Embed Size (px)
Citation preview

(CANCER RESEARCH 50. 6068-6074. September 15. 1990]
A Comparison of Synaptophysin, Chromogranin, and L-Dopa Decarboxylase as
Markers for Neuroendocrine Differentiationin Lung Cancer Cell Lines1
Sandra M. Jensen, Adi F. Gazdar, Frank Cuttitta, Edward K. Russell, and R. Ilona Linnoila2
National Cancer Institute-Navy Medical Oncology Branch, Clinical Oncology Program, Division of Cancer Therapy, National Cancer Institute Navy Hospital,Belhesda. Mary-land 20814
ABSTRACT
Synaptophysin is a M, 38,000 integral membrane glycoprotein expressed by a variety of normal and neoplastic neuroendocrine cells. \Vestudied Synaptophysin as an immunocytochemical marker for neuroendocrine differentiation in lung cancer and compared it to the immunocytochemical expression of chromogranin A, a marker for dense core(endocrine) granules, and the biochemical activity of i.-dopa decarboxyl-ase (DDC), the key amine-handling enzyme. Of the 250 cell linesavailable to us, we selected examples representative of the following celltypes: bronchial carcinoids (n = 4), small cell lung cancer (SCLC) (n =7), extrapulmonary small cell carcinomas (n = 4), and non-small celllung cancers (n = 18) whose neuroendocrine status had been previouslydetermined on the basis of electron microscopy and DDC activity. Wedemonstrated (a) there was a higher incidence of Synaptophysin thanchromogranin A immunoreactivity in carcinoid (100 versus 75%), classicSCLC (70 versus 50%), and variant SCLC (57 versus 29%) cell lines; (ft)3 of the 4 (75%) extrapulmonary small cell lung cancer cell linesexpressed Synaptophysin and chromogranin A; (c) 5 of the 7 (71%) non-small cell lung cancer cell lines previously shown to express multipleneuroendocrine markers were positive for Synaptophysin, chromograninA, and DDC activity; (</) none of the other 11 non-small cell lung cancercell lines expressed Synaptophysin or chromogranin A; and (<•)formalin
Fixation and paraffin embedding reduced Synaptophysin immunoreactivityin 11 of 14 (79%) of the cell lines, as compared to freshly preparedspecimens fixed in 95% ethanol. Western blot analysis using the synap-tophysin antibody (SY38) demonstrated immunoreactive proteins rangingfrom U, 43,000 to 45,000 in five representative cell lines. The concordance of expression of all three neuroendocrine markers was statisticallysignificant when values for all cell lines were totalled. Synaptophysin wasa more commonly expressed marker for variant SCLC cell lines, whichrarely showed DDC activity. We conclude that Synaptophysin may be amore sensitive and specific marker for neuroendocrine differentiation,when compared to chromogranin A and DDC in lung cancer cell lineswhich express only part of the neuroendocrine program.
INTRODUCTION
NE' tumors are a diverse group of neoplasms that usually
express multiple markers for NE differentiation. They are characterized by the presence of dense core granules, high concentrations of the key amine-handling enzyme L-DOPA decarbox-ylase (aromatic-L-amino acid decarboxylase, EC 4.1.1.28), andthe production of various hormones and neuropeptides (1, 2).
Received 1/20/89; accepted 6/18/90.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1The opinions and assertions contained herein are the private views of theauthors and are not to be construed as official or reflecting the views of theDepartment of the Navy, the Department of Health and Human Services, or theDepartment of Defense.
2To whom requests for reprints should be addressed, at NCI-Navy MOB,
Naval Hospital. Building 8, Room 5101. Bethesda. MD 20814.3The abbreviations used are: NE. neuroendocrine; SCLC-C, classic small cell
lung cancer; SCLC-V, variant small cell lung cancer; NSCLC, non-small cell lungcancer; ExPuSC, extrapulmonary small cell carcinoma; Syn, Synaptophysin; CgA,chromogranin A; dopa, dihydroxyphenylalanine: DDC. L-dopa decarboxylase;NSE, neuron-specific enolase; PAGE, polyacrylamide gel electrophoresis; PBS.phosphate-buffered saline; SCLC, small cell lung cancer: SDS, sodium dodecylsulfate.
NE tumors of the lung include bronchial carcinoid, SCLC-C,and SCLC-V cell types. It is important to distinguish SCLCfrom other lung cancers because of the better therapeutic response to cytotoxic therapy in patients (3). Among SCLC, thevariant subtype SCLC-V is associated with worse prognosisthan SCLC-C. Although considered NE tumors, SCLC-V celltypes typically lack dense core granules and may, therefore, bedifficult to diagnose (4-6).
ExPuSC is morphologically similar to SCLC and has beendescribed in a large variety of sites, including esophagus, stomach, pancreas, larynx, hypopharynx, salivary glands, nasal cavity and paranasal sinuses, thymus, small and large bowel, uterine cervix, endometrium, breast, prostate, urinary bladder, andskin. A significant number of, but not all, ExPuSC tumors havebeen shown to contain neurosecretory granules and produceectopie hormones (which can be demonstrated using immuno-histochemical techniques) (7-9). We included four ExPuSC celllines in our study because of their similarities to NE tumors ofthe lung, namely SCLC.
NSCLC do not express NE features and are usually diagnosedon the basis of histology alone. However, we and others havereported that some NSCLCs, diagnosed by conventional lightmicroscopy, express a variety of NE markers when immunocytochemical techniques are employed. This subset of NSCLCsthat express multiple NE markers has been named NSCLC-NE(10, 11).
Specific NE markers in lung cancers consist of a large numberof defined hormones and neuropeptides, both eutopic (i.e.,gastrin-releasing peptide, calcitonin, and serotonin) and ectopie(adrenocorticotropic hormone, vasopressin, and neurotensin).However, these NE products are seldom very useful for diagnosis, since their expression in tumors is quite variable and maybe subtype specific ( 10).
Among the currently available general NE markers are NSE,Leu-7, CgA, and DDC (2, 10). Immunocytochemical and enzymatic techniques have demonstrated these general NE markers to be useful investigative and diagnostic tools, but notwithout some limitations. NSE, the 7-subunit of the glycolyticenzyme enolase, was originally thought to be expressed exclusively in neurons and NE cells (12). It is now known that NSEis expressed in a variety of non-NE cells and tumors, which hasled many investigators to question its specificity (13-15). Leu-7, a differentiation antigen shown to be selectively expressedon human natural killer cells (16), is also expressed on SCLCcells and other benign and malignant NE and nerve cells,although the precise antigenic determinant of the antibody hasyet to be determined (17, 18). CgA was originally isolated fromthe chromaffin granules of the adrenal medulla (19) and hasbeen found to be expressed by a variety of normal and neoplastichuman polypeptide hormone-producing tissues (20-22). Al
though its function is not known, CgA is associated with theco-secretion of a wide variety of hormones, suggesting somerole in the secretory process (22). Because SCLCs have a paucity
6068
on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

SYNAPTOPHYSIN IN LUNG CANCER CELL LINES
of secretory granules (4), the use of CgA as a general NE markermay be limited only to those neoplasms containing more abundant dense core granules (5, 10, 23). These facts demonstratethe need for additional NE markers with greater sensitivity andspecificity.
Syn was originally isolated from bovine neurons and is a M,38,000 membrane-integrated glycosylated polypeptide to whicha monoclonal antibody (SY 38) has been raised (24). Syn hasbeen demonstrated in a variety of normal and neoplastic NEcells of neural and epithelial type (25, 26) and has been localizedwithin normal and neoplastic cells of the bronchopulmonarytract (27). While some results suggest that Syn immunoreactiv-ity is maintained in formalin-fixed paraffin-embedded specimens (26-28), others have reported variable immunoreactivity(29). Our goal in the present study was to define the sensitivityand specificity of Syn as a marker for NE differentiation inlung cancer cell lines and to compare it to other general NEmarkers.
MATERIALS AND METHODS
Cell Lines. Four bronchial carcinoid, 4 ExPuSC, 10 SCLC-C, 7SCLC-V, 11 NSCLC, and 7 NSCLC-NE cell lines were established,maintained, and characterized in our laboratory, as described previously(4, 5, 30, 31). Cell lines demonstrating substrate adherence were tryp-sinized by adding 10 ml of warm PBS to the flask and 5 ml of 0.05%trypsin to stop cell adherence. Once trypsinization was complete (5-10min), 10 ml of fresh serum-containing medium were added to neutralizethe trypsin. The cell suspension was centrifuged at 1200 rpm for 5 minand resuspended in fresh medium. Trypsinized cells were allowed torecover for 2-3 h prior to immunocytochemical staining. All cell lineswere washed twice in 10 ml of ice-cold PBS and cytocentrifuged ontopoly(L-lysine) (Sigma Chemical Company, St. Louis, MO)-coatedslides.
Immunocytochemical Staining. Immunocytochemical staining wasperformed by the avidin-biotin-peroxidase technique, using VectastainABC staining kits (Vector Laboratories, Burlingame, CA), followingthe vendor's instructions with a few modificai ions (10). Mouse mono
clonal anti-Syn (SY 38, lot 10644532-01) was purchased from Boehrin-ger Mannheim (Indianapolis, IN) and used at a 1:10 dilution. Mousemonoclonal anti-CgA (LK2H10) was a gift from Dr. Barry S. Wilson(University of Michigan Medical Center, Ann Arbor. MI) and was usedat a dilution of 1:100.
Immunoreactivity for Syn and CgA was tested following the fixationof freshly made cytospin preparations in methanol, acetone, and 95%ethanol. Fixation for 10 min in ice-cold 95% ethanol appeared to givethe best results (i.e., strongest staining with least background and goodpreservation of morphology). Furthermore, pelleted cells were fixed forl h at 4°Cin freshly prepared 4% neutral buffered paraformaldehyde
solution and rinsed with ice-cold PBS. The cell pellets were embeddedin paraffin, and 5-^m sections were mounted on gelatin-coated slides.
Cytopreparations of each cell line were stained with hematoxylin andeosin and reviewed to ensure proper cell distribution and morphology.Positive controls for immunocytochemistry in each assay includedparaffin-embedded sections of a Syn-positive carcinoid and a CgA-positive pheochromocytoma. Controls for the specificity of stainingincluded (a) using irrelevant antibody as a primary antibody and (b)omitting the primary antibody from the staining procedure. Both ofthese controls gave negative staining results.
Immunocytochemical staining results were scored independently bytwo of the authors (S. M. J. and R. I. L.) for the intensity of staining(0 = negative, 1 = weak, 2 = moderate, 3 = strong) and distribution ofpositive cells (0 = no positive cells, 1 = <10%, 2 = 10-50%, 3 = >50%of tumor cells positive). Discrepancies were resolved after joint review.
i,-Dopa Decarboxylase Assay. DDC assays were performed by amodification (32) of a previously published method (33). Freshly prepared cell pellet homogenates were clarified and frozen at -70°C untilanalysis. One unit of enzyme activity was defined as 1 nmol of 14CO2/
h. DDC values of >4 units/mg of soluble protein were scored positive,since minimal DDC activity can occur in most tissues, including normallung.
Western Blot Analysis. Tumor cell lines were evaluated by Westernblot analysis for expression of Syn, using a modified method of theTowbin and Gordon technique (34, 35). Whole-cell lysates were adjusted to equal protein concentrations and applied to the gel at 10-20Mgof protein/well. SDS-PAGE was performed under reduced conditions and fractionated products were electroblotted onto nitrocellulosemembranes. The membranes were then treated with anti-Syn antiserum(SY38, 0.1 mg/ml) at a dilution of 1:100, followed by exposure torabbit anti-mouse IgG (lot 17529; Cappel Laboratories, Inc., Cochran-ville, PA) at a dilution of 1:1000. Selective antibody deposition wasidentified with labeled '"I-Protein A. To confirm the specificity andcompetency of our immunostaining technique with the anti-Syn reagent, a parallel experiment was performed on identical cell lines, usingan indifferent isotypically matched monoclonal antibody, MOPC 21,at a concentration of 0.1 mg/ml.
RESULTS
The results for Syn staining distribution are summarized inTable 1. All carcinoids were positive for Syn and 3 of the 4 celllines had over 50% cells positive. Syn staining in small cellcarcinoma cells lines was more heterogeneous, with 14 of the21 cell lines positive. The 5 Syn-positive NSCLC cell lines allbelonged to the NSCLC-NE subtype.
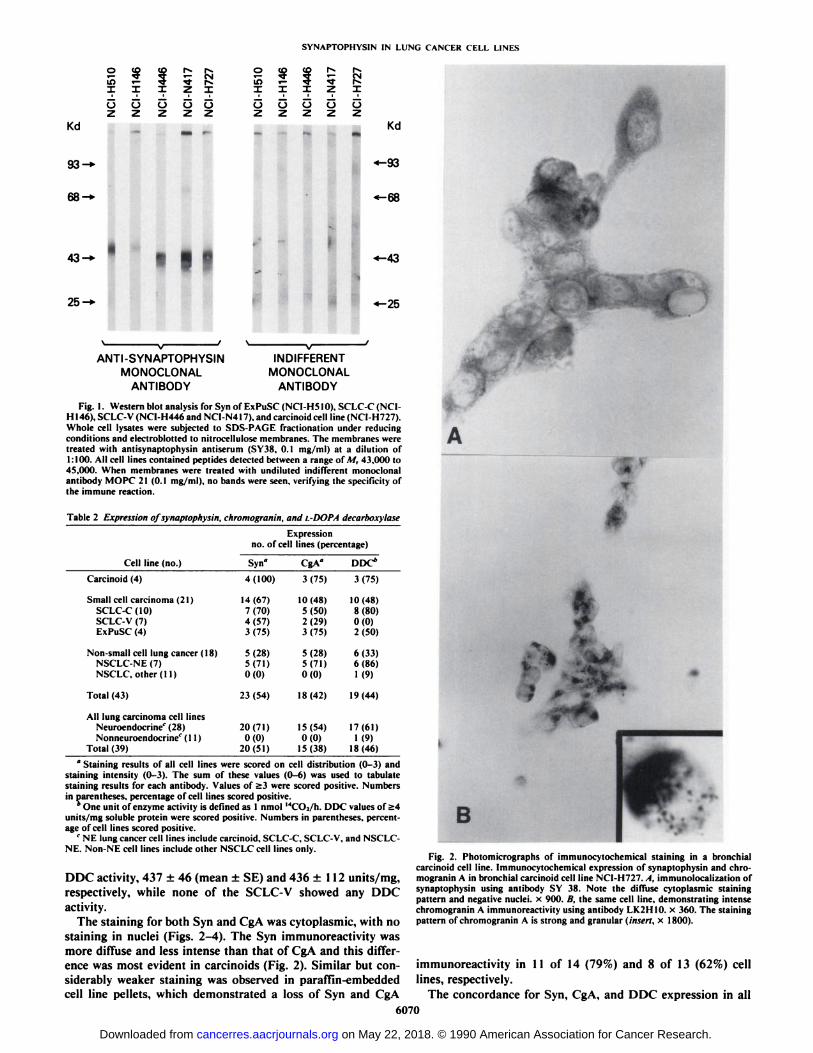
Western blot analysis was performed to demonstrate antibodyspecificity on representative cell lines including 1 carcinoid, 1SCLC-C, 2 SCLC-V, and 1 ExPuSC (Fig. 1). Physical characterization of these cell lines by SDS-PAGE revealed a distinctcellular component which expressed Syn immunoreactivity.This component migrated as a single band in Fig. 1, lanes Iand 2, and as a series of bands in Fig. 1, lanes 3-5, with adetermined molecular weight ranging from 43,000 to 45,000under the reduced conditions. The reactivity was specific forantibody SY38, since no immunoreactive bands were observedin any of the cell lines when indifferent mouse monoclonalantibody (MOPC 21) was substituted for SY38 (Fig. 1).
Syn, CgA, and DDC expression in all cell lines is summarizedin Table 2. Syn was expressed more commonly in carcinoids(100%) and SCLC (65%) than CgA and DDC. Four of 7 SCLC-V were positive for Syn, while only 2 of 7 were positive forCgA. In NSCLC cell lines the expression of Syn as well as thetwo other markers were restricted to the NSCLC-NE subtype,which showed average DDC levels of 273 ±43 units/mg. Asexpected, SCLC-C and carcinoid cell types showed the highest
Table 1 Expression of synaptophysin staining distribution in all cell typesThe distribution score for synaptophysin staining in each cell line ranges from
Oto 3;0 = no positive cells, 1 = <IO%, 2 = 10-50%. and 3 = 51-100% of tumorcells positive. These categories for distribution were used throughout the paper.Numbers in parentheses, percentage of cell lines scored positive.
Syn stainingno. of cell lines (percentage)
Cell line(no.)Carcinoid
(4)Small
cell carcinoma (21)SCLC-C (10)SCLC-V (7)ExPuSC(4)Non-small
cell lung cancer(18)
NSCLC-NE (7)NSCLC. other(11)Total
(43)00(0)7(33)
3 (30)3(43)
1(25)13(72)2(29)
11(100)20
(47)10(0)1
(5)1 (10)0(0)0(0)1
(5)1(14)
0(0)2(5)21(25)7
(33)3 (30)3(43)1(25)3(17)3
(43)0(0)1
1 (26)33
(75)6(29)
3 (30)1 (14)2(50)1(5)1
(14)0(0)10(23)
6069
on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
SYNAPTOPHYSIN IN LUNG CANCER CELL LINES
X X X Z X
G G G G Gz z z z z
S r- 5 5 r--X X X Z X
ö G G G ö
Kd
93-
43-
25-
•II
Kd
«-93
«-68
«-43
«-25
ANTI-SYNAPTOPHYSINMONOCLONAL
ANTIBODY
INDIFFERENTMONOCLONAL
ANTIBODY
Fig. l. Western blot analysis for Syn of ExPuSC (NCI-H510). SCLC-C (NC1-H146). SCLC-V (NCI-H446 and NC1-N417). and carcinoid cell line (NCT-H727).Whole cell Usâteswere subjected lo SDS-PAGE fractionation under reducingconditions and electroblotted to nitrocellulose membranes. The membranes weretreated with antisynaptophysin anliserum (SY38. O.I mg/ml) at a dilution of1:100. All cell lines contained peptides detected between a range of M, 43.000 to45.000. When membranes were treated with undiluted indifferent monoclonalantibody MOPC 21 (0.1 mg/ml). no bands were seen, verifying the specificity ofthe immune reaction.
Table 2 Expression of synaptophysin. chromogranin, and i.-liOPA Jecarboxylase
Expressionno. of cell lines (percentage)
Cell line(no.)Carcinoid
(4)Small
cell carcinoma (21)SCLC-C (10)SCLC-V (7)ExPuSC(4)Non-small
cell lung cancer (18)NSCLC-NE (7)NSCLC. other (11)Syn"4(100)14(67)
7(70)4(57)3(75)5(28)
5(71)0(0)CgA"3
(75)10(48)
5(50)2(29)3(75)5(28)
5(71)0(0)DDC*3
(75)10(48)
8(80)0(0)2(50)6
(33)6(86)1 (9)
Total (43) 23(54) 18(42) 19(44)
All lung carcinoma cell linesNeuroendocrine''(28) 20(71) 15(54) 17(61)Nonneuroendocrine' (II) 0(0) 0 (0) 1 (9)
Total (39) 20(51) 15(38) 18(46)" Staining results of all cell lines were scored on cell distribution (0-3) and
staining intensity (0-3). The sum of these values (0-6) was used to tabulatestaining results for each antibody. Values of >3 were scored positive. Numbersin parentheses, percentage of cell lines scored positive.
* One unit of enzyme activity is defined as I nmol MCO2/h. DDC values of >4
units/mg soluble protein were scored positive. Numbers in parentheses, percentage of cell lines scored positive.
' NE lung cancer cell lines include carcinoid, SCLC-C, SCLC-V, and NSCLC-NE. Non-NE cell lines include other NSCLC cell lines only.
DDC activity, 437 ±46 (mean ±SE) and 436 ±112 units/mg,respectively, while none of the SCLC-V showed any DDCactivity.
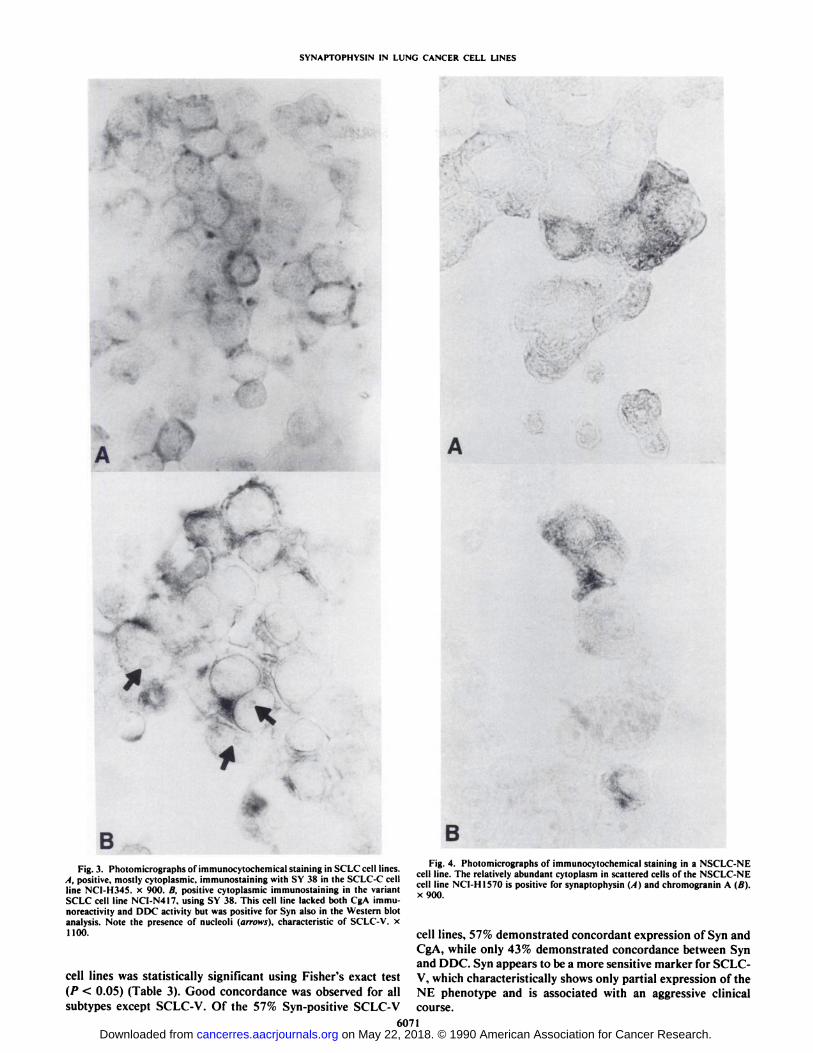
The staining for both Syn and CgA was cytoplasmic, with nostaining in nuclei (Figs. 2-4). The Syn ¡mmunoreactivity wasmore diffuse and less intense than that of CgA and this difference was most evident in carcinoids (Fig. 2). Similar but considerably weaker staining was observed in paraffin-embeddedcell line pellets, which demonstrated a loss of Syn and CgA
;T
*
B
Fig. 2. Photomicrographs of immunocytochemical staining in a bronchialcarcinoid cell line. Immunocytochemical expression of synaptophysin and chro-mogranin A in bronchial carcinoid cell line NCI-H727. A, immunolocalization ofsynaptophysin using antibody SY 38. Note the diffuse cytoplasmic stainingpattern and negative nuclei, x 900. A. the same cell line, demonstrating intensechromogranin A immunoreactivity using antibody LK2H10. x 360. The stainingpatlern of chromogranin A is strong and granular (insert, x 1800).
immunoreactivity in 11 of 14 (79%) and 8 of 13 (62%) celllines, respectively.
The concordance for Syn, CgA, and DDC expression in all6070
on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
SYNAPTOPHYSIN IN LUNG CANCER CELL LINES
Im
.
•I
A
B B•• •c-^i <~ n i- Fig. 4. Photomicrographs of immunocvtochemical slamine in a NSCLC-NEFig. 3. Photomicrographs of immunocvlochcmical staining in SC LC cell lines. .. ...... , .. , , ' , , . uri. x,c-,~i /- v.r-
6. . . . ' . . . , ,.., ,„. ., C-/-I/-/~ ¡i cell line. The relatively abundant cvloplasm in scattered cells of Ihc NSC LC-NEA. positive, mostly cvtoplasmic. mimunostaining with Si 3«in the SC LC-C cell .... *,,-, U««<«A¿--r i.-/ ji. -./»>x,;'H-,_.,.,., J „"„„„ .. , ...... cell line NCI-HI570 is positive for svnaptophvsin (A and chromoeranin A (B).line NCI-H345. x. 900. B, positive cvtoplasmic immunostainmg in the variant „..SCLC cell line NCI-N417. using SV 38. This cell line lacked both CgA immu-noreactivitv and DDC activity hut was positive for Syn also in the Western blotanalysis. Note the presence of nucleoli (arrows), characteristic of SCLC-V. X110°- cell lines, 57% demonstrated concordant expression of Syn and
CgA, while only 43% demonstrated concordance between Synand DDC. Syn appears to be a more sensitive marker for SCLC-
cell lines was statistically significant using Fisher's exact test V, which characteristically shows only partial expression of the
(P < 0.05) (Table 3). Good concordance was observed for all NE phenotype and is associated with an aggressive clinicalsubtypes except SCLC-V. Of the 57% Syn-positive SCLC-V course.
6071on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

SYNAPTOPHYSIN IN LUNG CANCER CELL LINES
Table 3 Concordant expression ofsynaptophysin, chromogranin A, and ¡.-dopadecarboxylase
Concordance is when both markers are present in a particular cell line. Thenumbers in parentheses represent percentage. Concordances were statisticallysignificant (P < 0.05) for all cell types (n = 43) using Fisher's exact test.
Expressionno. of cell lines (percentage)
Cell line(no.)Carcinoid
(4)Small
cell carcinoma (21)SCLC-C(IO)SCLC-V (7)ExPuSC(4)Non-small
cell lung cancer (18)NSCLC-NE (7)NSCLC, other(11)Total
(43)Syn"
andCgA°3(75)14
(67)6(60)4(57)
4(100)16(89)
5(71)11(100)33
(77)Syn"
andDDC*3(75)13(62)
7(70)3(43)
3(75)17(94)
6(86)11(100)33
(77)CgA"
andDDC*3
(75)14(67)
6(60)5(71)
3(75)17(94)
6(86)11(100)35(81)
" Staining results for all cell lines were scored on cell distribution (0-3) andstaining intensity (0-3). The sum of these values (0-6) was tabulated for eachantibody. Values of >3 were scored positive.
* One unit of enzyme activity is defined as 1 nmol l4CO2/h. DDC values of >4
units/mg soluble protein were scored positive.
DISCUSSION
Our findings indicate that Syn is a specific and sensitiveimmunocytochemical marker for NE differentiation in lungcancer cell lines and ExPuSC. We demonstrated that Synimmunoreactivity is expressed more often in bronchial carci-noid, SCLC-C, and SCLC-V cell lines, in comparison to immunoreactivity with CgA. These results suggest that Syn maybe a more sensitive biochemical marker for NE differentiationthan CgA in these subtypes. Antibodies to both Syn and CgAshowed positivity with the same frequency in ExPuSC andNSCLC-NE cell lines, while none of the other NSCLC celllines expressed either Syn or CgA.
When all cell types were considered, the concordant expression of two of the three NE markers (Syn, CgA, DDC) washighest for CgA and DDC (81%). These results are in agreementwith our earlier findings, which report excellent concordancebetween CgA and DDC in both SCLC and NSCLC lung tumors(3). Syn was expressed in 70% of the SCLC-C cell lines, whichcorrelates well with a previous study on tumor specimens (24).
Of the SCLC-V cell lines which are characterized by alteredmorphology and partial lack of the NE program (4, 5), 4 of 7(57%) expressed Syn, while only 2 of 7 (29%) stained positivelyfor CgA. The presence of Syn in SCLC-V was also confirmedby Western blot analysis. None of the SCLC-V cell lines werefound to express DDC. This is in agreement with previousfindings which report a loss of dense core granules and lack ofDDC activity in this SCLC subset (4, 5). Because patients withSCLC-V may respond less well to chemotherapy (6), the detection of these tumors is of clinical and biological interest.
We have recently shown that as many as 20% of NSCLCtumors, most commonly adenocarcinomas and large cell carcinomas, contain multiple NE markers (NSCLC-NE) (10). Preliminary results suggest that patients with NSCLC-NE tumorsmay respond more favorably to chemotherapy than patientswith other NSCLC tumors (11). We included a panel of 7NSCLC-NE cell lines in the present study and found that 4 of7 (57%) NSCLC-NE cell lines expressed all three NE markers,Syn, CgA, and DDC. Of interest, 2 of these were large cellcarcinoma cell lines, 1 bronchioloalveolar and 1 adenosqua-mous carcinoma. Thus, the addition of Syn as a marker for NEdifferentiation in NSCLC tumors diagnosed by conventionallight microscopy techniques may prove to be clinically impor
tant, since it has been suggested that NSCLC-NEs are initiallymore sensitive to chemotherapy than other NSCLCs (11, 36).
A marked loss of Syn immunoreactivity was demonstrated in11 of 14 (79%) of the formalin-fixed paraffin-embedded cellpellets (previously showing immunoreactivity in freshly prepared ethanol-fixed specimens). These results support previously reported findings that various fixation techniques canalter the immunocytochemical demonstrability of Syn (29).Moreover, it appears that cultured cells are even more likely tolose their immunoreactivity due to formalin fixation and paraffin embedding than routinely processed tissue samples collected in the practice of surgical pathology (28).
Our study on cell lines is in accordance with previouslypublished results showing that immunoreactive Syn is presentin vivo in most but not all NE tumors of the lung (25-28, 37,38). While many studies have included only a few cases, Gouldet al. (25) in their study on 270 tumors reported that up to 17of 19 (88%) pulmonary carcinoids and 6 of 14 (43%) NEcarcinomas of the lung were positive for Syn. Higher incidenceswere reported for lung NE carcinomas (88%) by Lee et al. (27)and for SCLC (79%) by Kayser et al. (28), who also noted that8 of 74 (10%) NSCLC were positive for Syn. In our in vitrostudy, 70% of SCLC-C and 67% of all small cell carcinoma celllines tested positive for Syn. In summary, these studies suggestthat, while most lung NE tumors contain Syn, some tumors donot express this protein. Presently, the biological or clinicalimplications of this are not fully understood.
The gene for human Syn has been previously cloned and wasdemonstrated to generate a single mRNA transcript whichencodes a 30,700-amino acid precursor protein (39). Althoughthe deduced molecular weight of this prepro-Syn molecule is33,312, multiple forms have been reported in a variety of humantissue, ranging in molecular weight from 38,000 to 40,000 (39).This reported heterogeneity in the calculated molecular weightof the Syn precursor, particularly evident in tumor samples, hasbeen attributed to the degree of glycosylation obtained duringposttranslational modification. Studies by Navone et al. (40)and Rehm et al. (41) have shown that chemical deglycosylationor inhibition of de novo glycosylation renders the Syn precursorof «34,000. Our biochemical characterization of Syn-immu-noreactive proteins in lung tumor cell lines revealed a slightlyhigher molecular weight form, ranging from 43,000 to 45,000.This is in accordance with the slightly lower electrophoreticmobility that has been reported for selected tumors, includinga bronchial carcinoid. paragangliomas, and retinoblastomas(26, 37, 42). Interestingly, the SCLC-C (NCI-HI46) and ExPuSC (NCI-H510) cell lines expressed a higher molecularweight form of M, 45,000, when compared to the SCLC-V(NCI-H446 and NCI-N417) and carcinoid (NCI-H727) celllines, in which a M, 43,000 product was noted. These datasuggest that, in addition to extensive glycosylation of the Synmolecule, there may be tumor selectivity associated with thisposttranslational modification step in lung and extrapulmonarytumor cell lines.
The function of Syn is not known. Syn is a hexameric proteincapable of forming transmembrane channels (43) and is one ofthe major calcium-binding proteins of the synaptic vesicle mem
brane, with the binding site on its cytoplasmic domain (41).Moreover, Syn is one of the major target proteins of endogenous tyrosine phosphorylation in highly purified synaptic vesicles, as well as in synaptosomes from rat forebrain (44). It canalso be phosphorylated by the c-src-encoded protein tyrosine
6072
on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

SYNAPTOPHYSIN IN LUNG CANCER CELL LINES
kinase.4 While Syn was initially localized in intracytoplasmic
clear vesicles, subsequently smaller amounts were also found inlarge dense core vesicles (40, 45-49). Thus, it can be speculatedthat Syn has a role in the intracellular vesicle transport system,in communication with membrane and cytoskeleton, or inexocytosis. In cancer cells, this may involve intracellular trafficking of growth factors and oncogenes.
In summary, we recommend that Syn be added to the panelof general NE markers currently in use for the following reasons: (a) Syn may be a more specific and sensitive biochemicalmarker for NE differentiation than CgA in carcinoid and classicSCLC cell types; (b) Syn is a more sensitive marker for NEneoplasms which lack part of the NE program (i.e., SCLC-V);(c) Syn may be helpful in identifying differentiating, and classifying small cell carcinomas of extrapulmonary or unknownorigin; and (</) Syn may be a clinically important tool inidentifying patients with NSCLC-NE neoplasms.
REFERENCES
1. Gould. V. E.. Linnoila. R. I.. Memoli. V. A., and Warren. W. Biology ofdisease—neuroendocrine components of the bronchopulmonary tract: hyper-plasias. dysplasias. and neoplasms. Lab. Invest., 49: 519-537, 1983.
2. Gazdar. A. F., Helman. L.. Israel, M. A., Russell. E. K.. Linnoila. R. I..Mulshine. J.. Schuller. H., and Park. J. G. Expression of neuroendocrinecell markers L-dopa decarboxylase. chromogranin A. and dense core granulesin human tumors of endocrine and non-endocrine origin. Cancer Res.. 48:4078-4082. 1988.
3. Gazdar, A. F.. and Linnoila. R. I. The pathology of lung cancer—changingconcepts and newer diagnostic techniques. Semin. Oncol., IS: 215-225. 1988.
4. Carney, D. N., Gazdar. A. F., Bepler, G., Guccion. J. G.. Marangos. P. J..Moody. T. W., Zweig, M. H., and Minna. J. D. Establishment and identification of small cell lung cancer cell lines having classic and variant features.Cancer Res.. 45: 2913-2923, 1985.
5. Gazdar, A. F., Carney. D. N., Nau, M. M., and Minna. J. D. Characterizationof variant subclasses of cell lines derived from small cell lung cancer havingdistinctive biochemical, morphological, and growth properties. Cancer Res.,«.•2924-2930,1985.
6. Radice. P. A.. Matthews, M. J.. Ihde. D. C.. Gazdar. A. F.. Carney. D. N.,Bunn. P. A., Cohen. N. H.. Fossieck, B. E.. Makuch. R. W.. and Minna. J.D. The clinical behavior of "mixed" small cell/large cell bronchogenic carcinoma compared to "pure" small cell subtypes. Cancer (Phila.). 50: 2894-
2902. 1982.7. Ibrahim. N. B. N., Briggs, J. C.. and Corbishley, C. M. Extrapulmonary oat
cell carcinoma. Cancer (Phila.), 54: 1645-1661. 1984.8. Richardson. R. L.. and Weiland. L. H. llndifferentiated small cell carcinomas
in extrapulmonary sites. Semin. Oncol., 9: 484-496, 1982.9. Gould. V. E.. Banner, B. F.. and Bacrwaldt. M. Neuroendocrine neoplasms
in unusual primary sites. Diagn. Histopathol.. 4: 263-277. 1981.10. Linnoila, R. I.. Mulshine. J., Steinberg, S. M., Fuña.K.. Matthews. M. J..
Cotelingam. J.. and Gazdar, A. F. Neuroendocrine differentiation in endocrine and nonendocrine lung carcinomas. Am. J. Clin. Pathol.. 90: 1-12,1988.
11. Linnoila. R. I., Jensen. S.. Steinberg, S.. Minna. J., Gazdar, A. F.. andMulshine. J. Neuroendocrine differentiation correlates with favorable response to chemotherapy in patients with non-small cell lung cancers (Abstract). Lung Cancer, 4: 33. 1988.
12. Schmechel, D., Marangos. P. J.. and Brightman. M. Neuron-specific enolaseis a molecular marker for peripheral and central neuroendocrine cells. Nature(Lond.). 276: 834-836. 1978.
13. PÃ hlman. S.. Esscher, T.. and Nilsson. K. Expression of -y-subunit of enolase,neuron specific enolase. in human non-endocrine tumors and derived celllines. Lab. Invest.. 54: 554-560. 1986.
14. Haimoto. J.. Takahashi. V., Koshikawa, T.. Nagura, H,. and Kato. K.Immunohistochemical localization of -y-enolase in normal human tissuesother than nervous and neuroendocrine tissues. Lab. Invest-, 52: 257-263.1985.
15. Schmechel. D. E. >-Subunit of the glycolytic enzyme enolase: nonspecific orneuron specific? Lab. Invest., 52: 239-242, 1985.
16. Abo. T., and Balch. C. M. A differentiation antigen of human NK and Kcells identified by a monoclonal antibody (HNK-1). J. Immunol.. 727: 1024-1029, 1981.
17. Bunn. P. A.. Linnoila. R. I.. Minna. J. D.. Carney, D., and Gazdar. A. F.Small cell lung cancer, endocrine cells of the fetal bronchus, and otherncuroendocrine cells express the Leu-7 antigenic determinant present onnatural killer cells. Blood. 65: 764-768. 1985.
18. Schuller-Petrovic. S., Gebhart. W., Lassmann. R. H.. Rumpold. H.. andKraft. D. A shared antigenic determinant between natural killer cells and
19.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44.
1A. Barnekow. personal communication.
nervous tissue. Nature (Lond.). 306: 179-181. 1983.O'Connor, D. T.. Frigon, R. P.. and Sokoloff. R. L. Human chromogranin
A: purification and characterization from catecholamine storage vesicles ofhuman pheochromocytoma. Hypertension (Dallas). 6: 2-12. 1984.O'Connor, D. T., Burton. D.. and Deftos. L. J. Immunoreactive human
chromogranin A in diverse polypeptide hormone producing human tumorsand normal endocrine tissues. J. Clin. Endocrinol. Metab.. 57: 1084-1086.1983.O'Connor. D. T.. Burton. D., and Deftos. L. J. Chromogranin A: immuno-
histology reveals its universal occurrence in normal polypeptide hormoneproducing endocrine glands. Life Sci.. 33: 1657-1663, 1983.Deftos, L. J., Linnoila, R. !.. Carney, D. N.. Burton. D. W., Leong. S. S.,O'Connor. D. T., Murray, S. S., and Gazdar. A. F. Demonstration of
chromogranin A in human neuroendocrine cell lines by immunohistologyand immunoassay. Cancer (Phila.). 62: 92-97, 1988.Wilson. B. S., and Lloyd, R. V. Detection of chromogranin in neuroendocrinecells with a monoclonal antibody. Am. J. Pathol.. 115: 458-468. 1984.Wiedenmann. B.. and Franke, W. W. Identification and localization ofsynaptophysin. an integral membrane protein of M, 38.000 characteristic ofpresynaptic vesicles. Cell. 41: 1017-1028. 1985.Gould. V. E., Wiedenmann. B., Lee, I., Schwechheimer. K., Dockhorn-Dworniczak. B.. Radoscvich, J.. Moll, R.. and Franke. W. W. Synaptophysinexpression in neuroendocrine neoplasms as determined by immunocytochem-istry. Am. J. Pathol., 126: 243-257. 1987.Wiedenmann. B.. Franke, W. W.. Kühn.C., Moll, R.. and Gould, V. E.Synaptophysin: a marker protein for ncuroendocrine cells and neoplasms.Proc. Nati. Acad. Sci. USA, 83: 3500-3504. 1986.Lee, I., Gould. V. E., Moll, R., Wiedenmann. B.. and Franke. W. W.Synaptophysin expressed in the bronchopulmonary tract: neuroendocrinecells, neuroepithelial bodies, and neuroendocrine neoplasms. Differentiation,34: 115-125, 1987.Kayser, K., Schmid. W.. Eben, W., and Wiedenmann. B. Expression ofneuroendocrine markers (neuron specific enolase. synaptophysin and bom-besin) in carcinoma of the lung. Pathol. Res. Pract., 183: 412-417. 1988.Hoog, A.. Gould. V. E.. Grimelius. L., Franke, W. W.. Falkmer, S.. andChejfec. G. Tissue fixation methods alter the immunohistochemical de-monstrability of synaptophysin. Ultrastruct. Pathol., 12: 673-678, 1988.Brower, M., Carney, D. N., Oie, H. K., Gazdar, A. F.. and Minna, J. D.Growth of cell lines and clinical specimens of human non-small cell lungcancer in a serum-free defined medium. Cancer Res.. 46: 798-806. 1986.Gazdar. A. F., Carney, D. N., Russell. E. K.. Sims, H. L., Baylin. S. B.,Bunn. P. A.. Guccion. J. G.. and Minna. J. D. Establishment of continuous,clonable cultures of small-cell carcinoma of the lung which have amineprecursor uptake and decarboxylation cell properties. Cancer Res., 40:3502-3507. 1980.Baylin. S. B.. Stevern, S. A., and Shakir, K. M. Association of diamineoxidase and ornithine decarboxylase with maturing cells in rapidly proliferating epithelium. Biochim. Biophys. Acta, 541: 415-419, 1978.Beaven. M. A., Wilcox, G.. and Terpstra. G. K. A microprocedure for themeasurement of I4CO¡release from ["Cjcarboxyl labelled amino acids. Anal.Biochem.. «4:638-641. 1978.Towbin, H., and Gordon. J. Immunoblotting and dot immunobinding—current status and outlook. J. Immunol. Methods. 72: 313-340, 1984.Cullata. F.. Fedorko, J.. Gu. J.. Lebacq-Verheyden, A. M., Linnoila. R. L,and Battey. J. F. Gastrin-releasing peplide gene-associated peptides areexpressed in normal human fetal lung and small cell lung cancer: a novelpeptide family found in man. J. Clin. Endocrinol. Metab., 67: 576-583,1988.Graziano, S., Mazid, R.. Newman. N.. Tatum. A.. Oler, A.. Mortimer, J. A.,Gullo. J. J., DiFino. S. M.. and Scalzo. A. J. The use of neuroendocrineimmunoperoxidase markers to predict chemotherapy response in patientswith non-smail-cell lung cancer. J. Clin. Oncol.. 7: 1398-1406, 1989.Miettinen, M. Synaptophysin and neurofilament proteins as markers forneuroendocrine tumors. Arch. Pathol. Lab. Med.. ///: 813-818. 1987.Wiedenmann. B., Kühn.C., Schwechheimer. K.. Waldherr, R., Raue. F..Brandeis. W., Kommerell, B.. and Franke. W. Synaptophysin identified inmétastasesof neuroendocrine tumors by immunocytochemistry and immu-noblotting. Am. J. Clin. Pathol., 88: 560-569, 1987.Leubc, R. E.. Kaiser. P.. Seiter, A.. Zimbelmann, R., Franke. W. W.. Rehm.H.. Knaus. P.. Prior. P., Betz, H.. Rcinke. H.. Beyrether. K.. and Wiedenmann. B. Synaptophysin: molecular organization and mRNA expression asdetermined from cloned cDNA. EM BO J., 6: 3261-3268, 1987.Navone, F.. Jahn. R.. Di Gioia. G.. Stukenbrok, H.. Greengard, P.. and DeCamilli. P. Protein p38: an integral membrane protein specific for smallvesicles of neurons and neuroendocrine cells. J. Cell Biol.. 103: 2511-2527.1986.Rehm. H., Wiedenmann. B., and Betz, H. Molecular characterization ofsynaptophysin. a major calcium binding protein of the synaptic vesiclemembrane. EMBO J.. 5: 535-541. 1986.Kivcla. T.. Tarkkanen. A., and Virtanen. I. Synaptophysin in the humanretina and retinoblastoma. Invest. Ophthalmol. Vis. Sci., 30: 212-219. 1989.Thomas. L.. Härtung.K.. Langosch. D.. Rehm. H.. et al. Identification ofsynaptophysin as a hexamcric channel protein of the synaptic vesicle membrane. Science (Washington, DC). 242: 1050-1053. 1988.Pang. D. T.. Wang, J. K. T., ValorÃa,F., Benfenati, F.. and Greengard, P.Protein tyrosine phosphorylation in synaptic vesicles. Proc. Nail. Acad. Sci.USA, 85: 762-766. 1988.
6073
on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

SYNAPTOPHYSIN IN LUNG CANCER CELL LINES
45. Wiedenniann. B.. Rehm. H.. Knicrim. M.. and Becker. C. Fractionalion of medulla. J. Neurochem.. 53: 1043-1049, 1989.synaptophysin-containing vesicles from rat brain and cultured PC12 phco- 48. Schilling. K.. and Grätzel. M. Quantification of p38/synaplophysin in highlychromocytoma cells. FEBS Lett., 240: 71-77, 1988. purified adrenal medullary chromaffin vesicles. FEBS Lett.. 233: 22-24,
46. Lowe. A. W., Madeddu. L., and Kelly, R. B. Endocrine secretory granules 1988.and neuronal synaptic vesicles have three integral membrane proteins in 49. Obendorf. D.. Schwarzenbrunner. L'.. Fischer-Colbrie. R., Laslop, A., and
common. J. Cell Biol.. 106: 51-59. 1988. Winkler. H. In adrenal medulla synaptophysin (protein p38) is present in47. Fournier. S.. Novas. M. L., and Trifaro. J. M. Subcellular distribution of chromaffin granules and in a special vesicle population. J. Neurochem., 51:
65,000 calmodulin-binding protein (p65) and synaptophysin (p38) in adrenal 1573-1580. 1988.
6074
on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1990;50:6068-6074. Cancer Res Sandra M. Jensen, Adi F. Gazdar, Frank Cuttitta, et al. Lung Cancer Cell LinesDecarboxylase as Markers for Neuroendocrine Differentiation in A Comparison of Synaptophysin, Chromogranin, and l-Dopa
Updated version
http://cancerres.aacrjournals.org/content/50/18/6068
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/50/18/6068To request permission to re-use all or part of this article, use this link
on May 22, 2018. © 1990 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























