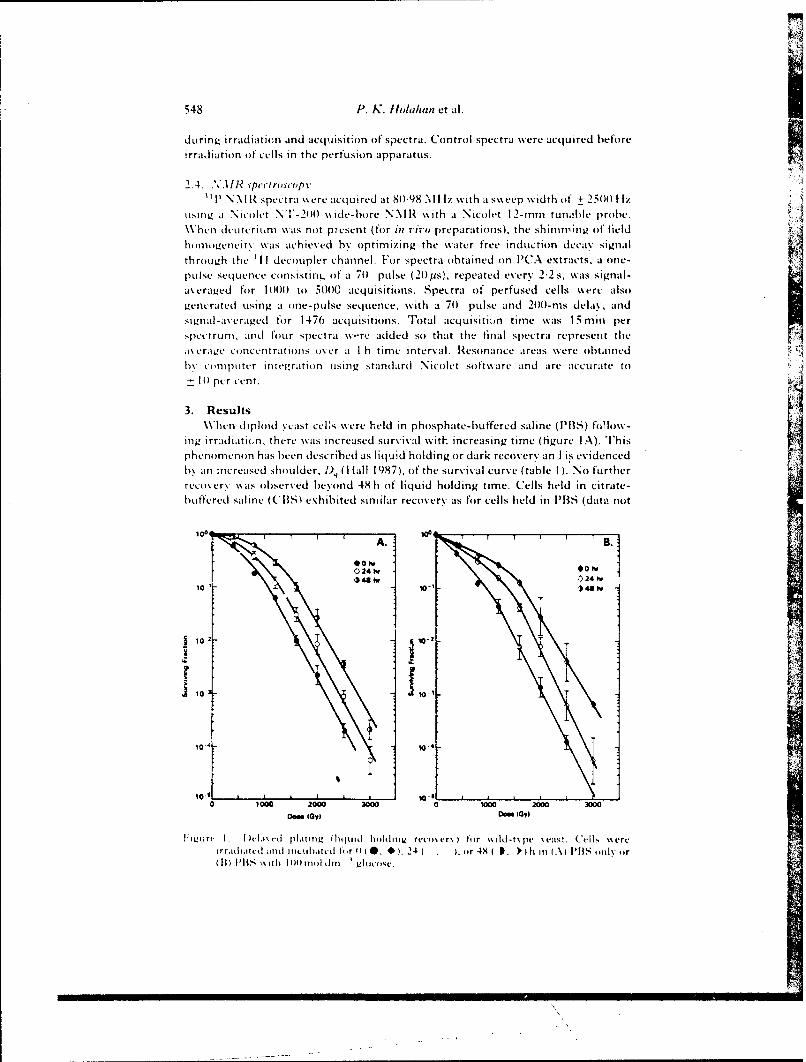
548 P. K. Iioltihin et al.
during irradiaticii and acquisition of spectra. Control spectra were acquired beforeirradiation of cells in the perfusion apparatus.
2.4. si) pectroscopy
TP NM R spectra %%ere acquired at 80-98 Mliz with at sweep width of + 2500 1 lzusing. at Nicolet NI!'- 200 w\ide-bore V\1I1 \with a Nicolet 12-mm tuna!ble probe.When deiireritm \ as not p, esent (for in vivo preparations), the shimming of field
homogeneiit\ wais achieved by optimizing the water free induction decay signal
through the 'IjI decoupler channel. For spectra obtained on PtCA extracts, a one-plse Sequence coInsISting of a 70 pulse (20 pus). repeated every 2-2 s, %wais signal-averaged for 1000f to 5000 acquisitions. Spectra of perfused cells were alsogenerated using at one-pulse sequence, with a 70 pulse and 2(X)-ms delay, andsignal-averaged for 1476 acquisitions. Tlotal acquisitio~n time was is mm per
spcctru~in aild four spectra %\-,re added so that th-e final spectra represent the*I\ era~e co ncentrat ions over at I hi time interval. Resonance areas were obtainedN,b\ comniptiter in teitration using standard N icolet software and are accurate to
-0per cent.
3. ResultsWhen diploid yeast cels w\ere held in phosphate-buffered saline (PBS) follow-
ing irradiatin.n, there wais increased Survival wvith~ increasing titne (figure IA). This Pphenomenon has been described ats liquild holding or dark recovery an .] is evidencedhv an increased shoulder, 1), (1 fall 1 987), of the survival curve (table I ). .No furtherrecovery \\as observed beyond 48 h oif liquid holding time. Cells held in citrate-buffered saie(CBS)5 exhibited similar recovery ats for cells held in PBlS (data not
l0~~ A. j001 24r
102 ]v 00. 4h
1104 11012 h
10o 10-14 1
10 2- 102
0 10010 2000 3000 0 i000 2000 3000Do". bOy) Don I0,
Iiinr 1. I )vI.i vd plaiiz (lipui holiitz' rccoivry) ir %O id-t~ pv '.es!. Cells \%ereirra~id iund ificiiud for 1) (0. *, 241 . , or 48 I , I h in (A PB 'S oily or

PhJosphates (irid r(idiati(,n ciinage 549
Tab.le I Rrco~ery of n ii)- tvpv Yeast ceilS (d1,r711 liqjuid hccldiniz re~ccmerv in PBS s" oh or\\ih0Llt gluecose or 2- decvxgIuceo.
T[ime after irracdiauionu (11)
4424 4X
1), I) I) I) ' RU R R
IM 2 ,50 525 250) 70 123 1-12 250 1025 15;7 1 2')I l ilt 4-I 4Mmol dill
LI'ducctf 2 25 520 225 X50 1 43 1II') 225 M(10(1 173 1-41PI) i + 5 illni'I d m
2-d~C0X\--g1LuCsCc 2010 370) 230' 250 (4) 0XX 23(1 25)) (190 0,48
D1), I) re in mitls of G\;~Reec\ cn r ratio I CiI( eits dose )c r de axed platinog ditided hv cse fo r inime-diare
plcr I l, hcr xcxiRixii 1 0 ') cr i ~i : RR, 1 IS i~cxrx ix i
shox\ n). W\hen 100mol dm 3glucose is present in the recovery medium, theShoulder of the Survival curve increases more rapidl\ with no effect on the slope(figure 1 13). Recovery was expressed as the recovery ratio, defined as the dose of'radiation after delayed plating divided by the dose after immediate plating to obtainan islisurvlval, e.g. 10(1 (table 1). T[his ratio demonstrates that there is greaterrecov ery \, hen 01lucose is present in the holding medium.
In figure 2, representative NN U spectra of both it, ivo and in vitro
prep)arations of cells are shown. T[his figure shows the effects of radiation at 2000( (ivKon the in vivs, cell suspension perfused with CBIS in the presence (If 100) mmol dmnglucose, lor both immediate and 24-h liqluid holding time on the N )cl spectra.)nl\ in the presence of glucose are the A' resonances easily resolvable tor in vivo
preparatioins (peaks 3, 4, and 8). PCA extracts of unirradiated cells were used for theidentificatioin of resonance signals. The Changes in NNIR observable phosphates inl'(A exti acts of , east (aCid-soluble phosphates) \\ere determined as a function (of
dose (figure .3). 'The data are plotted as a pt'rcen~ttg of unirradiated controls; thearea under each NN\ I R resonance signal x% as taken to be prolportional to the amountof phosphate present. All observable phosphate metabolite concentrations in-cased w ith increasirv (l ose. In particular, the sugar p! osphates (SIP) increased
o-f d afer504) 20)))) ( altho ugh at higher doses the increase was not ats
d rainal ic. 'The levls of middle phosphates (N\III) of long-chain polv-phospbatesand the Y, fl, acnd -,phosphates oIf ATP'l also x\ crc increased. Only the fl-A'l'I reson-!W
~e'.e ded a t rue indication of' A'TP'I levels, since the x and -, resonances[;ilso ci, ntaifh . co r' ributions frocm :2 and VADP)1 phosphates, respectively. TIhe
-APresonance (Icc cior'a panlel) ocx erlaips the resonance from the end phosphates(IT) of llong-chainl poklilospharcs. W\e tentatively attribute this increase in all
phlc cph itts \ Ith linicreasing (lose to he the radicition- induced disruptiol (If
hounod onn-el lular phosphate molecu les, seb ich enables more cot c[ lete extractioinofl phosphates by l'CA.
111 7 170 3i11) NN~ I R spectra were acqu ired for cells perfused with oxyvgenated (CBSO'ic ng li('ucd holding, recoverv for 24hb follox\ ing irradiation. .N I spectra were.icq ui red on ctlII, embhedded in iga rose th reads before irradiation as internalco ntro ls. IThe area under each res onance peak \\ias determined relative to the area
All
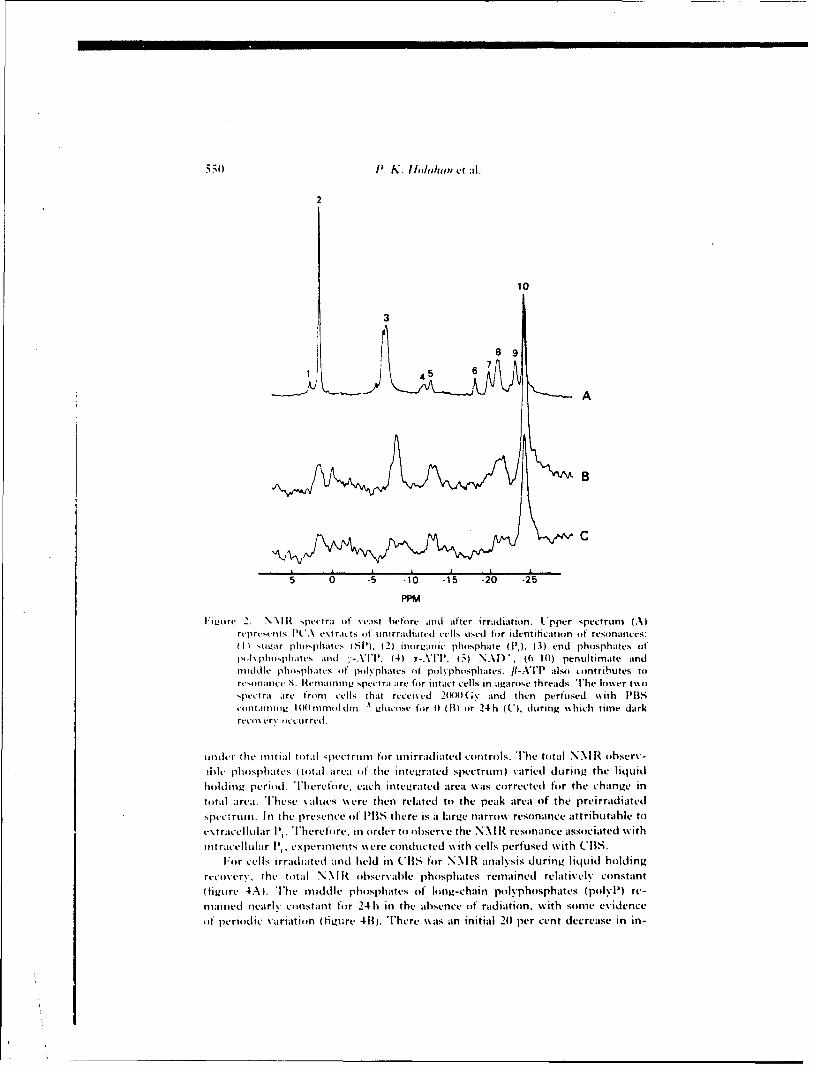
550 1'. K. l1oda/uip etal.
2
10
A
B
C
5 0 .5 -10 -15 .20 -25
PPM
F:iiort 2. \N \I spectra of %vast before and after irradiation. U.pper SPeCtrUm (A)rrt re-eos I CA ex~tracts of on i rrd imutd cells uised for identiticat ion of reso nances:(1 tii'ir phi .pha tcs ) S P1 (2) iorgan it phiosphate (1',). (3) end pho sph ates of
(i *tpiis~ltttandt 1-XT 4) Y-ATlP. (5;) NAI), (6 10) penuiltimate andmi ddlIe ph isphint s i f pi l vphattes of pioiiyphosphates. /1-A IP al so contlributes tore~ima nct- S. Retmainin g spectra are ti ittact cellIs in aga ruse threads. The b i'mer t%%oi,pt--t ra are- from tells that received 2(Wttt Gv- and then perfused with PBS5cmntatininu Itt) runiddin -1Ittciise fur 1) (11) iir 24 h (C), dutring, %hich time darkrvco i erv it or red.
kititur the initial total spectrum for mnirraidiated controls. The total NN H ohserv-ibie phosphates (total area of the integrated spectrumn) varied during the liquid
hiilding period. Therefiire. each integrated area was ciirrected for the change intiital area. Tlhese values wecre then related to the peak area of the preirradiatedspectrum. In the presence of PBS there is a large narrow% resonance attrihutale to
extracellular Ii. Therefore, in order to observe the N NI 11 resonance associated wkithtntracellular P~, experiments were cundueted with cells perfused with CBS.
Foir cells irradiated and held in CIBS for NM H analysis during liquid holdingrecoverv, the total N NI H ohservable phosphates remained relatively constant(tigure 4A). The middle phosphates otf long-chain polyphosphates (polyP) re-mained ntearly ciinstant for 24 h in the absence of radiation, with sonic evidenceof periodic variation (figure 413. TIhere \\as an initial 201 per cent decrease in in-
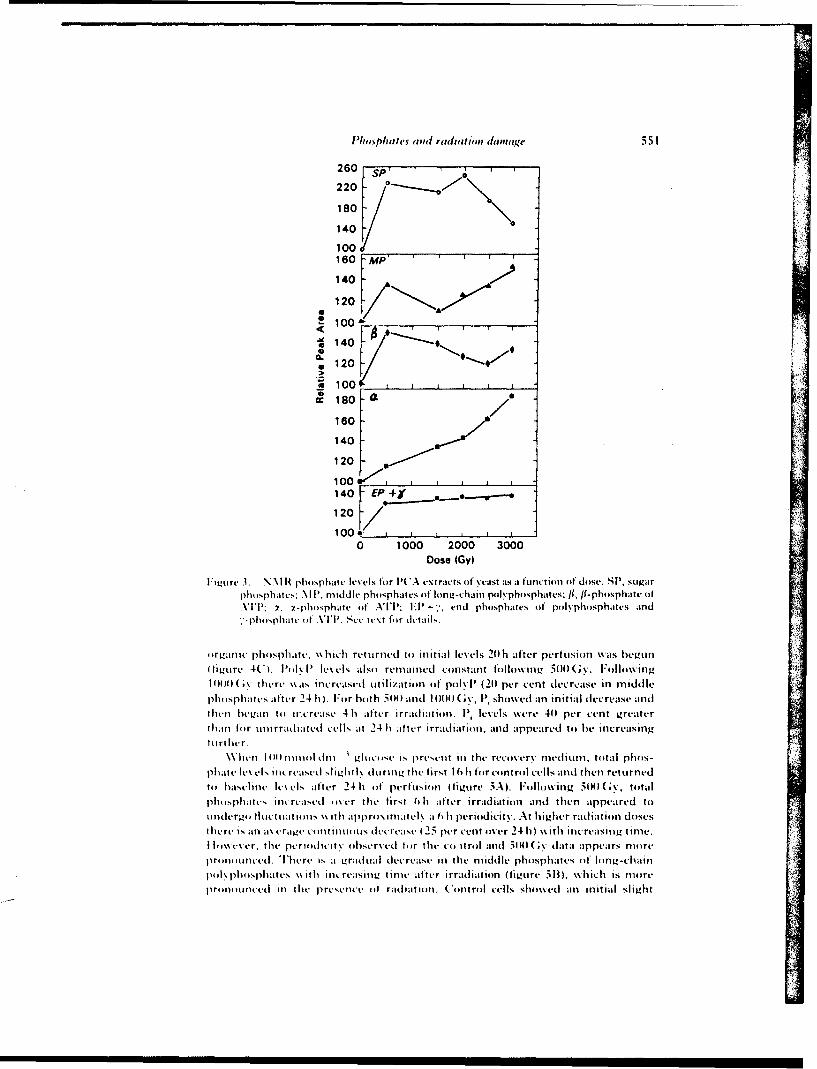
I'/iop/uitts (1,1(1 rtI(Ifiation damai~ge 551
260
220
180
140
100 _____________
160 Afp
1402
1202'
S100*
S140
00
cc180F 7160
140F
120
100U~t I140 E
120
1000 1000 2000 3000
Dose (GYl
l N1Ot . N I H phosphate levels for I'CA extracts of yeast its a function of dose. SP', sugarphosphatv : MlI '. middle phosphates of lung-chain puilvphosphattes; iII.-phosphite ofXII'; )!, x-phosphate of ATlP: l ~; end phosphates of ptulsphuusphates aInd
i ur.jimc pho sphaite, wh ich returnedc to initial lev-els 20 h after perfusion %%as begunItiLure 4(CI. l'ol\ l' e~cls also remaiined constant following 5(0 (i. Following
I 114(vthere \Nias increased utilization of pok I' (21) per cent decrease in middlephosphates alter 24 h . For hoth 541(1and 14000( sv, P'~showed an initial decrease andthten beg~an to increase 4hI after irradiat io n. Pj levels were 401 per cent greaterthan f(or tinirradiated cells at 24 hi after irraldiation, and appeared to he increasingfuriltr.
When 10III4I13 I1 ;nnt dni w~ .ltc N pruwn in the reco~ery medium, total phos-
phatele~ e.. icreasd sliht l during the first 16lt for conitrol cells and itheni returnedt'' baselinie Ic\ LIs after 24 h oft perfuasiotn (figure ;A). Foilltowing tota(v ~~lphlt sph at s ini rca sd t cer lhe firs t OhI after i rrad it ion and then appeared tounldergo) 1IuCtuitins \% ith aIpprtts.iniatvl\ a 6 h periodicity. At higher radiatiomn doseshert is an average tttintotus decrease I 2. pe-rcent uner 2410l \%itli increasing time.
I I imvecr, the periitdicit t bserved (or the to-itrtol and 541( (* data appears moireptmou intl . T here I.- aI grad nal ducrease InI the middle phosphaes ouf long-chain
1\ptl pliisate.s~ wtli inicreasitiLg time after irradiation (tigtire ilt), x~hich is nmtrep r tnctd i1III thte P rest- oetf radli at iton. Cotnt rtol cells slit ued ain in it ialI slight
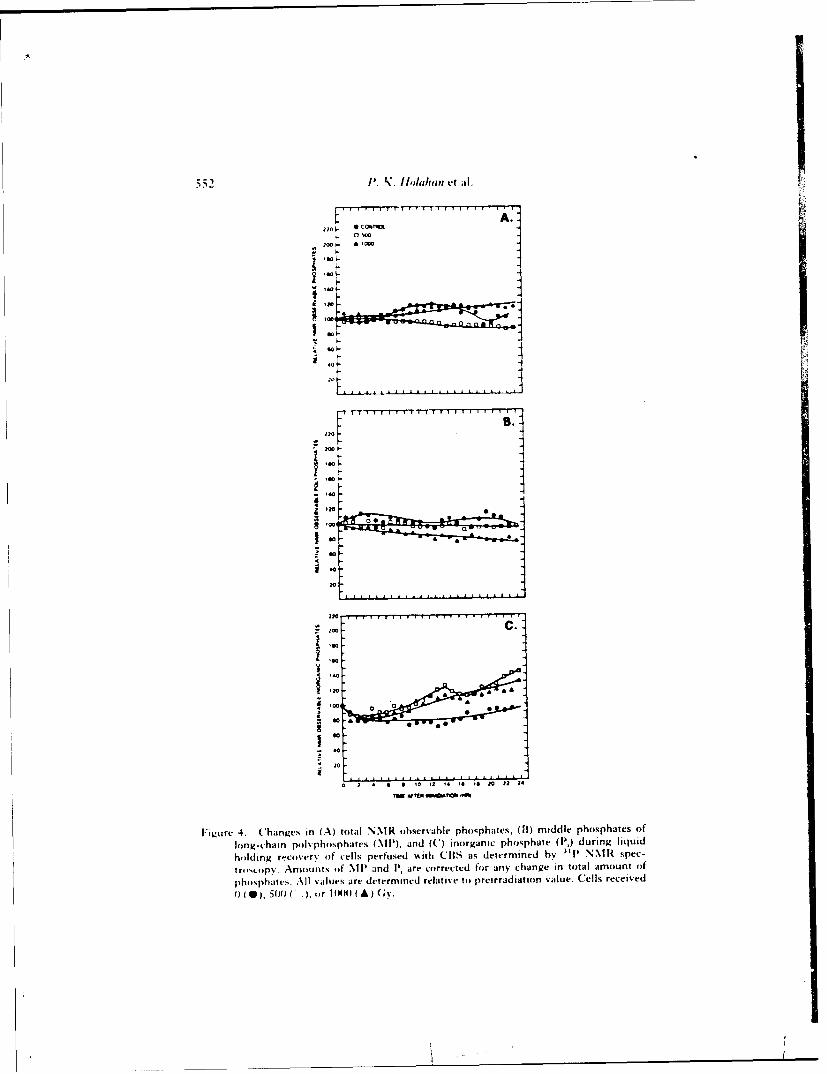
552 I'..?'. IbI,jhi, V't ;Il.
so
:0
' '
1 1
'40
Go.-01
2 '0
r.. .
220
0 A
w s
w.0
o 2 a a ' 0 'a 'a 0N 22 24
|Figure" 4. Changes in (A) total' NNIR observable phosphates, (11) middle phosphates of
long-chain polyphosphates (MP), and (C) inorganic phosphate ([',) during liquid
holding r.covery of cells perfused with CBS as determined by 3| P NMR spec-
troscopy. Amounts of MIP and I, are corrected for any change in total amount of
phosphates. All values are determined relative t) preirradiation value. Cells received
( (0),5(11 C 4, 5 or I(O (A) (;y.
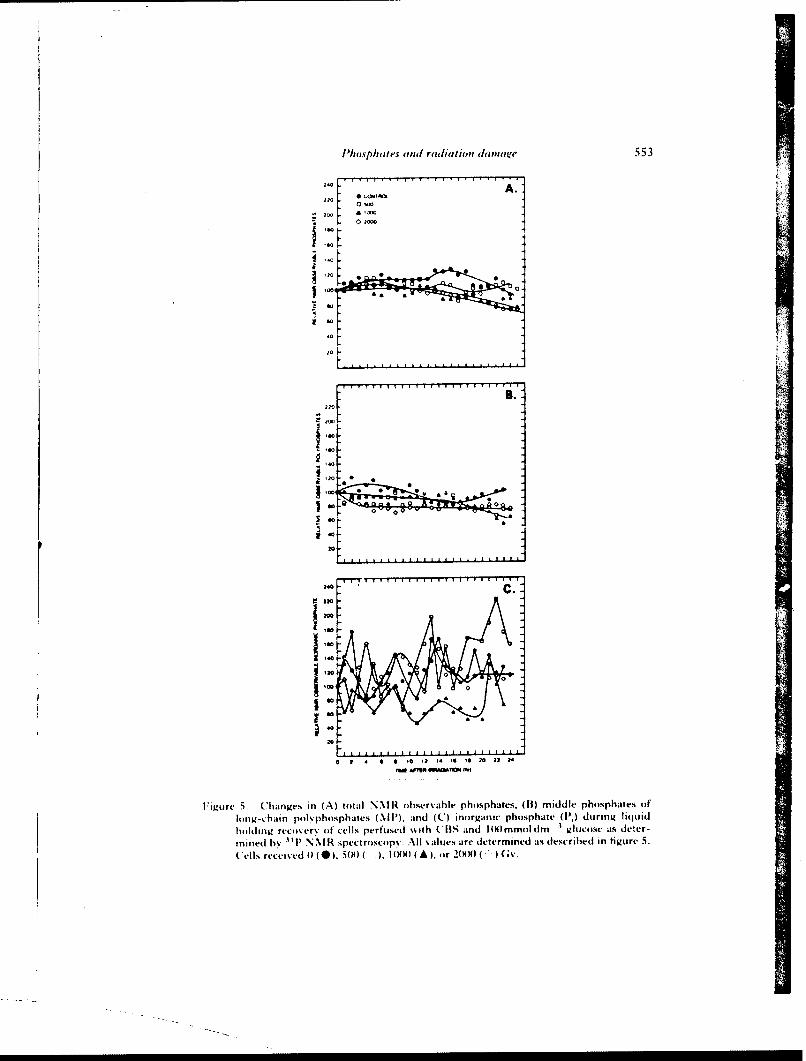
l'Jwsp/awes arn ralitiun dammiise 553
a
2. A
IN
2wFT 0 T
Ii~re .han~v in(A)to~iINM obeah hshf.(I)mdlphpht OIiinj.-chainp h-phshic (M',m C nr-ncposht I, uglqi
mintd boij)N
CmllsdreyeiTed R spectro~scopy. All %a I tes are determined astdescribed in tigture 5.Celsrecvv 4 0), 504) INK)lHH (A) fir 2(XHJ ( Gv'.
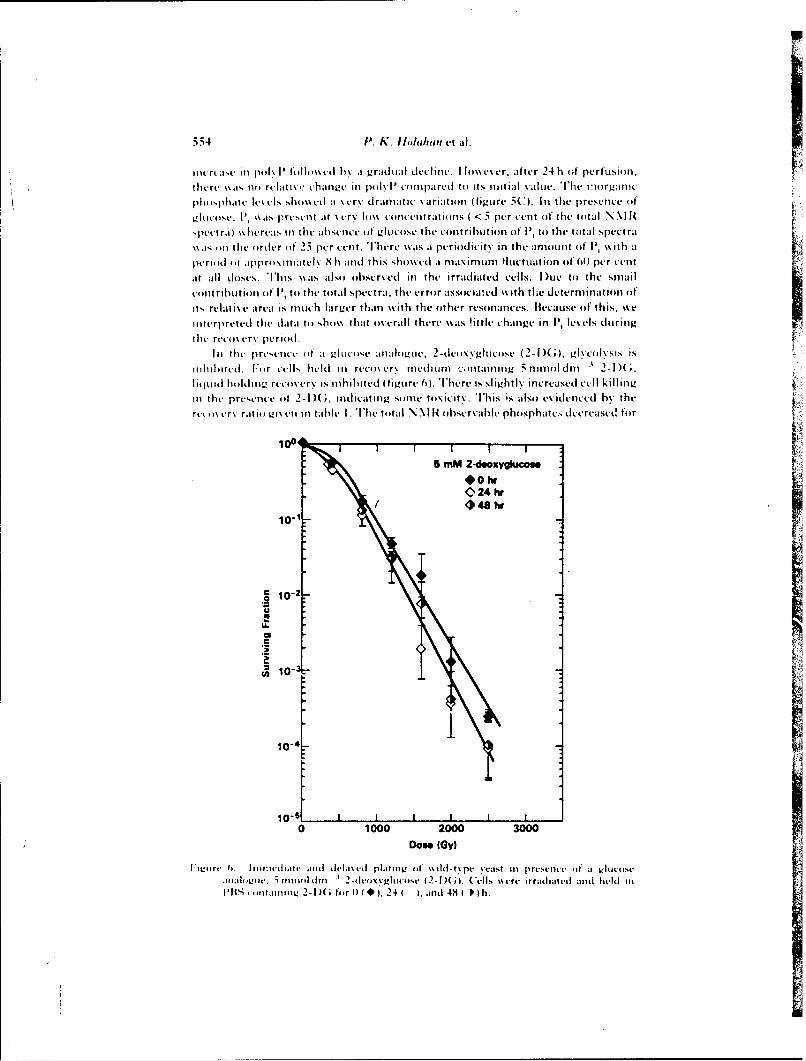
5.54 1. K. I1oh,/umn ut A.
int rvasc in pi )k V f Ii lI o%% d b\ a gra dta I detclIit i. I lo%\c\ cr, aftcr 24 h (it perfuLsion.
theure %\as. no rulanx , changeu in polklV comopared to its initial \ altic. The in~organic
phiosphatic uIs slhO\\ t % ~cry dramnatic \ariation (tigure 5C). In the presence of
iiik ose,. k its present at \ cr\ hmi~ concent ratnins ( < .5 per vent of the total N N\ ItR,puct raj \\ lurcas in the absence of gluco se the contribution of P, to the toital spectra
\\a, mi t he ordeur (it' 25 pur cent. There wais a periodicity in the amnount of P) w\ith at
period ol" apprommnatc\ S h and this sho\% d at nlaxinlum Hticttion oft 00 per cuntat all closes. TPhis \\ias also iibscr~ed in the irradiated culls. I uc to the smal
tont rihutiou ut, P to the total spectra. the crror assoviwed %%ith thu determination of
its rviati~v arva is much lartur than\with the oither resonanuces. llucausu of this, %%e
inoterpreted the data to ,hi '\ thast u ierall there \%ias !ittlv change~' in 1P levels dturing~the rt-mi~cr\ puriod.
In the presence (oft aglu1 o se anloueov 2-dlvio\\gkuuosu (2-I ) ;), gi'Cuivsis is
nibited. F.or tells huld in ruco~ur niudiUni '.ntaining' 5 ninildm 2-I )(
I iqoidbld Ii~rt-uo\ rv is inhibited (ititre 0). There is sI ithtiv incrucasud Cull killingin the presence of 2-I )(, indicating~ sonic toxicity. This is also evidenced by the
ruci i~ r\ rat ioii ic' u in table I . The total NN\I R observable phosphat. 4 dcruasud for
100 5 mM 2-deoxygiucos
C 24 hr/ *48 tw
10-1
.2 10-2-
u~10-5
0 1000 2000 3000Do** (Gy)
I:Iiitrc 6i. liiiu:it'd div anidda~d plat itn of it 1 ilt .pc vorast in p resence (if a g I um sea a Iil't., 511in no I im dirIiiiin - ( ) ('ei Is \%re irradiatd and t-it]u in
PitS i itaiiinv 2-l)( hir ( (* 24 t ). anti 48 1 Il th.

Phosphates and radiation damage 555
control cells but increased follow\ing 100)( v (figure 7A). After 200)(iy thereappear to he fluctuations on the order of 20 per cent with an 8 h periodicitv.I loo\ever, in another control experiment the total phosphates showed an initialincrease and then declined. In all experiments with 10(XI(;y, total phosphateseither remained constant or increased. NN1R analysis shows a decrease in themidd'e phosphates of long-chain polyphosphates (polyP) even in the absence ofradiation (figure 711). This decrease did not correlate with increasing radiation dose.The d..ta sho\% n for 14)))(v\ are representative of several experiments, although insome experiments the decrease in polyp was more pronounced. Although in thepresence of glucose there was no overall increase in inorganic phosphates (P i) as afunction of time, the presence of 2-deoxy'glucose in the recovery medium resulted
in a concomitant increase in Pi with decreasing polyP (figure 7C). The mostdramatic change in both polvP and li was seen following 2000 (v. Although notshow\n here, the N.IR spectrum displays a very prominent peak associated with 2-deo)x.\glucose-b)-phospliate. It accounts for approximately 25 per cent (determinedb1 ratio of areas) of all .ibservable phosphates by the end of the experiment.
"urthermore, in the presence f glucose it is possible to observe ATP reson-ances that are beneath the level of detection by N .l R in the absence of glucose. Therejativc amount of AP'I'i was determined from the area of the 3 [1 resonance signalf/i-.l'lI relative to the total amount of NMR observable phosphates (figure 8).
There are difficulties in assigning the area under resonance No. 8 (see figure 2) tothe A''P3 content of the cell since IlT also contributes to this peak. Judging by PCAextracts /i-All' contributes (to resonance No. 8) approximately 30 35 per cent;home\er, direct comparison to ii ivo data is difficult since extractability of ATP
and pol. phosphates could differ. In our discussion we assume that any variation inresonance No. 8 area is due to changes in ATP. Cells held in CBS containing1WOsmol dm 3 glucose \vith no radiation showed almost a three-fold increase inA''P le\cis, w%-hich plateaued after 8 h of liquid holding. Following irradiation there\\as less of an increase in AT' levels so that, with 2000v, fi-ATP levels remainedconstant. In cells exposed to 54)0(iy there was an initial decrease in the first 4hfollowved by an increase, which plateaued after 14h. With 10)(;%, ATP levelsin.creased o\ er the first 12 h and then decreased back to preirradiation levels. Cellsthat recei\cd 24)00(;v showed no change in AP'I' levels during 24h of liquidholding after an initial decrease in the first 2 h following irradiation.
4. DiscussionT'he present investigation \%as undertaken to examine the relationship between
hith-energy phosphates and the recovery from potentially lethal radiation damagein \east. Liquid holding recover\ in yeast has been shown to be altered by a varietyof aLents that moidify the enetrgy supply, such as 2-deoxyglucose, potassiumcyanide, and 2,4-dinitrophenol (latrick and Ilaynes 1964, Jam r' al. 1977).F~urthermore, if energy production is inhibited prior to irradiation, there is areduced capacity for cell survival (Seymour et al. 198;). In this study the additionof ,glucose to the extracellular medium resulted in greater recovery, as demonstrated
by a larizer recovery ratio (see table I ). In contrast, there wvas no recovery observedin the presence of 2-dcoixvglucose, and there was a reduction of the shoulder of thesurvival curvc (),). l'his decrease in 1). has been attributed to a de,-rease in theintracellular AlI'P concentration (Reinhard and i'ohlit 1976).
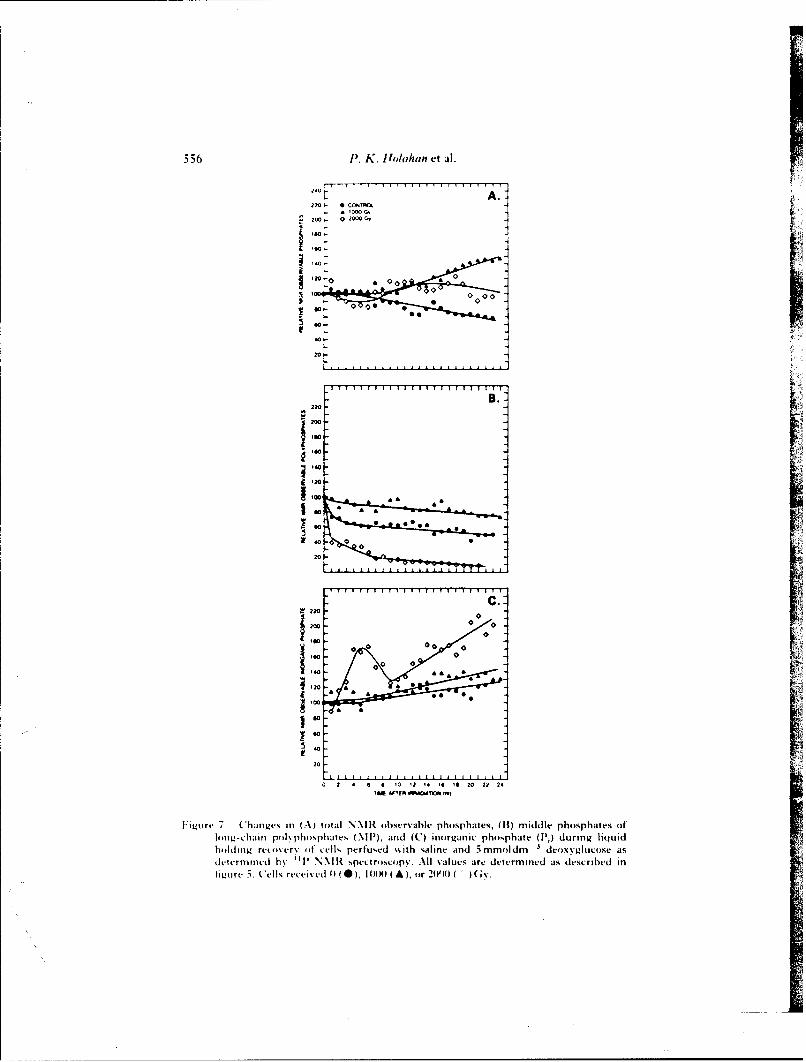
556 11. K. Houlahan et al.
210
220 - 0C iT00 *COG,
200 - ,O O C
101
'.0B.
220
so 0 0
0
00
20
Iw
so A000*~~0
l:Lw.7.(hn~'sin(A otlN\R beralephshae, B)mddephspaeso
Ioo~~~~-chiiiri~~~~ Io~bspae I~) In ( I Iorai ph.ht I'~ Iuin liquIidhi.l i ~re ie vutculspefuedwthsain ad m odm ' eoy~ucs.a
deeriedb T N secrscp. l vlesae etrindasdsciedi
2-S
ti~ore 5. Cels received0 0) 0X A.o 0O(I(~
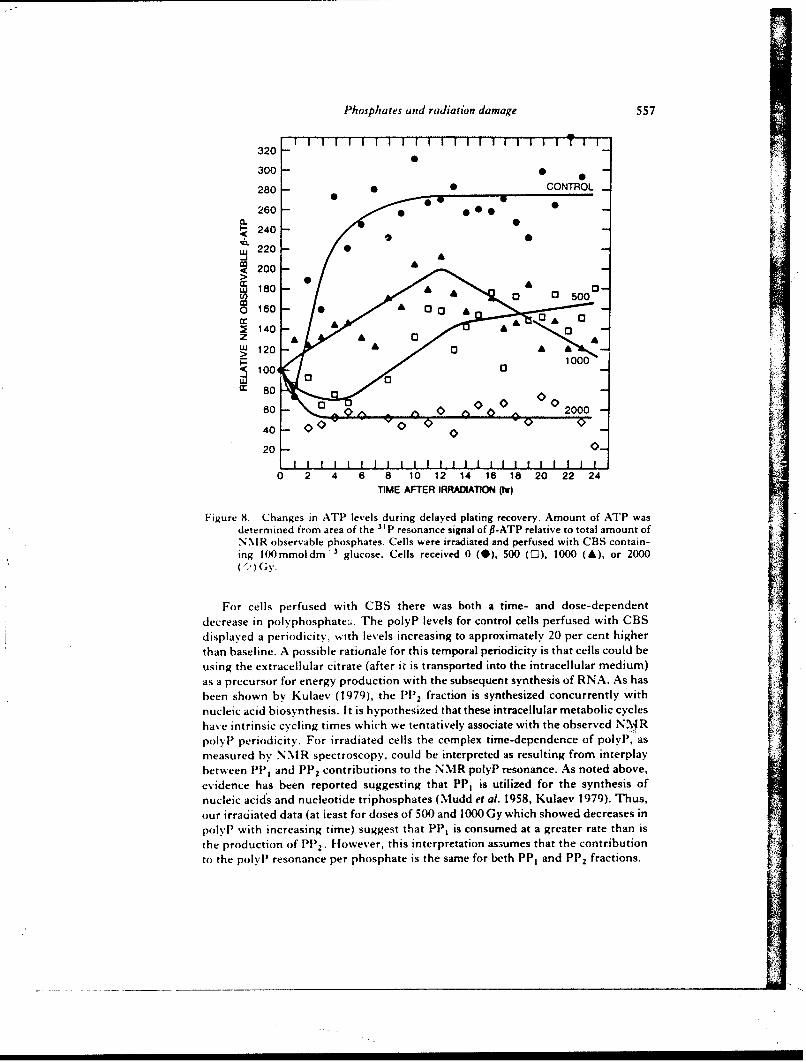
Phosphates and radiation damage 557
320
300280 -• CONTROL_
260- -a. -240 -
220-
200 -
W 180 2 4 6 5000 160 -0
z2 140
TIM AFE RAIAIN(
wu 120 aL>00
ing m d g 1000 (X 80
600
600 0 -
0 2000v
20F s-
0 2 4 6 8 10 12 14 16 18 20 22 24TIME AFTER IRRADATION (11")
Figure 8. Changes in ATFP levels during delayed plating recovery. Amount of ATP wasdetermined from area of the " P resonance signal of Pl-ATP relative to total amount ofN.N R observable phosphates. Cells were irradiated and perfused with CBS contain-ing 100~mmoldm - 3 glucose. Cells received 0 (0), 500 (0-), 1000 (A), or 2000
For cells perfused with CBS there was both a time- and dose-dependent
decrease in polyphosphate:;. The polyP levels for control cells perfused with CBSdisplayed a periodicity, with levels increasing to approximately 20 per cent higherthan baseline. A possible rationale for this temporal periodicity is that cells could beusing the extracellular citrate (after it is transported into the intracellular medium)as a precursor for energy production with the subsequent synthesis of RNA. As hasbeen shown by Kulaev (1979), the Pl) 2 fraction is synthesized concurrently withnucleic acid biosynthesis. It is hypothesized that these intracellular metabolic cycleshave intrinsic cycling times which we tentatively associate with the observed N\'JRpolylp periodicity. For irradiated cells the complex time-dependence of polyP, asmeasured by NMR spectroscopy, could be interpreted as resulting from interplaybetween PP, and PP 2 contributions to the NMR polyP resonance. As noted above,evidence has been reported suggesting that PP, is utilized for the synthesis ofnucleic acids and nucleotide triphosphates (Mudd et al. 1958, Kulaev 1979). Thus,our irradiated data (at least for doses of 500 and 1000 Gy which showed decreases inpolyp' with increasing time) suggest that PP1 is consumed at a greater rate than isthe production of PP2. However, this interpretation as3umes that the contributionto the polyp resonance per phosphate is the same for beth PP, and PP 2 fractions.

5 58 P". K. lahan et al.
Vhen gluc(ise was added to the reco\ ery medium there %%as again both a time-and dose-dependent decrease in the middle phosphates of the long-chain polv-phosphates. The absence of a concurrent increase in Ili suggests the followingexplanations for this decrease: (a) polyl) are being used for the direct phosphory-lation of glucose, (b) polyl are being used as a source of phosphates in the repair ofI)NA damage, or (c) polyP are being used in the synthesis of RNAs which nav code
for synthesis of repair enzymes. Evidence that the direct phosphorylation of glucoseis not the sole explanation conies from our data on phosphate metabolism in thepresence of 2-deoxyglucose a glucose analogue that results in the inhibition ofglycolysis. In the N.IR analysis of cells irradiated and held in rtvovery mediumcontaining 2-D(, there was a decrease in the middle phosphates of the long-chainpolyphosphates. I lowever, unlike cells that were held in glucose-containing recov-ery medium, there was a corresponding increase in li (figures 5C and 7C). Thisincrease in ), \was also greater than that observed with CBS alone and, in fact, didnot show the initial 20 per cent decrease. If polyp \ere being used solely for glucosephosphorylation, they would be used in the presence of both glucose and2-deoxyglucose to form glucose-6-phosphate and 2-deoxygl ucose-6-phosphate,respectively. In some lower species, polyphosphates can directly phosphorylateglucose with the enzyme polvphosphate glucokinase, but this enzyme has not beenidentified in yeast (Kulaev 1979). However, it does appear that polyphosphate isused to phosphorvlate glucose, although it may be an indirect reaction as chemicalenergy of polyP bonds do not appear to be directly available to the cells (vanStevenick 1968). PolviP may indirectly phosphorvlate AI)P or a carrier or enzymethat can mediate the transport of glucose into the cell (van Stevenick 1968). Theinorganic phosphate would therefore be expected to show similar changes in bothglucose and 2-deoxyglucose, a conclusion not supported by the data reported intigures 5 and 7. Therefore, polyP must be utilized as a phosphate source for morethai, glucose. Our experimental evidence supporting explanations (b) and (c) as alsobeing viable possibilities is reflected in the changes in P,. The increase in Ili duringrecovery in the presence of 2-deoxyglucose suggests that polyiP are also being usedas a phosphate source for )NA repair and or RNA synthesis. Polvphosphatesdecrease during )NA synthesis in the absence of extracellular Pi, suggesting thatpolyP are being used as a substitute phosphate source (Gillies et al. 1981). mRNAsynthesis has been shown to increase significantlv following irradiation, althoughthere \\as inhibition of protein synthesis (Skog et al. 1985). Therefore, althoughthere may be no increase in total repair enzymes, there are still increased nucleicacid synthesis and repair, which require phosphate. This interaction betweenpolyphosphats and other intracellular macromolecules is illustrated in figure 9.This is a modification of a hypothetical scheme outlined by Kulaev (1979) for highmolecular w\eight polyphiosphates in fungi . In this scheme we note the interdepen-lenCTies exhibited by polyphosphates. nucleotide triphosphates, nucleic acid bio-
synthesis, and ATIl.The ob-,rvation that the inhibition of recovery is greater in respiratory-
deticient mutants than wild-type yeast (Jain et al. 1982) suggests that energy is stillsupplied by thc respiratory pathw\ay. Furthermore, it has been shown that when
2-deoxyglucose is present in the recovery medium, both I)NA repair and recoveryfrom potentially lethal radiation damage are reduced (Jain et a/. 1977, Verma et al.I ()S2). From these investigations the authors concluded that )NA repair dependson .-TI . ()ur data indicated that, in the presence of glucose, A'rTP concentrations,ire increased, \\hich is supported by the work ofl Reinhard and l'ohlit (1976). A'Tl
/
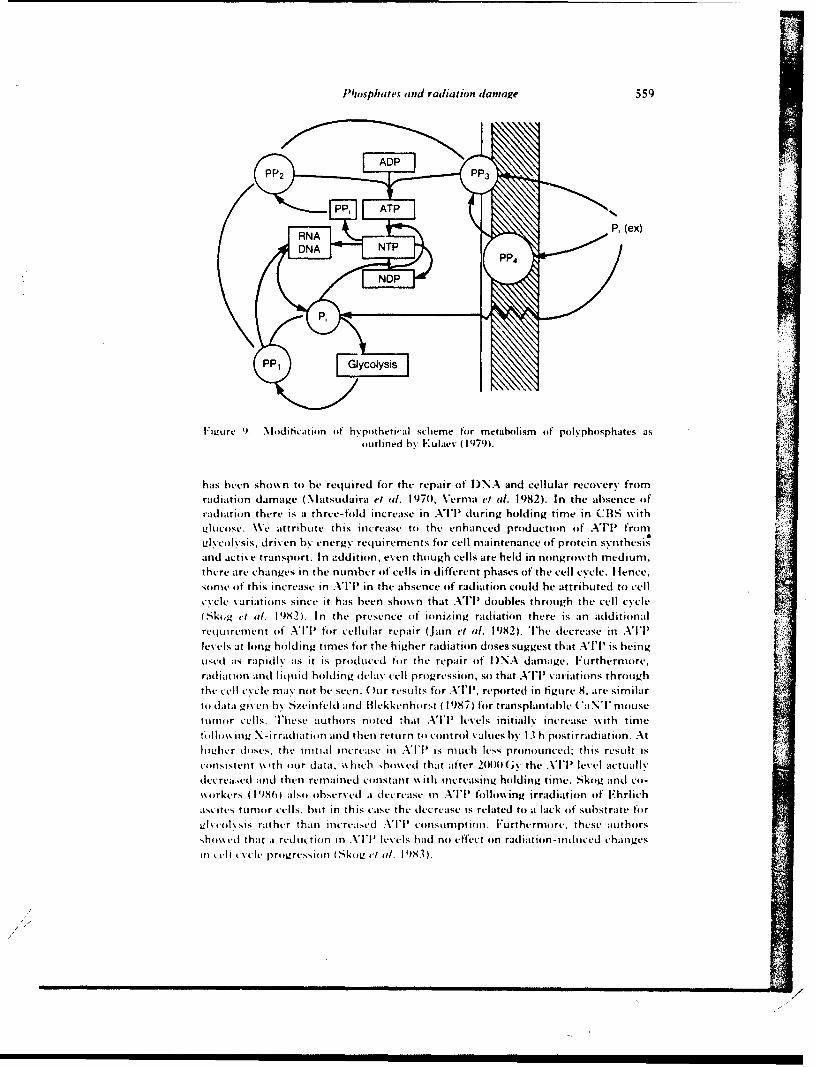
Phaosphates and radiation damage 559
PP4,
lIL'Urc 1). Mdiication of hypotheti'aI scheme tor metaboismn of polyphosphates asoutlined by PKulaev (1979).
has been shown to be required for the repair of I)NA and ceilular recovery tromradiation damage (\Iatsudlaira el al. 1970, Verma~ ei al. 1982). In the absence ofradiation there is a three-told increase in ATlP during holding time in CBS withLtlucosc. We attribute this increase to the enhanced production of ATP' from
Llycolysis, driven by energy requirements for cell maintenance o~f protein svyithcsisand active transport. In addition, even though cells are held in nongrowth medium,there are changes in the number of cells in different phases of the cell cycle. I fence.sonic of this increase in ATP'l in the absence oif radiation could he attributed to cellcy, cle variations since it has been shown that ATP doubles through the cell cycleSk,g el al. 1982. In the presence of ionizing radiation there is an additional
recluirement of ATP'l for cellular repair (Ja i etl. 1982). Tlhe (lecrease in Alllevelis at long holding times for the higher radiation doses suggest that ATP1 is beinguISed as-. rapidly' as it is produced tor the repair olf I) NA damage. Furthermore,radiation and liqjuid holding dela\ cell progressioln, s4) that ATP'l variations throughthe cell cy,%cle may not be seen. O ur results for ATP', reported in figure 8, are similartoi data -(iveun b\ Szeinfeld and Blekkenhiorst ( 1987) for transplantable (aN NmousehtlE Er cells. These authors noted that 'I' lvels initially increase with time
I IE EEA% ing N - irradiati, in and then return to control values by- 13 h postirradiation. Athighier doses. thty Initial increase iii NI is nmuch less pronoEunced: this result isconsistent w~ th our data, -,%hich .,hos ed that after 2()11011(;\ the ATP' level actually-di. rei... d and then remained co nstant %%ith increasing holding time. Sk-og and col-
E rkers (I19806 a lsoE bserved a decrease in AITP following irradiation of Ehrlich
Estites tunmor cells, hut in this case the d-crease is related to ak lack Elf Substrate for
&,l\co EIsis rat hur than increiaer ATP] colnstmptiion. Furthermo re, these authorsshiEmcd~ that a red uction in ATPl lvl had no effe'ct on radiation -induced changes
In eL c cl e pro gression (0ISkog v tit 1 983).
C.

560 1.* K. Ilolahan et all.
In addition, polyP may also be used as an energy source if the ATPI con-centration is low. P~olyphosphate can be used to phosphory late AlDPl to form ATP'l\-ia a reversible reaction catalyzed by polyphosphate kinase (Kulaiev 1979). \%hell notvilucose \\as present ats anl energy Source, 1) increased] as pots.] decreased (figure 4),suego.stinrg that polvphospbates wecre being used] as at supplemental energy sonurce inladdition toit aSoUrce of phosphates. In the presence of an external soiurce of i'nergv
.e. glulcose), the cell was able to synthesize and use ATP'l; hence polvT it s Usedprirnmirliv as at phosphate source. Since ATP'l was depleted in the presence of2.deoxvglucose there is aI possibility that polyP may. be used to provide someenergy. I Iowever, the cell may, not be able to obtain sufficient energy solely fromtpol\ phosphate degradation to allow for repair. It appears that yeasts areable to hydroly.ze polyP in the presence of 2-deox'.glucose but are unable to utilizeeither the free phosphates for phosphorylation or the energy release for damagerepair.
Inl concIlusion, weC note thalt there are limitations to cellular NN\I 11 spect roscopythat should be considered in the discussion of measured phosphate levels. NN\lspectroiscopy is unable to determine absolute concen trat ions of a particular phos-phate because there could be changes in the balance between immobile and mobilepopulations of phosphates. If phosphates become bound follow.ing radiation, thisW.ould also givec the appearance of at decrease in phosphate levels. I lowever, becauseinorganic phosphate increases with the decrease in polyphosphates, it appears thatpolvP are being hvdrolyzed rather than becoming bound. Furthermore, the-technique~ does not distinguish possible differences between irradiated cells com-mitted to full recovery (vitable cells) and those destined for reproductive cell death.Tlhus it remains to be determined if recovery of polyphosphates occurs in viablecellIs.
Acknowledge mentsTlhe authors appreciate the technical assistance of Ms Andrea Lunsford and
Ms C'olleen Loss in performing the numerous cell survival experiments, andalcknowledge the help of D~rs Chiwan Chen and Jack Cohen (NIH-) for inform-ing and teaching LIS the technique for viable cell preparation for NM R experiments.We also thank D~r Eric I lolwitt for many helpful discussions.
Referenceslox si.i 1). I .. , ad (OlIEN, j. S., I 983, NM H studies of perfused cells. Jouirnaliof liagnetir
Uvwimanc-, 52, 346 349).l' vX.\i.i.. 1). L ., (ohiiN. J . S.. arid Nhiiwoi., J. BI., 1 984, Continuous perfusion of
miammnalian cells emhedded in agaroose threads. Evperimental Cell Research, 154,521 .529.
Is \NKI-N1iiiR;-SIiWsi;FH. Ml., FR.'.'NIxiiiRs;, D)., 131.0(11 14 D)., and AiSAM~CZYK, C., 1 980,Hiepair of I N.A double-strand breaks in irradiated yeast cells under nongrowthconditions, Radiat ion Research, 82, 498 5 10.
(ir W. .J., Ufoiimo., K., i*N lloi iANiWH,j I., and S~IIIF'.,N, R. Gi., 1981 , 'T NNMHstudies of intracellular pl and phosphate mietahoismr during cell division cycle ot.Saccha r(m vies cerei -isiae. Proiceedings of the Na tonal .lcademY of Sciences, I .S. 1l., 78,2125 2121).
I u. .J., 1979, Radiphinlogv ifor, the Raedloist (New York: Ilarper & Row), p. 34.I I.si)i, F~.I .966, Inorganic pielvphosphates in biology: structure, metabolism, and
fuLnction. Bacterioloi,',al lRe'iei, 30, 772 794.JA(IO s N, L.., and (iOi N, J . S., 981I, Improved technique for investigation oif cell
nietab'idism by P N M' IR spectroschipy. flioscience Reports. 1, 141 1Sf0.

Phosphates an'd radiat ion dantage 561
i t\, V. K.. and Potol rr, W.i , 1973, 1Influence of energy netabolism oIn the repair of X-raydamnave In living cells. 11I Split-dose recovery. liquid-holding reactivation and
di~istundla~\ re.esli ttoaypp lioso es.liposik. 9, 1.55 1 65.
\I\,. V K.. I lot tI/. G. W., l'ottt I , WN., and Pomi.rr. S. C. 1977, Inhibition of unscheduled
I )NA ti tht-sts and repair ot pottially lethatl X-ray damage liv 2-deioxv-ii-glucose in
Cast l,, I tyatinnl J ounal ol Rad~liatio B0 iologv., 32, 1 75 ISOI
\\.V. K.. (t vi \, I., and Li \,s K.. 1 982. Energetics of cellular repair processes to a
rcspiratror\ -dfctient nmutaion of \.cast. Radiation Research,. 92, 403 473.Ki I \I1. S.. 1979, The Iioehe,,,istrv of Inorganic Pol)vphosphates (New~ York: John Wile\)
I \ I us I)k N I I , Ft to '\o, I., and 0 )t si K,%, If., 19701, Possible requirement of adenosinetriphosphate 10r the rejoining of X-ray-induced breaks in the D)NA of Ehrlich ascites-
tumnour cells. International Journal of Radiation Biology-. 17, 339- 347..N It III S., Yoiii \, A.. and Kow:, NI., 1958, Pol yphosphate as accumulator of phosphorus
and energy . Journal of Hactertology. 75, 224-235i.Gut.,. ; x ~ S., Siiw, R. G., and YsANINE T., 1977, 51P Nuclear magnetic
resonance- stutdies of Ehrlich asciteS tumnor cells. Proceedings of the National AcademYof Scienc es, I *.S.A., 74, 87 9)1.
(INo', G.L, Slit t.M N, W. G., Y \M\% Tl.,.u l.it~tsH~si.i., TI. R., I..xi, K.-I., BA.5ONOiFSKY,
JJand N I %H~ii R, J ., 1979, Phosphorus-31 tI Iiclear magnetic resonance studies of
oIld-t\ pe and g\ vcodvt ic path\% at\ mutants (if Vai-charomwces cerevsiae. Biochemist,.18, 4487 4499.
NI (ot i), K., SctiFtmg, WN.l1HIt'INE:uoo, 1. Mand KAPTEi,. R., 1983, In vivo "T
NM 1l studies on the role of the vacuole in phosphate metabolism in yeasts. Archives of.1icrobiology. 134, 270- 275.
Pi'm]HtK. M. IL., and HAsYNES. R. If., 1904, D~ark recoverv phenomena in \east. 11.Conditions that modify the reco vecry process. Radiation Research, 23, 564- 579.
P\ I IK, NI. I I., I I \NNES, R. It., and Lisivr '., R. It., 1964, Dark recovery phenorn.. *n yeast.1. Comparative effects %%ith various inactivating agents. Radiation Research, 21,144 163.
Ri. I~t mtRi, R. D)., and Po"i.lT, \%*.. 1976, Influence of intracellular adenosine- tri phosphateconcentration on survival of yeast cells following X-irradiation. Radiation andCe'llular Control Processes, edited by J. Kiefer. (Berlin: Springer- Verlag), pp. 117-1I23.
Sit IINY, J. M\., YAMANI-, TI., SHUL;MAN, R. ';., and ()GAWAs. h., 1975, High resolution 31t Pnuclear magnetic resonance studies of intact yeast cells. Proceedings of the National,lcade~,v of Sciences, U.S.A., 72, 4966-49701.
S,-Ny\,ol R, C'. If., \IitmRissii.i., C., and MIRTi~wY, M1. J., 1985, Glucose analogues alter theresponse of CIIO-KI cells to gamma irradiation. Acta Radiologica, Oncology-, 24,351 -356.
5Kmi;, S., TIimiiK.Ai I, If.. and Si \ius, G., 1982, Energy metabolism and ATIP turnover timeduring the cell cycle of Ehrlich ascites tumour cells. bFeperimtental C'ell Researchi, 141,23 29).
Stun;, S.,TRB 'iiKAsI I, It., and St NiM s, G., 19)83, Energy metaholism and ATP turnover timedluring the cell cycle in Roentgen irradiated Ehrlich ascites tumour cells. ActaRadiologica. Oncology, 22, 369 379).
Stoit;, S., 'l'iitiK~irI, If., and Nyi;..A.su, ()., 19)85, [INA and protein synthesis of irradiatedE'hrlich ascites tumnour cells. 11. Amount of m-RNA and in vitro, investigations. ActaRadiologica, Oncologv, 24, 549 553.
St ; S., Nosiiii.i., If , Esof(sso-N, A., IsjitiKArr, If., and NISHIDA, TU., 1986, Changes inenergy metabolism follo%%ing Roentgen irradiation in in tito griming Ehrlich ascitestumour cells studie %. by'P magnetic resonance spectroscopy. Ar'ta Radiologica.0?, .obogv. 25, 03 61)
SziiiNiF., D)., and LfiEKKENHOHST, G., 19987, Effect of X irradiation on adenosine triphos-phate and glucose-6-phosphate dehydrogenase in the CaNTr mouse tumnor. RadiationResearch. 110, 305 309.
%1. SIFAEtNINiiK. J., 1968, Tlransport and iransport -associated phosphory'larion of 2-deoxy-ii-glucitse in yeast. Biochimica et BiophYsica Acta, 163, 396- 394.
V1 RNIA, A., SICAsRMA, R.. and JAsIN, V. K., 1 9M2, Energetics of D)NA repair in LV-irradiated
peripheral blood leukocytes from chronic mveloid leukaemia patients. PhntochemistrYand PMiotobnoogv, 36, 627 632. r

562 Jo/wsp/uihx tand radiation damnage
\V iitI I (f It irg ranic p v nwh sph it t ant po I\ phospha1tes as so w rces (if energy.
I ,,A P) ;' N\Itphllsphatc. IL -IHeat oft hydlrol\ysiso 4 rntatphosphatc vstraictcd from'v.ld t s .)oo i-toild ,,i ohemifr v t Tkvtri42, 103 108.

Circulatory Sho~ck 25: 123-130) (1988)
ARNO POPICS KAOIOSOLOGV0N1111AiCH .61T1TU111
SR88-23
Alterations in BidirectionalTransm~ambrane Calcium Flux OccurWithout Changes in Protein Kinase CLevels in Rat Aorta During SepsisR.7- Linten, J.A. Carcillo, and B.L RothPhysiology Department, Armed Forces Radiobology Research Institute (R.ZL.), andSurgical Research Division, Naval M' dical Research Institute (J.AC., B.L.R.), Bethesda,Maryland, Department of Anesthesiology, Critical Care Medicine. Child Health andDevelopment, Children's Hospital Medical Center, George Washington University School ofMedicine and Health Sciences, Washington, D.C. (J. A. C.)
A depression in aonic contractility has been previously denikinstrated in rat incraperitoncalsepsis and during endiitoxeemia. In th!. study. we determined whether the inbili/aion(it extracellular Lalcium1 iuwing "Ca) and the release of intracellular calcium are alteredin septic rat aorta when compared it) %hati.-orarated controls. he concentration 01 proteinkinasce C was alsodeeminned hy using I III phorbol -I2.3-dibuvrawe(PD)But. We foundthat calcium inliv was unaltered uider basal conditions bui that the ability of norepi-nephnne i NE) to augment influx %%as sigrificantly, depressed t1' - .5:I1control vs. septic,572 -54 JSF vs. 428 - 301 ILioil ('a kg dry wt. aortal). Calcium influx stimulatedhy high K was unchanged in jortue between control and sep.ic animal'.. In the presenceot NI:. calcium11 eltIL11 (an indirect measurement of intracellular calcium release) was%igniticantly diminished iPi' - (El ) iti aortae amon septic rats. The comncentration of aorticprotein kinase C as assessed by PI)FBu binding 't;es was unaltered in septic rat% whencompared with controls. In conclusion, we found that during sepsis alpha,-adrenergicreceptor activatioii of) both caicium int! , and efflux hy NE is decreased: these alteatiOnScould be related *o the depressed aormc c-nractility i'b %.Ved in sepsis. o
Ktv 'x.rds: calcium influx, calcium effux, nomnepint'phrine
Suhmaittcd for publication F-ebruary 21). 1"X
Thewe findings were presci-ted at the Firsi International Shock (ancs. l997 and published in preliminaryformi 1291,
T'he opinions and asscrions contained herein .re the private mines mi the authors and should not be construedas reflecting the vieWS mm) the UT.S. Navy, the naval sersice at large, the D~epartment -4i lDdense. or theIkctense Nuclear Ag~ency
D~r. Roth', present address is R4mamni 5 -25 3. Stanflord ( Ini vcrsitv Medical C'enter. Stanford. ('A94.104.
Addre',s re 'rti '']JuCS1s i. r. .Z. itten. P'hysiology' I )partmcnmn. Armied iitrces Radmothiology ResearchInstitrile. ltcihesda. MD 201t4-5145,
c 1984 Aan R. Lim~. Inc.

124 Litten, Carcillo, and Roth
INTRODUCTION
Hivman sepsis and septic shock are frequent causes of death in clinical medicine.Septic shock is characterized by peripheral vasodilation with a decreased systemic vascularresistance and hvxptension 11-31. In an attempt to increase the vascular resistance andelevate bhxxl pressure. clinicians usually administer catecholamines and catecholaminederivatives. However, a diminished peripheral vascular responsiveness to catecholaminesis commonly observed 141. thus making this approach frequently ineffective.
Alterations in vascular contractility may either iavolve extrinsic factors (prostaglan-dins. opioid peptides, etc.) or may be intrinsic to vascular smooth muscle. During endotox-emia and sepsis, previous investigators have demonstrated a diminished ability of NE toinduce contraction in rat aorta [see ref. 3 for review]. We previously demonstrated a de-crease in the number of aortic alpha, -adrenoceptors and a depressed aortic phosphoinositide(PI) metabolism in this rat model of chronic sepsis [51. Since we have shown that activationof the alphat-adrenoceptors with NE results in a stimulation of Pi metabolism [6,71, itappears that signal transduction involving alpha,-adrenoceptors is altered in aortae fromseptic rats.
In this study we determined whether calcium mobilization was altered in aortae fromseptic rats during the stimulation of alpha,-adrenergic receptors by NE. Calcium influxthrough calcium channels and calcium efflux, an indication of intracellular calcium releaseby the sarcoplasmic reticulum [81, were measured in the rat aortae. Both processes in rataorta are stimulated by the activation of alpha,-adrenoceptors 191. In addition, we measuredthe levels of protein kinase C as determined by PDBu binding sites in aortae from septicrats. Activated protein kinase C has been shown to phosphorylate smooth muscle myosinlight chain 110, 111 as well as alter calcium influx in vascular smooth muscle 112.131 andinduce rat aortic contraction 1141. We have also shown that activation of protein kinase Cinhibits Pl hydrolysis 171, which could explain, in part. the diminished Pl turnover observedin the rat aorta during sepsis 151.
MATERIALS AND METHODS
Chronic Rat Sepsis Model
Male Sprague Dawley rats (250-300 g) were obtained from Taconic Farms, Ger-mantown. NY. Control and experimental animals were randomly subjected to sham sur-gery or cecal ligation with two-hole puncture. respectively, as previously described 115.161.Twenty-four hours after surgery the surviving animals were killed by decapitation. In ourlaboratory the model has a 20-.30% mortality rate. Surviving animals displayed signs ofsepsis originally described by Wichterman et al. 1151 including piloerection, a bloodydischaree from the nose and mucous membrane, bloody diarrhea, and lethargy.
"5Ca Influx
Calcium influx was measured as described by Meisheri and van Breemen 1171 withminor modifications 1131. Briefly, 4-mm thoracic aortic rings were incubated at 37°C for45 min in oxygenated physiological saline (PSS) of the following composition (in mM):NaCI (140), MgCI, (I), CaCI, (1.5), KCI (4.6). glucose (10). HEPES buffer (5). pH 7.4(buffer A). The rings were transferred to PSS (37CC) containing "Ca (2 i.Ci/ml) and var-ious experimental protocols as described in figure legends. The rings were then washed inice-cold PSS for 30 min and placed in a hypotonic 5.0 mM EDTA solution overnight at7-B
7i
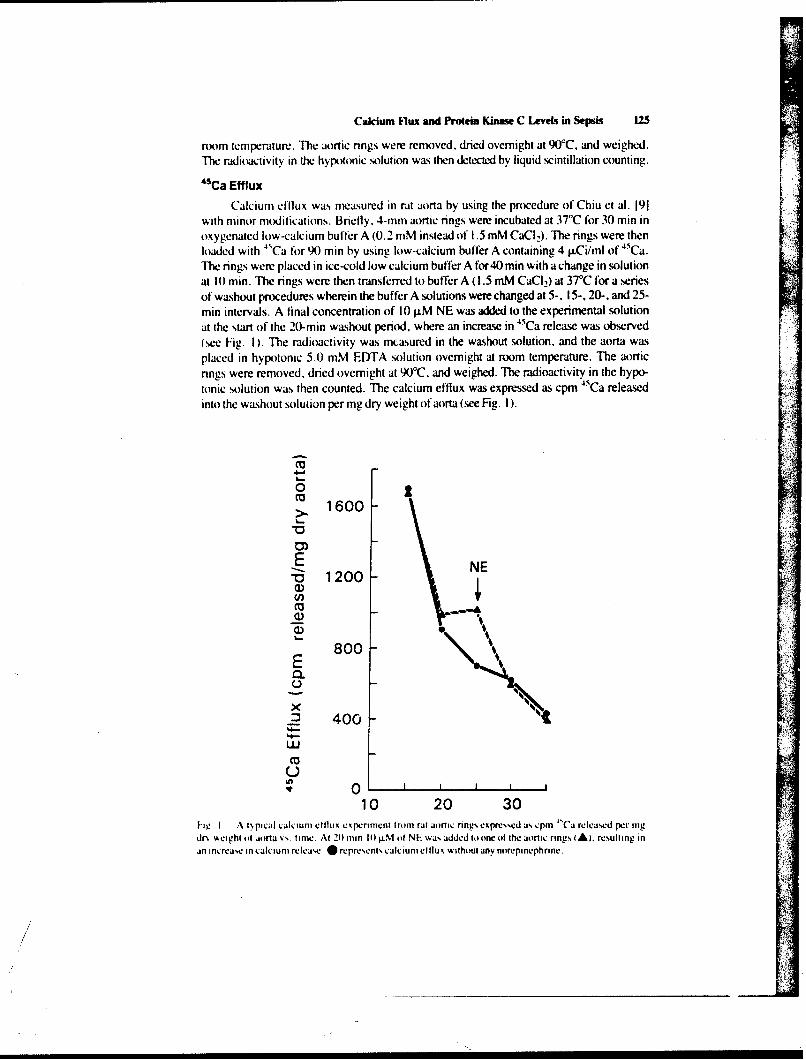
Calcium Flux and Proein Kinwe C Levels in Sepsis 12
room temperature. The aortic rings were removed, dried overnight at 90C, and weighed.The radioactivity in the hypotonic solution was then detected by liquid scintillation counting.
4Ca Efflux
Calcium efflux was measured in rat aorta by using the procedure of Chiu et al. 191with minor modifications. Briefly. 4-mm aortic rings were incubated at 37°C for 30 min inoxygenated low-calcium buffer A (0.2 mM instead of 1.5 mM CaC,). The rings were thenloaded with 4sCa for 9X) min by using low-calcium buffer A containing 4 AiCi/ml of 45Ca.The rings were placed in ice-cold low calcium buffer A for4O min with a change in solutionat 10 min. The rings were then transferred to buffer A (!.5 mM CaCI,) at 37C for a seriesof washout procedures wherein the buffer A solutions wee changed at 5-, 15-, 20-, and 25-min intervals. A final concentration of 10 j.M NE was added to the experimental solutionat the %tart of the 20-min washout period, where an increase in "Ca release was observed(see Fig. I). The radioactivity was mcasured in the washout solution, and the aorta wasplaced in hypotonic 5.0 mM EDTA solution overnight at room temperature. The aorticrings were removed, dried overnight at 9 0 C, "and weighed. The radioactivity in the hypo-tonic solution was then counted. The calcium efflux was expressed as cpm 45Ca releasedinto the washout solution per mg dry weight of aorta (see Fig. I).
o 11600
ENE" 1200C,,
_, 'S
8001.
U
x.2 400-
' O 1 I I I
10 20 30I-ig I A ipi.al calcium etllux experiment from rat aor c nng, expr,,d as cpm "Ca released per mgdrx ,eighio1 ajrta ,,, time. Al 21) mm Ii0 .M i NE wa, added Ioonc of the aortic rings A). reulting inan inrcae in calcium releas. represent calcium ellux without any norepinephrine.
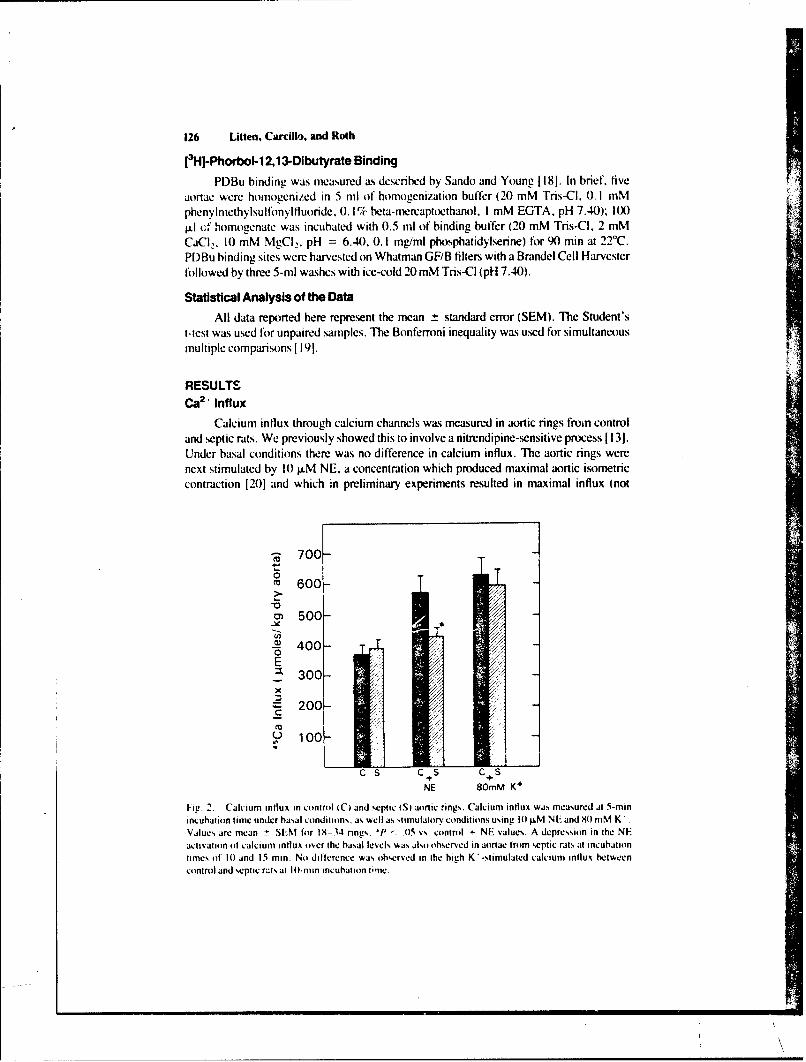
126 Utten. Carcillo, and Roth
[3H-Phorbol-1 2,1 3-Dibutyrate Binding
PDBu binding was measured a~s described by Sando and Young 11l81. In brief, fiveaortae were homogenized in 5 nil of homogenization buffer (20 mM Tris-CI. 0. 1 mMphenylIrnethylIsulIfonyl fluoride, 0. 1% bea-mercapmethanol. I mM EGTA. pH 7.140); 1(000± cf homogenate was incubated with 0.5 ml of binding buffer (20 mM Tris-CI. 2 mMCad 2, 10 mM MgC]2. pH = 6.40), 0.1I mg/mi phosphatidylserine) for 90 min at 22TC.PDBu binding sites were harvested on Whatman GF/B filters with a Brandel Cell Harvesterfollowed by three 5-mI washes with ice-cold 20 mM Tris-CI (pH1 7.40).
Statistical Analysis of the DataAll data reported here represent the mean ± standard error (SEM). The Student's
t-test was used for unpaired samples. T"he Bonferroni inequality was used for simultaneousmultiple comparisons 1191.
RESULTSCa2, Influx
Calcium influx through calcium channels was measured in aortic rings from controland septic rats. We previously showed this to involve a nitrendipine-sensitive process 1131.Under basal conditions there was no difference in calcium influx. The aortic rings werenext stimulated by 10 p.M NE. a concentration which produced maximal aortic isometriccontraction 1201 and which in preliminary experiments resulted in maximal influx (not
-R 700-0M 600->
o~500-
-~400-
E/
3 00-
2 00-
NE 80mM K+
Fig. 2. Calcium influx in control i C) and septic (St aortic ring%. Calcium influx was measured at 5-mmnincubation time under basal conditions, as %hell as stimulatory condition% using 1l. M NE and 90 mM K'.Values are mean -- SEM for 19-34 rings. J' - .05 vs. control + NE values. A depression in the NEactivation of calcium influx over the basal levels was also observed in aortae from septic rats at incubationtimes of 10) and 15 min. No difference was observed in the high K'-stimulated calcium influx betweencontrol and septic rut% at 10-nin incubation tine.
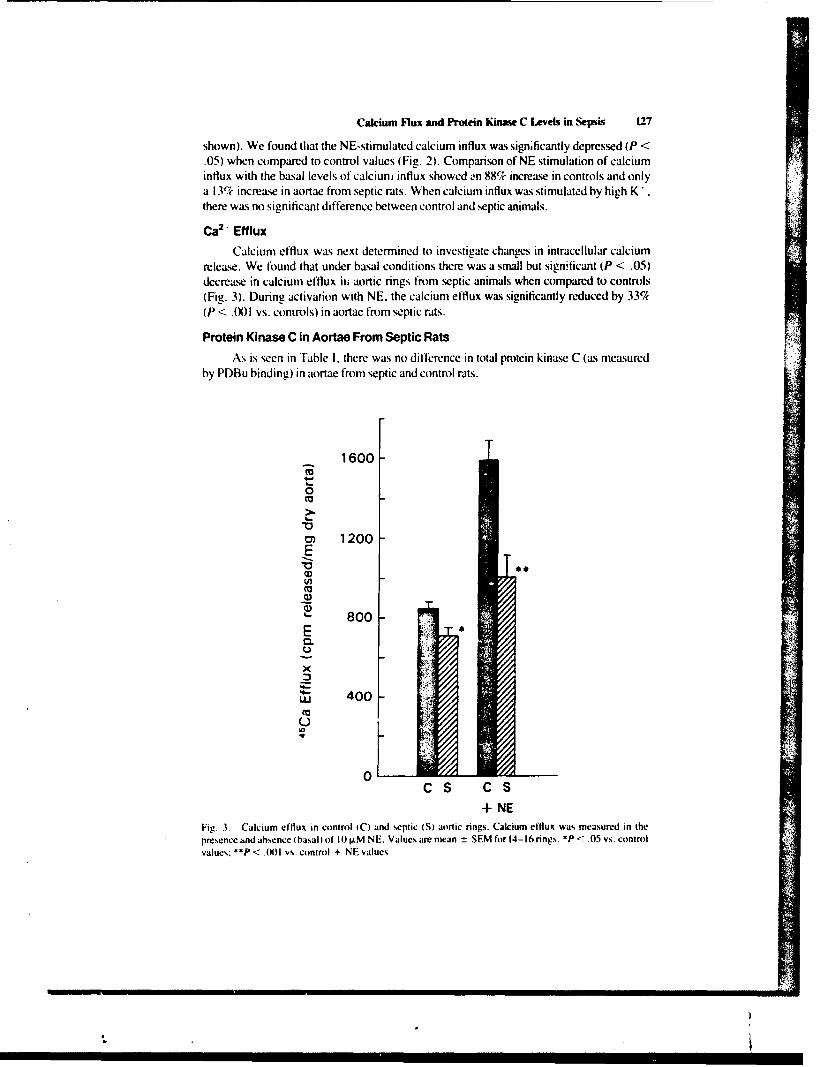
Calcium Flux ankd Pro~ein Kinase C Lee inSepsis 127
shown). We found that the NE-stimulated calcium influx was significantly depressed (P <.05) when compared to control values (Fig. 2). Comparison of NE stimulation of calciuminflux with the basal levels of calcium influx showed L-n 88% increase in controls and onlya 13%c increase in aortae from septic rats. When calcium influx was stimulated by high K*there was no significant difference between control and septic animals.
Ca2 'Efflux
Calcium efflux was next determined to investigate changes in intracellular calciumrelease. We found that under basal conditions there was a small but significant (P < .05)decrease in calcium efflux in~ aortic rings from septic animals when compared to controls(Fig. 3). During activation with NE. the calcium efflux was significantly reduced by 33%tP < .001l vs. controls) in aortae from septic rats.
Protein Kinase C in Aortae From Septic RatsAs is seen in Table 1. there was no difference in total protein kinase C (as measured
by PDBu binding) in aortae from septic and control rats.
1600-
(0
o~1200
800
U,
0.
C S C S+ NE
Fig. . Calcium efflux in control (C) and septic (S aortic rings. Calcium efflux was measured in thepresence kind absence (basal) of It) LM NE. Values are mean t SEM for 14-16 rings. *P <.05 vs. controlvalues; **P < .(X)l vs. control + NE values.

128 Litten, Carcillo, and Roth
TABLE I. 1 Hlphorbol-12.13-Dibutyrate Binding to Aortic Homogenate From Control andSeptic Rats*
Total PDBu binding 'ite(dpm ig
Control 34.(0X) 3.5()Septic 32.5() ± 4.250
'Value, are mean SEM for three ditffrent expenments. Data obtained are deeroibed in Materials andMethods There Aa%, no significant difference Iet-, en control and septic preparations.
DISCUSSION
The tindings in this study indicate significant alterations in alpha,-adrenergic recep-tor-mediated mobilization of extracellular calcium and the release of intracellular calciumin aortae from septic rats. Calcium influx was depressed by 25%. while calcium efflux wasreduced by 33(1( when compared with control values. At the same time, we found that theamount of protein kinase C as assessed by PDBu binding sites was unaltered in aortae fromseptic rats.
The decrease in calcium mobilization across the calcium channels could be related tothe observed depression in aortic contractility during sepsis. This effect could be causeddirectly by endoxotin or by other circulatory factors such as lymphokines or endothelium-releasing factorls). We previously showed that the activatior of protein kinase C by aphorbol ester increases the calcium influx in rat aorta [131 as well as altering the ability ofvasoactive agents to activate PI hydrolysis [21,221. Since levels of protein kinase C wereunchanged it is unlikely that changes in protein kinase C caused the observed change: amore likely explanation is the decrease in receptor levels in rat aorta, as we previouslydiscovered 151. We also found that P1 metabolism was depressed in the rat aorta duringsepsis 151, thus creating the possibility of decreased activation of protein kinase C by dia-cylglycerol (DAG). one of the released products of PI metabolism [23]. Future studiesusing specific antibodies against protein kinase C could verify this distinction betweenactivation of protein kinase C (i.e., by DAG) and the amount of the enzyme.
The decreased calcium efflux seen in sepsis suggests that intracellular calcium releasefrom the sarcoplasmic reticulum may be depressed in response to NE. This depressioncould be related to a decrease in inositol-1.4,5-trisphosphate ([P,), another product of PImetabolism which has been shown to cause calcium release from sarcoplasmic reticulumin vascular smo)th muscle [241. Indeed, recent studies by Chiu et al. 1251 in rat aorta aswell as our earlier studies 1211 indicate a good correlation between PI hydrolysis and intra-cellular calcium release. Furthermore, we found that levels of phosphatidylinositol-4.5-biphosphate (PIP,). the precursor to IP,. were decreased in aortae from septic rats 151.Alterations in calcium mobilization from sarcoplasmic reticulum have also been reportedin vascular smooth muscle during endotoxic shock. Soulsby et al. 1261 showed that theATP-dependent calcium uptake and cal-ium-stimulated ATPase activity were depressedby endotoxin in the microsomal-enriched fraction from canine aorta. Since the sarco-plasmic reticulum may be the major system controlling free cytoplasmic calcium in bloodvessels 18.271. changes in the release and uptake of calcium from the sarcoplasmic reticu-lum could he a major factor in explaining the depressed contractility in septic aorta.
Overall. we have demonstrated striking alterations in aortic alpha,-adrenergic recep-tor-mediated signal transduction during sepsis. We previously showed a decrease in alpha,-

Calcium Flux and Protein Kine C Lees in Sepsis 129
adrenergic receptor, and in the alpha.-adrenergic receptor-medizted PI hydrolysis 151.Furthermore, the decrease in calcium release from sarcoplasmic reticulum, as demon-strated by the decrease in calcium efflux, could be caused by a decrease in PIP, hydrolysis.Alpha 1-adrenergic receptor stimu lation of calcium influx across the calcium channels in theplasmalemnma was also depressed in sepsis. In addition, we have also shown an alterationof protein phosphorylation in aortae from septic rats 1281 which may reflect changes in theactivation. but not t amount. of protein kinase C. Thus these findings suggest that aportion of the diminished responsivity seen in rat aorta during sepsis could be rimlatcd to A
perturbations in this reccptor-mediated cascade.Finally. it is important to realize that, similar to these findings in aorta, Spitzer's
group found alterations in adrenergic and vasopressin receptor- mediated mobilization ofintracellular calcium in hepatocytes 1291. Also, analogous changes in alpha, -adrenergicreceptors were also observed in hepatocytes from endotoxemic rats 1301 as well as nearlythe same alterations in PI metabolism 13 11.
CONCLUSIONS
During experimental sepsis we found a depressed mobilization of extracellular cal-cium through calcium channels and depressed calcium efflux. indicating a diminished cal-cium release from the sarcoplasmic reticulum. The concentration of protein kinast- C wasnot altered. These findings along with our previous results suggest an alteration in aorticalpha, -adrenergic receptor- mediated signal transduction in sepsis.
ACKNOWLEDGMENTS
The authors appreciate the technical assistance of G.T. Gainey and the secretarialassistance of Marianne Owens and Mila M. Ly. This work was supported in part by theArmed Forces Radiobiology Research Institute. Defenise Nuclear Agency and Naval Med-ical Research Institute. Department of Defense, under Work Unit (X) 169 and Research TaskNumber MR(X) 1.0 1. 1032.
REFERENCESI les%. MLt. flastillo A. Greenfield Mi: Spctiruio tcard iovacular functions during grati-nceative sepsis.
Prot, Card ivac Di% 23:279- 29M. 19M 1.2. Ponieranhi K. Caw, L. H-elchen JR. Romwell PW: Vascular reactivity in endotoxin shock: Effect o1
Iioancor indomethicin Ireat~n Adi, Shock Re% 7:191. 1982.
31 (hernowk B. Roth HI.. Pharniacologic manipulation of the peripheral iasculature in shock: Clinical andesperoiintmal approaches. (irc Shock I X: 141 155. 1986.
4, (herrnom H. Raines r(;. Liake (R: Endogenous and exogenou% catecholamnines in critical care medi-cine. ('ri (are Med 11):41N-416. 1952.
S ( jrcillo JA. Li J. Venter X(. Roth III.: Molecular properties of altered alphal-adrenerLgic receptor% inrat intraperitoncal sepsis. (orc Shock 21:3012, 1 9X7.
6 cegan F. (Chcrno%4 H. Parrillo J. Roth lit.. Actis aton of* phosphatid% linosiloI turnoser in rat aorta byit -adrenc rf;it recepitoir stiimu la ioi n. 1: ur J tPharrnacol I 10i:3X9- 3%M. 1985.
7 MlcMillan M. Chernow B. Roth HI.: Phorhol esters inhihit alpha, -adrenergic receptor-stimulated phos-phoinomitde hydrolysis and contraction in rat aorta: Evidenct. fr a link between vasc.,ular contractionand phosphotnositide turnosr. Biochem Biophys Res (Commun 134:970- 974. 1986.
X Johns A. tLeiiten 1P. Yanamioto H. Hwoan.' K. sajn Breemen C: Calkium regulation in sa%4cular mtithmuscle ciintractilit-, Am J (CardiiI 59: 1 MA- 23A. I 9X7.

130 Litten, Carmillo, and Roth
9. Chiu AT. McCall DE. Thoolen M. Timinerman% P: Ca utilization in the constriction of rat aorta tolull and partial alpha- I adrenoceptor agonists. j Pharmacol Exp Ther 238:224-231. 1986.
10. Nishikawa M, Hidaka H. Adelstein R: Phosphorylation of smooth muscle heavy mtromyosin by cal-cium-activated. phospholipid-dependent protein kinase. J Biol Chem 258: 14M69-14072. 1983.
11. Nishikawa M. SelleriJR. Adelstein RS. Hidaka H: Protein kinase Cmodulates in vitro phosphorylationof the smooth muscle heavy mieromyosin by myosin light chain kinase. J Biol Chem 259:8808-88 14.
984.12. Sperti G. Colucci WS: Phorbol ester-stimulated bidirectional transmenib~ane calcium flux in A7r5
1.vascular smooth musc le cells. Mol Pharmacol 32:37-42. 1987.1.Litten RZ. Suba EA. Roth BL: Effects ofa phorbol esteron rat aortic contraction and calcium influx in
the presence and absence of BAY k 8644. Euri Pharmacol 144:185-191, 1987.14. Nakaki T, Roth BL. Chuang D-M. Costa E: Phasic and tonic components in 5-HT, receptor-mediated
rat aorta contraction: Participation oif Ca" - tnannels and phospholipase C. J Pharmacol Exp Ther2-34:442-4446. 1985.
15. Wichierman K. Baue AL Chaundry IH: Sepsis and sepic shock: A review of laboratory models and aproposal. J Surg Re% 29:1 89- 21. 1980.
l6. McMillan M. Chemow B. Roth BL: Alterations in hepatic alphal-adrenergic receptors in a rat model ofchronic sepsis. Circ Shock 19:185- 194. 1986.
17. Mcisheri K[). van Breemen C: Fltects ot 1-adrenergic stimulation oin calcium movements in rabbitaortic smooth muscle: Relationship with cyclic AMP. J Physiol (Lond) 331:429-441. 1982.
18. Sando JJ. Young MC: Identification of high-afhinity phortsil ester receptor in cytosol of EL4 thyi-nomnacells: Requirement for calcium magnesium. and phospholipids. Proc Nail Acad Sci USA 80:2642-2646. 1983.
19. Wallenstein S. Zucker CL. Fleiss JL: Some statistical methods useful in circulation research. Oirc Res47:1-9, 19801.
20. Suba EA. Roth BL: ProstaglanJ ins activate phosphoinositide metabol ism in rat aorta. Euri Pharmacol136:325-332. 1987.
21. Roth BL, Nakaki T, Chuang D-M. Costa E: 5 Hydroxytrvptamine, receptor% coupled to phospholipaseC in rat aorta: Modulation of phosphoinositol metabolism by phorbol ester. J Pharmacol Exp Ther238:480-485. 1986.
22. Roth BL. Chuang D-M: Minireview: Multiple mechanismsof serotonergic signal transduction. Life Sci41:10151-1064. 1987.
23. Nishizuka Y: The role of protein kinase C in cell surface signal transduction and tumor promotion.Nature 308:693- 698, 1984.
24. Somlyo AV, Bond M. Somlyo AP. Scarpa A: Inositol trisphosphate-induced calcium release and con-traction in vascular smooth muscle. Proc Natil Acad Sci 82:5231-5235. 1985.
25. Chius AT. Bo,.arth JM. Timmermans PBM: Relationship between phosphatidylinositol turnover andCa '' mobilization induced by alpha, -adrenoceptorstimulation in the rat aorta. J Pharmacol Exp Ther240:123- 127. 1987.
26. Soulsbs' ME. Bennett C'L. Hess ML: Canine arterial calcium transport during endotoxin shock. CireShock 7:139-148. 19801.
27. van Breeman C. Leekeman S. Leijien P. Yamamoto H. Loutenhiser R: The role of'superlicial SR inmodulating force development induced by Ca entry into arterial smooth muste. J Cardiovasc Pharma-col 8lSupp 8):SI I I-S 116. 1986.
28. Litten RZ. Carcillo JA, Roth BL: Vascular calcium metabolis m and protein phosphorvlation in ratintraperitoneal sepsis. Circ Shock 2 1:332. 19X7.
29. I~aciic IV. SpitieriA: Rat liver free cytosolic Ca> and glycogen phosphorylase in endotoxicosis andsepsis. AmiJ Physiol 25 I:R984-R995, 1996.
30. Roth BL, Spitter JA: Altered hepatic vasopressin and a1,-adrenergic recelitoi., after chronic endotoxininfusion. Am J Phsiit 252:E699- E70)2. 1987.
31. Spitter JA. Turco ER. IDeaciuc IV. Roth BL: Perturbation of transmembrane signaling mechanisms inacute and chronic endotiixemnia. In Schlag G. Redl H iedst: -First Vienna Shock Forum. Part A:Pathophysiological Role of Mediators and Mediator Inhibitors in Shock.** Progress in Clinical andBioilogical Research. Vol. 236A. New York: Alan R. Liss. Inc.. 1987. pp 401-41 H.
Trhe experiments described in this paper were pertormed in adherence to the NIH guidelines for the use of'experimental aniiiials.

Radiat. Phvs. Chem. Vol. 32. No. 3, pp. 349-353. 1988 AtitiiP0A000LV 0146-5724/88 S3.00+0.o0m. ada:..pp Inirun. anPl J. Rit t. I ppl Intrum. Part C Copyright C 1988 Perpmon Press pkinted in Great Britain. All nihts reserved 801111111 II 1U11111 T " 1o1--.
SRSS-24
MODELING RADICAL YIELDS IN ORIENTED DNA EXPOSEDTO HIGH-LET RADIATION
J. H. MILLER,' W. E. WILSON.' C. E. SWENIIERG,2 L. S. MyEi.s JR3 and D. C. CHARLTON
4
Pacific Northwest Laboratory, Richland, Wash., U.S.A.'Armed Forces Radiobiology Research Institute. Bethesda. Md. U.S.A.
'Uniformed Services University of the Health Sciences. Bethesda. Md, U.S.A.4Concordia University, Montreal. Quebec. Canada
(Received 16 June 1987)
Abstract-- Monte Carlo simulation of energy absorption in oriented fibers of DNA is used to model thedependence of free radical yields on the orientation of the fibers relative to a flux of ionizing radiation.We assume a large asymmetry in the thermal conductivity of the fibers that permits rapid transport of%,ibrational energy along a DNA molecule, but not between different molecules. Based on this assumption.our model predicts that thymine radical anions have a significantly greater probability of undergoing asecondary protonation reaction if they are produced by a flux of high-energy protons that is incidentparallel to the helical axis of DNA than if they are generated by a flux that is incident perpendicular tothe DNA molecules. These results are in qualitative agreement with experimental data on the yield of5.6-dihydrothymin-5-yl radica!s when samples of oriented DNA were exposed at 77 K to neutrons.
INTRODUCTION mine anion (T ) and guanine cation were the prin-cipal radical species present at 77 K. However, when
Understanding the mechanisms by which high-energy the neutron flux was approximately parallel to thecharged particles produce reactive chemical species in orientation of DNA molecules, the total yield ofbiologically significant materials is important for radicals was reduced and a new ESR signal character-assessing the health effects of low-level exposure to istic of the 5,6-dihydrothymin-5-yl (TH) radicalionizing radiation. This is particularly true for radi- predominated.ation with high linear-energy-transfer (LET) where The most obvious difference in the patterns ofcorrelations between track structure and target struc- energy absorption in the tw3 irradiation geometriesture can have a profound effect on the nature and is that charged particles that traverse the sampledistribution of damage. The role of nonhomogeneous nearly parallel to the helical axis of DNA have aprocesses in radiation chemistry was recently re- greater probability of multiple energy transfer to theviewed by Magee and Chatterjee."' Our research in same molecule than do particles incident on thethis area has focused on the use of detailed Monte sample perpendicular to the fiber orientation. ThisCarlo simulation of the spatial distribution of energy difference in the spatial distribution of energy absorp-absorbed from charged particles"' as a basis for tion should have no effect on the production of freeunderstanding the subsequent evolution of chem- radicals if energy and/or charge move between DNAically active species. In this paper we report the use molecules in a fiber as freely as they are transportedof this approach to investigate production of free along a single DNA chain. Hence the unusual orien-radicals in oriented DNA exposed to a flux of tation dependence of radical yields in this system mayhigh-energy protons. have important implications for intramolecular
Samples of oriented DNA fibers prepared by the energy and charge transfer in DNA.wet spinning technique have proven to be a valuable Arroyo et a.' speculated that the reduction ofexperimental tool for investigating the production of radical yields in the parallel irradiation geometry wasfree radicals in DNA by both u.v. light and ionizing due to enhanced recombination of precursor ionradiation." "' The recognition that spatially cor- pairs. This explanation seems plausible in light ofrelated electron gain and loss centers may be precur- recent measurements of conductivity by van Lith etsors of DNA double strand breaks"' has led to al,""' which suggest that the high mobility of electronsrenewed interest in this experimental system for in- in the ice-like water layer around a biopolymer couldvestigating radiation-induced DNA damage. Re- enhance intramolecular charge transfer. Arroyo et al.cently. Arroyo et al."' used this type of sample in attributed the appearance of TH" radicals in theconjunction with electron spin resonance (ESR) to parallel irradiation geometry to intramolecular trans-investigate the production of radicals at 77 K by fer oV triplet excitation produced in charge re-exposure to neutrons. When samples were irradiated combination. The long lifetime of triplet excitons atby a flux of neutrons incident perpendicular to the 77K"" certainly favors this mechanism. However,helical axis of DNA. ESR spectra showed that thy- due to the variation in triplet transfer rates," 21 triplet
349

350 J. H. MILLER et al
excitons tend to be restricted to the neighborhood of affinity, is beyond the scope of this paper. Rather wethymnine that is bounded by the nearest guanine or simply assume that the probability to form T in thecytosine on each side. Hence, in DNAs with roughly ith energy deposition event is proportional to theequal G-C and A-T content, the range of triplet amount of energy i, absorbed locally in that event.migration is not expected to be large. Ion quenching Hence, the initial concentration of T" is approxi-experiments'"' suggest a range of 5-30 base pairs. The mated by the equationshort lifetime of' singlet excitons'"' is probably themajor factor that limits the range of their migration. CT. (S, 0) = -T V 6 (4)
In this paper we present a model for the orientation
dependence of TH yields that is based on an assumed where g1" is the average yield of T, per unit ofas~mmetry in the transport of vibrational excitation absorbed energy and 6(x) is the Dirac delta function.in the oriented DNA samples. Asymmetry of phonon Using this approximation in equation (3) we canscattering rates in oriented DNA have been observed perform the space integration and obtain the result.using Raman spectroscopy: '" '' however, these low-frequency modes should not be very effective in Ifll(K)activating proton transfer to T" if the activation g -11. (5)energy is 0.2 0.7eV as suggested by the work of .9T
Grislund et al." Regardless of the mechanism, ourmodel is macroscopic in that asymmetric energy for the yield of TH" relative to the yield of itstransfer is approximated by the solution of thermal precursor T . This result is just a weighted averagediffusion equations wizh a coefficient of diffusion of the probability to convert T" to TH" over all thealong DNA that is 1000 times greater than the positions of energy deposition in a DNA moleculecoefficient for diffusion perpendicular to the helical from charged particles traversing the sample at aaxis. This model is developed in the next section of specified angle relative to the molecular orientation.the paper and our numerical results for the prod- The weighting factors express the relative probabilityuction of TH" radicals by proton irradiation at 0.3 that T' will be produced by a given energy absorp-and 1.0 MeV are discussed in the third section. tion event. .Conclusions and the potential for improved theory The rate of proton transfer used in equation (3) toand experiment are discussed in the final section. calculate PIH' depends upon the transient local tem-
perature T(x.t) through the relationTHEOR, k(x, t) = A exp[-Q T(x. t)], (6)
As suggested by Grdslund er al.."' we assume that where .4 is a constant of the order of ,ibration ratesthe major source of TH" radicals is the reaction (10" sec) and Q is the activation energy. If we
T' + XH* -. TH' + X. (I) completely neglect intermolecular transfer of vi-brational energy and make the additional assumption
where the proton donor. XH'. is probably water of that the heat capacity and thermal conductivity arehsdration In an excess of proton donor, this reaction independent of temperature, then T(x., t) can be,ill be pseudo first-order and the yield of radiation- obtained by solution of the one-dimensional thermalinduced TH radicals per DNA molecule is given by diffusion equation for a cluster of point sources. " ' 14)
the equation This result can be written in ihe form
Y,= 0 h: .CJ (V. 0)Pr 1 .x). (2) T(x. t) = T, + T,(-v. 1), (7)
where where T, is the ambient sample temperature,
PrH.) kc'.texp[- , k(x. t')di'Jdt 13) T(.c. t) =7. (l + r.. '.:expLOc -V, (9)
is the probabilits that a thymine anion formed at withposition v on an oriented DNA molecule, which weapproximate as a long cylinder of raj'us h = 10. A.
sAill be converted to TH by protona. on. The initial r 2D.. (9)concentration of T' . the rate of proturaiion. and theduration of the radiation-induced exccss temperature being the parallel diffusion relaxation time andare denoted by C, (v,0). k(x. t), and t,,. f0
respectisel,. TO = - 10)Modeling the pri . 'tion of T . which probably , 2nr'hAp(*
im.olhes the decay oi ghly excited states through being the initial excess temperature at x, due to,utoioni/ation channels follovked by capture of low- absorption of energy (,. The sample density, specificenergy secondary electrons in traps with high electron heat and coefficient of thermal diffusion parallel to
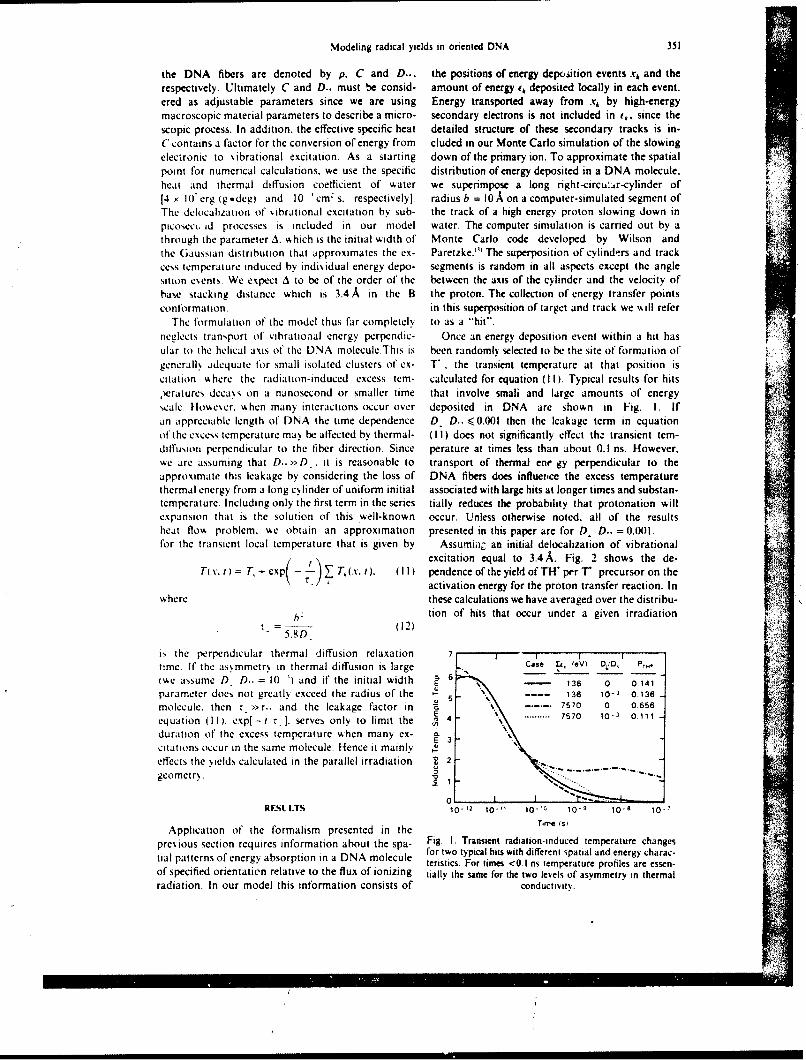
Modeling radical yields in oriented DNA 351
the DNA fibers are denoted by p. C and D.., the positions of energy deposition events x, and therespectively. Ultimately C and D.. must be consid- amount of energy i, deposited locally in each event.ered as adjustable parameters since we are using Energy transported away from xk by high-energymacroscopic material parameters to describe a micro- secondary electrons is not included in (,. since thescopic process. In addition, the effective specific heat detailed structure of these secondary tracks is in-C contains a factor for the conversion of energy from cluded in our Monte Carlo simulation of the slowingelectronic to vibrational excitation. As a starting down of the primary ion. To approximate the spatialpoint for numerical calculations, we use the specific distribution of energy deposited in a DNA molecule.heat and thermal diffusion coefficient of water we superimpose a long right-circu:ar-cylinder of[4 ) 10' erg (g*deg) and 10 'cm s. respectively], radius b = 10 A on a computer-simulated segment ofThe delocalization of vibrational excitation by sub- the track of a high energy proton slowing down inpicosect id processes is included in our model water. The computer simulation is carried out by athrough the parameter A. which is the initial width of Monte Carlo code developed by Wilson andthe Gaussian distribution that approximates the ex- Paretzke." The superposition of cylinders and trackcess temperature induced by individual energy depo- segments is random in all aspects except the anglesition events. We expect A to be of the order of the between the axis of the cylinder and the velocity ofbase stacking distance which is 3.4.A in the B the proton. The collection of energy transfer pointsconformation. in this superposition of target and track we %kill refer
The formulation of the model thus far completely to as a "hit"neglects transport of vibrational energy perpendic- Once an energy deposition event within a hit hasular to the helical axis of' the DNA molecule.This is been randomly selected to be the site of formation ofgenerally adequate for small isolated clusters of ex- T". the transient temperature at that position iscitation %here the radiation-induced excess tom- calculated for equation (Ill. Typical results for hits,Vratures decays on a nanosecond or smaller time that involve smali and large amounts of energyscale. However. when many interactions occur over deposited in DNA are shown in Fig. I. Ifan appreciable length of DNA the time dependence D. D.. <0.001 then the leakage term in equationof the excess temperature may be affected by thermal- ( I I ) does not significantly effect th. transient tem-diffusioni perpendicular to the fiber direction. Since perature at times less than about 0.1 ns. However,we are assuming that D.. >D it is reasonable to transport of thermal ene gy perpendicular to theapproximate this leakage by considering the loss of DNA fibers does influerice the excess temperaturethermal energy from a long cylinder of uniform initial associated with large hits at longer times and substan-temperature. Including only the first term in the series tially reduces the probability that protonation willexpansion that is the solution of this well-known occur. Unless otherwise noted. all of the resultsheat flo, problem. we obtain an approximation presented in this paper are for D, D.. = 0.001.for the transient local temperature that is given by Assumian an initial delocalization of vibrational
excitation equal to 3.4 A. Fig. 2 shows the de-Tcv. t= T, +- exp(- T, (.v. 0, 01) pendence of the yield of TH per T" precursor on the
activation energy for the proton transfer reaction. Inwhere these calculations we have averaged over the distribu-
tion of hits that occur under a given irradiation
= D (12)
is the perpendicular thermal diffusion relaxation 7 I
time. If the asmmetr, in thermal diffusion is large Case Zi, lev) D2 D,, PTH
(we assume D D.-.= 10 ') and if the initial width a 6 -- 136 0 0.141parameter does not greatly exceed the radius of the 5. 136 10-s 0.136
molecule. then r >>r.. and the leakage factor in ---..- 7570 0 0656
equation 111. exp(-t r_ . serves only to limit the a - 7570 1o- 0111
duration of the excess temperature when many cx- a 3E 3L
citations occur in the same molecule. Hence it mainlyeffects the yields calculated in the parallel irradiation g 2
geometry. 0RESL I.TS 10-12 10-11 10-'0 10-9 10-8 10-7
Application of the formalism presented in the rhme ts2
previous section requires information about the spa- Fig. I. Transient radiation-induced temperature changesfor two typical hits with different spatial and energy charac-til patterns of energy absorption in a DNA molecule teristics. For times <0.1 ns temperature profiles are essen-
of specified orientation relative to the flux of ionizing tially the same for the two levels of asymmetry in thermalradiation. In our model this information consists of conductivity.
iA
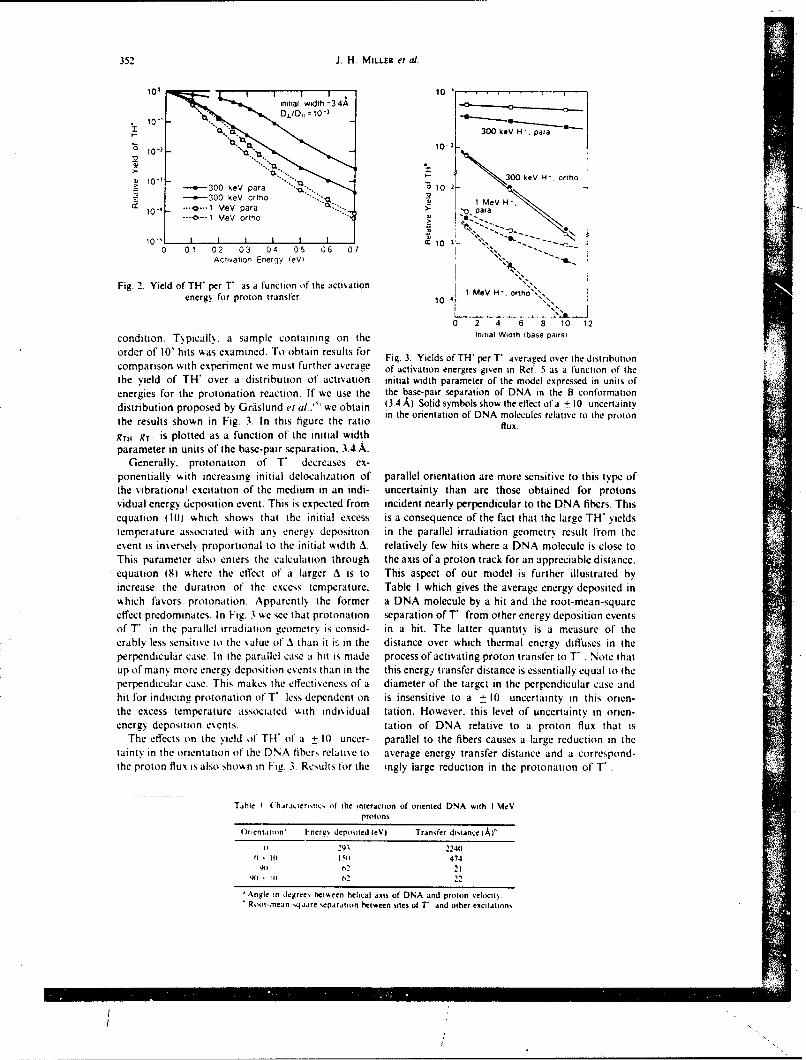
352 1. H. MILLER el al.
to0I 10-'inital wdth =3 4A• 0" t ~x. D1ID, = 10- 3
10 L- 300 keV H paraz "- 10- 2-
103 Z 10 2 - 300 keY H". ortho
- 300 keV para " 0 10 2- ' 300 key CrLh z...e
104 .---.-- l MeV para pI.-- MeV ortho10-1
0 0 11 0 12 0 13 0 14 0 15 0If6 07 ,a: ... 1-
AcTivation Energy ieVi
Fig. 2. Yield of TH" per T' as a function of the activation
energy for proton transfer. 10 1 MeV H'. ortho"s ,,5
0 2 4 6 8 10 12
condition. T~picall[, a sample containing on the Initial Width lbase pairsi
order of I0' hits was examined. To obtain results for Fig. 3. Yields of TH" per T' averaged over the distributioncomparison with experiment we must further average of activation energies given in Ref. 5 as a function of thethe yield of TH' over a distribution of activation initial width parameter of the model expressed in units ofenergies for the protonation reaction. If we use the the base-pair separation of DNA in the B conformationdistribution proposed by Grislund et al..' we obtain (3.4 A). Solid symbols show the effect of a + 10 uncertaintythe results shown in Fig. 3. In this figure the ratio in the orientation of DNA molecules relative to the proton
flux.
gTH' gT' is plotted as a function of the initial widthparameter in units of the base-pair separation, 3.4 A.
Generally. protonation of T" decreases ex-ponentially with increasing initial delocalization of parallel orientation are more sensitive to this type ofthe vibrationa! excitation of the medium in an indi- uncertainty than are those obtained for protonsvidual energy deposition event. This is expected from incident nearly perpendicular to the DNA fibers. Thisequation (10) which shows that the initial excess is a consequence of the fact that the large TH" yieldstempetature associated with any energy deposition in the parallel irradiation geometry result from theevent is inversely proportional to the initial width A. relatively few hits where a DNA molecule is close toThis parameter also enters the calculation through the axis of a proton track for an appreciable distance.equation (8) where the effect of a larger A is to This aspect of our model is further illustrated byincrease the duration of the excess temperature, Table I which gives the average energy deposited inwhich favors protonation. Apparently the former a DNA molecule by a hit and the root-mean-squareeffect predominates. In Fig. 3 we see that protonation separation of T from other energy deposition eventsof T' in the parallel irradiation geometry is consid- in a hit. The latter quantity is a measure of theerablv less sensitive to the vLalue of A than it iFs in the distance over which thermal energy diffuses in theperpendicular case. In the parallel case a hit is made process of activating proton transfer to T . Note thatup of many more energy deposition events than in the this energy transfer distance is essentially equal to theperpendicular case. This makes the effectiveness of a diameter of the targct in the perpendicular case andhit for inducing protonation of T less dependent on is insensitive to a + 10 uncertainty in this orien-the excess temperature associated with individual tation. However. this level of uncertainty in orien-energy deposition eents. tation of DNA relative to a proton flux that is
The effects on the yield of TH" of a + 10 uncer- parallel to the fibers causes a large reduction in thetainty in the orientation of the DNA fibers relative to average energy transfer distance and a correspond-the proton flux is also shown in Fig. 3. Results for the ingly large reduction in the protonation of T"
Table I (hirjcteri,tic, ot the interaction of oriented DNA with I MeVprotons
Orientation' Energ, depoited leVI Transfer distance i "
0i _91 2240(I - iii 511 474IM (2 21
IN)- 2 22
Angle n degrees hetween helical ats of DNA and proton velocit)Root-,nean-,quare separation between sites of T and other excitations
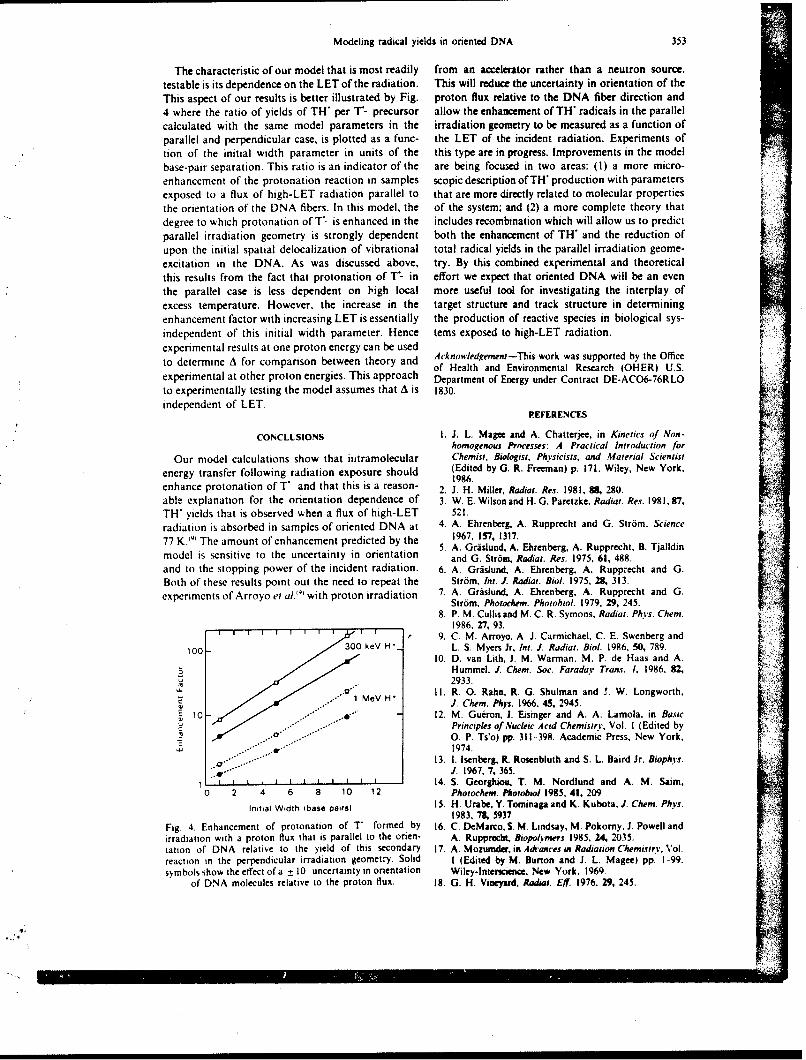
Modeling radical yields in oriented DNA 353
The characteristic of our model that is most readily from an accelerator rather than a neutron source.testable is its dependence on the LET of the radiation. This will reduce the uncertainty in orientation of theThis aspect of our results is better illustrated by Fig. proton flux relative to the DNA fiber direction and4 where the ratio of yields of TH" per T*- precursor allow the enhancement of TH" radicals in the parallelcalculated with the same model parameters in the irradiation geometry to be measured as a function ofparallel and perpendicular case, is plotted as a func- the LET of the incident radiation. Experiments of
tion of the initial width parameter in units of the this type are in progress. Improvements in the model
base-pair separation. This ratio is an indicator of the are being focused in two areas: (i) a more micro-enhancement of the protonation reaction in samples scopic description of TH" production with parametersexposed to a flux of high-LET radiation parallel to that are more directly related to molecular propertiesthe orientation of the DNA fibers. In this model, the of the system; and (2) a more complete theory thatdegree to which protonation of T*h is enhanced in the includes recombination which will allow us to predictparallel irradiation geometry is strongly dependent both the enhancement of TH" and the reduction ofupon the initial spatial delocalization of vibrational total radical yields in the parallel irradiation geome-excitation in the DNA. As was discussed above, try. By this combined experimental and theoreticalthis results from the fact that protonation of T in effort we expect that oriented DNA will be an eventhe parallel case is less dependent on high local more useful tool for investigating the interplay ofexcess temperature. However, the increase in the target structure and track structure in determiningenhancement factor with increasing LET is essentially the production of reactive species in biological sys-
independent of this initial width parameter. Hence tems exposed to high-LET radiation.experimental results at one proton energy can be usedto determine A for comparison between theory and Acknowledgement-This work was supported by the Office
n aof Health and Environmental Research (OHER) U.S.experimental at other proton energies. This approach Department of Energy under Contract DE-ACO6-76RLOto experimentally testing the model assumes that A 25 1830.independent of LET.
PEFERENCES
CONCLUSIONS I. J. L. Magee and A. Chatterjee. in Kinetics of Non-homogenous Processes: A Practical Introduction for
Our model calculations show that ititramolecular Chemist, Biologist, Physicists, and Material Scientist
energy transfer following radiation exposure should (Edited by G. R. Freeman) p. 171. Wiley, New York.1986.
enhance protonation of T" and that this is a reason- 2. J. H. Miller, Radiat. Res. 1981, S, 280.
able explanation for the orientation dependence of 3. W. E. Wilson and H. G. Paretzke, Radiat. Res. 1981, 37,TH" yields that is observed when a flux of high-LET 521.radiation is absorbed in samples of oriented DNA at 4. A. Ehrenberg, A. Rupprecht and G. Str6m. Science
77 K." The amount of enhancement predicted by the 1967, 157, 1317.e othe u 5. A. Grislund. A. Ehrenberg. A. Rupprecht, B. Tjalldin
model is .sensitive to the uncertainty in orientation and G. Str6m. Radiat. Res. 1975, 61, 488.and to the stopping power of the incident radiation. 6. A. Grislund, A. Ehrenberg. A. Rupprecht and G.Both of these results point out the need to repeat the Strom, Int. J. Radiat. Biol. 1975, 28, 313.
experiments of Arroyo et al."' with proton irradiation 7. A. Grislund, A. Ehrenberg, A. Rupprecht and G.Str6m, Phorochem. Photobiol. 1979, 29, 245.
8. P. M. Cullis and M. C. R. Symons, Radiat. Phvs. Chem.1986. 27, 93.
p 9. C. M. Arroyo, A. J. Carmichael, C. E. Swenberg and
lO0- 300 keV H*_ L. S. Myers Jr. Int. J. Radiat. Biol. 1986, 50, 789.10. D. van Lith, J. M. Warman, M. P. de Haas and A.
5Hummel, J. Chem. Soc. Faraday Trans. I. 1986, 32,.. 2933.
"11. R. 0. Rahn, R. G. Shulman and 3. W. Longworth,.-'" 1 MeV HJ. Chem. Phrs. 1966. 45, 2945.
E10 ." .12. M. Gueron, J. Eisinger and A. A. Lamola, in BasicPrinciples of Nucleic Acid Chemistry. Vol. I (Edited by0. P. Ts'o) pp. 311-398. Academic Press. New York,
-""" 1974.
.0"" .... 13. I. Isenberg. R. Rosenbluth and S. L. Baird Jr. Biophys.J. 1967, 7, 365.
1 14. S. Georghimo T. M. Nordlund and A. M. Saim,0 2 4 6 8 10 12 Photochem. Photobiol 1985. 41, 209.
Initial Width (base pairs) 15. H. Urabe, Y. Tominaga and K. Kubota. J. Chem. Phys.1983. 78, 5937.
Fig. 4. Enhancement of protonation of T" formed by 16. C. DeMarco, S. M. Lindsay, M. Pokorny, J. Powell andirradiation with a proton flux that is parallel to the orien- A. Rupprecht, Biopolymers 1985, 24, 2035.tation of DNA relative to the yield of this secondary 17. A. Mozumder. in A.ances in Radiation Chemistrv, Vol.reaction in the perpendicular irradiation geometry. Solid I (Edited by M. Burton and J. L. Magee) pp. 1-99.symbols show the effect of a = 10 uncertainty in orientation Wiley-lnterence New York. 1969.
of DNA molecules relative to the proton flux. 18. G. H. Vinepard. Radiot. Eff. 1976. 29, 245.

OmI$ADO SIO LOOVINF. J. HAfl1.1r. HIO.., 1988, vol.. 53, o. 6, 901-907 I811,1tuvU
Sfl88-25
Rapid communication
Stochastic model of free radical yields in oriented DNAexposed to densely ionizing radiation at 77K
J. 11..MILLERt, XV. E. \VILSONt, C. E. S\V ENBER(;++,
L. S. IYERS Jr§ and 1). E. CHARLTONr
SlPacific Northvest Laboratorv, P() Box 999, Richland, Vashington 99352,L.S.A.++ Armed Forces Radiobiology Research Institute,Bethesda, Maryland 20814-5145, U.S.A.§ Uniformed Services lniversity of the Health Sciences,Bethesda, Marvlatd U.S.A.
(oncordia University, .Montreal, Quebec, Canada
(Received 23 September 1987; revisio. received 6 Jania,y 1988;ac'ept'd I, JaptuarY 1988)
Monte Carlo simulation techniques were used to calculate the probability thatthymine radical anions (T'), formed by the sloving-do\n of high-energyprotons in oriented DNA, will undergo a secondary protonation reaction. Byassuming a large asymmetry in the thermal conductivity of oriented DNA fibreswe predict a significant enhancement of protonation ofT1 when the proton flux isincident on the sample parallel to the orientation of the DNA. These results are inqualitative agreement %kith experimental data on the production of TH radicalswhen oriented )N.A is exposed to fast neutrons.
Oriented DNA fibres prepared by the wet spinning technique of Rupprecht(1966) have been extensively used to investigate the production of free radicals by
ultraviolet (u.v.) light ((;rfislund et al. 1971) and gamma irradiation (Grfislund et al.1979, 198I, I !iittermann et al. 1984). Recently Arrovo and co-workers (1986) usedoriented Na DNA films in conjunction with electron paramagnetic resonance
(e.p.r.) to measure the orientation dependence of radicals produced y exposure at77K to fast neutrons from the TRIGA reactor at the Armed Forc,:s RadioiologyResearch Institute. For samples irradiated by a neutron flux perpendicular to thehelical axis of DNA, the e.p.r. spectra recorded at 77K were consistent with roughlyequal amounts of thymine anion (T-) and guanine cation, a result in agreel.ent with
the studies mentioned above using u.v. and gamma-irradiated samples. However,when the neutron flux was parallel to the oriented DNA fibres the total radical yield\as significantly reduced, and a signal characteristic of 5,6-dihydro thymin-5-vl('Ill) radicals became the predominant feature in the e.p.r. spectrum.
Since neutrons are absorbed in hydrogenous materials mainly through theproduction of recoil protons, sve are investigating these unusual findings through thestudy of free-radical production in oriented DNA by exposure t o proton fluxes. Thispaper examines quantitatively the hypothesis that intramolecular energy transfer is
responsible for the orientation dependence of radical yields reported by Arroyo et al.

qD R2 Rapid ioimnunitilion
(1986). Although the probability of multiple energy depositions in the same )NAmolecule is much greater for protons with velocity nearly parallel to the fibreorientation, this difference between the patterns of energy absorptior in the paralleland perpendicular case does not necessarily imply vn orientation dependence of free-
radical yields. For example, if each ion pair induced in DNA by the radiationexposure evolved independently, then the radiation chemistry of free radicals \\ o(idbe decoupled from the o% erall patterns of ionization in the sample. ILike%\ ise, if ion
pairs in different )NA molecules interact as strongly as ion pairs in the samemolecule, then th.. impact of this interaction on radical yields should be the same forparallel and perpendicular irradiation. This paper investigates the complex interplaybetween the track structure of densely ionizing radiation and the asymmetry ofenergy migration after deposition as a possible explanati~m for the unusual
observations reported by Arroyo el al. (1986).It is difficult to assess the degree to which secondary protons in the experiments
of Arroyo e! al. (1980) preserve the initial orientation of the neutron flux. it is also,
difficult to predict absolute yields of free radicals in a system as complex as DNAexposed to densely ionizing radiation. I lence we do not attempt to make quantitativecomparison l)et%\ cen the theory and experiment in this paper. The goal of this work isto calculate the probability that T- \% ill undergo a secondary reaction to form T1 I asa function of the energy and direction of the proton track in which the thymine
radical anion is produced. \We predict that this secondary reaction is significantlyenhanced in proton tracks with high stopping power that are nearlyN parallel to the)NA. This findinv supports the observation by Arroyo el al. (1986) that TlI- is the
predominant radical sptecies formed in the parallel irradiation geometry. Similararguments may also explain the observed reduction in total radical yields; however,to make quantitative predictions concerning the total radical yield requires moredetailed information about the thermally induced decay of primary radical speciesthan is currently available. This is particularly true with regard to the fate of radicalcations (I le et al. 1987).
Our model assumes the following: (a) The major source of TH' radikals isthermally activated protonation of'l, with water of hydration being the most likelyproton donor. This hypothesis is consistent with the findings ot Griislund et al.
(1975), \ ho iinvestigated the formation of TI I by anneihng of radicals produced bygamma irradiation of oriented )NA. (b) Intramolecular energy transfer is muchmore rapid than intermolecular energy transfer, even for two DNA molecules in thesame oriented D)NA fibr( In our macroscopic model this assumption is expressed asa large asymmetry in the coefficient of thermal diffusion. (c) Spatial pattern of energyabsorption in oriented DNA molecules can be approximated by the overlap of acylinder of uniform radius 10 A with the track of a high-energy proton (Wilsonand Paretzke 1981) that is slowing down in a homogeneous material of unit density.This approximation, which is based on inelastic scattering of protons and secondary
electrons by \water molecules, is assumed to be adequate for calculating the transientexcess temperature that activates the protonation of T!.
The production of T-" probably results from autoionization of highly excitedplasmon-like'states followed by capture of the low-energy secondary electrons atbase sites with high electron affinity. Since we cannot model these processes based oncomputer simulation of the slowing down of protons in water, we assume that theprobability of formin, T in an energy deposition event at position x i in a targetcylinder is proportional to the amount of energy i:i absorbed locally in that event.

Raipid imiiglin90 3
This approximation leads to the expression
t'lir th lit- \ oi-LI(I TI I relatt i\e to the \yield of its precursor T ,~here ['Tit( V is the
pritbahi lit\ of thermnall\ acti\ -atcd proton transfer at x, and the %\eigh ing factors i:,rctleet thet bias that inelastic coljisions " ith large eniergty transfer to the absorbing
Mtediunm are, more likel\ to he the type that would lead to f*ormationl of TIni an excess ol proton donors the protonation reaction will be pseudo-fi rst-order
Miid the pri babilit% that protonation \%Ill occur at position x is given by
Pill( %)= kCV. 1) ex F fk(x.i')dt'ldt (2)
'\ here ,,,i the duration of the radhiton-iniduced excess temperature. '[he rate otprotont fil,~ k( %,I), is giv en 1w
k(V, t) = .cx p t- Q T 1~) (3)
Michre . 1 is a constant, the order of vibrational rates, 10"s'- , Q is the activationeneriz for protonation. and T(.v, 1) is the transient temperature. WVe assume that hoth
the hevat capacity and thermal conductivity are temperature- independent and thatthertml diffusion parallel to the D)NA fibres (!),,) is rapid compared to diffusionperpeitndicular (/) - to the fibres. In the limit where only the shortest transverserelaxation time (r- = h2 5X))is included
~~here T, is thet ambient sample temperature, r, =A2. -'D,, is the longitudinal
thermal relaxation time, and T, is the excess temperature at position A and time I duetio abworption of energzy i:, at position x,. Tlhe sum over k includes all energydteposit i4irs in) the target cylinder that approximates a D)NA- molecule, except the onewivcd is the site of formation oif TF
B\~ ok ing the thermal diffusion equation in one dimension, one finds
T'= Tj, I + I r. lxp - 1 2 (X (-h) 2 2A-( I + I r, ) (5)
v hre
T'= 4: ((2,r')' 2h2A;p(') (6)
l Otlt ial excess tempe-rature at vi due to the absorption of energy iA,. '[he sample
dct-t Mtid spcific heat art denoted respectively by 1) and C . 1,1timately ('and D..
ni1' itb treated as adjustable parameters of the model since we are using macroscopicmiate-rial paranicttrs ito describe microscopic processes. As a starting point for ournumecrical calcuilations \\ use their \\ater values 4 x 10 () rg g-deg and I0f) -1cm 2 s 1rtespicticly. '[he initial width. A. of the Ga~ussian distribution that approx imates the
p~atili distribution of' excss temperature from an individual energy depositionc~ucot iti I)NA should not cecd a few~ base-pairs if the estimates given by Blrandt.11)d Ritchie( 19741 fo(rthe delocaliizaition oifeollective excitaitionis in the water apply toI ) NA.
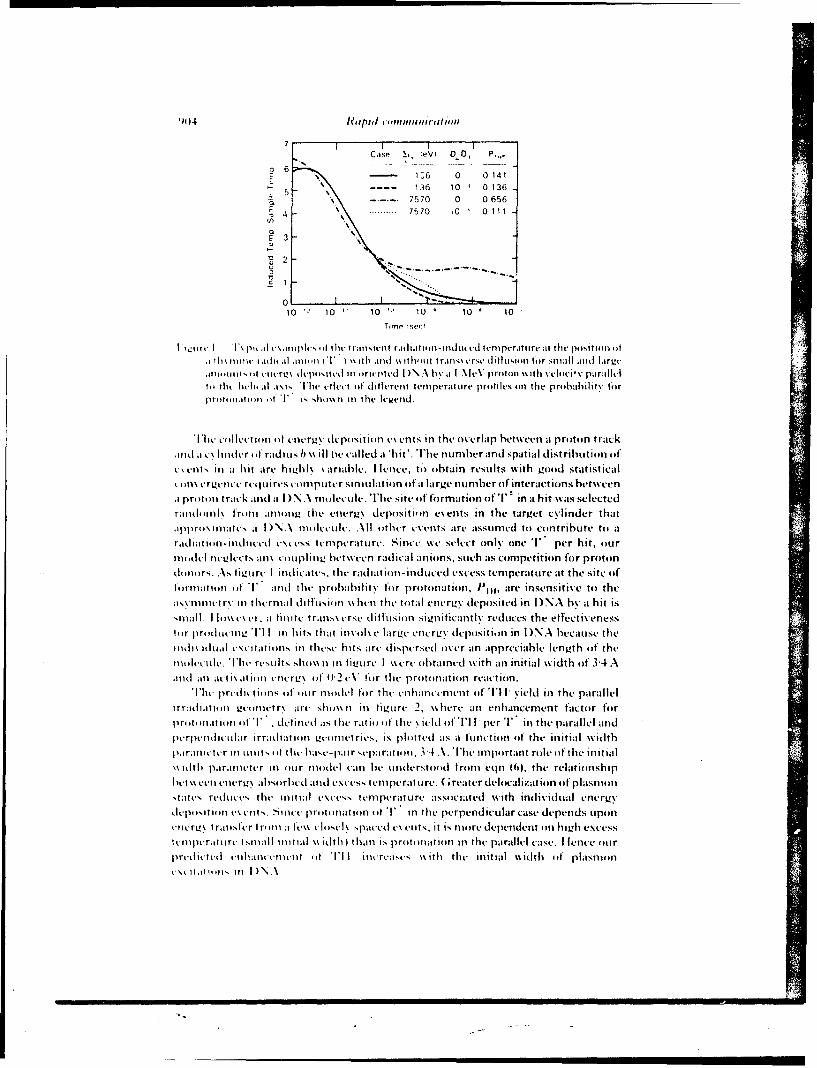
7 1 1Case (eV Di 0, P.
z6- 1A6 0 0 141
- -. 136 10 10 1365
7----7570 0 06564
E 3
010 10 10 11) 10 '~10
Time 1w,--
I w.ilit I TI pi, i .iiiijilis .1 )Ifit, traitsittlt raiitin-intiticed ttmlperature it the pIsitin ot.I 11i% 111i11t i.1iti .ii i (T I 4 ii it midl %% ithi'.iii Ir t-set titlisiii for mall andt I;Irzt-imint ofiil tiitict% ileposittil in oritrtcd DN .\ a I \IvV prolton %%tit %vlchwi parllt-1
T'. ti t hu, i al 1\1 s. The t tht0 t-it di titren t tern perattoart proijles (n the p ibahii ,ty fo
initi-i ot Tl i, slii inl the Irwend.
Ih litti llecItimili it oIntrZy depositioin events in the overlap between at prolton track
Midil a c\ Iiiitlr ofi raditis b %\il Il e called a *hit'. The number and spatial distribution oft
v.4 cts in at hit aire hiv.h I\ \ariable. I Irnc*, to obtain results with gooid statisticaltqin\ trgltt- rttlui res coimpuiter simullatio)n of a large number of interactions btween
A priitt in track and a I ) NA. molecule. The site ot formation oTlI in a hit was selectedlandmiiilk fromti afliiiL tile eilert' deposition events in the targ~et c\y.linder thatipprimt l\nattes at I)A mu lectle. All ither events aire assumred to contribute to at
r~il t on-inlieel \ccss tenmperatulre. Since %%e select onily one T per hit, ourtIll i neLIects in\ coupmphn! betwet radical anions, such as competition for protondono14 rs. As tiwiire I indicates, tilt- radiation-induced. ecess temiperature at the site oIftiriii ()f T andh tile probabilit\ tiir protonation. I'l,, are insensitive to theissniilet rx in t hernial tI itflsiu n %% hen the total etlerLv deposited in D)NA by at hit is'ITIi I. I Iic~r 454 e i the t ravilsvrse diffusioin sigmiiicatll reduces the effectivenesstiir pi)ItLuILL11' I I in hits that nlvuils larL.,c encrcv% deposition in D NA because the111(1is ilnil t'ii talt iiis In these hits ar- 4.1ispersed over an appreciable lengith of thenli itcle. 'I'lit res Ilts shi issii Ii fiviirt I wecre obtained with ain initial w.idth of 3-4A.Mitd an achi tion tnierL!% of tt2 e\ for the priotinatiion reactioin.
lilt lrtitiis of1 our nlnthel for t114 enhlancement of 'I'll- yield inl the parallelrrithiatiu ii vei nletr\ art, sl4ioS in~ fiiUre 2, xs here ain enhlanicement factor for
priiliiatioiint 1) T detintl ats the ratioft die %icld iii i'llI per T in thelparallel antdptripititliciifar ir-rad.iatii vctnn14trics, is. plnte4. as a ttfunction~ of the initial witdthimritllttcr-1 iii iiitS it tht'lat-ii taain 3-4 A\. Ilwiimphortant role 'it heinitial
ss idth parailtttr ill iiir umoe c41. an lbe untderstoo 44trom eqIn (01. the relationship
shatts retduces the- Iitial t\ctss temnperito rv assoceiated. wvith individual energyvisi)'tii in tv4.uts. Si iltt. pri it, iatii n of T in thet perpendicular case depends Ipi in
ti4rL'\ I rausttri Imn it a f'th 14sti\ paced cx .its, it is more depe~dent On Iisi~zh excessik4nilptraitiirt- Isuiali initial ss ldth than is priitimutiiin Ii the parallel case. I len4.e oitr
prutfit tth tnllauttvtitt ()t TI I increases %%titll tlt initial %\idth of plasniones li inii I )\A
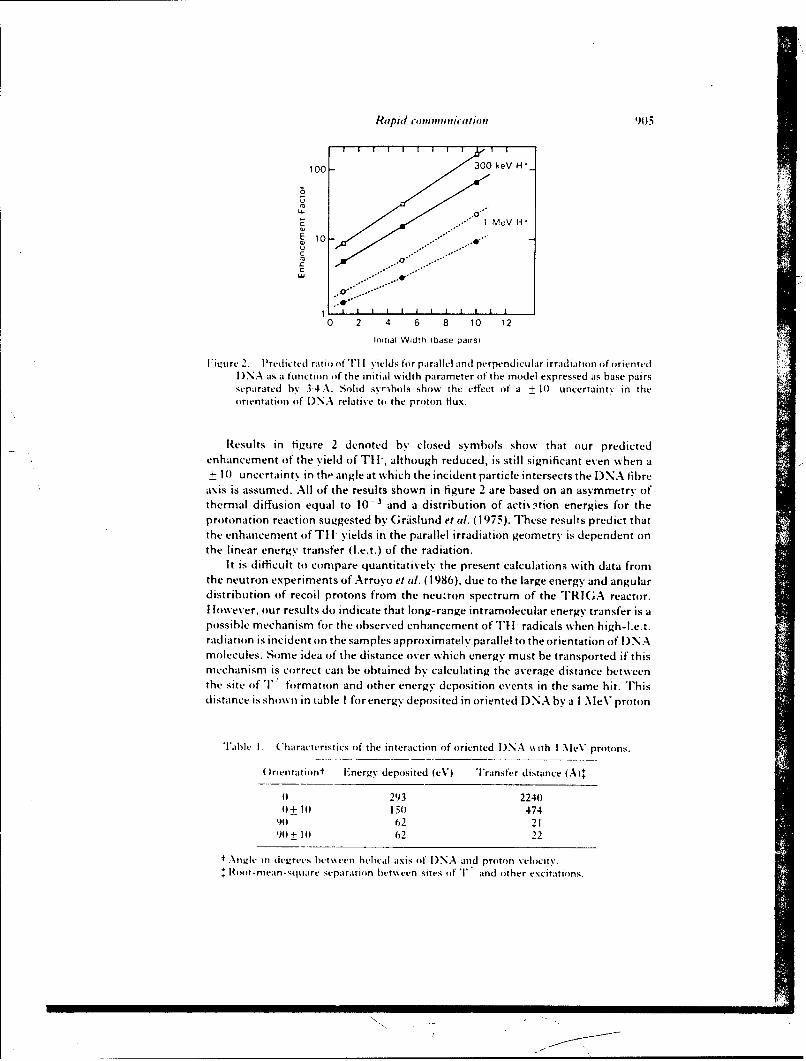
Rapid enini..icatio. 905=
100 300keV H*_
1MeV H,E !0o-"'
C .00
0 2 4 6 8 10 12
Initial Width (base pairs)
'iiure 2. Predicted ratio ot I I yields for parallel and perpendicular irradiation of oriented)NA as a function of the initial width parameter of the model expressed as base pairs
separated by 3"4 \. Solid syribols show the effect of a + 10 uncertainty in theorientation of )NA relative to the proton lux.
Results in figure 2 denoted by closed symbols show that our predictedenhancement of the yield of TI 1, although reduced, is still significant even when a+ 10 uncertainty in the angle at which the incident particle intersects the I)NA fibre
axis is assumed. All of the results shown in figure 2 are based on an asymmetry ofthermal diffusion equal to 10- and a distribution of activation energies for theprotonation reaction suggested by Griislund et al. (1975). These results predict thatthe enhancement of TI I yields in the parallel irradiation geometry is dependent onthe linear energy transfer (l.e.t.) of the radiation.
It is difficult to compare quantitatively the present calculations with data fromthe neutron experiments of Arroyo et al. (1986), due to the large energy and angulardistribution of recoil protons from the neutron spectrum of the "'lRIGA reactor.I lo\ever, our results do indicate that long-range intramolecular energy transfer is apossible mechanism for the observed enhancement of 11t- radicals when high-l.e.t.radiation is incident on the samples approximately parallel to the orientation of DNAmolecules. Some idea of the distance over which energy must be transported if thismechanism is correct can be obtained by calculating the average distance betweenthe site of 'F formation and other energy deposition events in the same hit. Thisdistance is shown in table I for energy deposited in oriented DNA by a I MeV proton
Table I. Characteristics of the interaction of oriented )NA %ith I MeV protons.
()rientation" Energy deposited (eV) Transfer distance (A)+
0 293 22400+ 10 150 474
9i 62 2190+10 62 22
" .-\iiele in devrees bt's een helical axis of I)NA and proton velocity.,oot-mean-square separation between sites of T and other excitations.

1141 R Iopiul connu nicaii
fil. A-S \%oUld be expected. ititramnolecular eniergy transfer distances in oiur niodelare comnparable to the diamieter of the D N A double helix %%hen the proton flux is
ticarl. perpenidicular to thle helical axis of' I )N A; however for the parallel case, thleiv eia~e cniir-t ranster distance is miany timies greater than the diamneter of thetin ilecu Ic.
)ur niacro scoiic ni ide I based onl thle soliiton of thermal di fuLsion eq lat ii nS,
does not pro% ide miuch insight into miole-Cular mechanismis of the assumned energymig~ration along D NA. Singlet excitoins are not a likely candidate for the miobileenergy state due to their short litetinie (( ;orgh iou 0 (1/. 1985) and miigration ofttriplet excitons in D)NA wvith inhologeneous base sequence is not expected toexceed 1 () 21) oif base pairs (Gueron el al. 19'74). Asymnmetry in phonion -scat teri ngrates has been detected by Rarnan spectroscopy of oriented D)NA (Urabe et al. 1983,1 )e\ larco el at. 1985), but these low%-frequency niodes are noit expected to be very
teti C y iii activat ing proitoni tranisfer toT .I I fence, although our model calculationsshow% that long-range enrgyj, mnigratitin Ii D)NA could account for the unlusual
orintaiondepndece il. I ields observed by Arrooet WI. (19'86), the mnolecularbasis of this hypothesis remnains Uncertain. We are Currently attemptitng to includeradical recornbination in our mnodel to investigate the orientation dependence of theti tal raidical yield. We are also starting experimients wvith direct proton irradiation of'oriented I )N.- to obtaini experimiental data wvith aI more wvell-deliiied proton flux forcoimparison with our miodel predictions.
AcknowledgementThle authors gratefuJly acknowledge support for this wvork by the Offbice of I Iealth
Miid Environmiental Research of the I. iited States [)epartnient oIf Energy tindercontract D E-.-\( '(I-76Rl. 1A 831), and by the I efense Nuclear Agency throughinteraigt-iicy transfer oif f~unds.
ReferencesA~(I ) Ii ( .\lC. .(* iiiwc I \1 I.A. J ., "M IN I WHI.. C. E'_ ai (IMl ius, Jr. L.. S., 1986o, Nettron-
784) 74.It114\\111 I W., iiid Ri i11 its, H. II.. 1974. Plrimaory prncess in the physical staize. P',vsica/
.1 Ic/raimoo iii Jaaiwiun, I//i'i, vditied h\ R . I ). Ciper anid W. V. WIioid Cl 'chliica Iitiltmriiiii Ceniter. L S.I:(*), pp. 201 41).
I CIii.(., S. M.,i~~ P.l'KI)NN . oP~i-i ., 3., anld Hiti ii., 95I iicrlivIiil tI('ti I il (M\ -Ite m ui iiii(les atii phas~e trariitioiis ot' L i- iiid Na-I I N. . ~'pdm o24, 2015 21(41.
04)141;11141 , S_. No)Iii I N(;, TI.1. arid S i m, A. \I.. 1985, PiteoIeciird tbuoresctie decayim e iisrceiiuii of ilt . cids at rimiii iemiiivraiire InI aqueous silti or.
I 'imhenisrv wnlidJih,/,,v 41, 2(19 212.1,61 .14 C III III;, V, 10 Hi-titi iilmd Sii~i .1971, lonuehaws railts irratitc~d 1)\.\ . Iiiii 0 ma v ci ioiuiia Attua. 254, 1 72 Isf'.
; i \, I -,1) A.1:ota \iti III,.HIi-i t llt Ai. inl Simm,~i G ., 1479. Uiv-uitucetl tree
radikl. mn lritiiictI [)\N.\ . I~imI'iIiand I',,iI,,,i'.29, 245 251.1,16,i 1 \11,i.loonI \iii lo;, A., RIHim initi,.~ Tj \.iui\, IW, miod Soioo~i. G;.. 1973, l:MR
k t i i l oI .0 rev r;Io ii a I vli-i -sifii III - i rranii t iic iud DIN. tidiimi Roetoi/c,61, OXN 5113.
(.it\-i I \i.Ii ini lIII,\. KoItNI I IN, W. inld IFi I i~ ~ . 91 ~htniiilmt(ret' radicAs III iiiit m ittud iiiailxiiiitt D NA. Ii'atito'.t Ii'cca,-ch, 88, 1 Ml.
;I It()\. W~, I $1i.Ii. . ;1i1d A. V.~i . 19)74, F\i imm sialte tit miitlic acids. Iivie
Amm ,iuiit Pic.p. pp, 31135

Rtipid copitinhitclitm <()7
I F, .0., Mai o:, W. I I., C..osr., ).l.., and S"A(;STI FN, E., 1987, ES Rand E N I)O studyit tic cotialne cation: secondary product in 5'-dC I P. J. .i11l/1if ( 'enicalPysics, 86,
I li ut iRt \N ,.. ', ur, K., 01 or., I I., K ri.-I, V., (;,;tmI), A. arid RHtttt ('tt, A.,I i4, S pcctific formation of'l cctron ga iIt and loss centres in X-irradtatcd Ioritnted tibvrs.
of 1 )NA at lo\ temperatures. Farair - Disctssiotu ( "m'/l',c+; Swoietiv, 78, 135 141).lit l'llft' i. A., I)66, Preparation (f oriented I)NA by \%et spinning. At ( ht,.owa
Samhditi,'., 20, 4)4 504.t'.mt. II., omitw.\;., Y., and Ktinor\, K.. 193, Epcrinicntal c~idncc itf collectivc
\ lratitons in I) )NA doihi, heli x (RIanman spe~ct rtmsct py ). Jourtmtl .4[( h']moa'I lHl ysims,, 78,
5t)37 5)39.
Vil ,4), W. '., and P.ARTM.T I. G., I( I1. Calculatin )f distributio)ns f etnerg y importedand ionization by fast prottms in nanometer sites. Raditiie Rust.trcl,. 87, 521 537.

Reprint Aao FoO Io- osMA " __
Publishers: S. Karger. Basel 9NNTIFI ? Ao"Pnnted in Swit,erland SR88-26
Bonavida, Gifford. Kirchner, Old (cds.), Tumor Necrosis Factor/Cachectin andRelated Cytokines. Int. Conf. Tumor Necrosis Factor and Related Cytotoxins, Heidelberg 1987,pp. 246-251 (Karger, Basel 1988)
In vivo Effects and Interactions of Recombi;,antInterleukin 1 and Tumor Necrosis Factor inRadioprotection and in Induction of Fibrinogen
Ruth Neta
Armed Forces Radiobiology Research Institute, Bethesda, Md.. USA
Although interleukin I (IL-I) and tumor necrosis factor (TNF) areboth produced by stimulated macrophage-monocytes, they are molecularlydistinct, act via separate receptors, but show striking resemblance in theirbiological activity. Both cytokines are pyrogenic [I], induce colony stimu-lating factor [2] and acute phase proteins [3], activate neutrophils [4, 5],reduce cytochrome P-450 functions [6, 7], and inhibit lipoprotein lipase [8,9]. Furthermore, IL- I and TNF have been reported to induce the release ofone another [1, 10]. Because of this mutual induction, the relative contri-bution of IL- I or TNF to the induction of a given activity becomes difficultto establish. In an attempt to determine whether these two cytokines actindependently, we have compared the effect of administrating them sepa-rately or in combination on radioprotection and on the induction of anacute phase reactant - fibrinogen. We now report that IL-I and TNF havesynergistic effects on radioprotection and on the levels of circulatingfibrinogen.
Materials and Methods
Mice. B6DZF inbred mice were obtained from Jackson Laboratories, Bar Harbor,Me. The mice were housed in the Veterinary Department Facility at the Armed ForcesRadiobiology Research Institute in cages, with Micro-Isolation unit tops, 10 mice/cage.Female mice, 8-12 weeks of age, were used for all experiments. Standard Lab chow andHCI acidified water (pH 2.4) were given ad libitum. All cage-cleaning procedures andinjections were carried out in a laminar flow unit.

Synergy of IL-A and TNF in Radioprotection 247
Cytokines. Human recombinant lL-la was generously provided by Immunex and byHoffmann-La Roche. The preparations were supplied in PBS pH 7.2 and 30 mMV Tris-HCI, 400 m.t NaCI, pH 7.8, respectively, and used on weight basis. Human recombinantTNFa, lot No. CP4026PO8, specific activity 9.6 X 106 units/mg in PBS was a generousgift from Biogen. All reagents were diluted to the desired concentration in 0.5 ml pyrogen-free saline just prior to the single intraperitoneal injection of mice, 20 h before irradiation.All cytokine preparations were assayed for LPS contamination in LAL assay and deter-mined to contain less than 0. 1 ng/inoculum.
Irradiation. Mice were placed in plexiglass containers and were given whole bodyirradiation at 40 rd/min by bilaterally positioned b"Co elements. The number of survivingmice was recorded daily for 30 days.
Fibrinogen Assa i. Mice were bled (retroorbitally) at 20 h after injection of cyto-kines and the plasma was collected and stored at -20"C. Assays for fibrinogen indiluted citrated plasma were performed by measuring the rate of conversion of fibrino-gen to fibrin in the presence of thrombin excess. The calibration was made using theSigma Diagnostic Kit. Measurements of fibrin clot formation were performed on afibrometer (Becton-Dickinson). The data are expressed as milligram fibrinogen per100 ml plasma.
Results
Radioprotection with hriL-la and hrTNFaWe have compared the effect of increasing doses of hrIL-la and of
hrTNFct on protection of lethally irradiated B6D2F, mice (LD 95/30 - 1,050rad). IL- la in doses ranging from 75 to 1,000 ng protected 80-85% of micefrom death. Although equivalent doses of hrTNFa had no radioprotectiveeffect, significant radioprotection was obtained with doses ranging from 5to 10 jig hrTNFa. The maximal degree of radioprotection achieved withthe optimal dose of hrTNFa (40-50%) was significantly less than thatobserved with hrIL-Ia. Therefore, hrTNFa is a less effective radioprotec-tor in mice than hrIL-la. The combination of hrIL-la and hrTNFaresulted in a synergistic radioprotective effect, at supralethal doses ofradiation, with a greater surviving number of mice than predicted from thesum of radioprotection obtained with each cytokine alone (fig. 1). Asupralethal dose of 1, 150 rad, rather than the usual dose of 1,050 rad, wasused in this experiment in order to reduce the radioprotective effects ofIL-I and TNF by themselves, and to reveal interactions between these twocytokines.
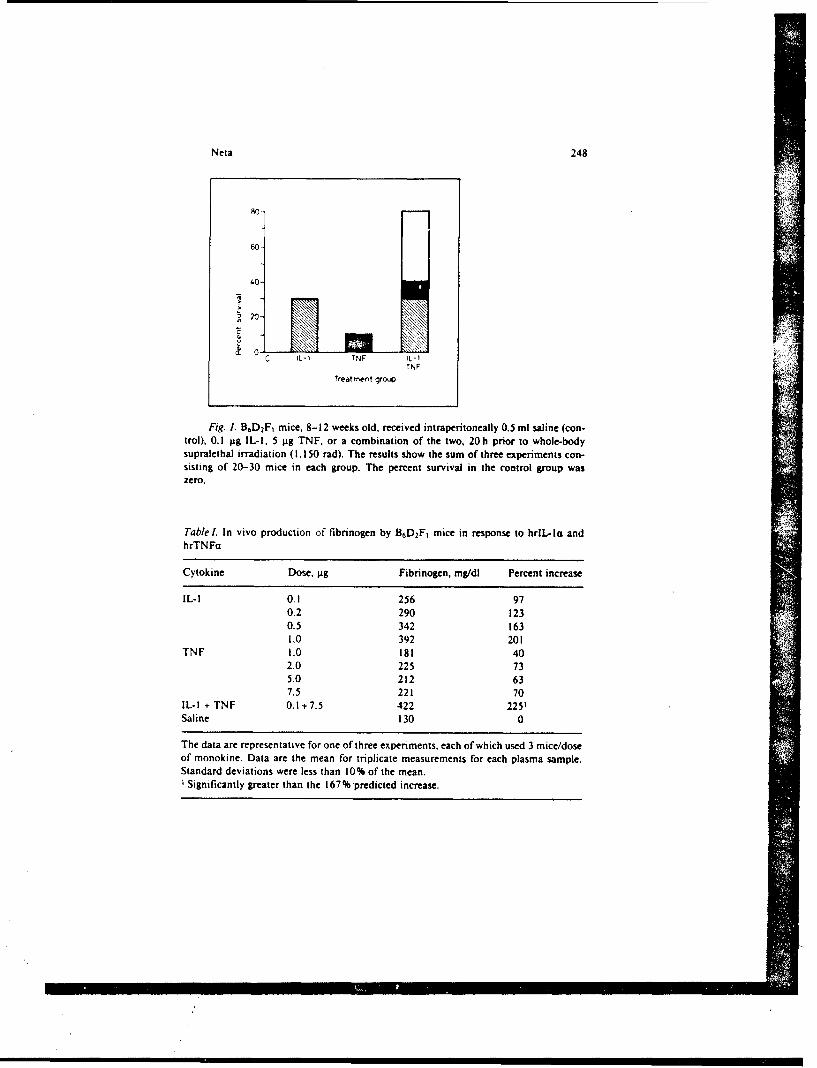
Neta 248
80-
60-
40-
~20 -
IL-i TNF IL-ITNF
Treatment group
Fig. 1. B.D 2F, mice, 8-12 weeks old, received intraperitoneally 0.5 ml saline (con-trol), 0.1 g±g IL-I, 5 pg TNF, or a combination of the two, 20h prior to whole-bodysupralethal irradiation (1.150 tad). The results show the sum of three experiments con-sisting of 20-30 mice in each group. The percent survival in the control group waszero.
Tablel. In vivo production of fibrinogen by B6D2F, mice in response to hrIL-la andhrTNFa
Cytokine Dose, p±g Fibrinogen, mg/dI Percent increase
IL-I 0.1 256 970.2 290 1230.5 342 1631.0 392 201
TNF 1.0 181 402.0 225 735.0 212 637.5 221 70
IL-I + TNF 0.1+7.5 422 225'Saline 130 0
The data are representative for one of three experiments, each of which used 3 mice/doseof monokine. Data are the mean for triplicate measurements for each plasma sample.Standard deviations were less than 10% of the mean.
Significantly greater than the 167% predicted increase.

Synergy of IL-I and TNF in Radioprotection 249
Induction of Fibrinogen with hrIL- Ia and/or hrTNFaThe levels of fibrinogen in the plasma of B6D2F1 mice 20 h following
administration of the two cytokines were also determined. The results intable I show that on a weight basis hrlL-la induces higher levels of fibrin-ogen than hrTNFa. Furthermore, an optimal dose of hrIL-la induces amuch higher increase in fibrinogen than the optimal dose of hrTNFa.Administration of hrlL-la and hrTNFa in combination resulted in asynergistic increase in production of fibrinogen.
Discussion
The overlapping biological effects and mutual induction of hrlL-laand hrTNFa make it difficult to attribute a given activity to one or theother of these two cytokines. Although our results do not establish theprecise contribution of each cytokine to radioprotection and fibrinogenproduction, the evidence suggests they have independent effects.
The requirement for much higher doses of hrTNFa than hrIL- I a can-not be attributed to lower cross-species activities of TNFa, since we alsorequire similarly higher doses of murine TNFa to obtain radioprotection[unpubl. observations]. Furthermore, the suggestion that the effect of TNFis mediated by IL-I is unlikely since the synergy of optimal doses of hrlL-lIa and hrTNFa in protecting mice against lethal doses of radiation sug-gests that the effect of these two cytokines is based on different pathways.Although the mechanism of action to achieve radioprotection remainsunknown, a number of the activities of IL-la and TNFa may be related tothe radioprotective effect. For example, induction of acute phase proteins,some of which (metallothionein and ceruloplasmin) with the capacity toscavenge free radicals, may contribute to radioprotection [I1, 12]. Al-though IL-la induction of bone marrow cell cycling [ 131 may present yetanother critical event in radioprotection, TNFa is not known to have thiscapability. In fact, TNFa has been reported to be inhibitory to hemato-poiesis [14, 15]. Several reports exist, however, that TNF has a role inhematopoietic differentiation [ 16, 17]. This differentiating effect of TNF ismost pronounced when TNF is used in conjunction with other cytokines.Whether this effect of TNFct on hematopoietic cells contributes to itsradioprotective effect remains to be established.
Elevated plasma levels of fibrinogen have been shown previously to bea consequence of stimulation of hepatocytes [181. The finding that com-

Neta 250
bined administration of IL-I and TNF results in more than an additive.."effect on fibrinogen production, again suggests that the combination of the
two signals (IL-I and TNF) induces distinct rather than overlappingresponses by hepatocytes.
IL-I and TNF are cytokines prominent in inflammation. As such theyhave a dual role of contributing to host defense as well as being involved inthe process of healing and repair. Unfortunately, their conjoint activities,if pronounced, may also be deleterious to the host. The effects of these twocytokines may be regulated and therapeutically useful after sufficientknowledge is acquired concerning their actions and interactions.
Acknowledgements
I wish to thank Dr. J.J. Oppenheim for critical review of the manuscript and SusanD. Douches for technical assistance. This work was supported by the Armed ForcesRadiobiology Research Institute, Defense Nuclear Agency, under research work unitMJB3148. The opinions or assertions contained herein are the private views of theauthor, no endorsement by the Defense Nuclear Agency has been given or should beinfeired. The research was conducted according to principles enunciated in the 'Guide forthe Care and Use of Laboratory Animals' prepared by the Institute of Laboratory AnimalResources, National Research Council.
References
I Dinarello, C.A.; Cannon, J.G.; Wolff, S.M.; Bernheim, H.A.; Beutler, B.; Cerami, A.;Figari, i.S.; Palladino, M.A.; O'Conner, J.V.: Tumor necrosis factor (cachectin) is anendogenous pyrogen and induces production of interleukin-!. J. exp. Med. 163:1433-1449 (1986).
2 Vogel, S.N.; Douches, S.D.; Kaufman, E.N.; Neta, R.: Induction of colony stimulat-ing factor in vivo by recombinant interleukin-la and recombinant tumor necrosisfactor-a. J. Immun. 138: 2143-2148 (1987).
3 Sipe, J.D.; Vogel, S.N.; Douches, S.D.; Neta, R.: Tumor necrosis factor/cachectin isa less potent inducer of serum amyloid A synthesis than interleukin-l. LymphokineRes. 6:93-101 (1987).
4 Klempner, M.S.: Dinarello, C.A.; Galin, J.I.: Human leukocytic pyrogen inducesrelease of specific granule contents from human neutrophils. J. din. Invest. 61:1330-1336 (1978).
5 Tsujimoto, M.; Yokota, S.; Vilcek, J.; Weissman, G.: Tumor necrosis factor pro-vokes superoxide anion generation from neutrophils. Biochem. biophys. Res. Com-mun. 137: 1094-1100 (1986).
6 Ghezzi. P.; Saccardo, B.; Bianchi, M.: Recombinant tumor necrosis factor depressescytochrome P-450-dependent microsomal drug metabolism in mice. Biochem. bio-phys. Res. Commun. 136: 316-321 (1986).
/

Synergy of IL-I and TNF in Radioprotection 251
7 Ghezzi, P.; Saccardo, B.; Villa, P.; Rossi, V.; Bianchi, M.; Dinarello, C.A.: Role ofinterleukin-l in the depression of liver drug metabolism by endotoxin. Infect.Immunity 54: 837-840 (1986).
8 Kawakami, M.; Cerami, A.: Studies of endotoxin-induced decrease in lipoproteinlipase activity. J. exp. Med. 154: 1631 (1981).
9 Beutler, B.A.; Cerami, A.: Recombinant interleukin-I suppresses lipoprotein lipaseactivity in 3T3-L' cells. J. Immun. 135: 3969-3971 (1985).
10 Philip, R.; Epstein, L.B.: Tumor necrosis factor as immunomodulator and mediatorof monocyte cytotoxicity induced by itself, gamma-interferon, and interleukin-l.Nature, Lond. 323. 86-89 (1986).
I1 Goldstein, I.M.; Charo, I.F.: Ceruloplasmin: an acute phase reactant and antioxi-dant; in Pick, Lymphokines, vol. 8, pp. 373-411 (Academic Press, New York1983).
12 Karin, M.: Metallothioneins: proteins in search of function. Cell 41: 9-I1 (1985).13 Neta, R.; Sztein, M.B.; Oppenheim, J.J.; Gillis, S.; Douches, S.D.: In vivo effects of
IL-I. I. Bone marrow cells are induced to cycle following administration of IL-I.Immun. 139: 1861-1866 (1987).
14 Peetre, C.; Gullberg. U.; Nilsson, E.; OIsson, I.: Effects of recombinant tumor necro-sis factor on proliferation and differentiation of leukemic and normal hemopoieticcells in vitro. J. clin. Invest. 78: 1694-1700 (1986).
15 Murase, T.; Hotta, T.; Saito, H.; Ohno, R.: Effect of recombinant human tumornecrosis factor on the colony growth of human leukemia progenitor cells and norma:hematopoietic progenitor cells. Blood 69: 467-472 (1987).
16 Trinchieri, G.; Kobayashi, M.; Rosen, M.; Loudon, R.; Murphy, M.; Perussia, 3.:Tumor necrosis factor and lymphotoxin induce differentiation of human myeloidcell lines in synergy with immune interferon. J. exp. Med. 164: 1206-1225(1986).
17 Munker, R.; Koeffler, P.: In vitro action of tumor necrosis factor on myeloid leu-kemic cells. Blood 69: 1102-1108 (1987).
18 Pepys, M.B.; Balz, M.L: Acute phase proteins with special reference to C-reactiveprotein and related proteins (pentaxins) and serum amyloid A protein. Adv. Immu-nol. 34: 141-212 (1983).
Ruth Neta, PhD, Department of Experimental Hematology,Armed Forces Radiobiology Research Institute, Bethesda, MD 20814 (USA)

AMMO "MIS1 RLOMOUM
SR88-27CONCISE REPORT
Cytokines in Therapy of Radiation Injury
By Ruth Neta and J.J. Oppenheim
Repeated injections or infusion of hematopoistic growth potential therspeutic agents in irradiated C3H-HeN mice. Afactors, such as interleukin-3 (IL-3). granulocyte macro- single intrapertoneal injection, administered within threephage-colony stimulating factor (GM-CSF). or granulocyte- hours after a lethal dose (LD)141o of irradiation that wouldcolony stimulating factor (G-CSF), accelerate restoration kill 95% of mice within 30 days, protected in a dose-of hematopoiesis in animals compromised by sublethal dependent manner up to 100% of mice from radiation-doses of cytotoxic drugs or irradiation. Previuus work by induced death due to hematopoietic syndrome. Significantthe investigators has shown that IL-1 induced circulating therapeutic effects were also achieved with a single doseCSF in normal mice and. when used after sublethal irradia- of IFN-,y or of TNF. In contrast. GM-CSF and G-CSF.tion. accelerated the recovery of endogenous splenic col- administered shortly after irradiation, had no effect in thenies. Therefore. IL-1. as well as IFN-y,. tumor necrosis doses used on mice survival.factor (TNF). G-CSF. and GM-CSF. were evaluated as e 1M98byGnw & Stratton, Inc.
A S ATTESTED BY the experience with recent nuclear hardwood chip contact bedding and provided with commercialaccidents, there is no effective treatment for patients rodent chow and acidified tap water (HCI to a pH of 2.5) ad libitum.
exposed to doses of radiation that result in fatal hemato- Animal holdingrooms were maintained at 70 ± 20F with 5O% t 10
poietic failure and/or secondary infections. Clinical difficul- relative humidity using at least ten air changes per hour of 100%
tics due to HLA mismatching, such as graft v host (GVH) conditioned fresh air. The mice were on a 12-hour light-dark full
reactions and graft rejection, minimize the successful use of spectrum lighting cycle with no twilight. Mice were 8 to 10 weeks ofage when used. All cage cleaning, handling, and injections werebone marrow transplantation (BMT) in these situations. carried out in a laminar flow clean air unit.
Therefore, agents that promote repair of bone marrow Cytokines. Human recombinant IL-la wasgenerously provideddamage and improve the recovery of the surviving fraction of by Immunex (Seattle) and by Hoffman-La Roche (Nutley. NJ).cells must be identified. The preparations were supplied in phosphate buffered saline (PBS)
The investigators have reported previously that the cyto- at pH 7.2 or in 30 mmol/L tris-HCI. 400 mmol/L NaCI, pH 7.8.kincs interleukin- I (I L- I ) and tumor necrosis factor (TN F), respectively, and used on a weiglht basis. Human recombinant TNFbut not IL-2, interferon (IFN)--y. or granulocyte macro- a. lot number CP4026PO8. specific activity 9.6 x 10'U/mginPBSphage-colony stimulating factor (GM-CSF), when adminis- was a generous gift from Biogen (Cambridge. MA). Murine recom-tared before lethal irradiation, protect mice from death binant GM-CSF was provided by Immunex as a lyophilized powdertred g bfoe lthal irraatio yndproe mic ehe with sucrose as a stabilizing agent. Human recombinant G-CSF,specific activity 9 x 10' U/mg was a gift from Amgen (Thousanduse of I1.-I following !ctha| irradiation (lethal dose Oaks, CA). Murine recombinant IFN--y, lot 4296, specific activity[LI)] ,,/) did not affect survival.' It has been reported that 6.8 x 10 U/mg was a gift from Genentech (San Francisco). Allthe hcmatopoictic growth factors, IL-3, GM-CSF, G-CSF, reagents were diluted to the desired concentration in pyrogen-frecand CSF-I, accelerate restoration of hematopoiesis in ani- saline just before the single intraperitoneal injection of 0.5 mL tomals compromised by suble'hal doses of radiation or by mice. one to three hours after irradiation. All cytokine preparationscytotoxic drugs.'" IL-I used after irradiation was also e'ec- were assayed for lipopolysaccharide (LPS) contamination in ative in accelerating hematopoietic recovery in sublethally(700 c(iy) irradiated mice.'0 Furthermore, IL-I and TNFboth induce the appearance of high titers of CSFs in thecirculation." The effect of administering a single intraperito- From the Department of Experimental Hematology. Armed
Forces Radiobialogy Research Institute. Bethesda: and Laboratoryneal dose of IL-I, TNF, GM-CSF, or G-CSF shortly after of Molecular Immunoregulation. National Cancer Institute. Fred-irradiation of mice has been examined. In addition, IFN-y erick. MD.was studied to determine if its antiproliferative effect renders Submitted April 5. 1988: accepted Mat, 25. 1988.it ineffective as a restorative agent. Supported h' the Armed Forces Radiobiologr Research Insti-
The investigators present data which show that a single tute. Defense Nuclear Agen., under research work unit MJB3148injection of II.-I, in a dose-dependent manner, promotes The opinions or assertions contained herein are the private views ofsurvival of C311/leN mice from a radiation dose that the autho . mi endorsement hV the Delense Nuclear Agency has beenresults in death of 95'; of control animals within 30 days given or should he injerred. The research was conducted according(II), 5:). IFN-"- had a similar effect, TNF also showed to principles enunciated in the Guide for the Care and Use oflimited therapeutic elicacy. while G-CSF and G M-CSF Laboratory Animals prepared bt, the Institute of Laboratory Ani-were not effective in promoting survival. mal Resources. National Research Council.,4ddres. reprint requests to Ruth Neta. PhD. Defense Nuclear
MATERIALS AND METHODS Agency Armed Forces Radiobiology Research Institute. Bethesda.R MD 20814.
.W(e. A total of 450 C311/lieN mice were purchased from The puhlicatior costs ojthis article were defrated in part hi "pageAnimal Genetics and Production Branch. NC I (Frederick, M D), fir charge par'ment. This article must therefore be herebt markeduse in these experiments. Mice were quarantined on arrival and ":advertiscment'inacccrdance with 8 .S.C. ectio. 1734sole toscreened for evidence of disease before being released from quaran- indicate this fact.tone They were maintained in an AAAI.AC accredited facility in 1 1988 by'(vrune A Stratton. Inc.plastic Micro-isolator cages (Lab Products, Mavwcxjd. NY) on O(X)6-4971/88/7203-OY)48S3.(O)
Blood. Vol 72. No 3 (Septembert. 1988: pp 1093-1095 1093
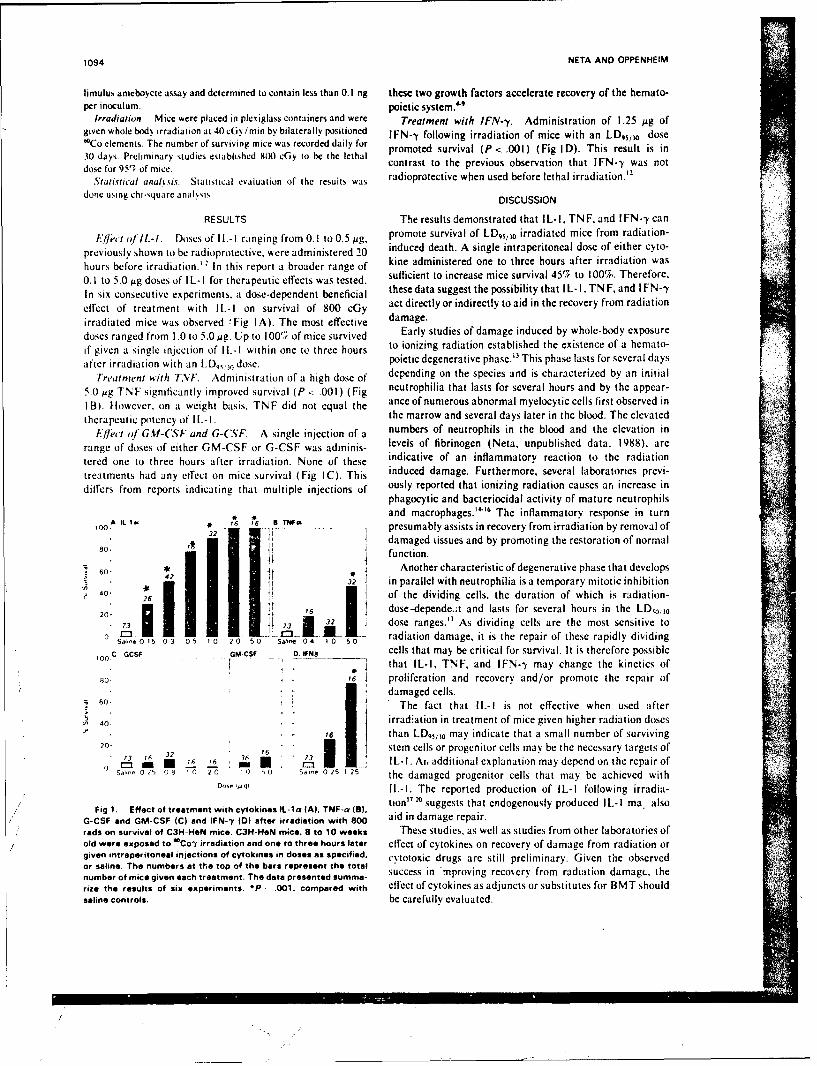
1094 NETA AND OPPENHEIM
limulus ameboycte assay and determined to contain less than 0.1 ng these two growth factors accelerate recovery of the hemato-per inoculum. poictic system. "
Irradiation. Mice were placed in plexiglass containers and were Treatment with IFN--y. Administration of 1.25 pg ofgiven whole bod, irradiation at 40 c(;v/min by bilaterally positioned IFN-y following irradiation of mice with an LDqsj dose'Co elements. The number of surviving mice was recorded daily for promoted survival (P < .001) (Fig I D). This result is in30 days. Preliminary studies established 800 cGy to be the lethal contrast to the previous observation that IFN--y was notdose for 95'T of mice. cort t the us bereaintha Iadi wason ot
Statistical analyvis. Statistical evaluation of the results was radioprotective when used before lethal irradiation. 2
done using chi-square anal.sis. DISCUSSION
RESULTS The results demonstrated that IL-I. TN F, and I FN--y canEileet of IL-I. Doses of I1.-I ranging from 0.1 to 0.5 tag. promote survival of LD,5 /30 irradiated mice from radiation-
previously shown to be radioprotective, were administered 20 induced death. A single intraperitoneal dose of either cyto-
hours before irradiation.' In this report a broader range of kine administered one to three hours after irradiation was
0. 1 to 5.0 pg doses of IL- I for therapeutic effects was tested. sufficient to increase mice survival 45% to 100%. Therefore,
In six consecutive experiments, a dose-dependent beneficial these data suggest the possibilitythat IL-I,TNF,and IFN--y
effect of treatment with IL-I on survival of 800 cGy act directly or indirectly to aid in the recovery from radiation
irradiated mice was observed 'Fig IA). The most effective damage.
doses ranged from 1.0 to 5.0 pg. Up to 1005, of mice survived Early studies of damage induced by whole-body exposure
if given a single injection of IL-I within one to three hours to ionizing radiation established the existence of a hemato-
after irradiation with an -D57, dose. poietic degenerative phase." This phase lasts for several days
Treatment with TNF. Administration of a high dose of depending on the species and is characterized by an initial
5.0 pg TNF significantly improved survival (P < .001 ) (Fig neutrophilia that lasts for several hours and by the appear-
I B). However, on a weight basis. TN F did not equal the ance of numerous abnormal myelocytic cells first observed in
therapeutic piotency of Ii.-I. the marrow and several days later in the blood. The elevated
Elject of GM-(SF and G-(S*F. A single injection of a numbers of neutrophils in the blood and the elevation in
range of doses of either GM-CSF or G-CSF was adininis- levels of fibrinogen (Neta, unpublished data, 1988). are
tered one to three hours after irradiation. None of these indicative of an inflammatory reaction to the radiation
treatments had any effect on mice survival (Fig IC). This induced damage. Furthermore, several laboratories previ-
dilfers from reports indicating that multiple injections of ously reported that ionizing radiation causes an increase inphagocytic and bacteriocidal activity of mature neutrophils
32 IL 1 -and macrophages."" The inflammatory response in turn,lo. * 2 6 i6 atwo presumably assists in recovery from irradiation by removal of
4 damaged tissues and by promoting the restoration of normal80 i
60. iifunction.
60. * Another characteristic of degenerative phase that develops3 in parallel with neutrophilia is a temporary mitotic inhibition
, 6 of the dividing cells, the duration of which is radiation-
2o. I i 32 dose-dependent and lasts for several hours in the LD,0o10
20 73 3 dose ranges.'" As dividing cells are the most sensitive toS 0 5 0 0. .2. radiation damage, it is the repair of these rapidly dividingS.1-e 0 15 0 3 0 5 1 0 2 0 5 0 Sathe 04, 1 0 5 0
100 C
GCSF GMCSF . "WN8 cells that may be critical for survival. It is therefore possible'00 that IL-I, TNF, and IFN--y may change the kinetics of
5o i6 proliferation and recovery and/or promote the repair of[] damaged cells.
60 The fact that IL-I is not effective when used after
40. irradiation in treatment of mice given higher radiation doses• Y6' than LD,5/3o may indicate that a small number of surviving
20. 6 "1 stem cells or progenitor cells may be the necessary targets of73 16 6 , ,'o , ,L , I IL-I. At, additional explanation may depend on the repair of
SM,, 0 2 06 1 20 0 0 S,,,e 0 25 1 25 the damaged progenitor cells that may be achieved with0n,, M'~ IL-I. The reported production of IL-I following irradia-
,, tion" 2° suggests that endogenously produced IL-I ma alsoFig 1. Effect of treatment with cytokines IL-Ia (A). TNF-a (),
G-CSF and GM-CSF (C) and IFN--y (D) after irradiation with 800 aid in damage repair.rads on survival of C3H-HeN mice. C3H-HON mice. a to 10 weeks These studies, as well as studies from other laboratories ofold were exposed to *Con' irradiation and one to three hours later effect of cytokines on recovery of damage from radiation orgiven intraperitoneal injections of cytokines in doses as specified, cytotoxic drugs are still preliminary. Given the observedor saline. The numbers at the top of the bars represent the totalnumber of mice given each treatment. The data presented summa-rize the results of six experiments. *P .001. compared with effect of cytokines as adjuncts or substitutes for BMT shouldsaline controls. be carefully evaluated.

CYTOKINES IN THERAPY OF RADIATION INJURY 1095
ACKNOWLEDGMENT recovery of the monkeys transplanted with autologous bone marrow.
We wish to thank Drs M.L. Patchen and T.J. MacVittie for Blood 70:1996, 1987
critical review of this manuscript. W.E. Jackson for statistical 10. Net R. Oppenheim JJ. Douches SD. Giclas PC, Imbra RI,analysis of the results, and S.D. Douches and M. White for technical Karin M: Radioprotection with Interleukin-1: Comparison withassis tance, other cytokines, in Cinadr B. Miller RG (eds): Progress in Immu-nology. San Diego, Academic Press, vol 6. 1986, p 900
REFERENCES 1I. Vogel SN, Douches SD, Kaufman EN, Neta R: Induction of
I. Neta R. Douches SD. Oppenheim JJ: laterleukin-I is a colony stimulating factor in vivo by recombinant Interleukin-I andradioprotector. J Immunol 136:2483. 1986 tumor necrosis factor. J Immunol 138:2143, 1987
2 Neta R. Oppenheim ii. Douches SD. Interdependence of the 12. Neta R, Vogel SN, Oppenheim JJ, Douches S!): Cytokines inradioprotective effects of huynan recombinant Interleukin- la. tumor radioprotection. Comparison of the radioprotectiv effects of IL- I to
IL-2, GM-CSF. and IFN-gamma. Lymphokine Res 5:105-110.necrosis factor , granulocyte colony stimulating factor and murine 1986recombinant granulocyte-macrophage coluny stimulating factor. J (suppl)Immunol 140:108. 1988 13. Bond VP, Flicdner TM, Archambeau JO: Mammalian3. Neta R. Douches SD, Oppenheim JJ Radioprotection by Radiation Lethality. A Disturbance in Cellular Kinetics. San Diego,etR.DuhsS.ppnemi Rairtetnby Academic Press. 1965 :
inlrluki-l i Godsei GBah i. igal (ds: Imuo- 14. Mukherjee AK. Starra A J: The role of the phagocyte in rinterleukin-1, in Goldstein G, Bach JF, Wigzal H (eds): Immuno- AcdmcPes15regulation by Characterized Polypptides. UCLA Symposia on 14 uhjeAKStra J:Teolofhepgcyeireulaon by Chlarcteried Piolog eptidr. LA Symposa o9 host-parasite interactions. XIV. Effects of concurrent X-irradiationMolecular and Cellular Biology. New York. Liss, 1986. pp 4294. Sieff CA: Hematopoietic growth factors. J Clin Invest on phagocytosis. J Reticuloendothelial Soc 5:134. 1968
79:1549, 1987 15. Geiger B, Gallily R: Effect of X-irradiation on various5. Groopman JE: Hematopoitic growth factors: From methyl functions of murine macrophages. Clin Exp Immunol 16:643, 1974cellulose to man. Cell 50:5 1987 16. Meyer OT, Dannenberg AM Jr: Radiation, infection, and
6. Cohen AM. Zsebo KM. Inou H. Hines D, BooneTC. Chazin macrophage function. II. Effect of whole body radiation on thevivo stimulation of granulopoic- number of pulronary alveolar macrophages and their levels of
sis by recombinant human granulocyte colony stimulating factor hydrolytic enzymes. J Reticuloendothelial Soc 7:79, 1970si bail recbant huma grAnu y coly s g f17. Geiger B, Fality R, Gery I: The effect of irradiation on theProc: Nail AXcad Sci USA 84:2484. 19877. Shimanura M, Kobayashi Y, Yno A, Urabe A. Okabe T, release of lymphocyte activating factor (LAF). Cell Immunol 7:177.7. SimauraM. obaash Y. no . UabeA. kab T. 1973
Komatsu Y. Itoh S, Takaku F: Effect of human recombinant
granulocyte colony stimulating factor on hematopoictic injury in 18. Granstein RD, Sander DN: Whole body exposure to ultravio-let radiation results in increased serum Intcrleukin-I activity inm ice induced by 5-fluoro uracil. Blood 69:353. 1987 h m n .L m h k n e : 8 ,1 8
8. Nienhuis AW. Donahue RE, Karlsson S, Clark SC, Agricola humans. Lymphokine Res 6:187. 1987B. Antinoff N, Pierce JE, Turner P. Anderson %WF, Nathan DG: 19. Ansel JC. Luger TA, Green 1: The effect of in vitro and inRecombinant human GM-CSF shortens the period of neutropenia vivo UV irradiation on the production of ETAF by human andafter autologous bone marrow transplantation in a primate model. J murine Keratenocytes. J Invest Dermatol 81:519, 1983Clin Invest 80:573. 1987 20. Ansel JC, Luger TA. Green I: Fever and increased serum
9. Monroy RL. Skelly RR. MacVittie TJ, Davis TA, Sauber JJ, IL-I activity as a systemic manifestation of acute phototoxicity inClark SC, Donahue RE: The effect of recombinant GM-CSFon the New Zealand white rabbits. J Invest Dcrmatol 89:32. 1987
II.

Brain Res earch. 4.% (1488 1 4Y - 5 h IN 96ANI 11111111u 49H ses icr S1 HM111111 4010
Effects of dithiothreitol, a sulfhydryl reducing agent, on CA,pyramidal cells of the guinea pig hippocamnpus in vitro
J. M. Tolliver* and T.C. PelimarPI %.il 'Ii ,eipiiryneil. A1 rond force% Rathohwhpgt, Re'searchc himiitue. Bethesda. MD1I 2(h14-5 /45 W .8 A.
1 Accepted 9 Fehruarv 1988$J
Wet D,~stithisithreitil. I lippcicarrpu%: Radioprotectant. Intracellular recording
I he ridioprotcctant . dithiothrectol (1) 1-1) has been shcissn to increase excitability in the hippocampal slice preparation. In the pres-cmt tuid%. intraccelular reciording tecchnique% %%ere used it) further examine the actions of DIT. Elect rophysiologicalI recordingts tromCA, p% rarnidal cells %%ere obtained prior ito. during and after D-l exposure. DTr caused a small depolarization withcut altering mcm-brrie resistance 1)1-1 induced spointaneous tiring and occasional hurst firine in normalIv silenlt neurons. These effects were accompa-nied h% a reductioin in spike lrcqucnc adaptation but no change in the alterhyperpoflarization following it train of act ion p )tent als.F-ollossilL 1) 1-1 exposure. irthodromic stimulation produced multiple firing. Subthreshold excitatory postssnaptlc potentials f [PSPsI%~ere sieniticantl% priilonged. Isola ting the (A, subfield, attenuated the prolongation of the EPSP by DTT. Recurrent inhibitory post-s naptic poitentials \%ere unattcted h% Dl . IThe actions of DTT are likely to result from DTT-induced reduction of disulfide bondsstice the reduced fiorm cit Dl I dies not cause a similar h\ percxcitability.
IN!I R( Ml Il ION agents in the hippocampal slice (for review see ref.6). A previous study showed that DTT increases hip-
The hippocampus is vulnerable to radiation dam- pocampal excitability. causing spontaneous anda~'e. FollowinL, exposure to X-radiation. spiking ac- evoked burst firing"'. Both sv'naptic and extrasvnap-itit% appears in elect roenrcephalographic recordings tic mechanisms were implicated. The present studyfrom this area of the brain'-' . Single unit record- uses intracellular recording techniques to examinenws in % i\i oshow% that neuronal firing patterns in the the mechanism(s) whereby DTT increases neuronalhippocampus are disrupted by radiation2. In vitro excitability. Prelim~nary results have been presentedstudtes tndtcate that :,-radtatton can tmpatr both syn- elsewhereaptic and extras\ naptic mechanisms in the hippocam-pus't. MATERIALS AND METHODS
Dithiothreitol (DT-T). at disulfide reducing agent'has been used as a radioprotectant in cellular and en- H-ippecampal slices (4(Xl-45(l om thick) were pre-tvme s\stemst 5)) It is thought to act both by pared from euthanized male Hartley guinea pies assea\,enging free radicals"'- and by donating hv- previously describe& ail.. Slices were incubated atdrogienl to damaged macromolecules 1; Many studies room temperature in oxygenated solution (see be-have demonstrated that DT-T affects neurotransmit- low) for at least 2 h to allow recovery from the dissec-ter systems includince tchinII> dopa- tion. One slice was then placed in a submerged slicemine' . opiates. norepinephrine"~ and his- chamber (Zbicz design)"'. The tissue was continu-tamtine'"1 Aill these transmitters are cffective ously superfused (0.8-1I.0) ml min) wih a solution
Present aiddress: tDrul! Entiirc.,ment Administratiion, Office of Disersion Control. Washington. D).C. 2ti537. U.S.A.(vrre'spndi'nie 1. Pellmar. Phsiiiliicy t0epartrnent. Armed Fiirces Radiobiology Research Institute. Bethesda. NI) 20814-5 145.
I*,S A\

54)
contlainintg (in nIl): Na('l 24 KCI 3.0, ('a'l, 2.4. rent. To study spike frequency adaptation. supra-MgS() 1.3. KIItIPO, 1.24. NaICO 26.0 and glu- threshold depolarizing current pulses of 7( ms dura-cOwe 10.01, ox~g.nated with 9)5' 05 C(), and tion were applied via the recording electrode at 0.05-maintained at 3o)' _ I ('. (. 1 Hz. An AHP was elicited every 10-20 s by inject-
I)TT. obtained ron ('albiochem (Lot numbers ing a train of 4 suprathreshold depolarizing current23315 3 and 410(0). \kas dissol ed in the bathing so- pulses (8 ms duration. 80 Hz). EPSPs were evoked bylulion mniediatcl% prior to use to gis e a final concen- stimulation (11-75 YA) of the stratum radiatum andtration of 5 nNl. After obtaining control data. the recurrent IPSPs by stimulation (25- (NIA) of the al-
sliceN %%ere supertused % ith the DF solution for 25 veus. In several experiments the CA, region was iso-in. [h 1is dlose and exposure time were chosen be- lated. CA, and CA. were removed by cutting the
cause they consistently increased excitability in the slice with a scalpel blade. No attempt was made to.eld potential recordings. Subsequent to D'" expo- verify histologically that all of the CA, and CA, re-sure. slices were superfused with normal solution for gion was removed in these experiments. The slicethe remainder of the experiment. Diata were col- was allowed to recover from the isolation procedurelcctcd for a niiniuil of 644 rllin after the initial expo- for at least 45 min before attempting to impale CA,sure to D I-l, neurons.
('oncentric. bipolar stainless steel electrodes wereused ito proside constant-current stimuli (up to 0.5 RESULTSmA. 2(M .1s duration. (.2 II) to hippocampal path-
avs. Cells in the CA, region of hippocampus were As previously reported " . field potential record-orthodromicallv activated b\ stimulation of afferent ings in field CA, of hippocampus revealed an in-fibers in the stratum radiatum. Antidronic potentials crease in excitability following exposure to 0.5 mM%%ere elicited by stimulation of the alveus. DT for 25 min. Twenty to 40 min after initial expo-
Intracellular recordings from CA, pyramidal cells sure to DTT. the baseline 'noise' level increased in\were obtained through electrodes filled with either 2 the extracellular recordings from stratum radiatum.NI KC(I (20-40 N1Q) or 4 NI potassium acetate This was followed by an increase in the amplitude of(7(- l() NIO ). A conventional bridge circuit (l)agan the orthodromic population spike and the emergenceS 1IM) allowCd potential recording and intracellular of multiple (3-7) peaks in the orthodromic field po-current in ection \ iai the same electrode. The bridge tential (Fig. IA). This effect was sustained for the du-%% as balanced frequently by passing pulses of current ration (1-2 h)of the experiment.and monitoring the potential on the oscilloscope. [he increased excitability produced by DTT wasI)ata "\ere recorded on ia (ould chart recorder and a further examined in CA, pyramidal cells with intra-lcktronix oscilloscope, and were digitized and cellular recording techniques. Prior to drug expo-storCd oil an .SI 11-113 minicomputer. InI all experi- sure, ('A, pyramidal cells were generally silent-ments. somatic field potentials were monitored con- spontaneous action potentials were rare. Twenty tocurrcntly using electrodes ( I - 10 NI ) filled \with 2 NI 4) min following initial exposure to 0.5 mM DTT.Na(Tl. Held potlenti,s s'erc amlplifiCd using it high- spontaneous activity was always evident (Fig. IB).gain dtiffercntia preamplifier. I hey were either pho- This effect coincided with the appearance of *noise*tographed directl\ off the oscilloscope or digiti/ed, in the extracellular recording. In approximately 60"iMid stored on ti I.Sl 11 -03 Iiinicomnputer. oif the cells. spontaneous doublets and burst firing oc-
[he etects ot I)1-1 on the tollo\\ ing clcctroph\sio- curred in an irregular pattern. Similar spontaneouslogical paraileters \were examined: ricnibrane po- bursting act'itv was observed in field potentialtential. input resistance, spike frequency aidaplttion. recordines".aftterlixperpolari/ation (AlP1). Cxcitator postsvn- [he intracellular changes were accompanied by aaplic ptcnti'iis (IPSPs,) and inhibitor% post%naptic smiall. but statistically significant depolarization ofpotcntials (II'.s%1,). Mcnliranc resistance wa,s calcu- the membrane potential. The mean ( + S. E.M. ) rest-lated frot th ncnbrane po'teitial hllange produced ing potential of all 25 cells before exposure to D1FT
% in ictions ot 0(.25 or 01.501 nA li crpoiri/ing cur- was -64.0 t |I.X mV. Sixty minutes after initial expo-

CONTROL 60 MIN DTT
1 my
20 msec
B~ .3m 4 mV
V 6 Vtv V=-57 mV t 2 sac
C L 20 mL1
20nisec
-- Fit! I [)'I alte red the orthodromie activity in CA, of the. hippocampal slice. A: orthodromie field potentials in response it) stimula-I ion ot t he striltuLM raditrm prior to and A) 'mini after beimnninge .xposure to DiT (1.5 mM). DYT increased t he amplitude of t he initIialpopulation spike and caused multiple spiking. B: tUtT indued spontaneous action potential firing. increased synaptic input andcaused sorne memblrane depolarization. At the arroA%. straturn ridiatumn was stimulated. Action potentials were truncated bys thechart reorder C: orthodrornie action potentials recorded from C A, pyramidal cell. Straight line indicates zero potential. The onset ofmultiple spikiirqi in the orthodroic field potential coincided with the appearance of multiple firing recorded intracellulars'.
sure to DFli the membrane potential wa-is depola- EPSPs were examined in 5 cells. Comparisons ofrized to -(flo ±h 11.7 tnV (Student's 1-test for paired EPSPs hefore and after DTT exposure were alwayssamples. it = 25. P' 01.015. WT did not significantly made at the same membrane potential. The rate ofalter the input resistatnce of CAI pyramidal cells. The rise and the amplitude of the early component of theinput resistatnce tin control was 41.7 ±4.7 MIQ and fol- EPSP were not significantly affected by DTT (Fig,.lowing exposure to IF was 44.01 5.10 NI2 (nt = 12. 2A). The mean amplitude in control was 6.2 ± (1.51)> .015). mV while after exposure to DIT was 5.9 ± 01.6 mV
Increased excitability of CA, pyramidal cells was (nt 5. P > 0.05). The rate of rise of the EPSP wasevident when thev were stimulated orthodromicalk%. 1.4 0 (.2 mV/ms in control and 1.5 ± 01.2 mV/ms af-In control. the stimulus applied to the stratumn radia- ter DIT exposure (n =5. P > 01.05). Similarly, thetumn was adjusted to elicit an EPSP with a single ac- extracellularly recorded population synaptic re-tion potential (Fig. IC. Control). After exposure to sponse showed no change in the initial slope"'. DFliPIT, the same orthodromic stimulus evoked multi- did. however. conststently. prolong the EPSP (Fig.ple (3-7) action potentials (Fig. IC). The onset of 2A). DPIT increased the mean duration from 401.5 ±multiple firing of pyramidal cells in response to or- 4.4 to 91.4 ± 13.3 ms (n = 4. P < 0.0)5) measuredthodromic stimulation Loincided with the appearance from the initial depolarization to the time it returnedof multiple spiking in the orthodromic field potential to the resting membrane potential. The shape of the(Fie. IA. C). Even when membrane potential was EPSP depended on stimulus strength. Stimulationmaintained at pre-DTIT levels, multiple spikes were with very low currents could evoke only a delayed de-elicited by orthodromic stimulation. Subthreshold polarizing potential with no initial EPSP. As the stim-
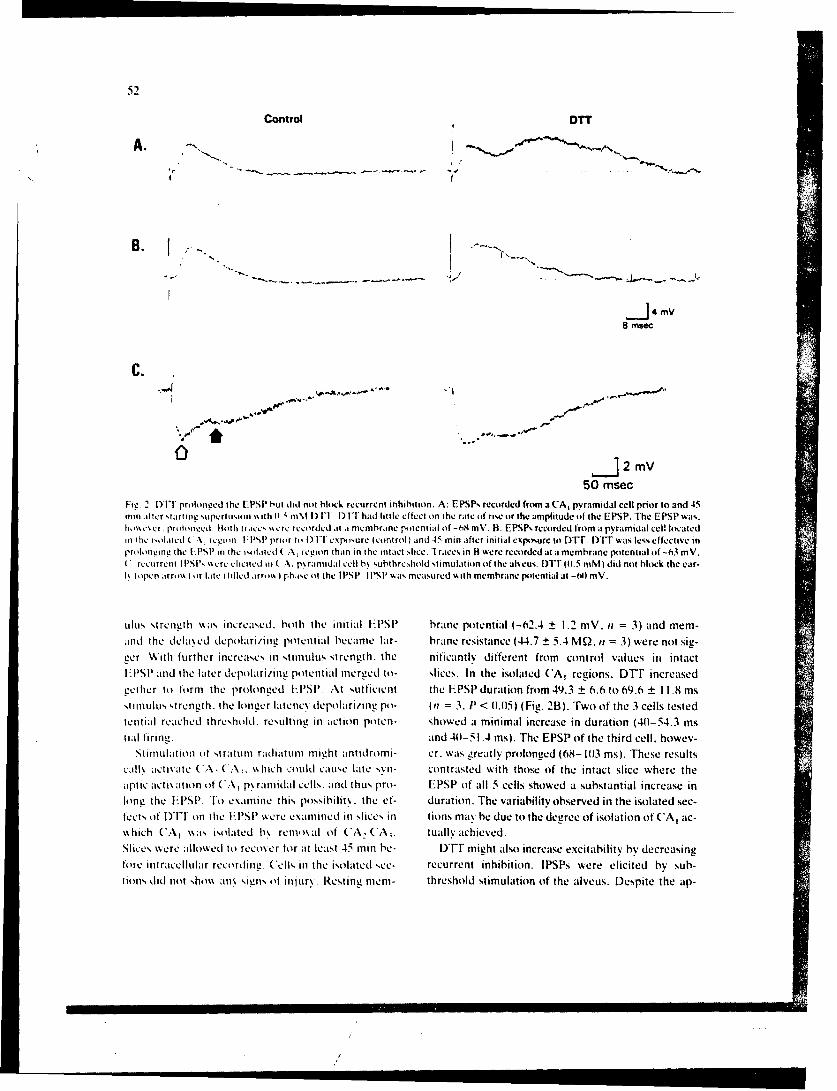
Control OTT
A.
B.
-J4 mV8 msec
C.
2 mV50 mnsec
Fig. 2. 1)11I pruolonged the EPSP but did not block recurrent inhibition. A: EPSPs recorded from a CA, pyramidal cell prior to and 45filn alIter startI iie upcriuitn in %it h 11 m; tin %f IT had littIle effect tin the rate of rise or the amplitude of the EPSP. Trhe EPSP was.hl0A'Actr proloncd Btoth traces %%ere recorded mt a membrane potential of -h8 mV. B: EPSP% recorded from at pyramidal cell locatedin the isolated (.A rc 'ion II'S!' prior to 1)1-1 expl)ure (control) and 45 min after initial expoisure it) Drr. DFIT was less effectiv~e inro~longing the L*PSP in the isolated ('A, region than in the Intact slice. 'Iraces in B were recorded asta memhrane potential of -63 mV.(recurrent I l'SI's %ere elicited tit( Al p\ rantidal cell b% ,utithreshold stimulation of the aireus. flIT M0.5 mNM) did not block the ear-
ls opilen iirrou I or late tilled irronik% phase (it the I l'SP. IPIS!' \%as measured with membraine potential at -Wl mV.
ulus strength was. tncreased. hoth the initial FPSP brane potential (-62.4 t 1.2 MV. it = 3) and mem-and the dcla .ed depolaiing potential became lar- brane resistance (44.7 ± 5.4 MQ. it = 3) were not sig-gecr. With further increases in stimulus strength. the nificanl~v different from control values ini intact1-PSP anid the later dcpolari/ini! potential mer~ed to- slices. In the isolated CA, regions. DTT increasedgecther to tormi the proloni!ed Fl'SP At sufficient the EPSP duration from 49.3 ± 6.6 to 69.6 ± 11 .8 mssltilus strer-I~th. the longer latenc\ depolariitg po- (It 3. P' < 01.05) (Fig. 213). Two oif the 3 cells testedtetitial reached threshold. restiti in action poten- showed a minimal increase in duration (401-54.3 mstial tiring. and 401-51.4 ins). The EPSP oif the third cell. howev-
Stimulation tit s ratuin radiatum might antidromi- er. wa-is greatly proloneed (68- 1013 mns). These resultscalk~ acti% ate C, C. ( \\~ hich could cause Late s\xn- contrasted with those of the tntact sltce where theaptic acti~atiotiot C~A praimidal cells. anid thus pro- EPSP of all 5 cells showed a substantial increase inlong the HPSP. To csamine this possibility, the ef- duration. The variability observed in the isolated sec-
let Do YI oth PPsreeaidinlcsin tions mav be due to the degree of isolation of CA, ac-s\ hich CA, %\as isolated h\ remotital of CA, CA,. tually achieved.Slices were alwdto recover for att least 45 min be- DT T might also increase excitability bw decreasingtore intracellular recoirding. Cells tin the isolated sec- recurrent inhibition. IPSPs were elicited by sub-(totns did not sho%\ aiti signs of injur\. Resting mnem- threshold stimulation of the alveus. Despite the ap-

53
CONTROL 60 MIN DTT AHP. The effects of DlT on the AllP produced by atrain of 4 action potentials were examined in 4 cells.As shown in Fig. 4. DT-I did not alter the early or late
phase of the AHP. In control, the late AHP was 6.75.N) mV while after exposure to DTT. the AHP
was 6.38± 1.55 mV (n = 4, P > 0l.05).
20 ' V DISCUSSION
1lw S ) 1 Iccrcacd spike Irequency adaptation recordedtroin (A, p.ranidiI cell. Spike Ircqucnc, adaptation 's ex- This study demonstrates that DT - increases theanimncd h'% ',timulatiig a cell \.ith depolari/ing current pul.c'. ol excitability of CA, pyramidal cells in the hippocam--m0 , dnilt ion Recordini. \.ere made prior to (Control) and pal slice. This increased excitability is manifested by(i mii iltcr t,.arom, t)1I cx o' ur.. Membrane potential re-miiincd'it--S mV the appearance of the following phenomena re-
corded from CA 1 pyramidal cells: (I) spontaneous
pearance ot increased excitability. DTI did not re- spiking and occasional spontaneous bursting activity:duct the amplitutide of the early or the late phase of (2) abnormal repetitive firing in response to ortho-the recurrent IPSP (early IPSP, control 5.4 0 t).6 dromic stimulation: and (3) a reduction in spike fre-nV. I)Vr 5.2 - \11.5 rV' late IPSP. control 2.3 + ((.6 quency adaptation. The appearance of these intra-mV. IYl-T 2.1 1.3 mV. ?i = 3. 1 > 1.t15) (Fig. 2C). cellular events coincides with the onset of changes inThe duration ot the IPSP also did not change follow- the orthodromic field potential recordings reportedine expo,,ure to I1- (control. 319.2 = 38.6 ms: previously".DTF. 311.8 ± 6(i.() nis. ;t = 3. P > t.05). A prolongation of the EPSP appears to be respon-
A depolarizing current pulse of 71) ms duration sible for the repetitive firing elicited by orthodromic
produced a train of action potentials that decreased stimulation following treatment with DTT. Onein frequency during the step (Fig. 3). This phenome- mechanism that could account for the increasednon. spike frequency adaptation. has been well de- EPSP duration is the blockade of inhibitory inputs toscribed in hippocampal neurons". Spike frequency the pyramidal cells. The convulsants bicuculline andadaptation was examined prior to and after DT ex- penicillin prolong the EPSP through this action"posure in 4 cells (Fig. 3). DTT decreased spike fre- In contrast to these agents DTT does not alter the re-quency adaptation in all cells examined. DT" more current IPSP. Feedforward inhibition was not tested,than doubled (2.29 ± (0.25 times. it = 3) the number however, and may be affected by DTT.of action potentials elicited h% 7(H1 ms depolarizing An alternative mechanism for EPSP prolongationstep. The increased number of action potentials did is inhibition of the reuptake of the excitatory neuro-not result from a change in the resting membrane po- transmitter. The transmitter released by Schaffertential or input resistance. There was no obvious collaterals at the CA, pyramidal cells is thought to bechange in the shape of the evoked action potentials. the excitatory amino acid. glutamate.5 3 ' . Considering
Spike frequency adaptation is. in part. due to the that DTT (0.5 mM) decreases y-aminobutyric acidtemporal summation of the calcium-dependent slow reuptake2 . it is feasible that through a similar mech-
Control 60 min DTT
Vm =-60 mV Vm -60 mV j 2.5 mV
1 sec
Fig, 4. t)tT did not alter the early or the late phase of the AHP following 4 action potentials evoked by a train of 4 suprathreshold cur-rent pue'. im,,. ,11 ltil traces were recorded prior to and N) min following the start of DTT supcrfusion.

54
anism. 1) 1-1 could impair the reuptake of excitatory larlv Terrarx' observed that a 3(0-min exposure to
amino acids. The multiphasic shape of the prolonged DTT caused a small depolarization in muscle. Such aTPSP followiing i)1I exposure is inconsistent with mechanism is not likely to explain all the actions of
thi, possibilit. I)IT. howe er. The oxidized form of DIT (trans-4.5
\ third mechanism for prolongation of the I-FPSP is dihydroxv- 1.2 dithiane. ()xDIT) was found in extra-thal 1)TT ,lters the interaction of the CA, and the cellular experiments to he ineffective in inducing(..\, (*A. ,,subfield,s. Stimulation o the ,tritum radia- multiple spiking in the orthodromic field potential"'.tur it) elicit an orthodromic response in CA, also an- Preliminary intracellular experiments indicate that.
tidrom:iall% acti\ates the CA, ('A region. If the cx- lthough OxDTT causes a similar depolarizationcitaibilitx of this region %\as cle\ated b\ DFl. then (3-8 mV). it does not induce burst firing.stimulation ot the stratum radiatun could result in a DTT decreased spike frequency adaptation. Thisrexerberating circuit between CA, and the CA, CA, effect would promote repetitive firing since normal,,tub1field,. Thus a rcactivation of CA, resulting from mechanisms to attenuate trains of actions potentialsthe delayod firing of the C..\ CA region w\ould pro- are impaired. The calcium-dependent potassium cur-
long the tlPSP" :'. Support for this hpipothesis comes rent contributes %ignificantly to regulation of firingfrom the tibser\ ation that DI seems to be less effec- frequency in hippocampal pyramidal cells" ' . A de-ti.e in prolonging the I-PSP in isolated CA, regions crease in this current, however, is unlikely to he thethan in intact slices. In addition, it was possible to mechanism since the AHP (resulting from the calci-stimulate the stratum radiatum at a sufficientlv low um-dependent potassium current"0i' ' -) is un-stimulu,. strength to aoke a long latency potential in affected by DTT.
the absence of an early EPSP. This longer latency po- In conclusion. DTT increases the excitability of thetential most likely represents activation of CA, re- hippocampus in vitro. This effect probably resultssuiting from the dela.ed firing of the CA. CA, region from DTT-induced reduction of disulfide bonds sincein response to the stimulation of the stratum radia- the reduced form. OxDTI. does not cause similarturn. hyperexcitability. DTT is known to interact with a
DTUT prohabl. does not increase excitability of number of neurotransmitter systems which are pres-CA, pyramidal cells by increasing transmitter release ent in the hippocampus. Although stimulation ofor by enhancing sensitivity of the receptor mediating stratum radiatum predominantly activates the Schaf-normal synaptic excitation. If these mechanisms fer collaterals and therefore an excitatory amino acidwere responsible for the increased excitability. DTT pathway, other inputs and other transmitter systemswould be expected to increase the amplitude and rate are likely to be activated. It is not unlikely that altera-of rise of the LPSP. Instead. DTT had minimal ef- tion of these systems would contribute to the dvsfunc-fects on these parameters. Although the neurotrans- tion of the hippocampus following DTT exposure.mitter mediating the EPSP produced by Schaffer col- The abnormal activity produced by DTT in neurallateral stimulation is uncertain, it is likely to be gluta- tissue would limit its usefulness as a radioprotectant.mate' '". To date. no effects of DTT on the excitatoryamino acid receptors have been reported. DTT does ACKNOWLEDGEMENTSnot alter the response of Onchidium esophageal gan-glia to applied i.-glutamate2 . Other sulfhydryl Supported by the Armed Forces Radiobiology Re-modifying reagents such as mercuric chloride, n- search Institute. Defense Nuclear Agency. underchloromercuribenzoate and n-ethylmaleimide de- work unit (10105. Views presented in this paper arepress rather than enhance glutamate and kainate re- those of the authors: no endorsement by the Defensesponses recorded from pyramidal neurons of the rat Nuclear Agency has been given or should be in-hippocampus -23 . ferred. Research was conducted according to the
Extracellular recordings suggested that DT" in- principles enunciated in the 'Guide for the Care andcreased excitability through both synaptic and extra- Use of Laboratory Animals' prepared by the Insti-synaptic mechanisms"'. Most neurons were some- tute of Laboratorv Animal Resources. National Re-what depolarized following exposure to DTT. Simi- search Council.

55
REFE RENCES oxygen and sullbvdrvl-containing compounds (on irradiatedtransforming DNA. II. Glutathione. cystetne. and cs'ste-
I Aleer. B.E. and Nicoll. R.A.. Epileptiform burst afterhv- amine. Int. J. Radiat. Biol.. 45 (1984) 615-62h.perpolarizatuin: a calcium dependent potassium potential 19 H'otson. J.R. and Prince. D.A.. A calcium-activated hyper-in hippoca mpal ('AI pv~ram idal cells. Science 211) (19~80)) poilarizat ion follows repetitive firing in h ippocampal neu-122- 124. rons. JiNeiurip/ivsiot.. 43 ( 1980) 4lN-4 19.
2 llassant. MI. and Court. L... Effect of whole-body irradia- 2(0 Iversen, L.L. and Johnston, G.A. R.. jABA uptake in rattion on the actiy tit rabbit hippocampal neurons. Radial. central nervous system: comparison sif uptake in slices andRei. 75) 197S) 593-ti)ti homogenates and the effects oif soime inhibitors. J. Veiiro-
3 Brown. D.A. and K%%iatkaiski. D)., A note (in the effect of chern.. IS( 1971 ) 1939- 1952.dithiothreitol (DPI) on the depolarization of isolated svm- 21 Karlin. A. and Bartel%. E.. Effects of blocking sulphydrvipath.,tic ganglia bs' carbachol and brimsiacetscholine. Br. groups and oif reducing disulphide bonds on the acets'lcho-J. Pharmnaco/.. 56 ( 197h) 128- 130. line-activated permeaility system sif the clectroplax. Bio-
4 Cleland. W.W.. Dithiothreitol. A nesw protective reagent c/tim. Biophss. Acta. 126 (1966) 525-535.for 511 groups. Btochemstrv. 3 1964) 4801-482. 22 Kato. M.. Oomura. Y. andlMaruhashi. J.. Effects of cmi-
5 Cotman. C.W. and Nadler. J.V.. Glutamate and aspartate call modification (in the i-glutamate receptors (in the (inc/i-as hippocampal transmitters: biochemiical and pharmaco- diun neurons. Jap. J. P/tcsiol.. 33 ( 1983) 535 -546.logical evidence. In P.J. Roberts. 1. Stoirm-Mthiscn and 23 Kiskin. N..Krishtal. O.A.. Tsvndrcnko. A.Ya. andG A. R. Johnson I Eds.)I. (lthamnate: Framisttoer in i/ue Cen- Akaike. N.. Are sulfhydryl groiups essential for function oftrat Nesrvouk .Svemn. Wilcv and Sons. New% York. 1981. pp. the glutamate-operated rcccptor-ionophore complex.117-154 .Veurosci. Lett.. 66 (1986) 30)5 -3 1).
ii Dingled inc. R., Flippocampus: synaptic pharmacology. In 24 Lucas. MI.. Hanoune. J. and Bockaert. J.. Chemical modifi-R. [)ingledine (Ed.)I. Brain S/ices, Plenum. New York. cation of the beta adrenergic receptor coiupled with adenyl-1984, op. 87- 112. ate cvclase by disulfide bridge-reducing agents. Vio. P/tar-
7 Dingledine. R. and Gjerstad. L..- Penicillin blocks hippo- ,naco/. - 14(1978) 227-236.campal i.p.s.p.s unmasking prolonged e.p.s.p.s. Brain Re- 25 Lukas. RiJ. and Bennett. EL.. Chemical modification ando'arch. 168 ( 1979) 205 -29. reactivity of sulfhydrsvls and dlisulfides (if rat brain nicotinic-
XS 1)inaledine. R. and (ijerstad. L.. -Reduced inhibition dur- like acetylcholine receptors. J. Biol. (Jie,,i. 255 (1981)ing eptleptiform activo it in the in vitro hippocampal slice. J. 5573-5577.P/rcdo/. (Lond. 1,305 1 1980)1297-313, 26 Madison, D.V. and Nicoll. R.A.. Control oif the repetitive
9 Dlingledine. R., Hynes. M.A. and King. G.L.. Involvement discharge of rat CAl pyramidal neurones in vitroi. J. Phv-iif N-methvl-io-aspartate receptors in epileptiform bursting jot. (Lond.), 354 (1984) 319- 33 1.in the rat hippocampal slice. J. Phvsio/. (ILond., 380 (1986) 27 Nlarzullo. G. and Hine. B.. Opiate receptor function may175-189. be modulated through an oxidation-reduction rn-chanism.
M) Donaldson. J. ,ind Hill. SiJ.. EnhaLncement (if histamine Science. 208 (1980) 1171- 1173.N -receptor agonist .ictivitv by I .4-dithiothreitol in guinea- 28 Pederson. S.E. and Ross. E.M.. Functional activation oifpig cerebellum !nd cerebral cortex. J. Nettroche, 47 beta-adrenergic receptoirs by thiols in the presence (it ab-)l98M6) 1470- 1482. sence of agonists. J. Biol. Chem.. 26( 1985) 14150- 14157.
11 Diinaldson. J. and Hfill. SiJ.. l.4-Dithiothreitol-induced al- 29 Pellmar. T.C.. Electrophysiological correlates oif peroxideicratin in histamine 11,-agonist binding in guinea-pig cere- damage in guinea pig hilppocampus in sitro. Brain Re-bellum and ccrcbral cortex. Eur. J. P/tar,,acol.. 129 (198h) searc h. 364 (1986) 377- 381.25, -3,1. 30) R a ng. H. P. and Ri tter. J. M.,The e ffects o f d isu Iphidle bon d
12 (iangloff. if.. Acute effccts of X-irradiation (in brain clee- reduction on the properties of cholinergie receptors in ehicktrical activitYi in cats and rabbits. In Effectvs of Ionizing Pa- muscle. Mo/. Pharmacol.. 7 (1971) 6201-63 1.dhilon on? the Nervous S~stern. International Atomic Ener- 31 Redpath. i.L.. Pulse radiolysis of dithiothreitol. Radiationgy Agency. Vienna. 1962. pp. 123- 135. Res.. 541(1973) 364-374.
13 (iustafsson. B. and Wigstrom. HI.. Esidence for two types 32 Redpath. i.L.. Radioprotection of enzyme and bacterialof afterhyperpolaiiation in (Al pyframidal cells in the hip- systems by dithiothreitol. Radiation Res., 55 (1973)pocampus. Brain Research. 2) 1198)14()2-468. IM~- 117.
14 llablitz. J.J.. Picrotoxin-induced epileptiform activity in 33 Schoenbrun. R. L.. Campeau. E. and Adley. W. R.. Elctro-hippocampus: role of endogenous versus synaptic factors, encephalographic and behavioral effects from x-irradiationJ .Neurophcso/.. '5 11(1984) 1011 1-10)27. oif the hippocampal system. In TiJ. Haley and R.S. Snider
IS Hale%. T.J. - Chanees induced in brain activity by low doses (Eds. I. Response of the Nerc-ous Syostemi to Ionizing Radia-tif X-irradiation. In Effects of /oitng Radiation on tie tion, Second International S 'yrposium. Little and Brown.Nertoouv Svstern. International. Atomic Energy Agency. Boston. 1964. pp. 411-428.Vienna. 1962. pp. 123- 135. 34 Sehwartzkroin. P.A. and Prince. D.A.. Changes in excita-
16 Hedlund. B. and Bartfai. T..- The importance oif thiol and tory and inhibitory potentials leading to epileptogenie ac-disulfide groiups in agonist and antagoinist binding to the tivity. Brain Research, 183 (19801)61-76,muearinic receptor. Mo. Pharmacol. - 15)(1979) 531-544. 35 Schwartzkroin. P.A. and Stafstrom. C.E.. Effects of
17 FHeld. K.D.. Interactiions of radioprotectors and oxygen in EGTA on the calcium-activated afterhyperpolarization incultured mammalian cells. 1. Dithiothreitol effects on radia- hippocampal CA3 pyramidal cells. Science, 210 (1981)tion-induced cell killine. Radiation Res., 101) (1985) 1125-1126.4241-433. 36 Storm-Mathisen. J.. Localization of transmitter candidates
18 H~eld. KI).. - Farrop. H.A. and Mlichael. B.D.. Effects of in the brain: the hippocampal formation as a model. Prog.

56
.VrurobtoI., 8 (1477) 1 19- 18 1. 41 Tolliver, J.M. and Pellrnar. T.C.. Ionizing radiation alter%37 Suen. E.T., Stefanini. E. and (lement-Cormier. Y.C.. Evi- neuronal excitability in hippxeampal slice% of the guinea
dence for essential thiol groups and disullide bonds in agon- pig. Radiation Res.. 112 (198/) 555-563.ist and antagonist binding to the dlopamnine receptor. Rio- 42 Wong. R. K.S. and Prince. D.A.. Atterpotential generationtlhemt. Biop/o... Rev. C onornn.. 96 ( 19801) 9i3-960(. in hippocampal pyramidal cells. J. Neurophitsm.., 45 (198 1)
38 Ferrar. D. A.. Effects (if dithiothrecil on end-plate cur- 86-97.rents. J. l'hiviol. (Ioc~.27h 1978) -44(3-417, 43 Wong. R.K.S. and Traub. R.D.. S 'nrchronized hurst dis-
39 Foliiser. JiM. and l'ellrnar. T.C.. lntraicelluLiar .inalss of charge in disinhihited hippocampal slice. 1. Initiation indiihiothreitol effects oin guiea pi ' CA]I ptyramidal cells. CA2 CA3 region. J. Nettrop/iVA1Il.. 49(1983)442-458..So Ve'urosu .4 hvtr . I-2(198h) h76. 44 Zhicz. K.L. and Weight. F.F. Transient iVoltage and calci-
4411 lolliser. J, %1. aind Pelimar. I.C.. Dithiothreitol elicits epi- urn d&pendent outward currents in hippocampAl CA3 pyra-leptiforrm actiN t in (Al of the guinea pig hippocampal midal neurons. J. Neuropmvuml. 41t(1985) (038- 11158.slice. Brain Reo'arc h. -4(1-4 1I987) 133- 141

AAM10 FONCIS NAOIOgIIOL6
AISEANCH IN4STITUTE
INFECTION AND IMMUNITY. Oct. 1988, p. 2650-2657 SCIENTIPIC AVPON, Vol. 56. No. 100o19L9567/8&10265-08$02.00/0Copyright 0 198. Afnercani Society for Microbiology SR88-29
Recombinant Interleukin-la and Recombinant Tumor NecrosisFactor a Synergize In Vivo To Induce Early Endotoxin
Tolerance and Associated Hematopoietic ChangesSTEFANIE N. VOGEL." ERIC N. KAUFMAN.' MICHELE D. TATE.' AND RUTH NETA2
Department of Microbiology, Uniformed Services University of the Health Sciences, 4301 Jones Bridge Road,' andDepartment of Experimental Hematology, Armed Forces Radiobiology Research Institute.2 Bethesda, Maryland 20814
Received 25 April 19MAccepted 11 July 1988
Endotoxin, the lipopolysacharlde (LPS) derived from gram-negative bacteria, Invokes a wide range ofresponses in susceptible hosts. It is known that virtually all resposes to LPS are mediated by the action ofmacrophage-derived cytokines (such as interleukln-I [IL-I], tumor necrosis factor (TNFI, and others) whicharm produced principally by macrophages and maximally within several hours of LPS administration. Onemanifestation of LPS administration which is not well understood is the phenomenon of "early endotoxintolerance." In response to a single sublethal Injection of LPS, experimental animals become refractory tochallenge with a homologous or beterologous LPS preparation 3 to 4 days later. Animals rendered tolerantexhibit mitigated toxicity and a reduced capacity to produce circulating cytokines (i.e., colony.stimulatingfactor or interferon) in response to the challenge LPS Injection. Previous studies have also shown that this stateof tramnsient, acquired hyporespoasiveness to LPS is accompanied by a marked Increase in the size of cells inthe bone r-'-row which are enriched in numbers of macrophage progenitors. In this study, we examined thecal;city ok r-combinant IL-I or recombinant TNF or both to Induce early endotoxin tolerance and Itsassociated bematopoietic changes. Neither cytokine alon was able to mmik LPS for induction of tolerance.Combined adminlstdon of recombinant IL-i and recombinant TNF dom which were not toxic whenadministered Individually led to synergistic toicity (a assessed by death or weight loss). However, within anontoxic range, the two cytokine synergized to Induce a significant reduction in the capacity to producecoky-stimulating factor in response to LPS, as well as the characteristic increme in bone marrow cell siz andmacrophage progenitors shown previously to he asociated with LPS-induced tolerance.
Endotoxin. the lipopolysaccharide (LPS) component of For instance, anti-IL-1 serum has been shown to blockgram-negative bacterial outer membranes, induces in vivo LPS-induced fever (8). Similarly, injection of anti-TNFmany of the pathophysiologic changes associated with sys- antibodies afforded significant protection against lethal chal-temic gram-negative bacterial infection (reviewed by S. N. lenge with LPS (4). Lastly, in mouse strains that bear theVogel and M. M. Hogan, in J. J. Oppenheim and E. Sher- defective allele for LPS responsiveness. Lps'. protein-freeach, ed.. Immunophysiology: Role of Cells and Cytokines in preparations of LPS fail to elicit the production of macro-Immunity and Inflammation, in press). Among these are phage-derived factors, either in vivo or in vitro (reviewed byfever, hypoglycemia, hypotension, shock, and even death. Vogel and Hogan (in press]).Several lines of evidence support the hypothesis that the A single injection of LPS results in the appearance of amacrophage is the principal cellular mediator of endotoxicity temporal hierarchy of factors within the serum (reviewed byand, more specifically, that LPS-induced, macrophage-de- Vogel and Hogan (in press]). The first group of cytokinesrived soluble factors are the direct mediators of endotoxic (i.e., IL-I, TNF, and interferon) appears maximally within 2phenomena. First, LPS-stimulated macrophages produce in h of injection, and they have been referred to as -earlyvitro many of the same soluble factors which circulate in the acute-phase reactants." By 4 to 8 h postinjection, circulatingserum following in vivo administration of LPS. These in- levels of these factors are greatly decreased. Production ofclude interleukin-1 (IL-1), tumor necrosis factor (TNF; also granulocyte-macrophage CSF activity is somewhat delayed,referred to as cachectin), interferon, colony-stimulating fac- with peak activity occurring at 4 to 6 h following LPS andtor (CSF), and prostaglandins of the E series. When purified declining to basal levels by 24 h. A third group of solubleand injected into experimental animals in the absence of factors which appears maximally in the serum 18 to 24 hLPS, many of these soluble factors have been shown to postinjection has been collectively referred to as "lateinduce one or more of the effects of in vivo LPS treatment.Agents which increase macrophage activation greatly in- acute-phase reactants" and includes C-reactive protein.
crease endotoxin sensitivity in vivo, even in the genetically serum amyloid A, fibrinogen, and others. This last group of
LPS-hyporesponsive C3H/HeJ mouse strain (23, 27). In- factors is produced primarily by hepatocytes and is inducedcreased sensitivity to LPS correlates well with quantifiable by the action of early acute-phase reactants on these cells.increases in levels of circulatingl LPS-induced factors (5, 13, For many years. it has been recognized that initial suble-
24, 27, 32). In studies in which antisera against specific thai exposure to LPS renders experimental animals refrac-cytokines have been administered prior to or simultaneously tory to a subsequent challenge with homologous or heterol-with LPS, many of its biologic effects have been mitigated. ogous LPS several days later. This effect was referred to as
"early-phase endotoxin tolerance" (reviewed in reference9). Until recently, very little was known about the mecha-
Corresponding author. nisms which underlie this phenomenon. Early studies dem-
2650

VOL. 56. 1988 IL-I AND TNF SYNERGIZE IN EARLY LPS TOLERANCE 2651
onstrated that macrophages derived from mice which had were maintained at -20*C, either in lyophilized form or asreceived a tolerance-inducing injection of LPS failed to intermediate stocks at high protein concentrations, beforerespond to subsequent LPS challenge in vitro to produce dilution in pyrogen-free saline just before injection.endogenous pyrogen or prostaglandins (7. 21). In this sense, Meurem of CSF activity in serum. Serum was testedmacrophages from mice rendered tolerant are phenotypi- for CSF activity in a bone marrow colony assay in semisolidcally similar to macrophages rendered refractory to LPS by agar as described previously (11). Briefly, serum was ob-pharmacological means (e.g.. by treatment with glucocorti- tained from pooled blood collected from mice injected (ascoids; 3. 28). Williams et al. (31) demonstrated that a splenic indicated) with saline, rIL-1. rTNF, or LPS. Serial dilutionsadherent cell population was necessary for abrogation of of serum were made in six-well tissue culture plates (Costar.early-phase endotoxin tolerance in transfer experiments. Cambridge. Mass.). Bone marrow cells were obtained fromsupporting the cellular nature of this phenomenon. In studies the femurs and tibias of C3H/HeJ mice and processed byperformed in this laboratory, it was demonstrated that density gradient centrifugation in lymphocyte separationearly-phase endotoxin tolerance is associated with alter- medium (Litton Bionetics, Kensington, Md.). The cells wereations in bone marrow-derived macrophage precursor pools then collected from the gradient interface and diluted to a10, 11). Specifically, cell-sizing profil,:s of bone marrow final concentration of 100 cells per ml in a mixture of tissue
cells from mice rendered tolerant showed enrichment for a culture medium and molten agar. One milliliter of the cellpopulation of cells significantly larger than control bone suspension was added to each of the wells, which containedmarrow populations. and by density gradient sedimentation 0.2 ml of the serum dilution. The wells were mixed byit was shown that the denser population of cells contained swirling and allowed to solidify. Cultures were incubated atincreased numbers of macrophage progenitors. The induc- 37rC (6% C02) for 7 days. at which time bone marrowtion, maintenance, and loss of these hematopoietic changes colonies (025 cells) were enumerated with an invertedcoincided temporally with the acquisition, maintenance, and microscope.loss of the tolerant state. These changes were observed in a Determinatin of the number of macrophage progenitorvanety of outbred and inbred mouse strains, including those cells in boat marrow. The number of macrophage progenitorwith defects or deficiencies within certain lymphoid cell cells was determined as described elsewhere (11). Briefly.subsets. For example. early-phase endotoxin tolerance, as mice from each experimental group were sacrificed, and thewell as the associated hematopoietic changes, were ob- bone marrow cells were obtained from the femurs and tibiasserved in athymic (nude). B-cell-deficient (xid), and splenec- by flushing with serum-free medium. The bone marrow cellstomized mice (12). As an additional control, LPS-hypore- were centrifuged and suspended in tissue culture medium.sponsive C3HiHeJ mice did not exhibit any of the Cell counts and cell-sizing profiles were obtained with ahematopoietic alterations observed in fully LPS-responsive model ZM Coulter Counter and a C1000 Channelyzermice rendered tolerant by injection of LPS (11). (Coulter Electronics, Inc., Hialeah, Fla.) calibrated as di-
In this study, we tested the possibility that cytokines rected by the manufacturer.which are normally produced in response to a tolerance- To determine the number of macrophage progenitors, ainducing dose of LPS may mediate the induction of early double-layer, semisolid agr colony assay was used asendotoxin tolerance or the accompanying hematopoietic described previously (11). An excess of partially purifiedalterations or both. Our findings indicate that combined murine macrophage CSF (CSF-1) was incorporated into thetreatment of mice with recombinant IL-la (rlL-1) and bottom layer of the assay system. Bone marrow cells (5 xrecombinant TNFa (rTNF) induces a significant, synergistic 10 or I x 105) were then suspended in a molten agar mediumlevel of toxicity, but at sublethal-dose ranges they induce a mixture and overlaid onto the CSF-1-containing layer. Col-significant degree of endotoxin tolerance, as well as changes onies (2!50 cells) were enumerated after 10 days in culture,in bone marrow cell-sizing profiles and enrichment for and the number of progenitors per IWs input bone marrowmacrophage progenitors previously associated with the tol- cells was calcuiated.erant state. Injecton schedule for inductim of early endotoxln toler-
ance. The protocol for induction of early endotoxin tolerancewas identical to that used in previous studies (10-12, 31).
MATERIALS AND METHCDS Briefly, mice were injected intraperitoneally with 25 ;Lg of E.Mice. Female C57BL/6J mice 5 to 6 weeks old were coli K235 LPS in a volume of 0.5 ml on day 0. Three days
purchased from Jackson Laboratory (Bar Harbor, Maine) later, mice were challenged with 25 Lg of LPS and bled 6 hand used within 2 weeks of their receipt. Mice were allowed later for measurement of CSF in serum. Controls includedaccess to food and acid water ad libitum. mice injected on days 0 and 3 with pyrogen-free saline and
Reagents. Protein-free LPS was prepared from Esche- mice injected on day 0 with saline and on day 3 with LPS.richia coil K235 by the phenol-water extraction method of Mice which received rlL-1 or rTNF or both were alsoMclntire et al. (14). Human rIL-lct (lot SM59) was the injected on day 0 with the indicated concentrations ofgenerous gift of Hoffmann-LaRoche Inc. (Nutley. N.J.) and cytokines but challenged on day 3 with saline or LPS.possessed a specific activity of 5 x 10' U/mg. The activity ofthis material was verified frequently in a standard thymocyte RES'LTScomitogenic assay (25) throughout the course of this study.The concentration of contaminating LPS in this rlL-1 prep- Toxicity of r[L-I an rTNF whe adminiswe individuallyaration was 1.5 ng1.3 x 10 U. Human rTNFu (lots NPI02 or in combleuam. In our previous studies, an early-phaseand NP200B) was the generous gift of Cetus Corporation endotoxin tolerance system was established in which injec-(Emeryville. Calif.) and possessed a specific activity of tion of 25 to 50 .g of LPS (approximately 0.05 to 0.1 50%approximately 2 x 107 U/mg. The activity of the rTNF was lethal dose) was shown to render mice significantly lessalso verified in a standard actinomycin D-treated L929 responsive to an LPS challenge-3 to 4 days later. Since LPSfibroblast cytotoxicity assay (29). rhe concentration of LPS has been shown to induce both IL-I and TNF concurrently,in the rTNF preparations was <0.03 ng/0.3 mg. All reagents and since both cytokines have been shown to mimic LPS
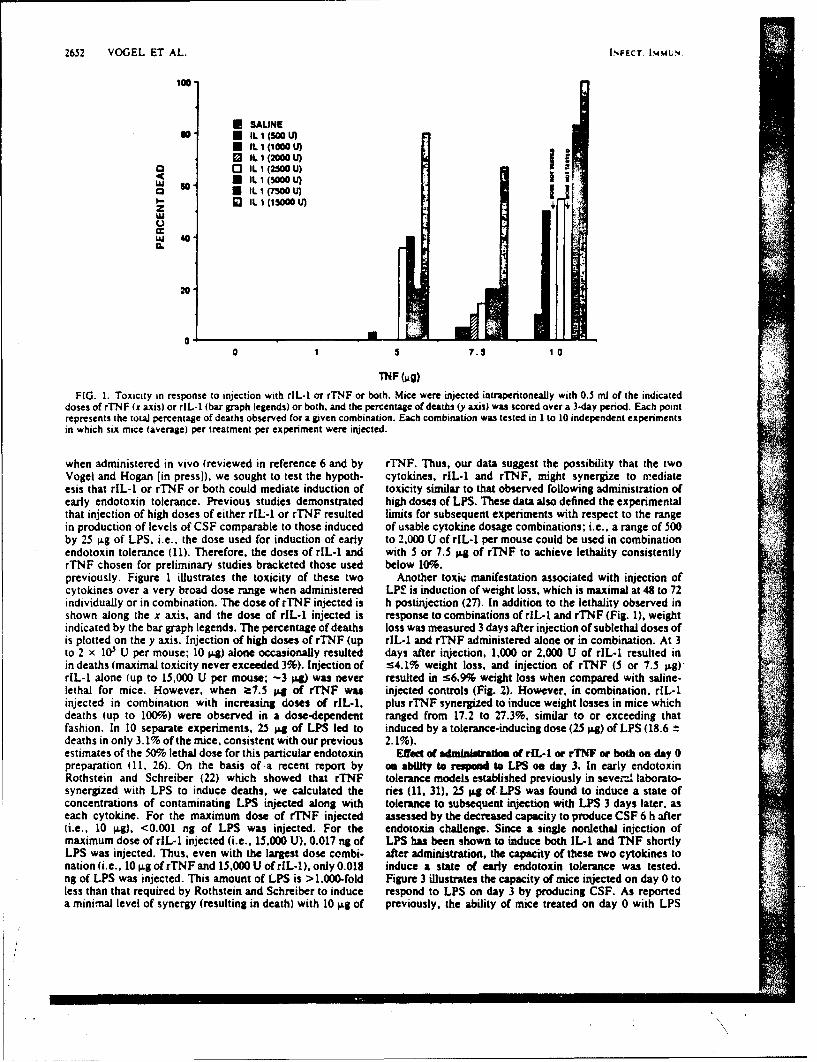
2652 VOGEL ET AL. INFECT iMU4N.
100
* SAUNEso, • IL 1 (5O0 U)
I t. 1 (1000 U
3 IL 1 (2000 U)0 I" IL 1 (2500 U)
< IL 1 (500 U)IL. I M US3 II. 1 (7500 U)
z
Wj 40'0
20
0 1 7.5 10
TNF (jig)
FIG. 1. Toxicity in response to injection with rIL-1 or rTNF or both. Mice were injected intraperitoneally with 0.5 ml of the indicateddoses of rTNF (x axis) or rlL-1 (bar graph legends) or both. and the percentage of deaths (y axis) was scored over a 3-day period. Each pointrepresents the total percentage of deaths observed for a given combination. Each combination was tested in I to 10 independent experimentsin which six mice (average) per treatment per experiment were injected.
when administered in vivo (reviewed in reference 6 and by rTNF. Thus, our data suggest the possibility that the twoVogel and Hogan [in press]), we sought to test the hypoth- cytokines, rlL-1 and rTNF, might synergize to mediateesis that rlL-l or rTNF or both could mediate induction of toxicity similar to that observed ollowing administration ofearly endotoxin tolerance. Previous studies demonstrated high doses of LPS. These data also defined the experimentalthat injection of high doses of either rIL-1 or rTNF resulted limits for subsequent experiments with respect to the rangein production of levels of CSF comparable to those induced of usable cytokine dosage combinations; i.e., a range of 500by 25 tg of LPS. i.e., the dose used for induction of early to 2,000 U of rIL-1 per mouse could be used in combinationendotoxin tolerance (11). Therefore, the doses of rlL-1 and with 5 or 7.5 ". of rTNF to achieve lethality consistentlyrTNF chosen for preliminary studies bracketed those used below 10%.previously. Figure 1 illustrates the toxicity of these two Another toxic manifestation associated with injection ofcytokines over a very broad dose range when administered LP! is induction of weight loss, which is maximal at 48 to 72individually or in combination. The dose of rTNF injected is h postwjection (27). In addition to the lethality observed inshown along the x axis, and the dose of rIL-1 injected is response to combinations of rIL-1 and rTNF (Fig. 1), weightindicated by the bar graph legends. The percentage of deaths loss was measured 3 days after injection of sublethal doses ofis plotted on the y axis. Injection of high doses of rTNF (up rlL-1 and rTNF administered alone or in combination. At 3to 2 x 10 U per mouse; 10 p4) alone occasionally resulted days after injection, 1,000 or 2,000 U of rIL-1 resulted inin deaths (maximal toxicity never exceeded 3%). Injection of !4.1% weight loss, and injection of rTNF (5 or 7.5 l.g)'rlL-1 alone (up to 15,000 U per mouse; -3 W was never resulted in s6,9% weight loss when compared with saline-lethal for mice. However, when a7.5 p.g of rTNF was injected controls (Fig. 2). However, in combination, rIL-1injected in combination with increasing doses of rIL-1, plus rTNF synergized to induce weight losses in mice whichdeaths (up to 100%) were observed in a dose-dependent ranged from 17.2 to 27.3%, similar to or exceeding thatfashion. In 10 separate experiments, 25 ".g of LPS led to induced by a tolerance-inducing dose (25 IJ.g) of LPS (18.6 tdeaths in only 3.1% of the mice, consistent with our previous 2.1%).estimates of the 50% lethal dose for this particular endotoxin Effect a adrnalntatio of r[L-1 or rTNF or both on day 0preparation Q1, 26). On the basis of a recent report by oe ability to respon to LPS on day 3. In early endotoxinRothstein and Schreiber (22) which showed that rTNF tolerance models established previously in sevei-.. laborato-synergized with LPS to induce deaths, we calculated the ries (11, 31), 25 "ag of LPS was found to induce a state ofconcentrations of contaminating LPS injected along with tolerance to subsequent injection with LPS 3 days later, aseach cytokine. For the maximum dose of rTNF injected assessed by the decreased capacity to produce CSF 6 h after(i.e., 10 lg), <0.001 ng of LPS was injected. For the endotoxin challenge. Since a single nonlethal injcction ofmaximum dose of rlL-1 injected (i.e., 15,000 U), 0.017 ng of LPS has been shown to induce both IL-1 and TNF shortlyLPS was injected. Thus, even with the largest dose combi- after administration, the capacity of these two cytokines tonation (i.e., 10 l.g of rTNF and 15,000 U of rlL-1), only 0.018 induce a state of early endotoxin tolerance was tested.ng of LPS was injected. This amount of LPS is >1.000-fold Figure 3 illustrates the capacity of mice injected on day 0 toless than that required by Rothstein and Schreiber to induce respond to LPS on day 3 by producing CSF. As reporteda minimal level of synergy (resulting in death) with 10 Ig of previously, the ability of mice treated on day 0 with LPS
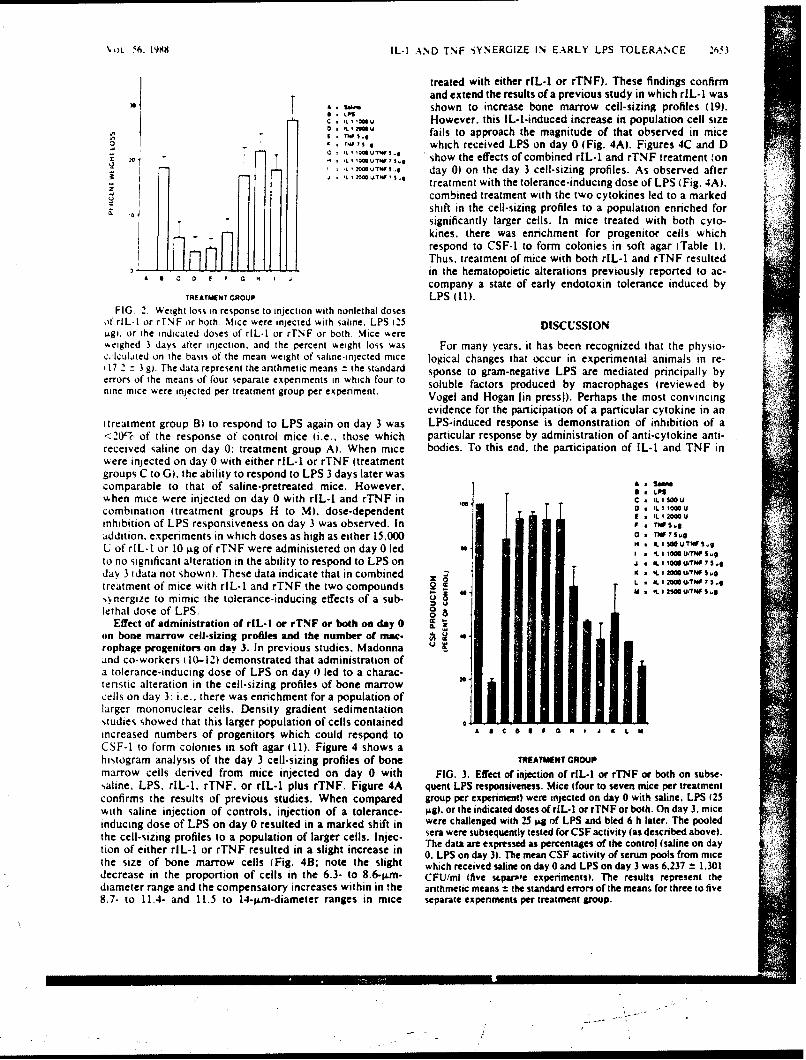
VOL. '6. 1998 IL-I AND TNF -5YNERGIZE IN EARLY LPS TOLERANCE 263
treated with either rIL-1 or rTNF). These findings confirmand extend the results of a previous study in which rIL-I was
sA - shown to increase bone marrow cell-sizing profiles (19).CI *Oi u However. this IL-i-induced increase in population cell size
o ,imU fails to approach the magnitude of that observed in miceE TI~"5.0S. •,,,which received LPS on day 0 (Fig. 4A). Figures 4C and D
I 1 ' * IS,, show the effects of combined rIL-1 and rTNF treatment (on=- J~~l -i .... .... 2=U., I..$.... day 0) on the day 3 cell-sizing profiles. As observed after "i.-L 1 2000 U.TFI 1 3 ,
treatment with the tolerance-inducing dose of LPS (Fig. 4A).
u combined treatment with the two cytokines led to a markedshift in the cell-sizing profiles to a population enriched forsignificantly larger cells. In mice treated with both cyto-kines, there was enrichment for progenitor cells whichrespond to CSF-1 to form colonies in soft agar ITable 1).Thus. treatment of mice with both rIL-1 and rTNF resulted
A a C in the hematopoictic alterations previously reported to ac-company a state of early endotoxin tolerance induced by
TREATMENT GROUP LPS (11).FIG. 2. Weight loss in response to injection with nonlethal doses
ot rlL-I or rlrNF or both. Mice were injected with saline. LPS f25 DISCUSSIONL.g. or the indicated doses of rtL-1 or rTNF or both. Mice wereweighed 3 days after injection, and the percent weight loss was For many years. it has been recognized that the physio-c lculated on the basis of the mean weight of saline-injected mice logical changes that occur in experimental animals in re-
17 2 - 3 g). The data represent the anthmetic means = the standard sponse to gram-negative LPS are mediated principally byerrors of the means of four separate experiments in which four to soluble factors produced by macrophages (reviewed bynine mice were injected per treatment group per experiment. Vogel and Hogan [n pressi). Perhaps the most convincing
evidence for the participation of a particular cytokine in anItreatment group B) to respond to LPS again on day 3 was LPS-induced response is demonstration of inhibition of a<20% of the response of control mice (i.e., those which particular response by administration of anti-cytokine anti-received saline on day 0: treatment group A). When mice bodies. To this end, the participation of IL-I and TNF inwere injected on day 0 with either rlL-1 or rTNF (treatmentgroups C to G), the ability to respond to LPS 3 days later wascomparable to that of saline-pretreated mice. However. A Salim
when mice were injected on day 0 with rIL-1 and rTNF in ,J * S.,0u
combination (treatment groups H to M). dose-dependent L 200U
inhibition of LPS responsiveness on day 3 was observed. In * 1.0.addition. experiments in which doses as high as either 15.000 G • 3 .9
U of rIL-1 or 10 lag of rTNF were administered on day 0 led IL 1 ,* U-70S.9I IL 1 1000l UITW~ 5 .
to no significant alteration in the ability to respond to LPS on a IL 1,io0U/rnv5 ,@day 3 1data not shownl. These data indicate that in combined - = N 1,9
treatment of mice with rlL-1 and rTNF the two compounds 2 L * It 1200 uTNs.
-,nergize to mimic the tolerance-inducing effects of a sub- M 1 urf, s e
lethal dose of LPS. 0Effect of administration of rIL-l or rTNF or both on day 0 C
on bone marrow cell-sizing proflies and the number of mac- .rophage progenitors on day 3. In previous studies. Madonna Uand co-workers (10-12) demonstrated that administration ofa tolerance-inducing dose of LPS on day i) led to a charac-tenstic alteration in the cell-sizing profiles of bone marrowcells on day 3. i.e.. there was enrichment for a population oflarger mononuclear cells. Density gradient sedimentationstudies showed that this larger population of cells containedincreased numbers of progenitors which could respond to A a C 0 It a J L M
CSF-1 to form colonies in soft agar (11). Figure 4 shows ahistogram analysis of the day 3 cell-sizing profiles of bone TREATMENT GROUPmarrow cells derived from mice injected on day 0 with FIG. 3. Effect of injection of rIL-1 or rTNF or both on subse-saline, LPS, rIL-I. rTNF. or rlL-1 plus rTNF. Figure 4A quent LPS responsiveness. Mice (four to seven mice per treatmentconfirms the results of previous studies. When compared group per experiment) were injected on day 0 with saline. LPS (25with saline injection of controls, injection of a tolerance- gg), or the indicated doses of rIL-1 or rTNF or both. On day 3. miceinducing dose of LPS on day 0 resulted in a marked shift in were challenged with 25 Ag of LPS and bled 6 h later. The pooled
the cell-sizing profiles to a population of larger cells. injec- sera were subsequently tested for CSF activity (as described above).The data are expressed as percentages of the control (saline on daytion of either rlL-1 or rTNF resulted in a slight increase in 0 LPS on day 3). The mean CSF activity of serum pools from mice
the size of bone marrow cells (Fig. 48; note the slight which received saline on day 0 and LPS on day 3 was 6.237 ± 1.301decrease in the proportion of cells in the 6.3- to 8.6-eIm- CFU/ml (five sLparple experiments). The results represent thediameter range and the compensatory increases within in the arithmetic means ± the standard errors of the means for three to five8.7- to 11.4- and 11.5 to 14-;&m-diameter ranges in mice separate experiments per treatment group.
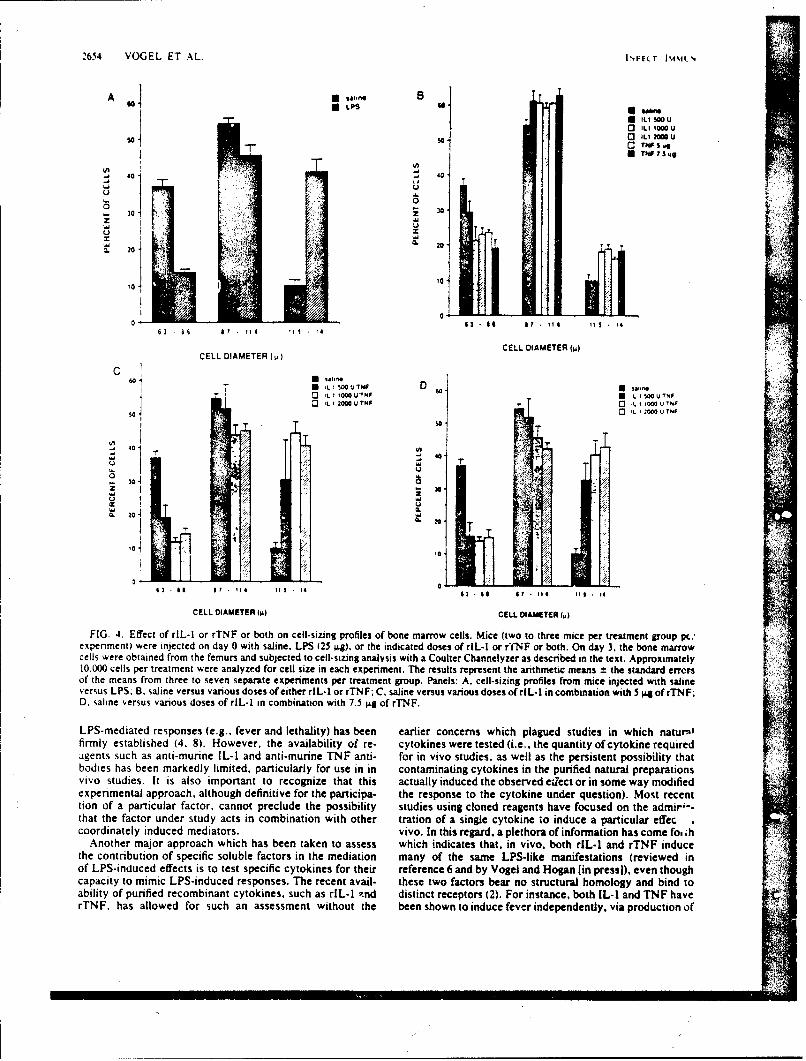
2654 VOGEL ET AL. I\FE.T l i N',
A T.lo
Sa u~
01 so C TN 5 u
(I
40 "J 40I~ "J
1 -i
.1 0
30 Z 30
Uw
~. 20 A 20
0 S 1• I 17 • 11 11 43 a s It 114 Ili 14
CELL DIAMETER (4)CELL DIAMETER (L)
C 60 D ,,60 U .LISOOUT4F D 6j•san
IL 1 00 VTNFU :LI:,500 U7NFo Ll 2000UTNF C3 LI 00QU'6"NFC3 IL 1 2000 U TNF'
00
Z 30UU
U 44o 304
I.-.
,oto
i 14 16 I 14 63 of 1 Ie 11 .14
CELL DIAMETER (l CELL DIAMETER (w)
FIG. 4. Effect of rlL-l or rTNF or both on cell-sizing profiles of bone marrow cells. Mice (two to three mice per treatment group pc.expenment) were injected on day 0 with saline. LPS (25 4g), or the indicated doses o r[L-1 or rfNF or both. On day 3. the bone marrowcells were obtained from the femurs and subjected to cell-sizing analysis with a Coulter Channelyzer as described in the text. Approximately10.000 cells per treatment were analyzed for cell size in each experiment. The results represent the arithmetic means = the standard errorsof the means from three to seven separate experiments per treatment group. Panels: A. cell-sizing profiles from mice injected with salineversus LPS; B. saline versus various doses of either rlL-1 or rTNF; C, saline versus various doses of rlL-1 in combination with 5 ig of rTNF:D. saline versus various doses of rlL-1 in combination with 7.5 gg of rTNF.
LPS-mediated responses (e.g., fever and lethality) has been earlier concerns which plagued studies in which naturaklfirmly established (4. 8). However, the availability of re- cytokines were tested (i.e.. the quantity of cytokine requiredagents such as anti-murine IL-I and anti-murine TNF anti- for in vivo studies, as well as the persistent possibility thatbodies has been markedly limited, particularly for use in in contaminating cytokines in the purified natural preparationsvivo studies. It is also important to recognize that this actually induced the observed eirect or in some way modifiedexperimental approach, although definitive for the participa- the response to the cytokine under question). Most recenttion of a particular factor, cannot preclude the possibility studies using cloned reagents have focused on the admir"--that the factor under study acts in combination with other tration of a single cytokine to induce a particular effeccoordinately induced mediators. vivo. In this regard, a plethora of information has come fo. ih
Another major approach which has been taken to assess which indicates that, in vivo. both rIL-1 and rTNF inducethe contribution of specific soluble factors in the mediation many of the same LPS-like manifestations (reviewed inof LPS-induced effects is to test specific cytokines for their reference 6 and by Vogel and Hogan [in pressi), even thoughcapacity to mimic LPS-induced responses. The recent avail- these two factors bear no structural homology and bind toability of purified recombinant cytokines, such as rIL-1 -.nd distinct receptors (2). For instance, both IL-I and TNF haverTNF, has allowed for such an assessment without the been shown to induce fever independently, via production of

P"
VOL. 56. 1988 IL-I AND TNF SYNERGIZE IN EARLY LPS TOLERANCE 2655
TABLE 1. Effect of rlL-! or rTNF or both on the numbers of ducing combinations of rIL-1 and rTNF also led to theCSF-1-responsive progenitors in bone marrow" hematopoietic alterations shown previously to be associated
Mean ItSEMI no. of CSF-I with LPS-induced early endotoxin tolerance (11). i.e., anTreatment (amt) prollettorm 10 bone marrow increase in the size of bone marrow cells (Fig. 4) with
cells 1P vaJue? concurrent enrichment for macrophage precursors (Table 1).Saline ............................................... 47 = 1.5 W ith respect to the latter, it is interesl ng that injection of 7.5rIL-I (1.000 U) ................................... 37 = 10.8(0.334) gofrTNFalone. butnot5 i.gofr.NF, led toa statisticallyrTNF (5 .g) ...................................... 52 = 7.8 (0.754) .Jgnif.:ant increase in the number of macrophage progenitorsrTNF (7.5 Lg) .................................... 67 = 3.5 (0.010) in the bone marrow. This may reflect the fact that high dosesrlL-I (i,000 U)-rTN F 1_ IS ) ................ 89 = 5 (0.002) ofTNF administered in vivo have been shown to induce lL-1rlL-1 1.000 U).-4TNF (.5 i.1) ..............121 ± 1.9 (0) production (8). Thus, rTNF-induced IL-1 may act synergis-
Bone marrow cells were denved onday3from miceltwotothree mice per tically with administered rTNF to increase the number oftreatment group per experiment) which had received saline or the indicated CSF-1-responsive progenitors.dose of rIL.! or rTNF or both on day 0. The results are expressed as the Taken collectively, these findings suggest that, in vivo.number of CSF-1-responsive progenitors per 10' bone marrow cells. The LPS-induced early endotoxin tolerance is mediated b,' theresults represent three separate experiments.
'An unpaired, two-tailed Student t test was used. synergistic action of LPS-induced IL-I and TNF. For mostf the studies performed, combined administration of IL-1
and TNF has been shown to result in an additive effect;prostaglandins (8). In this regard, they are both classical however, a precedent for synergy between these two factors"'endogenous pyrogens." In addition, at high doses, rTNF exists. For example. Beck et al. (1) showed that intradermalwas shown to induce IL-1 in vivo (8). This particular injection of natural IL-I. like LPS, led to attraction ofexample provides a good illustration of both cytokine redun- 'Cr-la! .[ed neutrophils to the site of injection. Movat et al.dancy (in the sense that two distinct cytokines can induce (17) confirmea these fir;ings by using rIL-1 and extendedthe same biological effect) and the potential for an inductiv. :hem by demonstrating that the combined action of riL-1 andcascade which, in turn, could prolong a given manifestation rTNF was synergistic in pruducing infiltration. Similar find-
in vivo. Injection of either rlL-1 or rTNF has been demon- ings were recently reported by Wankowicz et al. (30). Movatstrated in vivo to induce many of the same physiologic et al. (16) also demonstrated that 'ntradermal injection ofalterations set:n in response to LPS in addition to fever, such rIL-t and rTNF led to synergistic induction of a localas hypoglycemia, shock and death, increased resistance to Schwartzman reaction in rabbits challenged intravenouslyinfection. radioprotection. resistance to neoplasia, induction with endotoxin 18 h later. Neta et al. (18) have shown that atof CSF and late acute-phase reactants, and others (reviewed doses of radiation whi-.h cinsely approach 100% lethality,by Vogel and Hogan [in press]). the radioprotective effects of rIL-1 and rTNF are additive;
In this study, we sought to determine whether induction of however, at higher levels of irradiation, the radioprotectiorearly endotoxin tole-ance by LPS is mediated by soluble afforded by combined rlL-1 and rTNF injection was muchfactors produced in response to the tolerance-inducing injec- greater than would be pret;icted by summing the protectiontion (i.e.. the sublethal injection of LPS given on day 0 to afforded by injection of the cytokines individually. Forinduce a state of resistance to a subsequent challenge 3 to 4 induction of certain acute-phase reactants, such as serumdays later). In these studies, it was shown that neither rIL-1 amyloid P, simultaneous administratior of rIL-1 and rTNFnor rTNF injected individually (at doses which induced resulted in an additive response, however, combined admin-levels of circulating CSF in vivo comparable to that induced istration of these two cytokines led to synergistic inductionby a tolerance-inducing injection of LPS) induced a state of of fibnnogen (15). Recently. it was pointed out that IL-I andearly endotoxin tolerance. This was assessed by production TNF are strongly synergistic for generation of hypotensionof normal levels of CSF in response to LPS administered 3 and the capillary leak syndrome (6. 20). In studies performeddays later. Since IL-i and TNF are coordinately produced in with C3H/HeJ mice, synergistic protection from infection byresponse to LPS, we hypothesized that induction of toler- E. coli was afforded by treatment of mice with a combinedance depends upon the simultaneous presence of both solu- rlL-I and rTNF regimen (G. Saydoff. personal communica-ble factors. When adminisiered individually, rIL-1 and rTNF tion).rarely induced overt signs of toxicity; however, simulta- The mechanisms by which LPS induces a transient rever-neous administration of high doses of the two cytokines sal of sensitivity to homologous or heterologous challengeresulted in frank toxicity and often death. It is highly with LPS. i.e.. early endotoxin tolerance, are not very wellunlikely that this is due to synergy between rTNF and understood. Early studies in which peritoneal macrophagescontaminating LPS, since the maximum amount of contain- or Kupffer cells of animals rendered tolerant to endotoxininating LPS injected was >1.000-fold less than that shown were found to be refractory to stimulation with LPS in vitropreviously to induce minimal synergy with rTNF, resulting (7, 21) stron;ly suggested that tolerance was a function of ain death 122). Synergy between rTNF and rIL-1 was evident, failure to produce those soluble macrophage fac:ors, such aseven within a combined-dose range which rarely led to endogenous pyrogen or prostaglandins, shown previously todeaths, as measured by induction of weight loss greater than be associated with endotoxin-mediated toxicity. Subsequentor equivalent to that induced by 25 ixg of LPS. Within this studies by Williams et al. (31) strengthened the role ofsame sublethal-dose range, the two cytokines induced re- macrophages in the induction of tolerance by showing thatfractoriress to LPS challenge in a dose-dependent fashion injection of either a splenic adherent cell population or(Fig. 3). The amount of contaminating LPS injected with a peritoneal exudate macrophages along with splenic nonad-more typical tolerance-inducing dose combination (e.g., 7.5 herent cells was essential for overriding the inability toLg of rTNF and 2.000 U rIL-1) was <0.003 ng. Thus, it is produce CSF in recipients rendered tolerant to endotoxin.more likely that the synergistic toxicity observed by Roth- Lastly, in work by Madonna and co-workers (10-12). thestein and Schreiber (22) is due to LPS induction of IL-I. tolerance-inducing dose of LPS was followed by normalwhich in turn synergizes with rTNF. These tolerance-in- induction of macrophage-derived products, such as CSF and

2656 VOGEL ET AL. INFIEct. ISW4.
interferon, but resulted in a depressed capacity to produce 4. Studer, 1., 1. W. Milort. and A. C. Cerand. 1965. Passivethese same factors upon LPS challenge 3(0o4 days later. The immunization against cachectin-tumor necrosis factor protectskinetics of tolerance induction was correlated with acquisi- mice from lethal effect of endoloxin. Science 229-869.471.tion of a characteristic change in the cell-sizing pattern of the 5. CwwmI, W. A.. L J. Old, R. L. Kinud, S. Green. N. Fiore, amdbone maATOW towards larger cells. and within this increased S. Wllh1apu. 1975. An endotoxin induced seruim factor thatpopulation of larger cells a marked increase in the number of 367 ncos0o.mo.Po.Nal cd.S, SA7:66CSF-1-responsive macrophage progenitors was observed. It 6. DinaiwI.. C. A. 1988. Biobogy of interteukin-1. (Fed. Am. Soc.was therefore proposed that Ii) early endotoxin tolerance Exp. Biol. J. 2:108-115.results from a developmental blockade which results in 7. Dbiurtll. C. A.. P. T. Sodd, and E. Atkis. 1961. The role ofaccumulation of immature macrophages in the bone marrow the liver in the production of fever and in pyrogenic tolerance.which. in turn, limits the number of fully mature. LPS- Ttans. Assoc. Am. Physicians 81:334.responsive macrophages in the periphery or 60i initial expo- 8. D~fisarlKe C. A., J1. G. Canion. S. W. WoW, HI. A. De'mheim, 3.sure of mature macrophages to LPS renders them refractory Deuder, A. Ceran. 1. S. FIgal, M. A. Pallading, amid J. V.
of blckde r dwn O'Comi. 1986. Tumor necrosis factor (cachectin) is an endog-to subsequent stimulation because of a blcaeo on eos yo n nue production of interleukin 1. J. Exp.regulation of the LPS receptor. Med. 163:1433-1450.
The data presented in this report, that rlL-i and rTNF 9. Grknm S. E. 1913. Induction of endotoxin tolerance. p. 149-synergize to induce refractoriness to LPS, as well as the 178. In A. Nowotny fed.). Beneficial effects of endotoxins.hematopoietic changes reported previously. suggest that Plenum Publishing Corp.. New York.tolerance is induced in the absence of LPS and is mediated 10. Madonna G. S., J. E. Peterso. E. E. Rbil, andl S. N. Vogel.ndirectly, rather than by a mechanism which involves a 1986. Earty-phase endotoxin tolerance: induction by a detoxi-
blockade of the LPS receptor. Another possibility which tied lipid A derivative. monophosphoryl lipid A. Infect. Immun.might be invoked to explain the failure to respond to a 52:6-11.
chaleng inecton f LS i tht te iitil ijecionof1he . Madonnia, G. S., and S. N4. VogeL 1985. Early endotoxinchaleng inectin o LP isthattheiniialinjetio ofthe tolerance is associated with alterations in bone marrow-denivedtwo cytokines results in induction of a state of tachyphylaxis macrophage precursor pools. J. Immunrol. 135:3763-3771.to IL-i or TNF produced in response to the LPS challenge. 12. Madmna, G. S.. and S. N. Volell 1916. Induction of early-phaseIf this were so. then one would expect to observe diminished entdotoxin tolerance in athymic (nude) mice. B-cell-deficienttoxicity in the face of normal levels of circulating IL-i or (xi4 mice, and splenectomnized mice. Infect. Immun. 53:707-TNF produced in response to the LPS challenge. Prelimi- 710.nary experiments have indicated that, similar to the reduced 13. MaaueL D. N., R. N. Moore, and S. E. Mergenshapas. 1980.levels of CSF and interferon in circulation following LPS Endotoxin-induced tumor cvtotoxic factor. p. 141-143. In D.challenge, animals rendered endotoxin tolerant produced Schiessinger (ed.). Microbiology-198O. Ameencan Society forsignificantly less circulating TNF upon challenge than did 1.Microbiology. Washington, D.C.
conrolanmal (atano shwn) cnsitet wthearlier 1.Melate. F. C.. H. W. SlaerS, G. H. Barlow, R. ... Finley, radcontol nimls dat no shon),conistnt ithA. Y. Los. 1967. Chemcal. physical, and biological properties
findings (7. 21) that macrophages derived from animals of a lipopolysnechanide from &scherichia coil K235. Biochem-rendered tolerant produce depressed levels of soluble factors istry 4:2363-7372.in vitro (e.g.. endlogenous pyrogen and prostaglandins) when i5. Martessom, R. F., J. Shapiro. 3.-F. Urn,6 S. Domcea, and R.exposed to LPS. Thus, failure to produce adequate levels of Nets, 1918. Interaction of recombinat IL-i and recombinantcytokines is more likely to underlie the observed decrease in tumor necrosis factor in the induction of mouse acute phasetoxicity, rather than cytokine-induced tachyphylaxis to proteins. J. Immunol. 140-2260-2266.TNF. Confirmation of these conclusions will depend upon 16. Movie. H. Z., C. E. Durrowes, M. 1. Cyhaley, ased C. A.the availability of high-titered anti-murine IL-i and ani Dnarells, 1917. Role of complement. interleukin-1. and tumormurine TNF antibodies and the demonstration that injection necrosis factor in a local Shwartzman-like reaction. p. 69-78. In
of ethe regentabrgats iducton f LS-inuce toer- H. Movat led.). Leukocyte emipration and its sequelae. S.of ethe regentabrgats iducton f LS-inuce toer- Karger. Basel.
ance in vivo. As indicated above, the hematopoietic changes 17. Movie, H. Z., M. 1. Cybuhkty, 1. G. Cohldb, M. L. W. Chu. andobserved in response to a tolerance-inducing dose of LPS C. A. Dhiarel.. 1967. Acute inflammation in pram-negativeare only a correlate of early LPS-induced tolerance. The infection: central role of endotoxin. interleukin-i. tumor necro-demonstration that anti-IL-i or anti-TNF or both antibodies sis factor, andi the neutrophil leukocyte. Fed. Proc. 4L-97-104.also abrogate the hematopoictic responses to a tolerance- 18. Neta, L. J. J. Opaeh*W6 and S. D. Douhe. 1966. Interde-inducing dose of LPS would strengthen this relationship. pendeisce of the radioiprotective effiects of human reconibinant
inteileukin la. tumor necrosis factor a. grmnulocyte colony-ACKNOWLDGMENTSstimulating factor, and munine recombinant pnutlocyte-macro-ACKNOLEDGENTSphage coionry-stimulaiing factor. J. Immunol. 140:106-ill.
This work was supported by Uniformned Services University of 19. Neta, R., MI. 3. Szhim, J. J. Oppmenlnii S. G&k% and S. D.the Health Sciences protocol R07338 and the Armed Forves Radio- Domebe., 1967. The in vivo effects of nterleukin 1. 1. Bonebiology Research tnstitute. Defense Nuclear Agency, under re- marrow cells are induced to cycle after administration of inter-search work unit Mi 83148 (Uniformed Services University of the leukin 1.2J. Immunol. 130:.161-1666.Health Sciences protocol G27384). 20. O1tuawai S., .1. A. CGeinLd. 1 41iei L. 3. CosnI7, and
C. A. Dneef . 968. Interleukin I induces a shock-like state inLITERATURE CITED rabbits. Synergism with tumor necrosis factor and the effect of
1. Beck,. G.. G. S. Habdcht. J. L. Deaseb and F. Miller. 1986. cyclooxygenase inhibition. J. Clin. Invest. 81:1162-1172.Interleukin 1: a common endogenous mediator of inflammation 21. Rieahii L T., U. Sehada, 0. lduitai H. Folar, and 2. A.and the local Schwamsman. J. Immunol. 136:3025-3011. Pd. 1960. Prostaglandins in endotoxicosis, p. 66-72. In D.St Buder. 5., and A. Ceraml. 1915. Recombinant interieukin I Schiessinger led.). Microbiology-1900. Anmran Society forsuppresses lipoprotein ip~Ase activity in 3T3-L1 cells. J. Immu- Microbioloigy. Washington, D.C.net. 13S:3969.-3977. 22. Radmodai J. L., and I. S Abw 1977. Synergy between
3. Deuder, 3.. N. Kroebin, 1. Milork C. Lmuei, and A. Cerin. tumor necrosis factor and bacterial products cause hemor-1916. Control of cachectin (tumor necrosis factor) iynthesis: rhagi necrosis and lethal shock in normal miuce. Proc. NatI.mechanisms of endotoxin resistance. Science 232:977-960. Acrid. Sci. USA 115:607-611.

VOL. 56. 1968 IL-1 AND TNF SYNERGIZE IN EARLY LPS TOLERANCE 2657
23. Sueler, B. M., md G. W. Go . 1977. Characteristics of nective tissue, p. 225--234. In R. J. Genco and S. E. Mergenha-endotoxin.resiswint low-responder mice. p. 304-309. In D. geta (ed. Host-punaats interactions in peniodontal diseases.Schiessinger (ed.), Microbiology-1977. American Society tar Amencan Society tar Microbiology, Washigton, D.C.
-'MicrobWolgy Washington, D.C. 29. Wan. A. M., A. A. C iey, M. B. Lna, L. S. Lim. J.24. Urih. R. M.. R. K. Sbadducit. C. Sam&. and S. E. Mejo. Stricliar, I. N. VmnAndsel, L. Yonimoa. and D. F. Mark.
hogan. 1960. Colony-stimulating factor in nonspecific resistaince 1965. Molecular cloning of the complemientary DNA for humanand in increased susceptibility to endotoxin. p. 115-119. In D. tumor necrosis factor. Science 223:14%-154.Schiessinger (ed.). Microbiology-1980. American Society for 30. Wmnklowicit. Z., P. Megyais.ed A. JlueltuM 1968. SynergyMicrobiology. Washaington, D.C. between tumour necrosis factor m and interleukin-1 in the
25. Vogl. S. N.. S. D. Douches. E. N. Kanltsan. andl R. Nets. 1967. induction of polymorphonuclear leukocyte migration duringInduction of colony stimulating factor in vivo by recombinant inflammation. J. Leukocyte Biol. 43:349-356.interleulun la and tumor necrosis factor at. J. Immunoll. 13M 31. Wlflinms, Z., C. F. Hopg. and D. H. Ptuanok. 1963. Use of2143-2148. mice tolerant to lipopolysaccharide to demonstrate requirement
26. Vogel. S. N., K. E. Engilish. and A. D. O'Brien. 1962. Silica of cooperation between macrophages and lymphocytes to gen-enhancement of munne endotoxin sensitivity. Infect. Immun. erate lipopolysacharide-induced colony-stimulating factor in3111:681-685. vivo. Infect. Immun. 41:1-5.
27. Vogel, S. N.. R. N4. Mosee. J. D. Sip. and D. L. Roweh 32. Yasagnsir. J. S., andl W. R. Sdnebring. 1965. Interferon appear-1960. BCG-induced enhancement of endotoxin st isitivity in ance stimulaed by endotoxin, bacteria, or viruses in miceC3HIHeJ mice. I. In vivo studies. J. Immunol. 124:2004-2009. pre-treated with Eschericuia coli endoto:un or infected with
28. Wahl, S. 196'.. Mononuclear cell-mediated alterations in con- Mycobacterium tuberculosis. Nature (London) 208:456-458.





























