Embed Size (px)
Citation preview

of April 3, 2018.This information is current as
the TestisMamu-AG Is Expressed in the Placenta andNonclassical MHC Class I Molecule A Soluble Isoform of the Rhesus Monkey
Thaddeus G. GolosAndy F. Ryan, Richard L. Grendell, Daniel E. Geraghty and
http://www.jimmunol.org/content/169/2/673doi: 10.4049/jimmunol.169.2.673
2002; 169:673-683; ;J Immunol
Referenceshttp://www.jimmunol.org/content/169/2/673.full#ref-list-1
, 14 of which you can access for free at: cites 45 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2002 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

A Soluble Isoform of the Rhesus Monkey Nonclassical MHCClass I Molecule Mamu-AG Is Expressed in the Placenta andthe Testis1
Andy F. Ryan,2* Richard L. Grendell, 2* Daniel E. Geraghty,‡ and Thaddeus G. Golos3*†
The nonclassical MHC class I locusHLA-G is expressed primarily in the placenta, although other sites of expression have beennoted in normal and pathological situations. In addition, soluble HLA-G isoforms have been detected in the serum of pregnant andnonpregnant women as well as men. The rhesus monkey placenta expresses a novel nonclassical MHC class I molecule Mamu-AG,which has features remarkably similar to those of HLA-G. We determined that the rhesus placenta expressesMamu-AG mRNA(Mamu-AG5), retaining intron 4 as previously noted in HLA-G5. Immunostaining experiments with Ab 16G1 against the solubleHLA-G5 intron 4 peptide demonstrated that an immunoreactive protein(s) was present in the syncytiotrophoblasts of the chorionicvilli of the rhesus placenta, within villous cytotrophoblasts, and occasionally within cells of the villous stroma. TheMamu-AG5mRNA was readily detected in rhesus testis (although not in ejaculated sperm). Whereas an Ab against membrane-boundMamu-AG stained few cells, primarily in the interstitium of the testis, there was consistent immunostaining for Mamu-AG5 in cellswithin the seminiferous tubules, which was corroborated by localization ofMamu-AG mRNA by in situ hybridization. Whileprimary spermatocytes were negative, Sertoli cells, spermatocytes, and spermatids were consistently positive for 16G1 immuno-staining. The specific recognition of the soluble Mamu-AG isoform was confirmed by Western blotting of Mamu-AG5 expressedin heterologous cells. The results demonstrate that a soluble nonclassical MHC class I molecule is expressed in the rhesus monkeyplacenta and testis, and confirm and extend the unique homology between HLA-G and the rhesus nonclassical moleculeMamu-AG. The Journal of Immunology, 2002, 169: 673–683.
T he nonclassical MHC class I molecule HLA-G has beenshown to be expressed on the surface of invasive extra-villous trophoblasts of the human placenta, with potential
significance for regulation of the maternal immune response topregnancy (1–6). As with other MHC class I molecules, HLA-G isexpressed on the cell surface complexed with �2-microglobulinand interacts with a variety of receptors on cells of the innateimmune system, including NK cells and monocytes (7–11). One ofthe intriguing characteristics of the molecular biology of HLA-G isthe expression of an alternatively spliced transcript coding for atruncated, soluble molecule (12, 13), now designated HLA-G5.These studies have also identified intron 4-retaining isoforms lack-ing the �2 domain (HLA-G6). In these transcripts, the fourth in-tron is retained, and the reading frame of the intron results in aunique 21-aa carboxyl terminus following the fourth exon. A stopcodon in the intron 4 sequence deletes the 41 aa of the fifth andsixth exons, including the transmembrane domain. Initial studieswith an Ab (16G1) raised against the unique C-terminal extensiondetected the protein within the placenta (14), but not in peripheral
lymphocytes. In addition, several groups have now reported assaysfor detecting soluble HLA-G in peripheral blood and other bio-logical fluids (15–18). Unexpectedly, soluble HLA-G was detectednot only in pregnancy, but also in nonpregnant women as well asmales. Recent studies suggested that HLA-G5, one isoform of sol-uble HLA-G, can induce apoptosis in peripheral CD8� T cells viaa Fas-Fas ligand mechanism (19), can suppress an allogeneic pro-liferative T cell response in mixed lymphocyte cultures (20), andcan inhibit peripheral blood NK cell-mediated cytotoxicity (21).These results collectively suggest that while membrane-boundHLA-G may interact with resident leukocytes within the decidua,soluble HLA-G may act on blood cells in the peripheral circulationin both pregnant and nonpregnant individuals.
The consistent observation of soluble HLA-G in nonpregnantwomen as well as men raises a number of provocative questions,including the source(s) of this molecule as well as its function bothduring and outside the setting of pregnancy. To address these ques-tions in an experimental animal model, we have characterized theexpression of a nonclasical MHC class I molecule, Mamu-AG, aputative HLA-G homolog, in the rhesus monkey placenta.Mamu-AG shares a number of biochemical and molecular featuresof HLA-G, including a relatively low level of polymorphism, thepresence of alternatively spliced mRNAs and multiple isoelectricisoforms, and a high level of expression at the maternal-fetal in-terface (22–24). We have also previously shown that, as withHLA-G, the mRNA for Mamu-AG has restricted expression in ex-traplacental tissues (25). In the current study we have shown thatthe placenta as well as the testis in the rhesus monkey expresses atranscript that could encode a soluble Mamu-AG retaining thefourth intron, which contains a premature stop codon. Both pla-centa and testis demonstrate reproducible immunostaining withmAb 16G1, raised against the fourth intron-encoded C-terminal
*Wisconsin Regional Primate Research Center and †Department of Obstetrics andGynecology, University of Wisconsin, Madison, WI 53715; and ‡Fred HutchinsonCancer Research Center, Seattle, WA 98104
Received for publication October 26, 2001. Accepted for publication May 10, 2002.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants HD34215 andHD37120, and ES09090 to the Developmental Toxicology Center. This is Publication41-018 of the Wisconsin Regional Primate Research Center.2 A.F.R. and R.L.G. contributed equally to this work.3 Address correspondence and reprint requests to Dr. Thaddeus G. Golos, WisconsinRegional Primate Research Center, University of Wisconsin, 1223 Capitol Court,Madison, WI 53715-1299. E-mail address: [email protected]
The Journal of Immunology
Copyright © 2002 by The American Association of Immunologists, Inc. 0022-1767/02/$02.00
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

peptide of HLA-G. Expression in the testis was strikingly cell spe-cific, suggesting that the rhesus monkey will be an excellent modelto study the physiology of a soluble nonclasical MHC class Imolecule.
Materials and MethodsAnimals and tissues
Adult female rhesus monkeys (Macaca mulatta) used for timed matingswere from the colony maintained at Wisconsin Regional Primate ResearchCenter. Rhesus monkey placental tissues were obtained by cesarean sectionas we have previously described (26). Other tissues were obtained fromhealthy animals euthanized in other studies. All surgical procedures wereperformed in accordance with the National Institutes of Health Guide forthe Care and Use of Laboratory Animals and under approval of the Uni-versity of Wisconsin Graduate School animal care and use committee.
RT-PCR, cloning, and sequencing
In RT-PCR experiments, 1 �g total RNA was reverse transcribed to cDNAusing an oligo(dT) primer and Moloney murine leukemia virus reversetranscriptase (PerkinElmer, Foster City, CA). The cDNA was amplified for34 cycles, with each cycle consisting of 94°C for 30 s, 55°C for 30 s, and72°C at 1 min, in a reaction mixture containing 10 mM Tris-HCl, 1.5 mMMgCl2, 50 mM KCl, 0.25 mM of each dNTP, and 1.25 U AmpliTaq DNApolymerase (PerkinElmer). Primers used for the amplification and sequenc-ing of rhesus MHC class I cDNAs are depicted in Table I. Amplificationreactions using as template RT reactions to which no RNA or no reversetranscriptase had been added served as negative controls. The PCR prod-ucts were separated on 3% agarose gels, isolated with the GeneClean II kit(BIO 101, Vista, CA), and subcloned using a TA cloning kit (Invitrogen,Carlsbad, CA). Subcloned fragments were sequenced using an ABI 377automated sequencing machine and the BigDye Terminator mix (PE Ap-plied Biosystems, Foster City, CA).
Immunohistochemistry
Tissues collected at surgery or necropsy were immediately prepared forfrozen sections. After fixing in 2% paraformaldehyde for 4 h, the tissueswere washed with PBS twice for 20 min each time. They were then de-hydrated in 9% sucrose for 4 h and 20% sucrose overnight. The tissueswere then embedded in OCT mounting medium (Sakura Finetek U.S.A.,Torrance, CA) and frozen in isopentane cooled with dry ice and ethanol.The mouse mAbs 25D3, 16G1, and goat anti-�2-microglobulin (DAKO,Carpinteria, CA) were used at a concentration of 5 �g/ml. W6/32 was usedat 1 �g/ml. A mouse IgG1 � Ab (Sigma-Aldrich, St. Louis, MO) or goatpolyclonal IgG antiserum (Santa Cruz Biotechnology, Santa Cruz, CA)were used as negative controls at the same concentrations as the primaryAbs. Immunostaining was performed as previously reported (23, 24). Pep-tide blocking experiments were performed on placental and testicular sec-tions with 16G1 by incubating the Ab with its immunizing peptide or anonspecific peptide, GnRH, as a control. The Ab was mixed with eitherpeptide at molar concentrations of 4/1 and 10/1 (peptide/Ab) and incubatedfor 30 min at 0°C. During this time, the sections were blocked with animal
serum. After Ab neutralization, the mix was added to the sections for 1 h.16G1 alone and mouse IgG were used as additional controls. The remain-der of the procedure was performed as described above. Peroxidase andalkaline phosphatase kits were used along with diaminobenzidene, VectorNovaRed, Vector Red, and Vector Blue substrate kits (Vector Laboratories,Burlingame, CA). Some slides were counterstained with hematoxylin.
In situ hybridization (ISH)4
For ISH, placental and testicular tissues were prepared for paraffin sections.Tissues were fixed in 2% paraformaldehyde for 4 h and then embedded inparaffin. Ten-micrometer sections were cut and processed as previouslydescribed (23). A digoxigenin-labeled 141-bp RNA probe derived fromexon 2 of Mamu-AG, as described previously (23), was generated for ISH(digoxigenin labeling kit, Roche, Indianapolis, IN). Transcription of senseand antisense RNA was confirmed by dot blot and gel electrophoresis.Hybridization buffer containing 400 �g/ml probe was applied to sections,which were then covered with a glass coverslip to prevent drying, andincubated at 55°C for up to 40 h. Slides were washed extensively to ensurespecific hybridization (23). The slides were washed in 2� SSC with 0.1%SDS (0.3 M sodium chloride and 0.03 M sodium citrate) for 30 min, placedin 2� SSC at 60°C for 1 h, 0.2� SSC at room temperature for 15 min, and0.1� SSC at 60°C for 1 h. Sections were blocked with 25% goat serum inPBS for 30 min and then incubated with alkaline phosphatase conjugatedanti-digoxigenin (1/2000; Roche) for 2 h. Sections were washed in TBS for2, 10, and 10 min. Slides were incubated in detection buffer for 5 min andthen covered with the substrate BM-purple (Roche) overnight.
Recombinant Mamu-AG5 expression and Western blotting
The RNA for Mamu-AG5 was amplified using RT-PCR and primers inexons 2 and 6 containing the start codon and the stop codon, respectively(Table I, primers 6 and 7). The sequence for an enterokinase cleavage site(GACGATGACGATAAG) and hemagglutinin (HA) protein tag (TACCCATACGATGTTCCGGATTACGCTAGCCTC) were then added to the3� region of the transcript by a second round of PCR. The resulting cDNAwas directly sequenced and subcloned into pGEM vector (Promega, Mad-ison, WI) for subsequent transfection into 293 cells. 293 cells were tran-siently transfected according to a standard calcium phosphate transfectionprocedure, and cells were harvested after 48 h of culture, lysed, and pre-pared for Western blotting. Protein samples were prepared from culturedtransfected 293 cells by lysis in 25 mM Tris-phosphate (pH 7.8), 2 mMDTT, 2 mM 1,2-diaminocyclohexane-N,N,N�,N�-tetraacetic acid, 10%glycerol, and 1% Triton X-100 , (E1531; Promega). Samples were quan-titated using the Bradford assay. Discontinuous SDS-PAGE was performedusing a 10% acrylamide resolving gel and a 3.9% stacking gel. Fifty to 100�g cell extract was fractionated along with Rainbow markers (RPN 800;Amersham, Arlington Heights, IL) to estimate m.w. After electrophoresis,proteins were electroblotted onto a polyvinylidene difluoride membrane(Bio-Rad, Hercules, CA). The membrane was washed twice with TBS andblocked with a 0.2% nonfat dry milk/TBS solution overnight at 4°C. Fol-lowing blocking, the membrane was washed three times with a 0.1%
4 Abbreviations used in this paper: ISH, in situ hybridization; HA, hemagglutinin.
Table I. Primers used for soluble Mamu-AG RT-PCR
Primera Sequenceb Nucleotide No.c
1 5�-AGA ACA TGA AGA CCG CGA CAC AGA CCT A-3� 267–2942 5�-GAC CCC CCC AAG ACA AAT-3� 620–6373 5�-CTC ACC TTG AGA TGG GGT AAA G-3� 881–896d
4 5�-TGG GAA AAG AGG GGA AGG TGA GGG GTC C-3� 101–74e
5 5�-CAG CCT GAG AGT AGC TCC CGC C-3� 1043–10226 5�-ATG GCG GTC ATG GCG CCC CGA ACC-3� 2–257f 5�-AAG GTC TCC AGA GAG GCT CCG-3� 62–42e
a Primers location is schematically illustrated in Fig. 1A.b All primers are listed in the 5� to 3� orientation.c Nucleotide numbers are with respect to the sequence of Mamu-AG *0302 (Ref. 22, GenBank accession no. U84879).d The six nucleotides at the 3� terminus (GTAAAG) are derived from intron 4 of Mamu-AG (GenBank accession no.
AY059404).e The nucleotide numbers refer to the intron 4 sequence (GenBank accession no. AY059404).f For primer 7, the sequence shown reflects the intron 4 sequences used to amplify a Mamu-AG5 cDNA, numbered according
to intron 4 sequence (GenBank accession no. AY059404). The primer also contained this sequence at its 5� end: CGT ATG GTACT TAT CGT CAT CGTC, which encoded part of the HA epitope tag (nt 1–10) and the enterokinase recognition site (nt11–25).
674 SOLUBLE MHC IN RHESUS PLACENTA AND TESTIS
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Tween 20/TBS solution, then incubated with the anti-HA Ab (0.5 �g/ml;Roche) or 16G1 (5 �g/ml) for 2 h at room temperature. The membraneswere washed three times for 10 min each time and then incubated withalkaline phosphatase-conjugated goat anti-mouse secondary Ab (1/3000;Bio-Rad) for 1.5 h. The membrane was washed three times and incubatedwith Immun-Star chemiluminescent substrate solution (Bio-Rad) for 5 min,and immunoreactive bands were visualized by autoradiography.
ResultsRT-PCR reveals expression of a transcript encoding a solubleMamu-AG
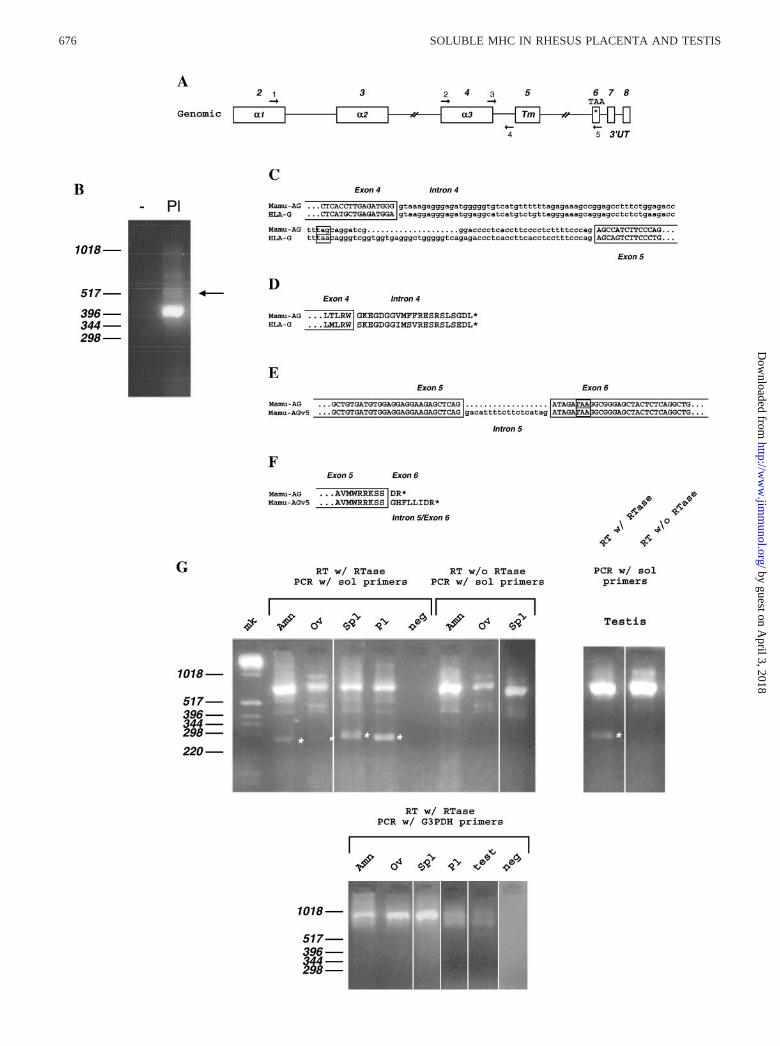
We investigated whether the placenta expresses a transcript forMamu-AG retaining intron 4, as with the soluble HLA-G tran-scripts previously reported in the human placenta and other tissues.First, we amplified reversed transcribed cDNA with primers 2 and5 located in the fourth and sixth exons, respectively, reasoning thatsince the mRNA was only detectable by RT-PCR in human pla-centa, we might find the same situation with the rhesus placenta.Fig. 1B demonstrates that several amplicons dependent on reversetranscription were obtained from rhesus placental RNA. Whereasthe major band seen in Fig. 1B primarily contained the expectedsequences of the fourth, fifth, and sixth exons, cloning and se-quencing of the band indicated by an arrow revealed the presenceof an intron highly homologous to the human HLA-G intron (seebelow). The intron sequence from this mRNA was confirmed torepresent intron 4 in its entirety by sequencing PCR product am-plified from rhesus genomic DNA (not shown). Most significantly,a stop codon was noted at the same location as that for solubleHLA-G (Fig. 1C). Since the intron 4-containing PCR ampliconmight be derived from a splicing intermediate, we sought to con-firm the splicing at exon-intron junctions 5� from the fourth intronregion in putative soluble Mamu-AG transcripts. For this experi-ment we designed downstream primer 4 within the fourth intron,and upstream primer 1 within the second exon (�1 domain) todetermine whether transcripts containing the fourth intron alsocontained other upstream introns. We cloned and sequenced sevenplacental cDNAs amplified by these primers from three differentplacentas, and none was found to contain intron 2 or 3 (not shown).We concluded that the soluble Mamu-AG cDNAs represent maturemRNAs retaining only the fourth intron. In addition, these clonesall contained exons 2, 3, and 4, demonstrating that in our handsonly a Mamu-AG mRNA homologous to HLA-G5 is detected inthe placenta.
The predicted amino acid sequences for the fourth intron ofMamu-AG and the fifth intron carboxyl terminus were determined.Alignment of Mamu-AG intron 4 peptide with that of HLA-Grevealed that the predicted amino acid sequences were 86% iden-tical between soluble MamuAG and soluble HLA-G intron 4-de-rived carboxyl termini (Fig. 1D). The amino acid substitutions be-tween rhesus and human intron 4 peptides were mostlyconservative. Sequencing of placenta-derived amplicons also re-sulted in the identification of an additional novel cDNA, whichretained 18 nucleotides derived from the 3� end of the fifth intron(Fig. 1E, Mamu-AGv5). All of these clones derived from placentalmRNA lacked intron 4. The entire intron 5 was also cloned andsequenced by PCR from rhesus genomic DNA, and was 442 bp inlength (not shown). The inclusion of the intron 5 fragment resultedin an insertion of a novel six-amino acid carboxyl-terminal exten-sion N-terminal to the two amino acids encoded by exon 6 (Fig.1F). We are not aware of any reports of a similar splice variant inhuman MHC class I mRNAs.
Soluble Mamu-AG mRNA in rhesus nonplacental tissues
Our previous studies of the expression of Mamu-AG demonstratedthat the mRNA is expressed in a variety of tissues, with the highest
nonplacental level of Mamu-AG mRNA in the testes (25). Thosestudies were conducted with a ribonuclease protection assay se-lective for the Mamu-AG � 1 domain (exon 2). We re-evaluated anumber of rhesus nonplacental tissues by RT-PCR with primers 3and 5 that would selectively amplify mRNAs containing the exon4/intron 4 junction. Fig. 1G presents representative results fromseveral tissues. As expected, placental tissues had abundant intron4-containing Mamu-AG mRNA, while in most tissues this isoformwas undetectable (eye, adrenal, lymph nodes, skeletal muscle,skin, heart, lung, liver, and pancreas; not shown) or barely detect-able (ovary, Fig. 1G; kidney and intestine, not shown). Severaltissues had a moderate, but consistent, expression of intron 4-con-taining mRNA, including amniotic membranes, testes, spleen (Fig.1G) and thyroid (data not shown).
Although to our knowledge the retention of intron 4 is unique inhumans to HLA-G and is not known in classical loci, the questionarose of whether the PCR fragments amplified represented rhesusclassical loci. Sequencing of multiple PCR-generated clones fromboth testicular and placental RNA indicated that all transcriptsfrom these tissues contained an entire intron 4, and all containedthe diagnostic TAA stop codon in exon 6. Four of the six testicularand one of three spleen (but none of 19 placental) clones se-quenced that retained intron 4 also contained 18 nt of the 3� end ofintron 5 discussed above. Since the stop codon was located withinintron 4, the intron 5 sequences do not contribute to the codingsequence for these nonplacental transcripts. Several ampliconsfrom thyroid and spleen also were found to contain the stop codondiagnostic for Mamu-AG (Fig. 2) (22, 27, 28). Surprisingly, onetranscript from spleen and two from thyroid were apparently de-rived from classical MHC class I molecule(s), since they did notcontain the diagnostic stop codon (Fig. 2). Further work will berequired to understand the significance of this observation.
Western blots confirm that Ab 16G1 recognizes Mamu-AG5
We wished to explore the expression of soluble Mamu-AG iso-forms in rhesus monkey tissues with the Ab 16G1 raised againstthe carboxyl-terminal region of soluble HLA-G encoded by intron4 sequences. To evaluate recognition of Mamu-AG5 by 16G1, aMamu-AG5 cDNA was cloned into an expression vector contain-ing sequences encoding an in-frame HA tag at the 3� end of theMamu-AG5 cDNA. 293 cells were transiently transfected with theconstruct, and cell extracts of transfected and naive cells wereanalyzed by Western blot. Parallel analysis was performed with16G1 and an Ab against the HA epitope (Fig. 3). Fig. 3A demon-strates that extracts of transfected, but not naı̈ve, cells expressedthree immunoreactive bands of �35–39 kDa as identified by theanti-HA Ab. Fig. 3B demonstrates that an identical trio of bandswas identified by Ab 16G1 in transfected, but not naı̈ve, cells.
Immunocytochemical localization of placental and testicularMamu-AG
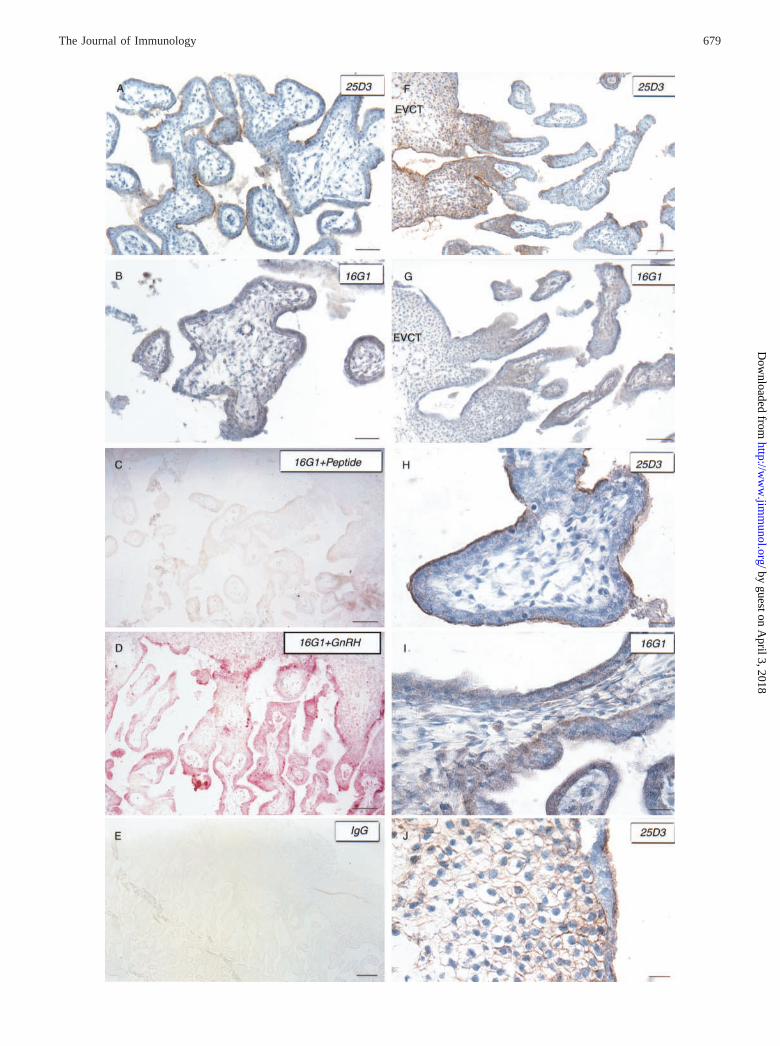
We next conducted immunocytochemical analysis for Mamu-AGin these tissues, evaluating three placentas and testes and epi-didymi from five different animals. We used an mAb (25D3)against Mamu-AG we have previously described (24), as well asan mAb against the intron 4 peptide of HLA-G (16G1) (12). Con-sistent with previous reports, 25D3 identified Mamu-AG on thevillous syncytial apical membranes (Fig. 4, A and H), in the prox-imal columns (Fig. 4F), and in the cytotrophoblastic shell delin-eating the maternal-fetal interface (Fig. 4J). Mesenchymal cells inthe core of the villi (Fig. 4H) as well as the maternal decidua anduterus were consistently negative.
675The Journal of Immunology
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
676 SOLUBLE MHC IN RHESUS PLACENTA AND TESTIS
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

By contrast, immunostaining with the 16G1 Ab was localized tothe syncytia, villous cytotrophoblasts, and some villous mesenchy-mal cells of day 36 placentas (Fig. 4, B, D, G, and I). Staining wasoften pronounced on the syncytial border toward the intervillousspace and the cytotrophoblastic edge facing the core of the villi(Fig. 4I), suggesting cytoplasmic localization. Diffuse staining wasalso observed throughout both villous syncytial and cytotropho-blastic cell layers (Fig. 4, B, D, and I). Large cells with abundantcytoplasm within the villi, presumably Hofbauer cells, frequentlydemonstrated positive staining with 16G1 (Fig. 4I). Blood vesselswithin the villous stroma were also positively stained (Fig. 4B).Extravillous cytotrophoblasts, notably in the cytotrophoblasticshell, were not stained by 16G1, unlike the results with 25D3(compare serial sections Fig. 4, F and G; also Fig. 4J). The specificityof immunostaining was demonstrated by the absence of immuno-staining when the 16G1 primary Ab was preincubated with theimmunizing peptide (Fig. 4C), but not with an unrelated peptide
(Fig. 4D). A nonspecific primary antiserum likewise gave no pos-itive staining (Fig. 4E).
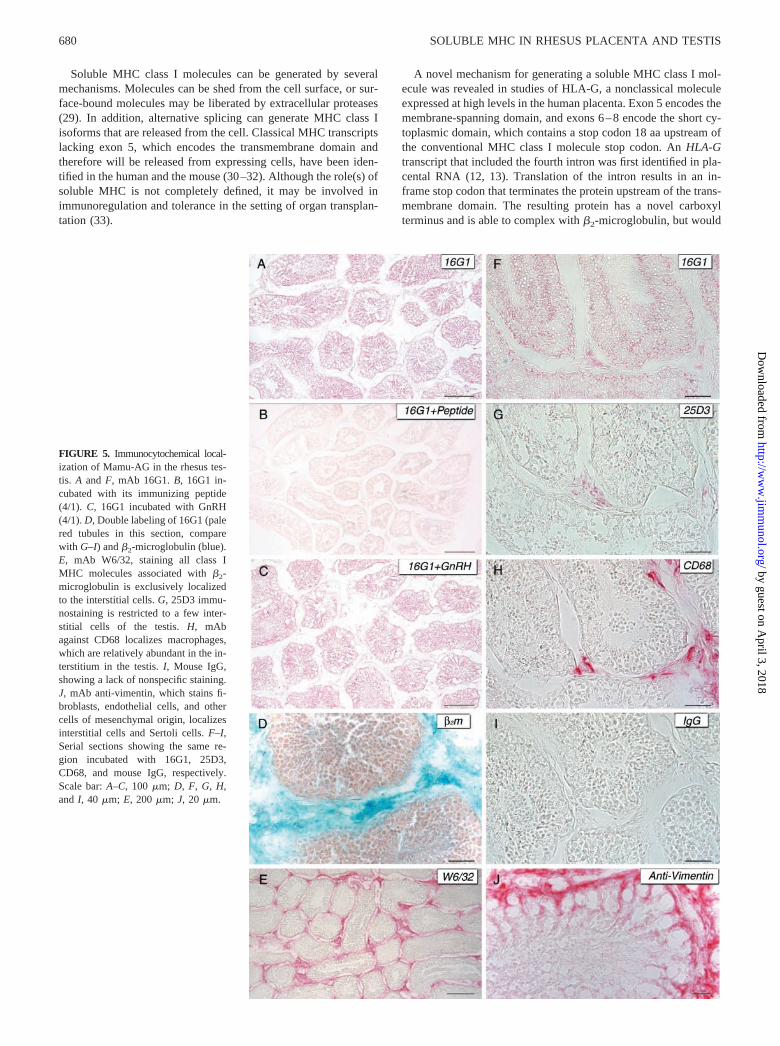
We next conducted similar immunostaining experiments withrhesus testes. 25D3 staining in the testis was minimal and waslocalized exclusively to occasional cells in the interstitium sur-rounding the seminiferous tubules (Fig. 5G). Germ cells and Ser-toli cells within the tubule were consistently negative. Althoughonly occasional interstitial cells were 25D3 positive, these cellswere found to be consistently positive on serial sections, occasion-ally appearing in small aggregates. Other sporadic cells were typ-ically within peritubular regions, and immunostaining of serial sec-tions suggested that at least some of these cells were positive forCD68, indicating their likely identity as macrophages (Fig. 5H).Serial sections (Fig. 5, F–H) demonstrate interstitial macrophagesthat appear to coexpress Mamu-AG, soluble Mamu-AG, andCD68. Epidydimal structures and mature sperm were not positivefor Mamu-AG with 25D3 (not shown).
FIGURE 2. Alignment of intron 4, exon 5, intron 5,and exon 6 nucleotide sequences cloned by RT-PCR fromrhesus placenta, testis, spleen, and thyroid. One represen-tative placental sequence is included, and only nucleo-tides that differ from a consensus sequence are shown.
FIGURE 1. Cloning and sequence analysis of soluble Mamu-AG mRNA. A, Schematic diagram of the exon organization of Mamu-AG, indicating the�1–�3 domains, transmembrane domain (Tm), stop codon (TAA), and PCR primer designations and approximate locations. B, RT-PCR amplification ofMamu-AG amplicons from rhesus placental (Pl) RNA. Primers in the fourth and fifth exons were used with placental RNA. Lane �, Amplifications whereno RNA was added to the RT reaction. The arrow indicates the band found to contain the amplicon harboring intron 4. C, Nucleotide sequence of Mamu-AGsplice variant retaining intron 4, aligned with the HLA-G intron 4 mRNA sequence. Portions of exons 4 and 5 and the putative stop codons in intron 4 areboxed. D, Predicted amino acid sequence of the intron 4-derived carboxyl terminal of soluble Mamu-AG aligned with soluble HLA-G. E, Alignment ofthe nucleotide sequence of Mamu-AG and Mamu-AG retaining a portion of intron 5 (Mamu-AGv5). F, Alignment of the carboxyl terminus of membrane-bound Mamu-AG (top) and Mamu-AGv5 (bottom). G, RT-PCR of Mamu-AG soluble isoform mRNA from rhesus nonplacental tissues. Primers at the exon4/intron 4 boundary and just 3� of the Mamu-AG TAA stop codon were used. Asterisks indicate sMamu-AG amplicons. mk, Size markers; Amn, amnioticmembrane; Ov, ovary; Spl, spleen; Pl, placenta; neg, no RNA in RT reaction. The three right-most reactions were conducted with RT reactions lackingRT, demonstrating that high m.w. bands arose from nonspecific PCR and/or genomic DNA. G, lower panel, RT-PCR of samples corresponding to upperpanel, amplified for G3PDH to demonstrate relative RNA loading.
677The Journal of Immunology
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

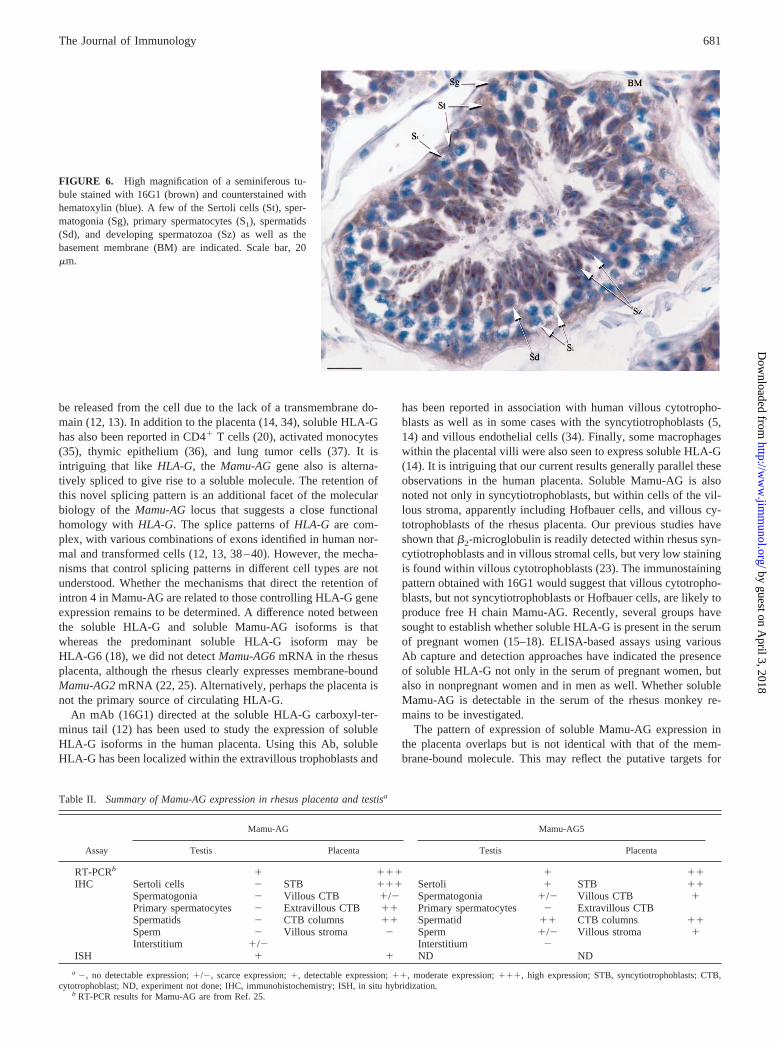
Analysis of rhesus testes with 16G1 revealed prominent immu-nostaining in the seminiferous tubules, with only occasional pos-itive cells in the interstitium (Fig. 5, A, C, and F). Staining wasnotably found in the mural aspect of the tubular compartment,through the entire cross-section of the seminiferous tubules, sug-gesting staining of Sertoli cells (e.g., Fig. 5F; see also Fig. 6).Individual Sertoli cells were identified by their position passingbetween germ cells and their pointed, irregular nuclei (Figs. 5J and6), with cytoplasmic processes extending toward the lumen andsurrounding the nuclei of advanced germ cells. Staining was notnoted in primary spermatocytes in early metaphase during meiosis,identified by their distinct speckled chromatin structure (S1, Fig.6). However, 16G1 immunostaining was abundant in late stageprimary and secondary spermatocytes and spermatids, with dis-tinctly condensed nuclei (Sd, Fig. 6). Variable staining was ob-served in spermatozoa and residual bodies. Spermatozoa undergo-ing spermiogenesis were positively stained, while those thatappeared to have completed the process and were free in the lumenwere generally not stained (Fig. 5F). Residual bodies free in thelumen were sometimes stained by 16G1. Staining was not ob-served in the epidydimi or in mature sperm. A summary of im-munostaining results is presented in Table II.
The discrepancy between 25D3 and 16G1 immunostaining inthe testis was both intriguing and puzzling. In particular, it is sur-prising that 25D3 did not detect Mamu-AG within the seminifer-ous tubules. Since the 25D3 Ab only recognizes surface-boundMamu-AG complexed with �2-microglobulin, it seemed possiblethat 16G1 immunostaining is detecting soluble Mamu-AG H chainthat is not complexed with �2-microglobulin. Immunostainingconfirmed this hypothesis (Fig. 5, D and E), since while abundant�2-microglobulin was identified within the interstitial tissue, theseminiferous tubules were devoid of immunostaining. Interest-ingly, Sertoli cells do not express �2-microglobulin.
We also evaluated several tissues previously shown to express lowlevels of Mamu-AG mRNA (25). Adrenal, spleen, ovary, kidney,heart, and pituitary samples did not reveal any Mamu-AG expressionby immunostaining with 25D3 or 16G1 (data not shown).
Mamu-AG ISH
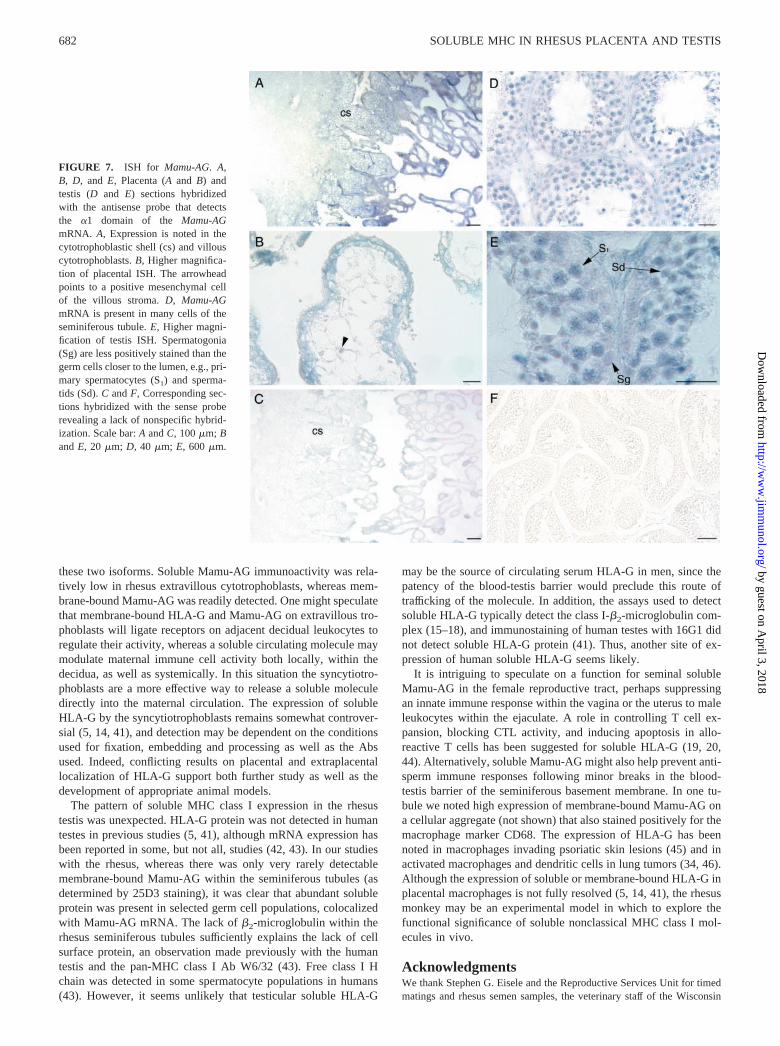
Immunocytochemical analyses indicated that the Mamu-AG tran-scripts we detected by RT-PCR were expressed in the seminiferoustubules. To confirm that the tubules contain Mamu-AG mRNA, weconducted ISH with a probe for the �1 domain, with which wehave previously defined conditions for locus-specific ISH (23). Aswe have previously reported, the villous syncytiotrophoblasts andtrophoblastic shell consistently expressed Mamu-AG mRNA (Fig.7A). In addition, villous cytotrophoblasts and occasional cellswithin the core of the villi were positive (Fig. 7B). Maternal de-cidua and uterine tissue were always negative, and no hybridiza-tion was observed with the sense probe (Fig. 7C).
ISH with rhesus testes localized Mamu-AG mRNA mainly in theseminiferous tubules (Fig. 7D). Most cells in the process of sper-matogenesis hybridized with the antisense probe. Cells occupyingthe mural compartment of the seminiferous tubule, primarily sper-matogonia and Sertoli cells, had relatively lower hybridization sig-nal compared with spermatocytes (Fig. 7D). Higher magnification(Fig. 7E) revealed that spermatogonia contained less Mamu-AGmRNA than the germ cells closer to the lumen, i.e., primary sper-matocytes and spermatids. Sertoli cells were difficult to identifywithout counterstaining during ISH. Hybridization with spermato-zoa was uncommon, but was occasionally noted. In general, all theseminiferous tubules in the testis sections demonstrated some de-gree of mRNA expression. A few of the surrounding interstitialcells of the testis, possibly macrophages or Leydig cells, werepositive for Mamu-AG mRNA (Fig. 7E). However, their stainingintensity was generally observed to be lower than that in germcells. Sections incubated with the sense probe revealed no stainingin testes (Fig. 7F).
Finally, we conducted RT-PCR for Mamu-AG mRNA exons4–6 with samples of ejaculated rhesus sperm. Cloning and se-quencing of amplified cDNAs revealed that both whole semen and“swim-up” fraction sperm contained Mamu-AG mRNA; howeverno transcripts containing intron 4 were detected.
DiscussionWe have demonstrated the expression of a mRNA encoding a sol-uble form of Mamu-AG, a nonclassical MHC class I moleculeexpressed at high levels in the rhesus monkey placenta. ThemRNA is expressed in the testes and several other nonplacentaltissues at moderate levels. Immunostaining with an Ab recognizingthe soluble HLA-G carboxyl-terminal peptide showed that the ex-pression of soluble Mamu-AG in the placenta overlapped but wasnot identical with that of cell surface Mamu-AG. While we foundonly occasional cells expressing cell surface Mamu-AG in the tes-tes, we also demonstrated widespread, although apparently cell-selective, expression of soluble Mamu-AG in the seminiferous tu-bules of the testes. Only a few other tissues had detectable solubleMamu-AG mRNA, and no immunostaining was noted outside theplacenta and testes in the tissues evaluated.
FIGURE 3. Western blot and immunocytochemical analysis of Mamu-AG5 expressed in heterologous cells. A, Lysates of Mamu-AG5-trans-fected 293 cells (lane 1) or naive cells (lane 2) analyzed by Western blotwith an Ab against the HA tag. B, Cell lysates from Mamu-AG5-trans-fected cells (lane 3, 50 �g; lane 4, 200 �g) or naive cells (lane 5) analyzedwith Ab 16G1 against soluble HLA-G.
FIGURE 4. Immunohistochemistry of Mamu-AG in the rhesus placenta. A, F, H, and J, mAb 25D3 immunostaining for membrane-bound Mamu-AG(brown), counterstained with hematoxylin (blue). Note the strong syncytial staining and the lack of localization in the core of villi (A and H). B, G, andI, mAb 16G1 immunostaining for intron 4 peptide (brown) counterstained with hematoxylin (blue). Note the diffuse staining pattern in the cytotrophoblastscompared with 25D3, including occasional cells in the core of the villi (B and I) and a villous stromal vessel as well (B). C, 16G1 preincubated with itsimmunizing peptide (4/1). Lack of staining demonstrates the specificity of 16G1 against its immunizing peptide. D, 16G1 incubated with a nonspecificpeptide, GnRH (4/1). Positive red staining on this section again reveals that 16G1 is sMamu-AG specific and not inhibited by an unrelated peptide. E,Section incubated with nonspecific mouse IgG. The lack of staining is comparable to that in C. F and G, Serial sections comparing the staining patternsof 25D3 and 16G1. Note that 25D3 clearly stains the extravillous cytotrophoblasts (EVCT), whereas 16G1 does not. J, Higher power view of F showingthe specific membrane staining of 25D3 on EVCT. Scale bar: A and B, 60 �m; C, D, F, and G, 100 �m; E, 200 �m; H–J, 20 �m.
678 SOLUBLE MHC IN RHESUS PLACENTA AND TESTIS
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
679The Journal of Immunology
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
Soluble MHC class I molecules can be generated by severalmechanisms. Molecules can be shed from the cell surface, or sur-face-bound molecules may be liberated by extracellular proteases(29). In addition, alternative splicing can generate MHC class Iisoforms that are released from the cell. Classical MHC transcriptslacking exon 5, which encodes the transmembrane domain andtherefore will be released from expressing cells, have been iden-tified in the human and the mouse (30–32). Although the role(s) ofsoluble MHC is not completely defined, it may be involved inimmunoregulation and tolerance in the setting of organ transplan-tation (33).
A novel mechanism for generating a soluble MHC class I mol-ecule was revealed in studies of HLA-G, a nonclassical moleculeexpressed at high levels in the human placenta. Exon 5 encodes themembrane-spanning domain, and exons 6–8 encode the short cy-toplasmic domain, which contains a stop codon 18 aa upstream ofthe conventional MHC class I molecule stop codon. An HLA-Gtranscript that included the fourth intron was first identified in pla-cental RNA (12, 13). Translation of the intron results in an in-frame stop codon that terminates the protein upstream of the trans-membrane domain. The resulting protein has a novel carboxylterminus and is able to complex with �2-microglobulin, but would
FIGURE 5. Immunocytochemical local-ization of Mamu-AG in the rhesus tes-tis. A and F, mAb 16G1. B, 16G1 in-cubated with its immunizing peptide(4/1). C, 16G1 incubated with GnRH(4/1). D, Double labeling of 16G1 (palered tubules in this section, comparewith G–I) and �2-microglobulin (blue).E, mAb W6/32, staining all class IMHC molecules associated with �2-microglobulin is exclusively localizedto the interstitial cells. G, 25D3 immu-nostaining is restricted to a few inter-stitial cells of the testis. H, mAbagainst CD68 localizes macrophages,which are relatively abundant in the in-terstitium in the testis. I, Mouse IgG,showing a lack of nonspecific staining.J, mAb anti-vimentin, which stains fi-broblasts, endothelial cells, and othercells of mesenchymal origin, localizesinterstitial cells and Sertoli cells. F–I,Serial sections showing the same re-gion incubated with 16G1, 25D3,CD68, and mouse IgG, respectively.Scale bar: A–C, 100 �m; D, F, G, H,and I, 40 �m; E, 200 �m; J, 20 �m.
680 SOLUBLE MHC IN RHESUS PLACENTA AND TESTIS
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
be released from the cell due to the lack of a transmembrane do-main (12, 13). In addition to the placenta (14, 34), soluble HLA-Ghas also been reported in CD4� T cells (20), activated monocytes(35), thymic epithelium (36), and lung tumor cells (37). It isintriguing that like HLA-G, the Mamu-AG gene also is alterna-tively spliced to give rise to a soluble molecule. The retention ofthis novel splicing pattern is an additional facet of the molecularbiology of the Mamu-AG locus that suggests a close functionalhomology with HLA-G. The splice patterns of HLA-G are com-plex, with various combinations of exons identified in human nor-mal and transformed cells (12, 13, 38–40). However, the mecha-nisms that control splicing patterns in different cell types are notunderstood. Whether the mechanisms that direct the retention ofintron 4 in Mamu-AG are related to those controlling HLA-G geneexpression remains to be determined. A difference noted betweenthe soluble HLA-G and soluble Mamu-AG isoforms is thatwhereas the predominant soluble HLA-G isoform may beHLA-G6 (18), we did not detect Mamu-AG6 mRNA in the rhesusplacenta, although the rhesus clearly expresses membrane-boundMamu-AG2 mRNA (22, 25). Alternatively, perhaps the placenta isnot the primary source of circulating HLA-G.
An mAb (16G1) directed at the soluble HLA-G carboxyl-ter-minus tail (12) has been used to study the expression of solubleHLA-G isoforms in the human placenta. Using this Ab, solubleHLA-G has been localized within the extravillous trophoblasts and
has been reported in association with human villous cytotropho-blasts as well as in some cases with the syncytiotrophoblasts (5,14) and villous endothelial cells (34). Finally, some macrophageswithin the placental villi were also seen to express soluble HLA-G(14). It is intriguing that our current results generally parallel theseobservations in the human placenta. Soluble Mamu-AG is alsonoted not only in syncytiotrophoblasts, but within cells of the vil-lous stroma, apparently including Hofbauer cells, and villous cy-totrophoblasts of the rhesus placenta. Our previous studies haveshown that �2-microglobulin is readily detected within rhesus syn-cytiotrophoblasts and in villous stromal cells, but very low stainingis found within villous cytotrophoblasts (23). The immunostainingpattern obtained with 16G1 would suggest that villous cytotropho-blasts, but not syncytiotrophoblasts or Hofbauer cells, are likely toproduce free H chain Mamu-AG. Recently, several groups havesought to establish whether soluble HLA-G is present in the serumof pregnant women (15–18). ELISA-based assays using variousAb capture and detection approaches have indicated the presenceof soluble HLA-G not only in the serum of pregnant women, butalso in nonpregnant women and in men as well. Whether solubleMamu-AG is detectable in the serum of the rhesus monkey re-mains to be investigated.
The pattern of expression of soluble Mamu-AG expression inthe placenta overlaps but is not identical with that of the mem-brane-bound molecule. This may reflect the putative targets for
FIGURE 6. High magnification of a seminiferous tu-bule stained with 16G1 (brown) and counterstained withhematoxylin (blue). A few of the Sertoli cells (St), sper-matogonia (Sg), primary spermatocytes (S1), spermatids(Sd), and developing spermatozoa (Sz) as well as thebasement membrane (BM) are indicated. Scale bar, 20�m.
Table II. Summary of Mamu-AG expression in rhesus placenta and testisa
Assay
Mamu-AG Mamu-AG5
Testis Placenta Testis Placenta
RT-PCRb � ��� � ��IHC Sertoli cells � STB ��� Sertoli � STB ��
Spermatogonia � Villous CTB �/� Spermatogonia �/� Villous CTB �Primary spermatocytes � Extravillous CTB �� Primary spermatocytes � Extravillous CTBSpermatids � CTB columns �� Spermatid �� CTB columns ��Sperm � Villous stroma � Sperm �/� Villous stroma �Interstitium �/� Interstitium �
ISH � � ND ND
a �, no detectable expression; �/�, scarce expression; �, detectable expression; ��, moderate expression; ���, high expression; STB, syncytiotrophoblasts; CTB,cytotrophoblast; ND, experiment not done; IHC, immunohistochemistry; ISH, in situ hybridization.
b RT-PCR results for Mamu-AG are from Ref. 25.
681The Journal of Immunology
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from
these two isoforms. Soluble Mamu-AG immunoactivity was rela-tively low in rhesus extravillous cytotrophoblasts, whereas mem-brane-bound Mamu-AG was readily detected. One might speculatethat membrane-bound HLA-G and Mamu-AG on extravillous tro-phoblasts will ligate receptors on adjacent decidual leukocytes toregulate their activity, whereas a soluble circulating molecule maymodulate maternal immune cell activity both locally, within thedecidua, as well as systemically. In this situation the syncytiotro-phoblasts are a more effective way to release a soluble moleculedirectly into the maternal circulation. The expression of solubleHLA-G by the syncytiotrophoblasts remains somewhat controver-sial (5, 14, 41), and detection may be dependent on the conditionsused for fixation, embedding and processing as well as the Absused. Indeed, conflicting results on placental and extraplacentallocalization of HLA-G support both further study as well as thedevelopment of appropriate animal models.
The pattern of soluble MHC class I expression in the rhesustestis was unexpected. HLA-G protein was not detected in humantestes in previous studies (5, 41), although mRNA expression hasbeen reported in some, but not all, studies (42, 43). In our studieswith the rhesus, whereas there was only very rarely detectablemembrane-bound Mamu-AG within the seminiferous tubules (asdetermined by 25D3 staining), it was clear that abundant solubleprotein was present in selected germ cell populations, colocalizedwith Mamu-AG mRNA. The lack of �2-microglobulin within therhesus seminiferous tubules sufficiently explains the lack of cellsurface protein, an observation made previously with the humantestis and the pan-MHC class I Ab W6/32 (43). Free class I Hchain was detected in some spermatocyte populations in humans(43). However, it seems unlikely that testicular soluble HLA-G
may be the source of circulating serum HLA-G in men, since thepatency of the blood-testis barrier would preclude this route oftrafficking of the molecule. In addition, the assays used to detectsoluble HLA-G typically detect the class I-�2-microglobulin com-plex (15–18), and immunostaining of human testes with 16G1 didnot detect soluble HLA-G protein (41). Thus, another site of ex-pression of human soluble HLA-G seems likely.
It is intriguing to speculate on a function for seminal solubleMamu-AG in the female reproductive tract, perhaps suppressingan innate immune response within the vagina or the uterus to maleleukocytes within the ejaculate. A role in controlling T cell ex-pansion, blocking CTL activity, and inducing apoptosis in allo-reactive T cells has been suggested for soluble HLA-G (19, 20,44). Alternatively, soluble Mamu-AG might also help prevent anti-sperm immune responses following minor breaks in the blood-testis barrier of the seminiferous basement membrane. In one tu-bule we noted high expression of membrane-bound Mamu-AG ona cellular aggregate (not shown) that also stained positively for themacrophage marker CD68. The expression of HLA-G has beennoted in macrophages invading psoriatic skin lesions (45) and inactivated macrophages and dendritic cells in lung tumors (34, 46).Although the expression of soluble or membrane-bound HLA-G inplacental macrophages is not fully resolved (5, 14, 41), the rhesusmonkey may be an experimental model in which to explore thefunctional significance of soluble nonclassical MHC class I mol-ecules in vivo.
AcknowledgmentsWe thank Stephen G. Eisele and the Reproductive Services Unit for timedmatings and rhesus semen samples, the veterinary staff of the Wisconsin
FIGURE 7. ISH for Mamu-AG. A,B, D, and E, Placenta (A and B) andtestis (D and E) sections hybridizedwith the antisense probe that detectsthe �1 domain of the Mamu-AGmRNA. A, Expression is noted in thecytotrophoblastic shell (cs) and villouscytotrophoblasts. B, Higher magnifica-tion of placental ISH. The arrowheadpoints to a positive mesenchymal cellof the villous stroma. D, Mamu-AGmRNA is present in many cells of theseminiferous tubule. E, Higher magni-fication of testis ISH. Spermatogonia(Sg) are less positively stained than thegerm cells closer to the lumen, e.g., pri-mary spermatocytes (S1) and sperma-tids (Sd). C and F, Corresponding sec-tions hybridized with the sense proberevealing a lack of nonspecific hybrid-ization. Scale bar: A and C, 100 �m; Band E, 20 �m; D, 40 �m; E, 600 �m.
682 SOLUBLE MHC IN RHESUS PLACENTA AND TESTIS
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

Regional Primate Research Center for surgical assistance, members of theGolos laboratory for critical reading of the manuscript, Carlos Quian-Su-arez (Georgetown University, Washington, D.C.) for advice on histologicalinterpretation, and Steven Busch for editorial assistance. We acknowledgethe assistance of George Flentke and the Microanatomy Core of the Uni-versity of Wisconsin Developmental Toxicology Center for helpful train-ing and advice for riboprobe preparation.
References1. Kovats, S., E. K. Main, C. Librach, M. Stubblebine, S. J. Fisher, and R. DeMars.
1990. A class I antigen, HLA-G, expressed in human trophoblasts. Science 248:220.
2. Loke, Y. W., and A. King. 2000. Decidual natural-killer-cell interaction withtrophoblast: cytolysis or cytokine production? Biochem. Soc. 28:196.
3. Hunt, J. S., M. G. Petroff, P. Morales, P. Sedlmayr, D. E. Geraghty, and C. Ober.2000. HLA-G in reproduction: studies on the maternal-fetal interface. Hum. Im-munol. 61:1113.
4. Le Bouteiller, P. 2000. HLA-G in the human placenta: expression and potentialfunctions. Biochem. Soc. Trans. 28:208.
5. Paul, P., N. Rouas-Freiss, P. Moreau, F. A. Cabestre, C. Menier, I. Khalil-Daher,C. Pangault, M. Onno, R. Fauchet, J. Martinez-Laso, et al. 2000. HLA-G, -E, -Fpreworkshop: tools and protocols for analysis of non-classical class I genes tran-scription and protein expression. Hum. Immunol. 61:1177.
6. Carosella, E. D., P. Moreau, S. Aractingi, and N. Rouas-Freiss. 2001. HLA-G: ashield against inflammatory aggression. Trends Immunol. 22:553.
7. Ponte, M., C. Cantoni, R. Biassoni, A. Tradori-Cappai, G. Bentivoglio, C. Vitale,S. Bertone, A. Moretta, and M. C. Mingari. 1999. Inhibitory receptors sensingHLA-G1 molecules in pregnancy: decidua-associated natural killer cells expressLIR-1 and CD94/NKG2A and acquire p49, an HLA-G1-specific receptor. Proc.Natl. Acad. Sci. USA 96:5343.
8. Cantoni, C., S. Verdiani, M. Falco, A. Pessino, R. Cilli, R. Conte, D. Pende,M. Ponte, M. S. Mikaelsson, L. Moretta, et al. 1998. p49, a putative HLA classI-specific inhibitory receptor belonging to the immunoglobulin superfamily. Eur.J. Immunol. 28:1980.
9. Colonna, M., J. Samaridis, M. Cella, L. Angman, R. L. Allen, C. A. O’Callaghan,R. Dunbar, G. S. Ogg, V. Cerundolo, and A. Relink. 1998. Human myelomono-cytic cells express an inhibitory receptor for classical and nonclassical MHC classI molecules. J. Immunol. 160:3096.
10. Navarro, F., M. Llano, T. Bellon, M. Colonna, D. E. Geraghty, andM. Lopez-Botet. 1999. The ILT2(LIR1) and CD94/NKG2A NK cell receptorsrespectively recognize HLA-G1 and HLA-E molecules co-expressed on targetcells. Eur. J. Immunol. 29:277.
11. Rajagopalan, S., and E. O. Long. 1999. A human histocompatibility leukocyteantigen (HLA)-G-specific receptor expressed on all natural killer cells. J. Exp.Med. 189:1093.
12. Fujii, T., A. Ishitani, and D. E. Geraghty. 1994. A soluble form of the HLA-Gantigen is encoded by a messenger ribonucleic acid containing intron 4. J. Im-munol. 153:5516.
13. Moreau, P., E. Carosella, M. Teyssier, S. Prost, E. Gluckman, J. Dausset, andM. Kirszenbaum. 1995. Soluble HLA-G molecule: an alternatively splicedHLA-G mRNA form candidate to encode it in peripheral blood mononuclear cellsand human trophoblasts. Hum. Immunol. 43:231.
14. Chu, W., M. E. Fant, D. E. Geraghty, and J. S. Hunt. 1998. Soluble HLA-G inhuman placentas: synthesis in trophoblasts and interferon-�-activated macro-phages but not placental fibroblasts. Hum. Immunol. 59:435.
15. Hamai, Y., T. Fujii, A. Miki, D. E. Geraghty, I. Harada, Y. Takai, S. Kozuma,O. Tsutsumi, and Y. Taketani. 1999. Quantitative assessment of human leukocyteantigen-G protein in amniotic fluid by a double-determinant enzyme-linked im-munosorbent assay using anti-human leukocyte antigen-G-specific antibody“87G.” Am. J. Reprod. Immunol. 41:293.
16. Fournel, S., M. Aguerre-Girr, A. Campan, L. Salauze, A. Berrebi, Y.-C. Lone,F. Lenfant, and P. Le Bouteiller. 1999. Soluble HLA-G: purification from eu-karyotic transfected cells and detection by a specific ELISA. Am. J. Reprod.Immunol. 42:22.
17. Rebmann, V., K. Pfeiffer, M. Passler, S. Ferrone, S. Maier, E. Weiss, andH. Grosse-Wilde. 1999. Detection of soluble HLA-G molecules in plasma andamniotic fluid. Tissue Antigens 53:14.
18. Hunt, J. S., L. Jadhav, W. Chu, D. E. Geraghty, and C. Ober. 2000. SolubleHLA-G circulates in maternal blood during pregnancy. Am. J. Obstet. Gynecol.183:682.
19. Fournel, S., M. Aguerre-Girre, X. Huc, F. Lenfant, A. Alam, A. Toubert, B. A.,and P. Le Bouteiller. 2000. Cutting edge: soluble HLA-G1 triggers CD95/CD95ligand-mediated apoptosis in activated CD8� cells by interacting with CD8.J. Immunol. 164:6100.
20. Lila, N., N. Rouas-Freiss, J. Dausset, A. Carpentier, and E. D. Carosella. 2001.Soluble HLA-G protein secreted by allo-specific CD4� T cells suppresses theallo-proliferative response: a CD4� T cell regulatory mechanism. Proc. Natl.Acad. Sci. USA 98:12150.
21. Marchal-Bras-Goncalves, R., N. Rouas-Freiss, F. Connan, J. Choppin, J. Dausset,E. D. Carosella, M. Kirszenbaum, and J. Guillet. 2001. A soluble HLA-G proteinthat inhibits natural killer cell-mediated cytotoxicity. Transplant. Proc. 33:2355.
22. Boyson, J. E., K. K. Iwanaga, T. G. Golos, and D. I. Watkins. 1997. Identificationof a novel MHC class I gene, Mamu-AG in the placenta of primate with aninactivated G locus. J. Immunol. 159:3311.
23. Slukvin, I. I., J. E. Boyson, D. I. Watkins, and T. G. Golos. 1998. The rhesusmonkey analogue of human lymphocyte antigen-G is expressed primarily in thevillous syncytiotrophoblast. Biol. Reprod. 58:728.
24. Slukvin, I. I., D. P. Lunn, D. I. Watkins, and T. G. Golos. 2000. Placental ex-pression of the nonclassical MHC class I molecule Mamu-AG at implantation inthe rhesus monkey. Proc. Natl. Acad. Sci. USA 97:9104.
25. Slukvin, I. I., D. I. Watkins, and T. G. Golos. 1999. Tissue distribution of themRNA for a rhesus monkey class Ib molecule Mamu-AG. Tissue Antigens 53:282.
26. Golos, T. G., R. R. Handrow, M. Durning, J. M. Fisher, and J. K. Rilling. 1992.Regulation of chorionic gonadotropin-� and chorionic somatomammotropin mes-senger ribonucleic acid expression by 8-bromo-adenosine 3�,5�-monophosphateand dexamethasone in cultured rhesus monkey syncytiotrophoblasts. Endocrinol-ogy 131:89.
27. Boyson, J. E., K. K. Iwanaga, J. A. Urvater, A. L. Hughes, T. G. Golos, andD. I. Watkins. 1999. Evolution of a new nonclassical MHC class I locus in twoOld World primate species. Immunogenetics 49:86.
28. Urvater, J. A., N. Otting, J. H. Loehrke, R. Rudersdorf, I. I. Slukvin,M. S. Piekarczyk, T. G. Golos, A. L. Hughes, R. E. Bontrop, and D. I. Watkins.2000. Mamu-I: a novel primate MHC class I B-related locus with unusually lowvariability. J. Immunol. 164:1386.
29. Demaria, S., L. D. DeVito-Haynes, R. D. Salter, W. J. Burlingham, andY. Bushkin. 1999. Peptide-conformed �2m-free class I heavy chains are inter-mediates in generation of soluble HLA by the membrane-bound metalloprotein-ase. Hum. Immunol. 60:1216.
30. Krangel, M. S. 1985. Unusual RNA splicing generates a secreted form ofHLA-A2 in a mutagenized B lymphoblastoid cell line. EMBO J. 4:1205.
31. Krangel, M. S. 1986. Secretion of HLA-A and -B antigens via an alternative RNAsplicing pathway. J. Exp. Med. 163:1173.
32. Ulker, N., K. D. Lewis, L. E. Hood, and I. Stroynowski. 1990. Activated T cellstranscribe an alternatively spliced mRNA encoding a soluble form of Qa-2 an-tigen. EMBO J. 9:3839.
33. Burlingham, W. J. 2000. Soluble MHC, immunoregulation, and tolerance: aprogress report. Hum. Immunol. 61:1316.
34. Blaschitz, A., F. Lenfant, V. Mallet, M. Hartmann, A. Bensussan, D. E. Geraghty,P. Le Bouteiller, and G. Dohr. 1997. Endothelial cells in chorionic fetal vesselsof first trimester placenta express HLA-G. Eur. J. Immunol. 27:3380.
35. Onno, M., G. Le Friec, C. Pangault, L. Amiot, V. Guilloux, B. Drenou,S. Caulet-Maugendre, P. Andre, and R. Fauchet. 2000. Modulation of HLA-Gantigens expression in myelomonocytic cells. Hum. Immunol. 61:1086.
36. Mallet, V., A. Blaschitz, L. Crisa, C. Schmitt, S. Fournel, A. King, Y. W. Loke,G. Dohr, and P. Le Bouteiller. 1999. HLA-G in the human thymus: a subpopu-lation of medullary epithelial but not CD83� dendritic cells expresses HLA-G asa membrane-bound and soluble protein. Intl. Immunol. 11:889.
37. Urosevic, M., M. O. Kurrer, J. Kamarashev, B. Mueller, W. Weder, G. Burg,R. A. Stahel, R. Dummer, and A. Trojan. 2001. Human leukocyte antigen Gup-regulation in lung cancer associates with high-grade histology, human leuko-cyte antigen class I loss and interleukin-10 production. Am. J. Pathol. 159:817.
38. Ishitani, A., and D. E. Geraghty. 1992. Alternative splicing of HLA-G transcriptsyields proteins with primary structures resembling both class I and class II an-tigens. Proc. Natl. Acad. Sci. USA 89:2669.
39. Riteau, B., N. Rouas-Freiss, C. Menier, P. Paul, J. Dausset, and E. D. Carosella.2001. HLA-G2, -G3, and -G4 isoforms expressed as nonmature cell surface gly-coproteins inhibit NK and antigen-specific CTL cytolysis. J. Immunol. 166:5018.
40. Kirszenbaum, M., P. Moreau, E. Gluckman, J. Dausset, and E. Carosella. 1994.An alternatively spliced form of HLA-G mRNA in human trophoblasts and ev-idence for the presence of HLA-G transcript in adult lymphocytes. Proc. Natl.Acad. Sci. USA 91:4209.
41. Blaschitz, A., H. Hutter, V. Leitner, S. Pilz, R. Wintersteiger, G. Dohr, andP. Sedlmayr. 2000. Reaction patterns of monoclonal antibodies to HLA-G inhuman tissues and on cell lines: a comparative study. Hum. Immunol. 61:1074.
42. Fiszer, D., M. Ulbrecht, N. Fernandez, J. P. Johnson, E. H. Weiss, andM. Kurpisz. 1997. Analysis of HLA class Ib gene expression in male gameto-genic cells. Eur. J. Immunol. 27:1691.
43. Hutter, H., and G. Dohr. 1998. HLA expression on immature and mature humangerm cells. J. Reprod. Immunol 38:101.
44. Kanai, T., T. Fujii, S. Kozuma, T. Yamashita, A. Miki, A. Kikuchi, andY. Taketani. 2001. Soluble HLA-G influences the release of cytokines from al-logeneic peripheral blood mononuclear cells in culture. Mol. Hum. Reprod.7:195.
45. Aractingi, S., N. Briand, C. Le Danff, M. Viguier, H. Bachelez, L. Michel,L. Dubertret, and E. D. Carosella. 2001. HLA-G and NK receptor are expressedin psoriatic skin. Am. J. Pathol. 159:71.
46. Pangault, C., G. Le Friec, S. Caulet-Maugendre, H. Lena, L. Amiot, V. Guilloux,M. Onno, and R. Fauchet. 2002. Lung macrophages and dendritic cells expressHLA-G molecules in pulmonary diseases. Hum. Immunol. 63:83.
683The Journal of Immunology
by guest on April 3, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from





























