Embed Size (px)
Citation preview

R E S E A R C H L E T T E R
Atransposable class I composite transposon carryingmph (methylparathion hydrolase) fromPseudomonas sp. strainWBC-3Min Wei1,2, Jun-Jie Zhang1, Hong Liu1, Shu-Jun Wang1, He Fu1,2 & Ning-Yi Zhou1
1State Key Laboratory of Virology, Wuhan Institute of Virology, Chinese Academy of Sciences, Wuhan, China; and 2Graduate School, Chinese Academy
of Sciences, Beijing, China
Correspondence: Ning-Yi Zhou, State Key
Laboratory of Virology, Wuhan Institute of
Virology, Chinese Academy of Sciences,
Wuhan 430071, China. Tel.: 186 27
87197655; fax: 186 27 87197655; e-mail:
Received 16 October 2008; accepted 5
December 2008.
First published online 7 January 2009.
DOI:10.1111/j.1574-6968.2008.01468.x
Editor: Hans-Peter Kohler
Keywords
IS6100 ; methyl parathion hydrolase;
Pseudomonas ; transposon.
Abstract
Pseudomonas sp. strain WBC-3 utilizes methyl parathion (O,O-dimethyl O-p-
nitrophenol phosphorothioate) or para-nitrophenol as the sole source of carbon,
nitrogen and energy. A gene encoding methyl parathion hydrolase (MPH) had
been characterized previously and found to be located on a typical class I
composite transposon that comprised IS6100 (Tnmph). In this study, the
transposability of this transposon was confirmed by transposition assays in two
distinct mating-out systems. Tnmph was demonstrated to transpose efficiently in a
random manner in Pseudomonas putida PaW340 by Southern blot and in Ralstonia
sp. U2 by sequence analysis of the Tnmph insertion sites, both exhibiting MPH
activity. The linkage of the mph-like gene with IS6100, together with the
transposability of Tnmph, as well as its capability to transpose in other phylogen-
etically divergent bacterial species, suggest that Tnmph may contribute to the wide
distribution of mph-like genes and the adaptation of bacteria to organopho-
sphorus compounds.
Introduction
Two types of organophosphorus degradation genes have
been identified in a number of bacterial strains isolated from
diverse geographical regions. The organophosphate degra-
dation gene (opd) was first identified in Flavobacterium sp.
ATCC27551 (Mulbry & Karns, 1989) and Pseudomonas
diminuta MG (Serdar et al., 1989), which were isolated from
the Philippines and Texas, respectively, in the late 1980s.
Recently, a methyl parathion (O,O-dimethyl O-p-nitrophe-
nol phosphorothioate) degradation gene (mph or mpd) was
reported in the Pseudomonas sp. strain WBC-3 (Liu et al.,
2005) and Plesiomonas sp. strain M6 (Zhongli et al., 2001)
isolated in China, but it showed only 12% identity to OPD
at the amino acid level (Liu et al., 2005). Subsequently, mph-
like genes have also been identified in seven other organo-
phosphorus pesticide-degrading strains of four different
genera at different locations in China (Zhang et al., 2006).
It has been demonstrated that bacterial genes for the
degradation of various xenobiotic compounds were often
located on transposable elements, resulting in the wide
distribution and rapid evolution of common catabolic path-
ways for microorganisms in the environment (Tan, 1999;
Top & Springael, 2003). opd was found to be flanked by one
copy of ISFlsp1 (a member of the IS21 family) and one copy
of a Tn3-like element encoding a transposase and a resolvase
in Flavobacterium sp., but no transposition events were
detected in a transposition study (Siddavattam et al., 2003).
However, because transposition is a complicated process,
this might be due to some unknown factors such as
nutritional or environmental conditions, or other genetic
factors present only in the natural host (Siddavattam et al.,
2003). opdA of Agrobacterium radiobacter P230 (Horne
et al., 2002), which shares 88% identity with opd at the
nucleotide sequence level, was located within an insertion
sequence derivative (TnopdA), whose transposase gene and
inverted repeat sequences are identical to those of IS6100
(Horne et al., 2003). It was demonstrated to be transposable
by an assay using an artificial transposable element with
disruption of the opdA gene (Horne et al., 2003), but no
attempt was made to investigate whether it jumped into the
recipients’ genome in a random manner.
Of all the strains that contain either type of organopho-
sphorus degradation genes, Pseudomonas sp. strain WBC-3
is the only one that is capable of utilizing methyl parathion
or para-nitrophenol as the sole source of carbon, nitrogen
and energy. The gene encoding methyl parathion hydrolase
(MPH, E.C.3.1.8.1), which catalyzes the hydrolyzation of
FEMS Microbiol Lett 292 (2009) 85–91 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved

methyl parathion to para-nitrophenol, was functionally
expressed (Liu et al., 2005), and the three-dimensional
structure of this enzyme was also resolved (Dong et al.,
2005). mph was localized to a typical class I composite
transposon (Tnmph), flanked by two isocopies of IS6100 at
each end in the same direction (Liu et al., 2005) (GenBank
accession no. AY251554), as shown in Fig. 1. In this study,
we report the transposability of this transposon and eluci-
date its potential role in horizontal gene transfer during the
evolution of microbial OPD.
Materials and methods
Bacterial strains, plasmids, and cultureconditions
The bacterial strains and plasmids used in this study are
listed in Table 1. Escherichia coli was routinely cultivated at
37 1C in Luria–Bertani (LB) media or on LB plates solidified
with 1.5% agar. Pseudomonas and Ralstonia strains were
cultured at 30 1C.
Streptomycin (1 mg mL�1 for selection of Pseudomonas
putida PaW340 or 100mg mL�1 for E. coli HB101), ampicillin
(100mg mL�1 for selection of Ralstonia sp. U2), tetracycline
(20mg mL�1), kanamycin (45mg mL�1), chloramphenicol
(34mg mL�1) or trimethoprim (50mg mL�1) was added to
the growth media, when necessary.
DNA manipulation and construction of plasmids
Established protocols were used for plasmid DNA prepara-
tion and manipulation (Sambrook et al., 1989). Genomic
DNA was isolated from strain PaW340 using the previously
described procedures (Ausubel, 2001). A 6.5 kb KpnI/Bam-
HI fragment carrying the complete Tnmph (Liu et al., 2005)
from strain WBC-3 was cloned into pEX18Tc to produce
pZWWM001. A kanamycin resistance gene (nptII), as a
selective marker, was amplified from plasposon pTnMod-
Okm (Dennis & Zylstra, 1998) and subsequently inserted
into the NotI site immediately downstream of mph in
pZWWM001 to produce pZWWM002, leaving the mph
gene intact. The physical map of pZWWM002 is illustrated
in Fig. 1. The KpnI/BamHI fragment of pZWWM002
carrying Tnmph with the nptII insertion was cloned into
pSTV29 to produce pZWWM003.
Transposition assay
The transposition of Tnmph in pZWWM002 was assayed by
biparental mating, as described previously (Williams et al.,
2002). Escherichia coli S17-1 (Simon et al., 1983) harboring
Fig. 1. Replicative transposition of Tnmph from
pZWWM002. Transposase-mediated fusion of
donor and target molecules generates a third
copy of IS6100 in the same orientation as the
original pair. The two possible results of trans-
position, depending on which IS6100 copy is
duplicated, are shown below the physical map
of pZWWM002. The solid and dotted lines in
pZWWM002 are, respectively, representative of
a 458-bp internal IS6100 fragment used as the
IS6100 probe (probe 1) and a 559-bp internal
fragment of sacB as the pEX18Tc probe (probe
2). In hybridization with the IS6100 probe, three
hybridized bands are expected: the 3.3 and
1.2-kb bands identical to those in the plasmid
pZWWM002 and a third one of variable size in
each independent transconjugant. In the case of
pEX18Tc probe, an 8.6-kb band was detected,
corresponding to that illustrated in Fig. 2b and d.
Primers P1, P2, P3 and P4 used for genome
walking are also indicated. mph, methyl para-
thion hydrolase gene; sacB, levansucrase gene;
tet, tetracycline resistant gene; nptII, kanamycin
resistant gene; P, PstI; K, KpnI; B, BamHI.
FEMS Microbiol Lett 292 (2009) 85–91c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
86 M. Wei et al.

pZWWM002 was used as the donor strain and the plasmid-
free strain PaW340 (Jeenes & Williams, 1982), in which
pEX18Tc or its derivatives do not replicate, was used as the
recipient strain. The resulting transconjugants were screened
on LB plates containing 1 mg mL�1 streptomycin (selection
for strain PaW340) and 45 mg mL�1 kanamycin (selection for
Tnmph with nptII). The frequency of conjugation plus
transposition was calculated as the ratio of transconjugants
to recipients.
The transposition of a Tnmph derivative (kanamycin
resistant, Kmr) was also examined using a mating-out
experiment, as described previously (Tsuda & Iino, 1987;
Sota et al., 2002). For this purpose, E. coli DH5a harboring
pZWWM003 and plasmid R388 (trimethoprim resistant,
Tpr) (Ward & Grinsted, 1982), a conjugal plasmid free of
transposon, was used as the donor to mate with E. coli
HB101 (streptomycin resistant, Smr) on a membrane filter.
The transposition frequency was expressed as the number of
KmrSmr transconjugants per Tpr Smr transconjugants and
the plasmids in the transconjugants were further character-
ized by restriction digestion.
Conjugation assays were also performed using Ralstonia
sp. strain U2 (Zhou et al., 2001), as the recipient. Constructs
based on pEX18Tc were confirmed not to replicate in U2
before this conjugation. The resulting transconjugants were
screened on LB plates containing 100 mg mL�1 ampicillin
(selection for strain U2) and 45mg mL�1 kanamycin (selec-
tion for Tnmph with nptII).
Southern hybridization analysis
Genomic DNA from the PaW340 strain and transconjugants
was digested with PstI and subjected to 0.7% agarose gel
electrophoresis before the separated DNA fragments were
transferred from agarose gels to a positively charged nylon
membrane (Boehringer Mannheim, Germany). DNA probes
were prepared by PCR amplification with pZWWM002 as
the template, and hybridization was performed using the
DIG High Prime DNA Labeling and Detection Starter Kit I
(Roche Diagnostics, Germany) according to the manufac-
turer’s instructions.
Analysis of Tnmph insertion sites
Regions flanking Tnmph insertion sites were obtained by the
method of genome walking (Siebert et al., 1995). As
illustrated in Fig. 1, there are two theoretically possible
results of transposition, and so two groups of walking
primers are needed. To avoid unspecific amplification,
primers were not IS6100 specific, but were immediately
upstream or downstream of IS6100 (as indicated in Fig. 1),
depending on which direction of genome walking was
conducted, and the resulting fragment would contain both
IS6100 and its flanking section. DNA sequences were
determined by Shanghai Sangon Biological Engineering
Technology & Services Co. Ltd (Shanghai, China).
Results and discussion
Tnmph transposition onto the chromosome ofstrains PaW340 and U2 from a suicide plasmid
A transposition frequency of 10�5 was observed after bipar-
ental mating between the donor E. coli S17-1 [pZWWM002]
carrying Tnmph with a Kmr cassette inserted and the
recipient P. putida PaW340. It seemed far higher than that
of the recombination of the plasmid with the recipient’s
Table 1. Strains or plasmids used in this study
Strain or plasmid Genotype or phenotype Reference or source
Strains
E. coli S17-1 TprSmr recA thi pro hsdR M1 RP4::2-Tc::Mu::Km Tn7 lpir lysogen Simon et al. (1983)
E. coli HB101 hsdS20 recA13 ara-14 proA2 lacY1 galK2 rpsL20 xyl-5 mtl-1 supE44 Ausubel et al. (1994)
E. coli DH5a F – recA1 gyrA96 thi-1 hsrdR17 supE44 relA1 deoRD(lacZYA-argF)U169 Woodcock et al. (1989)
Pseudomonas sp. WBC-3 p-Nitrophenol and methyl parathion utilizer, wild type Liu et al. (2005)
Pseudomonas putida PaW340 Mxy�Mtol� Strr Trp�MP� Williams & Murray (1974)
Ralstonia sp. strain U2 Naphthalene utilizer, AmprTcsKms Zhou et al. (2001)
Plasmids
pEX18Tc Tcr; oriT1 sacB1 gene replacement vector with MCS rom pUC18 Hoang et al. (1998)
R388 Tra1 Tpr Sur Ward & Grinsted (1982)
pSTV29 Cmr; cloning vector TaKaRa
pTnMod-Okm Kmr, source of neomycin phosphotransferase II gene (npt II) Dennis & Zylstra (1998)
pZWWM001 Tcr; pEX18Tc derivative carrying 6.5-kb KpnI and BamHI fragment
with the complete Tnmph
This study
pZWWM002 TcrKmr; pZWWM001 derivative carrying Kmr determinant at
unique NotI site immediately downstream of mph
This study
pZWWM003 CmrKmr; pSTV29 derivative carrying the KpnI/BamHI
fragment of pZWWM002
This study
FEMS Microbiol Lett 292 (2009) 85–91 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
87Methyl parathion catabolic transposon

genome, which was found to be lower than 10�8 in the
controls. This indicated that the kanamycin resistance of the
transconjugants resulted from transposition rather than
other recombination events.
Of the 500 Kmr transconjugants selected, all showed
resistance to tetracycline (Tcr), suggesting that pZWWM002
carrying Tnmph may have been entirely integrated into the
chromosome of the PaW340 strain (as illustrated in Fig. 1).
This was further confirmed by colony PCR to detect the
presence of mph, nptII (markers for Tnmph) and tet (tetra-
cycline-resistant gene, marker for pEX18Tc) genes in the
transconjugants obtained. Positive results were obtained in
each case among 20 transconjugants tested, indicating the
consistency between their genotypes and phenotypes. In
addition, all these transconjugants exhibited MPH activity,
as evidenced by the conversion of methyl parathion to
yellow-colored para-nitrophenol by these transconjugants.
The profiles of antibiotic resistance, diagnostic PCR analysis
and the presence of MPH activity indicated that cointegrate
molecules had formed between the donor plasmid
pZWWM002 and the target genomic DNA of the PaW340
strain.
Similar results have also been obtained for the U2 strain,
with a transposition frequency of 10�7. This is much higher
than that in the control, in which the frequency for the
recombination between the plasmid and the recipient’s
genome was found to be lower than 10�9.
Tnmph transposes in a random manner
To discriminate between SmrTcrKmr transconjugants either
as a result of transposition events or because of other
recombination events leading to plasmid integration, and
to whether the transposition occurred randomly, Southern
hybridization was performed on the PstI-digested genomic
DNA of transconjugants that were randomly selected from
the PCR diagnostically tested ones. This was conducted
twice in parallel, each with 10 transconjugants from an
independent conjugation experiment, using an internal
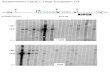
fragment of IS6100 or sacB as the probe (Fig. 2). Neither
probe hybridized to DNA from the control PaW340 strain
(lane 2 in Fig. 2a–d). In contrast, at least one extra fragment
with variable sizes was identified in 16 of the 20 transconju-
gants with the IS6100 probe (lanes 3–9 and 12 in Fig. 2a;
lanes 4–9, 11 and 12 in Fig. 2c), in addition to the two
original IS6100 fragments in pZWWM002 (lane 1 in Fig. 2a
and c). The sizes of the variable IS6100-hybridized frag-
ments differed between the transconjugants, indicating that
Fig. 2. Southern blot hybridization analysis of
the end products of Tnmph transposition in
Pseudomonas putida PaW340. Genomic DNA
(4 mg) was digested with PstI. After electrophor-
esis on a 0.7% agarose gel, the DNA was
transferred to a positively charged nylon mem-
brane and hybridized with DIG-labeled probes of
IS6100 (a and c) and sacB (b and d) (as described
in Fig. 1) at a hybridization temperature of 63
and 54 1C, respectively. Two independent South-
ern blots (a, b; c, d) were performed and in both
cases: lane 1, pZWWM002; lane 2, DNA from
strain PaW340; lanes 3–12, independent
SmrTcrKmr transconjugants.
FEMS Microbiol Lett 292 (2009) 85–91c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
88 M. Wei et al.

independent events had occurred at different genomic sites.
It also implied that there was no hot spot for the target site
of Tnmph, which was consistent with the characteristics of
IS6100-related transposable elements (Herron et al., 1999).
The hybridization with the sacB gene probe further con-
firmed the formation of a cointegrate of pZWWM002 in all
20 transconjugants, in 15 of which a single fragment
identical to the positive control in the PaW340 strain was
hybridized (lanes 3–12 in Fig. 2b; lanes 3, 4, 6, 10 and 11 in
Fig. 2d). For the other five samples, fragments shorter than
the control were observed (lanes 5, 7– 9 and 12 in Fig. 2d).
Interestingly, both phenomena were observed in one trans-
conjugant, with one band identical to the control and one
shorter than the control (lane 5 in Fig. 2d), indicating that
the recipients’ genome is in a dynamic state. This might be
due to some unknown genome rearrangements such as
deletion, recombination or reversion as a result of the activity
of IS6100. This unstable state of the recipients’ genome was
also found in our previous repetition of southern blot with
the transconjugants used in Fig. 2a and b (data not shown).
To detect the random transposition of Tnmph in Ralsto-
nia sp. U2 transconjugants (AmprTcrKmr), sequencing of
Tnmph insertion sites other than Southern blot was con-
ducted from seven transconjugants, using transposon-origi-
nated sequences as primers, as described in Materials and
methods. Sequence analysis could not only further confirm
the randomness of transposition but could also investigate
whether transposition occurred at a particular hotspot. The
result of insertion site sequencing revealed seven different
direct repeats flanking the insertion sites, as indicated in
Table 2.
These data indicate that the transconjugants were a result
of transposition, but not due to other recombination events
leading to plasmid integration. No transposition hotspot
could be detected, suggesting that this kind of transposition
occurred randomly. It is notable that in most transconju-
gants (six out of seven cases), an 8-bp direct repeat was
produced after transposition, which is consistent with the
characteristics of the IS6 family of insertion sequences, as
described in the ISfinder database (http://www-is.biotoul.fr)
(Siguier et al., 2006).
Observation of second transpositions
Second transpositions of IS6100 may have also occurred on
the chromosome, as suggested by the presence of more than
one extra copy of IS6100 detected in four transconjugants
(lanes 8, 9 and 12 in Fig. 2a; lane 4 in Fig. 2c). This is similar
to that seen in the study of the composite transposon DEH,
in which ISPpu12 transposed independently from DEH with
multiple insertions (Weightman et al., 2002; Williams et al.,
2002). Meanwhile, different hybridization patterns with
IS6100 for the same transconjugants were observed in
several cases during their propagation (data not shown),
implying its capability for second transposition. However,
only two copies of IS6100 were detected in four transconju-
gants (lanes 10 and 11 in Fig. 2a; lanes 3 and 10 in Fig. 2c).
This may be because either pZWWM002 had integrated into
the chromosome by abnormal recombination, or deletions
may have occurred after cointegrate formation, as proposed
in a study that used Tn610 (Martin et al., 1990).
Transposition of Tnmph from pSTV29 onto R388in a mating-out experiment
We also confirmed the Tnmph transposability using a well-
developed mating-out experiment (Tsuda & Iino, 1987; Sota
et al., 2002), although it does not identify whether random
insertions occurred during transposition. In this experi-
ment, plasmid pZWWM003, a pSTV29 derivative carrying
Tnmph inserted with the Kmr determinant, was constructed
and then used for the transposition assay. Conjugation
between the donor strain E. coli DH5a-containing plasmids
R388 and pZWWM003 and the recipient strain E. coli
HB101 yielded Kmr/Smr transconjugants at a transposition
frequency of 10�6 per R388 transfer. MPH activity was also
evident in all 10 randomly selected KmrSmr transconju-
gants. Subsequent restriction analyses revealed that each of
the transconjugants carried a plasmid of the same size,
which was presumably due to the formation of a cointegrate
between R388 and pZWWM003 in all cases.
Notably, Tnmph did not resolve from the target sites
during the transposition process after formation of the
cointegrate in either transposition assay. This is because the
Table 2. DNA sequences flanking the insertion sites after the random transposition of Tnmph in Ralstonia sp. U2 transconjugants
Transconjugant Sequence of insertion sites (50–30)
U2-01 GACCCGGCCGTGCTGGCGCAGGCCATCGACCGGGA-insertion site-GACCGGGATACCGGCGCGATCTTCCACCTGGCCGC
U2-02 GCGCGGCCATGGCGGTTTCGCCAGATTCGCTGGTC-insertion site-CGCTGGTCGATGCATGCTGGGCTTCGTCGATCATCA
U2-03 CGACAAGTACGCCCGGCTGCTCGCACGTTGCGAGG-insertion site-CGTTGCGAGGGCCTTCCGCCCATCCCGACCGCCGT
U2-04 AGGGTGTCGACGAGAACGCGCTGACCGTGCCGCAG-insertion site-TGCCGCAGCGCGCGCTGATCCGCAGCGCCCAGGCG
U2-05 GCCGGCACGCAACGACGACAGCCAGATCTATAACG-insertion site-CTATAACGAATATGTGTCGATGCAGTTGGCCCAGG
U2-06 GGACGTGCCGGTGGATGGCTTCTGGTCGGTCAGCG-insertion site-GGTCAGCGTCTACAACGCCAAGGGCTACTTCGAGA
U2-07 GTCAAGGAGTACGTGGTGTCGATCAAGGGCCCGTT-insertion site-GGCCCGTTGACCACGCCGGTTGGTGGCGGCATCCT
Underlined letters indicate the direct repeat produced after transposition.
FEMS Microbiol Lett 292 (2009) 85–91 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
89Methyl parathion catabolic transposon

IS6 family, to which IS6100 belongs, was distinguished by
the formation of a cointegrate molecule as the end point of
transposition, with the integration of the donor vector plus
one additional copy of the IS6100 sequence (Fig. 1) (Martin
et al., 1990; Guilhot et al., 1994; Smith & Dyson, 1995).
In contrast, in the two other functional class I transposons
identified, namely chlorobenzene dioxygenase transposon
(Tn5280), which comprised IS1066 (van der Meer et al.,
1991), and hydrolytic dehalogenases transposon DEH,
which comprised ISPpu12 (Weightman et al., 2002;
Williams et al., 2002), half the transconjugants acquired
the transposon without the integration of the full donor
plasmids.
Tnmph is structurally different from the opd or the opdA
transposable element, implying that diverse ways may have
been adopted in the evolution of these catabolic transposons
during the bacterial adaptation to organophosphorus com-
pounds. Interestingly, mph and opdA, two phylogenetically
divergent genes, are associated with similar transposable
elements, and in contrast, opd and opdA, two similar genes,
are associated with different transposons. This is yet another
example showing the divergent nature of recruitment
events, which bacteria have used to adapt themselves to
newly produced environmental contaminants.
In addition to the current case reported here, an mph-like
gene was also found in phylogenetically divergent bacterial
species and all were located in an identical Tnmph-like
organization (Zhongli et al., 2001; Liu et al., 2005; Zhang
et al., 2006). The linkage of mph to IS6100, their wide
dissemination in various phylogenetically divergent bacter-
ial species, as well as the transposability in a heterologous
host of Tnmph in this study, all suggest that Tnmph might
have played a role in horizontal gene transfer in bacterial
adaptation to organophosphorus compounds. If this is the
case, it is likely that strain WBC-3 has evolved from a para-
nitrophenol utilizer by acquiring the mph transposon
through a natural patchwork assembly, resulting in a com-
plete catabolic pathway for methyl parathion.
Acknowledgements
This work was supported by the National Natural Science
Foundation of China (grant no. 30570021), the National
High Technology Research and Development Program of
China (grant no. 2006AA10Z403) and the Knowledge In-
novation Program of the Chinese Academy of Sciences
(grant no. KSCX2-YW-G-009). We are also grateful to Dr
Masahiro Sota for supplying plasmids R388 and pSTV29.
Authors’contribution
M.W. and J.-J.Z. have contributed equally to this work.
References
Ausubel FM (2001) Preparation and analysis of DNA. Current
Protocols in Molecular Biology. John Wiley, New York.
Ausubel F, Kingston R, Moore D, Seidman J, Smith J & Struhl K
(1994) Current Protocols in Molecular Biology. John Wiley &
Sons Inc., New York.
Dennis JJ & Zylstra GJ (1998) Plasposons: modular self-cloning
minitransposon derivatives for rapid genetic analysis of gram-
negative bacterial genomes. Appl Environ Microb 64:
2710–2715.
Dong YJ, Bartlam M, Sun L et al. (2005) Crystal structure of
methyl parathion hydrolase from Pseudomonas sp. WBC-3. J
Mol Biol 353: 655–663.
Guilhot C, Otal I, Van Rompaey I, Martin C & Gicquel B (1994)
Efficient transposition in mycobacteria: construction of
Mycobacterium smegmatis insertional mutant libraries. J
Bacteriol 176: 535–539.
Herron PR, Evans MC & Dyson PJ (1999) Low target site
specificity of an IS6100-based mini-transposon, Tn1792,
developed for transposon mutagenesis of antibiotic-producing
Streptomyces. FEMS Microbiol Lett 171: 215–221.
Hoang TT, Karkhoff-Schweizer RR, Kutchma AJ & Schweizer HP
(1998) A broad-host-range Flp-FRT recombination system for
site-specific excision of chromosomally-located DNA
sequences: application for isolation of unmarked Pseudomonas
aeruginosa mutants. Gene 212: 77–86.
Horne I, Sutherland TD, Harcourt RL, Russell RJ & Oakeshott JG
(2002) Identification of an opd (organophosphate
degradation) gene in an Agrobacterium isolate. Appl Environ
Microb 68: 3371–3376.
Horne I, Qiu X, Russell RJ & Oakeshott JG (2003) The
phosphotriesterase gene opdA in Agrobacterium radiobacter
P230 is transposable. FEMS Microbiol Lett 222: 1–8.
Jeenes DJ & Williams PA (1982) Excision and integration of
degradative pathway genes from TOL plasmid pWW0. J
Bacteriol 150: 188–194.
Liu H, Zhang JJ, Wang SJ, Zhang XE & Zhou NY (2005) Plasmid-
borne catabolism of methyl parathion and p-nitrophenol in
Pseudomonas sp. strain WBC-3. Biochem Bioph Res Co 334:
1107–1114.
Martin C, Timm J, Rauzier J, Gomez-Lus R, Davies J & Gicquel B
(1990) Transposition of an antibiotic resistance element in
mycobacteria. Nature 345: 739–743.
Mulbry WW & Karns JS (1989) Parathion hydrolase specified by
the Flavobacterium opd gene: relationship between the gene
and protein. J Bacteriol 171: 6740–6746.
Sambrook J, Fritsch EF & Maniatis T (1989) Molecular Cloning: A
Laboratory Manual. Cold Spring Harbor Laboratory Press,
Cold Spring Harbor, NY.
Serdar CM, Murdock DC & Rohde MF (1989) Parathion
hydrolase gene from Pseudomonas diminuta MG: subcloning,
complete nucleotide sequence, and expression of the mature
portion of the enzyme in Escherichia coli. Nat Biotechnol 7:
1151–1155.
FEMS Microbiol Lett 292 (2009) 85–91c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
90 M. Wei et al.

Siddavattam D, Khajamohiddin S, Manavathi B, Pakala SB &
Merrick M (2003) Transposon-like organization of the
plasmid-borne organophosphate degradation (opd) gene
cluster found in Flavobacterium sp. Appl Environ Microb 69:
2533–2539.
Siebert PD, Chenchik A, Kellogg DE, Lukyanov KA & Lukyanov
SA (1995) An improved PCR method for walking in uncloned
genomic DNA. Nucl Acids Res 23: 1087–1088.
Siguier P, Perochon J, Lestrade L, Mahillon J & Chandler M
(2006) ISfinder: the reference centre for bacterial insertion
sequences. Nucl Acids Res 34: D32–D36.
Simon R, Priefer U & Puhler A (1983) A broad host range
mobilization system for in vivo genetic engineering:
transposon mutagenesis in gram negative bacteria. Nat
Biotechnol 1: 784–791.
Smith B & Dyson P (1995) Inducible transposition in
Streptomyces lividans of insertion sequence IS6100 from
Mycobacterium fortuitum. Mol Microbiol 18: 933–941.
Sota M, Endo M, Nitta K, Kawasaki H & Tsuda M (2002)
Characterization of a class II defective transposon carrying two
haloacetate dehalogenase genes from Delftia acidovorans
plasmid pUO1. Appl Environ Microb 68: 2307–2315.
Tan HM (1999) Bacterial catabolic transposons. Appl Microbiol
Biot 51: 1–12.
Top EM & Springael D (2003) The role of mobile genetic
elements in bacterial adaptation to xenobiotic organic
compounds. Curr Opin Biotech 14: 262–269.
Tsuda M & Iino T (1987) Genetic analysis of a transposon
carrying toluene degrading genes on a TOL plasmid pWW0.
Mol Gen Genet 210: 270–276.
van der Meer JR, Zehnder AJ & de Vos WM (1991) Identification
of a novel composite transposable element, Tn5280, carrying
chlorobenzene dioxygenase genes of Pseudomonas sp. strain
P51. J Bacteriol 173: 7077–7083.
Ward JM & Grinsted J (1982) Physical and genetic analysis of
the IncW group plasmids R388, Sa, and R7K. Plasmid 7:
239–250.
Weightman AJ, Topping AW, Hill KE, Lee LL, Sakai K, Slater JH &
Thomas AW (2002) Transposition of DEH, a broad-host-range
transposon flanked by ISPpu12, in Pseudomonas putida is
associated with genomic rearrangements and dehalogenase
gene silencing. J Bacteriol 184: 6581–6591.
Williams PA & Murray K (1974) Metabolism of benzoate and the
methylbenzoates by Pseudomonas putida (arvilla) mt-2:
evidence for the existence of a TOL plasmid. J Bacteriol 120:
416–423.
Williams PA, Jones RM & Shaw LE (2002) A third transposable
element, ISPpu12, from the toluene–xylene catabolic plasmid
pWW0 of Pseudomonas putida mt-2. J Bacteriol 184:
6572–6580.
Woodcock DM, Crowther PJ, Doherty J, Jefferson S, DeCruz E,
Noyer-Weidner M, Smith SS, Michael MZ & Graham MW
(1989) Quantitative evaluation of Escherichia coli host strains
for tolerance to cytosine methylation in plasmid and phage
recombinants. Nucleic Acids Res 17: 3469–3478.
Zhang R, Cui Z, Zhang X, Jiang J, Gu JD & Li S (2006) Cloning of
the organophosphorus pesticide hydrolase gene clusters of
seven degradative bacteria isolated from a methyl parathion
contaminated site and evidence of their horizontal gene
transfer. Biodegradation 17: 465–472.
Zhongli C, Shunpeng L & Guoping F (2001) Isolation of
methyl parathion-degrading strain M6 and cloning of the
methyl parathion hydrolase gene. Appl Environ Microb 67:
4922–4925.
Zhou NY, Fuenmayor SL & Williams PA (2001) nag genes
of Ralstonia (formerly Pseudomonas) sp. strain U2
encoding enzymes for gentisate catabolism. J Bacteriol
183: 700–708.
FEMS Microbiol Lett 292 (2009) 85–91 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
91Methyl parathion catabolic transposon