Embed Size (px)
Citation preview

A New Cold-Adapted, Organic Solvent Stable Lipasefrom Mesophilic Staphylococcus epidermidis AT2
Nor Hafizah Ahmad Kamarudin • Raja Noor Zaliha Raja Abd. Rahman •
Mohd Shukuri Mohamad Ali • Thean Chor Leow • Mahiran Basri •
Abu Bakar Salleh
Published online: 29 April 2014
� Springer Science+Business Media New York 2014
Abstract The gene encoding a cold-adapted, organic
solvent stable lipase from a local soil-isolate, mesophilic
Staphylococcus epidermidis AT2 was expressed in a pro-
karyotic system. A two-step purification of AT2 lipase was
achieved using butyl sepharose and DEAE sepharose col-
umn chromatography. The final recovery and purification
fold were 47.09 % and 3.45, respectively. The molecular
mass of the purified lipase was estimated to be 43 kDa.
AT2 lipase was found to be optimally active at pH 8 and
stable at pH 6–9. Interestingly, this enzyme demonstrated
remarkable stability at cold temperature (\30 �C) and
exhibited optimal activity at a temperature of 25 �C. A
significant enhancement of the lipolytic activity was
observed in the presence of Ca2?, Tween 60 and Tween 80.
Phenylmethylsulfonylfluoride, a well known serine inhibi-
tor did not cause complete inhibition of the enzymatic
activity. AT2 lipase exhibited excellent preferences
towards long chain triglycerides and natural oils. The
lipolytic activity was stimulated by dimethylsulfoxide and
diethyl ether, while more than 50 % of its activity was
retained in methanol, ethanol, acetone, toluene, and
n-hexane. Taken together, AT2 lipase revealed highly
attractive biochemical properties especially because of its
stability at low temperature and in organic solvents.
Keywords Staphylococcal lipase � Mesophilic �Purification � Cold-adapted � Organic solvent stable
Abbreviations
A600 Absorbance at 600 nm wavelength
ßME 2-mercaptoethanol
EDTA Ethylenediamineteraacetic acid
DMSO Dimethylsulfoxide
DTT Dithiothreitol
IPTG Isopropyl b-D-1thiogalactopyranoside
OD Optical density
PCR Polymerase chain reaction
PMSF Phenylmethylsulfonylfluoride
PVDF Polyvinylidene fluoride
SDS Sodium dodecyl sulfate polyacrylamide gel
electrophoresis
1 Introduction
Lipase (EC 3.1.1.3) has long been known for its diverse
functions in biocatalysis. The ability of lipase to carry out
hydrolysis and synthesis reactions in aqueous and non
aqueous media, respectively, offers many advantages for
biotechnology exploitation in various industries including
detergents, cosmetics and food industries.
Of many sources of lipases, microbial lipases have
notably attracted the global attention in a very rapid and
exciting rate compared to counterparts from higher
organisms. The enormous labors in exploiting lipases of
N. H. A. Kamarudin � R. N. Z. R. Abd. Rahman (&) �M. S. M. Ali � T. C. Leow � M. Basri � A. B. Salleh
Enzyme and Microbial Technology Research Centre,
Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
e-mail: [email protected]
N. H. A. Kamarudin � R. N. Z. R. Abd. Rahman �M. S. M. Ali � T. C. Leow � A. B. Salleh
Faculty of Biotechnology and Biomolecular Sciences,
Universiti Putra Malaysia, 43400 Serdang, Selangor, Malaysia
M. Basri
Faculty of Sciences, Universiti Putra Malaysia, 43400 Serdang,
Selangor, Malaysia
123
Protein J (2014) 33:296–307
DOI 10.1007/s10930-014-9560-3

microbial origin are mainly due to ease in genetic manip-
ulation and economic feasibility [1]. Staphylococcal lipases
(Family I.6), in particular, are relatively unique through
their distinct molecular organization which comprised of
pre-pro-sequence along with their large size (*70 kDa).
The mature active form (40–46 kDa) is generated by pro-
teolytic processing after secretion into the culture medium
[2]. Their unusual molecular properties have led many to
conduct detailed investigation on the molecular and bio-
chemical characteristics as well as the structural architec-
ture and potential biotechnological applications. While the
information is steadily expanding, knowledge on the
structure and function correlation of staphylococcal lipases
at molecular level, however, is still limited. This lack of
information is in part due to the availability of only a single
crystal structure from staphylococcal lipase family that is
derived from S. hyicus lipase [3].
Apart from their importance in biological function,
staphylococcal lipases have emerged as one of the valuable
industrial enzymes worth investigating. Therefore, the
search of staphylococcal lipase with new and attractive
properties remains a continuous pursuit. Lipase exhibiting
organic solvent stable property has been of prime interest
in many studies; driven by its versatility in catalyzing
synthesis reaction in low water environment. Lipases
especially those that can tolerate methanol and ethanol,
have the potential to improve biodiesel production while
others can be employed in food and pharmaceutical
industries by the synthesis of esters and intermediates [4,
5]. Enzymatic reaction in organic media also provides
attractive advantages in chemical reactions and biotech-
nological applications such as enhanced solubility of
hydrophobic substrates, exclusion of microbial contami-
nation and side reactions caused by water [6].
In addition, cold-adapted lipases have been discovered
as valuable enzymes for novel practical applications.
Enzyme reactions at low temperature offer many advan-
tages including reduced energy consumption and preven-
tion from loss of volatile compounds. A broad avenue in
industrial application can be generated by the exploitation
of cold-adapted lipases as additives in detergents for cold-
water washing, additives in food industries and environ-
mental bioremediations [7].
Besides its industrial importance, modern enzyme
studies have also focused on the understanding of the
structure and function relationship. Identification of the key
features that govern specific characteristics allows modifi-
cation of enzymes to circumvent certain limitations hence
producing enzymes with improved properties. This inves-
tigation will also facilitate determination of the structural
patterns from diverse sequences.
Owing to the overwhelming, non-stop progression in
lipase biotechnology, discovery of novel staphylococcal
lipase genes is of great important for better fundamental
understanding and industrial exploitation. Previously, the
gene encoding a lipase from a newly soil-isolated meso-
philic Staphylococcus epidermidis AT2 (AT2 lipase) has
been sequenced, cloned and expressed in E. coli system [8].
Herein, the purification and biochemical characteristics of
the purified staphylococcal lipase are described. It is worth
emphasized that very few lipases have been reported to
exhibit both cold-adapted and organic solvent stable
properties. In fact, this is by far, the first report on purified
recombinant staphylococcal lipase demonstrating such
unique and potentially marketable properties. Furthermore,
literature on cold-adapted lipases produced by mesophilic
bacterium is relatively few, hence, it is hoped that these
findings will expand the knowledge of lipases particularly
of those produced by Staphylococcus sp.
2 Materials and Methods
2.1 Bacterial Strains, Plasmid and Media
Staphylococcus epidermidis AT2 was isolated from con-
taminated soil at a car service area in Port Dickson,
Malaysia. The lipase gene was identified, sequenced and
cloned in pTrcHis2-TOPO expression vector (Invitrogen,
UK) [8]. E. coli Tuner (DE3) pLacI (Novagen, UK) was
used as expression host. The bacterial culture was culti-
vated in Luria–Bertani medium (10 g tryptone, 5 g yeast
extract, 5 g NaCl in 1 L at pH 7, Difco, USA), supple-
mented with 50 lg/mL of ampicillin (Sigma, USA).
2.2 Expression of AT2 Lipase Gene
The mature active form of AT2 lipase containing 390 amino
acids was cloned in pTrcHis2/AT2 expression vector in
E. coli Tuner (DE3) pLacI (complete open reading frame
(ORF) was submitted to GenBank and assigned with
Accession No. EU814893). E. coli Tuner (DE3) pLacI har-
boring recombinant pTrcHis2/AT2 was grown in 10 mL LB
broth overnight at 37 �C and subsequently sub-culture in
200 mL of LB broth for 3–4 h until OD at A600 = 0.5 for
isopropyl b-D-Thiogalactopyranoside (IPTG) induction.
Cells were induced with 60 lM of IPTG and further incu-
bated for 12–16 h. The induced culture was further incubated
for 12 h and harvested by centrifugation (10,0009g, 10 min,
and 4 �C). The supernatant was discarded, while pellet
(intracellular fraction) was used to obtain the soluble protein.
2.3 Purification of AT2 Lipase
The cell pellet of 600 mL culture was resuspended with
buffer comprising of 20 mM Tris–HCl and 1 M
Stable Lipase from Mesophilic Staphylococcus epidermidis 297
123

ammonium sulfate (pH 8) and lysed by sonication (Branson
Sonifier 250, Branson, USA). The lysed cells were centri-
fuged for 10,0009g, 10 min, 4 �C. The cleared crude cell
lysate was loaded into the first hydrophobic interaction
chromatography column (HIC); butyl sepharose 4 fast flow
(GE Healthcare, UK) which was equilibrated with the same
buffer at a flow rate of 1.0 mL/min. The target proteins
were eluted at 50 % of salt-free elution buffer (20 mM
Tris–HCl pH 8) by a descending linear gradient (20 column
volume, CV). The fractions containing active protein were
pooled and subjected to dialysis/buffer exchange against
20 mM Tris–HCl (pH 9). The dialyzed sample was loaded
into a second ion exchange chromatography column (IEX),
DEAE Sepharose Fast Flow (GE Healthcare, UK) at a flow
rate of 0.5 mL/min. The active, target protein was eluted at
approximately 10 % of elution buffer (20 CV of 0–0.2 M
NaCl linear gradient). For each purification step, fractions
were examined for lipase activity and monitored by SDS-
PAGE analysis.
2.4 Lipase Activity Assay
Lipase activity assay was conducted according to Kwon
and Rhee method (1986) [9] with slight modification. Olive
oil (C18:1) was used as substrate. The substrate was pre-
pared by mixing equal volume of olive oil and buffer at 1:1
ratio. Reaction mixture containing 1 mL of enzyme,
2.5 mL of substrate emulsion, was incubated in a water
bath shaker for 30 min at 25 �C with vigorous agitation,
200 rpm. After 30 min of incubation, the enzyme reaction
was terminated by adding 1 mL of 6 N HCl and the lib-
erated fatty acids were dissolved in 5 mL of isooctane. The
mixture was vigorously vortexed for 30–60 s and two
layers formed. Four ml of the upper layer was drawn out
into test tubes containing 1 mL of 5 % (w/v) copper acetate
pyridine pH 6.1 and mixed vigorously by vortexing. The
absorbance reading was taken at 715 nm and isooctane was
used as the blank. Enzyme activity was determined based
on oleic acid standard curve. One unit of activity is defined
as the rate of free fatty acids released in 1 min.
2.5 Protein Determination (Bradford Assay)
Protein concentration was determined by Bradford method
using bovine serum albumin (BSA) as a standard. The
absorbance reading was taken at 595 nm.
2.6 Electrophoresis (SDS-PAGE)
SDS-PAGE was carried out as described by Laemmli [10]
using 6 % of stacking gel and 12 % of separating gel. The
molecular weight of protein was estimated by comparison
with a broad range of protein molecular weight marker
(116–14.4 kDa; Thermo, UK).
2.7 Activity Staining and N-terminal Sequencing
Lipase activity staining was performed according to the
method outlined by Sommer et al. [11]. Sample was pre-
pared in non-denaturing condition. Following SDS-PAGE,
the unstained gel was immersed in 20 % (v/v) isopropanol
for 30 min on a rotary shaker, washed two to three times
with ultrapure water, and further overlaid on a tributyrin
agar (TB agar) for 30 min. For N-terminal sequencing,
sample was separated by SDS-PAGE and the gel was
electrophoretically transferred to PVDF membrane. The
target band was excised and subjected to Edman Degra-
dation by Applied Biosytems, Procise Sequencer (Life
Technologies, USA).
2.8 Effect of Temperature on Lipase Activity
and Stability
Optimal temperature for AT2 lipase was determined by
varying assay temperatures ranging from 15 to 50 �C at
5 �C intervals. Lipolytic activities at different temperature
were assayed for 30 min. For thermal stability/half-life
study, purified enzyme was pre-incubated at three different
temperatures (20, 25 and 30 �C) for 300 min in which, at
every 30 min intervals, sample was removed and the
lipolytic activity was determined at 25 �C. Sample at 0 min
(t = 0) was regarded as the control of the experiment
(100 %).
2.9 Effect of pH on Lipase Activity and Stability
The optimal pH of purified AT2 lipase was determined by
preparing substrate emulsion (olive oil) in 50 mM buffer of
different pH ranging from pH 4 to 12 at 1:1 ratio (olive oil
and buffer), and assayed at 25 �C for 30 min. Lipolytic
activity in 50 mM Tris–HCl (pH 8) was further selected as
the control reaction for stability study. To determine the pH
stability profile of AT2 lipase, purified enzyme was pre-
incubated in buffer of tested pHs for 30 min followed by
lipase activity assay at 25 �C, 30 min. Buffer systems used
were sodium acetate (pH 4–6), phosphate (pH 6–8), Tris–
HCl (pH 8–9), Glycine/NaOH (pH 9–11) and sodium
hydrogen phosphate, Na2HPO4/NaOH (pH 11–12).
2.10 Effects of Metal Ions on Lipase Activity
The effects of additional metal ions were investigated by
pre-incubating the purified enzyme with metal ions at two
concentrations (1 mM and 5 mM) for 30 min at 20 �C and
subsequently subjected to lipase assay. The metal ions used
298 N. H. A. Kamarudin et al.
123

were: K?, Na?, Mg2?, Ca2?, Mn2?, Cu2?, Co2?, Zn2? and
Fe3?. Control reaction corresponded to the non-treated
enzyme (without incubation with metal ions) and regarded
as 100 %.
2.11 Effect of Surfactants on Lipase Activity
To assimilate the effect of surfactants on AT2 lipase, the
enzyme was treated with 0.1 % and 0.5 % (v/v) of Tween
20, 40, 60, 80, Triton X-100, and SDS for 30 min at 20 �C
prior to lipase assay. Surfactant-free enzyme served as the
control reaction (100 %) and the remaining activities of the
treated enzymes were determined in comparison to the
control of the experiment.
2.12 Effect of Inhibitors on Lipase Activity
The influence of inhibitors, chelating agents and reducing
agents (phenylmethylsulfoxide (PMSF), pefabloc, pepsta-
tin A, ethylenediaminetetraacetic acid (EDTA), dithio-
threitol (DTT) and b-mercaptoethanol (bME)) on the
purified enzyme were investigated at two different con-
centrations (1 and 5 mM). AT2 lipase was pre-treated with
the respective inhibitors for 30 min prior to lipase assay.
Relative activities were measured against the lipolytic
activity of the non-treated sample which considered as
100 %.
2.13 Effect of Substrates on Lipase Activity
Triglycerides and natural oils were exploited to study the
effect of different carbon chain length substrates on AT2
lipase activity. Substrates were prepared by mixing buffer
(50 mM Tris–HCl pH 8) and triglycerides/oils at 1:1 ratio.
Reaction mixtures containing different type of substrates
were assayed at 25 �C for 30 min. Substrates used were
triacetin (C2:0), tributyrin (C4:0), triolein (C18:1), coconut
oil (C12:0), palm oil (C16:0), palm kernel oil (C16:0),
olive oil (C18:1), rice bran oil (C18:1), soy oil (C18:2),
corn oil (C18:2), sunflower oil (C18:2). Lipolytic activities
of AT2 lipase at different substrates were calculated in
relative to that obtained in olive oil which served as control
(100 %).
2.14 Effect of Organic Solvents on Lipase Activity
Fifteen organic solvents with different log P were tested on
AT2 lipase stability; DMSO (-1.3), methanol (-0.76),
acetonitrile (-0.33), ethanol (-0.24), acetone (-0.24),
1-propanol (0.28), ethyl acetate (0.68), diethyl ether (0.85),
chloroform (2.0), benzene (2.0), toluene (2.5), octanol
(2.9), xylene (3.1), n-hexane (3.5) and n-heptane (4.0).
Purified enzyme was pre-incubated with 25 % (v/v) of
organic solvent for 30 min at 20 �C with vigorous shaking
(150 rpm) in a water bath shaker. The treated enzymes, as
well as the organic solvent-free enzyme (which served as
the control of the experiment), were assayed for their
lipolytic activities. Residual activities were measured in
relative to the control reaction which was regarded as
100 %.
3 Results
3.1 Expression of AT2 Lipase
A lipase gene from Staphylococcus epidermidis AT2
(termed AT2 lipase) was previously sequenced and
expressed in E. coli system [8]. The deduced gene which
contained a single ORF comprising of 643 amino acids was
deposited in GenBank with accession number EU814893.
The nucleotide sequence of AT2 lipase is organized as a
pre-pro sequence, and this molecular organization is sim-
ilarly found in other deduced staphylococcal lipase genes.
The mature active form of AT2 lipase which translated to
390 amino acids was cloned and expressed in pTrcHis2-
TOPO vector in E. coli TOP10. The expression system is
designed for direct insertion of PCR product into the
plasmid vector via 30 thymidine (T) overhangs and requires
no ligase and post PCR procedures. The resulting lipolytic
activity obtained at its optimum condition (16 h, at 0.6 mM
of IPTG) was 5.58 U/mL which corresponded to 18-fold
increase in the activity compared to its wild type [8]. In the
present study, the expression of soluble AT2 lipase was
improved when E. coli Tuner (DE3) pLacI was used as
host. The lipolytic activity increased by 4.5-fold under its
optimized condition; 60 lM of IPTG, 12 h of post induc-
tion time yielding an optimal activity of 25.20 U/mL (data
not shown). Owing to its higher activity, E. coli Tuner
(DE3) pLacI harboring recombinant pTrcHis2/AT2 was
further used for the purpose of purification and character-
ization studies.
3.2 Purification of AT2 Lipase
Isolating a desired protein from a complex mixture requires
the employment of an ideal purification step. In the attempt
to purify AT2 lipase, the crude lysate was loaded into
hydrophobic interaction chromatography (HIC) butyl
sepharose fast flow column followed by ion exchange
chromatography (IEX) on DEAE Sepharose Fast Flow
exchanger. The efficiency of each purification protocol was
evaluated by measuring the total protein and specific
activity as presented in Table 1. HIC displayed a good
capturing strategy for the purification of AT2 lipase as
denoted by 92.47 % of recovery and almost a onefold
Stable Lipase from Mesophilic Staphylococcus epidermidis 299
123

increment of the purification fold. A bulk of impurities in
the cell-free supernatant was removed during sample
binding, leaving few proteins to tightly bind to the matrix.
Two separated broad peaks were detected during the elu-
tion fractionation. The second peak which eluted at
approximately 0.5 M of ammonium sulfate was identified
as the target protein through lipase activity assay and SDS-
PAGE analysis. Following HIC, the accomplishment of
IEX as the final polishing step was evidenced by marked
improvement in purity with 3.45 purification fold and a
final recovery of 47.09 % over the crude enzyme. A single
sharp peak was obtained at approximately 0.05 M NaCl of
the linear gradient and the fractions were further assayed
for confirmation.
To assimilate the purity level of AT2 lipase at every
stage of purification, SDS-PAGE, followed by activity
staining were conducted. Figure 1a denotes the SDS-PAGE
analysis for each steps of purification. The impurities in the
crude enzyme were removed by almost 70 % through HIC
as clearly observed in Lane 3. Meanwhile, a pure, single
band was detected on the gel after the final step of purifi-
cation (Lane 5). The molecular weight (MW) of the puri-
fied product was estimated to be 43 kDa, which agreed
with the calculated MW of AT2 lipase based on its primary
sequence analysis.
Besides the quantitative lipase measurement, activity
staining was performed to qualitatively ensure that the target
protein is biologically active. A clearing zone (halo) sur-
rounding the target protein band was observed when the gel
was placed on tributyrin agar for 30 min at 25 �C. Tributyrin
agar is one of the selective media for lipase detection where
tributyrin (C4:0) serves as the substrate. The active protein
hydrolyzed the substrates surrounding it resulting in the
formation of clearing zone. This signified and confirmed that
the purified product was a lipase (Fig. 1b).
The purity level of AT2 lipase was further determined
by Native PAGE analysis. Somewhat unexpected, no dis-
tinct band was observed on the gel, instead, a low mobility
band and smearing were detected (data not shown). Such
behavior is deduced to be related to the tendency of protein
to form aggregates or oligomer [12, 13]. The oligomeric
state of the enzyme was then determined by analytical ultra
centrifugation (AUC) based on sedimentation velocity
(SV) analysis. This method is used to probe the aggregation
behavior of proteins in solution by physical separation of
molecular species according to variation in mass and shape
Fig. 1 Analysis of the purified
product. a SDS-PAGE analysis
of AT2 lipase at different
purification steps. Lane 1
Standard protein marker, Lane 2
crude enzyme, Lane 3 AT2
lipase after hydrophobic
interaction chromatography
(HIC), Lane 4 pooled fractions
after dialysis, Lane 5 purified
AT2 lipase after ion exchange
chromatography (IEX).
b Activity staining of AT2
lipase on Tributyrin agar. Lane
1 purified AT2 lipase (clearing
zone was observed after 30 min
of incubation at 25 �C); Lane 2
Standard protein marker
Table 1 Flow sheet of AT2
lipase purification
a One Unit of activity
corresponds to the rate of free
fatty acids released in 1 min
using olive oil emulsion as
substrate
Step Volume
(mL)
Activitya
(U/mL)
Protein
(mg/mL)
Total
Activity
(U)
Specific
Activity
(U/mg)
Yield
(%)
Fold
Crude 30 36.81 0.88 1,104.30 41.83 100 1
Hydrophobic interaction
chromatography (butyl
sepharose fast flow)
28 36.47 0.45 1,021.16 81.04 92.47 1.94
Ion exchange chromatography
(DEAE sepharose fast flow)
12 43.34 0.30 520.08 144.47 47.09 3.45
300 N. H. A. Kamarudin et al.
123
[14]. Through this analysis, a dominant single peak with a
molecular mass of approximately 42.1 kDa was obtained,
suggesting the occurrence of a single protein species in the
solution thereby, reflecting the existence AT2 lipase in a
monomeric form. No corresponding peaks for any oligo-
mers were detected (data not shown).
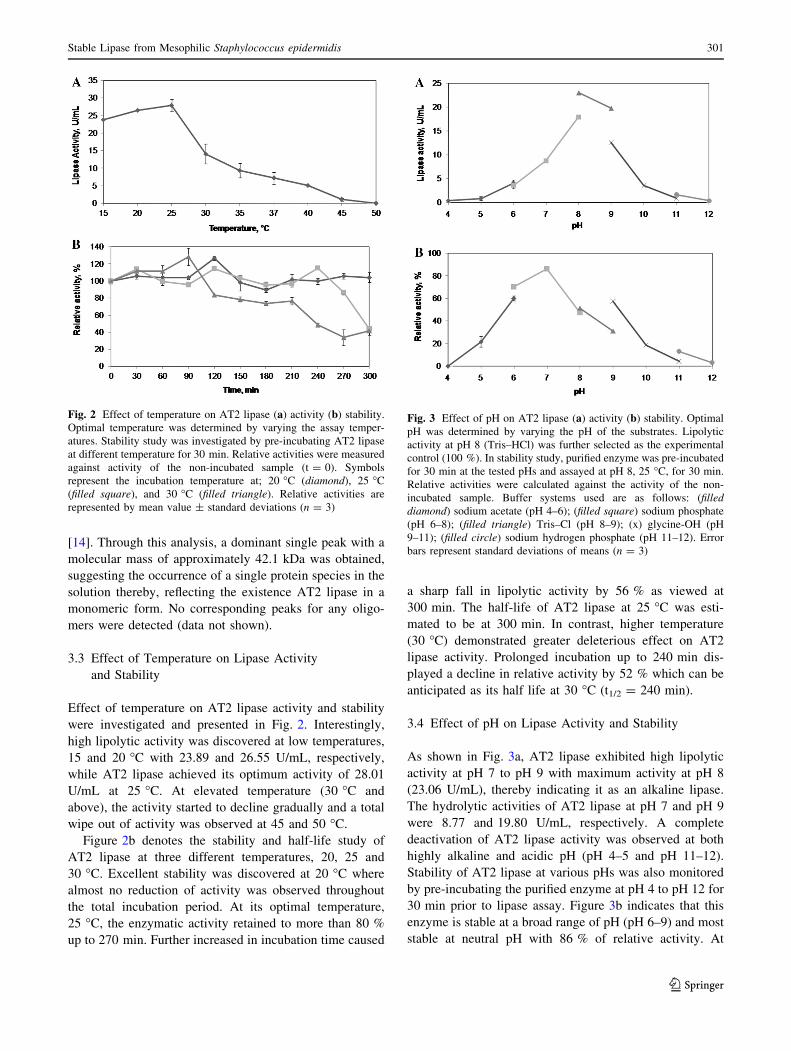
3.3 Effect of Temperature on Lipase Activity
and Stability
Effect of temperature on AT2 lipase activity and stability
were investigated and presented in Fig. 2. Interestingly,
high lipolytic activity was discovered at low temperatures,
15 and 20 �C with 23.89 and 26.55 U/mL, respectively,
while AT2 lipase achieved its optimum activity of 28.01
U/mL at 25 �C. At elevated temperature (30 �C and
above), the activity started to decline gradually and a total
wipe out of activity was observed at 45 and 50 �C.
Figure 2b denotes the stability and half-life study of
AT2 lipase at three different temperatures, 20, 25 and
30 �C. Excellent stability was discovered at 20 �C where
almost no reduction of activity was observed throughout
the total incubation period. At its optimal temperature,
25 �C, the enzymatic activity retained to more than 80 %
up to 270 min. Further increased in incubation time caused
a sharp fall in lipolytic activity by 56 % as viewed at
300 min. The half-life of AT2 lipase at 25 �C was esti-
mated to be at 300 min. In contrast, higher temperature
(30 �C) demonstrated greater deleterious effect on AT2
lipase activity. Prolonged incubation up to 240 min dis-
played a decline in relative activity by 52 % which can be
anticipated as its half life at 30 �C (t1/2 = 240 min).
3.4 Effect of pH on Lipase Activity and Stability
As shown in Fig. 3a, AT2 lipase exhibited high lipolytic
activity at pH 7 to pH 9 with maximum activity at pH 8
(23.06 U/mL), thereby indicating it as an alkaline lipase.
The hydrolytic activities of AT2 lipase at pH 7 and pH 9
were 8.77 and 19.80 U/mL, respectively. A complete
deactivation of AT2 lipase activity was observed at both
highly alkaline and acidic pH (pH 4–5 and pH 11–12).
Stability of AT2 lipase at various pHs was also monitored
by pre-incubating the purified enzyme at pH 4 to pH 12 for
30 min prior to lipase assay. Figure 3b indicates that this
enzyme is stable at a broad range of pH (pH 6–9) and most
stable at neutral pH with 86 % of relative activity. At
Fig. 2 Effect of temperature on AT2 lipase (a) activity (b) stability.
Optimal temperature was determined by varying the assay temper-
atures. Stability study was investigated by pre-incubating AT2 lipase
at different temperature for 30 min. Relative activities were measured
against activity of the non-incubated sample (t = 0). Symbols
represent the incubation temperature at; 20 �C (diamond), 25 �C
(filled square), and 30 �C (filled triangle). Relative activities are
represented by mean value ± standard deviations (n = 3)
Fig. 3 Effect of pH on AT2 lipase (a) activity (b) stability. Optimal
pH was determined by varying the pH of the substrates. Lipolytic
activity at pH 8 (Tris–HCl) was further selected as the experimental
control (100 %). In stability study, purified enzyme was pre-incubated
for 30 min at the tested pHs and assayed at pH 8, 25 �C, for 30 min.
Relative activities were calculated against the activity of the non-
incubated sample. Buffer systems used are as follows: (filled
diamond) sodium acetate (pH 4–6); (filled square) sodium phosphate
(pH 6–8); (filled triangle) Tris–Cl (pH 8–9); (x) glycine-OH (pH
9–11); (filled circle) sodium hydrogen phosphate (pH 11–12). Error
bars represent standard deviations of means (n = 3)
Stable Lipase from Mesophilic Staphylococcus epidermidis 301
123
slightly acidic environment (pH 6), 70 % of the residual
activity was retained while at pH 8 and 9, the activity
remained up to 52 and 58 %, respectively. AT2 lipase was
very unstable at extremely acidic and alkaline pH (pH 4–5
and pH 10–12).
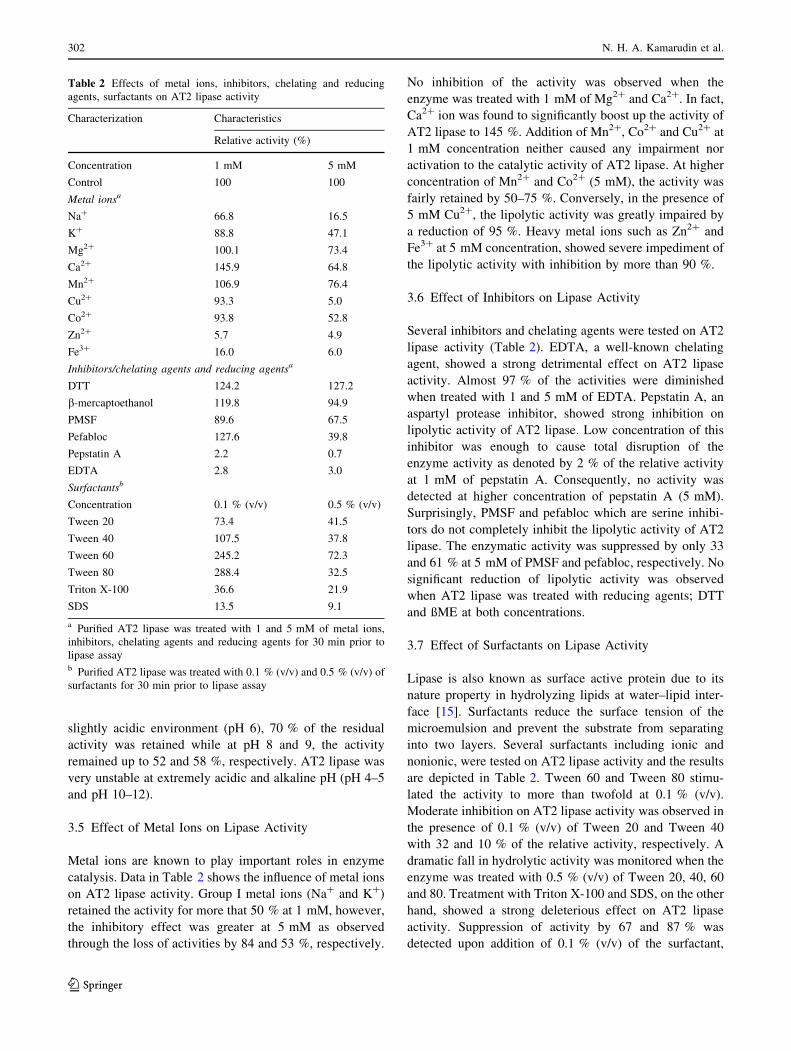
3.5 Effect of Metal Ions on Lipase Activity
Metal ions are known to play important roles in enzyme
catalysis. Data in Table 2 shows the influence of metal ions
on AT2 lipase activity. Group I metal ions (Na? and K?)
retained the activity for more that 50 % at 1 mM, however,
the inhibitory effect was greater at 5 mM as observed
through the loss of activities by 84 and 53 %, respectively.
No inhibition of the activity was observed when the
enzyme was treated with 1 mM of Mg2? and Ca2?. In fact,
Ca2? ion was found to significantly boost up the activity of
AT2 lipase to 145 %. Addition of Mn2?, Co2? and Cu2? at
1 mM concentration neither caused any impairment nor
activation to the catalytic activity of AT2 lipase. At higher
concentration of Mn2? and Co2? (5 mM), the activity was
fairly retained by 50–75 %. Conversely, in the presence of
5 mM Cu2?, the lipolytic activity was greatly impaired by
a reduction of 95 %. Heavy metal ions such as Zn2? and
Fe3? at 5 mM concentration, showed severe impediment of
the lipolytic activity with inhibition by more than 90 %.
3.6 Effect of Inhibitors on Lipase Activity
Several inhibitors and chelating agents were tested on AT2
lipase activity (Table 2). EDTA, a well-known chelating
agent, showed a strong detrimental effect on AT2 lipase
activity. Almost 97 % of the activities were diminished
when treated with 1 and 5 mM of EDTA. Pepstatin A, an
aspartyl protease inhibitor, showed strong inhibition on
lipolytic activity of AT2 lipase. Low concentration of this
inhibitor was enough to cause total disruption of the
enzyme activity as denoted by 2 % of the relative activity
at 1 mM of pepstatin A. Consequently, no activity was
detected at higher concentration of pepstatin A (5 mM).
Surprisingly, PMSF and pefabloc which are serine inhibi-
tors do not completely inhibit the lipolytic activity of AT2
lipase. The enzymatic activity was suppressed by only 33
and 61 % at 5 mM of PMSF and pefabloc, respectively. No
significant reduction of lipolytic activity was observed
when AT2 lipase was treated with reducing agents; DTT
and ßME at both concentrations.
3.7 Effect of Surfactants on Lipase Activity
Lipase is also known as surface active protein due to its
nature property in hydrolyzing lipids at water–lipid inter-
face [15]. Surfactants reduce the surface tension of the
microemulsion and prevent the substrate from separating
into two layers. Several surfactants including ionic and
nonionic, were tested on AT2 lipase activity and the results
are depicted in Table 2. Tween 60 and Tween 80 stimu-
lated the activity to more than twofold at 0.1 % (v/v).
Moderate inhibition on AT2 lipase activity was observed in
the presence of 0.1 % (v/v) of Tween 20 and Tween 40
with 32 and 10 % of the relative activity, respectively. A
dramatic fall in hydrolytic activity was monitored when the
enzyme was treated with 0.5 % (v/v) of Tween 20, 40, 60
and 80. Treatment with Triton X-100 and SDS, on the other
hand, showed a strong deleterious effect on AT2 lipase
activity. Suppression of activity by 67 and 87 % was
detected upon addition of 0.1 % (v/v) of the surfactant,
Table 2 Effects of metal ions, inhibitors, chelating and reducing
agents, surfactants on AT2 lipase activity
Characterization Characteristics
Relative activity (%)
Concentration 1 mM 5 mM
Control 100 100
Metal ionsa
Na? 66.8 16.5
K? 88.8 47.1
Mg2? 100.1 73.4
Ca2? 145.9 64.8
Mn2? 106.9 76.4
Cu2? 93.3 5.0
Co2? 93.8 52.8
Zn2? 5.7 4.9
Fe3? 16.0 6.0
Inhibitors/chelating agents and reducing agentsa
DTT 124.2 127.2
b-mercaptoethanol 119.8 94.9
PMSF 89.6 67.5
Pefabloc 127.6 39.8
Pepstatin A 2.2 0.7
EDTA 2.8 3.0
Surfactantsb
Concentration 0.1 % (v/v) 0.5 % (v/v)
Tween 20 73.4 41.5
Tween 40 107.5 37.8
Tween 60 245.2 72.3
Tween 80 288.4 32.5
Triton X-100 36.6 21.9
SDS 13.5 9.1
a Purified AT2 lipase was treated with 1 and 5 mM of metal ions,
inhibitors, chelating agents and reducing agents for 30 min prior to
lipase assayb Purified AT2 lipase was treated with 0.1 % (v/v) and 0.5 % (v/v) of
surfactants for 30 min prior to lipase assay
302 N. H. A. Kamarudin et al.
123
respectively. Meanwhile, at higher concentration of the
surfactants, greater inhibition on AT2 lipase activity was
observed.
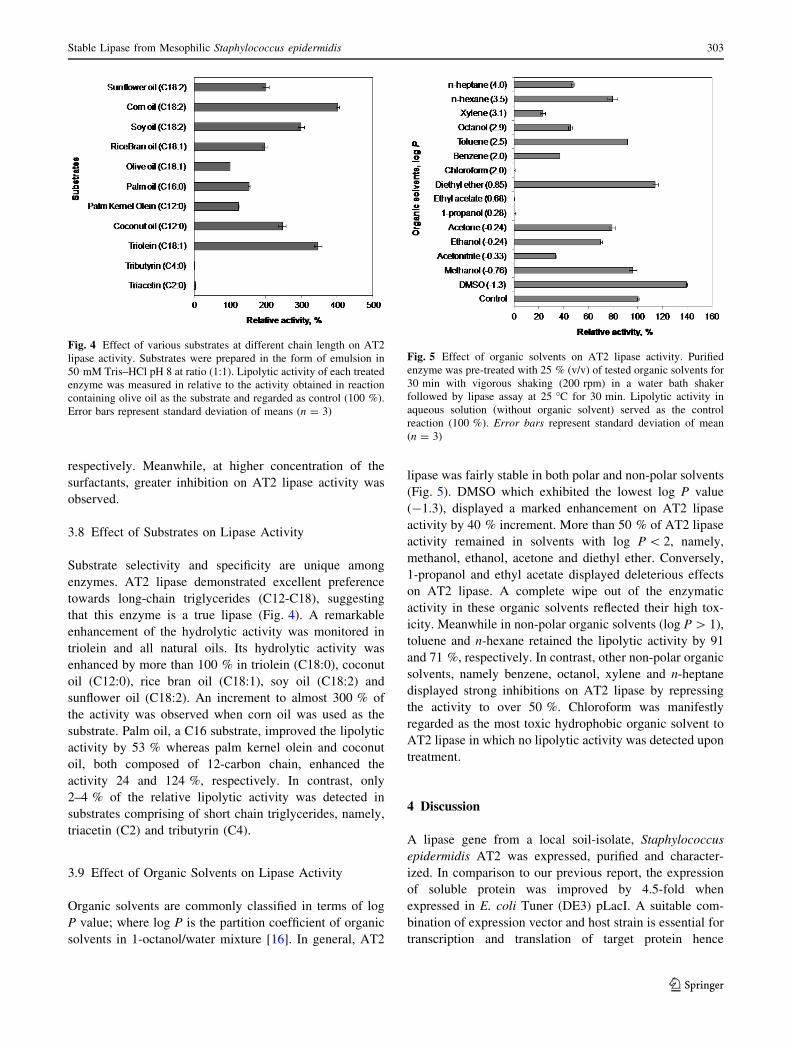
3.8 Effect of Substrates on Lipase Activity
Substrate selectivity and specificity are unique among
enzymes. AT2 lipase demonstrated excellent preference
towards long-chain triglycerides (C12-C18), suggesting
that this enzyme is a true lipase (Fig. 4). A remarkable
enhancement of the hydrolytic activity was monitored in
triolein and all natural oils. Its hydrolytic activity was
enhanced by more than 100 % in triolein (C18:0), coconut
oil (C12:0), rice bran oil (C18:1), soy oil (C18:2) and
sunflower oil (C18:2). An increment to almost 300 % of
the activity was observed when corn oil was used as the
substrate. Palm oil, a C16 substrate, improved the lipolytic
activity by 53 % whereas palm kernel olein and coconut
oil, both composed of 12-carbon chain, enhanced the
activity 24 and 124 %, respectively. In contrast, only
2–4 % of the relative lipolytic activity was detected in
substrates comprising of short chain triglycerides, namely,
triacetin (C2) and tributyrin (C4).
3.9 Effect of Organic Solvents on Lipase Activity
Organic solvents are commonly classified in terms of log
P value; where log P is the partition coefficient of organic
solvents in 1-octanol/water mixture [16]. In general, AT2
lipase was fairly stable in both polar and non-polar solvents
(Fig. 5). DMSO which exhibited the lowest log P value
(-1.3), displayed a marked enhancement on AT2 lipase
activity by 40 % increment. More than 50 % of AT2 lipase
activity remained in solvents with log P \ 2, namely,
methanol, ethanol, acetone and diethyl ether. Conversely,
1-propanol and ethyl acetate displayed deleterious effects
on AT2 lipase. A complete wipe out of the enzymatic
activity in these organic solvents reflected their high tox-
icity. Meanwhile in non-polar organic solvents (log P [ 1),
toluene and n-hexane retained the lipolytic activity by 91
and 71 %, respectively. In contrast, other non-polar organic
solvents, namely benzene, octanol, xylene and n-heptane
displayed strong inhibitions on AT2 lipase by repressing
the activity to over 50 %. Chloroform was manifestly
regarded as the most toxic hydrophobic organic solvent to
AT2 lipase in which no lipolytic activity was detected upon
treatment.
4 Discussion
A lipase gene from a local soil-isolate, Staphylococcus
epidermidis AT2 was expressed, purified and character-
ized. In comparison to our previous report, the expression
of soluble protein was improved by 4.5-fold when
expressed in E. coli Tuner (DE3) pLacI. A suitable com-
bination of expression vector and host strain is essential for
transcription and translation of target protein hence
Fig. 4 Effect of various substrates at different chain length on AT2
lipase activity. Substrates were prepared in the form of emulsion in
50 mM Tris–HCl pH 8 at ratio (1:1). Lipolytic activity of each treated
enzyme was measured in relative to the activity obtained in reaction
containing olive oil as the substrate and regarded as control (100 %).
Error bars represent standard deviation of means (n = 3)
Fig. 5 Effect of organic solvents on AT2 lipase activity. Purified
enzyme was pre-treated with 25 % (v/v) of tested organic solvents for
30 min with vigorous shaking (200 rpm) in a water bath shaker
followed by lipase assay at 25 �C for 30 min. Lipolytic activity in
aqueous solution (without organic solvent) served as the control
reaction (100 %). Error bars represent standard deviation of mean
(n = 3)
Stable Lipase from Mesophilic Staphylococcus epidermidis 303
123

producing soluble protein at high level. AT2 lipase was
further purified by two-step column chromatography; HIC
butyl sepharose and IEX DEAE sepharose. In many cases
of lipase purification, HIC was successfully employed as a
sole or part of the purification strategies. The strong
binding affinity of AT2 lipase towards the matrix may in
part due to the occurrence of approximately 21.3 % of total
hydrophobic residues (leucine, isoleucine, valine, phenyl-
alanine) in the protein; which interacted with the hydro-
phobic 4-linear alkyl chain ligand (butyl sepharose) besides
being aided with high ionic strength in the mobile phase.
IEX was employed based on its powerful capability in
separating biomolecules with minor difference in charge
properties. A pure, single band of AT2 lipase was observed
through SDS-PAGE analysis after DEAE Sepharose chro-
matography. Evidently, IEX has been widely used in many
purification strategies. S. haemolyticus lipase was purified
using a series of IEX chromatography techniques. DEAE
Sepharose CL-6B, CM Sepharose CL-6B, and Resource Q
column were applied respectively in the attempt to obtain
highly purified lipase [18]. Sakinc et al. (2007) [19]
reported on the use of DEAE Sepharose CL-6B as a final
polishing strategy in purification of S. saprophyticus lipase.
AT2 lipase was found prone to aggregation as observed
in Native PAGE analysis. Smears on Native PAGE are
often associated with incomplete solubility of protein while
low mobility band corresponds to oligomer [12, 13]. Fur-
ther investigation on the oligomeric state of AT2 lipase by
AUC showed that this enzyme existed in monomeric form
(data not shown). Staphylococcal lipases have been
claimed to exhibit a complex structure and its tendency to
form aggregates which is driven by the hydrophobic force
from high content of hydrophobic residues [20].
Temperature is one of the most crucial factors in
mediating enzyme-catalyzed reaction rates. It is therefore a
primary prerequisite to determine the optimal temperature
of an enzyme for its best function. Interestingly, in this
study, S. epidermidis AT2 - a mesophile, produced a cold-
adapted lipase that exhibited optimal activity at a temper-
ature of 25 �C. In fact, the enzyme was catalytically active
and stable at temperature below 30 �C, while little or no
activity was detected at higher temperature. It is therefore
clearly evidenced the heat-sensitivity of AT2 lipase, a
distinguishing feature of a cold-loving enzyme. Cold-active
enzymes are commonly produced by psychrophiles as part
of their adaptation and survival strategies in cold climate.
There are very few reports on mesophilic microorganisms
producing cold-adapted enzymes. Of the limited examples,
Cai et al. [21] reported on the production of two cold-
adapted lipases (Lipase-A and Lipase-B) from Geotrichum
sp. SYBC WU-3, a mesophile. Recently, a cold-adapted
lipase from Streptomyces strain which exhibited stability at
temperature below 25 �C was reported [22]. It has been
generally accepted that the stability of enzyme at cold
temperature is governed by the nature of protein structures
and conformations. Conformational flexibility remains the
dominating theory in cold adaptation however the issue on
either local or global flexibility is still under argument [23].
Enzymes adopt specific strategies to become catalytically
active in the extreme environment such as through reduced
number of disulphide bridge and large proportion of sur-
face-exposed charged residues [24], decreased in proline
content [25], and increased number of glycine, methionine
and loops [26].
AT2 lipase was identified as an alkaline lipase where it
exhibited its optimal activity at pH 8. Alkaline lipases are
important in detergent industries. Most staphylococcal
lipases share similar preference towards alkaline pH (pH
8.5) including S. haemolyticus lipase and S. simulans lipase
[18, 27]. In contrast, S. aureus NCTC8530 lipase favored
slightly acidic pH (pH 6.5) [28]. In terms of stability, this
enzyme was found stable at a broad range of pH (pH 6–9).
Similarly, lipase from S. saprophyticus, showed stability at
pH 6–9; below or above this range, the activity was greatly
inactivated [19]. pH is important in maintaining a proper
protonation form of active site of an enzyme as the site
composed of charge residues which involve in substrate
binding. Changes in pH will also lead to disruption of ionic
bonds that hold the enzyme’s tertiary structure.
Nature chooses metals for their biological function. For
instance, Mg2? and Ca2?, both divalent charged metal
ions, are involved in substrate activation and electrostatic
stabilization of enzyme [29].Ca2? was found to strongly
stimulate the activity of AT2 lipase. Enhancement of the
lipolytic activity by Ca2? has been observed in many
staphylococcal lipases as well as other bacterial lipases. In
S. hyicus lipase crystal structure, the role of Ca2? has been
proposed to be involved in stabilizing the three-dimen-
sional structure rather than directly involved in enzyme
catalysis [3]. Primary sequence comparison between AT2
lipase and S. hyicus lipase revealed that the calcium
binding site was highly conserved thereby similar reason
might explain the functional role of Ca2? in AT2 lipase.
Meanwhile, the toxic effect of Zn2? and Fe3? on AT2
lipase might be generated by excess amount of these heavy
metals.
A complete inhibition of AT2 lipase activity was
observed when treated with EDTA. This total deleterious
consequence reflected the essential role of metal ions in
enzyme structure and catalysis. Hence, AT2 lipase can be
classified as a metal-dependent enzyme. Such characteristic
is highly in line to the activation of AT2 lipase in the
presence of metal ion, particularly, Ca2? as described
earlier. Many lipases are inhibited by this heavy metal ion
scavenger for their similar dependency towards Ca2? such
as that in S. saprophyticus and S. warneri lipase 2 [19, 30].
304 N. H. A. Kamarudin et al.
123

Besides EDTA, pepstatin A also demonstrated severe
impairment on AT2 lipase. The probable reason for the
pronounced inhibitory effect of pepstatin A may be
attributed by its potent action on aspartate residues par-
ticularly the catalytic aspartate and the aspartate residues
involve in Ca2? binding site. Insensitivity of AT2 lipase
towards PMSF was found to be another interesting finding.
Lipase which active site composed of catalytic serine is
theoretically sensitive towards serine inhibitors. These
inhibitors bind covalently at the active serine and cause
inactivation. However, notably, with very few exceptions,
some lipases demonstrated resistancy towards inactivation
by PMSF such as that observed in S. hyicus lipase [31]. In
the case of palmitoyl protein thioesterase (PPTI), the
occupancy of PMSF in the catalytic region was restricted
by the narrow substrate-binding cleft [32]. Presumably, low
sensitivity AT2 lipase towards PMSF and pefabloc may be
explained by the limited accessibility of aromatic sulfonyl
fluoride to the active site. Lacking of cysteine residues
which possibly involved in disulphide bond formation, this
enzyme was not inhibited by reducing agents, DTT and
ßME.
The hydrolytic activity of AT2 lipase was remarkably
enhanced in the presence of 1 % (v/v) of Tween 60 and
Tween 80. Both surfactants were likely to stabilize the
substrate emulsion and trigger the lid displacement of AT2
lipase through hydrophobic interaction between the sur-
factant and hydrophobic residues, giving access for more
substrates to interact at the active site. The latter was
deduced as surfactants are known to resemble lipase sub-
strate [33]. However, at higher concentration of the sur-
factants (0.5 % (v/v), a dramatic fall in the lipolytic
activity of AT2 lipase was detected. Such decline in
activity may be caused by treatment of surfactants near or
above the critical micellar concentration (CMC). At this
point, surfactants were in saturation and restricting the
absorption of lipase at the oil/water interface resulting in
enzyme deactivation [34]. Ionic surfactant, SDS, showed a
total inhibition on AT2 lipase activity. Deleterious effect
by SDS may be attributed by the formation of additional
hydrophobic interactions which resulted in complete
unfolding of the tertiary structure and altered surface
hydrophobicity [34].
AT2 lipase showed strong preferential specificity for
higher chain length triglycerides and natural oils. Long
chain substrates which are less water soluble seem to be
essential in stimulating the lipase-substrate interaction
through the formation of oil/water interface between the
substrates and enzyme. A greater accessibility for the
substrate to enter the active site was triggered by strong
hydrophobic interaction between the lid and substrate.
Chain length selectivity of staphylococcal lipases differs
from one another. S. aureus lipase was reported to favor
short chain triacylglycerols but poorly hydrolyzed medium
and long chain triglycerides as well as phospholipids [28].
S. warneri lipase 2 was capable of hydrolyzing short chain
substrates and phospholipids [30].
Stability of enzyme in organic solvents is regarded as an
extra-valuable property for various biotechnology applica-
tions. AT2 lipase was found stable in both hydrophilic and
hydrophobic organic solvents. The enzyme displayed sta-
bility not only in methanol, ethanol and acetone but the
lipolytic activity was also enhanced in the presence of
DMSO and diethyl ether. Stimulation of activity in DMSO
has been observed to associate with glycine-rich enzyme
[35]. Classically, solvents with log P \ 2 are not favorable
for biocatalysis due to their inactivation effects towards
biocatalysts [16]. Therefore, the capability of AT2 lipase to
tolerate solvents such as methanol and ethanol is highly
important for industrial applications especially in biodiesel
production. Exception was observed on 1-propanol and ethyl
acetate whereby these solvents showed complete inhibition
to the enzymatic activity. The notion on stripping water
molecules off the protein surface by hydrophilic organic
solvents may explain the inactivation effect by these two
solvents. Optimal amount of water molecules is of necessity
to lubricate the enzyme for increased activation [36].
Purified AT2 lipase displayed variable results when
treated with water-immiscible organic solvents. The enzyme
was catalytically active in toluene and n-hexane mixture.
Hydrophobic organic solvents have been regarded as more
superior and favorable in the enzyme-solvent interaction
mainly by maintaining the solvation shell of the protein [37].
Furthermore, in most of the transesterification reactions for
biosynthesis of esters, hexane is the most favoured solvent.
Hence AT2 lipase is also a good candidate for the biocatal-
ysis of esters in food and flavouring industries, cosmetics,
and pharmaceuticals manufacturing. In contrast, benzene,
octanol, xylene, n-heptane and chloroform inhibited the
enzymatic activity of AT2 lipase. A plausible explanation for
the instability of AT2 lipase in these solvents is in their
involvement in penetrating and attacking the active centre of
lipase which is hydrophobic in nature.
Few staphylococcal lipases (wild-type) have been
claimed as organic solvent stable enzymes. For instance,
extracellular lipase from S. caseolyticus EX-17 displayed
enhancement in activity by 1.5-fold in the presence of n-
hexane [38]. Stability of S. saprophyticus M36 lipase in
organic solvents was achieved in 25 % (v/v) of toluene,
benzene, p-xylene and n-hexane in which residual activity
remained in the range of 80-90 % [39]. Crude lipase from
S. epidermidis CMST-Pi 1 lipase conserved its activity in
hexadecane, tetradecane and dodecane [40]. Clearly, AT2
lipase is unique by having stability in both polar and non
polar organic solvents compared to other staphylococcal
lipases.
Stable Lipase from Mesophilic Staphylococcus epidermidis 305
123

Stability of lipase in organic solvents is deduced to
correlate with the distribution of surface charged resi-
dues and surface property of the enzyme [35]. In
agreement, Ogino et al. [41] reported that the surface
residues of the molecule were important for the solvent
stable nature of PST-01 protease. There is however
relatively few published studies on the mechanism of
enzyme adaptation towards organic solvents thereby
limiting our understanding on this subject matter. No
clear correlation and structural pattern have yet been
observed as different enzymes adopt different adapta-
tion strategies. A detailed investigation on the 3D
structure will permit better understanding on the struc-
ture and function relationship.
5 Conclusion
AT2 lipase displays new and exciting biochemical prop-
erties exclusively as a cold-adapted, organic solvent stable
enzyme produced by a mesophilic bacterium. To the best
of our knowledge, no staphylococcal lipase exhibiting both
cold-adapted and organic solvent stability properties has
been reported so far. This finding may serve as useful
information for further exploration on the structural basis
and commercial application in modern biotechnology such
as in cold water washing, food industries and production of
intermediates.
Acknowledgments We thank Prof Dr. Andrew H.-J Wang and Dr.
Lee Cheng-Chung from Institute of Biological Chemistry, Academia
Sinica, Taiwan for AUC analysis. This project was supported by
Ministry of Science and Technology, Malaysia [02-01-04 SF 1024].
Conflict of interest The authors declare that there is no conflict of
interest.
References
1. Gupta R, Gupta N, Rathi P (2004) Bacterial lipases: an overview
of production, purification and biochemical properties. Appl
Microbiol Biotechnol 64:763–781
2. Mosbah H, Sayari A, Mejdoub H, Dhouib H, Gargouri Y (2005)
Biochemical and molecular characterization of Staphylococcus
xylosus lipase. Biochim Biophys Acta 1723:282–291
3. Tiesinga JJW, van Pauderoyen G, Nardini M, Ransac S, Dijkstra
BW (2007) Structural basis of phospholipase activity of Staphy-
lococcus hyicus lipase. J Mol Biol 371:447–456
4. Houde A, Kademi A, Leblanc D (2004) Lipases and their
industrial applications. Appl Biochem Biotechnol 118:155–170
5. Hasan F, Shah AA, Hameed A (2006) Industrial applications of
microbial lipases. Enzyme Microb Technol 39:235–251
6. Doukyu N, Ogino H (2010) Organic solvent tolerant enzymes.
Biochem Eng J 48:270–282
7. Joseph B, Ramteke PW, Thomas G (2008) Cold active microbial
lipases: some hot issues and recent developments. Biotechnol
Adv 26:457–470
8. Rahman RNZRA, Kamarudin NHA, Yunus J, Salleh AB, Basri M
(2010) Expression of an organic solvent stable lipase from
Staphylococcus epidermidis AT2. Int J Mol Sci 11:3195–3208
9. Kwon DY, Rhee JS (1986) A simple and rapid colorimetric
method for determination of free fatty acids for lipase assay.
J Am Oil Chem Soc 63:89–92
10. Laemmli UK (1970) Cleavage of structural proteins during the
assembly of the head of bacteriophage T4. Nature 227:680–685
11. Sommer P, Bormann C, Gotz F (1997) Genetic and biochemical
characterization of a new extracellular lipase from Streptomyces
cinnamomeus. Appl Environ Microbiol 63:3553–3560
12. Nybo K (2012) Troubleshooting forum, molecular biology and
techniques: native PAGE. Biotechniques 52:20–21
13. Arakawa T, Philo JS, Kita Y (2001) Kinetic and thermodynamic
analysis of thermal unfolding of recombinant erythropoietin.
Biosci Biotechnol Biochem 65:1321–1327
14. Arakawa T, Philo JS, Ejima D, Tsumoto K, Arisaka F (2006)
Aggregation analysis of therapeutic proteins, part 1. General
aspects and techniques for assessment. BioProcess Int 4:42–43
15. Skagerlind P, Folmer B, Jha BK, Svensson M, Holmberg K
(1998) In: Lindman B, Ninham BW (eds) The colloid science of
lipids, lipase-surfactant interactions. Steinkopff Publisher,
Dresden
16. Laane C, Boeren S, Vos K, Veeger C (1987) Rules for the
optimization of biocatalysis in organic solvents. Biotechnol
Bioeng 30:81–87
17. van Oort MG, Deveer AM, Dijkman R, Tjeenk ML, Verheij HM,
de Haas GH, Wenzig E, Gotz F (1989) Purification and substrate
specificity of Staphylococcus hyicus lipase. Biochemistry
28:9278–9285
18. Oh BC, Kim HK, Lee JK, Kang SC, Oh TK (1999) Staphylo-
coccus haemolyticus lipase: biochemical properties, substrate
specificity and gene cloning. FEMS Microbiol Lett 179:385–392
19. Sakinc T, Klein B, Gattermann GS (2007) Biochemical charac-
terization of the surface-associated lipase of Staphylococcus
saprophyticus. FEMS Microbiol Lett 274:335–341
20. Tyski S, Hryniewicz W, Jeljaszewicz J (1983) Purification of and
some properties of the extracellular lipase. Biochim Biophys Acta
749:312–317
21. Cai Y, Wang L, Liao X, Ding Y, Sun J (2009) Purification and
partial characterization of two new cold adapted lipases from
Geotrichum sp. SYBC.WU-3. Process Biochem 44:786–790
22. Simkhada JR, Yoo HY, Cho SS, Choi YH, Kim SW, Park DH,
Yoo JC (2012) A novel cold-adapted lipase, LP28, from a mes-
ophilic Streptomyces strain. Bioprocess Biosyst Eng 35:217–225
23. Feller G (2013) Psychrophilic enzymes: from folding to function
and biotechnology. Scientifica 512840. doi:10.1155/2013/512840
24. Alquati C, De Gioia L, Santarossa G, Alberghina L, Fantucci P,
Lotti M (2002) The cold-active lipase of Pseudomonas fragi.
Heterologous expression, biochemical characterization and
molecular modeling. Eur J Biochem 269:3321–3328
25. Aghajari N, Feller G, Gerday C, Haser R (1998) Structures of the
psychrophilic Alteromonas haloplanctis a-amylase give insights
into cold adaptation at a molecular level. Structure 6:1503–1515
26. Fu J, Leiros H-KS, De Pascale D, Johnson KA, Blencke HM,
Landfald B (2013) Functional and structural studies of a novel
cold-adapted esterase from an Arctic intertidal metagenomic
library. App Microbiol Biotechnol 97:3965–3978
27. Sayari A, Agrebi N, Jaoua S, Gargouri Y (2001) Biochemical and
molecular characterization of Staphylococcus simulans lipase.
Biochimie 83:863–871
28. Simons JW, Adams H, Cox RC, Dekker N, Gotz F, Slotboom AJ,
Verheij HM (1996) The lipase from Staphylococcus aureus.
Expression in Escherichia coli, large-scale purification and
comparison of substrate specificity to Staphylococcus hyicus
lipase. Eur J Biochem 242:760–769
306 N. H. A. Kamarudin et al.
123

29. Andreini C, Bertini I, Cavallaro G, Holliday GL, Thornton JM
(2008) Metal ions in biological catalysis: from enzyme databases
to general principles. J Biol Inorg Chem 13:1205–1218
30. van Kampen MD, Rosenstein R, Gotz F, Egmond MR (2001)
Cloning, purification and characterisation of Staphylococcus
warneri lipase 2. Biochim Biophys Acta 1544:229–241
31. Tjeenk ML, Bulsink YBM, Slotboom AJ, Verheij HM, de Haas
GH, Demleitner G, Gotz F (1994) Inactivation of Staphylococcus
hyicus lipase by hexadecylsulfonyl fluoride: evidence for an
active site serine. Protein Eng 7:579–583
32. Das AK, Bellizzi JJ III, Tandel S, Bieh E, Clardy J, Hofmann SL
(2000) Structural basis for the insensitivity of a serine enzyme
(palmitoyl-protein thioesterase) to phenylmethylsulfonyl fluoride.
J Biol Chem 275:23847–23851
33. Mogensen JE, Sehgal P, Otzen DE (2005) Activation, inhibition,
and destabilization of Thermomyces lanuginosus lipase by
detergents. Biochemistry 44:1719–1730
34. Delorme V, Dhouib R, Canaan S, Fotiadu F, Carriere F, Cavalier
J-F (2011) Effects of surfactants on lipase structure, activity, and
inhibition. Pharm Res 28:1831–1842
35. Chakravorty D, Parameswaran S, Dubey VK, Patra S (2012)
Unraveling the rationale behind organic solvent stability of
lipases. Appl Biochem Biotechnol 167:439–461
36. Lousa D, Baptista AM, Soares CM (2013) A molecular per-
spective on nonaqueous biocatalysis: contribution from simula-
tion studies. Phys Chem Chem Phys 15:13723–13736
37. Rahman RN, Geok LP, Basri M, Salleh AB (2005) An organic
solvent tolerant protease from Pseudomonas aeruginosa strain K.
Enzyme Microb Technol 36:749–757
38. Volpato G, Rodrigues RC, Heck JX, Ayub MAZ (2008) Pro-
duction of organic solvent tolerant lipase by Staphylococcus
caseolyticus EX17 using raw glycerol as substrate. J Chem Tech
Biotechnol 83:821–828
39. Fang Y, Lu Z, Lv F, Bie X, Liu S, Ding Z, Xu W (2006) A newly
isolated organic solvent tolerant Staphylococcus saprophyticus
M36 produced organic solvent-stable lipase. Curr Microbiol
53:510–515
40. Esakkiraj P, Rajkumarbharathi M, Palavesam A, Immanuel G
(2010) Lipase production by Staphylococcus epidermidis CMST-
Pi 1 isolated from the gut of shrimp. Ann Microbiol 60:37–42
41. Ogino H, Uchiho T, Doukyu N, Yasuda M, Ishimi K, Ishikawa H
(2007) Effect of exchange of amino acid residues of the surface
region of the PST-01 protease on its organic solvent-stability.
Biochem Biophys Res Commun 13:1028–1033
Stable Lipase from Mesophilic Staphylococcus epidermidis 307
123





























