-
8/6/2019 Transposon in Maize
1/11
a # fi Acta Genetica Sinica , June 2006, 33 (6): 477487 ISSN
0379-4 172
Mutator Transposon in Maize and MULES in the Plant GenomeDIAO
Xian-Min''a', Dam on Lisch21. National Millet Improvement Center of
China, Institute of Millet Crops. Hehei Academ y of Agricultural
and Forestry Sciences,Shijiazhuang 05003 , China;2. University of
California at Berkeley, Department of Plant and Microbial Biology,
11 1 Koshland Hall , CA 94720, USA
Abstract: Mutator ( M u ) is by far the most mutagenic plant
transposon. The high frequency of transposition and the tendency
toinsert into low copy sequences for such transposon have m ade it
the primary means by which genes are m utagenized in maize
(&amays L.). Mus like elements (MULEs) are widespread among
angiosperms and multiple-diverged functional variants can be
presentin a single genome. MULEs often capture genetic sequences.
These Pack-MuLEs can m obilize thousands of gene fragments,
whichmay have had a significant impact on host genome evolution.
There is also evidence that MULEs can move between
reproductivelyisolated species. Here we present an overview of the
discovery, features and utility of Mu transposon. Classification of
Mu elementsand future directions of related research are also
discussed. Understanding Mu will help us elucidate the dynamic
genome.Key words: Mutator; transposon; genome evolution; MULE;
Pack-MULE
Transposons or "controlling elements" were dis-covered by B
arbara McClintock in maize (Zeamays L.)in the early 1950s, but the
theory of transposition wasnot widely accepted until 1970s.
Transposable ele-ments of one type or another have been found in
allorganisms, including all plants that have been investi-gated.
Transposons make up over 50% DNA of thegenome in many species with
large genomes. Trans-poson can rearrange genomes and alter
individual genestructure and expression as a consequence of
transpo-sition, insertion, excision and chromosome breakage.Many
transposon system have been studied in China,such as Ac/Ds, Sp dd
Sp m, Tourist , Stowaway in maize,Mariner in soybean (Glycine max
L.) and Tam inSnapdragon"72'.However, little is known about
Muta-ror (simplified as Mu ), which is the hot point of trans-poson
research in America. Murator transposon wasdiscovered by Robertson
in 1978 from a maize linethat yielded diverse mutations at a
frequency muchhigher than the spontaneous mutation frequency ' 3341
.Itwas found that these mutations were caused by theinsertion of Mu
transposons. M u elements have be-
come the major tool of gene discovery in maize func-tional
genome research and other related fields. Herewe review the
discovery, properties, genetic applica-tion of Mu transposon and
its role in plant gene andgenome evolution.1 The Discovery ofMu
"kansposon
A maize line, known as Mutator, was sent to D.S . Robertson of
Iowa State University from the Uni-versity of Wisconsin for
detailed analysis of its fre-quent mutations. Genetic experiment by
Robertsondemonstrated that not only the heritable mutationfrequency
of this line was very high, but a great dealof somatic variation
was also observed. Further re-search showed that these mutations
were not causedby the previously discovered transposons such
asAc/Ds and S p d d S p m . In ACIDSand S p d d S p m lines,only a
few autonomous elements are present and theysegregate as a
Mendelian element. Transpositions ofthese elements, when they
occurred, could be de-tected as changes in the segregation ratios
of activity.
Received: 2005-09-2 1 ;Accepted: 2005- 10-08This work was
supported by the National Natural Science Foundation of China
(No.30370766).@ Corresponding author. E-mail: xm diao@ya
hoo.com.cn; Tel: +86-3 1-8767 0697
-
8/6/2019 Transposon in Maize
2/11
478 BffYfw Acta Genetica Sinica Vo1.33 No.6 2006In contrast, 90%
of the progeny of an outcross of aMutator line carried active
transposons and were con -sistent with a very high duplication
rate. Thoseprogenies that did lose activity appeared to do so dueto
epigenetic silencing, rather than the segregation ofan autonomous,
or controlling element. This wasconfirmed in many later experiments
[jq6'.he com-plexity and epigenetic regulation features of this
pu-tative transposon system d elayed the discovery of themeans by
which the system was regulated.
In the 1980s, using this Mututor line as mutagenfor gene tagging
became popular. A new kind oftransposon inserted into an allele of
alcohol dehydro-genase 1 and was cloned by Bennetzen et ~ 1 ' ~ '
.ow weknow that this transposon is a nonautonomo us memberof the Mu
transposon family, which was named M u l .Characterization of Mu1
and using it as a probe notonly identified a new kind of transposon
but alsoopened a new era of efficient maize functional
genomeresearch. This new transposon was named Mututorbecause it
came from Mutator maize line. Schnable etul. [*I identified a
Cy/rcy transposon system from anindependent maize line with similar
transposion andgenetic properties of Mu system, which was
laterdemonstrated to be Mu7 of the Mu family. In the early1990s, a
minimal version of the Mutator line was usedto isolate the
regulatory transposon for the Mututorsystem. This regulator, which
was isolated independ-ently in three different laboratories, was
designated asMuDR, in honor of Don Robertson. The isolation ofthe
autonomous two-element, MUDR 9, lo], the creationof a minimal
Mututor line [ ' 'I and an engineered res-cue-Mu element have made
it possible for a sys-tematically elucidation of this super
transposo n family.
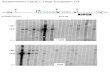
2 The Components and Gene Structure ofMuDR and Mu Transposon
FamilyAll m aize Mu-transposable elements are regulated
by MuDR, which is the autonomous element; MuDRcarries two genes,
mudrA and mudrB (Fig. 1). ThemudrA transcript encodes a 120 kDa
protein, MU RA,which is the transposase of the family. MU RA
containsa domain with high similarity to several bacterial
transposase, which was used as evidence for its veryearly origin
I 3 ' . The mudrB major transcript encodes a23 kDa protein (MURB)
that is not similar to any se-quences in public database outside m
aize and its closerelatives. Although the precise function of MURB
re-mains enigmatic, both deletion derivatives and trans-genic
experiments suggest that MURB is required forM u insertions,
especially germinally transmitted inser-tions L'4 ' . Only two
maize lines with active Mu DR ele-ments have been identified so
far, but all maize linescarry MuDR elements derivatives, or
homologousMuDR sequences (hMuDRs), whose coding sequencesare
80%-99% identical to those of MuDR. Surprisingly,these hMuDRs can
be expressed at the transcriptionlevel though they are not
associated with Mututor ac-tivity and they might play an
as-yet-unknown functionin Mututor regulation j1 .
Unlike Ac/Ds an d S p d d S p m transpson systems,which mostly
comprise of the autonomous elementand its deletion derivatives, the
Mu transposon systemcontains complex family elements,
includingautonomous M u D R , homologous M u D R elements andmany
variants with similar TIRs. All Mu transposonelements share similar
-170-220 bp TIRs; the lengthof direct repeat sequence flanlung the
inserted ele-ments is 9 bp. Many Mu elements contain
additionaldirect or inverted repeat sequen ces. Although
deletionderivatives are the most common source of Ds ele-ment from
Ac and of dSpm elements from Spm, mostMu elements internal
sequences are unrelated to thetranposase-coding sequence of Mu DR,
or to eachother (Fig. I ) . In most cases, the internal sequen ces
ofM u elements are part of a host gene and it seems thatMu TIRs can
capture host functional sequence by sofar unknown mechanism. These
kinds of elements arecalled Puck-MuLEs I s 1 . In some species,
such as rice(Oryza sativu L.), there can be thousands of
inde-pendently derived Puck-MuLEs.
Jittery is a newly isolated and characterizedmember of M u
element, whose transposase-codingsequence is only a 3.9 kb gene
with high identity tomudrA but not to mudrB. The length of direct
hostsequence duplication flanking the Jittery insertions isalso 9
bp, but its 177 bp TIRs are unrelated to that of
-
8/6/2019 Transposon in Maize
3/11
DIAO Xian-Min ef al.: Murator Transposon in Maize and MULES n
the Plant Genome 479
Fig. 1 The Mutator transposon family in maize and the engineered
RescueMuAll classes of Mu elements share similar terminal inverted
repeats (TIRs; Black ends), but each class has a unique internal
sequencethat shares similarity to various host functional protein
coding sequences, as indicated in the figure. Mu1 is similar to
MRS-A(Mu-related sequence A). Mu3 is similar to an Arubidopsis
protein and Mugo nashi protein. Mu4 includes a sequence 95%
identicalto a maize expressed sequence, accession No . BG466445.
Mu7 is similar to Arubidopsis protein accession No. NP-192120.
Mu8contains a sequence with similarity to pgpl protein from
Aruhidopsis. RescueMu is an engineered element based on Mu1 used as
agene-tagging tool, into which unique tag sequence, pBluescript and
selectable maker w ere inserted. For MuDR, the position of mudrAand
mudrB are indicated. The figure is a modified version from Lisch
(2002) [I7.
M u D R . Despite limited investigation has been doneon the
transposition activity of Jittery, the results sofar obtained
demonstrated that i t is an autonomoustransposable element.
Jittery, like M u D R causes ahigh frequency of late somatic
reversion, but like de-letion derivatives that lack mudrB, it is
not associatedwith new insertions I 6 .
In addition to the members of Mu elements foundin maize, many M
u-like elements (MULES)have beenidentified in both dicots and
monocots. Mu transposonfamily is characterized by the presence of
many diver-sified and potentially functional variants, and its
veryhigh levels of activity in maize, which,complicatedanalysis of
its origin and evolution.
3 Transposition Activity ofMuData related to Mu transposition
activity are
largely a product of investigations on maize, althoughMULES in
Arnbidopsis had been confirmed to be ac-tive in D D M I mutant
background I . Two main typesof transpositions occur with maize Mu
elements. Oneis somatic excision, or excision and insertion late
dur-ing development in somatic cells and tissues, the other
one is insertions occurring in germinal cells, which isthe
source of heritable new mutations in Murutor lines.M u excisions
have been investigated extensively withanthocyanin reporter
alleles, such as a 1-mum2, e-cause pigmented spots can easily be
visualized andquantified. Most somatic excisions occur in the
latedevelopment stage, within the last two or three celldivisions
of a given lineage, though early excisionscan occasionally occur.
The most visible result of so-matic excision is revertant sectors,
resulting from theexcision of the inserted transposon and
restoration ofthe a1 pigmentation function. Two kinds of
somaticexcision have been documented for Mu elements, thecut-only
and the cut-and-paste. The excised trans-posons of the cut-only
form were failed in insertinginto other sites of the host genome.
Cut-and-pastetransposition is the typical mode of transposition
ofDN A transposons and is often associated with reinser-tion
elsewhere in the genome. But excision of A d D sand S p d d S pm
elements typically result in minorchanges to the host sequence
duplication created ontransposon insertion. In contrast, Mu element
excisioncreate deletions and additions of the host gene, *
In contrast to high frequency of somatic revertant
-
8/6/2019 Transposon in Maize
4/11
480 Acta Genetica Sinica Vo1.33 No.6 2006
mutations, germinally transmitted revertants are ex-tremely rare
in Mututor lines, which suggest that exci-sions are prevented by
some kind of mechanism incells that give rise to In minim al
Mututorlines with just o ne copy of Mu1 initially inserted intothe
ul-mum2 allele and one copy of MuDR, from10%-20%of progeny contain
more than one copyof Mu1 or MuDR"'] and many experiments con-formed
that the existing M u insertion segregate in aMendelian fasion.
These experiments reveal that newgerminal insertions must happen
without the loss ofthe existing Mu insertion; in another word, som
e kindof duplicative or replicative MuDR transposition hap-pens
during the process of de~elopment[~'g 1 . Thoughgerminal
duplicative insertion can occur throughoutthe development, which
mostly begins late in sporo-phyte cell divisions, where
transposition events createsmall clusters of gam etes carrying the
same mutations.Germinal transposition continue through meiosis
tothe last mitotic divisions of the gametophytes, and theafter
meiosis insertion generates sperm with d ifferentmutations
[33221.The frequency of germinally transmit-ted insertions depends
on the Mututor line used, butthere are clear position effects on
both cis and transactivity of individua l MuDR elem ents "I
Many Mu-induced mutations are suppressible.The mutant phenotype
in these cases arises only in thepresence of active MuDR elements,
probably due tosteric effects introduced by the presence of the
trans-posase bound to the transposon TIRs. Consistent withthis
model, many suppressible Mu insertions are inpromoter regions.
However, M u insertion into intronscan also be suppressible 1231,
and it is likely that Musuppression will turn out to be more
complex than asimple model of steric hindrance would suggest.
Many investigations have demonstrated that Mutransposons
generally insert into single copy orlow-copy-number regions of the
genom e whichmakes it a powerful tool for maize functional gene
tag-ging. By analyzing the sequences that flanking the 88RescueMu
insertions, Dietrich and co-authors demon-strated that 69%of these
sequences were genes and only4% were repetitive
retrotranspo~ons[~~~.much recentexperiment also with RescueMu shows
that the rate of
single copy insertion is about 66%,which is consistentwith the
previous result Both Ac/Ds and S p d d S p mtend to transpose to
genetically linked sites. In contrast,Mututor transposes to
unlinked sites, an advantage forwhole genome mutagenesis
applications. However,Hardeman and ChandleJ271ound that certain
classes ofMu elements predominantly targeted certain genes,which
implies that some M u elements may have inser-tion affinities. The
5WTR region or the promoter regionof 818 gene show ed a strong
preference for Mu elementinsertion, 62 of 75 insertions of g18 gene
targeted in thisregion, suggesting that 5'UTR targeting might be a
fea-ture of Mu insertion. But introns and other regions ofgenes are
also often targeted by Mu elements'251. naly-sis of 339 target site
duplications (TSDs) created by Muinsertions also showed some degree
of sequence prefer-ence, the weak consensus for Mu insertion
wasCTCB(G/C)(A/C)(G/A)(A/G)C. Furthermore, se-quences immediately
linked to TSDs also showed con-servation; the consensus sequence of
5' of the TSD isCCT and that of the 3' of the TSD is AGG
Mu-targetedsequences were found to be GC rich relative to the
restof the maize genome 2" However, many M u insertionsites do not
have the consensus sequence.
Four types of assays are usually used to demonstrateMututor
system activity, including examination of somaticinstability of
reporter alleles (such as sectored leaves,kernels or anthers),
special enzym es to detect m ethylationof diagnostic restriction
sites in the TIRs Hinf I for Mu1and Sac I for MuDR), the detection
of new insertions inprogeny plants, and an elevated forward
mutation fre-quency resulting from new germinal insertion^'^'.
Whichmethod should be used depends on needs of the user.Early
observations of Mututor suggested that the loss ofactivity was due
to epigenetic silencing, rather than sim-ple segregation of a
regulatory transposon. Two modelsfor silencing had been proposed:
ectopic paring betweenhomologous TIRs and posttranscriptional
RNA-basedsilencing '52g'. Until recently, the detailed initiation
andprocess of Mu family silencing still remains enigmatic.
Adominant locus that can initiate Mututor silencing, Mukiller
(Muk), was cloned. This locus was dem onstrated tobe an inverted
duplication of a partially deleted autono-mous MuDR element. Muk
produces a d r A hairpin
-
8/6/2019 Transposon in Maize
5/11
DIAO Xian-Min et al .:MutatorTmsposon in Maize and MULEY n the
Plant Genome 48 1
transcript that is processed into smal l RNAs that targetsmudrA
for silencing. This, in turn, esults in transcrip-tional silencing
of mudrA, and subsequently mudrB. Mukprovides the first example of
a natural occurrence of de-rivative that is able to initiate
heritable silencing of anactive transposon family '299". This
discovery not onlyclarified our understanding of M u epigenetic
silencing,but also gave us new tools for investigation of
bothtranscriptional and posttranscriptional regulation ofgene
expression.
It is suggested that the initial silencing of Mu ac -tivity is
followed by detectable TIRs methylati~n'~'.Indeed, 5'-methylation
of cytosines within TIRs is adiagnostic feature of Mu transposons
silencing state.In typical Mutator lines or complex lines, both Mu
DRand nonautonomous element, such as M u l , can bemethylated
gradually in the progeny of a linage or atdifferent development
stage of an individual plant.The methylation of MuDR is accompanied
by tran-scriptional silencing and loss of activity [ sx31 i .
Inminimal lines with single copy of Mu DR, both MuDRand the
nonautonomous elements are unmethylated.Methylation of
nonautonomous elements occurs ifMuDR element is lost due to genetic
segregation. Insuch cases that Mu DR is restored genetically, the
me-thylation of nonautonomous elements is lost, sug-gesting that it
represents a default state that occurs inthe absence of the
transposase"'. 321 . This default me-thylation is dependent on at
least two mutations thatwere discovered due to their effects on
paramutation.In mop1 (mediator of paramutation) mutants,
para-mutatable alleles of several color genes are epigen-etically
activated and nonautonomous Mu elementsTable 1 List of someMu
agging cloned m aize functional genes
are hypomethylated. Mu DR elements that had beensilenced by Mu
kil ler are also hypomethylated in amop1 mutant background. If this
element is main-tained in a mutant background for multiple
genera-tions, one of the two genes encoded by Mu DR, thetransposase
mudrA becomes reactivated. Further, thesecond gene, mudrB, remains
methylated and si-lenced, suggesting that, although the two genes
hadboth been silenced by Muk, maintenance of that si-lenced state
is mediated by different factors'331.Si-lenced M u elements in
typical lines can also be reac-tivated by gamma and U V radiation
of seeds and pol-len, and radiation treatments are more efficient
onlines within one or two generations of silencing thanthose lines
that had kept many generations of silencestate'". Singer et al. ["I
found that Mutatur-like ele-ments ( M U L E S ) n Arabidopsis
genome become de-methylated and active in the
chromatin-remodelingmutant ddml (Decrease in DNA Methylation),
whichleads to loss of heterochromatic DNA methylation.
4 Genetic Application of M u TransposonTransposon tagging is one
of the main methods
used in functional genome research. A d D s taggingsystem has
not only been successfully used in maizebut also in rice and
Arabidupsis [34i. The advantages ofM u tagging are that it moves to
any chromosome, itcauses a very high mutation rate, insertions tend
to beinto or near genes, and most loci appear to be poten-tial
targets. Using this transposon system, many maizegenes have been
cloned and characterized, some ofwhich are listed in Table 1.
No. Gene or allele Possible function Database code Reference1 su
I Kernel developm ent related AY290 402 WI2 Rough sheath 1 Leaf
sheath development related L44133 P6 13 APETALA2 Spikelet
development related AF048900 WI4 Ligueless3 Architecture of ligule
AF4 57 125 [3815 Knotted1 Leaf development AY3 12169 P I6
Viviparous1 Induction of embryo development NO01635 ~4017 A d h l
Oxygen starvation response X04049 [71
-
8/6/2019 Transposon in Maize
6/11
482 B.ff%#@Acta Genetica Sinica Vo1.33 No.6 2006Targeted
mutagenesis was the initial application
in typical Mutator lines, because it is relatively easyfor
scientists to recover mutations in well-studiedgenes that con fer
visible phenotypes [51. For loci witha recessive loss-of-function
allele, wild-type plant withactive Mu were crossed to the
homozygous recessivetester stock, mutant individuals in the F1
progenywould be expected to carry a Mu-tagged allele fromthe
Mu-active parent. For loci with dominantgain-of-function alleles,
Mu-active lines homozygousfor the dominant allele need to be
established first, andthen crossed to a normal test line. Normal
individualsof the F, most likely contained a Mu-inserted
disrup-tion of the dominant allele. Because Mu causes a
highmutation rate, large populations of Mu active lineshave been
screened to find new phenotypes of interest.
Three big project of Mu-tagging maize func-tional genome
research had been carried out in theUnited States. The Trait
Utility System for Corn(TUSC) was implemented by Pioneer Hi-Bred,
Inc.,which was based on DNA samples of -45 000Mu-active individual
plants and the available ofmaize EST sequences. Using a primer
running outfrom the Mu TIR sequence and a primer designed onthe
basis of an EST of interest, population screeningcan be used to
find individuals with a Mu insertioninto or near the gene of
interest. Progeny analysis canthen be used to determine which
plants transmitted aheritable mutant allele. Maize-Targeted
Mutagenesis(MTM) is a US National Science Foundation fundedproject
for public maize research service. With thesame strategy to TUSC,
MTM uses PCR to screenfrom a population of -46 000 M u active
plants andprovide users with seed for phenotypic characteriza-tion.
RescueMu was another project carried out byjoint co-operation of
Stanford University, UC Berke-ley and other institutes, which
involves screeningprogeny of RescueMu transformed maize plants.
Thisapproach has the virtue of combining genomic se-quencing and
functional genomics into a single step,in which RescueMu insertions
provide not only amutation but also a cloned allele for sequencin
g.
Since M u transposon had been effectively used
in maize functional genome research, it would beadvantageous if
we use this system in other plantspecies for gene tagging as well.
Th e most direct wayto do this is to create a heterologous M u
transposontagging in other species. Unfortunately, althoughmudrA
and mudrB have been successfully trans-formed into rice, and the
transformed genes can beinherited for a few generations,
transcription ofmudrA an d mudrB was not detected
(unpublisheddata). So we need more knowledge about Mu epige-netic
regulation and transferring gene silencing.Transposition active
MULEs have been observed inArabidopsis D D M l mutant I and a
fungal speciesFusurium oxysporum [411, suggesting that
transposi-tionally active M u elements do exist in organisms
inaddition to maize. Thus, screening for active andmutagenic MULEs
may be another option for M utagging in species su ch as rice.
5 Occurrence and Classification of M uElements in Other Plant
Genom esIt has been demonstrated that homologs ofmudrA exist in
many plant genomes, but homologs
of mudrB have only been observed i n Z . luxurians,Z.
diploperennis and a few other species closely re-lated to
maize[421.Mu-like elements are genericallyreferred to as M U L E S
[ ] .Autonomous MULEs showvarying degrees of similarity with mudrA
in maize.So far intact element of MULEs , including TIRs andTSDs,
had been cloned and characterized fromArubidopsis, Brassic napus,
rice, sorghum (Sorghumbicolor L.), sugar cane (Saccharum oficinarum
L.),wheat (Triticum aestivum L.), barley (Hordeum vul-gare L.),
foxtail millet (Seturia italica L.) and a fewbamboo species
(Bambuseae) species[532-44. Lisch etal . [421 loned and
characterized mudrA sequencesfrom 28 plant species. Cluster
analysis based on theunclear tide identity of the cloned fragments
showeda discrepancy in the relationship between phylogenyof the
sample species and the evolution phylogeny ofthose MULEs . Eisen et
al. I 3 found that the sequenceof transposase mudrA was similar to
transposase ofsome insertion sequence (IS) in a group of
bacteria
-
8/6/2019 Transposon in Maize
7/11
DIAO Xian-Min e f al.: MufalorTransposon in Maize and MULES n
the Plant Genome 48 3
species such as Mycobacterium bovis. These suggestthat the M u
transposon family might have had a veryearly origin. The genomes of
fungus do have MULEsand even a transpositional active one w as
characterizedI 4 I 1 . To date no MULEs have been reported in
animals.
Not only do we know that MULEs are ubiqui-tous in plant genomes
so far tested, but analysis ofpublic EST data demonstrate that
MULEs in manyplants, including rice, wheat, sugar cane and
barleyare transcripted. Thus, transcriptional active MULEsmaybe a
common property of plant gen~mes'~*].os tMULEs contain mudrA genes
that are well divergedfrom that in maize, but many of those MULEs
havegood open-reading frames with evidence for selectionat the
amino acid level, consistent with continuedfunctionality.
Conservation of TIR sequences witheach other and targeted side
duplication (TSD ) is alsoconstant with continued transposition
activity. Manyduplicated copies of one kind MULEs were found inthe
sequenced rice genome (unpublished observation),and we found
several MULEs of Setaria viridis werehighly polymorphic among
different genotypes of thesame species by Southern blot
(unpublished data),suggesting activity in this species. Database
searchusing the protein sequence of MURA reveals thatproteins with
high similarity to MURA exist inArabodopsis, potato (Salanum
tuberosum L.), tomato(Lycopersicon esculentum M.) and many other
plantspecies [ 3 2 1 .Yoshida 1451 found a rice MULE expressedin
callus subcultured with praline and even found atranscriptionally
active M U L E in rice somaclonallines. Three instances of
transpositionally activeMULEs have been reported, including Jittery
inmaize, AtMul in Arabidopsis DDM mutant and Hop1in the fungus
Fusarium oxysporum. Jittery resemblesMurator in the length of the
element's TIRs, the sizeof the target site duplication, and the
makeup of itstransposase, but differs from MuDR in that it encodesa
single MURA-like protein. Jittery also differs fromMutator elements
in the high frequency with which itexcises to produce germinal
revertants and in its lowcopy number in most maize lines and maize
relativesexamined. However, Jittery cannot be considered as abona
fide transposon in its present host line, because
i t does not reinsert in the genome [ I 6 ' . Together,
thesedata suggest that MULEs may be transpositionallyactive now or
in the near past in many plant genomes.Thus, it is quite likely
that active Mu systems in otherplant species may be as useful as
Mutator does inmaize.
MULEs can be widely diverged from each othereven they come from
the same genome. Jittery, whichis an active M U L E in maize, is m
ore similar to MULESin rice and Arabidopsis than it is to MuDR
[16]. Re-markably, two of MULEs in Arabidopsis with highsimilarity
to Jittery appear to be host functional genesrather than
transposons; mutations in those genes causedefects in the
far-red-light response pathway , Inthis case, it appears that the
transposase has been re-cruited by the host to perform a novel
function.
The distribution of any given group of MULEs inthe grasses is
patchy, suggesting the possibility ofhorizontal gene transfer. Diao
and colleagues'491 ounda MULE from Setaria with high similarity to
a MULEfrom rice; the nucleotide identity between the twoelements is
as high as 90% including correspondingintron sequences. Given the
50-60 million years ofevolution divergence that separate Setaria
and rice'501,the high degree of similarity of non-coding
sequencescan only be explained by horizontal transfer[491. his
isthe first well-documented examp le of horizontal trans-fer of any
nuclear-encoded genes between higherplants. It is clear that the
evolution of M u super familyis markedly different from their
hosts.
MULEs form a highly com plex and broadly diver-sified family of
transposon. All Mu-related elements sofar found in plant genomes
can be classified into fourgroups: the first includes MuDR and
hMuDRs as statedin the second part of the review. Many of these
elementscontain point m utations but are transc riptionally
ex-pressed and may p lay a role in the epigene tic regulationof the
family '51. The second group includes elementsthat have mudrA
homologs and long distinct TIRs fromMuDR, such as Jittery in maize
and Sf4 in Setaria. Thethird group of Mu super family includes
those elementsthat share similar TIRs with a MULE encoding mudrAbut
that carry internal sequences that are unrelated to the
[46-481
-
8/6/2019 Transposon in Maize
8/11
484 B%%%! Acta Genetica Sinica Vo1.33 No.6 2006transposase.
Examples of these include some of thenonautonmous elements in
maize. Their internal se-quences are captured fragments of host
genes. Theseelements are known as Pack-MULES, and they can
bepresent in large numbers in any given genome"5,5',521.The fourth
group includes transposons that canymudrA-like genes but lack long
TIRs. Individual in-stances of this class of e lemen ts may
represent cooptedmudrA genes, genes such as Farl in Arabidopsis
[46,471.6 Mu Elements in Plant Genome and G ene
EvolutionThe genom e of all plant species is in a state of
dy-namic equilibrium between evolution and stability, but
of course stability is always temporary. The diverse ac-tivity
of transposons has been demonstrated to be animportant factor
affecting on the evolution of genes andgenomes 11953v541.ransposon
can induce gene silencingor reactivation, gene or genome
restructuring includingdeletion, duplication, reversion and
translocation. Theaccumulation of such changes undoubtedly has had
aprofound impact on genome evolution .Mu transposons are the most
active, most com-plex and m ost ubiquitous DNA transposable element
inplant genomes, suggesting a special role in gene andgenome
evolution. Unlike Ac/Ds and SpddSpm,whose activity result in
relatively minor changes to thehost sequence, Mu activity can
result in a wide rangeof changes [I2]. It has demonstrated that
MuDR ele-ments in different loci can show distinct frequencies
oftransposition, suggesting that this class of elements
isparticularly sensitive to their local environment. It isclear
that Mu elements can evolve a novel function,such as in the case of
the Arabidopsis Farl gene .The most recent discovery of the impact
of M u ele-ments on plant genome evolution is Pack-MULES,which
contain fragments of genes. With systematic andcareful analysis,
Jiang er aL[553561und more than 3 OOOPack-MULES in the rice genome,
and Pack-MULESalso had been seen in maize and Arabidopsis. Thelarge
number of Pack-MULES and their presence inmultiple genomes suggest
that they may be a majorcomponent of most or all flowering plant
genomes.
W 4 7 1
Gene fragments from different rice genes were foundtogether in
-23% of Pack-MULES in rice, suggesting ameans by which hybrid genes
could be created due tothe activity of a transposon. At least 5%
ofPack-MULES were found to be expressed, as evi-denced by
full-length cDNAs, with an identical DNAsequence match. H ence, by
the criterion of expressionat the RNA level, many of these
Pack-MULES are al-ready new genes. The ability of Pack-MULES to
cap-ture gene fragments combined with the possibility ofhorizontal
transfer as documented by Diao er ~ 1 . ' ~ ~ 'also suggest a means
by which gene fragments couldbe moved between species.
Although we have made some progress in under-standing of the
MuDR/Mu superfamily, many interestingpuzzles remains to be explored
for future research. Themost intriguing questions include the
detailmechanism ofMu transposition and the epigenetic regulation of
Musilencing and reactivation. The solution for such questionswill
help us establish Mu tagging system in other plantsthan maize and w
idening its application in plant genetics.The frequent
transposition and interaction with host ge-nomes make the evolution
study of transposons difficult,but analysis of this fascinating
super family of elem entsand a clear understanding of their
evolution will certainlyhelp us understand the dynamics of gene and
genomeevolution. Pack-MULE structure is ubiquitous in plantgenomes
and predicted to be a mechansism for the crea-tion of novel but we
know nothing about themechanism of fragments acquisition and so far
no in-stances of newly created genes in this way have beenreported.
As stated by Shapiro of Chicago Un i~e rsit y"~ '"transposable
elements are the key to a 21st centuryview of evolution", advances
in our understanding ofthe MULE super family will for certainly
help us to il-luminate both transposon and genome evo
lution.Acknowledgement: We thank Hui Zhi of NationalMillet
Improvement Center of China for figurepreparation.References:[ l ]
Bennetzn J L. Transposable element contributions to
plant gene and genome evolution. Plunf Mol Biol, 2000,
-
8/6/2019 Transposon in Maize
9/11
DIAO Xian-Min et a l . :Mutator Transposon n Maize and MULES n
the Plan t Genome 485
42(1) : 25 1-269.[2] SanMiguel P, Bennetzn J L. Evidence that a
recent in-
crease i n maize genome size was caused by the m
assiveamplification of intergene retrotransposons. An n Bor
Res,1998,82(SuppIem ent A) : 37-44.Robertoson D S. Mutator activity
in maize: timing of itsactivation in ontogeny. Science, 198 , 2
13(4515) :1515-1517.Robertoson D S. Genetic studies on the loss of
M u muta-tor activity in maize. Genetics, 1986, 1 13(3) :
765-773.
Walbot V, Rudenko G N. MuDWMu transposons of maize.In: Aig N L,
Craigie R, Gellert M, Lambowitz A, eds.Mobile DNA 11. American
Society for Microbiology,Washington, DC. 2002, 533-564.Martienssen
R, Baron A. Coordinate suppression of mu-tations caused by
Robertsons mutator transposons inmaize. Genetics, 1994, 136 (3) :
1157- 1 170.Bennetzen J L, Swanson J, Taylor W C, Freeling M.
DNAinsertion in the first intron of maize Adhl affects
messagelevels: cloning of progenitor and mutant Adhl alleles.Proc
Nut1 Acad Sci USA, 1984,8 ( I 3) : 4 125-41 28.Schnable P S,
Peterson P A. The Mutator-related C ytransposable element of Zea
mays L. behaves as anear-Mendelian factor. Genetics, 1988, 120(2):
587-596.Chomet P, Lisch D, Hardeman K J, Chandler V L, Freel-ing M.
Identification of a regulatory transposon that con-trols the
Mutator transposable element system in maize.Genetics, 1991, 129( )
: 26 1-270.
[ lo] Hershberger R J , Warren C A , Walbot V. Genetics Muta-tor
activity in maize correlates with the presence and ex-pression of
the Mu transposable element Mu9. Proc NailAcad Sci USA ,
1991,88(22) : 10198-10202.
[ 1 I ] Lisch D, Chomet P, Freeling M. Genetic
characterizationof the Mutator system in maize: behavior and
regulationof Mu transposons in a minimal line. Genetics,
1995,139(4) 1777-1 796.Raizada M N, Nan G L, Walbot V. Somatic and
germinalmobility of the RescueMu transposon in transgenic m
aize.Plant Cell, 2001, 13(7) : 1587-1608.Eisen J A, Benito M I,
Walbot V. Sequence similarity ofputative transposases links the
maize Mutator autono-mous element and a group of bacterial
insertion se-quences. Nucleic Acids Res, 1994, 22( 13) :
2634-2636.
[14] Lisch D, Girard L, Donlin M, Freeling M. Functionalanalysis
of deletion derivatives of the maize transposonMuDR delineates
roles for the MURA and MURB pro-teins. Genetics, 1999, 151 1 ) :
331 -34 I .
[15] Jiang N, Bao Z R, Zhang X Y, Eddy S R, Wessler S
R.Pack-MULE transposable elements mediate gene evolu-
tion in plants. Nature, 2004,43 l(7008) : 569-573.[I61 Xu Z N,
Yan X H, Maurais S , Fu H H, OBrien D (3,
Mottinger J, Dooner H K. Jittery, a Mutator distant rela-tive
with a paradoxical mobile behavior: excision withoutreinsertion.
Plant Cell, 2004, 16(5) : 1 105- I I 14.
[I71 Lisch D. Mututor transposons. Trends Plant Sci, 2002,7( 11
) : 498-504.
[181 Singer T, Yordan C, Martienssen R A. Robertsons Muta-tor
transposons in A. thaliana are regulated by the
chro-matin-remodeling gene Decrease in DNA Methylation( D D M I )
.G e n e s D e v , 2001, 15(5) : 591-602.
[I91 Nordborg M, Walbot V. Estimating allelic diversity
gen-erated by excision of different transposon types. TheorAppl
Genet, 1995,90(6): 771-775.
[20] Britt A B, Walbot V. Germinal and somatic products
ofexcision of Mu1 from the Bronze-1 gene of Zea mays.Mol Gen Genet,
1991,227(2) : 267-276.
[21] Chandler V L, Hardeman K J. The Mu elements of Zeamays. A
dv Genet, 1992, 30 : 77-122.
[22] Robertoson D S. The timing of M u activation in
maize.Genetics, 1989, 94(4) : 969-978.
[23] Greene B, Walko R, Hake S. Mutator insertions in anintron
of the maize Knotted1 gene result in dominantsuppressible
mutations. Genetics, 1994, 138(4) :1275-1285.Fernandes J, Dong Q,
Schneider B, Morrow D J, Nan G L,Brendel V, Walbot V. Genome-wide
mutagenesis of Zeamays L. using RescueMu transposons. Genome Biol,
2004,5(10) :R82Dietrich C R, Cui F, Packila M L, Li J, Ashlock D
A,Nikolau B J, Schnable P S. Maize Mu transposons aretargeted to
the 5 untranslated region of the g18 gene andsequences flanking Mu
target-site duplications exhibitnonrandom nucleotide composition
throughout the ge-nome. Genetics, 2002, 160(2) : 697-716.
[26] Fernandes J, Dong Q F, Schneider B, Morrow D J, Nan GL,
Brendel V, Walbot V. Genome-wide mutagenesis ofZea mays L. using
RescueMu transposons. Genome Bid,2004,5( 10): R82. 1-R82.20.
[27] Hardeman K J, Chandler V L. Two maize genes are
eachtargeted predominantly by distinct classes of M u
elements.Genetics, I 993, 135(4) : 1I4 - I 50.
[28] Bennetzen J L, Springer P S, Cresse A D, Hendrichkx
H.Specificity and regulation of the Mutator transposableelement
system in maize. Crit Rev Plant Sci,1993, 12(1) :57-95.[29] Slotkin
R K, Freeling M, Lisch D. Mu killer causes theheritable
inactivation of the Mutator family of transposableelements in Zea
mays. Genetics, 2003, 16x2) : 78 1-797.
-
8/6/2019 Transposon in Maize
10/11
486 %@%H Acta Genetica Sinica Vo1.33 No.6 2006[30] Slotkin R K,
Freeling M, Lisch D. Heritable transposon
silencing initiated by a naturally occuring transposon in-verted
duplication. Nat Genet, 2005, 37(6) : 6 4 - 6 4 4 .
[31] Rudenko G N, W albot V. Expression and
posttranscriptionalregulation of maize transposable element MuDR
and its de-rivatives. Plan t Cell, 2001, 13(3) : 553-570.
[32] Lisch D, Chomet P, Freeling M. Genetic characterizationof
the Mutaor system in maize: behavior and regulationof Mu transposon
in a minimal line. Genetics, 1995,139(4) : 1777- 1796.
[33] Woodhouse M R, Freeling M, Lisch D. The mop1 (me-diator of
paramutationl) mutant progressively reactivatesone of the two genes
encoded by the MuDR transposon inmaize. Genetics, 2006, 172(1) :
79-92.
[34] Zhu Z G, Fu Y P, Xiao H, Hu G C, Si H M,Yu Y H, Sun ZX.
Ac/Ds transposition activity in transgenic rice popula-tion and DNA
flanking sequence of Ds insertion sites.Acta Bot Sin, 2001,45 (1) :
102-107.
[35] Dinges J R, Colleoni C, Myers A M, James M G Mo-lecular
structure of three mutations at the maize sugary1locus and their
allele-apecific phenotypic effects1 . PlantPhysiol, 2001, 125(3) :
1406-1418.
[36] Girard L, Freeling M. Mutator-suppressible alleles of
roughsheath1 and liguleless3 in maize reveal multiple mechanismsof
suppression. Genetics, 2000, 154( ) :437-446.
[37] Chuck G, Meeley R B, Hake S. The control of maizespikelet
meristem fate by the A PETALA2-like gene inde-terminate spikeletl.
Genes Dev, 1998, 12(8) : 1145-1154.
[38] Fowler J E, Muehlbauer G J, Freeling M. Mosaic analysisof
the ligueless3 mutant phenotype in maize by coordi-nate suppression
of Mutaror-insertion alleles. Genetics,1996, 143(1) : 489-503.
[39] Greene B, Walko R, Hake S. Mutator insertion in an in-tron
of the maize knotted1 gene result in dominant sup-pressible
mutations. Genetics, 1994, 138(4) : 1275-1285.
[40] McCarty D R, Carson C B , Stinard P S, Robertson D
S.Molecular analysis of viviparous 1: an abscisic acid-in-sensitive
mutant of maize. Plant Cell, 1989, l(5) :523-532.
[41] Chalvet F, Grimaldi C, Kaper F, angin T, Daboussi M J.Hop,
an active Mutator-like element in the genome of thefungus Fusarium
oxysporum. Mol Biol Evol, 2003, 20(8) :1362-1375.
[42] Lisch D, Freeling M, Langham R J , Choy M Y.
Mutatortransposase is widespread in the grasses. Plant
Physiol,2001, 125(3) : 1293-1303.
[43] Rossi M , A raujo P C, de Jesus E M, Varani A M,vanSIuys M
A. Comparative analysis of Mutator-like trans-posases in sugarcane.
Mol Gene Gen, 2004, 272(2) :194-203.
[44] Mao L , Wood T C, Yu Y, Budiman M A, Tomkins J, WooS,
Sasinowski M, P resting G, Frisch D, Goff S, Dean R A,Wing R A.
Rice transposable elements: a survey of 73000sequence
tagged-connectors. Genome Res, 2000, 1O(7) :982-990.
[45] Yoshida S, Tamaki K, Watanabe K, Fujino M,Nakamura C.A
maize M uDR-like element expressed in rice callus sub-cultured with
praline. Hereditas, 1998, 129(1) : 95-99.Hudson M, Ringli C, Boylan
M T, Quail P H. The FAR1locus encodes a novel nuclear protein
specific to phyto-chrome A signaling. Genes Dev, 1999, 13(15) :20
17-2027.Wang H, Deng X W. Arabidopsis FHY3 defines a keyphytochrome
signaling component directly interactingwith its homologous partner
FARI. EMBO J , 2002,21(6) : 1339-1349.Hudson M E, Lisch D R, Quail
P H. The FHY3 and FAR1genes encode transposes-related proteins
involved inregulation of gene expression by the
phytochromeA-signaling pathway. Pla nt J , 2003, 34(4) :
453-471.Diao X M, Freeling M, Lisch D. Horizontal transfer of
aplant transposon. Public Library of Sciences, Biology,2006,4( 1) :
119-128.Kellogg E A. Evolutionary history of the grasses.
PlantPhysiol, 2001, 125(3) : 1198-1205.Lisch D. Pack-MU LES: theft
on a massive scale. Bioes-says, 2005,27(4) : 353-355.Bennetzen J L.
Transposable elements, gene creation andgenome rearrangement in
flowering plants. Curr OpinGenet Dev, 2005, 15(6) : 62
1-627.Fedoroff N . Transposons and genome evolution in plants.Proc
Natl Acad Sci USA, 2000 ,97 (13) : 7002-7007.Shapiro J A.
Transposable elements as the key to a 21stcentury view of
evolution. Genetica, 1999, 107(1-3) :17 1-179.Talbert L E, Chandler
V L. Characterization of a highlyconserved sequence related to
Murator transposable ele-ments in maize. M ol Biol Evol, 1988, 5(5)
: 519-529.Yu Z, Wright S I, Bureau T E. Mutator-like elements
inArabidopsis thaliana: structure, diversity and
evolution.Genetics, 2000, 156(4) : 2019-2031.
-
8/6/2019 Transposon in Maize
11/11
DIAO Xian-Min et al .:MutatorTransposon n Maize and MULESin the
Plant Genom e 487
/3 Z a ' Damon Lisch'











![Transposon-Mediated Alteration of TaMATE1B Expression in ...Transposon-Mediated Alteration ofTaMATE1B Expression in Wheat Confers Constitutive Citrate Efflux from Root Apices[W] Andriy](https://img.pdfslide.us/doc/110x75/5f88f725203f795e6c4d9d31/transposon-mediated-alteration-of-tamate1b-expression-in-transposon-mediated.jpg)