Embed Size (px)
Citation preview

Vol. 172, No. 8
A Gene Required for Very Short Patch Repair in Escherichia coli IsAdjacent to the DNA Cytosine Methylase Gene
ANJUM SOHAIL,"2 MARGARET LIEB,3 MUBASHER DAR,4 AND ASHOK S. BHAGWATl.4*Cold Spring Harbor Laboratory, Cold Spring Harbor, New York 117241; Centre for Advanced Molecular Biology,
University of the Punjab, Lahore-20, Pakistan2; University of Southern California School of Medicine, Los Angeles,California 900333; and Department of Chemistry, Wayne State University, Detroit, Michigan 482024
Received 14 March 1990/Accepted 9 May 1990
Deamination of 5-methylcytosine in DNA results in T/G mismatches. If unrepaired, these mistches canlead to C-to-T transition mutations. The very short patch (VSP) repair process in Escherichia coli counteractsthe mutagenic process by repairing the mismatches in favor of the G-containing strand. Previosly we haveshown that a plasmid containing an l1-kilobase fragment from the E. coli chromosome can complement achromosomal mutation defective in both cytosine methylation and VSP repair. We have now mapped theregions essential for the two phenotypes. In the process, we have constructed plasmids that complement thechromosomal mutation for methylation, but not for repair, and vice versa. The genes responsible for thesephenotypes have been identified by DNA sequence analysis. The gene essential for cytosine methylation, dem,is predicted to code for a 473-amino-acid protein and is not required for VSP repair. It is similar to other DNAcytosine methylases and shares extensive sequence similarity with its isoschizomer, EcoRl methylase. Thesegment of DNA essential for VSP repair contains a gene that should code for a 156-amino-acid protein. Thisgene, named vsr, is not essential for DNA methylation. Remarkably, the 5' end of this gene appears to overlapthe 3' end of dcm. The two genes appear to be transcribed from a common promoter but are in differenttranslational registers. This gene arrangement may assure that Vsr is produced along with Dcm and mayminimize the mutagenic effects of cytosine methylation.
Base mismatches can arise in DNA for a variety ofreasons. These include replication errors and the action ofchemical mutagens that react with DNA (for a review, seereference 30). An additional cause for the generation of basemismatches is the inherent instability of cytosine. Underphysiological conditions, cytosine deaminates to uracil atappreciable rates (24). The resulting uracil/guanine mis-matches are repaired by a process which, in the first step,involves the removal of uracil (23). 5-Methylcytosine is moreunstable than cytosine (10, 24), and its deamination results inthe creation of thymine/guanine (T/G) mismatches. If notcorrectly repaired prior to replication, C-to-T transitionsoccur. Cytosine methylation sites in DNA have been foundto be "hotspots" for such mutations in Escherichia coli (7;M. Lieb, unpublished results).The only DNA cytosine methylase in E. coli K-12, Dcm,
transfers a methyl group to position 5 of the internal cytosinein the sequence 5'-CCWGG-3', where W is A or T (29). E.coli also contains a base mismatch correction process thatcorrects T/G mismatches in sequence contexts such as ClAGG/GjTCC and CITGG/GGACC to C/G (21). This pro-cess, called very short patch (VSP) repair, reduces themutagenic effect of cytosine methylation (M. Lieb, unpub-lished results). It is likely that this process arose in E. coli toreduce the mutagenicity of 5-methylcytosine.Marinus and Morris used a chemical mutagen to obtain a
series of E. coli mutations, dcm-1 through dcm-11, that weredefective in the methylation of cytosines in DNA (28). Oneof these mutations, dcm-6, was later shown to lead to theloss of VSP repair in the cell as well (14, 20, 40). This led tothe conclusion that Dcm was required for VSP repair andmight be a bifunctional protein. We recently showed that aplasmid containing an 11-kilobase fragment of chromosomal
* Corresponding author.
DNA from wild-type E. coli complements both the methyl-ation and repair defects of dcm-6 (22). This is consistent withthe suggestion that Dcm plays a direct role in VSP repair. Inthat study, we also tested the gene for another methylase,EcoRII, for its ability to complement dcm-6 for its repairdefect. EcoRII methylase is part of the EcoRII restriction-modification system and has sequence specificity identical tothat of Dcm (29, 35). Hence, the gene that codes for theEcoRII methylase complements dcm-6 for methylation. In-terestingly, it did not restore VSP repair in the cell (22). Asa result of these observations, it was concluded that theability of EcoRII to methylate Dcm/EcoRII sites is notsufficient for that enzyme to have a role in VSP repair. Wereport here that this is true of Dcm as well. Contrary toprevious reports, dcm plays no direct role in VSP repair.Instead, a gene adjacent to it is required for this process. Inturn, this gene appears to play no role in DNA methylation.(A preliminary report of this work was published [4] as the
summary of a paper presented at the New England BioLabsWorkshop on Biological DNA Modifications, Gloucester,Mass., 20 to 23 May 1988.)
MATERIALS AND METHODS
Bacterial strains. GM1 [F- thr-l ara-14 leuB6 A(gpt-proA)62 lac Yl tsx-33 supE44 galK2 hisG4 rflhbD mgl-S1rpsL31 kdgKSJ xyl-S mtl-l metBI thi-1]; its derivativescarrying various dcm alleles; GM30 (thr-J hisG4 leuB6 rpsLara-14 supE44 lacYl tonA31 tsx-78 galK2 galE2 xylS thi-lmtl-i) and GM31 (i.e., GM30 dcm-6) were kindly providedby M. G. Marinus (University of Massachusetts School ofMedicine, Amherst, Mass.). RP4182 [A(supD-dcm-fla) trpgal rpsL] was provided by J. S. Parkinson (University ofUtah, Salt Lake City, Utah).Assay for methylation of Dcm sites. The ability of a
bacterial strain to methylate Dcm/EcoRII sites was assessed
4214
JOURNAL OF BACTERIOLOGY, Aug. 1990, p. 4214-42210021-9193/90/084214-08$02.00/0Copyright © 1990, American Society for Microbiology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.

GENE REQUIRED FOR VSP REPAIR IN E. COLI 4215
in two different ways. Plasmid DNAs from the strains were
routinely digested with EcoRII endonuclease (Bethesda Re-search Laboratory, Rockville, Md.), and the result wasanalyzed by electrophoresis on an agarose gel. Methylationof the Dcm/EcoRII sites results in the protection of theplasmid DNA against cleavage. Quantitation of the methyl-ase activity was done for selected strains by determining thenumber of 3H atoms transferred from [3H]adenosyl-L-methi-onine (Dupont, NEN Research Products, Boston, Mass.) tochromosomal DNA from RP4182 as described before (5).
Assay for VSP repair. The percentage of cI+ progenyamong N+ O+ recombinants resulting from a cross ofbacteriophage mutants Nam53 cIam6 O+ and N+ cI CP7Oam29 was used to quantitate VSP repair as describedbefore (21). Mutation am6 changes a CCAGG sequence toCTAGG; mutation CP7 is a T-to-A transversion located 20base pairs (bp) to the left of am6 on the X map. Briefly,bacteria were infected with a 1:1 mixture of the two mutantsto give a total multiplicity of infection of 10. After a 15-minadsorption period at 37°C, broth was added and incubationwas continued for a total of 100 min, followed by the additionof CHCl3 and vortexing. Recombinants that are N+ O+ wereselected by plating the phage progeny on a lawn of Supobacteria; N+ O+ phage that are also cI+ form turbid plaques,while phage that carry the am6 or the CP7 mutation in cI
form clear plaques. N+ O+ recombinants that are also cI+require three crossover events and are expected to be veryrare. However, during the recombinational events leading toN+ O+ phage, heteroduplex regions span the intervening cI
gene, resulting in am6/+ (T/G) mismatches. Correction ofam6 (T) to (C) results in excess N+ cI+ O recombinants.Therefore, the fraction of cI+ among the recombinant prog-eny phage is a measure of VSP repair in the host.Recombinant DNA work and DNA sequencing. Standard
techniques were used in the construction of Bal3l-mediateddeletions, deletion of restriction fragments, and constructionof M13 phage clones (26). Inserts in the M13 clones were
sequenced by using appropriate primers by the technique ofSanger et al. (34).
RESULTS
VSP repair and Dcm phenotypes can be unlinked. Threeresearch groups have independently reported that E. coliK-12 cells carrying the dcm-6 mutation are deficient in VSPrepair (14, 20, 40). dcm-6 is 1 of the 11 independent alleles ofdcm isolated by Marinus and Morris by chemical mutagen-esis of the strain GM1 (28). The nature of the mutation(s) isnot known for any of the alleles. Three of the dcm mutantalleles have been lost; we have tested five of the remainingalleles for their effect on VSP repair. The results are sum-marized in Table 1.When a bacteriophage X cross diagnostic for VSP repair
was done in GM1, the percentage of cI+ recombinantsobtained was about 1.8% (Table 1). This number is similar tothat obtained in another dcm' strain, GM30 (Table 1) (20).In GM31, the dcm-6 derivative of GM30, this number wassixfold lower (Table 1) (20), indicating a lower level of VSPrepair. GM1 derivatives containing the alleles dcm-9 ordcm-10 showed a similarly low level of repair activity (Table1). These mutations caused 10- and 12-fold reductions,respectively, in the Dcm activity in the cell (28). In contrast,GM1 derivatives carrying the alleles dcm4 or dcm-7 showedthe same level of repair activity as GM1 and hence are VSPrepair proficient (Table 1). Surprisingly, the derivative withthe allele dcm-J showed slightly higher VSP repair activity
TABLE 1. VSP repair phenotypes of dcm mutations
Strain Allele % of cI+ recombinantsa
GM30 dcm' 2.2 ± 0.6GM31 dcm-6 0.36 ± 0.15GM1 dcm' 1.8 ± 0.3GM5 dcm-1 6.0 ± 1.9GM8 dcm4 2.0 + 0.5GM11 dcm-7 2.5 ± 0.4GM13 dcm-9 0.32 ± 0.19GM14 dcm-1O 0.49 ± 0.07RP4182 Adcm 0.08 ± 0.03
a Among N+ O+ progeny (see Materials and Methods). Each value is anaverage of data from three or more experiments.
than GM1. Mutations dcm-J, dcm4, and dcm-7 cause 6-,500-, and 2-fold reductions, respectively, in the Dcm activityin the cell (28). It is clear that there is no direct correlationbetween the Dcm activity and VSP repair activity in thesestrains. From these data we concluded that, although Dcmand VSP repair are tightly linked to each other, the twophenotypes can be unlinked.
It should also be noted that there may not be a completeloss of VSP repair activity in dcm-6-, dcm-9-, or dcm-10-containing strains. RP4182 is a strain of E. coli K-12 thatcarries a large deletion at about 43' on the chromosome andis missing several genes including dcm (2, 5, 17). It consis-tently produced three- to fourfold fewer cI+ recombinants ina similar cross (Table 1). It is possible that this is a result ofthe differences in the genetic backgrounds of GM1 andRP4182. It is more likely that there is some residual VSPrepair in the strains containing the dcm-6, dcm-9, or dcm-10alleles, and the reason for the lower numbers of cI+ recom-binants in RP4182 is that the deletion in its chromosomeeliminates a gene(s) essential for VSP repair.Dcm and VSP repair phenotypes can be separated biochem-
ically. We have shown that a multicopy plasmid, pDCM1,that contains an 11-kilobase-pair DNA fragment from the E.coli chromosome (5), complements the dcm-6 mutation forVSP repair as well as methylation at the Dcm sites (22).Another plasmid, pDCM3, containing an overlapping chro-mosomal fragment (5), also complements this mutation formethylation and repair (data not shown). pDCM4 was cre-ated by deleting a HindlIl fragment within pDCM3 (5). Thisplasmid also complements the dcm-6 mutation for both thephenotypes, i.e., it has the phenotype methylation-positiveVSP repair-positive (M+ Vsr+) (22).
Bal3l deletions were generated from three unique sites inpDCM4 (BamHI, StuI, and Sall), and the resulting deriva-tives were tested for methylation and VSP repair (Fig. 1).Short deletions that removed sequences from the right sideof the chromosomal insert had little effect on either pheno-type (pDCM22, pDCM23, and pDCM33). Larger leftwarddeletions from the BamHI site showed two new phenotypes,M+ Vsr- and M- Vsr- (Fig. 1). All constructs that had morethan about 2.0 kilobases ofDNA removed from the right endwere Vsr- (i.e., pDCM8, pDCM9, and pDCM21). One ofthese plasmids, pDCM21, fully complemented the dcm-6mutation for methylation. Another, pDCM8, showed a re-duced level of methylation activity. DNA prepared fromRP4182 containing this plasmid was only partially resistantto the EcoRII endonuclease (data not shown). Deletionsfrom the right end that removed greater than 2.7 kilobasepairs of the insert were defective in methylation as well as inVSP repair (pDCM9; data not shown).The Vsr- phenotype could also be created by the insertion
VOL. 172, 1990
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.
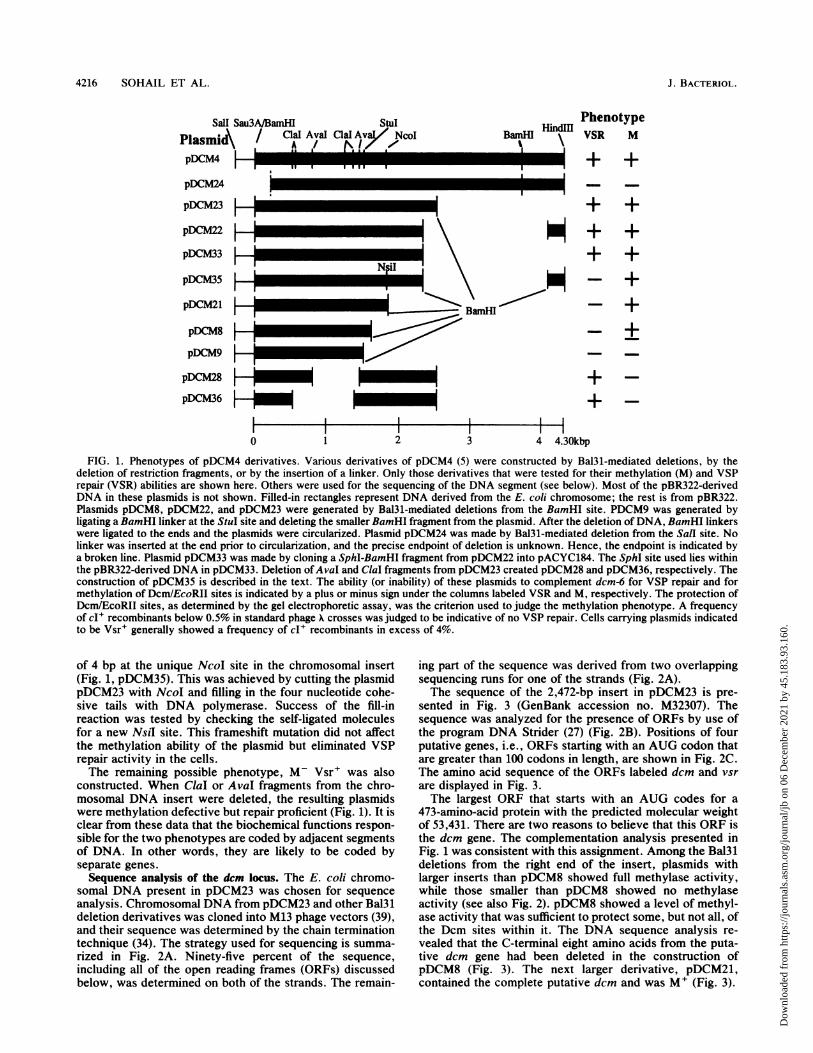
4216 SOHAIL ET AL.
Sail Sau3A/BamHIPlasmi / Cial AvalpDCM4
pDCM24
pDCM23 HpDCM22 1
pDCM33
pDCM35 1
pDCM21
pDCM48
pDCM9pDCM28pDCM36 I
StulClalAY NcOI
l\i.
FIG. 1. Phenotypes of pDCM4 derivatives. Various derivatives of pDCM4 (5) were constructed by Bal3l-mediated deletions, by thedeletion of restriction fragments, or by the insertion of a linker. Only those derivatives that were tested for their methylation (M) and VSPrepair (VSR) abilities are shown here. Others were used for the sequencing of the DNA segment (see below). Most of the pBR322-derivedDNA in these plasmids is not shown. Filled-in rectangles represent DNA derived from the E. coli chromosome; the rest is from pBR322.Plasmids pDCM8, pDCM22, and pDCM23 were generated by Bal3l-mediated deletions from the BamHI site. PDCM9 was generated byligating a BamHI linker at the StuI site and deleting the smaller BamHI fragment from the plasmid. After the deletion ofDNA, BamHI linkerswere ligated to the ends and the plasmids were circularized. Plasmid pDCM24 was made by Bal31-mediated deletion from the SalI site. Nolinker was inserted at the end prior to circularization, and the precise endpoint of deletion is unknown. Hence, the endpoint is indicated bya broken line. Plasmid pDCM33 was made by cloning a SphI-BamHI fragment from pDCM22 into pACYC184. The SphI site used lies withinthe pBR322-derived DNA in pDCM33. Deletion ofAvaI and ClaI fragments from pDCM23 created pDCM28 and pDCM36, respectively. Theconstruction of pDCM35 is described in the text. The ability (or inability) of these plasmids to complement dcm-6 for VSP repair and formethylation of Dcm/EcoRII sites is indicated by a plus or minus sign under the columns labeled VSR and M, respectively. The protection ofDcm/EcoRII sites, as determined by the gel electrophoretic assay, was the criterion used to judge the methylation phenotype. A frequencyof cI+ recombinants below 0.5% in standard phage crosses was judged to be indicative of no VSP repair. Cells carrying plasmids indicatedto be Vsr+ generally showed a frequency of cI+ recombinants in excess of 4%.
of 4 bp at the unique NcoI site in the chromosomal insert(Fig. 1, pDCM35). This was achieved by cutting the plasmidpDCM23 with NcoI and filling in the four nucleotide cohe-sive tails with DNA polymerase. Success of the fill-inreaction was tested by checking the self-ligated moleculesfor a new NslI site. This frameshift mutation did not affectthe methylation ability of the plasmid but eliminated VSPrepair activity in the cells.The remaining possible phenotype, M- Vsr+ was also
constructed. When ClaI or AvaI fragments from the chro-mosomal DNA insert were deleted, the resulting plasmidswere methylation defective but repair proficient (Fig. 1). It isclear from these data that the biochemical functions respon-sible for the two phenotypes are coded by adjacent segmentsof DNA. In other words, they are likely to be coded byseparate genes.Sequence analysis of the dem locus. The E. coli chromo-
somal DNA present in pDCM23 was chosen for sequenceanalysis. Chromosomal DNA from pDCM23 and other Bal3ldeletion derivatives was cloned into M13 phage vectors (39),and their sequence was determined by the chain terminationtechnique (34). The strategy used for sequencing is summa-rized in Fig. 2A. Ninety-five percent of the sequence,including all of the open reading frames (ORFs) discussedbelow, was determined on both of the strands. The remain-
ing part of the sequence was derived from two overlappingsequencing runs for one of the strands (Fig. 2A).The sequence of the 2,472-bp insert in pDCM23 is pre-
sented in Fig. 3 (GenBank accession no. M32307). Thesequence was analyzed for the presence of ORFs by use ofthe program DNA Strider (27) (Fig. 2B). Positions of fourputative genes, i.e., ORFs starting with an AUG codon thatare greater than 100 codons in length, are shown in Fig. 2C.The amino acid sequence of the ORFs labeled dcm and vsr
are displayed in Fig. 3.The largest ORF that starts with an AUG codes for a
473-amino-acid protein with the predicted molecular weightof 53,431. There are two reasons to believe that this ORF isthe dcm gene. The complementation analysis presented inFig. 1 was consistent with this assignment. Among the Bal3Mdeletions from the right end of the insert, plasmids withlarger inserts than pDCM8 showed full methylase activity,while those smaller than pDCM8 showed no methylaseactivity (see also Fig. 2). pDCM8 showed a level of methyl-ase activity that was sufficient to protect some, but not all, ofthe Dcm sites within it. The DNA sequence analysis re-vealed that the C-terminal eight amino acids from the puta-tive dcm gene had been deleted in the construction ofpDCM8 (Fig. 3). The next larger derivative, pDCM21,contained the complete putative dem and was M+ (Fig. 3).
I I I I I I
BamHI Hind\lPhenotypeVSR M
+ +
+ +
++
BamHI
I1 2 3
l43+4 4.3Okbp
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.

GENE REQUIRED FOR VSP REPAIR IN E. COLI 4217
-t-,
A.0
-242b2472 bp
4-4 4-4 4<< +-
4-
dcm lllll
Rdcmse
I EcoRII Methylase
1
2
3
-1
-2
-3
> < EcoRII Endonuclease |
FIG. 2. (A) Sequencing scheme. The lengths of different sequencing runs are indicated by arrows. Arrows with solid arrowheads indicateruns from M13-specific primer, while the arrows with empty arrowheads indicate runs from primers synthesized from the sequencedetermined in the earlier sequencing runs. (B) ORFs in the sequence. ORFs in the sequence were analyzed by use of DNA Strider (27) ona Macintosh IIcx computer. The six panels represent the six possible registers for the ORFs in the sequence. Full-height bars within eachpanel represent the position of a stop codon in the sequence, while bars with half the height of the panel represent positions of AUG codons.(C) Possible genes in the sequence. Positions of ORFs that contain more than 100 codons and that start with an AUG codon are indicated.(D) EcoRII genes. Arrangement of the EcoRII endonuclease and methylase genes in relation to the dcm-vsr arrangement is shown. TheEcoRII methylase is positioned below the dcm gene so that the homologous regions between the two genes are aligned.
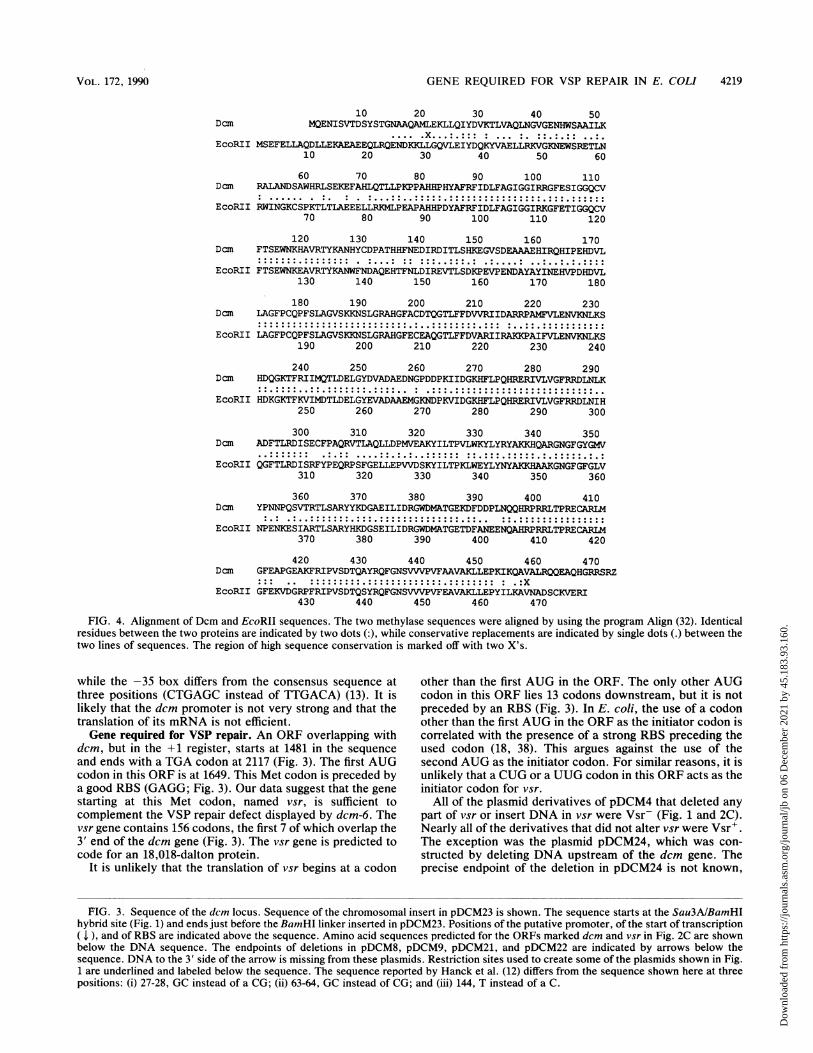
The second reason for the assignment of this ORF as dcmis the similarity of its sequence to the EcoRII methylasesequence. EcoRII is known to methylate the same sequenceas Dcm, and the genes for EcoRII reside on a natural E. coliplasmid (29, 35). Figure 4 shows an alignment of the pre-dicted amino acid sequence of the two proteins. Over a444-amino-acid stretch, 68.2% residues were identical be-tween the two proteins. The sequence conservation wasseen at the DNA level as well. In a 1,427-bp segment thatstarts about 40 bp before the initiator AUG codon of dcmand ends 40 bp before the termination codon of dcm, therewas 66.1% identity between the two sequences (data notshown). Clearly, Dcm and EcoRII methylase are closeevolutionary relatives.A map of the E. coli chromosome constructed with eight
restriction enzymes has been published by Kohara et al.(16). We aligned the map predicted by our sequence withthat presented by these researchers (data not shown). Thisalignment predicts that the dcm gene is transcribed counter-clockwise around the E. coli map and that the segment ofDNA required for Dcm and VSP repair activities should lieon the phage A clones 8D10 and 25D8 of Kohara et al. (16).The sequence of the insert in the clone 25D8 was recently
reported by Hanck et al. (12). The sequence reported bythese authors agrees with the sequence reported here exceptat three positions. These are noted in the legend to Fig. 3.These differences do not affect any of the ORFs discussed
above and may represent sequence polymorphism betweendifferent strains of E. coli. While we used DNA from thestrain DH1 (X- F- recAl supE44 gyrA96 hsdRJ7 endAI thi-JrfbDJ? relAl? spoTI?), the phage A clones of Kohara et al.(16) were constructed with DNA from W3110 [A- F- IN(rrnD-rrnE)J]. Hanck et al. (12) did not discuss the functionsof the various ORFs in their sequence.The PIR/NBRF data base (March 1989 release, 3,002,182
residues in 11,158 sequences) was searched with the FASTAprogram (25, 32) to find sequences similar to Dcm. All eightDNA cytosine methylases in the data base were found by theprogram, and these sequences scored between 1,666 (EcoRIImethylase) and 84 (DdeI methylase) against Dcm. Dcmscored 2,365 against itself. No other protein in the data basescored more than 74. Ten conserved sequence motifs wererecently identified in all of the DNA cytosine methylases(33), and Dcm protein contains all of these motifs. It is clearthat Dcm is similar to other cytosine methylases but is unlikeother proteins for which the primary sequence is known.The initiation codon for the dcm gene is preceded by
GGUG, a possible ribosome-binding site (RBS) which ispreceded by -10 and -35 boxes for a promoter. Neither theRBS nor the putative promoter is closely related to theconsensus sequences for these elements. The RBS consistsof only four of the possible six conserved bases. The -10box for the promoter is different from the consensus se-
quence at one position (TGTAAT instead ofTATAAT) (13),
B. 1
2
3-1
-2
-3
C.
D.
VOL. 172, 1990
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.

4218 SOHAIL ET AL. J. BAcTERIOL.
ULA - Ln 0 LA 0 Ln 0 Ln 0 Ln 0 L 0
(N ONCm) 0 (N LA r- 0 (N LA r- 0 (N LA r 0 (N LA)
r-ilw LA OD 0 (4N Cm) LA D r- 0 (Ncl
(N H 4C~-N (N (N C)
C)uH< a -4 u -
H - u H- FH E- .u - -4H u *
uuO H Ou O O4U >H H 0co~ C) HC) 0 0u u u u <W.O OO W0EHO0 H u U<EH0
u up u E-4 > H 40 Hu > 4E- u O<OH H O
O( O O O4U04Z 0U E u 8 p~) H o-< 0O=HHu OH u u4O0 E-O O 0 U H>O Hu
o4 O PC H HO O u 04m H '-4 CN 0~004
C) uul 0 0 E- PC 0a4H> 0 O 4 0 04 0W. H E04
u H~0OA H 0 0 H4 uEO0 0 W. HOp
400 H 4 u PCcHHO 0 OE-0 0 HO4 H -4
O~~~~~~~O~~~~~~~O0~~~z- E-O0:H PC>4H 0- 0 O3 HHE (N
0040 0 0 HE-4 HE- 0 0-O-4 EHE-4
H UE-4 -4 uO lH H u0H 0 e4>4EH uH H
4 O0 HH4H 0 0 uOc 0D H- 0-O 0IA 0 00 0 0 0
H
E-
OHE O 0
E-4
0 H>O 00 0OH04E- 0
H HH u0 Wu E-4 H H u4H 04u H4 PC H H
E-iU~4 U E-4 U u 0 H E-1 HHuu< -
u u 8 u E-0 .- >C -AC)
H- 4O 0 H 4lO0 0 HH 00 4 0 0 4 Ol <
4 E-4 E-4~~~~0Oc )4 H
HC)M: H0EHu4C u u HO H
0E> 4(mOH H-4 H E-AH0u 4 uH 0 000 0EuO0 p(H 0 u
U 0 E-4 H EH 0>0PCH>4LDC 0 H 0 H >uE u
O 00E-0OH H>W 0 0 0 HO 0H C
%( HH ) u~ H> O )-4O HE- H> H 0 E-H4Hu u LD uu uuuu0O 0(1)0E-0-- 0 uEO C E0
C)4H0 uE0 004 E-4 Hz EHH H> OO u <
Ou ~ O H 0 ()uu u> 0 U 00 H H> 0 u >40a 00E- iO H4 4 H 00u O 0 P0 uaO 0 uH 0~ 0 4 0
O 00 H H O O 0 (D0 OA 0 0 O( 004u o
H O uE LO1)HD ~ H:u a-0 H H0u()C)E- uUE0 H
O HO (.<W O 1)E-4Hn-1H Hu E- E-4 H 9O H 0000O U 0<OZHC)~ E- -4 C0 0 0u H- u 0H u C
OHu HE-1E-j0 ( H H u-) H> 00 C) ) H
U00400WHuzLD E0 0.4 E- 004 0 HHHg
u Uu H E- 4O H. C
EH
rL40 1)
ZO 0 O- C ) O
LA EC) H
E-4 Pz oo)0 ~
< OCO C)u H P
0zu
OHH >
H C)O C)
0 =H u u0 OAu EO C) C)- C)<
< HO H
mOO 0 00 0 H H> u-1 0 E-H H H4
0 O 0 1)
PC E-H >
-4 ( PC 0 C0 0 C)u E-4 C4E-
UE-O H -0 HHOc HHO 00U 0 E-O H E-HO
OE H H 000z0 UHu- HH o< >4 H-4 0 O C) EHH uH HOOu Z
W EO (10 0
j4E- < u u E4o
uO Ou O
u (L )0-H> HE- (u u00 0 OHuU 0
O 00E 0 0 00 H> OH4 a 0 0o O uC
4 E- 00
Hu 0 H
0H
E0444 0 4 00u c
O 0 Hu E" 4
u(O O H4 E- u aH EL40 H(LE- 0 0(/) 0- uE-)O
u 4 HIZC) LH HC 0) (u W4H Hu LDuO0 0
E-uHE( u E-X !)
0 04
0 H >04 0EO4OC)
U E-4 Ou 0oc= C) 0< HME- o WCOct u
H E0-1O4 044E- 0a 0E- E-H 0E-
OOHH-4 a0 40 a 0- 00uU
E-UE4 t 0D o&4H o( U0 E0 0> H
s<< 4
E-HOOH4a4 OHO)D u 4 4E4
E-4 E-4u~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~0~
HH 0 00HuFu HH 00 E-4 0 WOH O4. 0 H C 0-4 E-OO H ~ E- H Ol)H OH u u
C) OHHO)
C)E
H4 0 E
04 0-4 E
OHr&C) O uC)
-4E-Oo0 0004 0- Oc 0u u CuE)4H 0 uuH 0
u E-QO rZHE- HHO 0U 04 0l u 00U 0
E- U : -4 H- H4 H4 H. HC4 0 E0 4 u E8 oO C
E-4E-4o to u ) 0 OH 0E- 0 H 0 O H
E-4 u P u E- u 14H 84O ~0 0 E-4> CH4 < OHC. uH H H4r4 ) cu Uu0L)u H 4~H >E-0 E0 H E-)EOH--H 0 H >4 0-> >40 0-H 0 0O uH E-O 0 0E- u0 O 0 H a 1O 0 EO4 H>OO HO(
u 4E-O 0 0H404 ( c "ou4E0-0 o<0.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.
GENE REQUIRED FOR VSP REPAIR IN E. COLI 4219
Dcm
EcoRII
Dcm
EcoRII
Dcm
EcoRII
10 20 30 40 50MQENISVTDSYSTGNAAQAMLEKLLQIYDVKTLVAQLNGVGENHWSAAILK
MSEFELLAQDLLEKAEAEEQLRQENDKKLLGQVLEIYDQKYVAELLRKVGKNEWSRETLN10 20 30 40 50 60
60 70 80 90 100 110RALANDSAWHRLSEKEFAHLQTLLPKPPAHHPHYAFRFIDLFAGIGGIRRGFESIGGQCV
RWINGKCSPKTLTLAEEELLRKMLPEAPAHHPDYAFRFIDLFAGIGGIRKGFETIGGQCV70 80 90 100 110 120
120 130 140 150 160 170FTSEWNKHAVRTYKANHYCDPATHHFNEDIRDITLSHKEGVSDEAAAEHIRQHIPEHDVL
FTSEWNKEAVRTYW.NWFNDAQEHTFNLDIREVTLSDKPEVPENDAYAYINEHVPDHDVL130 140 150 160 170 180
180 190 200 210 220 230Dcm LAGFPCQPFSLAGVSKKNSLGRAHGFACDTQGTLFFDVVRIIDARRPAMFVLENVKNLKS
EcoRII LAGFPCQPFSLAGVSKKNSLGRAHGFECEAQGTLFFDVARIIRAKKPAIFVLENVKNLKS190 200 210 220 230 240
240 250 260 270 280 290Dcm HDQGKTFRIIMQTLDELGYDVADAEDNGPDDPKIIDGKHFLPQHRERIVLVGFRRDLNLK
EcoRII HDKGKTFKVINDTLDELGYEVADAAEMGKNDPKVIDGKHFLPQHRERIVLVGFRRDLNIH250 260 270 280 290 300
300 310 320 330 340 350Dan ADFTLRDISECFPAQRVTLAQLLDPMVEAKYILTPVLWKYLYRYAKKHQARGNGFGYGMV
EcoRII QGFTLRDISRFYPEQRPSFGELLEPVVDSKYILTPKLWEYYLYAKKAAGNGFGFGLV310 320 330 340 350 360
360 370 380 390 400 410Dcn YPNNPQSVTRTLSARYYKDGAEILIDRGWDMATGEKDFDDPLNQQHRPRRLTPRECARM
EcoRII NPENKESIARTLSARYHDGSEILIDRGWDMTGETDFANEENQAHRPRRLTPREI370 380 390 400 410 420
Dan
EcoRII
420 430 440 450 460 470GFEAPGEAKFRIPVSDTQAYRQFGNSVVVPVFAAVAKLLEPKIKQAVALRQQEAQHGRRSRZ...... . . .... ....GFEKVDGRPFRIPVSDTOSYROFGNSVVVPVFE'AVAKLLEPYILKAVNADSCKVE,RT
430 440 450 460 470
FIG. 4. Alignment of Dcm and EcoRII sequences. The two methylase sequences were aligned by using the program Align (32). Identicalresidues between the two proteins are indicated by two dots (:), while conservative replacements are indicated by single dots (.) between thetwo lines of sequences. The region of high sequence conservation is marked off with two X's.
while the -35 box differs from the consensus sequence atthree positions (CTGAGC instead of TTGACA) (13). It islikely that the dcm promoter is not very strong and that thetranslation of its mRNA is not efficient.Gene required for VSP repair. An ORF overlapping with
dcm, but in the +1 register, starts at 1481 in the sequenceand ends with a TGA codon at 2117 (Fig. 3). The first AUGcodon in this ORF is at 1649. This Met codon is preceded bya good RBS (GAGG; Fig. 3). Our data suggest that the genestarting at this Met codon, named vsr, is sufficient tocomplement the VSP repair defect displayed by dcm-6. Thevsr gene contains 156 codons, the first 7 of which overlap the3' end of the dcm gene (Fig. 3). The vsr gene is predicted tocode for an 18,018-dalton protein.
It is unlikely that the translation of vsr begins at a codon
other than the first AUG in the ORF. The only other AUGcodon in this ORF lies 13 codons downstream, but it is notpreceded by an RBS (Fig. 3). In E. coli, the use of a codonother than the first AUG in the ORF as the initiator codon iscorrelated with the presence of a strong RBS preceding theused codon (18, 38). This argues against the use of thesecond AUG as the initiator codon. For similar reasons, it isunlikely that a CUG or a UUG codon in this ORF acts as theinitiator codon for vsr.
All of the plasmid derivatives of pDCM4 that deleted anypart of vsr or insert DNA in vsr were Vsr- (Fig. 1 and 2C).Nearly all of the derivatives that did not alter vsr were Vsr+.The exception was the plasmid pDCM24, which was con-structed by deleting DNA upstream of the dcm gene. Theprecise endpoint of the deletion in pDCM24 is not known,
FIG. 3. Sequence of the dcm locus. Sequence of the chromosomal insert in pDCM23 is shown. The sequence starts at the Sau3AIBamHIhybrid site (Fig. 1) and ends just before the BamHI linker inserted in pDCM23. Positions of the putative promoter, of the start of transcription( I ), and of RBS are indicated above the sequence. Amino acid sequences predicted for the ORFs marked dcm and vsr in Fig. 2C are shownbelow the DNA sequence. The endpoints of deletions in pDCM8, pDCM9, pDCM21, and pDCM22 are indicated by arrows below thesequence. DNA to the 3' side of the arrow is missing from these plasmids. Restriction sites used to create some of the plasmids shown in Fig.1 are underlined and labeled below the sequence. The sequence reported by Hanck et al. (12) differs from the sequence shown here at threepositions: (i) 27-28, GC instead of a CG; (ii) 63-64, GC instead of CG; and (iii) 144, T instead of a C.
VOL. 172, 1990
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.

4220 SOHAIL ET AL.
but, based on restriction analysis, it is expected to remove200 bp or more from the chromosomal insert. It is likely thatthe deletion removes the dcm promoter. Hence, the ob-served lack of both the methylation and the repair abilities inthis plasmid may mean that the two genes are normallytranscribed from a common promoter. While no promoter-like sequences were found within 200 bp upstream of theinitiator AUG for vsr, an appropriately placed RBS could belocated (Fig. 3). It is possible that the vsr gene is transcribedfrom the dcm promoter but is translated independent of dcm.The deletion data presented above are not consistent with arole for either of the remaining ORFs longer than 100 codonsin length (Fig. 2C, arrows Y and Z) in either VSP repair or inthe methylation of Dcm sites.
DISCUSSION
We have shown that dcm, the gene that codes for the onlyDNA cytosine methylase in E. coli K-12, is not essential forthe mismatch correction process called VSP repair. We haveidentified a gene required for this repair process by comple-mentation and DNA sequence analysis. This gene, vsr, liesdownstream of dcm, and the two genes appear to be tran-scribed from a common promotor. Interestingly, the twogenes appear to have a 7-codon overlap.The lack of direct participation by dcm in VSP repair is
suggested by two pieces of data. At least three null alleles ofdcm show normal VSP repair activity. Also, deleting roughlytwo-thirds of the dcm gene does not eliminate the ability ofthe plasmid carrying the gene to complement a chromosomalmutation for VSP repair. Instead, a separate gene, vsr, isrequired for VSP repair. Three research groups have sug-gested that the observation that there is no VSP repair indcm-6 background implies that Dcm plays a direct role in therepair process (15, 20, 40). This appears to be incorrect.There are several possible reasons why dcm-6 and two
other mutations of dcm are pleiotropic. The dcm mutationswere generated with the chemical mutagen nitrosoguanidinewhich is known to introduce multiple mutations (11). Be-cause of the close proximity of dcm and vsr genes, separatemutations in these genes are likely to remain together insubsequent strain constructions. It is also possible that themutations are the result of insertion sequences, which showpolar effects. In addition, any mutations in the commonpromoter for the two genes are expected to affect expressionof both the genes. Finally, if the two genes do overlap in themanner suggested by the DNA sequence data, any mutationsin the region of overlap between dcm and vsr are likely toaffect both phenotypes. Sequence analysis on these dcmalleles should allow us to distinguish between the variouspossibilities.
E. coli contains several DNA repair systems that act onmismatched DNA. The VSP repair system is unique amongthem in requiring a specific sequence context for mismatchesthat are corrected. T/G mismatches are corrected by VSPrepair only if they occur within the sequence context of Dcmmethylation. If the T/G mismatch occurs in a sequencewhere its correction to C/G would lead to a sequence that isnot a normal substrate for Dcm (or is not a subset thereof),it is not subject to correction (19, 21). Mismatches such asIAGG/GTCC and CIAG/G(iTC, which are corrected byVSP repair (21), are likely to arise due to random errors inDNA replication. Hence, the result of their correction infavor of the guanine-containing strand is expected to beneutral in terms of mutations. This suggests that the VSPrepair process functions to minimize C-to-T mutations that
can arise as a result of the deamination of 5-methylcytosinein DNA. Hence, the repair must involve some protein(s) thatconfers the observed sequence specificity upon the process.If Dcm did have a direct role in VSP repair, it would be thelogical candidate for the job. In the absence of such a role forDcm, some other protein(s) must serve this function.
It is instructive to compare the gene arrangements in thedcm-vsr and the EcoRII restriction-modification systems(Fig. 2C and D) (3, 37). Although the methylase genes in thetwo systems are close evolutionary relatives, the EcoRIIendonuclease gene is unrelated to vsr (or to its inversecomplement). The close proximity of the endonuclease andmethylase genes in the EcoRII system assures the coordi-nate transfer of the two genes between cells, which is auseful feature for a restriction-modification system. Thedcm-vsr gene arrangement appears to allow a coordinateregulation of the two genes. E. coli appears to have shuffledtwo homologous methylase genes into different genetic con-texts to achieve different biological goals.Mismatch correction processes appear to proceed in one
of two ways. In one, illustrated by the dam-dependentprocess in E. coli (31), a set of proteins recognize themismatch and introduce a nick on the 5' side of an unmeth-ylated GATC on the "incorrect" strand. Several hundrednucleotides of the nicked strand are replaced by PolIII, andthe nick is sealed by a DNA ligase. Alternatively, the firststep in the correction process can be the removal of theincorrect base by an endoglycosylase. A nonspecific apu-rinic/apyrimidinic endonuclease can then introduce the nickrequired for the subsequent steps. An example of the latterpathway is the MutY-dependent mismatch correction sys-tem of E. coli (1).Although the biochemical steps involved in VSP repair
and the role of vsr in them remain to be elucidated, Vsr maybe the most interesting protein in this repair process. Theonly other genes known to be involved in VSP repair aremutL, mutS, and polA (9, 14, 20, 40; M. Lieb, unpublishedresults). Remarkably, the requirement for the products ofmutL and mutS, which are known to be involved in mis-match recognition in the dam-dependent process, can beovercome by the introduction of the vsr gene on a multicopyplasmid (M. Lieb, unpublished results). This suggests a rolefor Vsr in mismatch recognition. It should be noted, how-ever, that the Vsr protein does not strongly resemble anyprotein in the PIRINBRF data base. Specifically, it does notresemble MutH, MutS, or HexA, proteins known to beinvolved in other mismatch correction processes in E. coliand Streptococcus pneumonia.Gene overlap can be a means to gene regulation. It is often
a way of producing small amounts of a protein or ofproducing a protein late in the life cycle. Examples of thisphenomenon include the lysis genes in the viruses related toOX174 and the transposase of ISI (36; for recent reviews,see references 6 and 8). The dcm gene may play an indirectrole in VSP repair by controlling the expression of vsr. Asthe two genes appear to be transcribed from a commonpromoter, Vsr protein will be made only when Dcm isproduced. This is an advantageous arrangement becauseVSP repair is required only when there is cytosine methyl-ation of DNA in the cell. Elucidation of the regulatorymechanism controlling the expression of dcm-vsr should tellus a great deal about how E. coli has adapted to the presenceof 5-methylcytosine in its DNA.
J. BACTERIOL.
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.

GENE REQUIRED FOR VSP REPAIR IN E. COLI 4221
ACKNOWLEDGMENTS
This work was initiated while A.S. and A.S.B. were in thelaboratory of R. J. Roberts (Cold Spring Harbor Laboratory, ColdSpring Harbor, N.Y.), and they thank him for his support andencouragement.
This work was supported by National Science Foundation grantsPCM-8217553 (to R.J.R.), DMB-8516062 (to M.L.), and DMB-8806771 (to A.S.B.) and Public Health Service grant GM25034 (toM.L.) from the National Institutes of Health.
LITERATURE CITED1. Au, K. G., S. Clark, J. H. Miller, and P. Modrich. 1989.
Escherichia coli mutY gene encodes an adenine glycosylaseactive on G-A mispairs. Proc. Natl. Acad. Sci. USA 86:8877-8881.
2. Bale, A., M. d'Alarcao, and M. G. Marinus. 1979. Characteri-zation of DNA adenine methylation mutants of Escherichia coliK-12. Mutat. Res. 59:157-165.
3. Bhagwat, A. S., B. Johnson, K. Weule, and R. J. Roberts. 1990.The primary sequence of the EcoRII endonuclease and proper-ties of its fusions with b-galactosidase. J. Biol. Chem. 265:767-733.
4. Bhagwat, A. S., A. Sohail, and M. Lieb. 1988. A new geneinvolved in mismatch correction in Escherichia coli. Gene74:155-156.
5. Bhagwat, A. S., A. Sohail, and R. J. Roberts. 1986. Cloning andcharacterization of the dcm locus of Escherichia coli K-12. J.Bacteriol. 166:751-755.
6. Cattaneo, R. 1989. How 'hidden' reading frames are expressed.Trends Biochem. Sci. 14:165-167.
7. Coulondre, C., J. H. Miller, P. J. Farabaugh, and W. Gilbert.1978. Molecular basis of substitution hotspots in Escherichiacoli. Nature (London) 274:775-780.
8. Craigen, W. J., and C. T. Caskey. 1987. Translational frame-shifting: where will it stop? Cell 50:1-2.
9. Dzidic, S., and M. Radman. 1989. Genetic requirements forhyper-recombination by very short patch mismatch repair:involvement of Escherichia coli DNA polymerase I. Mol. Gen.Genet. 217:254-256.
10. Ehrlich, M., K. F. Norris, R. Y.-H. Wang, K. C. Kuo, and C. W.Gehrke. 1986. DNA cytosine methylation and heat-induceddeamination. Biosci. Rep. 6:387-393.
11. Guerola, N., J. L. Ingraham, and E. Cerda-Omedo. 1971.Induction of closely linked multiple mutations by nitrosoguani-dine. Nature (London) New Biol. 230:122-125.
12. Hanck, T., N. Gerwin, and H. Fritz. 1989. Nucleotide sequenceof the dcm locus of Escherichia coli K-12. Nucleic Acids Res.17:5844.
13. Harley, C. B., and R. P. Reynolds. 1987. Analysis of E. colipromoter sequences. Nucleic Acids Res. 15:2343-2361.
14. Jones, M., R. Wagner, and M. Radman. 1987. Mismatch repairand recombination in E. coli. Cell 50:621-626.
15. Jones, M., R. Wagner, and M. Radman. 1987. Mismatch repairof deaminated 5-methyl-cytosine. J. Mol. Biol. 194:155-159.
16. Kohara, Y., K. Akiyama, and K. Isono. 1987. The physical mapof the whole E. coli chromosome: application of a new strategyfor analysis and sorting of a large genomic library. Cell 50:495-508.
17. Kondoh, H., and H. Ozeki. 1976. Deletion and amber mutants offla loci in Escherichia coli K-12. Genetics 84:403-421.
18. Kozak, M. 1983. Comparison of initiation of protein synthesis inprocaryotes, eucaryotes, and organelles. Microbiol. Rev. 47:1-45.
19. Lieb, M. 1985. Recombination in the I repressor gene: evidencethat very short patch (VSP) mismatch correction restores a
specific sequence. Mol. Gen. Genet. 199:465-470.20. Lieb, M. 1987. Bacterial genes mutL, mutS, and dcm participate
in repair of mismatches at 5-Methylcytosine sites. J. Bacteriol.169:5241-5246.
21. Lieb, M., E. Alien, and D. Read. 1986. Very short patch repairin phage lambda: repair sites and length of repair tracts.Genetics 114:1041-1060.
22. Lieb, M., and A. S. Bhagwat. 1988. Very short patch mismatchrepair activity associated with gene dcm is not conferred by aplasmid coding for EcoRII methylase. J. Bacteriol. 170:4967-4968.
23. Lindahl, T. 1974. An N-glycosidase from Escherichia coli thatreleases free uracil from DNA containing deaminated cytosineresidues. Proc. Natl. Acad. Sci. USA 71:3649-3653.
24. Lindahl, T., and B. Nyberg. 1974. Heat-induced deamination ofcytosine residues in deoxyribonucleic acid. Biochemistry 13:3405-3410.
25. Lipman, D. J., and W. R. Pearson. 1985. Rapid and sensitiveprotein similarity searches. Science 227:1435-1441.
26. Maniatis, T., E. F. Fritsch, and J. Sambrook. 1982. Molecularcloning: a laboratory manual. Cold Spring Harbor Laboratory,Cold Spring Harbor, N.Y.
27. Marck, C. 1988. 'DNA Strider': a 'C' program for the fastanalysis ofDNA and protein sequences on the Apple Macintoshfamily of computers. Nucleic Acids Res. 16:1829-1836.
28. Marinus, M. G., and N. R. Morris. 1973. Isolation of deoxyri-bonucleic acid methylase mutants of Escherichia coli K-12. J.Bacteriol. 114:1143-1150.
29. May, M. S., and S. Hattman. 1975. Deoxyribonucleic acid-cytosine methylation by host- and plasmid-controlled enzymes.J. Bacteriol. 122:129-138.
30. Modrich, P. 1987. DNA mismatch correction. Annu. Rev.Biochem. 56:435-466.
31. Modrich, P. 1989. Methyl-directed DNA mismatch correction.J. Biol. Chem. 264:6597-660.
32. Pearson, W. R., and D. J. Lipman. 1988. Improved tools forbiological sequence analysis. Proc. Natl. Acad. Sci. USA 85:2444-2448.
33. Posfai, J., A. S. Bhagwat, G. Posfai, and R. J. Roberts. 1989.Predictive motifs derived from cytosine methyltransferases.Nucleic Acids Res. 17:2421-2435.
34. Sanger, F., S. Nicklen, and A. R. Coulson. 1977. DNA sequenc-ing with chain-terminating inhibitors. Proc. Natl. Acad. Sci.USA 74:5463-5467.
35. Schlagman, S., S. Hattman, M. S. May, and L. Berger. 1976. Invivo methylation by Escherichia coli K-12 mec+ deoxyribonu-cleic acid-cytosine methylase protects against in vitro cleavageby the RII restriction endonuclease (R * EcoRII). J. Bacteriol.126:990-996.
36. Sekine, Y., and E. Ohtsubo. 1989. Frameshifting is required forproduction of the transposase encoded by insertion sequence 1.Proc. Natl. Acad. Sci. USA 86:4609-4613.
37. Som, S., A. S. Bhagwat, and S. Friedman. 1987. Nucleotidesequence and expression of the gene encoding the EcoRIImodification enzyme. Nucleic Acids Res. 15:313-332.
38. Stormo, G. D. 1986. In W. S. Reznikoff and L. Gold, Maximiz-ing gene expression, p. 195-224. Butterworth Publishers, Stone-ham, Mass.
39. Yanisch-Perron, C., J. Viera, and J. Messing. 1985. ImprovedM13 phage cloning vectors and host strains: nucleotide se-quences of the M13mpl8 and pUC19 vectors. Gene 33:103-119.
40. Zell, R., and H. Fritz. 1987. DNA mismatch-repair in Esche-richia coli counteracting the hydrolytic deamination of 5-methyl-cytosine residues. EMBO J. 6:1809-1815.
VOL. 172, 1990
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/j
b on
06
Dec
embe
r 20
21 b
y 45
.183
.93.
160.




























