Embed Size (px)
Citation preview

http://dx.doi.org/10.1016/j.jmb.2012.08.007 J. Mol. Biol. (2012) 423, 624–631
Contents lists available at www.sciencedirect.com
Journal of Molecular Biologyj ourna l homepage: ht tp : / /ees .e lsev ie r.com. jmb
ArfA Recruits RF2 into Stalled Ribosomes
Yoshihiro Shimizu
Laboratory for Cell-Free Protein Synthesis, Quantitative Biology Center (QBiC), RIKEN, 2-2-3,Minatojima-minamimachi, Chuo-ku, Kobe, Hyogo 650‐0047, Japan
Received 1 June 2012;received in revised form2 August 2012;accepted 15 August 2012Available online21 August 2012
Edited by R. L. Gonzalez
Keywords:YaeJ;ArfA;YhdL;release factor;translation
E-mail address: [email protected] used: ArfA, alterna
factor; RF1, release factor 1; RF2, relrelease factor 3.
0022-2836/$ - see front matter © 2012 E
During translation in Escherichia coli, the ribosome rescue factor YaeJ and thealternative ribosome rescue factor (ArfA, previously called YhdL) canrelease stalled ribosomes from mRNA. Here, I used a reconstituted cell-freeprotein synthesis system to examine YaeJ- and ArfA-dependent recycling ofstalled ribosomes, in which mRNA lacks in-frame stop codons. It is shownthat YaeJ alone could recycle the ribosome but that ArfA required thepresence of release factor 2 (RF2). Furthermore, I show that RF2 binds tostalled ribosomes only in the presence of ArfA, demonstrating that ArfArecruits RF2 into the A site of the ribosome to facilitate peptidyl-tRNAhydrolysis. It is also demonstrated that the efficiency of the ArfA-dependentprocess decreases rapidly with an increase in mRNA length downstream ofthe A site of the ribosome whereas YaeJ function is maintained on mRNAwith sufficient length. From the results, I discuss differences of in vivo rolesof these two systems in addition to the well-known tmRNA-dependenttrans-translation system.
© 2012 Elsevier Ltd. All rights reserved.
Introduction
At the final step of translation, the terminationcodon moves to the A site of the ribosome and thesynthesized protein is released by peptidyl-tRNAhydrolysis. In bacteria, hydrolysis is catalyzed byclass 1 release factors including release factor 1(RF1) and release factor 2 (RF2) through a specificGGQ amino acid motif.1 Recent structural analysesusing co-crystals composed of the ribosome andclass 1 release factors have clearly demonstratedthe molecular mechanisms of this process at theatomic level.1–3
Occasionally, a translating ribosome encounterssituations related to aberrant protein release. Threepathways for handling these situations are known ineukaryotes: nonsense-mediated decay for mRNAsthat have premature stop codons, nonstop decay formRNAs that lack stop codons, and no-go decay for a
p.tive ribosome rescueease factor 2; RF3,
lsevier Ltd. All rights reserve
stalled ribosome during elongation.4 In all threepathways, class 1 or class 2 release factors or theirhomologs are involved. For example, eRF1 and eRF3mediate nonsense‐mediated decay,5 eRF3 homologSki7 mediates nonstop decay,6 and eRF1 homologDom34 and eRF3 homolog Hbs1 aid no‐go decay.7
These factors bind to the ribosome in aberrantsituations and are necessary for subsequent mRNAdegradation to rescue the ribosome for a new roundof protein synthesis.Stalled ribosome rescue mechanisms in eubacteria
are distinct from those in eukaryotes. The process oftrans-translation is a well-characterized systemwhere tmRNA, a hybrid transfer messenger RNA,bypasses the normal translation machinery.8,9 Inthis process, tmRNA recognizes a stalled ribosomeon mRNA that lacks a stop codon and is accom-modated in the A site of the ribosome as alanyl-tmRNA. After A site entry, translation is switchedfrom the incomplete mRNA to the open readingframe of tmRNA, which contains a stop codon. Thepeptide sequence encoded by the tmRNA andattached at the C-terminus of the synthesizedprotein acts as a proteolysis tag for the SspB–ClpXP protein degradation system.10 The stalledribosome is subsequently recycled for a new round
d.
625ArfA recruits RF2 into stalled ribosomes
of protein synthesis and the synthesized proteinsare degraded immediately, which contributes to themaintenance of the translation process and thequality of cellular proteins.Recently, two alternative pathways for handling
ribosomes in aberrant situations in Escherichia colihave been described. YaeJ, which contains a GGQmotif similar in structure to domain 3 of class 1release factors, has been reported to be associatedwith the ribosome and hydrolyze peptidyl-tRNA invitro.11 Additionally, alternative ribosome rescuefactor (ArfA, previously called YhdL), a smallprotein containing only 72 amino acids, has beenidentified as a factor required for E. coli growth inthe absence of the trans-translation system.12 Thisprotein associates with the ribosome and is neces-sary for peptidyl-tRNA hydrolysis.12,13 Those stud-ies have suggested that E. coli cells have multiplepathways for handling stalled ribosomes duringprotein synthesis.In the present study, I analyzed YaeJ and ArfA in a
reconstituted cell-free protein synthesis system. Ishow that these factors are able to recycle theribosome on mRNA without a stop codon. Howev-er, the presence of RF2 was found to be essential forthe ArfA-dependent, but not YaeJ-dependent, ribo-some recycling. Additionally, I show that RF2, butnot RF1, binds to the ribosome in the presence ofArfA. These studies suggest that ArfA acts as anadaptor protein for RF2 binding in the absence ofstop codons in the A site of the ribosome.
(a)
(b)
mRNA with a stop codon
5' -
5' -
mRNA with a stop codon:
mRNA without a stop codon:
Syn
thes
ized
pep
tide
( 1.2
1.0
0.8
0.6
0.4
0.2
0
mRNA without a stop codon
RF1, RF2, RF3, RRF
YaeJ
ArfA
+ --+ ++ +-- -
+ - + - ++-- --
-- -- -
1.4
1.6
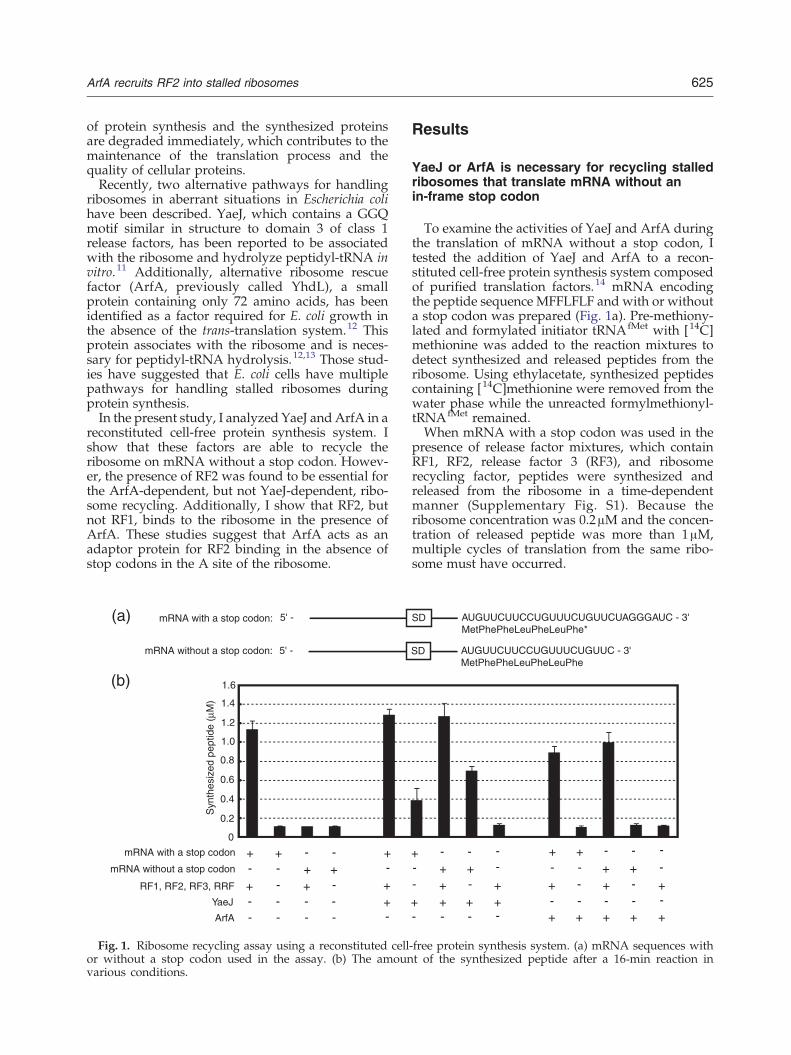
Fig. 1. Ribosome recycling assay using a reconstituted cellor without a stop codon used in the assay. (b) The amounvarious conditions.
Results
YaeJ or ArfA is necessary for recycling stalledribosomes that translate mRNA without anin-frame stop codon
To examine the activities of YaeJ and ArfA duringthe translation of mRNA without a stop codon, Itested the addition of YaeJ and ArfA to a recon-stituted cell-free protein synthesis system composedof purified translation factors.14 mRNA encodingthe peptide sequence MFFLFLF and with or withouta stop codon was prepared (Fig. 1a). Pre-methiony-lated and formylated initiator tRNAfMet with [14C]methionine was added to the reaction mixtures todetect synthesized and released peptides from theribosome. Using ethylacetate, synthesized peptidescontaining [14C]methionine were removed from thewater phase while the unreacted formylmethionyl-tRNAfMet remained.When mRNA with a stop codon was used in the
presence of release factor mixtures, which containRF1, RF2, release factor 3 (RF3), and ribosomerecycling factor, peptides were synthesized andreleased from the ribosome in a time-dependentmanner (Supplementary Fig. S1). Because theribosome concentration was 0.2μM and the concen-tration of released peptide was more than 1μM,multiple cycles of translation from the same ribo-some must have occurred.
AUGUUCUUCCUGUUUCUGUUCUAGGGAUC - 3' MetPhePheLeuPheLeuPhe*
SD
AUGUUCUUCCUGUUUCUGUUC - 3' MetPhePheLeuPheLeuPhe
SD
+ + +-- - -- -
+ +- - + +-- -- + - + + - + - ++ + + +
+ + + + +- -- --- -- -
-free protein synthesis system. (a) mRNA sequences witht of the synthesized peptide after a 16‐min reaction in
(a)
RF2
RF2 conc.
ArfA
1 2 3 4 5 6 7 8 9 10 11 12 13
626 ArfA recruits RF2 into stalled ribosomes
Next, I investigated the contribution of ArfA andYaeJ on peptide synthesis under various conditions(Fig. 1b). In the case of mRNA with a stop codon,the presence of release factor mixtures was neces-sary for efficient peptide synthesis. In the absence ofrelease factor mixtures, ArfA was not effective forthe peptide synthesis and the presence of YaeJresulted in only a small amount of peptidesynthesis. The peptide synthesis in the presence ofYaeJ may have been caused by YaeJ-mediatedrecycling of ribosomes stalled at the stop codon.In the case of the mRNA without a stop codon, thesynthesized peptide yield increased with YaeJ aloneand was further improved by the presence ofrelease factors. By contrast, peptide synthesis inthe presence of ArfA was inefficient unless releasefactors were present.
RF2 is necessary for ArfA-dependent stalledribosome rescue
These results agree with a previous finding inwhich the synthesized protein was released from thestalled complex by YaeJ, but not by ArfA.13 Takentogether, these data suggest that ArfA requires class1 release factors for peptidyl-tRNA hydrolysis andrelease of the synthesized protein. To determinewhich release factor is required, I measured synthe-sized peptide concentrations from ArfA-dependentrescued ribosomes in the presence of RF1 or RF2(Fig. 2). The inclusion of RF1 did not lead toaccumulation of a released peptide, whereas theaddition of RF2 produced a significant amount ofpeptide. This suggests that RF2 aids ArfA-depen-dent ribosomal rescue.
RF1
Syn
thes
ized
pep
tide
(µM
)
1.2
1.0
0.8
0.6
0.4
0.2
0
RF2
ArfA
+ --++-+ -
+ ++ -+
++
1.4
Fig. 2. Effect of RF1 and RF2 on ArfA-dependentribosome rescue. The amount of synthesized peptidefrom mRNA without a stop codon after a 16‐min reactionis shown.
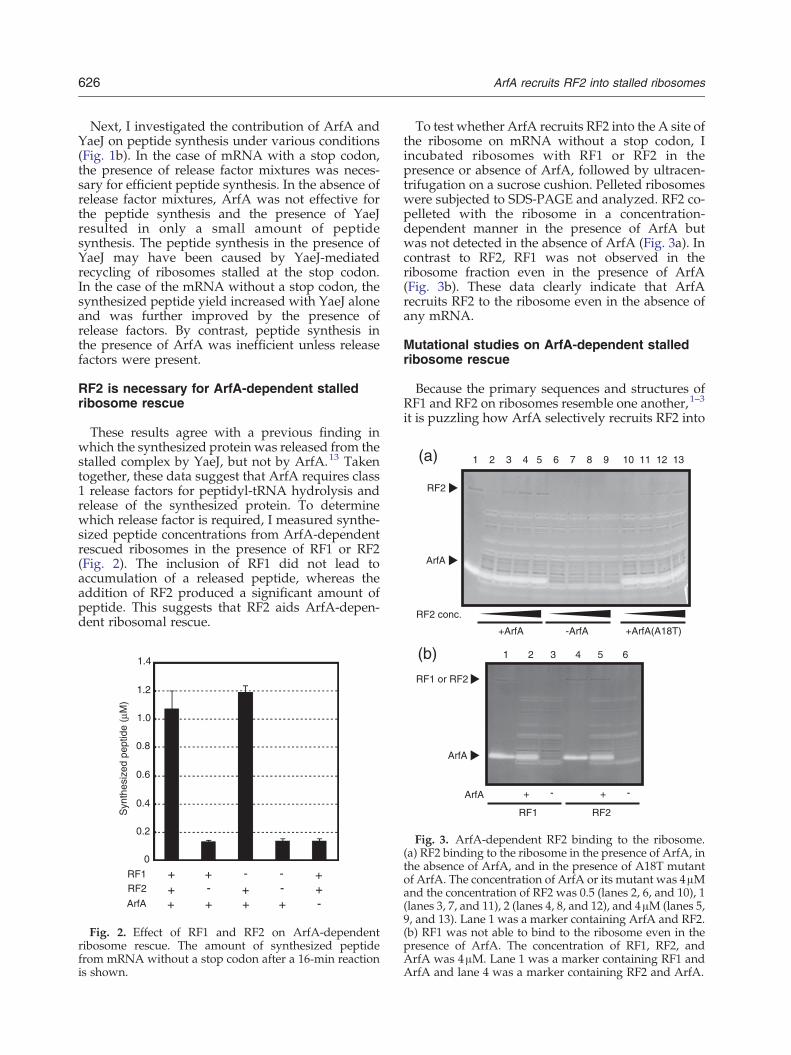
To test whether ArfA recruits RF2 into the A site ofthe ribosome on mRNA without a stop codon, Iincubated ribosomes with RF1 or RF2 in thepresence or absence of ArfA, followed by ultracen-trifugation on a sucrose cushion. Pelleted ribosomeswere subjected to SDS-PAGE and analyzed. RF2 co-pelleted with the ribosome in a concentration-dependent manner in the presence of ArfA butwas not detected in the absence of ArfA (Fig. 3a). Incontrast to RF2, RF1 was not observed in theribosome fraction even in the presence of ArfA(Fig. 3b). These data clearly indicate that ArfArecruits RF2 to the ribosome even in the absence ofany mRNA.
Mutational studies on ArfA-dependent stalledribosome rescue
Because the primary sequences and structures ofRF1 and RF2 on ribosomes resemble one another,1–3
it is puzzling how ArfA selectively recruits RF2 into
(b)
+ArfA -ArfA
RF1 or RF2
ArfA
RF1 RF2
ArfA + +- -
1 2 3 4 5 6
+ArfA(A18T)
Fig. 3. ArfA-dependent RF2 binding to the ribosome.(a) RF2 binding to the ribosome in the presence of ArfA, inthe absence of ArfA, and in the presence of A18T mutantof ArfA. The concentration of ArfA or its mutant was 4μMand the concentration of RF2 was 0.5 (lanes 2, 6, and 10), 1(lanes 3, 7, and 11), 2 (lanes 4, 8, and 12), and 4μM (lanes 5,9, and 13). Lane 1 was a marker containing ArfA and RF2.(b) RF1 was not able to bind to the ribosome even in thepresence of ArfA. The concentration of RF1, RF2, andArfA was 4μM. Lane 1 was a marker containing RF1 andArfA and lane 4 was a marker containing RF2 and ArfA.
627ArfA recruits RF2 into stalled ribosomes
the ribosome. To answer this question, I testedseveral RF1 and RF2 chimera proteins with theirdomains swapped.15,16 Since the domain 1 of RF2 islonger than that of RF1 and the sequence of domain1 is not well conserved between RF1 and RF2, Itested an RF1 variant in which domain 1 is swappedto that of RF2 but still functions as RF1. 16
Additionally, although ArfA interacts with the 50Ssubunit of the ribosome,12 the possibility remainsthat it discriminates between the PAT motif of RF1and the SPF motif of RF2, which both directlyinteract with the stop codon of mRNA in the Asite.1–3 In a hybrid protein with different combina-tion of domains of RF1 and RF2, it has been shownthat RF1 and RF2 function can be exchanged byswapping domain 4 (Supplementary Fig. S2a).15,16
A hybrid protein with domain 4 from RF1 (ΨRF1)functioned as RF1 and a hybrid protein withdomain 4 from RF2 (ΨRF2) functioned as RF2.Furthermore, in a hybrid protein (ΨRF) withdomain 4 that has an N-terminal sequence fromRF2 and a C-terminal sequence from RF1, functionexchange can be achieved by merely exchanging thePAT motif and the SPF motif in the middle cassetteregion (Supplementary Fig. S2a). Thus, I testedΨRF1, ΨRF2, and ΨRF with the SPF motif(ΨRFwSPF), but the purification of ΨRF2 wasunsuccessful because of aggregate formation. Un-fortunately, these mutant proteins were not able tobind to the ribosome even in the presence of ArfA(Supplementary Fig. S2b). Therefore, further muta-tional or structural studies are necessary to elucidatethe basis of the discrimination.It has been reported that A18T mutant of ArfA
cannot mediate peptidyl-tRNA hydrolysis on stalled
(a)
(b)
SD
Syn
thes
ized
pep
tide
(µM
)
1.2
1.0
0.8
0.6
0.4
0.2
0
(c
+0 +1 +2 +3 +6 +9 +15 +25
1.4YaeJ-dependent ribosome rescue
Fig. 4. Effect of mRNA downstream of the A site of the ribThe sequence of a series of mRNA used in the assay. (b and c)ribosome rescue, respectively. The amount of synthesized pep
ribosomes although it can bind to the ribosome.12,13
Because it seems that the mutant cannot recruit RF2into the ribosome, I tested the mutant. Unexpected-ly, the results showed that RF2 can bind to theribosome even in the presence of the mutant ArfA(Fig. 3a), whereas ribosome recycling is significantlyreduced (Supplementary Fig. S3). These resultssuggest that simply recruiting RF2 into the ribosomeis not sufficient for efficient peptidyl-tRNA hydro-lysis on stalled ribosomes. Instead, appropriatepositioning of RF2 in the A site of the ribosome byArfA or concerted action of RF2 and ArfA may benecessary for the ArfA-dependent peptidyl-tRNAhydrolysis on stalled ribosomes.
YaeJ, but not ArfA, can function despite theexistence of mRNA downstream of the A siteof the ribosome
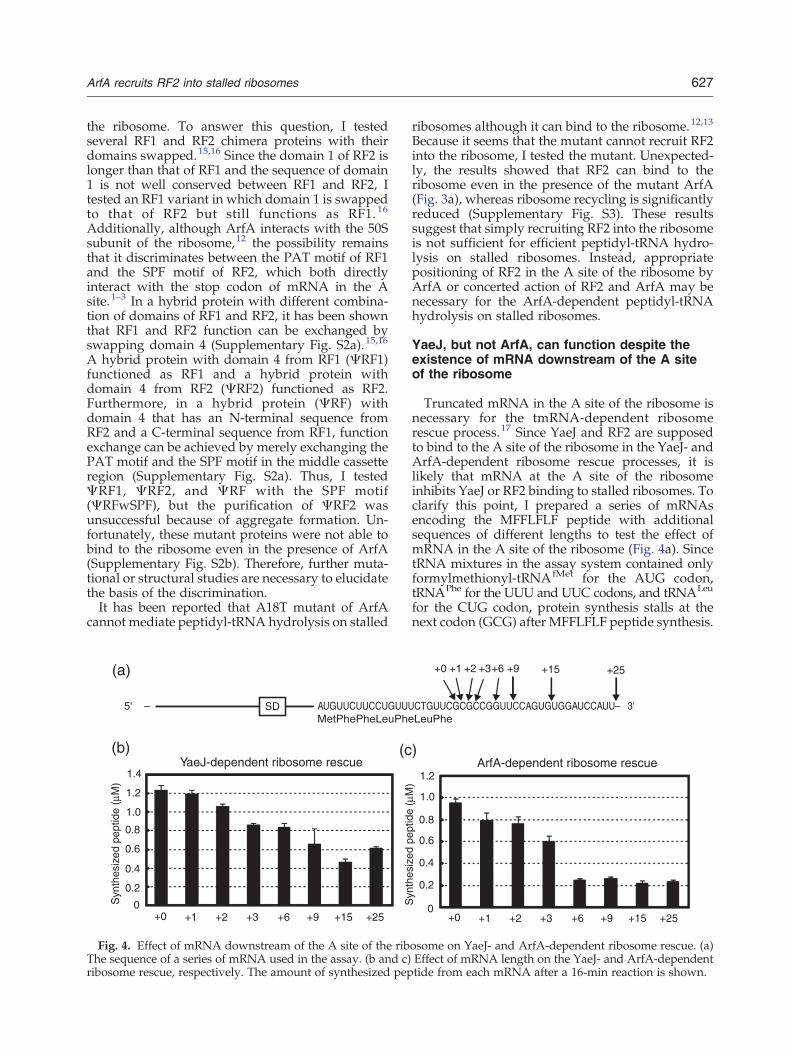
Truncated mRNA in the A site of the ribosome isnecessary for the tmRNA-dependent ribosomerescue process.17 Since YaeJ and RF2 are supposedto bind to the A site of the ribosome in the YaeJ- andArfA-dependent ribosome rescue processes, it islikely that mRNA at the A site of the ribosomeinhibits YaeJ or RF2 binding to stalled ribosomes. Toclarify this point, I prepared a series of mRNAsencoding the MFFLFLF peptide with additionalsequences of different lengths to test the effect ofmRNA in the A site of the ribosome (Fig. 4a). SincetRNA mixtures in the assay system contained onlyformylmethionyl-tRNAfMet for the AUG codon,tRNAPhe for the UUU and UUC codons, and tRNALeu
for the CUG codon, protein synthesis stalls at thenext codon (GCG) after MFFLFLF peptide synthesis.
+0 +1 +2 +3+6 +9 +15 +25
)
Syn
thes
ized
pep
tide
(µM
)
1.2
1.0
0.8
0.6
0.4
0.2
0+0 +1 +2 +3 +6 +9 +15 +25
ArfA-dependent ribosome rescue
osome on YaeJ- and ArfA-dependent ribosome rescue. (a)Effect of mRNA length on the YaeJ- and ArfA-dependenttide from each mRNA after a 16‐min reaction is shown.

628 ArfA recruits RF2 into stalled ribosomes
In mixtures containing YaeJ, the amount of thesynthesized peptide gradually decreased accordingto the length of mRNA downstream of the last UUCcodon (Fig. 4b). However, the concentration of thesynthesized peptide was higher than that of theribosome (0.2μM) even when the longest mRNAwas used, which suggests that YaeJ can facilitatemultiple ribosomal turnovers in this reaction. Bycontrast, in mixtures with ArfA, the amount ofsynthesized peptide decreased to background levelwhen mRNA contained more than six basesadditional sequence downstream of the last UUCcodon (Fig. 4c). These results suggest that mRNA inthe A site significantly inhibits ArfA-dependent RF2binding, whereas YaeJ can bind to the ribosome andfunction even when mRNA is lodged at the A site.
Discussion
In addition to the structuredN-terminal domain ofYaeJ, which contains a GGQ motif and resemblesdomain 3 of class 1 release factors, YaeJ possesses aC-terminal unstructured basic region. A previousbiochemical study showed that this C-terminalregion is essential for YaeJ binding to the ribosome.11
Recently, a crystal structure of E. coli YaeJ bound tothe Thermus thermophilus 70S ribosome in complexwith an initiator tRNAfMet and a short mRNAfragment revealed that the N-terminal domain ofYaeJ is located in the A site of the ribosome next tothe P-site tRNAfMet.18 The GGQ motif of YaeJ ispositioned in the peptidyl-transferase center adja-cent to the CCA end of tRNAfMet, suggesting that themotif is responsible for peptidyl-tRNA hydrolysis onthe ribosome. TheC-terminal region of YaeJ forms anα‐helix and is accommodated in the mRNA channeldownstream of the A site.In the present study, I show that YaeJ hydrolyzes
the peptidyl-tRNA and recycles ribosomes stalled atthe 3′-terminus end on mRNAs without a stopcodon (Fig. 1) and ribosomes stalled in the middle ofmRNAs absent of any corresponding tRNAs (Fig. 4).The latter result is in accordance with the previousbiochemical data demonstrating that YaeJ hydro-lyzes peptidyl-tRNA on the ribosome stalled by arare codon cluster in vitro.11 However, the efficiencyof the ribosome recycling decreases according to thelength of mRNA downstream of the A site (Fig. 4).Because the C-terminal region of YaeJ overlaps withthe mRNA path through the ribosome in the crystalstructure, the presence of mRNA downstream of theA site may occlude interaction between YaeJ and theribosome. Nevertheless, ribosomes stalled on longmRNAs were still recycled by YaeJ (Fig. 4). Thissuggests that mRNA does not completely inhibitYaeJ binding to the ribosome. Flexibility of mRNAmay allow YaeJ to avoid a steric clash when bindingto ribosomes stalled on mRNA.18
In contrast with YaeJ, although ArfA can bind tothe ribosome, it alone cannot hydrolyze peptidyl-tRNA on the ribosome.13 I demonstrate that RF2 isnecessary for ArfA-dependent peptidyl-tRNA hy-drolysis (Fig. 2) and that RF2 can bind to theribosome in the presence of ArfA (Fig. 3), presum-ably through direct interaction between ArfA andRF2 on the ribosome or other mechanisms such as astructural change in the ribosome due to ArfAbinding. These results strongly suggest that ArfAserves as an adaptor protein to recruit RF2 into the Asite of the ribosome in the absence of a stop codon inorder to hydrolyze the peptidyl-tRNA. Additional-ly, mRNA downstream of the A site significantlyinhibited the ribosome recycling, suggesting that thepresence of the codon in the A site of the ribosomeprevents RF2 binding to the ribosome (Fig. 4).Considering that RF2 entry into the A site of theribosome depends on the presence of UAA or UGAcodon in the A site, it is probable that inhibition iscaused by a mismatch between the sense codon ofthe mRNA and the SPF motif of RF2, which plays arole in stop codon recognition during normaltermination process.The primary sequences and structures of RF1 and
RF2 on ribosomes resemble one another.1–3 How-ever, there are some differences between twostructures. The main difference is seen in thepositioning of domain 1 of RF1 and RF2 in theribosome. The position of RF2 domain 1 is closer tothe A site than RF1 domain 1, placing it in contactwith the L11 stalk.2,3 Thus, it can be argued thatArfA recognizes this structural difference and that iswhy RF2 is selectively recruited into the ribosomeby ArfA. Another possibility is that ArfA mimics thecodon and discriminates between the PAT motif ofRF1 and the SPF motif of RF2, although this is lesslikely because ArfA is reported to interact with the50S subunit of the ribosome.12 Since neither the RF1variant with the RF2 domain 1 nor the chimeraprotein with the SPF motif is recruited into theribosome by ArfA (Supplementary Fig. S2), othermechanistic scenario should be considered. Furtherstudies are necessary to understand the basis ofdiscrimination by ArfA.A major difference between the YaeJ- and ArfA-
dependent ribosome rescue processes becomesapparent when the ribosome stalls in the middle ofmRNA during protein synthesis. The efficiency ofthe ArfA-dependent process decreases rapidly withan increase in mRNA length downstream of the Asite of the ribosome whereas YaeJ function ismaintained on mRNA with sufficient length(Fig. 4). This suggests that YaeJ may cover a widerrange of ribosomal situations than ArfA includingribosomes stalled by rare codon clusters.11 Howev-er, the broad specificity of YaeJ may disturb normaltranslation processes by unfavorable YaeJ binding tothe translating ribosome. By contrast, the ArfA-

629ArfA recruits RF2 into stalled ribosomes
dependent ribosome rescue system is less likely todisturb normal translation processes because RF2may act as a discriminating sensor of a translating orstalled ribosome.An analogous mechanism has been also identified
in the tmRNA-dependent trans-translation system.It has been shown by in vivo and in vitro experimentsthat the tagging reaction by tmRNA does not takeplace on a ribosome with mRNA downstream of theA site.17,19,20 The recently elucidated crystal struc-ture composed of a tmRNA fragment, SmpB,elongation factor Tu, and the ribosome well explainsthe basis for this mechanism.9 In this structure, theC-terminal region of SmpB extends into the spacenormally occupied by mRNA, leaving space forabout four or five nucleotides after the P site codon.Therefore, the presence of mRNA downstream ofthe A site of the ribosome would clash with the C-terminal region of SmpB, which suggests that thisregion functions as a discriminating sensor of atranslating or stalled ribosome.Thus, the roles of tmRNA- and ArfA-dependent
ribosome rescue systems are similar and comple-mentary strategies for cells to escape stalledtranslation. Interestingly, the expression of ArfAis tightly regulated by the system involving trans-translation. 21 This suggests that the ArfA-dependent system may play a backup role to thetmRNA-dependent system. Additionally, thetmRNA rescue system is linked to the SspB–ClpXPprotein degradation system for the maintenance ofthe quality of cellular proteins,10 which suggests amore prevailing role for the tmRNA rescue systemin the translation maintenance.In contrast to these two systems, YaeJ can rescue
stalled ribosomes with mRNA present downstreamof the A site, albeit with decreased efficiency.Therefore, the YaeJ system may be a third way incells for handling ribosomal situations that are notcovered by ArfA and tmRNA systems. However, thepresence of YaeJ may occasionally be harmful to thecells because normal translation processes can bedisturbed. Overexpression of YaeJ has been reportedto suppress the lethal phenotype of the ssrA arfAdouble mutant whereas the normal expression levelof YaeJ is not sufficient to suppress it.13 Additionally,recent proteome analyses with single-molecule sen-sitivity in single cells reported the expression level ofYaeJ and SmpB as 0.526 and 14.608 protein mole-cules/cell, respectively.22 These observations areconsistent with the putative role of the YaeJ system.
Materials and Methods
Preparation of protein and RNA samples
The components used in the reconstituted cell-freeprotein synthesis system were purified as previously
described.23 E. coli tRNALeu, tRNAPhe, and the initiatortRNAfMet were transcribed using T7 RNA polymerasefollowed by purification with anion‐exchange chroma-tography. The genes encoding YaeJ and ArfA with a His-tag at their N-terminus were cloned into pET16b (MerckMillipore) and transformed into an E. coli BL21/DE3strain. Overexpression and purification of YaeJ and ArfAwere carried out similarly to protein components for thecell-free protein synthesis system.23 The 12 C-terminalamino acid residues of ArfA were removed becausethese residues result in poor expression and theirdeletion does not affect ArfA activity.12 The plasmidsencoding A18T mutant of ArfA and several mutants ofclass 1 release factors (RF1 or RF2) were generated byusing PfuUltra II Fusion HS DNA Polymerase (AgilentTechnologies) with appropriate DNA primers andpurified as described above.
In vitro translation experiments
DNA templates used in the reconstituted cell-freeprotein synthesis system were prepared as follows: Byusing pURE1 vector (BioComber) containing a T7 promot-er sequence and the ribosome binding site (Shine–Dalgarno sequence) as the template, DNA templateswere amplified by PCR with the T7 promoter primer 5′-GCGCGTAATACGACTCACTATAG-3′ and appropriatereverse primers. The amplified DNA templates werepurified by Wizard SV Gel and PCR Clean-Up System(Promega). Pre-methionylated and formylated initiatortRNAfMet with [14C]methionine were prepared as follows:Reaction mixtures (250μl) containing 50mMHepes–KOH,pH7.6, 100mM potassium glutamate, 13mM magnesiumacetate, 2mM spermidine, 1mM DTT, 1mM ATP, 10mg/mL 10-formyl-5,6,7,8-tetrahydrofolic acid, 12.5μCi [14C]methionine (American Radiolabeled Chemicals, Inc.),1600pmol initiator tRNAfMet, and sufficient amounts ofmethionyl-tRNA formyltransferase and methionyl-tRNAsynthetase were incubated at 37 °C for 20min. Afterincubation, tRNA was purified and precipitated by theacidic phenol extraction and isopropanol precipitation.The precipitated formylmethionyl-tRNAfMet (fMet-tRNAfMet)was dissolved in water and immediately used in thefollowing translation experiments: Reaction mixtures(50μl) containing 50mM Hepes–KOH, pH7.6, 100mMpotassium glutamate, 13mM magnesium acetate, 2mMspermidine, 1mMDTT, 2mMATP, 2mMGTP, 1mMUTP,1mM CTP, 10mM creatine phosphate, 0.1mM phenylal-anine, 0.1mM leucine, 100 pmol tRNALeu, 100 pmoltRNAPhe, 0.2μM ribosome, 4μg/mL leucyl-tRNA synthe-tase, 16.5μg/mL phenylalanyl-tRNA synthetase, 10μg/mL initiation factor 1, 40μg/mL initiation factor 2, 10μg/mL initiation factor 3, 50μg/mL elongation factor G,100μg/mL elongation factor Tu, 50μg/mL elongationfactor Ts, 10μg/mL RF1, 10μg/mL RF2, 10μg/mL RF3,10μg/mL ribosome recycling factor, 10μg/mL YaeJ,10μg/mL ArfA, 10μg/mLT7 RNA polymerase, 1.1μg/mL nucleotide diphosphate kinase, 3μg/mL myokinase,4μg/mL creatine kinase, 1μg/mL pyrophosphatase,108μg/mL PrlC, and 0.5 pmol DNA template were pre-incubated at 37°C for 5min. It should be noted that E. colioligopeptidase A (PrlC) was added to the reactionmixtures to digest and dissolve the synthesized peptidein the ethyl acetate efficiently. The peptide synthesis

630 ArfA recruits RF2 into stalled ribosomes
reactionwas started by the addition of 200 pmol [14C]fMet-tRNAfMet into the reaction. At specified times, aliquots(10μl) were withdrawn and mixed with 10μl 1N HCl toterminate the reaction. The peptide containing [14C]methionine was extracted with ethyl acetate and radioac-tivity was measured with a liquid scintillation counter.
Ribosome binding analysis of class 1 release factors
Reaction mixtures (50μl) containing 50mM Hepes–KOH, pH7.6, 100mM potassium glutamate, 5mMmagnesium acetate, 2mM spermidine, 1mM DTT, 1μMribosome, and specific amounts of ArfA and RF1 or RF2were incubated at 37°C for 10min. After incubation, themixtures were loaded onto 300μl of a 20% sucrose buffer(10mM Hepes–KOH, pH7.6, 60mM ammonium chlo-ride, 5mM magnesium acetate, 20% sucrose, and 7mM2-mercaptoethanol), followed by ultracentrifugation at60,000rpm (162,000g) for 3h using an S140AT rotor(Hitachi). The supernatant was discarded and thepelleted ribosome was dissolved in 100μl of water. Theresultant samples (3μl) were subjected to 15% SDS-PAGE, stained with InVision His-tag In-gel Stain(Invitrogen), and visualized by Typhoon FLA 7000 (GEHealthcare).
Acknowledgements
I thank Dr. Hiroki R. Ueda for helpful discussionsand Dr. ArthurMillius and Dr. Peter Karagiannis forcritically reading the manuscript. I am thankful tothe RIKEN Center for Developmental Biology (CDB)for the use of the bio-imaging analyzer and in-struments related to the radioisotope work. Thiswork was supported by the RIKEN QuantitativeBiology Center.
Supplementary Data
Supplementary data to this article can be foundonline at http://dx.doi.org/10.1016/j.jmb.2012.08.007
References
1. Korostelev, A. A. (2011). Structural aspects of trans-lation termination on the ribosome. RNA, 17,1409–1421.
2. Laurberg, M., Asahara, H., Korostelev, A., Zhu, J.,Trakhanov, S. & Noller, H. F. (2008). Structural basisfor translation termination on the 70S ribosome.Nature, 454, 852–857.
3. Korostelev, A., Asahara, H., Lancaster, L., Laurberg,M., Hirschi, A., Zhu, J. et al. (2008). Crystal structureof a translation termination complex formed withrelease factor RF2. Proc. Natl Acad. Sci. USA, 105,19684–19689.
4. Atkinson, G. C., Baldauf, S. L. & Hauryliuk, V. (2008).Evolution of nonstop, no-go and nonsense-mediatedmRNA decay and their termination factor-derivedcomponents. BMC Evol. Biol. 8, 290.
5. Isken, O. & Maquat, L. E. (2007). Quality control ofeukaryotic mRNA: safeguarding cells from abnormalmRNA function. Genes Dev. 21, 1833–1856.
6. van Hoof, A., Frischmeyer, P. A., Dietz, H. C. &Parker, R. (2002). Exosome-mediated recognition anddegradation of mRNAs lacking a termination codon.Science, 295, 2262–2264.
7. Becker, T., Armache, J. P., Jarasch, A., Anger, A. M.,Villa, E., Sieber, H. et al. (2011). Structure of the no-gomRNA decay complex Dom34-Hbs1 bound to astalled 80S ribosome. Nat. Struct. Mol. Biol. 18,715–720.
8. Dulebohn, D., Choy, J., Sundermeier, T., Okan, N. &Karzai, A. W. (2007). Trans-translation: the tmRNA-mediated surveillance mechanism for ribosome res-cue, directed protein degradation, and nonstopmRNA decay. Biochemistry, 46, 4681–4693.
9. Neubauer, C., Gillet, R., Kelley, A. C. & Ramakrishnan,V. (2012).Decoding in the absence of a codon by tmRNAand SmpB in the ribosome. Science, 335, 1366–1369.
10. Levchenko, I., Seidel, M., Sauer, R. T. & Baker, T. A.(2000). A specificity-enhancing factor for the ClpXPdegradation machine. Science, 289, 2354–2356.
11. Handa, Y., Inaho, N. & Nameki, N. (2011). YaeJ is anovel ribosome-associated protein in Escherichia colithat can hydrolyze peptidyl-tRNA on stalled ribo-somes. Nucleic Acids Res. 39, 1739–1748.
12. Chadani, Y., Ono, K., Ozawa, S., Takahashi, Y., Takai,K., Nanamiya, H. et al. (2010). Ribosome rescue byEscherichia coli ArfA (YhdL) in the absence of trans-translation system. Mol. Microbiol. 78, 796–808.
13. Chadani, Y., Ono, K., Kutsukake, K. & Abo, T. (2011).Escherichia coli YaeJ protein mediates a novel ribo-some-rescue pathway distinct from SsrA- and ArfA-mediated pathways. Mol. Microbiol. 80, 772–785.
14. Shimizu, Y., Inoue, A., Tomari, Y., Suzuki, T.,Yokogawa, T., Nishikawa, K. & Ueda, T. (2001).Cell-free translation reconstituted with purified com-ponents. Nat. Biotechnol. 19, 751–755.
15. Ito, K., Uno, M. & Nakamura, Y. (2000). A tripeptide‘anticodon’ deciphers stop codons in messenger RNA.Nature, 403, 680–684.
16. Nakamura, Y. & Ito, K. (2002). A tripeptide discrim-inator for stop codon recognition. FEBS Lett. 514,30–33.
17. Ivanova, N., Pavlov, M. Y., Felden, B. & Ehrenberg, M.(2004). Ribosome rescue by tmRNA requires truncat-ed mRNAs. J. Mol. Biol. 338, 33–41.
18. Gagnon, M. G., Seetharaman, S. V., Bulkley, D. &Steitz, T. A. (2012). Structural basis for the rescue ofstalled ribosomes: structure of YaeJ bound to theribosome. Science, 335, 1370–1372.
19. Hayes, C. S. & Sauer, R. T. (2003). Cleavage of the Asite mRNA codon during ribosome pausing providesa mechanism for translational quality control. Mol.Cell, 12, 903–911.
20. Yamamoto, Y., Sunohara, T., Jojima, K., Inada, T. &Aiba, H. (2003). SsrA-mediated trans-translation playsa role in mRNA quality control by facilitatingdegradation of truncated mRNAs. RNA, 9, 408–418.

631ArfA recruits RF2 into stalled ribosomes
21. Chadani, Y., Matsumoto, E., Aso, H., Wada, T.,Kutsukake, K., Sutou, S. & Abo, T. (2011). trans-Translation-mediated tight regulation of the ex-pression of the alternative ribosome-rescue factorArfA in Escherichia coli. Genes Genet. Syst. 86,151–163.
22. Taniguchi, Y., Choi, P. J., Li, G. W., Chen, H., Babu,M., Hearn, J. et al. (2010). Quantifying E. coli proteomeand transcriptome with single-molecule sensitivity insingle cells. Science, 329, 533–538.
23. Shimizu, Y. & Ueda, T. (2010). PURE technology.Methods Mol. Biol. 607, 11–21.