Embed Size (px)
DESCRIPTION
Explains how to use algae and aquatic biomass for a sustainable production of 2nd generation biofuels
Citation preview

AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 1 of 258
Proposal full title: Algae and aquatic biomass for a sustainable production of 2nd generation biofuels
Proposal acronym: AquaFUELs
Type of funding scheme: Cooperation
Theme 5 Energy
Taxonomy, Biology and Biotechnology
Name of the coordinating person: Dr. Raffaello Garofalo Coordinator email: ebb@ebb‐eu.org Coordinator phone: +32 2 7632477 Coordinator fax: +32 2 7630457
REV Date Organisation Beneficiaries involved
Dissemination level
FINAL 20/05/2011 Natascia Biondi, Mario Tredici
UNIFI UNIFI PU

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 2 of 258
Table of contents
1 INTRODUCTION................................................................................................................................................. 5 1.1 IMPORTANCE OF ALGAE AND AQUATIC BIOMASS FOR BIOFUELS......................................................................... 6
1.1.1 Suitability of algae as biomass producers ..................................................................................................... 6 1.1.2 Sustainability, the strategic advantage of algal biofuels ............................................................................... 6
1.2 RATIONALE OF THE DOCUMENT ......................................................................................................................... 8 1.3 TARGET GROUPS ................................................................................................................................................ 8 1.4 PROBLEMS INCURRED ........................................................................................................................................ 9 1.5 COMMON ERRONEOUS "MYTH" .......................................................................................................................... 9 2 CRITERIA FOR STRAIN SELECTION............................................................................................................ 10 2.1 PRODUCTIVITY................................................................................................................................................. 10 2.2 ROBUSTNESS.................................................................................................................................................... 10 2.3 HARVESTABILITY............................................................................................................................................. 10 2.4 BIOMASS COMPOSITION ................................................................................................................................... 10 2.5 PROCESSABILITY / EXTRACTABILITY................................................................................................................ 11 2.6 ADDED VALUE OF CO-PRODUCTS ..................................................................................................................... 11 2.7 LOCAL ORIGIN OF STRAINS............................................................................................................................... 11 3 BIOLOGY OF ALGAE....................................................................................................................................... 12 3.1 CYANOBACTERIA............................................................................................................................................. 12 3.2 CHLOROPHYTA (GREEN ALGAE)...................................................................................................................... 15 3.3 RHODOPHYTA (RED ALGAE)............................................................................................................................ 17 3.4 HETEROKONTOPHYTA...................................................................................................................................... 18
3.4.1 Phaeophyceae (Brown algae)...................................................................................................................... 19 3.4.2 Eustigmatophyceae ..................................................................................................................................... 21 3.4.3 Other classes ............................................................................................................................................... 21
3.5 LABYRINTHULEA (PHYLUM HETEROKONTA) ................................................................................................... 21 3.6 BACILLARIOPHYTA (DIATOMS)........................................................................................................................ 22 3.7 HAPTOPHYTA................................................................................................................................................... 24 3.8 DINOPHYTA (DINOFLAGELLATES) ................................................................................................................... 24 3.9 OTHER ALGAL GROUPS..................................................................................................................................... 26 4 BIOTECHNOLOGY OF ALGAE ...................................................................................................................... 28 4.1 INTRODUCTION ................................................................................................................................................ 28 4.2 CULTIVATION SYSTEMS ................................................................................................................................... 29
4.2.1 Open ponds ................................................................................................................................................. 30 4.2.2 Photobioreactors ......................................................................................................................................... 31
Main photobioreactors designs ...............................................................................................................................................32 Polyethylene bags and vertical columns.............................................................................................................................32 Tubular PBR ......................................................................................................................................................................33 Flat photobioreactors (panels) ............................................................................................................................................35
4.2.3 Sustainability of different cultivation systems............................................................................................ 35 4.3 HARVESTING METHODS.................................................................................................................................... 36 4.4 BIOTECHNOLOGY OF THE MAJOR MICROALGAL GROUPS .................................................................................. 37
4.4.1 Cyanobacteria ............................................................................................................................................. 37 4.4.2 Chlorophyta (Green Algae)......................................................................................................................... 38 4.4.3 Rhodophyta (Red Algae) ............................................................................................................................ 39 4.4.4 Heterokontophyta........................................................................................................................................ 39 4.4.5 Labyrinthulea (phylum Heterokonta).......................................................................................................... 40 4.4.6 Bacillariophyta (Diatoms)........................................................................................................................... 40 4.4.7 Haptophyta.................................................................................................................................................. 41 4.4.8 Dinophyta (Dinoflagellates)........................................................................................................................ 41
4.5 BIOTECHNOLOGY AND USES FOR MACROALGAE .............................................................................................. 41 4.6 BIOTECHNOLOGY OF OTHER AQUATIC BIOMASS ............................................................................................. 43 5 SYMBOLOGY.................................................................................................................................................... 44 6 REFERENCES.................................................................................................................................................... 45 7 PROKARYOTIC MICROALGAE ..................................................................................................................... 51 7.1 CYANOBACTERIA............................................................................................................................................. 51
7.1.1 Arthrospira sp. (common name spirulina) .................................................................................................. 51 7.1.2 Phormidium sp............................................................................................................................................ 54 7.1.3 Anabaena sp................................................................................................................................................ 57 7.1.4 Synechococcus sp........................................................................................................................................ 60 7.1.5 Synechocystis sp.......................................................................................................................................... 62

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 3 of 258
8 EUKARYOTIC MICROALGAE........................................................................................................................ 64 8.1 CHLOROPHYTA ................................................................................................................................................ 64
8.1.1 Ostreococcus sp. ......................................................................................................................................... 64 8.1.2 Tetraselmis sp ............................................................................................................................................. 66 8.1.3 Botryococcus braunii .................................................................................................................................. 70 8.1.4 Chlamydomonas reinhardtii ....................................................................................................................... 72 8.1.5 Haematococcus pluvialis ............................................................................................................................ 76 8.1.6 Dunaliella sp............................................................................................................................................... 79 8.1.7 Chlorococcum sp. ....................................................................................................................................... 83 8.1.8 Neochloris oleoabundans............................................................................................................................ 86 8.1.9 Scenedesmus sp........................................................................................................................................... 92 8.1.10 Desmodesmus sp. ........................................................................................................................................ 97 8.1.11 Chlorella sp. ............................................................................................................................................... 99 8.1.12 Parietochloris incisa................................................................................................................................. 109 8.1.13 Prototheca sp. ........................................................................................................................................... 111
8.2 RHODOPHYTA ................................................................................................................................................ 113 8.2.1 Porphyridium cruentum ............................................................................................................................ 113
8.3 BACILLARIOPHYTA ........................................................................................................................................ 115 8.3.1 Benthic diatoms (Amphora; Amphiprora; Cylindrotheca; Navicula; Nitzschia) ...................................... 115
Amphora sp. ..........................................................................................................................................................................115 Amphiprora hyalina ..............................................................................................................................................................116 Cylindrotheca sp. ..................................................................................................................................................................116 Navicula sp. ..........................................................................................................................................................................117 Nitzschia dissipata ................................................................................................................................................................118
8.3.2 Phaeodactylum tricornutum...................................................................................................................... 126 8.3.3 Chaetoceros muelleri ................................................................................................................................ 126 8.3.4 Cyclotella cryptica .................................................................................................................................... 136 8.3.5 Odontella aurita........................................................................................................................................ 139 8.3.6 Skeletonema sp.......................................................................................................................................... 141 8.3.7 Thalassiosira pseudonana ........................................................................................................................ 143
8.4 EUSTIGMATOPHYCEAE (PHYLUM HETEROKONTOPHYTA) .............................................................................. 146 8.4.1 Monodus subterraneus.............................................................................................................................. 146 8.4.2 Nannochloropsis sp................................................................................................................................... 148
8.5 HAPTOPHYTA................................................................................................................................................. 153 8.5.1 Isochrysis sp.............................................................................................................................................. 153 8.5.2 Pavlova sp................................................................................................................................................. 156
8.6 DINOPHYTA ................................................................................................................................................... 159 8.6.1 Crypthecodinium cohnii............................................................................................................................ 159
8.7 LABYRINTHULOMYCETES .............................................................................................................................. 162 8.7.1 Schizochytrium sp. .................................................................................................................................... 162 8.7.2 Thraustochytrium sp. ................................................................................................................................ 164 8.7.3 Ulkenia sp. ................................................................................................................................................ 166
9 MACROALGAE............................................................................................................................................... 167 9.1 CHLOROPHYTA .............................................................................................................................................. 167
9.1.1 Caulerpa sp............................................................................................................................................... 167 Caulerpa racemosa ...............................................................................................................................................................167 Caulerpa taxifolia .................................................................................................................................................................168
9.1.2 Ulva sp. ..................................................................................................................................................... 170 Ulva lactuca ..........................................................................................................................................................................170 Ulva rigida ............................................................................................................................................................................170
9.1.3 Cladophora sp. ......................................................................................................................................... 178 9.1.4 Codium sp. ................................................................................................................................................ 180
Codium fragile ......................................................................................................................................................................180 Codium parvulum..................................................................................................................................................................180
9.2 RHODOPHYTA ................................................................................................................................................ 183 9.2.1 Chondrus crispus ...................................................................................................................................... 183 9.2.2 Mastocarpus stellatus ............................................................................................................................... 185 9.2.3 Grateloupia turuturu................................................................................................................................. 187 9.2.4 Palmaria palmata ..................................................................................................................................... 189 9.2.5 Solieria chordalis...................................................................................................................................... 191
9.3 PHAEOPHYCEAE (PHYLUM HETEROKONTOPHYTA) ........................................................................................ 193 9.3.1 Alaria esculenta ........................................................................................................................................ 193 9.3.2 Undaria pinnatifida .................................................................................................................................. 195

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 4 of 258
9.3.3 Ascophyllum nodosum .............................................................................................................................. 197 9.3.4 Fucus sp. ................................................................................................................................................... 199
Fucus serratus.......................................................................................................................................................................199 Fucus spiralis........................................................................................................................................................................199 Fucus vesiculosus..................................................................................................................................................................200
9.3.5 Himanthalia elongata ............................................................................................................................... 202 9.3.6 Cystoseira sp............................................................................................................................................. 204
Cystoseira baccata................................................................................................................................................................204 Cystoseira tamariscifolia ......................................................................................................................................................205
9.3.7 Halidrys siliquosa ..................................................................................................................................... 207 9.3.8 Sargassum muticum .................................................................................................................................. 208 9.3.9 Laminaria, Saccharina, Saccorhiza.......................................................................................................... 210
Laminaria sp. ........................................................................................................................................................................210 Laminaria digitata ...........................................................................................................................................................210 Laminaria hyperborea......................................................................................................................................................211 Laminaria ochroleuca ......................................................................................................................................................212
Saccharina latissima .............................................................................................................................................................212 Saccorhiza polyschides .........................................................................................................................................................213
10 OTHER AQUATIC BIOMASS ........................................................................................................................ 221 10.1 EGERIA DENSA ................................................................................................................................................ 221 10.2 EICHHORNIA CRASSIPES................................................................................................................................... 223 10.3 ELODEA CANADENSIS ...................................................................................................................................... 226 10.4 LAGAROSIPHON MAJOR.................................................................................................................................... 228 10.5 LEMNA MINOR ................................................................................................................................................. 231 11 CONCLUDING REMARKS............................................................................................................................. 236 ANNEX I ................................................................................................................................................................... 237

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 5 of 258
1 Introduction
Algae are a group of organisms that have been generally described as photoautotrophic unicellular or multicellular, mainly water dwelling organisms lacking complex morphological organization. Historically, the prokaryotic blue‐green algae, or cyanobacteria (Class Cyanophyceae), are often included in discussing microalgae, and indeed some cyanobacterial species (Arthrospira or spirulina) hold a prominent position in the biotechnological exploitation of microalgae.
There are several main groups of microalgae differing in biochemical constituents, ultrastructure, and life cycle. Some of the characteristics traditionally used for algae classification are the nature of their photosynthetic pigments, storage products, cell wall, presence or absence of flagella and the number of membranes surrounding the chloroplast.
More recently classification has been based on comparisons of specific DNA sequences, leading to major revisions in classification of many groups of alga. Recent molecular genetic studies confirmed that photoautotrophic eukaryotes belong to several highly diverse groups of organisms and are the result of different and independent events of secondary endosymbioses. As a consequence algae belong to genetically widely diverting groups of organisms often closer related to nonphotosynthetic organisms than to more distant algal clades (Fig. 1). This fact requires due attention when developing tools such as transformation or genetic engineering etc for microalgae.
The most recent results on algal taxonomy, are summarized in detail by the Tree of Life project (http://tolweb.org/tree/) and AlgaeBase (http://www.algaebase.org/), both providing up to date taxonomic information concerning classification of algal species, that is continuously being updated and revised in light of newest results obtained by molecular genetic approaches such as DNA sequence comparisons.
Figure 1 Phylogenetic tree of the eukaryotic organisms (modified from Tree of Life Project http://tolweb.org/tree/).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 6 of 258
Microalgae reproduction occurs primarily by vegetative (asexual) cell division, although sexual reproduction can occur in many species under appropriate growth conditions.
1.1 Importance of algae and aquatic biomass for biofuels
1.1.1 Suitability of algae as biomass producers
Microalgae are considered fast‐growing photosynthetic organisms and have been reported to reach short term maximal summer productivities of 50 ‐ 60 g per m2 per day in CO2 enriched raceway ponds in Hawaii and California. This corresponds to transformation of 5‐6% of incoming light energy into biomass. Such numbers, as well as productivity data from lab scale experiments have promoted the reputation of microalgae as prime candidates for providing unlimited amounts of cheap biomass as food, fodder or energy. Furthermore many algal strains can produce large amounts of oil or lipid like storage products that can easily be converted into biodiesel (Sheehan et al., 1998). This has been used by some to combine maximal biomass productivity with maximal oil content, yielding phantastic oil productivity numbers that have been exploited for funding intensive research on algal biofuels production. Potential oil productivities of over 100 tons/ha per year were initially predicted. However, none of the large scale long term experiments ever reached the high productivity projections. Current productivity obtained in large scale operations range from 40 – 60 tons of algal biomass production per ha and year, with conservative projections anticipating up to 100 tons of biomass, or 30 tons of biodiesel per ha and year in subtropical or tropical, sunny climates (Scott et al., 2010).
Five groups of microalgae were classified as high priority for biofuel production by the US microalgal species program ASP (Sheehan et al., 1998): diatoms (Class Bacillariophyceae), green algae (Class Chlorophyceae), golden‐brown algae (Class Chrysophyceae), prymnesiophytes or haptophytes (Class Prymnesiophyceae), and eustigmatophytes (Class Eustigmatophyceae). However, different classes of macroalgae, as well as further yet less studied microalgal groups may turn out to be equally relevant for successful biomass production from algae.
Other aquatic biomass such as water lentils (Lemna), water hyacinth, Elodea and others have also been considered for potential biofuel production due to their significant productivity and their usefulness in treating polluted nutrient rich water bodies.
1.1.2 Sustainability, the strategic advantage of algal biofuels
Land Use ‐ Current biofuels such as oil from soy bean, palm, and rape seed, or ethanol from corn or wheat, suffered from serious setbacks revealed by recent analysis showing their adverse ecological impact and low greenhouse gas reduction potential. A recent statement by UNEP Director, A. Steiner, reads: "biofuels from palm oil grown by Indonesia might never be deemed to be sustainable", due to ongoing destruction of tropical forests for expanding palm oil production. In addition, with yields of less than 500‐5,000 L of biodiesel per hectare (Johnston et al., 2009), those crops require enormous areas of scarce arable land, water and fertilizer and are generally highly work intensive. Life cycle assessments (LCA) indicate that no, or very low, reductions in greenhouse gas emissions can be achieved using such biofuels (Zah et al., 2007), and if they are being produced following conversion of natural ecosystems their GHG emissions surpass those of fossil fuels for years to come (Fargione et al., 2008). The major impact, land use, is often not adequately considered if the strategic implications of expanding biofuels production are taken into account (Searchinger et al., 2008):
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 7 of 258
Supplying the world's 2030 liquid fuel demand from dedicated biofuel crops would consume all or most of the available land in the appropriate climatic zones including most remaining natural ecosystems even if the most productive biofuels crops such as palm oil, Miscanthus cellulosic ethanol or sugar cane ethanol or an optimized mix of those crops were planted. Even if a doubling of yields is achieved in the next 20 years, around half of the worlds remaining intact ecosystems will have to be sacrificed to cover the projected liquid fuel demand.
Only algae can provide sufficient liquid fuels on a few percent of available dryland areas. Algal biomass if managed properly, may be produced on unproductive desert land, under utilization of ocean or waste water by exploiting and recycling of waste nutrients from municipal and agricultural sources, since no health concerns need to be considered for biofuel production. Algal biofuel production is thus complementary to ongoing efforts to grow lignocellulosic biomass in areas with good soil and water resources, because the microalgae are projected to be grown in those areas where the lignocellulosic or oil seeds crops will not perform well (Brown, M. Lewis, “Biodiesel from Microalgae: Complementarity in a Fuel Development Strategy”, NREL, http://www.nrel.gov/docs/legosti/old/5715.pdf ). Algae may produce sufficient biofuels on less than 10% of available drylands even under current productivity estimates, often using the most unproductive areas like salt flats or degraded dryland soils. They may also deliver additional environmental services such as wastewater treatment or exhaust gas detoxification.
Table 1 ‐ Comparison of land use impact of various biofuel crops. *For all crops it is assumed that 50% of biomass energy is used for process energy and all nutrients will be recycled to the maximum possible, according to the state of the art of sugar cane ethanol production. In Miscanthus this 50% are removed from the claimed ethanol yield, since no leftover biomass is available for process energy. The crops marked in italics, as well as algae, are experimental as no actual production in the large scale has been demonstrated. (Land areas are derived from Ito and Oikawa, 2004; biofuel productivities are derived from Johnston et al., 2009).
Land type Area (mio km2)
Natural Productivity (tons of carbon fixed per
hectare and year)
Crop and biofuel yield (tons per ha / GJ
per ha)
% area of corresponding
ecosystem required to cover 2030
demand
Tropical and subtropical evergreen forest
10.5 10.7 Palmoil (5 / 189)
110%!!!
Tropical and subtropical dry forest
4.7 7.67 Jatropha‐ oil (1.5 / 56.7 )
765%!!
Tropical Savanna, Woodland 6.7 6.65
Cane‐ethanol (4.34 / 116)
270 %
Mid lattitude forests, abandoned croplands
14 5.30
Miscanthus cellulosic ethanol* (4.4 / 120)
95 %
Warm Shrubland/grassland or desert
33 1 – 3.50 Algae‐ oil (20 / 756)
5.4 – 8.2 %
Other Environmental Services ‐ Interestingly, even under desert conditions the water footprint of algal biomass production is lower than that of irrigated maize ethanol production, calculated on the water input per unit of energy created. Our estimates suggest that it requires less than 0.5 million liters of water to produce one 1 ton of dry algal biomass, and this may be waste‐ or seawater. It can take about 3 million

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 8 of 258
liters of water to produce 1 ton of rice and about 2 million liters to produce 1 ton of soybeans (www.clw.csiro.au/issues/water/water_for_food.html and www.gdrc.org/uem/footprints/water‐footprint.html). However, significant investments into (waste)‐water and CO2 infrastructure would be required to achieve the necessary global algae biomass production potential.
Properly planned algal biomass production facilities may recover and reuse most of the nutrients applied, minimizing eutrophication impact, and this in contrast to intensive agriculture where nutrient runoff and escaping nitric oxide pose serious problems.
In fact, algae have been shown to be able to treat successfully any kind, even the most problematic, forms of wastewater. Pesticide use in algal cultivation is expected to be minimal. Even a small proportion of the algal biomass required for energy purposes would provide sufficient protein to replace all the soybean cultivation capacity installed for feed production, resulting in reduced deforestation or 'negative indirect land use changes' (Searchinger et al., 2008).
While algal biomass production may never achieve the low production costs of other agricultural commodities, full accounting of above and additional environmental services may result in a balance favouring the algal fuels. This point requires intensive investigation progressing far beyond currently used LCA models (Stephenson et al., 2010), starting with defining those production parameters and system boundaries that will actually deliver the above mentioned environmental advantages.
1.2 Rationale of the document This document summarizes in short current views and prospects on the potential contribution of algae to biofuel production.
Recent predictions and calculations, both at the EU level and in the US (A USDA Regional Roadmap to Meeting the Biofuels Goals of the Renewable Fuels Standard by 2020, USDA Strategic Biofuels Production report, June 2010), do not incorporate a significant algae potential into their projections for the next 10 years. This is dictated by the fact that algae experts and external observers disagree about the true potential of algal biofuels production relating to economic and environmental sustainability, and any given time frame for achieving competitive algal biofuel production is speculative at the best.
Given the high complexity of algal taxonomy and evolutionary relationships, this document was conceived as an instrument to place the algae that have arisen an interest for biofuel production within the correct frame. The list of algae proposed is based on the literature concerning biofuel production, on the commercially produced algae and on the feedback from the questionnaire in deliverable 1.1. Detailed description of biotechnology is provided only for pivotal taxa or group of taxa, that are actually produced at least at pilot scale. These taxa represent the reference model for those taxa that are not currently exploited.
It was beyond the scope of this document to propose any kind of new or revised taxonomy of algae. The classification reported is based on AlgaeBase (http://www.algaebase.org/) and Tree of Life project (http://tolweb.org/tree/).
The other aquatic biomasses species reported are all invasive weeds that have been proposed as biofuel crops. The classification reported is based on ITIS Catalogue of Life 2010 (http://www.catalogueoflife.org/annual‐checklist/2010/search/all) and US Department of Agriculture PLANTS Database (http://plants.usda.gov/).
1.3 Target groups Since the major gaps in knowledge concerning algal biofuels are the lack of operating pilot scale and commercial algal biofuels production facilities, the target audience for this report are all interested

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 9 of 258
stakeholders in Europe’s energy, agricultural and environmental policy, such as policy makers, NGOs, research infrastructures, interested industries and financial institutions. The conclusion of this report should be an appeal for rapid and significant investments into algae related biomass production in the form of public or privately managed pilot or production facilities with the aim of testing a multitude of operational parameters for increasing yields and sustainability while reducing production costs of algal biomass as much as possible. Taking into account the long term multi trillion Euro per year energy and by‐product market, this report aims at convincing all involved potential stakeholders and investors to designing long term strategies for funding an algae based fuel and biomass development program that may provide the necessary insight on their undoubtedly huge potential.
1.4 Problems incurred It is a disturbing fact that today no results on true sustained biomass production yields for biofuels production are available, and no functional biofuel production plant is accessible anywhere around the world. Thus all recent publications in the field, be it yields, economics or LCA, remain pure speculation and demand greatest care in their interpretation.
During the last 10 years algal biofuels companies, driven by large investments from venture capital, have aimed to demonstrate a potential for rapidly achieving economic profitability. This trend lead to high degrees of secrecy surrounding mysterious processes developed whose technical and scientific soundness cannot be confirmed nor discarded due to lacking access to raw data and facilities. The summary presented below therefore relies on publications and patents released by mostly academic institutions and a few open minded companies feeling that little in terms of technology and biology in the process of algal biomass production deserves this degree of secrecy. Nevertheless it cannot be excluded that certain secret breakthroughs may have been achieved recently that would put the state of the art significantly ahead of what is being presented here.
1.5 Common erroneous "myth" In contrast to often voiced opinions, algae are not significantly more efficient in biomass production than other plants grown under optimal conditions. The most common error is comparing biomass doubling time or specific growth rates, which indicate the rate of biomass accumulation under exponential growth conditions, where indeed algae and cyanobacteria may multiply several times per day. However, those conditions are possible under very low biomass densities only that are not applicable to large scale algal cultivation since actual biomass produced per day is in fact lower than in cultures with higher biomass densities, where all the incoming light is captured by algae and used for photosynthesic biomass production.
If such doubling times of exponentially growing cultures are being applied to denser cultures (which could be done with heterotrophic organisms by increasing the food input) in fact very easily fantastic daily growth rates can be assumed. However other than in heterotrophic culture conditions the one and only energy source for growth of algae is incoming light energy that is transformed with an efficiency of around 3% into biomass. Under absolutely optimized conditions in terms of temperature, light intensity, mixing and CO2 supply, higher photosynthetic efficiencies of up to 7% may be achieved, however under exponential increase in bioreactor and maintenance costs that are generally claimed not to be covered by increased yield, and also require far higher energy inputs leading to a negative ratio between energy input and gain in form of algal biomass for example in tubular photobioreactors (Jorquera et al. 2010).
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 10 of 258
2 Criteria for strain selection
The objective was to agree on criteria for both micro‐ and macro algae on which species can be selected for their suitability for biofuel production. These criteria should ideally be quantitatively measurable. For all criteria one should keep an outdoor, large scale system in mind, because for lab (scale) experimentations different criteria may apply.
2.1 Productivity This includes productivity of biomass and of specific biomass components (e.g. lipids). In order to be able to compare algae with traditional crops and with each other in terms of productivity, the most objective criterion is to use photosynthetic efficiency as a measure for productivity. Basically this means the % of available light (energy) that is converted into biomass or specific biomass components. In this way all species and all cultivation systems can be compared.
2.2 Robustness This is a rather vague term which includes resistance to many extreme conditions. This criterion can be best assessed using table in which the resistance to these several conditions is scored.
Table 2‐ Conditions of robustness.
Condition Relevant for Range pH Reduce risk of infection
CO2 transfer i.e. <4 and >10
Oxygen concentration Closed photobioreactors >20% Temperature Outdoor cultivation
Open water cultivation Large range to accommodate day/night and seasonal fluctuation (e.g. 10 – 40 °C)
Salinity Cultivation in fresh / sea / brackish water Reduce risk of infection
e.g 0‐10% salinity
Organic contaminants Ability to grow on wastewater / flue gas
Concentration of organic contaminants that still allows good growth
2.3 Harvestability For microalgae this will mainly be the sedimentation rate and the possibilities for induced‐ or auto flocculation. For macroalgae this includes the possibilities for mechanical harvesting or harvesting by hand.
2.4 Biomass composition This should include a breakdown of the total biomass composition in:
• total caloric value of the biomass (for burning it),
• % lipids and lipid composition (for biodiesel),

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 11 of 258
• % starch and carbohydrate composition (for bio ethanol and to identify higher value byproducts (i.e. agar),
• % protein and protein composition (soluble/insoluble for food/feed purposes),
• presence of heavy metals or toxins (specification).
2.5 Processability / extractability This should include relevant aspects for biorefinery, such as the cell volume, thickness/toughness of the cell wall and the presence of tough fibers (macroalgae) and the moisture content. A measure for this could be the energy input per gram of dry weight necessary for full biorefinery.
2.6 Added value of coproducts Does the organism produce any by‐ or co‐product that have an intrinsic added value, such as carotenoids. This is important to reduce the costs of the final biofuel product. Here a specification of the compounds and their expected added value per gram of dry biomass should be indicated.
2.7 Local origin of strains The use of locally selected strains may be of significance both for ease of management and for reasons of sustainability Based on criteria of the 'Roundtable on Sustainable Biofuels' (http://rsb.epfl.ch/). Non‐native potentially invasive biofuels crops should not be used in open cultivation systems, and adherence to this rule will require the identification and use of locally isolated algal strains. Furthermore such strains may have unique adaptations to the local climate, water and possible parasites that imported or even laboratory grown strains may not have.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 12 of 258
3 Biology of algae
Algae are an assemblage of organisms that have been generally described as photoautotrophic unicellular or multicellular organisms lacking complex morphological organization, and as such have been reclassified several times in recent biology according to technical advances on the basis of differences in subcellular organization, or later of molecular genetic characteristics that will allow, when sufficient data will be available, precise determination of evolutionary relationships. Historically, algae included also cyanobcateria, prokaryotic oxygenic phototrophs.
Many microalgae grow quite rapidly and their reproduction occurs primarily by vegetative (asexual) cell division, although sexual reproduction can occur in many species under appropriate growth conditions. There are several main groups of microalgae, which differ primarily in pigment composition, biochemical constituents, ultrastructure, and life cycle. Five groups were of primary importance: diatoms (Bacillariophyta), green algae (Chlorophyta), Prymnesiophyta or Haptophyta and Eustigmatophyta together with the prokaryotic blue‐green algae, or cyanobacteria.
Recent molecular genetic studies confirmed that photoautotrophic eukaryotes belong to several highly diverse groups of organisms and are the result of different and independent events of secondary endosymbioses. As a consequence algae are a genetically widely diverting group of organisms, a fact that will require due attention when developing tools such as transformation or genetic engineering etc.
3.1 Cyanobacteria Cyanobacteria are prokaryotic photoautotrophic microorganisms that can be found in almost every environment, from oceans to freshwater to bare rock to soil. Though the prokaryotic Cyanobacteria (commonly referred to as blue‐green algae) were traditionally included as "algae" in older textbooks, many modern sources regard this as outdated as they are now considered to be bacteria.
Classification ‐ The cyanobacteria were traditionally classified by morphology according to the International Code of Botanical Nomenclature into five orders: Chroococcales, Pleurocapsales, Oscillatoriales, Nostocales and Stigonematales. Starting from the 1970s cyanobacteria have been classified also according to the International Code of Nomenclature of Bacteria and they were treated in the Bergey’s Manual of Systematic Bacteriology Volume 3 of the 1989 edition, then updated in volume I of the edition of 2004, where cyanobacterai are subdivided in five subsections, I‐V, corresponding to the orders of the Botanical Code, except that the Prochlorales have been included within the cyanobacteria and precisely in the I subsection (Komárek and Anagnostidis, 1986; Anagnostidis and Komárek, 1988; Castenholz and Waterbury, 1989; Komárek and Anagnostidis, 1989; Anagnostidis and Komárek, 1990; Castenholz, 2001;Oren, 2004; Wilmotte and Herdman, 2001; Herrero and Flores (eds), 2008).
Cell structure ‐ The first two subsections include unicellular cyanobacteria (Castenholz and Waterbury, 1989; Castenholz, 2001). The members of Chroococcales are unicellular cyanobacteria that reproduce by binary fission or budding. Cells are coccoid or rod shaped and can vary in length from 0.5 to 30 μm. Division can occur in one to three successive planes, so that cells can be single or in colonies. The classic taxonomic criterion has been the cell morphology and the plane of cell division. In

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 13 of 258
Pleurocapsales, cyanobacteria reproduce by multiple fission which generates small spherical cells named baeocytes, that can be motile or not according to the genus (Herdman and Rippka, 1988b). In unicellular forms there is only multiple fission, whereas in colonial forms after binary fission on different planes some of the cells undergo multiple fission.
The remaining sections include filamentous cyanobacteria. In Oscillatoriales (Castenholz and Waterbury, 1989; Castenholz, 2001), the cells are uniseriately arranged and do not form specialized cells (akinetes and heterocysts). They reproduce by binary fission in a single plane. Filament diameter varies from 0.4 to 100 μm. Outside the cell wall a sheath may be present. In this case terminal hormogonia of short length can glide out of the sheath and eventually form new sheaths. New filaments (or trichomes) are originated from fragmentation in correspondence to a dead cell or certain cells (necridial cells) are purposely destined to die. In Nostocales and Stigonematales (Castenholz and Waterbury, 1989; Castenholz, 2001) the cells have the ability to differentiate cells like heterocysts and akinetes. Nostocales are filamentous cyanobacteria dividing only by binary fission in one plane, though some genera produce false branching. Filaments may be composed of cells of uniform diameter or by cells with decreasing diameter towards the end of the filament (tapering trichomes). Heterocysts may be terminal and intercalary or only terminal in different genera. Motile trichomes (hormogonia) can be formed for dispersion in some genera (Herdman and Rippka, 1988b). Resistance cell (akinetes) may be formed under unfavorable conditions (Herdman and Rippka, 1988a). Stigonematales, unlike Nostocales, includes species with truly branched trichomes. Within this class there is the maximum degree of complexity and differentiation of all the cyanobacterial groups. Longitudinal or oblique cell division occurs in addition to tranverse division, so that periodic true branching and, in some cases, multiseriate trichomes are formed. Hormogonia may be formed, even if reproduction occurs mainly by random breakage of the trichome. Akinets can be produced in some genera. Heterocysts are both intercalary and terminal. Cell diameter varies within a “trichome” as secondary branches are usually narrower.
In cyanobacteria (Castenholz and Waterbury, 1989), generation times are usually higher than 24 h, though some unicellular and oscillatorian strains can duplicate in 4 h. Some genera can have complex morphogenetic cycles including filamentation, aseriate phases, dispersal through hormogonia and akinetes production.
Cyanobacteria share the basic cell characteristics with the other Bacteria (Stanier and Cohen‐Bazire, 1977; Castenholz and Waterbury, 1989). The cell wall is of the Gram‐negative type, though the peptidoglycan layer is considerably thicker than in the other Gram‐negative bacteria. Many cyanobacteria have a sheath or glycocalix or capsule or gel, mucilage or slime outside the outer membrane of the cell wall, mainly composed of polysaccharides. Some sheaths can have a microfibrillar structure and can become laminated with aging of the trichome. Some cyanobacteria (and many hormogonia) can contain gas‐vesicles that allow buoyancy in the water column.
Cyanobacteria have an elaborate and highly organized system of internal membranes which function in photosynthesis (thylakoids) (Stanier and Cohen‐Bazire, 1977; Castenholz and Waterbury, 1989). The lipophilic pigments chlorophyll a (both reaction centers and antenna) and photosynthetic carotenoids are located within the thylakoids, while the hydrophilic antenna pigments (allophycocyanin‐APC‐, phycocyanin ‐PC‐ and, where they are present, phycoerythrin –PE‐ or phycoerythrocyanin ‐PEC) are located in the phycobilisomes which are attached to the outside of the thylakoid membranes. The phycobilisome is haemidiscoidal and is composed of stacks of biliproteins in

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 14 of 258
the order (from inside to outside) APC, PC, PE or PEC. Genera belonging to the former group of the Prochlorales lack phycobilisomes and have chlorophyll b as antenna pigment. The cyanobacterium Acaryochloris marina has been reported to contain chlorophyll d instead of chlorophyll a as light harvesting pigment, so that its photosynthetic process depends on far‐red light (710‐718 nm) (Miyashita et al., 2003).
The reserve carbohydrate is glycogen (Stanier and Cohen‐Bazire, 1977; Castenholz and Waterbury, 1989). Cyanobacteria contain also cyanophycin, a nitrogen reserve polymer made of arginine and aspartic acid, polyphosphate granules and carboxisomes, that are a cell reserve of the photosynthesis key enzyme rubisco (ribulose‐1,5‐biphosphate carboxylase). Some cyanobacteria also contain poly‐β‐hydroxybutyrate granules.
Physiology ‐ Photosynthesis in cyanobacteria (Wolk, 1973; Stanier and Cohen‐Bazire, 1977; Castenholz and Waterbury, 1989)uses water as an electron donor and produces oxygen as a by‐product. This water‐oxidizing process is accomplished by coupling the activity of photosystem (PS) II and I (Z‐scheme). Under anaerobic conditions some genera (belonging to the I and III subsections) are able to carry out anoxygenic photosynthesis using only PS I to carry out cyclic photophosphorylation and obtain ATP, and using electron donors other than water (hydrogen sulfide or thiosulphate) (Cohen et al., 1975; Garlick et al., 1977). Carbon dioxide is reduced to form carbohydrates via the Calvin cycle. During respiration reduced NADP is obtained through the pentose phosphate cycle. The plasma membrane contains only components of the respiratory chain, while the thylakoid membrane hosts both respiratory and photosynthetic electron transport.
Most cyanobacteria are obligate photoautotrophs, but some species can grow as heterotrophs in the dark at the expense of glucose, fructose or sucrose. Under anaerobic conditions, some species can perform lactate fermentation (Oren and Shilo, 1979).
Nitrogen fixation occurs both in heterocystous cyanobacteria and in some non‐hetrocystous cyanobacteria. To avoid contact of nitrogenase with oxygen (and then its permanent inactivation) these latter cyanobacteria adopt a temporal separation between the photosynthetic and the nitrogen fixation processes (Bergman et al., 1997). Increased respiration rates allow to control the oxygen concentration inside the cell, due to diffusion, necessary to carry out cell metabolism. In heterocystous forms, the nitrogen fixation process is spatially separated from the oxygenc photosynthesis. Nitrogen fixation is carried out in specialized cells, the heterocysts (Adams and Duggan, 1999). These have many characteristics that allow to reduce diffusion of oxygen, such as a thick cell wall surrounded by a complex external envelope and a reorganization of the photosynthetic apparatus: lack of PS II to avoid internal oxygen production, presence of PS I to obtain ATP through cyclic photophosphorilation. Reducing power is obtained from vegetative cells in the form of sugars. Molecular nitrogen is fixed into ammonia and immediately converted to organic form, usually as glutamine. As nitrogen fixation is a very energy‐consuming process, nitrogenese is produced and heterocysts are differentiated only in the absence of combined nitrogen in the environement surrounding the cell.
Ecology ‐ Cyanobacteria are the only group of organisms that are able to reduce nitrogen and carbon in aerobic conditions, a fact that may be responsible for their evolutionary and ecological success (Whitton and Potts (eds), 2000). They contribute significantly to global ecology and the oxygen cycle. The large amounts of oxygen in the atmosphere originally derive from the activities of ancient cyanobacteria. The tiny marine cyanobacterium Prochlorococcus was discovered in 1986 (Chisholm et

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 15 of 258
al., 1988) and, together with the picoplanktonic cyanobacteria, accounts for up to half of the primary production of waters, from oligotrophic open ocean to estuarine ecosystems.
Due to their ability to fix nitrogen in aerobic conditions they are often found as symbionts (Rai et al. (eds), 2002) with a number of other groups of organisms such as fungi (lichens), corals, pteridophytes (Azolla), angiosperms (Gunnera), protists (including some diatoms), and sponges. The rice paddies of Asia rely on nitrogen‐fixing cyanobacteria as fertilizers, both free biomass and symbiont to the fern Azolla.
They can occur as planktonic cells (thanks to the buoyancy ability) or form phototrophic biofilms in freshwater and marine environments, they occur in damp soil, or even temporarily moistened rocks in deserts (Whitton and Potts (eds), 2000). Some live in the fur of sloths, providing a form of camouflage. Aquatic cyanobacteria are probably best known for the extensive and highly visible blooms that can form in both freshwater and marine environments. The association of toxicity with such blooms has frequently led to the closure of recreational waters when blooms are observed. Certain cyanobacteria produce cyanotoxins (Chorus and Bartram, 1999) including neurotoxins, hepatotoxins, cytotoxins, and endotoxins. Examples of cyanotoxins are anatoxin‐a, anatoxin‐as, aplysiatoxin, saxitoxin, cylindrospermopsin, microcystins, nodularin. These toxins can be dangerous to humans and animals. Several cases of human poisoning have been documented. Recent studies suggest that significant exposure to high levels of BMAA a non‐proteic aminoacid produced by many cyanobacteria could be among the causes of neurodegenerative diseases such as Amyotrophic Lateral Sclerosis.
Genome sequencing ‐ The unicellular cyanobacterium Synechocystis sp. PCC6803 was the third prokaryote and first photosynthetic organism whose genome was completely sequenced (Kaneko et al. 1996) . It continues to be an important model organism. Today over 40 complete cyanobacterial genomes are known (www.ncbi.nlm.nih.gov). The smallest genomes have been found in Prochlorococcus spp. (1.7 Mb) and the largest in Nostoc punctiforme (9 Mb). Those of Calothrix spp. are estimated at 12‐15 Mb, as large as yeast.
Relationship to chloroplasts ‐ Chloroplasts found in eukaryotes (algae and plants) likely evolved from an endosymbiotic relation with cyanobacteria. This endosymbiotic theory is supported by various structural and genetic similarities. Primary chloroplasts are found among the "true plants" or green plants as well as among the red algae and glaucophytes, marine species which contain phycobilins. It now appears that these chloroplasts probably had a single origin, in an ancestor of the clade called Primoplantae. Other algae likely took their chloroplasts from these forms by secondary endosymbiosis or ingestion.
3.2 Chlorophyta (Green Algae) The green algae are a large group of algae from which the embryophytes (higher plants) emerged (Jeffrey et al., 2004). The group including both green algae and embryophytes is monophyletic (and often just known as kingdom Plantae). The green algae include unicellular and colonial flagellates, usually but not always with two flagella per cell, as well as various colonial, coccoid, and filamentous forms. In the Charales, the closest relatives of higher plants, full differentiation of tissues occurs (Thomas, 2002). There are about 6,000 species of green algae. Many species live most of their lives as single cells, while other species form colonies or long filaments.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 16 of 258
Some species of green algae, particularly of genera Trebouxia and Pseudotrebouxia (Trebouxiophyceae), can be found in symbiotic associations with fungi to form lichens. In general the fungal species that partner in lichens can not live on their own, while the algal species is often found living in nature without the fungus.
Prasinophyceae are a class of primitive eukaryotic marine green algae (Sym and Pienaar, 1993). Their best known genus is Ostreococcus, which is considered to be the smallest (ca. 0.95 μm) free‐living eukaryote and which has been detected in marine samples around the world (Courties et al., 1994). Prasinophyceae are thought to have low cellular complexity, that is, they possess single, multiple or no flagella and contain only a single chloroplast and a single mitochondrion. They also have very small genomes for a eukaryote (about 12 Mbp), and the genomes of two Ostreococcus species, taurii and lucimarinus, have been completely sequenced. It has been suggested that a prasinophyceae‐like flagellate was the ancestor to Chlorophyta and Streptophyta (Kapraun, 2007). A study of photosynthetic gene‐sequence diversity (rbcL) in the Gulf of Mexico indicated that Prasinophyceae are particularly prevalent at the Subsurface Chlorophyll Maximum (SCM) (Warwick et al., 2003) and several different ecotypes of Ostreococcus have been detected in the environment (Guillou et al., 2004). These ecotypes are distinguished by their adaptation to light intensities.
The Chlorophyceae are one of the classes of green algae, distinguished mainly on the basis of ultrastructural morphology. For example the chlorophycean CW clade, and chlorophycean DO clade, are defined by the arrangement of their flagella. Members of the CW clade have flagella that are displaced in a "clockwise" (CW, 1–7 o'clock) direction eg. Chlamydomonadales. Members of the DO clade have flagella that are "directly opposed" (DO, 12–6 o'clock) e.g. Sphaeropleales.
They share many similarities with the higher plants, including the presence of asymmetrical flagellated cells, the breakdown of the nuclear envelope at mitosis, and the presence of phytochromes, flavonoids, and the chemical precursors to the cuticle (Raven et al., 2005).
Cell structure ‐ Almost all forms have chloroplasts. These contain chlorophylls a and b, giving them a bright green colour (as well as the accessory pigments beta carotene and xanthophylls), and have stacked thylakoids (van den Hoek et al., 1995). All green algae have mitochondria with flat cristae. The storage product for members of this group is true starch, amylose, and amylopectin (α‐1,4‐linked polyglucans), and is found inside the chloroplasts. The starch (seen as whitish granules with the TEM) can often be observed surrounding the pyrenoid, a distinct spherical structure embedded in the chloroplast. There may be more than one pyrenoid or the prenoid is not always present (e.g., Ankistrodesmus and Tetraedron) or the pyrenoid is lacking. In most representative taxa, the cells are surrounded by a cellulose cell wall (Wehr and Sheath, 2003). Some taxa may also have chitin or sporopollenin deposited on the wall. This gives added strength and is thought to help prevent desiccation. Some taxa have wall ornamentation, such as scales, a rough texture, thick walls with distinct layers, warts, ridges, and spines. The Volvocales usually have cell walls, loricae, or gelatinous matrices and the main component of the cell walls is glycoprotein, rather than cellulose. The flagellated green microalgae can have from one to eight isokont flagella.
The Chlorophyta macroalgae share the following common characteristics: flagella of swimming cells in pairs or multiples of two; stellate structure linking nine pairs of microtubules at basal body transition zone; thylakoids single or stacked; plastid with two membranes without periplastid endoplasmic reticulum; starch inside plastid; glycolate dehydrogenase present; cell wall, when present, of cellulose; cell division without phragmoplast.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 17 of 258
Origin ‐ The chloroplasts of green algae are bound by a double membrane, so presumably they were acquired by direct endosymbiosis of cyanobacteria. A number of cyanobacteria show similar pigmentation, but this appears to have arisen more than once, and the chloroplasts of green algae are no longer considered closely related to such forms. Instead, the green algae probably share a common origin with the red algae.
Phylogeny ‐ The orders outside the Chlorophyta are often grouped as the division Charophyta, which is paraphyletic to higher plants, together comprising the Streptophyta. Sometimes the Charophyta is restricted to the Charales, and a division Gamophyta is introduced for the Zygnematales and Desmidiales. In older systems the Chlorophyta may be taken to include all the green algae, but taken as above they appear to form a monophyletic group. One of the most basal green algae is the flagellate Mesostigma, although it is not yet clear whether it is sister to all other green algae, or whether it is one of the more basal members of the Streptophyta.
Reproduction ‐ Most green algae can proliferate vegetatively by cell division, often the mother cell can divide into up to 16 offspring before releasing them. They often can profliferate sexually whereby haploid algae cells of opposing mating type (containing only one copy of their DNA) can fuse with other haploid cells to form diploid zygotes. They can also follow a reproduction cycle called alternation of generations. Reproduction varies from fusion of identical cells (isogamy) to fertilization of a large non‐motile cell by a smaller motile one (oogamy). However, these traits show some variation, most notably among the basal green algae, called prasinophytes.
Some taxa produce motile cells (planospores). Planospores may be asexual zoospores or sexual gametes. Aplanospores (nonmotile cells) may be also produced.
When filamentous algae conjugate, they form bridges between cells, and leave empty cell walls behind that can be easily distinguished under the light microscope.
The species of Ulva are reproductively isomorphic, the diploid vegetative phase is the site of meiosis and releases haploid zoospores, which germinate and grow producing a haploid phase alternating with the vegetative phase.
3.3 Rhodophyta (Red Algae) The Rhodophyta are a distinct eukaryotic lineage characterized by the accessory photosynthetic pigments phycoerythrin, phycocyanin and allophycocyanin arranged in phycobilisomes, and the absence of flagella and centrioles (Woelkerling, 1990). This is a large assemblage of between 2500 and 6000 species in about 670 largely marine genera (Woelkerling, 1990) that predominate along the coastal and continental shelf areas of tropical, temperate and cold‐water regions (Lüning, 1990). Red algae are ecologically significant as primary producers, providers of structural habitat for other marine organisms, and their important role in the primary establishment and maintenance of coral reefs. Red algae are common and widespread, and ecologically important.
Cell structure ‐ Red algae have a number of general characteristics that in combination distinguish them from other eukaryotic groups:
• absence of flagella and centrioles, • floridean starch as a storage product and the storage of starch in the cytoplasm, • phycoerythrin, phycocyanin, and allophycocyanin as accessory pigments,

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 18 of 258
• unstacked thylakoids in plastids, • no chloroplast endoplasmic reticulum.
The rhodophyta exhibit the following common characteristics: they are unicellular to multicellular (up to 1 m), mostly free‐living but in some cases parasitic or symbiotic, with chloroplasts containing phycobilins. Cell walls are made of cellulose with mucopolysaccharides (mainly agars and carrageenans) penetrated in many red algae by pores mostly blocked by proteins (complex referred to as pit connections). Their mitochondria have flat cristae sometimes associated with forming faces of dictyosomes. Thylakoids are single, with phycobilisomes, plastids with peripheral thylakoid. During mitosis, the nuclear envelope mostly remains intact but some microtubules of spindle extend from noncentriolar polar bodies through polar gaps in the nuclear envelope.
Phylogeny ‐ Traditionally the red algae were divided into two Classes the Bangiophyceae and Florideophyceae. Alternatively a single Class, the Rhodophyceae and two Subclasses, Bangiophycidae and Florideophycidae are used. Based on ultrastructure and molecular evidence the Bangiophyceae is now accepted as a paraphyletic group, while the Florideophyceae is considered to be monophyletic based on two synapomorphic characters ‐ presence of a filamentous gonimoblast and tetrasporangia (Garbary and Gabrielson, 1990 [and references within], Ragan et al., 1994).
Reproduction ‐ They usually have separated phases of vegetative growth and sexual reproduction.
3.4 Heterokontophyta The Heterokontophyta are a major line of eukaryotes. Most are algae, ranging from the giant multicellular kelp to the unicellular forms. The name heterokonts refers to the motile life cycle stage, in which the flagellate cells possess two different shaped flagella (Leipe et al., 1994; Patterson, 1989).
Cell structure ‐ Heterokont algae are surrounded by four membranes, which are counted from the outermost to the innermost membrane. The first membrane is continuous with the host's chloroplast endoplasmic reticulum, or cER. The second membrane presents a barrier between the lumen of the endoplasmic reticulum and the primary endosymbiont or chloroplast, which represents the next two membranes, within which the thylakoid membranes are found. This arrangement of membranes suggest that heterokont chloroplasts were obtained from the reduction of a symbiotic red algal eukaryote, which had arisen by evolutionary divergence from the monophyletic primary endosymbiotic ancestor that is thought to have given rise to all eukaryotic photoautotrophs. The chloroplasts usually contain chlorophyll a and chlorophyll c, and usually the accessory pigment fucoxanthin, giving them a golden‐brown or brownish‐green color.
Many heterokonts are unicellular flagellates, and most others produce flagellate cells at some point in their life‐cycle, for instance as gametes or zoospores. The name heterokont refers to the characteristic form of these cells, which typically have two unequal flagella. The anterior or tinsel flagellum is covered with lateral bristles or mastigonemes, while the other flagellum is whiplash, smooth and usually shorter, or sometimes reduced to a basal body. The flagella are inserted subapically or laterally, and are usually supported by four microtubule roots in a distinctive pattern.
Mastigonemes are manufactured from glycoproteins in the cell's endoplasmic reticulum before being transported to its surface. When the tinsel flagellum moves, these create a backwards current, pulling the cell through the water or bringing in food. The mastigonemes have a peculiar tripartite

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 19 of 258
structure, which may be taken as the defining characteristic of the group, thereby including a few protists that do not produce cells with the typical heterokont form. They have been lost in a few lines.
Origins ‐ Most basal heterokonts are colorless. This suggests that they diverged before acquisition of chloroplasts within the group. Fucoxanthin‐containing chloroplasts are also found among the haptophyta. These two groups may have a common ancestry, and possibly also a common phylogenetic history with cryptophyta. This may be interpreted as suggesting that the ancestral heterokont was an alga, and all colorless groups arose through loss of the secondary endosymbiont and its chloroplast.
Phylogeny ‐ As noted above, classification varies considerably. Originally the heterokont algae were treated as two divisions, first within the kingdom Plantae and later the Protista.
In this scheme, however, the Chrysophyceae are paraphyletic to both other groups. As a result, various members have been given their own classes and often divisions. Recent systems often treat these as classes within a single division, called the Heterokontophyta, Chromophyta or Ochrophyta. This is not universal, however ‐ for instance Round et al. (1990) treat the diatoms as a division.
The discovery that oomycetes and hyphochytrids are related to these algae, rather than fungi as previously thought, has led many authors to include them among the heterokonts. Should it turn out that they evolved from colored ancestors, the group would be paraphyletic in their absence. Once again, however, usage varies. Patterson (1999) named this extended group the stramenopiles, characterized by the presence of tripartite mastigonemes, mitochondria with tubular cristae, and open mitosis. He used the stramenopiles as a prototype for a classification without Linnaean ranks. Their composition has been essentially stable, but their use within ranked systems varies. Cavalier‐Smith (1981) treats the heterokonts as identical in composition with the stramenopiles; this is the definition followed here. He has proposed placing them in a separate kingdom Chromalveolata, together with the haptophytes, cryptomonads and alveolates. This is one of the most common revisions to the five‐kingdom system, but has not been generally adopted, partly because some biologists doubt their monophyly. A few treat the Chromalveolata as identical in composition with the heterokonts, or list them as a kingdom Stramenopila.
3.4.1 Phaeophyceae (Brown algae)
The class of the Phaeophyceae (Guiry and Guiry, 2007), or brown algae, is a large group of mostly marine multicellular algae, including many seaweeds of colder Northern Hemisphere waters. Brown algae are unique among heterokonts in developing into multicellular forms with differentiated tissues, but they reproduce by means of flagellate spores and gametes, which closely resemble other heterokont cells. Genetic studies show their closest relatives to be the yellow‐green algae.
They play an important role in marine environments both as food, and for the habitats they form. For instance Macrocystis, a member of the Laminariales or kelps, may reach 60 m in length, and forms prominent underwater forests. Another example is Sargassum, which creates unique habitats in the tropical waters of the Sargasso Sea. Many brown algae such as members of the order Fucales are commonly found along rocky seashores. Some members of the class are used as food for humans. Worldwide there are about 1500‐2000 species of brown algae. Some species are of sufficient commercial importance, such as Ascophyllum nodosum, that they have become subjects of extensive research in their own right (Senn, 1987; van den Hoek, 1995).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 20 of 258
Algal structure ‐ Filamentous, syntagmatic or parenchymatous; cell wall present, containing alginate compounds and cellulose; plasmodesmata or pores between cells in parenchymatous forms; chloroplasts with girdle lamella; outer chloroplast endoplasmic reticulum membrane with direct membrane connection to the outer nuclear envelope membrane; plastid DNA with ring‐type genophore; eyespots present or absent; plastid pigments include chlorophylls a and c1 and c2, fucoxanthin, and violaxanthin; swimming cells with two flagella usually inserted laterally, one anteriorly directed, one posteriorlydirected; usually four microtubular kinetosome roots but no striated kinetosome root (rhizoplast); flagellar transitional helix typically with 6 gyres located above the major transitional plate; no paraflagellar rod; little to substantial tissue differentiation occurring in parenchymatous forms.; macroscopic or microscopic, some polysiphonous; some form crusts, cushions or are hollow and others grow to form large leathery fronds (Jones, 1962).
Evolutionary history ‐ Phaeophyta evolved from the phaeothamniophyceae between 150 and 200 million years ago. Claims that earlier (Ediacaran) fossils are brown algae have since been dismissed (Loeblich, 1974; Medlin et al., 1997; Lee, 2008). The lineages of brown algae diverged in the following order, from oldest to youngest: Dictyotales; Sphacelariales; Cutleriales; Desmarestiales; Ectocarpales; Laminarales; Fucales. Their occurrence as fossils is rare due to their generally soft‐bodied habit, and scientists continue to debate the identification of some finds. Only a few species of brown algae deposit significant quantities of minerals in or around their cell walls. Other algal groups, such as the red algae and green algae have a number of calcareous members, which are more likely to leave evidence in the fossil record than the soft bodies of most brown algae. Miocene fossils of a soft‐bodied brown macro algae, Julescrania, have been found well‐preserved in Monterey Formation diatomites, but few other dubiously assigned fossils, particularly of older specimens are known in the fossil record (Coyer et al., 2001).
Life cycle ‐ The life cycle shows great variability from one group to another. However the life cycle of Laminaria consists of the diploid generation, that is the large kelp well known to most people. It produces sporangia from specialised microscopic structures, these divide meiotically (meiosis) before they are released. As they are haploid there are equal numbers of male and female spores (Thomas, 2002). With the exception of the Fucales all brown algae have a life cycle which consists of an alternation between haploid and diploid forms.
Ecology ‐ Brown algae have adapted to a wide variety of marine ecological niches including the tidal splash zone, rock pools, the whole intertidal zone and relatively deep near shore waters. They are an important constituent of some brackish water ecosystems, and four species are restricted to life in fresh water (Lee, 2008). A large number of Phaeophyceae are intertidal or upper littoral, and they are predominantly cool and cold water organisms that benefit from nutrients in up welling cold water currents and inflows from land; Sargassum being a prominent exception to this generalisation. Brown algae growing in brackish waters are almost solely asexual (Lee, 2008).
Chemistry ‐ Brown algae have a δ13C value between −‐20.8‰ – −10.5‰, in contrast with red algae and greens. This reflects their different metabolic pathways (Fletcher et al., 2004).
They have cellulose walls with alginic acid; fucoidin also important in amorphous section of cell walls. A few species (of Padina) calcify with aragonite needles (Lee, 2008).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 21 of 258
3.4.2 Eustigmatophyceae
Eustigmatophyceae (Hibberd, 1981; Andersen et al., 1998) are a small group (7 genera; 12 species) of eukaryotic algae that includes marine, freshwater and soil‐living species (van den Hoek et al., 1995). In terms of ecology, eustigmatophyceae occur as photosynthetic autotrophs across a range of systems. Most genera live in freshwater (Fawley, 2007) or in soil, although Nannochloropsis (Guiry and Guiry, 2007) contains marine species of picophytoplankton (2 → 4 μm).
Cell structure ‐ All eustigmatophyceae are unicellular, with coccoid cells and polysaccharide cell walls. They contain one or more yellow‐green chloroplasts, which contain chlorophyll a and the accessory pigments violaxanthin and β‐carotene. Eustigmatophyte zoids (gametes) possess a single or pair of flagella, originating from the apex of the cell. Unlike other heterokontophyta, eustigmatophyceae zoids do not have typical photoreceptive organelles (or eyespot); instead, an orange‐red eyespot outside of the chloroplast is located at the anterior end of the zoid.
3.4.3 Other classes
Yellow‐green algae or Xanthophyceae are an important group of heterokont algae. Most live in freshwater, but some are found in marine and soil habitats. They vary from single‐celled flagellates to simple colonial and filamentous forms. Xanthophyceae chloroplasts contain the photosynthetic pigments chlorophyll a, chlorophyll c, β‐Carotene, and the xanthophylls vaucheriaxanthin, duatoxantin and diadinoxanthin. Unlike other heterokonts, their chloroplasts do not contain fucoxanthin, which accounts for their lighter colour. Its storage polysaccharide is chrysolaminarin. Xanthophyceae cell walls are produced of cellulose and hemicellulose. They appear to be the closest relatives of the brown algae (Stace, 1991). Recent ultrastructural and molecular phylogenetic DNA (nuclear and plastid) research shows that the Mischococcales might be paraphyletic, and the Tribonematales and Botrydiales polyphyletic, and suggests two orders at most be used until the relationships within the division are sorted (Adl et al., 2005).
The golden algae or Chrysophyceae are a large group of algae found mostly in freshwater. Originally they were taken to include all such forms except the diatoms and multicellular brown algae, but since then they have been divided into several different groups based on pigmentation and cell structure. They are now usually restricted to a core group of closely related forms, distinguished primarily by the structure of the flagella in motile cells, also treated as an order Chromulinales. It is possible membership will be revised further as more species are studied in detail. Most members are unicellular flagellates, with either two visible flagella, as in Ochromonas, or sometimes one, as in Chromulina. Most genera have no cell covering, some have loricae or shells. Some members are generally amoeboid, with long branching cell extensions, though they pass through flagellate stages as well. Other members are non‐motile. Cells may be naked and embedded in mucilage, such as Chrysosaccus, or coccoid and surrounded by a cell wall, as in Chrysosphaera. A few are filamentous or even parenchymatous in organization, such as Phaeoplaca.
3.5 Labyrinthulea (phylum Heterokonta) Thraustochytrids are exclusively marine heterotrophic protists that feed non‐phagotrophically as saprobes, epibionts on algae (micro and macroalgae) or more rarely as parasites of microalgae (such as Skeletonema) and animals. These unicellular eukaryotic protists are a common component of marine

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 22 of 258
microbial consortia. Whether to include or not this group among algae is still object of debate in the scientific community, but due to their phylogenetic vicinity and to the fact that products from these organism are on the market as “algal products” they are discussed in this document.
Cell structure – Thraustochytriidae present an ectoplasmic net (EN) that forms a branched network of plasma membrane extensions, associated with an organelle termed the bothrosome or sagenogenetosome (sagenogen) at the periphery of the cell. The EN contributes to the increased surface area of the cell and contains hydrolytic enzymes that are surface‐bound or are secreted into the surrounding medium, helping in the digestion of organic material. The EN also attaches the cells to surfaces and, in the case of thraustochytrids, penetrates organic particles (Raghu Kumar, 2002). Their cell walls are composed of non‐cellulosic scales and contain sulphated polysaccharides, predominantly of galactose or fucose, and proteins (Leander and Porter, 2001; Raghu Kumar, 2002). Vegetative stages of thraustochytrids consist of single cells which are globose to subglobose, measuring 4 to 20 µm in diameter, mostly growing epibiontically on various substrata.
Phylogeny – The Labyrinthulomycota are a group of microorganisms of uncertain taxonomic collocation. They were placed among fungi due to their functional ecology and morphology. They were then placed in the group of Oomycetes due to the presence of biflagellate zoospores. (Raghu Kumar, 1996). Cavalier Smith et al. (1994) includes the Labyrinthulomycetes in the phylum Heterokonta, based on the analysis of the 18S rRNA. The Class Labyrinthulea is then subdivided in the two sub‐classes Thraustochytridae and Labyrinthulidae.
Reproduction – The Labyrinthulomycetes possess simple, asexual life cycles. Most thraustochytrids reproduce by means of zoospores which possess a long anterior, tinsel flagellum and a short, posterior, whiplash flagellum. The mode of production of zoospores varies between genera and forms the major taxonomic criterion (Raghu Kumar, 2002). The cytoplasmic contents of the mature cell, the sporangium, divide directly into zoospores in the genus Thraustochytrium. The cytoplasm escapes as one amoeboid mass, prior to zoospore division in the genus Ulkenia. The genus Schizochytrium is characerised by successive bipartition of a vegetative cell, resulting in a cluster of cells, each of which develops into a zoosporangium or zoospore. Species within the genera are primarily defined by the number of proliferating bodies and the size and shape of the sporangia.
3.6 Bacillariophyta (Diatoms) Despite the abundance and diversity of diatoms in nature, few species are cultured in aquaculture or for biotechnology relevant products (Lebeau and Robert, 2003). Further, only a handful of diatoms have been studied and often only one or a few strains without any information on intraspecific variation. There is a need to identify new diatom strains with as much positive characteristics as possible or to breed or select for improved strains. Therefore, a basic knowledge of physiology, ecology and taxonomy is important.
Diatoms are the most species‐rich and productive group of eukaryotic algae. Over a comparatively short evolutionary time (< 250 Ma) (Sims et al., 2006; Sorhannus, 2007), they have diversified into hundreds of genera and perhaps 200.000 extant species (Mann, 1999). They are extremely abundant in all aquatic ecosystems, occurring in the plankton and benthos of marine and fresh waters (as freeliving organisms or as endosymbionts in e.g. dinoflagellates and foraminifers), and in terrestrial environments, such as damp soils and moist surfaces of rocks and plants, from the tropics to the polar

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 23 of 258
regions (Round et al., 1990). Diatoms have global ecological significance in the carbon and silicon cycles and probably provide 20‐25% of global photosynthetic fixation of carbon and contribute up to 40% of the total oceanic primary production (Field et al., 1998; Falkowski et al., 1998).
Cell structure ‐ Diatoms are unicellular (10‐200 µm), although they can also exist as colonies. The hallmark of the diatom cell is its intricately shaped and ornamented silica cell wall, called the frustule, which consist of two overlapping thecae (Greek diatomos, meaning ‘cut in half ’), each in turn consisting of a valve and a number of hoop‐like or segmental girdle bands. The frustules show a wide diversity in shape, form and ornamentation, which has been the basis of the traditional diatom taxonomy. This cell wall is encased in an organic matrix that is rich in proteins and sugars.
Major pigments of diatoms are chlorophylls a and c, β‐carotene, fucoxanthin, diatoxanthin and diadinoxanthin. Diatoms are primarily photosynthetic. A few, however, are obligate heterotrophs, while others can live heterotrophically in the absence of light, provided an appropriate organic carbon source is available. The main storage compounds of diatoms are lipids (TAGs) and a β‐1,3‐linked carbohydrate known as chrysolaminarin.
Life cycle ‐ Diatoms have a diplontic life cycle with a prolonged vegetative phase during which the cells divide mitotically. Their unique cell wall structure (two overlapping halves) and division pattern in which new cell wall components are formed within the parental cell, cause diatoms to gradually reduce their cell size in the course of the mitotic part of the life cycle (MacDonald‐Pfitzer rule) (Round et al., 1990). Cell size is restored through the development of a special expanding cell called the auxospore, which normally results from sexual reproduction (Chepurnov et al., 2004). Sexual events are vital to re‐establish the initial cell size and to avoid becoming too small for survival (Chepurnov et al., 2004). As genetic recombination is also achieved through sexual reproduction, the obligatory nature of sex in their life cycle may be linked with the evolutionary and ecological success of diatoms.
Classification ‐ Diatoms are recognized as a single well‐defined group at the class or divisional (called Bacillariophyta) level (Mann and Evans, 2007; Kooistra et al., 2007).
Diatoms have historically been divided into two classes or orders, the centric and the pennate diatoms. Most centric diatoms have valves with a radial symmetry (circular or shortly elliptical, triangular, or polygonal) while pennate diatoms are mostly elongate and have a bilateral symmetry. Sexual reproduction in centrics is usually oogamous, with motile uniflagellate sperm. Pennate diatoms are usually isogamous with amoeboid gametes which may be differentiated biochemically and behaviourally (Chepurnov et al., 2004). The pennate diatoms include araphid and raphid forms. The raphe, a slit opening in the cell wall, allow for active gliding along surfaces (Round et al., 1990). A recent re‐evaluation of diatom phylogeny (Medlin and Kaczmarska, 2004) has divided diatoms into three classes: the Coscinodiscophyceae includes radial centric diatoms, the Mediophyceae includes multipolar centrics plus some radial centric diatoms, and the Bacillariophyceae includes pennate diatoms. Members of these classes differ with regards to cell shape, presence/absence of a raphe (in pennate diatoms), structure and arrangement of the Golgi apparatus and chloroplast pyrenoid and mode of sexual reproduction (Medlin and Kaczmarska, 2004).
Traditionally, classification at the order level and below was almost exclusively based on morphological features of the siliceous cell wall (Round et al., 1990). Species boundaries have largely been based on discontinuities in morphological and ultrastructural characters of the frustule. The advent of molecular genetic studies have shown that many morphologically‐defined species are, in

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 24 of 258
fact, assemblages of cryptic species; that is, assemblages of genetically and biologically distinct, albeit often closely related species. Cryptic diversity has been discovered throughout the diatom diversity (reviewed in e.g. Mann and Evans, 2007; Kooistra et al., 2007).
The prevalence of cryptic species has important consequences for understanding patterns in diatom diversity and biogeography, their evolutionary history and their relationship to environmental conditions. Good knowledge of species limits is also a prerequisite for further studies at the species level and below, including within‐species genetic, physiological and biochemical variation, which can have important consequences in commercial uses of diatoms.
Genome sequencing ‐ The entire genomes of the centric diatom, Thalassiosira pseudonana (34 Mb), (Armbrust et al., 2004) and the pennate diatom, Phaeodactylum tricornutum (27 Mb), (Bowler et al., 2008) have been sequenced. A draft sequence is now available for the polar species Fragilariopsis cylindrus (80 Mb) and the toxic coastal species Pseudo‐nitzschia multiseries (300 Mb) (Armbrust, 2009).
One of the outcomes of the sequencing projects thus far is recognition of the unique combination of genes and metabolic pathways that distinguish diatoms from the evolutionarily distinct plant and animal lineages. Enormous amounts of diversity are encapsulated within diatoms. For example, T. pseudonana and P. tricornutum probably diverged from one another only about 90 million years (Ma) ago, yet their genomes are about as different as those of mammals and fish, which diverged about 550 Ma ago (Bowler et al., 2008).
3.7 Haptophyta The Haptophyta, also named Prymnesiophyta, are sometimes classified as Haptophyceae (Satoh et al., 2009). However, although the phylogenetics of this group has become much more well understood in recent years, there remains some dispute over which taxon level is most appropriate.
Cell structure ‐ The chloroplasts are pigmented similarly to those of the heterokonts (Anderson, 2004), but the structure of the rest of the cell is different, so they may be separate lines, whose chloroplasts are derived from similar endosymbionts. The cells typically have two slightly unequal flagella, both of which are smooth, and a unique organelle called haptonema, which is superficially similar to a flagellum but differs in the arrangement of microtubules and in its use. The name comes from the Greek hapsis, touch, and nema, thread. The mitochondria have tubular cristae.
Examples and classification ‐ The best‐known haptophytes are coccolithophores, which have an exoskeleton of calcareous plates called coccoliths. Coccolithophores are some of the most abundant marine phytoplankton, especially in the open ocean and are extremely abundant as microfossils. Other planktonic haptophytes include Chrysochromulina and Prymnesium, which periodically form toxic marine algal blooms, and Phaeocystis, the blooms of which can produce unpleasant foam which often accumulates on beaches. Both molecular and morphological evidence supports their division into five orders.
3.8 Dinophyta (Dinoflagellates) Dinophyta or dinoflagellates are common organisms in all types of aquatic ecosystems. Roughly half of the species in the group are photosynthetic (Gaines and Elbrächter, 1987), the other half is exclusively

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 25 of 258
heterotrophic and feeds via osmotrophy and phagotrophy. As a consequence, they are prominent members of both the phytoplankton and the zooplankton of marine and freshwater ecosystems. They are also common in benthic environments and in sea ice. Approximately 4500 species assigned to more than 550 genera have been described, nearly three quarters of the genera and more than half of the species being fossil. Of the ca. 2000 living species, more than 1700 are marine and about 220 are from freshwater (Taylor et al., 2008). Between the years 2000 and 2007 three new dinoflagellate families, 22 new genera, and 87 new species were described (Centre of Excellence for Dinophyte Taxonomy CEDiT). Recent molecular analyses have shown that there are large numbers of undescribed dinoflagellate species in environments like marine picoplankton (e.g. Moreira and López García, 2002; Worden, 2006) or as symbionts (‘zooxanthellae’) in many types of protists and invertebrates like corals (Coffroth and Santos, 2005).
The photosynthetic and mixotrophic (utilising both inorganic carbon and organic carbon sources) species are very important players in oceanic carbon cycles, and some cause harmful (toxic) algal blooms when cell densities reach exceedingly high levels (Taylor, 1987). Many photosynthetic dinoflagellates are also consumers of bacteria and other microeukaryotes (Stoecker, 1999).
Cell structure – Dinoflagellates have two flagella with independent beating pattern that confers a peculiar rotattory swimming whirling motion. Flagella are inserted apically (desmokont type) or they emerge from a region close to the midpoint ventral side of the cell (dinokont type) (Barsanti and Gaultieri, 2006). Around the cell they present a layer of flat vescicels surrounding the cells: these can be empty or filled with cellulose (armored dinoflagellates). They possess chlorophylls a, b, c1 and c2 and fucoxanthin, peridinin, dinoxanthin. Dinoflagellates have novel cytoskeletal and nuclear features (e.g. permanently condensed chromosomes) that make them very distinctive among eukaryotes (Fensome et al., 1999). Dinoflagellates morphological features include:
• A system of abutting membranous sacs, called “alveoli”, positioned beneath the plasma membrane (synapomorphy); the alveoli are filled with cellulosic material.
• Distinct micropores through the cell surface that function in pinocytosis (synapomorphy).
• Extrusive organelles.
• Closed mitosis.
• Tubular mitochondrial cristae. Complex organelles found in the group include structures reminiscent of a full‐fledged vertebrate
eye (but in a unicellular organism that lacks a brain), nematocysts comparable to those of cnidarians, and a bewildering array of plastid types in the photosynthetic forms. Dinoflagellates exist as plasmodia (i.e. multinucleate organisms), biflagellated cells, coccoid stages and even, in one small group, as cell arrays that approach multicellularity.
Genetically, dinoflagellates are also unique. The nucleus of a large majority of dinoflagellates (the so‐called dinokaryotes) is so different from other eukaryotic nuclei that it has been given its own name, the dinokaryon. Dinokarya lack nucleosomes, and DNA content is orders of magnitude larger than that of other eukaryotic cells, for example those of humans. These dinokarya divide via a unique form of mitosis. Recent research is starting to show just how unique dinoflagellate genetic systems are. For example, gene products of all dinoflagellate nuclei (not only dinokarya) are processed in a unique way: a spliced leader is trans‐spliced to all mRNA molecules. The genomes of plastids and mitochondria of the group are also unique: they are atomized (i.e. the genome is split into very small

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 26 of 258
fragments), and gene content is much, much lower than that of comparable organelles in other organisms.
Ecological significance ‐ Dinoflagellates are perhaps best known as causers of harmful algal blooms (webpages about this topic: ISSHA, WHOI, IOC). About 75‐80% of toxic phytoplankton species are dinoflagellates (Cembella, 2003), and they cause “red tides” that often kill fish and/or shellfish either directly, because of toxin production, or because of effects caused by large numbers of cells that clog animal gills, deplete oxygen, etc. (Smayda, 1997). Dinoflagellate toxins are among the most potent biotoxins known. They often accumulate in shellfish or fish, and when these are eaten by humans they cause diseases like paralytic shellfish poisoning (PSP), neurotoxic shellfish poisoning (NSP), diarrheic shellfish poisoning (DSP) and ciguatera (Lehane and Lewis, 2000). They also have been linked to major human health concerns, especially in estuarine environments (Pfiesteria). Some syndinians, notably Hematodinium, are parasites of economically‐significant crustacean species.
The main ecological significance of dinoflagellates lies elsewhere, though. They are second only to diatoms as marine primary producers. As phagotrophic organisms they are also important components of the microbial loop in the oceans and help to channel significant amounts of energy into planktonic food webs. As zooxanthellae, dinoflagellates have a pivotal role in the biology of reef‐building corals.
3.9 Other algal groups Euglenophyta (or euglenoids) are one of the best‐known groups of flagellates, commonly found in freshwater especially when it is rich in organic materials, with a few marine and endosymbiotic members. Many euglenids have chloroplasts and produce energy through photosynthesis, but others feed by phagocytosis or strictly by diffusion. Euglenoids are distinguished mainly by the presence of a pellicle, which is composed of proteinaceous strips underneath the cell membrane, supported by dorsal and ventral microtubules. This varies from rigid to flexible, and gives the cell its shape, often giving it distinctive striations. In many euglenoids the strips can slide past one another, causing an inching motion called metaboly. Otherwise they move using the flagella.
Cryptophyta are aquatic unicellular photosynthetic eukaryotes that inhabit both marine and freshwater environments. Plastids are very diverse in pigmentation. In addition to being important from an ecological perspective, the cryptophytes are of pivotal significance in terms of our understanding of endosymbiosis and the evolution of plastids. This is because cryptophyta acquired photosynthesis by the process of secondary (i.e., eukaryote‐eukaryote) endosymbiosis, having engulfed and assimilated a red algal endosymbiont at some time during their evolutionary past (Archibald and Keeling, 2002; Bhattacharya et al., 2003; Gould et al., 2008). As a result, cryptophyta are extremely complex from a genetic and cell perspective, having a four‐membrane‐bound plastid and four distinct DNA‐containing cellular compartments, the plastid, mitochondrion, host nucleus and endosymbiont nucleus, the latter being referred to as the nucleomorph. The cryptophyte nucleomorph genome is highly reduced in structure and coding capacity, and the focus of ongoing research aims at understanding the pattern and process of secondary endosymbiosis (Gilson and McFadden, 2002; Archibald, 2007).
Glaucophyta, also known as glaucocystophytes or glaucocystids, are a small group of freshwater microscopic algae (Keeling, 2004). The glaucophytes are of interest to biologists studying the development of chloroplasts, because some studies suggest that they may be similar to the original

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 27 of 258
alga type which led to green plants and red algae (Keeling, 2004, Kim and Graham, 2008). The chloroplasts of glaucophytes are known as cyanelles. Unlike other eukaryotic plastids they have a peptidoglycan layer which is believed to be a relic of the endosymbiotic origin of plastids from cyanobacteria (Keeling, 2004). Glaucophytes contain the photosynthetic pigment chlorophyll a (Keeling, 2004). Along with red algae and cyanobacteria they harvest light via phycobilisomes, structures consisting largely of phycobiliproteins. Glaucophytes have mitochondria with flat cristae, and undergo open mitosis without centrioles. Motile forms have two unequal flagella, which may have fine hairs and are anchored by a multilayered system of microtubules, both of which are similar to forms found in some green algae. There are only 13 species of glaucophytes known, none of which are particularly common in nature.
Chlorarachniophyta are green amoebas living in sea water sample. The green color is caused by chloroplasts, which are permanently housed in the amoeba cell. Although the chloroplasts contain chlorophylls a and b, chlorarachniophyta are phylogenetically distinct from other chlorophyll b‐containing eukaryotes (Chlorophyta and Euglenophyta), and constitute an independent phylum, which was established only two decades ago (Hibberd and Norris, 1984). The group consists of four genera and five species, and there are also several species waiting to be named (Ishida et al., 1999; 2000).

AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 28 of 258
4 Biotechnology of algae
4.1 Introduction
Edible blue‐green microalgae, including the cyanobacteria Nostoc and Arthrospira (formerly Spirulina), have been part of the human diet since many centuries. However, the modern era of microalgal biotechnology begins in the early 1950s, when the first algae mass cultures in photobioreactors (PBR) were attempted. Today, between 8,000 and 10,000 tons of algae biomass are produced annually (Pulz and Gross, 2004; Becker, 2007), mainly for use as human food supplements and animal feed. Table 3 shows the present status of commercial production and the main application areas of microalgae (Tredici et al. , 2009). Table 4 shows the comparison of several algae production systems.
Microalgae biomass is either harvested from natural habitats or obtained through more or less controlled cultivation processes carried out mainly in large open ponds or lagoons. Commercial production in photobioreactors (PBR) is limited to a few hundreds of tons (see below).
Table 3 ‐ Commercial production of microalgae (from Tredici al., 2009).
The commercial production of microalgae is mainly limited to species belonging to the genera Arthrospira, Chlorella and Dunaliella that, due to their high growth rate or of a selective growth medium that limits contamination, can be grown in large open ponds. These systems are easier to operate, less expensive, and more durable than large closed reactors (Tredici, 2004). However, the majority of microalgae does not require a specific growth environment or a selective medium and can not be cultivated for prolonged periods in outdoor open systems because of contamination. Photobioreactors provide a closed environment for the culture (limiting direct fall‐out and invasion by unwanted species) and a better control of culture parameters (pH, temperature, pO2, etc.) which ensure the dominance of the desired species (Tredici, 2004).
To exploit the large biodiversity of microalgae and cyanobacteria, low‐cost and scalable PBR are a necessity, either to be used alone or in combination with open ponds, in this latter case to produce the inocula required to maintain the selected species in the ponds.
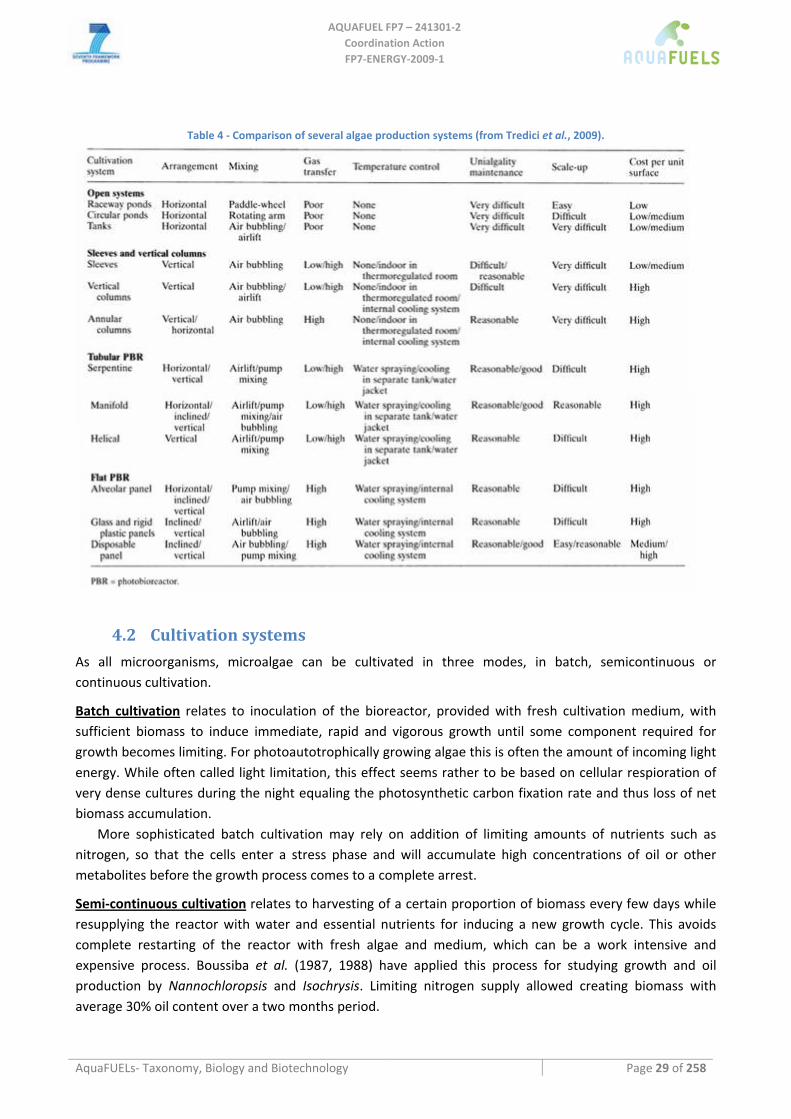
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 29 of 258
Table 4 ‐ Comparison of several algae production systems (from Tredici et al., 2009).
4.2 Cultivation systems As all microorganisms, microalgae can be cultivated in three modes, in batch, semicontinuous or continuous cultivation.
Batch cultivation relates to inoculation of the bioreactor, provided with fresh cultivation medium, with sufficient biomass to induce immediate, rapid and vigorous growth until some component required for growth becomes limiting. For photoautotrophically growing algae this is often the amount of incoming light energy. While often called light limitation, this effect seems rather to be based on cellular respioration of very dense cultures during the night equaling the photosynthetic carbon fixation rate and thus loss of net biomass accumulation.
More sophisticated batch cultivation may rely on addition of limiting amounts of nutrients such as nitrogen, so that the cells enter a stress phase and will accumulate high concentrations of oil or other metabolites before the growth process comes to a complete arrest.
Semi‐continuous cultivation relates to harvesting of a certain proportion of biomass every few days while resupplying the reactor with water and essential nutrients for inducing a new growth cycle. This avoids complete restarting of the reactor with fresh algae and medium, which can be a work intensive and expensive process. Boussiba et al. (1987, 1988) have applied this process for studying growth and oil production by Nannochloropsis and Isochrysis. Limiting nitrogen supply allowed creating biomass with average 30% oil content over a two months period.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 30 of 258
Similarly the bioreactor facility at Qetura (Israel) relies on semi continuous cultivation for production of green Haematococcus biomass, while the second growth stage, the stress phase inducing astaxanthin biosynthesis, is handled as batch culture.
Continuous cultivation: continuous cultivation is widely used for cultivation of heterotrophic organisms whereby all reactor parameters as well as cell density are tightly supervised and controlled and growing biomass is continuously harvested to maintain optimal growth conditions for extended periods of time. While this mode can yield the best possible biomass yields, production of high concentrations of oil or carotenoids requiring stress conditions is not possible.
4.2.1 Open ponds
Open shallow ponds (Fig. 2) mixed and aerated by any given method have been the method of choice so far for producing biotechnologically relevant amounts of algal biomass. CO2 is inserted as required. Cultures are mixed in circular ponds using circulation ponds, or in the oval so called raceway ponds using paddle wheels to move the medium around. Major producers of Arthrospira (Spirulina) and Dunaliella in California, Hawaii, Australia and Israel have been relying on this method for over 30 years, taking advantage of the fact that the growth media of those two algal species (basic or highly saline respectively) are unsuitable for competing algal species. Currently, commercial cultivation of microalgae is limited to a few microalgal species that are cultivatedin open ponds. A raceway pond is an economic open system for microalgae cultivation. It consists of acircuit of parallel tunnels placed at ground level, in which the microalgae suspension flows gently and is moved by a paddle wheel. The system is built in concrete or ditch dug into the ground and may be lined with white plastic sheets. Evaporation is significant as well as temperature fluctuations, and rainfalls can dilute theavailable nutrients.
High level of contamination by predators and other fast growing heterotrophs have restricted thecommercial production of algae in open culture systems to only those organisms that can grow under extreme conditions. Examples are Dunaliella salina (high salinity) and Arthrospira platensis (Spirulina) (high alkalinity). However, BGU is maintaining an open Nannochloropsis culture outdoors without any significant contamination problems, and Seambiotic has similarly been able to successfully maintain Nannochloropsis cultures even in close vicinity of the ocean.
The Algal Biofuels Program of the DOE (Sheehan et al., 1998) has also concluded that open raceway ponds would be the method of choice for algal biomass production, whereby spontaneously developing local algal populations might be the biomass of choice for simplifying the production process.
Figure 2 ‐ Examples of microalgae open cultivation systems: a) Board mixed ponds, UNIFI, Italy (1980); b) Pilot raceway pond, Necton, Portugal (1996); c) Raceway ponds, Hainan DIC, China (1996); d) Cyanotech, Hawaii, USA (2008); e) Flue gas fixation

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 31 of 258
project of Seambiotic Ltd, Israel; f) Raceway pond,s Earthrise farms, California, USA (1982); g) Raceways of 2000 m2, designed by
BGU and in operation at Eilat; Israel; h) Ponds in operation at Parry Agro Industries, India (total ponds area 53 ha) (2006).
4.2.2 Photobioreactors Photobioreactors (PBRs) (some examples are reported in Fig. 3) can be defined as culture systems for phototrophs in which a great proportion of the light does non impinge directly on the culture surface, but has to pass through the transparent reactor’s walls to reach the cultivated cells. Consequently, PBRs do not allow, or strong limit, direct exchange of gases and contaminants with the environment (Tredici, 2004).
Reactors used for the cultivation of microalgae are either naturally or artificially illuminated, and may be open or closed systems. Open systems with naturally illuminated large surface areas include essentially raceways ponds. PBRs are closed cultivation systems to grow microalgae under photo‐autotrophic conditions. Several types of photobioreactors (PBRs) have been designed and experimented with since the late 1940s, when algal cultures were first considered the ideal solar technology to produce in a cost‐effective manner biomass and protein on a large scale and save the hungry part of humanity (Tredici et al., 2010). They can be classified on the basis of both design and mode of operation. In design terms, the main categories of reactors are (1) flat or tubular; (2) horizontal, inclined, vertical or spiral; and (3) manifold or serpentine. The most used designs are flat‐plates (vertical or inclined), tubular reactors (serpentine or manifold with horizontal, vertical or inclined arrangement), air‐bubbled plastic bags and annular columns. Some of them, especially in small‐scale, are artificially illuminated with fluorescent or other types of lamps. The two major categories of PBRs recently proposed for biofuels at relatively large scale are flat and tubular reactors (Tredici et al., 2010).
As PBR are more expensive to install and operate, intensive study is being performed in an effort to reduce their cost and thus facilitate their use especially for low value products like algal oils. All parameters (e.g. nutrients, light regime, gas exchange) are maintained to realize optimal culture conditions. Costs of PBRs are much higher, but contamination is much lower compared to open systems (Tredici, 2004). PBR design and engineering is still a very active field of research today, since closed culture systems are necessary to grow photosynthetic microbes and exploit them as a source of aquaculture feeds, food additives, specialty chemicals, cosmetics and are preferred by industry as research tools for biofuel production.
In air‐bubbled plastic bags and annular columns the cost of operation may be very high due to the need of mixing to provide the necessary mass transfer. This operation requires more than 100 W.m‐3 (approximately 2000 MJ ha‐1 day‐1) or up to 50% of the energy stored in the biomass [Bassi et al., Unpublished Data].
Within all the photobioreactor types described, tubular PBRs are the most popular design (Chisti, 2007; Tredici et al., 2010). Their operation costs are normally significantly higher than those of ponds. Mixing in tubular reactors to achieve sufficient fluid velocities and Reynold’s numbers may require even higher energy inputs. Besides, there is the need for cooling, which is generally provided by water (even seawater) spraying or by insertion of a cooling serpentine in the culture.
Flat Panel Reactors consist of a rectangular transparent box with a depth of only 1 to 5 cm. Accordingly, this reactor type allows growth of high cell densities. Vertical reactors intercept sun rays at large angles and dilute light compared with horizontal ponds. Besides, their rear surface receives mostly diffuse and reflected radiation of low intensity. For this reason, vertical PBRs are more efficient than horizontal ponds in terms of solar‐energy utilization (Tredici et al., 2010).
One of the main problems associated with PBRs culture is that the formation of biofilms of bacteria, or the cultivated algae themselves, at the transparent walls of the tubes considerably reduce light

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 32 of 258
penetration. Biofilms are minimized by maintaining highly turbulent flow, using small particles and regularly cleaning with hydrogen peroxide.
PBRs have many advantages over open ponds, some of them were referred in the most recent article of Tredici (Tredici et al., 2010), and are described below:
• they are closed, making it easier to maintain an unialgal culture and reducing evaporation;
• due to a higher surface‐to‐volume ratio, PBRs may attain higher volumetric productivities thanponds and adopt higher cell concentrations, which reduce the costs for medium preparationand handling and that for harvesting;
• PBRs provide a more accurate control of culture parameters;
• the ultimate and most important advantage of PBRs, however, is that they permit to cultivatealgal species that cannot be grown in open ponds.
Photobioreactors pay heavily for their advantages. In general, PBRs are much more expensive to build than ponds, but simple low‐cost systems can also be designed. Most constrains require further investigation, specially low‐cost materials for construction and lower energy requirements, but also overheating, bio‐fouling, oxygen accumulation, difficulty in scaling up and cell damage by shear stress.
Figure 3 ‐ Examples of microalgae close cultivation systems: a‐b) Green Wall Panels and tubular photobioreactors from UNIFI ‐ Università degli Studi di Firenze and F&M ‐ Fotosintetica & Microbiologica s.r.l., Italy; c) Flat Panel
photobioreactor from Necton, S.A., Portugal; d‐e) Tubular photobioreactor with flue gas fluxuation from AlgaFuel, S.A., Portugal; f‐g) Tubular photobioreactors from Alga Technologies, Israel; h) Flat Panel at Ben Gurion University –
BGU‐, Israel.
Main photobioreactors designs
Polyethylene bags and vertical columns
Polyethylene bags (sleeves) suspended from a framework or supported within a mesh frame and mixed by air bubbling are the most common cultivation devices used in hatcheries for the production of algal biomass (Fig. 4). From 50 to 500 L in volume, such reactors are mostly used indoors with artificial illumination. They are currently used by different companies (e.g. Ketura Kibbutz, Israel; GreenSea, France; NOVAgreen GmbH, Germany) for the cultivation of selected species for the cosmetic, food or pharmaceutical markets. Although sleeve reactors are inexpensive, their low surface‐to‐volume ratio (S/V), bio‐fouling and the need of a very large number of units for large‐scale production limit their applications.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 33 of 258
The company Proviron developed inflatable bags with multiple vertical panes (ProviAPT) (http://www.proviron.com/algae/GB/algae_solution_working.php) (Fig. 4). The big, translucent, plastic bag is filled with water containing the multiple vertical panes. The algae grow in these panes while CO2 rich air is bubbled through. Due to the water the reactor is self supporting and the water buffers the temperature efficiently between day and night. It is a continous sytem with continous harvesting.
Vertical columns are made of rigid transparent cylinders (typically 2‐2.5 m in height and 30‐50 cm in diameter), with mixing achieved by air bubbling or by an airlift. They are extensively used in hatcheries and suffer from the same limitations of sleeves. After the first design by Helm, vertical cylinders internally illuminated have been developed at the University of Florence. The reactor, named annular column (Fig. 4), is made of two concentric transparent cylinders of different diameter sealed at the base to form a culture chamber with much higher S/V compared to completely filled columns. Lamps or fluorescent tubes can be placed inside the inner cylinder. Annular columns of 30‐230 L in volume are currently commercialized by Fotosintetica & Microbiologica S.r.l. (Italy), a spin‐off company of the University of Florence. The small size and relatively high cost of this reactor limit its use for large scale production.
Figure 4 ‐ Polyethylene bags and Annular Columns. Top left: Polyethylene bags at Necton, SA – Algarve, Portugal; Top right: Annular Columns at Department of Agricultural Biotechnology of the University of Florence, Italy;
Bottom: Inflatable bags with multiple vertical panes (Proviron, Belgium).
Tubular PBR
Most of the PBR adopted in commercial plants are of the tubular type. This category can be subdivided into three main sub‐groups: i) serpentine, ii) manifold and iii) helical PBR (Fig. 5, 6).
Serpentine PBR, firstly designed by Tamiya in the 1950s, consist of straight tubes connected by U‐bends to form a flat loop (the photostage) that may be arranged either vertically or horizontally.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 34 of 258
Figure 5 ‐ Horizontal tubular photobioreactors. Mera Pharmaceuticals, Inc – USA (left); Serpentine photobioreactor. AlgaeLink NV ‐ The Netherlands (right).
Manifold PBR are made of parallel tubes connected at the ends by two manifolds. The main advantages of these systems over serpentine reactors are the reduction of head losses and lower oxygen concentrations, two factors that facilitate scale‐up to industrial size. A horizontal manifold reactor is used by Algatechnologies Ltd (Israel) for the commercial production of H. pluvialis. The Near‐Horizontal Tubular Reactor (NHTR), a manifold reactor with tubes inclined from 5 to 20° to the horizontal so that mixing could be achieved by air bubbling was developed by Tredici and co‐workers (Tredici et al., 1999). A 20‐m long NHTR was experimented with at the University of Hawaii (USA). Two small NHTR units are currently used by ENI SpA (Italy) to develop CO2 abatement strategies with microalgae (Pedroni et al., 2004). A similar design, the triangular airlift reactor, consisting of a series of riser tubes, gas separators and downcomer tubes arranged in a triangular configuration was used by GreenFuel Technologies Co (USA) for the abatement of greenhouse gases. Rather common, both at pilot scale and in commercial plants, are manifold photobioreactors arranged fence‐like.
Figure 6 ‐ First on the left: Triangular Photobioreactor. Green Fuel Technologies Co ‐ USA. Middle and right figures: BioFence systems. Varicon Aqua Solutions Ltd ‐ UK
The BioFence, developed by Applied Photosynthetics Ltd (UK) in the late 1990s, consists of an array of transparent tubes racked together in a fence‐like structure in which the culture suspension is circulated by a centrifugal pump or by an airlift. BioFence systems are currently commercialized by Varicon Aqua Solutions Ltd (UK). Biofence photobioreactors from 10 to 35 000 L are also distributed by B Braun Biotech International GmbH (Germany). Industrial scale plants based on this design are operated by Bioprodukte Prof Steinberg GmbH (Germany) for the production of Chlorella and by Algatechnologies Ltd (Israel) for the growth of H. pluvialis.
Helical photobioreactors (biocoils), consisting of small‐diameter flexible tubes wound around an upright structure have also been much experimented and commercialized in the past.
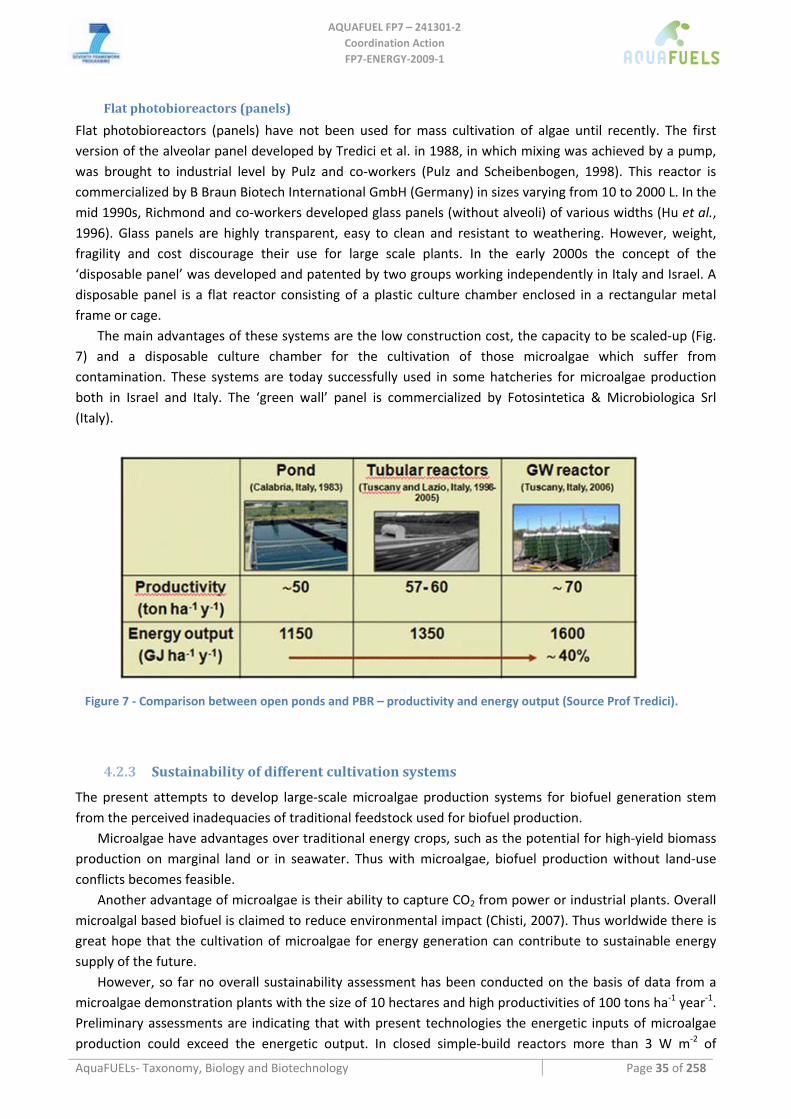
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 35 of 258
Flat photobioreactors (panels)
Flat photobioreactors (panels) have not been used for mass cultivation of algae until recently. The first version of the alveolar panel developed by Tredici et al. in 1988, in which mixing was achieved by a pump, was brought to industrial level by Pulz and co‐workers (Pulz and Scheibenbogen, 1998). This reactor is commercialized by B Braun Biotech International GmbH (Germany) in sizes varying from 10 to 2000 L. In the mid 1990s, Richmond and co‐workers developed glass panels (without alveoli) of various widths (Hu et al., 1996). Glass panels are highly transparent, easy to clean and resistant to weathering. However, weight, fragility and cost discourage their use for large scale plants. In the early 2000s the concept of the ‘disposable panel’ was developed and patented by two groups working independently in Italy and Israel. A disposable panel is a flat reactor consisting of a plastic culture chamber enclosed in a rectangular metal frame or cage.
The main advantages of these systems are the low construction cost, the capacity to be scaled‐up (Fig. 7) and a disposable culture chamber for the cultivation of those microalgae which suffer from contamination. These systems are today successfully used in some hatcheries for microalgae production both in Israel and Italy. The ‘green wall’ panel is commercialized by Fotosintetica & Microbiologica Srl (Italy).
Figure 7 ‐ Comparison between open ponds and PBR – productivity and energy output (Source Prof Tredici).
4.2.3 Sustainability of different cultivation systems
The present attempts to develop large‐scale microalgae production systems for biofuel generation stem from the perceived inadequacies of traditional feedstock used for biofuel production.
Microalgae have advantages over traditional energy crops, such as the potential for high‐yield biomass production on marginal land or in seawater. Thus with microalgae, biofuel production without land‐use conflicts becomes feasible.
Another advantage of microalgae is their ability to capture CO2 from power or industrial plants. Overall microalgal based biofuel is claimed to reduce environmental impact (Chisti, 2007). Thus worldwide there is great hope that the cultivation of microalgae for energy generation can contribute to sustainable energy supply of the future.
However, so far no overall sustainability assessment has been conducted on the basis of data from a microalgae demonstration plants with the size of 10 hectares and high productivities of 100 tons ha‐1 year‐1. Preliminary assessments are indicating that with present technologies the energetic inputs of microalgae production could exceed the energetic output. In closed simple‐build reactors more than 3 W m‐2 of

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 36 of 258
electrical energy are used for operating the cultivation. This comes close to the amount actually harvested at the end. However, it has to be taken into account, that in nowadays commercial algae cultivation, a positive energy‐balance is not targeted due to the high‐prices achieved for the products.
LCA and sensitive analysis can help to identify at which points of the production chain technological progress is needed and how the demonstration plant could be designed in a more sustainable way. A long dispute concerning the optimal modes of cultivation for algal biofuels production may have been laid to rest during the last two years thanks to a number of analyses determining the environmental life cycle impact of different cultivation modes.
Several LCA analyses agree in one major conclusion, that even under optimistic yield assumptions tubular photobioreactors require far more energy to run than the algal biomass being produced. More simple panel reactors fare somewhat better, but still are unable to return the amount of energy required to construct and run them. On the other hand open raceway ponds assuming yields of 20 g m‐2 or more can return significantly more energy in the form of biomass than has to be invested for building , running and maintaining the ponds (Joquera et al., 2010; Stephenson et al., 2010).
This difference becomes even more pronounced when taking into account that above analyses have not incorporated energy expenses for cooling or heating which can be tremendous in photobioreactors, but are not required in open ponds.
4.3 Harvesting methods
Algae harvesting and dewatering processes may account for 20‐30% of the total production cost for algal biomass (Uduman et al., 2010; Pienkos et al., 2009). In order to remove large quantities of water and process large algal biomass volumes, a harvesting method involving several steps is required (Mata et al., 2010). The concentration of algal biomass is typically 0.5‐1.0 g L‐1 in open ponds and can reach 5‐10 g L‐1 in closed systems. At 1g L‐1 algal biomass, 1000 kg of water must be processed to obtain 1kg of dry biomass.
Most common harvesting methods include sedimentation, centrifugation, filtration, ultra‐filtration, sometimes with an additional flocculation step or with a combination of flocculation‐flotation. The choice of which harvesting technique to use depends on the species of microalgae and the final product desired. Microalgal properties that simplify harvesting are large cell size, high specific gravity compared to the medium and reliable autoflocculation. Moreover the optimum harvesting method should have minimum energy requirements and be as economical as possible. The main harvesting techniques were recently described by Uduman et al. (2010), and are summarized in Table 5.
Centrifugation is seen as the most efficient recovery technique, yet the energy and capital costs associated are unappealing (Uduman et al., 2010). Filter presses operating under pressure or vacuum can be used to recover large quantities of biomass, but for some applications filtration can be relatively slow which may be unsatisfactory. Alternatively, membrane microfiltration and ultra‐filtration are other possible alternatives to conventional filtration for recovering algal biomass, which are more suitable for fragile cells and small‐scale production processes. Flocculation is also a very efficient technique that is currently applied in industry. Microstrainers are an attractive harvesting method because of their mechanical simplicity and availability in large unit sizes (Mata et al., 2010). Nevertheless, a significant engineering research effort aimed at developing more cost‐effective algal harvesting techniques will be required.
BGU and Algatech (Israel) have designed successful harvesting and processing procedures for Haematococcus biomass that are based on sedimentation in large funnels and works without energy requirements at over 90% efficiency, creating a concentrated cell slurry that can be further dewatered and dried. Flocculation of Nannochloropsis and Isochrysis has also been successfully applied (Boussiba et al., 1987, 1988). Evodos (The Netherlands), leader of WP7, has established a 100% subsidiary, called ‘Evodos
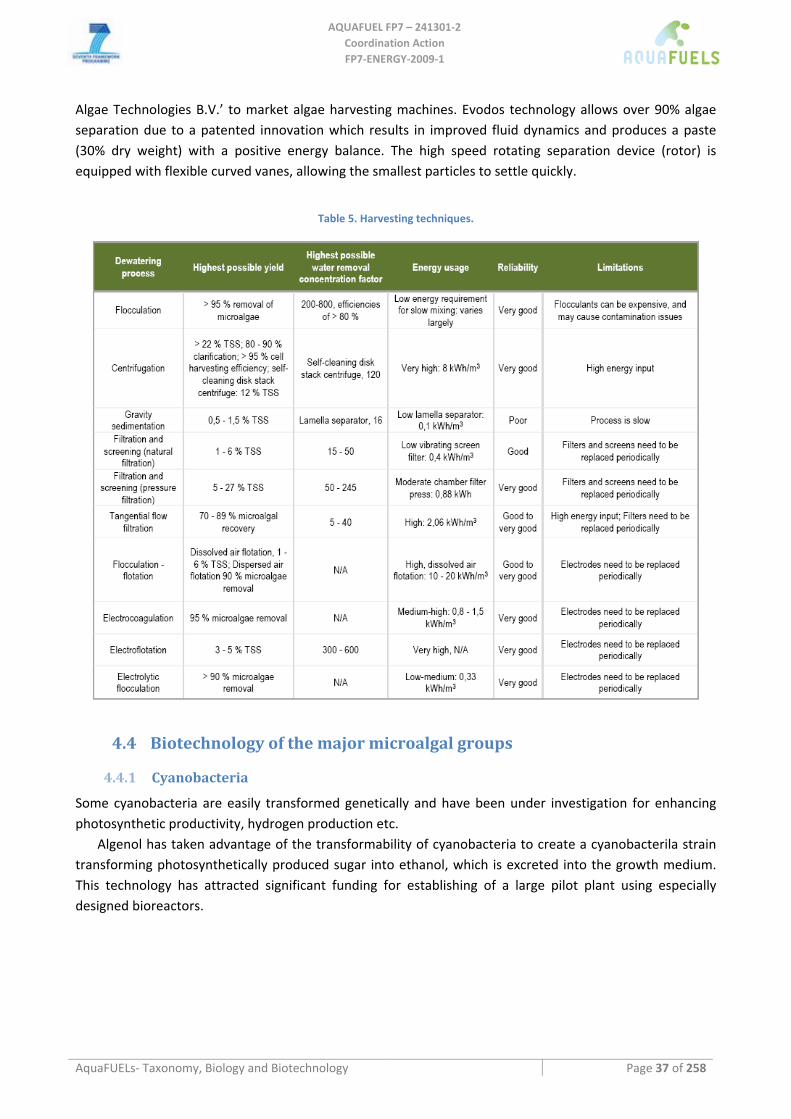
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 37 of 258
Algae Technologies B.V.’ to market algae harvesting machines. Evodos technology allows over 90% algae separation due to a patented innovation which results in improved fluid dynamics and produces a paste (30% dry weight) with a positive energy balance. The high speed rotating separation device (rotor) is equipped with flexible curved vanes, allowing the smallest particles to settle quickly.
Table 5. Harvesting techniques.
4.4 Biotechnology of the major microalgal groups
4.4.1 Cyanobacteria
Some cyanobacteria are easily transformed genetically and have been under investigation for enhancing photosynthetic productivity, hydrogen production etc.
Algenol has taken advantage of the transformability of cyanobacteria to create a cyanobacterila strain transforming photosynthetically produced sugar into ethanol, which is excreted into the growth medium. This technology has attracted significant funding for establishing of a large pilot plant using especially designed bioreactors.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 38 of 258
Figure 8‐ http://www.algenolbiofuels.com/overview.htm.
An interesting application of the transformability techniques was cloning and expression of BTi toxins
into Anaebaena for creating a biological mosquito control agent, though release of this organism into nature has not been licensed so far (Xiaoqiang et al., 1997).
Cyanobacteria can be thermophilic and occur in all watery environments including toxic hot‐springs. This ability has been exploited to establish a revolutionary exhaust gas scrubbing methodology with thermophilic cyanobacteria isolated from Yellowstone hotsprings growing directly on panels lit by sun light concentrated and delivered via light guides, in chambers fed by exhaust gas (http://www.greencarcongress.com/2005/12/greenshift_lice.html).
Recent Researches have also hinted at cyanobacteria possible application to the generation of clean and green energy via converting sunlight directly into electricity.
A number of important advances have occurred in cyanobacterial biotechnology in the recent years. World wide attention is drawn towards cyanobacteria for their possible use in mariculture, as food, feed, fuel, fertilizers, colourants, and for the production of various secondary metabolites including vitamins, toxins, enzymes, pharmaceuticals and pharmacological probes, and for pollution abatement (Tredici et al., 2009). Only a few cyanobacterial strains have been well‐characterized or exploited commercially. Some cyanobacteria are sold as food, notably Aphanizomenon flos‐aquae and Arthrospira platensis (Spirulina). Spirulina is being grown commercially (and marketed as dietary supplements) mostly in open raceway ponds in Asia and USA, while short pre‐commercial cultivation was also performed by Ben Gurion University for Koor industries in Israel (Vonshak, personal communication).
4.4.2 Chlorophyta (Green Algae)
A significant number of green algal species are being exploited for production of pigments (Dunaliella, Haematococcus) or neutraceuticals (Chlorella), and biological remediation (Chlorella, Scenedesmus), whereby the vast metabolic capacity of green algae holds a great promise for identification of further commercially viable production processes. Furthermore, Chlamydomonas reinhardtii is the most advanced tool for studying molecular, genetic and biochemical aspects of microalgae.
Chlorella, Dunaliella and Haematococcus provide the vast majority of algal biomass produced worldwide, exclusively as nutraceutical or health products. Chlorella is being cultivated in open ponds in Asia (e. g. http://www.taiwanchlorella.com/) or in Europe's largest closed photobioreactor facility in Klotze, Germany (http://www.algomed.de/index.php). The protein, vitamin and mineral rich dry biomass is

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 39 of 258
marketed as food additive. The capacity of Dunaliella to survive in Dead Sea water and to accumulate high concentrations of β‐carotene and glycerol has early on caught the attention of scientists, and a range of production facilities in open lagoons, e. g. in Australia (http://www.nutrakol.com/pdf/Dunaliella.pdf), Asia or in raceway ponds in Eilat, Israel (http://www.wondercare.co.in/nature.htm) have been operating successfully. Haematococcus pluvialis is producing large amounts of the highly valued pigment astaxanthin that is a potent antioxidant with significant health benefits reported. Astaxanthin accumulation requires cultivation under nutrient stress, and major producers today are found in Asia, Australia and Hawaii (Fig. 2 d), where the algae are cultivated in raceway ponds (http://www.cyanotech.com/), or Israel, where astaxanthin is produced in a large photobioreactor facility in Qetura (Fig. 3 f/g; http://www.algatech.com/).
Other production processes for various pigments from species such as Scenedesmus (lutein) Chlorococcum (astaxanthin) or Chlorella zofingensis (both) (Del Campo et al., 2004) have been investigated but not achieved commercial deployment.
In all instances described, alternative products either from land plants or from chemical synthesis are significantly cheaper to produce than the algal product, so that to date the only market niche for such algal products is the health market.
A significant number of species or genera are being considered as biofuel feedstock, due to the fact that numerous green algae species are very fast growing and are able to accumulate high concentrations of oil useful for biodiesel production. About half the strains tested by Hu et al. (2008) are green algae, as are 11 out of 30 tested by Rodolfi et al. (2009) Their high lipid productivity, accessability to genetic engineering and capability to grow rapidly in the dark fermenting organic substrates has made Chlorella the primary platform for custom made biofuels production via heterotrophic fermentation by biofuels company Solazyme.
4.4.3 Rhodophyta (Red Algae)
Several Red Algae species have significant biotechnology potential. Some of them are economically important as source of agars and carrageenans. For this reason, extensive farming and natural harvest of red algae occurs in numerous areas of the world (see macroalgal biotechnology).
The unicellular red algae Porphyridium is producing significant amounts of LC‐PUFA and medicinally active polymers. Up to 44% of its total fatty acids consist of the high value long chain PUFA EPA (Cohen and Cohen, 1991). Besides red microalgae in general, Porphyridium cruentum produces highly sulfonated extracellular polysaccharides with strong and specific activity against animal viruses such as Herpes (Huheihel et al., 2002).
4.4.4 Heterokontophyta
This group include a variety of algal divisions of highest significance to biotechnology, with significance to aquaculture, food and biofuels industries as well as the nutraceutical market.
Among the microalgae the most important group from the biotechnological point of view is represented by the eustigamotphytes. Nannochloropsis is considered a promising alga for industrial applications because of its ability to accumulate high levels of polyunsaturated fatty acids (eicosapentaenoic acid). It is mainly used as feed for fish larvae and rotifers (see detailed form at the end of the document). Nannochloropsis is also used as an ingredient in cosmetic products (Tredici et al., 2009). Nannochloropsis has also been proposed as feedstocks for biodiesel production because of its ability to accumulate up to 60% lipid under nitrogen starvation (Rodolfi et al., 2009). Monodus is a freshwater eustigamtophyte which is also rich in eicosapentaenoic acid (EPA). It has been cultivated to study its EPA production under laboratory conditions and outdoors in different types of photobiorectors under different

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 40 of 258
conditions. In Spain productivities of 0.16 and 0.54 g L‐1 d‐1 were obtained in a 57‐L‐bubble column and in a 75‐L‐ helical photobioreactor, respectively, with a maximum EPA productivity of 9 mg L‐1 d‐1 (Lu et al., 2002). In Israel, Hu et al. (1997) in a flat inclined modular photobioreactor made of four 14‐L units, obtained a biomass productivity for Monodus of about 1.4 g L‐1 d‐1 and an EPA productivity of 59 mg L‐1 d‐1 at the optimal cell density of 4 g L‐1. For its EPA content Monodus subterraneus is used for the preparation of nutraceutical products together with the DHA‐rich primnesiophyte Isochrysis galbana by Nikken Sohonsha Co. (Japan). Recently, a freshwater xantophyte alga, Trachydiscus minutus, has been proposed as a source of EPA due to its large content of this fatty acid (up to >35% of total fatty acids) (Řezanka et al., 2010).
4.4.5 Labyrinthulea (phylum Heterokonta)
Thraustochytrids are a commercial source of polyunsaturated fatty acids, in particular doicosahexaenoic acid (DHA). DHA‐rich oils are produced through fermentation from the marine thraustochytrids Schizochytrium and Ulkenia by Martek Co. (USA) and Nutrinova (Germany), respectively, for nutritional supplements and animal feeds (Lewis et al., 1999; Tredici et al., 2009). During commercial production there are two phases: first, biomass production and then lipid induction. Glucose is generally the preferred substrate to grow these organimsms, though there is a lot of variability among strains concerning the best carbon and nitrogen source. Dried Schizochytrium has GRAS status for use as feed to broiler chickens and laying hen feed in order to enhance DHA in the meat and eggs (Raghukumar, 2008). Schizochytrium cells are produced by Aquafauna, Inc (USA) to be included in feed for aquaculture, in particular to enrich in DHA rotifers and Artemia. Most thraustochytrids produce one or more of other PUFAs, namely arachidonic acid, docosapentaenoic acid, eicosapentaenoic acid and docosatetraenoic acid and, although they can be considered as a source of these molecules, there is not an actual exploitment. The DHA biosynthetic pathway in thraustochytrids has presently attracted attention because of efforts by many researchers to produce recombinant plants or yeasts capable of producing PUFAs. Thraustochytrids are also considered a potential source of carotenoids such as β‐carotene and the xanthophylls astaxanthin and canthaxanthin (Raghukumar, 2008).
4.4.6 Bacillariophyta (Diatoms)
Diatoms are used for various biotechnological applications, mainly at the laboratory scale, and in aquaculture at a higher scale for commercial outlets. Examples of applications are: (1) silicon production originating from frustules, for technological applications in nanotechnology, pollution remediation or as food in aquaculture thanks to the lipid‐ and amino‐acid‐rich algal content, (2) intracellular metabolites that accumulated in cells, e.g. lipids, particularly eicosapentaenoic acid (EPA) for pharmaceutical applications, or amino acids for cosmetic applications, and (3) extracellular metabolites released into the medium, e.g. various pigments (for chicken and fish feeds) and antibiotics (Lebeau and Robert, 2003).
In diatoms, the major fatty acids are C16:0 and C16:1 (Hu et al., 2008). The most nutritionally relevant biomolecules produced by diatoms are very‐long‐chain polyunsaturated fatty acids (vlc‐PUFAs) like arachidonic acid (AA) (C20:4ω6), eicosapentaenoic acid (EPA) (C20:5ω3), docosahexaenoic acid (DHA) (C22:6ω3) and other omega‐3 fatty acids (Bozarth et al., 2009).
The average total lipid content of an oleaginous diatom is 22.7% DCW when maintained under normal growth conditions, whereas a total lipid content of 44.6% DCW is achievable under stress conditions (Hu et al., 2008). A general trend towards accumulation of lipids, particularly TAG, in response to nitrogen deficiency has been observed in various algal taxa including diatoms. However, in diatoms, silicon is an equally important nutrient that affects cellular lipid metabolism (Roessler, 1988). Phosphorus limitation

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 41 of 258
also results in increased lipid content, mainly TAG (Reitan et al., 1994). Typically, high light intensity decreases total polar lipid content with a concomitant increase in the amount of neutral storage lipids. High light also alters fatty acid synthesis to produce more of the saturated and mono‐unsaturated fatty acids that mainly make up neutral lipids. In general, the total lipid content of cells increase with age. An increase in TAGs is often observed during stationary phase (Hu et al., 2008).
4.4.7 Haptophyta
Economically important haptophytes include Pavlova lutheri and Isochrysis sp., which are widely used in the aquaculture industries due to their high docosahexaenoic acid (DHA) content. They are commercially produced by several companies to be sold as feed for bivalve shellfish larvae and post‐larvae, shrimp larvae and rotifer enrichemnt as concentrates (e.g. 64 to 90 g L‐1), alone or mixed with other algal species (Tredici et al., 2009). Due to the high DHA content, Isochrysis galbana is used, mixed with Monodus subterraneus, for the production of nutraceuticals by Nikken Sohonsha Co. (Japan). Recently, exctracts of Isochrysis sp. have been patented for application in cosmetics, as ingredients for skin and hair health products (Herrmann et al., 2009).
The suitability of Isochrysis for producing oil for biodiesel production has been confirmed in pilot sized raceway ponds in Israel (Boussiba et al., 1988).
4.4.8 Dinophyta (Dinoflagellates)
A few photoautotroph Dinoflagellate species have been evaluated for lipid productivity. The most prominent applications for Dinoflagellates involves heterotrophic species that are grown in fermentors for production of PUFA‐rich oil or biofuels. Commercial cultivation of the heterotrophic dinoflagellate Cryptechodinium cohnii to obtain a docosahexaenoic acid (DHA)‐rich oil to be used as an ingredient in nutraceutical products is carried out in fermentors by Martek Co. in the US.
Dinoflagellates are among the major toxin producers known. Besides the huge negative impact of these toxins on the ecosystems when they are produced in large amounts from algae blooms, an application can be foreseen. Dinoflagellate, as well as cyanobacteria, toxins are used in medical studies in attempts to understand their modes of action and assess their therapeutic potential or that of their analogs. Saxitoxins, are considered safe, effective, long‐acting topical anesthetics (Garcia Camacho et al., 2007). The saxitoxin related tetrodotoxin, a non‐peptide neurotoxin, is being developed by WEX Pharmaceuticals Inc. (www.wexpharma.com) for treatment of cancer‐related pain as an alternative to opioids. Okadaic acid is considered a model potent neurotoxin for analyzing the therapeutic effects of atypical antipsychotic drugs in treatment of cognitive impairment and neuropathological changes of schizophrenia and other neurodegenerative diseases (Garcia Camacho et al., 2007).
Dinoflagelltes produces many bioactive molecules with interesting anticancer properties, that presently have been evidenced only at lab screening phase.
4.5 Biotechnology and uses for macroalgae
Current uses and biotechnology of Macroalgae far exceeds those of Microalgae, with Macroalgae being harvested for food, feed or biocompounds (agar) all across the world, with major concentrations in East Asia. Three distinct algal classes, Chlorophyta, Rhodophyta, and Phaeophyta, have macroalgae representatives, and all classes are being exploited economically.
Macroalgae based fuels hold great promise, directly related to the potential to produce more biomass per unit area in a year than any other form of biomass. The main Chlorophyta species relevant to biofuel uses are belonging to the genus of Ulva and Caulerpa. The red algae species relevant to biofuel uses mainly

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 42 of 258
belong to the order of Gigartinales, Halymeniales and Palmariales. The brown algae relevant to biofuel uses are mainly belonging to the order of Fucales (including representant of the following families: Alariaceae, Fucaceae, Himanthaliaceae, Sargasseae), Laminariales and Tilopteridales.
Sewage can be treated with algae, reducing the need for greater amounts of toxic chemicals than are already used. Algae can be used to capture fertilizers in runoff from farms. When subsequently harvested, the enriched algae itself can be used as fertilizer. Agricultural Research Service scientists found that 60‐90% of nitrogen runoff and 70‐100% of phosphorus runoff can be captured from manure effluents using an algal turf scrubber (ATS). Scientists developed the ATS, which are shallow, 100‐foot raceways of nylon netting where algae colonies can form, and studied its efficacy for three years. They found that algae can readily be used to reduce the nutrient runoff from agricultural fields and increase the quality of water flowing into rivers, streams, and oceans. The enriched algae itself also can be used as a fertilizer. Researchers collected and dried the nutrient‐rich algae from the ATS and studied its potential as an organic fertilizer. They found that cucumber and corn seedlings grew just as well using ATS organic fertilizer as they did with commercial fertilizers. Air‐dried algae from an algal turf scrubber (ATS) captured most of the nitrogen and phosphorus in manure runoff (United States Department of Agriculture, Agricultural Research Service, www.ars.usda.gov/is/AR/archive/may10/algae0510.htm).
Several red macroalgae species are used as food, such as Dulse (Palmaria palmata) and Porphyra in the British Isles, or nori in Asia. Cultivation is relatively simple and began in Japan more than 300 years ago. Major products from macroalgae today are: Agar, a gelatinous substance derived from red algae, has a number of commercial uses. Alginates: Between 100,000 and 170,000 wet tons of Macrocystis are harvested annually in California for alginate extraction and abalone feed. Fertilizer: For centuries seaweed has been used as a fertilizer. This kind of ore they often gather and lay on great heapes, where it heteth and rotteth, and will have a strong and loathsome smell; when being so rotten they cast on the land, as they do their muck, and thereof springeth good corn, especially barley. After spring‐tydes or great rigs of the sea, they fetch it in sacks on horse backes, and carie the same three, four, or five miles, and cast it on the land, which doth very much better the ground for corn and grass. Nutrition: Naturally growing seaweeds are an important source of food, especially in Asia. They provide many vitamins including: A, B1, B2, B6, niacin and C, and are rich in iodine, potassium, iron, magnesium and calcium.
Algae are national foods of many nations: China consumes more than 70 species, including fat choy, a cyanobacterium considered a vegetable; Japan, over 20 species; Ireland, dulse; Chile, cochayuyo. Laver is used to make "laver bread" in Wales where it is known as bara lawr; in Korea, gim; in Japan, nori and aonori. It is also used along the west coast of North America from California to British Columbia, in Hawaii and by the Māori of New Zealand. Sea lettuce and badderlocks are a salad ingredient in Scotland, Ireland, Greenland and Iceland. Dulse is a good source of minerals and
vitamins, contains all trace elements and has a high protein content. Dulse is commonly used in Ireland, Iceland, Atlantic Canada and the Northeast United States. Dulse can be used both fresh or sun‐dried (ground to flakes or powder). Finely diced, it can also be used as a flavour enhancer in place of monosodium glutamate.
Stabilizing substances: Carrageenan, from the red alga Chondrus crispus, is used as a stabiliser in milk products.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 43 of 258
4.6 Biotechnology of Other Aquatic Biomass
Aquatic vascular plants can represent attractive biomass production alternatives especially when integrated into treatment of eutrophicated water bodies. A large number of often invasive water plants, such as Water Hyacinth, Lemna, various reeds, Elodea, Egeria etc become a pest in nutrient rich waters clogging up waterways and suffocating native flora and fauna. On the other hand such plants provide high productivity under significant nutrient removal capacity. Lacking success with microalgae has caused former algae pioneer PetroAlgae to switch technology to cultivation of "microcrops" (Lemna) on nutrient rich lagoons (http://www.petroalgae.com/index.php), offering certain advantages such as a mature technology, easier harvesting (Fig. 9) and process control at the expense of somewhat reduced productivity.
Figure 9 ‐ Removal of algae blooms and water hyacinth at Hertebeesport dam, South Africa (http://www.dwa.gov.za/harties/).
Remediation efforts of polluted water bodies in general can provide significant amounts of free waste
biomass from such water plant material (see Hartebeesport dam http://www.dwa.gov.za/harties/) that may be utilized for fuels production. Alternatively, biological water treatment using constructed wetlands can also provide significant amounts of waste biomass that may be utilized for biofuels production.
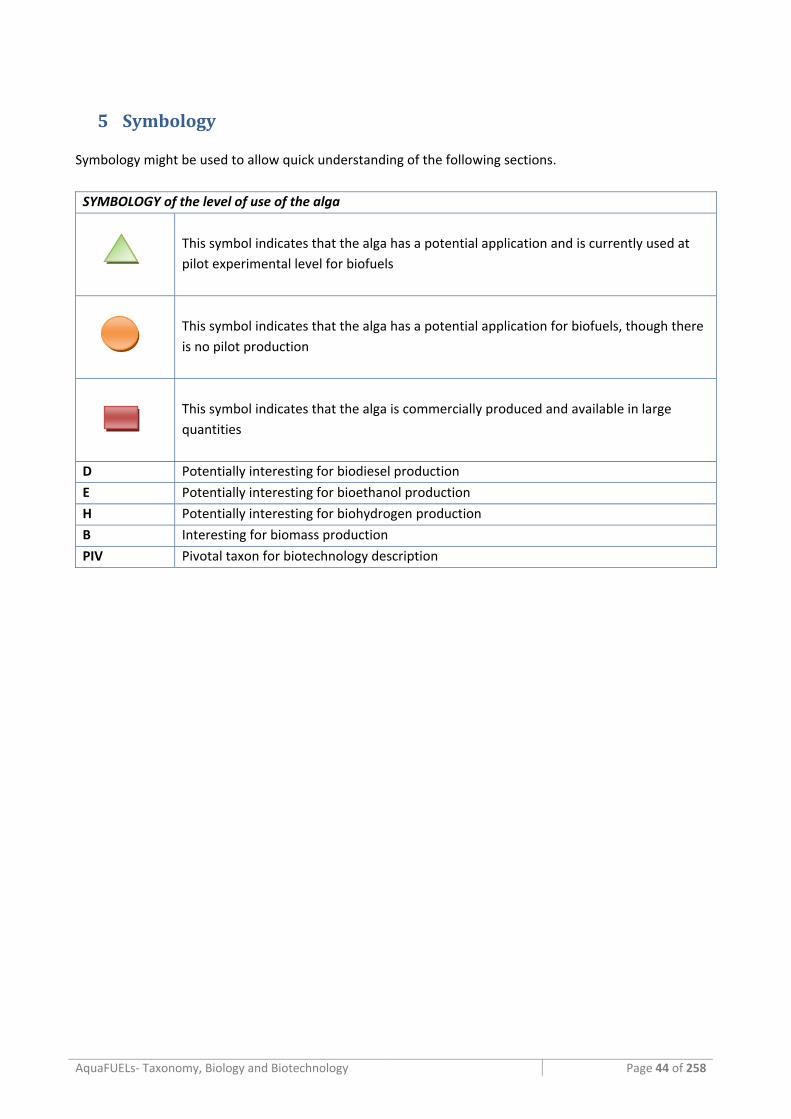
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 44 of 258
5 Symbology
Symbology might be used to allow quick understanding of the following sections.
SYMBOLOGY of the level of use of the alga
This symbol indicates that the alga has a potential application and is currently used at pilot experimental level for biofuels
This symbol indicates that the alga has a potential application for biofuels, though there is no pilot production
This symbol indicates that the alga is commercially produced and available in large quantities
D Potentially interesting for biodiesel production
E Potentially interesting for bioethanol production
H Potentially interesting for biohydrogen production
B Interesting for biomass production
PIV Pivotal taxon for biotechnology description

AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 45 of 258
6 References
Adams D.G., Duggan P.S. (1999) Heterocyst and akinete differentiation in cyanobacteria. Tansley Review No. 107. New Phytologist 144: 3‐33.
Adl S.M., Simpson A.G.B., Farmer M.A., et al. (2005) The new higher level classification of eukaryotes with emphasis on the taxonomy of protists. Journal of Eukaryotic Microbiology 52: 399–451.
Anagnostidis K., Komárek J. (1988) Modern approach to the classification system of cyanophytes. Teil 3‐ Oscillatoriales. Archiv für Hydrobiologie/Algological Studies 50‐53: 327–472.
Anagnostidis K., Komárek J. (1990) Modern approach to the classification system of cyanophytes. Teil 5‐ Stigonematales. Archiv für Hydrobiologie/Algological Studies 59: 1–73.
Andersen R.A., Brett R.W., et al. (1998) Phylogeny of the Eustigmatophyceae based upon 18s rDNA, with emphasis on Nannochloropsis. Protist 149: 61‐74.
Anderson R.A. (2004) Biology and systematics of heterokont and haptophyte algae. American Journal of Botany 91: 1508‐1522.
Archibald J.M. (2007) Nucleomorph genomes: structure, function, origin and evolution. BioEssays 29: 392‐402. Archibald J.M., Keeling P.J. (2002) Recycled plastids: a green movement in eukaryotic evolution. Trends in Genetics 18:
577‐584. Armbrust E.V. (2009) The life of diatoms in the world's oceans. Nature 459: 185‐192 Armbrust E.V., Berges J.A., Bowler C,. et al. (2004) The genome of the diatom Thalassiosira pseudonana: Ecology,
evolution, and metabolism. Science 306: 79‐86. Barsanti L., Gualtieri P. (2006) Algae: Anatomy, Biochemistry, and Biotechnology. CRC Press Taylor & Francis Group,
Boca Raton.
Bergman B., Gallon J.R., Rai A.N., Stal L.J. (1997) N2 fixation by non‐hetrocystous cyanobacteria. FEMS Microbiol. Rev., 19: 139‐185
Bhattacharya D., Yoon H.S., Hackett J.D. (2003) Photosynthetic eukaryotes unite: Endosymbiosis connects the dots. BioEssays 26: 50‐60.
Boussiba S., Vonshak A., Cohen Z., Avissar Y., Richmond A. (1987) Lipid and biomass production by the halotolerant microalga Nannochloropsis salina. Biomass 12: 37‐47
Boussiba S, Sandbank E., Shelef G., Cohen Z., Vonshak A., Ben‐Amotz A., Arad S., Richmond A. (1988) Outdoor cultivation of the marine microalga Isochrysis galbana in open reactors. Aquaculture 72: 247‐253.
Boussiba S., Wu X.Q., Ben‐Dov E., Zarka A., Zaritsky A. (2000) Nitrogen‐fixing cyanobacteria as gene delivery system for expressing mosquiticidal toxins of Bacillus thuringensis ssp. israelensis. Journal of Applied Phycoogy. 12: 461‐467.
Bowler C., Allen A.E., Badger J.H., et al. (2008) The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 456: 239‐244.
Bozarth A., Maier U.G., Zauner S. (2009) Diatoms in biotechnology: modern tools and applications. Applied Microbiology and Biotechnology 82: 195‐201.
Brennan L., Owende P. (2010) Biofuels from microalgae‐A review of technologies for production, processing, and extractions of biofuels and co‐products. Renewable and Sustainable Energy Reviews 14: 557‐577.
Castenholz R.W., Waterbury J.B. (1989) Group I. Cyanobacteria. In: Staley J.T., Bryant M.P., Pfennig N., Holt J.G. (eds.) Bergey's Manual of Systematic Bacteriology, Vol. 3. Williams & Wilkins, Baltimore, pp. 1710–1727.
Castenholz R.W. (2001) Phylum BX. Cyanobacteria. In: Boone D.R., Castenholz R.W. (eds.) Bergey's Manual of Systematic Bacteriology, 2nd Edition, vol. 1. Springer, New York, pp. 473–487.
Cavalier‐Smith T. (1981) Eukaryote kingdoms: seven or nine? Biosystems 14: 461–481.
Cavalier‐Smith T., Chao E.E. (1997) Sarcomonad ribosomal RNA sequences, rhizopod phylogeny, and the origin of euglyphid amoebae. Archiv für Protistenkunde. 147: 227‐236.
Cavalier‐Smith T., Allsopp M.T.E.P., Chao E. E. (1994) Thraustochytrids are Chromists, not Fungi: 18s rRNA Signatures of Heterokonta. Philosophical Transactions: Biological Sciences, 346: 387‐397.
Cembella A.D. (2003) Chemical ecology of eukaryotic microalgae in marine ecosystems. Phycologia 42: 420‐447. Chepurnov V.A., Mann D.G., Sabbe K., Vyverman W. (2004) Experimental studies on sexual reproduction in diatoms.
International Review of Cytology 237: 91–154.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 46 of 258
Chisholm S.W., Olson R.J., Zettler E.R., Goericke R., Waterbury J.B., Welschmeyer N.A. (1988) A novel free‐living prochlorophyte abundant in the oceanic euphotic zone. Nature 334: 340‐343.
Chorus I., Bartram J. (1999) Toxic cyanobacteria in water: A guide to their public health consequences, monitoring and management. World Health Organization.
Chisti Y (2007) Biodiesel from microalgae. Biotechnology Advances 25: 294‐306. Cohen Z., Cohen S. (1991) Preparation of eicosapentaenoic acid (EPA) concentrate from Porphyridium cruentum.
Journal of American Oil Chemists’ Society 68: 16 ‐19. Cohen Y., Padan E., Shilo M. (1975) Facultative anoxygenic photosynthesis in the cyanobacterium Oscillatoria
limnetica. Journal of Bacteriology 123: 855‐861. Coffroth M.A., Santos S.R. (2005) Genetic diversity of symbiotic dinoflagellates in the genus Symbiodinium. Protist 156:
19‐34. Courties C., Vaquer A., Troussellier M., Lautier J., Chrétiennot‐Dinet M.J., Neveux J., Machado C., Claustre H. (1994)
Smallest eukaryotic organism. Nature 370: 255.
Coyer J.A., Smith G.J., Anderson R.A. (2001) Evolution of Macrocystis spp. (Phaeophyta) as determined by ITS1 and ITS2 sequences. Journal of Phycology 37: 574–585.
Del Campo J.A,. Rodríguez H., Moreno J., Vargas M.A., Rivas J., Guerrero M.G. (2004) Accumulation of astaxanthin and lutein in Chlorella zofingiensis (Chlorophyta). Appied Microbiology and Biotechnology 64: 848‐54.
Falkowski P.G., Barber R.T., Smetacek V.V. (1998) Biogeochemical controls and feedbacks on ocean primary production. Science 281: 200–207.
Fargione J., Hill J., Tilman D., Polasky S., Hawthorne P. (2008) Land clearing and the biofuel carbon debt. Science 319: 1235‐1238.
Fast N.M., Xue L., Bingham S., Keeling P.J. (2002) Re‐examining alveolate evolution using multiple protein molecular phylogenies. Journal of Eukaryotic Microbiology 49: 30‐37.
Fawley K.P. (2007) Observations on the diversity and ecology of freshwater Nannochloropsis (Eustigmatophyceae), with descriptions of new taxa. Protist 158: 325‐336.
Fensome R.A., Taylor F.J.R., Norris G., Sarjeant W.A.S., Wharton D.I., Williams G.L. (1993) A classification of living and fossil dinoflagellates. Micropaleontology Special Publication 7, Sheridan Press, Hanover, Pennsylvania, USA.
Field C.B., Behrenfeld M.J., Randerson J.T., Falkowski P.G. (1998) Primary production of the biosphere: integrating terrestrial and oceanic components. Science 281: 237–240.
Fletcher B.J., Beerling D.J., Chaloner W.G. (2004) Stable carbon isotopes and the metabolism of the terrestrial Devonian organism Spongiophyton. Geobiology 2: 107.
Garbary D.J., Gabrielson P.W. (1990) Taxonomy and evolution. In: Cole K.M., Sheath R.G. (eds.) Biology of the red algae. Cambridge University Press, Cambridge, 477‐498.
Garcia Camacho F., Gallardo Rodríguez J., Sánchez Mirón A., Cerón García M.C., Belarbi E.H., Chisti Y., Molina Grima E. (2007) Biotechnological significance of toxic marine dinoflagellates. Biotechnology Advances 25: 176–194.
Garlick S., Oren A., Padan E. (1977) Occurrence of facultative anoxygenic photosynthesis among unicellular and filamentous cyanobacteria. Journal of Bacteriology 129: 623‐629.
Gilson P.R., McFadden G.I. (2002) Jam packed genomes‐a preliminary, comparative analysis of nucleomorphs. Genetica 115: 13‐28.
Gould S.B., Waller R.F., McFadden G.I. (2008) Plastid evolution. Annual Reviews in Plant Biology 59: 491‐517. Griffiths M.J., Harrison S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel
production. Journal of Applied Phycology 21: 493‐507 Guillou L., Eikrem W., Chrétiennot‐Dinnet M.J., Le Gall F., Massana R., Romari K., Pedros‐Alio C., Vaulot D. (2004)
Diversity of picoplanktonic Prasinophyceae assessed by direct SSU rDNA sequencing of environmental samples and novel isolates retrieved from oceanic and coastal marine ecosystems. Protist 155: 193–214.
Guiry M.D., Guiry G.M. (2007) AlgaeBase version 4.2, World‐wide electronic publication, National University of Ireland, Galway AND SUCCESSIVE MODIFICATIONS.
Herdman M., Rippka R. (1988a) Cellular differentiation: akinetes. In: Packer L., Glazer A.N. (eds) Cyanobacteria. In: Colowock S.P., Kaplan N.O. (eds) Methods in Enzimology, 167: 222‐232.
Herdman M., Rippka R. (1988b) Cellular differentiation: hormogonia and baeocytes. In: Packer L., Glazer A.N. (eds) Cyanobacteria. In: Colowock S.P., Kaplan N.O. (eds) Methods in Enzimology, 167: 232‐242.
Herrero A., Flores E. (eds) (2008) The cyanobacteria: Molecular biology, genetics and evolution. Caister Academic Press, Norfolk.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 47 of 258
Herrmann M., Joppe H., Pertile P., Zanella L. (2010) Extracts of Isochrysis sp. Patent Application No. US2010/0080761 A1.
Hibberd D.J. (1981) Notes on the taxonomy and nomenclature of the algal classes Eustigmatophyceae and Tribophyceae (Synonym Xanthophyceae). Botanical Journal of the Linnean Society 82: 93‐119.
Hibberd D.J., Norris R.E. (1984) Cytology and ultrastructure of Chlorarachnion reptans (Chlorarachniophyta divisio nova, Chlorarachniophyceae classis nova). Journal of Phycology 20: 310‐330.
Hoek C. van den, Mann D.G., Jahns H.M. (1995) Algae An introduction to phycology. Cambridge University Press, Cambridge.
Hu Q., Guterman H., Richmond A. (1996) Physiological characteristics of Spirulina platensis (Cyanobacteria) cultured at ultrahigh cell densities. Journal of Phycology 32: 1066–1073.
Hu Q., Zhengyu H., Cohen Z., Richmond A. (1997) Enhancement of eicosapentaenoic acid (EPA) and y‐linolenic acid (GLA) production by manipulating algal density of outdoor cultures of Monodus subterraneus (Eustigmatophyta) and Spirulina platensis (Cyanobacteria). European Journal of Phycoogy. 32: 81‐86.
Hu Q., Sommerfeld M., Jarvis E., et al. (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. Plant Journal 54: 621‐639.
Huheihel M., Ishanu V., Tal . Arad (Malis) S. (2002) Activity of Porphyridium sp. polysaccharide against herpes simplex viruses in vitro and in vivo. Journal of Biochemical and Biophysical Methods 50: 189‐200.
Ishida K., Green B.R., Cavalier‐Smith T. (1999) Diversification of a Chimaeric Algal Group, the Chlorarachniophytes: Phylogeny of Nuclear and Nucleomorph Small Subunit rRNA Genes. Molecular Biology and Evolution 16: 321‐331.
Ishida K., Ishida N., Hara Y. (2000) Lotharella amoeboformis sp. nov.: a New Species of Chlorarachniophytes from Japan. Phycological Research 48: 221‐230.
Ito A., Oikawa T.(2004) Global mapping of terrestrial primary productivity and light‐use efficiency with a process‐based model. In: Shiyomi M., Kawahata H., Koizumi H., Tsuda A., Awaya Y. (eds) Global environmental change in the ocean and on land, TerraPub, Tokyo, pp. 343–358.
Jones W.E. (1962) A key to the genera of the British seaweeds. Field Studies 1: 1 ‐32. Johnston M., Foley J.A., Holloway T., Kucharik C., Monfreda C. (2009) Resetting global expectations from agricultural
biofuels. Environmental Research Letters 4: 019801 doi: 10.1088/1748‐9326/4/1/019801.
Jorquera O., Kiperstok A., Sales E.A., Embiruçu M., Ghirardi M.L. (2010) Comparative energy life‐cycle analyses of microalgal biomass production in open ponds and photobioreactors. Bioresource Technology 101: 1406‐1413.
Kaneko T., Sato S., Kotani H., et al. (1996) Sequence analysis of the genome of the unicellular cyanobacterium Synechocystis sp. strain PCC6803. II. Sequence determination of the entire genome and assignment of potential protein‐coding regions. DNA Research 3: 109‐136.
Kapraun D.F. (2007) Nuclear DNA content estimates in green algal lineages: chlorophyta and streptophyta. Annals of Botany 99: 677‐701.
Keeling P.J. (2004) Diversity and evolutionary history of plastids and their hosts. American Journal of Botany 91: 1481‐1493.
Kim E, Graham L. (2008) EEF2 analysis challenges the monophyly of Archaeplastida and Chromalveolata. PLoS ONE 3: e2621.
Komárek J., Anagnostidis K. (1986) Modern approach to the classification systém of cyanophytes. Teil 2‐ Chroococcales. Archiv für Hydrobiologie/Algological Studies 43: 157‐226.
Komárek J., Anagnostidis K. (1989) Modern approach to the classification system of cyanophytes. Teil 4‐ Nostocales. Archiv für Hydrobiologie/Algological Studies 56: 247‐345.
Kooistra W.H.C.F., Gersonde R., Medlin L.K., Mann D.G. (2007) The origin and evolution of the diatoms: their adaptation to a planktonic existence, In: Falkowski P.G., Knoll A.H. (eds) Evolution of Primary Producers in the Sea, Elsevier, pp. 207–249.
Leander C.A., Porter D. (2001) The Labyrinthulomycota is comprised of three distinct lineages. Mycologia 93: 459‐464. Lebeau T., Robert J.M. (2003) Diatom cultivation and biotechnologically relevant products. Part I: Cultivation at various
scales. Applied Microbiology and Biotechnology 60: 612‐623 Lee R.E. (2008) Phycology, 4th edition, Cambridge University Press. Lehane L., Lewis R.J. (2000) Ciguatera: recent advances, but the risk remains. International Journal of Food
Microbiology 61: 91‐125.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 48 of 258
Leipe D.D., Wainright P.O., Gunderson J.H., Porter D., Patterson D.J., Valois F., Himmerich S., Sogin M.L. (1994) The stramenopiles from a molecular perspective: 16S‐like rRNA sequences from Labyrinthuloides minuta and Cafeteria roenbergensis. Phycologia 33: 369‐377.
Lewis T. E., Nichols P.D., McMeekin T.A. (1999) The biotechnological potential of thraustochytrids. Marine Biotechnology 1: 580‐587.
Loeblich A.R. (1974) Protistan phylogeny as indicated by the fossil record. Taxon 23: 277‐290. Lu C., Acién Fernández F.G., Cañizares Guerrero E., Hall D.O., Molina Grima E. (2002) Overall assessment of Monodus
subterraneus cultivation and EPA production in outdoor helical and bubble column reactors. Journal of Applied Phycology 14: 331‐342.
Lüning K. (1990) Seaweeds: Their environment, biogeography, and ecophysiology. Wiley, New York.
Mann D.G. (1999) The species concept in diatoms (Phycological Reviews 18). Phycologia 38: 437‐495. Mann D.G., Evans K.M. (2007) Molecular genetics and the neglected art of diatomics, In: Brodie J., Lewis J.M. (eds)
Unravelling the algae – the past, present and future of algal molecular systematics, CRC Press, London, pp. 231–265.
Mata T.M., Martins A.A., Caetano N.S. (2010) Microalgae for biodiesel production and other applications: A review. Renewable and Sustainable Energy Reviews 14: 217‐232.
Medlin L.K., Kooistra W.H.F., Potter D., Saunders G.W., Andersen R.A. (1997) Phylogenetic relationships of the 'golden algae' (haptophytes, heterokont chromophytes) and their plastids, In: Bhattacharya D. (ed) Origins of algae and their plastids, Special edition of Plant Systematics and Evolution, Suppl.11, pp. 187–219.
Medlin L.K., Kaczmarska I. (2004) Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 43: 245‐270.
Miyashita H., Ikemoto H., Kurano N., Miyachi S., Chihara M. (2003) Acaryochloris marina gen. et sp. nov. (cyanobacteria), an oxygenic photosynthetic prokaryote containing chl d as a major pigment. Journal of Phycology 39: 1247‐1253.
Moreira D., López‐García P. (2002) The molecular ecology of microbial eukaryotes unveils a hidden world. Trends in Microbiology 10: 31‐38.
Oren, A. (2004). A proposal for further integration of the cyanobacteria under the Bacteriological Code. International Journal of Systematic and Evolutionary Microbiology 54: 1895‐1902.
Oren A., Shilo M. (1979) Anaerobic heterotrophic dark metabolism in the cyanobacterium Oscillatoria limnetica: sulphur respiration and lactate fermentation. Archives of Microbiology 122: 77‐84.
Patterson D.J. (1989) Stramenopiles: chromophytes from a protistan perspective, In: Green J.C., Leadbeater B.S.C., Diver W.L. (eds), The chromophyte algae problems and perspectives, Clarendon Press, Oxford, pp. 357‐379.
Pedroni P.M., Lamenti G., Prosperi G., Ritorto L., Scolla G., Capuano F., Valdiserri M. (2004) Enitecnologie R & D project on microalgae biofixation of CO2: Outdoor comparative tests of biomass productivity using flue gas CO2 from a NGCC power plant. Proceedings of Seventh International Conference on Greenhouse Gas Control Technologies (GHGT‐7), 5–9 September 2004, Vancouver, Canada.
Pulz O (2001) Photobioreactors: Production systems for phototrophic microorganisms. Applied Microbiology and Biotechnology 57: 287‐293.
Pulz O, Gross W (2004) Valuable products from biotechnology of microalgae. Applied Microbiology and Biotechnology 65, 635‐648.
Pulz O, Scheibenbogen K (1998) Photobioreactors: Design and performance with respect to light energy input. In: Scheper T (ed.) Advances in Biochemical Engineering/Biotechnology, Springer‐Verlag, Berlin, pp. 123‐152.
Ragan M.A., Bird C.J., Rice E.L., Gutell R.R., Murphy C.A., Singh R.K. (1994) A molecular phylogeny of the marine red algae (Rhodophyta) based on the nuclear small‐subunit rRNA gene. Proceedings of the National Academy of Sciences of the USA 91: 7276‐7280.
Raghukumar S. (1996) Morphology, taxonomy and ecology of tharustochytrids and labyrinthulids, the marine counterparts of zoosporic fungi. In: Dayal R. (ed.) Advances in zoosporic fungi. M D Publications Pvt Ltd, New Delhi, 35‐58.
Raghukumar S. (2002) Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids), European Journal of Protistology 38: 127–145.
Rai A.N., Bergman B., Rasmussen U. (eds.) (2002) Cyanobacteria in symbiosis. Kluwer Academic Publishers, Dodrecht.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 49 of 258
Ramachandra T.V., Mahapatra D.M., Karthick B., Gordon R. (2009) Milking diatoms for sustainable energy: Biochemical engineering versus gasoline‐secreting diatom solar panels. Industrial and Engineering Chemistry Research 48: 8769‐8788.
Raven P.H., Evert R.F., Eichhorn S.E. (2005) The Biology of Plants 7th edition, W. H. Freeman and Company, New York.
Reitan K.I., Rainuzzo J.R., Olsen Y. (1994) Effect of nutrient limitation on fatty‐acid and lipid‐content of marine microalgae. Journal of Phycology 30: 972‐979.
Rodolfi L., Chini Zittelli G., Bassi N., et al. (2009) Microalgae for oil: strain selection, induction of lipid synthesis and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Řezanka T., Petránková M., Cepák V., Přibyl P., Sigler K., Cajthaml T. (2010) Trachydiscus minutus, a new biotechnological source of eicosapentaenoic acid. Folia Microbiologica 55: 265–269.
Roessler P.G. (1988) Changes in the activities of various lipid and carbohydrate biosynthetic enzymes in the diatom Cyclotella cryptica in response to silicon deficiency. Archives of Biochemistry and Biophysics 267: 521–528.
Round F.E., Crawford R.M., Mann D.G. (1990) The Diatoms – Biology and Morphology of the Genera. Cambridge University Press, Cambridge, UK.
Satoh M., Iwamoto K., Suzuki I., Shiraiwa Y. (2009) Cold stress stimulates intracellular calcification by the coccolithophore, Emiliania huxleyi (Haptophyceae) under phosphate‐deficient conditions. Marine Biotechnology 11: 327‐333.
Scott E.L., Kootstra A.M.J., Sanders J.P.M. (2010) Perspectives on bioenergy and biofuels. In: Singh, O.V and S.P. Harvey (eds.) Sustainable Biotechnology. Elsevier, The Netherlands, pp: 179‐194.
Searchinger T., Heimlich R., Houghton R.A., Dong F., Elobeid A., Fabiosa J., Tokgoz S. , Hayes D. Yu T.‐H. (2008) Use of U.S. croplands for biofuels increases greenhouse gases through emissions from landuse change. Science 319: 1238‐1240.
Senn T.L. (1987) Seaweed and Plant Growth. Clemson University, Clemson. Sheehan J., Dunahay T., Benemann J., Roessler P. (1998) A Look Back at the U.S. Department of Energy’s Aquatic
Species Program—Biodiesel from Algae. Close‐out Report. NREL/TP‐580‐24190, 295 pp. Sims P.A., Mann D.G., Medlin L.K. (2006) Evolution of the diatoms: Insights from fossil, biological and molecular data.
Phycologia 45: 361‐402. Smayda T.J. (1997) What is a bloom? A commentary. Limnology and Oceanography 42: 1132‐1136. Sorhannus U. (2007) A nuclear‐encoded small‐subunit ribosomal RNA timescale for diatom evolution. Marine
Micropaleontology 65: 1‐12. Stace C. A. (1991) Plant Taxonomy and Biosystematics. Cambridge University Press. Stanier R.Y., Cohen‐Bazire G. (1977) Phototrophic prokaryotes: the cyanobacteria. Annual Reviews of Microbiology 31:
225‐234. Stephenson A.L., Kazamia E., Dennis J.S., Howe C.J., Scott S.A., Smith A.G. (2010) Life‐cycle assessment of potential
algal biodiesel production in the United Kingdom: A comparison of raceways and Aair‐lift tubular bioreactors. Energy Fuels 24: 4062‐4077.
Sym S.D., Pienaar R.N. (1993) The class Prasinophyceae. Progress in Phycological Research 9: 281‐376. Taylor F.J.R., Hoppenrath M., Saldarriaga J.F. (2008) Dinoflagellate diversity and distribution. Biodiversity Conservation
17: 407‐418. Thomas D. (2002) Seaweeds. The Natural History Museum, London.
Tredici M.R. (2004) Mass Production of Microalgae: Photobioreactors. In: Richmond A (ed.) Handbook of Microalgal Culture. Blackwell Science Ltd, Oxford, pp 178‐214.
Tredici M.R. (2010) Photobiology of microalgae mass cultures: understanding the tools for the next green revolution. Biofuels 1: 143‐162.
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G., (eds.) New Technologies in Aquaculture: Improving production efficiency, quality and environmental management. Woodhead Publishing Ltd, Cambridge, UK, and CRC Press LLC, Boca Raton, FL, USA., pp. 610‐676.
Tredici M.R., Chini Zittelli G., Rodolfi L. (2010) Photobioreactors In: Flickinger M.C., Anderson S. (eds) Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology. John Wiley & Sons, Inc., Hoboken, NJ, USA. Vol 6, pp. 3821‐3838.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 50 of 258
Uduman N., Qi Y., Danquah M.K., Forde G.M., Hoadley A. (2010) Dewatering of microalgal cultures: A major bottleneck to algae‐based fuels. Journal of Renewable and Sustainable Energy 2: 012701 doi:10.1063/1.3294480.
Warwick B, Paul J.H., Campbell L., Griffin D., Houchin L., Fuentes‐Ortega A., Müller‐Karger F. (2003) Vertical structure of the phytoplankton community associated with a coastal plume in the Gulf of Mexico.Marine Ecology Progress Series 251: 87‐101.
Wehr J.D., Sheath R.G. (eds) (2003) Freshwater algae of North America: Ecology and Classification. Elsevier Inc, Amsterdam.
Whitton B.A., Potts, M. (2000) The ecology of cyanobacteria. Their diversity in time and space. Kluwer Academic Publishers, Dodrecht.
Wijffels R.H., Barbosa M.J. (2010) An outlook on microalgal biofuels. Science 329: 796‐799. Wilmotte A., Herdman M. (2001) Phylogenetic relationships among the cyanobacteria based on 16S rRNA sequences.
In: Boone D.R., Castenholz R.W. (eds.) Bergey's Manual of Systematic Bacteriology, 2nd Edition, vol. 1, Springer, New York, pp. 487–493.
Woelkerling W.J. (1990) An introduction. In: Cole K.M., Sheath R.G. (eds.) Biology of the red algae. Cambridge University Press, Cambridge, pp. 1‐6.
Wolk P.C. (1973) Physiology and cytological chemistry of blue‐green algae. Bacteriological Reviews 37: 32‐101.
Worden A.Z. (2006) Picoeukaryote diversity in coastal waters of the Pacific Ocean. Aquatic Microbial Ecology 43: 165‐175.
Xiaoqiang W., Vennison S.J., Huirong L., Ben‐Dov E., Zaritsky A., Boussiba S. (1997) Mosquito larvicidal activity of transgenic Anabaena strain PCC 7120 expressing combinations of genes from Bacillus thuringiensis subsp. Israelensis. Applied and Environmental Microbiology 63: 4971‐4974.
Zah R., Böni H., Gauch M., Hischier R., Lehmann M., Wäger P. (2007) Life cycle assessment of energy products: Environmental assessment of biofuels. http://www.bioenergywiki.net/images/8/80/Empa_Bioenergie_ExecSumm_engl.pdf.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 51 of 258
7 Prokaryotic microalgae
7.1 Cyanobacteria
7.1.1 Arthrospira sp. (common name spirulina)
Figure 10 ‐ Arthrospira jenneri
Picture from http://www‐cyanosite.bio.purdue.edu
SYMBOLS: B, PIV
TAXONOMY
Botanical Code
Phylum Cyanobacteria Class Cyanophyceae Order Oscillatoriales Family Phormidiaceae Genus Arthrospira Species
Microbiological Code
Group Cyanobacteria Subsection 3 Family 3.1 Genus Arthrospira
Important Species
A. fusiformis, A. jenneri, A. maxima, A. platensis
BIOLOGY Arthrospira (Spirulina) is a microscopic blue‐green alga in the shape of a spiral coil, living both in sea and fresh water. Spirulina is the common name for human and animal food supplements produced primarily from two species of cyanobacteria: Arthrospira platensis, and Arthrospira maxima. Though referred to as 'algae' because they are aquatic organisms capable of photosynthesis, cyanobacteria are not related the eukaryotic algae (Vonshak, 1997).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 52 of 258
Arthrospira is cultivated around the world, and is used as a human dietary supplement, as well as a whole food, and is available in tablet, flake, and powder form. It is also used as a feed supplement in the aquaculture, aquarium, and poultry industries (Ciferri, 1983).
Arthrospira are free‐floating filamentous cyanobacteria characterized by cylindrical, multicellular trichomes in an open left‐hand helix. They occur naturally in tropical and subtropical lakes with high pH and high concentrations of carbonate and bicarbonate. Arthrospira platensis occurs in Africa, Asia and South America, whereas Arthrospira maxima is confined to Central America. These species were once classified in the genus Spirulina. They are in fact Arthrospira; nevertheless, the older term Spirulina remains in use for historical reasons. Spirulina was found in abundance at Lake Texcoco by French researchers in the 1960s, but there is no reference to its use there as a daily food source after the 16th century. The first large‐scale Spirulina production plant, run by Sosa Texcoco, was established there in the early 1970s.
Spirulina may have an even longer history in Chad, as far back as the 9th century Kanem Empire. It is still in daily use today, dried into cakes called dihé, harvested from small lakes and ponds around Lake Chad (Abdulqader et al., 2000).
Biochemical composition of algae and aquatic biomass: main constituents
Spirulina contains an unusually high amount of protein with, between 55% and 77% of dry weight (Babadzhanov et al., 2004; Tokusoglu et al., 2003). It is a complete protein source containing all essential amino acids, though with reduced amounts of methionine, cysteine, and lysine when compared to the proteins of meat, eggs, and milk. It is, however superior to typical plant protein, such as that from legumes. Spirulina is rich in gamma‐linolenic acid (GLA), and also contains alpha‐linolenic acid (ALA), linoleic acid (LA), stearidonic acid (SDA). Spirulina also contains vitamin B1 (thiamine), B2 (riboflavin), B3 (nicotinamide), B6 (pyridoxine), B9 (folic acid), vitamin C, vitamin D, vitamin A, and vitamin E. Spirulina is a rich source of potassium, and also contains calcium, chromium, copper, iron, magnesium, manganese, phosphorus, selenium, sodium, and zinc. Spirulina contains many pigments, including chlorophyll a, beta‐carotene, echinenone, myxoxanthophyll, zeaxanthin, canthaxanthin, diatoxanthin, 3'‐hydroxyechinenone, beta‐cryptoxanthinand, oscillaxanthin, plus the phycobiliproteins c‐phycocyanin and allophycocyanin (Leema et al., 2010)
BIOTECHNOLOGY
Culture media
Spirulina can live in a wide range of media with elevated salinity and high alkalinity. See http://www.antenna.ch/en/malnutrition/grow‐your‐own‐spirulina.html) for an exemplary growth medium.
Cultivation methods
Since Spirulina cultivation does not include a stress phase, the algae can be cultivated in semi continuous mode with a daily, constant amounts being harvested maintaining the culture in a stable state. The optimal temperature for growth is 35‐38 °C while the minimum is 15‐20 °C.
Production system
Spirulina is being produced in natural open water bodies of suitable temperature, salinity and alkalinity, and in raceway ponds cultivation using appropriate growth media. Most cultivated Spirulina is produced in open‐channel raceway ponds, with paddle‐wheels used to agitate the waterculture. The largest commercial producers of Spirulina are located in the United States, Thailand, India, Taiwan, China, Pakistan, Burma

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 53 of 258
(a.k.a. Myanmar) and Chile (Borowitzka, 1992; 1999; Vonshak (ed.), 1997). Paddle wheels speed in the order of 20 cm s‐1 has been recommended.
Spirulina cultivation may be carried out in closed and open systems. The first one involve laboratory and pilot‐scale photobioreactors (Vonshak (ed.), 1997; Tredici and Chini Zittelli, 1998), not used in industrial production.
CO2 supply is not normally required as the high pH facilitates uptake of CO2 from the atmosphere, and sodium sodium bicarbonate may serve as carbon source.
Harvesting methods and Biomass processing
Spirulina is forming large aggregates and can be harvested easily using nets. Filtration is simply accomplished by passing the culture through a fine weave cloth, using gravity as the driving force. Synthetic fibre cloth (especially polyamide or polyester) with a mesh size of about 30 to 50 microns is the preferred filtering mediumdevice. Supporting the filtration cloth by a fine net will accelerate somewhat the filtration and protect the cloth against rupturing, but a simple bag made from the cloth works well also. The biomass is dried usually spray‐dried and transformed to pills directly.
Scaling up limitation
For a health product contaminating contamination by small animals can pose a serious problem. Since Spirulina prefers temperatures above 30 degrees centigrade, reduced temperatures during winter even in subtropical climates require temporary breaks in the production process.
HIGHLIGHTS IN BIOTECHNOLOGY
Spirulina is one of only two algal species (together with Chlorella) that has been cultivated for decades in a variety of ponds and bioreactors.
Production price is estimated at $ 5 per kg, and dried algae are sold as food and feed additives only.
References
Abdulqader, G., Barsanti, L., Tredici, M. (2000) Harvest of Arthrospira platensis from Lake Kossorom (Chad) and its household usage among the Kanembu. Journal of Applied Phycology 12: 493‐498.
Babadzhanov A.S., Abdusamatova N., Yusupova F.M., Faizullaeva N., Mezhlumyan L..G., Malikova M.K.. (2004) Chemical composition of Spirulina platensis cultivated in Uzbekistan. Chemistry of Natural Compounds. 40: 276‐279.
Borowitzka M.A. (1992) Algal biotechnology products and processes ‐ matching science and economics. Journal of Applied Phycology 3: 267‐279.
Borowitzka M.A. (1999) Commercial production of microalgae: ponds, tanks, tubes and fermenters. Journal of Biotechnology 70: 313‐321.
Ciferri O. (1983) Spirulina, the edible microorganism. Microbiological Reviews 47: 551‐578. Garrity G.M., Boone D.R., Castenholz R.W. (eds) (2004) Bergey’s Manual of Systematic Bacteriology, Second Edition, Volume 1, The
Archaea and the deeply branching and phototrophic Bacteria, Springer. Leema J.T.M., Kirubagaran R., Vinithkumar N.V., Dheenan P.S., Karthikayulu S. (2010) High value pigment production from
Arthrospira (Spirulina) platensis cultured in seawater. Bioresource Technology 101: 9221‐9227. Tokusoglu O., Unal M.K. (2003) Biomass nutrient profiles of three microalgae: Spirulina platensis, Chlorella vulgaris, and Isochrysis
galbana. Journal of Food Science 68: 1144‐1148. Vonshak, A. (ed.) (1997) Spirulina platensis (Arthrospira): Physiology, Cell‐biology and Biotechnology. London: Taylor & Francis,
1997.
.
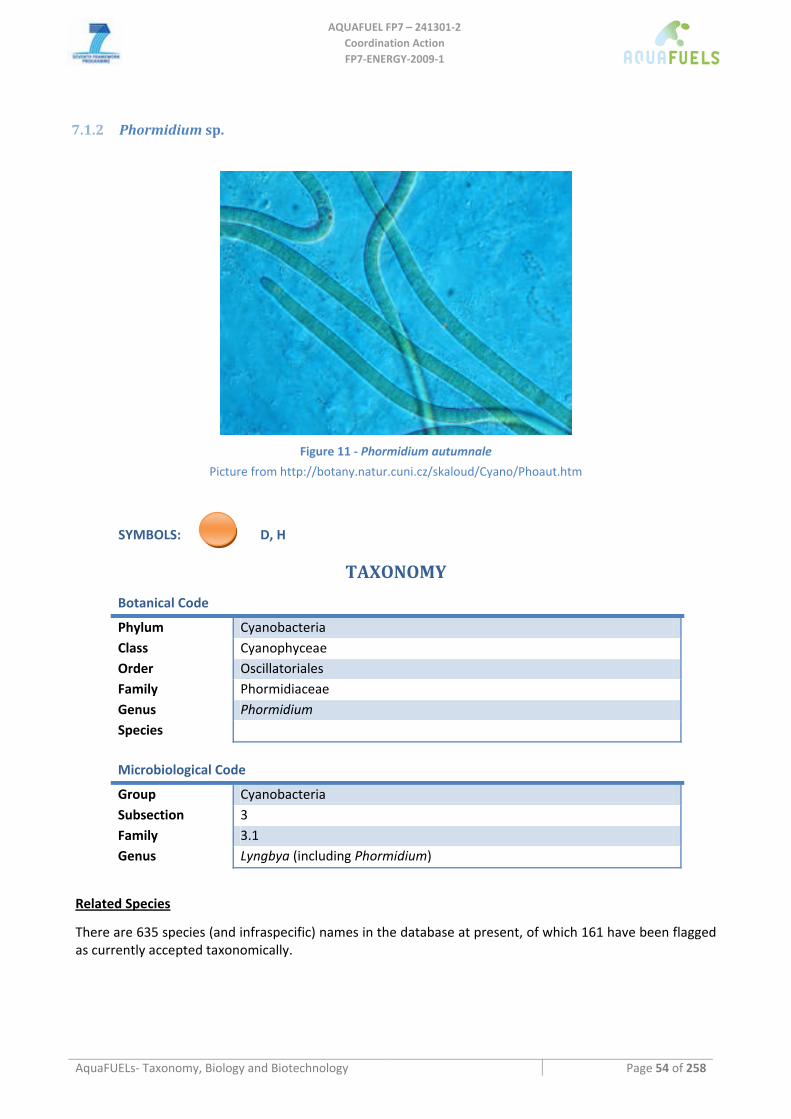
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 54 of 258
7.1.2 Phormidium sp.
Figure 11 ‐ Phormidium autumnale
Picture from http://botany.natur.cuni.cz/skaloud/Cyano/Phoaut.htm
SYMBOLS: D, H
TAXONOMY
Botanical Code
Phylum Cyanobacteria Class Cyanophyceae Order Oscillatoriales Family Phormidiaceae Genus Phormidium Species
Microbiological Code
Group Cyanobacteria Subsection 3 Family 3.1 Genus Lyngbya (including Phormidium)
Related Species
There are 635 species (and infraspecific) names in the database at present, of which 161 have been flagged as currently accepted taxonomically.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 55 of 258
BIOLOGY
Trichomes are isopolar, more or less straight, coiled or waved, usually 2‐14 μm wide, uniserial, never branched, composed of cylindrical up to slightly barrel‐shaped cells, more or less isodiametrical or slightly shorter or longer than wide, constricted or unconstricted at the cross walls, not attenuated and bent or screw‐like twisted towards the ends, motile (waving, creeping, oscillations) within and out of sheaths. End cells widely rounded, attenuated or pointed, sometimes with calyptra. Sheaths develop facultatively in different frequencies only in suboptimal conditions, or in dependence on changing environmental factors, or regularly in all conditions. Sheaths are tube‐like, firm, colourless, joined to the trichomes, not layered, opened at the ends, containing always only one trichome. Cell content usually blue‐green, rarely brownish, pinkish or violet, sometimes modifications with stable PE:PC ratio occur; thylakoids situated perpendicularly to the cell wall (radially in a cross section). Cell division crosswise, perpendicularly to the long axis of a trichome, daughter cells grow up to the original size before the next division. All cells capable of division with the exception of apical ones. Reproduction by hormogonia, which separate at the end parts of trichomes by help of necridic cells or by fragmentation of whole trichomes with necridic cells.
Rarely solitary filaments, usually in mats on different aeric or water substrates (soil, wet rocks, mud, water plants, stones and woods in both stagnant and streaming waters), some species occur in the marine littoral. Several species are known from extreme habitats (thermal springs, desert soils, etc.), few of them take part in the biolithogenic processes and form travertine crusts in limestone water biotopes.
BIOTECHNOLOGY
Phormidium has been proposed by many authors for treatment of different effluents (swine, aquaculture, municipal, industrial) due to its ability to remove nutrients and to degrade pollutants as phenols and hydrocarbons (Cañizares‐Villanueva et al., 1994; Dumas et al., 1998; Satheesh Kumar et al., 2009; Shashirekha et al., 1997).
Phormidium extracts have shown antimicrobial activity in many screenings (Biondi et al., 2008; Rodríguez‐Meizoso et al., 2008) and several bioactive molecules have been individuated, such as anti‐HIV glycolipids and hierridin, a molecule with antiplasmodial activity (Gustafson et al., 1989; Papendorf et al., 1998). A bioactive Antarctic P. priestleyi was cultivated under continuous illumination at low intensity in a 10‐L bubbled reactor obtaining productivies of 95 mg L‐1 day‐1 (Biondi et al., 2008). At present, extracts from Phormidium persicinum have been included in cosmetic products as skin rejuvenator (Morvan and Valee, 2007, 2010).
Though the studies have all been limited to laboratory scale, Phormidium have been considered for hydrogen production (Patel and Madamwar, 1994; Prabaharan and Subramanian, 1996) and also for biodiesel production (Francisco et al., 2010).
References
Biondi N., Tredici M.R., Taton A., Wilmotte A., Hodgson D.A., Losi D., Marinelli F. (2008) Cyanobacteria from benthic mats of Antarctic lakes as a source of new bioactivities. Journal of Applied Microbiology 105: 105‐115.
Cañizares‐Villanueva R.O., Ramos A., Corona A.I., Monroy O., De La Torre M., Gomez‐Lojero C., Travieso L. (1994) Phormidium treatment of anaerobically treated swine wastewater. Water Research 28: 1891‐1895.
Dumas A., Laliberté G., Lessard P., de la Noüe J. (1998) Biotreatment of fish farm effluents using the cyanobacterium Phormidium bohneri. Aquacultural Engineering 17: 57‐68.
Francisco E.C., Neves D.B., Jacob‐lopes E., Franco T.T. (2010) Microalgae as feedstock for biodiesel production: Carbon dioxide sequestration, lipid production and biofuel quality. Journal of Chemical Technology and Biotechnology 85: 395‐403.
Gomont M. (1892 '1893'). Monographie des Oscillariées (Nostocacées homocystées). Annales des Sciences Naturelles, Botanique, Series 7 16: 91‐264, Plates 1‐7. [BIOLOGY section]
Garrity G.M., Boone D.R., Castenholz R.W. (eds) (2004) Bergey’s Manual of Systematic Bacteriology, Second Edition, Volume 1, The Archaea and the deeply branching and phototrophic Bacteria, Springer. [BIOLOGY section]
Gustafson K.R., Cardellina II J.H., Fuller R.W., Weislow O.S., Kiser R.F., Snader K.M., Patterson G.M.L, Boyd M.R (1989) AIDS‐antiviral sulfolipids from cyanobacteria (blue‐green algae). Journal of the National Cancer Institute 81: 1254‐1258.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 56 of 258
Komarek J. (1992) in: http://www.cyanodb.cz/Phormidium [BIOLOGY section] Morvan P.Y., Vallee R. (2007) Effect of micro‐algal extracts on thioredoxin expression in human skin cells and their protection for
skin. IFSCC Magazine, 10(2): 119‐126. Morvan P.Y., Vallee R. (2010) New solution for brighter and younger skin. PersonalCare Magazine, June 2010. Papendorf O., Konig G.M., Wright A.D. (1998) Hierridin B and 2,4‐dimethoxy‐6‐heptadecyl‐phenol, secondary metabolits from the
cyanobacterium Phormidium ectocarpi with antiplasmodial activity. Phytochemistry 49: 2383‐2386. Patel S., Madamwar D. (1994) Photohydrogen production from a coupled system of Halobacterium Halobium and Phormidium
valderianum. International Journal of Hydrogen Energy 19: 733‐738. Prabaharan D., Subramanian G. (1996) Oxygen‐free hydrogen production by the marine cyanobacterium Phormidium valderianum
BDU 20041. Bioresource Technology 57: 111‐116. Rodríguez‐Meizoso I., Jaime L., Santoyo S., Cifuentes A., García‐Blairsy Reina G., Señoráns F.J., Ibáñez E. (2008) Pressurized fluid
extraction of bioactive compounds from Phormidium species. Journal of Agricultural and Food Chemistry 56: 3517‐3523. Satheesh Kumar M., Muralitharan G., Thajuddin N. (2009) Screening of a hypersaline cyanobacterium, Phormidium tenue, for the
degradation of aromatic hydrocarbons: naphthalene and anthracene. Biotechnology Letters 31: 1863‐1866. Shashirekha S., Uma L., Subramanian G. (1997) Phenol degradation by the marine cyanobacterium Phormidium valderianum BDU
30501. Journal of Industrial Microbiology & Biotechnology 19: 130‐133.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 57 of 258
7.1.3 Anabaena sp.
Figure 12 ‐ Anabaena siamensis 40x
Picture from UNIFI
SYMBOLS: H
TAXONOMY
Botanical Code
Phylum Cyanobacteria Class Cyanophyceae Order Nostocales Family Nostocaceae Genus Anabaena Species
Microbiological Code
Group Cyanobacteria Subsection 4 Family 4.1 Genus Anabaena
Species
There are 536 species (and infraspecific) names in the database at present, of which 80 have been flagged as currently accepted taxonomically.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 58 of 258
BIOLOGY
Anabaena trichomes are untapered with constrictions at the cross‐walls. They can be straight, curved or helical. The cells are cylindrical, spherical or barrel‐shaped and, usually, broader than longer with a width of 2‐10 μm. The terminal cell may be round, tapered or conical. Heterocysts are intercalary, terminal or both and thei shape, usually spherical to barrel‐shaped, may be conical when terminal. Akinetes are formed with a species‐dependent position in the trichome. No trichome sheath, but often a mucilaginous covering, is present. Trichomes are usually motile. Reproduction is by trichome fragmentation. Gas vesicles are present in a number of species, especially planktonic.
Anabaena is a main component of freshwater and saline lake phytoplankton. Marine species also occur, but never dominate phytoplankton community. Several Anabaena species are able to produce toxins: the neurotoxins anatoxin‐a (A. flos‐aquae, A. circinalis, A. mendotae, A. planktonica, Anabaena sp.), anatoxin‐a(S) (A. flos‐aquae), saxitoxin (A. circinalis), and BMAA (A. variabilis, Anabaena sp.) and the hepatotoxins microcystin‐LR (A. flos‐aquae, A. circinalis, A. lemmermannii, A. subcylindrica, Anabaena sp.) and cylindrospermopsin (A. bergii, A. lapponica).
Anabaena has been the model organism for studies on cyanobacterial nitrogen fixation, including heterocyst structure and nitrogen fixation genes. This genus includes both obligate photoautotrophs and facultative heterotrophs.
BIOTECHNOLOGY
Besides toxins Anabaena can produce a number of bioactive molecules, such as enzyme inhibitors (e.g., circinamide, anabaenopeptilides), cardioactive molecules (e.g., anabaenopeptins, pawainaphycin C), and antimicrobial agents (e.g., bastadins, laxaphycins, spiroidesin), that may have a potential interest for the pharmaceutical industry.
Freshwater Anabaena strains are cultivated in Allen and Arnon (1955) growth medium or BG110 (Rippka et al., 1979), while marine strains are usually cultivated in ASP‐2 medium without nitrogen (Van Baalen, 1962).
Outdoor cultures of Anabaena have been performed, to verify the production potential of biotechnologically interesting polysaccharides, in a 1‐m2 deep open pond obtaining biomass productivities (with a 10‐cm deep culture) from 9 to 23 g m‐2 day‐1 depending on the season, with a photosynthetic efficiency around 2.4% and in a 55‐L airlift tubular photobioreactor, obtaining volumetric productivities of 0.4 g L‐1 day‐1 correspondint to 9‐10g m‐2 day‐1 in winter (Moreno et al., 2003). Hu et al. (1996) studied the effect of light path length, mixing mode, cell density and dilution rate on productivity of Anabaena siamensis using a flat inclined modular photobioreactor, obtaining with air bubbling at the optimal cell density of 3.2 ± 0.7 g L‐1 maximum productivities of ca 1.43 g L‐1 day‐1 with a light path of 2.6 cm.
Research on hydrogen production by Anabaena started in the 1970s (Benemann and Weare, 1974) and after numerous laboratory researches (Lopes‐Pinto et al., 2002), has led to some small scale experiments outdoors. In a 4.35‐L volume photobioreactor made of 10‐mm internal diameter PVC tubing with A. variabilis mutant (PK84) Tsygankov et al. (2002) obtained a very low efficiency of conversion from light energy into biomass (maximum 0.82%) and into hydrogen (maximum 0.094%). With the same photobioreactor, Lindlblad et al. (2002) obtained a maximum conversion efficiency of 0.042% using the hydrogen uptake deficient mutant AMC 414 of Anabaena PCC 7120. Using a gas‐tight box with outside dimensions of 136.8 x 108.6 x 13 cm and internal dimensions of 128.4 x 100.2 x 13 cm, containg 20 L of culture and 8 L of glass beads and ca 158‐L of gas phase, Smith and Lambert (1981) obtained with Anabaena cylindrica a sustained hydrogen production for over one month.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 59 of 258
References
Allen M.B., Arnon D.I. (1955) Studies on nitrogen‐fixing blue‐green algae. 1. Growth and nitrogen‐fixation by Anabaena cylindrica Lemm. Plant Physiology 30: 366‐372.
Benemann J.R., Weare N.M.(1974) Hydrogen evolution by nitrogen‐fixing Anabaena cylindrica cultures. Science 184: 174‐175. Bornet, É. and Flahault, C. (1886 '1888'). Revision des Nostocacées hétérocystées contenues dans les principaux herbiers de France.
Annales des Sciences Naturelles, Botanique, Septième Série 7: 177‐262. [BIOLOGY section] Burja A.M., Banaigs B., Abou‐Mansour E., Burgess J.G., Wright P.C. (2001) Marine cyanobacteria – a prolific source of natural
products. Tetrahedron 57: 9347‐9377. Castenholz R.W. (1989) Family I. Nostocaceae. Genus I. Anabaena Bory de St. Vincent 1822.. In: Staley J.T., Bryant M.P., Pfennig N.,
Holt J.G. (eds.) Bergey’s Manual of Systematic Bacteriology, Volume 3, Williams & Wilkins, Baltimore, pp. 1783‐1785. [BIOLOGY section]
Garrity G.M., Boone D.R., Castenholz R.W. (eds) (2004) Bergey’s Manual of Systematic Bacteriology, Second Edition, Volume 1, The Archaea and the deeply branching and phototrophic Bacteria, Springer. [BIOLOGY section]
Hu Q., Guterman H., Richmond A. (1996) A Flat Inclined Modular Photobioreactor for outdoor mass cultivation of photoautotrophs. Biotechnology and Bioengineering, 51: 51‐60.
Jaiswal P., Kumar Singh P., Prasanna R. (2008) Cyanobacteria bioactive molecules ‐ an overview of their toxic properties. Canadian Journal of Microbiology 54: 701‐717.
Lindblad P., Christensson K., Lindberg P., Fedorov A., Pinto F., Tsygankov A. (2002) Photoproduction of H2 by wildtype Anabaena PCC 7120 and a hydrogen uptake deficient mutant: from laboratory experiments to outdoor culture. International Journal of Hydrogen Energy 27: 1271‐1281.
Lopes Pinto F.A., Troshina O., Lindblad P. (2002) A brief look at three decades of research on cyanobacterial hydrogen evolution International Journal of Hydrogen Energy 27: 1209‐1215.
Moreno J., Vargas M.A., Rodríguez H., Rivas J., Guerrero M.G. (2003) Outdoor cultivation of a nitrogen‐fixing marine cyanobacterium, Anabaena sp. ATCC 33047. Biomolecular Engineering 20: 191‐197.
Rippka R., Deruelles J., Waterbury J.B., Herdman M., Stanier R.Y. (1979) Generic assignments strain histories and properties of pure cultures of cyanobacteria. Journal of General Microbiology 111: 1‐61.
Sivonen K., Börner T. (2008) Bioactive compounds produced by cyanobacteria. In: Herreo A., Flores E. (eds.) The Cyanobacteria. Molecular Biology, Genetics and Evolution, Caister Academic Press, Norfolk (UK), pp. 159‐191.
Smith G.D., Lambert G.R. (1981) An outdoor biophotolytic system using the cyanobacterium Anabaena cylindrica B629. Biotechnology and Bioengineering 23:. 213‐220.
Tsygankov A.A., Fedorov A.S., Kosourov S.N., Rao K.K. (2002) Hydrogen production by cyanobacteria in an automated outdoor photobioreactor under aerobic conditions. Biotechnology and Bioengineering 80: 777‐783.
Van Baalen C. (1962) Studies on marine blue‐green algae. Botanica Marina 4: 197‐201.
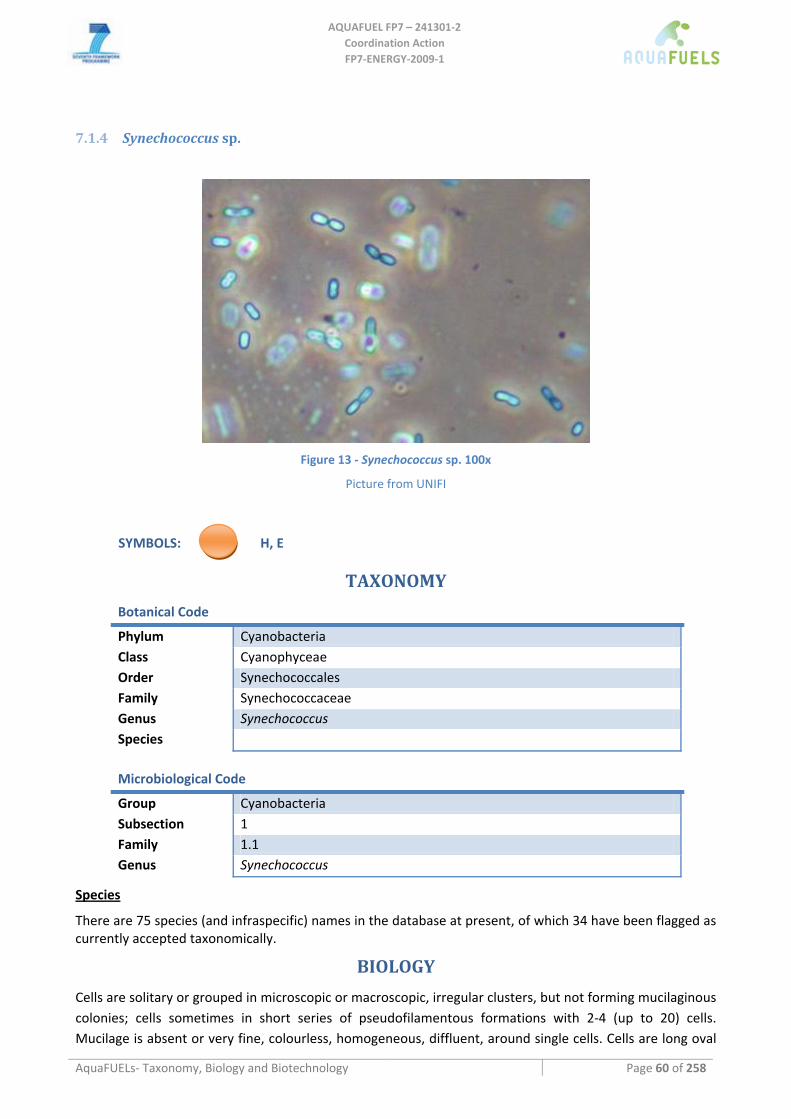
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 60 of 258
7.1.4 Synechococcus sp.
Figure 13 ‐ Synechococcus sp. 100x
Picture from UNIFI
SYMBOLS: H, E
TAXONOMY
Botanical Code
Phylum Cyanobacteria Class Cyanophyceae Order Synechococcales Family Synechococcaceae Genus Synechococcus Species
Microbiological Code
Group Cyanobacteria Subsection 1 Family 1.1 Genus Synechococcus
Species
There are 75 species (and infraspecific) names in the database at present, of which 34 have been flagged as currently accepted taxonomically.
BIOLOGY
Cells are solitary or grouped in microscopic or macroscopic, irregular clusters, but not forming mucilaginous colonies; cells sometimes in short series of pseudofilamentous formations with 2‐4 (up to 20) cells. Mucilage is absent or very fine, colourless, homogeneous, diffluent, around single cells. Cells are long oval

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 61 of 258
or cylindrical, sometimes several times longer than wide, straight, arcuate or sigmoid, 1.5 up to more than 20 μm long and 0.4‐6 (up to 11) μm wide. Thylakoids are parietal. Cells divide by binary fission, transversely (always in one plane in successive generations, perpendicular to the longitudinal axis) into two isomorphic or different (after asymmetric binary fission) daughter cells, which occasionally remain joined for longer period in pairs. Different cyanobacteria commonly designated as Anacystis nidulans and Agmenellum quadruplicatum belong to this genus (=Synechococcus nidulans). The Synechococcus‐group is a provisional assemblage which can be defined as unicellular coccoid to rod‐shaped cyanobacteria that divide by binary fission in a single plane. It is subdivided into several clusters. Most of the Synechococus‐group members are obligate photoautotrophs.
Synechococcus species occur in ecologically extreme conditions (e.g. thermophilic Synechococcus lividus, S. bigranulatus). Several species grow within mats and colonies of other algae, or form fine colonies on wet substrates (mud, wood, stones, etc.) Interesting picoplanctic or planktic species were found in oceans as well as in freshwater reservoirs (e.g., S. rhodobactron).
BIOTECHNOLOGY
Synechococcus has been investigated as a hydrogen producer (Kumazawa and Mitsui, 1994; Lopes‐Pinto et al., 2002). Its main biotechnological interest is its transformability, that allowed insertion of genes from other bacteria, in order to enchance production of naturally occurring molecules such as poly‐β‐hydroxybutyrate (Miyake et al., 2000; Nishioka et al., 2001), interesting for bio‐plastic production, or exogenous molecules such as ethanol (Deng and Coleman, 1999) or isobutyraldehyde (Atsumi et al., 2009), a precursor of butanol ‐a potential fuel ‐, that could be obtained directly from CO2 and sunlight.
References
Atsumi S., Higashide W., Liao J.C. (2009) Direct photosynthetic recycling of carbon dioxide to isobutyraldehyde. Nature Biotechnology 27: 1177‐1182.
Deng M.D., Coleman J.R. (1999) Ethanol synthesis by genetic engineering in cyanobacteria. Applied and Environmental Microbiology 65: 523–528.
Garrity G.M., Boone D.R., Castenholz R.W. (eds) (2004) Bergey’s Manual of Systematic Bacteriology, Second Edition, Volume 1, The Archaea and the deeply branching and phototrophic Bacteria, Springer. [BIOLOGY section]
Herdman M., Castenholz R.W., Iteman I., Waterbury J.B., Rippka R. (2001) The Archaea and the deeply branching and phototrophic bacteria. In: Boone D.R., Castenholz R.W. (eds.) Bergey's Manual of Systematic Bacteriology, 2nd edn. Springer Verlag: Heidelberg. [BIOLOGY section]
Komárek J. (1992) in: http://www.cyanodb.cz/Synechococcus [BIOLOGY section] Komárek J., Anagnostidis K. (1998) Cyanoprokaryota 1. Teil: Chroococcales. ‐ In: Ettl H., Gärtner G., Heynig H., Mollenhauer D. (eds.)
Süsswasserflora von Mitteleuropa 19/1, Gustav Fischer, Jena‐Stuttgart‐Lübeck‐Ulm. [BIOLOGY section] Kumazawa S., Mitsui A. (1994) Efficient hydrogen photoproduction by synchronously grown cells of a marine cyanobacterium,
Synechococcus sp. Miami BG 043511, under high cell density conditions. Biotechnology and Bioengineering 44: 854‐858. Lopes Pinto F.A., Troshina O., Lindblad P. (2002) A brief look at three decades of research on cyanobacterial hydrogen evolution
International Journal of Hydrogen Energy 27: 1209‐1215. Miyake M., Takase K., Narato M., Khatipov E., Schnackenberg J., Shirai M., Kurane R., Asada Y. (2000) Polyhydroxybutyrate
production from carbon dioxide by cyanobacteria. Applied Biochemistry and Biotechnology 84–86: 991‐1002. Nägeli C. (1849) Gattungen einzelliger Algen, physiologisch und systematisch bearbeitet. Neue Denkschriften der Allg.
Schweizerischen Gesellschaft für die Gesammten Naturwissenschaften 10: i‐viii, 1‐139, pls I‐VIII. [BIOLOGY section] Nishioka M., Nakai K., Miyake M., Asada Y., Taya M. (2001) Production of poly‐β‐hydroxybutyrate by thermophilic cyanobacterium,
Synechococcus sp. MA19, under phosphate‐limited conditions. Biotechnology Letters 23: 1095‐1099.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 62 of 258
7.1.5 Synechocystis sp.
Figure 14 ‐ Synechocystis sp. PCC 6308
Picture from www.ibvf.cartuja.csic.es/
SYMBOLS: H, E
TAXONOMY
Botanical Code
Phylum Cyanobacteria Class Cyanophyceae Order Synechococcales Family Merismopediaceae Genus Synechocystis Species
Microbiological Code
Group Cyanobacteria Subsection 1 Family 1.1 Genus Synechocystis
Species
There are 30 species (and infraspecific) names in the database at present, of which 25 have been flagged as currently accepted taxonomically.
BIOLOGY
Cells are solitary, spherical or widely oval, after division hemispherical and for a short time the two cells remain attached together. Cells are without mucilage or with narrow, fine, colourless and usually diffluent and indistinct mucilaginous envelopes. In several strains the cell walls contain a special «S‐layer» with characteristical hexagonal substructure. It never forms colonies. Cell division occurs by binary fission

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 63 of 258
(mainly by pinching) into two morphologically equal daughter cells, which reach the original globular shape before next division; cells always divide in two perpendicular planes in successive generations. If mucilaginous envelopes are formed around the cells, they split together with dividing cells. Synechocystis has been widely investigated from the physiology and genetic point of view and some strains (e.g., Synechocystis PCC 6803) are among the most well‐known cyanobacteria.
Several species are planktonic in freshwater reservoirs or in saline or sea waters, other grow in pools of thermal and mineral springs, of brackish swamps or in moors.
BIOTECHNOLOGY
Its biotechnology is very similar to that of Synechoccoccus. It is a potential producer of polyhydroxyalkanoates (Sudesh et al., 2001; Wu et al., 2001). It has been studied for hydrogen production (Lopes‐Pinto et al., 2002; Angermayr et al., 2009) as well as, being easily transformable, for ethanol production after being genetic engineered (Dexter and Fu, 2009).
References
Angermayr A., Hellingwerf K.J., Lindblad P., Teixeira de Mattos M.J. (2009) Energy biotechnology with cyanobacteria. Current Opinion in Biotechnology 20: 257‐263.
Dexter J., Fu P. (2009) Metabolic engineering of cyanobacteria for ethanol production. Energy and Environmental Science 2: 857‐864.
Garrity G.M., Boone D.R., Castenholz R.W. (eds) (2004) Bergey’s Manual of Systematic Bacteriology, Second Edition, Volume 1, The Archaea and the deeply branching and phototrophic Bacteria, Springer. [BIOLOGY section]
Komárek J. (1992) in: http://www.cyanodb.cz/Synechocystis [BIOLOGY section] Komárek J., Anagnostidis K. (1998) Cyanoprokaryota 1. Teil: Chroococcales. ‐ In: Ettl H., Gärtner G., Heynig H., Mollenhauer D. (eds.)
Süsswasserflora von Mitteleuropa 19/1, Gustav Fischer, Jena‐Stuttgart‐Lübeck‐Ulm. [BIOLOGY section] Lopes Pinto F.A., Troshina O., Lindblad P. (2002) A brief look at three decades of research on cyanobacterial hydrogen evolution
International Journal of Hydrogen Energy 27: 1209‐1215. Sauvageau C. (1892) Sur les algues d'eau douce recoltées en Algérie pendant le session de la societé botanique en 1982. Bulletin de
la Société Botanique de France 34: 104‐128. [BIOLOGY section] Sudesh K., Taguchi K., Doi Y. (2001) Can cyanobacteria be a potential PHA producer? RIKEN Review 42: 75‐76. Wu G.F., Wu Q.Y., Shen Z.Y. (2001) Accumulation of poly‐β‐hydroxybutyrate in cyanobacterium Synechocystis sp. PCC6803.
Bioresource Technology 76: 85‐90.
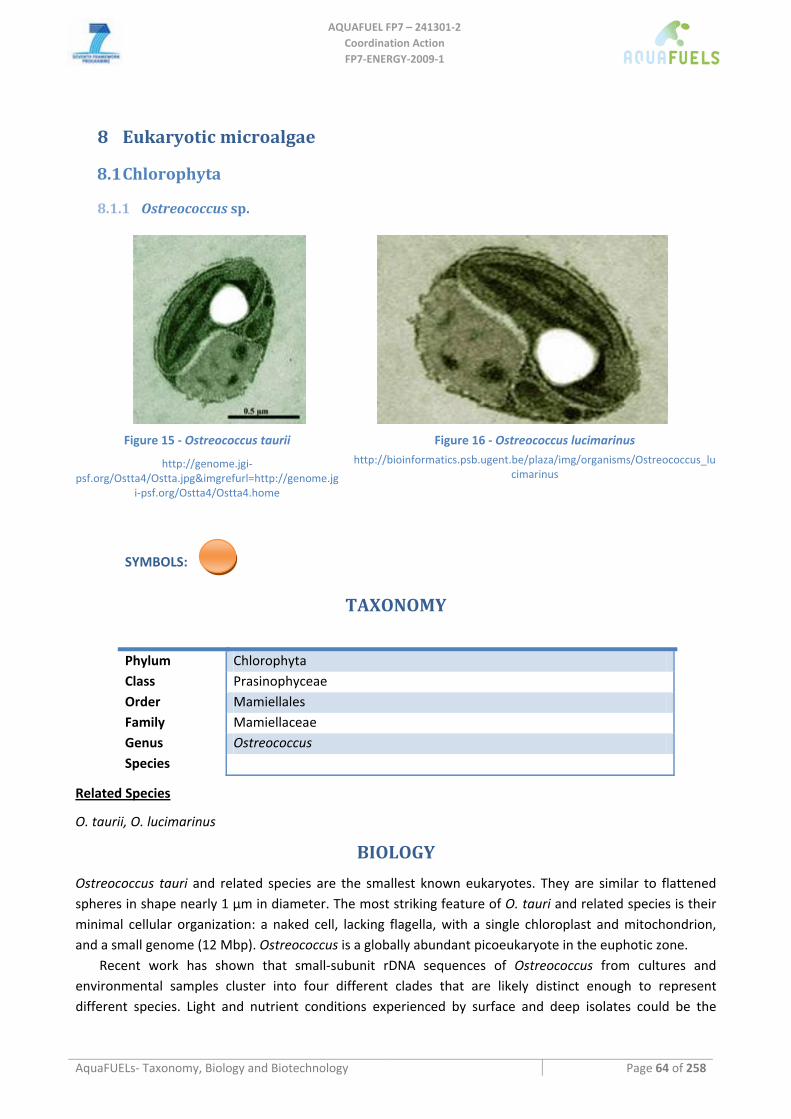
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 64 of 258
8 Eukaryotic microalgae
8.1 Chlorophyta
8.1.1 Ostreococcus sp.
Figure 15 ‐ Ostreococcus taurii
http://genome.jgi‐psf.org/Ostta4/Ostta.jpg&imgrefurl=http://genome.jg
i‐psf.org/Ostta4/Ostta4.home
Figure 16 ‐ Ostreococcus lucimarinus http://bioinformatics.psb.ugent.be/plaza/img/organisms/Ostreococcus_lu
cimarinus
SYMBOLS:
TAXONOMY
Phylum Chlorophyta Class Prasinophyceae Order Mamiellales Family Mamiellaceae Genus Ostreococcus Species
Related Species
O. taurii, O. lucimarinus
BIOLOGY
Ostreococcus tauri and related species are the smallest known eukaryotes. They are similar to flattened spheres in shape nearly 1 μm in diameter. The most striking feature of O. tauri and related species is their minimal cellular organization: a naked cell, lacking flagella, with a single chloroplast and mitochondrion, and a small genome (12 Mbp). Ostreococcus is a globally abundant picoeukaryote in the euphotic zone.
Recent work has shown that small‐subunit rDNA sequences of Ostreococcus from cultures and environmental samples cluster into four different clades that are likely distinct enough to represent different species. Light and nutrient conditions experienced by surface and deep isolates could be the

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 65 of 258
driving factors behind their genetic divergence. Comparative analysis of Ostreococcus sp. will help to understand niche differentiation in unicellular eukaryotes and evolution of genome size in eukaryotes.
BIOTECHNOLOGY
At present, no biotechnological development has been reported for this alga. However, it one of the most studied algae for genetic analysis (Hallmann, 2007) and it is important for the possibilities to have an insight in metabolic and genetic pathways of metabolite production and growth processes.
References
Palenik B., Grimwood J., Aerts A. et al. (2007) The tiny eukaryote Ostreococcus provides genomic insights into the paradox of plankton speciation. Proceedings of the National Academy of Sciences of the USA 104: 7705‐7710. [BIOLOGY section]
Rodríguez F., Derelle E., Guillou L., Le Gall F., Vaulot D., Moreau H. (2005) Ecotype diversity in the marine picoeukaryote Ostreococcus (Chlorophyta, Prasinophyceae). Environmental Microbiology 7: 853‐859. [BIOLOGY section]
Hallmann A. (2007) Algal transgenics and biotechnology. Transgenic Plant Journal 1: 81‐98.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 66 of 258
8.1.2 Tetraselmis sp
Figure 17 ‐ Tetraselmis chuii
CNRS, Station Biologique de Roscoff
http://planktonnet.awi.de
SYMBOLS: D, E, PIV
TAXONOMY
Phylum Chlorophyta Class Prasinophyceae Order Chlorodendrales Family Chlorodendraceae Genus Tetraselmis Species
Related Species
T. alacris, T. apiculata, T. ascus, T. astigmatica, T. chuii, T. convolutae, T. cordiformis, T. desikacharyi, T. gracilis, T. hazeni, T. impellucida, T. inconspicua, T. levis, T. maculata, T. marina, T. micropapillata, T. rubens, T. striata, T. suecica, T. tetrabrachia, T. tetrathele, T. verrucosa, T. wettsteinii.
BIOLOGY
Structural and morphological features
Tetraselmis is a marine green flagellate. The four flagella of this alga, inserted in an anterior depression of the cell, are covered by scales of different types: pentagonal, rod‐shaped and hair scales and, only in some strains, knotted scales (Barsanti and Gualtieri, 2006). Most species of the genus usually are encountered as solitary, free‐swimming, thecate cells. Cell wall or theca is formed by the fusion of scales similar to those found covering the flagella (Nozaki, 2003). Cells are ovoid or ellipsoidal, somewhat compressed equatorially. In the case of Tetraselmis suecica cells have an average size of 10 x 8 µm and a weight of about 200 pg. The chloroplast is single, cup shaped with one pyrenoid (in the species were it is present) and a stigma. The asexual reproduction is by bipartition within the theca, while sexual reproduction is unknown

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 67 of 258
(Nozaki, 2003). It is a very common component in inshore marine environments, tide pools in particular, but there are also seven freshwater species (Nozaki, 2003). Tetraselmis is a very robust microorganism able to resist to extreme pH, salinity and temperature and to adapt to rapid changes in environmental conditions. This feature makes it particularly suitable for outdoor mass cultivation. Among the 50 species known, the most widely used are the marine T. suecica, T. chui and T. tetrathele.
Gross composition under optimal and stressed conditions
Tetraselmis suecica has a high protein content (up to 40‐50%). Carbohydrate is about 20% and lipid about 20% of the cell dry weight. Under nutrient stress (nitrogen or phosphorus deprivation) Tetraselmis suecica accumulates carbohydrates.
Renaud et al. (1999) report for Tetraselmis sp. a content of 26‐30% protein, 13‐14% lipid, 8‐9% carbohydrate and 14‐17% ash as well as about 60% of polyunsaturated fatty acids over the total fatty acid content.
BIOTECHNOLOGY Tetraselmis offers a valuable source of protein, bioactive compounds, antioxidants, vitamins, sterols and polyunsaturated fatty acids for human and animal consumption. One the most important applications of Tetraselmis is in aquaculture for rearing zooplankton and larval stages of marine fish, bivalve molluscs and crustaceans (Muller‐Feuga et al., 2003; Tredici et al., 2009). The genus has been found to have antibacterial activity towards important aquaculture pathogens (Austin et al., 1992) and it was also proposed as probiotic (Tredici et al., 2009, Irianto and Austin, 2002). Due to its high content of good quality protein (40‐50% d.wt), Tetraselmis suecica biomass could represent an alternative ingredient for animal feed. Because of its high content of vitamin E (0.13‐0.25 g kg‐1), Tetraselmis has also been proposed as a source of this vitamin as preservative in foods, additive in animal feed and sunscreen in cosmetics (Carballo‐Cárdenas et al., 2003). Active ingredients extracted from this microalga are currently used in the development of novel cosmetic formulations influencing growth of human hair and/or pigmentation of human skin (Pertile et al., 2010). An emerging use of Tetraselmis is for carbon biofixation in combination with biofuels production (biodiesel, bioethanol) (Tredici, 2010).
Culture Media
F medium (Guillard and Ryther, 1962) is widely used for cultivation of Tetraselmis. The microalga can be grown also in natural seawater integrated with nutrients.
Cultivation methods
In general Tetraselmis is cultivated under autotrophic conditions. Some species have also heterotrophic or mixotrophic capacity. Intensive cultivation of Tetraselmis has been carried out in open “raceway” ponds and in different kinds of closed photobioreactors (PBR). In Hawaii in 24‐m2 flumes Laws and Berning (1991) obtained a productivity of 15‐20 g C m‐2 day‐1 with photosynthetic efficiencies of 9‐10% with Tetraselmis suecica. In pilot‐scale open ponds in Southern Italy, Tetraselmis tetrathele has reached productivities of about 30 g m‐2 day‐1 during the summer, with photosynthetic efficiencies in the PAR region around 5% (Materassi et al., 1983). A parallel cultivation of Tetraselmis suecica in pilot‐scale open ponds and near‐horizontal tubular reactors has shown similar productivities for the two systems, about 26 g m‐2 day‐1 (Pedroni et al., 2004). Current methods for its culture rely on batch, semi‐continuous or continuous cultivation. At present, culture methods used in hatcheries for Tetraselmis production use mainly polyethylene bags and transparent glass‐fibre cylinders (up to 500 L) usually kept indoors with artificial light (Fulks and Main, 1991). Advances have recently been made in the field of photobioreactor technology

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 68 of 258
(Tredici et al., 2010) that improved microalgae productivity in comparison to the traditional culture systems. Among these systems annular columns (Chini Zittelli et al., 2006) and disposable flat panels (Tredici et al., 2010) have been and are currently used in laboratory and at pilot‐scale outdoors. During the summer in Central Italy in an experiment reproducing a full scale plant arrangement, in 120‐L annular columns at a daily dilution rate of 40%, Tetraselmis suecica attained an average volumetric productivity of 0.46 g l‐1 day‐1, and an overall areal productivity of 36.3 g m‐2 day‐1 with a photosynthetic efficiency in the PAR region of 9.4% (Chini Zittelli et al., 2006). Tetraselmis has been also grown at industrial scale under heterotrophic conditions in fermenters with yields in excess of 100 g L‐1 day‐1 (Day et al., 1991).
Harvesting methods
Tetraselmis settles spontaneously and can be harvested in funnels, though centrifugation is a faster and a more efficient harvesting method.
Upscaling limitations
Although Tetraselmis is a very robust microorganism able to resist to extreme pH, salinity and temperature and to adapt to the rapid changes in environmental conditions, to obtain high productivities high amounts of energy are generally required for culture mixing due to the high sedimentation rate of the microalga.
HIGHLIGHTS IN BIOTECHNOLOGY
Major characteristics of this genus are robustness and high productivity in outdoor mass cultures
References
Austin B., Bauder E., Stobie M.B.C. (1992) Inhibition of bacterial fish pathogens by Tetraselmsis suecica. Journal of Fish Diseases 15: 55‐61.
Barsanti L., Gualtieri P. (2006) Algae. Anatomy, Biochemistry, and Biotechnology. Taylor & Francis, Boca Raton. Carballo‐Cárdenas E.C., Tuan P.M., Janssen M., Wijffels R.H. (2003) Vitamin E (α‐tocopherol) production by marine microalgae
Dunaliella tertiolecta and Tetraselmis suecica in batch cultivation. Biomolecular Engineering 20: 139‐147. Chini Zittelli G., Rodolfi L., Biondi N., Tredici M.R. (2006) Productivity and photosynthetic efficiency of outdoor cultures of
Tetraselmis suecica in annular columns. Aquaculture 261: 932–943. Day J.G., Edwards A.P., Rodgers G.A. (1991) Development of an industrial‐scale process for the heterotrophic production of
microalgal mollusc feed. Bioresource Technology 38: 245‐249. Fulks W., Main K.L. (eds.) (1991) Rotifer and Microalgae Culture Systems. Proceedings of a U.S.‐Asia Workshop. The Oceanic
Institute, Honolulu, Hawaii, USA. Guillard R.R.L., Ryther J.H. (1962). Studies of marine planktonic diatoms. I. Cyclotella nana (Hustedt) and Detonula confervacea
(Cleve). Canadian Journal of Microbiology 8: 229‐239. Irianto A., Austin B. (2002) Probiotics in aquaculture Journal of Fish Diseases 25: 633‐642. Laws E.A., Berning J.L. (1991) A Study of the energetics and economics of microalgal mass culture with the marine chlorophyte
Tetraselmis suecica: Implications for use of power plant stack gases. Biotechnology and Bioengineering 37: 936‐947. Materassi R., Tredici M.R., Milicia F., Sili C., Pelosi E., Vincenzini M., Torzillo G., Balloni W., Florenzano G., Wagener K. (1983)
Development of a production size system for the mass culture of matine microalgae. In: Palz W., Pirrwitz D. (eds) Energy from biomass. Riedel Publishing Company, Boston, Series E, vol. 5, pp. 150‐158.
Muller‐Feuga A., Robert R., Cahu C., Robin J., Divanach P. (2003) Uses of microalgae in aquaculture. In: Stottrup, J.G., McEvoy, L.A. (Eds.), Live Feeds in Marine Aquaculture. Blackwell, Oxford, pp. 253–299.
Nozaki H. (2003) Flagellated green algae. In: Wehr J.D., Sheath R.G. (eds.) Freshwater algae of North America. Ecology and classification. Elsevier, Amsterdam, pp. 225‐252.
Pedroni P.M., Lamenti G., Prosperi G., Ritorto L., Scolla G., Capuano F., Valdiserri M. (2004) Enitecnologie R & D project on microalgae biofixation of CO2: outdoor comparative tests of biomass productivity using flue gas CO2 from a NGCC power plant. Proceedings of Seventh International Conference on Greenhouse Gas Control Technologies (GHGT‐7), 5–9 September 2004, Vancouver, Canada.
Pertile P., Zanella L., Hermmann M., Holger J., Gaebler S. (2010) Extracts of Tetraselmis sp. US patent 2010/0143267. Renaud S.M., Thinh L.‐V., Parry D.L. (1999) The gross chemical composition and fatty acid composition of 18 species of tropical
Australian microalgae for possible use in mariculture. Aquaculture 170: 147‐159.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 69 of 258
Throndsen, J. (1996) The planktonic marine flagellates. In: Tomas, C.R. (ed) Identifying marine phytoplankton. Academic Press, San Diego, pp. 591‐730
Tredici M.R. (2010) Photobiology of microalgae mass cultures: understanding the tools for the next green revolution. Biofuels 1, 143‐162.
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G., (eds.) New Technologies in Aquaculture: Improving production efficiency, quality and environmental management. Woodhead Publishing Ltd, Cambridge, UK, and CRC Press LLC, Boca Raton, FL, USA, pp. 610‐676.
Tredici M.R., Chini Zittelli G., Rodolfi L. (2010) Photobioreactors In: Flickinger M.C., Anderson S. (eds) Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology. John Wiley & Sons, Inc., Hoboken, NJ, USA. Vol 6, pp.3821–3838.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 70 of 258
8.1.3 Botryococcus braunii
Figure 18 ‐ Botryococcus braunii
http://erenovable.com/wp‐content/uploads/2007/02/BotryococcusbrauniiAlga.jpg
SYMBOLS: D
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Chlorococcales Family Dictyosphaeriaceae West 1916, Botryococcaceae (Wille 1909) Genus Botryococcus Species Botryococcus braunii
Related Species
There are 15 species (and infraspecific) names in the database at present, of which 12 have been flagged as currently accepted taxonomically.
B. australis, B. balkachicus, B. braunii, B. calcareous, B. canadensis, B. comperei, B. coorongianus, B. fernandoi, B. giganteus, B. neglectus, B. pila, B. protuberans, B. sudeticus, B. terribilis, B. terricola.
BIOLOGY
Botryococcus braunii is a green colonial microalga. Colonies, under microscope observation, exhibit a typical morphology characterised by a botryoid organisation of individual pyriform‐shaped cells held together by a refringent matrix containing lipids. Oil droplets can be excreted from the matrix by the pressure of a coverglass. Ultrastructural studies reveal that the matrix surrounding the basal part of the cells consists of outer walls originating from successive cellular divisions. Furthermore, the bulk of B. braunii hydrocarbons are stored in these outer walls Colonies frequently are compounded by interconnecting strands of tough mucilage between clusters of cells. Cells have a cup‐shaped plastid with a naked pyrenoid‐like body. Found in phytoplankton and metaphyton of ponds and lakes. Geographically

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 71 of 258
widespread, it is rarely abundant, though it has the ability to form blooms, sometimes enduring over many years, like in the Darwin Reservoir in Australia.
Botryococcus braunii produces various types of ether lipids and hydrocarbons up to 60% of biomass. According to hydrocarbon production, B. braunii is subclassified into three chemical races. Algae in race A produce essentially n‐alkadiene and triene hydrocarbons, odd‐carbon‐numbered from C23 to, algae in race B produce triterpenoid hydrocarbons, C30–C37 botryococcenes and C31–C34 methylated squalenes and algae in race L produce a single tetraterpenoid hydrocarbon, lycopadiene. 18S rRNA sequences of four strains of B. braunii belonging to the three chemical races established that these strains formed a monophyletic group.
BIOTECHNOLOGY
Most studies concerning the influence of various factors on the production of biomass and hydrocarbons were carried out in the laboratory. In the three chemical races of B. braunii, the hydrocarbon productivity, as well as the ether lipid synthesis, was shown to be optimal during the exponential phase of growth and does not occur in nitrogen‐ and phosphorous deficient media (Metzger and Largeau, 2005).
Entrapment of B. braunii colonies in calcium alginate beads exhibits some interesting advantages by comparison to free suspension cultures: enhancement in chlorophyll photosynthetic activity, protection against photoinhibition towards high irradiance and an increase in hydrocarbon production despite a decrease in the rate of biomass production; however, the lack of stability of calcium alginate beads over a long period is not favourable to their use in cultures on a large scale (Metzger and Largeau, 2005).
There are no reports on successful large‐scale cultivation. Experimental cultures under natural illumination in tubular photobioreactors, immersed in water for cooling, have been conducted up to a volume of 200 L (Gudin and Chaumont, 1984). In 1000‐L raceway ponds, average biomass productivity of 89 mg L‐1 day‐1 were achieved, with a lower hydrocarbon content compared to indoor cultures, with B. mahabali (Dayananda et al., 2010). Outdoor cultures are easily contaminated by other algae (Metzger and Largeau, 2005). Small scale commercial production of B. braunii is carried out in Portugal by A4F Algafuel S.A.
Botryococcus is able to grow on different types of effluents under laboratory conditions (Metzger and Largeau, 2005; Shen et al., 2008), although very slowly: e.g., on secondary treated domestic sewage its productivity was of about 30 mg L‐1 day‐1 (Sawayama et al., 1994; Sydney et al., 2011).
References
Dayananda C., Kumudha A., Sarada R., Ravishankar G.A. (2010) Isolation, characterization and outdoor cultivation of green microalgae Botryococcus sp. Scientific Research and Essays 5: 2497‐2505.
Gudin C., Chaumont D. (1984) Solar biotechnology study and development of tubular solar receptors for controlled production of photosynthetic cellular biomass for methane production and specific exocellular biomass. In: Palz W., Pirrwitz D. (eds.) Energy from biomass, serie E, vol 5. Reidel, Dordrecht, pp 184‐193.
Metzger P., Largeau C. (2005) Botryococcus braunii: a rich source for hydrocarbons and related ether lipids. Applied Microbiology and Biotechnology 66: 486‐496. [Also for BIOLOGY section]
Sawayama S., Inoue S., Yokoyama S. (1994) Continuous culture of hydrocarbon‐rich microalga Botryococcus braunii in secondarily treated sewage. Applied Microbiology and Biotechnology 41: 729‐731.
Shen Y., Yuan W., Pei Z., Mao E. (2008) Culture of microalga Botryococcus in livestock wastewater. Transactions of the ASABE (American Society of Agricultural and Biological Engineers) 51: 1395‐1400.
Sydney E.B., da Silva T.E., Tokarski A., Novak A.C., de Carvalho J.C., Woiciecohwski A.L., Larroche C., Soccol C.R. (2011) Screening of microalgae with potential for biodiesel production and nutrient removal from treated domestic sewage. Applied Energy: doi:10.1016/j.apenergy.2010.11.024.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 72 of 258
8.1.4 Chlamydomonas reinhardtii
Figure 19‐ Chlamydomonas reinhardtii
http://sandwalk.blogspot.com/2007/10/genome‐of‐chlamydomonas‐reinhardtii.html
SYMBOLS: H, PIV
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Volvocales Family Chlamydomonadaceae Genus Chlamydomonas Species Chlamydomonas reinhardtii
Related Species
NCBI Nucleotide Sequences As of 2 June 2009, nucleotide sequence data are available at http://www.ncbi.nlm.nih.gov/Genbank for 265,283 samples identified as Chlamydomonas.
Taxonomic identifications of these samples to genus (or species) levels unaccompanied by explicit indication of voucher specimens may not be verifiable on morphological/anatomical grounds and consequently be of little or no taxonomic value. Numbers of names and species: there are 1163 species (and infraspecific) names in the database at present, of which 382 have been flagged as currently accepted taxonomically.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 73 of 258
BIOLOGY
Structural and morphological features Chlamydomonas reinhardtii is a unicellular green alga about 10‐ 20 μm in diameter with two flagella. The cell wall is made of hydroxyproline‐rich glycoproteins. C. reinhardtii has a single, large, cup‐shaped chloroplast, a large pyrenoid, and a stigma ("eyespot") that senses light.
C. reinhardtii is primarily used as a model organism in biology. C. reinhardtii can grow photoautotrophicallyt, or heterotrophically in the dark when supplied with acetate. This has been exploited for creation of thousands of mutants affected in the photosynthetic process (www.chlamy.org).
Due to its sexual and vegetative growth, heterotroph and photoautotroph growth cycles, genetic mapping of photosynthetic markers became possible early on and made Chlamydomonas a prime tool for studying the genetics of photosynthesis, among others revealing maternal inheritance of chloroplast encoded genes (Harris, 1989).
The alga as such is not a prime target for biofuels production. However, access to the numerous mutants and genetic transformation technologies have made this alga a prime target for biofuels research (mainly for biohydrogen).
Vegetative cells of the reinhardtii sspecies are haploid and have 17 small chromosomes. Haploid gametes develop under nitrogen starvation. Two mating types mt(+) and mt(‐) then fuse to form a diploid zygote. The zygote undergoes meiosis and releases four flagellated haploid cells that resume the vegetative life cycle.
Under ideal growth conditions, cells may sometimes undergo two or three rounds of mitosis before the daughter cells are released from the old cell wall into the medium. Thus, a single growth step may result in 4 to 16 daughter cells per mother cell. In presence of acetate the cells grow significantly faster than with light alone. The cell cycle of this unicellular green algae can be synchronized by alternating periods of light and dark, a process widely applied to study regulation of gene expression during the diurnal cycle (e. g., Leu et al., 1990; Breidenbach et al., 1990).
The attractiveness of the alga as a model organism has recently increased with the release of several genomic resources (see Chlamydomonas Center, http://www.chlamy.org/), which also administrates thousands of mutant strains, plasmids and other resources for genetic and molecular research. In addition to genomic sequence data there is a large supply of expression sequence data available as cDNA libraries and expressed sequence tags (ESTs). cDNA libraries are available onlineand a BAC library can be purchased from the Clemson University Genomics Institute. There are also two databases of >50 000 and >160 000 ESTs available online.
Gross composition under optimal and stressed conditions
The variability of algal biochemical composition has been impressively demonstrated in C. reinhardtii during its diurnal growth cycle. During the early light period most cellular mass is protein, during the second half of the light period the alga accumulates up to 50% starch. While oil droplets in Chlamydomonas were not originally reported, recent studies revealed up to 20 % TAG under nitrogen starvation.
BIOTECHNOLOGY Clamydomonas is not being investigated for mass cultivation and biomass production, though Hu et al. (2008) have included two Chlamydomonas species in their comparative study on lipid productivity with C. applanta showing higher productivity than C. reinhardtii. However, the wide knowledge on its genetics and cell biology has allowed revealing numerous mechanisms of interest to biofuels production.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 74 of 258
Analysis of starch deficient mutants revealed the tight link between carbon allocation to starch or lipid biosynthesis. Identification of mirna in Chlamydomonas has revealed their usefulness in upregulation of lipid biosynthesis (Maor, personal communication).
Clamydomonas has the potential to produce large amounts of hydrogen because it can directly split water into hydrogen and oxygen using the enzyme hydrogenase. It was discovered that when the alga is deprived of essential sulfate salts, it no longer maintains the protein complex necessary for photosynthetic production of oxygen and instead switches to the hydrogen‐producing metabolic pathway. However, the alga can not grow in a sulfur deprived condition for long time before it needs to revert to the oxygen producing mode (Torzillo et al., 2009; Ghirardi et al., 2010). Detailed understanding of the hydrogen production process and the hydrogenase complex of Chlamydomonas have allowed creating mutants with enhanced hydrogen production capacity.
Significant efforts have been undertaken to investigate expression of recombinant proteins in Chlamydomonas reinhardtii (Mayfield et al., 2003). Among the proteins expressesd successfully are antibody molecules, bovine mammary‐associated serum amyloid that may be expressed successfully both in the cytoplasm or chloroplast. (e.g. Mayfield et al., 2007).
Culture media
C. reinhardiii can be cultivated in a large variety of synthetic growth media such as sager‐granick, high salt (sueoka), tap, in presence or absence of acetate (Harris, 1989, p 31 on). Presence of acetate significantly enhances the growth rate under diurnal light regime.
Production system
Possible biohydrogen production has been investigated and modelled in a study to nrel NREL (Wade and Amos, 2004), though no actual field tests and up scaling have been performed as actual studies have been performed on the lab scale so far.
For the production of recombinant proteins rather small well controlled photobioreactors are suggested rather than large outdoors cultivation systems.
HIGHLIGHTS IN BIOTECHNOLOGY
• Early genetic work, genetic transformation and fully sequenced genome make this alga the favorite for exploring the power of genetic engineering in algal biotechnology (Harris, 1989).
• Major breaktroughs are algae with increased hydrogen production, reduced chlorophyll etc. Mutants with increased oil content due to impaired starch synthesis (Li et al., 2010), and application of mirna miRNA technology for enhanced oil production (Maor, personal communication).
• Furthermore the algae has been used to express and produce recombinant medically relevant peptides and proteins (Mayfield et al., 2003, 2007).
References Breidenbach E., Leu S., Michaels A., Boschetti A. (1990) Synthesis of EF‐Tu and distribution of tis mRNA between stroma and
thylakoids during the cell cycle of Chlamydomonas reinhardii. Biochimica et Biophysica Acta 1048: 209‐216. Ghirardi M.L., Kosourov S., Maness P., Smolinski S., Seibert M. (2010) Algal Hydrogen production. In: Flickinger M.C., Anderson S.
(eds.) Encyclopedia of Industrial Biotechnology: Bioprocess, Bioseparation, and Cell Technology. John Wiley & Sons, Inc., Hoboken, NJ, USA. Vol.1, pp. 184‐198.
Harris E. (1989) The Chlamydomonas Sourcebook: A Comprehensive Guide to Biology and Laboratory Use.Academic Press, San Diego.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 75 of 258
Hu Q., Sommerfeld M., Jarvis E., Ghirardi M., Posewitz M., Seibert M., Darzins A. (2008) Microalgal triacylglycerols as feedstocks for biofuels production: perspectives and advances. The Plant Journal 54: 621‐639.
Leu S., White D., Michaels A. (1990) Cell cycle‐dependent transcriptional and post‐transcriptional regulation of chloroplast gene expression inChlamydomonas reinhardtii. Biochimica et Biophysica Acta 1049: 311‐317.
Li Y., Han D., Hu G., Dauvillee D., Sommerfeld M., Ball S., Hu Q. (2010) Chlamydomonas starchless mutant defective in ADP‐glucose pyrophosphorylase hyper‐accumulates triacylglycerol. Metabolic Engineering 12: 387‐391.
Mayfield S.P., Manuell A.L., Chen S., Wu J., Tran M., Siefker D., Muto M., Marin‐Navarro J. (2007) Chlamydomonas reinhardtii chloroplasts as protein factories. Current Opinion in Biotechnology 18: 126‐133.
Mayfield S.P., Franklin S.E., Lerner R.A. (2003) Expression and assembly of a fully active antibody in algae. Proceedings of the National Academy of Science of the USA 100: 438‐442.
Molnar A., Bassett A., Thuenemann E., Schwach F., Karkare S., Ossowski S., Weigel D., Baulcombe D. (2009) Highly specific gene silencing by artificial microRNAs in the unicellular alga Chlamydomonas reinhardtii. The Plant Journal 58: 165‐174.
Torzillo G., Scoma A., Faraloni C., Ena A., Johanningmeier U. (2009) Increased hydrogen photoproduction by means of a sulfur deprived Chlamydomonas reinhardtii D1 protein mutant. International Journal of Hydrogen Energy 34: 4529‐4536.
Wade A., Amos A. (2004) Updated cost analysis of photobiological hydrogen production from Chlamydomonas reinhardtii green algae. Milestone completion report, NREL/mp‐560‐35593.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 76 of 258
8.1.5 Haematococcus pluvialis
Figure 20 ‐ Green and astaxanthin containing H. pluvialis cells (light microscope).
D. Reinecke, BGU
SYMBOLS: B, PIV
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Volvocales Family Haematococcaceae Genus Haematococcus Species Haematococcus pluvialis
Related Species
There are 15 species (and infraspecific) names in the database at present, of which 7 have been flagged as currently accepted taxonomically.
H. allmanii, H. buetschlii, H. capensis, H. carocellus, H. droebakensis var. fastigatus, H. droebakensis, H. grevillei, H. insignis, H. lacustris, H. murorum, H. pluvialis, H. salinus, H. sanguineus, H. thermalis, H. zimbabwiensis.
BIOLOGY
Structural and morphological features
Haematococcus pluvialis is a medium to large unicellular green algae (10 – 100 μm) with a large single cloroplast, two flagellae. Young cells are motile, ageing cells (palmelloids) lose their mobility. Under stress the algae accumulates the carotenoid asthaxanthin in cytoplasmic oil globules and enters a resting stage (Droop, 1954). The cell wall thickens and alters its chemical composition during maturation.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 77 of 258
Biochemical composition of algae and aquatic biomass main constituents
During steady state growth the algae is rich in protein, with less than 10% lipid content, displaying the typical green algal fatty acid composition with significant amounts of short (C16, C18) polyunsaturated fatty
acids mainly in the chloroplast lipids, chlorophylls a and b and photosynthetic carotenoids such as β‐carotene.
Under stress the alga accumulates initially up to 50% starch, then initiates synthesis of triacylglycerol (TAG) accumulating to 40% of cell weight in cytoplasmic oil globules, and up to 4% of the ketocarotenoid astaxanthin (Boussiba et al., 1992, 1999).
Growth kinetics and efficiencies
Haematococcus is considered a slow growing alga. Productivity in outdoors systems has been reported by Huntley and Redalje (2007), implying a photosynthetic energy conversion efficiency of 3%.
BIOTECHNOLOGY
Culture Media
BG11 (Rippka et al., 1979), mBG11 or similar synthetic growth media.
Cultivation methods
Haematococcus is cultivated both in open ponds or closed bioreactors. The tubular photobioreactor facility at Qetura is the largest operating photobioreactor facility for microalgae production. Large scale outdoors cultivation in two stage mode, photobioreactor for green cells and open ponds for production of red cells was tested in Hawaii and yielded a long term growth average of 38 tons per hectare and year with 25% oil content, or 422 GJ ha‐1 year‐1 (Huntley and Redalje, 2007).
Production system
Production is managed as semicontinuous or batch cultivation in two stages. The first stage is a nutrient sufficient green stage that can be handled as continuous or semicontinuous culture (Boussiba et al., 1997; Huntley and Redalje, 2007), while the second stage under nutrient limitation for accumulation of astaxanthin is necessarily a batch cultivation where all the resulting biomass is harvested. One step cultivation has been proposed but not commercially deployed (Garcia‐Malea et al., 1999).
Harvesting methods
Haematococcus, specifically stressed resting cells settle spontaneously and can easily be harvested in large funnels or sedimentation ponds. However centrifugation yields higher recovery of biomass faster.
Biomass processing
The biomass is dried,dried; cells are broken by bead mills or other suitable technologies. Astaxanthin containing oil is being extracted, e. g. by supercritical CO2 extraction, and sold.
Upscaling limitations
Contaminations, competitors, cold or heat stress can significantly reduce productivity outdoors, and in bioreactors heating or cooling are required for maintaining satisfactory productivities. Being a freshwater microalgae, Haematococcus cultivation is frequently hampered by other, fast growing microalgae such as

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 78 of 258
Scenedesmus or Chlorella, zooplankton or even funghi which can drastically reduce the culture performance.
HIGHLIGHTS IN BIOTECHNOLOGY
Astaxanthin production at the tubular bioreactor facility at Algatech Qetura (http://www.algatech.com/, Boussiba et al., 1997) may be considered the most advanced microalgal production process having reached full commercialization. This production facility has been running successfully for eight years uninterrupted under continuous incorporation of innovations for increasing productivity and product quality.
References
Boussiba S., Bing W., Yuan J.P., Zarka A., Chen F. (1999) Changes in pigments profile in the green alga Haematococcus pluvialis exposed to environmental stresses. Biotechnology Letters 21: 601‐604.
Boussiba S., Fan L., Vonshak A. (1992) Enhancement and determination of astaxanthin accumulation in green alga Haematococcus pluvialis. Methods in Enzymology. New York: Academic Press, pp. 386‐371.
Boussiba S., Vonshak A., Cohen Z., Richmond A. Ben‐Gurion University of the Negev, Israel (1997). A procedure for large‐scale production of astaxanthin from Haematococcus WO 97/28274.
Droop M. (1956a) Haematococcus pluvialis and its allies. I. The Sphaerellaceae. Revue Algologique 2: 53‐70. Droop M. (1956b) Haematococcus pluvialis and its allies. II. Nomenclature in Haematococcus. Revue Algologique 3: 182‐192. Elliott A. (1934) Morphology and life history of Haematococcus pluvialis. Archiv für Protistenkunde 82L.: 250‐272. Garcia‐Malea M.C., Acien F.G., Del Rio E., Fernandez J.M., Ceron M.C., Guerrero M.G., Molina‐Grima E. (2009) Production of
astaxanthin by Haematococcus pluvialis: Taking the onestep system outdoors. Biotechnology and Bioengineering 102: 651‐657.
Hepperle D., Nozaki H., Hohenberger S., Huss V.A., Morita E., Krienitz L.. (1998) Phylogenetic position of the Phacotaceae within the Chlamydophyceaeas revealed by analysis of 18S rDNA and rbcL sequences. Journal of Molecular Evolution 47: 420‐30.
Huntley M.E., Redalje D.J. (2007) CO2 mitigation and renewable oil from photsynthetic microbes: A new appraisal. Mitigation and Adaptation Strategies for Global Change 12: 573‐608.
Kobayashi M., Kurimura Y., Kakizono .T, Nishio N., Tsuji Y. (1997) Morphological changes in the life cycle of the green alga Haematococcus pluvialis. Journal of Fermentation and Bioengineering 84: 94‐97.
Rippka R., Deruelles J., Waterbury J.B., Herdman M., Stanier R.Y. (1979). Generic assignment, strains histories and properties of pure cultures of cyanobacteria. Journal of General Microbiology 111: 1‐61.
Triki A., Maillard P., Gudin C. (1997) Gametogenesis in Haematococcus pluvialis Flotow (Volvocales, Chlorophyta). Phycologia 36: 190‐194.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 79 of 258
8.1.6 Dunaliella sp.
Figure 21 – Dunaliella sp. © D.J. Patterson
http://starcentral.mbl.edu/msr/rawdata/viewable/dunaliella_1097788928_esmlaw.jpg; Left:
Figure 22 ‐ Dunaliella tertiolecta Roscoff Culture Collection Strain # 6
CNRS Station Biologique de Roscoff www.sb‐roscoff.fr/Phyto/gallery/main.php?g2_itemId=371
SYMBOLS: B, PIV
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Volvocales Family Dunaliellaceae Genus Dunaliella Species
Related Species
There are 28 species (and infraspecific) names in the database at present, of which 23 have been flagged as currently accepted taxonomically.
D. acidophila, D. assymetica, D. baasbeckingii, D. bardawil, D. bioculata, D. carpatica, D. cordata, D. euchlora, D. gracilis, D. granulata, D. lateralis, D. maritima, D. media, D. minuta, D. parva, D. peircei, D. polymorpha, D. primolecta, D. pseudo‐salina, D. quartolecta, D. ruineniana, D. salina, D. terricola, D. tertiolecta, D. turcomanica, D. viridis var. palmelloides, D. viridis, D. viridis f. euchlora.
BIOLOGY Structural and morphological features
Dunaliella is a genus of algae, specifically belonging to the Dunaliellaceae family. Dunaliella sp. are motile, unicellular, rod to ovoid shaped (9 ‐ 11 µm) green algae, Chlorophyta, which are common in marine waters.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 80 of 258
The genus Dunaliella has marine and halophilic representatives. Freshwater species have also been described. Dunaliella also has a very wide pH tolerance ranging from pH 1 (D. acidophila) (Gimmler et al., 1989) to pH 11 (D. salina). In fact, D. salina is one of the most environmentally tolerant eukaryotic organisms known and can cope with a salinity range from seawater (= 3% NaCl) to NaCl saturation (= 31% NaCl), and a temperature range from <0 °C to >38 °C (Ginzburg, 1987; Borowitzka, 1988).
Dunaliella is morphologically similar to Chlamydomonas, with the main difference being the absence of a cell wall in Dunaliella. It has two flagella of equal length and a single, cup‐shaped chloroplast, which in the marine and halophilic species has a central pyrenoid (Borowitzka, 1988). D. salina is a halophile microalga and was originally identified in sea salt fields. It is different from all other green algae cells because it lacks a cell wall and is wrapped with an extremely thin elastic membrane. The lack of cell wall allows the cell to change its volume with changing osmotic pressure. In the Dead Sea it was first discovered in 1941. Another strain of interest to biotechnology is Dunaliella tertiolecta. D. tertiolecta is a marine microalga with a cell size of 10 – 12 µm. D. tertiolecta is a fast growing strain in sea water, reported to have oil yield of about 37%, though comparative growth rates and lipid productivities have not been reported.
Biochemical composition
In order to survive under such extreme conditions, Dunaliella synthesizes different compounds in a very high concentration. Its resistance against high salt concentrations (35%) is linked to its ability to synthesize and accumulate glycerol up to 10% of its dry weight. Due to this it can maintain shape and functions under high osmotic pressures. In order to grow in an environment of high temperature and intense sunlight, Dunaliella synthesizes beta‐carotene up to 6 of its dry weight. Studies show that the function of beta‐carotene is to protect the algae cell from damages caused by intense radiation.
Proteins accumulate up to 60% of the dry cell weight and contain most amino acids. Proteins and amino acids are important ingredients in cosmetic preparations. They are used in order to create a contaminating layer on the skin surface while nourishing the skin cells.
The carbohydrates include mono‐sugars (glucose, glactose, mannose, xylose, ribose, rhamnose), di‐sugars and 1,4 polysaccharides ‐ glucosen and starch. The sugars, especially the poly‐sugars are used as stabilizers. They thicken and give the product a smooth and gentle texture. They absorb large quantities of water and grant the product with moisturizing traits. Attached to the negative electrical charge of the poly‐sugars are electrolytes, released in a controlled way to the skin cells. The presence of polysaccharides in the cosmetic product allows this controlled release of the active substances in the product and offers an efficient treatment of skin diseases without using substances which risk the user in side‐effects.
Lipids accumulate to 6‐18% of the dry cell weight depending on growth conditions. Fatty acids include palmitic acid, 3 – trans acid hexadecanoic, linoleic acid and arachidic acid. Beta‐carotene can accumulate up to 6% of the cell's dry weight. Glycerol assembles up to 10% of the dry cell weight. Glycerol includes monogalacto glycerol, digalacto glycerol and diacyl glycerol.
In addition to high level of beta‐carotene, Dunaliella contains thiamine, pyridoxine, riboflavin, nicotinic acid, biotin and tocopherol (vitamin E). β ‐carotene, produced from Dunaliella is composed of two isomers: all trans and cis‐9, in contrast to beta‐carotene produced from carrots or synthetic beta‐carotene. β‐carotene seems to act as photo‐protective ‘sun‐screen’ to protect the chlorophyll and the cell DNA from the high irradiance which characterizes the normal habitat of D. salina. It has also been proposed that β‐carotene also acts as a “carbon sink” to store the excess carbon produced during photosynthesis under conditions where growth is limited but photosynthetic carbon fixation must continue.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 81 of 258
BIOTECHNOLOGY Since establishment of a first pilot plant for Dunaliella cultivation for β‐carotene production in the USSR in 1966, the commercial cultivation of Dunaliella for the production of β‐carotene throughout the world is now one of the success stories of halophile biotechnology (Ben Amotz and Avron, 1982).
Although technically the production of glycerol from Dunaliella was shown to be possible, economic feasibility is low and no biotechnological operation presently exists. β ‐carotenoid production by Dunaliella is also not competitive with other resources, so that Dunaliella production to date is limited to the nutraceuticals market. For the accumulation of high concentrations of β ‐carotene and glycerol, the algae is cultivated at extreme salinity with reported very low productivity of about 2 g m‐2 day‐1, which is not competitive for biofuels production.
Production system, harvesting and processing
The larger the individual ponds used to grow the algae, the smaller the productivity of the system seems to be. In D. salina the optimum salinity for growth lies between 18 and 22% NaCl whereas the optimum salinity for carotenoid production is >27% NaCl (Borowitzka et al., 1984). Thus the optimum yield of β‐carotene per unit volume and time is achieved at about 24% NaCl.
Two strategies can be applied to maximize the production of β‐carotene. One of these strategies is a two‐stage growth process in which the algae are first grown at a low salinity (= 15% NaCl) in nutrient‐rich medium to maximize biomass production, and then transferred to a high salinity, low nutrient medium in order to induce β‐carotene production (Borowitzka et al., 1984). A similar, two‐stage production process was proposed by Chen and Chi (1981) for glycerol production from Dunaliella.
NBT Eilat cultivates Dunaliella in open raceway ponds in hyper saline water obtained after passages through evaporation ponds and achieves productivities of about 2 g m‐2 day‐1. A complete description of different production processes and sites, including harvesting, processing and economics of the process can be downloaded from: http://www.wind‐sea‐algae.org/wsapresentations/day1/Ami%20BenAmotz%20WSA%20April%202009.pdf
Culture media
The most commonly used medium for culture of Dunaliella is Modified Johnsons Medium (Borowitzka, 1988). However, these algae can also be grown in a wide range of other media including Guillard's f/2 medium (Guillard and Ryther, 1962), modified ASP medium (McLachlan and Yentsch, 1959) and enriched seawater (Rao and Chauhan, 1984).
Upscaling limitations
At low salinities protozoa such as the ciliate Fabrea salina and the amoeba Heteroamoeba sp. can invade the culture and very rapidly decimate the algal production.
Lower salinities generally favour the growth of the non‐carotenogenic Dunaliella species (D. viridis, D. minuta and D. parva) that can overgrow D. salina and drastically reduce the β‐carotene productivity of the pond (Burford and Borowitzka, 1987).
A two‐stage process requires greater capital and running costs and thus may make the process uneconomic.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 82 of 258
HIGHLIGHTS IN BIOTECHNOLOGY
Dunaliella was the first microalga cultivated in large scale for production of a high value product (β‐carotene) rather than bulk biomass. Its cultivation was favoured by the hypersaline medium which reduces the number of potential contaminants during long term open pond cultivation.
References
Ben‐Amotz A., Avron M. (1982) The potential use of Dunaliella for the production of glycerol, β‐carotene and high‐protein feed. In: San Pietro A. (ed.) Biosaline research: A look to the future. Plenum Pub. Corp., New York, pp. 207‐214.
Borowitzka M.A. (1988) Algal growth media and sources of cultures. In: Borowitzka M.A,. Borowitzka L.J. (eds.) Micro‐algal Biotechnology. Cambridge University Press, Cambridge, pp. 456‐465.
Borowitzka L.J., Borowitzka M.A., Moulton T.P. (1984) The mass culture of Dunaliella for fine chemicals: from laboratory to pilot plant. Hydrobiologia 116/117: 115‐121.
Burford M.A., Borowitzka L.J. (1987) Competition between Dunaliella species at high salinity. Hydrobiologia 151/152: 107‐116. Chen B.J., Chi C.H. (1981) Process development and evaluation for algal glycerol production. Biotechnology and Bioengineering 23:
1267‐1287. Cifuentes A. (1992) Growth and carotenogenesis in eight strains of Dunaliella salina Teodoresco from Chile. Journal of Applied
Phycology 4: 111‐118. Gimmler H, Weis U., Weiss C., Kugel H., Treffny B. (1989) Dunaliella acidophila (Kalina) Masyuk ‐ an alga with a positive membrane
potential. New Phytologist 113: 175‐184. Ginzburg M.(1987) Dunaliella: a green alga adapted to salt. Advances in Botanical Research 14: 93‐183. Guillard R.R.L., Ryther J.H. (1962). Studies of marine planktonic diatoms. I. Cyclotella nana (Hustedt) and Detonula confervacea
(Cleve). Canadian Journal of Microbiology 8: 229‐239. Jimenez C., Pick U. (1993) Differential reactivity of β‐carotene isomers from Dunaliella bardawil toward oxygen radicals. Plant
Physiology 101: 385‐390. McLachlan J., Yentsch C.S. (1959) Observations on the growth of Dunaliella euchlora in culture. The Biological Bulletin 116: 461‐471. Rabbani S., Beyer P., Von Lintig J., Hugueney P., Kleinig H. (1998) Induced beta‐carotene synthesis driven by triacylglycerol
deposition in the unicellular alga Dunaliella bardawil. Plant Physiology 116: 1239‐1248. Rao P.S.N., Chauhan V.D. (1984) On occurrence and growth of Dunaliella from India. I. Enriched seawater for mass culture of the
alga. Phykos 23: 33‐37.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 83 of 258
8.1.7 Chlorococcum sp.
Figure 23 ‐ Chlorococcum sp.
UTEX 819, D. Reinecke, BGU SYMBOLS: H, D, E
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Chlorococcales Family Chlorococcaceae Genus Chlorococcum Species
Related Species
There are 76 species (and infraspecific) names in the database at present, of which 34 have been flagged as currently accepted taxonomically.
C. acidum, C. aegyptiacum, C. botryoides, C. choloepodis, C. citriforme, C. costatozygotum, C. diplobionticum, C. dissectum, C. echinozygotum, C. elbense, C. elkhartiense, C. ellipsoideum, C. hypnosporum, C. infusionum, C. isabeliense, C. lobatum, C. macrostigmatum, C. minimum, C. minutum, C. novae‐angliae, C. oleofaciens, C. olivaceum, C. pamirum, C. pinguideum, C. polymorphum, C. pseudodictyosphaerium, C. pyrenoidosum, C. refringens, C. salinum, C. schizochlamys, C. schwarzii, C. submarinum, C. tatrense, C. vacuolatum.
BIOLOGY
Vegetative cells solitary or in temporary groups of indefinite form, never embedded in gelatin. Cells ellipsoidal to spherical and vary in size. Cell walls smooth. The chloroplast is cup‐shaped, parietal, with or without a peripheral opening and has one or more pyrenoids. Cells uniucleate, or multinucleate just prior to zoosporogenesis. Reproduction by zoospores, aplanospores, or isogametes. Motile cells have two equal flagella and remain ellipsoidal for a time after motility ceases.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 84 of 258
This free‐living genus is cosmopolitan. Though primarily an edaphic alga, it has been reported from such diverse habitats as hot springs in Central Asia and soils collected in Antarctica. Aquatic, marine, and aerial isolates have been recorded.
BIOTECHNOLOGY
Several strains of Chlorococcum have been tested as potential sources of astaxanthin (Zhang and Lee, 1997; Liu and Lee, 2000; Masojídek et al., 2000; Ma and Chen, 2001). Outdoor cultures to verify astaxanthin yield have been performed. An enhanced astaxanthin producing mutant was cultivated in a tubular‐loop photobioreactor consisting of two inclined panels with 34 Pyrex glass tubes each (each tube approximately 1 m long and 1.2‐cm in diameter), reaching biomass productivities up to 0.3 g L‐1 day‐1 and ketocarotenoid productivities up to 3.4 mg L‐1 day‐1 (Zhang and Lee, 1999). In a horizontal 50‐L tubular photobioreactor Masojídek et al. (2000) obatined growth rates about 0.13 h‐1, four times that of Haematococcus, but with 20 times lower secondary carotenoid content. In 10‐L bubbled tubes Tredici and co‐workers (unpublished data) obtained productivities of 0.20 g L‐1 day‐1 outdoors and 0.27 g L‐1 day‐1 indoors under contnuos illumination, with temperature reaching at mid day values up to 45 and 36 °C, respectively.
In a flat‐plate photobioreactor under artificial light of 2000 μmol photons m‐2 s‐1, a ultra‐high density culture of the marine Chlorocococcum littorale reached a productivity of 380 ± 20 mg l‐1 h‐1, with 1‐cm light path length, a value 2.4 and 6.4 times higher than those obtained in the 2‐ and 4‐cm reactors. Culture denisites as high as 84 g L‐1 were reached and daily CO2 fixation rate was 16.7 g L
‐1 (Hu et al., 1998). Chlorococcum has been investigated as hydrogen producer (Schnackenberg et al., 1995; Ueno et al.,
1999), but hydrogen yields are much lower than those obtained with Chlamydomonas and Scenedesmus spp. (Winkler et al., 2002). Chlorococcum was also proposed for bioethanol production via dark fermentation of starch (Ueno et al., 1998; Harun and Danquah, 2011; Harun et al., 2011) and was investigated as a source of lipid for biodiesel production (Rodolfi et al., 2009; Halim et al., 2011).
References
Algaebase: http://www.algaebase.org/search/genus/detail/?genus_id=37477 [BIOLOGY section] Halim R., Gladman B., Danquah M.K., Webley P.A. (2011) Oil extraction from microalgae for biodiesel production. Bioresource
Technology 102: 178‐185. Harun R., Danquah M.K. (2011) Influence of acid pre‐treatment on microalgal biomass for bioethanol production. Process
Biochemistry 46: 304‐309. Harun R., Jason W.S.Y., Cherrington T., Danquah M.K. (2011) Exploring alkaline pre‐treatment of microalgal biomass for bioethanol
production. Applied Energy: doi:10.1016/j.apenergy.2010.10.048. Hu Q., Kurano N., Kawachi M., Iwasaki I., Miyachi S. (1998) Ultrahigh‐cell‐density culture of a marine green alga Chlorococcum
littorale in a flat‐plate photobioreactor. Applied Microbiology and Biotechnology 49: 655‐662. Liu B.H., Lee Y.K. (2000) Secondary carotenoids formation by the green alga Chlorococcum sp. Journal of Applied Phycology 12: 301‐
307. Ma R.Y.N., Chen F. (2001) Enhanced production of free trans‐astaxanthin by oxidative stress in the cultures of the green microalga
Chlorococcum sp. Process Biochemistry 36: 1175‐1179. Masojídek J., Torzillo G., Kopecký J., Koblížek M., Nidiaci L., Komenda J., Lukavská A., Sacchi A. (2000) Changes in chlorophyll
fluorescence quenching and pigment composition in the green alga Chlorococcum sp. grown under nitrogen deficiency and salinity stress. Journal of Applied Phycology 12: 417‐426.
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Schnackenberg J., Ikemoto H., Miyachi S. (1995) Relationship between oxygen‐evolution and hydrogen‐evolution in a Chlorococcum strain with high CO2‐tolerance. Journal of Photochemistry and Photobiology B: Biology 28: 171‐174.
Ueno Y., Kurano N., Miyachi S. (1998) Ethanol production by dark fermentation in the marine green alga, Chlorococcum littorale. Journal of Fermentation and Bioengineering 86: 38‐43.
Ueno Y., Kurano N., Miyachi S. (1999) Purification and characterization of hydrogenase from the marine green alga, Chlorococcum littorale. FEBS Letters 443: 144‐148.
Winkler M., Hemschemeier A., Gotor C., Melis A., Happe T. (2002) [Fe]‐hydrogenases in green algae: photo‐fermentation and hydrogen evolution under sulfur deprivation. International Journal of Hydrogen Energy 27: 1431‐1439.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 85 of 258
Zhang D.H., Lee Y.K., Ng M.L., Phang S.M. (1997) Enhanced accumulation of secondary carotenoids in a mutant of the green alga, Chlorococcum sp. Journal of Applied Phycology 9: 147‐155.
Zhang D.H., Lee Y.K. (1999) Ketocarotenoid production by a mutant of Chlorococcum sp. in an outdoor tubular photobioreactor. Biotechnology Letters 21: 7‐10.
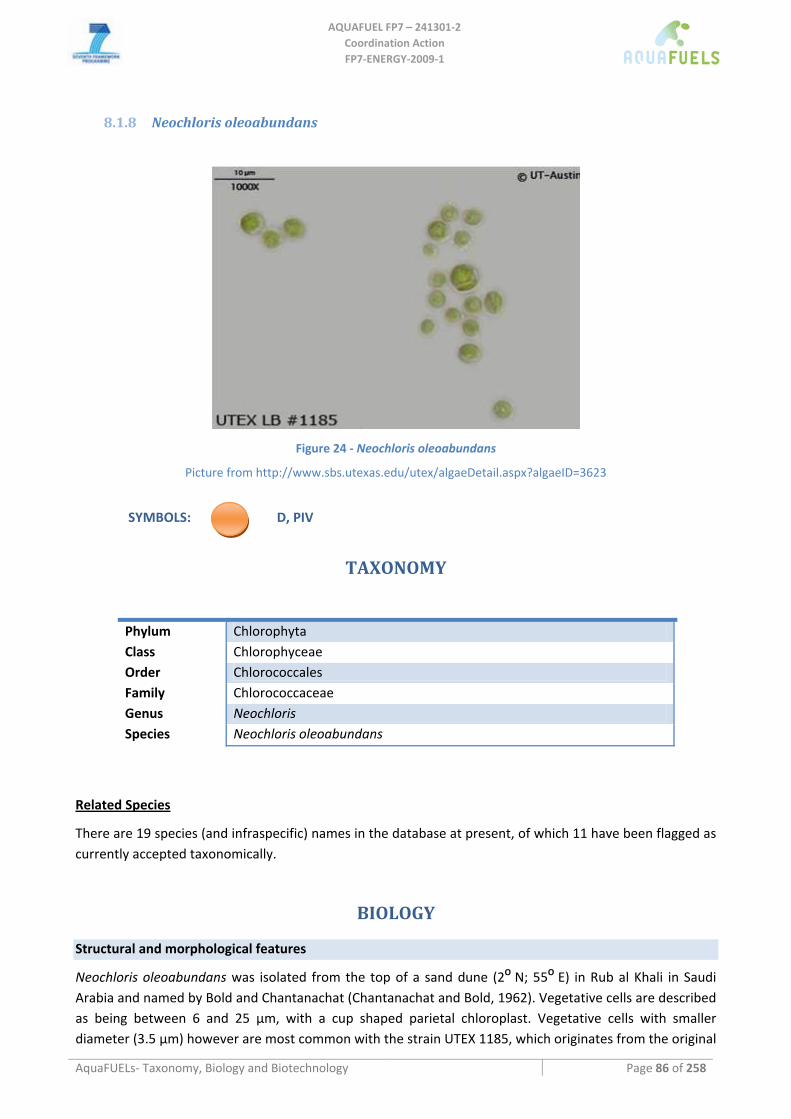
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 86 of 258
8.1.8 Neochloris oleoabundans
Figure 24 ‐ Neochloris oleoabundans
Picture from http://www.sbs.utexas.edu/utex/algaeDetail.aspx?algaeID=3623
SYMBOLS: D, PIV
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Chlorococcales Family Chlorococcaceae Genus Neochloris Species Neochloris oleoabundans
Related Species
There are 19 species (and infraspecific) names in the database at present, of which 11 have been flagged as currently accepted taxonomically.
BIOLOGY
Structural and morphological features
Neochloris oleoabundans was isolated from the top of a sand dune (2O N; 55O E) in Rub al Khali in Saudi Arabia and named by Bold and Chantanachat (Chantanachat and Bold, 1962). Vegetative cells are described as being between 6 and 25 μm, with a cup shaped parietal chloroplast. Vegetative cells with smaller diameter (3.5 μm) however are most common with the strain UTEX 1185, which originates from the original
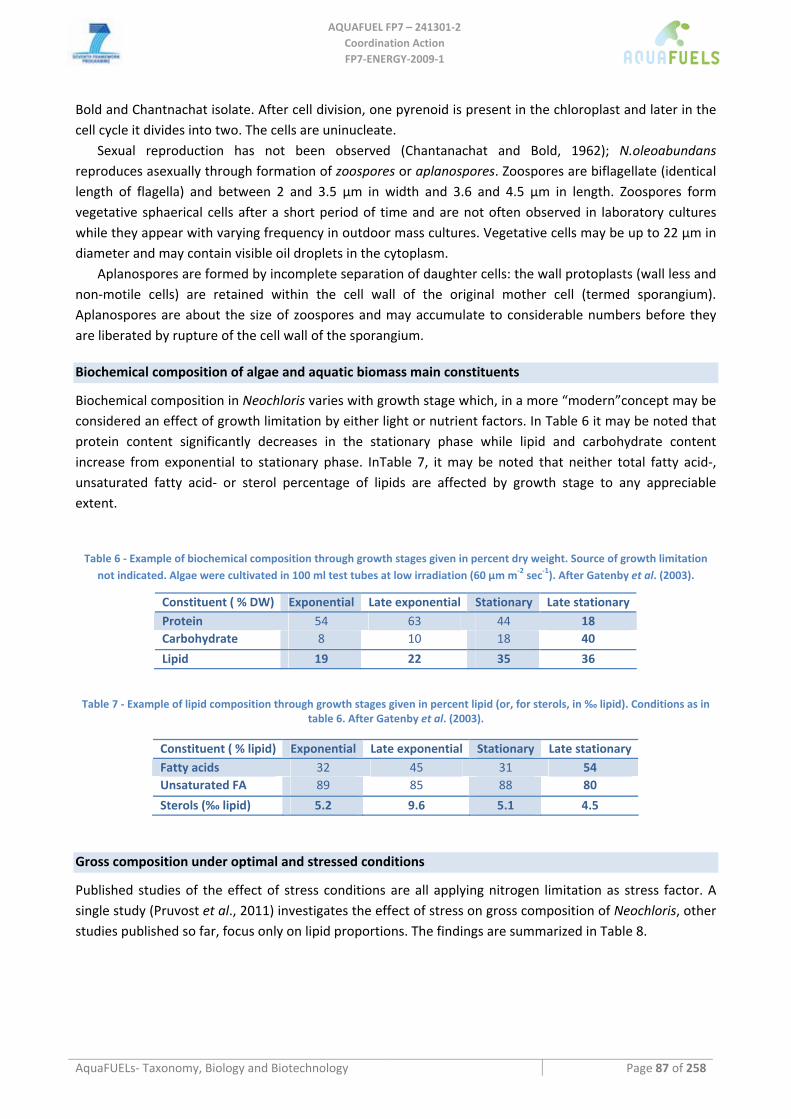
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 87 of 258
Bold and Chantnachat isolate. After cell division, one pyrenoid is present in the chloroplast and later in the cell cycle it divides into two. The cells are uninucleate.
Sexual reproduction has not been observed (Chantanachat and Bold, 1962); N.oleoabundans reproduces asexually through formation of zoospores or aplanospores. Zoospores are biflagellate (identical length of flagella) and between 2 and 3.5 μm in width and 3.6 and 4.5 μm in length. Zoospores form vegetative sphaerical cells after a short period of time and are not often observed in laboratory cultures while they appear with varying frequency in outdoor mass cultures. Vegetative cells may be up to 22 μm in diameter and may contain visible oil droplets in the cytoplasm.
Aplanospores are formed by incomplete separation of daughter cells: the wall protoplasts (wall less and non‐motile cells) are retained within the cell wall of the original mother cell (termed sporangium). Aplanospores are about the size of zoospores and may accumulate to considerable numbers before they are liberated by rupture of the cell wall of the sporangium.
Biochemical composition of algae and aquatic biomass main constituents
Biochemical composition in Neochloris varies with growth stage which, in a more “modern”concept may be considered an effect of growth limitation by either light or nutrient factors. In Table 6 it may be noted that protein content significantly decreases in the stationary phase while lipid and carbohydrate content increase from exponential to stationary phase. InTable 7, it may be noted that neither total fatty acid‐, unsaturated fatty acid‐ or sterol percentage of lipids are affected by growth stage to any appreciable extent.
Table 6 ‐ Example of biochemical composition through growth stages given in percent dry weight. Source of growth limitation not indicated. Algae were cultivated in 100 ml test tubes at low irradiation (60 μm m‐2 sec‐1). After Gatenby et al. (2003).
Constituent ( % DW) Exponential Late exponential Stationary Late stationary Protein 54 63 44 18 Carbohydrate 8 10 18 40
Lipid 19 22 35 36
Table 7 ‐ Example of lipid composition through growth stages given in percent lipid (or, for sterols, in ‰ lipid). Conditions as in table 6. After Gatenby et al. (2003).
Constituent ( % lipid) Exponential Late exponential Stationary Late stationary Fatty acids 32 45 31 54 Unsaturated FA 89 85 88 80
Sterols (‰ lipid) 5.2 9.6 5.1 4.5
Gross composition under optimal and stressed conditions
Published studies of the effect of stress conditions are all applying nitrogen limitation as stress factor. A single study (Pruvost et al., 2011) investigates the effect of stress on gross composition of Neochloris, other studies published so far, focus only on lipid proportions. The findings are summarized in Table 8.
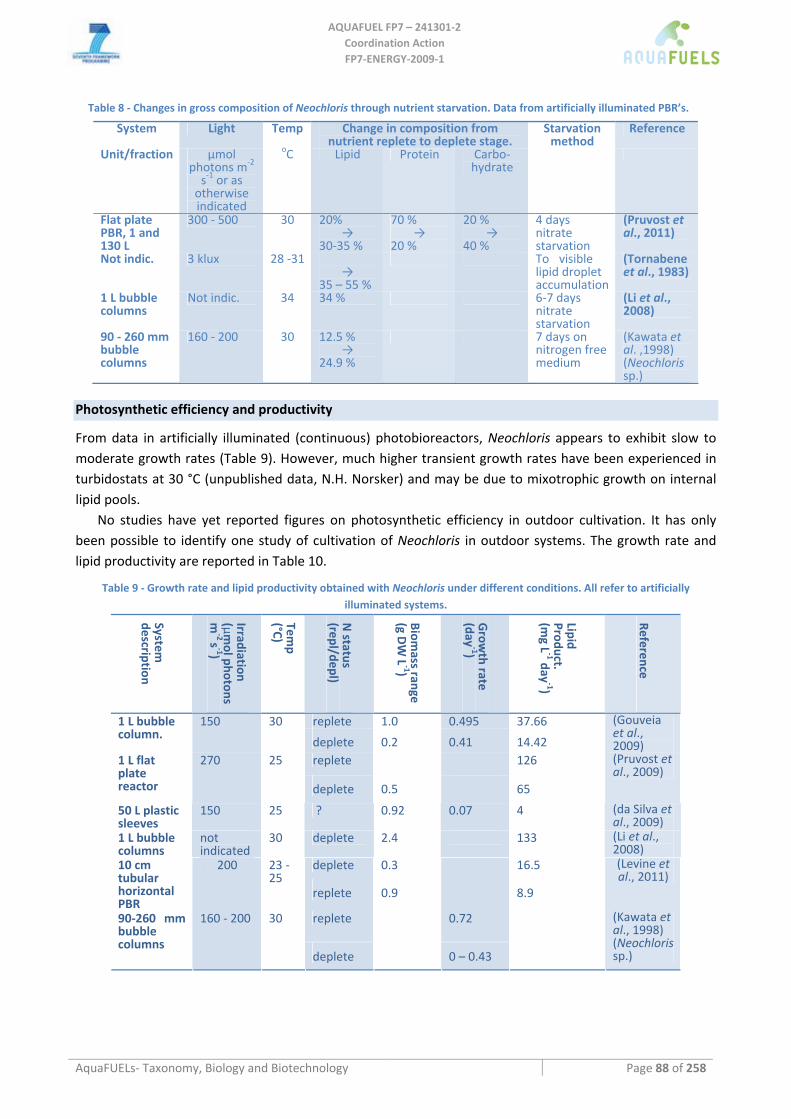
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 88 of 258
Table 8 ‐ Changes in gross composition of Neochloris through nutrient starvation. Data from artificially illuminated PBR’s.
System Light Temp Change in composition from nutrient replete to deplete stage.
Starvation method
Reference
Unit/fraction μmol photons m‐2 s‐1 or as otherwise indicated
oC Lipid Protein Carbo‐ hydrate
Flat plate PBR, 1 and 130 L
300 ‐ 500 30 20% →
30‐35 %
70 % →
20 %
20 % →
40 %
4 days nitrate starvation
(Pruvost et al., 2011)
Not indic. 3 klux 28 ‐31 →
35 – 55 %
To visible lipid droplet accumulation
(Tornabene et al., 1983)
1 L bubble columns
Not indic. 34 34 % 6‐7 days nitrate starvation
(Li et al., 2008)
90 ‐ 260 mm bubble columns
160 ‐ 200 30 12.5 % →
24.9 %
7 days on nitrogen free medium
(Kawata et al. ,1998) (Neochloris sp.)
Photosynthetic efficiency and productivity
From data in artificially illuminated (continuous) photobioreactors, Neochloris appears to exhibit slow to moderate growth rates (Table 9). However, much higher transient growth rates have been experienced in turbidostats at 30 °C (unpublished data, N.H. Norsker) and may be due to mixotrophic growth on internal lipid pools.
No studies have yet reported figures on photosynthetic efficiency in outdoor cultivation. It has only been possible to identify one study of cultivation of Neochloris in outdoor systems. The growth rate and lipid productivity are reported in Table 10.
Table 9 ‐ Growth rate and lipid productivity obtained with Neochloris under different conditions. All refer to artificially illuminated systems.
System
description
Irradiation (μm
ol photons m
‐2 s‐1)
Temp
(°C)
N status
(repl/depl)
Biomass range
(g DW L
‐1)
Grow
th rate (day
‐1)
Lipid Product. (m
g L‐1 day
‐1)
Reference
replete 1.0 0.495 37.66 1 L bubble column.
150 30
deplete 0.2 0.41 14.42
(Gouveia et al., 2009)
replete 126 1 L flat plate reactor
270 25
deplete 0.5 65
(Pruvost et al., 2009)
50 L plastic sleeves
150 25 ? 0.92 0.07 4 (da Silva et al., 2009)
1 L bubble columns
not indicated
30 deplete 2.4 133 (Li et al., 2008)
deplete 0.3 16.5 10 cm tubular horizontal PBR
200 23 ‐ 25
replete 0.9 8.9
(Levine et al., 2011)
replete 0.72 90‐260 mm bubble columns
160 ‐ 200 30
deplete 0 – 0.43
(Kawata et al., 1998) (Neochloris sp.)
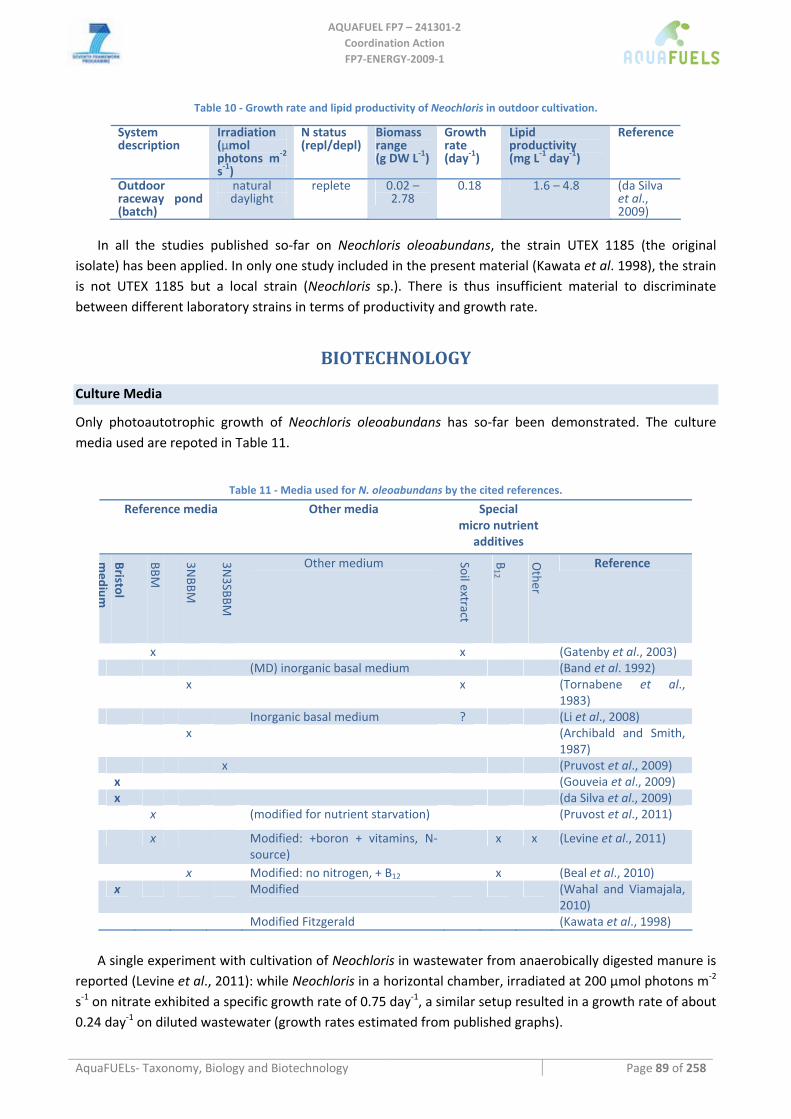
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 89 of 258
Table 10 ‐ Growth rate and lipid productivity of Neochloris in outdoor cultivation.
System description
Irradiation (μmol photons m‐2
s‐1)
N status (repl/depl)
Biomass range (g DW L‐1)
Growth rate (day‐1)
Lipid productivity (mg L‐1 day‐1)
Reference
Outdoor raceway pond (batch)
natural daylight
replete 0.02 – 2.78
0.18 1.6 – 4.8 (da Silva et al., 2009)
In all the studies published so‐far on Neochloris oleoabundans, the strain UTEX 1185 (the original isolate) has been applied. In only one study included in the present material (Kawata et al. 1998), the strain is not UTEX 1185 but a local strain (Neochloris sp.). There is thus insufficient material to discriminate between different laboratory strains in terms of productivity and growth rate.
BIOTECHNOLOGY
Culture Media
Only photoautotrophic growth of Neochloris oleoabundans has so‐far been demonstrated. The culture media used are repoted in Table 11.
Table 11 ‐ Media used for N. oleoabundans by the cited references.
Reference media Other media Special micro nutrient
additives
Bristol medium
BBM
3NBBM
3N3SBBM
Other medium Soil extract
B12
Other
Reference
x x (Gatenby et al., 2003) (MD) inorganic basal medium (Band et al. 1992) x x (Tornabene et al.,
1983) Inorganic basal medium ? (Li et al., 2008) x (Archibald and Smith,
1987) x (Pruvost et al., 2009) x (Gouveia et al., 2009) x (da Silva et al., 2009) x (modified for nutrient starvation) (Pruvost et al., 2011)
x Modified: +boron + vitamins, N‐ source)
x x (Levine et al., 2011)
x Modified: no nitrogen, + B12 x (Beal et al., 2010) x Modified (Wahal and Viamajala,
2010) Modified Fitzgerald (Kawata et al., 1998)
A single experiment with cultivation of Neochloris in wastewater from anaerobically digested manure is
reported (Levine et al., 2011): while Neochloris in a horizontal chamber, irradiated at 200 μmol photons m‐2 s‐1 on nitrate exhibited a specific growth rate of 0.75 day‐1, a similar setup resulted in a growth rate of about 0.24 day‐1 on diluted wastewater (growth rates estimated from published graphs).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 90 of 258
Cultivation methods
Only one experiment cultivating Neochloris in open systems has been reported. The raceway area was 2.5 m2, the depth of 15 cm and it was agitated with a paddlewheel to create a flow velocity of 25 cm s‐1. The culture was diluted after 20 and 36 days of cultivation (da Silva et al., 2009). Specific growth rate during the first 20 days was 0.18 day‐1. Other systems have been used for indoor use and among these:
• bubble columns of various diameter up to 26 cm (Kawata et al. 1998; Li et al. 2008; Gouveia et al. 2009)
• flat panel reactors, 1 ‐130 L (Pruvost et al., 2011) • shaken flasks (Wahal and Viamajala, 2010)
• plastic bags (da Silva et al., 2009; Levine et al., 2011)
Production systems
At present time (2010), Neochloris has not reached a state of commercial production. Neither has any information about testing of production systems in pilot scale been published.
Harvesting methods
No studies of harvesting methods for Neochloris have yet been published. Speculating about harvest methods, it is worth noting that the cell size of Neochloris oleoabundans is rather small (average diameter assuming sphaerical cells 3 µm for vegetatively growing cultures) and high energy input for separation by, for instance, centrifugation could be assumed. However, it is generally observed that Neochloris sediments rather readily in culture flasks and may therefore be suitable for flocculation.
Flotation or foam separation are other potentially attractive harvesting technologies as Neochloris cultures are prone to foaming.
Biomass processing
There are no published studies on processing Neochloris biomass for any purpose. For potential use for biodiesel purposes, it is worth noting that a substantial part of the lipid fraction may be constituted by triacylglycerols: 80 % with lipids constituting 35 – 54 % of the biomass (Tornabene et al., 1983). A recent study of oil formation optimization demonstrated a TAG yield of 50 % of total lipids or 18% of DW (Pruvost et al., 2009). Relatively high specific TAG productivities by Neochloris is one of the main motives for focussing on Neochloris for biodiesel production purposes.
Scaling up limitation
Production of Neochloris for biodiesel is here considered only for photoautotrophic production methods. Conversion of carbohydrate sources to biodiesel by heterotrophic production is being considered with other algae species, but a heterotrophic growth potential of Neochloris has not been established.
Due to the lack of experience with outdoor cultivation of Neochloris, it is very difficult to pin point limitations to scale up.
Two studies have dealt with outdoor production. In da Silva et al. (2009) production of Neochloris in a 2.5 m 2 raceway pond resulted in high biomass densities (2.8 g DW L‐1) at the end of a 20 days batch growth run with specific growth rate of 0.18 day‐1 . Lipid productivity at the peak was 4.8 g m‐2 day‐1 (July, Portugal) which must be considered promising if such rates can be sustained over longer periods. It is likely that the lipid productivity can be enhanced by reducing the biomass density in the pond as high average light intensities are required to obtain high lipid productivity and the other design parameter, culture depth which determines the average light intensity in the culture, cannot be reduced in raceway ponds for

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 91 of 258
practical reasons. This will, on the other hand, add to the harvesting costs and may add to the vulnerability of the culture to contamination by other algae or protozoans and zooplankton.
In Levine et al. (2011) the use of wastewater from biogas production as fertilizer was studued, but overgrowth by other algae was encountered and closed reactors were suggested for the production.
For these reasons, the development of photobioreactors with low installation and operating costs is believed to be the most promising technologies for a scale up strategy. References
Arce G., Bold H.C. (1958) Some Chlorophyceae from Cuban soils. American Journal of Botany 45: 492‐503. Archibald P.A., Smith V.J. (1987) Notes on variation on physiological attributes between aquatic and edaphic species of the
chlorophycean algal genus Neochloris. Transactions of the American Microscopical Society 106: 179‐182. Band C.J., Arredondovega B.O., Vazquezduhalt R., Greppin H. (1992) Effect of a salt‐osmotic upshock on the edaphci microalga
Neochloris oleoabundans. Plant Cell and Environment 15: 129‐133. Beal C.M., Webber M.E., Ruoff R.S., Hebner R.E. (2010) Lipid analysis of Neochloris oleaoabundans by liquid state NMR.
Biotechnology and Bioengineering 106: 573‐583. Chantanachat S., Bold H.C. (1962) Phycological Studies. II. Some algae from arid soils. University of Texas Publication (6218). da Silva T., Reis A., Medeiros R., Oliveira A., Gouveia L. (2009) Oil production towards biofuel from autotrophic microalgae
semicontinuous cultivations monitorized by flow cytometry. Applied Biochemistry and Biotechnology 159: 568‐578. Gatenby C.M., Orcutt D.M., Kreeger D.A., Parker B.C., Jones V.A., Neves R.J. (2003) Biochemical composition of three algal species
proposed as food for captive freshwater mussels. Journal of Applied Phycology 15: 1‐11. Gouveia L., Marques A., da Silva T., Reis A. (2009) Neochloris oleoabundans UTEX #1185: a suitable renewable lipid source for
biofuel production. Journal of Industrial Microbiology and Biotechnology 36: 821‐826. Kawata M., Nanba M., Matsukawa R., Chihara M., Karube I. (1998) Isolation and characterization of a green alga Neochloris sp. for
CO2 fixation. Studies in Surface Science and Catalysis 114: 637‐640. Levine R.B., Costanza‐Robinson M.S., Spatafora G.A. (2011) Neochloris oleoabundans grown on anaerobically digested dairy manure
for concomitant nutrient removal and biodiesel feedstock production. Biomass and Bioenergy 35: 40‐49. Li Y., Horsman M., Wang B., Wu N., Lan C. (2008) Effects of nitrogen sources on cell growth and lipid accumulation of green alga
Neochloris oleoabundans. Applied Microbiology and Biotechnology 81: 629‐636. Pruvost J., Van Vooren G., Cogne G., Legrand J. (2009) Investigation of biomass and lipids production with Neochloris oleoabundans
in photobioreactor. Bioresource Technology 100: 5988‐5995. Pruvost J., Van Vooren G., Le Gouic B., Couzinet‐Mossion A., Legrand J. (2011) Systematic investigation of biomass and lipid
productivity by microalgae in photobioreactors for biodiesel application. Bioresource Technology 102: 150‐158. Starr R.C. (1955) A comparative study of Chlorococcum meneghini and other spherical, zoospore‐producing genera of the
Chlorococcales. Indiana University Publications Science 20: 1‐111. Starr R.C. (1978) Culture collection of algae at the University‐of‐Texas at Austin. Journal of Phycology 14: 47‐100. Tornabene T.G., Holzer G., Lien S., Burris N. (1983) Lipid composition of the nitrogen starved green alga Neochloris oleoabundans.
Enzyme and Microbial Technology 5: 435‐440. Wahal S., Viamajala S. (2010) Maximizing algal growth in batch reactors using sequential change in light intensity. Applied
Biochemistry and Biotechnology 161: 511‐522.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 92 of 258
8.1.9 Scenedesmus sp.
Figure 25 ‐ Scenedesmus sp. Local isolate Sde Boker, Israel
D. Reinecke, BGU SYMBOLS: B, D, E, PIV
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Sphaeropleales Family Scenedesmaceae Genus Scenedesmus Species
Related Species
There are 418 species (and infraspecific) names in the database at present, of which 87 have been flagged as currently accepted taxonomically.
BIOLOGY Scenedesmus is a freshwater medium to large size unicellular green alga often appearing in tetrads with four to 16 elongated cells connected (cenobia), although cells can also appear as individual in oval form.
Scenedesmus is versatile in production of oil and secondary metabolites such as carotenoids. Multiple species produce different carotenoids during logarithmic growth or stress, though cellular levels never exceed 1% of dry weight. It is relevant that major carotenoid is usually lutein. Exponentially growing cells have high protein content (up to 50%), less than 10% lipids, the rest variable amounts of starch and cell wall components. Under nutrient stress Scenedesmus species can accumulate high amounts of storage lipids (TAG) in cytoplasmic oil globules. Fatty acid composition under logarithmic growth includes significant amounts of short polyunsaturated fatty acids such as gamma linolenic and stearidonic acid. Under stress, TAG accumulates mostly C16 and C18 unsaturated and some monounsaturated fatty acids.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 93 of 258
Growth kinetics and efficiencies
Scenedesmus is among the most vigorously growing green algae and outcompetes most other algal species under high nutrient conditions, e. g. in wastewater. It can cause serious contamination problems when cultivating slower growing algae such as Haematococcus. It is among the faster growing and highest oil producing strains tested by Rodolfi et al. (2009) or Hu et al. (2008).
Growth rates and productivity in laboratory main groups
Maximum specific growth rates as higher than 0.12 h‐1 has been reported for Scenedesmus sp. Lower specific growth rates, of 0.04 h‐1, has been reported for Scenedesmus obliquus. The most relevant aspect of this strain is its tolerance to high temperature, including 40 °C, its maximum growth rate being obtained at temperatures of 30‐35 °C. At laboratory scale biomass productivities of 0.9 g∙L‐1∙day‐1 has been reported for Scenedesmus almeriensis. High productivity requires the use of high irradiance, but Scenedesmus has demonstrate to be resistant to irradiances higher than 1700 μmol photons∙m‐2∙s‐1 without photoinhibition. Scenedesmus is also tolerant to impulsion using centrifugal pumps, in addition to aeration, no mechanical damage being reported by this phenomenon. Concerning pH, Scenedesmus tolerates wide ranges of pH, from 5 to 10, although optimal pH is in the range of 7.5‐8.0. It is particularly relevant the tolerance of Scenedesmus to alkaline pH for wastewater and flue flue gas depuration.
Photosynthetic efficiency and productivity outdoors
Due to the fact that Scenedesmus is highly robust and fast growing, it has been produced outdoors using both open systems and closed photobioreactors. In open raceways, biomass productivities higher than 0.5 g∙L‐1∙day‐1 has been reported, while biomass productivities up to 1.2 g∙L‐1∙day‐1 have been obtained using closed photobioreactors. Overall mean annual productivity of 0.6 g∙L‐1∙day‐1 has been obtained in pilot scale tubular photobioreactors (30 m3) with Scenedesmus almeriensis. This value is about 3% solar efficiency. Solar efficiencies from 1‐3% has have been reported for Scenedesmus.
Gross composition under optimal and stressed conditions
Under optimal growth conditions the major component of Scenedesmus biomass are proteins, 40‐50% d.wt., next being carbohydrates. The lipid content under adequate growth conditions is lower than 15%, woith maximum values of 10% corresponding to fatty acids. It is relevant that under optimal conditions the carotenoid content, especially lutein, increases up to 1% in some species. Scenedesmus cells can be stressed by nutrient depletion, nitrogen or phosphorous, which triggers accumulation of lipids. However, no lipid content higher than 30% are obtained in these conditions.
BIOTECHNOLOGY
Culture Media
Scenedesmus grows well in modified BG11 medium, but also in other growth media, in nutrient rich wastewater etc, and apparently may also exploit organic molecules for photoheterotrophic growth. It has been cultivated using commercial fertilizers outdoors, and in brackish water. It is tolerant to the use of nitrate, ammonia or urea as nitrogen source, thus Scenedesmus is adequate for removal of inorganic nitrogen from effluents.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 94 of 258
Cultivation methods
Scenedesmus can be cultivated in either discontinuous (batch) or continuous mode. In discontinuous mode the productivity is lower, thus usually semi‐continuous operation being utilized. No large variation in the composition of the biomass is observed when cultivated in different modes. In continuous mode the optimal dilution rate is in the range from 0.3‐0.4 day‐1.
Production system
Scenedesmus are not damaged by aeration or impulsion centrifugal pumps, thus it has been cultivated in most of culture systems developed, including open and closed photobioreactors. Fouling is an important issue when cultivating this strain. Under adverse conditions, but also under optimal conditions, cells of Scenedesmus fix on the reactor’s walls, reducing light availability inside the culture. When closed reactors are used it is obligatory to implement self‐cleaning systems. This is not necessary when using open raceways.
Harvesting methods
Although Scenedesmus cells are larger than other microalgae cells, their settling velocity by gravity is low, in the range of 10‐6 m∙s‐1. Thus, Scenedesmus cells cannot be harvested by natural sedimentation. It is necessary the use of centrifugation or filtration operations. Cells of Scenedesmus are easily harvested by centrifugation: a slurry containing 15% d.wt is obtained under continuous operation, and a paste containing 30% d.wt. is obtained under discontinuous operation. Although filtration can be performed, it is not usually carried out. Natural sedimentation can be performed by previous flocculation. The use of anionic polyelectrolyte at doses below 0.1 mg/L has been demonstrated to be useful to pre‐concentrate the culture by twenty times.
Biomass processing
Biomass of Scenedesmus can be used as biomass for feeding animals or fishes, mainly for its high protein content. The fatty acid profile is not highly valuable because no PUFAs are present. The only especially valuable components present in the biomass of Scenedesmus are carotenoids. Lutein contents up to 1% d.wt. has have been reported, it being useful as nutraceutical for human and animals. Lutein can be extracted using organic solvents, although new processes using supercritical fluids or ethanol‐water mixtures have been proposed.
Scaling up limitation
No limitations are directly related with the culture of Scenedesmus, different than the production of other strains. For scale‐up purposes the tolerance of this strain to high temperatures, up to 45 °C, is highly relevant.
HIGHLIGHTS IN BIOTECHNOLOGY
Scenedesmus is highly robust and has been used successfully used to treat such problematic waste effluents as olive mill wastewater, biogas or municipal land fill effluents which are toxic to most microorganisms including bacteria. A biotechnology for production and purification of lutein from Scenedesmus has been developed based on cultivation in tubular photobioreactors and extraction and purification of the carotenoid from cellular lysate, though the technology is not currently commercially applied.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 95 of 258
References
Abeliovich A., Dikbuck S. (1977) Factors affecting infection of Scenedesmus obliquus by a Chytridium sp. in sewage oxidation ponds. Applied Environmental Microbiology 34: 832‐836.
Allard B., Templier J. (2000) Comparison of neutral lipid profile of various trilaminar outer cell wall (TLS)‐containing microalgae with emphasis on algaenan occurrence. Phytochemistry 54: 369‐380.
Benemann J.R., Koopman B.L., Murry M., Weissman J.C., Eisenberg D.M., Oswald W.J. (1977) Species control in large‐scale algal biomass production. Final report. United States.
Blokker P., van den Ende H., de Leeuw J.W., Versteegh G.J.M., Damste J.S.S. (2006) Chemical fingerprinting of algaenans using RuO4 degradation. Organic Geochemistry 37: 871‐881.
Buchheim M.A., Michalopulos E.A., Buchheim J.A. (2001) Phylogeny of the Chlorophyceae with special reference to the Sphaeropleales: a study of 18S and 26S rDNA data. Journal of Phycology 37: 819‐835.
Cepak V., Pribyl P. (2006) The effect of colour light on production of zooids in 10 strains of the green chlorococcal alga Scenedesmus obliquus. Czech Phycology 6: 127‐133.
Cepak V., Pribyl P., Vitova M., Zachleder V. (2007) The nucleocytosolic and chloroplast cycle in the green chlorococcal alga Scenedesmus obliquus (Chlorophyceae, Chlorococcales) grown under various temperatures. Phycologia 46: 263‐269.
Czygan F. (1968) Secondary carotenoids in green algae II. Studies on biogenesis. Archiv für Mikrobiologie 62: 209‐236. de Morais M.G., Costa J.A.V. (2007) Biofixation of carbon dioxide by Spirulina sp. and Scenedesmus obliquus cultivated in a three‐
stage serial tubular photobioreactor. Journal of Biotechnology 129: 439‐445. Dean A.P., Sigee D.C., Estrada B., Pittman J.K. (2010) Using FTIR spectroscopy for rapid determination of lipid accumulation in
response to nitrogen limitation in freshwater microalgae. Bioresource Technology 101: 4499‐4507. Disch A., Schwender J., Muller C., Lichtenthaler H.K., Rohmer M. (1998) Distribution of the mevalonate and glyceraldehyde
phosphate/pyruvate pathways for isoprenoid biosynthesis in unicellular algae and the cyanobacterium Synechocystis PCC 6714. Biochemical Journal 333: 381‐388.
Gonzalez Lopez CV, Ceron Garcia MdC, Fernandez FG, Segovia Bustos C, Chisti Y, Fernandez Sevilla JM. 2010. Protein measurements of microalgal and cyanobacterial biomass. Bioresource Technology 101(19):7587‐7591.
Greger M., Johansson M. (2004) Aggregation effects due to aluminum adsorption to cell walls of the unicellular green alga Scenedesmus obtusiusculus. Phycological Research 52: 53‐58.
Grewe C., Menge .S, Griehl C. (2007) Enantioselective separation of all‐E‐astaxanthin and its determination in microbial sources. Journal of Chromatography A 1166: 97‐100.
Gurbuz F., Ciftci H., Akcil A. (2008) Biodegradation of cyanide containing effluents by Scenedesmus obliquus. Journal of Hazardous Materials 162: 74‐79.
Gutman J., Zarka A., Boussiba S. (2009) The host‐range of Paraphysoderma sedebokerensis, a chytrid that infects Haematococcus pluvialis. European Journal of Phycology 44: 509 ‐ 514.
Hanagata N., Dubinsky Z. (1999) Secondary carotenoid accumulation in Scenedesmus komarekii (Chlorophycea, Chlorophyta). Journal of Phycology 35: 960‐966.
Heide H., Kalisz H.M., Follmann H. (2004) The oxygen evolving enhancer protein 1 (OEE) of photosystem II in green algae exhibits thioredoxin activity. Journal of Plant Physiology 161: 139‐149.
Ho S.H., Chen W.M., Chang J.S. (2010) Scenedesmus obliquus CNW‐N as a potential candidate for CO2 mitigation and biodiesel production. Bioresource Technology 101: 8725‐8730.
Hu Q. ,Sommerfeld M. (2004). Selection of high performance microalgae for bioremediation of nitrate‐contaminated roundwater. Technical Report for Grant Number. School of Life Sciences. Arizona State University.
Humbeck K. (1990) Light‐dependent carotenoid biosynthesis in mutant C‐6D of Scenedesmus obliquus. Photochemistry and Photobiology 51: 113‐118.
Kim M.K., Park J.W., Park C.S., Kim S.J., Jeune K.H., Chang M.U., Acreman J. (2006) Enhanced production of Scenedesmus spp. (green microalgae) using a new medium containing fermented swine wastewater. Bioresource Technology 98: 2220‐2228.
Krienitz L., Wirth M. (2006) The high content of polyunsaturated fatty acids in Nannochloropsis limnetica (Eustigmatophyceae) and its implication for food web interactions, freshwater aquaculture and biotechnology. Limnologica ‐ Ecology and Management of Inland Waters 36: 204‐210.
Kuck U., Jekosch K., Holzamer P. (2000) DNA sequence analysis of the complete mitochondrial genome of the green alga Scenedesmus obliquus: evidence for UAG being a leucine and UCA being a non‐sense codon. Gene 253: 13‐18.
Lee J.Y., Yoo C., Jun S.Y., Ahn C.Y., Oh H.M. (2010) Comparison of several methods for effective lipid extraction from microalgae. Bioresource Technology 101 Suppl S1: S75‐S77.
Lopez‐Rodas V., Agrelo M., Carrillo E., Ferrero L.M., Larrauri A., Martin‐Otero L., Costas E. (2001) Resistance of microalgae to modern water contaminants as the result of rare spontaneous mutations. European Journal of Phycology 36: 179‐190.
Lurling M. (2006) Effects of a surfactant (FFD‐6) on Scenedesmus morphology and growth under different nutrient conditions. Chemosphere 62: 1351‐1358.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 96 of 258
Ma J.Y., Xu L.G., Wang S.F., Zheng R.Q., Jin S.H., Huang S.Q., Huang Y.J. (2002) Toxicity of 40 herbicides to the green alga Chlorella vulgaris. Ecotoxicology and Environmental Safety 51: 128‐132.
Mallick N., Mohn F.H., Soeder C.J., Grobbelaar J.U. (2002) Ameliorative role of nitric oxide on H2O2 toxicity to a chlorophycean alga Scenedesmus obliquus. Journal of General and Applied Microbiology 48: 1‐7.
Masojidek J., Torzillo G., Koblizek M., Kopecky J., Bernardini P., Sacchi A., Komenda J. (1999) Photoadaptation of two members of the Chlorophyta Scenedesmus and Chlorella in laboratory and outdoor cultures: changes in chlorophyll fluorescence quenching and the xanthophylls cycle. Planta 209: 126‐135.
Matsunaga T., Matsumoto M., Maeda Y., Sugiyama H,. Sato R., Tanaka T. (2009) Characterization of marine microalga, Scenedesmus sp. strain JPCC GA0024 toward biofuel production. Biotechnology Letters 31: 1367‐1372.
Ordog V., Stirk W., Lenobel R., Bancirova M., Strnad M., van Staden J., Szigeti J., Nemeth L. (2004) Screening microalgae for some potentially useful agricultural and pharmaceutical secondary metabolites. Journal of Applied Phycology 16: 309‐314.
Orosa M., Torres E., Fidalgo P., Abalde J. (2000) Production and analysis of secondary carotenoids in green algae. Journal of Applied Phycology 12: 553‐556.
Proctor V. (1957) Studies of algal antibiosis using Haematococcus and Chlamydomonas. Limnology and Oceanography 2: 125‐139. Proctor V. (1957) Some controling factors in the distribution of Haematococcus pluvialis. Ecology 38: 457‐462. Qiang L., Junda L. (2010) Effects of nitrogen source and concentration on biomass and oil production of a Scenedesmus rubescens
like microalga. Bioresource Technology 102: 1615‐1622. Qin S., Liu G.X., Hu Z.Y. (2008) The accumulation and metabolism of astaxanthin in Scenedesmus obliquus (Chlorophyceae). Process
Biochemistry 43: 795‐802. Radmer R., Behrens P., Arnett K. (1987) Analysis of the productivity of a continuous algal culture system. Biotechnology and
Bioenginering 29: 488‐492. Rausch T. (1981) The estimation of micro‐algal protein content and its meaning to the evaluation of algal biomass I. Comparison of
methods for extracting protein. Hydrobiologia 78: 237‐251. Sánchez J.F., Fernández‐Sevilla J.M., Acién F. G., Cerón M.C., Pérez‐Parra J., Molina‐Grima E. (2008) Biomass and lutein productivity
of Scenedesmus almeriensis: influence of irradiance, dilution rate and temperature. Applied Microbiology and Biotechnology 79: 719–729.
Tripathi U., Sarada R., Ravishankar G.A. (2001) A culture method for microalgal forms using two‐tier vessel providing carbon dioxide environment: Studies on growth and carotenoid production. World Journal of Microbiology & Biotechnology 17: 325‐329.
Vandamme D., Foubert I., Meesschaert B., Muylaert K. (2010) Flocculation of microalgae using cationic starch. Journal of Applied Phycology 22: 525‐530.
Voltolina D., Cordero B, Nievesc M., Soto L.P. (1998) Growth of Scenedesmus sp. in artificial wastewater. Bioresource Technology. 68: 265‐268.
Xia J.R., Gao K.S. (2002) Effects of CO2 enrichment on microstructure and ultrastructure of two species of freshwater green algae. Acta Botanica Sinica 44: 527‐531.
Xin L., Hong‐ying H., Jia Y. (2010) Lipid accumulation and nutrient removal properties of a newly‐isolated freshwater microalga, Scenedesmus sp. LX1, growing in secondary effluent. New Biotechnology 27: 59‐63.
Xin L., Hong‐ying H., Jia Y., Yin‐hu W. (2010) Enhancement effect of ethyl‐2‐methyl acetoacetate on triacylglycerols production by a freshwater microalga, Scenedesmus sp. LX1. Bioresource Technology 101: 9819‐9821.
Yang Y., Gao K.S. (2003) Effects of CO2 concentrations on the freshwater microalgae, Chlamydomonas reinhardtii, Chlorella pyrenoidosa and Scenedesmus obliquus (Chlorophyta). Journal of Applied Phycology 15: 379‐389.
Yoo C., Jun S. Y., Lee J.Y., Ahn C.Y., Oh H.M. (2010) Selection of microalgae for lipid production under high levels carbon dioxide. Bioresource Technology 101 Suppl. 1: S71‐S74.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 97 of 258
8.1.10 Desmodesmus sp.
Figure 26 ‐ Desmodemus sp.
Picture from UNIFI
SYMBOLS: H, D, E
TAXONOMY
Phylum Chlorophyta Class Chlorophyceae Order Sphaeropleales Family Scenedesmaceae Genus Desmodesmus Species
Related Species
There are 72 species (and infraspecific) names in the database at present, of which 16 have been flagged as currently accepted taxonomically.
D. communis, D. costato‐granulatus, D. bicellularis, D. serratus, D. denticulatus, D. lefevrei, D. arthrodesmiformis, Desmodesmus sp. Hegewald 1987‐51, D. subspicatus, D. hystrix, D. opoliensis, D. pannonicus, D. perforatus, D. pirkollei, Desmodesmus sp. CL1, D. maximus, D. tropicus, D. komarekii, D. multivariabilis, D. pleiomorphus, D. fennicus, D. armatus.
BIOLOGY
Desmodesmus used to be the most species‐rich subgenus of Scenedesmus, but it was given genus status based on 18S and ITS2 rDNA phylogenies. The large genetic distance between the two subgenera and their clear distinct cell wall ultrastructure supported retention of the Scenedesmus Meyen for non‐spiny organisms and formation of a genus Desmodesmus (Chodat) An, Friedl et Hegewald for those which could

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 98 of 258
bear spines. Desmodesmus appears as unicells or coenobia 2–4–8–16‐celled, with long axes of cells parallel, laterally adjoined, and arranged in a single linear or alternating series. Cells are ellipsoidal to ovoid, and spines usually are present on the terminal cells and/or medial cells, but may be entirely absent. The cell wall may have ridges, warts, or nets. The chloroplast is parietal, usually with one pyrenoid.
Desmodesmus is an extremely common genus, occasionally abundant, found in the phytoplankton of ponds and lakes. They are cosmopolitans and able to withstand harsh conditions, such as periods with strong grazing pressure.
The relatively low number of studies with Desmodesmus can be explained from investigators still being unaware of the division of the old genus Scenedesmus into the new genera Scenedesmus and Desmodesmus.
BIOTECHNOLOGY
Little scientific literature is available on Desmodesmus, although its application are similar to that reported for Scenedesmus. Part of the literature available about Scenedesmus actually refers to Desmodesmus.
Virginia Coastal Energy Research Consortium cultivated a Desmodemus strain for biodiesel production studies in a 0.4 ha open pond during about a year with low and variable biomass concentrations and quite low lipid yields (Stubbins, 2009).
References
Algaebase: http://www.algaebase.org/search/genus/detail/?genus_id=45397 [BIOLOGY section] Lürling M. (2003) Phenotypic plasticity in the green algae Desmodesmus and Scenedesmus with special reference to the induction
of defensive morphology. Annales de Limnologie ‐ International Journal of Limnology 39: 85‐101. [BIOLOGY section] Shubert L.E. (2003) Nonmotile coccoid and colonial green algae. In: Wehr J.D., Sheath R.G. (eds.) Freshwater Algae of North
America: Ecology and Classification. Elsevier Science, pp. 253‐309. [BIOLOGY section] Stubbins A. (2009) Virginia Coastal Energy Research Consortium Final Report: Algal Biodiesel Studies, July 2007 to September 2009. Vanormelingen P., Hegewald E., Braband A., Kitschke M., Friedl T., Sabbe K., Vyverman W. (2007) The systematics of a small
spineless Desmodesmus species, D. costato‐granulatus (Sphaeropleales, Chlorophyceae), based on its rDNA sequence analyses and cell wall morphology. Journal of Phycology 43: 378‐396. . [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 99 of 258
8.1.11 Chlorella sp.
Figure 27‐ Chlorella emersonii, left picture control culture, right oicture N‐starved culture
D. Reinecke, BGU
SYMBOLS: B, D, E, H, PIV
TAXONOMY
Phylum Chlorophyta Class Trebouxiophyceae Order Chlorellales Family Chlorellaceae Genus Chlorella Species
Related Species
Numbers of names and species: There are 71 species (and infraspecific) names in the database at present, of which 32 have been flagged as currently accepted taxonomically.
C. acuminata, C. angustoellipsoidea, C. anitrata, C. anitrata var. minor, C. antartica, C. aureoviridis, C. autotrophica, C. botryoides, C. candida, C. capsulata, C. communis, C. conductrix, C. conglomerata, C. desiccata, C. ellipsoidea var. minor, C. ellipsoidea, C. emersonii var. rubescens, C. emersonii var. globosa, C. emersonii, C. faginea, C. fusca var. rubescens, C. fusca var. vacuolata, C. fusca, C. glucotropha, C. homosphaera, C. kessleri, C. kolkwitzii, C. lobophora, C. luteo‐viridis var. lutescens, C. luteo‐viridis, C. marina, C. miniata, C. minor, C. minutissima, C. mirabilis, C. mucosa, C. mutabilis, C. nocturna, C. oocystoides, C. ovalis, C. parasitica, C. parva, C. peruviana, C. protothecoides, C. protothecoides var. mannophila, C. pyrenoidosa var. tumidus, C. pyrenoidosa, C. pyrenoidosa var. duplex, C. regularis var. minima, C. reisiglii, C. reniformis, C. rugosa, C. saccharophila, C. saccharophila var. ellipsoidea, C. salina, C. sorokiniana, C. spaerckii, C. sphaerica, C. stigmatophora, C. subsphaerica, C. terricola, C. trebouxioides, C. variabilis, C. vulgaris f. minuscula, C. vulgaris f. suboblonga, C. vulgaris f. globosa, C. vulgaris var. viridis, C. vulgaris var.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 100 of 258
autotrophica, C. vulgaris var. tertia, C. vulgaris, C. zofingiensis.
BIOLOGY
Structural and morphological features
Chlorella is a large and diverse genus of small unicellular green algae of highest relevance to multiple aspects of biotechnology.
Cells are round or ellipsoidal in shape. Smooth rigid cellulosic cell wall contains contains glucosamine (chitosan). Nucleus is single and eccentric, chloroplast single and parietal, pyrenoid single and covered with starch envelope. Pyrenoid stroma penetrated with 2 or 3 closely appressed thylakoids. Only asexual reproduction by autospores is known, autospores (2‐16 per mother cell) are released by rupture of parental cell wall. Chlorella, essentially cosmopolitan, occurs in both freshwater and marine habitats. Traditionally, the genus Chlorella has been comprised over 100 species; nevertheless, 10 well established species were recognized by chemotaxonomic methods (Kessler and Huus, 1992). Based on modern polyphasic approach, only four: C. vulgaris Beijerinck, C. lobophora Andreyeva, C. sorokiniana Shihira & Krauss, and C. kessleri Fott & Nováková (Huus et al., 1999) or even three: C. vulgaris, C. lobophora and C. sorokiniana (Krienitz et al., 2004) “true” Chlorella species are recognized recently.
Strains of speciel interest
Chlorella are fast growing freshwater, in some cases marine, water algae, reported to accumulate high concentration of oil under stress (Demirbas, 2009). Furthermore Chlorella strains can express a variety of carotenoids, among them astaxanthin. Some species is characterized by a very high growth rate (µmax = 0.20/h) and tolerance to a high culture temperature (40oC). Its desirable technological properties (resistance to shear stress, low adhesion to a surface of the bioreactor, low tendency to form aggregates) are expected to offer significant advantages for its use in large‐scale production bioreactors (Doucha and Lívanský, 2009). Its ability to grows under high CO2 permits with a direct supply of flue gas containing up to 40% (v/v) of CO2 (Doucha and Lívanský, 2005; Douskova et al., 2009, 2010).
Chlorella vulgaris is a fast growing species and some strains can accumulate very high concentrations of lipids under stress (Francisco et al., 2010; Hsieh and Wu, 2009; Ly et al., 2010; Liang et al., 2009) while another othersones accumulate high amount of starch (Doušková et al., 2010). Oil production was tested also in other Chlorella species and strains as termophilic alga Chlorella sorokiana, or heterotrophically grown Chlorella protothecoides (Xu et al., 2006). Chlorella zofingiensis, can accumulate astaxanthin and lutein (Del Campo et al., 2004; Liu et al., 2010a,b; Yp et al., 2004).
Biochemical composition and biomass main constituents
The biochemical composition and the essential aminoacid content of Chlorella biomass are repoted in Tables 12 and 13.
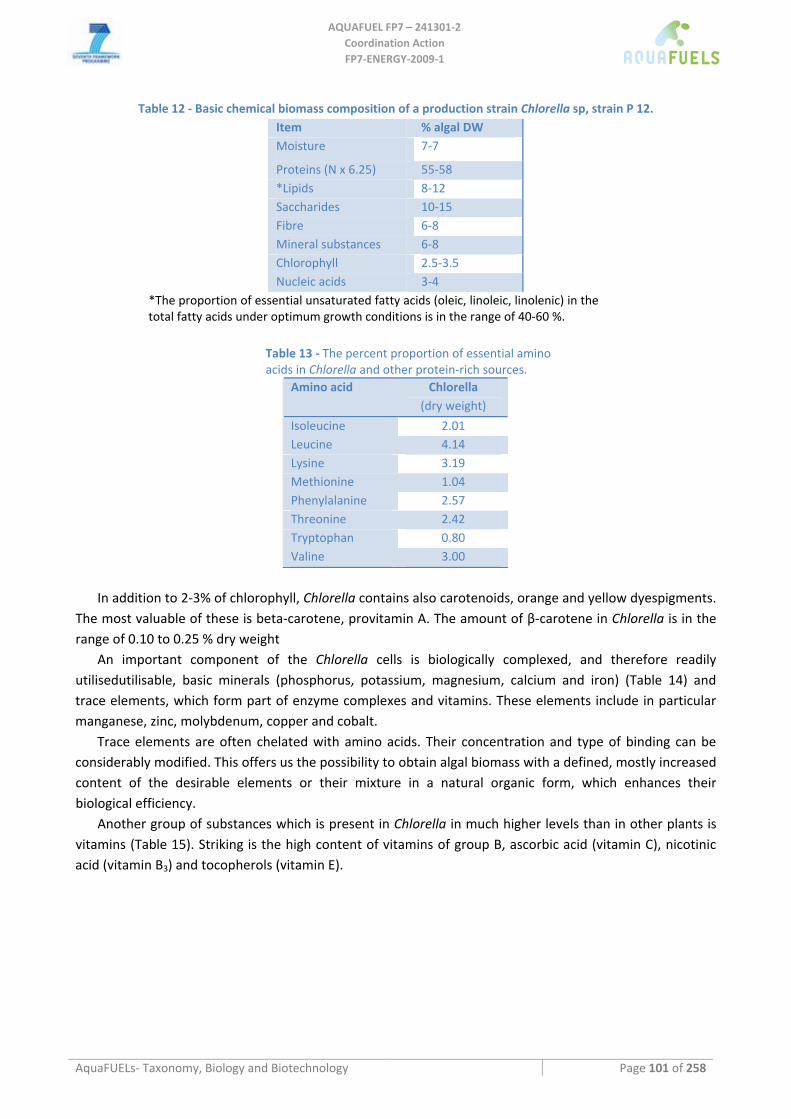
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 101 of 258
Table 12 ‐ Basic chemical biomass composition of a production strain Chlorella sp, strain P 12. Item % algal DW Moisture 7‐7
Proteins (N x 6.25) 55‐58 *Lipids 8‐12 Saccharides 10‐15 Fibre 6‐8 Mineral substances 6‐8 Chlorophyll 2.5‐3.5 Nucleic acids 3‐4
*The proportion of essential unsaturated fatty acids (oleic, linoleic, linolenic) in the total fatty acids under optimum growth conditions is in the range of 40‐60 %.
Table 13 ‐ The percent proportion of essential amino acids in Chlorella and other protein‐rich sources.
Amino acid Chlorella (dry weight)
Isoleucine 2.01 Leucine 4.14 Lysine 3.19 Methionine 1.04 Phenylalanine 2.57 Threonine 2.42 Tryptophan 0.80 Valine 3.00
In addition to 2‐3% of chlorophyll, Chlorella contains also carotenoids, orange and yellow dyespigments.
The most valuable of these is beta‐carotene, provitamin A. The amount of β‐carotene in Chlorella is in the range of 0.10 to 0.25 % dry weight
An important component of the Chlorella cells is biologically complexed, and therefore readily utilisedutilisable, basic minerals (phosphorus, potassium, magnesium, calcium and iron) (Table 14) and trace elements, which form part of enzyme complexes and vitamins. These elements include in particular manganese, zinc, molybdenum, copper and cobalt.
Trace elements are often chelated with amino acids. Their concentration and type of binding can be considerably modified. This offers us the possibility to obtain algal biomass with a defined, mostly increased content of the desirable elements or their mixture in a natural organic form, which enhances their biological efficiency.
Another group of substances which is present in Chlorella in much higher levels than in other plants is vitamins (Table 15). Striking is the high content of vitamins of group B, ascorbic acid (vitamin C), nicotinic acid (vitamin B3) and tocopherols (vitamin E).
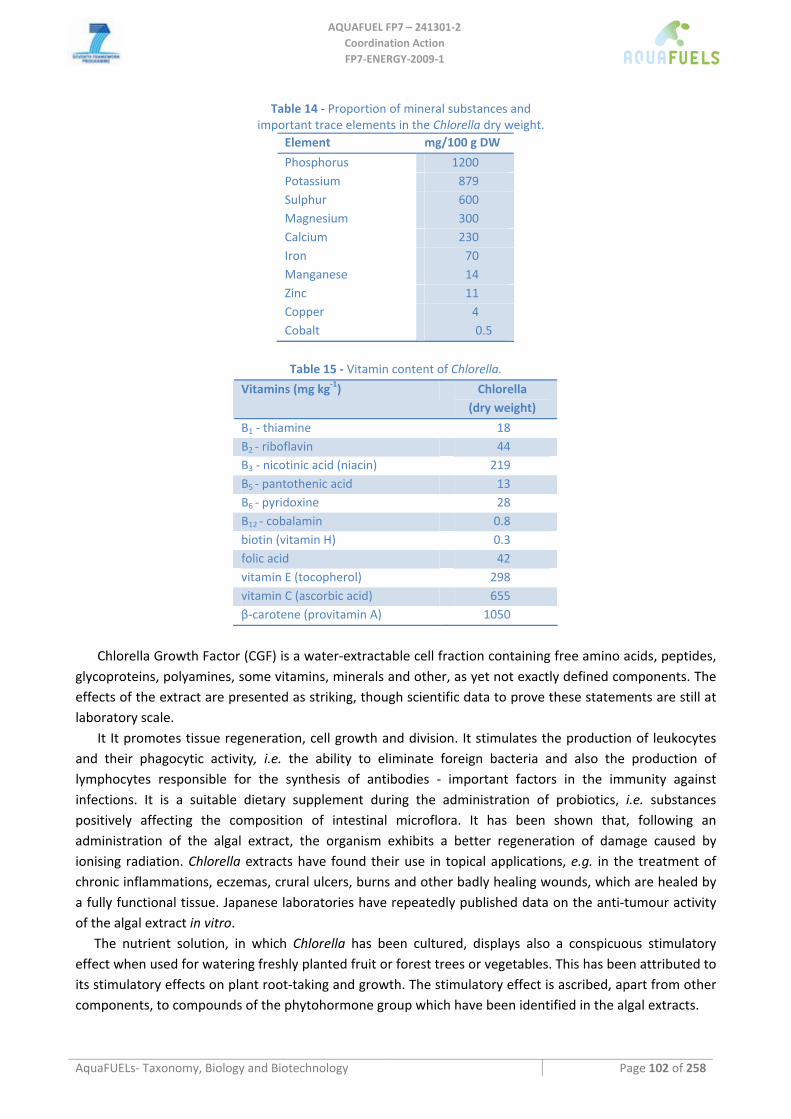
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 102 of 258
Table 14 ‐ Proportion of mineral substances and important trace elements in the Chlorella dry weight.
Element mg/100 g DW
Phosphorus 1200 Potassium 879 Sulphur 600 Magnesium 300 Calcium 230 Iron 70 Manganese 14 Zinc 11 Copper 4 Cobalt 0.5
Table 15 ‐ Vitamin content of Chlorella.
Vitamins (mg kg‐1) Chlorella (dry weight)
B1 ‐ thiamine 18 B2 ‐ riboflavin 44 B3 ‐ nicotinic acid (niacin) 219 B5 ‐ pantothenic acid 13 B6 ‐ pyridoxine 28 B12 ‐ cobalamin 0.8 biotin (vitamin H) 0.3 folic acid 42 vitamin E (tocopherol) 298 vitamin C (ascorbic acid) 655 β‐carotene (provitamin A) 1050
Chlorella Growth Factor (CGF) is a water‐extractable cell fraction containing free amino acids, peptides,
glycoproteins, polyamines, some vitamins, minerals and other, as yet not exactly defined components. The effects of the extract are presented as striking, though scientific data to prove these statements are still at laboratory scale.
It It promotes tissue regeneration, cell growth and division. It stimulates the production of leukocytes and their phagocytic activity, i.e. the ability to eliminate foreign bacteria and also the production of lymphocytes responsible for the synthesis of antibodies ‐ important factors in the immunity against infections. It is a suitable dietary supplement during the administration of probiotics, i.e. substances positively affecting the composition of intestinal microflora. It has been shown that, following an administration of the algal extract, the organism exhibits a better regeneration of damage caused by ionising radiation. Chlorella extracts have found their use in topical applications, e.g. in the treatment of chronic inflammations, eczemas, crural ulcers, burns and other badly healing wounds, which are healed by a fully functional tissue. Japanese laboratories have repeatedly published data on the anti‐tumour activity of the algal extract in vitro.
The nutrient solution, in which Chlorella has been cultured, displays also a conspicuous stimulatory effect when used for watering freshly planted fruit or forest trees or vegetables. This has been attributed to its stimulatory effects on plant root‐taking and growth. The stimulatory effect is ascribed, apart from other components, to compounds of the phytohormone group which have been identified in the algal extracts.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 103 of 258
Gross composition under optimal and stressed conditions
A complex treatment of agricultural waste including the following major steps: anaerobic fermentation of suitable waste, cogeneration of the obtained biogas and growth of microalgae consuming the CO2 from biogas or flue gas, was verified under field conditions in a pilot‐scale photobioreactor. Laboratory analyses of the produced microalgae confirmed that it meets the strict EU criteria for relevant contaminants level in foodstuff (Doušková et al., 2009; 2010a; Kaštánek et al., 2010).
The freshwater alga Chlorella, a highly productive source of starch, might substitute for starch‐rich terrestrial plants in bioethanol production. Cheap enhanced starch biomass can be produced from highly productive Chlorella cultures grown in suitable outdoor photobioreactors in which the photosynthetic carbon dioxide source is derived from combustion of organic waste, fermentation processes or other sources (Doucha et al., 2005; Douskova et al., 2009; Mann et al., 2009). This characteristic enhances the ecological and economic impact of the proposed technology, because of its potential to bioremediate carbon dioxide emissions from different CO2 sources including waste incinerators, power stations, limekilns, cogeneration units, etc. in situ.
BIOTECHNOLOGY
Culture media
Chlorella was one of the first algae isolated as a pure culture by Beijerinck in 1890. Since the half of the last century, attention has been drawn towards its potential for autotrophic mass cultivation. To this purpose, many mineral media were developed based in principle on the chemical composition of the algal cells. The basic inorganic elements used are: N, P, K, Mg, Ca, S, Fe, Cu, Mn, Mo and Zn (Krauss, 1958; O´Kelly, 1968). Many formulas are used for the cultivation of this genus ‐ see Hama and Miyachi (1988). For high‐yielded production of Chlorella outdoors an optimized composition of nutrient solution was proposed by Doucha and Lívanský (2006).
The mineral medium was based on the mean content of P, N, K, Mg, and S in algal biomass and had the following initial composition (mg/L): 1100 (NH2)2CO, 237 KH2PO4, 204 MgSO4.7H2O, 40 C10H12O8N2NaFe, 88 CaCl2, 0.83 H3BO3, 0.95 CuSO4.5H2O, 3.3 MnCl2.4H2O, 0.17 (NH4)6Mo7O24.4H2O, 2.7 ZnSO4.7H2O, 0.6 CoSO4.7H2O, 0.014 NH4VO3 in distilled water.
Cultivation of Chlorella heterotrophically in fermenters is also used (Lee et al., 1997). Heterotrophic culture may provide a cost effective, large‐scale alternative method for cultivation of some microalgae that can utilize organic carbon substances as their sole carbon and energy source (Chen and Chen, 2006). Nutrient solution containing glucose or acetate as a source of carbon for intensive growth of Chlorella in fermenter was described by Endo and Shirota (1972) and lately by Doucha and Lívanský (Cz. patent. 288638, 2001). Technology of heterotrophic Chlorella cultivation has been commercially used for production of biomass (Doucha and Lívanský, 2011) for food and feed as well as for production of biomass enriched by organically bound selenium (Doucha et al., 2009;, Skřivan et al., 2006; Trávníček et al., 2008) or as a source of lipids for production of biodiesel (Xiong et al., 2008).
Cultivation methods and production systems
There is still a noticeable discrepancy between the extent of commercially operated algal cultures and the potential of algae. Since the first experiments with large‐scale algal cultures in the 1950s (Burlew, 1953) many types of culture equipment have been developed (Stengel, 1970; Richmond and Becker, 1986; Tredici, 2004). For most products of microalgal mass cultivation outdoor open circular or “raceway” ponds with a 15‐30 cm layer of algal suspension are the most used technology for the growth of algae
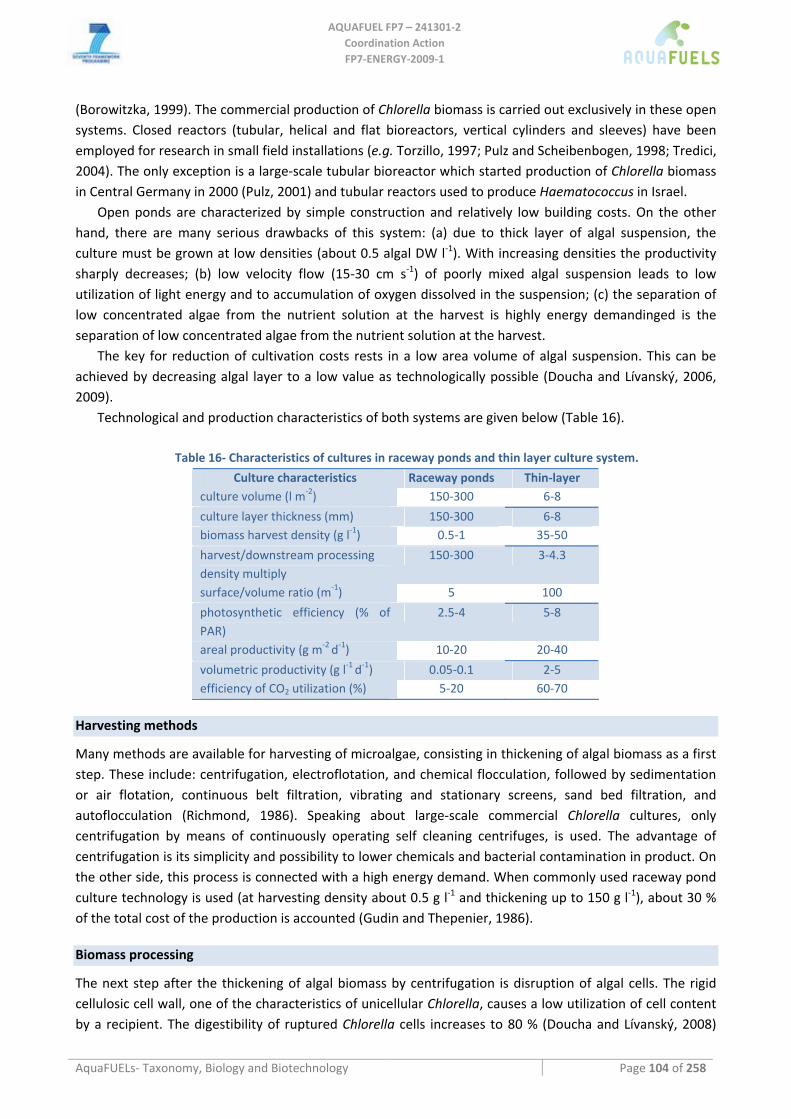
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 104 of 258
(Borowitzka, 1999). The commercial production of Chlorella biomass is carried out exclusively in these open systems. Closed reactors (tubular, helical and flat bioreactors, vertical cylinders and sleeves) have been employed for research in small field installations (e.g. Torzillo, 1997; Pulz and Scheibenbogen, 1998; Tredici, 2004). The only exception is a large‐scale tubular bioreactor which started production of Chlorella biomass in Central Germany in 2000 (Pulz, 2001) and tubular reactors used to produce Haematococcus in Israel.
Open ponds are characterized by simple construction and relatively low building costs. On the other hand, there are many serious drawbacks of this system: (a) due to thick layer of algal suspension, the culture must be grown at low densities (about 0.5 algal DW l‐1). With increasing densities the productivity sharply decreases; (b) low velocity flow (15‐30 cm s‐1) of poorly mixed algal suspension leads to low utilization of light energy and to accumulation of oxygen dissolved in the suspension; (c) the separation of low concentrated algae from the nutrient solution at the harvest is highly energy demandinged is the separation of low concentrated algae from the nutrient solution at the harvest.
The key for reduction of cultivation costs rests in a low area volume of algal suspension. This can be achieved by decreasing algal layer to a low value as technologically possible (Doucha and Lívanský, 2006, 2009).
Technological and production characteristics of both systems are given below (Table 16).
Table 16‐ Characteristics of cultures in raceway ponds and thin layer culture system.
Culture characteristics Raceway ponds Thin‐layer culture volume (l m‐2) 150‐300 6‐8
culture layer thickness (mm) 150‐300 6‐8 biomass harvest density (g l‐1) 0.5‐1 35‐50
harvest/downstream processing density multiply
150‐300 3‐4.3
surface/volume ratio (m‐1) 5 100
photosynthetic efficiency (% of PAR)
2.5‐4 5‐8
areal productivity (g m‐2 d‐1) 10‐20 20‐40
volumetric productivity (g l‐1 d‐1) 0.05‐0.1 2‐5 efficiency of CO2 utilization (%) 5‐20 60‐70
Harvesting methods
Many methods are available for harvesting of microalgae, consisting in thickening of algal biomass as a first step. These include: centrifugation, electroflotation, and chemical flocculation, followed by sedimentation or air flotation, continuous belt filtration, vibrating and stationary screens, sand bed filtration, and autoflocculation (Richmond, 1986). Speaking about large‐scale commercial Chlorella cultures, only centrifugation by means of continuously operating self cleaning centrifuges, is used. The advantage of centrifugation is its simplicity and possibility to lower chemicals and bacterial contamination in product. On the other side, this process is connected with a high energy demand. When commonly used raceway pond culture technology is used (at harvesting density about 0.5 g l‐1 and thickening up to 150 g l‐1), about 30 % of the total cost of the production is accounted (Gudin and Thepenier, 1986).
Biomass processing
The next step after the thickening of algal biomass by centrifugation is disruption of algal cells. The rigid cellulosic cell wall, one of the characteristics of unicellular Chlorella, causes a low utilization of cell content by a recipient. The digestibility of ruptured Chlorella cells increases to 80 % (Doucha and Lívanský, 2008)

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 105 of 258
compared to 15‐25 % for unruptured cells (Becker, 1984). To open the cell, methods of freezing, alcalic alkaline and organic solvents, osmotic shocks, sonication, high‐pressure homogenization and bead milling were tested (Molina Grima et al., 2004). For large‐scale processing of Chlorella cultures disintegration of cell walls by bead mills is mostly used (Middelberg, 1995; Doucha and Lívanský, 2008).
Spray drying as the end step of the downstream processing process is the most extended method for dehydration of ruptured Chlorella cells. Algae biomass is dispersed into very fine droplets, whose surface temperature in the course of several seconds of drying does not surpass 60 oC. Thus, the process is very considerate and the quality of the product is high.
Scaling up
The first commercial production of Chlorella and, some years later, blue‐green Spirulina cultures started in Japan and Taiwan in the 1960’s. Nowadays, large‐scale production plants are located also in the USA, China, India, Thailand, Indonesia, Germany and other countries (Borowitzka, 1996; Lee, 1997; Pulz , 2001; Borowitzka 1996; Tredici, 2004; Spolaore et al., 2006). Their per year ‐ world production is estimated to be about 8000 tons (Spirulina) and 5000 tons (Chlorella). Most of the products have been used in human nutrition.
High growth rate, high photosynthetic efficiency, relatively high content of energy‐rich chemicals on one side and experience with large‐scale culture and downstream processing technologies concentrate in last decades increasing attention on microalgae as a feedstock for biofuels. Today projects dealing with algae are focused almost entirely on biodiesel production. Nevertheless, algal strains containing higher amount of lipids are characterized by low growth rate. Slow growth increases the operational costs and demands cultivation in closed bioreactors whose building is expensive.
On the other side for economical production of bioethanol, relatively cheap biomass of high‐yielded Chlorella cultures, containing enhanced amount of starch grown in suitable open bioreactors is perspective solution. To produce starch economically, conditions for culturing starch‐enriched algae in dense cultures must be attained. Using a thin layer algal suspension in outdoor cultures, linear growth continues up to very high biomass concentrations (about 40 g/L) enabling easy and cheap harvesting and processing (Doucha and Lívanský, 2006, 2009). However, the content of starch in the biomass is low (15% of DW or less).
The conditions under which starch content increases in commercially produced algal biomass to a level that would be viable for bioethanol production can be achieved if the processes and events during which starch is extensively degraded are slowed down, or stopped completely, while the factors supporting starch synthesis, namely light intensity, remain sustainable (Figure 27) (Brányiková et al., 2011). Similarly, the Chlorella species producing oil as their energy reserves rather than starch can be similarly treated to markedly increase their oil content (Figure 28).
An increase in the production of starch in sulphur‐limited culture up to a maximum of 50% starch content of algal biomass (DW) was demonstrated under field conditions using the outdoor scale up, thin‐layer solar photobioreactor.
While use of algae with enriched starch content is conventional for bioethanol production, another attractive exploitation of starch from algae might be the production of hydrogen, which may be realized in the near future (Miura et al., 1982; Tsygankov et al., 2002; Melis et al., 2004; Chochois et al., 2009; Melis et al. 2004; Miura et al. 1982; Tsygankov et al. 2002). Sulphur limitation could be one of the ways to support hydrogen production (Melis et al., 2000; Zhang et al., 2002). It has been shown recently that some strains of Chlorella can produce and accumulate significant volume of hydrogen gas under anaerobic conditions and sulphur deprivation such as it is reported in literature using C. reinhardtii (Chader et al., 2009).
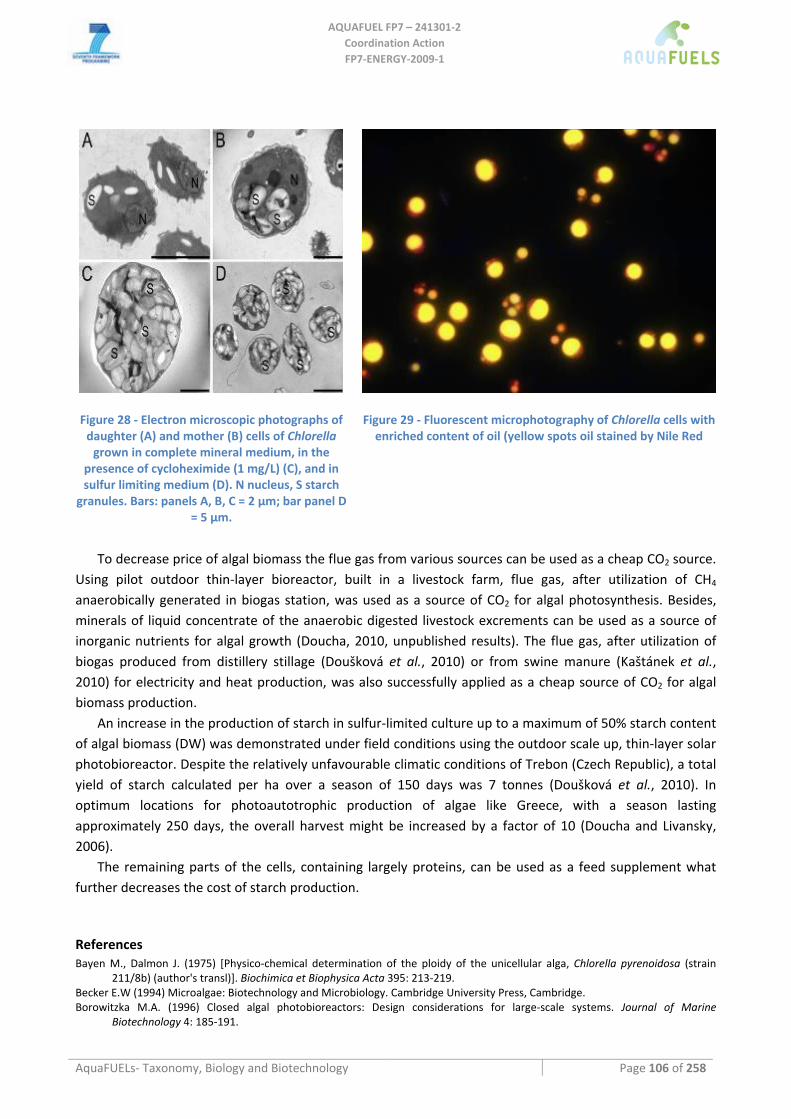
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 106 of 258
Figure 28 ‐ Electron microscopic photographs of daughter (A) and mother (B) cells of Chlorella grown in complete mineral medium, in the
presence of cycloheximide (1 mg/L) (C), and in sulfur limiting medium (D). N nucleus, S starch
granules. Bars: panels A, B, C = 2 μm; bar panel D = 5 μm.
Figure 29 ‐ Fluorescent microphotography of Chlorella cells with enriched content of oil (yellow spots oil stained by Nile Red
To decrease price of algal biomass the flue gas from various sources can be used as a cheap CO2 source.
Using pilot outdoor thin‐layer bioreactor, built in a livestock farm, flue gas, after utilization of CH4 anaerobically generated in biogas station, was used as a source of CO2 for algal photosynthesis. Besides, minerals of liquid concentrate of the anaerobic digested livestock excrements can be used as a source of inorganic nutrients for algal growth (Doucha, 2010, unpublished results). The flue gas, after utilization of biogas produced from distillery stillage (Doušková et al., 2010) or from swine manure (Kaštánek et al., 2010) for electricity and heat production, was also successfully applied as a cheap source of CO2 for algal biomass production.
An increase in the production of starch in sulfur‐limited culture up to a maximum of 50% starch content of algal biomass (DW) was demonstrated under field conditions using the outdoor scale up, thin‐layer solar photobioreactor. Despite the relatively unfavourable climatic conditions of Trebon (Czech Republic), a total yield of starch calculated per ha over a season of 150 days was 7 tonnes (Doušková et al., 2010). In optimum locations for photoautotrophic production of algae like Greece, with a season lasting approximately 250 days, the overall harvest might be increased by a factor of 10 (Doucha and Livansky, 2006).
The remaining parts of the cells, containing largely proteins, can be used as a feed supplement what further decreases the cost of starch production.
References Bayen M., Dalmon J. (1975) [Physico‐chemical determination of the ploidy of the unicellular alga, Chlorella pyrenoidosa (strain
211/8b) (author's transl)]. Biochimica et Biophysica Acta 395: 213‐219. Becker E.W (1994) Microalgae: Biotechnology and Microbiology. Cambridge University Press, Cambridge. Borowitzka M.A. (1996) Closed algal photobioreactors: Design considerations for large‐scale systems. Journal of Marine
Biotechnology 4: 185‐191.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 107 of 258
Borowitzka M.A. (1999) Commercial production of microalgae: ponds, tanks, tubes and fermenters. Journal of Biotechnology 70: 313‐321.
Brányiková I., Maršálková B., Doucha J., Brányik T., Bišová K., Zachleder V., Vítová V. (2011) Microalgae ‐ novel highly‐efficient starch producers. Biotechnology and Bioengineering 108: 766‐776.
Burlew J.S. (ed.) (1953) Algal Culture from Laboratory to Pilot Plant. Carnegie Institute of Washington, DC. Chader S., Hacene H., Agathos S.N. (2009) Study of hydrogen production by three strains of Chlorella isolated from the soil in the
Algerian Sahara. International Journal of Hydrogen Energy 34: 4941‐4946. Chen G.Q., Chen F. (2006) Growing phototrophic cells without light. Biotechnology Letters 28: 607‐616. Chochois V., Dauvillée D., Beyly A., Tolleter D., Cuiné S., Timpano H., Ball S., Cournac L., Peltier G. (2009) Hydrogen production in
Chlamydomonas: photosystem II‐dependent and independent pathways differ in their requirement for starch metabolism. Plant Physiology 151: 631‐640
Del Campo J.A., Rodriguez H., Moreno J., Vargas M.A., Rivas J., Guerrero M.G. (2004) Accumulation of astaxanthin and lutein in Chlorella zofingiensis (Chlorophyta). Applied Microbiology and Biotechnology 64: 848‐854.
Demirbas A. (2009) Production of biodiesel from algae oils. Energy Sources, Part A: Recovery, utilization, and environmental effects 31: 163‐168.
Doucha J., Lívanský K. (2001) Method of controlled cultivation of algae in heterotrophic mode of nutrition. Czech Patent 288638. Doucha J., Lívanský K. (2006) Productivity, CO2/O2 exchange and hydraulics in outdoor open high density microalgal (Chlorella sp.)
photobioreactors operated in a Middle and Southern European climate. Journal of Applied Phycology 18: 811‐826. Doucha J., Lívanský K. (2009) Outdoor open thin‐layer microalgal photobioreactor: potential productivity. Journal of Applied
Phycology 21: 111‐117. Doucha J., Lívanský K. (2008) Influence of processing parameters on disintegration of Chlorella cells in various types of
homogenizers. Applied Microbiology and Biotechnology 81: 131‐440. Doucha J, Lívanský K. (2011) Production of high‐density Chlorella culture grown in fermenters. Journal of Applied Phycology DOI
10.1007/s10811‐010‐9643‐2. Doucha J., Straka F., Livansky K. (2005) Utilization of flue gas for cultivation of microalgae (Chlorella sp.) in an outdoor open thin‐
layer photobioreactor. Journal of Applied Phycology 17: 403‐412. Doucha J., Lívanský K., Kotrbáček V., Zachleder V. (2009) Production of Chlorella biomass enriched by selenium and its use in animal
nutrition: a review. Applied Microbiology and Biotechnology 83: 1001‐1008. Doušková I., Doucha J., Livansky K., Machat J., Novak P., Umysova D., Zachleder V., Vitova M. (2009) Simultaneous flue gas
bioremediation and reduction of microalgal biomass production costs. Applied Microbiology and Biotechnology 82: 179‐185. Doušková I., Kaštánek F., Maléterová Y., Kaštánek P., Doucha J., Zachleder V. (2010) Utilization of distillery stillage for energy
generation and concurrent production of valuable microalgal biomass in the sequence: Biogas‐cogeneration‐microalgae‐products. Energy Conversion and Management 51: 606‐611.
Endo H., Shirota M. (1972) Studies on the heterotrophic growth of Chlorella in a mass culture. Proceedings IV IFS: Fermentation Technology Today, pp. 533‐541.
Francisco É.C., Neves D.B., Jacob‐Lopes E., Franco T.T. (2010) Microalgae as feedstock for biodiesel production: Carbon dioxide sequestration, lipid production and biofuel quality. Journal of Chemical Technology and Biotechnology 85: 395‐403.
Guckert J.B., Cooksey K.E. (1990) Triglyceride accumulation and fatty acid profile changes in Chlorella (Chlorophyta) during high pH‐induced cell cycle inhibition. Journal of Phycology 26: 72‐79.
Gudin C., Thepenier C. (1986) Bioconversion of solar energy into organic chemicals by microalgae. Advances in Biotechnology Processes 6: 73‐110.
Hsieh C.H., Wu W.T. (2009) Cultivation of microalgae for oil production with a cultivation strategy of urea limitation. Bioresource Technology 100: 3921‐3926.
Huss V.A.R., Frank C., Hartmann E.C., Hirmer M., Kloboucek A., Seidel B.M., Wenzeler P., Kessler E. (1999) Biochemical taxonomy and molecular phylogeny of the genus Chlorella sensu lato (Chlorophyta). Journal of Phycology 35: 587‐598.
Kaštánek F., Šabata S., Šolcová O., Maléterová .Y, Kaštánek .P, Brányiková I., Kuthan K., Zachleder V. (2010) In‐field experimental verificatiom of cultuvation of microalgae Chlorella sp. using the flue gas from a cogeneration unit as a source of carbon dioxide. Waste Management and Research 28: 961‐966.
Kessler E., Huss V.A.R. (1992) Comparative physiology and biochemistry and taxonomic assignment of the Chlorella (Chlorophyceae) strains of the Culture Collection of the University of Texas at Austin. Journal of Phycology 28: 550‐553.
Krauss R.W. (1958) Physiology of the fresh‐water algae. Annual Review of Plant Physiology 9: 207‐244. Krienitz L.., Hegewald E.H., Hepperle D., Huss V.A.R., Rohr T., Wolf M. (2004) Phylogenetic relationship of Chlorella and
Parachlorella gen. nov. (Chlorophyta, Trebouxiophyceae). Phycologia 43: 529‐542. Krusteva N.G., Tomova N.G., Georgieva M.A. (1984) Allosteric regulation of NAD(NADP)‐dependent glyceraldehyde‐3‐phosphate
dehydrogenase from Chlorella by α‐amino acids, dithiothreitol and ATP. FEBS Letters 171: 137‐140. Lee Y.K. (1997) Commercial production of microalgae in the Asia‐Pacific rim. Journal of Applied Phycology 9: 403‐411. Lee Y.K. (2001) Microalgal mass culture systems and methods: Their limitation and potential. Journal of Applied Phycology 13: 307‐
315. Li X., Xu H., Wu Q. (2007) Large‐scale biodiesel production from microalga Chlorella protothecoides through heterotrophic
cultivation in bioreactors. Biotechnology and Bioengineering 98: 764‐771. Liang Y., Sarkany N., Cui Y. (2009) Biomass and lipid productivities of Chlorella vulgaris under autotrophic, heterotrophic and
mixotrophic growth conditions. Biotechnology Letters 31: 1043‐1049. Liu J., Huang J., Fan K.W., Jiang Y., Zhong Y., Sun Z., Chen F. (2010a) Production potential of Chlorella zofingienesis as a feedstock for
biodiesel. Bioresource Technology 101: 8658‐8663.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 108 of 258
Liu J., Zhong Y., Sun Z., Huang J., Sandmann G., Chen F. (2010b) One amino acid substitution in phytoene desaturase makes Chlorella zofingiensis resistant to norflurazon and enhances the biosynthesis of astaxanthin. Planta 232: 61‐67.
Mann G., Schlegel M., Schumann R., Sakalauskas A. (2009) Biogas‐conditioning with microalgae. Agronomy Research 7: 33‐38. Melis A., Seibert M., Happe T. (2004) Genomics of green algal hydrogen research. Photosynthesis Research 82: 277‐288. Melis A., Zhang L., Forestier M., Ghirardi M.L., Seibert M. (2000) Sustained photobiological hydrogen gas production upon
reversible inactivation of oxygen evolution in the green alga Chlamydomonas reinhardtii. Plant Physiology 122: 127‐135. Middelberg A.P.J. (1995) Process‐scale disrruption of microorganisms. Biotechnology Advances 13: 491‐555. Miura Y., Yagi K., Shoga M., Miyamoto K. (1982) Hydrogen production by a green alga, Chlamydomonas reinhardtii, in an alternating
light/dark cycle. Biotechnology and Bioengineering 24: 1555‐1563. Molina Grima E., Fernández F.G.A., Medina A.R. (2004) Downstream processing of cell‐mass and products. In: Richmond A. (ed.)
Handbook of Microalgal Culture: Biotechnology and Applied Phycology. Blackwell Science, pp 215‐251. O´Kelly J.C. (1968) Mineral nutrition of algae. Annual Review of Plant Physiology 19: 89‐112. Oh‐hama T., Miyachi S. (1988) Chlorella. In: Borowitzka M.A., Borowitzka L.J. (eds.), Micro‐algal Biotechnology. Cambridge
University Press, Cambridge, pp. 3‐26. Pulz O. (2001) Photobioreactors: Production systems for phototrophic microorganisms. Applied Microbiology and Biotechnology 57:
287‐293. Pulz O., Scheibenbogen K. (1998) Photobioreactors: Design and performance with respect to light energy input. In: Scheper T. (ed.)
Advances in Biochemical Engineering/Biotechnology, Springer‐Verlag, Berlin, pp. 123‐152. Richmond A. (ed.) (1986) CRC Handbook of Microalgal Mass Culture. CRC Prsss, Boca Raton, Florida. Richmond A., Becker E.W. (1986) Technological aspects of mass cultivation – a general outlook. In: Richmond A. (ed.) CRC
Handbook of Microalgal Mass Culture. CRC Press, Boca Raton, Florida, pp 245‐263. Rodolfi L., Chini Zitteli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: Strain selection,
induction of lipid synthesis and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Sauer N., Tanner W. (1989) The hexose carrier from Chlorella. cDNA cloning of a eukaryotic H+‐cotransporter. FEBS Letters 259: 43. Skřivan M., Šimáně J., Dlouhá G., Doucha J. (2006) Effect of dietary sodium selenite, Se‐enriched yeast and Se‐enriched Chlorella on
egg Se concentration, physical parameters of eggs and laying hen production. Czech Journal of Animal Science 51: 163‐167. Spolaore P., Joannis‐Cassan C., Duran E., Isambert A. (2006) Commercial application of microalgae. Journal of Bioscience and
Bioengineering 101: 87‐96. Stengel E. (1970) Anlagentype und Verfahren der technischen Algenmassenproduktion. Berichte der Deutschen Botanische
Gesellschaft 83: 589‐606. Syrett P.J., Thomas E.M. (1973) The assay of nitrate reductase in whole cells of Chlorella: Strain differences and the effect of cell
walls. New Phytologist 72: 1307‐1310. Torzillo G. (1997) Tubular bioreactors. In: Vonshak A (ed.) Spirulina platensis (Arthrospira): Physiology, Cell Biology and
Biotechnology. Taylor & Francis, London, pp 101‐115. Trávníček J., Racek J., Trefil L., Rodinová H., Kroupová V., Illek J., Doucha J., Písek L. (2008) Activity of glutathione peroxidase (GSH‐
Px) in the blood of ewes and their lambs receiving the selenium‐enriched unicellular alga Chlorella. Czech Journal of Animal Science 53: 292–298.
Tredici M.R. (2004) Mass Production of Microalgae: Photobioreactors. In: Richmond A. (ed.) Handbook of Microalgal Culture. Blackwell Science Ltd, Oxford, pp 178‐214.
Tsygankov A., Kosourov S., Seibert M., Ghirardi M.L. (2002) Hydrogen photoproduction under continuous illumination by sulfur‐deprived, synchronous Chlamydomonas reinhardtii cultures. International Journal of Hydrogen Energy 27: 1239‐1244.
Xiong W., Li X., Xiang F., Wu Q. (2008) High density fermentation of micro alga Chlorella protothecoides in bioreactor for microbio‐diesel production. Applied Microbiology and Biotechnology 78: 29‐36.
Xu H., Miao X., Wu Q. (2006) High quality biodiesel production from a microalga Chlorella protothecoides by heterotrophic growth in fermenters. Journal of Biotechnology 126: 499‐507.
Yp J.M., Cheng L.H., Xu X.H., Zhang L., Chen H.L. (2010) Enhanced lipid production of Chlorella vulgaris by adjustment of cultivation conditions. Bioresource Technology 101: 6797‐6804.
Yp P.F., Wong K.H., Chen F. (2004) Enhanced production of astaxanthin by the green microalga Chlorella zofingiensis in mixotrophic culture. Process Biochemistry 39: 1761‐1766.
Zhang L., Happe T., Melis A. (2002) Biochemical and morphological characterization of sulfur‐deprived and H2‐producing Chlamydomonas reinhardtii (green alga). Planta 214: 552‐561.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 109 of 258
8.1.12 Parietochloris incisa
Figure 30 – Parietochloris incisa in balanced growth (left) and nitrogen starved (right)
I. Khozin, BGU
SYMBOLS: D, PIV
TAXONOMY
Phylum Chlorophyta Class Trebouxiophyceae Order Chlorellales Family Chlorellaceae Genus Parietochloris Species Parietochloris incisa
Related Species
Numbers of names and species: There are 6 species (and infraspecific) names in the database at present.
P. alveolaris, P. bilobata, P. cohaerens, P. incisa, P. ovoidea, P. pseudoalveolaris.
BIOLOGY Parietochloris incisa is a unicellular, oleaginous, freshwater alga. The alga was isolated from the slopes of a snow mountain in Japan (Watanabe et al., 1996), an alpine environment which is characterized by a broad temperature range, UV radiation and light levels that can be extremely high. Such environments were found to be a natural habitat for phototrophic microorganisms that accumulate high polyunsaturated fatty acids (PUFA) concentrations, including sea ice diatoms, dinoflagellates, and green algae. Indeed P. incisa was found to accumulate unusually high content of the long chain PUFA, arachidonic acid (AA) (Bigogno et al., 2002a).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 110 of 258
Biochemical composition
Under nitrogen starvation, the fatty acid content in P. incisa is over 35% of dry weight; AA constitutes about 60% of total fatty acids, and over 90% of cell AA is deposited in TAG (Khozin‐Goldberg et al., 2002). In the lipid‐linked pathway of its biosynthesis, AA is mainly exported to TAG and accumulated in cytoplasmic lipid bodies (Bigogno et al., 2002b). The pathway of AA biosynthesis involves stepwise desaturation and elongation of oleic acid via the so‐called ω6 pathway. The genes encoding for ∆12, ∆6, ∆5 desaturases and ∆6 PUFA elongase were cloned and their functions were validated in heterologous system (Iskandarov et al., 2009, 2010).
Furthermore, under optimal growth temperature (25 °C), AA is partially converted to the valuable PUFA eicosapentaenoic acid (EPA, 20:5ω3). This conversion is more pronounced when alga is exposed to low temperature (10‐15oC). The conversion of ω6 precursors into ω3 fatty acids is catalyzed by a group of enzymes called ω3 fatty acid desaturases (FAD) that differ by fatty acid substrate preference and cellular localization. A Δ5‐desaturase‐deficient mutant isolated at BGU (Cohen et al., 2009) is able to accumulate the high value PUFA DGLA (20:3ω6).
BIOTECHNOLOGY
Growth medium
mBG11
Growth rate
Growth rate in panel reactors and continuous illumination under nitrogen starvation: 160 mg/l per day accumulating 6% DGLA and 18% TAG.
Pilot scale scale production
The DGLA production process using the P. incisa mutant has been tested at BGU at the pilot scale using 50 – 200 l flatpanel reactors in greenhouse or climate controlled growth rooms. DGLA content 10% of total dry biomass was achieved after cultivation in N‐deficient medium demonstrating a commercial production capability.
HIGHLIGHTS IN BIOTECHNOLOGY
P. incisa and its mutant are unique among algae in being able to accumulate high concentrations of PUFA in TAG in cytoplasmic oil globules. Identification of several desaturase and elongase genes may allow breakthroughs in engineering more efficient PUFA producing algae.
References
Bigogno C., Khozin‐Goldberg I., Boussiba S., Vonshak A., Cohen Z. (2002a) Lipid and fatty acid composition of the green oleaginous alga Parietochloris incisa, the richest plant source of arachidonic acid. Phytochemistry 60: 497‐503.
Bigogno C., Khozin‐Goldberg I., Adlerstein D., Cohen Z. (2002b) Biosynthesis of arachidonic acid in the oleaginous microalga Parietochloris incisa (Chlorophyceae): radiolabeling studies. Lipids 37: 209‐216.
Cohen Z., Khozin‐Goldberg I., Boussiba S., Vonshak A.; Ben‐Gurion University of the Negev Research and Development Authority, assignee. 2009 19.02.2009. Over‐Production of dihomo gamma linolenic acid by a Mutant Strain of Parietochloris incisa. IL.
Iskandarov U., Khozin‐Goldberg I., Ofir R., Cohen Z. (2009) Cloning and characterization of the ω6 polyunsaturated fatty acid elongase from the greenm Parietochloris incisa. Lipids 44: 545‐554.
Watanabe S., Hirabayashi S., Boussiba S., Cohen Z., Vonshak A., Richmond A. (1996) Parietochloris incisa comb.nov. (Trebouxiophyceae, Chlorophyta). Phycological Research 44: 107‐108.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 111 of 258
8.1.13 Prototheca sp.
Figure 31 ‐ Light microscopic view showing Prototheca moriformis
Lactophenol cotton blue mount fixation. Picture from http://content9.eol.org/content/2009/11/25/02/75871_small.jpg
SYMBOLS: D
TAXONOMY
Phylum Chlorophyta Class Trebouxiophyceae Order Chlorellales Family Chlorellaceae Genus Prototheca Species
Related species
There are 21 species (and infraspecific) names in the database at present, of which 13 have been flagged as currently accepted taxonomically.
P. blaschkeae, P. chlorelloides, P. ciferrii, P. crieana, P. filamenta, P. hydrocarbonea, P. kruegeri, P. moriformis, P. moriformis var. betulina, P. portoricensis, P. portoricensis var. trisporus, P. salmonis, P. segbwema, P. stagnorum, P. trispora, P. ulmea, P. viscosa, P. wickerhamii, P. zopfii var. portoricensis, P. zopfii var. hydrocarbonea, P. zopfii.
BIOLOGY
The genus Prototheca is composed of microscopic achlorophyllous organisms with a life cycle similar to that of the genus Chlorella. The species typically produce thick‐walled cells (sporangia) which, at ma‐turity, divide by irregular cleavage forming 2‐15 aplanospores (endo‐spores). Following rupture of the sporangial wall, freed aplanospores enlarge and repeat the cycle. One to three percent of the sporangia cleave to form 2‐3 thick‐walled resting cells (hypnospores). No sexual cycle has been observed.
Prototheca can use dextrose, levulose, galactose, ethanol, n‐butanol, iso‐butanol, iso‐pentanol, hexanol and glycerol and some species assimilate sucrose, trehalose, n‐propanol and n‐pentanol. In general, alcohols are assimilated similarly in both liquid and vapor phases; n‐pentanol is assimilated only in the vapor phase. Species of Prototheca require thiamine.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 112 of 258
Numerous studies have reported a pathogenic potential for P. wickerhamii and P. zopfii. The cases of human protothecosis are predominantly caused by P. wickerhamii and occur as local (predominantly cutaneous) and systemic infections mainly in immune‐compromised patients, e.g. patients infected with HIV or treated with glucocorticoids. P. blaschkeae were isolated from some cases of onychomycosis. Canine protothecosis is caused by P. wickerhamii and P. zopfii, and is characterized by similar clinical symptoms as in humans. Worldwide, P. zopfii has been identified to induce a therapy‐resistant inflammation of the mammary gland in dairy cows.
BIOTECHNOLOGY
P. zopfii and P. moriformis have been proposed, together with Chlorella protothecoides, as organisms for production of ascorbic acid (Running et al., 2002). In a laboratory scale experiment (1‐14 L fermentors) P. zopfii at 35 °C at pH 7 produced 37.8 mg L‐1 of ascorbic acid, mainly intracellular, with a biomass preodcution of 27 g L‐1, while at pH 4/5 it produced 73 mg L‐1 of extracellular ascorbic acid and 55 g L‐1 of biomass in 29 h; similarly P. moriformis produced in 30 h at pH 4 162 mg L‐1 of extrcellular ascorbic acid and 42 g L‐1 of biomass (Huss et al., 1995; Running, 1999).
P. zopfii is able to utilise crude oil hydrocarbons (aromatics, cyclic and branched alkanes) as well as pure n‐alkanes (Ueno et al., 2002, 2008).
Solazyme Inc. (San Francisco, USA) has recently patented several processes to obtain oil from microrganisms including Prototheca, that contains lipids with a higher degree of saturation compared to other algae and has the advantage of lacking pigments, also through genetic engineering to increase production (Dillon et al., 2010; Franklin et al., 2010, 2011).
References
Arnold P., Ahearn D.G. (1972) The systematics of the genus Prototheca with a description of a new species P. filamenta. Mycologia 64: 265‐275. [BIOLOGY section]
Dillon H.F., Elefant D., Day A.G., Franklin S., Wittenberg J. (2010) Fractionation of oil‐bearing microbial biomass. WO2010/138620. Franklin S., Somanchi A., Espina K., Rudenko G., Chua P. (2011) Renewable chemical production from novel fatty acids feedstocks.
US Patent No. 7,883,882 B2. Franklin S., Somanchi A., Espina K., Rudenko G., Chua P. (2010) Manufactoring of tailored oils in recombinant heterotrophic
microorganisms. WO2010/063031 A2. Huss J.R., Running J.A., Skatrud T.J. (1995) L‐ascorbic acid production in microorganisms. WO95/21933. Running J.A. (1999) Process for the production of ascorbic acid with Prototheca. US Patent No. 5,900,370. Running J.A., Severson D.K., Schneider K.J. (2002) Extracellular production of L‐ascorbic acid by Chlorella protothecoides, Prototheca
species, and mutants of P. moriformis during aerobic culturing at low pH. Journal of Industrial Microbiology and Biotechnology 29: 93‐98.
Sudman M.S., Kaplan W. (1973) Identification of the Prototheca Species by immunofluorescence. Applied and Environmental Microbiology 25: 981‐990. [BIOLOGY section]
Ueno R., Urano N., Wada S., Kimura S. (2002) Optimization of heterotrophic culture conditions for n‐alkane utilization and phylogenetic position based on the 18S rDNA sequence of a thermotolerant Prototheca zopfii strain. Journal of Bioscience and Bioengineering 94: 160‐165.
Ueno R., Wada S., Urano N. (2008) Repeated batch cultivation of the hydrocarbon –degrading, micro‐algal strain Prototheca zopfii RND16 immobilised in polyurethane foam. Canadian Journal of Microbiology 54: 66‐70.
von Bergen M., Eidner A., Schmidt F., Murugaiyan J., Wirth H., Binder H., Maier T., Roesler U. (2009) Identification of harmless and pathogenic algae of the genus Prototheca by MALDI‐MS. Proteomics Clinical Applications 3: 774‐784. [BIOLOGY section]
Wolff G., Plante I., Lang B. F., Kück U., Burger G. (1994) Complete sequenceof the mitochondrial DNA of the chlorophyte alga Prototheca wickerhamii: Gene content and genome organization. Journal of Molecular Biology 237: 75‐86. [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 113 of 258
8.2 Rhodophyta
8.2.1 Porphyridium cruentum
Figure 32 ‐ Light microscopic view showing Porphyridium cruentum strain CCALA 415.
Picture from www.butbn.cas.cz/ccala/col_images/415.jpg
SYMBOLS: D
TAXONOMY
Phylum Rhodophyta Class Porphyridiophyceae Order Porphyridiales Family Porphyridiaceae Genus Porphyridium Species Porphyridium cruentum
Related species
There are 7 species (and infraspecific) names in the database at present, of which 3 have been flagged as currently accepted taxonomically.
P. aerugineum, P. cruentum, P. griseum, P. marinum, P. purpureum, P. sordidum, P. violaceum.
BIOLOGY
Porphyridium is composed of spherical to ovoid unicells with a stellate chloroplast and prominent central pyrenoid. The cell diameter is 5‐10 μm in the exponential phase, 7‐16 μm in the stationary phase. Cells are solitary, but often grouped into irregular colonies with an ill‐defined mucilaginous matrix. Species are distinguished by chloroplast color. The chloroplasts of freshwater species contain single thylakoids with phycobilisomes (granules consisting of the accessory pigments) on both sides. The phycobilisomes of blue‐colored species, such as Porphyridium aerugineum, tend to be hemidiscoidal in shape and predominated by the blue pigment phycocyanin . In contrast, the phycobilisomes of the red‐colored Porphyridium purpureum

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 114 of 258
are larger and hemispherical and composed mostly of the red pigment phycoerythrin. Reproduction occurs by cell division. Porphyridium forms gelatinous coatings on various surfaces; it is widespread in freshwaters, brackish environments. Species of Porphyridium can form gelatinous crusts on moist soils and decaying wood . In these habitats, these species are reasonably desiccation resistant and shade tolerant.
BIOTECHNOLOGY
The marine microalga Porphyridium cruentum is of increasing interest as source of valuable compounds like phycoerythrin, sulfated exopolysaccharides, superoxide‐dismutase, and polyunsaturated fatty acids with applications in the food, pharmaceutical and cosmetic industries (Fábregas et al., 1998; Dillon et al., 2007). Phycoerythrin is used as fluorescent dye in immunoassays (Bermejo Román et al., 2002), sulfated polysaccharides are known inhibit viruses (Huheihel et al., 2002) and show hypocholesterolemic activity in rats (Dvir et al., 2000, 2009). P. cruentum is also considered to be one of the most important sources of the polyunsaturated fatty acids eicosapentaenoic acid (20:5 ω 3, EPA) and arachidonic acid (20:4ω6, ARA) (Cohen and Cohen 1991; Guil‐Guerrero et al., 2001).
In a 220‐L ailift tubular photobioreactor P. cruentum reached average biomass productivities of 1.76 g L‐1 day‐1 with 2% biomass dry weight of ARA , 1.7% palmitic acid and 1.1% EPA (Rebelloso Fuentes et al., 1999), similar to those obtained by Camacho Rubio et al. (1999).
Porphyridium have been tested in screening for algal strains suitable for biodiesel production or considered as a potential interesting alga based on its biochemical composition (Griffiths and Harrison, 2009; Rodolfi et al., 2009), however up to now no in depth studies on this topic have been carried out.
References
Bermejo Román R., Alvárez‐Pez J.M., Acién Fernández F.G., Molina Grima E. (2002) Recovery of pure B‐phycoerythrin from the microalga Porphyridium cruentum. Journal of Biotechnology 93: 73–85.
Camacho Rubio F., Acién Fernández F.G., Sánchez Pérez J.A., García Camacho F., Molina Grima E. (1999) Prediction of dissolved oxygen and carbon dioxide concentration profiles in tubular photobioreactors for microalgal culture. Biotechnology and Bioengineering 62: 71‐86.
Cohen Z., Cohen S. (1991) Preparation of eicosapentaenoic acid (EPA) concentrate from Porphyridium cruentum. Journal of the American Oil Chemists' Society 68: 16‐19.
Cohen Z., Vonshak A., Richmond A. (1988) Effect of environmental conditions on fatty acid composition of the red alga Porphyridium cruentum: correlation to growth rate. Journal of Phycology 24: 328‐332. . [BIOLOGY section]
Dillon H.F., Somanchi A., Rao K., Jones. P.J.H. (2007) Nutraceutical compositions form microalgae and related methods of production and administration. WO2007/136428 A2.
Dvir I., Stark A.H., Chayoth R, Madar Z., Malis Arad S. (2009) Hypocholesterolemic effects of nutraceuticals produced from the red microalga Porphyridium sp in rats. Nutrients 1: 156‐167.
Dvir I., Chayoth R., Sod‐Moriah U., Shany S., Nyska A., Stark A.H., Madar Z., Malis Arad S. (2000) Soluble polysaccharide and biomass of red microalga Porphyridium sp. alter intestinal morphology and reduce serum cholesterol in rats. British Journal of Nutrition 84: 469‐476.
Fabregas J., Garcia D., Morales E., Domínguez A., Otero A. (1998) Renewal rate of semicontinuous cultures of the microalga Porphyridium cruen turn modifies phycoerythrin, exopolysaccharide and fatty acid productivity. Journal of Fermentation and Bioengineering 86: 477‐481.
Griffiths M.J., Harrison S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Journal of Applied Phycology 21: 493‐507.
Guil‐Guerrero J.L., Belarbi E.H., Rebolloso‐Fuentes M.M. (2001) Eicosapentaenoic and arachidonic acids purification from the red microalga Porphyridium cruentum. Bioseparation 9: 299–306.
Huheihel M., Ishanu V., Tal J., Malis Arad S. (2002) Activity of Porphyridium sp. polysaccharide against herpes simplex viruses in vitro and in vivo. Journal of Biochemical and Biophysical Methods 50: 189‐200.
Rebolloso Fuentes M.M., García Sánchez J.L., Fernández Sevilla J.M., Acién Fernández F.G., Sánchez Pérez J.A., Molina Grima E. (1999) Outdoor continuous culture of Porphyridium cruentum in a tubular photobioreactor: quantitative analysis of the daily cyclic variation of culture parameters. Journal of Biotechnology 70: 271‐288.
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Sheath R.G. (2003) Red algae. In: Wehr J.D., Sheath R.G. (eds.) Freshwater Algae of North America: Ecology and Classification. Elsevier Science, pp. 197‐224. [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 115 of 258
8.3 Bacillariophyta
8.3.1 Benthic diatoms (Amphora; Amphiprora; Cylindrotheca; Navicula; Nitzschia)
TAXONOMY
Amphora sp.
Figure 33 ‐ Amphora
coffeaeformis.
All rights reserved Seambiotic © 008
Figure 34 ‐ Light microscopic picture of Amphora sp.
Copyright 1995‐2010 Protist Information Server
Figure 35 ‐ Light microscopic picture of Amphora coffeaeformis.
Copyright © 2010 Monash University; arts.monash.edu.au
SYMBOLS: D, PIV
Phylum Bacillariophyta Class Bacillariophyceae Order Thalassiophysales Family Thalassiophysales Genus Amphora Species
Related species
Amphora is a very large and heterogeneous genus. There are 1032 species names in the algae database at present, of which 170 have been flagged as currently accepted taxonomically.
A. coffeaeformis, A. coffeaeformis punctata, A. coffeaeformis linea, A. coffeaeformis tenuis, A. coffeaeformis taylori, A. delicatissima, A. delicatissima capitata.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 116 of 258
Amphiprora hyalina
Figure 36 ‐ Light microscopic picture of Amphiprora sp.
Picture from www.plingfactory.de/
Figure 37‐ Light microscopic picture of Amphiprora sp.
Picture from www.mikroskopie‐ph.de/
SYMBOLS: D, PIV
Phylum Bacillariophyta Class Bacillariophyceae Order Naviculales Family Amphipleuraceae Genus Amphiprora Species Amphiprora hyalina
Related species
There are 230 species names in the algae database at present, of which 29 have been flagged as currently accepted taxonomically. Synonym: Amphiprora paludosa var. hyalina (Eulenstein ex Van Heurck) Cleve 1894.
Cylindrotheca sp.
Figure 38 ‐ Light microscopic picture of Cylindrotheca
sp.
Picture courtesy of PAE (UGent)
Figure 39 ‐ Scanning electron microscopic picture of Cylindrotheca sp.
Picture from ocean.inha.ac.kr/l3.htm
SYMBOLS: D, PIV

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 117 of 258
Phylum Bacillariophyta Class Bacillariophyceae Order Bacillariales Family Bacillariaceae Genus Cylindrotheca Species
Related species
There are 6 species names in the algae database at present, of which 2 have been flagged as currently accepted taxonomically.
Potentially important for biofuel: • Cylindrotheca fusiformis Reimann & Lewin 1964 • Cylindrotheca closterium (Brébisson ex Kützing) Grunow in van Heurck 1882
Navicula sp.
Figure 40 ‐ Light microscopic picture of Navicula
gregaria
Picture courtesy of PAE (UGgent)
Figure 41 ‐ Light microscopic picture of Navicula lanceolata
Picture courtesy of Petr Znachor
SYMBOLS: D, PIV
Phylum Bacillariophyta Class Bacillariophyceae Order Naviculales Family Naviculaceae Genus Navicula Species
Related species
Navicula is a very large and heterogeneous genus. There are 6714 species names in the algae database at present, of which 890 have been flagged as currently accepted taxonomically.
Potentially important for biofuel
Navicula acceptata, Navicula saprophila, Navicula pelliculosa.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 118 of 258
Nitzschia dissipata
Figure 42 ‐ Light microscopic picture of Nitzschia dissipata
Picture from http://craticula.ncl.ac.uk/
Figure 43 ‐ Raster electron microscopic picture of Nitzschia sp.
Picture from http://ina.beyer‐privat.net/
SYMBOLS: D, PIV
Phylum Bacillariophyta Class Bacillariophyceae Order Bacillariales Family Bacillariaceae Genus Nitzschia Species Nitzschia dissipata
Related species
There are 1024 species names in the algae database at present, of which 321 have been flagged as currently accepted taxonomically.
Potentially important for biofuel:
N. communis, N. frustulum, N. palea.
BIOLOGY Structural and morphological features
Benthic diatoms are all raphid, pennate diatoms. Amphora sp. The benthic Amphora species appear epiphytic, epilithic or epipelic (in or attached to sediments). The fouling A. coffeaeformis is a common marine species.
Cells of Amphora are solitary, sometimes sessile but usually motile, almost always lying in girdle view and then appearing elliptical or lanceolate, with truncate ends (Round et al., 1990). Amphora species have typical asymmetrical valve morphology. Its dorsiventral frustule resembles ‘‘a third of an orange’’ (Hendey, 1964) with both raphe systems on the same (ventral) side of the cell. Cells usually having 1 or 2, sometimes many, plastids which are extremely diverse in position, shape and structure (Round et al., 1990). Cell length and width varies with species, roughly ranging from 14 – 55 µm and 2.5 – 9 µm respectively (Sala et al., 1998).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 119 of 258
Amphiprora hyalina Amphiprora (currently placed in the genus Entomoneis) is a benthic genus, epipelic of brackish marine sediments; occasionally in freshwater. A. hyalina is a marine species.
Amphiprora/Entomoneis cells are solitary and twisted about the apical axis (twisted frustules) usually lying in girdle view and then appearing bilobate. This torsion of the cell means that valves or whole frustules can present a great variety of aspects depending on exactly how they lie relative to the observer. Cells contain one plate‐like plastid or two plastids, one on each side of the median transapical plane. A variety of pore and raphe structures is found in Entomoneis. The raphe is median, sigmoid, on a raised keel forming a wing (Round et al., 1990).
Cylindrotheca sp. Cyclindrotheca is benthic and widely distributed in the epipelon (living on (or in) fine sediments) of marine habitats; rarely occurring in freshwater. The genus is very abundant in coastal waters worldwide (Round et al., 1990).
Cylindrotheca cells are solitary, long and narrow, needle‐like and only weakly silicified. The frustules are twisted about the apical axis, consequently the cells rotate as they move through the sediment. Cells contain two or many chloroplasts that are plate‐like or discoid (Round et al., 1990).
Navicula sp. All Navicula are benthic diatoms. Cells of Navicula sensu lato diatoms have naviculoid (boat‐shaped) cells that may exist singly or in ribbons. Navicula is Latin for "small ship". The valves are symmetrical both apically and transapically, and may have rounded, acute, or capitate ends. The central area is often distinctly expanded. They contain two girdle‐appressed plastids, one on either side of the apical plane. The raphid system is well developed with a raphe on each valve which makes cells highly motile.
Nitzschia dissipata Nitzschia comprises both planktonic and benthic species. N. dissipata is a benthic species and was recorded in freshwater (e.g. Aboal et al., 2003; Day et al., 1995; Krammer and Lange‐Bertalot, 1988; Roberts et al.; 2004) as well as in coastal waters (Tadros and Johansen, 1988). Nitzschia is one of the most difficult genera for species identification and many features are only seen by electron microscopy (e.g. Mann, 1986; Trobajo et al., 2004, 2006). Nitzschia cells are usually linear to lanceolate and may be solitary or colony forming. Most species have two chloroplasts, one in each end of the cell. Each valve possesses a raphe that is more or less eccentric and supported by fibulae (=bridges of silica between portions of the valve on either side of the raphe, giving a ladder‐like appearance). The two raphes of a frustule are positioned diagonally opposite (nitzschioid). Valve striae (= lines with small holes) are usually uniseriate.
Biofilm formation
Benthic diatoms are the most common benthic microalgae, which are abundant in many soft‐sediment aquatic habitats (estuaries, shallow subtidal seas, coral reef flats, lakes, and rivers) and can contribute up to 50% of the total autotrophic production in some ecosystems. They form biofilms, a matrix of cells, sediments and extracellular polymeric substances (EPS) (Underwood and Paterson, 2000). It is known that diatom mucilages are rich in polysaccharides and proteoglycans and are secreted through the channels and pores in the diatom frustule. In some benthic species, these compounds produce structures (tubes, pads and stalks) that are used for attachment to surfaces and often contribute to biofouling problems. Epipelic diatoms do not produce permanent structures but secrete large quantities of extracellular mucilages that are involved in motility (Underwood and Paterson, 2000). Motility is an essential adaptation for photosynthetic organisms in these environments, allowing cells to migrate into the illuminated (photic) zone of sediment near the surface after periods of sediment mixing or deposition. In diatoms, this motility

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 120 of 258
is generated by the production of extracellular polymeric substances (EPS), primarily polysaccharides (Underwood et al., 2004). The mechanism of movement in diatoms is unique for microbial cells and relies on the extrusion of mucilage through a slit in the surface of the silica frustule (cell wall). This slit, known as the raphe, may be present on only a single valve of the frustule (monoraphid) or on both valves (biraphid) (Underwoos and Paterson, 2000).The production of EPS in the aquatic environment is ecologically significant because EPS and other carbohydrate‐rich exudates can be used by bacteria, meiofauna, and macrofauna as a carbon source and EPS can increase the stability of sediments (Underwood et al., 2004). The production of extracellular carbohydrates shows some dependency on environmental conditions, for example, irradiance and nutrient conditions. The production of some extracellular carbohydrates ceases in darkness, but studies on axenic cultures of benthic diatoms and natural sediment assemblages have shown production of EPS in dark as well as under illuminated conditions. Continued EPS production in darkness indicates that EPS production is not directly coupled to the photosynthetic production of carbohydrates (Underwood et al., 2004). Nutrient limitation can also increase extracellular carbohydrate production in benthic diatoms: this production of extracellular carbohydrates is assumed to be due to unbalanced metabolism, with cells releasing fixed carbon in excess of their energetic requirements, because of growth being prevented by nutrient (N, P) limitation (Underwood et al., 2004). Diatom EPS consists of a relatively undefined complex mixture of proteins, proteoglycans and carbohydrates (Underwood and Paterson, 2000).There is evidence that benthic diatoms produce a number of different types of EPS, which vary in structure and sugar composition, and that the production of these EPS depends on environmental conditions and the nutrient status of the cells (Underwood et al., 2004). The biosynthetic pathway for these carbohydrates and the mechanisms causing changes in EPS composition are yet to be elucidated. Glucans either may be the precursor of EPS or may act as the photoassimilate carbon store, providing energy for EPS synthesis during periods of darkness.This latter hypothesis is supported by the significant correlation found between glucan catabolism and EPS production in the dark in a number of benthic species. This provides indirect evidence that glucans are involved in the production of EPS (Underwood et al., 2004). Microphytobenthic biofilms can have high rates of photosynthesis and a significant proportion of their photo‐assimilated carbon is released into the environment as extracellular carbohydrates (Underwood and Paterson, 2000).
Biochemical composition under optimal and stressed conditions
Amphora sp.: The biochemical composition vary among Amphora species as indicated by the diverse values reported by different studies (de la Peña, 2007; Gordon et al., 2006; Khatoon et al., 2009; Sheehan et al., 1998). In addition, different culture conditions result in significant variations in growth and the biochemical composition of the cells of the same strain as shown by de la Peña (2007). He showed that the proximate chemical composition (protein, carbohydrates, fatty acid content, chlorophyll a) of Amphora sp. is highly dependent on light intensity, the culture location (indoor‐outdoor) and the type of enrichment used (de la Peña, 2007). A higher protein and carbohydrate content of Amphora sp. was noted in cultures located inside the laboratory compared to cultures grown outside (probably due to the more regulated cultural conditions inside like constant irradiance and temperature). Lipid content ranged from 26 – 81% depending on culture site and nutrients used (de la Peña, 2007). Amphora is rich in total lipids and fatty acids with a high amount of polyunsaturated fatty acids (PUFAs) especially EPA and a high amount of essential amino acids (Gordon et al., 2006; Khatoon et al., 2009). Griffiths and Harrison (2009) calculated the average total lipid content for Amphora sp. from available literature data.
• 51% cdw (cell dry weight) under nutrient replete laboratory conditions • 40% cdw in outdoor ponds

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 121 of 258
Amphiprora hyalina: A. paludosa var. hyalina contains a high amount of EPA (28%) (Correa‐Reyes et al., 2009). Proximate composition (dry weight percentage) in A. paludosa var. hyalina (Correa‐Reyes et al., 2009):
• 11.33% ± 0.36 protein • 20.96% ± 0.94 nitrogen free extracts • 8.10% ± 0.49 lipids • 59.61% ± 1.01 ash
The average total lipid content for Amphiprora hyalina calculated from available literature data by Griffiths and Harrison (2009) is:
• 22% cdw under nutrient replete laboratory conditions • 28% cdw under N deficient laboratory conditions • 37% cdw under Si deficient conditions
Cylindrotheca sp.: The average total lipid content for Cylindrotheca sp. calculated from literature by Griffiths and Harrison (2009) is:
• 27% cdw under nutrient replete conditions • 27% cdw under N deficient conditions
Navicula sp.: The average total lipid content for N. acceptata calculated from literature by Griffiths and Harrison (2009) is:
• 33% cdw under nutrient replete conditions • 35% cdw under N deficient conditions • 46% cdw under Si deficient conditions
N. saprophila contains large quantities of EPA and is considered a potential source of this important fatty acid (Kitano et al., 1997). It was shown that EPA production was enhanced under mixotrophic conditions in CO2 enriched (about 2%) atmosphere in the presence of acetate as compared with photoautotrophic conditions (Kitano et al., 1997; 1998). The biomass of a freshwater N. saprophila strain has the following composition (Pilny, 2009):
grown in WC medium medium with half strength phosphorus and half strength nitrogen (urea as a source)
• Protein 46.03 % 38.82 % • Lipid 28.57 % 18.24 % • Crude fiber < 15.4 % < 11.86 % • Ash 11 % 11.86 %
Saline strains of N. saprophila have been found to produce significant amounts of carbohydrate in some media (Barclay et al., 1986). The average total lipid content for N. saprophila calculated from literature by Griffiths and Harrison (2009):
• 24% cdw under nutrient replete conditions • 51% cdw under N deficient conditions • 49% cdw under Si deficient conditions
Chelf (1990) showed that nitrogen concentration was the variable with the greatest effect on neutral lipid accumulation in N. saprophila. Nitrogen deficiency led to a higher synthesis of lipids.
Nitzschia dissipata: Various Nitzschia species have high oil content (Chisti, 2007; Griffiths and Harrison, 2009; Mata et al., 2010) and have been suggested for production of EPA (Spolaore et al., 2006). EPA production in Nitzschia sp. was enhanced under mixotrophic conditions in the presence of acetate as compared with photoautotrophic conditions (Kitano et al., 1997). Some Nitzschia species are cultured

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 122 of 258
heterotrophically (e.g. Barclay et al., 1994; Wen and Chen, 2003). In the ASP program different Nitzschia species were isolated (a.o. N. dissipata, N. palea, N. communis, N. pusilla, etc.) (Sheehan et al., 1998). Preliminary screening experiments indicated that N. dissipata had the best growth rates and lipid accumulation potential (Sheehan et al., 1998). The average total lipid content for N. dissipata calculated from literature by Griffiths and Harrison (2009):
• 28% cdw under nutrient replete conditions • 46% cdw under N deficient conditions • 47% cdw under Si deficient conditions
BIOTECHNOLOGY
Benthic diatoms (Amphora, Amphiprora, Cylindrotheca, Navicula, Nitzschia) are commonly used in abalone culture, where they act as inductors for larval settlement and as food for the early juvenile stages. They excrete extracellular polymeric substances that play an important role in abalone larval settlement (Brown et al., 1997; Pulz and Gross, 2004; Carbajal‐Miranda et al., 2005; see refs in de la Peña, 2007; Correa‐Reyes et al., 2009).
Species of the genus Nitzschia occur in nearly every diatom assemblage in fresh, brackish, and marine habitats. Many of them are considered to be important indicators of organic pollution and high nutrient loads, making them important for water quality studies and biomonitoring (Lange‐Bertalot, 1979; Van Dam et al., 1994).
For their high lipid content benthic diatoms have been considered as sources for biodiesel, though experiments were carried only at laboratory scale (mainly in flasks), obtaining quite low growth and productivity (Griffith and Harrison, 2009; Rodolfi et al., 2009). Navicula saprophila was genetically transformed to optimize lipid production (Dunahay et al., 1995).
Culture Media
For marine and brackish strains the most common medium used is medium F or f/2 added with Silicon (Guillard and Ryther, 1962) and adjusted in salinity according to necessity. For freshwater strains media like Chu (Gerlogg et al., 1950) or BG11 (Rippka et al., 1979) added with Silicon can be used.
Cultivation methods
The benthic diatoms, in general, have to be grown in photobioreactors purposely designed, because of their tendency to form biofilms. The simplest solution is represented by the benthic algae growth chamber (BAGC), which was used to grow a mixed benthic algae community on dairy manure by Wilkie and Mulbry (2002). The culture medium (220 L) was recirculated from a tank to the chamber by a submersible recirculation pump while algal growth was supported by screens of 96:5 x 96:5 cm with 3 x 4 mm mesh, for a total growing area of 0.93 m2.
Another possible solution was that by Lebeau et al. (2000), who proposed a photobioreactor design for immobilised microalgal cells. The aim of this photobioreactor was to cultivate microalgae, especially benthic diatoms that suffer from bioturbation. In these photobioreactors, the surface/volume ratio of the matrix (agar or alginate) was maximised to offer the maximal contact between microalgae and nutrient/light. The immobilized cell photbioreactor consisted of 10‐L glass and stainless steel cylinders. The inoculum was entrapped in a tubular agar gel layer, then iserted into the cylinder containing the liquid culture medium. The reactor was tested with Haslea ostrearia for the production of the pigment marennine (Rossignol et al., 2000).
Another solution is to multiply the adhesion surface inside the reactor. This has been achieved in different ways. The simplest is that proposed by de la Peña (2007) that cultivated Amphora in a 8 L

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 123 of 258
rectangular acrylic glass aquaria containing 5 L of culture medium. Ten pieces of 12× 12×12 cm2 area acrylic glass plates were installed in each aquarium to serve as diatom settlement plates. The plates were provided with orange polyvinyl stand pipes to keep them in an upright and slanting position. A two‐point mild aeration was provided by glass‐wool filtered air to provide an effective gas exchange and to allow the alga to settle onto the acrylic glass plates. A more complex device was proposed by Avendaño‐Herrera and Riquelme (2007), that used a new system called the Tanaka photobioreactor, that consists of a borosilicate tube measuring 40 x 4 cm, with a 4.5 x 1 cm side‐arm effluent tube at the bottom and a PVC stopper at the top which has an opening to allow the entry of a 5 mm diameter glass rod. The interior of the tube contains an array of about 350 sterilized polyethylene bristles in a ‘‘bottle brush’’ arrangement measuring 30x 4 cm, providing an extensive surface area for adhesion. They used the reactor to cultivate Navicula veneta with associated bacteria.This configuration was slightly modified by Silva‐Aciares and Riquelme (2008) with a reactor called ‘‘bristles photobioreactor’’ (PBB). PBB is constructed of transparent acrylic plastic 1 m in length with an inside diameter of 18 cm and outside diameter of 19 cm. The top of the tube is closed by a PVC cap whith a connector for the introduction of the air line. The base of the tube is closed. Within the body of the PBB, throughout the entire length of the tube, there is a structure resembling a large ‘‘bottle brush’’ bearing PVC bristles measuring 17.5 cm. The bristles and the structure were designed as supports for the growth of the (adherent) benthic diatoms. PVC bristles were the best material for this purpose in comparison with a number of other types of plastic materials tested. The circulation of the cell suspension throughout the photobioreactor is obtained using an airlift system. They cultivated Nitzschia, Amphora, Navicula and Cylindrotheca closterium obtaining volumetric productivities in the range 0.12‐0.28 g L‐1 d‐1, with Nitzschia being the best and one Navicula the worse performer.
On an industrial scale, SBAE Industries nv (Belgium) has developed and patented an outdoor technology called DiaForce for production of benthic diatoms. DiaForce pumps water through channels filled with artificial carriers on which the diatoms attach. DiaForce technology is used for the production of diatom polycultures.
Some species, like some Cylindrotheca, can be grown also in bubbled photobioreactors.
Harvesting methods
Benthic (attached) freshwater algae may have an advantage over planktonic (suspended) algae in the ease of separation and recovery of algal biomass from an aqueous stream. Avendaño‐Herrera and Riquelme (2007) report that harvesting of the N. veneta cultures was carried out at the end of each culture cycle (each 7 days). As much as possible of the diatom (>85%) was dislodged from the bristles within the photobioreactor using a 1‐min agitation of the glass rod, draining the reactor through the sidearm. To avoid handling of the photobioreactor, the remaining diatom‐bacterial biomass (approximately 15% of the total) was used to start a new culture cycle. Microalgae were harvested manually from the PBB, dislodging the diatoms by vigorous agitation of all the PVC bristles in the photobioreactor column for a period of 2 min (Silva‐Aciares and Riquelme, 2008). SBAE’s triangle carriers facilitate the harvesting process as algal biomass is directly collected, by lifting them from the water. This concentrates the biomass x 100 and reduces the cost compared to centrifugation of the algae out of the water.
Upscaling limitations
All the systems proposed, except SBAE’s one, have been developed in relation to aquaculture necessities, so they are of rather difficult scaling‐up. To exploit benthic diatoms as source of biodiesel, it will be necessary to design novel photobioreactors, specific for their cultivation, that can be scaled up.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 124 of 258
References
Aboal M., Alvarez Cobelas M., Cambra J., Ector L (2003) Floristic list of non‐marine diatoms (Bacillariophyceae) of Iberian Peninsula, Balearic Islands and Canary Islands. Updated taxonomy and bibliography. Diatom Monographs 4: 1‐639.
Avendaño‐Herrera R.E., Riquelme C.E. (2007) Production of a diatom‐bacteria biofilm in a photobioreactor for aquaculture applications. Aquaculture Engineering 36: 97‐104.
Barclay W., Johansen J.R., Chelf P., Nagle N., Roessler P., Lemke P. (1986) Microalgae culture collection 1986‐1987. Solar Research Institute. Golden, Colorado.
Barclay W.R., Maeger K.M., Abril J.R. (1994) Heterotrophic production of long chain omega‐3 fatty acids utilizing algae and algae‐like microorganisms. Journal of Applied Phycology 6: 123‐129.
Brown M.R., Jeffrey S.W., Volkman J.K., Dunstan G.A. (1997) Nutritional properties of microalgae for mariculture. Aquaculture 151: 315‐331.
Carbajal‐Miranda M.J., Sanchez‐Saavedra M.D., Simental J.A. (2005) Effect of monospecific and mixed benthic diatom cultures on the growth of red abalone postlarvae Haliotis rufescens (Swainson 1822). Journal of Shellfish Research 24: 401‐405.
Chelf P. (1990) Environmental control of lipid and biomass production in two diatom species‐Journal of Applied Phycology 2: 121‐129.
Chisti Y. (2007) Biodiesel from microalgae. Biotechnology Advances 25:, 294‐306. Correa‐Reyes J.G., Sanchez‐Saavedra M..D, Viana M.T., Flores‐Acevedo N., Vasquez‐Pelaez C. (2009) Effect of eight benthic diatoms
as feed on the growth of red abalone (Haliotis rufescens) postlarvae. Journal of Applied Phycology 21: 387‐393. Day S.A., Wickham R.P., Entwisle T.J., Tyler P.A. (1995) Bibliographic check‐list of non‐marine algae in Australia. Flora of Australia
Supplementary Series 4: VII + 276. de la Peña M.R. (2007) Cell growth and nutritive value of the tropical benthic diatom, Amphora sp., at varying levels of nutrients
and light intensity, and different culture locations. Journal of Applied Phycology 19: 647‐655. Dunahay T.G., Jarvis E.E., Roessler P.G. (1995) Genetic transformation of the diatoms Cyclotella cryptica and Navicula saprophila.
Journal of Phycology 31: 1004‐1012. Ezzat A.I. (1983) Effects of some common pesticides on growth and metabolism of the unicellular algae Skeletonema costatum,
Amphiprora paludosa, and Phaeodactylum tricornutum. Aquatic Toxicology 3: 1‐14. French M.S. (1988) The effects of copper and zinc on growth of the fouling diatoms Amphora and Amphiprora. Biofouling 1: 3‐18. Gerlogg G.C., Fitzgerald G.P., Skoog F. (1950) The isolation, purification, and culture of blue‐green algae. American Journal of
Botany 37: 216‐218. Gordon N., Neori A., Shpigel M., Lee J., Harpaz S. (2006) Effect of diatom diets on growth and survival of the abalone Haliotis discus
hannai postlarvae. Aquaculture 252: 225‐233. Griffiths M.J., Harrison S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production.
Journal of Applied Phycology 21: 493‐507. Guillard R.R.L.., Ryther J.H. (1962) Studies of marine planktonic diatoms. I. Cyclotella nana (Hustedt) and Detonula confervacea
(Cleve). Canadian Journal of Microbiology 8: 229‐239. Hendey N.I. (1964) An introductory account of the smaller algae of British coastal waters. Part V. Bacillariophyceae (Diatoms). Her
Majesty’s Stationery Office, London. Khatoon H., Banerjee S., Yusoff F., Shariff M. (2009) Evaluation of indigenous marine periphytic Amphora, Navicula and Cymbella
grown on substrate as feed supplement in Penaeus monodon postlarval hatchery system. Aquaculture Nutrition 15: 186‐193.
Kitano M., Matsukawa R., Karube I. (1997) Changes in eicosapentaenoic acid content of Navicula saprophila, Rhodomonas salina and Nitzschia sp. under mixotrophic conditions. Journal of Applied Phycology 9: 559‐563.
Kitano M., Matsukawa R., Karube .I (1998) Enhanced eicosapentaenoic acid production by Navicula saprophila. Journal of Applied Phycology 10: 101‐105.
Krammer K., Lange‐Bertalot H. (1988) Susswasserflora von Mitteleuropa. Bacillariophyceae. 2/2. VEB Gustav Fischer Verlag, Stuttgart, Germany..
Lange‐Bertalot H. (1979) Pollution tolerance of diatoms as a criterion for water quality estimation. Nova Hedwigia 64: 285‐304. Lebeau .T, Gaudin P., Mignot L., Junter G.A. (2000) Continuous marennine production by agar‐entrapped Haslea ostrearia using a
tubular photobioreactor with internal illumination. Applied Microbiology and Biotechnology 54: 634‐640. Mann D.G. (1986) Nitzschia subgenus Nitzschia. Notes for a monograph of the Bacillariaceae, 2nd ed. In Ricard M. (ed). Proceedings
of the 8th International Diatom Symposium. Koeltz Scientific Books, Koningstein, pp. 215‐226. Mata T.M., Martins A.A., Caetano N.S. (2010) Microalgae for biodiesel production and other applications: A review. Renewable and
Sustainable Energy Reviews 14: 217‐232. Pilny J. (2009) Biofuels from algae: Physiological characterization of candidate diatom species, University of South Bohemia, Czech
Republic. Pulz O., Gross W. (2004) Valuable products from biotechnology of microalgae. Applied Microbiology and Biotechnology 65: 635‐
648. Rippka R., Deruelles J., Waterbury J. B., Herdman M., Stanier R. Y. (1979) Generic assignments, strain histories and properties of
pure cultures of cyanobacteria. Journal of General Microbiology 111: 1‐61. Roberts S, Sabater S, Beardall J (2004) Benthic microalgal colonization in streams of differing riparian cover and light availability .
Journal of Phycology 40, 1004‐1012. Rodolfi L., Chini Zittelli G., Bassi N., Padovani N., Biondi N., Bonini G, Tredici MR. (2009) Microalgae for oil: strain selection,
induction of lipid synthesis and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 125 of 258
Round F.E., Crawford R.M., Mann D.G. (1990) The diatoms ‐ biology and morphology of the genera. Cambridge University Press, Cambridge, UK.
Sala S.E., Sar E.A., Ferrario M.E. (1998) Review of materials reported as containing Amphora coffeaeformis (Agardh) Kützing in Argentina. Diatom Research 13: 323‐336.
Sheehan J., Dunahay T., Benemann J., Roessler P. (1998) A look back at the U.S. Department of Energy's Aquatic Species Program—biodiesel from algae.
Silva‐Aciares F.R., Riquelme C.E. (2008) Comparisons of the growth of six diatom species between two configurations of photobioreactors. Aquaculture Engineering 38: 26‐35
Spolaore P., Joannis‐Cassan C., Duran E., Isambert A. (2006) Commercial applications of microalgae. Journal of Bioscience and Bioengineering 101: 87‐96.
Tadros M.G., Johansen J.R. (1988) Physiological characterization of 6 lipid‐producing diatoms from the southeastern United‐States. Journal of Phycology 24: 445‐452.
Trobajo R., Cox E.J., Quintana X.D. (2004) The effects of some environmental variables on the morphology of Nitzschia frustulum (Bacillariophyta), in relation its use as a bioindicator. Nova Hedwigia 79: 433‐445.
Trobajo R., Mann D.G., Chepurnov V.A., Clavero E., Cox E.J. (2006) Taxonomy, life cycle, and auxosporulation of Nitzschia fonticola (Bacillariophyta). Journal of Phycology 42: 1353‐1372.
Underwood G.J.C., Paterson D.M. (2000) The importance of extracellular carbohydrate production by marine epipelic diatoms. Advances in Botanical Research 40: 183‐240.
Underwood G.J.C., Boulcott M., Raines C.A., Waldron K. (2004) Environmental effects on exopolymer production by marine benthic diatoms: dynamics, changes in composition, and pathways of production. JournL of Phycology 40: 293‐304.
Van Dam H., Mertens A., Sinkeldam J. (1994) A coded checklist and ecological indicator values of freshwater diatoms from the Netherlands. Netherlands Journal of Aquatic Ecology 28: 117‐133.
Wen Z.Y., Jiang Y., Chen F. (2002) High cell density culture of the diatom Nitzschia laevis for eicosapentaenoic acid production: fed‐batch development. Process Biochemistry 37: 1447‐1453.
Wen Z.Y., Chen F. (2003) Heterotrophic production of eicosapentaenoic acid by microalgae. Biotechnology Advances 21: 273‐294. Wilkie A.C., Mulbry W.W. (2002) Recovery of dairy manure nutrients by benthic freshwater algae. Bioresource Technology 84: 81‐
91.
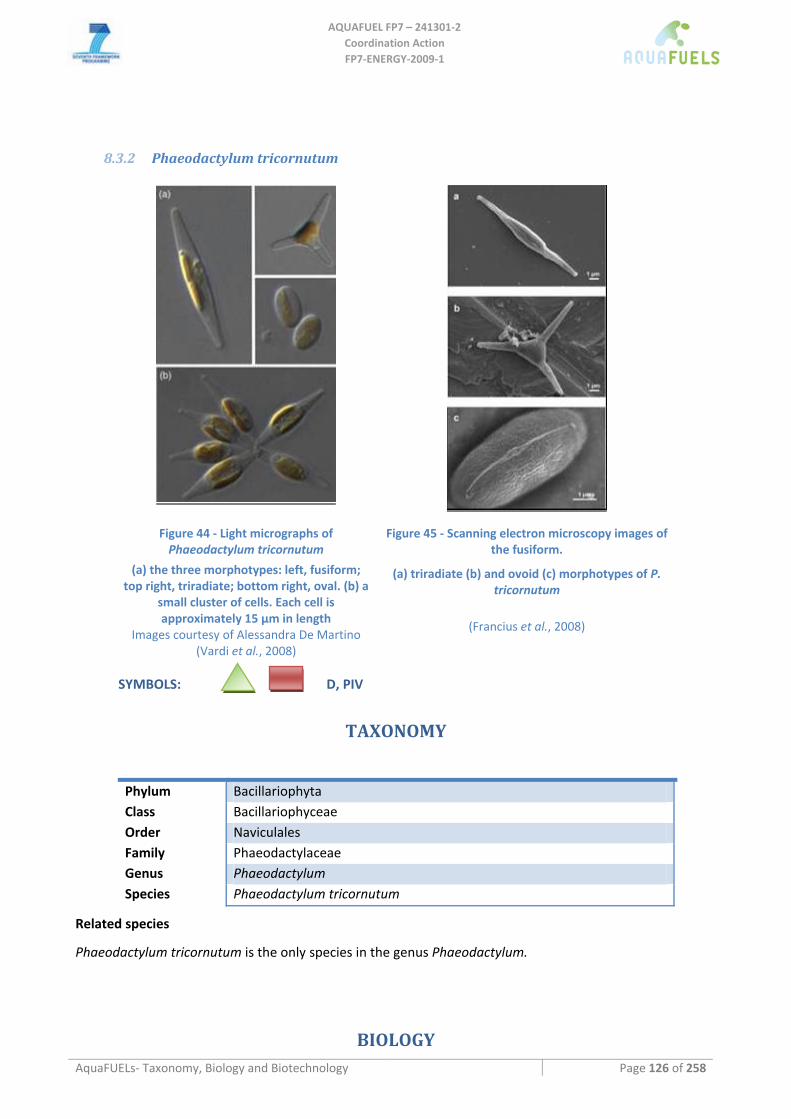
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 126 of 258
8.3.2 Phaeodactylum tricornutum
Figure 44 ‐ Light micrographs of Phaeodactylum tricornutum
(a) the three morphotypes: left, fusiform; top right, triradiate; bottom right, oval. (b) a
small cluster of cells. Each cell is approximately 15 μm in length
Images courtesy of Alessandra De Martino (Vardi et al., 2008)
Figure 45 ‐ Scanning electron microscopy images of the fusiform.
(a) triradiate (b) and ovoid (c) morphotypes of P. tricornutum
(Francius et al., 2008)
SYMBOLS: D, PIV
TAXONOMY
Phylum Bacillariophyta Class Bacillariophyceae Order Naviculales Family Phaeodactylaceae Genus Phaeodactylum Species Phaeodactylum tricornutum
Related species
Phaeodactylum tricornutum is the only species in the genus Phaeodactylum.
BIOLOGY

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 127 of 258
General description
Phaeodactylum belongs to the pennate diatoms (Bacillariophyceae). It is a member of the raphid pennate clade (Medlin and Kaczmarska, 2004). Examination of intraspecies genetic diversity based on rDNA internal transcribed spacer 2 (ITS2) sequences and amplified fragment length polymorphism (AFLP) analyses indicate four different genotypes within the species (De Martino et al., 2007).
Phaeodactylum tricornutum is fast growing and has emerged as a model system for physiological, biochemical, and molecular studies mainly because of its ease of culture and the ability to be routinely genetically transformed (Scala et al., 2002, Montsant et al., 2005).
It was chosen as the second diatom for whole genome sequencing after the centric diatom Thalassiosira pseudonana. The completed P. tricornutum genome, sequenced by the Joint Genome Institute (JGI), is approximately 27.4 megabases (Mb) in size and is predicted to contain fewer genes than T. pseudonana (Bowler et al., 2008). P. tricornutum shares 57% of its genes with T. pseudonana. A remarkably high number of P. tricornutum predicted genes appears to have been transferred between diatoms and bacteria and are likely to provide novel metabolic capacities (Bowler et al., 2008).
Structural and morphological features
Phaeodactylum tricornutum is unicellular or forms small cell clusters. Cells are circa 3 µm wide and 8 to 20 µm long (Lewin et al., 1958), and contain a single plastid (Round et al., 1990). Unlike other diatoms it can exist in different morphotypes (i.e. fusiform, triradiate or oval) (Borowitzka and Volcani, 1978). This plasticity is related to the atypical nature of the cell wall, which is only poorly silicified compared with other diatoms. In addition, Phaeodactylum does not exhibit the size reduction– restitution cycle that is so unique for the diatoms. The species appears to be unique in that it does not have an obligate requirement for silicic acid. The ovoid form is the only morphotype which is able to synthesize true single silica valves when grown in the presence of silicic acid. Siliceous frustules were never observed in fusiform or triradiate cells, or in oval cells grown in the absence of silicic acid (De Martino et al., 2007). Fusiform and triradiate morphotypes are characterized by cell walls possessing almost exclusively organic components (De Martino et al., 2007; Francius et al., 2008).
Physiological characteristics
Although not considered to be of great ecological significance, P. tricornutum has been found in several locations around the world, typically in coastal areas with wide fluctuations in salinity as well as in inland waters (Rushforth et al., 1988).
The ability of the species to adapt to changing environmental conditions could be related to the pleiomorphic character of the cells and the different morphotypes are thought to be adapted for survival in different habitats. It is shown that the three morphotypes are physiologically different. The oval morphotype appears to be induced as an acclimation response to suboptimal growth conditions (De Martino et al., 2007). The oval and triradiate morphotypes may represent distinct eco‐phenotypes each one specifically adapted to growth in particular conditions (De Martino et al., 2007). In contrast to oval cells, triradiate cells appear to be much more sensitive to stress and rapidly disappear (or convert to oval ⁄ fusiform cells) when growth conditions are suboptimal (De Martino et al., 2007).
Fusiform and triradiate forms are common when grown in liquid media. Fusiform cells tend to transform into the ovoid morphotype when they are transferred on a solid medium (agar) while the reverse transformation occurs upon transfer in liquid medium (Lewin et al., 1958). It has also been noted that fusiform and triradiate cells are more buoyant than oval cells (Lewin et al., 1958) and so would be expected to be better adapted to a planktonic lifestyle than oval cells. De Martino et al. (2007) suggested that P.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 128 of 258
tricornutum should predominantly occur in benthic communities because oval and round cells tend to aggregate and sink and can adhere strongly to surfaces. Very little is known on the nature and biological role of the triradiate–fusiform transition (Francius et al., 2008).
Biochemical composition
P. tricornutum is extensively used as a food source for the aquaculture industry because of its ease of cultivation and its rich oil content. However, different culture conditions result in significant variations in the biochemical composition of the cells and, therefore, in their nutritious value.
On average, the biomass of P. tricornutum contains (Rebolloso‐Fuentes et al., 2001): • 36,4% crude protein, • 26,1% available carbohydrates, • 18,0% lipids, • 15,9% ash.
The biomass produced at low external irradiance was richer in protein and eicosapentaenoic acid (Rebolloso‐Fuentes et al., 2001).
The total lipid content and the fatty acid and lipid class (polar membrane lipids vs neutral storage lipids) composition are dependent on different chemical (nutrient starvation, salinity, growth‐medium pH) and physical stimuli (temperature and light intensity). In addition growth phase and/or aging of the culture has an effect on oil content and composition (Hu et al., 2008). This hampers rigorous comparison across experiments under different conditions (Griffiths and Harrison, 2009).
Griffiths and Harrison (2009) calculated the average total lipid content for P. tricornutum from available literature data:
• 21% cdw (cell dry weight) under nutrient replete conditions, • 26% cdw under N deficient conditions.
Other literature mention values for total oil content of: • 31% dw (Sheehan et al., 1998), • 18.7 % biomass dw (Rodolfi et al., 2009), • 18 – 57% dw (Mata et al., 2010).
The fatty acid composition and the partitioning into storage lipids in P. tricornutum is intensely studied, e.g. (Alonso et al., 1998, 2000; Rebolloso‐Fuentes et al., 2001; Tonon et al., 2002; Meiser et al., 2004; Alonso et al., 1998; Liang et al., 2006; Meiser et al., 2004; Ohse et al., 2009; Rebolloso‐Fuentes et al., 2001; Tonon et al., 2002; Yu et al., 2009).
In Phaeodactylum, the very‐long‐chain fatty acids (vlc PUFA) arachidonic acid (AA) (C20:4ω6), eicosapentaenoic acid (EPA) (C20:5x3) and docosahexaenoic acid (DHA) (C22:6ω3) are the major fatty acid species accounting for approximately 30% of the total fatty acid content (Hu et al., 2008).
P. tricornutum is able to produce the nutritionally relevant EPA (3.9–5% CDW) in high proportion to the total fatty acid content and has important advantages as a potential commercial producer of EPA because it is fast‐growing with low amounts of other PUFAs, such as DHA and AA, which has important advantages in simplifying recovery. (Lebeau and Robert, 2003; Meiser et al., 2004). Unlike in most algal species/strains examined where TAGs are composed primarily of C14–C18 fatty acids (saturated or mono‐unsaturated) in P. tricornutum, partitioning of very‐long‐chain (>C20) into TAGs have been observed (Hu et al., 2008). The TAG yield from P. tricornutum is about 14% of total dry weight (Yu et al., 2009).
Phosphorus limitation result in increased lipid content, mainly TAG, in P. tricornutum (Reitan et al., 1994). Phosphorus deprivation was found to result in a higher relative content of 16:0 and 18:1 and a lower relative content of 18:4x3, 20:5x3 and 22:6x3 (Reitan et al., 1994).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 129 of 258
A decrease of the nitrogen concentration caused the galactolipid (GL) fraction to decrease from 21 to 12%. Conversely, both neutral lipids (NLs) and phospolipids (PLs) increased from about 73 to 79% and from 6 to 8%, respectively. TAG was the lipid class with the highest increase, from 69 to 75% (Alonso et al., 2000).
Unlike in some other diatoms, culture age had almost no influence on the total fatty acid content in P. tricornutum that remained around 11% of dry weight (Alonso et al., 2000). Conversely, culture age had a greater impact on lipid classes, producing changes in amounts of triacylglycerols (TAG) which ranged between 43% and 69%, and galactolipids (GLs) that oscillated between 20% and 40%. In general, the content of polar lipids of the biomass decreased with culture age (Alonso et al., 2000). .
Carotenoids in Phaeodactylum tricornutum are low, ranging from 0.115 to 0.45 g/100g dry biomass. In P. tricornutum, chlorophylls are the main pigments (1.17 – 2.87g) (Rebolloso‐Fuentes et al., 2001).
Growth kinetics and productivity
Growth rate and biomass productivity are influenced by environmental conditions, available resources and choice of culture system.
In general, P. tricornutum shows good growth at temperatures between 15 and 25 °C. For most isolates, growth ceases at temperatures above 30 °C. In relation to the appropriate conditions for the cultures, some researchers have found in this species a reduction in the photosynthesis activity of nearly 75% at pH values above 9.0 and under 5.5. On the other hand, it has also been informed that, for restrictive light conditions, the specific growth rate decreases at a pH above 9.0. The influence of illumination in microalgae growth rates is one of the most controversial factors. According to some investigators, P. tricornutum uses a photo adaptation strategy, developing an increase of the cellular content of chlorophyll under limiting light conditions. This effect causes that the photosynthetic efficiency, and consequently the specific growth rate, changes very little for a wide irradiance range (reviewed in Perez et al., 2008).
The influence of several culture conditions on the specific growth rate of P. tricornutum was studied in batch cultures by Perez et al. (2008). They calculated an optimum pH of 7,8 and a specific growth rate of 0.064 h−1 was achieved for certain nitrate conditions and illumination. However, for pH values between 6 and 9 growth was favorable and did not show great variation. The temperature optimum was achieved at 20.4 °C in aerated cultures and at 22.3 °C in non‐aerated cultures. Better adaptation to low temperatures than high ones has been obtained. The experiments carried out with different irradiances drive to a simple Monod’s equation for the irradiance influence on growth, with semi‐saturation irradiance of 10.2 and 6.8 µmol photons m‐2 s‐1 for aerated and non‐aerated cultures, respectively. Furthermore, aeration affects to the growth of P. tricornutum, with higher growth rates under aerated conditions (Perez et al., 2008).
Analysis of biomass productivity, lipid content and their combination to yield lipid productivity has been done across literature data by Griffiths and Harrison (2009). Lipid productivity is a critical variable for evaluating algal species for biodiesel production, because lipid content does not correlate directly with lipid productivity.
Average literature data for P. tricornutum grown in laboratory under nutrient replete conditions Laboratory averages under nutrient replete conditions from literature for P. tricornutum (Griffiths and Harrison, 2009):
• biomass growth (average doubling time): 25 h • biomass productivity on a volumetric basis: 0.34 g L‐1 day‐1 • biomass productivity on a surface area basis: 20 g m‐2 day‐1 • lipid productivity calculated from laboratory biomass productivity (in grams per litre per day) and
nutrient replete lipid content in nutrient replete conditions: 72 mg L‐1 day‐1 • lipid productivity reported directly in the literature: 45 mg L−1 day−1

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 130 of 258
Rodolfi et al. (2009) showed a biomass productivity of 0.24 g L−1 day−1 and a lipid productivity of 44.8 mg L−1 day−1 for P. tricornutum F&M‐M40 strain cultivated in 250‐mL flasks.
BIOTECHNOLOGY Paheodactylum tricornutum is widely used in aquaculture as feed for penaid shrimp larve, freshwater prawn larvae, bivalve larvae and postlarvae and marine zooplankton (Tredici et al., 2009). Phaeodactylum has been proposed as a source of eicosapentaenoic acid (EPA, 20:5ω3) (Veloso et al., 1991; Molina Grima et al., 1994). It is a species of interest for biodiesel production. Phaeodactylum extracts are used in cosmetics (Nizard et al., 2007; http://www.cosmeticsdatabase.com/). Fatty acids from Phaeodactylum have shown antibacterial activity (Desbois et al., 2009) and they were found in higher amounts in the fusiform than in the oval cell form (Desbois et al., 2010). Antibacterial and anti‐inflammatory activities were found in polysaccharides (Guzman‐Murillo and Ascencio, 2000; Guzman et al., 2003), while galactolipids were found to induce apoptosis in mammalian engineered cell lines (Andrianasolo et al., 2008).
Growth medium
P. tricornutum is usually grown in Mann and Myers (1968) medium.
Cultivation systems and methods
In Spain, biomass productivity in an outdoor 200‐L airlift tubular photobioreactor operated in continuous was 1.2 – 1,.9 g L−1 day−1 (19 ‐ 32 g m−2 day−1), depending on dilution rate and superficial liquid velocity (Molina et al., 2001). In the same location, in a 75‐L helical tubular photobioreactor biomass productivies up to 1.3 g L−1 day−1 with a photosynthetic efficiency up to 15% were obtained (Hall et al., 2003). In Germany, in an airlift flat panel system with 33‐L modules, a biomass productivy of 0.53‐0.73 g L‐1 day‐1 was obtained in mid‐year months, while on an annual base, an average of 27 mg L‐1 day‐1 of EPA was achieved (http://www.igb.fraunhofer.de/www/gf/umwelt/algen/en/Eicosapentaensaeure_en.html). In Portugal, in 2.2‐m2 ponds, average productivities of 4 g (ash free dry weight) m‐2 day‐1 were achieved, with an EPA productivity of 0.15 g m‐2 day‐1 (Veloso et al., 1991). EPA productivities up to 47.8 mg L‐1 day‐1 were achieved in the airlift tubular photobioreactor, with biomass productivity up to 2.57 g L‐1 day‐1 (Molina Grima et al., 1994).
Phaeodactylum can also grow mixotrophically on different carbon sources, among which glycerol (the by‐product of biodiesel production) yielding higher biomass and EPA productions than those obtained in photoautotrophic conditions (Céron‐Garcia et al., 2005). In three 60‐L reactors, a bubble column and two airlift photobioreactors, outdoors an increase in biomass and EPA productivity was achived by adding glycerol (Fernández Sevilla et al., 2004).
Harvesting methods
Phaeodactylum tricornutm can be harvested by centrifugation, e.g. at at 1800× g (Ibáñez Gonzáles et al., 1998). Flocculation was studied by Veloso et al. (1991) and calcium hydroxide was found to be the best flocculant at concentrations of 30‐100 mg L‐1. The addition of chitosan did not improved the efficiency of flocculation.
References
Alonso D.L., Belarbi E.H., Fernandez‐Sevilla J.M., Rodriguez‐Ruiz J., Molina Grima E. (2000) Acyl lipid composition variation related to culture age and nitrogen concentration in continuous culture of the microalga Phaeodactylum tricornutum. Phytochemistry 54: 461‐471.
Alonso D.L., Belarbi E.L., Rodriguez‐Ruiz J., Segura C.I., Gimenez A. (1998) Acyl lipids of three microalgae. Phytochemistry 47: 1473‐1481.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 131 of 258
Andrianasolo E.H., Haramaty L., Vardi A., White E., Lutz R., Falkowski P. (2008) Apoptosis‐inducing galactolipids from a cultured marine diatom, Phaeodactylum tricornutum. Journal of Natural Products 71: 1197‐1201.
Borowitzka M.A., Volcani B.E. (1978) The polymorphic diatom Phaeodactylum tricornutum ‐ ultrastructure of its morphotypes. Journal of Phycology 14: 10‐21.
Bowler C., Allen A.E., Badger J.H., et al. (2008) The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 456: 239‐244.
Céron Garcia M.C., Sánchez Mirόn A., Fernández Sevilla J.M., Molina Grima E., Garcia Camacho F. (2005) Mixotrophic growth of the microalga Phaeodactylum tricornutum ‐ Influence of different nitrogen and organic carbon sources on productivity and biomass composition. Process Biochemistry 40: 297‐305.
De Martino A., Meichenin A., Shi J., Pan K.H., Bowler C. (2007) Genetic and phenotypic characterization of Phaeodactylum tricornutum (Bacillariophyceae) accessions. Journal of Phycology 43: 992‐1009.
Desbois A.P., Mearns‐Spragg A., Smith V.J. (2009) A fatty acid from the diatom Phaeodactylum tricornutum is antibacterial against diverse bacteria including Multi‐resistant Staphylococcus aureus (MRSA). Marine Biotechnology 11: 45‐52.
Desbois A.P., Walton M., Smith V.J. (2010) Differential antibacterial activities of fusiform and oval morphotypes of Phaeodactylum tricornutum (Bacillariophyceae). Journal of the Marine Biological Association of the United Kingdom 90: 769‐774.
Fernández Sevilla J.M., Cerón García M.C., Sánchez Mirón A., Belarbi el H., García Camacho F., Molina Grima E. (2004) Pilot‐plant‐scale outdoor mixotrophic cultures of Phaeodactylum tricornutum using glycerol in vertical bubble column and airlift photobioreactors: studies in fed‐batch mode. Biotechnology Progress 20: 728‐736.
Francius G., Tesson B., Dague E., Martin‐Jezequel V., Dufrene Y.F. (2008) Nanostructure and nanomechanics of live Phaeodactylum tricornutum morphotypes. Environmental Microbiology 10: 1344‐1356.
Griffiths M.J., Harrison S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Journal of Applied Phycology 21: 493‐507.
Guzman S., Gato A., Lamela M., Freire‐Garabal M., Calleja J.M. (2003) Anti‐inflammatory and immunomodulatory activities of polysaccharide from Chlorella stigmatophora and Phaeodactylum tricornutum. Phytotherapy Research 17: 665‐670.
Guzman‐Murillo M.A., Ascencio F. (2000) Anti‐adhesive activity of sulphated exopolysaccharides of microalgae on attachment of red sore disease‐associated bacteria and Helicobacter pylori to tissue culture cells. Letters in Applied Microbiology 30: 473‐478.
Hall D.O., Acién Fernández F.G., Cañizares Guerrero E., Krishna Rao K., Molina Grima E. (2003) Outdoor helical tubular photobioreactors for microalgal production: Modeling of fluid‐dynamics and mass transfer and assessment of biomass productivity. Biotechnology and Bioengineering 82: 62‐73.
Hu Q., Sommerfeld M., Jarvis E., Ghirardi M., Posewitz M., Siebert M., Darzins A. (2008) Microalgal triacylglycerols as feedstocks for biofuel production: perspectives and advances. The Plant Journal 54: 621‐639.
Ibáñez González M.J., Robles Medina A., Molina Grima E., Giménez Giménez A., Carstens M., Esteban Cerdán L. (1998) Optimization of fatty acid extraction from Phaeodactylum tricornutum UTEX 640 biomass. Journal of the American Oil Chemists’ Society 75: 1735‐1740.
Lebeau T., Robert J.M. (2003a) Diatom cultivation and biotechnologically relevant products. Part I: Cultivation at various scales. Applied Microbiology and Biotechnology 60: 612‐623.
Lebeau T., Robert J.M. (2003b) Diatom cultivation and biotechnologically relevant products. Part II: Current and putative products. Applied Microbiology and Biotechnology 60: 624‐632.
Lewin J.C., Lewin R.A., Philpott D.E. (1958) Observations on Phaeodactylum tricornutum. Journal of General Microbiology 18: 418‐426.
Liang Y., Beardall J., Heraud P. (2006) Effects of nitrogen source and UV radiation on the growth, chlorophyll fluorescence and fatty acid composition of Phaeodactylum tricornutum and Chaetoceros muelleri (Bacillarlophyceae). Journal of Photochemistry and Photobiology B‐Biology 82: 161‐172.
Mann J.E., Myers J. (1968) On pigments, growth, and photosynthesis of Phaeodactylum tricornutum. Journal of Phycology 4: 349‐355.
Mata T.M., Martins A.A., Caetano N.S. (2010) Microalgae for biodiesel production and other applications: A review. Renewable and Sustainable Energy Reviews 14: 217‐232.
Medlin L.K., Kaczmarska I. (2004) Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 43: 245‐270.
Meiser A., Schmid‐Staiger U., Trosch W. (2004) Optimization of eicosapentaenoic acid production by Phaeodactylum tricornutum in the flat panel airlift (FPA) reactor. Journal of Applied Phycology 16: 215‐225.
Molina E, Fernández J., Acién F.G., Chisti Y. (2001) Tubular photobioreactor design for algal cultures. Journal of Biotechnology 92: 113‐131.
Molina Grima E., García Camacho F., Sánchez Pérez J.A., Urda Cardona J., Acién Fernández F.G., Fernández Sevilla J.M. (1994) Outdoor chemostat culture of Phaeodactylum tricornutum UTEX 640 in a tubular photobioreactor for theproduction of eicosapentaenoic acid. Biotechnology and Applied Biochemistry 20: 279‐290.
Monsant A., Jabbari K., Maheswari U., Bowler C. (2005) Comparative genomics of the pennate diatom Phaeodactylum tricornutum. Plant Physiology 137: 500–513.
Nizard C., Friguet B., Moreau M., Bulteau A.L., Saunois A. (2007) Use of Phaeodactylum algae extract as cosmetic agent promoting the proteasome activity of skin cells and cosmetic composition comprising same. WO02/080876‐US Patent No. 7,220,417.
Ohse S., Derner R.B., Ozόrio R.Á., Braga M.V. da C., Cunha P., Lamarca C.P., dos Santos M.E. (2009) Production of biomass and carbon, hydrogen, nitrogen and protein contents in microalgae. Ciencia Rural 39, 1760‐1767.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 132 of 258
Perez E.B., Pina I.C., Rodriguez L.P. (2008) Kinetic model for growth of Phaeodactylum tricornutum in intensive culture photobioreactor. Biochemical Engineering Journal 40: 520‐525.
Rebolloso‐Fuentes M.M., Navarro‐Pérez A., Ramos‐Miras J.J., Guil‐Guerrero J.L (2001) Biomass nutrient profiles of the microalga Phaeodactylum tricornutum. Journal of Food Biochemistry 25: 57‐76.
Reitan K.I., Rainuzzo J.R., Olsen Y. (1994) Effect of nutrient limitation on fatty‐acid and lipid‐content of marine microalgae. Journal of Phycology 30: 972‐979.
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Round F.E., Crawford R.M., Mann D.G. (1990) The diatoms ‐ biology and morphology of the genera. Cambridge University Press, Cambridge, UK.
Rushforth S.R., Johansen J.R., Sorensen D.L. (1988) Occurrence of Phaedactylum tricornutum in the Great Salt Lake, Utah, USA. Great Basin Naturalist 48: 324‐326.
Scala S., Carels N., Falciatore A., Chiusano M.L., Boweler C. (2002) Genome properties of the diatom Phaeodactylum tricornutum. Plant Physiology 129: 993‐1002.
Sheehan J., Dunahay T., Benemann J., Roessler P. (1998) A look back at the U.S. Department of Energy's Aquatic Species Program—biodiesel from algae.
Tonon T., Harvey D., Larson T.R., Graham .IA. (2002) Long chain polyunsaturated fatty acid production and partitioning to triacylglycerols in four microalgae. Phytochemistry 61: 15‐24.
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G., (eds.) New Technologies in Aquaculture: Improving production efficiency, quality and environmental management. Woodhead Publishing Ltd, Cambridge, UK, and CRC Press LLC, Boca Raton, FL, USA. pp. 610–676.
Vardi A., Thamatrakoln K., Bidle K.D., Falkowski P.G. (2008) Diatom genomes come of age. Genome Biology 9: 245. Veloso V., Reis A., Gouveia L., Fernandes H.L., Empis J.A., Novais J.M. (1991) Lipid production by Phaeodactylum tricornutum.
Bioresource Technology 38: 115‐119. Yu E.T., Zendejas F.J., Lane P.D., Gaucher S., Simmons B.A., Lane T.W. (2009) Triacylglycerol accumulation and profiling in the model
diatoms Thalassiosira pseudonana and Phaeodactylum tricornutum (Baccilariophyceae) during starvation. Journal of Applied Phycology 21: 669‐681.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 133 of 258
8.3.3 Chaetoceros muelleri
Figure 46 ‐ Light microscopic picture
of Chaetoceros muelleri.
Instant Algae®
Figure 47 ‐ Scanning electron microscopic picture of Chaetoceros
sp.
Picture form SDSU Center For Inland Water
Figure 48 ‐ Scanning electron microscopic picture of a resting
spore of Chaetoceros sp.
Picture form SDSU Center For Inland Waters
SYMBOLS: D
TAXONOMY
Phylum Bacillariophyta Class Mediophyceae Order Chaetocerotales Family Chaetocerotaceae Genus Chaetoceros Species Chaetoceros muelleri
Related species
Chaetoceros muelleri var. subsalsum Johansen & Rushforth 1985. The taxonomic relationships are not adequately resolved in Chaetoceros (see 1.5) and several cryptic species within C. muelleri are conceivable (Johansen et al., 1990). Potentially important for biofuel: Chaetoceros calcitrans
BIOLOGY
The diatom genus Chaetoceros is among the largest genera of diatoms and comprises several hundreds of species. The genus belongs to the bi‐ or multipolar centric class (Mediophyceae), with Chaetoceros having a bipolar valve outline. The cells occur in chains held together by long spines called setae that extend from the corners of the cells. Many species form resting spores, which are often abundant in the fossil record. Therefore, this genus is important within the disciplines of marine biology, marine geology, oceanography, and aquaculture. Chaetoceros is usually divided in 2 subgenera which are further divided in numerous sections:

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 134 of 258
• subgenus Phaeoceros with species characterized by numerous chloroplasts throughout the cell and robust spiny setae containing chloroplasts.
• subgenus Hyalochaete consisting of species having many parietal plastids and thin setae lacking chloroplasts.
The evolutionary history of the genus Chaetoceros is poorly known, and the classification largely based on observations of the morphology and ultrastructure of some species. Molecular data will be necessary to unravel phylogenetic relationships.
Cells of Chaetoceros are joined in chains that are coiled, curved or straight. Occasionally the cells are solitary. Cells are narrowly to broadly elliptical in valve view and rectangular in girdle view and are united by fusion or interlocking of setae produced from the valve. They contain one or more small plate‐like plastids. Cell width (apical axis) varies with species, roughly ranges from <10 µm to 50 µm. Vegetative cells/colonies are weakly silicified. It can be difficult to identify Chaetoceros taxa because they are notoriously variable in morphology. C. muelleri is also a variable taxon, both in vegetative cells and resting spores and it has been reported under more than 10 different specific names. Most populations of non‐marine, non‐colonial Chaetoceros species have been reported as C. muelleri. Two different forms of C. muelleri have been described: the nominate form C. muelleri var. muelleri is characterized by the frequent presence of a small process on the valve face associated with chain formation (chains of only 2‐4 cells) and smoothly curved setae, while C. muelleri var. subsalsum lacks valvar processes and consequently does not form colonies. Both forms have no ornamentation on their resting spores.
Proximate composition (dry weight percentage) in C. muelleri is 43.1% crude protein, 17.1 % carbohydrates, 21.5% lipids and 18.3% ash.
Chaetoceros is an important marine planktonic genus and some species are major bloom formers in both oceanic and coastal habitats. Only a few species have been recorded in fresh water. Chaetoceros muelleri has been reported mainly from brackish waters of varying salinities and temperatures in Europe and North America. The species is only rarely reported from freshwater habitats and has never been recorded from true marine systems.
BIOTECHNOLOGY
Chaetoceros muelleri is used in the aquaculture industry, primarily as feed for bivalve shellfish and shrimp (Brown et al., 1997). This is due to its fatty acid profile, comprising 5–20% eicosapentaenoic acid (EPA, C20:5ω3), 0.2–1% docosahexaenoic acid (DHA, C22:6ω3) and < 0.2% arachidonic acid (AA, C20:4ω6) of the total fatty acid content and its appropriate size for feeding of the larvae (e.g. 5–8 µm) (Brown et al., 1997).
Chaetocers has been evaluated as a source for biodiesel production, although only at laboratory scale. The average total lipid content for C. muelleri is about 19% cell dry weight under nutrient replete conditions, 27% under N deficient conditions, 36% under Si deficient conditions (Griffiths and Harrison, 2009). The high lipid content of the algae of this genus (up to 40%) is counterbalanced by a low biomass productivity (70 mg L‐1 day‐1) that makes the exploitation of this alga difficult (Rodolfi et al., 2009).
Outdoor mass cultivation of Chaetoceros is usually carried out in the hatcheries, often indoors, in cylinders and tanks that do not allow high productivities (Tredici et al., 2009). In 3000‐L tanks, biomass productivity of C. muelleri ranged from 22.5 to 45.7 mg L‐1day‐1 indoors, while outdoors the mean productivity varied from 29.0 to 69.7 mg L‐1day‐1 in winter and spring, respectively (Lόpez‐Elías et al., 2005). In a 350‐L Flat‐Plate photobioreactor outdoors, Zhang and Richmond (2003) obtained with C. muelleri var.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 135 of 258
subsalsum productivities as high as 0.15 g L‐1 day‐1 with a daily harvest of 10% and a light path length of 20 cm.
References
Blinn D.W. (1984) Growth‐responses to variations in temperature and specific conductance by Chaetoceros muelleri (Bacillariophyceae). British Phycological Journal 19: 31‐35. [BIOLOGY section]
Brown M.R., Jeffrey S.W., Volkman J.K., Dunstan G.A. (1997) Nutritional properties of microalgae for mariculture. Aquaculture 151: 315‐331.
Gran H.H. (1897) Botanik. Prophyta: Diatomaceae, Silicoflagellata og Cilioflagellata. Den Norske Nordhavs‐Expedition 1876–1878. 7, pp. 1‐36. [BIOLOGY section]
Griffiths M.J., Harrison S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Journal of Applied Phycology 21: 493‐507.
Hasle G.R., Syvertsen E.E. (1997) Marine diatoms. In: Tomas C.R. (ed.) Identifying Marine Phytoplankton, Academic Press, San Diego, pp. 5‐385. [BIOLOGY section]
Jaime‐Ceballos B.J., Hernández‐Llamas A., Garcia‐Galano T., Villarreal H. (2006) Substitution of Chaetoceros muelleri by Spirulina platensis meal in diets for Litopenaeus schmitti larvae. Aquaculture 260: 215‐220. [BIOLOGY section]
Johansen J., Rushforth S. (1985) A contribution to the taxonomy of Chaetoceros muelleri Lemmermann (Bacillariophyceae) and related taxa. Phycologia 24: 437‐447. [BIOLOGY section]
Lόpez‐Elías J.A., Voltolina D., Enríquez‐Ocaña F., Gallegos‐Simental G. (2005) Indoor and outdoor mass production of the diatom Chaetoceros muelleri in a mexican commercial hatchery. Aquacultural Engineering 33: 181‐191.
Medlin L.K., Kaczmarska I. (2004) Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 43: 245‐270. [BIOLOGY section]
Ostenfeld C.H. (1903) Plankton from the sea around the Færöes. In: Warming, E. (ed.) Botany of the Færöes, Nordisk Forlag, Copenhagen, pp. 588‐611. [BIOLOGY section]
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Round F.E., Crawford R.M., Mann D.G. (1990) The diatoms ‐ biology and morphology of the genera, Cambridge University Press, Cambridge, UK. [BIOLOGY section]
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G. (eds.) New technologies in aquaculture: Improving production efficiency, quality and environmental management, Woodhead Publishing, Cambridge, pp. 610‐676.
Zhang C.W., Richmond A. (2003) Sustainable, high‐yielding outdoor mass cultures of Chaetoceros muelleri var. subsalsum and Isochrysis galbana in vertical plate reactors. Marine Biotechnology 5: 302‐310.
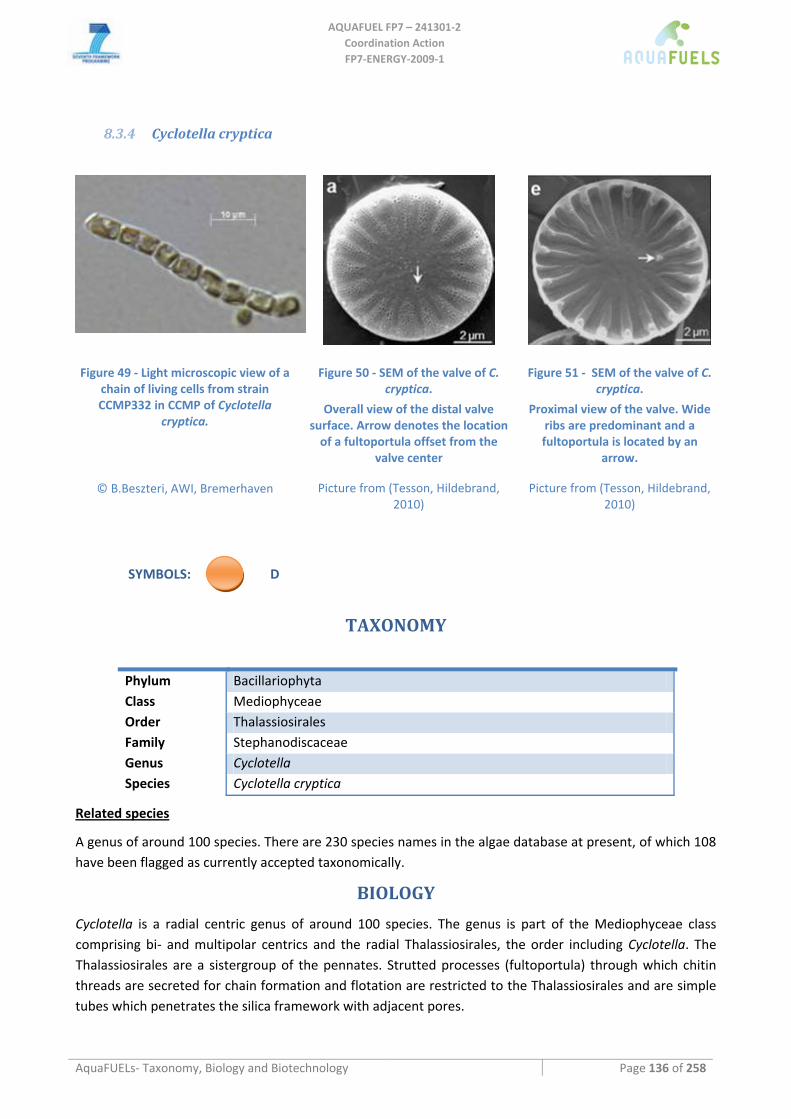
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 136 of 258
8.3.4 Cyclotella cryptica
Figure 49 ‐ Light microscopic view of a chain of living cells from strain CCMP332 in CCMP of Cyclotella
cryptica.
© B.Beszteri, AWI, Bremerhaven
Figure 50 ‐ SEM of the valve of C. cryptica.
Overall view of the distal valve surface. Arrow denotes the location of a fultoportula offset from the
valve center Picture from (Tesson, Hildebrand,
2010)
Figure 51 ‐ SEM of the valve of C. cryptica.
Proximal view of the valve. Wide ribs are predominant and a fultoportula is located by an
arrow. Picture from (Tesson, Hildebrand,
2010)
SYMBOLS: D
TAXONOMY
Phylum Bacillariophyta Class Mediophyceae Order Thalassiosirales Family Stephanodiscaceae Genus Cyclotella Species Cyclotella cryptica
Related species
A genus of around 100 species. There are 230 species names in the algae database at present, of which 108 have been flagged as currently accepted taxonomically.
BIOLOGY
Cyclotella is a radial centric genus of around 100 species. The genus is part of the Mediophyceae class comprising bi‐ and multipolar centrics and the radial Thalassiosirales, the order including Cyclotella. The Thalassiosirales are a sistergroup of the pennates. Strutted processes (fultoportula) through which chitin threads are secreted for chain formation and flotation are restricted to the Thalassiosirales and are simple tubes which penetrates the silica framework with adjacent pores.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 137 of 258
Cyclotella cells are short, drum‐shaped, free‐living or forming filaments, chains or rarely clusters, united by mucilage. They have numerous discoid plastids. C. cryptica exhibits very stiff, thin, crystalline, and chitinous fibril appendages that attach to the pores of its valves. The valves are almost flat to slightly concentrically undulate. The diameter of C. cryptica ranges from 5–25 µm. C. cryptica has been shown to display extreme morphological plasticity depending on salinity. Its frustules show the morphology considered characteristic of C. meneghiniana, a closely related species, when grown in low salinity medium, whereas they produced the typical C. cryptica morphology at higher salinities. C. cryptica can produce auxospores from vegetative cells.
C. cryptica is a planktonic species occurring in marine and brackish environments. The distribution of C. cryptica is widespread. In marine and brackish environments, C. cryptica requires NO3‐ as its source of nitrogen. It is capable of heterotrophic growth in the dark on glucose. This suggests it can survive in bottom waters or muds with high glucose content for extended periods of time. It probably recommences photoautotrophic growth higher in the water column when environmental conditions improve. It is also known to grow photoheterotrophically.
BIOTECHNOLOGY
Cyclotella species are cultured for aquaculture (brine shrimp, Artemia) (Lebeau and Robert, 2003), mainly due to its ability to prodcuce EPA. Wood et al. (1999) compared the growth of C. cryptica on different organic carbon substrates and on medium without organic carbon sources: growth was stimulated by supplementation with glucose and glycerol and was inhibited by acetate, and grew better on nitrate than on ammonia. The fatty acid composition seemed to be affected principally in the degree of unsaturation, that was lower with organic carbon. Under heterotrophic conditions in 19‐L carboys Pahl et al. (2010) reached 2.1 g L‐1 after 12 days of cultivation.
The first successful transformation of a microalgal strain with potential for biodiesel production was achieved in 1994, with successful transformation of the diatoms Cyclotella cryptica and Navicula saprophila (Dunahay et al., 1995). This opened ways for genetic engineering for enhanced lipid production.
In C. cryptica a dramatic increase in the lipid content of the cultures was seen under N‐deficient conditions in cells grown at 30°C: the total lipids, as a percentage of AFDW, increased from 15% to 44%, and the increase in total lipids was due to increases in both the neutral lipid and polar lipid fractions (Sheehan et al., 1998). A Cyclotella species, exhibited an increase of lipid content of more than 40% of dry weight upon Si limitation. However, lipid productivity (9 g m‐2 day‐1), was not significantly different between Si‐deficient and the Si‐sufficient controls, because of the high productivity of the Si sufficient culture (Sheehan et al., 1998).
Biomass productivity for C. cryptica grown in outdoor ponds in California was about 30 g m‐2 day‐1 with a photosynthetic efficiency of 7.0% (Sheehan et al., 1998).
References
Dunahay T.G., Jarvis E.E., Roessler P.G. (1995) Genetic transformation of the diatoms Cyclotella cryptica and Navicula saprophila. Journal of Phycology 31: 1004‐1012.
Herth W., Zugenmaier P. (1977) Ultrastructure of the chitin fibrils of the centric diatom Cyclotella cryptica. Journal of Ultrastructure Research 61: 230‐239. [BIOLOGY section]
Lebeau T., Robert J.M. (2003) Diatom cultivation and biotechnologically relevant products. Part I: Cultivation at various scales. Applied Microbiology and Biotechnology 60: 612‐623.
Medlin LK., Kaczmarska I. (2004) Evolution of the diatoms: V. Morphological and cytological support for the major clades and a taxonomic revision. Phycologia 43: 245‐270. [BIOLOGY section]
Pahl S.L., Lewis D.M., Chen F., King K.D. (2010) Growth dynamics and the proximate biochemical composition and fatty acid profile of the heterotrophically grown diatom Cyclotella cryptica. Journal of Applied Phycology 22: 165‐171.
Round F.E., Crawford R.M., Mann D.G. (1990) The diatoms ‐ biology and morphology of the genera, Cambridge University Press, Cambridge, UK. [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 138 of 258
Schultz M.E. (1971) Salinity‐related polymorphism in the brackish‐water diatom Cyclotella cryptica. Canadian Journal of Botany 49: 1285‐1289. [BIOLOGY section]
Sheehan J., Dunahay T., Benemann J., Roessler P. (1998) A look back at the U.S. Department of Energy's Aquatic Species Program—biodiesel from algae.
Tesson B., Hildebrand M. (2010) Dynamics of silica cell wall morphogenesis in the diatom Cyclotella cryptica: Substructure formation and the role of microfilaments. Journal of Structural Biology 169: 62‐74. [BIOLOGY section]
White A.W. (1974) Growth of two facultatively heterotrophic marine centric diatoms. Journal of Phycology 10: 292‐300. [BIOLOGY section]
Wood B.J.B., Grimson P.H.K., German J.B., Turner M. (1999) Photoheterotrophy in the production of phytoplankton organisms. Journal of Biotechnology 70: 175‐183.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 139 of 258
8.3.5 Odontella aurita
Figure 52 ‐ Light microscopic view showing Odontella aurita
Picture from Algaebase.com (Karl Bruun)
SYMBOLS: D
TAXONOMY
Phylum Bacillariophyta Class Mediophyceae Order Triceratiales Family Triceratiaceae Genus Odontella Species Odontella aurita
Related species There are 76 species (and infraspecific) names in the database at present, of which 11 have been flagged as currently accepted taxonomically. Synonym and related species:
• Odontella aurita var. minima (Grunow) De Toni • Odontella subaequa Kützing
BIOLOGY
Odontella aurita cells are oblong in girdle view, with long 'spines' and raised apical elevations. It often forms chains linked by the processes, with the tubular spines crossing each other. Cells have many plastids, small and discoid. The diameter varies from 10 to 100 µm. Valves are elliptical or lanceolate, with no separation into face and mantle. Valve face is plain or with fine granules, spinules or spines, sometimes with two ridges (which may be fibriate) running on either side delimiting an elliptical area in the centre. At each end there is an elevation, sometimes low and blunt, elsewhere horn‐like, which bears an ocellus. Wall is loculate, with fine external pores and round internal foramina. The edge of the valve mantle is sometimes recurved so that a groove runs around just above the free edge. The spines, which are very variable in length, are actually the exit tubes of the rimoportulae, and are placed in the centre of the valves or close to

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 140 of 258
the bases of the elevations, diagonally opposite each other; they can have small apical spinules. Internally the rimoportulae are sessile and lie in slight depressions. Copulae are split, with ligulae and clustered fimbriae along the advalvar edge; areolae are simple, in vertical rows. The valvocopula is modified to fit the 'sculptured' edge of the valve mantle. O. aurita is a marine alga, planktonic or epiphytic. It is very abundant throughout the oceans.
BIOTECHNOLOGY
O. aurita contains 22% of eicosapentaenoic acid (EPA, 20:5ω3) and 0.9‐1.7% of docosahexaenoic acid (DHA, 22:6ω3) with respect to toal fatty acid content. It has been approved for human consumption in 2002 by the European Community and is mainly sold in form of dried biomass as a dietary supplement (Tredici et al., 2009).
O. aurita is cultivated in open ponds in the Vandée Region in France. It develops from the end of April until half October. With stable growing conditions the average doubling time is 2.6 days and the specific growth rate is of 31% day‐1. O. aurita is harvested mechanically by sedimentation or by filtration, followed by centrifugation. The algal paste is then frozen, and is lyophilised (freeze‐dried) before marketing.
O. aurita is a rich source of the carotenoid fucoxanthin (Moreau et al., 2006). This molecule exhibited cytostatic activity and this effect could have important implications for the application of this kind of microalgae in food manufacturing and the formulation of ocular implant products used in cataract treatment (Moreau et al., 2006).
Odontella aurita is also used in cosmetic industry (Patt, 2008; Redziniak and Donguy, 2009). Although this alga is commercially produced and it is reported to have lipid contents of about 7‐13%
under nutrient replete conditions, no studies have been carried out to assess its biodiesel production potential.
References
Algaebase: http://www.algaebase.org/search/genus/detail/?genus_id=44427 [BIOLOGY section] Moreau D., Tomasoni C., Jacquot C., Kaas R., Le Guedes R., Cadoret J.P., Muller‐Feuga A., Kontiza I., Vagias C., Roussis V., Roussakis
C. (2006) Cultivated microalgae and the carotenoid fucoxanthin from Odontella aurita as potent anti‐proliferative agents in bronchopulmonary and epithelial cell lines. Environmental Toxicology and Pharmacology 22: 97‐103.
Patt L.M. (2008) Methods and compositions for preventing and treating aging or photodamaged skin. US Patent No. 7,384,916. Redziniak G., Donguy D. (2009) Use of fat‐soluble extract of Odontella aurita for restructuring skin, composition for use and
cosmetic method using the extract. Japanese Patent JP2009029806. Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other
uses. In: Burnell G., Allan G. (eds.) New technologies in aquaculture: Improving production efficiency, quality and environmental management, Woodhead Publishing, Cambridge, pp. 610‐676.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 141 of 258
8.3.6 Skeletonema sp.
Figure 53 ‐ Light microscopic view showing Skeletonema costatum
Picture from Algaebase.com (Karl Bruun)
SYMBOLS: D
TAXONOMY
Phylum Bacillariophyta Class Mediophyceae Order Thalassiosirales Family Skeletonemaceae Genus Skeletonema Species
Related species
There are 21 species (and infraspecific) names in the database at present, of which 20 have been flagged as currently accepted taxonomically.
BIOLOGY
Cells are generally colonial and contain one or two cup‐shaped chloroplasts. Cells joined by long marginal processes to form filaments, which appear in the light microscope like short beads joined by numerous fine threads. Valves circular; valve face convex to flat; mantles deep. Valves with a prominent network of costae externally, becoming pseudoloculate near the margin. Internally with distinct cribra on the flat or slightly ribbed surface. A single ring of processes occurs around the top of the mantle. These are closely associated with a ring of fultoportulae, the external openings of which are short tubes hidden in the bases of the processes. The processes are semi‐circular in cross‐section and expand at their apices to form 'knuckles' which interlock with the processes from the adjacent cell; they are sometimes much longer than the cell and can interlock with either one or two processes of the sibling valve. Apical axis is 2‐21 µm while
pervalvar axis is 2‐61 µm. Discrimination among species of the Skeletonema costatum group is very difficult.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 142 of 258
Skeletonema occurs in coastal waters throughout the world where it can be an extremely common diatom. Several species are reported in various oceanic regions but all these need re‐investigation. In addition several fossil species have been described.
BIOTECHNOLOGY
Skeletonema is widely used in aquaculture as bivalve larvae and postlarvae and for penaeid shrimp larve (Tredici et al., 2009). Gross composition is about 40% protein, 20% carbohydrates and 20% lipid and, among fatty acids, about 6% eicosapentaenoic acid (EPA, 20:5ω5) and 2% docosahexaenpic acid (DHA, 22:6ω3) (Brown et al., 1989; Rodolfi et al., 2009).
Skeletonema is worldwide cultivated in hatcheries using low productivity systems suc as tanks, sleeves and cylinders (Tredici et al., 2009). Biomass productivities are usually very low (in the order of 104 cells ml‐1 day‐1, corresponding to tens of milligrams dry weight per liter per day) (Laing, 1991; Popovich et al., 2011). Outdoors in 1000 m3 cultivation volume, made of 20 units of 50‐m3 4‐m depth and 28 m2 surface area, 12 tons of dry biomass are produced yearly (Kitto et al., 1999). Weiss (2008) reports a biomass productivity of about 20 g m‐2 day‐1 cultivating S. costatum in ponds with waste CO2 and turbine cooling seawter. Skeletonema can be harvested through auto‐flocculation and sedimentation (Weiss, 2008).
An extract from the peptidic fraction of Skeletonema costatum containing the active principle named grevilline is used in cosmetics as a skin treatment for erythema (Rocquet and Reynaud, 2008).
Skeletonema has been listed among possible interesting strains for biodiesel production (Rodolfi et al., 2009; Popovich et al., 2011), although at present no data about pilot scale cultivation for this purpose are available.
References
Algaebase: http://www.algaebase.org/search/genus/detail/?genus_id= 43754 [BIOLOGY section] Brown M.R., Jeffrey S.W., Garland C.D. (1989) Nutritional aspects of microalgae used in mariculture; a literature review. CSIRO
Marine Laboratories, Report 205. Kitto M.R., Regunathan C., Rodrigues A. (1999) An industrial photosynthetic system for Skeletonema costatum in arid regions.
Journal of Applied Phycology 11: 391‐397. Laing I. (1991) Cultivation of marine, unicellular algae. Laboratory Leaflet No. 67, Ministry of Agriculture, Fisheroes and Food, UK. Popovich C.A., Damiani C., Constenla D., Leonardi P.I. (2011) Lipid quality of the diatoms Skeletonema costatum and Navicula
gregaria from the South Atlantic Coast (Argentina): evaluation of its suitability as biodiesel feedstock. Journal of Applied Phycology: DOI 10.1007/s10811‐010‐9639‐y.
Rocquet C., Reynaud R. (2008) A natural way to relieve the skin from erythema: grevilline. Cosmetic Science Technology 2008: 129‐136.
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Stazione Zoologica di Napoli [BIOLOGY section]: http://www.szn.it/SZNWeb/cmd/ShowArchiveItem?TYPE_ID=SPECIE&ITEM_ID=6883&LANGUAGE_ID=1&_languageId_=2
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G. (eds.) New technologies in aquaculture: Improving production efficiency, quality and environmental management, Woodhead Publishing, Cambridge, pp. 610‐676.
Weiss H. (2008) Method for growing photosynthetic organisms. Patent WO 2008/107896 A2.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 143 of 258
8.3.7 Thalassiosira pseudonana
Figure 54 ‐ Light microscopic view showing
Thalassiosira pseudonana culture
Picture courtesy of SBAE
Figure 55 ‐ Scanning electron micrograph showing the intricate cell‐wall pattern of T. pseudonana
Photo by Nils Kröger, Georgia Institute of Technology
SYMBOLS: D
.
TAXONOMY
Phylum Bacillariophyta Class Mediophyceae Order Thalassiosirales Family Thalassiosiraceae Genus Thalassiosira Species Thalassiosira pseudonana
Related species There are 186 species names in the algae database at present, of which 82 have been flagged as currently accepted taxonomically. Potentially important for biofuel: T. weissflogii
BIOLOGY
Thalassiosira is a radial centric genus including more than 100 species. The genus is part of the Mediophyceae class comprising bi‐ and multipolar centrics and the radial Thalassiosirales to which Thalassiosira belongs. Fultoportula or strutted processes are the defining character of the order Thalassiosirales. A molecular phylogeny showed the the paraphyletic nature of the families Thalassiosiraceae, Skeletonemaceae, and Stephanodiscaceae within the order and of the genus Thalassiosira, as defined presently. Some species of Thalassiosira may be closer relatives of Stephanodiscus, whereas others are more closely related to some species presently assigned to the genus Cyclotella (both from the family Stephanodiscaceae), as compared with other species of the genus Thalassiosira.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 144 of 258
T. pseudonana was chosen as the first eukaryotic marine phytoplankton for whole genome sequencing. T. pseudonana is a model for diatom physiology studies, belongs to a genus widely distributed throughout the world's oceans, and has a relatively small genome (34 mega base pairs). In spite of the fact that the centric and pennate lineages have only been diverging for 90 million years, their genome structures are dramatically different and a substantial fraction of genes (40%) are not shared by these representatives of the two lineages. T. pseudonana can also be genetically transformed.
Generally Thalassiosira cells contain numerous chloroplasts; cells are disc‐ or drumshaped, occurring as single cells, as chains (connected by a central or several organic threads), or in mucilage colonies. Organic threads usually extrude from strutted processes. The circular, areolated valves possess one or sometimes several central and always marginal strutted processes. They can also be distributed over the whole valve. Resting spores known for some species. The morphological identification of Thalassiosira species is mainly based on ultrastructure details like the number and location of the rimoportulae (tubes on valve connecting the diatom protoplast with the outside) and fultoportulae processes on the valve. Colony configuration and the form of the cells also provide characters for species identification under the light microscope. However, for identification of most species examination of the frustules in the electron microscope is essential. T. pseudonana is a relatively small species (cell diameter 2‐15 µm) with variable valve morphology. The cell diameter is larger than the pervalvar axis. T. pseudonana showed no evidence of the size reduction–restitution cycle that is so unique for the diatoms.
T. pseudonana is a cosmopolitan species living in both fresh and coastal waters.
BIOTECHNOLOGY
Thalassiosira species are cultured in aquaculture because their high nutritional value (Lebeau and Robert, 2003). T. pseudonana is used as feed for bivalve mollusks larvae (Brown et al., 1997; Duerr et al., 1998). The biomass of T. pseudonana contains 30 ‐ 46% protein, 6 ‐ 36% carbohydrates, 21 ‐ 31% lipids depending on harvest stage and on light photoperiod and irradiance level (Brown et al., 1996).
Thalassiosira genes have already contributed to research and commercial efforts to produce vlcPUFAs in transgenic crop plants (Tonon et al., 2004, 2005).
Thalassiosira has been evaluated as a potential biodiesel producer only at laboratort scale. The average total lipid content for T. pseudonana calculated from literature by Griffiths and Harrison (2009) is of 16% cell dry weight under nutrient replete conditions and 26% under N deficient conditions. Cells in logarithmic
phase have high proportions of polar lipids (79 to 89% of total lipid) and low triacylglycerol (≤10% of total lipid). Cells in stationary phase contain less polar lipid (48 to 57% of total lipid) and more triacylglycerol (22 to 45% of total lipid), with an increase in saturated and monounsaturated fatty acids and a decrease in PUFAs (Brown et al., 1996; Zhukova, 2004). Yu et al. (2009) showed a TAG yield from T. pseudonana of about 14–18% of total dry weight. Silicate‐starved cells accumulated an average of 24% more TAGs than those starved for nitrate; however, the chemotypes of the TAGs produced were generally similar regardless of the starvation condition employed. Under laboratory conditions, Rodolfi et al. (2009) found for T. pseudonana low biomass (80 mg L‐1 day‐1) and lipid (17.4 mg L‐1 day‐1) productivities.
References
Armbrust E.V., Berges J.A., Bowler C., et al. (2004) The genome of the diatom Thalassiosira pseudonana: Ecology, evolution, and metabolism. Science 306: 79‐86. [BIOLOGY section]
Bowler C., Allen A.E., Badger J.H., et al. (2008) The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 456: 239‐244. [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 145 of 258
Brown M.R., Dunstan G.A., Norwood S.J., Miller K.A. (1996) Effects of harvest stage and light on the biochemical composition of the diatom Thalassiosira pseudonana. Journal of Phycology 32: 64‐73.
Brown M.R., Jeffrey S.W., Volkman J.K., Dunstan G.A. (1997) Nutritional properties of microalgae for mariculture. Aquaculture 151: 315‐331.
Duerr E.O., Molnar A., Sato V. (1998) Cultured microalgae as aquaculture feeds. Journal of Marine Biotechnology 7: 65‐70. Griffiths M.J., Harrison S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production.
Journal of Applied Phycology 21: 493‐507. Hasle G.R. (1976). Examination of diatom type material: Nitzschia delicatissima Cleve, Thalassiosira minuseula Kraaske, and
Cyclotella nana Hustedt. British Phycological Journal 11: 101‐110. [BIOLOGY section] Hasle G.R., Heimdal B.R. (1970) Some species of the centric diatom genus Thalassiosira studied in the light and electron
microscopes. Nova Hedwigia 31: 559‐597. [BIOLOGY section] Hasle G.R., Syvertsen E.E. (1997) Marine diatoms. In: Tomas C.R. (ed.) Identifying Marine Phytoplankton, Academic Press, San
Diego, pp. 5‐385. [BIOLOGY section] Hoppenrath M., Beszteri B., Drebes G., Halliger H., Van Beusekom J.E.E., Janisch S., Wiltshire K.H. (2007) Thalassiosira species
(Bacillariophyceae, Thalassiosirales) in the North Sea at Helgoland (German bight) and sylt (North Frisian Wadden Sea) ‐ A first approach to assessing diversity. European Journal of Phycology 42: 271‐288. [BIOLOGY section]
Kaczmarska I., Beaton M., Benoit A.C., Medlin L.K. (2005) Molecular phylogeny of selected members of the order Thalassiosirales (Bacillariophyta) and evolution of the fultoportula. Journal of Phycology 42: 121‐138. [BIOLOGY section]
Lebeau T., Robert J.M. (2003) Diatom cultivation and biotechnologically relevant products. Part I: Cultivation at various scales. Applied Microbiology and Biotechnology 60: 612‐623.
Poulsen N., Chesley P., Kröger N. (2006) Molecular genetic manipulation of the diatom Thalassiosira pseudonana (Bacillariophyceae). Journal of Phycology 42: 1059–1065. [BIOLOGY section]
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Round F.E., Crawford R.M., Mann D.G. (1990) The diatoms ‐ biology and morphology of the genera, Cambridge University Press, Cambridge, UK. [BIOLOGY section]
Tonon T., Harvey D., Qing R, Li Y., Larson T.R., Graham I.A. (2004) Identification of a fatty acid Δ11‐desaturase from the microalga Thalassiosira pseudonana. FEBS Letters 563: 28‐34.
Tonon T., Qing R.W., Harvey D., Li Y., Larson T.R., Graham I.A. (2005) Identification of a long‐chain polyunsaturated fatty acid acyl‐coenzyme a synthetase from the diatom Thalassiosira pseudonana. Plant Physiology 138: 402‐408.
Yu E.T., Zendejas F.J., Lane P.D., Gaucher S., Simmons B.A., Lane T.W. (2009) Triacylglycerol accumulation and profiling in the model diatoms Thalassiosira pseudonana and Phaeodactylum tricornutum (Baccilariophyceae) during starvation. Journal of Applied Phycology 21: 669‐681.
Zhukova N.V. (2004) Changes in the lipid composition of Thalassiosira pseudonana during its life cycle. Russian Journal of Plant Physiology 51: 702‐707.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 146 of 258
8.4 Eustigmatophyceae (phylum Heterokontophyta)
8.4.1 Monodus subterraneus
Figure 56 ‐ Light microscopic view showing Monodus sp.
Picture from UNIFI
SYMBOLS: D
TAXONOMY
Phylum Heterokontophyta Heterokontophyta Class Xanthophyceae Eustigmatophyceae Order Mischococcales ‐‐‐‐‐ Family Pleurochloridaceae Monodopsidaceae Genus Monodus Monodospis Species Monodus subterraneus Monodospis subterranea
Related species
This genus is of uncertain collocation. Most of the algae collection display Monodus under the Eustigmatophyceae, but several taxonomies still report this genus under the Xanthophyceae. Pigments are similar to Eustigmatophyceae while its cell structure is different from the standard eustimatophyte structure. There are 17 species (and infraspecific) names in the database at present. Synonym:
• Monodus subterraneus = Monodopsis subterranea • Monodus ovalis = Pseudocharaciopsis ovalis
BIOLOGY
Monodus has been previously classified as a xanthophyte, then it has been moved to eustigmatophytes, but it has intermediate characters. Cells are solitary, 5–10 μm long and less than 10 μm in diameter. Cells are free‐floating, spherical, ovoid, elliptical or cylindrical in shape. Chloroplast is terminal, yellow‐green, with embedded pyrenoid. Reproduction occurs by autospores and zoospores; aplanospores and cysts have also been noted. In Monodus subterraneus (syn. Monodopsis subterranea) zoospores are not present. Monodus

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 147 of 258
is euplanktonic and metaphytic in dystrophic, mesotrophic, and eutrophic ponds, pools, seeps, and lakes often in association with Sphagnum, sedges, and/or Typha.
BIOTECHNOLOGY
Monodus subterraneus has been cultivated at pilot plant for its ability to produce high amounts of eicosapentaenoic acid (EPA, 20:5ω5), which synthesis in this alga has been widely studied (Khozin‐Goldberg et al., 2002; Liu and Lin, 2005; Khozin‐Goldberg and Cohen, 2006). At present, only one ditary supplement conatianing Monodus for EPA is commercialised.
Under artificial illumination, in a cultivation system consisting of a helical tubular photobioreactor (‘Biocoil’) constructed from transparent PVC tubing to contain a total volume of 4.5 liter, Lu et al. (2001) found that biomass and EPA productivities were 1.3‐1.7 g L‐1 day‐1 and 44‐56 mg L‐1 day‐1, respectively.
Outdoors, M. subterraneus was cultivated in a flat, inclined modular photobioreactor (FIMP), consisting of a series of four individual 14‐L glass reactors measuring 70 cm high, 90 cm long and 2.8 cm wide, Qiang et al. (1997) obtained a maximal EPA productivity of 58.9 mg L‐1 day‐1 and a biomass productivity of 1.55 g L‐1 day‐1 at the optimal cell density of 4 g L‐1. Lu et al. (2002) cultivated M. subterraneus in a 75‐L helical reactor and in a 57‐L bubble column obtaining low biomass productivity in both photobioreactors, although productivity was higher in the helical reactor than in the bubble column regardless of the operational conditions, with maximum values of 0.54 and 0.16 g L‐1 d‐1, respectively, and a maximum EPA productivity of 9 mg L‐1 d‐1.
Monodus has been tested in laboratory as a potential strain for biodiesel production (Rodolfi et al., 2009).
References
Algaebase: http://www.algaebase.org/search/genus/detail/?genus_id= 45848 [BIOLOGY section] Khozin‐Goldberg I., Cohen Z. (2006) The effect of phosphate starvation on the lipid and fatty acid composition of the fresh water
eustigmatophyte Monodus subterraneus. Phytochemistry 67: 696‐701. Khozin‐Goldberg I., Didi‐Cohen S., Shayakhmetova I., Cohen Z. (2002) Biosynthesis of eicosapentaenoic acid (EPA) in the freshwater
eustigmatophute Monodus subterraneus (Eustigmatophyceae). Journal of Phycology 38: 745‐756. Liu C.P., Lin L.P. (2005) Morphology and eicosapentaenoic acid production by Monodus subterraneus UTEX 151. Micron 36: 545‐
550. Lu C., Rao K., Hall D., Vonshak A. (2001) Production of eicosapentaenoic acid (EPA) in Monodus subterraneus grown in a helical
tubular photobioreactor as affected by cell density and light intensity. Journal of Applied Phycology 13: 517‐522. Lu C., Acién Fernández F.G., Cañizares Guerrero E., Hall D.O., Molina Grima E. (2002) Overall assessment of Monodus subterraneus
cultivation and EPA production in outdoor helical and bubble column reactors. Journal of Applied Phycology 14: 331‐342. Ott D.W., Oldham‐Ott C.K. (2003) Eustigmatophyte, Raphidophyte, and Tribophyte Algae. In: Wehr J.D., Sheath R.G. (eds.)
Freshwater Algae of North America: Ecology and Classification, Elsevier, Rotterdam, pp. 423‐469. [BIOLOGY section] Qiang H., Zhengyu H., Cohen Z., Richmond A. (1997) Enhancement of eicosapentaenoic acid (EPA) and γ‐linolenic acid (GLA)
production by manipulating algal density of outdoor cultures of Monodus subterraneus (Eustigmatophyta) and Spirulina platensis (Cyanobacteria). European Journal of Phycology 32: 81‐86.
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Whittle S.J, Casselton P.J. (1975) The chloroplast pigments of the algal classes Eustigmatophyceae and Xanthophyceae. I. Eustigmatophyceae. European Journal of Phycology 10: 179‐191. [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 148 of 258
8.4.2 Nannochloropsis sp.
Figure 57 ‐ Light microscopic view showing Nannochloropsis salina
Picture from www.sb‐roscoff.fr/Phyto/gallery/main.php?g2_itemId=323
SYMBOLS: D, PIV
TAXONOMY
Phylum Heterokontophyta Class Eustigmatophyceae Order Eustigmatales Family Monodopsidaceae Genus Nannochloropsis Species
Related species
There are 6 species (and infraspecific) names in the database at present, all taxonomically accepted.
Species of interest for biofuel:
N. gaditana, N. granulata, N. limnetica, N. oceanica, N. oculata, N. salina.
BIOLOGY Structural and morphological features
Nannochloropsis cells (Karlson et al., 1996; Fawley and Fawley, 2007) are non‐motile, spherical to ovoid, 2–4 μm in diameter, with a single chloroplast lacking a pyrenoid and containing chlorophyll a. Major carotenoids are violaxanthin, vaucheraxanthin and neoxanthin, besides β‐carotene. Among others, algae of this genus can synthesize low amounts of canthaxanthin and astaxanthin. Zoospores are not present, then reproduction is by autospores (2 to 6). Species attribution is mainly carried out by genetic analysis (rbcL and 18S rDNA). Cells present the so‐called “red body”, a primary characteristic of the vegetative cells of many eustigmatophytes, that is an eyespot located extraplastidially and not surrounded by membranes. The composition of the cell wall is still unclear. Small refractile bodies can also be observed within the cytoplasm and can be both mobile (Brownian motion) or immobile. The genus Nannochloropsis includes

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 149 of 258
both marine and freshwater species, though the biotechnology of this alga is at present limited to the marine species.
Biochemical composition of algae and aquatic biomass main constituents
Under artificial light in a bubble‐mixed 29‐L photobioreactor, Rebelloso Fuentes et al. (2001) have found an average dry biomass composition of: 29% protein, 36% carbohydrate, 18% lipid, 9% ash, and plamitic, palmitoleic, oleic and eicosapentaenoic acid (on average 2.2%) as major fatty acids. Mineral composition was on average (mg/100g d.wt): 659 Na, 533 K., 972 Ca, 316 Mg, 136 Fe, 103 Zn, 35 Cu, 3.4 Mn, 529 S. Brown et al. (1998) report a different composition for Nannochloropsis CS‐246 tested for oyster rearing: 17% protein, 23% carbohydrate, 26% lipid, 16% ash.
Nannochloropsis is widely cultivated for its high content of eicosapentaenoic acid (EPA; 20:5 ω3), present in amounts up to 4‐5 % of the biomass, and because of its small cells (2‐3 μm diameter) as feed for rotifers in the “green water” technique. Modulation of fatty acid composition by culture parameters such as light intensity, light‐dark cycles, temperature, salinity and nutrients has been widely investigated and reviewed by Sukenik (1999). Patterns of variation in the lipid class and fatty acid composition during batch cultivation of N. oculata have also been investigated (Hodgson, et al. 1991). Generally, the higher the biomass productivity, the higher is EPA productivity (Chini Zittelli et al. 1999).
Nannochloropsis has been also proposed as source of lipid for biodiesel production because it is able to reach over 60% lipids after nitrogen starvation (Shifrin and Chrisholm, 1980; Rodolfi et al., 2009) mainly as TAGs containing saturated and monounsaturated fatty acids (Bondioli et al., 2010).
BIOTECHNOLOGY Culture Media
Nannochloropsis grows in seawater, with maximal growth rate in the salinity range from 25 to 30 g/L, but it can tolerate salinities between 10 and 35 g/L (Renaud and Parry, 1994). Seawater sources can be both natural and artificial seawater; the last one made either from single salts or from a mixture of seawater salts available on the market. F/2 or F medium (Guillard and Ryther, 1962) is widely used for cultivation of Nannochloropsis.In industrial‐scale microalgae plants, where culture volumes and thus amounts of culture medium to be prepared and treated before discharge are considerable and highly expensive, particularly when artificial seawater is used, the possibility of reutilizing the “exhaust” growth medium after biomass separation would be of high relevance. However, Richmond and Zou (1999) report the presence of inhibitory substances, released from the cells into to culture medium which may reduce growth and limit the degree of recycling. Moreover it was observed (Rodolfi et al., 2003) that the accumulation of metabolites produced by the microalga, together with particulate material such as cell walls released after cell division and bacteria, causes growth cessation after two weeks. Reutilization of culture medium seams possible only after treatment with activated charcoal filters.
Cultivation methods and production systems
In hatcheries, Nannochloropsis is mainly cultivated indoors, in batch and semi‐batch, in polyethylene bags, cylinders (Fulks and Main, 1991) and annular columns. Experiments with artificial or mixed (artificial and natural) light sources aiming to evaluate biomass productivity in different culture conditions have been carried out in alveolar and glass panels, annular columns, biofence systems and a new devised helical tubular reactor with continuous harvesting regimen (Chini Zittelli et al., 2000, 2003; Zou et al., 2000; Sandnes et al., 2005; Briassoulis et al., 2010).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 150 of 258
Cultivation in large scale outdoor raceway ponds up to 3000 m2 (20 cm depth) has been carried out in Eilat, Israel (Sukenik et al., 1993) by daily harvesting 10‐50% of pond volume. Average productivity of 7 and 22 g m‐2 were recorded in winter and in summer, respectively (Sukenik, 1991). The authors reported a culture collapse after 2 months of cultivation because of grazers and contaminating algal species. At present in Israel Seambiotic cultivated Nannochloropsis in raceways pond using flue gas to produce biomass as feedstock for biodiesel. In Japan Nannochloropsis oculata is extensively cultivated in tanks kept outdoors with seawater and chemical fertilizers (Okauchi, 1991). Decrease in cell density was recorded in the rainy season and the phenomenon was mainly attributed to predation by protozoa, partially solved by chlorination and application of terramycin. Boussiba et al. (1987) found effective the use of urea and NH4
+ as nitrogen source (instead of NO3
‐) at a concentration of 2‐5 mM against contaminating diatoms and predators.
Chini Zittelli et al. (1999) cultivated Nannochloropsis outdoors in an air‐mixed near horizontal tubular reactor with dimensions of up to 30 m2 (600 L of culture) kept in semicontinuous regimen. With a reactor of this type but of a smaller size, a productivity of about 15 g m2 day‐1 was obtained in June. Outdoor Nannochloropsis cultivation was also performed by using a 500 L glass plate reactor (Zhang et al., 2001) and Green‐Wall Panels. In the first reactor productivities of 10 and 14.2 g m2 illuminated surface area per day were obtained in winter and in summer, respectively. In the second reactor productivities from 9 to 20 g m2 per ground area per day, depending on cultivation season and on panel orientations, were achieved (Tredici, unpublished data).
Nannochloropsis has been cultivated in a 2800 liters tubular photobioreactor (50 m2 overall surface area) at the “Estación Experimental de Cajamar Las Palmerillas”(Spain). In February, with a daily dilution rate of about 35% (five days per week), the average biomass concentration was 1.2 g L‐1 and the volumetric productivity about 0.42 g L‐1 day‐1 (Prof. Acien personal communication).
Nannochloropsis has also been proposed as source of oil for biodiesel production. Rodolfi et al. (2009) have suggested a “two phase strategy” (a first‐stage cultivation of the microalga in the presence of nitrogen carried out in closed photobioreactors, followed by a second stage of nitrogen starvation, preferably carried out in open ponds), and have estimated a productivity of 20 ton of lipids per ha per year in the Mediterranean region. Lipid content increase after nitrogen starvation was not observed in N. salina (Boussiba et al., 1987), thus the selection of the right strain is of crucial importance for this application.
Harvesting methods
Due to Nannochloropsis small cell size, the most adequate system for biomass recovery is centrifugation, that gives a product (paste) with 25‐30% dry material. To reduce the volume to be centrifuged, autoflocculation can be used. The pH increase, consequent to the interruption of CO2 inlet in the culture exposed to sunlight, can cause cell aggregation and thus sedimentation, which allows to concentrate the culture 10‐20 times. The same occurs by addition of NaOH. Flocculation of Nannochloropsis by using cationic starch was not effective (Vandamme et al., 2010).
From experiments carried out at lab scale with two algae, Nannochloropsis and Scenedesmus, a potential reduction of harvesting costs of 20% was proposed as a result of pre‐treatment to centrifugation with submergible microfilatration or ultrafiltration membranes (Gori et al., 2010).
Biomass processing
Umdu et al. (2009) found that lipid transesterification with heterogeneous catalysts (Al2O3 supported CaO and MgO catalyst) was more active than with pure CaO and MgO in the production of biodiesel from N. oculata.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 151 of 258
Patil et al. (2011) demonstrated a one‐step process for direct liquefaction and conversion of wet Nannochloropsis biomass (50% lipid) to biodiesel under supercritical methanol conditions, a process allowing the simultaneous extraction and transesterification of the biomass. The optimal conditions for the process were: ratio between wet algae and methanol around 1:9, 255 °C and 25 min reaction time. These milder conditions compared to those necessary for pyrolisis seem to prevent formation of by‐products. Other processes to obtain oil and biodiesel from Nannochloropsis biomass, such as catalytic Mcgyan® process, hydrothermal liquefaction and gasification of the biomass, direct pyrolisis and catalytic pyrolysis of Nannochloropsis residue have been developed (Krohn et al. 2011, Brow et al., 2010, Biller and Ross, 2011; Pan et al., 2010)
Scaling up limitation
1) Low tolerance to temperatures above 30°C (Sukenik et al., 2009; Sandnes et al., 2005). Temperatures above 30 °C can cause reduction in productivity and formation of cell aggregates which favour contamination by bacteria and grazers (Rodolfi et al., 2003) and further depress productivity. Accurate (and expensive) cooling of the culture is thus necessary.
2) Small cell size allows less energy consumption for mixing, but more energy is required for separating the cells from the culture medium.
3) Thick cell wall, that allows mixing of the culture even with centrifugal pumps, but increases difficulty of cell breakage, which is necessary for oil extraction.
HIGHLIGHTS IN BIOTECHNOLOGY • Nannochloropsis is able to accumulate lipids in amounts higher than 50% of dry biomass after
nitrogen starvation • Nannochloropsis is able to accumulate lipids mainly as TAG and saturated and monounsaturated
fatty acids, more suitable for biodiesel production • After oil extraction the residue is a valuable PUFA‐ rich fraction that can be used as feed or food
ingredient
References
Biller P., Ross A.B. (2011) Potential yields and properties of oil from the hydrothermal liquefaction of microalgae with different biochemical content. Bioresource Technology 102: 215‐225.
Bondioli P., Della Bella L., Rivolta G., Casini D., Prussi M., Chiaramonti D., Chini Zittelli G., Bassi N., Rodolfi L., Tredici M. (2010) Oil production by the marine microalga Nannochloropsis sp. F&M‐M24. Proceedings of the 18th European Biomass Conference and Exhibition, 3‐7 May, Lyon, France.
Boussiba S., Vonshak A., Cohen Z., Avissar Y., Richmond A. (1987) Lipid and biomass production by the halotolerant microalga Nannochloropsis salina. Biomass 12: 37‐47.
Briassoulis D., Panagakig P., Chionidis M., Tzenos D., Lalos A., Tsinos C., Berberidis K., Jacobsen A. (2010) An experimental helical‐tubular photobioreactor for continuous production of Nannochloropsis sp. Bioresource Technology 101: 6768‐6777.
Brown M.R., McCausland M. A., Kowalsi K. (1998) The nutritional value of four Australian microalgal strains fed to Pacific oyster Crassostrea gigas spat. Aquaculture 165: 281‐293.
Brown T.M., Duan P., Savage P.E. (2010) Hydrothermal liquefaction and gasification of Nannochloropsis sp. Energy Fuels 24: 3639‐3646.
Chini Zittelli G., Lavista F., Bastianini A., Rodolfi L., Vincenzini M., Tredici M.R. (1999) Production of eicosapentaenoic by Nannochloropsis . cultures in outdoor tubular photobioreactors. Journal of Biotechnology 70: 299‐317.
Chini Zittelli G., Pastorelli R., Tredici M.R. (2000) A modular flat panel photobioreactor (MFPP) for indoor mass cultivation of Nannochloropsis sp. under artificial illumination. Journal Applied Phycology 12: 521‐526.
Chini Zittelli G., Rodolfi L., Tredici M.R. (2003) Mass cultivation of Nannochloropsis sp. in annular reactors. Journal Applied Phycology 15: 107‐114.
Fawley K.P., Fawley M.W. (2007) Observations on the diversity and ecology of freshwater Nannochloropsis (Eustigmatophyceae), with descriptions of new taxa. Protist 158: 325‐336.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 152 of 258
Fulks M., Main K.L. (eds) (1991) Rotifers and microalgae culture systems. Proceedings of a US‐Asia Workshop. The Oceanic Institute, Honolulu.
Gori R., Munz G., Lubello C., Daddi D., Biondi N., Tredici M.R. (2010) Energy demand and economic evaluation of solid‐liquid separation systems used for harvesting and concentration of cultivated microalgae. Venice 2010, Third International Symposium on Energy from Biomass and Waste Proceedings.
Guillard R.R.L., Ryther J.H. (1962) Studies of marine planktonic diatoms. I. Cyclotella nana (Hustedt) and Detonula confervacea (Cleve). Canadian Journal of Microbiology 8: 229‐239.
Hodgson P.A., Henderson R.J., Sargent J.R., Leftley J.W. (1991) Patterns of variation in the lipid class and fatty acid composition of Nannochloropsis oculata (Eustigmatophyceae) durting batch culture. I The growth cycle. Journal Applied Phycology 3: 169‐181.
Karlson B., Potter D., Kuylenstierna M., Dersen M.A. (1996) Ultrastructure, pigment composition, and 18S rRNA gene sequence for Nannochloropsis granulata sp. nov. (Monodopsidaceae, Eustigmatophyceae), a marine ultraplankter isolated from the Skagerrak, northeast Atlantic Ocean. Phycologia 35: 253‐260.
Krohn B.J.K., McNeff C.V., Yan B., Nowlan D. (2011) Production of algae‐based biodiesel using the continuous catalytic Mcgyan® process. Bioresource Technology 102: 94‐100.
Okauki M. (1991) The status of phytoplankton production in Japan. In: Fulks M., Main K.L. (eds) Rotifers and microalgae culture systems. Proceedings of a US‐Asia Workshop. The Oceanic Institute, Honolulu.
Pan P., Hu C., Yang W., Li Y., Dong L., Zhu L., Tong D., Qing R., Fan Y. (2010) The direct pyrolysis and catalytic pyrolysis of Nannochloropsis sp. residue for renewable bio‐oils. Bioresource Technology 101: 4593‐4599.
Patil P.D., Gude V.G., Mannarswamy A., Deng S., Cooke P., Munsun‐McGee S., Rhodes I., Lammers P., Nirmalakhandan N. (2011) Optimization of direct conversion of wet algae to biodiesel under supercritical methanol conditions. Bioresource Technology 102: 118‐122.
Rebelloso‐Fuentes M.M., Navarro‐Pérez A., García‐Camacho F., Ramos‐Miras J.J., Guil‐Guerrero J.L. (2001) Biomass nutrient profiles of the microalga Nannochloropsis. Journal of Agriculture and Food Chemistry 49: 2966‐2972.
Renaud S.M., Parry D.L. (1994) Microalgae for use in tropical aquaculture. II Effect of salinity on growth, gross chemical composition and fatty acid composition of three species of marine microalgae. Journal of Applied Phycology 6: 347‐356.
Richmond A., Zou N. (1999) Efficient utilisation of high photon irradiance for mass production of photoautotrophic micro‐organisms. Journal of Applied Phycology 11: 123‐127
Rodolfi L., Chini Zittelli G., Barsanti L., Rosati G., Tredici M.R. (2003) Growth medium recycling in Nannochloropsis mass cultivation. Biomolecular Engineering : 243‐248.
Rodolfi L., Chini Zittelli G., Bassi N., Padovani N., Biondi N., Bonini G, Tredici MR. (2009) Microalgae for oil: strain selection, induction of lipid synthesis and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Sandnes J.M., Källqvist T., Wenner D., Gislerød H.R. (2005) Combined influence of light and temperature on growth rates of Nannochlorpsis oceanica: linking cellular responses to large‐scale biomass production. Journal of Applied Phycology 17: 515‐525.
Shifrin M.S., Chrisholm S.W. (1980) Phytoplankton lipids: environmental influences on production and possible commercial applications. In: Shelef G., Soeder C.J. (eds.) Algae Biomass, Elsevier North Holland Biomedical Press, Amsterdam, pp. 624‐645.
Sukenik A. (1999) Production of eicosapentaenoic acid by the marine eustigmatophyte Nannochloropsis. In: Cohen Z. (ed.) Chemicals from Microalgae, Taylor & Francis Ltd, London, pp. 41‐ 56.
Sukenik A., Zmora O., Carmeli Y. (1993) Biochemical quality of masrine unicellular algae with special enphasis on lipid composition. II, Nannochloropsis sp. Aquaculture 117: 313‐326
Sukenik A., Beardall J., Kromkamp J.C., Kopeck J., Masojídek J., van Bergeik S., Gabai S., Shaaham E., Yamshon A. (2009) Photosyntetic performance of outdoor Nannochloropsis mass cultures under a wide range of environmental conditions. Aquatic Microbial Ecology 56: 297‐308.
Umdu E. S., Tuncer M., Seker E. (2009) Transestrification of Nannochloropsis oculata microalga’s lipid to biodiesel on Al2O3 supported CaO and MgO catalysts. Bioresource Technology 100: 2828‐2831.
Vandamme D., Foubert I., Meesschaert B., Muylaert K. (2010) Flocculation of microalgae using cationic starch. Journal of Applied Phycology 22: 525‐530.
Zhang C.W., Zmora O., Kopel R., Richmond A. (2001) An industrial flat plate galls reactor for mass production of Nannochloropsis sp. (Eustigmatophyceae). Aquaculture 195: 35‐49.
Zou N., Zhang C.W., Cohen Z, Richmond A. (2000) Production of cell mass and eicosapentaenoic acid (EPA) in ultrahigh cell density cultures of Nannochloropsis sp. (Eustigmatophyceae). European Journal of Phycology 35: 127‐133.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 153 of 258
8.5 Haptophyta
8.5.1 Isochrysis sp.
Figure 58 ‐ Light microscopic view showing Isochrysis sp.
Picture from UNIFI
SYMBOLS: D
TAXONOMY
Phylum Haptophyta Class Prymnesiophyceae Order Isochrysidales Family Isochrysidaceae Genus Isochrysis Species
Related species
There are 3 species names in the database at present, all taxonomically accepted.
Species of interest for biofuel:
Isochrysis galbana
BIOLOGY
Isochrysis presents slightly elongated motile cells with a capacity to change shape, a stigma, and two smooth apical flagella once to twice the length of the cells. Parke has mentioned for strains of I. galbana, the type‐species, the possibility of an important benthic phase, and, furthermore, the presence of haptophycean scales on the cell‐body of the motile cells. These scales are very difficult to detect, being embedded in a thick mucilage matrix. There is also a very reduced haptonema. I. galbana cell has two

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 154 of 258
chloroplasts found in two sides of the cell, and osmiophilic granules with median electron density in the chloroplasts.The pyrenoid is situated in the chromatophore.
Two Isochrysys species are mainly benthic: I. maritima and I. litoralis. Isochrysis maritima cells can be seen to be hemispherical, 3 x 6 μm, forming regular cubic masses of four to eight cells (typical aspect of the young culture). The cell‐walls are pectic. A small red stigma is present and the swarmers have a positive phototactic reaction. Each cell contains one or two parietal yellow‐green chloroplasts without any detectable pyrenoids. Preliminary thin‐sectioning though, has revealed an intraplastidial slightly bulging pyrenoid. Each cell contains a chrysolaminarin vacuole, fat globules, and peripheral muciferous bodies which are visible only when stained. In older cultures, the cells become globose and surrounded by various concentric mucilage envelopes. Asexual reproduction occurs in this species by division of the non‐motile individuals and also by production of swarmers. The latter are liberated from thick‐walled sporangia. The swarmers are elongated (3 x 4.5‐5 μm) and show a capacity to change shape. Two flagella rise anteriorly; they are slightly unequal, 6 and 8 μm in length. The method of swimming of this species is highly characteristic: the flagella behave homodynamically, movement of the individuals, generally in a backward direction, being rapid, with rotation of the body round the long axis. In I. litoralis young cells are non‐motile, hemispherical (4‐‐6 x 6‐8 μm ) or round (5 μm) while older cells are spherical varying in size (5‐15 μm). It has a thick pectic envelope. There are two parietal golden‐yellow chromatophores with one intraplastidial pyrenoid per chromatophore. Asexual reproduction occurs by division of the non‐motile cells and by production of elongated swarmers (4 x 7 μm) with two apical subequal (7‐8 and 8‐9 μm) homodynamic flagella. Swimming movements are similar to those of Isochrysis maritima. Cells are covered with one layer of oval plate scales (0.3 x 0.18 μm) with a thickened margin and radiating ridges.
BIOTECHNOLOGY
Isochrysis is commonly used in the aquaculture industry (Tredici et al., 2009). For its high docosahexaenoic acid (DHA, 22:6ω3) content it is often used to enrich zooplankton such as rotifers or Artemia. It is a primary algae used in shellfish hatcheries (oysters, clams, mussels, and scallops) and in some shrimp hatcheries.
Isochrysis has been proposed a s a souce of DHA for nutraceuticals and dietary supplements (Liu and Lin, 2001; Poisson and Ergan, 2001; ) although at present only one commercial product containg Isochrysis is in the market. Isochrysis sp. extracts have been atented for use in cosmetics (Herrmann et al., 2010).
Chini Zittelli et al. (2004) outdoors in 120‐L annular columns obtained with Isochrysis sp. a volumetric productivity of 0.31 g L‐1 day‐1, while in a full‐scale arrangement the overall areal productivity (including both the area occupied by the columns and the space between two rows of columns, as described by Tredici, 2004) was of 32.1 g m‐2 day‐1. In a 2.5‐m2 pond the production rate of Isochrysis galbana biomass was 29.5 g m‐2 day‐1 and that of lipids 6.5 g m‐2 day‐1, while, in a 100‐m2 pond, the average productivities of biomass and of lipids were 23.5 g m‐2 day‐1 and 5.6 g m‐2 day‐1, respectively (Boussiba et al., 1988). Harvesting can be achieved by means of flocculation with FeCl3 and dissolved air flotation (Boussiba et al., 1988).
Isochrysis has been tested at laboratory scale for biodiesel production (Rodolfi et al., 2009); however the culture carried out by Boussiba et al. (1988), although not intended for biodiesel production, evidenced the suitability of this alga as biodiesel feedstock.
References
BillardC., Gayral P. (1972) Two new species of Isochrysis with remarks on the genus Ruttnera. European Journal of Phycology 7: 289‐297. [BIOLOGY section]
Boussiba S., Sandbank E., Shelef G., Cohen Z., Vonshak A., Ben‐Amotz A., Arad S., Richmond A. Outdoor cultivation of the marine microalga Isochrysis galbana in open reactors. Aquaculture 72: 247‐253.
Chini Zittelli G., Somigli S., Rodolfi L., Tredici M.R. (2004) Outdoor mass cultivation of Isochrysis sp. in annular columns. 1er Congreso Latinoamericano sobre Biotecnologia Algal, Argentina 2004. ISBN No. 987‐1130‐32‐5, pp. 45‐48.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 155 of 258
Herrmann M., Joppe H., Pertile P., Zanella L. (2010) Extracts of Isochrysis sp. Patent Application No. US2010/0080761 A1. Liu C.P., Lin L.P. (2001) Ultrastructural study and lipid formation of Isochrysis sp. CCMP1324. Botanical Bulletin of Academia Sinica
42: 207‐214. Poisson L., Ergan F. (2001) Docosahexaenoic acid ethyl esters from Isochrysis galbana. Journal of Biotechnology 91: 75‐81. Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection,
induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G. (eds.) New technologies in aquaculture: Improving production efficiency, quality and environmental management, Woodhead Publishing, Cambridge, pp. 610‐676.
Tredici M.R. (2004) Mass production of microalgae: photobioreactors. In: Richmond A. (ed.) Handbook of Microalgal Cultures, Biotechnology and Applied Phycology, Blackwell, Oxford, pp. 178‐214.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 156 of 258
8.5.2 Pavlova sp.
Figure 59 ‐ Light microscopic view showing Pavlova lutheri
Picture from http://www.eol.org/
SYMBOLS: D
TAXONOMY
Phylum Haptophyta Class Pavlovophyceae Order Pavlovales Family Pavlovaceae Genus Pavlova Species
Related species
There are 12 species names in the database at present, 11 of which are taxonomically accepted. Species of interest for biofuel:
P. gyrans, P. lutheri, P. pinguis, P. salina.
BIOLOGY
All species are known to produce motile swarmers though in a number of species they do not always constitute the dominant phase (P. ennorea, P. granifera, P. noctivaga, P. virescens), which is instead the non‐motile palmelloid stage.
The motile cells are elongate (P. gyrans, P. granifera, P. pinguis, P. salina, P. virescens) though others may be typically broadly oval, rhombic, cordate or, indeed, almost circular (P. helicata, P. Iutheri). However, considerable variation can be observed in the shape of all species and some (e.g., P. gyrans) can display marked metaboly. The appendages are always inserted laterally or anterio‐laterally on one of the flattened faces of the cell which by convention is referred to as the ventral face. In all species the flagella arise from a depression of varying depth. The long anteriorly directed flagellum beats with a characteristic sinusoidal S‐

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 157 of 258
shaped wave action either in a flat plane or describing a helical path as in, for example, P. helicata. In length it varies from species to species ranging from 6‐10 μm (P. calceolata) to 11‐20 μm (P. helicata). It carries an investiture of long fine hairs and small electron‐dense bodies the latter arranged in a regular helix. Variations are found from species to species in the presence or absence of the bodies, their morphology and their arrangement. The bodies are reffered to as scales (knob‐scales) by analogy with the Golgi‐produced scales of other members of the Prymnesiophyceae. Compared with the long flagellum, the short or posterior flagellum is relatively uniform in length and external morphology. The range of length recorded is between 2.5 and 6.0 μm and distally there is a tapered tip. The short flagellum usually beats with a rather stiff, inflexible action and is often directed towards the cell posterior and held close to the cell‐body. P. salina and P. helicata differ from other species of Pavlova in that the short flagellum is reduced to a stump about 0.2 μm long. The haptonema is always short and difficult to see in the living cell. It does not coil neither does it contract. In some species there may be a long terminal "filopodium" but even without “filopodia” the haptonema is frequently attenuated distally. Normally the surface of the haptonema is naked but in the case of P. lutheri there is a superficial covering of small knob‐scales. All species are "naked" in that there is no cell wall although the non‐motile cells may be invested with layers of mucilage. Although some species have been recorded as possessing two chloroplasts, there is usually only one consisting of two lateral lobes extending round the cell both ventrally and dorsally approximately forming a V‐ or H‐shape. Some members of the Pavlovales are unusual amongst the Prymnesiophyceae in that they have stigmata. In P. gyrans, P. granifera and P. pinguis the conspicuous orange‐red stigma consists of a concave layer of lipid globules, 6‐50 in number and situated adjacent to the chloroplast membrane close to the flagellar insertion. Within the Pavlovales, conspicuous pyrenoids are found only in P. gyrans, P. granifera, P. pinguis. The cells of all species usually contain two discrete colourless bodies which are presumed to constitute the reserve metabolite, a β 1‐3 glucan.
The available records show that the Pavlovales is, perhaps, a very successful goup having representatives in oceanic, coastal, brackish and fresh water habitats. Some species are markedly euryhaline which is clearly of advantage in brackish habitats. Individual species may also be widely distributed geographically.
BIOTECHNOLOGY
Pavlova is an important alga in aquaculture, where it is used to feed bivalve larvae and post‐larvae, penaeid shrimp larvae, freshwater prawn larvae and marine zooplankton (Tredici et al., 2009). It contains high amounts of both eicosapentaenoic (EPA, 20:5ω3) and docosahexaenoic (DHA, 22:6ω3) acids.
It is usually cultivated in low efficient photobioreactors in the hatcheries (Tredici et al., 2009). A comparison study carried out by Ponis et al. (2006) using standard hatchery techniques (a 10‐L carboy) and a 4‐L Flat Alveolar Panel, both under continuous artificial illumination. The volumetric productivity of P. lutheri in the FAP was on average 0.25 g dry weight L‐1 day‐1 during the batch growth phase and increased to 0.53 g L‐1 day‐1 when a semicontinuous regimen was adopted, while in the carboys the average volumetric productivity was 0.10 g L‐1 day‐1.
No pilot scale studies concenring biodiesel production has been up to now carried out. At laboratory scale, both P. salina and P. lutheri showed a quite high lipid content (31 and 35%, respectively) (Rodolfi et al., 2009), similar to that obtained in the Flat Alveolar Panel (Ponis et al., 2006).
References
Green J.C. (1980) The fine structure of Pavlova pinguis Green and a preliminary survey of the order Pavlovales (Prymnesiophyceae). European Journal of Phycology 15: 151‐191. [BIOLOGY section]
Ponis E., Parisi G., Le Coz J.R., Robert R., Chini Zittelli G., Tredici M.R. (2006) Effect of the culture system and culture technique on biochemical characteristics of Pavlova lutheri and its nutritional value for Crassostrea gigas larvae. Aquaculture Nutrition 12: 322‐329.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 158 of 258
Rodolfi L., Chini Zittelli G., Bassi N., Padovani G., Biondi N., Bonini G., Tredici M.R. (2009) Microalgae for oil: strain selection, induction of lipid synthesis, and outdoor mass cultivation in a low‐cost photobioreactor. Biotechnology and Bioengineering 102: 100‐112.
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G. (eds.) New technologies in aquaculture: Improving production efficiency, quality and environmental management, Woodhead Publishing, Cambridge, pp. 610‐676.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 159 of 258
8.6 Dinophyta
8.6.1 Crypthecodinium cohnii
Figure 60 ‐ Light microscopic view showing Crypthecodinium cohnii.
Picture from http://researchpages.net/media/resources/2009/07/26/Crypthe2.jpg
SYMBOLS: D
TAXONOMY
Phylum Dinophyta Class Dinophyceae Order Gonyaulacales Family Crypthecodiniaceae Genus Crypthecodinium Species Crypthecodinium cohnii
Related species
There is only species Cryptechodinium cohnii, synonym Cryptechodinium setense.
BIOLOGY
Crypthecodinium cohnii was first described as Glenodinium cohnii, then transferred to the genus Gyrodinium, and finally Crypthecodinium cohnii was introduced, that apparently represents a supraspecies encompassing several biological species.
Two forms of C. cohnii have been reported, swimming cells and cysts with different dimensions. The swimming cells show velocities of the order of 1 km.year−1. The motile swimming cells have two unequal flagella. The cingulum sometimes almost completely girdles the cell and sometimes is much displaced and does not form a complete loop around the body. C. cohnii has a very delicate theca (15–20 μm) whose plates are mainly composed of cellulose and are barely visible, unless using a special staining method. During cell locomotion, the theca deforms. The theca is contained in relatively few alveoli with a pattern that can be determined (thecal plate tabulation) and used for taxonomic purposes. The cysts are solitary

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 160 of 258
and ovoid in shape and can stay in a dormant/survival stage or start dividing (vegetative cysts). The non‐motile cysts contain greater levels of lipids, including docosahexaenoic acid (DHA, 22:6ω3), than the swimmer form of the microalga. C. cohnii has both asexual and sexual reproduction. It has been reported that the latter is induced by nutrient depletion and has been observed in dense, rapidly growing cultures. The gametes form groups of 3–8 cells which move around each other until two cells, anisogametes or isogametes, establish contact with their ventral sides and begin to fuse. Then one of the two transverse flagella is shed, and the developing zygote, with two longitudinal flagella, swims away. Some time later the zygote encysts. The cysts divide and release normally 2, but sometimes 4–8, dinoflagellates. The new dinoflagellates are motile as soon as they leave the cyst. C. cohnii is an obligatory heterotrophic. Carbon and other nutrients are obtained from living or decomposing seaweeds. It might have parasitic capabilities in or on macrophytes, as motile cells appear to have a peduncle, an organelle employed by other dinoflagellates on phagotrophy. Some strains are phagotrophic: the cell content of the prey is ingested by a feeding tube, leaving the prey’s cell coat in the medium. It grows well on organic carbon substrates such as glucose, dextrose, ethanol, acetic acid, sodium acetate, carob pulp syrup.
This microalga is brackish, littoral and neritic. It is often present among macrophytes, particularly Fucus spp. Crypthecodinium cohnii‐like dinoflagellates have been obtained from various habitats including open ocean beaches, polluted brackish bays and estuaries, steaming mangrove swamps and frozen harbors. Nevertheless, the strains of essentially globally dispersed dinoflagellates resembling C. cohnii are not members of the same gene pool.
BIOTECHNOLOGY
C. cohnii can accumulate relatively high amounts of lipids (up to 20% by weight) with DHA contents up to 30–50% of the total fatty acids and no other polyunsaturated fatty acids present above 1% (Couto et al., 2010). For DHA production, the limiting nutrient should be nitrogen since cell growth and division is halted due to the lack of this nutrient for de novo protein and nucleotide synthesis, and the supplied carbon is converted into storage lipids (triacylglycerols) rich in DHA. Therefore, industrial C. cohnii fermentations are usually a carbon‐fed batch and progresses in two stages: the first is the active growth phase during which the lipid content of the biomass is low; once the nitrogen source is depleted, carbon is continuously supplied to the fermenter (Couto et al., 2010). In most of the commercial cultivation processes, glucose is used as the carbon source and energy source, as it represents an easily accessible feedstock for many industrial fermentation processes (Couto et al., 2010).The maximum overall volumetric productivity of DHA reported on glucose is 19 mg L‐1 h‐1, while in a fed‐batch cultivation with pure ethanol as feed, 83 g dry biomass L‐1, 35 g total lipid L‐1 and 11.7 g DHA L‐1 were produced in 220 h, with an overall volumetric productivity of DHA was 53 mg L‐1 h‐1; with acetic acid as carbon source, DHA productivities of up to 45 mg L‐1 h‐1 were achieved (de Swaaf et al., 2003). DHA from C. cohnii is added to many commercial products, including infant formulas (Tredici et al., 2009). About 240 tonnes of DHA oil per year are produced in th USA at a price og 43 € g‐1 (Brennan and Owende, 2010).
C. cohnii has been considered also as a biodiesel feedstock (Griffiths and Harrison, 2009). The fatty acid composition of the remaining lipid fraction from the biomass leftover with lower content in polyunsaturated fatty acids could be adequate for further uses as feedstock for biodiesel, contributing to the economy of the overall process suggesting an integrated biorefinery approach (Couto et al., 2010).
References
Brennan L., Owende P. (2010) Biofuels from microalgae—A review of technologies for production, processing, and extractions of biofuels and co‐products. Renewable and Sustainable Energy Reviews 14: 557‐577.
Couto R.M., Calado Simões P., Reis A., Lopes Da Silva T., Martins V.H., Sánchez‐Vicente Y. (2010) Supercritical fluid extraction of lipids from the heterotrophic microalga Crypthecodinium cohnii. Engineering in Life Sciences 10: 158‐164.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 161 of 258
de Swaaf M.E., Pronk J.T., Sijtsma L. (2003) Fed‐batch cultivation of the docosahexaenoic‐acid‐producing marine alga Crypthecodinium cohnii on ethanol. Applied Microbiology and Biotechnology 61: 40‐43.
Griffiths M.J., Harrison S.T.L. (2009) Lipid productivity as a key characteristic for choosing algal species for biodiesel production. Journal of Applied Phycology 21: 493‐507.
Mendes A., Reis A., Vasconcelos R., Guerra P., Lopes da Silva T. (2009) Crypthecodinium cohnii with emphasis on DHA production: a review. Journal of Applied Phycology 21: 199‐214. [BIOLOGY section]
Tredici M.R., Biondi N., Chini Zittelli G., Ponis E., Rodolfi L. (2009) Advances in microalgal culture for aquaculture feed and other uses. In: Burnell G., Allan G. (eds.) New technologies in aquaculture: Improving production efficiency, quality and environmental management, Woodhead Publishing, Cambridge, pp. 610‐676.
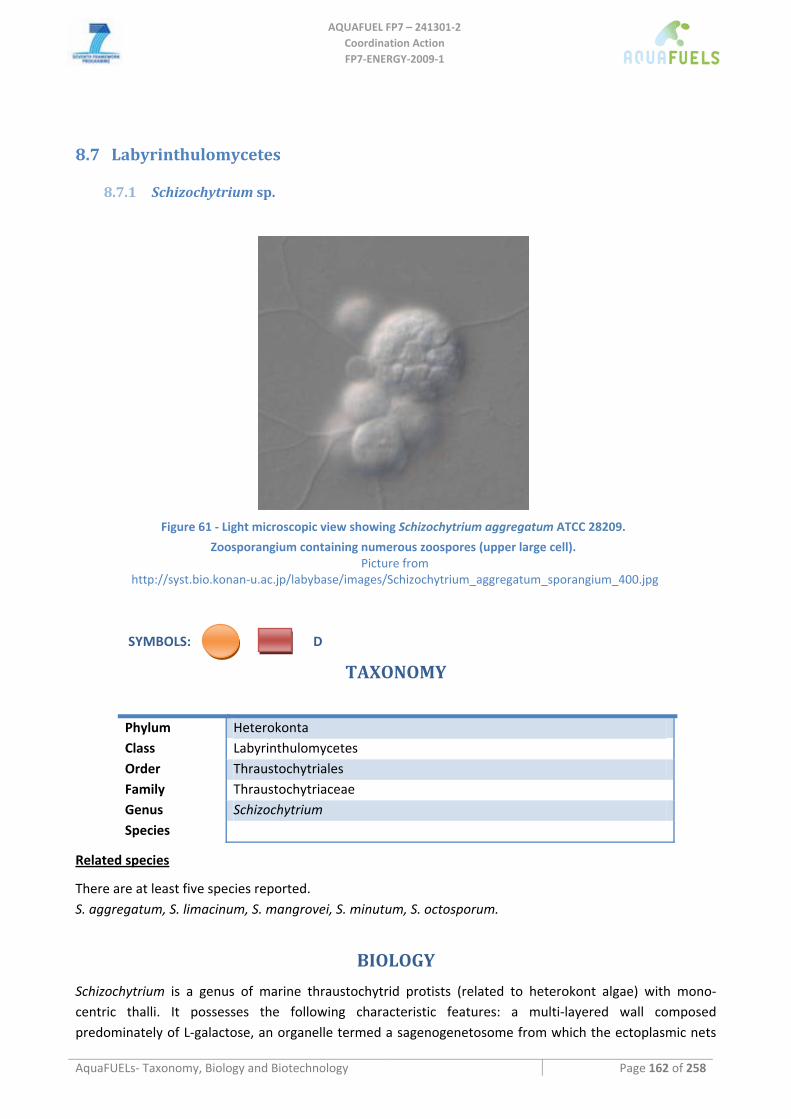
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 162 of 258
8.7 Labyrinthulomycetes
8.7.1 Schizochytrium sp.
Figure 61 ‐ Light microscopic view showing Schizochytrium aggregatum ATCC 28209.
Zoosporangium containing numerous zoospores (upper large cell). Picture from
http://syst.bio.konan‐u.ac.jp/labybase/images/Schizochytrium_aggregatum_sporangium_400.jpg
SYMBOLS: D
TAXONOMY
Phylum Heterokonta Class Labyrinthulomycetes Order Thraustochytriales Family Thraustochytriaceae Genus Schizochytrium Species
Related species
There are at least five species reported. S. aggregatum, S. limacinum, S. mangrovei, S. minutum, S. octosporum.
BIOLOGY
Schizochytrium is a genus of marine thraustochytrid protists (related to heterokont algae) with mono‐centric thalli. It possesses the following characteristic features: a multi‐layered wall composed predominately of L‐galactose, an organelle termed a sagenogenetosome from which the ectoplasmic nets

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 163 of 258
arise and biflagellate heterokont zoospores in many of described genera. Schizochytrium is characerised by successive bipartition of a vegetative cell, resulting in a cluster of cells, each of which develops into a zoosporangium or zoospore.It is an osmoheterotroph. They are ubiquitously distributed and can be found on the algal surface, in estuarine habitats, seawater and saline soil.
BIOTECHNOLOGY
DHA Algal Oil is produced via heterotrophic fermentation using Schizochytrium. DHA Algal Oil is intended to be used as a direct food ingredient to increase dietary intake of the long chain omega‐3 fatty acid DHA. Schizochytrium sp. has been utilized in aquaculture applications, including enrichment of DHA in Artemia and rotifers used to feed larval fish and shrimp. A product for aquaculture applications has been successfully utilized for over 7 years as an excellent, stable dietary source of DHA in shrimp larvaculture and finfish (red seabream, Japanese flounder) culture with no adverse effects (Hammond et al., 2001). Dried Schizochytrium sp. microalgae has also been determined to be generally recognized as safe (GRAS) for use as a DHA‐rich ingredient in broiler chicken and laying hen feed at levels up to 2.8 and 4.3%, respectively and, since 1997, DHA‐enriched eggs from hens fed a diet containing approximately 1% DRM are now commercially marketed in the United States, Mexico, Germany, Spain, Portugal, Benalux countries, Italy, Norway, and Israel (Hammond et al., 2001).
Under glucose and nitrogen fed‐batch culture conditions and with control of glucose, pH and oxygen levels, some Schizochytrium strains have been shown to grow to biomass densities of 200 g L‐1 in short fermentation cycles of 90–100 h, accumulating 40–45 g L‐1 DHA (Chi et al., 2009).
The oleaginous S. limacinum has capability of producing significant amounts of total lipid and DHA when growing in a variety of carbon sources such as glucose, fructose, as well as complex carbon sources such as sweet sorghum juice (Ethier et al., 2011). The total lipid contained in the biomass can be used as a source of biodiesel production. S. limacinum has been identified as capable of utilizing glycerol as a carbon source, which makes it possible to convert waste glycerol from the biodiesel industry to DHA (Chi et al., 2009). The highest biomass productivity of 3.88 g L‐1 day‐1 was obtained by feeding with 60 g L‐1 of glycerol, while the highest DHA productivity of 0.52 g L‐1 day‐1 was obtained at 90 g L‐1 of glycerol due to the higher DHA content (Ethier et al., 2011). In addition to DHA, S. limacinum also contains a high level of total fatty acid (ca 50% of dry biomass) and is a suitable feedstock for producing biodiesel via the direct transesterification method (Johnson and Wen, 2009). The biodiesel prepared was subjected to ASTM standard tests: parameters such as free glycerol, total glycerol, acid number, soap content, corrosiveness to copper, flash point, viscosity, and particulate matter met the ASTM standards, while the water and sediment content, as well as the sulphur content did not pass the standard and require further investigations (Johnson and Wen, 2009).
References
Chi Z., Liu Y., Frear C., Chen S. (2009) Study of a two‐stage growth of DHA‐producing marine algae Schizochytrium limacinum SR21 with shifting dissolved oxygen level. Applied Microbiology and Biotechnology 81: 1141‐1148.
Ethier S., Woisard K., Vaughan D., Wen Z. (2011) Continuous culture of the microalgae Schizochytrium limacinum on biodiesel‐derived crude glycerol for producing docosahexaenoic acid. Bioresource Technology 102: 88‐93.
Hammond B.G., Mayhew D.A., Holson J.F., Nemec M.D., Mast R.W., Sander W.J. (2001) Safety Assessment of DHA‐Rich Microalgae from Schizochytrium sp. II. Developmental Toxicity Evaluation in Rats and Rabbits. Regulatory Toxicology and Pharmacology 33: 205‐217.
Johnson M.B., Wen Z. (2009) Production of biodiesel fuel from the microalga Schizochytrium limacinum by direct transesterification of algal biomass. Energy and Fuels 23: 5179‐5183.
Kamlangdee N., Fan K.W. (2003) Polyunsaturated fatty acids production by Schizochytrium sp. isolated from mangrove. Songklanakarin Journal of Science and Technology 25: 643‐650. [BIOLOGY section]
Raghukumar S. (2002) Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids). European Journal of Protistology 38: 127‐145. [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 164 of 258
8.7.2 Thraustochytrium sp.
Figure 62 ‐ SEM image showing Tharustochytrium.
Picture from tolweb.org/tree/ToLimages/thrastosem2.png
SYMBOLS: D
TAXONOMY
Phylum Heterokonta Class Labyrinthulomycetes Order Thraustochytriales Family Thraustochytriaceae Genus Thraustochytrium Species
Related species
There are at least 17 species within this genus.
T. aggregatum, T. antarcticum, T. arudimentale, T. aureum, T. benthicola, T. caudivorum, T. gaertneri, T. globosum, T. indicum, T. kerguelense, T. kinnei, T. motivum, T. pachydermum, T. proliferum, T. roseum, T. rossii, T. striatum.
BIOLOGY
Thraustochytrium is a genus of marine thraustochytrid protists (heterotrophic organisms related to heterokont algae). Vegetative stages of thraustochytrids consist of single cells which are globose to subglobose, measuring 4 to 20 μm in diameter, mostly growing epibiontically on various substrata. Most thraustochytrids reproduce by means of zoospores which possess a long anterior flagellum and a short posterior flagellum. In Thraustochytrium the cytoplasmic contents of the mature cell, the sporangium, divide directly into zoospores. It lives mainly on detritus.
BIOTECHNOLOGY

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 165 of 258
Biotechnology of Thraustochytrium is similar to that of Schizochytrium, although no strain of this genus have been at present commercially produced. DHA production rate is similar to that of many Schizochytrium strains (Raghukumar, 2008).
All oils produced by Thraustochytrium sp. are >99% in the triacylglycerol form and <0.05% of free fatty acids (Scott et al., 2011). Using raw glycerol as carbon source produced comparable results for biomass, oil and DHA to those obtained when glucose was used (Scott et al., 2011).
References
Raghukumar S. (2002) Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids). European Journal of Protistology 38: 127‐145. [BIOLOGY section]
Raghukumar S. (2008) Thraustochytrid marine protists: Production of PUFAs and other emerging technologies. Marine Biotechnology 10: 631‐640.
Scott S.D., Armenta R.E., Berryman K.T., Norman. A.W. (2011) Use of raw glycerol to produce oil rich in polyunsaturated fatty acids by a thraustochytrid. Enzyme and Microbial Technology 48: 267‐272.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 166 of 258
8.7.3 Ulkenia sp.
SYMBOLS: D
TAXONOMY
Phylum Heterokonta Class Labyrinthulomycetes Order Thraustochytriales Family Thraustochytriaceae Genus Ulkenia Species
Related species
There are at least 6 species within this genus. U. amoeboidea, U. minuta, U. profunda, U. radiata, U. sarkariana, U. visurgensis.
BIOLOGY
Ulkenia is a genus of marine thraustochytrid protists (heterotrophic organisms related to heterokont algae). In Ulkenia the sporangium is formed from the mature thallus by the escape of the protoplast from the cell wall s one amoeboid mass, and thesucessive cleavage of the naked protoplast into zoospores. Ulkenia amoeboidea produces spores that assume the shape of an amoeba and move with a slow sinusoidal movement, then these cells turn into cells which produce zoospores. It is found on decaying biomass of macroalagae and in marine animal guts.
BIOTECHNOLOGY
DHA‐rich oil from Ulkenia sp., containing typically 45% DHA, is obtained through hetrotrophic fermentation by a German Company and is used as a food ingredient in such foods as breads, cakes and biscuits, breakfast cereals, cream cheese, modified milk and milk products, beverages, functional drinks (Food Standards Australia and New Zealand, 2005).
Ulkenia, as well as other thraustochytrids, has been proposed as a biodiesel feedstock due to its high lipid content (Fisher et al., 2008).
References
Raghukumar S. (1996) Morphology, taxonomy and ecology of Thraustochytrids and Labyrinthulids, the marine counterparts of zoosporic fungi. In: Dayal R. (ed.) Advances in zoosporic fungi, MD Publications PVT Ltd, New Delhi, pp. 35‐58. [BIOLOGY section]
Raghukumar S. (2002) Ecology of the marine protists, the Labyrinthulomycetes (Thraustochytrids and Labyrinthulids). European Journal of Protistology 38: 127‐145. [BIOLOGY section]
Fisher L., Nicholls D., Sanderson K. (2008) Production of biodiesel. WO2008/067605 A1. Food Standards Australia and New Zealand (2005) Final assessment report. Application A522. DHA‐rich micro‐algal oil from Ulkenia
sp. as a novel food.
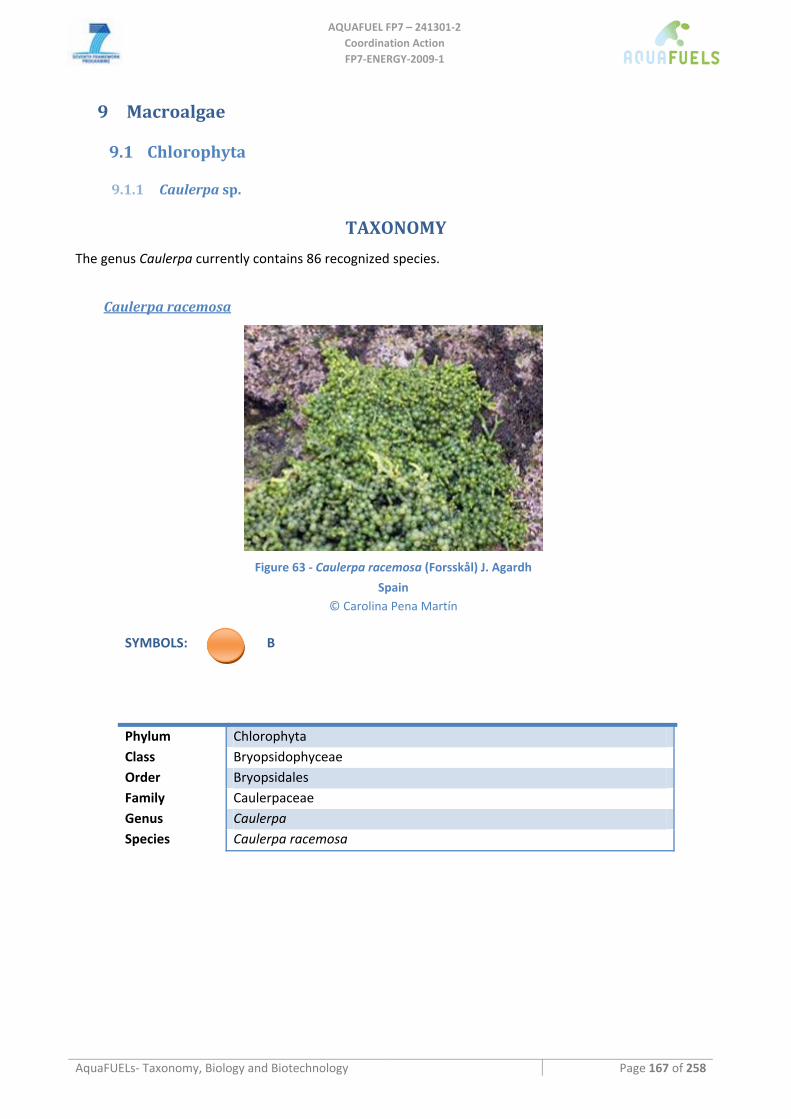
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 167 of 258
9 Macroalgae
9.1 Chlorophyta
9.1.1 Caulerpa sp.
TAXONOMY
The genus Caulerpa currently contains 86 recognized species.
Caulerpa racemosa
Figure 63 ‐ Caulerpa racemosa (Forsskål) J. Agardh
Spain © Carolina Pena Martín
SYMBOLS: B
Phylum Chlorophyta Class Bryopsidophyceae Order Bryopsidales Family Caulerpaceae Genus Caulerpa Species Caulerpa racemosa

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 168 of 258
Caulerpa taxifolia
Figure 64 ‐ Caulerpa taxifolia (M. Vahl) C. Agardh
Cala D'Or, Mallorca, Balearic Islands, Spain; taken at a depth of 7m on a dead rhizome of Posidonia oceanica. © Eduardo Infantes Oanes
SYMBOLS: B
Phylum Chlorophyta Class Bryopsidophyceae Order Bryopsidales Family Caulerpaceae Genus Caulerpa Species Caulerpa taxifolia
BIOLOGY Thallus composed of horizontal stolon anchored by colorless rhizoids, bearing erect photosynthetic fronds (assimilators) of extremely diverse morphology including thread‐like, blade‐like, pinnate, spongy and vesicular structures. Radial branching regarded as primitive, bilateral as more recent, interpretations which are supported by ultrastructure of chloroplasts. Reduction of light also results in change of erect portions from radial to bilateral symmetry. In species such as C. racemosa morphology has been shown to vary with habitat, resulting in description of numerous varieties or forms (or even species) which probably should be regarded as ecads. Growth apical and indeterminate. Extent of development of stolons and rhizoids depends on substratum; stolons can reach to 12 m.m‐2, but generally are more ramified in epilithic specimens; entire alga can reach to 1m. in length. Thallus composed of coenocytic filaments or siphons; walls principally of ‐1,3 xylan, with numerous trabeculae (branching cylindrical ingrowths of the wall) traversing the lumen. When wounded, as by grazing fish, blade or rhizome exude a yellowish sticky mass which hardens to a wound plug of carbohydrate in a few minutes. Genus heteroplastic. Large chloroplasts with pyrenoid and starch grains seem to be more primitive, being replaced by small chloroplasts lacking them. Amyloplasts mixed with chloroplasts. Isolated chloroplasts survive and divide in vitro for more than 2 weeks. Reproduction probably primarily by fragmentation of stolon. Holocarpic anisogamous sexual reproduction described for some species. Meiosis, where examined, occurs at gametogenesis with formation of gametes in unmodified areas of the thallus, without separation by cross walls; gametes liberated in gelatinous extrusions through superficial papillae. Some species of Caulerpa produce the poison caulerpicin, making their use as a food resource hazardous.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 169 of 258
Geographic distribution is global tropical to subtropical, with some species extending into the Mediterranean Sea and temperate regions of Australia. Highest species diversity occurs in S. Australia. Vertical range includes the intertidal to at least ‐50m; substrata include unconsolidated sand of seagrass meadows and hard surfaces such as coral rock.
BIOTECHNOLOGY Caulerpa racemosa is used in Bangladesh, Japan, South Pacfic Islands, Vietnam as food and in the Philippines as medicine (Zemke‐White and Ohno, 1999). In this latter country, C. taxifolia is also used, both as food and medicine. Here the annual production of Caulerpa is 810 t dry weight, all form cultured alga (Zemke‐White and Ohno, 1999).
References
Algaebase: http://www.algaebase.org/search/genus/detail/?genus_id=32944&‐session=abv4:96D919560808f1B19AmHOH2E91A9 Beach K.S., Borgeas H.B., Smith C.M. (2006) Ecophysiological implications of the measurement of transmittance and reflectance of
tropical macroalgae. Phycologia 45: 450‐457. Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere,
A.R.G. Gantner Verlag. Ruggell, pp. 1‐596, 266 pls. Falcão C., Menezes de Széchy M.T. (2005) Changes in shallow phytobenthic assemblages in southeastern Brazil, following the
replacement of Sargassum vulgare (Phaeophyta) by Caulerpa scalpelliformis (Chlorophyta). Botanica Marina 48: 208‐217. Guiry M.D., Guiry, G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 31 March 2010. Hodgson L.M., Pham Huu T., Lewmanomont K., McDermid K.J. (2004) Annotated checklist of species of Caulerpa and Caulerpella
(Bryopsidales, Caulerpaceae) from Vietnam, Thailand and the Hawaiian Islands. In: Abbott I.A., McDermid K.J. (eds.) Taxonomy of Economic Seaweeds with reference to the Pacific and other locations, Volume IX., pp. 21‐38.
Kraft G.T. (2007) Algae of Australia. Marine benthic algae of Lord Howe Island and the southern Great Barrier Reef, 1. Green algae, Australian Biological Resources Study & CSIRO Publishing,.Canberra & Melbourne, pp. i‐vi, 1‐347, 110 text‐figs; 11 pls.
Lam D.W., Zechman F.W. (2006) Phylogenetic analyses of the Bryopsidales (Ulvophyceae, Chlorophyta) based on RUBISCO large subunit gene sequences. Journal of Phycology 42: 669‐678.
Littler D.S., Littler M.M. (1997) An illustrated flora of the Pelican Cays, Belize. Bulletin of the Biological Society of Washington 9: 1‐149.
Nuber N., Gornik O., Lauc G., Bauer N., Zuljevic A., Papes D., Zoldos V. (2007) Genetic evidence for the identity of Caulerpa racemosa (Forsskal) J. Agardh (Caulerpales, Chlorophyta) in the Adriatic Sea. European Journal of Phycology 42: 113‐120.
Pedroche F.F., Silva P.C., Aguilar‐Rosas L.E., Dreckmann K.M., Aguilar‐Rosas R. (2005) Catálogo de las algas marinas bentónicas del Pacífico de México. I. Chlorophycota. pp. i‐viii, 17‐146. Ensenada, México: Universidad Autónoma de Baja California.
Phillips J.A. (2009) Reproductive ecology of Caulerpa taxifolia (Caulerpaceae, Bryopsidales) in subtropical eastern Australia. European Journal of Phycology 44: 81‐88.
Robledo D., Freile‐Pelegrín Y. (2005) Seasonal variation in photosynthesis and biochemical composition of Caulerpa spp. (Bryopsidales, Chlorophyta) from the Gulf of Mexico. Phycologia 44: 312‐319.
Serio D., Alongi G., Catra M., Cormaci M., Furnari G. (2006) Changes in the benthic algal flora of Linosa Island (Straits of Sicily, Mediterranean Sea). Botanica Marina 49: 135‐144.
Skelton P.A., South G.R. (2004) New records and notes on marine benthic algae of American Samoa ‐ Chlorophyta & Phaeophyta. Cryptogamie Algologie 25: 291‐312.
Stam W.T., Olsen J.L., Zaleski S.F., Murray S.N., Brown K.R., Walters L.J. (2006) A forensic and phylogenetic survey of Caulerpa species (Caulerpales, Chlorophyta) from the Florida coast, local aquarium shops, and e‐commerce: establishing a proactive baseline for early detection. Journal of Phycology 42: 1113‐1124.
Terrados J., Marba N. (2006) Is the vegetative development of the invasive chlorophycean, Caulerpa taxifolia, favored in sediments with a high content of organic matter? Botanica Marina 49: 331‐338.
Varela‐Alvarez E., Andreakis N., Lago‐Leston A., Pearson G.A., Serrao E.A., Procaccini G., Duarte C.M., Marba N. (2006) Genomic DNA isolation from green and brown algae (Caulerpales and Fucales) for microsatellite library construction (Note). Journal of Phycology 42: 741‐745.
Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐376.
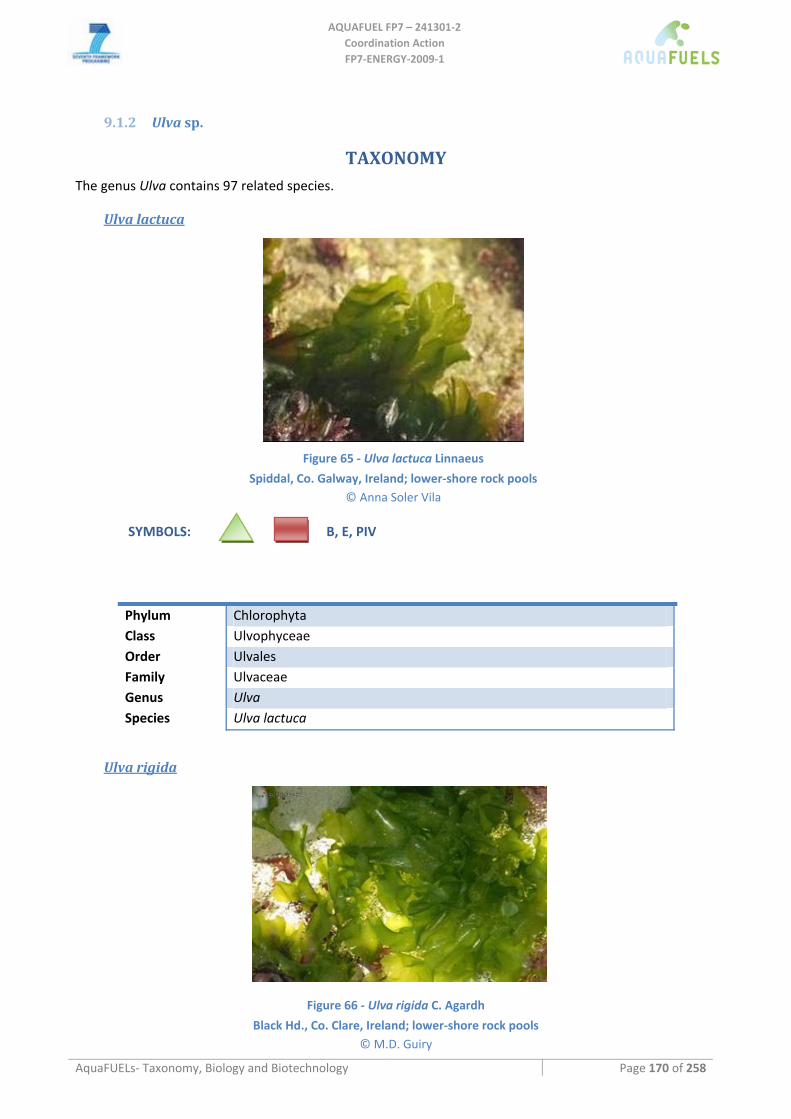
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 170 of 258
9.1.2 Ulva sp.
TAXONOMY The genus Ulva contains 97 related species.
Ulva lactuca
Figure 65 ‐ Ulva lactuca Linnaeus
Spiddal, Co. Galway, Ireland; lower‐shore rock pools © Anna Soler Vila
SYMBOLS: B, E, PIV
Phylum Chlorophyta Class Ulvophyceae Order Ulvales Family Ulvaceae Genus Ulva Species Ulva lactuca
Ulva rigida
Figure 66 ‐ Ulva rigida C. Agardh
Black Hd., Co. Clare, Ireland; lower‐shore rock pools © M.D. Guiry

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 171 of 258
SYMBOLS: B, E, PIV
Phylum Chlorophyta Class Ulvophyceae Order Ulvales Family Ulvaceae Genus Ulva Species Ulva rigida
BIOLOGY General description
This is a small genus of marine and brackish water green algae. It is edible and is often called 'Sea Lettuce'. Species with hollow, one‐layered thalli were formerly included in Enteromorpha, but it is widely accepted now that such species should be included in Ulva.
The thallus of ulvoid species is flat and blade‐like and is composed of two layers of cells. There is no differentiation into tissues; all the cells of the plant are more or less alike except for the basal cells, which are elongated to form attachment rhizoids. Each cell contains one nucleus and has a cup‐shaped choroplast with a single pyrenoid.
Ulva undergoes a very definite alternation of generations. Biflagellate isogametes are formed by certain cells of the haploid, gametangial plant. These are liberated and fuse in pairs to form a diploid zygote which germinates to form a separate diploid plant called the sporophyte; this resembles the haploid gametangial plant in outward appearance. Certain cells of the sporophyte undergo meiosis and form zoospores in sporangia; these zoospores are quite different to the gametes in that they form quadriflagellate zoospores (with 4 flagella). These are released, swim around for a time, settle and germinate to form the haploid gametangial thallus. Note that the haploid gametes are capable of settling and germinating without fusion to form a haploid thallus directly; most Ulva populations reproduce by this form of parthenogenesis and sexual reproduction is not very common. (Algaebase. 2010)
Biochemical composition The biochemical composition of wild macroalgae is subject to great spatial and seasonal variations. In Table 17 composition of Ulva sp. is reported. Table 17 ‐ Biochemical composition of Ulva sp. in Europe, except for pigment values. Composition expressed as % dry weight except for Water and ash. FW= Fresh weight. 1‐Ortiz et al. (2006). 2‐ Chakraborty and Santra (2008). 3‐
Briand and Morand (1997). Ulva sp. Water (%FW) 88.41 ‐ 65 to 833 Ash 111 – 17 to 353 Total Carbohydrate
61.51 ‐ 41 to 613
Uronic acids 7.7‐11.53 Glucose 3.9‐9.33 Mannose 0.7‐1.23 Rhamnose 16.6‐23.43 Xylose 2.9‐5.33

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 172 of 258
Ulva sp. Protein 27.21 ‐ 10.3 to 17.23 Lipid 0.31 – 1.8 to 3.53 Iodine 18‐2203 K Na 0.9‐5.93 Ca 1.4‐5.63 Mg 2.0‐2.73 S 2.8‐43 P 0.13‐0.243 Vitamin (mg.kg‐1) A
C: 3802 – 3.1 to 7893 B1: 0.42‐3.98
3 B12: 0.040‐0.064
3 E: 8.7‐37.53
Pigments (Chl a: 2.06 mg.g‐1 / Chl b: 1.35 mg.g‐1/ carotenoids: 19.55 μg.g‐1) 2
BIOTECHNOLOGY
Culture media / Cultivation methods / Growth kinetics and productivity Ulva sp. has been cultivated worldwide in an integrated polyculture, inline with either sewage treatment plants or aquaculture farms on land. Ulva sp. is also cultivated on Nori nets for food purposes in Japan. Table 18 represents the various cultivation systems and their productivity around the world.
Table 18. Ulva productivity in various cultivation systems around the world. Species Location/Seas
on (or culture duration)
Production Keywords Reference
Tank / Pond / Raceway Cultivation
Ulva lactuca USA (Florida) /
8 months 19 g DW m‐2 day‐1 (average over 8 months)
Outdoor cultivation DeBusk et al., 1986
Ulva lactuca Greece (Aegean sea)
9,4 g FW m‐2 day‐1 (8 days only) in shallow tanks
Associated with sewage treatment plant. Low salinity, not aimed at biomass production.
Tsagkamilis et al., 2010
Tank / Pond / Raceway Cultivation: Integrated Multitrophic aquaculture
Ulva lactuca Israel up to 376 g FW m‐2 day‐1 Biofiltration; fishpond effluents; yield; chemical analysis
Msuya and Neori, 2008
Sparus aurata
Ulva rotundata 33.6 g DW m‐2 day‐1 and GR 7.5% day‐1
Gracilariopsis longissima
Spain (Andalucia) / Spring
10.2 g DW m‐2 day‐1 and GR 6.5% day‐1
Biofiltration; stocking density; yield; biomass evolution; nitrogen; phosphorus
Hernandez et al., 2005
Sparus aurata Haliotis discus hannai
Israel / Winter‐Spring FCR= 2.66‐25 with Ulva
lactuca
Biofiltration; recirculation; ammonia toxicity
Schuenhoff et al., 2003
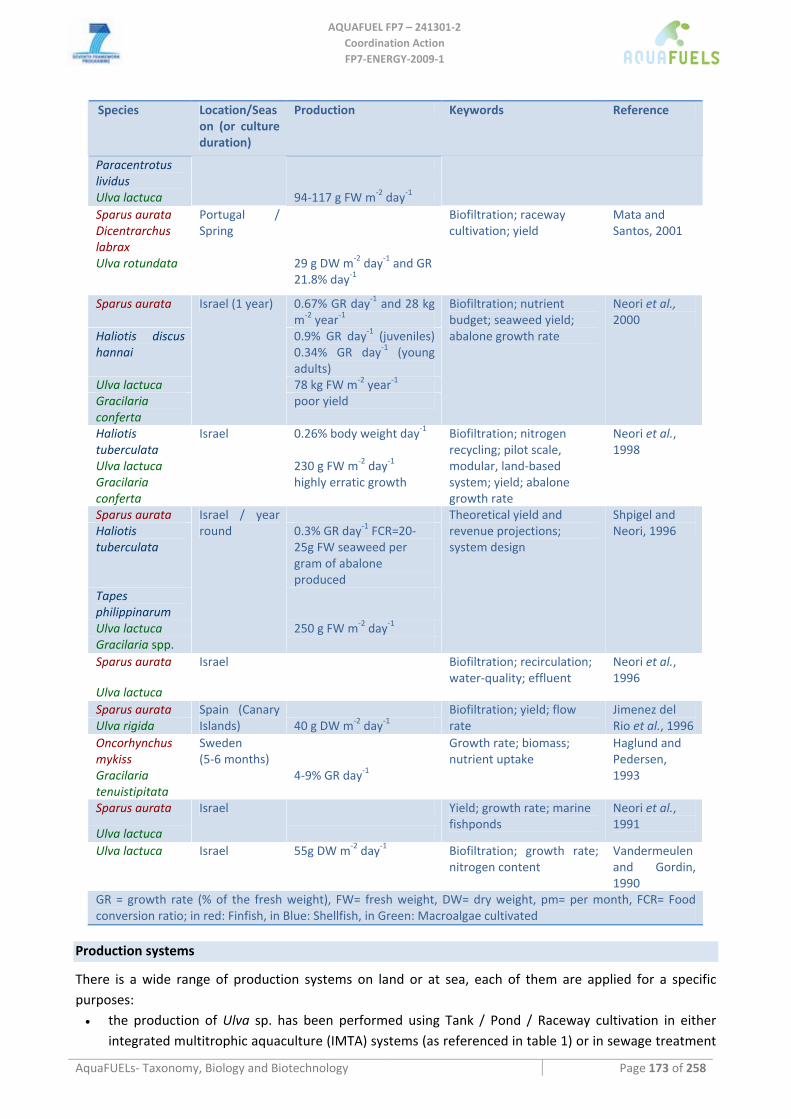
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 173 of 258
Species Location/Season (or culture duration)
Production Keywords Reference
Paracentrotus lividus
Ulva lactuca 94‐117 g FW m‐2 day‐1 Sparus aurata Dicentrarchus labrax
Ulva rotundata
Portugal / Spring
29 g DW m‐2 day‐1 and GR 21.8% day‐1
Biofiltration; raceway cultivation; yield
Mata and Santos, 2001
Sparus aurata 0.67% GR day‐1 and 28 kg m‐2 year‐1
Haliotis discus hannai
0.9% GR day‐1 (juveniles) 0.34% GR day‐1 (young adults)
Ulva lactuca 78 kg FW m‐2 year‐1 Gracilaria conferta
Israel (1 year)
poor yield
Biofiltration; nutrient budget; seaweed yield; abalone growth rate
Neori et al., 2000
Haliotis tuberculata
0.26% body weight day‐1
Ulva lactuca 230 g FW m‐2 day‐1 Gracilaria conferta
Israel
highly erratic growth
Biofiltration; nitrogen recycling; pilot scale, modular, land‐based system; yield; abalone growth rate
Neori et al., 1998
Sparus aurata Haliotis tuberculata
0.3% GR day‐1 FCR=20‐25g FW seaweed per gram of abalone produced
Tapes philippinarum
Ulva lactuca 250 g FW m‐2 day‐1 Gracilaria spp.
Israel / year round
Theoretical yield and revenue projections; system design
Shpigel and Neori, 1996
Sparus aurata
Ulva lactuca
Israel
Biofiltration; recirculation; water‐quality; effluent
Neori et al., 1996
Sparus aurata Ulva rigida
Spain (Canary Islands) 40 g DW m‐2 day‐1
Biofiltration; yield; flow rate
Jimenez del Rio et al., 1996
Oncorhynchus mykiss
Gracilaria tenuistipitata
Sweden (5‐6 months)
4‐9% GR day‐1
Growth rate; biomass; nutrient uptake
Haglund and Pedersen, 1993
Sparus aurata
Ulva lactuca
Israel
Yield; growth rate; marine fishponds
Neori et al., 1991
Ulva lactuca Israel 55g DW m‐2 day‐1 Biofiltration; growth rate; nitrogen content
Vandermeulen and Gordin, 1990
GR = growth rate (% of the fresh weight), FW= fresh weight, DW= dry weight, pm= per month, FCR= Food conversion ratio; in red: Finfish, in Blue: Shellfish, in Green: Macroalgae cultivated
Production systems
There is a wide range of production systems on land or at sea, each of them are applied for a specific purposes: • the production of Ulva sp. has been performed using Tank / Pond / Raceway cultivation in either
integrated multitrophic aquaculture (IMTA) systems (as referenced in table 1) or in sewage treatment

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 174 of 258
plant as biofilters (Giaccone et al., 1976; Guist and Humm, 1976; Ryther et al., 1979; Tsagkamilis et al., 2010), hence reducing the impact of fish farming on risks of eutrophication. The cultivated material originates from the vegetative growth of wild populations.
• Ulva sp. is also cultivated on suspended nets (similar to Porphyra, Nori, cultivation) for food purposes in Japan. Nets are spread on wild population of Ulva sp. and used as collectors. Besides, research programs have been developed to ensure reliable and consistent crop (Critchley in Werner et al., 2004). Ulva intestinalis (previously known as Enteromorpha intestinalis) cultivation has also been established from protoplasts (Rusig and Cosson, 2001). The authors claim that the plants regenerated from protoplasts may also be used as seedstock to facilitate propagation for macroalgal culture.
• Nevertheless, most biomass is harvested from the wild after blooming events, therefore no production system is required.
Harvesting methods • Tank / Pond / Raceway Cultivation
Either with nets or manually. • Nets cultivation
Although Ulva and Porphyra exhibit rather similar morphological characteristics (flat and blade‐like shape) and are cultivated on similar nets, harvesting methods differs. Ulva sp. is harvested manually leaving about 2 cm of the blade on the net to allow regeneration and a monthly growth/harvesting cycle (Perez, 1997).
• Blooms Ulva sp. can also be mechanically harvested from the wild, mostly during blooms (“green tides”) where the material is collected on sandy beaches in large quantities (several thousands of tons). In that case bulldozer, raking machines, sifting machines, scraping machines and baler (Hay press) have been used according to the scale and thickness of the deposit on the beach (up to 1 m high) (Briand in Guiry and Blunden, 1991). The existence of a subtidal and floating blooming populations of Ulva sp. has also been collected from boat equipped with conveyor belt‐collector in france and from special reaping machines in the Odense fjord in Denmark. Morand and Merceron (1999) reported a recurrent collection of over 50 000 m3 per year for Brittany (France) alone, which represents about 2000 truck arrivals and departures with associated hydrological and ecological impact (sand removal, erosion, etc).
Biomass processing Once harvested, the biomass is processed according to the end product. Different levels of quality are required whether the material will be used for feed and food purposes, fertilizer or energy purposes. Paper production was also one of the answers to resolve Macroalgal blooms of Ulva sp.
• Energy
Macroalgae are stored in a tank where the acidogenic fermentation occurs. The liquefaction juices from the storage of Ulva sp. are collected by draining and pressing the biomass. The juice is then transferred to another bioreactor for methanogenic fermentation (anaerobic digestion) (Morand and Merceron, 2006). The cake resulting from the pressing process can be used as organic enriching or fertilizing agent in agriculture.
Matsui et al. (2006) describe their field test plant process as divided in four parts: Pre‐treatment, fermentation, biogas storage and generation. In pre‐treatment part, seaweeds are smashed and diluted with water to suppress the effect of salt and make appropriate slurry. Collected Ulva sp. contained foreign

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 175 of 258
bodies such as sand, which do not affect fermentation directly but decrease available volume of tanks. Thus they are washed by water and foreign bodies were removed before they were used for fermentation tests. In fermentation part, there are two processes (pre‐fermentation and methane fermentation) for a higher efficiency of fermentation. The seaweed slurry is treated by pre‐fermentation (acid production) and use of methane fermentation as substrate. Capacity of a pre‐fermentation tank is 5 kL. In the methane fermentation process biogas is produced. Capacity of a methane fermentation tank is 30 kL. The methane fermentation tank contains porous matrix inside for inmobilizing bacterial cells. Biogas is refined (de‐sulfur) and stored in a gasholder (30 kl). Residue of methane fermentation is dehydrated and used as fertilizer. Biogas is mixed with city gas (natural gas) and fed to the gas engine generator (a co‐generation system). A gas mixer of the engine was improved to mix with biogas and city gas. Electricity (10 kW) generated by the gas engine is used for electric equipments of the plant. Heat (23 kW) from the engine is used for heating‐up energy for fermentation tanks. An equipment of deodorization using microorganisms is set.
• Fertilizer
The harvested biomass can be directly spread on fields as soil additive, but results in a subsequent stream and aquifer pollution. Besides, the possibility of spreading is limited by the field availability sufficiently close to the sea.
Composting algae with another substrate allows increasing the area of utilization, but not sufficiently because of the transportation cost and the low value of compost (Morand and Merceron, 2004).
• Food and feed purposes
The biomass coming from blooms is dried and milled for use in premixed poultry feeds (Brault et al., 1983). However, animal feeding only uses small amounts of algae, interesting as food supplement (i.e. β‐carotene) (Morand and Merceron, 2004). Human food requires highly clean material, which is often incompatible with harvesting methods of blooms, therefore only Ulva sp. cultivated on nets in Japan are commercially used for food purposes after drying, milling and conditioning. Scaling up limitations
Ulva sp. are relatively robust species with great tolerance to low salinity and temperature changes. The main scaling up limitations concerns the cultivation in tanks, ponds and raceways where land availability is scarce.
Other than that Hanisak (1987) mentions that Ulva sp. exhibits high yields but were not sustainable for significant periods as the species would become reproductive and shed spores. As each cell in the thallus can become reproductive, it was not unusual for an entire culture to sporulate and be lost overnight, leaving only empty cells. A search was initiated for sterile strains, as Ulva in particular lends itself to digestion with high methane yields because of its favourable carbohydrate and protein content.
References
Bliding C. (1969 "1968") A critical survey of European taxa in Ulvales, Part II. Ulva, Ulvaria, Monostroma, Kornmannia. Botaniska Notiser 121: 535‐629, 47 figs.
Brault D., Briand X., Golven P. (1983) “Les marees vertes”: premier bilan concernant les essais de valorisation. In: Bases biologiques de l’aquaculture, Ifremer, Montpellier, pp. 33‐42.
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeer, A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Briand X, Morand P. (1997) Anaerobic digestion of Ulva sp. 1. relationship between Ulva composition and methanisation. Journal of Applied Phycology 9: 511‐524.
Brodie J., Maggs C.A., John D.M. (2007) Green seaweeds of Britain and Ireland, British Phycological Society, London, pp. i‐xii, 1‐242, 101 figs.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 176 of 258
Burrows E.M. (1991) Seaweeds of the British Isles. Volume 2. Chlorophyta, Natural History Museum Publications, London, pp. xi 238, 60 figs, 9 plates.
Chakraborty S., Santra S.C. (2008) Biochemical composition of eight benthic algae collected from Sunderban. Indian Journal of Marine Sciences 37: 329‐332.
Dawes C.J., Mathieson A.C. (2008) The seaweeds of Florida, University Press of Florida, Gainesville, Florida, pp. i‐ viii, 1‐ 592, pls I‐LI. DeBusk T.A., Blakeslee M., Ryther,J.H. (1986) Studies on outdoor cultivation of Ulva lactuca L.. Botanica Marina 29: 381‐386. Giaccone G., Princi M., Rizzi Longo L. (1976) Riposte morfologiche e fisiologiche di alghe marine in coltura all’inquinamento di
liquami urbani e industriali. Ingeneria ambientale 5: 572‐582. Guist G.G., Humm H.J. (1976) Effects of sewage effluent on growth of Ulva lactuca. Florida Scientist 39: 267‐71. Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 20 October 2010. Guiry M.D., Blunden G. (1991) Seaweed resources in Europe: uses and potential, John Wiley & Sons, Chichester; New York, pp. xi,
432. Hanisak D.M. (1987) Cultivation of Gracilaria and other macroalgae in Florida for energy production. In: Bird K.T., Benson P.H. (eds.)
Seaweed cultivation for renewable resources. Developments in aquaculture and fisheries science, 16. Elsevier, Amsterdam, pp. 191‐218.
Hayden H.S., Waaland J.R. (2004) A molecular systematic study of Ulva (Ulvaceae, Ulvales) from the northeast Pacific. Phycologia 43: 364‐382.
Hernandez I., Fernandez‐Engo M.A., Perez‐Llorens J.L., Vergara J.J. (2005) Integrated outdoor culture of two estuarine macroalgae as biofilters for dissolved nutrients from Sparus aurata waste waters. Journal of Applied Phycology 17: 557‐567.
Jimenez del Rio M., Ramazanov Z., Garcia‐Reina G. (1996) Ulva rigida (Ulvales, Chlorophyta) tank culture as biofilters for dissolved inorganic nitrogen from fishpond effluents. Hydrobiologia 326/327: 61‐66.
Kraft G.T. (2007) Algae of Australia. Marine benthic algae of Lord Howe Island and the southern Great Barrier Reef, 1. Green algae, Australian Biological Resources Study & CSIRO Publishing, Canberra & Melbourne, pp. i‐ivi, 1‐347, 110 text‐figs; 11 pls.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Loughnane C.J., McIvor L.M., Rindi F., Stengel D.B., Guiry M.D. (2008) Morphology, rbcL phylogeny and distribution of distromatic
Ulva (Ulvophyceae, Chlorophyta) in Ireland and southern Britain. Phycologia 47: 416‐429. Mata L., Santos R. (2001) Cultivation of Ulva rotundata (Ulvales, Chlorophyta) in raceways, using semi‐intensive fishpond effluents:
yield and biofiltration. Proceedings of the 17th International Seaweed Symposium. Matsui T., Amano T., Koike Y., Saiganji A., Saito H. (2006) Sustainable nonfuel products/production systems from biomass
resources. Paper presented at 2006 American Institute of Chemical Engineers (AIChE) Conference, San Francisco, California (www.aiche.org) Session 412 ‐ http://aiche.confex.com/aiche/2006/techprogram/S3516.HTM.
Morand P., Briand X., Charlier R.H. (2006) Anaerobic digestion of Ulva sp.3. Liquefaction juices extraction by pressing and technico‐economic budget. Journal of Applied Phycology 18: 741‐755.
Morand P., Merceron M. (2004) Coastal eutrophication and excessive growth of macroalgae. In: Pandalai S.G. (ed.) Recent Research Developments in Environmental Biology 1. Research Signpost, Trivandrum, India, pp. 395‐449.
Morand, P., Merceron, M. (2005) Macroalgal population and sustainability. Journal of Coastal Research 21: 1009‐1020. Msuya F.E., Neori A. (2008) Effect of water aeration and nutrient load level on biomass yield, N uptake and protein content of the
seaweed Ulva lactuca cultured in seawater tanks. Journal of Applied Phycology 20: 1021‐1031. Neori A., Ragg N.L.C., Shpigel M. (1998) The integrated culture of seaweed, abalone, fish and clams in modular intensive land‐based
systems: II. Performance and nitrogen partitioning within an abalone (Haliotis tuberculata) and macroalgae culture system. Aquacultural Engineering 17: 215‐239.
Neori A., Shpigel M., Ben‐Ezra D. (2000) A sustainable integrated system for culture of fish, seaweed and abalone. Aquaculture 186: 279‐291.
Norris J.N. (2010) Marine algae of the Northern Gulf of California: Chlorophyta and Phaeophyceae, Smithsonian Contributions to Botany 94: i‐x, 1‐276.
Ortiz J., Romero N., et al. (2006) Dietary fiber, amino acid, fatty acid and tocopherol contents of the edible seaweeds Ulva lactuca and Durvillaea Antarctica. Food Chemistry 99: 98‐104.
Pedroche F.F., Silva P.C., Aguilar‐Rosas L.E., Dreckmann K.M., Aguilar‐Rosas R. (2005) Catálogo de las algas marinas bentónicas del Pacífico de México. I. Chlorophycota, Universidad Autónoma de Baja California, Ensenada, México, pp. i‐viii, 17‐146.
Perez R (1997) Ces algues qui nous entourent: conception actuelle, rôle dans la biosphère, utilisations, culture, IFREMER Plouzané, France, pp. xi 272.
Ryther J.H., De Boer J.A., Lapointe B.E. (1979) Cultivation of seaweeds for hydrocolloids, waste treatment and biomass for energy conversion. Jensen A., Stein J.R. (eds.) Proceedings of the IXth International seaweed symposium, Science Press, Princeton, pp. 1‐16.
Rusig A.M., Cosson J. (2001) Plant regeneration from protoplasts of Enteromorpha intestinalis (Chlorophyta, Ulvophyceae) as seedstock for macroagal culture. Journal of Applied Phycology 13: 103‐108.
Scagel R.F. (1957) An annotated list of the marine algae of British Columbia and northern Washington (including keys to genera). Bulletin, National Museum of Canada 150: vi ‐ 289.
Shhuenhoff A., Shpigel M., Lupatsch I., Ashkenazi A., Msuya F.E., Neori A. (2003) A semi‐recirculating, integrated system for the culture of fish and seaweed. Aquaculture 221: 167‐181.
Shpigel M., Neori A.(1996) The integrated culture of seaweed, abalone, fish and clams in modular intensive land‐based systems: 1. Proportions of size and projected revenues. Aquacultural Engineering 15: 313‐326.
Smith G.M. (1944) Marine algae of the Monterey Peninsula, Stanford University Press, Stanford, pp. ix, 622, 98 pls.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 177 of 258
Tsagkamilis P., Danielidis D., Dring M., Katsaros C. (2010) Removal of phosphate by the green seaweed Ulva lactuca in a small‐scale sewage treatment plant (Ios Island, Aegean Sea, Greece). Journal of Applied Phycology 22: 331‐339.
Vandermeulen H., Gordin H. (1990) Ammonium uptake using Ulva (Chlorophyta) in intensive fishpond systems: mass culture and treatment of effluent. Journal of Applied Phycology 2: 363‐374.
Werner A., Clarke C., Kraan S. (2004) Strategic review of the feasibility of seaweed aquaculture in Ireland. NDP Marine RTDI Desk Study Series, DK/01/008.2004
Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 178 of 258
9.1.3 Cladophora sp.
Figure 67 ‐ Cladophora glomerata © Fabio Rindi (left) and Cladophora rupestris (Linnaeus) Kützing © Michael D.
Guiry (right). Ireland
SYMBOLS: B
TAXONOMY
Phylum Chlorophyta Class Ulvophyceae Order Cladophorales Family Cladophoraceae Genus Cladophora Species Cladophora glomerata / rupestris
Related species
The genus Cladophora contains 180 related species.
BIOLOGY Thalli of uniseriate branched filaments with apical and/or intercalary growth. Branches sparse to profuse with branches inserted laterally below apex of cell or apically on cell (a pseudodichotomy). If attached, branching rhizoids arise from basal cell and other cells in basal region, or simple discoid holdfast produced. Chloroplasts parietal, either densely packed discoid and/or united in a reticulum. Pyrenoids in many chloroplasts, bilenticular, flanked by two bowl‐shaped starch bodies. Cells multinucleate, nuclei dividing more or less synchronously with nuclear membrane remaining intact. Cell wall polysaccharide mainly crystalline cellulose, forming numerous lamellae of microfibrils in crossed fibrillar pattern. Asexual reproduction by biflagellate or quadriflagellate zoospores only method of reproduction in some species, some species only reproducing by thallus fragmentation. Akinetes produced by most species under unfavourable conditions; these are swollen, thick‐walled, filled with starch. Thick walled rhizoids and basal portions of main axes also perennating devices. Sexual reproduction by regular alternation of generations producing biflagellate isogametes and quadriflagellate zoospores. Meiosis precedes spore production. Zoidangia with 1‐3 pores, cruciate zooids released under pressure. Zoids of C. rupestris with well developed

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 179 of 258
layer of granules outside the plasmalemma and inside the 'fibrous‐layer' which may be involved in cell wall synthesis.
Cladophora is cosmopolitan in temperate and tropical regions, occurring in freshwater, brackish and marine conditions. The various species are found intertidally in wave‐exposed to very sheltered habitats, in brackish pools, lagoons and mudflats, and in more or less eutrophic freshwater streams and lakes with pH > 7. Certain unattached species can be aggregated into spherical mats, 2‐10 cm in diameter, called "Cladophora Balls". Cladophora glomerata can be a rapid colonizer of rocky stream beds and at temperatures > 15 °C readily produces zoospores and new vegetative growth following winter dormancy or severe floods. Cell walls of the freshwater species C. glomerata include silica, an essential nutrient for this species. Calcium is also an essential element and CaCO3 depositions occur in older cells. In sea‐water, Cladophora normally absorbs bicarbonate rather than CO2 for photosynthesis.
BIOTECHNOLOGY
In Thailand, Cladophora edible freshwater alga, is known as Kai. It is abundant in the Nan and Mekong rivers in the Northern part of Thailand. The local people around these rivers collect it for domestic consumption and it is sold in the markets and is now cultured by using wastewater from fish rearing (Khuantrairong and Traichaiyaporn, 2011).
Cladophora glomerata is generally considered as a good bioindicator of heavy metals in aquatic bodies (Whitton et al., 1989).
References
AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=37 Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 02 July 2010. John D.M. (2002) Order Cladophorales (=Siphonocladales). In: John D.M., Whitton B.A., Brook,A.J. (eds.) The Freshwater Algal Flora
of the British Isles. An identification guide to freshwater and terrestrial algae, Cambridge University Press, Cambridge, pp. 468‐470.
Khuantrairong T., Traichaiyaporn S. (2011) The nutritional value of edible freshwater alga Cladophora sp. (Chlorophyta) grown under different phosphorus concentrations. International Journal of Agriculture & Biology 13: 297‐300.
Thomas D.N., Collins J.C., Russell G. (1988) Interactive effcts of temperature and salinity upon net photosynthesis of Cladophora glomerata (L.) Kütz. C. rupestris (L.) Kütz. Botanica Marina 31: 73‐77.
Whitton B. A., Burrows I.G., Kelly M.G. (1989) Use of Cladophora glomerata to monitor heavy metals in rivers. Journal of Applied Phycology 1: 293‐299.
Wiencke C., Davenport J. (1987) Respiration and photosynthesis in the intertidal alga Cladophora rupestris (L.) Kütz. under fluctuating salinity regimes. Journal of Experimental Marine Biology and Ecology 114: 183‐197.
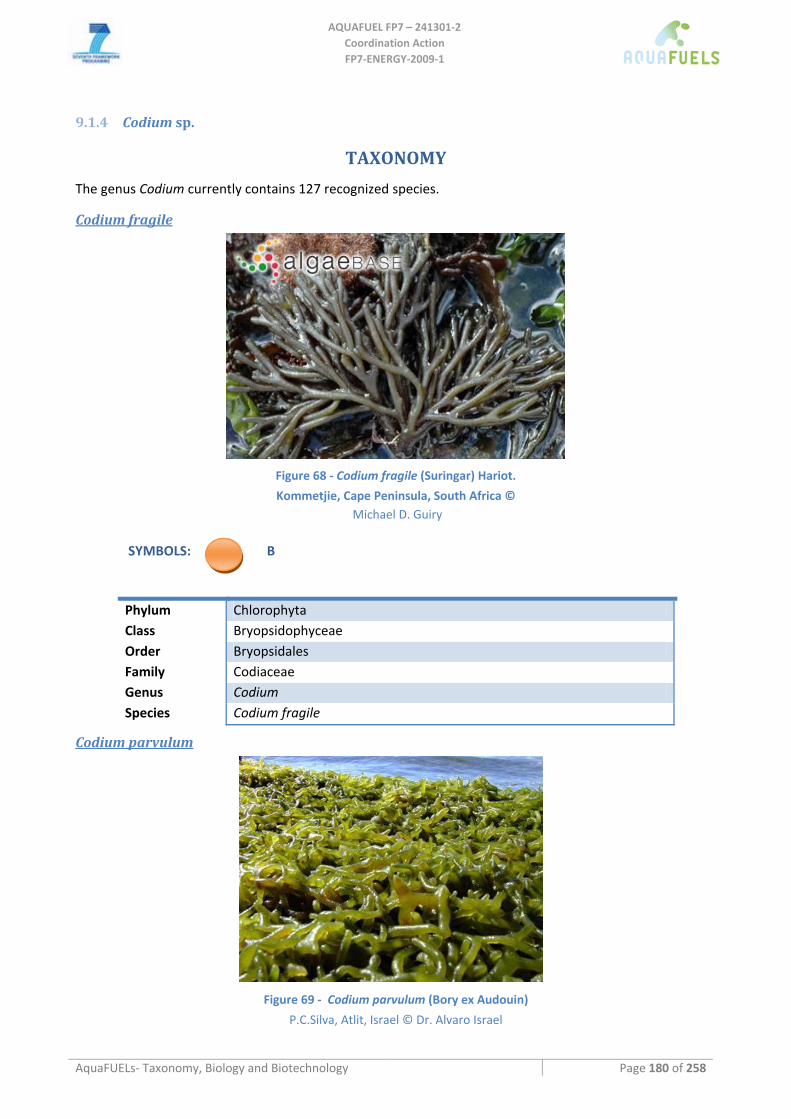
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 180 of 258
9.1.4 Codium sp.
TAXONOMY
The genus Codium currently contains 127 recognized species.
Codium fragile
Figure 68 ‐ Codium fragile (Suringar) Hariot.
Kommetjie, Cape Peninsula, South Africa © Michael D. Guiry
SYMBOLS: B
Phylum Chlorophyta Class Bryopsidophyceae Order Bryopsidales Family Codiaceae Genus Codium Species Codium fragile
Codium parvulum
Figure 69 ‐ Codium parvulum (Bory ex Audouin)
P.C.Silva, Atlit, Israel © Dr. Alvaro Israel

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 181 of 258
SYMBOLS: B
Phylum Chlorophyta Class Bryopsidophyceae Order Bryopsidales Family Codiaceae Genus Codium Species Codium parvulum
BIOLOGY
Thallus spongy, anchored to rocks or shells by a weft of rhizoids, varying in size from 1 cm to 10 m long. Habit varying widely applanate, pulvinate, digitaliform, globular, petaloid, membraniform, or dichotomously branched; erect or repent; branches wholly terete or variously flattened, at times anastomosing. Internal structure composed of a colorless medulla of densely intertwined siphons and a green palisade‐like layer of vesicles called utricles. Organelles, including innumerable nuclei and discoid chloroplasts (but no amyloplasts) are confined to a layer of cytoplasm appressed to a wall of which mannan is an important constituent. Chloroplasts lack pyrenoids; carotenoid pigments include siphaxanthin and siphonein. Incomplete septa (plugs) formed by centripetal deposition of wall material. Utricles arise primarily by enlargement of sympodial branches of medullary siphons, secondarily by budding or by production of additional utricle‐forming medullary siphons from basal portion of existing utricles. Mature utricles cylindrical or clavate, the apical wall usually thickened and often ornamented in a pattern characteristic of particular species. Rhizoidal siphons, which become buried in the medulla, also produced from basal portion of utricles. Colorless hairs, each with a basal plug, produced by utricles shortly below their apices, caducous at the plug, which remains as a prominent scar. Gametangia produced laterally by utricles, each with a basal plug above a short pedicel; fusiform to ovoid, the contents cleaving into biflagellate gametes following meiosis. Gametes extruded in a gelatinous mass through apical rupture. Male gametes contain only one or two chloroplasts, female gametes several times larger, with numerous chloroplasts, the two types produced on the same thallus (monoecious) or more often on different thalli (dioecious). Zygote develops into amorphous prostrate vesicle that produces erect elongate vesicles; these in turn initiate primary utricle‐producing siphons which eventually consolidate into a multiaxial thallus. Asexual reproduction by parthenogenesis, fragmentation, or the cutting off of modified aborted gametangia. Despite the ubiquity of Codium very little is know about its biology. The life history outlined above is generalized from fragmentary studies. Codium parvulum has been recently described as blooming in Nothern shores of Israel.
BIOTECHNOLOGY
Codium fragile is used as food in Korea and the Philippines, in the latter of which other species of Codium are also used. These are used also in other countries such as Argentina, Hawaii, Israel and Indonesia (Zemke‐White and Ohno, 1999). Codium in Korea is produced in an amount of 150 kg dry weight year‐1 (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=39&sk=0

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 182 of 258
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere, A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Israel A., Einav R., Silva P.C., Paz G., Chacana M.E., Douek J. (2010) First report of the seaweed Codium parvulum (Chlorophyta) in Mediterranean waters: recent blooms on the northern shores of Israel. Phycologia 49: 107‐112.
Kusakina J., Snyder M., Kristie D.N., Dadswell M.J. (2006) Morphological and molecular evidence for multiple invasions of Codium fragile in Atlantic Canada. Botanica Marina 49: 1‐9.
Lam D.W., Zechman F.W. (2006) Phylogenetic analyses of the Bryopsidales (Ulvophyceae, Chlorophyta) based on RUBISCO large subunit gene sequences. Journal of Phycology 42: 669‐678.
Mondragon J., Mondragon J. (2003) Seaweeds of the Pacific Coast. Common marine algae from Alaska to Baja California, Sea Challengers, Monterey, California, pp. iv, 5‐97.
Pedroche F.F., Silva P.C., Aguilar‐Rosas L.E., Dreckmann K.M., Aguilar‐Rosas R. (2005) Catálogo de las algas marinas bentónicas del Pacífico de México. I. Chlorophycota, Universidad Autónoma de Baja California, Ensenada, México, pp. i‐viii, 17‐146.
Provan J., Booth D., Todd N.P., Beatty G.E., Maggs C.A. (2008) Tracking biological invasions in space and time: elucidating the invasive history of the green alga Codium fragile using old DNA. Diversity and Distributions A Journal of Conservation Biogeography 14: 343‐354.
Scagel R.F. (1957) An annotated list of the marine algae of British Columbia and northern Washington (including keys to genera). Bulletin, National Museum of Canada 150: vi ‐ 289.
Schorie, D., Selig U., Schubert H. (2009) Species and synonym list of the German marine macroalgae based on historical and recent records. Rostock. Meeresbiologie Beitraeg 21: 7‐135.
Silva P.C. (1955) The dichotomous species of Codium in Britain. Journal of the Marine Biological Association of the United Kingdom 34: 565‐577.
Smith G.M. (1944) Marine algae of the Monterey Peninsula, Stanford University Press, Stanford, pp. ix, 622, 98 pls.. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 183 of 258
9.2 Rhodophyta
9.2.1 Chondrus crispus
Figure 70 ‐ Chondrus crispus (Stackhouse)
New Quay, Co. Clare, Ireland; plant to 60 mm long; on rock © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Rhodophyta Class Florideophyceae Order Gigartinales Family Gigartinaceae Genus Chondrus Species Chondrus crispus
Related species
C. armatus, C. canaliculatus, C. elatus, C. giganteus, C. giganteus f. flabellatus, C. nipponicus, C. ocellatus f. crispus, C. ocellatus f. parvus, C. ocellatus, C. ocellatus f. aequalis, C. ocellatus f. crispoides, C. pinnulatus f. conglobatus, C. pinnulatus, C. verrucosus, C. yendoi f. subdichotomus, C. yendoi, C. yendoi f. fimbriatus, C. crispus var. lonchophorus, C. crispus var. filiformis.
BIOLOGY Cartilaginous, dark purplish‐red, red, yellowish or greenish fronds to 150 mm high, gametophyte plants are often iridescent under water when in good condition. Stipe compressed, narrow, expanding gradually to a flat, repeatedly dichotomously branched frond, in tufts from a discoid holdfast. Axils rounded, apices blunt or subacute, frond thicker in centre than margins. Very variable in breadth of segments. Very variable in branching, colour and thickness. Cystocarps are embedded and aggregated distally or confined to proliferations. Cystocarps lack a surrounding filamentous hull, the major generic defining character of

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 184 of 258
Chondrus. Male plants are often said to be extremely rare, but are reported to occur in normally expected proportions in at least some populations of the most commerically valuable species, C. crispus. Tetrasporangia occur in branched chains formed laterally on cells of the inner cortex.
C. crispus is almost exclusively restricted to rock and ranges from intertidal to 24 m depths, being also tolerant of lowered salinities. It occurs in the north Atlantic from New Jersey to Spain and possibly Morocco and the Cape Verde Islands, as well as Japan.
BIOTECHNOLOGY
This genus is economically important, currently in eastern Canada and France, and formerly in Britain and Ireland. Much emphasis has been devoted to its cultivation in outdoor tanks as raw material for commercial utilization (Chen and Taylor, 1978; Simpson et al., 1978; Simpson and Shacklock, 1979; Shacklock and Croft, 1981; Bidwell et al., 1985). Bidwell et al. (1984, 1985) devised and tested outdoor tank methods generally applicable to Chondrus culture both within and outside of its native range, but conclude that world prices do not allow economically viable cultivation in eastern Canada where the largest natural populations are found. C. crispus is used for carrageenan production in France, Spain and US, and as food in Ireland and France, while in Japan C. ocellatus is used as food (Zemke‐White and Ohno, 1999). Canada produces 10,000 t dry weight per year of Chondrus, France 1,260, Japan 500, Spain 300, US 120, Portugal 30 and Ireland 3 (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=2 Bates C.R., Saunders G.W., Chopin T. (2005) An assessment of two taxonomic distinctness indices for detecting seaweed
assemblage responses to environmental stress. Botanica Marina 48: 231‐243. Bidwell R.G.S., Lloyd N.D.H., McLachlan J. (1984) The performance of Chondrus crispus (Irish moss) in laboratory simulations of
environments in different locations. Proceedings of the International Seaweed Symposium 6: 292‐294. Bidwell R.G.S., McLachlan J., Lloyd N.D.H. (1985) Tank cultivation of Irish moss, Chondrus crispus Stackh. Botanica Marina 28: 87‐97. Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere,
A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls. Chen L.C.M., Taylor A.R.A. (1978) Medullary tissue culture of the red alga Chondrus crispus. Canadian Journal of Botany 56: 883‐
886. Dixon P.S., Irvine L.M. (1977) Seaweeds of the British Isles. Volume 1. Rhodophyta. Part 1. Introduction, Nemaliales, Gigartinales, .
British Museum (Natural History), London, pp. xi ‐ 252, 90 figs. Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 31 March 2010. Hommersand M.H., Guiry M.D., Fredericq S., Leister G.L. (1993) New perspectives in the taxonomy of the Gigartinaceae
(Gigartinales, Rhodophyta). Proceedings of the International Seaweed Symposium 14: 105‐120, 41 figs. Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Mikami H. (1965) A systematic study of the Phyllophoraceae and Gigartinaceae from Japan and its vicinity. Memoirs of the Faculty
of Fisheries Hokkaido University 5: 181‐285. Shacklock P.F., Croft G.B. (1981) Effect of grazers on Chondrus crispus in culture. Aquaculture 22: 331‐342. Silva P.C., Basson P.W., Moe R.L. (1996) Catalogue of the benthic marine algae of the Indian Ocean. University of California
Publications in Botany 79: 1‐1259. Simpson F.J., Shacklock P.F. (1979) The cultivation of Chondrus crispus. Effect of temperature on growth and carrageenan
production. Botanica Marina 22: 295‐298. Simpson F.J., Neish A.C., Shacklock P.F., Robson D.R. (1978) The cultivation of Chondrus crispus. Effect of pH on growth and
production of carrageenan. Botanica Marina 21: 229‐235. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 185 of 258
9.2.2 Mastocarpus stellatus
Figure 71 ‐ Mastocarpus stellatus (Stackhouse)
Guiry Muigh Inis, Co. Galway, Ireland; on rock © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Rhodophyta Class Florideophyceae Order Gigartinales Family Phyllophoraceae Genus Mastocarpus Species Mastocarpus stellatus
Related species
M. jardinii, M. pacificus, M. papillatus, M. yendoi.
BIOLOGY
Cartilaginous, purplish brown fronds, often in dense tufts, arising from a discoid holdfast, to 200 mm high. Narrow compressed stipe expands into strap‐like blade, usually inrolled to form a channel, with thickened margins. Repeatedly dichotomously branched, axils acute. Upper part of frond with papillae to 10 mm or more long on surfaces and margins on female plants. Male plants lack papillae and are generally rare. Tetrasporophyte a purplish‐black crust (Petrocelis‐phase).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 186 of 258
M. stellatus occurs on rocks in lower intertidal, often in large continuous mats, widespread and abundant. Both the gametophyte and tetrasporophytic stages are characteristic of high to mid intertidal rocks on open coasts, although it also occurs subtidally.
M. stellatus occurs in the north Atlantic Ocean from Russia to Portugal and from Morocco to Mauritania and from Rhode Island to Newfoundland.
BIOTECHNOLOGY
M. stellatus is used as a source of carrageenan in Spain and Portugal and as food in Ireland. Annual production is of 600 t dry weight in Spain, 70 in Portugal and 5 in Ireland (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/species/detail/?species_id=24&sk=0&from=results http://www.algaebase.org/search/genus/detail/?genus_id=20
Bates C.R., Saunders G.W., Chopin T. (2005) An assessment of two taxonomic distinctness indices for detecting seaweed assemblage responses to environmental stress. Botanica Marina 48: 231‐243.
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere, A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Guiry M.D., West J.A. (1983) Life history and hybridization studies on Gigartina stellata and Petrocelis cruenta (Rhodophyta) in the North Atlantic. Journal of Phycology 19: 474‐494.
Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 31 March 2010.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Silva P.C. (1952) A review of nomenclatural conservation in the algae from the point of view of the type method. University of
California Publications in Botany 25: 241‐323. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376. Zuccarello G.C., Schidlo A., McIvor L., Guiry M.D. (2006) A molecular re‐examination of speciation in the intertidal red alga
Mastocarpus stellatus (Gigartinales, Rhodophyta) in Europe. European Journal of Phycology 40: 337‐344.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 187 of 258
9.2.3 Grateloupia turuturu
Figure 72 ‐ Grateloupia turuturu Yamada with Bifurcaria bifurcata
Finisterre, Galicia, Spain, 2008 © Ignacio Bárbara
SYMBOLS: B
TAXONOMY
Phylum Rhodophyta Class Florideophyceae Order Halymeniales Family Halymeniaceae Genus Grateloupia Species Grateloupia turuturu
Related species
The genus Grateloupia contains 87 related species
BIOLOGY
Grateloupia is compressed to foliose, linear to lanceolate, rarely unbranched, usually branched proliferously, to one or more orders, in one or more planes. Stipitate. Holdfast a basal disc. Lubricous to leathery. Medulla filamentous, with rhizoids. Inner cortex of anastomosing stellate cells, outer cortex of anticlinal files of more or less isodiametric cells, progressively smaller toward cuticle. Female reproductive structures in ampullae, sparingly branched usually to only two orders. Carpogonia terminating a 2‐celled branch on the primary ampullar filament. Connecting filaments can fuse with a succession of auxiliary cells in separate ampullae. A single, outwardly directed gonimoblast initial produces a compact, embedded cystocarp with many small carposporangia. Small auxiliary fusion cell formed. Carpostome usually present. Spermatangia superficial, sometimes in nemathecial sori. Tetrasporophyte isomorphic. Tetrasporangia cruciate, attached subbasally to intermediate cortical layer.
Grateloupia is distributed in warm temperate (to tropical) waters throughout the world. G. turuturu is a marine species.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 188 of 258
BIOTECHNOLOGY
G. filicina is used as food in Indonesia and Japan (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=114 Barreiro R., Quintela M., Bárbara I., Cremades J. (2006) RAPD differentiation of Grateloupia lanceola and the invasive Grateloupia
turuturu (Gigartinales, Rhodophyta) in the Iberian Peninsula. Phycologia 45: 213‐217. D'Archino R., Nelson W.A., Zuccarello G.C. (2007) Invasive marine red alga introduced to New Zealand waters: first record of
Grateloupia turuturu (Halymeniaceae, Rhodophyta). New Zealand Journal of Marine and Freshwater Research 41: 35‐42. De Clerck O., Gavio B., Fredericq S., Bárbara I., Coppejans E. (2005) Systematics of Grateloupia filicina (Halymeniaceae,
Rhodophyta), based on rbcl sequence analyses and morphological evidence, including the reinstatement of G. minima and the description of G. capensis sp. nov. Journal of Phycology 41: 391‐410.
Figueroa F.L., Korbee N., de Clerck O., Bárbara I., Gall E.A.R. (2007) Characterization of Grateloupia lanceola (Halymeniales, Rhodophyta), an obscure foliose Grateloupia from the Iberian Peninsula, based on morphology, comparative sequence analysis and mycosporine‐like amino acid composition. European Journal of Phycology 42: 231‐242.
Gavio B., Fredericq S. (2002) Grateloupia turuturu (Halymeniaceae, Rhodophyta) is the correct name of the non‐native species in the Atlantic known as Grateloupia doryphora. European Journal of Phycology 37: 349‐360.
Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 31 March 2010.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Verlaque M., Brannock P.M., Komatsu T., Villalard‐Bohnsack M., Marston M. (2005) The genus Grateloupia C. Agardh
(Halymeniaceae, Rhodophyta) in the Thau Lagoon (France, Mediterranean): a case study of marine plurispecific introductions. Phycologia 44: 477‐496.
Xia B.M. (2004) Flora algarum marinarum sinicarum Tomus II Rhodophyta No. III Gelidiales Cryptonemiales Hildenbrandiales, Science Press, Beijing, pp. i‐xxi, 1‐203, pls I‐XIII.
Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 189 of 258
9.2.4 Palmaria palmata
Figure 73 ‐ Palmaria palmata (Linnaeus) Kuntze
Spiddal, Co. Galway, Ireland; lower intertidal © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Rhodophyta Class Florideophyceae Order Palmariales Family Palmariaceae Genus Palmaria Species Palmaria palmata
Related species
P. callophylloides, P. decipiens, P. georgica, P. hecatensis, P. marginicrassa, P. mollis, P. moniliformis, P. stenogona.
BIOLOGY
Thallus with erect stipitate fronds arising from a basal disc, blades flattened, becoming cartilaginous with age, simple, dichotomously or palmately divided, frequently with marginal, palmately divided proliferations; construction multiaxial, cortex of 2‐5 layers of small pigmented cells increasing in number rather than size as the thallus matures to form a meristoderm, medulla compact with 2‐5 layers of large, rounded, loosely coherent, almost colourless cells with strings of bead‐like chloroplasts. Gametangial plants dioecious; spermatangia formed in large, irregular, tortuous sori over most of the surface of erect, frondose blades similar in morphology to the tetrasporangial plants; carpogonia occurring as single cells on young plantlets only; tetrasporangial plant developing directly from the fertilized carpogonium and overgrowing

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 190 of 258
the carpogonial plant, carposporophyte lacking. Tetrasporangia large, in extensive, irregular, tortuous sori generally covering much of the surface of the young frond when mature, formed in a terminal position from cortical cells, interspersed with modified, pigmented, sterile filaments, regenerating repeatedly from a basal generative stalk cell. Spores regularly cruciately or decussately arranged.
Marine. On rock, mussels and epiphytic on several algae, intertidal (at all levels but particularly near low water) and shallow subtidal , especially on upper part of Laminaria hyperborea stipes (to a depth of 5 m), widely distributed, abundant.
BIOTECHNOLOGY
Palmaria palmata is a food item of ancient usage in the North Atlantic. This species is variously known as dulse, dillisk, duilleasg or duillisg (Irish Gaelic), the narrower forms being called creannach (Irish Gaelic), soll (Icelandic), sou‐söll, søl, blôm (Norwegian), goémon à vaches (French), and darusu (Japanese). Dulse is a good source of vitamins and minerals, particularly when compared to higher‐plant vegetables (Morgan et al., 1980), and is still eaten in Scotland, Ireland, Iceland, Brittany and Norway, although, in many of these areas, its use is only occasional. Dulse contains large amount of several unusual carbohydrates including an unusual short‐chained one, floridoside, and this can form up to 30% of the dry weight. Scotland, Norway, Iceland and eastern Canada all produce small amounts of dulse for human consumption. In Ireland about 20 dry tonnes are sold each year, while in Canada 100 t are produced yearly (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=37291 http://www.algaebase.org/search/species/detail/?species_id=1
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere, A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 31 March 2010.
Irvine L.M., Guiry M.D. (1983) Palmariales. In: Irvine L. (ed.) Seaweeds of the British Isles. Volume 1. Rhodophyta, Part 2A Cryptonememiales (sensu stricto), Palmariales, Rhodymeniales, Vol.1 (2A), British Museum, London, pp. 65‐98.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Morgan K.C., Shacklock P.F., Simpson F.J. (1980) Some aspects of the culture of Palmaria palmata in greenhouse tanks. Botanica
Marina 23: 765‐770. Morgan K.C., Wright J.L.C., Simpson F.J. (1980) Review of chemical constituents of the red alga Palmaria palmata (dulse). Economic
Botany 34: 27‐50. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 191 of 258
9.2.5 Solieria chordalis
Figure 74 ‐ Solieria chordalis (C.Agardh) J.Agardh
© Stephanie Bondu (UBO‐Brest, France)
SYMBOLS: B
TAXONOMY
Phylum Rhodophyta Class Florideophyceae Order Gigartinales Family Solieriaceae Genus Solieria Species Solieria chordalis
Related species
S. anastomosa, S. dichotoma, S. dura, S. filiformis, S. jaasundii, S. pacifica, S. robusta, S. tenuis.
BIOLOGY
Plants of the largest species reach 30 cm in length. Thalli are erect or recumbent from a crustose or fibrous holdfast, terete to compressed, and irregularly to subdichotomously branched. Apices are multiaxial, the successive axial cells producing single periaxial derivatives either in orthostichous rows or rotated 120 degrees in a zig zag fashion. The medulla is broad and laxly filamentous with abundant rhizoids, and is surrounded by a cortex of progressively smaller subisodiametric cells. Carpogonial branches are 3‐celled, at times bearing sterile cells on the basal cell, and emit a single non‐septate connecting filament from fertilized carpogonia. Auxiliary cells are prominently situated in an "auxiliary cell complex" prior to diploidization. Cystocarps are deeply embedded in the axes, often in clusters, and consist of an ostiolate pericarp with a filamentous inner hull surrounding a carposporophyte in which a large central fusion cell

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 192 of 258
gives rise peripherally to short chains of gonimoblast cells bearing single, terminal carposporangia. Tetrasporangia are laterally pit‐connected to bearing cells.
S. chordalis is a marine species.
BIOTECHNOLOGY
Algae of the genus Solieria are used as food in Myanmar (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=33013 Bondu S., Cocquempot B., Deslandes E., Morin P. (2008) Effects of salt and light stress on the release of volatile halogenated
organic compounds by Solieria chordalis: a laboratory incubation study. Botanica Marina 51: 485‐492. Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere,
A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls. De Masi F., Gargiulo G.M. (1982) Solieria chordalis (C. Ag.) J. Ag. (Rhodophyta, Gigartinales) en Méditerranée. Allionia 25: 109‐111. Deslandes E., Floc'h J.Y., Bodeau‐Bellion C., Brault D., Braud, J.P. (1985) Evidence for (iota)‐carrageenans in Solieria chordalis
(Solieriaceae) and Calliblepharis jubata, Calliblepharis purpureum (Rhodophyllidaceae). Botanica Marina 28: 317‐318. Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 30 June 2010. Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 193 of 258
9.3 Phaeophyceae (phylum Heterokontophyta)
9.3.1 Alaria esculenta
Figure 75 ‐ Alaria esculenta (Linnaeus) Greville
Spiddal, Co. Galway, Ireland. Fronds exposed at low water; 16‐mm lens © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Alariaceae Genus Alaria Species Alaria esculenta
Related species
A. angusta, A. crassifolia, A. crispa, A. elliptica, A. esculenta f. latifolia, A. esculenta f. angustifolia, A. fragilis, A. grandifolia, A. marginata, A. ochotensis, A. paradisea, A. praelonga, A. pylaiei.
BIOLOGY
Plants with olive or yellow‐brown fronds to 4 m long and 25 cm wide. Attached by a root‐like holdfast at the base from which a narrow flexible stipe arises which continues into the leafy part of the plant as a distinct mid‐rib. The reproductive structures, apparent as dark‐brown areas, are confined to unbranched leafy appendages borne on the stipe, usually in two rows. This is the only kelp‐like plant in Ireland and Britain with a distinct midrib and is the only one with sporangia borne at the base of the frond in special leaflets called sporophylls.
Generally growing on rock in very exposed places, often forming a band at low water and in the shallow subtidal, but also occurring in tidal pools in the lower shore.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 194 of 258
BIOTECHNOLOGY
A. esculenta, as well as other Alaria species, is used as food in European, North American and Asian countries (Zemke‐White and Ohno, 1999). For further information on Alaria biotechnology see paragraph Biotechnology for Laminaria, Saccharina and Saccorhiza.
References
AlgaeBase: http://www.algaebase.org/search/species/detail/?species_id=82 Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere,
A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls. Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 31 March 2010. Lane C.E., Saunders G.W. (2005) Molecular investigation reveals epi/endophytic extrageneric kelp (Laminariales, Phaeophyceae)
gametophytes colonizing Lessoniopsis littoralis thalli. Botanica Marina 48: 426‐436. Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. [1]‐215. Widdowson T.B. (1971) A taxonomic revision of the genus Alaria Greville. Syesis 4: 11‐49. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 195 of 258
9.3.2 Undaria pinnatifida
Figure 76 ‐ Undaria pinnatifida (Harvey) Suringar
Venice, Italy; on chains of vaporetto stop © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Alariaceae Genus Undaria Species Undaria pinnatifida
Related species
U. crenata, U. undarioides
BIOLOGY
Life history diplohaplontic with alternation of large sporophyte bearing unilocular meiosporangia with paraphyses (sori) and microscopic dioecious and oogamous, heteromorphous gametophytes (for detail, see Laminaria). Haploid chromosome number is 30 in U. pinnatifida. Sporophyte annual, appearing in winter and disintegrating the following autumn. Sporophyte composed of holdfast with haptera, stipe, and blade. Meristematic region situated at transition zone between stipe and blade. Stipe compressed at base to flattened above, with wings (greatly expanded to narrow) which are more or less undulato‐plicated and with sori or sterile. Blade linear to rounded or with pinnate lobes, with midrib or thickened fascia. Cryptostomata and dot‐like mucilage glands present. Sori develop in summer on both surfaces of wings (sporophylls), or on both sides of midrib or fascia, or at the same time on sporophylls and blade. In culture,

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 196 of 258
sori of U. pinnatifida are formed in longday conditions, probably induced by high water temperatures. Zoospores of U. pinnatifida germinate between 13 and 24°C gametophytes grow well at 15‐23°C (Saito 1956). Survival range of gamtophytes is ‐1 to 28°C. Optimal temperatures for formation and growth of U. pinnatifida sporophytes is 10‐20°C but at 5 and 25°C sporophytes are still formed. Structure of sporophyte as in Laminaria composed of photosynthetic meristoderm, parenchymatic cortex and central medulla. In U. pinnatifida parthenosporophytes with normal morphology may develop at low rates releasing only female determined zoospores when maturing.
Genus originally endemic to the northwestern Pacific, growing subtidally on rocks in warm‐temperate waters of Japan, Korea and China. Recently, U. pinnatifida was introduced to France, New Zealand and Tasmania, probably via oyster cultures and ships. Since then the species is expanding its distribution range.
BIOTECHNOLOGY
Genus of great economic importance as food source in Japan and Korea, especially U. pinnatifida (trade name: Wakame). Total annual yield from natural harvest and cultivation sites of Undaria spp. was about 130,000 tons fresh weight in 1967. Long‐line culture started in the early 1960's and meanwhile production increased to about 30% of the wild harvest in Japan. In recent years, first progress in tissue culture, protoplast isolation and cryopreservation of U. pinnatifida was achieved. Annual productions in t dry weight are: Australia 6, China 20,000, Japan 18,310 (all from culture) and Korea 83,398 (all from culture) (Zemke‐White and Ohno, 1999). For further information on Undaria biotechnology see pargraph on Biotechnology for Laminaria, Saccharina and Saccorhiza.
References
Adams N.M. (1997) Common seaweeds of New Zealand, Canterbury University Press, Christchurch, pp. 1‐48, 48 pls. AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=32939 Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 31 March 2010. Lane C.E., Mayes C., Druehl L.D., Saunders G.W. (2006) A multi‐gene molecular investigation of the kelp (Laminariales,
Phaeophyceae) supports substantial taxonomic re‐organization. Journal of Phycology 42: 493‐512. Pedroche P.F., Silva P.C., Aguilar Rosas L.E., Dreckmann K.M., Aguilar Rosas R. (2008) Catálogo de las algas benthónicas del Pacífico
de México II. Phaeophycota, Universidad Autónoma Metropolitana and University of California Berkeley, Mexicali & Berkeley, pp. i‐vi, 15‐146.
Sliwa C., Johnson C.R., Hewitt C.L. (2006) Mesoscale dispersal of the introduced kelp Undaria pinnatifida attached to unstable substrata. Botanica Marina 49: 396‐405.
Yoshida T. (1998) Marine algae of Japan. Uchida Rokakuho Publishing, Tokyo, pp. 25 ‐ 1222. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 197 of 258
9.3.3 Ascophyllum nodosum
Figure 77 ‐ Ascophyllum nodosum (Linnaeus) Le Jolis
Céibh an tSrutháin, an Cheathrú Rua, Co. na Gaillimhe; 10.5‐mm lens © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Fucaceae Genus Ascophyllum Species Ascophyllum nodosum
Related species
The genus Ascophyllum only includes Ascophyllum nodosum. A free floating variety is known as Ascophyllum nodosum var. mackayi (Turner) Cotton.
BIOLOGY
This is a brown seaweed that is closely related to Fucus. It forms a single bladders centrally in long, flattened strap‐like fronds. The fronds hang downwards, draping sheltered intertidal rocks. Many fronds grow from the base and the plant generally regenerates new fronds from the base when one of the larger fronds are damaged. There is evidence that clumps can be over 400 years old and may be even older. Ascophyllum is currently confined to the North Atlantic basin, but plants have been found growing in San Francisco Bay, but the species does not persist there. The plants are used as packing for shellfish from the North Atlantic and when discarded may take hold.
BIOTECHNOLOGY

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 198 of 258
Ascophyllum nodosum is used in agriculture as a fertilizer and plant growth promoter (France, Canada, China, Iceland, US) and for alginate production (Ireland, Norway, UK) (Zemke‐White and Ohno, 1999). All the annual production is harvested from the wild. Annual production is: Ireland 8,999 t dry weight, Norway 6,632, Iceland 4,400, UK 3,500, China 3,000, Canada 2,500, France 1,700 and US 280 (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/species/detail/?species_id=5 Baardseth E. (1970) Synopsis of biological data on knobbed wrack Ascophyllum nodosum (Linnaeus) Le Jolis. FAO Fisheries Synopsis
38 (Rev. 1). Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere,
A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls. Cho G.Y, Rousseau F., Reviers B. De, Boo S.M. (2006) Phylogenetic relationships within the Fucales (Phaeophyceae) assessed by the
photosystem I coding psaA sequences. Phycologia 45: 512‐519. Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 31 March 2010. Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. McHugh D.J. (2003) A guide to the seaweed industry. FAO Fisheries Technical Paper 441. Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐
376.
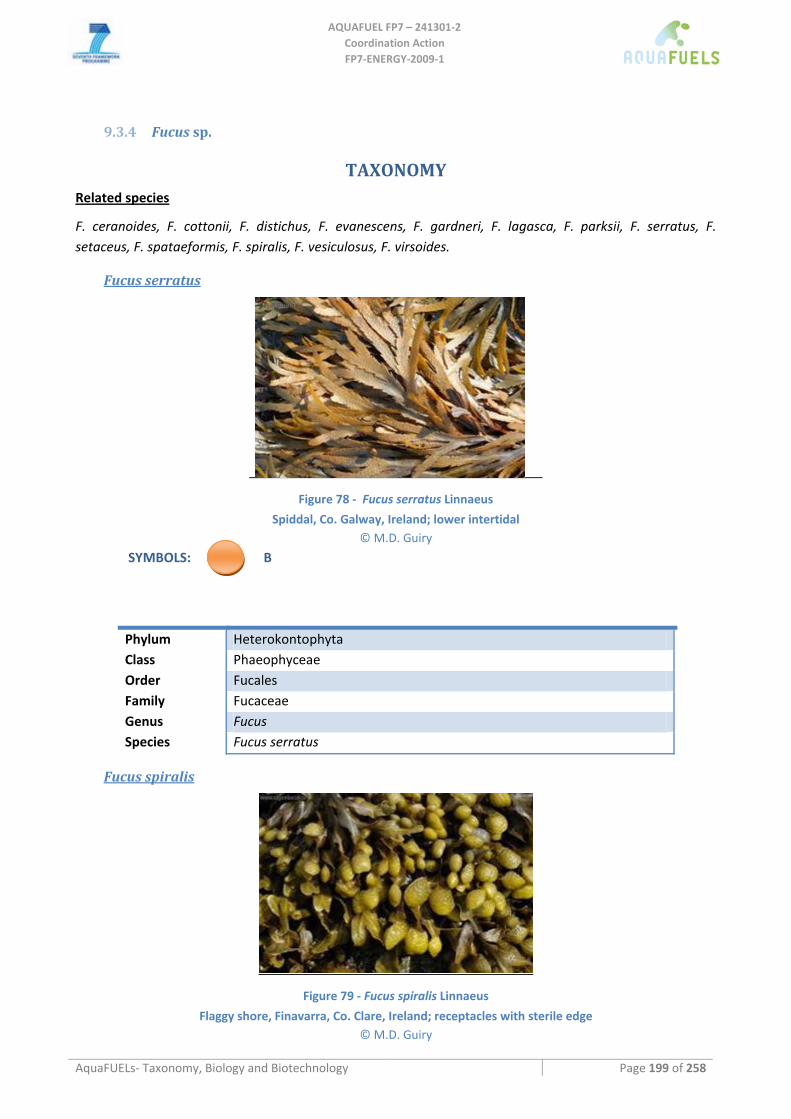
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 199 of 258
9.3.4 Fucus sp.
TAXONOMY
Related species
F. ceranoides, F. cottonii, F. distichus, F. evanescens, F. gardneri, F. lagasca, F. parksii, F. serratus, F. setaceus, F. spataeformis, F. spiralis, F. vesiculosus, F. virsoides.
Fucus serratus
Figure 78 ‐ Fucus serratus Linnaeus
Spiddal, Co. Galway, Ireland; lower intertidal © M.D. Guiry
SYMBOLS: B
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Fucaceae Genus Fucus Species Fucus serratus
Fucus spiralis
Figure 79 ‐ Fucus spiralis Linnaeus
Flaggy shore, Finavarra, Co. Clare, Ireland; receptacles with sterile edge © M.D. Guiry
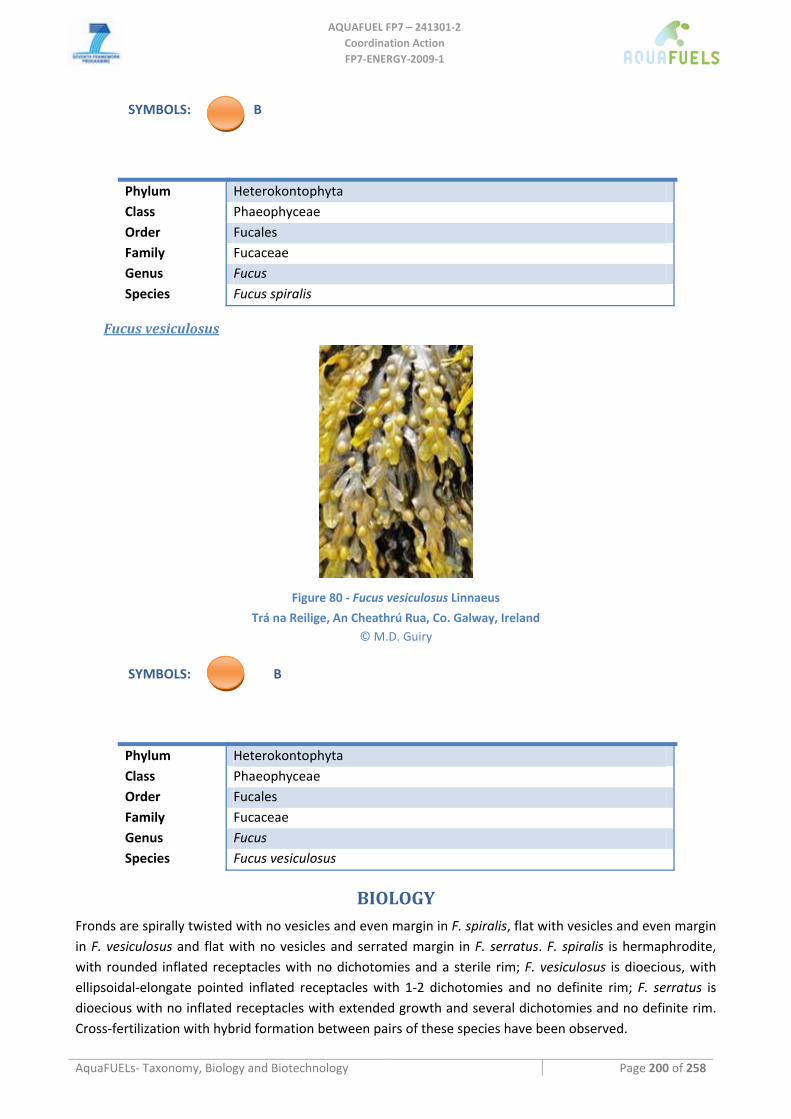
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 200 of 258
SYMBOLS: B
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Fucaceae Genus Fucus Species Fucus spiralis
Fucus vesiculosus
Figure 80 ‐ Fucus vesiculosus Linnaeus
Trá na Reilige, An Cheathrú Rua, Co. Galway, Ireland © M.D. Guiry
SYMBOLS: B
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Fucaceae Genus Fucus Species Fucus vesiculosus
BIOLOGY
Fronds are spirally twisted with no vesicles and even margin in F. spiralis, flat with vesicles and even margin in F. vesiculosus and flat with no vesicles and serrated margin in F. serratus. F. spiralis is hermaphrodite, with rounded inflated receptacles with no dichotomies and a sterile rim; F. vesiculosus is dioecious, with ellipsoidal‐elongate pointed inflated receptacles with 1‐2 dichotomies and no definite rim; F. serratus is dioecious with no inflated receptacles with extended growth and several dichotomies and no definite rim. Cross‐fertilization with hybrid formation between pairs of these species have been observed.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 201 of 258
F. spiralis is normally found in the highest position on the shore, where it may be left exposed for several days during neap tides, F. vesiculosus occupies the mid‐tide zone in competition with Ascophyllum nodosum and is alternatively exposed‐submerged, while F. serratus occupies the lowest level, that is more rarely exposed.
BIOTECHNOLOGY
Several Fucus species are used as food (France, Portugal, Alaska), for agriculture uses (France, Canada), and as alginate source (Ireland). Annual production of Fucus is of 80 t dry weight in Ireland, 2 in France and 0.04 in Portugal (Zemke‐White and Ohno, 1999).
References
Billard E., Daguin C., Pearson G., Serrao E., Engel C., Valero M. (2005) Genetic isolation between three closely related taxa: Fucus vesiculosus, F. spiralis and F. ceranoides (Phaeophyceae). Journal of Phycology 41: 900‐905.
Billard E., Serrao E.A., Pearson G.A., Engel C.R., Destombe C., Valero M. (2006) Analysis of sexual phenotype and prezygotic fertility in natural populations of Fucus spiralis, F. vesiculosus (Fucaceae, Phaeophyceae) and their putative hybrids. European Journal of Phycology 40: 397‐407.
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere, A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Brawley S.H., Coyer J.A., Blakeslee A.M., Hoarau G., Johnson L.E., Byers J.E., Stam W.T., Olsen J.L. (2009) Historical invasions of the intertidal zone of Atlantic North America associated with distinctive patterns of trade and emigration. Proceedings of the National Academy of Sciences of the United States of America 106: 8239‐8244.
Burrows E.M., Lodge S. (1951) Autoecology and the species problem in Fucus. Journal of the Marine Biological Association of the United Kingdom 30: 161‐176.
Cerantola S., Breton F., Ar Gall E., Deslandes E. (2006) Co‐occurrence and antioxidant activities of fucol and fucophlorethol classes of polymeric phenols in Fucus spiralis. Botanica Marina 49: 347‐351.
Coleman M.A., Brawley S.H. (2005) Are life history characteristics good predictors of genetic diversity and structure? A case study of the intertidal alga Fucus spiralis (Heterokontophyta, Phaeophyceae). Journal of Phycology 41: 753‐762.
Gabrielson P.W., Widdowson T.B., Lindstrom S.C. (2004) Keys to the seaweeds and seagrasses of Oregon and California. Phycological Contribution 6: iv ‐ 181.
Garbary D.J., Brackenbury A., McLean A.M., Morrison D. (2006) Structure and development of air bladders in Fucus and Ascophyllum (Fucales, Phaeophyceae). Phycologia 45: 557‐566.
Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 31 March 2010.
Larsen A., Sand‐Jensen K. (2005) Salt tolerance and distribution of estuarine benthic macroalgae in the Kattegat‐Baltic Sea area. Phycologia 45: 13‐23.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Mathieson A.C., Dawes C.J., Wallace A.L., Klein A.S. (2006) Distribution, morphology, and genetic affinities of dwarf embedded
Fucus populations from the Northwest Atlantic Ocean. Botanica Marina 49: 283‐303. Moss B.L. (1950) Studies in the genus Fucus. II. The anatomical structure and chemical composition of receptacles of Fucus
vesiculosus from three contrasting habitats. Annals of Botany 14: 395‐410. Moss B.L. (1950) Studies in the genus Fucus III. Structure and development of the attaching discs of Fucus vesiculosus. Annals of
Botany 14: 411‐419. Nägeli C. (1849) Gattungen einzelliger Algen, physiologisch und systematisch bearbeitet. Neue Denkschriften der Allg.
Schweizerischen Gesellschaft für die Gesammten Naturwissenschaften 10(7): i‐viii, 1‐139, pls I‐VIII. Pearson G., Lago‐Leston A., Valente M., Serrao E. (2006) Simple and rapid RNA extraction from freeze‐dried tissue of brown algae
and seagrasses. European Journal of Phycology 41: 97‐104. Peters A.F., Marie D., Scornet D., Kloareg, B., Cock J.M. (2004) Proposal of Ectocarpus siliculosus (Ectocarpales, Phaeophyceae) as a
model organism for brown algal genetics and genomics. Journal of Phycology 40: 1079‐1088. Perrin C., Daguin C., Van de Vliet M., Engel C.R., Pearson G.A., Serráo E.A. (2007) Implications of mating system for genetic diversity
of sister algal species: Fucus spiralis and Fucus vesiculosus (Heterokontophyta, Phaeophyceae). European Journal of Phycology 42: 219‐230.
Powell H.T. (1960) The typification of Fucus spiralis L. British Phycological Bulletin 2: 17. Serrão E.A., Alice L.A., Brawley S.H. (1999) Evolution of the Fucaceae (Phaeophyceae) inferred from nrDNA‐ITS. Journal of Phycology
35: 382‐394. Wallace A.L., Klein A.S., Mathieson A.C. (2004) Determining the affinities of salt marsh fucoids using microsatellite markers:
evidence of hybridization and introgression between two species of Fucus (Phaeophyta) in a Maine estuary. Journal of Phycology 40: 1013‐1027.
Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 202 of 258
9.3.5 Himanthalia elongata
Figure 81 ‐ Himanthalia elongata (Linnaeus) S.F. Gray
Les Glénan islands, Finistère, Bretagne, France. © Benoit Queguineur
SYMBOLS: B
TAXONOMY
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Himanthaliaceae Genus Himanthalia Species Himanthalia elongata
Related species
The genus Himanthalia only includes the specie Himanthalia elongata.
BIOLOGY
The growth form of Himanthalia differs from that of any other member of the Fucales. The young plants are first recognizable on the shore as small vesicles, pale greenish‐ brown in colour. These vesicles flatten out and after I‐2 years growth they form the small leathery disc‐shaped structure which is commonly referred to as the 'button' stage. From these 'buttons' the long thongs arise, and in some habitats they may reach a length of about 2 m or more. The 'button' is generally interpreted as equivalent to the vegetative thallus of the other Fucales, while the thongs correspond to the fertile receptacles. Gametes are shed over a long period, they are released from late July, on through the winter. In Himanthalia only one vegetative meristem is ever produced. The single apical cell of this divides many times; each new daughter cell becoming the apical cell of a reproductive apex. Each of these has deter‐ minate growth. By the time that the gametes are shed the identity of the apical meristem has been lost and the receptacle subsequently

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 203 of 258
disintegrates. There is no meristem left on the plant to continue e vegetative growth, and once an apical cell is lost the plant is unable to regenerate a new one.
Himanthalia grows at, or near low water level, so that in some habitats the plants are uncovered at low spring tides, whereas in other localities they may never be exposed.
BIOTECHNOLOGY
Himnathalia elongata is used as food (MacArtain et al., 2007).
References
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere, A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Cho G.Y, Rousseau F., Reviers B. De, Boo S.M. (2006) Phylogenetic relationships within the Fucales (Phaeophyceae) assessed by the photosystem I coding psaA sequences. Phycologia 45: 512‐519.
Gallardo Garciá T., Pérez‐Ruzafa I.M. (2001) Himanthalia Lyngb. In: Gómez Garreta, A. (ed.) Flora phycologica iberica Vol. 1 Fucales., Universidad de Murcia, Murcia, pp. 69‐71.
Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 31 March 2010.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Moss B. (1969) Apical meristems and growth control in Himanthalia elongata (S. F. Gray). New Phytologist 68: 387‐397. MacArtain P., Gill C.I.R., Brooks M., Campbell R., Rowland I.R.(2007) Nutritional value of edible seaweeds. Nutrition Reviews 65:
535‐543. Setchell W.A. (1931) Some early algal confusions. University of California Publications in Botany 16: 351‐366.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 204 of 258
9.3.6 Cystoseira sp.
TAXONOMY Related species
C. abies‐marina, C. algeriensis, C. amentacea, C. baccata, C. barbata, C. barbatula, C. brachycarpa, C. compressa, C. corniculata, C. crassipes, C. crinita, C. crinitophylla, C. dubia, C. elegans, C. foeniculacea, C. funkii, C. geminata, C. hakodatensis, C. helvetica, C. humilis, C. hyblaea, C. indica, C. jabukae, C. mauritanica, C. mediterranea, C. montagnei, C. myrica, C. neglecta, C. nodicaulis, C. occidentalis, C osmundacea, C. pelagosae, C. planiramea, C. platyclada, C. sauvageauana, C. schiffneri, C. sedoides, C. setchellii, C. sonderi, C. spinosa, C. squarrosa, C. susanensis, C. tamariscifolia, C. trinodis, C. usneoides, C. wildpretii, C. zosteroides.
Cystoseira baccata
Figure 82 ‐ Cystoseira baccata (S.G. Gmelin) P.C. Silva
Finavarra, Co. Clare, Ireland; MLWN lagoon © M.D. Guiry
SYMBOLS: B
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Sargassaceae Genus Cystoseira Species Cystoseira baccata

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 205 of 258
Cystoseira tamariscifolia
Figure 83 ‐ Cystoseira tamariscifolia (Hudson) Papenfuss
Les Glénan islands, Finistère, Bretagne, France. © Benoit Queguineur
SYMBOLS: B
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Sargassaceae Genus Cystoseira Species Cystoseira tamariscifolia
BIOLOGY
C. baccata plants are usually solitary, 1 m or more in length, attached by a thick, conical attaching disc. Axis simple or branched, up to 1 m in length, flattened, about 1 x 0.4 cm in transverse section; apex smooth and surrounded during periods of active growth by incurred young laterals. Lateral branch systems distichous, alternate, radially symmetrical, profusely branched in a repeatedly pinnate fashion and bearing sparse, filiform, occasionally bifurcate appendages on the branches of higher orders; deciduous, leaving decurrent bases which give an irregular, zigzag outline to the axis. Cryptostomata lacking. Aerocysts present in axes of branches of higher order, sometimes in chains; seasonal, particularly numerous in autumn. Receptacles 1‐5 cm long, formed from axes of ultimate ramuli, irregularly nodose and bearing simple, filiform appendages.
BIOTECHNOLOGY
No biotechnological use known, except at research level (contaminant biosorption, production of bioactive metabolites).
References
AlgaeBase: http://www.algaebase.org/search/species/detail/?species_id=92

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 206 of 258
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere, A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 31 March 2010.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Sánchez I., Fernández C., Arrontes J. (2005) Long‐term changes in the structure of intertidal assemblages following invasion by
Sargassum muticum (Phaeophyta). Journal of Phycology 41: 942‐949.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 207 of 258
9.3.7 Halidrys siliquosa
Figure 84 ‐ Halidrys siliquosa (Linnaeus) Lyngbye
Spanish Point, Co. Clare, Ireland; lower intertidal pools © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Sargassaceae Genus Halidrys Species Halidrys siliquosa
Related species
H. dioica, H. murmanica
BIOLOGY
Thallus to 2 m long, compressed tough and leatherly, attached by strong holdfast. Branches distichous and regularly alternate. Air bladders, up to 5 cms long, are oblong and stalked towards the ends of the branches. The air bladders are divided by cross walls visible when sectioned longitudinally. Common. Low littoral rock pools and upper sub‐littoral. Widespread around the British Isles. Europe: Portugal, Atlantic coasts of Spain and France, Netherlands, Baltic Sea, Norway and the Faroes.
BIOTECHNOLOGY
No biotechnological use known, except at research level (production of bioactive metabolites).
References
Braune W. (2008). Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere., A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org; searched on 31 March 2010.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Morton O., Picton B.E. (2010) Halidrys siliquosa. In: Encyclopedia of Marine Life of Britain and Ireland
http://www.habitas.org.uk/marinelife/species.asp?item=ZR7160

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 208 of 258
9.3.8 Sargassum muticum
Figure 854 ‐ Sargassum muticum (Yendo) Fensholt
Finavarra, Co. Clare, Ireland; © M.D. Guiry
SYMBOLS: B
TAXONOMY
Phylum Heterokontophyta Class Phaeophyceae Order Fucales Family Sargassaceae Genus Sargassum Species Sargassum muticum
Related species
The genus Sargassum contains 345 related species.
BIOLOGY
Thallus to 10‐200 cm or more in length, with one to a few simple, terete to compressed, stipes 1‐20 cm long arising from a discoid‐conical holdfast. Stipes bearing radially or distichously borne, long primary branches, produced seasonally from the stipe apices and subsequently deciduous, leaving scars or other residues on the stipe. Primary branches 10 cm to 200 cm or more long, distichously, tristichously or radially branched with a terete, angular, compressed or three‐sides axes; basal laterals simple or branched, compressed and relatively narrow to (in most species) leaf‐like, (1‐)3‐15(‐25) mm broad, entire ot with dentate margins; upper laterals usualy branched, with slender, compressed to terete, ramuli. Air bladders (vesicles) normally present, subspherical to ovoid, petiolate, mutic or apiculate, replacing ramuli or axilliary to the laterals.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 209 of 258
Growth from a 3‐sided cell in an apical depression. Structure of a central medulla of elonagate cells in the stipe and branch axes, with a core of isodiametric cells and a surface of phaeoplastic meristoderm, active in the larger branched. Reproduction. Thallus monoecious or dioecious. Receptacles unisexual or bisexual, developed in axils of laterals or ramuli, simple or usually in branched clusters, terete or compressed, smooth, verrucose or spinous, with scattered conceptacles and ostioles, growing apically with conceptacles maturing below; conceptacles unisexual or bisexual.
BIOTECHNOLOGY
Sargassum is used as food in many countries in Asia as well as in Hawaii. It is also used for the production of alginate and, in few countries, for agricultural purposes. Annual production , all from wild harvesting, is of 5,000 t dry weight in the Philippines, 2,249 in India and 400 in Vietnam (Zemke‐White and Ohno, 1999).
References
AlgaeBase: http://www.algaebase.org/search/genus/detail/?genus_id=77 Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere,
A.R.G. Gantner Verlag., Ruggell, pp. 1‐596, 266 pls. Cho G.Y, Rousseau F., Reviers B. De, Boo S.M. (2006) Phylogenetic relationships within the Fucales (Phaeophyceae) assessed by the
photosystem I coding psaA sequences. Phycologia 45: 512‐519. Connan S., Delisle F., Deslandes E., Gall E.A. (2006). Intra‐thallus phlorotannin content and antioxidant activity in Phaeophyceae of
temperate waters. Botanica Marina 49: 39‐46. Farnham W.F., Fletcher R.L., Irvine L.M. (1973) Attached Sargassum found in Britain. Nature 243: 231‐232. Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 31 March 2010. Kilar J.A., Hanisak M.D., Yoshida T. (1992) On the expression of phenotypic variability: why is Sargassum so taxonomically difficult?.
In: Abbott I.A. (ed.) Taxonomy of Economic Seaweeds with reference to Pacific and Western Atlantic species, Vol.3, California Sea Grant College, La Jolla, California, pp. 95‐117.
Lee K., Yoo S.A. (1992) Korean species of Sargassum subgenus Bactrophycus J. Agardh (Sargassaceae, Fucales In: Abbott I.A. (ed.) Taxonomy of Economic Seaweeds with reference to Pacific and Western Atlantic species, Vol.3, California Sea Grant College, La Jolla, California, pp. 139‐147
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. Pedroche P.F., Silva P.C., Aguilar Rosas L.E., Dreckmann K.M., Aguilar Rosas R. (2008) Catálogo de las algas benthónicas del Pacífico
de México II. Phaeophycota, Universidad Autónoma Metropolitana and University of California Berkeley, Mexicali & Berkeley, pp. i‐vi, 15‐146.
Phillips N. (1995) Biogeography of Sargassum (Phaeophyta) in the Pacific basin. Fucales In: Abbott I.A. (ed.) Taxonomy of Economic Seaweeds with reference to Pacific and Western Atlantic species, Vol.5, California Sea Grant College, La Jolla, California, pp. 107‐145.
Stiger V., Horiguchi T., Yoshida T., Coleman A.W., Masuda M. (2003) Phylogenetic relationships within the genus Sargassum (Fucales, Phaeophyceae), inferred from it ITS nrDNA, with an emphasis on the taxonomic revision of the genus. Phycological Research 51: 1‐10.
Tseng C.K., Yoshida T., Chiang Y.M. (1985) East Asiatic species of Sargassum subgenus Bactrophycus J.Agardh (Sargassaceae, Fucales), with keys to the sections and species. In: Abbott I.A., Norris J.N. (eds.) Taxonomy of Economic Seaweeds with reference to some Pacific and Caribbean species, Vol.I, California Sea Grant College, La Jolla, California, pp. 1‐14.
Varela‐Alvarez E., Andreakis N., Lago‐Leston A., Pearson G.A., Serrao E.A., Procaccini G., Duarte C.M., Marba N. (2006) Genomic DNA isolation from green and brown algae (Caulerpales and Fucales) for microsatellite library construction (Note). Journal of Phycology 42: 741‐745.
Yoshida T., Stiger V., Horiguchi T. (2000) Sargassum boreale sp. nov. (Fucales, Phaeophyceae) from Hokkaido, Japan. Phycological Research 48: 125‐132.
Zemke‐White W.L., Ohno M. (1999) World seaweed utilisation: an end‐of‐century summary. Journal of Applied Phycology 11: 369‐376.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 210 of 258
9.3.9 Laminaria, Saccharina, Saccorhiza
TAXONOMY
Laminaria sp.
Related species
L. abyssalis, L. agardhii, L. angustata, L. appressirhiza, L. bongardiana f. taeniata, L. bongardiana f. sub simplex, L. brasiliensis, L. bulbosa var. nodipes, L. bulbosa var. gymnopoda, L. bulbosa var. brevipes, L. bulbosa var. linearis, L. bullata, L. complanata, L. cordata, L. digitata, L. digitata var. ligulata, L. digitata var. lyrata, L. digitata var. pseudosaccharina, L. digitata var. elliptica, L. digitata var. bifida, L. ephemera, L. farlowii, L. hyperborea f. cucullata, L. hyperborea, L. inclinatorhiza, L. japonica f. membranacea, L. japonica f. longipes, L. longipes, L. multiplicata, L. nigripes, L. ochroleuca, L. pallida, L. platymeris, L. rodriguezii, L. ruprechtii, L. saccharina var. vividissima, L. saccharina var. vittata, L. saccharina var. vividissima, L. setchellii, L. sinclairii, L. solidungula, L. yezoensis.
Laminaria digitata
Figure 86 ‐ Laminaria digitata (Hudson) J.V. Lamouroux
Spiddal, Co. Galway, Ireland; plants on rocks at MLWS © M.D. Guiry
SYMBOLS: B, PIV
Phylum Heterokontophyta Class Phaeophyceae Order Laminariales Family Laminariaceae Genus Laminaria Species Laminaria digitata
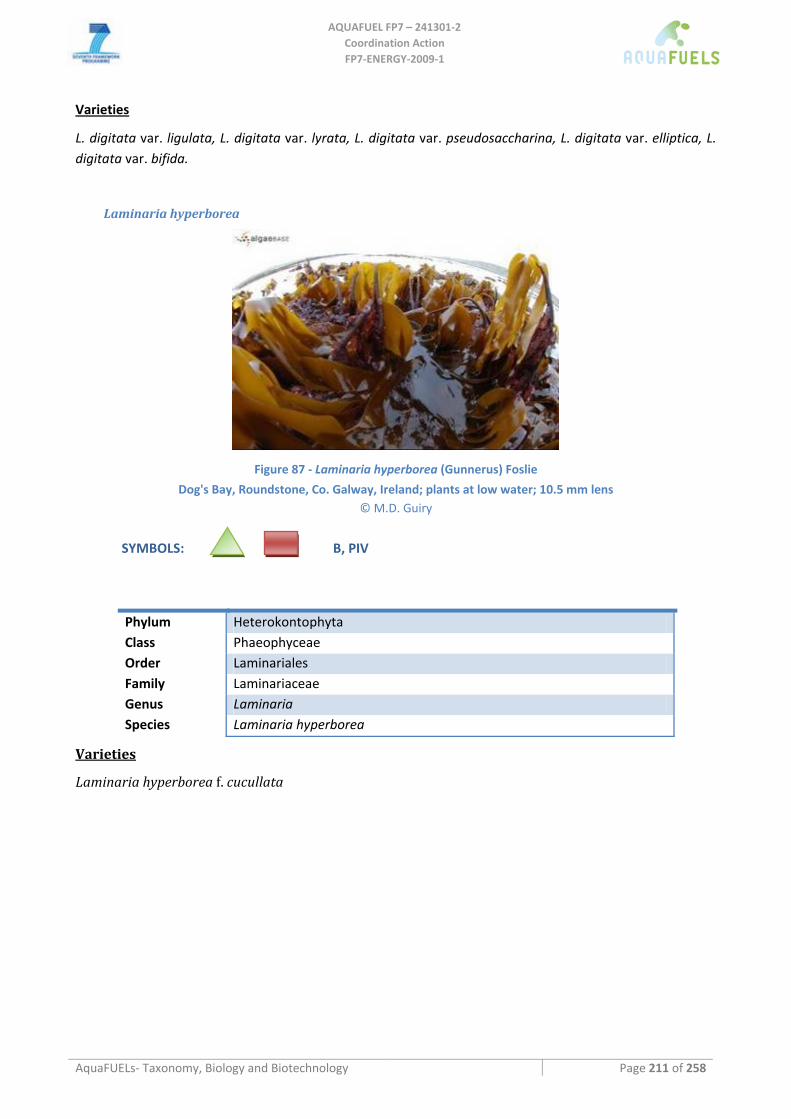
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 211 of 258
Varieties
L. digitata var. ligulata, L. digitata var. lyrata, L. digitata var. pseudosaccharina, L. digitata var. elliptica, L. digitata var. bifida.
Laminaria hyperborea
Figure 87 ‐ Laminaria hyperborea (Gunnerus) Foslie
Dog's Bay, Roundstone, Co. Galway, Ireland; plants at low water; 10.5 mm lens © M.D. Guiry
SYMBOLS: B, PIV
Phylum Heterokontophyta Class Phaeophyceae Order Laminariales Family Laminariaceae Genus Laminaria Species Laminaria hyperborea
Varieties
Laminaria hyperborea f. cucullata

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 212 of 258
Laminaria ochroleuca
Figure 88 ‐ Laminaria ochroleuca Bachelot de la Pylaie
Spain, Galicia, A Coruña, Ría de A Coruña, 2004, lower intertidal © Ignacio Bárbara
SYMBOLS: B, PIV
Phylum Heterokontophyta Class Phaeophyceae Order Laminariales Family Laminariaceae Genus Laminaria Species Laminaria ochroleuca
Saccharina latissima
Figure 89 ‐ Saccharina latissima (Linnaeus) C.E. Lane, C. Mayes, L.D. Druehl & G.W. Saunders
New Quay, Co. Clare, Ireland; plants at MLWS © M.D. Guiry

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 213 of 258
SYMBOLS: B, PIV
Phylum Heterokontophyta Class Phaeophyceae Order Laminariales Family Laminariaceae Genus Saccharina Species Saccharina latissima
Related species
S. angustata, S. angustata subsp. siberica, S. bongardiana f. taeniata, S. bongardiana f. subsimplex, S. bongardiana f. subsessilis, S. bongardiana f. bifurcata, S. bongardiana, S. cichorioides f. sikotanensis, S. cichorioides f. sachalinensis, S. cichorioides f. coriacea, S. cichorioides, S. cichorioides f. sinuicola, S. coriacea, S. crassifolia, S. dentigera, S. groenlandica, S. gurjanovae f. lanciformis, S. gurjanovae, S. gyrata, S. japonica, S. japonica f. diabolica, S. japonica f. longipes, S. kurilensis, S. longicruris, S. longipedalis, S. longissima, S. ochotensis, S. plana, S. religiosa, S. sculpera, S. sessilis, S. yendoana.
Saccorhiza polyschides
Figure 90 – Saccorhiza polyschides (Lightfoot) Batters
Cooliva Quay, Co. Clare, Ireland; plants on sandy rocks at low water © M.D. Guiry
SYMBOL: B, PIV
Phylum Heterokontophyta Class Phaeophyceae Order Tilopteridales Family Phyllariaceae Genus Saccorhiza Species Saccorhiza polyschides

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 214 of 258
Related species
S. dermatodea
BIOLOGY
General Description of Major divisions and classes
Laminaria and Saccharina are two small genera of large brown seaweeds commonly called kelps. The order of the Laminariales share an almost identical life history: The sporophyte is the dominant phase and the gametophyte is a microscopic phase. There is therefore an alternation of sporophyte and gametophyte. Sporophytes of both genera are differentiated into holdfast (below), stipe and lamina; the gametophyte is undifferentiated and is filamentous and creeping.
Sexual reproduction is oogamous; antherozoids (male gametes) are produced by a male gametophyte and eggs are produced in oogonia by the female gametophyte. The female liberates a pheromone called lamoxirene, a low molecular weight (C11), volatile hydrocarbon which, in addition to attracting the sperm, initiates their release from the antheridia of the male gametophyte. Fertilization takes place with the egg partially extruded from the oogonium and the zygote develops in situ to form a sporophyte (the macroscopic kelp). The sporophyte develops sporangia on the surface of the blades in unilocular sporangia (called such because they are not divided by cross walls or locules); the contents of these sporangia divide meiotically and form numerous flagellated zoospores which are haploid. These zoospores swim away and eventually settle and develop into the gametangial thalli. The life history is therefore heteromorphic; the gametophytes and the sporophyte have a different morphology.
Laminaria and Saccharina species are found on rocky shores at low tide and in the subtidal to depths of 8‐30 m in the north Atlantic and north Pacific; some species occur at depths of up to 120 m (Mediterranean and Brazil), but this requires extraordinary water clarity.
Saccorhiza polyschides sporophytes are annual, a single stipe attached initially by a small disc, sometimes with a few short haptera, soon covered by a discoid circumferential swelling (rhizogen) of the stipe, which forms downward‐growing hapteroid protuberances that surround the initial holdfast, succeeded by one or more whorls of haptera; all haptera unbranched. Stipe flattened, broadening to form a single lanceolate or expanded and split blade. Hair‐pits on young blades, deciduous with age; costae lacking. Medulla containing long, thick‐walled longitudinal conducting cells (solenocysts) and lateral connecting cells (allelocysts) rather than sieve tubes. Mucilage ducts absent. Sporangia sori discontinuous patches at base of blades, paraphyses lacking a hyaline appendage, zoospores with a red eyespot. Gametophytes of S. polyschides dioecious and sexually dimorphic. Marine, littoral to 19 m, distributed either in the western Mediterranean and the Atlantic from the Tropic of Cancer to southern Norway (Algaebase, 2010).
Biochemical composition
Table 5 ‐ Biochemical composition of Phaeophyceae relevant for biotechnology. Laminaria
hyperborea (lamina + stipe)
Laminaria digitata (lamina)
Saccorhiza polyschides (lamina)
Saccharina latissima
References
Water (%FW) 77‐89 73‐90 90.9‐93.0 Baardseth & Haug, 1953; Indergaard & Minsaas, 1991; Jensen et al., 1985
Ash 16‐37 13.8‐37.6 26.58 Jensen & Haug, 1956; Ruperez, 2002; Adams et al., 2011; Sanchez‐Machado et al., 2004
Total Carbohydrate 61 Kuppers & Kremer, 1978
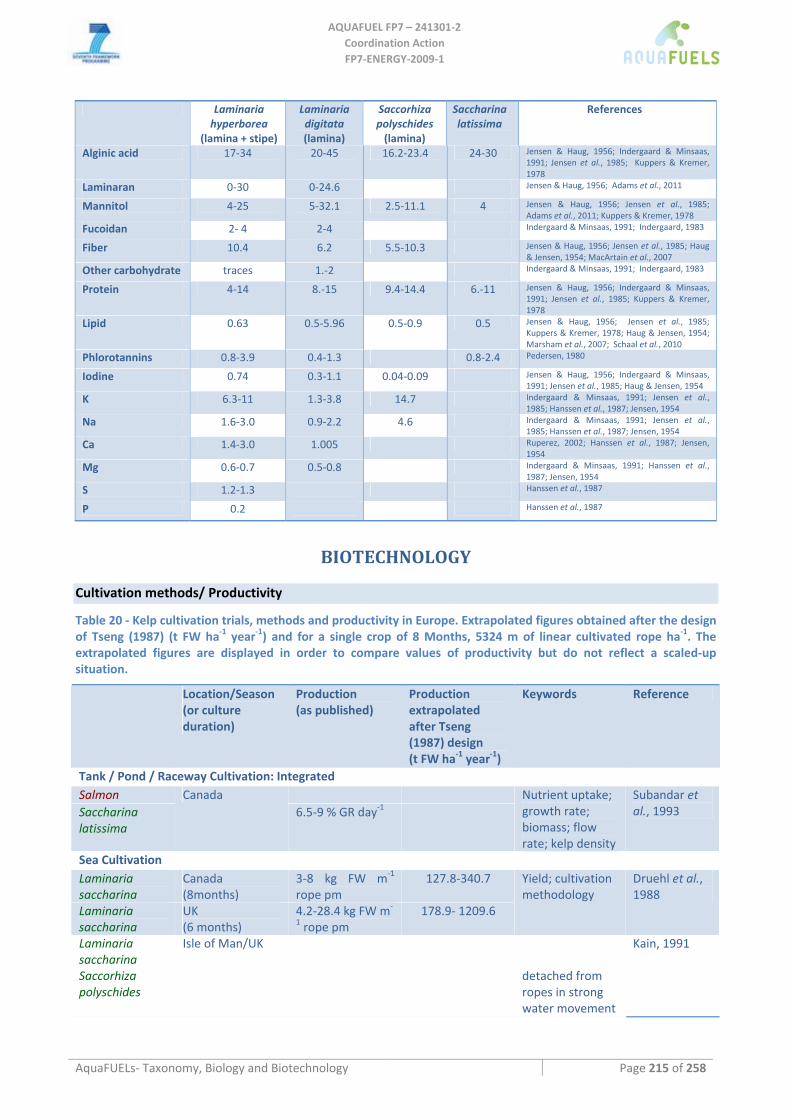
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 215 of 258
BIOTECHNOLOGY
Cultivation methods/ Productivity
Table 20 ‐ Kelp cultivation trials, methods and productivity in Europe. Extrapolated figures obtained after the design of Tseng (1987) (t FW ha‐1 year‐1) and for a single crop of 8 Months, 5324 m of linear cultivated rope ha‐1. The extrapolated figures are displayed in order to compare values of productivity but do not reflect a scaled‐up situation.
Laminaria hyperborea
(lamina + stipe)
Laminaria digitata (lamina)
Saccorhiza polyschides (lamina)
Saccharina latissima
References
Alginic acid 17‐34 20‐45 16.2‐23.4 24‐30 Jensen & Haug, 1956; Indergaard & Minsaas, 1991; Jensen et al., 1985; Kuppers & Kremer, 1978
Laminaran 0‐30 0‐24.6 Jensen & Haug, 1956; Adams et al., 2011
Mannitol 4‐25 5‐32.1 2.5‐11.1 4 Jensen & Haug, 1956; Jensen et al., 1985; Adams et al., 2011; Kuppers & Kremer, 1978
Fucoidan 2‐ 4 2‐4 Indergaard & Minsaas, 1991; Indergaard, 1983
Fiber 10.4 6.2 5.5‐10.3 Jensen & Haug, 1956; Jensen et al., 1985; Haug & Jensen, 1954; MacArtain et al., 2007
Other carbohydrate traces 1.‐2 Indergaard & Minsaas, 1991; Indergaard, 1983
Protein 4‐14 8.‐15 9.4‐14.4 6.‐11 Jensen & Haug, 1956; Indergaard & Minsaas, 1991; Jensen et al., 1985; Kuppers & Kremer, 1978
Lipid 0.63 0.5‐5.96 0.5‐0.9 0.5 Jensen & Haug, 1956; Jensen et al., 1985; Kuppers & Kremer, 1978; Haug & Jensen, 1954; Marsham et al., 2007; Schaal et al., 2010
Phlorotannins 0.8‐3.9 0.4‐1.3 0.8‐2.4 Pedersen, 1980
Iodine 0.74 0.3‐1.1 0.04‐0.09 Jensen & Haug, 1956; Indergaard & Minsaas, 1991; Jensen et al., 1985; Haug & Jensen, 1954
K 6.3‐11 1.3‐3.8 14.7 Indergaard & Minsaas, 1991; Jensen et al., 1985; Hanssen et al., 1987; Jensen, 1954
Na 1.6‐3.0 0.9‐2.2 4.6 Indergaard & Minsaas, 1991; Jensen et al., 1985; Hanssen et al., 1987; Jensen, 1954
Ca 1.4‐3.0 1.005 Ruperez, 2002; Hanssen et al., 1987; Jensen, 1954
Mg 0.6‐0.7 0.5‐0.8 Indergaard & Minsaas, 1991; Hanssen et al., 1987; Jensen, 1954
S 1.2‐1.3 Hanssen et al., 1987
P 0.2 Hanssen et al., 1987
Location/Season (or culture duration)
Production (as published)
Production extrapolated after Tseng (1987) design (t FW ha‐1 year‐1)
Keywords Reference
Tank / Pond / Raceway Cultivation: Integrated Salmon Saccharina latissima
Canada 6.5‐9 % GR day‐1
Nutrient uptake; growth rate; biomass; flow rate; kelp density
Subandar et al., 1993
Sea Cultivation Laminaria saccharina
Canada (8months)
3‐8 kg FW m‐1 rope pm
127.8‐340.7
Laminaria saccharina
UK (6 months)
4.2‐28.4 kg FW m‐
1 rope pm 178.9‐ 1209.6
Yield; cultivation methodology
Druehl et al., 1988
Laminaria saccharina
Saccorhiza polyschides
Isle of Man/UK
detached from ropes in strong water movement
Kain, 1991
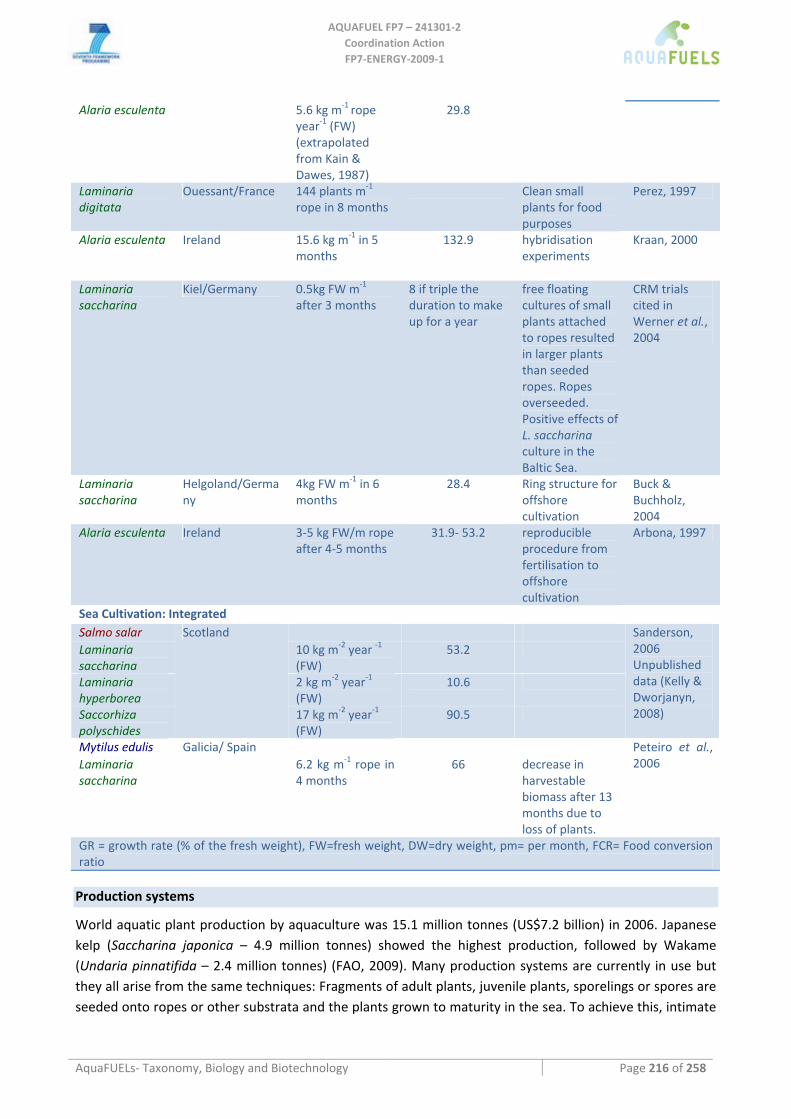
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 216 of 258
Production systems
World aquatic plant production by aquaculture was 15.1 million tonnes (US$7.2 billion) in 2006. Japanese kelp (Saccharina japonica – 4.9 million tonnes) showed the highest production, followed by Wakame (Undaria pinnatifida – 2.4 million tonnes) (FAO, 2009). Many production systems are currently in use but they all arise from the same techniques: Fragments of adult plants, juvenile plants, sporelings or spores are seeded onto ropes or other substrata and the plants grown to maturity in the sea. To achieve this, intimate
Alaria esculenta 5.6 kg m‐1 rope year‐1 (FW) (extrapolated from Kain & Dawes, 1987)
29.8
Laminaria digitata
Ouessant/France 144 plants m‐1 rope in 8 months
Clean small plants for food purposes
Perez, 1997
Alaria esculenta Ireland 15.6 kg m‐1 in 5 months
132.9 hybridisation experiments
Kraan, 2000
Laminaria saccharina
Kiel/Germany 0.5kg FW m‐1 after 3 months
8 if triple the duration to make up for a year
free floating cultures of small plants attached to ropes resulted in larger plants than seeded ropes. Ropes overseeded. Positive effects of L. saccharina culture in the Baltic Sea.
CRM trials cited in Werner et al., 2004
Laminaria saccharina
Helgoland/Germany
4kg FW m‐1 in 6 months
28.4 Ring structure for offshore cultivation
Buck & Buchholz, 2004
Alaria esculenta Ireland 3‐5 kg FW/m rope after 4‐5 months
31.9‐ 53.2 reproducible procedure from fertilisation to offshore cultivation
Arbona, 1997
Sea Cultivation: Integrated Salmo salar Laminaria saccharina
10 kg m‐2 year ‐1 (FW)
53.2
Laminaria hyperborea
2 kg m‐2 year‐1 (FW)
10.6
Saccorhiza polyschides
Scotland
17 kg m‐2 year‐1
(FW) 90.5
Sanderson, 2006 Unpublished data (Kelly & Dworjanyn, 2008)
Mytilus edulis Laminaria saccharina
Galicia/ Spain 6.2 kg m‐1 rope in 4 months
66 decrease in harvestable biomass after 13 months due to loss of plants.
Peteiro et al., 2006
GR = growth rate (% of the fresh weight), FW=fresh weight, DW=dry weight, pm= per month, FCR= Food conversion ratio

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 217 of 258
knowledge of both the biology and life history of the plants is critical. For example, kelps cannot be grown from fragments as there is a high level of specialization and fragments of sporophytes do not regenerate.
• In Asia Culture of sporelings The first step consists in the collection of the zoospores onto the seeding cords. To do this, fronds with mature sporangial sori are subjected to partial drying in the air and then placed in a small container with seawater. The liberated zoospores readily attach themselves to the seeding cords. The gametophytes and early sporophytes are cultured in water of 8–10°C in glasshouse for about three months, by which time the juvenile sporophytes are 2–3 cm in length. Transplantation When the water temperature has dropped to about 20 °C, sporeling cords are removed from the glasshouse and on floating rafts. In a month or so, the sporelings will have grown to juveniles of 10–15 cm or longer. These juvenile sporophytes will be eventually brought to the transplanting room and placed in tanks filled with seawater. Due to their fast growth on the cords they are thinned. Plants are removed from the original sporeling cord and inserted in the twists of the kelp ropes, at a density of about 30 juveniles to each rope of 2 m. Culture methods There are two basic floating‐raft kelp cultivation methods. One is the hanging‐kelp rope (also called single‐raft) cultivation method. The other is the horizontal kelp rope (also called double‐raft) cultivation method. The floating line, is about 60 meters long floated at the surface by buoys generally made of glass or plastics and anchored terminally by anchoring lines to wooden pegs driven into the sea bottom. Each kelp rope has about 30 plants twisted in it and is about 2 m in length. In the single‐raft method, the kelp ropes are hung down from floating line and weighted down by a small piece of stone. In the double‐raft method, the two kelp ropes are linked or tied together at one end and the other ends tied to floating lines. The hanging kelp rope method has the advantage of better water movement but has the defect of uneven growth of kelps. The horizontal kelp rope method gives an even growth of kelp. However, it has the disadvantage of being more resistant to water motion. Generally, the single‐raft method is better adapted to comparatively clearer water regions, and the double‐raft method to turbid regions with lower water transparency, such as the Zhejiang coast (Feijiu, 1988). The use of fertilizer is still in place but now limited to the first months of development, the association with fish/shellfish farming allows to greatly reduce the adjunction of nutrient. Several type of association are applied: Saccharina japonica + Undaria pinnatifida Saccharina japonica + Mytilus sp. Saccharina japonica + Haliotis (Abalone) Saccharina japonica + Undaria pinnatifida + Mytilus sp. Saccharina japonica + Undaria pinnatifida + Haliotis Saccharina japonica + Undaria pinnatifida + Haliotis + Mytilus sp. The productivity increases by 45% and the income rises by 15 to 20% when compared to monoculture (Perez, 1997).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 218 of 258
• In Europe Production systems have been investigated for research purposes in various parts of Europe since the late 1970’s. The technique of “free‐living” culture of the microscopic gametophytes has been developed in Ifremer in 1984. This technique allows the farmer to seed the ropes on demand, and after a shortened pre‐cultivation period in the hatchery, the ropes with young sporelings can be transferred at sea. The design of hollow cylindrical collectors have also simplified and hence reduced the time needed to set the culture at sea. Production at a commercial scale is set up for Undaria pinnatifida in France where it peaked at 100t DW in 1995 (FAO, 2002), and reaches a saturated market for food purposes.
In Ireland, cultivation trials have used hybridization of North‐Atlantic strains of Alaria esculenta in order to improve productivity as well as protein content to target the food and feed market (Kraan et al., 2000) Trials with rings structures compared to longlines have also taken place to propose a design for offshore seaweed aquaculture (Buck and Buchholz, 2004).
The use of polyculture (Integrated Multi‐Trophic Aquaculture), has been performed for some time, however recent research has focused on the use of seaweed as biofilters to take advantage of nutrient load from fish farming.
Harvesting methods
• In Asia
Harvesting takes place when the fronds are mature. The time for harvesting is important to kelp farmers. Since Saccharina is sold on the market on the basis of dry weight, and since the wet weight to dry weight ratio changes from month to month, the criterion for selecting harvest time must take into consideration the highest per‐unit area production rate plus the lowest wet‐to‐dry ratio.
Harvesting is done manually, the kelp ropes are detached from the floating line, and collected in small boats, many of which are towed in a long line by a motor boat to the shores.
• In Europe
Manual harvesting of longlines of kelp is not economically viable in a highly competitive area due to the high cost of the workforce in Western Europe. Therefore mechanization and minor adaptation of mussels harvesting boats to kelps longline is necessary.
Biomass processing
• In Asia
When the boats reach the wharf or shore, the plants are transported to land and dried under the sun. Formerly the kelp was sold on the market only in its crude dried form, but recently small package of shredded and seasoned forms with different flavours have been introduced to the market and have been very well reveived by the Chinese people.
• In Europe
In some cases, the crop was used for the food market. Hence the crop was dried, sometimes flavoured and sold as a snack or sea‐vegetables. (Laminaria saccharina and Alaria esculenta in the Island of Man, UK, Undaria pinnatifida in Brittany, France) (Werner et al, 2004).
Biomass resulting from cultivation trials for research purposes has mostly lead to knowledge, skills and data production on seaweed aquaculture. However, some European projects currently uses the biomass produced for anaerobic digestion.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 219 of 258
Scaling up limitations
There are no serious technological barriers (Kain, 1991) to scaling up seaweed aquaculture, as it has already been demonstrated spectacularly in Asia. However, limitations in a European context rather lie in the selection of adequate sites (both regarding the requirement of the targeted seaweed species and the competition with other coastal resource users), the development of mechanical harvesting techniques and assessment of the environmental impact of large‐scale aquaculture. Fei (2004) advocates for large‐scale seaweed cultivation as a way of solving eutrophication under the following conditions: (a) Large‐scale cultivation could be conducted within the region experiencing eutrophication; (b) Fundamental scientific and technological problems for cultivation should have been solved; (c) Cultivation should not impose any harmful ecological effects; (d) Cultivation must be economically feasible and profitable.
References
Adams J.M.M., Ross A.B., Anastasakis K., Hodgson E.M., Gallagher J.A., Jones J.M., Donnison I.S. (2011) Seasonal variation in the chemical composition of the bioenergy feedstock Laminaria digitata for thermochemical conversion. Bioresource Technology 102: 226‐234. (Special Issue: Biofuels ‐ II: Algal Biofuels and Microbial Fuel Cells)
Arbona J.F. (1997) A routine method for mass cultivation of Alaria esculenta (Greville 1830). M. Sc. Aquaculture, National University of Ireland, Galway.
Baardseth E., Haug, A. (1953) Individual variation of some constituents in brown algae, and reliability of analytical results. Reports of the Norwegian Institute of Seaweed Research 2: 1‐23.
Braune W. (2008) Meeresalgen. Ein Farbbildführer zu den verbreiteten benthischen Grün‐ Braun‐ und Rotalgen der Weltmeere. A.R.G. Gantner Verlag, Ruggell, pp. 1‐596, 266 pls.
Buck B.H., Buchholz C.M. (2004) The offshore‐ring: A new system design for the open ocean aquaculture of macroalgae. Journal of Applied Phycology 16: 355‐368.
Connan S., Delisle F., Deslandes E., Gall E.A. (2006) Intra‐thallus phlorotannin content and antioxidant activity in Phaeophyceae of temperate waters. Botanica Marina 49: 39‐46.
Dieck I.T., Oliveira Filho E.C. de (1993) The section Digitatae of the genus Laminaria (Phaeophyta) in the northern and southern Atlantic: crossing experiments and temperature response. Marine Biology 115: 151‐160.
Druehl L.D., Baird R., Lindwall A., Lloyd K.E., Pakula S. (1988) Longline cultivation of some Laminariaceae in British Columbia, Canada. Aquaculture and Fish Management 19: 253–263.
FAO (2009) The State of World Fisheries and Aquaculture, FAO Fisheries and Aquaculture Department, Food And Agriculture Organization of the United Nations. Rome. http://www.fao.org/docrep/011/i0250e/i0250e00.htm
Fei, X. (2004) Solving the coastal eutrophication problem by large scale seaweed cultivation. Hydrobiologia 512: 145‐151. Guiry M.D., Guiry G.M. (2010) AlgaeBase. World‐wide electronic publication, National University of Ireland, Galway.
http://www.algaebase.org; searched on 31 March 2010. Guiry M.D. (2010) Seaweed.ie. World‐wide electronic publication, National University of Ireland, Galway. http://www.seaweed.ie;
searched on 19 November 2010. Hanssen J.F., Indergaard M., Ÿstgaard K., Bêvre O.A., Pedersen T.A., Jensen A. (1987) Anaerobic digestion of Laminaria spp. and
Ascophyllum nodosum and application of end products. Biomass 14: 1‐13. Haug A., Jensen A. (1954) Seasonal variations on the chemical composition of Alaria esculenta, Laminaria saccharina, Laminaria
hyperborea and Laminaria digitata from Northern Norway. Reports of the Norwegian Institute of Seaweed Research 4. Indergaard M., Minsaas J. (1991) Animal and human nutrition. In: Guiry M.D., Blunden G. (eds.) Seaweed Resources in Europe: Uses
and Potential, John Wiley & Sons, Chichester, pp. 21‐64. Jensen A. (1954) Om tang og tare i husdyrf6ret. Medlemsblad for den Norske Veterinaer‐ forening 6: 1‐23. Jensen A., Haug A. (1956) Geographical and seasonal variation in the chemical composition of Laminaria hyperborea and Laminaria
digitata from the Norwegian coast. . Reports of the Norwegian Institute of Seaweed Research 14: 1‐18. Jensen A., Indergaard M., Holt T.J. (1985) Seasonal variation in the chemical composition of Saccorhiza polyschides (Laminariales,
Phaeophyceae). Botanica Marina 28: 375‐381. Kain (Jones) J.M. (1971) Synopsis of biological data on Laminaria hyperborea. FAO Fisheries Synopsis No 87(Rev. 1): 74 pp.. Kain J.M. (1991) Cultivation of attached seaweeds. In: Guiry M.D., Blunden G. (eds.) Seaweed Resources in Europe: Uses and
Potential, John Wiley & Sons, Chichester, pp. 309‐377. Kelly M., Dworjanyn S. (2008) The Potential of Marine Biomass for Anaerobic Biogas Production, The Crown Estate, London. Kraan S., Tramullas A.V., Guiry M.D. (2000) The edible brown seaweed Alaria esculenta (Phaeophyceae, Laminariales):
hybridization, growth and genetic comparisons of six Irish populations. Journal of Applied Phycology 12: 577‐583. Kraan S (2000) The genus Alaria (Alariaceae, Phaeophyceae) explored: phylogenetics, hybridization and aquaculture. PhD thesis.
National University of Ireland, Galway. Kuppers U., Kremer B.P. (1978) Longitudinal profiles of carbon dioxide fixation capacities in marine macroalgae. Plant Physiology
62: 49‐53 Lane C.E., Saunders,G.W. (2005) Molecular investigation reveals epi/endophytic extrageneric kelp (Laminariales, Phaeophyceae)
gametophytes colonizing Lessoniopsis littoralis thalli. Botanica Marina 48: 426‐436.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 220 of 258
Lane C.E., Mayes C., Druehl L.D., Saunders G.W. (2006) A multi‐gene molecular investigation of the kelp (Laminariales, Phaeophyceae) supports substantial taxonomic re‐organization. Journal of Phycology 42: 493‐512.
Loiseaux‐de Goër S., Noailles M.C. (2008) Algues de Roscoff, Editions de la Station Biologique de Roscoff, Roscoff, pp. 1‐215. MacArtain P., Gill C.I.R., Brooks M., Campbell R., Rowland I.R. (2007) Nutritional value of edible seaweeds. Nutrition Reviews 65:
535‐543. Marsham S., Scott G.W., Tobin M.L. (2007) Comparison of nutritive chemistry of a range of temperate seaweeds. Food Chemistry
100 :1331–1336 McHugh D.J. (2003) A guide to the seaweed industry. FAO Fisheries Technical Paper 441 Morrissey J., Kraan S., Guiry M.D. (2001) A guide to commercially important seaweeds on the Irish Coast,. Bord Iascaigh Mhara, Dun
Laoghaire. Norton T.A. (1970) Synopsis of biological data on Sacchoriza polyschides. FAO Fisheries Synopsis 83: 11‐93. Papenfuss G.F. (1950) Review of the genera of algae described by Stackhouse. Hydrobiologia 2: 181‐208. Pedersen A. (1980) Fenolinnhold i brunalger (Phaeophyceae) som funksjon av vekstype og salinitet. Ph. D. thesis. Inst. Marinbiol.,
Univ. Bergen. Perez R. (1997) Ces algues qui nous entourent: conception actuelle, rôle dans la biosphère, utilisations, culture, Edition IFREMER
Plouzané, France. Peteiro C., Salinas J.M., Freire Ó., Fuertes C. (2006) Cultivation of the autoctonous seaweed Laminaria saccharina off the galician
coast (NW): production and features of the sporophytes for an annual and biennial harvest. Thalassas 22: 45‐52. Roeder V., Collén J., Rousvoal S., Corre E., Leblanc C., Boyen C. (2005) Identification of stress gene transcripts in Laminaria digitata
(Phaeophyceae) protoplast cultures by expressed sequence tag analysis. Journal of Phycology 41: 1227‐1235. Ruperez P. (2002) Mineral content of edible marine seaweeds. Food Chemistry 79: 23‐26. Indergaard M. (1983) The aquatic resource. I. The wild marine plants: a global bioresource. In: Cote W.A. (ed.) Biomass utilization,
Plenum Publishing Corporation, pp. 137‐168. Sánchez I., Fernández C., Arrontes J. (2005) Long‐term changes in the structure of intertidal assemblages following invasion by
Sargassum muticum (Phaeophyta). Journal of Phycology 41: 942‐949. Sanchez‐Machado D.I., Lopez‐Cervantes J., Lopez‐Hernandez J., Paseiro‐Losada P. (2004) Fatty acids, total lipid, protein and ash
contents of processed edible seaweeds. Food Chemistry 85: 439‐444. Schaal G., Riera P., Leroux C. (2010) Trophic ecology in a Northern Brittany (Batz Island, France) kelp (Laminaria digitata) forest, as
investigated through stable isotopes and chemical assays. Journal of Sea Research 63: 24‐35. Subandar A., Petrell R.J., Harrison P.J. (1993) Laminaria culture for reduction of dissolved inorganic nitrogen in salmon farm
effluent. Journal of Applied Phycology 5: 455‐463. Feijiu W. (1988) Marine phytoculture in China. http://www.fao.org/docrep/field/003/AB719E/AB719E02.htm. Werner A., Clarke C., Kraan S. (2004) Strategic review of the feasibility of seaweed aquaculture in Ireland. NDP Marine RTDI Desk
Study Series, DK/01/008.2004.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 221 of 258
10 Other aquatic biomass
10.1 Egeria densa
Figure 91 ‐ Egeria densa Planch.
© Jeff Schardt of the Florida Fish and Wildlife Conservation Commission
SYMBOLS: B
TAXONOMY
Phylum Magnoliophyta Class Liliopsida Order Alismatales Family Hydrocharitaceae Genus Egeria Species Egeria densa
BIOLOGY
Egeria densa is a submersed plant, like a large Elodea (up to three times the size of Elodea). Egeria grows either rooted in the bottom or free‐floating. Stems are up to 2 m or more long and 3 mm thick and may be highly branched. Wherever branches occur, two nodes (stem joints) are compacted closely together, giving the impression that the node is doubled. Fine, unbranched white roots form at the double nodes, and only fragments containing this double node can grow into new plants. Leaves, like those of Elodea, are in whorls, but typically whorls of 4 or 5, along stems. The leaves are not strongly recurved and are 10‐30 mm long, and up to 4mm wide; 0.5 mm below the leaf tip the width is 0.5‐1 mm. Flowers are about 2 cm. across and have 3 petals. They occur on a short stalk about an inch above the water and are produced primarily in the spring through early summer, but occasionally appear later in the growing season. This plant only flowers in warmed water (native areas). Elsewhere reproduction is vegetative.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 222 of 258
It grows in ponds, reservoirs, gravel pits, ditches, swamps and canals. It is most common in eutrophic water, and will persist in dried‐up water bodies. Occasionally it is found on completely dry ground. It is known to have some frost tolerance.
Egeria is native to South America. It was introduced by the aquarium trade and currently is the top‐selling aquatic plant for use in aquaria as "oxygenators". It has been sold under several names including "oxygenating plant", elodea, and Anacharis.
Egeria densa has 22% ash, 20% protein, 29% cellulose.
BIOTECHNOLOGY
Egeria densa has been tested for removal of nutrients and heavy metals from waters and as raw material to produce cheap and good quality organic fertilizers.
References
Caro Lara I., Romero Otalora Z., Lora Silva R. (2009) Production of organic fertilizers with elodea (Egeria densa) present on the Fuquene lagoon. Revista U.D.C.A Actualidad & Divulgación Científica 12: 91‐100. [BIOTECHNOLOGY section]
Champion P.D., Tanner C.C. (2000) Seasonality of macrophytes and interaction with flow in a New Zealand lowland stream. Hydrobiologia 441: 1‐12. [BIOLOGY section]
Dadds N., Bell S. (n.a.) Invasive non‐native plants associated with fresh waters. A Guide to their identification. Prepared on behalf of Plantlife, Royal Botanic Garden Edinburgh, Scottish Natural Heritage, Scottish Environment Protection Agency, Scottish Water. [BIOLOGY section]
Módenes A.N., Pietrobelli J.M.T. de A., Espinoza‐Quiñones F.R. (2009) Cadmium biosorption by non‐living aquatic macrophytes Egeria densa. Water Science and Technology 60: 293‐300. [BIOTECHNOLOGY section]
Feijoó C., García M.E., Momo F., Toja J. (2002) Nutrient absorption by the submerged macrophyte Egeria densa Planch.: effect of ammonium and phosphorus availability in the water column on growth and nutrient uptake. Lirnnetica 21: 93‐104. [BIOTECHNOLOGY section]
FBIS Freshwater Biodata Information System New Zealand (2005) [BIOLOGY section]: https://secure.niwa.co.nz/fbis/validate.do?search=common [Accessed 5 August 2005]
Florence J., Chevillotte H., Ollier C., Meyer J.Y. (2007) Egeria densa. Base de données botaniques Nadeaud de l'Herbier de la Polynésie française (PAP). Available from: http://www.herbier‐tahiti.pf/Selection_Taxonomie.php?id_tax=5052 [Accessed 26 March 2008] [BIOLOGY section]
Getsinger K.D., Dillon C.R. (1984) Quiescence, growth and senescence of Egeria densa in Lake Marion. Aquatic Botany 20: 329‐338. [BIOLOGY section]
Haramotoa T., Ikusima I. (1988) Life cycle of Egeria densa Planch., an aquatic plant naturalized in Japan. Aquatic Botany 30: 389‐403. [BIOLOGY section]
ITIS (Integrated Taxonomic Information System) (2002) Online Database. Egeria densa. http://www.itis.gov/servlet/SingleRpt/SingleRpt?search_topic=TSN&search_value=38972 [Accessed March 2005] [BIOLOGY section]
Kay S., Hoyle S.(1999) Aquatic weed fact sheet. Brazilian elodea or Egeria, Egeria densa. NC State University, College of Agriculture and Life Science, Crop Sceince Department. [BIOLOGY section]
Little E.C.S. (1979) Handbook of utilization of aquatic plants. FAO Fisheries Technical Paper No. 187, Rome. [BIOLOGY section] Macdonald I.A.W., Reaser J.K., Bright C., Neville L.E., Howard G.W., Murphy S.J., Preston G. (eds.) (2003) Invasive alien species in
southern Africa: national reports & directory of resources. Global Invasive Species Programme, Cape Town, South Africa. [BIOLOGY section]
Roy B., Popay I., Champion P., James T., Rahman A. (2004) An Illustrated Guide to Common Weeds of New Zealand, Egeria densa, 2nd Edition, New Zealand Plant Protection Society. [BIOLOGY section]
USDA‐GRIN (Germplasm Resources Information Network) (2003) Egeria densa. National Genetic Resources Program [Online Database] National Germplasm Resources Laboratory, Beltsville, Maryland. [BIOLOGY section]
USDA‐NRCS (Natural Resource Conservation Service) (2002) Egeria densa. The PLANTS Database Version 3.5 [Online Database] National Plant Data Center, Baton Rouge, LA. [BIOLOGY section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 223 of 258
10.2 Eichhornia crassipes
Figure 92 ‐ Eichhornia crassipes (Mart.) Solms (1803).
Illustration by Ohze
SYMBOLS: B, E
TAXONOMY
Phylum Magnoliophyta Class Liliopsida Order Commelinales/Liliales Family Pontederiaceae Genus Eichhornia Species Eichhornia crassipes
Related species
E. azurea, E. diversifolia, E. heterosperma, E. paniculata, E. paradoxa
BIOLOGY
Eichhornia crassipes is a perennial aquatic herb; rhizome and stems are normally floating, with rooting at the nodes, with long black pendant roots. Leaves are usually with inflated spongy petioles, the leaf blades are circular to reniform, 4‐12 cm wide. Inflorescence is a contracted panicle, 4‐15 cm long, with several flowers; perianth is lilac, bluish‐ purple, or white, the upper lobe bearing a violet blotch with a yellow center. Stamens are 6; stalk of the inflorescence soon becomes goose‐necked, forcing the dead flowers under the water; capsule is dehiscent, surrounded by the perianth, membraneous, many‐seeded. In general, seed production in temperate populations was found to be half that of tropical populations, mainly due to differences in levels of insect visitation. Seeds are 11–14,‐winged, 1.1–2.1 × 0.6–0.9 mm. The diploid number of chrmomosome is 32.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 224 of 258
Water hyacinth is reported to tolerate annual precipitation of 8.2 to 27.0 dm (mean of 8 cases = 15.8), annual temperature of 21.1 to 27.2°C (mean of 5 cases = 24.9), and estimated pH of 5.0 to 7.5. Leaves are killed by frost, and plants cannot tolerate water temperatures >34°C. Water hyacinth is reported to tolerate grazing. Native to Brazil, now growing in most tropical and subtropical countries. More than 50 countries in which water hyacinth is weed have been listed. It grows in ponds, ditches, canals, calm waters of rivers at an elevation of 0‐300 m and a depth range of 2 m. Engineers have estimated that the Panama Canal would be impassable within three years without continuous aquatic weed control measures.
Fresh plant contains 95.5% moisture. On a dry matter basis, it is 75.8% organic matter, 1.5% N, and 24.2% ash. The ash contains 28.7% K2O, 1.8% Na2O, 12.8% CaO, 21.0% Cl, and 7.0% P2O5. Protein is bout 12‐18% of dry matter. Protein contains, per 100 g, 0.72 g methionine, 4.72 g phenylalanine, 4.32 g threonine, 5.34 g lysine, 4.32 g isoleucine, 0.27 g valine, and 7.2 g leucine. 23‐28% of dry weight is cellulose.
On a dry matter basis, standing crop is 12.8 t ha‐1, maximum productivity 14.6 g m‐2 day‐1, maximum yield 54.7 g m‐2 day‐1.
BIOTECHNOLOGY
Water hyacinth would be ideally suited for nutrient removal systems. Aquatic plants, when growing in water containing ample quantities of N, P and K, will exploit the situation by “luxury consumption” of these elements, far in excess of what they need for healthy growth. As it floats on the surface and is not rooted, harvesting is facilitated. By continuous harvesting the population could be kept in a rapidly expanding phase during which uptake rates of nutrients are at their highest. Waters beneath dense stands are anaerobic so additional N would be lost by denitrification. There would be considerable microbial activity beneath the hyacinths and nutrients would be absorbed by these organisms. However, considerable organic matter would reach the water by the loss of root fragments that probably have a fairly high biological oxygen demand (BOD) and it might prove necessary to use conventional sewage holding ponds to reduce the BOD prior to final release. Water hyacinth roots naturally absorb pollutants, including such toxic chemicals as lead, mercury, and strontium 90, as well as some organic pollutant, in concentrations 10,000 times that in the surrounding water.
Water hyacinths have been used as human food in the Philippines in war time. The soft white bud of the plant was eaten either raw, or as a salad, or as an ingredient in vegetable dishes. It was called ‘repollo’ (cauliflower). Under conditions of relative food abundance it is considered unlikely that it would still be used as food.
Dried and cleansed plants, can be used as fertilizer, poultry feed, additives to cattle‐feed, and plant mulch. Water hyacinth protein shows deficient levels in only two of the essential amino acids, valine and methionine, as compared to the FAO reference pattern. A diet containing an adequate protein level will be balanced in lysine if it contains 4.2% of that amino acid in its protein content. Corn is deficient in lysine, containing only 0.8%. However water hyacinth contains 5.3% lysine, while milk contains 7.8%. It is evident that water hyacinth could serve to improve the lysine content of a corn diet. Leaf protein which contains 6.3% lysine has been reported to be an effective supplement for barley in pig rations. As a feed for ruminants, water hyacinth should be administered as a combination of the lamina , where the protein and fibre mainly are, and the petiole where the carbohydrate are mainly located. Animals eating fresh water hyacinth as sole ingredient of the diet could not ingest sufficient dry matter, quite apart from the imbalance of the nutrients. On a dry matter basis water hyacinth is better than straw but a little low in protein to compare with hay. The physical structure of the plant is not suitable for hay or silage making and the product would not have much nutritional value. The nutritional value could be increased by mixing with molasses. The hyacinth is rich in minerals, but it would be simple to incorporate any necessary food

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 225 of 258
additives in the processing. Pig farming using water hyacinth grown in ponds fertilized with human and animal waste is well established in Malaysia. The hyacinth can be fed fresh after removal of the roots. Some people boil the plant. Pigs readily eat the hyacinth and thrive on it. Some farmers have machines for slicing the plants into small pieces. The high vitamin content (A, B and C) and easy availability of the plant are important advantages. But the high moisture content and the possibility of the material being contaminated with pathogens seem to deter the extensive practice of fresh plant feeding. Consequently most farmers (70–80%) prefer to cook the sliced hyacinth with other feed ingredients for 5 to 6 hours and feed the boiled material. Silage is also fed to the pigs. Good compost can be made from hyacinth.
Attempts have been made to utilize the plant as a raw material for paper, plastics and other commercial products, but so far no industry based on water hyacinth appears to have been established. The fibrous stem, constituting about 40% of the whole plant, is suitable for paper making. The addition of jute or cotton fibres to the extent of 8–10% on the weight of the pulp is considered necessary as the paper prepared from the stems alone is translucent. A plastic material suitable for the production of moulded articles and boards has been prepared from water hyacinth. In Asia, the plants are collected at the beginning of the cold weather, left to dry and the dry material used along with jute sticks as fuel. The ashes are subsequently used as manure. The possibility of using the dried weed for the production of power gas and power alcohol has been considered. Three methods have been suggested, viz. saccharification by acid digestion and subsequent fermentation, gasification by air and steam with recovery of ammonia, and bacterial fermentation and utilization of the evolved gas for power production. Potassium chloride (0.1 ton of KCl per ton of dry hyacinth) is recovered in all the processes. Starting from 1 ton of dried water hyacinth, 59 L of ethyl alcohol and 0.2 tons of residual fibre (8.1 MJ) are obtained by the first process. Gasification by air and steam gives, per ton of dried material, 37‐ 53 kg of ammonium sulphate and 1133 m3 of gas (0.16 MJ) containing hydrogen, 16.6%; methane, 4.8%; carbon monoxide, 21.7%; carbon dioxide, 4.1%; and nitrogen, 52.8%. Bacterial fermentation gives per ton of material 750 m3 of gas (0.63 MJ) containing: methane, 51.6%; hydrogen, 25.4%; carbon dioxide, 22.1%; and oxygen, 1.2%. The commercial possibilities of the processes have not been proved.
References
Anonymous (1976) Water hyacinths soak up pollution. BioScience 26: 234. [BIOTECHNOLOGY section] Bailey L.H. (1949) Manual of Cultivated Plants. Macmillan, New York. Duke J. Handbook of energy crops. http://www.hort.purdue.edu/newcrop/duke_energy/dukeindex.html [BIOTECHNOLOGY
section] Gohl B. (1981) Tropical feeds. Feed information summaries and nutritive values. FAO Animal Production and Health Series 12. FAO,
Rome. [BIOLOGY section] Little E.C.S. (1979) Handbook of utilization of aquatic plants. FAO Fisheries Technical Paper No. 187, Rome. [BIOTECHNOLOGY
section] Hammer R.L. (1996) Eichhornia crassipes. In Randall J.M., Marinelli J. (eds.) Invasive Plants: Weeds of the Global Garden. Brooklyn
Botanic Garden Inc., New York, p. 99. [BIOLOGY section] Holm L.G., Plunknett D.L., Pancho J.V., Herberger J.P. (1977) The world's worst weeds. University Press of Hawaii, Honolulu.
[BIOLOGY section] Holm L.G., Pancho J.V., Herberger J.P., Plucknett, D.L. (1979) A geographical atlas of world weeds. John Wiley & Sons, New York.
[BIOLOGY section] Integrated Taxonomic Information System (ITIS) catalogue of life 2010 http://www.catalogueoflife.org/annual‐
checklist/2010/search/all/key/eichhornia/match/1 (accessed on the 10th of June 2010) [BIOLOGY section] Matai S., Bagchi D.K. (1980) Water hyacinth: a plant with prolific bioproductivity and photosynthesis. In: Gnanam A., Krishnaswamy
S., Kahn J.S. (eds.) Proceedings of the International Symposium on Biological Applications of Solar Energy. MacMillan Co. of India, Madras, pp. 144‐148. [BIOLOGY section]
Penfound W.T., Earle T.T. (1948) The biology of the water hyacinth. Ecological Monographs 18: 449‐472. [BIOLOGY section] Reed C.F. (1970) Selected weeds of the United States. Agriculture Handbook 366. USDA, Washington, DC. [BIOLOGY section] USDA, NRCS( 2001) The PLANTS Database, Version 3.1. (http://plants.usda.gov). National Plant Data Center, Baton Rouge, LA
70874‐4490 USA.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 226 of 258
10.3 Elodea canadensis
Figure 93 ‐ Elodea canadensis Michaux (1803).
Illustration by C. A. M. Lindman
SYMBOLS: B
TAXONOMY
Phylum Magnoliophyta Class Liliopsida Order Hydrocharitales Family Hydrocharitaceae Genus Elodea Species Elodea canadensis
Related species
E. bifoliata, E. callitrichoides, E. granatensis, E. nuttallii, E. potamogeton, E. spinosa.
BIOLOGY
Elodea canadensis is a submerged aquatic perennial freshwater herb, usually firmly rooted to the bottom mud and producing a thick green mat below the water surface. Short thread like stolons giving rise to slender vertical stems to 3 m long. Leaves are dark green oblong‐linear, formed at intervals of 3‐25 mm along the stem, in groups of three, each 6‐12 mm long and about 1‐5 mm wide, usually with forward pointing teeth on the margins. Roots are filamentous, rising from the nodes along the stolons. Flowers are white or pale purple with three sepals and petals. They are solitary, forming in the axils of the leaves and growing towards the surface on threadlike stalks about 30 cm long.. Fruits are capsules less than 1 cm in length. It is dispersed by seeds and fragments via water currents. It is a dioecious plant flowering from June to August. Pollination occurs near the water surface and pollen is distributed by wind and water currents. Vegetative reproduction by fragments is very common. Mass development has been reported multiple

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 227 of 258
times in the last century. It is preyed upon by a high number of freshwater organisms including fish and birds.
It is diffuse in surface standing waters, and surface running waters, like shallow lakes, ponds, pools, ditches and streams with slow moving water. It grows up to 3 m water depth and, in exceptional cases, up to 16 m in depth. It tolerates pH values from 6.0 to 7.5 and temperatures from 1 to 25 °C.
It originates from North American inland waters. The first European record was reported from Ireland in 1836. It subsequently became widespread in north and central European countries. After a rapid colonization of northern Europe the populations declined due to the introduction of Elodea nuttallii. Today the population is stable. As it can be very dominant, it competes for nutrients and space with other plants. It can bioaccumulate nutrients and modify the habitat by reducing water movement. The species is known to outcompete other plants. During dense blooms, impairs boating, fishing, swimming, and water skiing. Clogging of water intake pipes of power plants and other industries were reported.
Elodea is anchored to the bottom deposits of a lake or river by adventitious roots, but nutrients are probably absorbed mainly by leaves and stems in contact with the free water. Although the concentration of dissolved salts in the surrounding water will largely control the density and composition of the plant, the evidence indicated a difference in composition between species, even in the same lake, as well as seasonal changes. It follows that if this water weed is to be exploited commercially, preliminary plant analyses are needed to determine the best species and season for harvesting.
Crude protein is about 12%, ash 28% and fiber 16%.
BIOTECHNOLOGY
To obtain 1 ton of dry Elodea canadensis 14 tons of wet material would have to be processed. E. canadensis is an excellent food for cattle and pigs, when fed to sheep it was found to be unpalatable but it was accepted when mixed with pasture (1:5 dry matter basis). It appears to contain all the vitamins in at least as high an amount as alfalfa. Of more interest is the fact that the biological value of the protein in Elodea is about 70% that of alfalfa and the digestibility is apparently better. Cystine seems to be the limiting amino acid in Elodea as well as in alfalfa. E. canadensis can be used as a non‐expensive supplemental food in order to increase growth and survival in summerling noble crayfish, A. astacus, that has the potential to consume this macrophyte in nature.
It shows the typical productivity of submerged macrophytes in temperate zones as 2–7 tons dry weight ha‐1 year‐1 and 17–59 tons fresh weight ha‐1 year‐1.
References
Bowmer H., Kathleen S.W., Jacobs L., Sainty G.R. (1995) Identification, biology and management of Elodea canadensis, Hydrocharitaceae. Journal of Aquatic Plant Management 33: 13‐19. [BIOLOGY section]
Champion P.D., Hofstra D.E., Clayton J.S. (2007) Border control for potential aquatic weeds. Stage 3. Weed risk management. Science & Technical Publishing New Zealand Department of Conservation. [BIOLOGY section]
D’Agaro E., Renai B., Gherardi F.(2004) Evaluation of the American waterweed (Elodea canadensis Michx.) as supplemental food for the noble crayfish, Astacus astacus. Bulletin Français de la Pêche et de la Pisciculture 372‐373: 439‐445. [BIOTECHNOLOGY section]
Department of Agriculture and Food, Australia: http://agspsrv95.agric.wa.gov.au/dps/version02/01_plantview.asp? [BIOLOGY section]
Gollasch S. (2006) Elodea canadensis. DAISIE‐Delivering Alien Invasive Species Inventories for Europe. http://www.europe‐aliens.org/pdf/Elodea_canadensis.pdf (accessed on the 10th of June 2010) [BIOLOGY section]
Integrated Taxonomic Information System (ITIS) catalogue of life 2010 http://www.catalogueoflife.org/annual‐checklist/2010/search/all/key/elodea/match/1 (accessed on the 10th of June 2010) [BIOLOGY section]
Kews World Checklist of Selected Plant Families http://apps.kew.org/ (accessed on the 10th of June 2010) [BIOLOGY section] Little E.C.S. (1979) Handbook of utilization of aquatic plants. FAO Fisheries Technical Paper No. 187, Rome. [BIOTECHNOLOGY
section]

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 228 of 258
10.4 Lagarosiphon major
Figure 93 ‐ Lagarosiphon major (Ridley) Moss
© Control of aquatic invasive species in Ireland (CAISIE Life+ project)
SYMBOLS: B
TAXONOMY
Phylum Magnoliophyta Class Liliopsida Order Alismatales Family Hydrocharitaceae Genus Lagarosiphon Species Lagarosiphon major
Related species
L. cordofanus, L. hydrilloides, L. ilicifolius, L. madagascariensis, L. muscoides, L. rubellus, L. steudneri, L. verticillifolius.
BIOLOGY
L. major is a rhizomatous, perennial, submerged aquatic plant. It reaches its maximum growth in clear water up to a depth of 6.5 m, but may only grow to 1 m in murky water. It has numerous threadlike roots, which are adventitious and, along with rhizomes, anchor it to the bottom. Stems, which can reach the surface, are brittle and sparsely branched, 3‐5 mm in diameter and curved towards the base (J‐shaped). The leaves are 5‐20 mm long and 2‐3 mm wide, and occur in alternate spirals along the stem. They generally have tapered tips curving downwards towards the stem, except in low alkalinity water where they are straight. The three‐petalled female flowers are very small, clear‐white on the surface, and grow on very thin white to almost translucent filament‐like stalks. Neither the male flower, which floats freely to the

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 229 of 258
surface, nor fruit or seeds have been recorded outside of its native range. Since the species is dioecious (sexes on different plants) both must be present for sexual reproduction. Only female plants are known outside of the native range of this species. All reproduction in introduced regions is therefore asexual primarily by fragmentation or local growth by rhizomatous spread.
L. major grows best in clear, still or slow‐moving fresh water with silty or sandy bottoms. It prefers the cooler waters of the temperate zone, with optimum temperatures of 20‐23 °C and a maximum temperature of around 25 °C. It can live in high and low nutrient levels and grows best under conditions of high light intensity. It also tolerates relatively high pH (i.e. alkaline conditions). Growth of L. major is greatest in sheltered areas protected from wind, waves and currents.
It is native of southern Africa, is found in high mountain streams and ponds. It has spread throughout the world as an aquarium plant and is also known as an 'oxygen plant'. However, dense infestations can actually consume more oxygen than they produce, and reduce water quality and available oxygen.
BIOTECHNOLOGY
L. major and other aquatic species grown in small outdoor tanks can be used successfully to assess the effects of crop‐protection products on non‐target aquatic flora. The possible utilization by harvesting for stock feed in New Zealand lakes was evaluated; however, the use of the plants as fodder was thought to be unsuitable because of the content of arsenic accumulated by the plants from the thermal waters that enter the lakes.
References
Airo S., Sconfietti R. (1995) In situ experiments on productivity of aquatic macrophytes in a pond. Rivista‐di‐Idrobiologia 34: 147‐156.
CAISIE life+project (2010) control of aquatic invasive species in Ireland. www.caisie.ie. Accessed on the 06th of October 2010. Champion P.D., Tanner C.C. (2000) Seasonality of macrophytes and interaction with flow in a New Zealand lowland stream.
Hydrobiologia 441: 1‐12. Coffey B.T., Clayton J.S. (1987) Submerged macrophytes of Lake Pupuke Takapuna New Zealand. New‐Zealand Journal of Marine
and Freshwater Research 21: 193‐198. CONABIO (2008) Sistema de información sobre especies invasoras en México. Especies invasoras ‐ Plantas. Comisión Nacional para
el Conocimiento y Uso de la Biodiversidad. Fecha de acceso. www.conabio.gob.mx/invasoras/index.php/Especies_invasoras_‐_Plantas
Conservatoire Botanique National De Mascarin (BOULLET V. coord.) (2007) Lagarosiphon major. Index de la flore vasculaire de la Réunion (Trachéophytes) : statuts, menaces et protections. ‐ Version 2007.1 (mise à jour 12 juin 2007).
Cook C.D.K. (2004) Aquatic and Wetland Plants of Southern Africa. Backhuys Publishers, The Netherlands. de Carvalho R.F., Bromilow R.H., Greenwood R. (2007) Uptake of pesticides from water by curly waterweed Lagarosiphon major and
lesser duckweed Lemna minor. Pest Management Science 63:789–797. [BIOTECHNOLOGY section] Egloff F. (1975) New and noteworthy species of Swiss flora. Bulletin de la Societe Botanique Suisse 84: 333‐342. Global Invasive Species Database: http://www.issg.org/database/species/ecology.asp?si=403&fr=1&sts=sss [BIOLOGY section] and
[BIOTECHNOLOGY section] ITIS (Integrated Taxonomic Information System) (2005) Online Database Lagarosiphon major.
http://www.itis.gov/servlet/SingleRpt/SingleRpt?search_topic=TSN&search_value=565981 James C.S., Eaton J.W., Hardwick K. (1999) Competition between three submerged macrophytes, Elodea canadensis Michx, Elodea
nuttallii (Planch.) St John and Lagarosiphon major (Ridl.) Moss. Hydrobiologia 415: 35‐40. Little E.C.S. (1979) Handbook of utilization of aquatic plants. FAO Fisheries Technical Paper No. 187, Rome. [BIOTECHNOLOGY
section] Rattray M.R. (1995) The relationship between P, Fe and Mn uptakes by submersed rooted angiosperms. Hydrobiologia 308: 117‐
120. Rattray M.R., Howard‐Williams C., Brown J.M. (1994) Rates of early growth of propagules of Lagarosiphon major and Myriophyllum
triphyllum in lakes of differing trophic status. New Zealand Journal of Marine and Freshwater Research 28: 235‐241. Riis T., Biggs B.J., Flanagan M. (2003) Seasonal changes in macrophyte biomass in South Island lowland streams, New Zealand. New
Zealand Journal of Marine and Freshwater Research 37: 381‐388. Roy B., Popay I., Champion P. James T., Rahman A. (2004) An Illustrated Guide to Common Weeds of New Zealand 2nd Edition.
Lagarosiphon major oxygen weed. New Zealand Plant Protection Society. State of Queensland (2004) Lagarosiphon major description. The State of Queensland (Department of Natural Resources and
Mines). Strickland R., Harding J., Shearer L. (2000) The Biology of Lake Dunstan. Cawthron Report No. 563; Contact Energy Limited.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 230 of 258
Symoens L., Triest S. (1983) Monograph of the African genus Lagarosiphon. Bullettin du Jardin Botanique National de la Belgique 53: 441‐488.
University of Florida (2001) Lagarosiphon major (Ridley) Moss. Non‐Native Invasive Aquatic Plants in the United States, Center for Aquatic and Invasive Plants, University of Florida and Sea Grant.
Wells R.D., De‐Winton M.D., Clayton J.S. (1997) Successive macrophyte invasions within the submerged flora of Lake Tarawera, central North Island, New Zealand. New Zealand Journal of Marine and Freshwater Research 31: 449‐459.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 231 of 258
10.5 Lemna minor
Figure 95 ‐ Lemna minor Linnaeus
Robert H. Mohlenbrock @ USDA‐NRCS PLANTS Database / USDA NRCS. 1995. Northeast wetland flora: Field office guide to plant species. Northeast National Technical Center, Chester
SYMBOLS: B, E, PIV
TAXONOMY
Phylum Magnoliophyta Class Liliopsida Order Arales Family Lemnaceae Genus Lemna Species Lemna minor
Related species
L. aequinoctialis, L. disperma, L. gibba, L. japonica, L. minuta, L. obscura, L. perpusilla, L. tenera, L. trisulca, L. turioni fera, L.mna valdiviana, L. yungensis.
BIOLOGY General Description of Major divisions and classes:
The Lemnaceae is a monocotyledonous family of 4 genera: Spirodela, Lemna, Wolffia and Wolfiella, and 37 species. All Lemnaceae species are small aquatic plants, commonly called duckweeds (Lemna and Spirodela species) and water meals (Wolffia species). The majority of research involving these plants has been done with only a few species; primarily L. gibba and L. minor, Spirodela polyrrhiza and Spirodela punctata, and to a lesser extent, Wolffia globosa. For the majority of species, little is known of their biology and the ability to extrapolate any of the technological applications developed for the better studied species (Stomp, 2005).

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 232 of 258
Physiological characteristics
The plants are perennials, growing anywhere there is water and sun. These plants require little mechanical support or vascular tissue (the smallest members of the family completely lack xylem and phloem) so most cells resemble maturing leaves expending very little photosynthetic energy on plant structures. As a result, vegetative propagation of these plants has high biomass yield potential.
Structural and morphological features
Members of the family Lemnaceae are tiny aquatic monocots that range in size from 1.5 cm long (Spirodela polyrhiza) to less than a millimeter (Wolffia globosa).
All members of the Lemnaceae are small, free‐floating, fresh‐water plants whose geographical range spans the entire globe (Landolt, 1986). Members of the Lemnaceae are the most morphologically reduced plants known. Plants of Spirodela and Lemna species consist of a frond, a root or roots (the number of roots are species specific) and, when present, a flower. The morphology of Wolffia species is further reduced, with plants consisting of tiny fronds devoid of roots and producing single flowers. Wolfiella species are more varied in morphology.
Biochemical composition
Table 21 ‐ Biochemical composition of Duckweed. Composition expressed as % dry weight except for Water and ash. From Landolt and Kandeler (1987a), except for starch (Cheng and Stomp, 2009).
Duckweed (Lemnaceae) Water (%FW) 86‐97 Ash 12.0‐27.6 Total Carbohydrate 14.1‐43.6 Crude fiber 5.7‐16.2 Starch 6‐75 (some strains in lab condition)
Protein 6.8‐ 45.0 Lipid 1.8 ‐9.2 K 0.03‐7.0 Na 0.03‐1.3 Ca 0.18‐4.5 Mg 0.04‐2.8 Cu 0.2‐10‐3 ‐ 3.2 Si 0.41‐5.35 S 0.33‐7.0 P 0.03‐2.8
The protein content of duckweeds is one of the highest in the plant kingdom, but it is dependent on growth conditions. Typically duckweeds are rich in leucine, threonine, valine, isoleucine and phenylalanine. They tend to be low in cysteine, methionine, and tyrosine.
Duckweed growth can be optimized to produce high levels of protein or high levels of starch. The plant's dry weight accumulation varies by species and growth conditions and ranges from 6 to 20% of fresh weight (Landolt and Kandeler, 1987b; Tillberg, 1979). Protein content of a number of duckweed species grown under varying conditions has been reported to range from 15 to 45% dry weight (Chang, 1977; Porath, 1979; Appenroth, 1982). These values place the protein content of dry duckweed biomass between alfalfa meal (20%) and soybean meal (41.7%) (Hillman, 1961). We routinely grow duckweed on dilute swine wastewater and get 30 to 35% protein of dry duckweed. Duckweed starch content is dependent on growth conditions, e.g., pH, phosphate concentration (Tasseron‐De‐Jong, 1971; McLaren and Smith, 1976) and
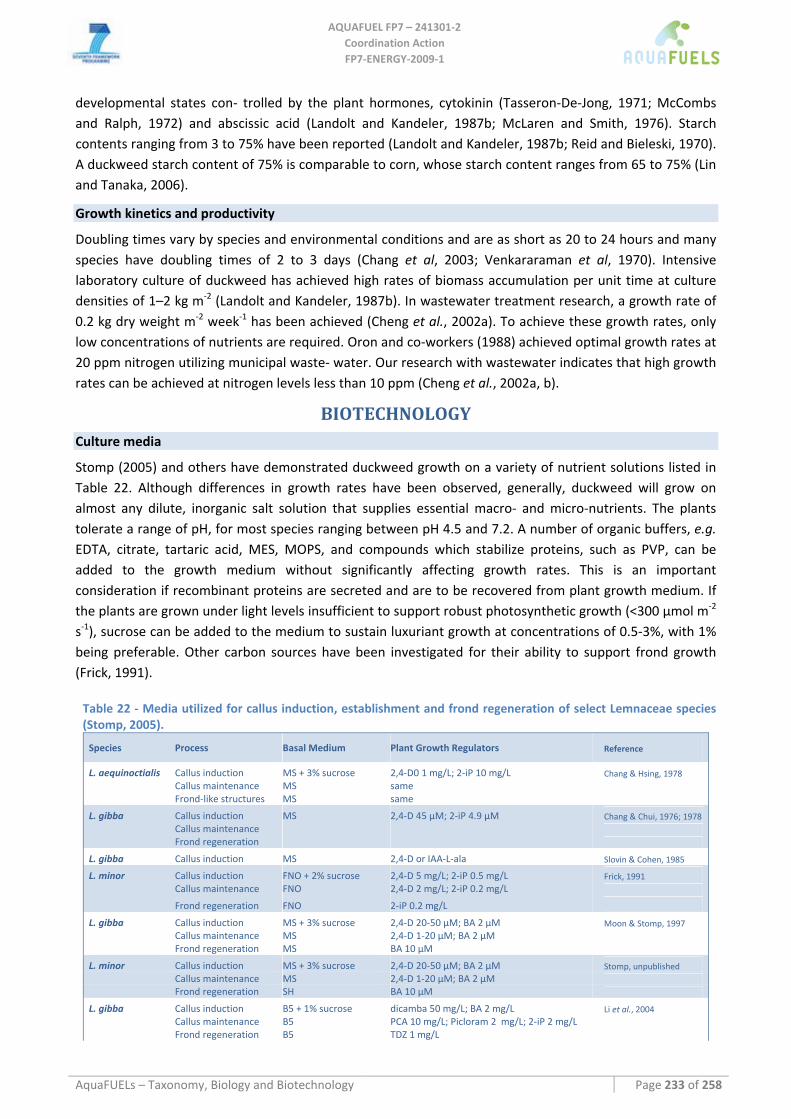
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 233 of 258
developmental states con‐ trolled by the plant hormones, cytokinin (Tasseron‐De‐Jong, 1971; McCombs and Ralph, 1972) and abscissic acid (Landolt and Kandeler, 1987b; McLaren and Smith, 1976). Starch contents ranging from 3 to 75% have been reported (Landolt and Kandeler, 1987b; Reid and Bieleski, 1970). A duckweed starch content of 75% is comparable to corn, whose starch content ranges from 65 to 75% (Lin and Tanaka, 2006).
Growth kinetics and productivity
Doubling times vary by species and environmental conditions and are as short as 20 to 24 hours and many species have doubling times of 2 to 3 days (Chang et al, 2003; Venkararaman et al, 1970). Intensive laboratory culture of duckweed has achieved high rates of biomass accumulation per unit time at culture densities of 1–2 kg m‐2 (Landolt and Kandeler, 1987b). In wastewater treatment research, a growth rate of 0.2 kg dry weight m‐2 week‐1 has been achieved (Cheng et al., 2002a). To achieve these growth rates, only low concentrations of nutrients are required. Oron and co‐workers (1988) achieved optimal growth rates at 20 ppm nitrogen utilizing municipal waste‐ water. Our research with wastewater indicates that high growth rates can be achieved at nitrogen levels less than 10 ppm (Cheng et al., 2002a, b).
BIOTECHNOLOGY Culture media
Stomp (2005) and others have demonstrated duckweed growth on a variety of nutrient solutions listed in Table 22. Although differences in growth rates have been observed, generally, duckweed will grow on almost any dilute, inorganic salt solution that supplies essential macro‐ and micro‐nutrients. The plants tolerate a range of pH, for most species ranging between pH 4.5 and 7.2. A number of organic buffers, e.g. EDTA, citrate, tartaric acid, MES, MOPS, and compounds which stabilize proteins, such as PVP, can be added to the growth medium without significantly affecting growth rates. This is an important consideration if recombinant proteins are secreted and are to be recovered from plant growth medium. If the plants are grown under light levels insufficient to support robust photosynthetic growth (<300 μmol m‐2 s‐1), sucrose can be added to the medium to sustain luxuriant growth at concentrations of 0.5‐3%, with 1% being preferable. Other carbon sources have been investigated for their ability to support frond growth (Frick, 1991).
Table 22 ‐ Media utilized for callus induction, establishment and frond regeneration of select Lemnaceae species (Stomp, 2005).
Species Process Basal Medium Plant Growth Regulators Reference
L. aequinoctialis Callus induction MS + 3% sucrose 2,4‐D0 1 mg/L; 2‐iP 10 mg/L Chang & Hsing, 1978 Callus maintenance MS same Frond‐like structures MS same
L. gibba Callus induction MS 2,4‐D 45 μM; 2‐iP 4.9 μM Chang & Chui, 1976; 1978 Callus maintenance Frond regeneration
L. gibba Callus induction MS 2,4‐D or IAA‐L‐ala Slovin & Cohen, 1985
L. minor Callus induction FNO + 2% sucrose 2,4‐D 5 mg/L; 2‐iP 0.5 mg/L Frick, 1991 Callus maintenance FNO 2,4‐D 2 mg/L; 2‐iP 0.2 mg/L
Frond regeneration FNO 2‐iP 0.2 mg/L
L. gibba Callus induction MS + 3% sucrose 2,4‐D 20‐50 μM; BA 2 μM Moon & Stomp, 1997 Callus maintenance MS 2,4‐D 1‐20 μM; BA 2 μM Frond regeneration MS BA 10 μM
L. minor Callus induction MS + 3% sucrose 2,4‐D 20‐50 μM; BA 2 μM Stomp, unpublished Callus maintenance MS 2,4‐D 1‐20 μM; BA 2 μM Frond regeneration SH BA 10 μM
L. gibba Callus induction B5 + 1% sucrose dicamba 50 mg/L; BA 2 mg/L Li et al., 2004 Callus maintenance B5 PCA 10 mg/L; Picloram 2 mg/L; 2‐iP 2 mg/L Frond regeneration B5 TDZ 1 mg/L
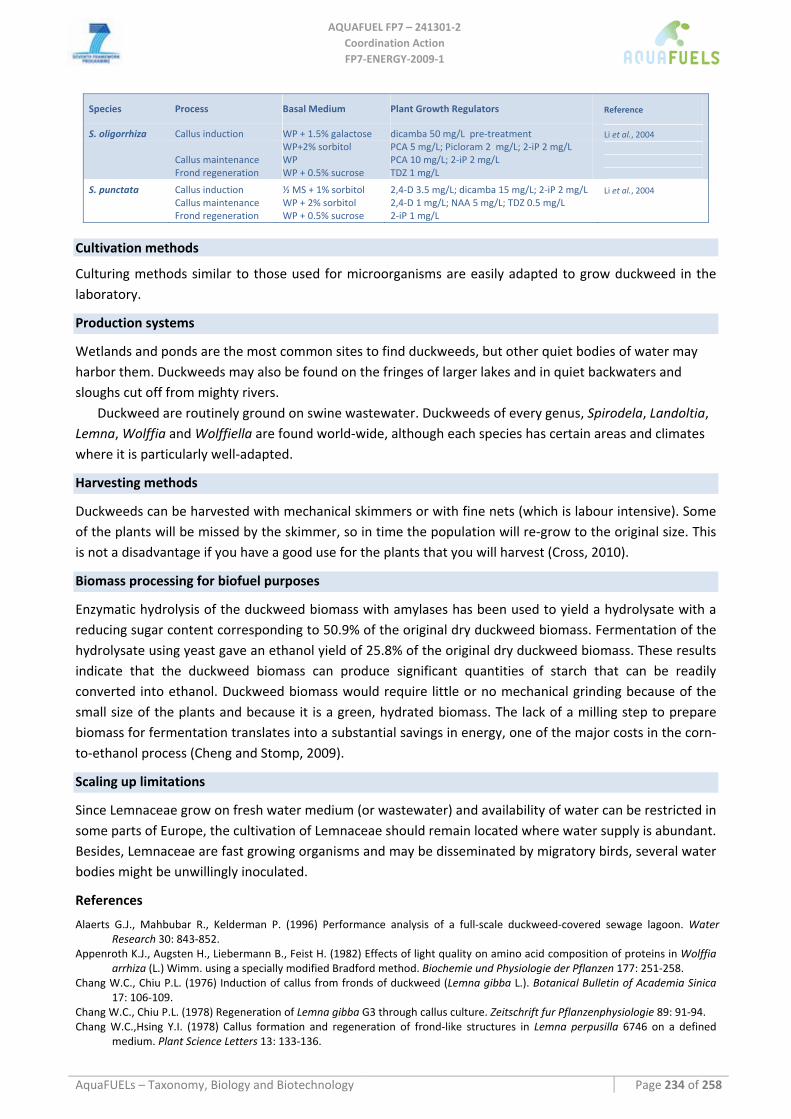
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 234 of 258
Species Process Basal Medium Plant Growth Regulators Reference
S. oligorrhiza Callus induction WP + 1.5% galactose dicamba 50 mg/L pre‐treatment Li et al., 2004 WP+2% sorbitol PCA 5 mg/L; Picloram 2 mg/L; 2‐iP 2 mg/L Callus maintenance WP PCA 10 mg/L; 2‐iP 2 mg/L Frond regeneration WP + 0.5% sucrose TDZ 1 mg/L
S. punctata Callus induction ½ MS + 1% sorbitol 2,4‐D 3.5 mg/L; dicamba 15 mg/L; 2‐iP 2 mg/L Li et al., 2004 Callus maintenance WP + 2% sorbitol 2,4‐D 1 mg/L; NAA 5 mg/L; TDZ 0.5 mg/L Frond regeneration WP + 0.5% sucrose 2‐iP 1 mg/L
Cultivation methods
Culturing methods similar to those used for microorganisms are easily adapted to grow duckweed in the laboratory.
Production systems
Wetlands and ponds are the most common sites to find duckweeds, but other quiet bodies of water may harbor them. Duckweeds may also be found on the fringes of larger lakes and in quiet backwaters and sloughs cut off from mighty rivers.
Duckweed are routinely ground on swine wastewater. Duckweeds of every genus, Spirodela, Landoltia, Lemna, Wolffia and Wolffiella are found world‐wide, although each species has certain areas and climates where it is particularly well‐adapted.
Harvesting methods
Duckweeds can be harvested with mechanical skimmers or with fine nets (which is labour intensive). Some of the plants will be missed by the skimmer, so in time the population will re‐grow to the original size. This is not a disadvantage if you have a good use for the plants that you will harvest (Cross, 2010).
Biomass processing for biofuel purposes
Enzymatic hydrolysis of the duckweed biomass with amylases has been used to yield a hydrolysate with a reducing sugar content corresponding to 50.9% of the original dry duckweed biomass. Fermentation of the hydrolysate using yeast gave an ethanol yield of 25.8% of the original dry duckweed biomass. These results indicate that the duckweed biomass can produce significant quantities of starch that can be readily converted into ethanol. Duckweed biomass would require little or no mechanical grinding because of the small size of the plants and because it is a green, hydrated biomass. The lack of a milling step to prepare biomass for fermentation translates into a substantial savings in energy, one of the major costs in the corn‐to‐ethanol process (Cheng and Stomp, 2009).
Scaling up limitations
Since Lemnaceae grow on fresh water medium (or wastewater) and availability of water can be restricted in some parts of Europe, the cultivation of Lemnaceae should remain located where water supply is abundant. Besides, Lemnaceae are fast growing organisms and may be disseminated by migratory birds, several water bodies might be unwillingly inoculated.
References
Alaerts G.J., Mahbubar R., Kelderman P. (1996) Performance analysis of a full‐scale duckweed‐covered sewage lagoon. Water Research 30: 843‐852.
Appenroth K.J., Augsten H., Liebermann B., Feist H. (1982) Effects of light quality on amino acid composition of proteins in Wolffia arrhiza (L.) Wimm. using a specially modified Bradford method. Biochemie und Physiologie der Pflanzen 177: 251‐258.
Chang W.C., Chiu P.L. (1976) Induction of callus from fronds of duckweed (Lemna gibba L.). Botanical Bulletin of Academia Sinica 17: 106‐109.
Chang W.C., Chiu P.L. (1978) Regeneration of Lemna gibba G3 through callus culture. Zeitschrift fur Pflanzenphysiologie 89: 91‐94. Chang W.C.,Hsing Y.I. (1978) Callus formation and regeneration of frond‐like structures in Lemna perpusilla 6746 on a defined
medium. Plant Science Letters 13: 133‐136.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 235 of 258
Chang S.M., Yang C.C., Sung S.C. (1977) The cultivation and the nutritional value of Lemnaceae. Bullettin of the Institute of Chemistry, Academia Sinica 24: 19‐30.
Cheng J.J., Stomp A.M. (2009) Growing duckweed to recover nutrients from wastewaters and for production of fuel ethanol and animal feed. CLEAN – Soil, Air, Water 37: 17‐26.
Cheng J., Bergmann B.A., Classen J.J., Stomp A.M., Howard J.W. (2002a) Nutrient recovery from swine lagoon water by Spirodela punctata. Bioresource Technology 81: 81‐85.
Cheng J., Landesman L., Bergmann B.A., Classen J.J., Howard J.W., Yamamoto Y.T. (2002b) Nutrient removal from swine lagoon liquid by Lemna minor 8627. Transactions of the ASAE 45: 1003‐1010.
Cross J.W. (2010) The Charms of Duckweed. http://www.mobot.org/jwcross/duckweed.htm (accessed 10 Nov. 2010). Den Hartog C. (1975) Thoughts about the taxonomical relationships within the lemnaceae. Aquatic Botany 1: 407‐416. Frick H. (1991) Callogenesis and carbohydrate utilization in Lemna minor. Journal of Plant Physiology 137: 397‐401. Hillman W.S. (1961) The Lemnaceae or duckweeds. A review of the descriptive and experimental literature. Botanical Review 27,:
221‐287. ITIS (2010) http://www.itis.gov/servlet/SingleRpt/SingleRpt accessed on the 01/07/2010 Landolt E., Kandeler R. (1987a) Biosystematic investigations in the family of duckweeds (Lemnaceae). Veroff. Geobot. Inst. ETH,
Zurich. vol. 2, pp. 42‐43. Landolt E and Kandeler, R. (1987b). The family of Lemnaceae – a monographic study. Volume 2. Biosystematic investigations in the
family of duckweeds (Lemnaceae) (vol. 4). Veroff. Geobot. Inst. ETH, Zurich. Li J., Jani M., Vunsh R., Vishnevetsky J., Hannania U., Flaishman M., Perl A., Edelman M. (2004) Callus induction and regeneration in
Spirodela and Lemna. Plant Cell Reports 22: 457‐464. Lin Y., Tanaka S. (2006) Ethanol fermentation from biomass resources: current state and prospects. Applied Microbiology and
Biotechnology 69: 627‐642. McCombs P.J.A., Ralph R.K. (1972) Protein, nucleic acid and starch metabolism in the duckweed, Spirodela oligorrhiza, treated with
cytokinins. Biochemistry Journal 129: 403‐417. McLaren J.S., Smith H. (1976) The effect of abscisic acid on growth, photosynthetic rate and carbohydrate metabolism in Lemna
minor. New Phytologist 76: 11‐20. Moon H.K., Stomp A.M. (1997) Effects of medium components and light on callus induction, growth and frond regeneration in
Lemna gibba (duckweed). In Vitro Cellular and Develepmental Biology‐Plant 33: 20‐25. Oron G., de‐Vegt A., Porath D., (1988) Nitrogen removal and conversion by duckweed grown on wastewater. Water Research 22:
179‐184. Porath D., Hepher B., Koton A. (1979) Duckweed as an aquatic crop: evaluation of clones for aquaculture. Aquatic Botany 7: 273‐
278. Reid M.S., Bieleski R.L. (1970) Response of Spirodela oligorrhiza (Rockville) to phosphorus deficiency. Plant Physiology 46: 609‐613. Slovin J.P. Cohen J.D. (1985) Generation of variant lines of Lemna gibba G3 via tissue culture, for use in selecting auxin metabolism
mutants. Plant Physiology 77(4 suppl.): 11. Stomp A.M. (2005) The duckweeds: A valuable plant for biomanufacturing. In: El‐Gewely M.R. (ed.) Biotechnology Annual Review,
Elsevier, Rotterdam, pp. 69‐99. Tasseron‐De‐Jong J., Veldstra H. (1971) Investigations on cytokinins. Part 2: Interaction of light and cytokinins as studied in Lemna
minor. Physiologia Plantarum 24: 239‐241. Tillberg E., Holmvall M., Ericsson T. (1979) Growth cycles in Lemna gibba cultures and their effects on growth rate and
ultrastructure. Physiologia Plantarum 46: 5‐12. Venkararaman R., Seth P.N., Maheshwari S.C. (1970) Studies on the growth and flowering of a short‐day plant, Wolffia
microscopica. I. General aspects and induction of flowering by cytokinins. Zeitschrift fur Pflanzenphysiologie 62: 316‐327.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 236 of 258
11 Concluding remarks
Algae have been recognized as valuable resource and exploited by humans for thousands of years. The last century has seen intensive research efforts to exploit the huge potential of micro‐ and macro algae and other aquatic plants for food, fodder, energy and biomaterials, as well as environmental applications such as waste water treatment and bioremediation that are summarized in the document above.
While a significant industry based on exploitation of macroalgae biomass has developed, microalgal biomass production remains restricted to a few high value nutraceutical products such as pigments (β‐carotene from Dunaliella and astaxanthin from Haematococcus) or dried microalgal biomass (Chlorella, Spirulina).
The huge efforts invested recently in investigating the potential of microalgae for biofuels production have significantly advanced our understanding in algal biology, cultivation, harvesting and biomass processing, but no commercially viable sustainable production pathway exists and in consequence no algal biofuels production facility is online today.
Difficulties to be overcome are mostly in reducing production and processing costs of algal biomass, by means of progress in engineering and culture management, and by selecting and improving strains for maximal productivity at minimal costs. Dozens of algal species and strains hold promise in this respect, and no strain may be singled out for extraordinary suitability under the wide variety of cultivation conditions imaginable. Many algae may besides fuel provide high value products or protein for improving the economic balance of the production process. Recent advances in genomics and genetic engineering, demonstrated in Chlamydomonas, Phaeodactylum and others, holds promise for targeted improvements of algal strains to adapt them specifically for enhanced growth and productivity under selected conditions. Alternatively great opportunities may reside in utilization of algal biomass cultivation for environmental applications such as waste water treatment or remediation, under recovery of valuable nutrients and biomass production.
All possible schemes proposed have been demonstrated in the small scale, but intensive research efforts are still necessary to further our understanding on algal biology and production processes. Any significant advance in biofuels production is dependent on massive funding for the establishment of multiple, different and diverse algal biomass production facilities of sufficient size for allowing gaining the necessary insights and data towards reaching sustainable and profitable production processes.

AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs‐ Taxonomy, Biology and Biotechnology Page 237 of 258
ANNEX I
AVAILABILITY of ALGAE
The main Culture Collections of alage and cyanobacteria are listed in the table below:
Collection + website CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/ CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/56 UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php PCC (Pasteur Culture Collection of Cyanobacteria) http://www.pasteur.fr/ip/easysite/go/03b‐000012‐00g/collection‐of‐cyanobacteria‐pcc/ ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/ RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC BCCM (Belgian Co‐ordinated Collections of Micro‐organisms) Diatom Collection at the Ghent University F&M Culture Collection, Italy http://www.femonline.it/ CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm CCCM (Canadian Center for the Culture of Microorganisms) including: NEPCC (North East Pacific Culture Collection) FWAC (Freshwater Algal Culture Collection) http://botany.ubc.ca/cccm CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/ SERI microalgae culture collection http://www.tpub.com/content/altfuels05/3814/38140091.htm FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en Japan ‐ NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
The availability of each microalga and cyanobacterium considered in the document is reported in the following section. No availability was found for Parietochloris incisa. Note that some strains are shared between collections. Note also that many strains are owned by Research Institutes and Universities and are not deposited in Culture Collections, so that their availability is not known.
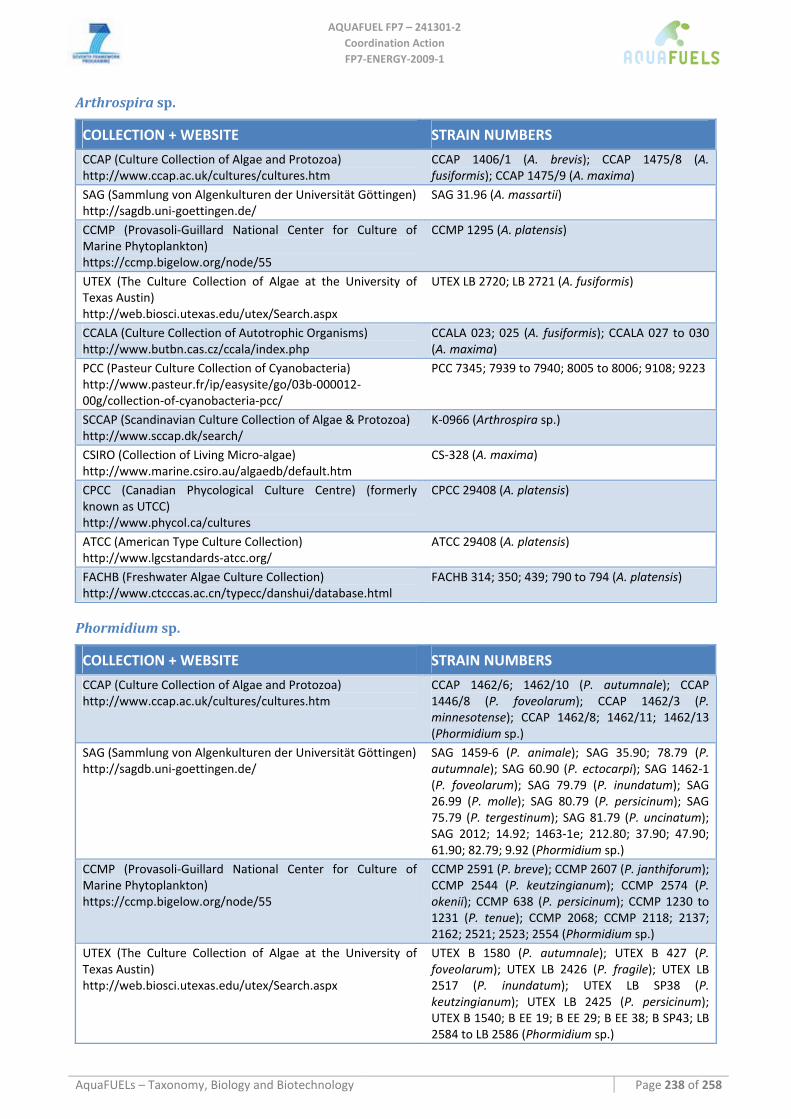
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 238 of 258
Arthrospira sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1406/1 (A. brevis); CCAP 1475/8 (A. fusiformis); CCAP 1475/9 (A. maxima)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 31.96 (A. massartii)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 1295 (A. platensis)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX LB 2720; LB 2721 (A. fusiformis)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 023; 025 (A. fusiformis); CCALA 027 to 030 (A. maxima)
PCC (Pasteur Culture Collection of Cyanobacteria) http://www.pasteur.fr/ip/easysite/go/03b‐000012‐00g/collection‐of‐cyanobacteria‐pcc/
PCC 7345; 7939 to 7940; 8005 to 8006; 9108; 9223
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐0966 (Arthrospira sp.)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐328 (A. maxima)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 29408 (A. platensis)
ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 29408 (A. platensis)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 314; 350; 439; 790 to 794 (A. platensis)
Phormidium sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1462/6; 1462/10 (P. autumnale); CCAP 1446/8 (P. foveolarum); CCAP 1462/3 (P. minnesotense); CCAP 1462/8; 1462/11; 1462/13 (Phormidium sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 1459‐6 (P. animale); SAG 35.90; 78.79 (P. autumnale); SAG 60.90 (P. ectocarpi); SAG 1462‐1 (P. foveolarum); SAG 79.79 (P. inundatum); SAG 26.99 (P. molle); SAG 80.79 (P. persicinum); SAG 75.79 (P. tergestinum); SAG 81.79 (P. uncinatum); SAG 2012; 14.92; 1463‐1e; 212.80; 37.90; 47.90; 61.90; 82.79; 9.92 (Phormidium sp.)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 2591 (P. breve); CCMP 2607 (P. janthiforum); CCMP 2544 (P. keutzingianum); CCMP 2574 (P. okenii); CCMP 638 (P. persicinum); CCMP 1230 to 1231 (P. tenue); CCMP 2068; CCMP 2118; 2137; 2162; 2521; 2523; 2554 (Phormidium sp.)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX B 1580 (P. autumnale); UTEX B 427 (P. foveolarum); UTEX LB 2426 (P. fragile); UTEX LB 2517 (P. inundatum); UTEX LB SP38 (P. keutzingianum); UTEX LB 2425 (P. persicinum); UTEX B 1540; B EE 19; B EE 29; B EE 38; B SP43; LB 2584 to LB 2586 (Phormidium sp.)
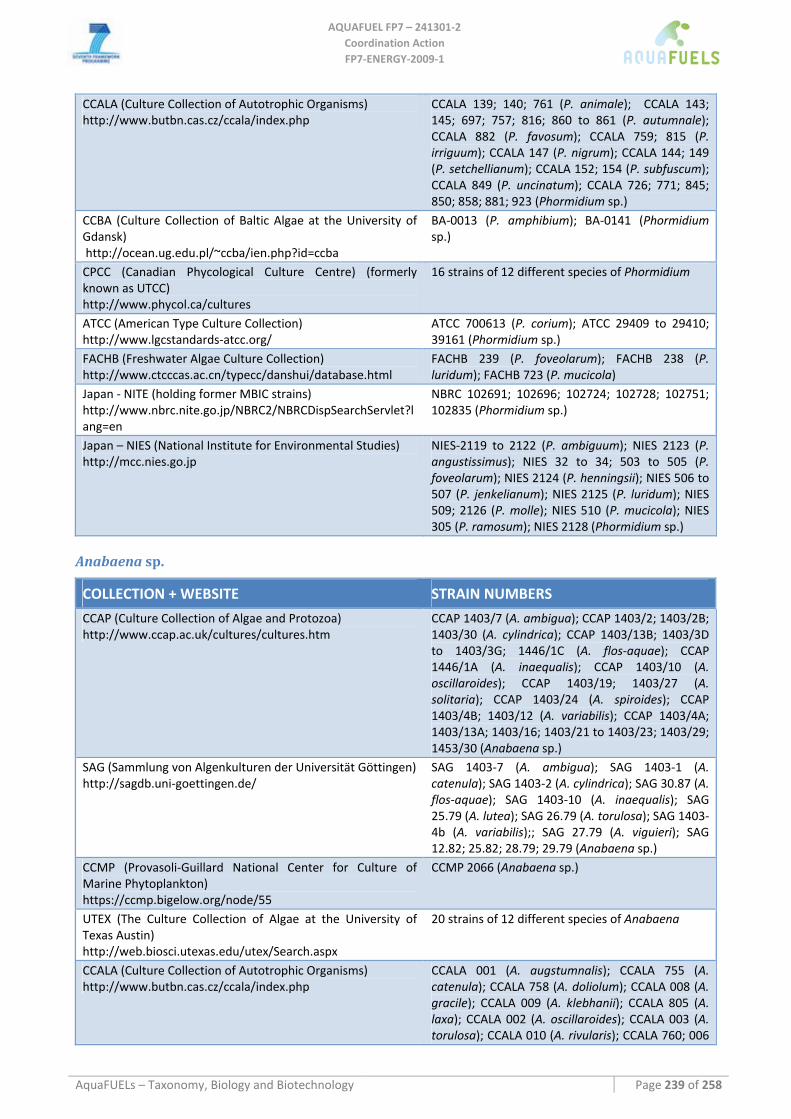
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 239 of 258
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 139; 140; 761 (P. animale); CCALA 143; 145; 697; 757; 816; 860 to 861 (P. autumnale); CCALA 882 (P. favosum); CCALA 759; 815 (P. irriguum); CCALA 147 (P. nigrum); CCALA 144; 149 (P. setchellianum); CCALA 152; 154 (P. subfuscum); CCALA 849 (P. uncinatum); CCALA 726; 771; 845; 850; 858; 881; 923 (Phormidium sp.)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA‐0013 (P. amphibium); BA‐0141 (Phormidium sp.)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
16 strains of 12 different species of Phormidium
ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 700613 (P. corium); ATCC 29409 to 29410; 39161 (Phormidium sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 239 (P. foveolarum); FACHB 238 (P. luridum); FACHB 723 (P. mucicola)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102691; 102696; 102724; 102728; 102751; 102835 (Phormidium sp.)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES‐2119 to 2122 (P. ambiguum); NIES 2123 (P. angustissimus); NIES 32 to 34; 503 to 505 (P. foveolarum); NIES 2124 (P. henningsii); NIES 506 to 507 (P. jenkelianum); NIES 2125 (P. luridum); NIES 509; 2126 (P. molle); NIES 510 (P. mucicola); NIES 305 (P. ramosum); NIES 2128 (Phormidium sp.)
Anabaena sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1403/7 (A. ambigua); CCAP 1403/2; 1403/2B; 1403/30 (A. cylindrica); CCAP 1403/13B; 1403/3D to 1403/3G; 1446/1C (A. flos‐aquae); CCAP 1446/1A (A. inaequalis); CCAP 1403/10 (A. oscillaroides); CCAP 1403/19; 1403/27 (A. solitaria); CCAP 1403/24 (A. spiroides); CCAP 1403/4B; 1403/12 (A. variabilis); CCAP 1403/4A; 1403/13A; 1403/16; 1403/21 to 1403/23; 1403/29; 1453/30 (Anabaena sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 1403‐7 (A. ambigua); SAG 1403‐1 (A. catenula); SAG 1403‐2 (A. cylindrica); SAG 30.87 (A. flos‐aquae); SAG 1403‐10 (A. inaequalis); SAG 25.79 (A. lutea); SAG 26.79 (A. torulosa); SAG 1403‐4b (A. variabilis);; SAG 27.79 (A. viguieri); SAG 12.82; 25.82; 28.79; 29.79 (Anabaena sp.)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 2066 (Anabaena sp.)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
20 strains of 12 different species of Anabaena
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 001 (A. augstumnalis); CCALA 755 (A. catenula); CCALA 758 (A. doliolum); CCALA 008 (A. gracile); CCALA 009 (A. klebhanii); CCALA 805 (A. laxa); CCALA 002 (A. oscillaroides); CCALA 003 (A. torulosa); CCALA 010 (A. rivularis); CCALA 760; 006
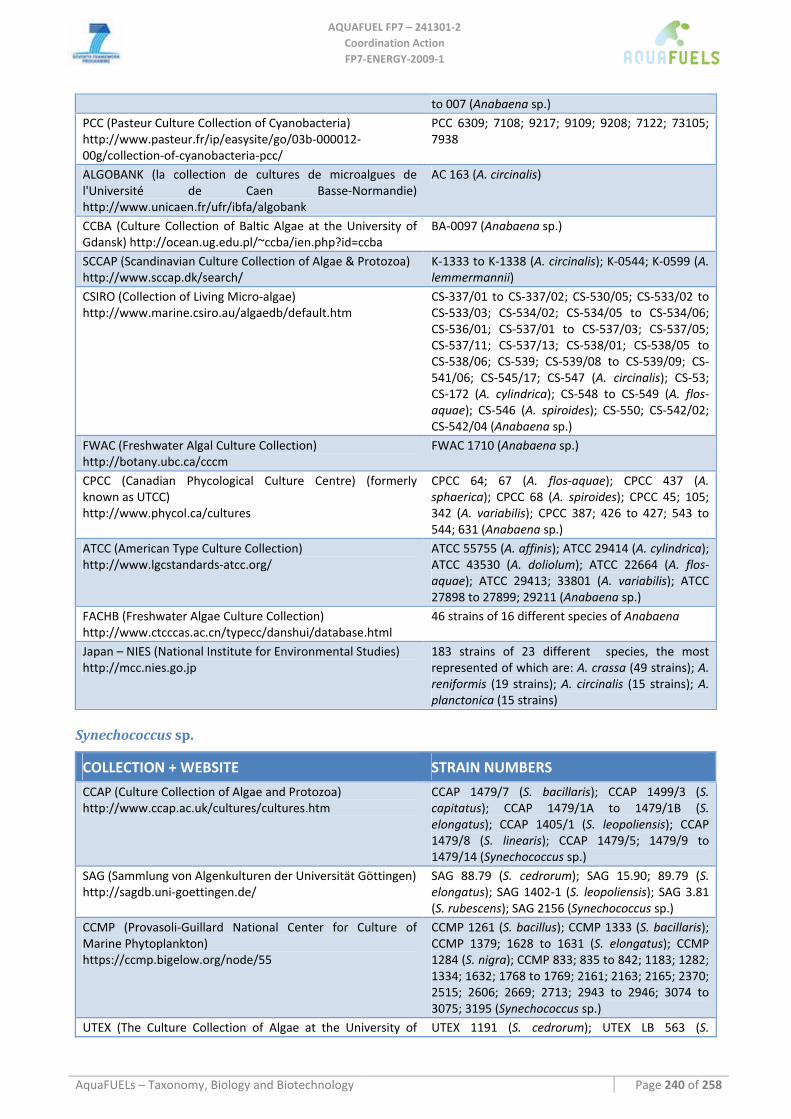
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 240 of 258
to 007 (Anabaena sp.) PCC (Pasteur Culture Collection of Cyanobacteria) http://www.pasteur.fr/ip/easysite/go/03b‐000012‐00g/collection‐of‐cyanobacteria‐pcc/
PCC 6309; 7108; 9217; 9109; 9208; 7122; 73105; 7938
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 163 (A. circinalis)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA‐0097 (Anabaena sp.)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1333 to K‐1338 (A. circinalis); K‐0544; K‐0599 (A. lemmermannii)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐337/01 to CS‐337/02; CS‐530/05; CS‐533/02 to CS‐533/03; CS‐534/02; CS‐534/05 to CS‐534/06; CS‐536/01; CS‐537/01 to CS‐537/03; CS‐537/05; CS‐537/11; CS‐537/13; CS‐538/01; CS‐538/05 to CS‐538/06; CS‐539; CS‐539/08 to CS‐539/09; CS‐541/06; CS‐545/17; CS‐547 (A. circinalis); CS‐53; CS‐172 (A. cylindrica); CS‐548 to CS‐549 (A. flos‐aquae); CS‐546 (A. spiroides); CS‐550; CS‐542/02; CS‐542/04 (Anabaena sp.)
FWAC (Freshwater Algal Culture Collection) http://botany.ubc.ca/cccm
FWAC 1710 (Anabaena sp.)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 64; 67 (A. flos‐aquae); CPCC 437 (A. sphaerica); CPCC 68 (A. spiroides); CPCC 45; 105; 342 (A. variabilis); CPCC 387; 426 to 427; 543 to 544; 631 (Anabaena sp.)
ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 55755 (A. affinis); ATCC 29414 (A. cylindrica); ATCC 43530 (A. doliolum); ATCC 22664 (A. flos‐aquae); ATCC 29413; 33801 (A. variabilis); ATCC 27898 to 27899; 29211 (Anabaena sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
46 strains of 16 different species of Anabaena
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
183 strains of 23 different species, the most represented of which are: A. crassa (49 strains); A. reniformis (19 strains); A. circinalis (15 strains); A. planctonica (15 strains)
Synechococcus sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1479/7 (S. bacillaris); CCAP 1499/3 (S. capitatus); CCAP 1479/1A to 1479/1B (S. elongatus); CCAP 1405/1 (S. leopoliensis); CCAP 1479/8 (S. linearis); CCAP 1479/5; 1479/9 to 1479/14 (Synechococcus sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 88.79 (S. cedrorum); SAG 15.90; 89.79 (S. elongatus); SAG 1402‐1 (S. leopoliensis); SAG 3.81 (S. rubescens); SAG 2156 (Synechococcus sp.)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 1261 (S. bacillus); CCMP 1333 (S. bacillaris); CCMP 1379; 1628 to 1631 (S. elongatus); CCMP 1284 (S. nigra); CCMP 833; 835 to 842; 1183; 1282; 1334; 1632; 1768 to 1769; 2161; 2163; 2165; 2370; 2515; 2606; 2669; 2713; 2943 to 2946; 3074 to 3075; 3195 (Synechococcus sp.)
UTEX (The Culture Collection of Algae at the University of UTEX 1191 (S. cedrorum); UTEX LB 563 (S.
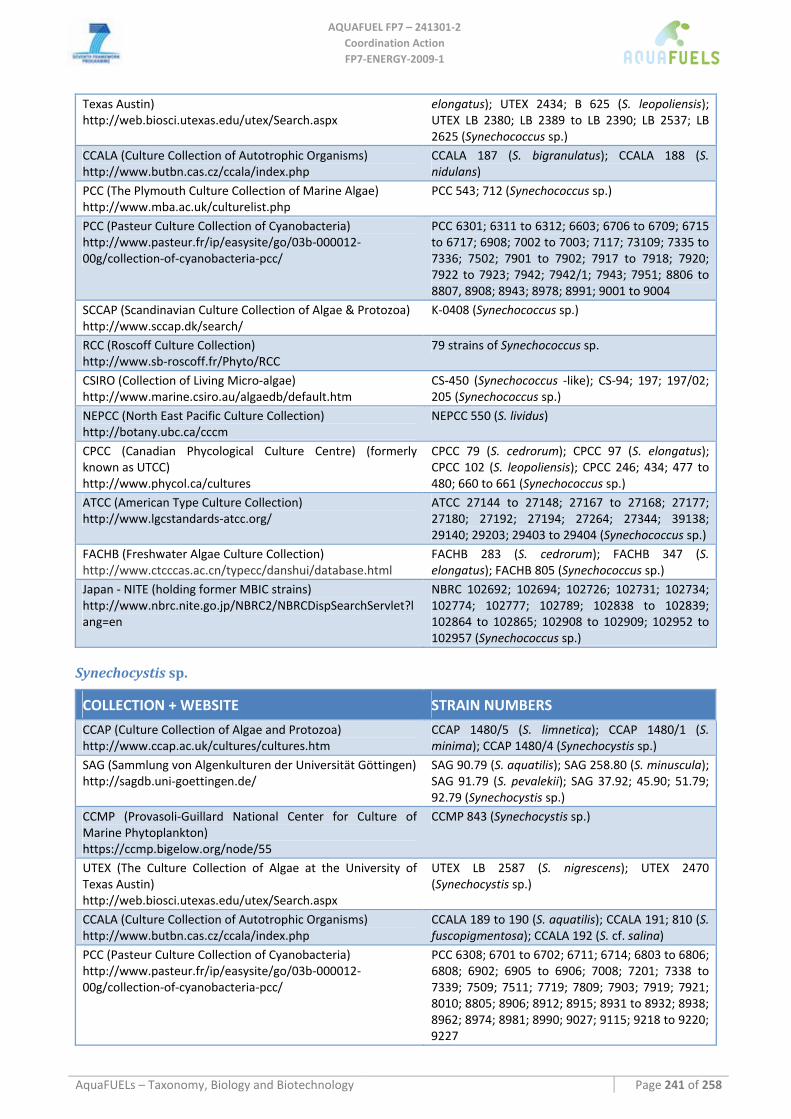
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 241 of 258
Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
elongatus); UTEX 2434; B 625 (S. leopoliensis); UTEX LB 2380; LB 2389 to LB 2390; LB 2537; LB 2625 (Synechococcus sp.)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 187 (S. bigranulatus); CCALA 188 (S. nidulans)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 543; 712 (Synechococcus sp.)
PCC (Pasteur Culture Collection of Cyanobacteria) http://www.pasteur.fr/ip/easysite/go/03b‐000012‐00g/collection‐of‐cyanobacteria‐pcc/
PCC 6301; 6311 to 6312; 6603; 6706 to 6709; 6715 to 6717; 6908; 7002 to 7003; 7117; 73109; 7335 to 7336; 7502; 7901 to 7902; 7917 to 7918; 7920; 7922 to 7923; 7942; 7942/1; 7943; 7951; 8806 to 8807, 8908; 8943; 8978; 8991; 9001 to 9004
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐0408 (Synechococcus sp.)
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
79 strains of Synechococcus sp.
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐450 (Synechococcus ‐like); CS‐94; 197; 197/02; 205 (Synechococcus sp.)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 550 (S. lividus)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 79 (S. cedrorum); CPCC 97 (S. elongatus); CPCC 102 (S. leopoliensis); CPCC 246; 434; 477 to 480; 660 to 661 (Synechococcus sp.)
ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 27144 to 27148; 27167 to 27168; 27177; 27180; 27192; 27194; 27264; 27344; 39138; 29140; 29203; 29403 to 29404 (Synechococcus sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 283 (S. cedrorum); FACHB 347 (S. elongatus); FACHB 805 (Synechococcus sp.)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102692; 102694; 102726; 102731; 102734; 102774; 102777; 102789; 102838 to 102839; 102864 to 102865; 102908 to 102909; 102952 to 102957 (Synechococcus sp.)
Synechocystis sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1480/5 (S. limnetica); CCAP 1480/1 (S. minima); CCAP 1480/4 (Synechocystis sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 90.79 (S. aquatilis); SAG 258.80 (S. minuscula); SAG 91.79 (S. pevalekii); SAG 37.92; 45.90; 51.79; 92.79 (Synechocystis sp.)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 843 (Synechocystis sp.)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX LB 2587 (S. nigrescens); UTEX 2470 (Synechocystis sp.)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 189 to 190 (S. aquatilis); CCALA 191; 810 (S. fuscopigmentosa); CCALA 192 (S. cf. salina)
PCC (Pasteur Culture Collection of Cyanobacteria) http://www.pasteur.fr/ip/easysite/go/03b‐000012‐00g/collection‐of‐cyanobacteria‐pcc/
PCC 6308; 6701 to 6702; 6711; 6714; 6803 to 6806; 6808; 6902; 6905 to 6906; 7008; 7201; 7338 to 7339; 7509; 7511; 7719; 7809; 7903; 7919; 7921; 8010; 8805; 8906; 8912; 8915; 8931 to 8932; 8938; 8962; 8974; 8981; 8990; 9027; 9115; 9218 to 9220; 9227
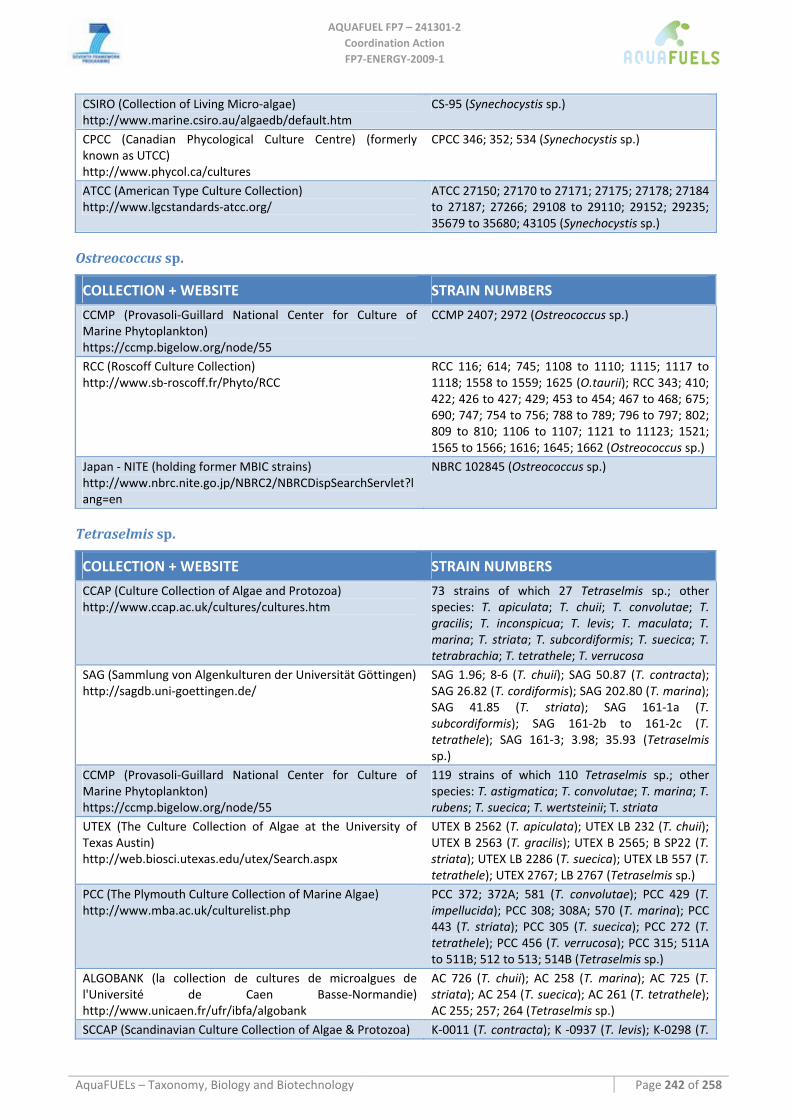
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 242 of 258
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐95 (Synechocystis sp.)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 346; 352; 534 (Synechocystis sp.)
ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 27150; 27170 to 27171; 27175; 27178; 27184 to 27187; 27266; 29108 to 29110; 29152; 29235; 35679 to 35680; 43105 (Synechocystis sp.)
Ostreococcus sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 2407; 2972 (Ostreococcus sp.)
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
RCC 116; 614; 745; 1108 to 1110; 1115; 1117 to 1118; 1558 to 1559; 1625 (O.taurii); RCC 343; 410; 422; 426 to 427; 429; 453 to 454; 467 to 468; 675; 690; 747; 754 to 756; 788 to 789; 796 to 797; 802; 809 to 810; 1106 to 1107; 1121 to 11123; 1521; 1565 to 1566; 1616; 1645; 1662 (Ostreococcus sp.)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102845 (Ostreococcus sp.)
Tetraselmis sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
73 strains of which 27 Tetraselmis sp.; other species: T. apiculata; T. chuii; T. convolutae; T. gracilis; T. inconspicua; T. levis; T. maculata; T. marina; T. striata; T. subcordiformis; T. suecica; T. tetrabrachia; T. tetrathele; T. verrucosa
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 1.96; 8‐6 (T. chuii); SAG 50.87 (T. contracta); SAG 26.82 (T. cordiformis); SAG 202.80 (T. marina); SAG 41.85 (T. striata); SAG 161‐1a (T. subcordiformis); SAG 161‐2b to 161‐2c (T. tetrathele); SAG 161‐3; 3.98; 35.93 (Tetraselmis sp.)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
119 strains of which 110 Tetraselmis sp.; other species: T. astigmatica; T. convolutae; T. marina; T. rubens; T. suecica; T. wertsteinii; T. striata
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX B 2562 (T. apiculata); UTEX LB 232 (T. chuii); UTEX B 2563 (T. gracilis); UTEX B 2565; B SP22 (T. striata); UTEX LB 2286 (T. suecica); UTEX LB 557 (T. tetrathele); UTEX 2767; LB 2767 (Tetraselmis sp.)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 372; 372A; 581 (T. convolutae); PCC 429 (T. impellucida); PCC 308; 308A; 570 (T. marina); PCC 443 (T. striata); PCC 305 (T. suecica); PCC 272 (T. tetrathele); PCC 456 (T. verrucosa); PCC 315; 511A to 511B; 512 to 513; 514B (Tetraselmis sp.)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 726 (T. chuii); AC 258 (T. marina); AC 725 (T. striata); AC 254 (T. suecica); AC 261 (T. tetrathele); AC 255; 257; 264 (Tetraselmis sp.)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) K‐0011 (T. contracta); K ‐0937 (T. levis); K‐0298 (T.
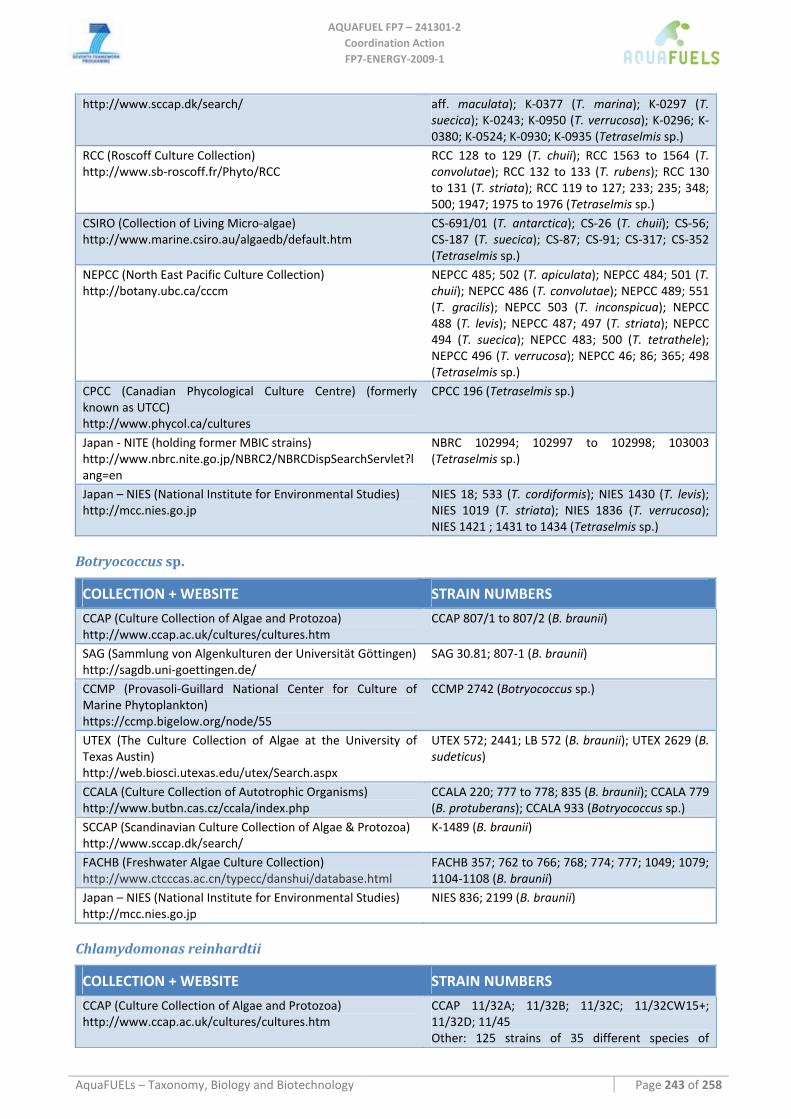
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 243 of 258
http://www.sccap.dk/search/ aff. maculata); K‐0377 (T. marina); K‐0297 (T. suecica); K‐0243; K‐0950 (T. verrucosa); K‐0296; K‐0380; K‐0524; K‐0930; K‐0935 (Tetraselmis sp.)
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
RCC 128 to 129 (T. chuii); RCC 1563 to 1564 (T. convolutae); RCC 132 to 133 (T. rubens); RCC 130 to 131 (T. striata); RCC 119 to 127; 233; 235; 348; 500; 1947; 1975 to 1976 (Tetraselmis sp.)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐691/01 (T. antarctica); CS‐26 (T. chuii); CS‐56; CS‐187 (T. suecica); CS‐87; CS‐91; CS‐317; CS‐352 (Tetraselmis sp.)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 485; 502 (T. apiculata); NEPCC 484; 501 (T. chuii); NEPCC 486 (T. convolutae); NEPCC 489; 551 (T. gracilis); NEPCC 503 (T. inconspicua); NEPCC 488 (T. levis); NEPCC 487; 497 (T. striata); NEPCC 494 (T. suecica); NEPCC 483; 500 (T. tetrathele); NEPCC 496 (T. verrucosa); NEPCC 46; 86; 365; 498 (Tetraselmis sp.)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 196 (Tetraselmis sp.)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102994; 102997 to 102998; 103003 (Tetraselmis sp.)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 18; 533 (T. cordiformis); NIES 1430 (T. levis); NIES 1019 (T. striata); NIES 1836 (T. verrucosa); NIES 1421 ; 1431 to 1434 (Tetraselmis sp.)
Botryococcus sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 807/1 to 807/2 (B. braunii)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 30.81; 807‐1 (B. braunii)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 2742 (Botryococcus sp.)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX 572; 2441; LB 572 (B. braunii); UTEX 2629 (B. sudeticus)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 220; 777 to 778; 835 (B. braunii); CCALA 779 (B. protuberans); CCALA 933 (Botryococcus sp.)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1489 (B. braunii)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 357; 762 to 766; 768; 774; 777; 1049; 1079; 1104‐1108 (B. braunii)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 836; 2199 (B. braunii)
Chlamydomonas reinhardtii
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 11/32A; 11/32B; 11/32C; 11/32CW15+; 11/32D; 11/45 Other: 125 strains of 35 different species of
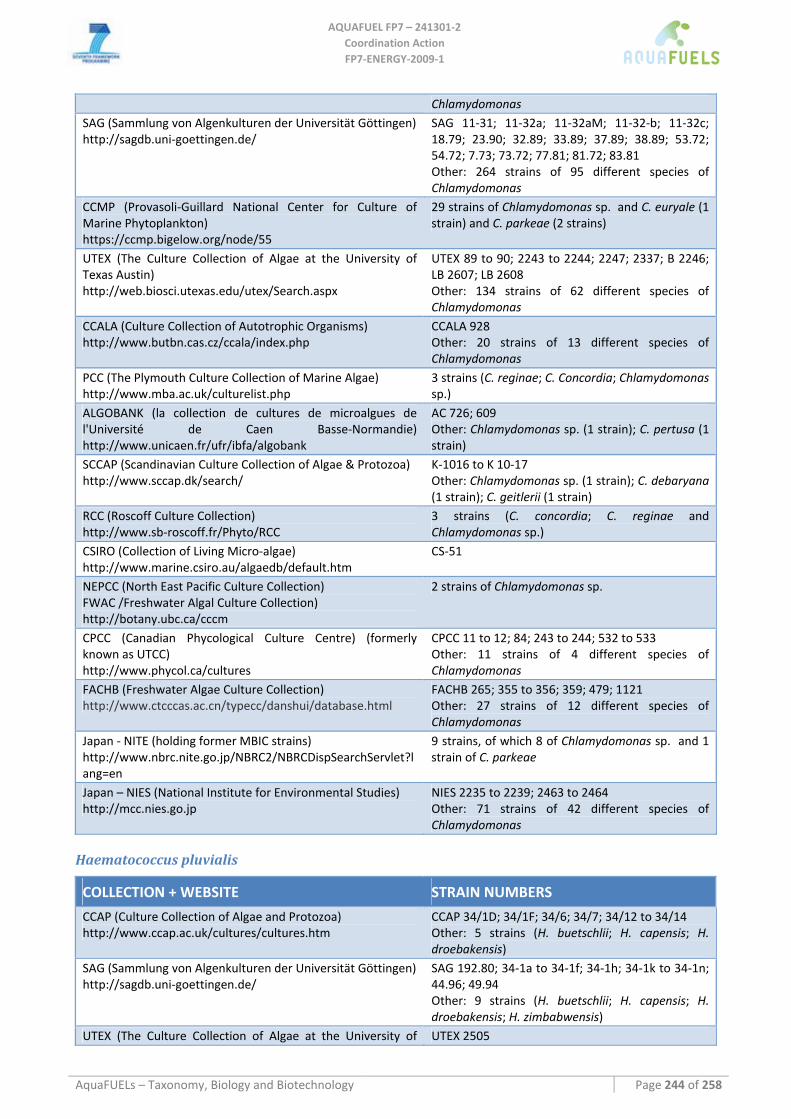
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 244 of 258
Chlamydomonas SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 11‐31; 11‐32a; 11‐32aM; 11‐32‐b; 11‐32c; 18.79; 23.90; 32.89; 33.89; 37.89; 38.89; 53.72; 54.72; 7.73; 73.72; 77.81; 81.72; 83.81 Other: 264 strains of 95 different species of Chlamydomonas
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
29 strains of Chlamydomonas sp. and C. euryale (1 strain) and C. parkeae (2 strains)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX 89 to 90; 2243 to 2244; 2247; 2337; B 2246; LB 2607; LB 2608 Other: 134 strains of 62 different species of Chlamydomonas
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 928 Other: 20 strains of 13 different species of Chlamydomonas
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
3 strains (C. reginae; C. Concordia; Chlamydomonas sp.)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 726; 609 Other: Chlamydomonas sp. (1 strain); C. pertusa (1 strain)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1016 to K 10‐17 Other: Chlamydomonas sp. (1 strain); C. debaryana (1 strain); C. geitlerii (1 strain)
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
3 strains (C. concordia; C. reginae and Chlamydomonas sp.)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐51
NEPCC (North East Pacific Culture Collection) FWAC /Freshwater Algal Culture Collection) http://botany.ubc.ca/cccm
2 strains of Chlamydomonas sp.
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 11 to 12; 84; 243 to 244; 532 to 533 Other: 11 strains of 4 different species of Chlamydomonas
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 265; 355 to 356; 359; 479; 1121 Other: 27 strains of 12 different species of Chlamydomonas
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
9 strains, of which 8 of Chlamydomonas sp. and 1 strain of C. parkeae
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 2235 to 2239; 2463 to 2464 Other: 71 strains of 42 different species of Chlamydomonas
Haematococcus pluvialis
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 34/1D; 34/1F; 34/6; 34/7; 34/12 to 34/14 Other: 5 strains (H. buetschlii; H. capensis; H. droebakensis)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 192.80; 34‐1a to 34‐1f; 34‐1h; 34‐1k to 34‐1n; 44.96; 49.94 Other: 9 strains (H. buetschlii; H. capensis; H. droebakensis; H. zimbabwensis)
UTEX (The Culture Collection of Algae at the University of UTEX 2505
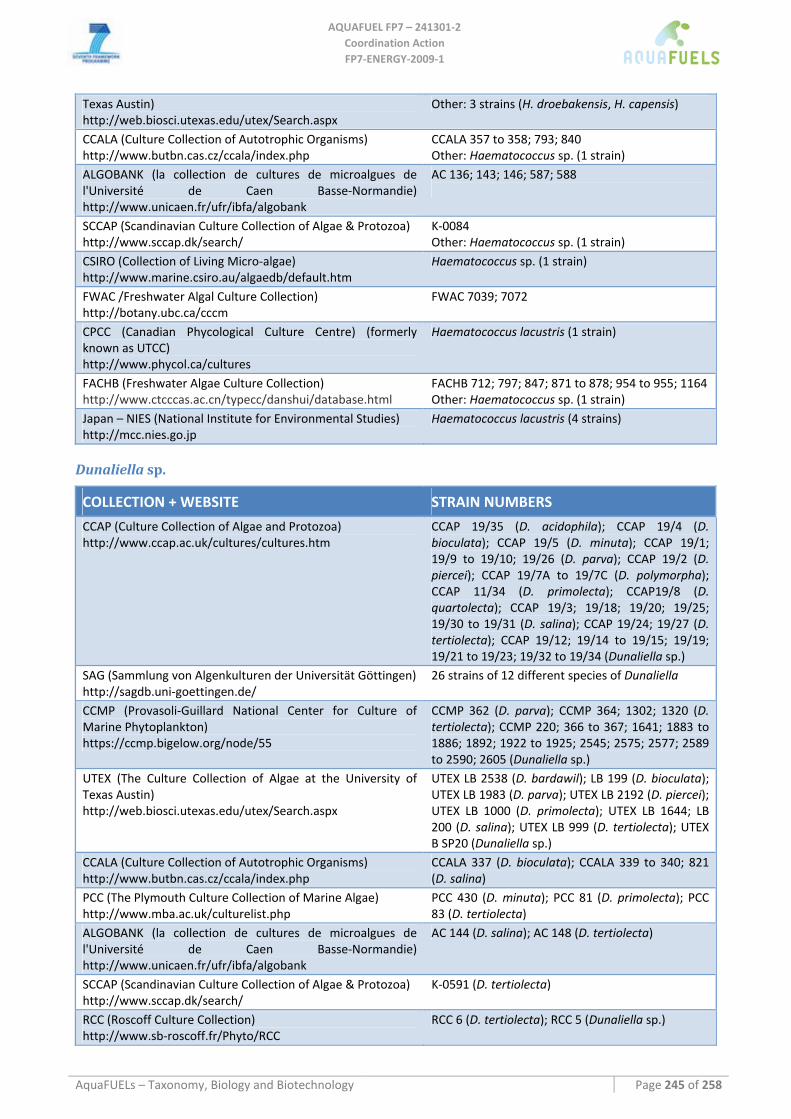
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 245 of 258
Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
Other: 3 strains (H. droebakensis, H. capensis)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 357 to 358; 793; 840 Other: Haematococcus sp. (1 strain)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 136; 143; 146; 587; 588
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐0084 Other: Haematococcus sp. (1 strain)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
Haematococcus sp. (1 strain)
FWAC /Freshwater Algal Culture Collection) http://botany.ubc.ca/cccm
FWAC 7039; 7072
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
Haematococcus lacustris (1 strain)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 712; 797; 847; 871 to 878; 954 to 955; 1164 Other: Haematococcus sp. (1 strain)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
Haematococcus lacustris (4 strains)
Dunaliella sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 19/35 (D. acidophila); CCAP 19/4 (D. bioculata); CCAP 19/5 (D. minuta); CCAP 19/1; 19/9 to 19/10; 19/26 (D. parva); CCAP 19/2 (D. piercei); CCAP 19/7A to 19/7C (D. polymorpha); CCAP 11/34 (D. primolecta); CCAP19/8 (D. quartolecta); CCAP 19/3; 19/18; 19/20; 19/25; 19/30 to 19/31 (D. salina); CCAP 19/24; 19/27 (D. tertiolecta); CCAP 19/12; 19/14 to 19/15; 19/19; 19/21 to 19/23; 19/32 to 19/34 (Dunaliella sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
26 strains of 12 different species of Dunaliella
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 362 (D. parva); CCMP 364; 1302; 1320 (D. tertiolecta); CCMP 220; 366 to 367; 1641; 1883 to 1886; 1892; 1922 to 1925; 2545; 2575; 2577; 2589 to 2590; 2605 (Dunaliella sp.)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX LB 2538 (D. bardawil); LB 199 (D. bioculata); UTEX LB 1983 (D. parva); UTEX LB 2192 (D. piercei); UTEX LB 1000 (D. primolecta); UTEX LB 1644; LB 200 (D. salina); UTEX LB 999 (D. tertiolecta); UTEX B SP20 (Dunaliella sp.)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 337 (D. bioculata); CCALA 339 to 340; 821 (D. salina)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 430 (D. minuta); PCC 81 (D. primolecta); PCC 83 (D. tertiolecta)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 144 (D. salina); AC 148 (D. tertiolecta)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐0591 (D. tertiolecta)
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
RCC 6 (D. tertiolecta); RCC 5 (Dunaliella sp.)
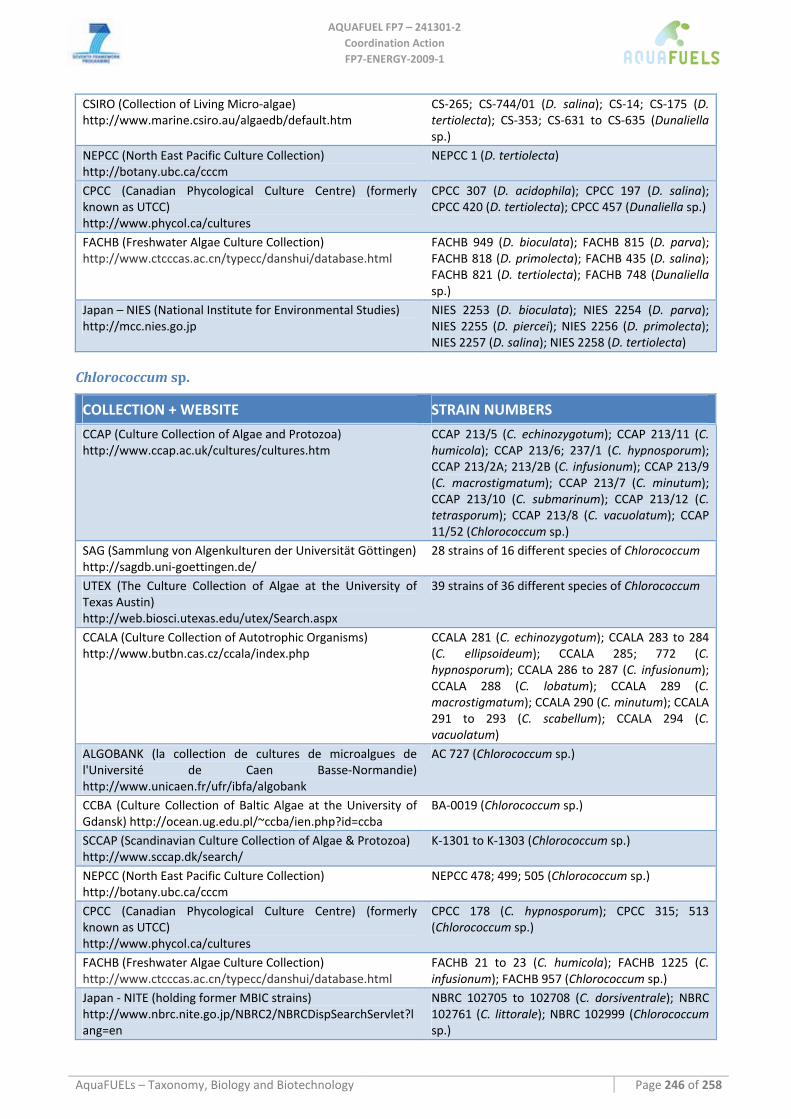
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 246 of 258
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐265; CS‐744/01 (D. salina); CS‐14; CS‐175 (D. tertiolecta); CS‐353; CS‐631 to CS‐635 (Dunaliella sp.)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 1 (D. tertiolecta)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 307 (D. acidophila); CPCC 197 (D. salina); CPCC 420 (D. tertiolecta); CPCC 457 (Dunaliella sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 949 (D. bioculata); FACHB 815 (D. parva); FACHB 818 (D. primolecta); FACHB 435 (D. salina); FACHB 821 (D. tertiolecta); FACHB 748 (Dunaliella sp.)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 2253 (D. bioculata); NIES 2254 (D. parva); NIES 2255 (D. piercei); NIES 2256 (D. primolecta); NIES 2257 (D. salina); NIES 2258 (D. tertiolecta)
Chlorococcum sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 213/5 (C. echinozygotum); CCAP 213/11 (C. humicola); CCAP 213/6; 237/1 (C. hypnosporum); CCAP 213/2A; 213/2B (C. infusionum); CCAP 213/9 (C. macrostigmatum); CCAP 213/7 (C. minutum); CCAP 213/10 (C. submarinum); CCAP 213/12 (C. tetrasporum); CCAP 213/8 (C. vacuolatum); CCAP 11/52 (Chlorococcum sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
28 strains of 16 different species of Chlorococcum
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
39 strains of 36 different species of Chlorococcum
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 281 (C. echinozygotum); CCALA 283 to 284 (C. ellipsoideum); CCALA 285; 772 (C. hypnosporum); CCALA 286 to 287 (C. infusionum); CCALA 288 (C. lobatum); CCALA 289 (C. macrostigmatum); CCALA 290 (C. minutum); CCALA 291 to 293 (C. scabellum); CCALA 294 (C. vacuolatum)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 727 (Chlorococcum sp.)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA‐0019 (Chlorococcum sp.)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1301 to K‐1303 (Chlorococcum sp.)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 478; 499; 505 (Chlorococcum sp.)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 178 (C. hypnosporum); CPCC 315; 513 (Chlorococcum sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 21 to 23 (C. humicola); FACHB 1225 (C. infusionum); FACHB 957 (Chlorococcum sp.)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102705 to 102708 (C. dorsiventrale); NBRC 102761 (C. littorale); NBRC 102999 (Chlorococcum sp.)
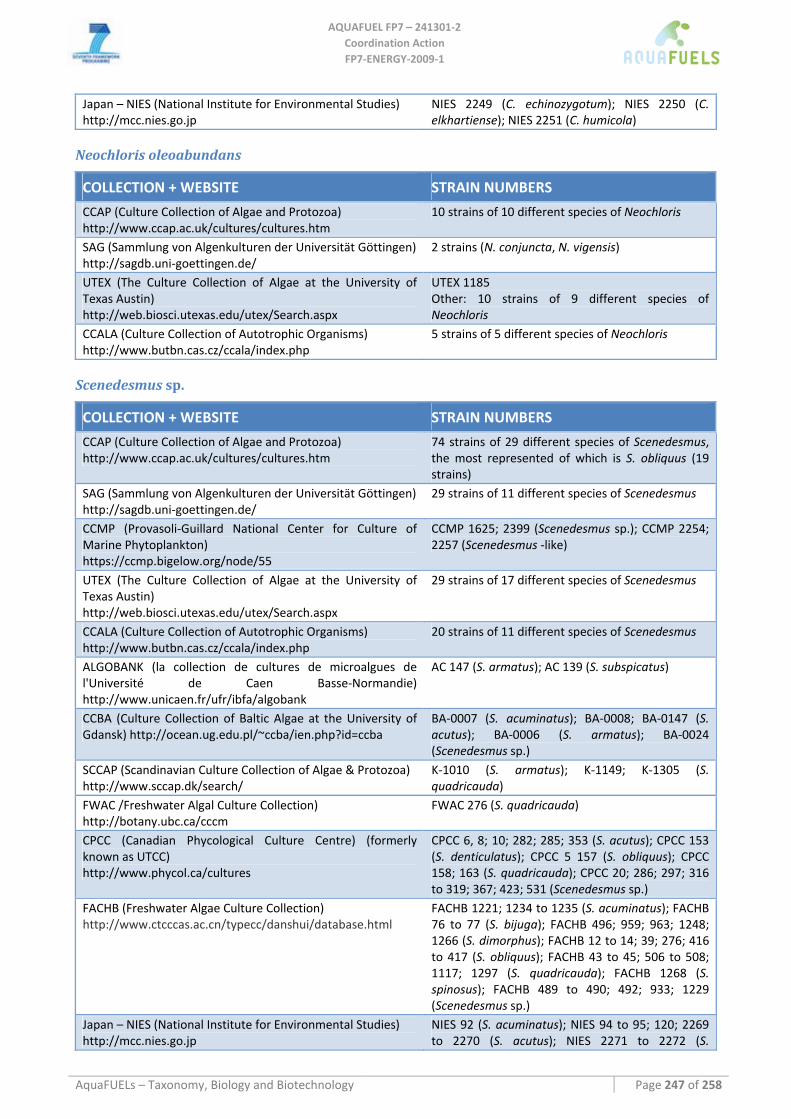
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 247 of 258
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 2249 (C. echinozygotum); NIES 2250 (C. elkhartiense); NIES 2251 (C. humicola)
Neochloris oleoabundans
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
10 strains of 10 different species of Neochloris
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
2 strains (N. conjuncta, N. vigensis)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX 1185 Other: 10 strains of 9 different species of Neochloris
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
5 strains of 5 different species of Neochloris
Scenedesmus sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
74 strains of 29 different species of Scenedesmus, the most represented of which is S. obliquus (19 strains)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
29 strains of 11 different species of Scenedesmus
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 1625; 2399 (Scenedesmus sp.); CCMP 2254; 2257 (Scenedesmus ‐like)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
29 strains of 17 different species of Scenedesmus
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
20 strains of 11 different species of Scenedesmus
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 147 (S. armatus); AC 139 (S. subspicatus)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA‐0007 (S. acuminatus); BA‐0008; BA‐0147 (S. acutus); BA‐0006 (S. armatus); BA‐0024 (Scenedesmus sp.)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1010 (S. armatus); K‐1149; K‐1305 (S. quadricauda)
FWAC /Freshwater Algal Culture Collection) http://botany.ubc.ca/cccm
FWAC 276 (S. quadricauda)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 6, 8; 10; 282; 285; 353 (S. acutus); CPCC 153 (S. denticulatus); CPCC 5 157 (S. obliquus); CPCC 158; 163 (S. quadricauda); CPCC 20; 286; 297; 316 to 319; 367; 423; 531 (Scenedesmus sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 1221; 1234 to 1235 (S. acuminatus); FACHB 76 to 77 (S. bijuga); FACHB 496; 959; 963; 1248; 1266 (S. dimorphus); FACHB 12 to 14; 39; 276; 416 to 417 (S. obliquus); FACHB 43 to 45; 506 to 508; 1117; 1297 (S. quadricauda); FACHB 1268 (S. spinosus); FACHB 489 to 490; 492; 933; 1229 (Scenedesmus sp.)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 92 (S. acuminatus); NIES 94 to 95; 120; 2269 to 2270 (S. acutus); NIES 2271 to 2272 (S.
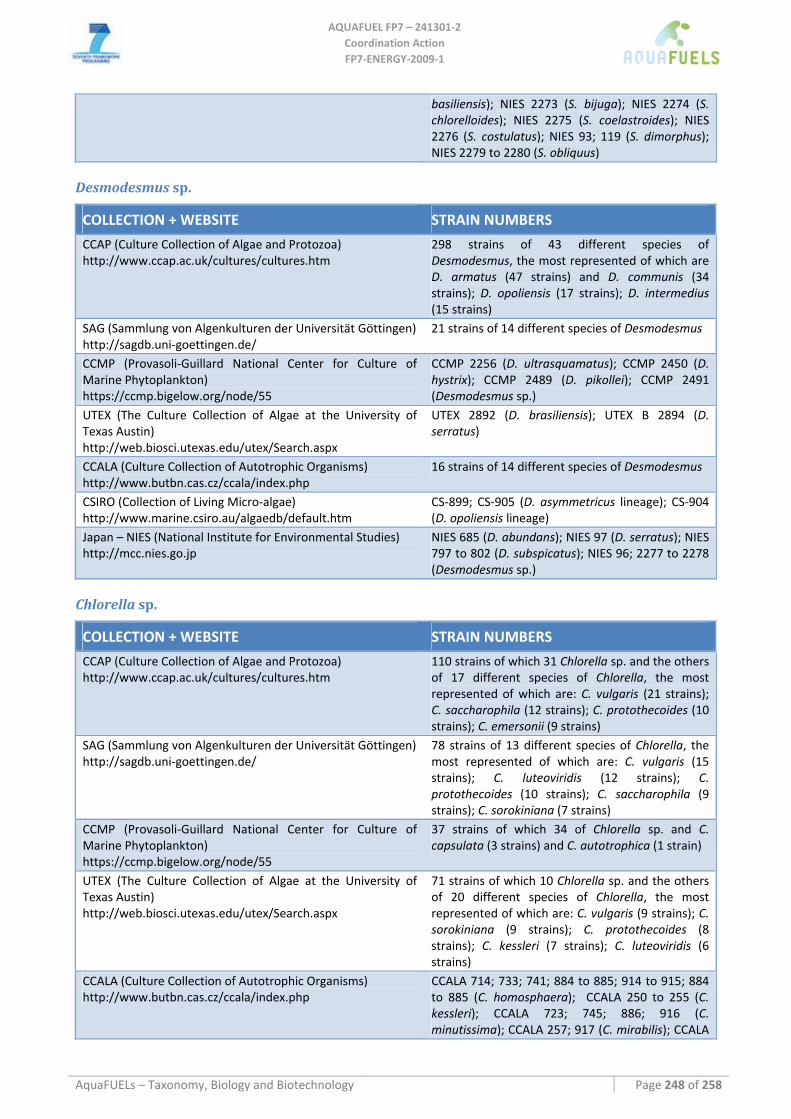
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 248 of 258
basiliensis); NIES 2273 (S. bijuga); NIES 2274 (S. chlorelloides); NIES 2275 (S. coelastroides); NIES 2276 (S. costulatus); NIES 93; 119 (S. dimorphus); NIES 2279 to 2280 (S. obliquus)
Desmodesmus sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
298 strains of 43 different species of Desmodesmus, the most represented of which are D. armatus (47 strains) and D. communis (34 strains); D. opoliensis (17 strains); D. intermedius (15 strains)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
21 strains of 14 different species of Desmodesmus
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 2256 (D. ultrasquamatus); CCMP 2450 (D. hystrix); CCMP 2489 (D. pikollei); CCMP 2491 (Desmodesmus sp.)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX 2892 (D. brasiliensis); UTEX B 2894 (D. serratus)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
16 strains of 14 different species of Desmodesmus
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐899; CS‐905 (D. asymmetricus lineage); CS‐904 (D. opoliensis lineage)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 685 (D. abundans); NIES 97 (D. serratus); NIES 797 to 802 (D. subspicatus); NIES 96; 2277 to 2278 (Desmodesmus sp.)
Chlorella sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
110 strains of which 31 Chlorella sp. and the others of 17 different species of Chlorella, the most represented of which are: C. vulgaris (21 strains); C. saccharophila (12 strains); C. protothecoides (10 strains); C. emersonii (9 strains)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
78 strains of 13 different species of Chlorella, the most represented of which are: C. vulgaris (15 strains); C. luteoviridis (12 strains); C. protothecoides (10 strains); C. saccharophila (9 strains); C. sorokiniana (7 strains)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
37 strains of which 34 of Chlorella sp. and C. capsulata (3 strains) and C. autotrophica (1 strain)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
71 strains of which 10 Chlorella sp. and the others of 20 different species of Chlorella, the most represented of which are: C. vulgaris (9 strains); C. sorokiniana (9 strains); C. protothecoides (8 strains); C. kessleri (7 strains); C. luteoviridis (6 strains)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 714; 733; 741; 884 to 885; 914 to 915; 884 to 885 (C. homosphaera); CCALA 250 to 255 (C. kessleri); CCALA 723; 745; 886; 916 (C. minutissima); CCALA 257; 917 (C. mirabilis); CCALA
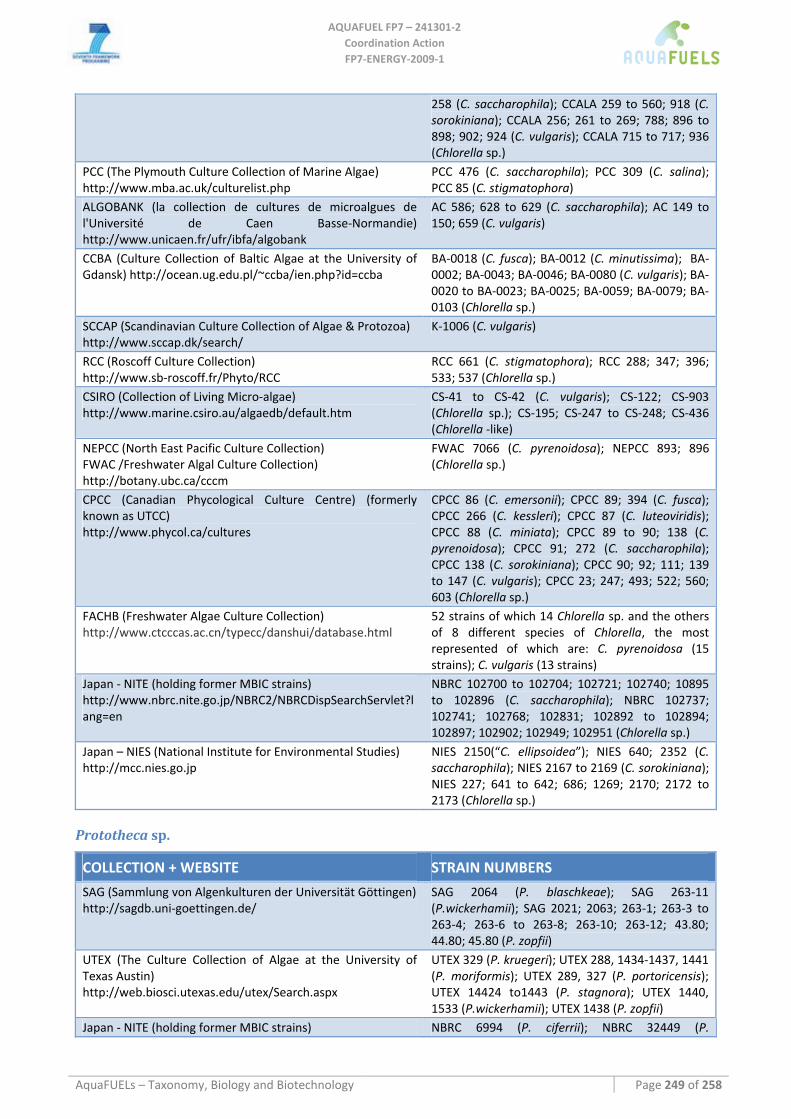
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 249 of 258
258 (C. saccharophila); CCALA 259 to 560; 918 (C. sorokiniana); CCALA 256; 261 to 269; 788; 896 to 898; 902; 924 (C. vulgaris); CCALA 715 to 717; 936 (Chlorella sp.)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 476 (C. saccharophila); PCC 309 (C. salina); PCC 85 (C. stigmatophora)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 586; 628 to 629 (C. saccharophila); AC 149 to 150; 659 (C. vulgaris)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA‐0018 (C. fusca); BA‐0012 (C. minutissima); BA‐0002; BA‐0043; BA‐0046; BA‐0080 (C. vulgaris); BA‐0020 to BA‐0023; BA‐0025; BA‐0059; BA‐0079; BA‐0103 (Chlorella sp.)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1006 (C. vulgaris)
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
RCC 661 (C. stigmatophora); RCC 288; 347; 396; 533; 537 (Chlorella sp.)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐41 to CS‐42 (C. vulgaris); CS‐122; CS‐903 (Chlorella sp.); CS‐195; CS‐247 to CS‐248; CS‐436 (Chlorella ‐like)
NEPCC (North East Pacific Culture Collection) FWAC /Freshwater Algal Culture Collection) http://botany.ubc.ca/cccm
FWAC 7066 (C. pyrenoidosa); NEPCC 893; 896 (Chlorella sp.)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 86 (C. emersonii); CPCC 89; 394 (C. fusca); CPCC 266 (C. kessleri); CPCC 87 (C. luteoviridis); CPCC 88 (C. miniata); CPCC 89 to 90; 138 (C. pyrenoidosa); CPCC 91; 272 (C. saccharophila); CPCC 138 (C. sorokiniana); CPCC 90; 92; 111; 139 to 147 (C. vulgaris); CPCC 23; 247; 493; 522; 560; 603 (Chlorella sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
52 strains of which 14 Chlorella sp. and the others of 8 different species of Chlorella, the most represented of which are: C. pyrenoidosa (15 strains); C. vulgaris (13 strains)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102700 to 102704; 102721; 102740; 10895 to 102896 (C. saccharophila); NBRC 102737; 102741; 102768; 102831; 102892 to 102894; 102897; 102902; 102949; 102951 (Chlorella sp.)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES 2150(“C. ellipsoidea”); NIES 640; 2352 (C. saccharophila); NIES 2167 to 2169 (C. sorokiniana); NIES 227; 641 to 642; 686; 1269; 2170; 2172 to 2173 (Chlorella sp.)
Prototheca sp.
COLLECTION + WEBSITE STRAIN NUMBERS
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 2064 (P. blaschkeae); SAG 263‐11 (P.wickerhamii); SAG 2021; 2063; 263‐1; 263‐3 to 263‐4; 263‐6 to 263‐8; 263‐10; 263‐12; 43.80; 44.80; 45.80 (P. zopfii)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX 329 (P. kruegeri); UTEX 288, 1434‐1437, 1441 (P. moriformis); UTEX 289, 327 (P. portoricensis); UTEX 14424 to1443 (P. stagnora); UTEX 1440, 1533 (P.wickerhamii); UTEX 1438 (P. zopfii)
Japan ‐ NITE (holding former MBIC strains) NBRC 6994 (P. ciferrii); NBRC 32449 (P.
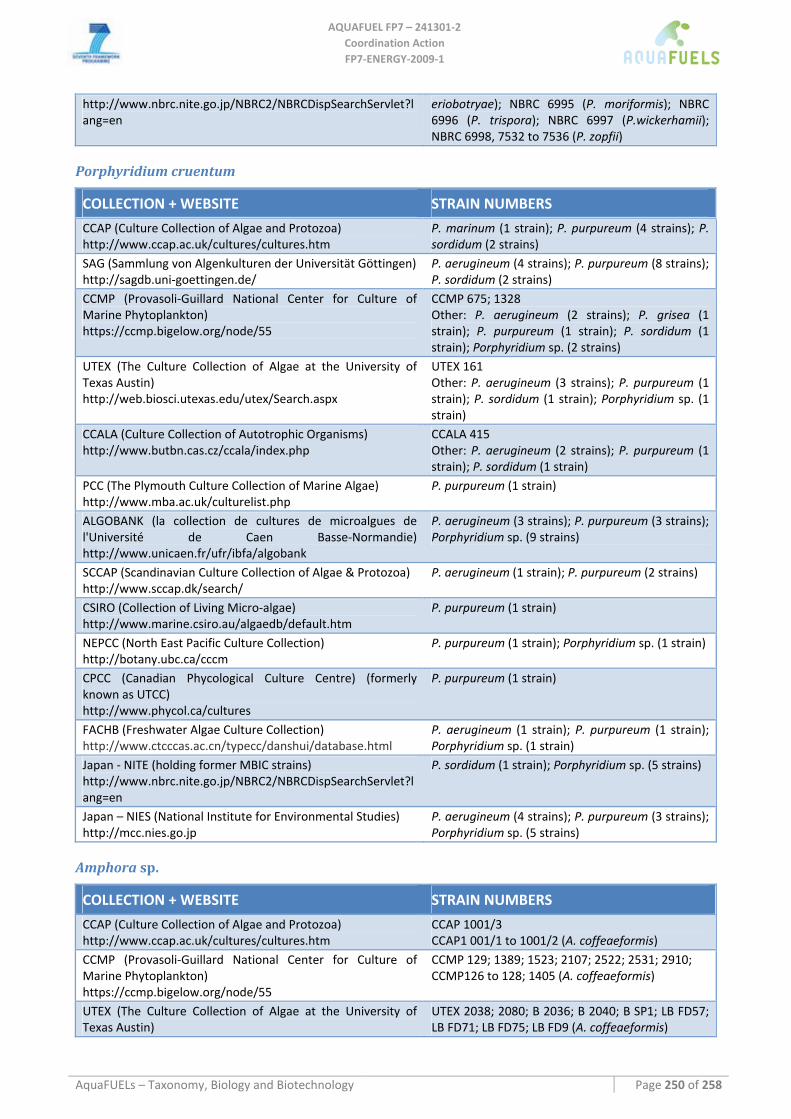
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 250 of 258
http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
eriobotryae); NBRC 6995 (P. moriformis); NBRC 6996 (P. trispora); NBRC 6997 (P.wickerhamii); NBRC 6998, 7532 to 7536 (P. zopfii)
Porphyridium cruentum
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
P. marinum (1 strain); P. purpureum (4 strains); P. sordidum (2 strains)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
P. aerugineum (4 strains); P. purpureum (8 strains); P. sordidum (2 strains)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 675; 1328 Other: P. aerugineum (2 strains); P. grisea (1 strain); P. purpureum (1 strain); P. sordidum (1 strain); Porphyridium sp. (2 strains)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX 161 Other: P. aerugineum (3 strains); P. purpureum (1 strain); P. sordidum (1 strain); Porphyridium sp. (1 strain)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 415 Other: P. aerugineum (2 strains); P. purpureum (1 strain); P. sordidum (1 strain)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
P. purpureum (1 strain)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
P. aerugineum (3 strains); P. purpureum (3 strains); Porphyridium sp. (9 strains)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
P. aerugineum (1 strain); P. purpureum (2 strains)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
P. purpureum (1 strain)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
P. purpureum (1 strain); Porphyridium sp. (1 strain)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
P. purpureum (1 strain)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
P. aerugineum (1 strain); P. purpureum (1 strain); Porphyridium sp. (1 strain)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
P. sordidum (1 strain); Porphyridium sp. (5 strains)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
P. aerugineum (4 strains); P. purpureum (3 strains); Porphyridium sp. (5 strains)
Amphora sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1001/3 CCAP1 001/1 to 1001/2 (A. coffeaeformis)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 129; 1389; 1523; 2107; 2522; 2531; 2910; CCMP126 to 128; 1405 (A. coffeaeformis)
UTEX (The Culture Collection of Algae at the University of Texas Austin)
UTEX 2038; 2080; B 2036; B 2040; B SP1; LB FD57; LB FD71; LB FD75; LB FD9 (A. coffeaeformis)
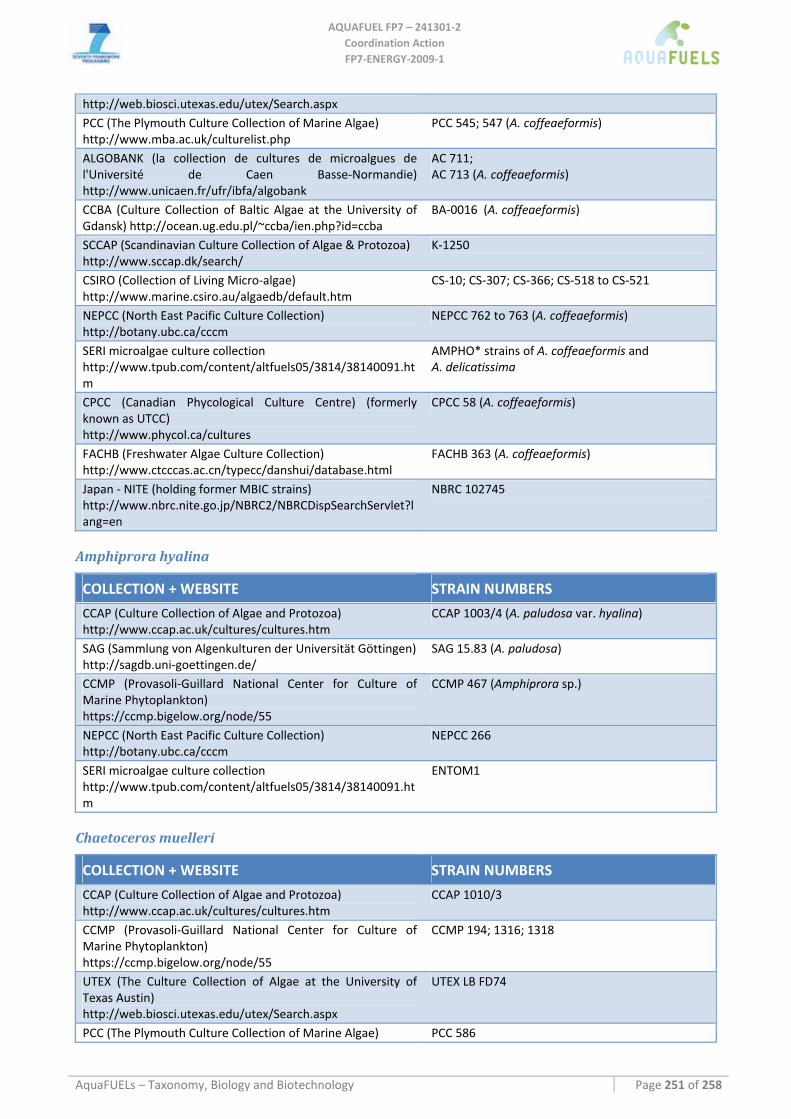
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 251 of 258
http://web.biosci.utexas.edu/utex/Search.aspx PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 545; 547 (A. coffeaeformis)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 711; AC 713 (A. coffeaeformis)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA‐0016 (A. coffeaeformis)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1250
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐10; CS‐307; CS‐366; CS‐518 to CS‐521
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 762 to 763 (A. coffeaeformis)
SERI microalgae culture collection http://www.tpub.com/content/altfuels05/3814/38140091.htm
AMPHO* strains of A. coffeaeformis and A. delicatissima
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 58 (A. coffeaeformis)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 363 (A. coffeaeformis)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102745
Amphiprora hyalina
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1003/4 (A. paludosa var. hyalina)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 15.83 (A. paludosa)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 467 (Amphiprora sp.)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 266
SERI microalgae culture collection http://www.tpub.com/content/altfuels05/3814/38140091.htm
ENTOM1
Chaetoceros muelleri
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1010/3
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 194; 1316; 1318
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX LB FD74
PCC (The Plymouth Culture Collection of Marine Algae) PCC 586
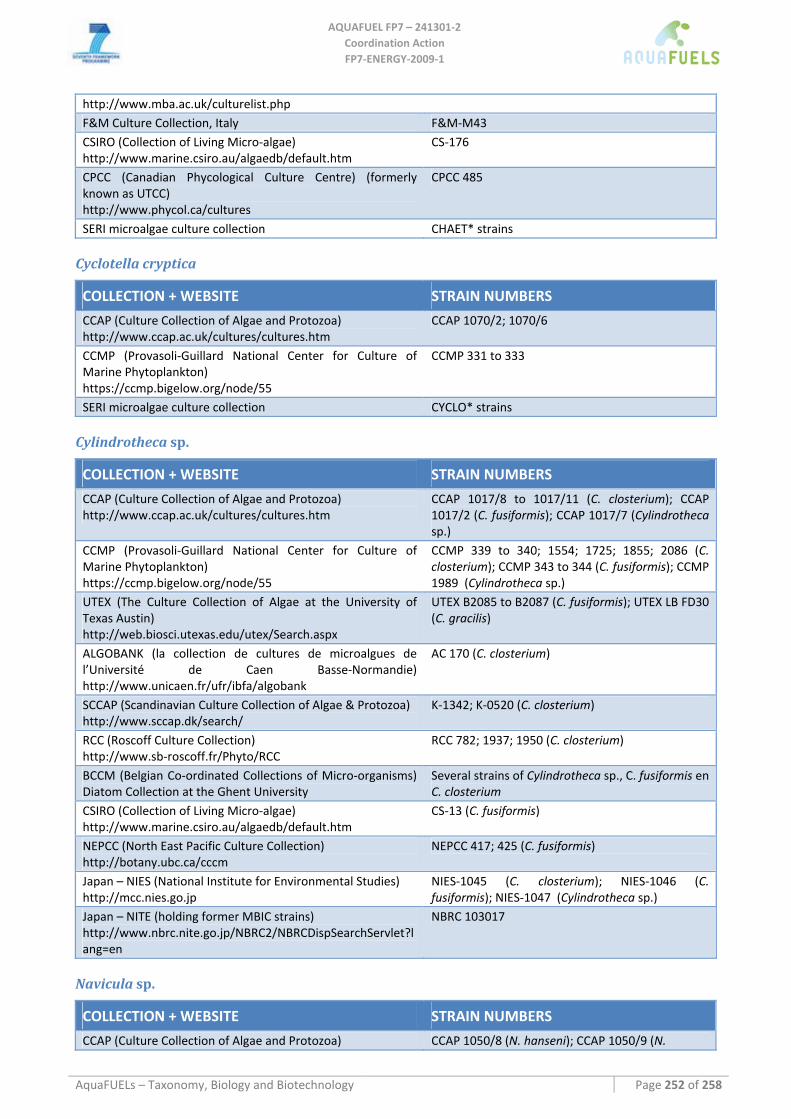
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 252 of 258
http://www.mba.ac.uk/culturelist.php F&M Culture Collection, Italy F&M‐M43 CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐176
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 485
SERI microalgae culture collection CHAET* strains
Cyclotella cryptica
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1070/2; 1070/6
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 331 to 333
SERI microalgae culture collection CYCLO* strains
Cylindrotheca sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1017/8 to 1017/11 (C. closterium); CCAP 1017/2 (C. fusiformis); CCAP 1017/7 (Cylindrotheca sp.)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 339 to 340; 1554; 1725; 1855; 2086 (C. closterium); CCMP 343 to 344 (C. fusiformis); CCMP 1989 (Cylindrotheca sp.)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX B2085 to B2087 (C. fusiformis); UTEX LB FD30 (C. gracilis)
ALGOBANK (la collection de cultures de microalgues de l’Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 170 (C. closterium)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1342; K‐0520 (C. closterium)
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
RCC 782; 1937; 1950 (C. closterium)
BCCM (Belgian Co‐ordinated Collections of Micro‐organisms) Diatom Collection at the Ghent University
Several strains of Cylindrotheca sp., C. fusiformis en C. closterium
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐13 (C. fusiformis)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 417; 425 (C. fusiformis)
Japan – NIES (National Institute for Environmental Studies) http://mcc.nies.go.jp
NIES‐1045 (C. closterium); NIES‐1046 (C. fusiformis); NIES‐1047 (Cylindrotheca sp.)
Japan – NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 103017
Navicula sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) CCAP 1050/8 (N. hanseni); CCAP 1050/9 (N.
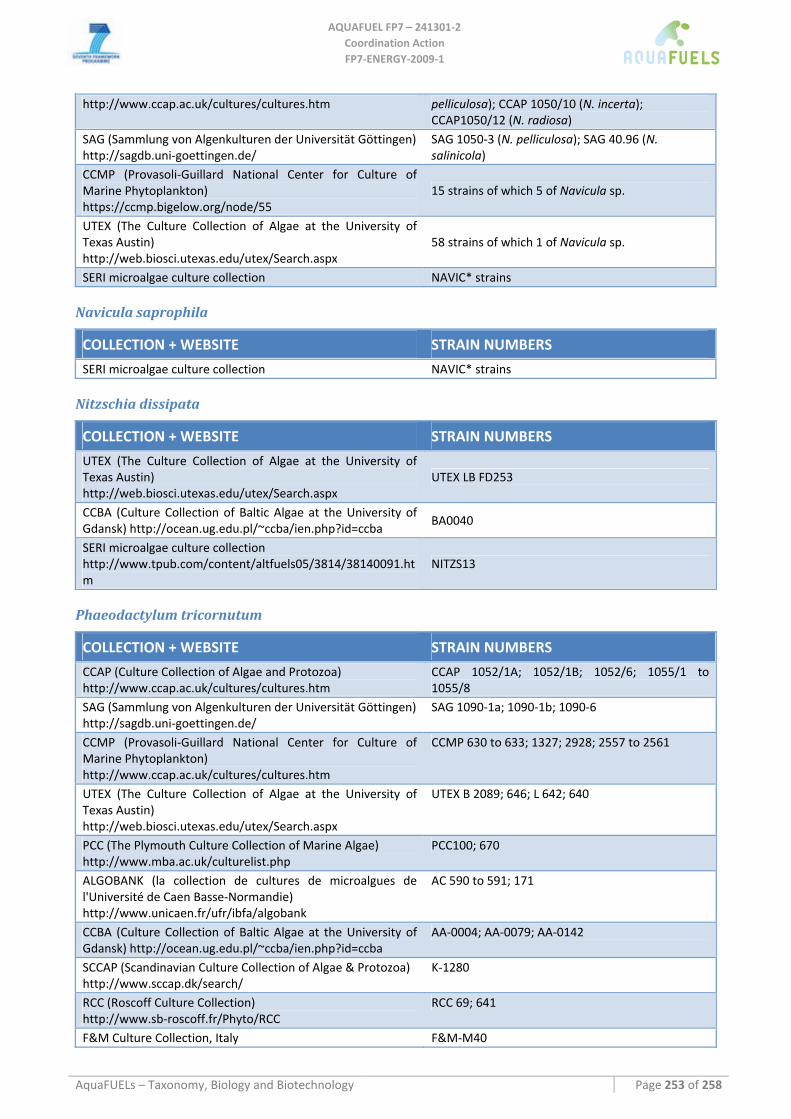
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 253 of 258
http://www.ccap.ac.uk/cultures/cultures.htm pelliculosa); CCAP 1050/10 (N. incerta); CCAP1050/12 (N. radiosa)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 1050‐3 (N. pelliculosa); SAG 40.96 (N. salinicola)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
15 strains of which 5 of Navicula sp.
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
58 strains of which 1 of Navicula sp.
SERI microalgae culture collection NAVIC* strains
Navicula saprophila
COLLECTION + WEBSITE STRAIN NUMBERS
SERI microalgae culture collection NAVIC* strains
Nitzschia dissipata
COLLECTION + WEBSITE STRAIN NUMBERS
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX LB FD253
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA0040
SERI microalgae culture collection http://www.tpub.com/content/altfuels05/3814/38140091.htm
NITZS13
Phaeodactylum tricornutum
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1052/1A; 1052/1B; 1052/6; 1055/1 to 1055/8
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 1090‐1a; 1090‐1b; 1090‐6
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) http://www.ccap.ac.uk/cultures/cultures.htm
CCMP 630 to 633; 1327; 2928; 2557 to 2561
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX B 2089; 646; L 642; 640
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC100; 670
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 590 to 591; 171
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
AA‐0004; AA‐0079; AA‐0142
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1280
RCC (Roscoff Culture Collection) http://www.sb‐roscoff.fr/Phyto/RCC
RCC 69; 641
F&M Culture Collection, Italy F&M‐M40
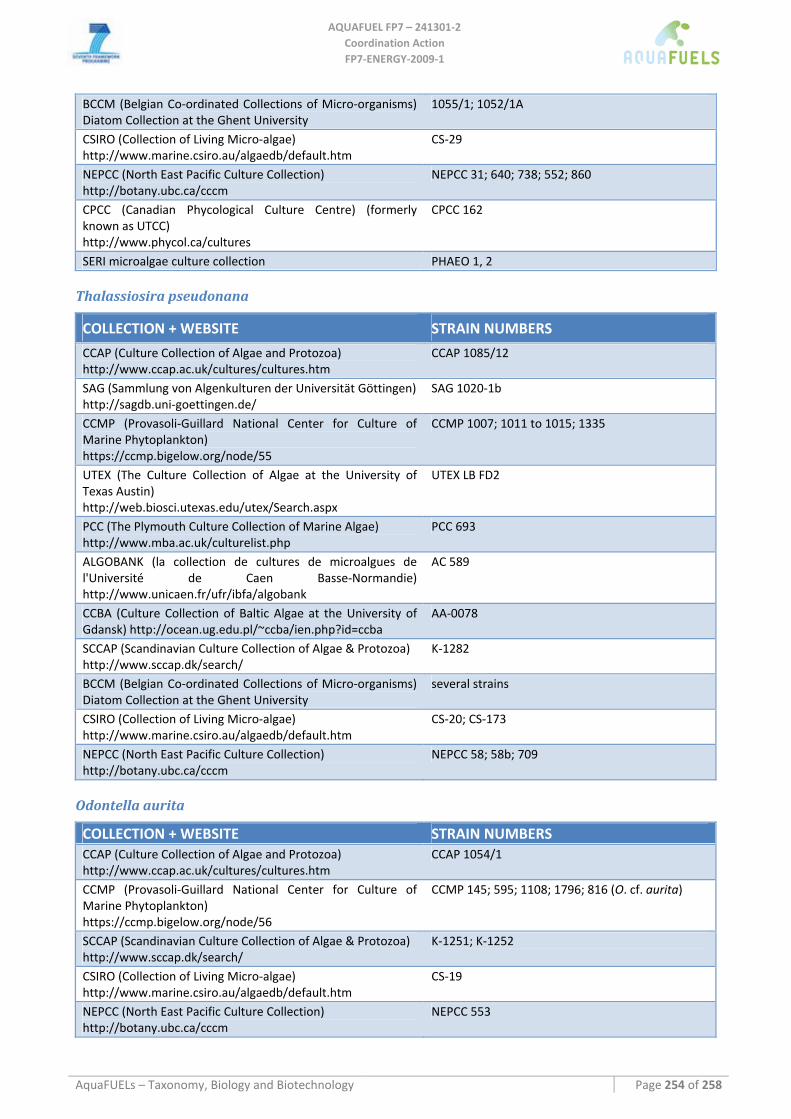
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 254 of 258
BCCM (Belgian Co‐ordinated Collections of Micro‐organisms) Diatom Collection at the Ghent University
1055/1; 1052/1A
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐29
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 31; 640; 738; 552; 860
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 162
SERI microalgae culture collection PHAEO 1, 2
Thalassiosira pseudonana
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1085/12
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 1020‐1b
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/55
CCMP 1007; 1011 to 1015; 1335
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX LB FD2
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 693
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 589
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
AA‐0078
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1282
BCCM (Belgian Co‐ordinated Collections of Micro‐organisms) Diatom Collection at the Ghent University
several strains
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐20; CS‐173
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 58; 58b; 709
Odontella aurita
COLLECTION + WEBSITE STRAIN NUMBERS CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1054/1
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/56
CCMP 145; 595; 1108; 1796; 816 (O. cf. aurita)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1251; K‐1252
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐19
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 553
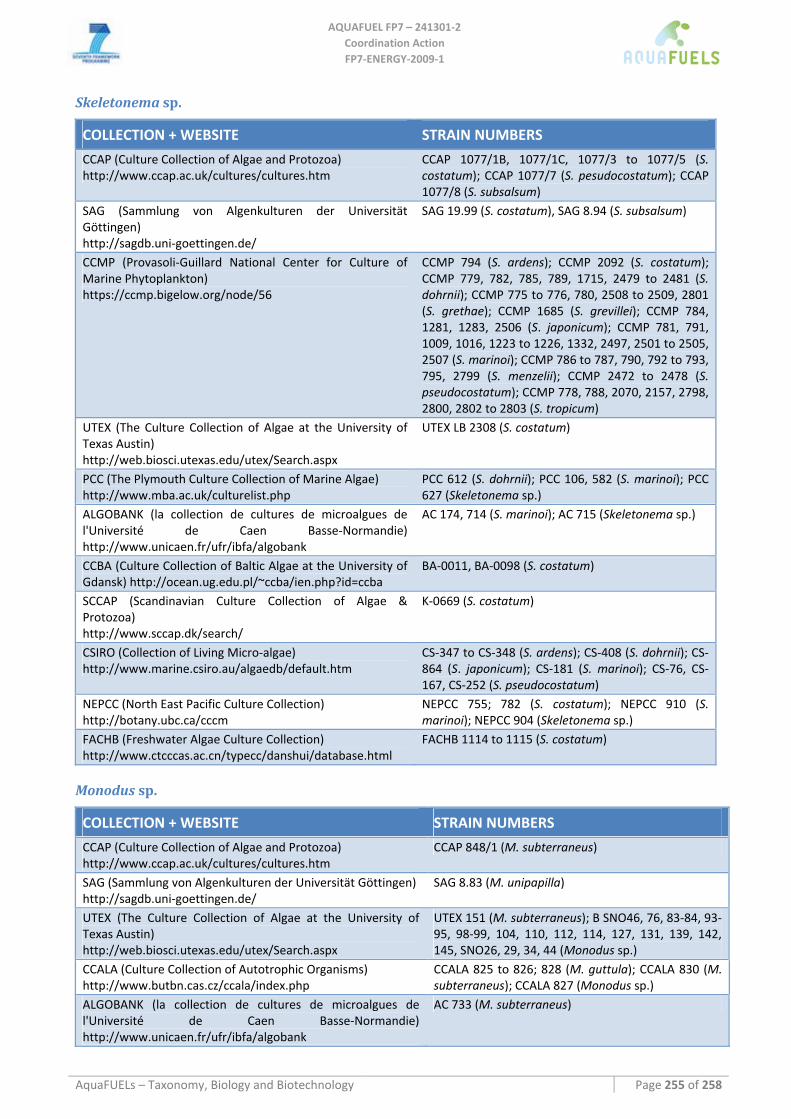
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 255 of 258
Skeletonema sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 1077/1B, 1077/1C, 1077/3 to 1077/5 (S. costatum); CCAP 1077/7 (S. pesudocostatum); CCAP 1077/8 (S. subsalsum)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 19.99 (S. costatum), SAG 8.94 (S. subsalsum)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/56
CCMP 794 (S. ardens); CCMP 2092 (S. costatum); CCMP 779, 782, 785, 789, 1715, 2479 to 2481 (S. dohrnii); CCMP 775 to 776, 780, 2508 to 2509, 2801 (S. grethae); CCMP 1685 (S. grevillei); CCMP 784, 1281, 1283, 2506 (S. japonicum); CCMP 781, 791, 1009, 1016, 1223 to 1226, 1332, 2497, 2501 to 2505, 2507 (S. marinoi); CCMP 786 to 787, 790, 792 to 793, 795, 2799 (S. menzelii); CCMP 2472 to 2478 (S. pseudocostatum); CCMP 778, 788, 2070, 2157, 2798, 2800, 2802 to 2803 (S. tropicum)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX LB 2308 (S. costatum)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 612 (S. dohrnii); PCC 106, 582 (S. marinoi); PCC 627 (Skeletonema sp.)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 174, 714 (S. marinoi); AC 715 (Skeletonema sp.)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
BA‐0011, BA‐0098 (S. costatum)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐0669 (S. costatum)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐347 to CS‐348 (S. ardens); CS‐408 (S. dohrnii); CS‐864 (S. japonicum); CS‐181 (S. marinoi); CS‐76, CS‐167, CS‐252 (S. pseudocostatum)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 755; 782 (S. costatum); NEPCC 910 (S. marinoi); NEPCC 904 (Skeletonema sp.)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 1114 to 1115 (S. costatum)
Monodus sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 848/1 (M. subterraneus)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 8.83 (M. unipapilla)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
UTEX 151 (M. subterraneus); B SNO46, 76, 83‐84, 93‐95, 98‐99, 104, 110, 112, 114, 127, 131, 139, 142, 145, SNO26, 29, 34, 44 (Monodus sp.)
CCALA (Culture Collection of Autotrophic Organisms) http://www.butbn.cas.cz/ccala/index.php
CCALA 825 to 826; 828 (M. guttula); CCALA 830 (M. subterraneus); CCALA 827 (Monodus sp.)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 733 (M. subterraneus)
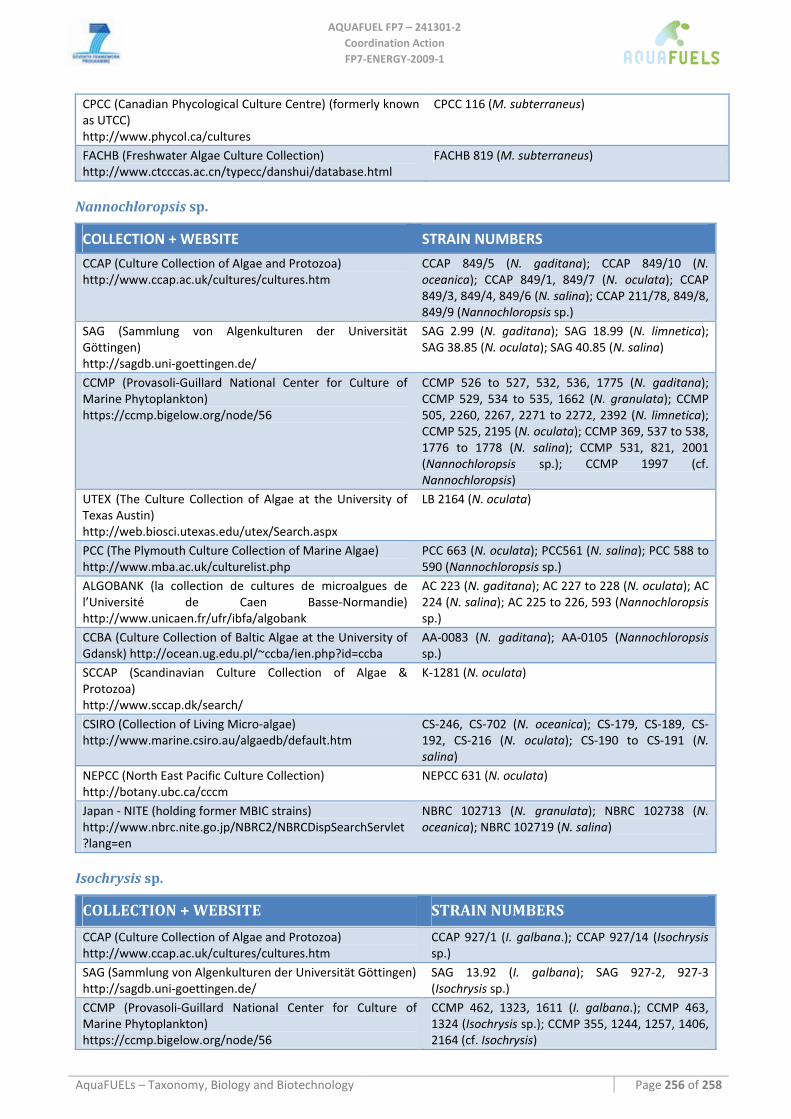
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 256 of 258
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 116 (M. subterraneus)
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 819 (M. subterraneus)
Nannochloropsis sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 849/5 (N. gaditana); CCAP 849/10 (N. oceanica); CCAP 849/1, 849/7 (N. oculata); CCAP 849/3, 849/4, 849/6 (N. salina); CCAP 211/78, 849/8, 849/9 (Nannochloropsis sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 2.99 (N. gaditana); SAG 18.99 (N. limnetica); SAG 38.85 (N. oculata); SAG 40.85 (N. salina)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/56
CCMP 526 to 527, 532, 536, 1775 (N. gaditana); CCMP 529, 534 to 535, 1662 (N. granulata); CCMP 505, 2260, 2267, 2271 to 2272, 2392 (N. limnetica); CCMP 525, 2195 (N. oculata); CCMP 369, 537 to 538, 1776 to 1778 (N. salina); CCMP 531, 821, 2001 (Nannochloropsis sp.); CCMP 1997 (cf. Nannochloropsis)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
LB 2164 (N. oculata)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 663 (N. oculata); PCC561 (N. salina); PCC 588 to 590 (Nannochloropsis sp.)
ALGOBANK (la collection de cultures de microalgues de l’Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 223 (N. gaditana); AC 227 to 228 (N. oculata); AC 224 (N. salina); AC 225 to 226, 593 (Nannochloropsis sp.)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
AA‐0083 (N. gaditana); AA‐0105 (Nannochloropsis sp.)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1281 (N. oculata)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐246, CS‐702 (N. oceanica); CS‐179, CS‐189, CS‐192, CS‐216 (N. oculata); CS‐190 to CS‐191 (N. salina)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 631 (N. oculata)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102713 (N. granulata); NBRC 102738 (N. oceanica); NBRC 102719 (N. salina)
Isochrysis sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 927/1 (I. galbana.); CCAP 927/14 (Isochrysis sp.)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 13.92 (I. galbana); SAG 927‐2, 927‐3 (Isochrysis sp.)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/56
CCMP 462, 1323, 1611 (I. galbana.); CCMP 463, 1324 (Isochrysis sp.); CCMP 355, 1244, 1257, 1406, 2164 (cf. Isochrysis)
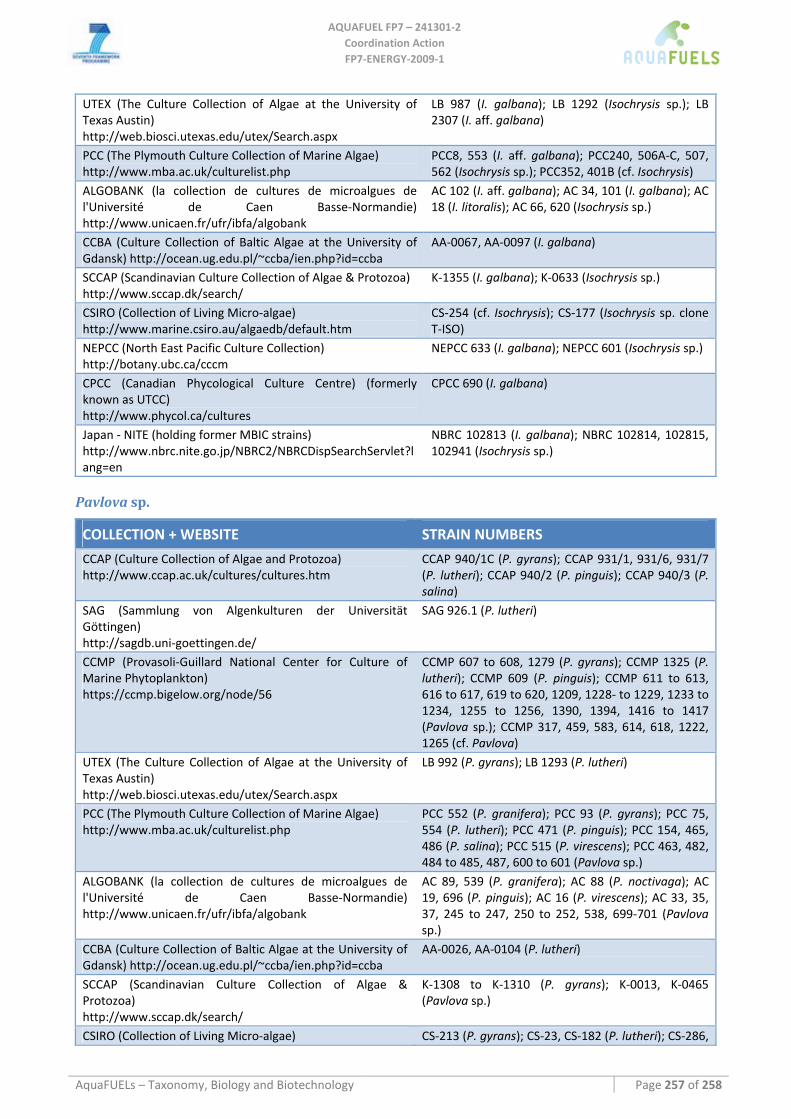
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 257 of 258
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
LB 987 (I. galbana); LB 1292 (Isochrysis sp.); LB 2307 (I. aff. galbana)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC8, 553 (I. aff. galbana); PCC240, 506A‐C, 507, 562 (Isochrysis sp.); PCC352, 401B (cf. Isochrysis)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 102 (I. aff. galbana); AC 34, 101 (I. galbana); AC 18 (I. litoralis); AC 66, 620 (Isochrysis sp.)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
AA‐0067, AA‐0097 (I. galbana)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1355 (I. galbana); K‐0633 (Isochrysis sp.)
CSIRO (Collection of Living Micro‐algae) http://www.marine.csiro.au/algaedb/default.htm
CS‐254 (cf. Isochrysis); CS‐177 (Isochrysis sp. clone T‐ISO)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 633 (I. galbana); NEPCC 601 (Isochrysis sp.)
CPCC (Canadian Phycological Culture Centre) (formerly known as UTCC) http://www.phycol.ca/cultures
CPCC 690 (I. galbana)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102813 (I. galbana); NBRC 102814, 102815, 102941 (Isochrysis sp.)
Pavlova sp.
COLLECTION + WEBSITE STRAIN NUMBERS
CCAP (Culture Collection of Algae and Protozoa) http://www.ccap.ac.uk/cultures/cultures.htm
CCAP 940/1C (P. gyrans); CCAP 931/1, 931/6, 931/7 (P. lutheri); CCAP 940/2 (P. pinguis); CCAP 940/3 (P. salina)
SAG (Sammlung von Algenkulturen der Universität Göttingen) http://sagdb.uni‐goettingen.de/
SAG 926.1 (P. lutheri)
CCMP (Provasoli‐Guillard National Center for Culture of Marine Phytoplankton) https://ccmp.bigelow.org/node/56
CCMP 607 to 608, 1279 (P. gyrans); CCMP 1325 (P. lutheri); CCMP 609 (P. pinguis); CCMP 611 to 613, 616 to 617, 619 to 620, 1209, 1228‐ to 1229, 1233 to 1234, 1255 to 1256, 1390, 1394, 1416 to 1417 (Pavlova sp.); CCMP 317, 459, 583, 614, 618, 1222, 1265 (cf. Pavlova)
UTEX (The Culture Collection of Algae at the University of Texas Austin) http://web.biosci.utexas.edu/utex/Search.aspx
LB 992 (P. gyrans); LB 1293 (P. lutheri)
PCC (The Plymouth Culture Collection of Marine Algae) http://www.mba.ac.uk/culturelist.php
PCC 552 (P. granifera); PCC 93 (P. gyrans); PCC 75, 554 (P. lutheri); PCC 471 (P. pinguis); PCC 154, 465, 486 (P. salina); PCC 515 (P. virescens); PCC 463, 482, 484 to 485, 487, 600 to 601 (Pavlova sp.)
ALGOBANK (la collection de cultures de microalgues de l'Université de Caen Basse‐Normandie) http://www.unicaen.fr/ufr/ibfa/algobank
AC 89, 539 (P. granifera); AC 88 (P. noctivaga); AC 19, 696 (P. pinguis); AC 16 (P. virescens); AC 33, 35, 37, 245 to 247, 250 to 252, 538, 699‐701 (Pavlova sp.)
CCBA (Culture Collection of Baltic Algae at the University of Gdansk) http://ocean.ug.edu.pl/~ccba/ien.php?id=ccba
AA‐0026, AA‐0104 (P. lutheri)
SCCAP (Scandinavian Culture Collection of Algae & Protozoa) http://www.sccap.dk/search/
K‐1308 to K‐1310 (P. gyrans); K‐0013, K‐0465 (Pavlova sp.)
CSIRO (Collection of Living Micro‐algae) CS‐213 (P. gyrans); CS‐23, CS‐182 (P. lutheri); CS‐286,
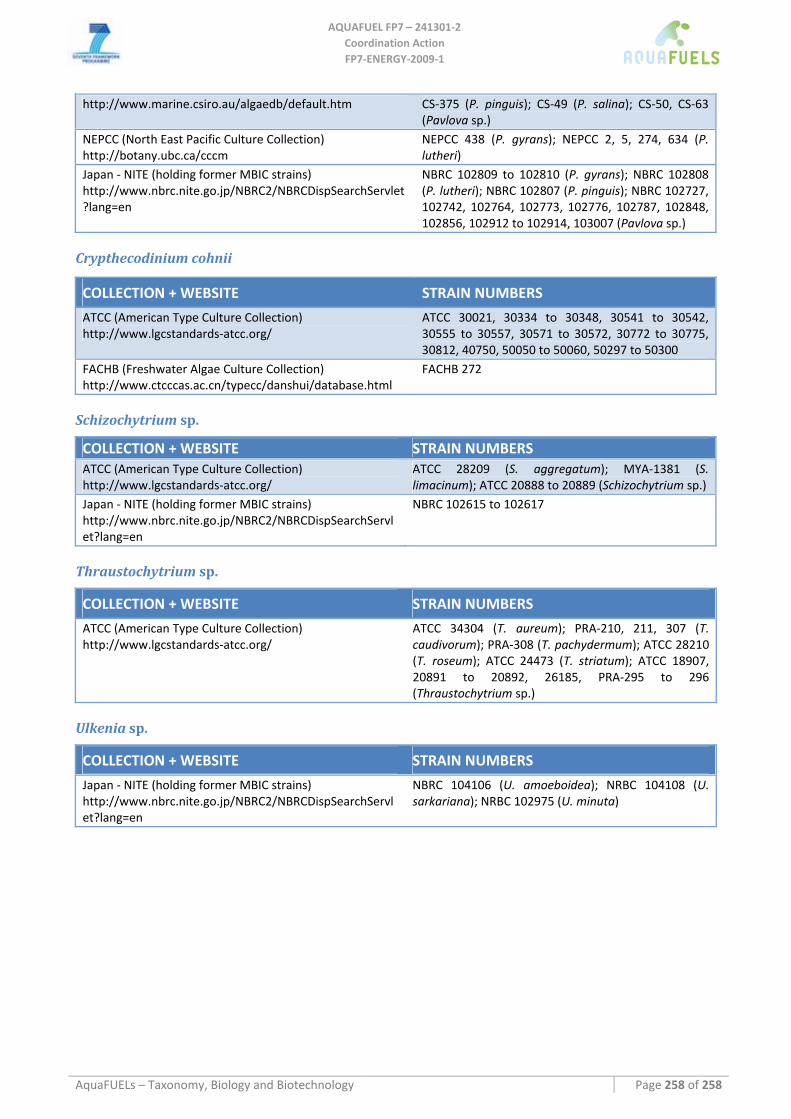
AQUAFUEL FP7 – 241301‐2
Coordination Action FP7‐ENERGY‐2009‐1
AquaFUELs – Taxonomy, Biology and Biotechnology Page 258 of 258
http://www.marine.csiro.au/algaedb/default.htm CS‐375 (P. pinguis); CS‐49 (P. salina); CS‐50, CS‐63 (Pavlova sp.)
NEPCC (North East Pacific Culture Collection) http://botany.ubc.ca/cccm
NEPCC 438 (P. gyrans); NEPCC 2, 5, 274, 634 (P. lutheri)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102809 to 102810 (P. gyrans); NBRC 102808 (P. lutheri); NBRC 102807 (P. pinguis); NBRC 102727, 102742, 102764, 102773, 102776, 102787, 102848, 102856, 102912 to 102914, 103007 (Pavlova sp.)
Crypthecodinium cohnii
COLLECTION + WEBSITE STRAIN NUMBERS
ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 30021, 30334 to 30348, 30541 to 30542, 30555 to 30557, 30571 to 30572, 30772 to 30775, 30812, 40750, 50050 to 50060, 50297 to 50300
FACHB (Freshwater Algae Culture Collection) http://www.ctcccas.ac.cn/typecc/danshui/database.html
FACHB 272
Schizochytrium sp.
COLLECTION + WEBSITE STRAIN NUMBERS ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 28209 (S. aggregatum); MYA‐1381 (S. limacinum); ATCC 20888 to 20889 (Schizochytrium sp.)
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 102615 to 102617
Thraustochytrium sp.
COLLECTION + WEBSITE STRAIN NUMBERS
ATCC (American Type Culture Collection) http://www.lgcstandards‐atcc.org/
ATCC 34304 (T. aureum); PRA‐210, 211, 307 (T. caudivorum); PRA‐308 (T. pachydermum); ATCC 28210 (T. roseum); ATCC 24473 (T. striatum); ATCC 18907, 20891 to 20892, 26185, PRA‐295 to 296 (Thraustochytrium sp.)
Ulkenia sp.
COLLECTION + WEBSITE STRAIN NUMBERS
Japan ‐ NITE (holding former MBIC strains) http://www.nbrc.nite.go.jp/NBRC2/NBRCDispSearchServlet?lang=en
NBRC 104106 (U. amoeboidea); NRBC 104108 (U. sarkariana); NRBC 102975 (U. minuta)















![Algae Cultures to Biofuels[1]](https://img.pdfslide.us/doc/110x75/577d36271a28ab3a6b925455/algae-cultures-to-biofuels1.jpg)













