Embed Size (px)
Citation preview

Accepted Manuscript
Analysis of liver and gill miRNAs of Larimichthys crocea against Cryptocryon irritanschallenge
Ying Qiao, Yong Mao, Jun Wang, Ruanni Chen, Yong-Quan Su, Jia Chen, Wei-QiangZheng
PII: S1050-4648(16)30660-X
DOI: 10.1016/j.fsi.2016.10.027
Reference: YFSIM 4255
To appear in: Fish and Shellfish Immunology
Received Date: 17 July 2016
Revised Date: 12 October 2016
Accepted Date: 15 October 2016
Please cite this article as: Qiao Y, Mao Y, Wang J, Chen R, Su Y-Q, Chen J, Zheng W-Q, Analysis ofliver and gill miRNAs of Larimichthys crocea against Cryptocryon irritans challenge, Fish and ShellfishImmunology (2016), doi: 10.1016/j.fsi.2016.10.027.
This is a PDF file of an unedited manuscript that has been accepted for publication. As a service toour customers we are providing this early version of the manuscript. The manuscript will undergocopyediting, typesetting, and review of the resulting proof before it is published in its final form. Pleasenote that during the production process errors may be discovered which could affect the content, and alllegal disclaimers that apply to the journal pertain.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Analysis of liver and gill miRNAs of Larimichthys crocea against Cryptocryon irritans challenge
Ying Qiao 1,2, Yong Mao1,2
,Jun Wang2, Ruanni Chen 2, Yong-Quan Su1,2*, Jia Chen1,Wei-Qiang
Zheng1
1. State Key Laboratory of Large Yellow Croaker Breeding (Fujian Fuding Seagull Fishing Food
Co., Ltd), 352103, Fujian, China
2.College of Ocean and Earth Science, Xiamen University, 361005, China * Corresponding author. Tel. +86 13606015589. E-mail address: [email protected] (Y, Q, Su).
Abstract
The white-spot disease caused by marine ciliate Cryptocryon irritans hindered the
sustainable development of large yellow croaker Larimichthys crocea industry. Better
understandings about the parasite-host interactions in the molecular level will facilitate the
prevention of mass mortality of the L. crocea caused by white-spot disease. MicroRNAs (miRNAs)
are a class of small RNA molecules about 20-22 nucleotides which post-transcriptionally regulated
many protein-coding genes and involved in many biological processes, especially in
host-pathogen responses. In this study, we identified known and novel miRNAs in the gill and
liver of L. crocea challenged by C. irritans by high throughput sequencing using Solexa
technology. The data were further studied to screen differentially expressed miRNAs, and predict
their target genes. The differential expression (p < 0.05) between libraries was observed in 103
miRNAs in liver tissue, among which 65 and 38 were conserved and novel miRNAs, 67 and 36
were up- and down-regulated miRNAs. While in gill tissue, 122 significant differentially
expressed miRNAs were identified, among which 83 and 39 were conserved and novel miRNAs,
79 and 43 were up- and down-regulated miRNAs. In addition, these differentially expressed
miRNAs target a series of genes which involved in many important biological processes including
immune response. Here via deep sequencing, we for the first time characterize L. crocea miRNAs
in response to C. irritans challenge, the results should help for better understandings about the
immune response of the L. crocea under C. irritans challenge.
Keywords:Larimichthys crocea ;Cryptocryon irritans ;MicroRNA (miRNA); Immune
response ;Deep sequencing;
1. Introduction
Large yellow croaker (Larimichthys crocea) (Perciformes: Sciaenidae), is known as one of
the most important commercial fishes in southeastern China, and the Large yellow croaker
industry is now one of the biggest industries in Chinese marine culture[1]. Nowadays the industry
has suffered significant economic losses due to various diseases. The cryptocaryonosis(white spot
disease), which is caused by the marine ciliate parasite Cryptocaryon irritans, can lead to as much
as 75% mortality of the large yellow croaker and other cultured marine fishes[2]. Hence, the
cryptocaryonosis is a serious constraint to aquaculture development of the marine fish.
MicroRNAs (miRNAs), typically ~22 nt in length, are considered playing various important
roles in many fundamental biological processes in animals and plants[3-6]. miRNAs are believed

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
to negatively regulate gene expression at the post-transcriptional level, they act their functions by
binding with the 3’- untranslated region(UTR) of the target gene mRNA or by mediating the
degradation of the target gene[7, 8]. miRNAs have attracted many research attention since the first
miRNA lin-14 was reported in Caenorhabditis elegans, which is demonstrated to act by negatively
regulating the level of LIN-14 protein[9]. To date, total of 21 of the database containing 28,645
entries representing hairpin precursor miRNAs, expressing 35,828 mature miRNA products, in
223 species are available(ftp://mirbase.org/pub/mirbase/CURRENT/README). All the miRNAs
participated in diverse biological processes, including stress responses[10, 11], metabolism[12,
13], morphogenesis[14], cell proliferation[15], apoptosis[16, 17] and host immune responses[18].
However, few researches have been done on the miRNAs functions in the host-parasite
interactions in the marine fishes.
Here in our study, in order to investigate large yellow croaker miRNA and expand novel
insights into host-parasite interactions in L. crocea, we analyzed the miRNA transcriptome from
liver and gill of L. crocea under control and challenge with the parasite C. irritans by deep
sequencing technology. The conserved and novel miRNAs were identified, the differentially
expressed miRNAs were screened and the corresponding target mRNAs were predict via
bioinformatics analysis.
2. Materials and methods
2.1. C. irritans challenge and sample collection
Healthy large yellow croakers L. crocea of nearly uniform size (weight: 5.47 ± 1.06 g, length:
8.81 ± 0.64 cm) were obtained from commercial local fish farm at Ningde, Fujian Province, China.
The fishes were maintained in aerated tank of 3 tons at 26-27 ℃ in running seawater. The fishes
were acclimatized for about 7 days prior to challenge by C. irritans.
After acclimatization for 7 d, 8 of L. crocea were randomly selected, anesthetized and
sacrificed to collect the gill and liver as the control group(G-c or L-c). For the C. irritans challenge,
the rest fishes were immersed in a suspension containing the newly hatched theronts of C. irritans,
at dose of 6,000 theronts per fish for 3 h. after challenge, the fishes were maintained in the running
seawater tank as described above. 12 h after challenge, the fishes were anesthetized and sacrificed,
the gill and liver were collected as the C. irritans infect/sensitive group(G-s or L-s). all the tissues
were immediately preserved in in RNAfixer (BioTek Corporation) at -80 ℃ for the further RNA
extraction.
2.2 RNA extraction
Total RNA of the gill and liver tissues were extracted from the control group and infected
group using TRIzol Reagent (Invitrogen) following the manufacturer’s instructions and
RNase-free DNase I (TaKaRa) was employed to remove DNA contaminants. The purified RNA
quantity and quality were examined using Nanodrop spectrophotometer(LabTech, USA), only the
RNAs with A260/A280 from 1.8 to 2.0 and A260/A230 from 2.0 to 2.5 were used for the
subsequent analyses. The integrity of the RNA samples were evaluated using Agilent 2100
bioanalyzer (Agilent Technologies) and only values of 28S/18S ≥ 0.7 and RIN ≥ 7.0 were
considered qualified for the subsequent small RNA library construction.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
2.3 miRNA library construction and deep-sequencing
miRNA fragments of 18-30 nt were enriched and purified from more than 7 µg of total RNA
from each group using 15% Tris-borate-EDTA (TBE) urea gel electrophoresis. Then a pair of
adaptors were ligated to both 5′ and 3′ termini of the miRNA fragments; the miRNA molecules
were amplified by RT-PCR using adaptor primers for 17 cycles and products of 62-75 nt(miRNA +
adaptor derived cDNA) were isolated from TBE PAGE gels. The purified cDNA fragments were
clustered and sequenced on Illumina sequencing platform (Illumina HiSeq™ 2500) using the
single-end technology by GENE DENOVO Corporation (Guangzhou, China).
2.4 Preprocessing of raw data and miRNA annotation
The raw data were pre-processed to exclude low-quality reads(length <20 nt and ambiguous
N) as well as 5′, 3′ adapter and poly(A) sequences. The filtered clean reads were aligned against
various databases, including genebank and Rfam to remove noncoding RNA, such as rRNA,
scRNA,snoRNA,snRNA and tRNA. Then obtained sequences were compared with exon and
intron in the L. crocea genome to classify mRNA degradation products and the repeat associate
miRNA sequences.
The filtered sequences were analyzed by BLAST search against Sanger miRBase 21.0 by
allowing at most two mismatches outside of the seed region[19]. Small RNAs that matched known
miRNAs of other animal species in miRBase were identified as conserved miRNAs and the
secondary structures of pre-miRNA hairpin were drawn using RNAfold[20].
2.5 Novel miRNAs identification
The non-annotated sequences in all libraries were searched against the L. crocea genome
using the MIREAP_v0.2 program with default parameters[21]. MIREAP is a tool which can be
used to identify both known and novel microRNAs from small RNA libraries deeply sequenced by
Solexa/454/Solid technology(https://sourceforge.net/projects/mireap). It takes into account
miRNA biogenesis, sequencing depth and structural features to improve the sensitivity and
specificity of miRNA identification.
2.6 Differential expression of miRNA
Different expression levels in miRNAs between the control and infection samples(G-c/G-s,
L-c/L-s) were determined using the program DEGseq as Zhu et al [22]described
(http://www.bioconductor.org/packages/2.6/bioc/html/DEGseq). The expression level of all
miRNAs(conserved and novel miRNAs) were normalized by dividing the absolute read count with
the total read count of the sample and multiplying the result by 1,000,000. Herein the specific
miRNA was considered significantly differentially expressed when the express quantity changes
more than 2 times and the p-value obtained by this method was less than 0.05.
2.7 Target gene prediction and Functional enrichment of differentially expressed miRNAs
The miRNA target genes of differentially expressed miRNAs were predicted using three
methods: RNAhybrid(v2.1.2)+svm_light(v6.01), Miranda(v3.3a) and TargetScan(Version: 7.0),
then the intersection of the three prediction results was taken as the final result of target gene
prediction of differentially expressed miRNAs.
To determine the main biological functions, different expression genes (DEGs) were

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
annotated by terms of the Gene Ontology (GO) database (http://www.geneontology.org/) using
Blast2GO[23] according to their numeric orders in nr database. After getting GO annotations of
every DEGs, GO functional classifications for all the DGEs were obtained via WEGO
software[24]. GO enrichment analysis of functional significance terms in the GO database was
applied using hypergeometric test to find significantly enriched GO terms in DEGs comparing to
the genome.
2.8 Real-Time PCR
In order to confirm the expression of microRNAs identified by the miRNA-seq, 3 of
miRNAs were randomly selected in each tissue. mir-459-x, mir-1-z, mir-458-y were selected in
liver tissue and mir-122-x, mir-142-x,mir-144-y were selected in gill tissue for analysis by
real-time PCR. Total RNA extracts were reverse-transcripted by TransScript Green miRNA
Two-Step qRT-PCR SuperMix (TransGen Biotech, China). The RT products were amplified by
TransStart Tip Green qPCR SuperMix(TransGen Biotech, China) on an ABI Quantstudio 6 Flex
system (Applied Biosystems, Foster City, CA, USA), using a specific primer and a U6 gene as the
universal primer. All the primers used in the real-time PCR reactions were summarized in
supplementary file 6. 2-∆∆CT calculation were employed for the relative quantification of the
miRNAs.
3. Results and discussion
We analyzed the miRNAs expression in liver and gill tissues of the C. irritans infected L.
crocea.Four separate libraries of miRNAs were generated from the liver (L-c/L-s) and gill
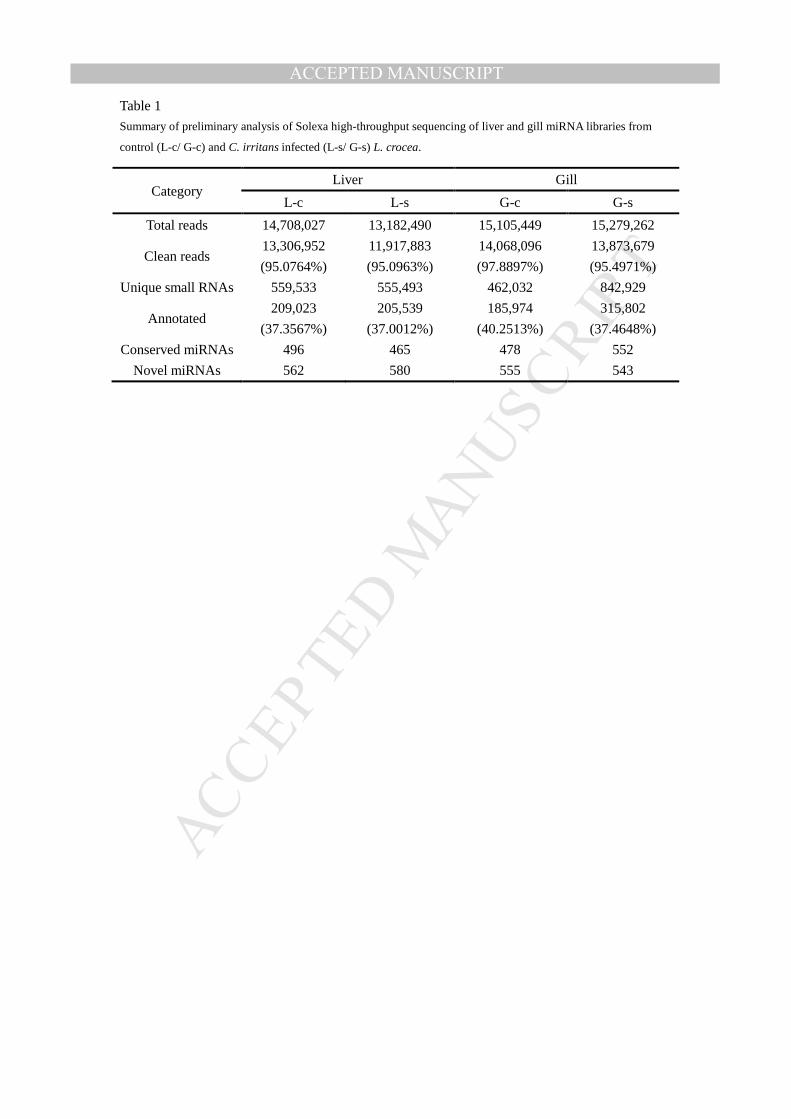
(G-c/G-s) tissues of large yellow croaker Larimichthys crocea. We obtained 14,708,027 and
13,182,490 raw reads from the L-c and L-s samples, respectively;meanwhile 15,105,449 and
15,279,262 raw reads were obtained from the G-c and G-s samples. The low quality sequences
(sequences more than one base which quality values lower than twenty, or reads containing N )
and the empty adapter sequences were discard; 13,306,952 and 11,917,883 clean reads were
obtained from L-c and L-s samples, and counted 95.0764% and 95.0963% of the total reads in
liver tissue; 14,068,096 and 13,873,679 clean reads were obtained from G-c and G-s samples,
counted 97.8897% and 95.4971% total reads in gill tissue. Then a total of 209,023 reads
accounting for 37.3567% of 559,533 unique small RNAs in L-c, 205,539 reads accounting for
37.0012% of 555,493 unique small RNAs in L-s; 185,974 reads accounting for 40.2513% of
462,032 unique small RNAs in G-c, and 315,802 reads accounting for 37.4648% of 842,929
unique small RNAs in G-s could be annotated by searching against genebank and Rfam database
(Table 1).
Length distribution of the four group miRNAs(L-c/L-s and G-c/G-s) were analyzed. 22 nt of
small RNAs were enriched as a peak, followed by 23 nt and 21 nt in all sample groups. Previous
studies have demonstrated the similar size distribution modes of miRNA in animals. In most
species, miRNAs of 22 nt long is the most typical phenomenon, such as in Paralichthys
olivaceus[4], Blattella germanica[25] and Culex pipiens[26].
The clean miRNA data were analyzed via BLAST search against miRBase 21.0 to identify
conserved miRNAs. All the conserved miRNAs from the liver and gill samples were sorted in
descending order of their expression values (TPM/ tags per million) and the top 30 conserved
miRNAs were listed, respectively (supplementary file 1). 20 conserved miRNAs (mir-21-x, let-7-x,

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
mir-26-x, mir-200-y, mir-199-x, mir-146-x, mir-143-y, mir-22-y, mir-181-x, mir-30-x, mir-142-y,
mir-125-x, mir-19-y, mir-199-y, mir-101-y, mir-126-y, mir-15-x, mir-16-x, mir-100-x, mir-29-y)
ranging from 21 to 23 nt were common to both liver and gill libraries. The mir-21-x and let-7-x
were top 2 of highly expressed miRNAs, and they were demonstrated in regulation of cell growth,
differentiation, apoptosis, immune response, and tumor metastasis[27, 28]. The mir-122-x were
observed as a unique and also the most abundant miRNA in liver library, this is similar with the
previous research results that the mir-22 as a liver-specific microRNA, playing an important
functions in the physiological process and disease development of the liver[29, 30].
For the typical hairpin structure of miRNAs comparing to other small RNAs, the candidate
novel miRNAs were identified by mapping to genome of L. crocea to predict secondary
structures[31]. Finally, 802 candidate novel miRNAs mapped to the L. crocea genome data were
identified from the liver and gill libraries, and top 30 expressed novel miRNAs were listed,
respectively (supplementary file 2). The length of novel miRNAs ranged from 20 to 24 nt, and 15
miRNAs (novel-m0431-5p, novel-m0235-5p, novel-m0120-5p, novel-m0127-3p, novel-m0384-3p,
novel-m0529-5p, novel-m0390-5p, novel-m0461-5p, novel-m0126-5p, novel-m0481-3p,
novel-m0120-3p, novel-m0305-3p, novel-m0528-3p, novel-m0215-5p and novel-m0108-5p) were
common to both liver and gill libraries. However, the specific role of these highly abundant
miRNAs have yet to be determined.
Here we focused on the functions of miRNAs in immune responses of anti-parasite infection.
miRNAs expressions are demonstrated to be altered in Japanese flounder (Paralichthys olivaceus)
infected with megalocytivirus[4]. Thus we believe differentially expressed miRNAs may play
important roles in immune response of scallop against C. irritans infections. Deep sequencing is
useful for identifying differential expressed miRNAs by measuring the sequencing frequencies[14,
32]. Here total of 122 and 103 differential expressed miRNAs were identified from liver and gill
libraries under C. irritans challenge. Among of the 122 differential expressed miRNAs in liver, 79
and 43 miRNAs were up- and down-regulated, 83 and 39 miRNAs were conserved and novel
miRNAs, respectively; but in 103 differential expressed miRNAs in gill, 67 and 36 miRNAs were
up- and down-regulated, 65 and 38 miRNAs were conserved and novel miRNAs. All the
differential expressed miRNAs were sorted in descending order by fold-change values, then top 20
of the up- and down-regulated miRNAs in both libraries were listed (supplementary file 3).
For all the differentially expressed miRNAs, we employed three prediction methods :
RNAhybrid(v2.1.2)+svm_light(v6.01),Miranda(v3.3a) and TargetScan(Version: 7.0) to predict the
target genes, and the intersection of the three prediction results was considered as the final result.
Then Gene ontology (GO) for the target genes of miRNAs were analyzed via using Blast2GO,
providing three type of information for each genes: “Biological Process”, “Cellular Component”
and “Molecular Function”. In the “biological processes” categories of liver libraries, 20 subtypes
were classified and most abundant genes were involved in cellular processes, biological regulation
and single-organism process. In the “Cellular Component” category, 16 subtypes were annotated
and most abundant genes were involved in cell, cell part and organelle. In addition, 13 subtypes
were annotated with “Molecular Function” which contained binding and catalytic activity as its
most abundant genes (Fig. 1). Similarly, the Gene ontology analysis of gill libraries the showed
the same results, but its contains 21, 19 and 11 subtypes in “Biological Process”, “Cellular
Component” and “Molecular Function” respectively (Fig. 2).
KEGG (Kyoto Encyclopedia of Genes and Genomes) database were used to predict the

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
functions of unigenes[33, 34]. 8614 genes and 218 differentially expressed genes (DEGs) were
classified into 121 KEGG pathways in liver library, 8614 genes and 491 DEGs were classified into
140 KEGG pathways in gill library. The predicted immune-related KEGG pathway analysis of
targeted genes in liver and gill tissues were listed in supplementary file 4 and supplementary file 5.
The results suggest some immune pathways involving in the immune response of L. crocea
against C. irritans infection, such as: RIG-I receptor signaling pathway, endocytosis, phagosome,
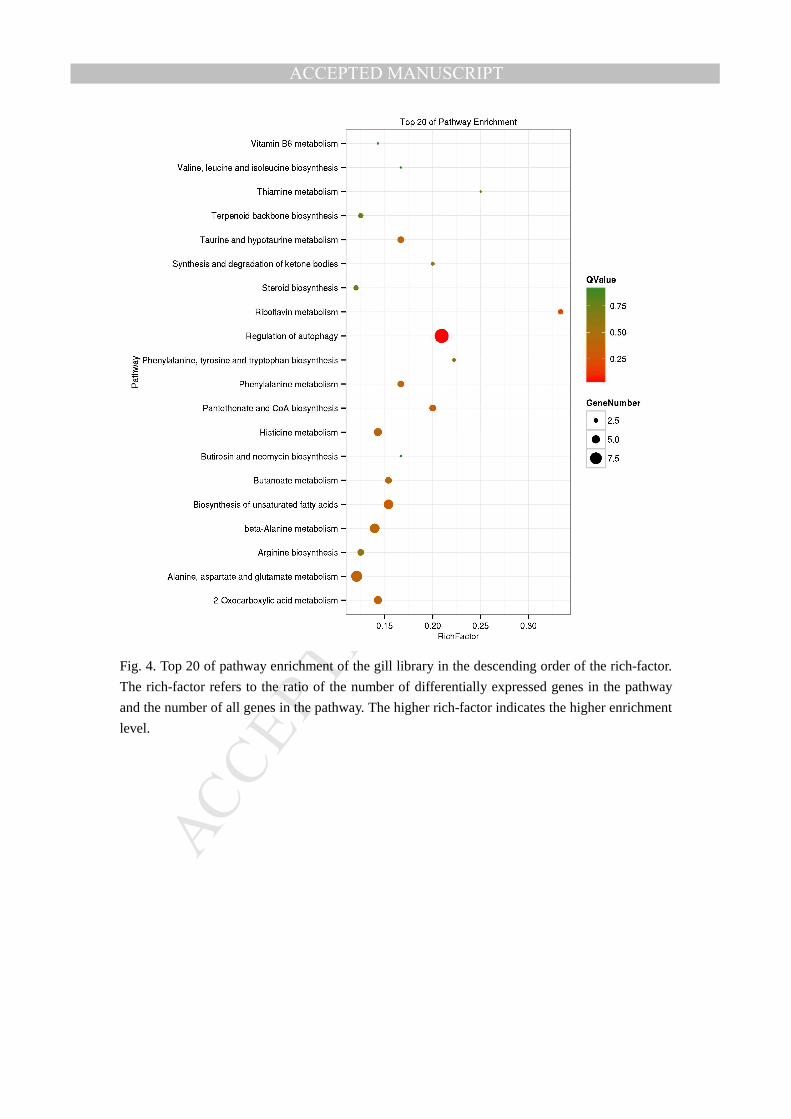
Toll-like receptor signaling pathway and Jak-STAT signaling pathway, etc. As we mapped the top
20 of pathway enrichment in the descending order of the rich-factor, we found the RIG-I receptor
signaling pathway (ko number: ko04622) seems to play an more important role than other immune
pathways in the immune process of liver (Fig. 3); but in the gill library of the L. crocea, regulation
of autophagy pathway (ko number: ko04140)was considered a more important pathway in the
immune response process (Fig. 4). This may suggests two of different immune response models in
the liver and gill tissues of L. crocea under C. irritans challenge.
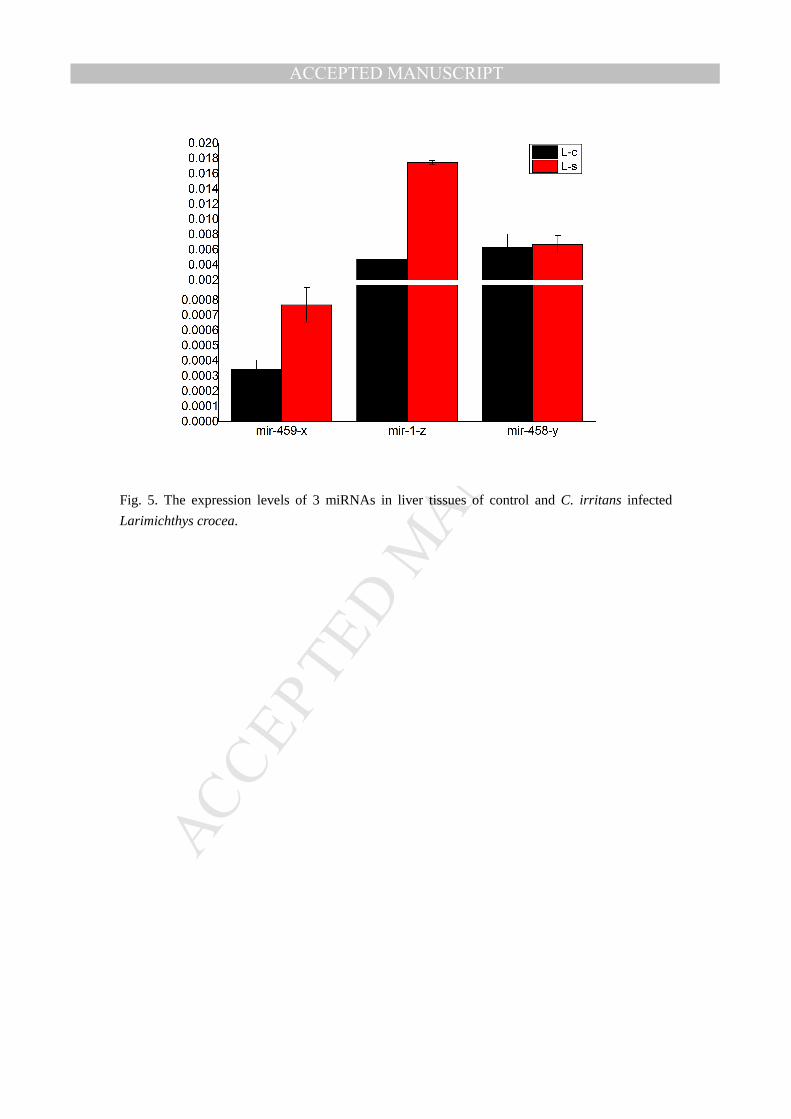
Real-time PCR is an acknowledged direct method for the precise measurement of mRNAs
and miRNAs. Here in our study, we use Real-time PCR to confirm the miRNAs expression
between the control and infected tissues of the Larimichthys crocea. The expression of mir-459-x,
mir-1-z, and mir-458-y gene were analyzed in liver groups(L-c and L-s)(Fig. 5) and mir-122-x,
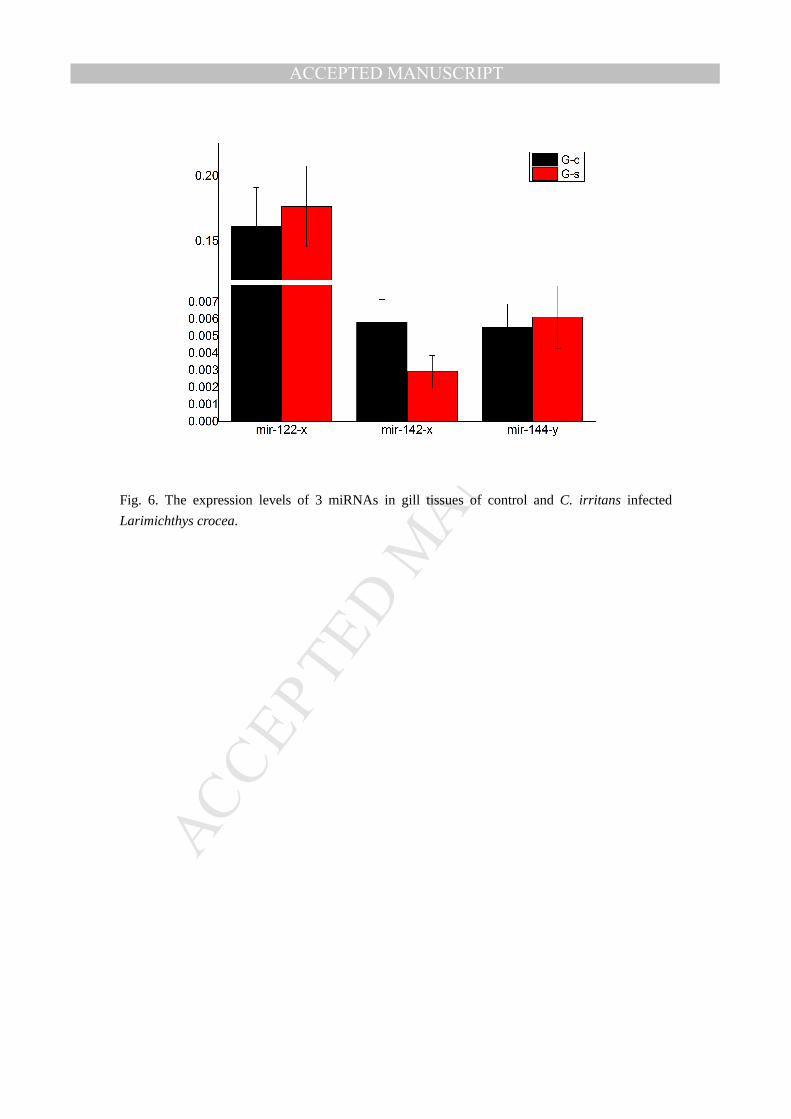
mir-142-x,mir-144-y genes were analyzed in gill(G-c and G-s)(Fig. 6). The expression trends of
the miRNAs in liver and gill tissues were roughly the same as the result of the miRNA-seq and the
result suggest the accuracy of the miRNA-seq. As such, the filteration and validation of the
immune –regulation related miRNAs still need more subsequent experimental evidence for the
conclusive results.
Acknowledgements
This study was financially supported by National Natural Science Foundation of China (Grant Nos.
31372504 and 41476118), Fujian science and Technology Department (Grant Nos.
2015I11010045 and 2015N2002-3) and Major projects of science and Technology Department of
Fujian Province(Grant No. 2016NZ0001-4).
References
1. Liu JF, Han KH. Current development situation and countermeasure of large yellow crocker
industry in China. Journal of Fujian Fisheries. 2011 1:1-5.
2. Niu SF, Jin Y, Xu X, Qiao Y, Wu Y, Mao Y, et al. Characterization of a novel piscidin-like antimicrobial
peptide from Pseudosciaena crocea and its immune response to Cryptocaryon irritans. Fish & shellfish
immunology. 2013 35:513-24.
3. Ruan L, Bian X, Ji Y, Li M, Li F, Yan X. Isolation and identification of novel microRNAs from
Marsupenaeus japonicus. Fish & shellfish immunology. 2011 31:334-40.
4. Bao-cun Zhang, Jian Zhang, Sun L. In-depth profiling and analysis of host and viral microRNAs in
Japanese flounder (Paralichthys olivaceus) infected with megalocytivirus reveal involvement of
microRNAs in host-virus interaction in teleost fish. BMC Genomics. 2014 1:1-15.
5. Singh N, Srivastava S, Shasany AK, Sharma A. Identification of miRNAs and their targets involved

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
in the secondary metabolic pathways of Mentha spp. Computational Biology and Chemistry. 2016
64:154-62.
6. Gupta OP, Sharma P, Gupta RK, Sharma I. Current status on role of miRNAs during plant–fungus
interaction. Physiological and Molecular Plant Pathology. 2014 85:1-7.
7. Hannon GJ. RNA interference. Nature. 2002 418:244-51.
8. Mattick JS, Makunin IV. Small regulatory RNAs in mammals. Human Molecular Genetics. 2005
14:R121-R32.
9. Lee RC, Feinbaum RL, Ambros V. The C. elegans heterochronic gene lin-4 encodes small RNAs
with antisense complementarity to lin-14. Cell. 1993 75:843-54.
10. Leung AKL, Sharp PA. MicroRNA Functions in Stress Responses. Molecular Cell. 2010 40:205-15.
11. Yan B, Zhao L-H, Guo J-T, Zhao J-L. miR-429 regulation of osmotic stress transcription factor 1
(OSTF1) in tilapia during osmotic stress. Biochem Bioph Res Co. 2012 426:294-8.
12. Wagschal A, Najafi-Shoushtari SH, Wang L, Goedeke L, Sinha S, deLemos AS, et al. Genome-wide
identification of microRNAs regulating cholesterol and triglyceride homeostasis. Nat Med. 2015
advance online publication.
13. Yang Z, Cappello T, Wang L. Emerging role of microRNAs in lipid metabolism. Acta Pharmaceutica
Sinica B. 2015 5:145-50.
14. Cristino AS, Tanaka ED, Mercedes R, Maria-Dolors P, Xavier B. Deep Sequencing of Organ- and
Stage-Specific microRNAs in the Evolutionarily Basal Insect Blattella germanica (L.) (Dictyoptera,
Blattellidae). Plos One. 2011 6:508-.
15. Jingjing L, Xu W, Xiaoning Y, Yunpeng L, Ying S, Jianlin R, et al. miRNA423-5p regulates cell
proliferation and invasion by targeting trefoil factor 1 in gastric cancer cells. Cancer Letters. 2014
347:98-104.
16. Hammond SM. MicroRNAs as oncogenes. Current Opinion in Genetics & Development. 2006
16:4-9.
17. Jovanovic M, ., Hengartner MO. miRNAs and apoptosis: RNAs to die for. Oncogene An
International Journal. 2006 25:págs. 6176-87.
18. Ma F, Xu S, Liu X, Zhang Q, Xu X, Liu M, et al. The microRNA miR-29 controls innate and adaptive
immune responses to intracellular bacterial infection by targeting interferon-[gamma]. Nat Immunol.
2011 12:861-9.
19. Kozomara A, Griffiths-Jones S. miRBase: integrating microRNA annotation and deep-sequencing
data. Nucleic acids research. 2010:gkq1027.
20. Hofacker IL. RNA secondary structure analysis using the Vienna RNA package. Current protocols
in bioinformatics. 2009:12.2. 1-.2. 6.
21. Chen X, Li Q, Wang J, Guo X, Jiang X, Ren Z, et al. Identification and characterization of novel
amphioxus microRNAs by Solexa sequencing. Genome Biol. 2009 10:R78.
22. Zhu E, Zhao F, Xu G, Hou H, Zhou L, Li X, et al. mirTools: microRNA profiling and discovery based
on high-throughput sequencing. Nucleic acids research. 2010 38:W392-W7.
23. Conesa A, Götz S, García-Gómez JM, Terol J, Talón M, Robles M. Blast2GO: a universal tool for
annotation, visualization and analysis in functional genomics research. Bioinformatics. 2005
21:3674-6.
24. Ye J, Fang L, Zheng H, Zhang Y, Chen J, Zhang Z, et al. WEGO: a web tool for plotting GO
annotations. Nucleic Acids Research. 2006 34:W293-W7.
25. Cristino AS, Tanaka ED, Rubio M, Piulachs M-D, Belles X. Deep Sequencing of Organ- and

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Stage-Specific microRNAs in the Evolutionarily Basal Insect <italic>Blattella germanica</italic> (L.)
(Dictyoptera, Blattellidae). Plos One. 2011 6:e19350.
26. Hong S, Guo Q, Wang W, Hu S, Fang F, Lv Y, et al. Identification of differentially expressed
microRNAs in Culex pipiens and their potential roles in pyrethroid resistance. Insect biochemistry and
molecular biology. 2014 55:39-50.
27. S Duygu S, Donoghue MTA, Charles S. miR-21 as a key regulator of oncogenic processes.
Biochemical Society Transactions. 2009 37:918-25.
28. Chae Won K, Mai-Tram V, Kyeung KH, Hyun Hee L, Nal Ae Y, Byung Ju L, et al. Ectopic
over-expression of tristetraprolin in human cancer cells promotes biogenesis of let-7 by
down-regulation of Lin28. Nucleic Acids Research. 2012 40:3856-69.
29. Yingzhuo Y, Peng K, Jie G, Chunlin X, shimei W, xia W. Research progress in liver-specific miR-122
in regulating liver diseases. J Clin Hepatol. 2014:473-7.
30. Ding X, Ding J, Ning J, Yi F, Chen J, Zhao D, et al. Circulating microRNA-122 as a potential
biomarker for liver injury. Molecular Medicine Reports. 2012 5:1428-32.
31. Victor A, Bonnie B, Bartel DP, Burge CB, Carrington JC, Xuemei C, et al. A uniform system for
microRNA annotation. Rna-a Publication of the Rna Society. 2003 9:277-9.
32. Creighton CJ, Reid JG, Gunaratne PH. Expression profiling of microRNAs by deep sequencing.
Briefings in Bioinformatics. 2009 10:490-7.
33. Ogata H, Goto S, Sato K. Fujibuchi W, Bono H, Kanehisa M: KEGG: Kyoto Encyclopedia of Genes
and Genomes. Nucleic Acids Research. 1999 volume 28:27-30(4).
34. Ogata H, ., Goto S, ., Sato K, ., Fujibuchi W, ., Bono H, ., Kanehisa M, . KEGG: kyoto encyclopedia
of genes and genomes. Nucleic Acids Research. 2000 28:29-34(6).
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Table 1
Summary of preliminary analysis of Solexa high-throughput sequencing of liver and gill miRNA libraries from
control (L-c/ G-c) and C. irritans infected (L-s/ G-s) L. crocea.
Category Liver Gill
L-c L-s G-c G-s
Total reads 14,708,027 13,182,490 15,105,449 15,279,262
Clean reads 13,306,952
(95.0764%)
11,917,883
(95.0963%)
14,068,096
(97.8897%)
13,873,679
(95.4971%)
Unique small RNAs 559,533 555,493 462,032 842,929
Annotated 209,023
(37.3567%)
205,539
(37.0012%)
185,974
(40.2513%)
315,802
(37.4648%)
Conserved miRNAs 496 465 478 552
Novel miRNAs 562 580 555 543

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Fig. 1. Gene ontology (GO) term of DEGs genes in L-c vs L-s. DEGs count for the liver of L.
crocea gene ontology classifications into biological process (green), cellular component (red),
molecular function (blue) categories.
Fig. 2. Gene ontology (GO) term of DEGs genes in G-c vs G-s. DEGs count for the gill of L.
crocea gene ontology classifications into biological process (green), cellular component (red),
molecular function (blue) categories.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Fig. 3. Top 20 of pathway enrichment of the liver library in the descending order of the rich-factor.
The rich-factor refers to the ratio of the number of differentially expressed genes in the pathway
and the number of all genes in the pathway. The higher rich-factor indicates the higher enrichment
level.
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Fig. 4. Top 20 of pathway enrichment of the gill library in the descending order of the rich-factor.
The rich-factor refers to the ratio of the number of differentially expressed genes in the pathway
and the number of all genes in the pathway. The higher rich-factor indicates the higher enrichment
level.
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Fig. 5. The expression levels of 3 miRNAs in liver tissues of control and C. irritans infected
Larimichthys crocea.
MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
Fig. 6. The expression levels of 3 miRNAs in gill tissues of control and C. irritans infected
Larimichthys crocea.

MANUSCRIP
T
ACCEPTED
ACCEPTED MANUSCRIPT
We analyzed the liver and gill miRNAs of L. crocea under C. irritans challenge using miRNA-seq.