Embed Size (px)
Citation preview

Molecular Microbiolcgy (1995) 16(1), 45-55
system of Escherichia coli
Dzwokai Wla, ' David N. Cook,^ Wlarie Albert!,^NingG. Pon,^ Hiroshi Nikaido' '* and John E.'Department of Chemistry, University of Gatlfornia,Berkeley, California 94720. USA.^Division of Structural Biology. Lawrence BerkeleyLaboratory, Berkeley. California 94720. USA.'^Sferitech inc., 2525 Stanwell Drive, Suite 300, Concord,California 94520, USA.'^Department of Molecular and Cell Biology, University ofCalifornia, Berkeley, California 94720, USA.
Summary
Defined mutations of acrA or acrB (formerly acrE)genes increased the susceptibility of Escherichia colito a range of small inhibitor molecules. Deletion ofacrAB increased susceptibility to cephalothin andcephaloridine, but the permeability of these |i-lactamsacross the outer membrane was not increased. Thisfinding is inconsistent with the eariier hypothesisthat acrAB mutations increase drug susceptibility byincreasing the permeability of the outer membrane,and supports our model that acrAB codes for amulti-drug efflux pump. The natural environment ofan enteric bacterium such as E. coli is enriched inbile salts and fatty acids. An acrAB deletion mutantwas found to be hypersusceptible to bile salts and todecanoate. In addition, acrAB expression was ele-vated by growth in 5mM decanoate. These resuitssuggest that one major physiological function ofAcrAB is to protect E. coli against these and otherhydrophobic inhibitors. Transcription of acrAB isincreased by other stress conditions including 4%ethanol, 0.5 Wl NaCl, and stationary phase in Luria-Bertani medium. Finally, acrAB expression wasshown to be increased in mar (multiple-antibiotic-resistant) mutants.
Introduction
Mutations in the acrA locus are known to render Escheri-chia coli hypersusceptible to basic dyes, detergents andantibiotics (Nakamura, 1965; 1968), The observation that
Received 23 September t994; revised 30 November 1994; accepted13 December, 1994. 'For correspondence, Tel. (510) 642 2027; Fax(510)643 9290,
acrA mutants are hypersusceptibte to a broad spectrumof agents, which have different structures and differentintracellular targets, suggested a general role for theacrA locus tn determining the intrinsic drug resistance ofE. coll. One explanation is that acrA mutations increasethe permeability of the outer membrane of E. coli. Inseveral studies to date that have addressed this ques-tion, however, an unambiguous change has not beendetected in the chemical composition of the outer mem-brane of acrA strains (see Sukupolvi and Vaara, 1989),
We have previously cloned the acrA locus by comple-menting the drug-hypersusceptible phenotype of an acrAmutant, N43 (Ma efai, 1993a; also see Xu etai, 1993),Sequence analysis has revealed at least two genes,acrA and acrB (formerly acrE), which are located within asingle operon. Gene acrA appeared to encode a periptas-mic fipoprotein whose amino-terminus was anchored tothe inner membrane, and gene acrB appeared to encodean integral inner membrane protein with 12 transmem-brane segments. The predicted topologies of AcrA andAcrB were supported by the construction of phoA fusionsto acrA and acrB (Ma ef ai. 1993a). These predictionsgained further credence by the comparison with severalAcrAB homologues found in E. co//or Pseudomonas aeru-ginosa {Se\iier etai. 1993; Poole efai. 1993), Strain N43contained an \S2 element inserted near tine 5' end of acrA,and a plasmid containing only the acrA* allele was able tocomplement the drug hypersusceptibility ot N43, There-fore, AcrA is required for the intrinsic drug resistance ofE, coli. Based on operon organization and sequence com-parison with other transporter proteins, we have furthersuggested that AcrB contributes to the intrinsic drug resis-tance as a component of an energy-dependent drug effluxpump. Direct experimental evidence for the involvement ofAcrB has been lacking, however.
Initial experiments strongly suggested that AcrABpumped out drugs such as acriflavine (Ma et ai. 1993a).However, there was much residual efflux activity lefteven in the acrA mutant N43, presumably because of theexistence of other multi-drug efflux pumps with overlap-ping specificities. Consistent with this hypothesis, threeAcrAB homologues — AcrEF (formerly EnvCD). OrfAB,and AcrD — have now been identified in E. coli (for areview, see Ma et ai. 1994). Mutations in orfAB resultin drug hypersusceptibiiity (G- Storz, personal covnniuni-cation). Moreover, drug-hypersusceptibility of an acrAB
© 1995 Biackweil Science Ltd

46 a Ma et al.
mutant can be suppressed by mutations mapping close toacrEP, which presumably increase the expression ofacrEFgenes (Xu etai, 1993). In addition, evidence hasrecently been presented suggesting the existence ofmulti-drug efflux pumps of another class, I.e. EmrAB(Lomovskaya and Lewis, 1992).
The proposal that AcrAB encodes a drug-efflux pump hasbeen supported by several recent discoveries. First, byfunctional complementation, Poole ef ai (1993) havecloned a putative effiux system (the MexAB-OprK sys-tem) for the secretion of siderophores during iron starva-tion from P. aeruginosa. MexAB is homologous to AcrAB,and mutations in the genes encoding MexAB-OprK alsomake P. aeruginosa hypersusceptible to hydrophobic anti-biotics and inhibitors. Second, Dinh ef ai (1994) have pro-posed that AcrA belongs to the family of 'membranefusion proteins' (MFP), Many members of this family areknown to function in conjunction with inner membrane trans-porters to pump out peptides, proteins, carbohydrates ordrugs across the two membranes of Gram-negative bao-teria. EmrA is also a member of the MFP family.
Although AcrAB may be hypothesized to contribute tothe intrinsic drug resistance of E. coll, most of its knownsubstrates, such as antibiotics and cationic dyes, are notfound in the natural environment of E, coll. It seems likelythat the AcrAB efflux system may have evolved for otherpurposes. In this regard, it is noteworthy that the naturalenvironment of enterfc bacteria such as E. co//is enrichedin bile salts and fatty acids (Lenlner, 1981). Both bile saltsand fatty acids are hydrophobic compounds that inhibitthe growth of Gram-positive bacteria (Freese etai, 1973).However, Gram-negative bacteria such as E. coll areknown to be intrinsically resistant to bile salts and fattyacids. Traditionally, this resistance has primarily beenattributed to the oufer membrane, which is the majorbarrier for the entry of hydrophobic inhibitors into the cell(Sheu ef al., 1973). However, recent studies have shownthat the outer membrane of Gram-negative bacteria canonly slow down the influx of hydrophobic inhibitors, andthe equilibration time for many hydrophobic inhibitorsacross the outer membrane is much shorter than thedoubting time of the bacteria (Plesiat and Nikaido, 1992).As a result, Gram-negative bacteria need additionalmechanism(s), such as active efflux, for protection againstthese hydrophobic inhibitors.
We show here that AcrB, as well as AcrA, is necessaryfor the intrinsic resistance of E coli to hydrophobic anti-biotics and detergents. Our measurernent of the perme-ability of |ii-lactams across the outer membrane of anacrAB null mutant was inconsistent with the earlier hypo-thesis that acrAB mutations increase drug susceptibilityby increasing the permeability of the outer membraneand supports the more recent modei that acrAB encodesa drug-eftlux pump. Moreover, the acrAB null mutant is
hypersusceptible to both bile salts and decanoate (a CIOfatty acid), suggesting that one physiological function ofAcrAB is to protect E. coli against these naturally occur-ring hydrophobic inhibitors. Finally, we report that trarvscription of acrAB is increased not only by decanoatebut also by conditions of general stress, and in marR(multiple-antibiotic-resistant) mutants.
Results
Both AcrA and AcrB are required for the intrinsicdrug resistance of E. coii
In order to delineate the role of AcrB, we have constructedthree mutants (Fig. 1) and determined their drug suscep-tibilities (Table 1). Mutants J2M100 or J2M130 contairiedthe insertion of a 1.3 kb kanamycin-resistance gene car-tridge near the 5' end of acrB or acrA, respectively. Formutant JZM120, a large portion of acrAt3 was deletedand replaced by the same 1.3 kb gene cartridge. Asshown in Table 1, all three mutants are hypersusceptibieto novobiocin, erythromycin and SDS, when comparedwith their isogenic parent sirain (JC7623). These results
JC7&23acrA iurB
acrB JZM130
ggliaiH^SSi::;:;:!:::
ilcrA JZM too
Mscl
JZM 120
Fig, •!. The construction of acrAB mutants, twtutations of acrABwere tirst introduced into ptasmid pUCK151A, wtiich contained thewitd-tyiDe acrAB genes. Mutations were Ihen moved to E. coliJC7623 by tiomotocjous recombination (tor detaits, ^ee theExperimentai procedures). Strain JZM100 or J2M130 contains theinsertion ot •1,3 kb Tn903kan' gene cartridge at the Nae\ site ofacrB or the Msc\ site of acrA, respectivety. t=or JZtV!i20, theMsc\-Hpa] fragmenl of acrAB was deleted and replaced by the1,3kb Tr\903kan' gene cartridge.
C) 1995 Biackweil Science Ltd, Molecular fiiiicroblology. 16, 45-55

stress-induced efflux system ot Esctterichia coli 47
Tabie 1. Comparison of lhe drug susceptibilityin LB medium between the acrAB mutants andIheir isogenic wild lypes.
Compound
NovobiocirtEr/thromycinSDSSodium cholaleSodium taurodeoxychoiate
JC7623
64128
>10000
JZtVitOO
<2<2
<64
MtC (ll
JZM120
<2<2
<64
gml ') Foi':
JZM130
<2<2
<64
K4401
256256
>10000>10000
KZM120
<28
6420481024
indicate that AcrB, as welt as AcrA, is required for theintrinsic drug resistance of E. coll. We also transducedthe acfAB null mutation from JZM1S0 into K4401 orW4680. two E. coll K-12 strains with no or few knownmutations (Tabte 2), The transductants. named KZM120or WZM120, retained the drug hypersusceptibility.
Permeability of the outer membrane of the acrAB nullmutant
Null mutations of aciAB make E. coli slightly more sus-ceptible to J3-lactams. Minitnum inhibitory concentrations(MICs) of cephalothin and cephaloridine for KZM120were lower than those for K4401 (4 versus 8f.tgml ^ forcephalothin: 2 versus 4(.rgmr' for cephaloridine). Bothcephalothin and cephatoridine are amphiphitic [S-lactams
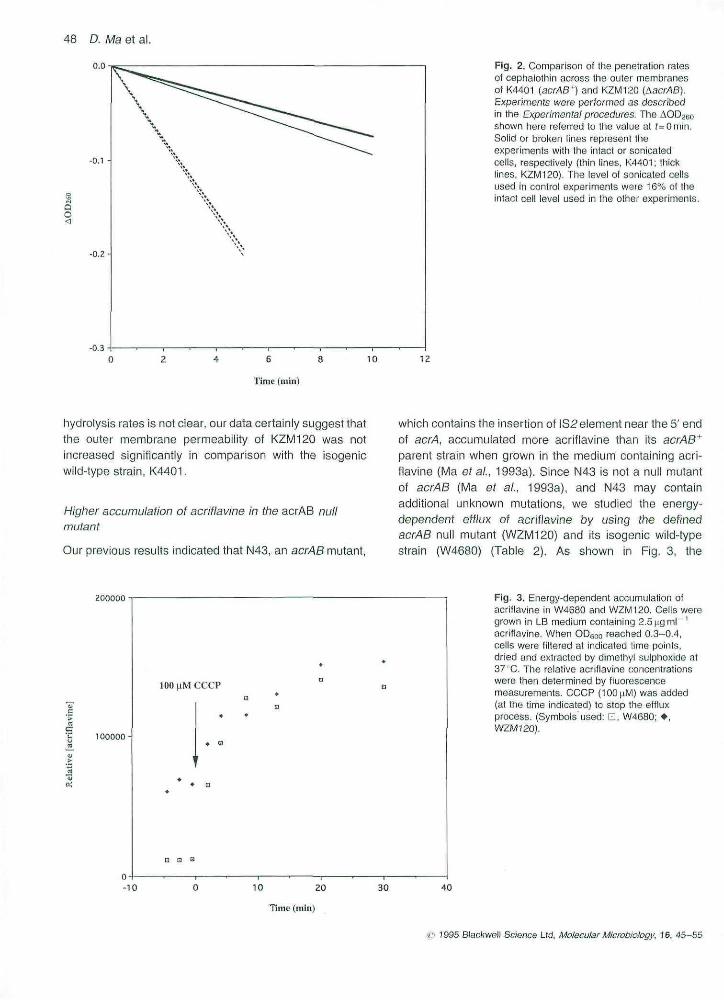
that are hydrolysed by common |3-lactamases. InpBR322-containing E. coll ceiis, the rate of their hydro-lysis, which is catalysed by the TEM p-lactamase encodedby pBR322, is limited by the rate of their penetration acrossthe outer membrane (Zimmermann and Rosselel, 1977).We can therefore measure the permeabilities of cephalo-thin and cephaloridine across the outer membrane byfollowing their hydr'olysis t>y intact celts. It was found thatthe rates of cephalothin hydrolysis were very similar inKZM120 and K4401 (Fig, 2). If anything, the AacrABmutant KZM120 appeared to hydiolyse cephalothinslightly more slowly than K4401, and this difference inhydrolysis rate disappeared when sonicated ceils wereused (Fig. 2), Similar slight differences were also observedwhen cephaloridine was used as the substrate. Althoughthe significance of the observed small differences in the
Table 2. Strains or piasmids.Strain/Plasmid Description
Strain
JC7623
JZtVilOOJZM 120JZM 130K4401KZM120GC4468JHC1069JHC1072
JHC1113
W4680WZM120
Piasmid
pUCK151ApZMlOO
pZM120
pZMI 30
pNN387
argE3 hisG4 leuB6\(gpt~proAj62 thr-1 tiv-tara 14 galK2 lacVI mil-1 xyi-t kdgK5itsx33 recB21 recC22 sbcB15 supE44rpsL31 rae
Same as JC7623; acrBv.Tn903kan'Same as JC7623; AacrAB and TnQOSkan'Same as JC7623; acrA::Tn903kan'No known mutation (Inr?)Same as K440t; AacrAB::Tn903kan'F AiacU1G9 rpsLSame as GC4468; cfxB1 zdd-2207v.Jn10kan'Same a.? GC4468; soxOI zdd-
Same as GC4468; marR1 zdd-2207v.Tn10kan'
F - MacZ39 rpsL45 or rpsL 110 me!B4Same as W4680; A.acrABv.Tn903i<an'
Kushner e/a/. (1971)
This woikThis workThis workClowes and Hayes (1968}This workGreenberg e! al. (1990)Greenberg el ai (1991)Greenberg el al. (1991)
GieentDerg etai. (1991)
Cook and Lederbcrg (1962)This work
pUC19 derivative oontaining tiie acrAB genes Ma et al. (1993a)1,3 kb Tn903kan' inserted at Msc\ of This work
pUCK151A1,3kb Tr}903kan' inserted to /Wscl-/-/pat-
restricted ptJCK151A1,3 kb Tn903kan' inserted al Nae\ of
PUCK151AA single-copy ptasmid containing the
promoterless lacZY and the Tn9 cam' genecartridge
pNW387 derivative comaining 2,2 kb DNAupstream of acrA
This work
This work
Ettedge and Davis (1989)
This wotK
1995 Blacttwell Science Ltd, Molecular Microbiology. 16, 45-56
48 D, Ma et ai
0.0
-0.1 -
-0,2-
-0,3 4
Fig. 2. Comparison of the penetration ratesof cephaiothin across the outer membranesof K4401 (acres ' ) and KZM120 (AacrAB).Experiments were performed as describedin the Experimental procedures. The .AOD^yoshown bere referred to the value at f=Omin,Solid or broken lines represent theexperiments with the intaci or sonicatedcells, respectively (Ihin lines. K4401; thicklines. kZIVt120), The levei of sonicated cellsused in control experiments were 16% of theintact ceil level used in the other experiments.
^ (iitin)
hydrotysis rates is not clear, our data certainly suggest thatthe outer membrane permeability of KZM120 was notincreased significantly in comparison with the isogenicwild-type strain, K4401.
Higher accumulation of acriftavine In the acrAB nullmutanf
Our previous results indicated that N43, an acrAB ritutant.
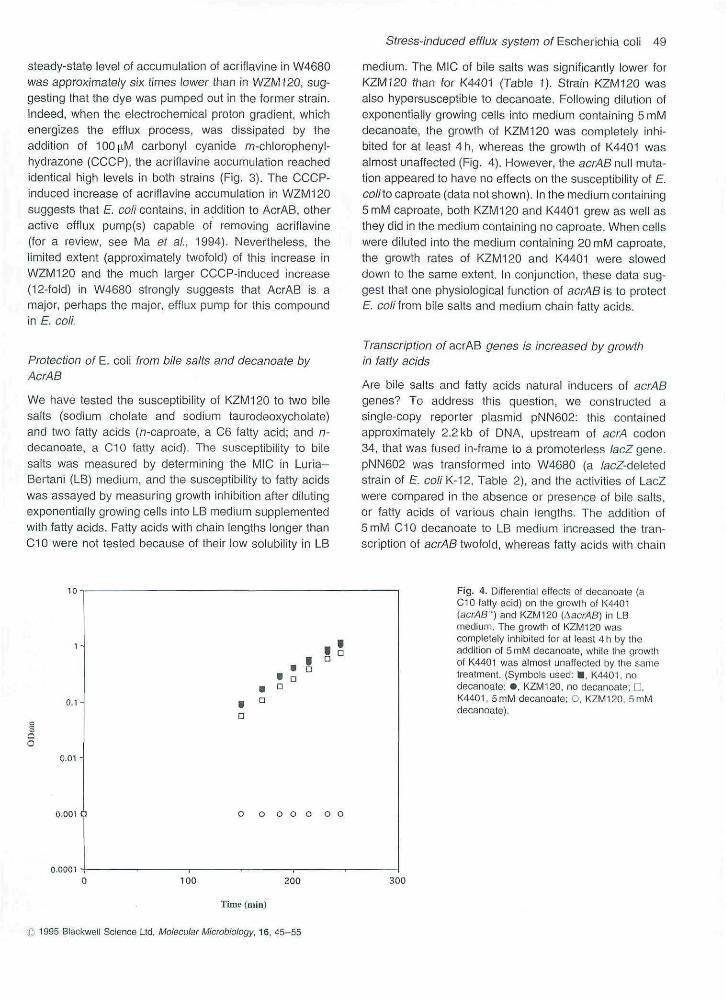
which contains the insertion of IS2 element near the 5'endof acrA, accumulated more acriflavine than its acrAB''parent strain when grown in the medium containing acri-flavine (Ma et ai, 1993a), Since N43 is not a null mutantof acrAB (Ma e ai, 1993a), and N43 may containadditional unknown mutatioris, we studied the energy-dependent efflux of acriflavine by using the definedacrAB null mutant (WZM120) and its isogenic wiid-typestrain (W4680) (Table 2), As shown in Fig. 3, the
-10
Fig. 3. Energy-dependent accumulation ofacriflavine in W4680 and WZW/120. Cetls weregrown in LB medium containing 2,5|i,gmt~'acriftavine. When ODgoo reached 0,3-0,4,cetts were tiltered at indicated lime points,dried and extracted by dimethyl sulphoxide at37 C, Ttie relative acritlavine concentrationswere then determined by fiuortiscencemeasurements. CCCP (100 jiM) was added(at the time indicated) to stop the etfluxprocess. (Symbols used: P, W46aO; • ,WZM12a).
Time (niin)
'_' 1995 Bl£ici(well Science Ltd, Molecular Microbiology, 16, 45-55
stress-induced efflux sysfem of Escherichia coti 49
steady-state level of accumulation of acrifiavine in W4680was approximately six times lower than in WZM120, sug-gesting that the dye was pumped out in the former strain.Indeed, when the electrochemical proton gradient, whichenergizes ihe efflux process, was dissipated by theaddition of 100|.iM carbonyl cyanide m-chlorophenyl-hydrazone (CCCP), the acriflavine accumulation reachedidentical high levels in both strains (Fig. 3). The CCCP-induced increase of aoriflavine accumulation in WZM120suggests that E. co//contains, in addition to AcrAB, otheractive efflux purTip(s) capable of removing acriflavine(for a review, see Ma ef ai, 1994). Nevertheless, thelimited extent (approximately twofold) of this increase inWZM120 and the much larger CCCP-induced increase(12-fold) in W4680 strongly suggests that AcrAB is amajor, perhaps the major, efflux pump for this compoundin H, coll.
Protection of E. coii from bile salts and decanoate byAcrAB
We have tested the susceptibility of KZM120 to two bilesalts (sodium cholate and sodium taurodeoxychoiate)and two fatty acids (n-caproate, a C6 fatty acid; and n-decanoate, a CIO fatty acid). The susceptibility to bilesalts was measured by determining the MIC in Luria-Bertani (LB) medium, and the susceptibility to fatty acidswas assayed by measuring growth inhibition after dilutingexponentially growing cells into LB medium supplementedwith fatty acids. Fatty acids with chain lengths longer thanC10 were not tested because of their low solubiiity in LB
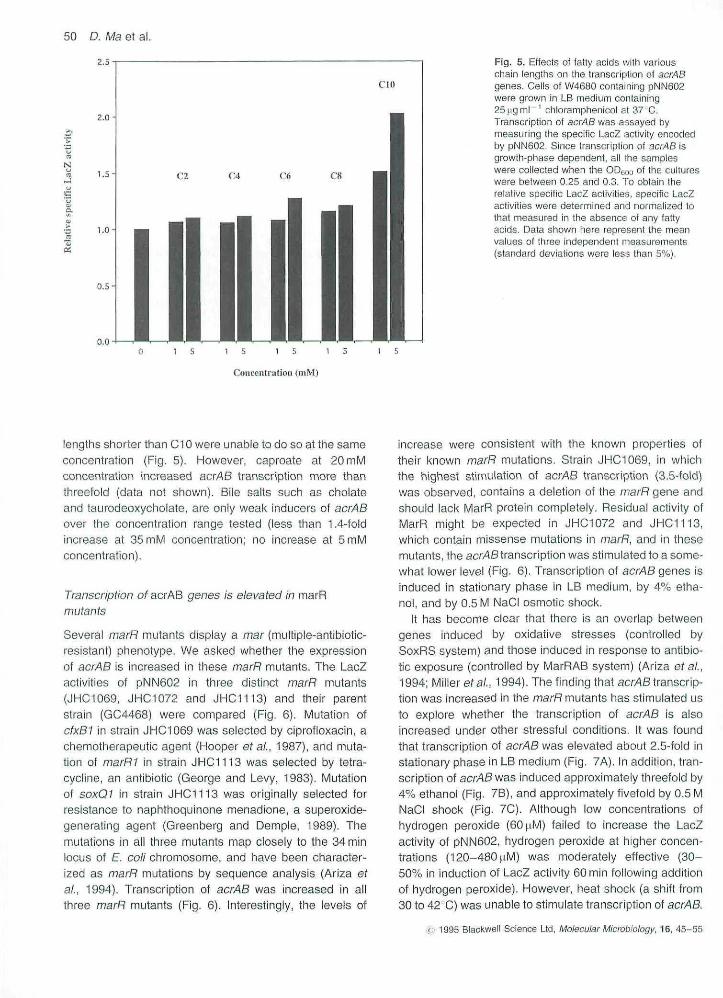
medium. The MtC of bile salts was significantly lower forKZM120 than for K4401 (Tabie 1), Strain KZM120 wasalso hypersusoeptible to decanoate. Following dilution ofexponentially growing cells into medium containing 5mMdecanoate, the growth of KZM120 was completely inhi-bited for at least 4 h, whereas lhe growth of K4401 wasalmost unaffected (Fig. 4). However, the acrAB null muta-tion appeared to have no effects on the susceptibility of Eco//to caproate (data not shown). In the medium containing5mM caproate, both KZM120 and K4401 grew as well asthey did in the medium containing no caproate. When cellswere diluted into the medium containing 20 mM caproate,the growth rates of KZM120 and K4401 were sloweddown to the same extent. In conjunction, these data sug-gest that one physiological function of acrAB is to protectE. CO//from bile salts and medium chain fatty acids.
Transcription of acrAB genes is Increased by growthin fatly acids
Are bile salts and fatty acids riatural inducers of acrABgenes? To address this question, we coristructed asingle-copy reporter plasmid pNN602: this containedapproximatety 2.2 kb of DNA, upstream of acrA codon34, that was fused in-frame to a promoterless tacZ gene.pNN602 was transformed into W4680 (a /acZ-deletedstrain of E. coti K-12, Table 2). and the activities of LacZwere compared in the absence or presence oi bile salts,or fatty acids of various chain lengths. The addition of5mM CIO decanoate to LB medium increased the tran-scription of acrAB twofold, whereas fatty acids with chain
o . t -
0.01 -
o.oot p o o o o o o o
0.0001 -^0 100 200
Time (min)
C- t995 Biackweil Science Ltd, Molecular Microbioiogy. 16, 45-55
300
Fig. 4. Differential effects ot decanoate (aC10 fatty acid) on Ihe growth of K4'101{acrAB') and KZM120 (.\acrAB) in LBmedium. The growth of KZM120 wascompletely inhibited for al least 411 by theaddition of 5mM decanoate, while the growthof K4401 was almost unaffected by the sametreatment. (Symbols used: • , K4401, nodecanoate; • . KZM120, no decanoate; • ,K440t, 5mM decanoate; O, KZM120, 5mMdecanoate).
50 D. Ma et al
2.5
2.0 •
5 1 5 1 5
Conceiilralion (niMJ
Fig. 5. bffecis of fatly acids with variouschain lengths on lhe transcription of acrABgenes. Cells ot W4fi80 containing pNN602were grown in LB medium containing25|.igmr ' chtoramphenicol at 37-C,Transcription ot acrAB was assayed bymeasuring the specific LacZ activity encodedby pNN602, Since transcription of aciAB isgrowth-phase dependent, all ihe sampleswere collected when the ODt;^^ of the cultureswere between 0,25 and 0,3, To obtain Iheretative specific LacZ activitie.s, specific LacZactivities were determined and normalised tothat measured in the absence ot any fattyacids. Data siiown here represent the meanvatues of three independent measurements(standard deviations were less than 5%).
lengths shorter than C10 were unable to do so at the sameconcentration (Fig. 5). However, caproate at 20 mMconcentration increased acrAB transcription more thanthreefold (data not shown). Bile salts such as oholateand taurodeoxychoiate, ate only weak inducers of acrABover the concentration range tested (less than 1.4-foldincrease at 35 mM concentration; no increase at 5mMconcentration).
Tianscription of acrAB genes is elevated in tTiarRmutants
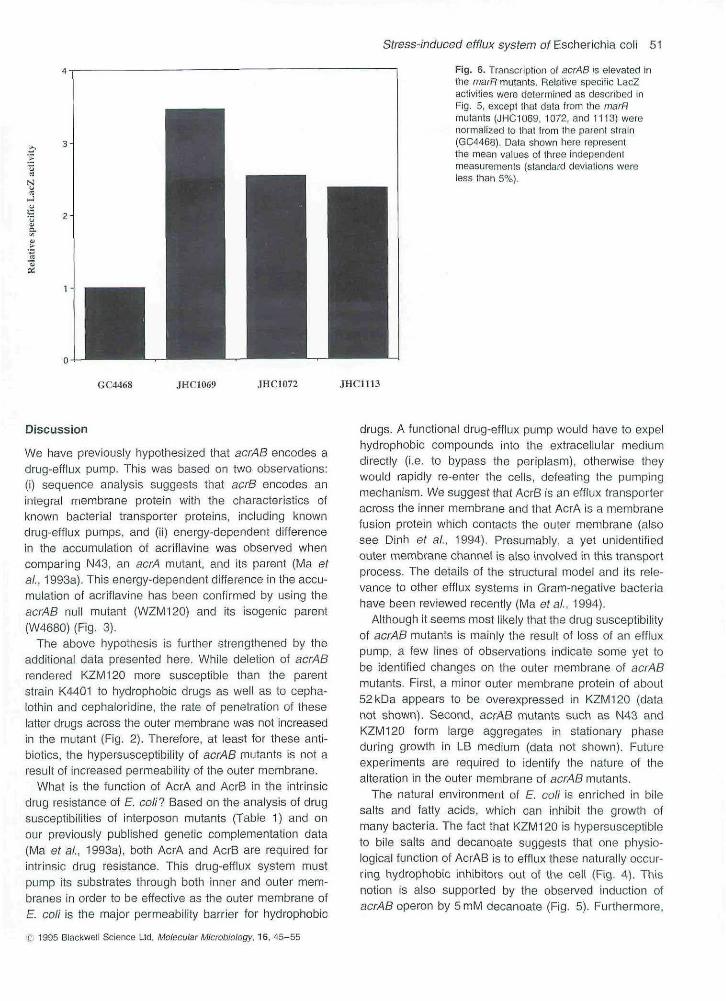
Several marR mutants display a mar (multipie-antibiotio-resistant) phenotype. We asked whether the expressionof acrAB is increased in these marR mutants. The LacZactivities of pNN602 in three distinct marR mutants(JHC1069, JHC1072 and JHC1113) and their parentstrain (GC4468) were compared (Fig. 6). Mutation ofcfxBI in strain JHC1069 was selected by ciprofloxacin, achemotherapeutio agent (Hooper et ai. 1987), and muta-tion of marR1 in strain JHC1113 was selected by tetra-cyciine, an antibiotic (George and Levy, 1983). Mutationof soxQ1 in strain JHC1113 was originally selected forresistance to naphthoquinone trtenadione, a superoxide-generating agent (Greenberg and Demple, 1989), Themutations in all three mutants map closely to the 34 minlocus of E. coll chromosome, and have been character-ized as marR iTtutations by sequence analysis (Ariza efai, 1994). Transcription of acrAB was increased in ailthree marR mutants (Fig, 6), Interestingly, the levels of
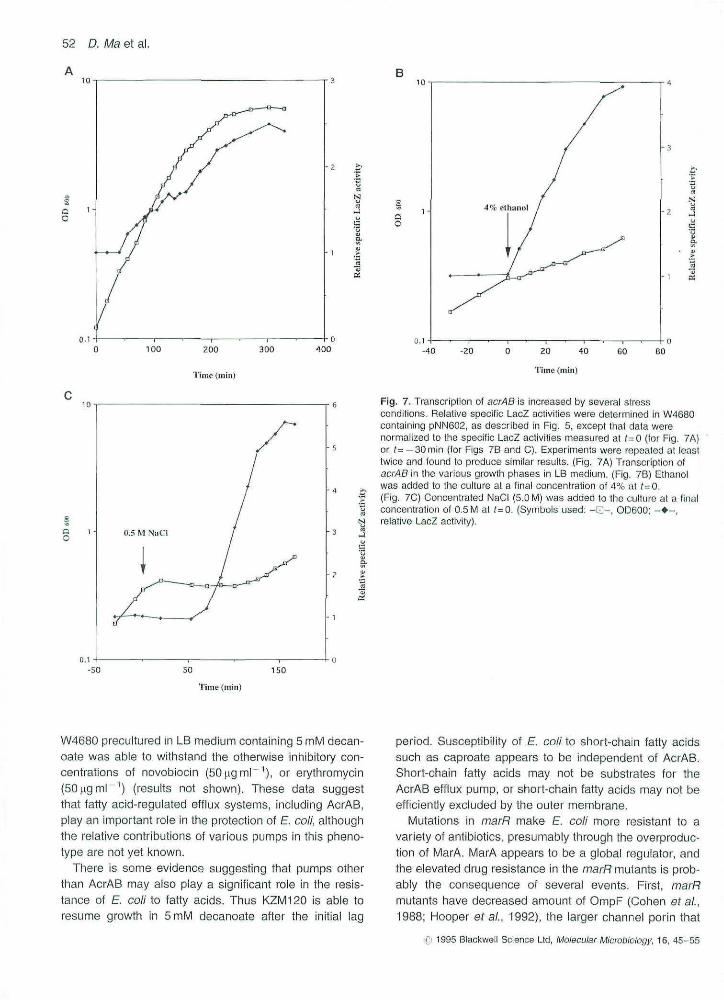
increase were consistent with the known properties oftheir known marR mutations. Strain JHC1069, in whichthe highest stimulation of acrAB transcription (3.5-fold)was observed, contains a deletion of the marR gene andshould iack MarR protein completely. Residual activity ofMarR might be expected in JHC1072 and JHC1113,which contain missense mutations in marR, and in thesemutants, the acrAS transcription was stimulated to a some-what lower level (Fig. 6). Transcription of acrAB genes isinduced in stationaiy phase in LB medium, by 4% etha-nol, and by 0.5 M NaCI osmotic shock.
It has become clear that there is an overlap betweengenes induced by oxidative stresses (controlled bySoxRS system) and those induced in response to antibio-tic exposure (controlled by MarRAB system) (Ariza efai,1994; Miller e/^/,, 1994). The finding that acMS transcrip-tion was increased in the /nar/? mutants has stimulated usto explore whether the transcription of eicrAB is alsoincreased under other stressful conditions. It was foundthat transcription of acrAB was elevated about 2.5-fold instationary phase in LB mediurn (Fig. 7A). In addition, tran-scription of acr/ABwas induced approximately threefold by4% ethanol (Fig, 7B), and approximately fivefold by 0,5 UNaCI shock (Fig, 7C). Although iow concentrations othydrogen peroxide (60).tM) failed to Increase the LacZactivity of pNN602, hydrogen peroxide at higher concen-trations (120-480 pM) was moderately effective (30-50% in induction of LacZ activity 60 min following additionof hydrogen peroxide). However, heat shock (a shift from30 to 42 C) was unable to stimulate transcription of acrAB.
•I:. 1995 Blackwell Science Ltd, Molecular Microbiotogy. 16, 45-55
stress-induced efilux system of Escherichia coli 51
Fig, 6. Transcription of acrAB is elevated inthe marR mutants. Relative specific LacZactivities were determined as described inFig, 5, except that data from the rnarRmutants (JtHC1069, 1072, and 1113) werenormalised to that from the parent strain(GC4468), Data shown here representthe mean vatues of three independentmeasurements (standard deviations weretess than 5%),
GC44fi8 JHC1069 JIICiO72
Discussion
We have previousty hypothesized that acrAB encodes adrug-efflux pump. This was based on two observations:(I) sequence analysis suggests that acrB encodes anintegral membrane protein with the characteristics ofknown bacterial transporter proteins, including knowndrug-efftux pumps, and (ii) energy-dependent differencein the accumulation of acrifiavine was observed whencomparing N43, an acrA mutant, and its parent (Ma etai, 1993a). This energy-dependent difference in the accu-mulation of acriflavine has been confirmed by using theacrAB null mutant (WZM120) and its isogenic parent(W4680) (Fig. 3),
The above hypothesis is further strengthened by theadditional data presented here. While deletion of acrABrendered KZM120 more susceptible than the parentstrain K4401 to hydrophobic drugs as well as to cepha-tothin and cephaloridine, the rate of penetration of thesetatter dtugs across the outer membrane was not increasedin the mutant (Fig, 2), Therefore, at teast for these anti-biotics, the hypersusceptibility of acrAB miitants is not aresult of increased pertTieability of the outer merTibrane,
What is the function of AcrA and AcrB in the intrinsicdrug resistance of E. coll? Based on the analysis of drugsusceptibilities of interposon mutants (Table 1) and onour previously published genetic complementation data(Ma et ai, 1993a), both AcrA and AcrB are required forintrinsic drug resistance. This dtug-eff1ux system mustpump its substrates through both inner and outer mem-branes in order to be effective as the outer membrane ofE. coli is the major permeability barrier for hydrophobic
drugs, A functional drug-effiux pump would have to expelhydrophobic compounds into the extracellular mediumdirectly (i,e. to bypass the periplasm), otherwise theywould rapidly re-enter the cells, defeating lhe pumpingmechanism. We suggest that AcrB Is an efffux transporteracross the inner merTibrane and that AcrA is a tiiembranefusion protein which contacts the outer membrane (alsosee Dinh ef ai, 1994). Presumably, a yet unidentifiedouter membrane channel is also involved in this transportprocess. The details of the structural model and its rele-vance to other effiux systems in Gram-negative bacteriahave been reviewed recently (Ma etai, 1994),
Although it seems most likeiy that the drug susceptibilityof acrAB mutants is mainly the result of loss of an effluxpurTip, a few lines of observations indicate sorTie yet tobe identified changes on the outer membrane of acrABmutants. First, a minor outer membrane protein of about52 kDa appears to be overexpressed in KZM120 (datanot shown). Second, acrAB mutants such as N43 andKZM120 form targe aggregates in stationaiy phaseduring growth in LB mediutTi (data not shown). Futureexperiments are required to identify the nature of thealteration in the outer membrane of acrAB mutants.
The natural environment of E. coli is enriched in bilesalts and fatty acids, which can inhibit the growth ofmany bacteria. The fact that KZM120 is hypersusceptibleto bile salts and decanoate suggests that one physio-logical function of AcrAB is to efflux these naturally occur-ring hydrophobic inhibitors out of the cell (Fig, 4). Thisnotion is also supported by the obsewed induotion otacrAB operon by 5mM decanoate (Fig, 5), Furthermore,
•(:• 1995 Biackweil SciencG Ltd. Molecular Micrabioiogy. 16, 45-55
-40 80
Fig. 7. Transcription of acrAB is increased by several stressconditions. Relative specific LacZ activities were determined in W4680containing pNN602, as described in Fig, 5, except that data werenormalized to ihe specific LacZ activities measured at f=0 (tor Fig, 7A)or (= - 3 0 min (tor Figs 7B and C), Experiments were repeated at ieasttwice and tound to produce similar results. (Fig. ?A) Transcription ofacrAB in the various growth phases in LB medium, (Fig, 7B) Ethanotwas added to the culture at a final concentration of 4% at (=0.(Fig, 7C) Concentrafed NaCI (5.0 M) was added to lhe cuiture at a finalconcentration of 0.5 M at ; -G . (Symbols used: - Q - , OD600; - • - ,relative LacZ activity).
-SO
linii; (min)
W4680 precultured in LB medium containing 5 mM decan-oate was able to withstand the otherwise inhibitory con-centrations of novobiocin (50|.igmr^), or erythromycin(50Mgmi ') (results not shown). These data suggestthat fatty acid-regulated efflux systerTis, including AcrAB,play an important role in the protection of £, coll, althoughthe relative contributions of various pumps in this pheno-type are not yet known.
There is some evidence suggesting that purrtps otherthan AcrAB may also play a significant role in the resis-tance of E. coti to fatty acids. Thus KZM120 is able toresume growth in 5mM decanoate after the initial lag
period. Susceptibility of E, coli to short-chain fatty acidssuch as caproate appear's to be independent of AcrAB.Short-chain fatty acids may not be substrates for theAcrAB efflux pump, or short-ohain fatty acids may not beefficiently excluded by the outer membrane.
Mutations in marR make E. coti more resistant Eo avariety of antibiotics, presumably through the overpr-oduc-tion of MarA. MarA appears fo be a global regulator, andthe elevated drug resistance in the marR mutants Is prob-ably the consequence of several events. First, marRmutants have decreased amount of OmpF (Cohen ef ai,1988; Hooper ef a/., 1992), the larger channel porin that
•C-. 1995 Blackwell Science Ltd, Molecular Microbiology. 16, 45-55

Sfress-induced efflux system of Escherichia coli 53
facilitates the passage of some antibiotics across the outermembrane (Nikaido and Vaara, 1985), Second, marRmutants seem to have enhanced ability to pumpout antibiotics (McMurry et ai, 1994). Our finding that tran-scription of acrAB is increased in the marR mutants(Fig, 6) thus identifies one such mar/^-regulated effluxpump, the identity of which has remained unknown sincethe discovery of marR mutants in the early 1980s(George and Levy, 1983). At this stage, the mechanismof the regulation of acrAB by MarR is unclear: While tran-scription of the mar operon is strongly induced by anti-biotics such as tetracyciine and chloramphenicol whenassayed by Northern blot (Hachier et ai, 1991; Cohen efai, 1993), our studies using acrAv.lacZ fusion did notreveal any significant increase in the transcription ofacrAB upon exposure to a wide range of totracycline con-centrations (data not shown). Interpretation of these data iscomplicated by the fact that tetracycline inhibits translationof mRNA which would reduce the production of LacZprotein in our experiments. Northern biot analysis mayhelp to clarify this question. Transcription of acrAB isalso subject to the regulation by another gene product,AcrR. The gene for AcrR is located upstream of acrAB,and is transcribed in the opposite direction. Pan andSpratt (1994) have pointed out that the sequence of AcrRsuggests that it belongs to a family of bacterial transcrip-tional regulators. Our recent studies have shown thatAcrR is a repressor of the acrAB operon (Ma ef ai, inpreparation). Understanding these overlapping regulatorycircuits will be a challenge.
Several stress conditions (i.e, 4% ethanol shock, 0.5 MNaCI osmotic shock and the stationary phase in LBmedium) also induced transcription of aorAB (Fig. 7A-C). At least three AcrAB homologues, including AcrEF,OrfAB, and AcrD, have been identified in E coli Althoughexact functions of these homologues are not known, pre-liminary studies indicate that their expression may bestress-regulated (for a review, see Ma ef ai, 1994).Furthermore, genetic analysis suggests a functional over-lap between AcrAB and AcrEF (Xu et ai. 1993). This familyof efflux proteins may therefore contribute in a verygeneral way to the survival of E. coll under stress condi-tions. The presence of multiple efilux systems with over-lapping specificities may allow E. coli to respond moreprecisely to environmental changes through differ'entglobal regulatory circuits. Cross-resistance to stress is awell-known phenomenon in E coll. and certain genesbelong to more than one global regulon. For example,stationary phase E. coli cultures are more resistant toosmotic shock, heat shook and hydrogen peroxide expo-sure (Jenkins efai, 1988; 1991), Mutants of mar/? displayelevated antioxidant defences in addition to increasedantibiotic resistance (Ariza et ai, 1994: Miller ef aL,1994). Gene micF, encoding a regulatory antisense RNA
affecting the expression of ompP (Mizuno ef ai, 1984;Andersen and Delihas, 1990), is an element of at leastfour distinct regulons (reviewed by Demple, 1991). Genezwf, encoding glucose-6-phosphate dehydrogenase, isregulated by both SoxRS (Greenberg et ai. 1990), aprimary regulator responding to superoxides, and MarA,a primary regulator responding to antibiotics (Greenberget aL, 1991). Our study here suggests that aciAB alsobelongs to this group of genes that are regulated by mul-tiple signals and pathways.
Experimental procedures
Bacterial strains and growth condifions
The E. coli strains used in this study were all derivatives ofK-12, and ar'e listed in Table 2. Alt bacteriat ceils were grownat 37' C with shaking at 200 r,p,m, in LB medium (containing10g f " ' of Bacto Tiyptone, 5 g l " ' of Bacto Yeast Extract,and l O g l " ' of NaCl) unless indicated otherwise. The anti-biotics streptomycin (lOOMQml"^). kanamycin (25|,igmi '),ampicillin ('lOOi.igmr ^), and chioramphenicot (25ngrTit ')were used for selective media.
Construction of piasmids
Piasmids used in this study are iisted in Table 2, Theconstruction of pUCK15iA has been described previously(Ma ef ai. 1993a). For pZtVllOO or pZM130, fhe 1,3kbEcoRI fragment from pUC4K (Phartnacia Inc) was end-filtedby Ktenow fragmetit and inserted Into the Nael site (nearthe 5' end of acrB) or the Msc\ site (near the 5' end of aciA)of pUGKISIA, respectivety. The 1.3 kb EcoRI fragmentcontains the kanamycin-resistance gene cartridge fromTn903. For pZM120, the 3 kb Msc\-Hpa\ fragment wasdeleted from pUCK151A and reptaced by the above 1,3kbkanamycin-resistance gene cartridge. To construct ptMN602,a two-step cloning strategy was adopted. The first step wasthe insertion of a 2,2kb Msc\ fragment from pBR151B (Maef a/., 1993a) into the SnaBi site of pDC401 to makepDC602, The 2,2 kb MscI fragment contains the regionupstream of the 34fh codon of the acrA gene, PlasmidpDC401 is a muitipte-copy-number reporter vector whichcontains the promoterless tacZ and the termination signalsfor both transcription and transtation (Ma efai., 1993b), Thesecond step was to move the acrAB promoter region frompDC602 to pNN387, a single-copy-number reporter vectorwhich contains the promoterless lacZY genes (Eltedge andDavis, 1989), Ptasmid pDC602 was restricted with BamH\,end-filied with Ktenow fragment and restricted again withSsfl, The resulting 4.2 kb DNA fragment, which containedthe acrAB promoter region and the 5' portion of lacZ. wasisotated by get etectrophoiesis, Plasmid pNN387 wasrestricted with Hind\\\, end-fitled with Klenow fragment andrestricted again with Ssfl. The resulting 11kb DNA frag-ment, which contains the 3' portion of lacZ and the repticon,was isotated by get electrophoresis. Piasmid pNN602 wasthen made by ligating the 4,2 kb fragment from pDC602 intothe 11 kb fragment from pNN387.
>f;". 1995 Biackweil Science Ltd, Molecular Microbiology, 16, 45-55

54 D. Ma et al.
Mufagenesls of aoiAB genes by homologous
recombination
Plasmid pZM100 was restricted by Dra\ (cuts within aciA) andHpa\ (cuts within acrB). Piasmids pZM120 and pZM130 wererestricted by Dra! (cuts within acrA) and Xbal (cuts within thepotytinker of pUCK151A), Restricted ptasmid DNA wasprecipitated by ethanot, resuspended in water and used totransform competent JC7623 cetts, which were preparedwith ice-cotd CaCt^ as previousty described (Satnbrook efai, 1989), tHomotogous recombinants were setected andpurified on LB plates containing 25 | . igmr ' kanamycin.When necessary. acr,4S mutations were transduced intoK4401 using P^clr100 phage according to the protocoldescribed by Miller (1992). All insertions and deletions wereconfirmed by PCR amptification of genotnic DNA.
described by Miller (1992) with the following modifications.Samples, which contained the equivalent amount of cells as1 mi of the cutture at an CDROO of 0.3, were r'emoved from37 C and stored on ice before use. Celts were spun downand resuspended in 0,5 mt of Z buffer (Mitter, 1992), Totuene(20|.il) was then added into each resuspended sample,which was then vortexed for 10 s. Samples were teft in thesafety cabinet at room temperature for 45 min to allowevaporation of toluene. For the LacZ assay, 100|.il of4 m g m t " ' ONPG (o-nitrophenyl (i-n-gatactopyranoside) wasadded after the samples were preincubated at 28 C for5 tnin. Typically, the reaction was allowed t:o proceed for40min and stopped by the addition of 0,25mt of I MNa2CO3. After centrifugation, the absorptions of the super-natants at 420 nm and 550 nm were measured, and LacZactivities calculated.
Drug susceptibility measurements
The MICs of antibiotics and detergents were determined byserial twofold ditutions in LB medium using 5 x 10" exponen-tial phase cetts per mt as inocuta, Cett growth was measuredat ODeoo after 18h incubation on a shaker (200r.p,m.) at37 C, An ODc5on of less than 0,05 was considered to be nega-tive. Ati drugs used in this experiment were bought from SigmaChemical Co.
Measurement of the permeability of ll-lactams acrossthe outer membrane
Strains transformed with pBR322 were grown overnight in LBmedium containing 50Mgmt"^ ampicittin and 5mM MgCI?, A1:100 dilution was made into 30 mt of this media, prewarmedto 37 C. After the ODeoo reached 1.0, 25 mt of cuiture wasrernoved. Att the fottowing procedures were done at roomtemperature, Cetls were spun down, washed in 25 mt cf10ml\/l sodium phosphate buffer (pH6.0) containing 5mMMgCI^ and resuspended in the same buffer at a density of6mg of dry weight cetls per ml. An 18,75(il portion (equiva-lent to 0.15 mg dry weight) of the above celt suspension wasthen added into 481.25|.it of the buffer containing 1 mM fi-lactam under investigation. The mixture was immediatetytransferred into a cuvette with a 1 mm light path, and theabsorption at 260 nm was recorded continuousty (Nikaido efai. 1983). Negtigibte amounts of [Vtactamase were reteasedinto the extracettuiar medium during the above treatments.As a control, the hydrolysis of j3-lactams was measured froma sonicated cell suspension.
Energy-dependent accumulafion of acriftavine
The procedures were the same as those described previously(Ma etai. 1993a) except that the relative amount of acriftavinereiafned in ceils was measured by exciting samples al: 468 nmand monitoring emission at 499 nm with a spectrofluorometer(Fluorolog #1909, Spex Industries, Inc.).
Beta-galactosidase assays
Beta-galactosidase (LacZ) activity was assayed essentially as
Note added in proof
The strain listed as K4401 in this paper corresponds tc thewitd-type K-12 strain; K4401 is a strain number used intern-atty by the F. co//Genetic Stock Center,
Acknowledgements
We thank B. Dempfe for providing £, co//strains (JIHC1069,JHC1072, atid JHC1113) and C. Lynch for technical assis-tance. This work was supported by gr'ants FD 8R1 GM41911A-03-NF-03/92 and At-09644 from the Nationat tnsti-tules of tHealth, and by the tJS Department of Energy underContract DE-AC03-76SF00098.
References
Andersen, J,, and Delihas, N. (1990) micP RNA binds to 5'end of o/npFmRNA and to a protein from Escherichia coli.Biochemlstiy 29: 9249-9256.
Ariza, R,R., Cohen, S.P., Bachhawat, N,, Levy, S,B,, andDemple, B, (1994) Repr'essor mutations in the marRABoperon that activate oxidative stress genes and muttlpteantibiotic resistance in Escherichia coli J Bacteriot 176:143-148.
Cohen, S,P,, McMurray, L,rvl,, and Levy, S,B, (1988) matAlocus causes decreased expression of ompF porin inmultiple-antibiotic-resistant (mar) mutants of Escherichiacoli J Bacferiol 170: 5416-5422,
Cohen, S.P,, Hachier, H,, and Levy, S,B, (1993) Genetic andfunctional analysis of the multiple antibiotic resistance(mar) locus in Escherichia coli J Baeteriol 175: 1484-1492.
Clowes, R.C, and Hayes, W, (1968) Experiments in MicrobialGenetics. Oxfotd: Biackweit Scientific Pubticaticns.
Cook. A,, and Lederberg, J. (1962) Recombin;3tion studies oftactose nonfermenting mutants of Escherichia coli K-12.Genef/cs47: 1335-1353.
Dempte, B, (1991) Regulation of bacterial oxidative stressgenes, Annu Rev Ganet 25: 315-337.
Dinh, T,, Paulsen, t,T,, Saier, M.H. (1994) A family ofextracytoplasmic proteins ihat allow transport of largemotecules across the outer membranes of Gram-negativebacteria. J Baeteriol 176; 3825-3831,
1995 Blackwell Science Ltd, Molecular Microbioiogy, 16. 45-55

SfresS'induced efffux system of Escherichia coli 55
Eiledge, S.J,, and Davis, R,W. (1989) Position and densityeffects on repression by stationary and mobile DNA-binding proteins. Genes Dev 3: 185-197.
Freese. E., Sheu, C.W., and Galliers, E, (1973) Function oflipophilic acids as antimicrobiat food additives. Nature 241:321-325,
Geor'ge, A.M., and Levy, S.B, (1983) Gene in the majorcctransduction gap of the Escherichia coli K-12 linkagemap r-equired for the expression of chromosomal resis-tance to telracycline and other antibiolics. J Sacferio/155-.541-548,
Greenberg, J.T., and Demple, B, (1989) A global r'esponseInduced in Escherichia coli by redox-cycfing agentsoverlaps with thaf induced by peroxide stress, J Baeteriol171: 3933-3939.
Greenberg, J.T., Monach, P., Chou, J.H., Josephy, P.D., andDemple, B, (1990) Positive control of a giobal antioxidantdefense regulon activated by superoxide-generatingagents in Escherichia coll. Proc Naft Acad Sci USA 87:6181-6185.
Greenberg. J.T,, Chou, J,H,, Monach, P,, and Demple, B.(1991) Activation ot oxidative stress genes by mutations atthe soxQ/cfxB/marA tocLis of Escherichia coli. J Baeteriol173: 4433-4439,
Hachier, H., Cohen, S,P., and Levy, S.B. (1991) marA. aregulated locus which controls expression of chrorTiosomalmultiple antibiotic resistance in Escherichia coli J Bacferiol173: 5532-5538.
Hooper, D.C., Wolfson. J.S,. Ng, E.Y,, and Swartz, M.N,(1987) Mechanisms of action and resistance to ciproflox-acin. Am J/Wec/82 (Suppl 4A); 12-20.
Hooper. D.C., Wolfson, J.S., Bozza, M.A,, and Ng, E.Y,(1992) Genetic and regulation of outer membrane proteinexpression by quinolone resistance toci nfxB, nfxC. andcfxB. Antimicrob Agents Chemother 36: 1151-1154.
Kushner, S.R., Templin, A,, Nagaishi, H,, and Ctark, A,J.(1971) Genetic recombination in Eschetlchia coli: the roteof exonuclease t, Proc Natl Acad Set USA 68: 824-827,
Jenkins, D,E., Schuttz. J.E,, and Martin, A, (1988) Starvation-induced cross protection against heat or H2O2 chattenge inEscherichia coti J Sacter/o/170: 3910-3914,
Jenkins, D.E,. Auger, E.A,, and Martin, A. (1991) Rote ofRpoH, a heat shock regulator protein, in Escherichia coticarbon starvation protein synthesis survival. J Bacteriot173: 1992-1996,
Leritner, C. (ed.) (1981) Bile. In Geigy Scienmc Tables. 8thedn. Vol. I. Basle: Ciba-Geigy Corp., pp. 139-146.
Lomovskaya, O,. and Lewis, K, (1992) emr, an Escherichiaco//locus for muitidrug resistance. Proc Natl Acad Sci USA89: 8938-8942,
Ma, D,, Cook, D,N,, Alberti, M,, Pon, N,G,, Nikaido, H,, andHearst, J.E, (1993a) Motecutar ctoning and characteriza-tion of acrA and acrE genes ot £sc(ienchia coh. J Bacterio!175: 6299-6313.
Ma, D., Cook, D.N., O'Brien, D.A., and Hearst, J,E. (1993b)Analysis of the promoter and regulatory sequence of anoxygen-regulated bch operon in R. capsutatus by site-directed mutagenesis. jeacfer /oM75: 2037-2045.
Ma, D., Cook, D,N,, Hearst, J.E,, Nikaido, H. (1994) Effluxpumps and drug resistance in Gram-negative bacteria.Trends Microbioi 12: 489-493.
McMur'ry, L.M,, George, A,M., and Levy, S.B. (1994) Activeefflux of chloramphenicol in susceptible Esct~ieiichia colistrains and in multiple-antibiotic-resistant [mar) mutants.Anfiinicrob Agents Chemother 38: 542-546.
Milter. J.H, (1992) A Short Course in Bacteria! Genetics: aLaboratoiy Manuat and Handbook for Bschenchla coti andRelated Bacteria. Cotd Spring Harbor, New York: ColdSpring Harbor Laboratory Press.
Milter, P.F., Gambino, L.F., Sutavik, M.C, and Gracheck, S,J.(1994) Genelic relationship between soxRS and mar locipromoting multiple antibiotic resistance in Escherichfa coliAntimiorob Agents Chemother 3B: 1773-1779.
Mizuno, T,, Chou, M.-Y,, and tnouye, M, (1984) A uniquemechanism regutating gene expression: transtational intii-bition by a comptemenlary RNA tr-anscript (micRNA), ProcNatl Acad Sci USA 81 : 1966-1970.
Nakamura, H, (1965) Gene-cotilrolied i-esistance to acrrflavinand other basic dyes in Escherichia coli J Baeteriol 9Q: 8 -14.
Nakamura, H. (1968) Genetic determination of resistance toacriftavin, phenyt alcohot. and sodium dodecyt sutphate inEscherictiia eoti. J Bacteriot 96: 987-996,
Nikaido, H,, Rosenberg, E.Y,, and Foutds, J, (1983) Porinchannets in Eschertetm eott: studies with |.!-iactarns inintact ceils. J Baeteriol 153: 232-240.
Nit^aido, H,, and Vaara, M, (1985) Motecutar basis of bacteriatouter membrane permeability, Mierobiot Rev 45: 1-32.
Pan, W., and Spratt, B.D, (1994) Regulation of the perme-ability of the gonococcat cett envelope by the Mtr system.Mol Microbiol^^: 769-755,
Ptesiat, P,, and Nikaido, H, (1992) Outer membranes ofGram-negative bacteria ere permeabte to steroid probes.Mol Microbioi 6: 1323-1333,
Pooie. K., Krebes, K,, Mcnally, C , and Neshat, S. (1993)Muttipte antibiotic resistance in Pseudomonas aeruginosa-evidence for involvement of ati efflux operon. (1993) JBaeteriot 175: 7363-7372.
Sambrook, J., Fritsch, E.F,, and Maniatis. T, (1989) MotecutarCloning: A Lahoratory Manuai 2nd edn, Cotd SpringHarbor, New York. Cold Spring Harbor Laboratoiy Pr'ess,
Seiffen D., Ktein, J.R,, and Ptapp, R, (1993) EnvC, a newtipoprotein of the cytoptasmic membrane of Esctierlehiaeoli EEMS Microbioi Lett 107: 175-178.
Sheu. C.W., and Freese, E. (1973) Lipopotysaccharide layerprotection of Gram-negative bacteria against inhibition bylong-chain fatty acids, J Bacterial 115: 869-B75.
Sukupoivi, S., and Vaara, M. (1989) Salmonetta typhimuriumand Escherichia coli mutants witti increased outer mem-brane permeabltity to hydrophobic compounds. BiochimBiophys Aefa 988: 377-387.
Xu, J,, Niltes. M.L., and Bertrand, K.P. (1993) TheEseheriehia coli acrAB and acrEP genes: a new family ofmuUidiug effiux pumps. In Absfr 93rd Gen Meet Am SocMicrobioi 1993. Washington, D.C: Ariierican Society forMicrobiotogy, Abstr. K-169, p, 290.
Zimmermann, W., and Rosselet, A, (1977) Function of theouter membrane of Eschertchia coii as a permeabititybarrier to beta-lactam antibiotics, Antimicrob AgentsChemofher12: 368-372,
Cl' 1995 Biackweil Science Ltd, Molecular Microbioiogy, 16, 45-55






























