Embed Size (px)
Citation preview

Investigative Ophthalmology & Visual Science, Vol. 30, No. 2, February 1989Copyright © Association for Research in Vision and Ophthalmology
Synapfophysin in the Human Retinaand Retinoblastoma
An Immunohisrochemical and Western Blotting Study
Tero Kivela,* Ahfi Tarkkanen,* and Ismo Virtanen-|-
Fifty-four formalin-fixed and paraffin-embedded intraocular retinoblastoma specimens and threehuman eyes enucleated because of orbital tumors were studied for the presence of synaptophysin, aneuron-associated integral membrane glycoprotein of presynaptic vesicles, by using the monoclonalantibody SY38. Normal human brain was used as control. In the human retina, synaptophysin-likeimmunoreactivity was present in both plexiform layers, but could not be detected in neuronal peri-karya. However, in reactive retinas present in retinoblastoma eyes, synaptophysin was often observedin perikarya and processes of photoreceptors. Positive neoplastic cells were found in 45 of the 54retinoblastomas. Differentiated tumors tended to contain greater numbers of positive cells than undif-ferentiated ones, a third of which were entirely negative. Identical immunoreactivity was seen in frozenspecimens from human retina and from three retinoblastomas. Using Western blotting, a majorpolypeptide comigrating with human brain synaptophysin was detected in human retina, and a similarbut slightly slower migrating band in retinoblastoma. The results support a primarily neuronal originfor this tumor and point to the possibility that synaptic elements, previously observed in a few cases,may be more frequent in retinoblastoma than had been thought. Invest Ophthalmol Vis Sci 30:212-219, 1989
Synaptophysin is an integral membrane protein ofunknown function originally purified from presyn-aptic vesicles of central nervous system neurons.1"3 Itis a calcium-binding glycoprotein having a relativemolecular weight of 38 kD and a tendency to formhomooligomers.1'2'4 Antibodies to synaptophysinreact with the synaptic regions of the mammalianbrain, retina and spinal cord, where this antigen ispresent in nerve terminals but not in glial or stromalcells.1"3 It is also typical of peripheral neuromuscularjunctions15 and of the majority of cells belonging tothe diffuse neuroendocrine system.1"36 Importantly,synaptophysin appears to provide a useful marker forthe detection of most neuronal and neuroendocrineneoplasms.23-6"10
The possibility that retinoblastoma, the most com-mon intraocular malignant tumor of children, wouldoriginate from multipotential precursor cells of reti-
From the 'Department of Ophthalmology, Helsinki UniversityCentral Hospital, and the tDepartments of Anatomy and Pathol-ogy, University of Helsinki, Helsinki, Finland.
Supported by a grant from the Emil Aaltonen Foundation.Submitted for publication: September 28, 1987; accepted March
7, 1988.Reprint requests: Tero Kivela, MD, Department of Ophthalmol-
ogy, Helsinki University Central Hospital, Haartmaninkatu 4C,SF-00290 Helsinki, Finland.
nal neurons and glia has recently been put forward onthe basis of cell culture studies.""13 In contrast tothis suggestion, however, malignant tumor cells insurgical retinoblastoma specimens do not generallyexhibit glial properties as judged by immunohisto-chemistry.14"21 Although most retinoblastomas arepositive for neuron-specific enolase,15"1720"23 neuro-filament proteins have so far been shown only in aminority of tumors,1821 and it has thus been difficultto provide direct evidence for their neuronal nature.To clarify this important issue further, the presentstudy was undertaken to determine the distributionof synaptophysin in human retina and retinoblas-toma.
Materials and Methods
Histological Specimens
During the years 1962-1987 a total of 71 retino-blastoma specimens from 62 patients have been ex-amined in the Ophthalmic Pathology Laboratory,Department of Ophthalmology, Helsinki UniversityCentral Hospital. Enough tumor for study was stillpresent in 54 formalin-fixed, paraffin-embedded eyesfrom 50 patients. The histopathological characteris-tics of these retinoblastomas are depicted in Table 1.In addition to light microscopically normal-appear-
212
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018

SYNAPT0PHY5IN IN RETINODLASTOMA / Kivela er ol 213
ing parts of the retinas present in the retinoblastomaspecimens, retinas from three identically processednormal human eyes enucleated because of a pre-viously untreated retrobulbar tumor were studied.Formalin-fixed and paraffin-embedded human brainobtained from uninvolved brain areas of an oligo-dendroglioma specimen was used as synaptophysin-positive control. Sections (5 fim thick) were cut fromthe specimens and mounted on chromium-gelatin-treated glass slides to ensure adherence (0.05 g potas-sium chromium(III)sulphate dodecahydrate and 0.5g gelatin in 100 ml distilled water).
Material from three undifferentiated retinoblas-tomas was taken from a file of frozen sections. Partsof these tumors had been frozen immediately afterenucleation, sections (5 fim thick) cut with a cryomi-crotome, air-dried, and stored at —20°C. Frozen sec-tions of undetached human retina, obtained fromthree eyes enucleated from patients with malignantchoroidal melanoma, were also selected from this file.The frozen material was used for indirect immuno-fluorescence microscopy.
Immunohistochemical Staining
Formalin-fixed and paraffin-embedded specimenswere deparaffinized in xylene, rehydrated in an eth-anol series, washed in phosphate-buffered saline(PBS, pH 7.4), and treated with 0.4% pepsin (2500FlP-U/g; E. Merck, Darmstadt, FRG) in 0.01 N hy-drochloric acid at 37°C for 15 min. In preliminarystainings, this procedure was found to reduce back-ground and to enhance the intensity of the specificreaction. Endogenous peroxidase activity was de-stroyed with a 30 min treatment in methanol con-taining 0.5% hydrogen peroxide. The sections werethen incubated with normal horse serum (VectastainABC Kit for Mouse IgG; Vector Laboratories, Bur-lingame, CA; diluted 1:50) in a moist chamber for 30min at room temperature. All immunoreagents werediluted with PBS containing 2% (w/v) bovine serumalbumin (BSA; E. Merck, Darmstadt, FRG). Betweenevery step, the sections were washed for three 10 minchanges in PBS.
The primary mouse monoclonal IgGl antibody tosynapthophysin (Clone SY38) was commercially ob-tained (Lot 10644526-01; Boehringer MannheimBiochemica, Mannheim, FRG) and used at a proteinconcentration of 2 fig/ml (dilution 1:5). An unrelatedmurine monoclonal IgGl antibody (Anti-MonkeyMyelin Basic Protein; Lot 701163; Hybritech Inc.,San Diego, CA) was used as a negative control (pro-tein concentration 4 fig/ml; dilution 1:100). Incuba-tion with the primary antibodies was carried out in amoist chamber at 37°C for 60 min.
Table 1. Distribution of the retinoblastomas studiedinto different histopathological groups
Flexner-Wintersteinerrosettes
Infiltration into thechoroid
Infiltration into theoptic nerve
Unilateral
41%(n = 14)
26%(n = 9)
18%(n = 6)
N = 34
Bilateral
65%(n = 13)
25%(n = 5)
20%(n = 4)
N = 20
Total
50%(n = 27)
26%(n = 14)
19%(n = 10)
N = 54
Subsequently the sections were incubated withbiotinylated horse anti-mouse IgG antiserum (Vec-tastain ABC Kit; diluted 1:200) and then with theavidin-biotinylated peroxidase complex (VectastainABC Kit reagents A and B, both diluted 1:160 andmixed 30 min prior to use) in a moist chamber for 30min at 37°C. The peroxidase reaction was developedwith 40 mg of 3-amino-9-ethylcarbazole (Sigma, St.Louis, MO; dissolved in 12 ml of N,N-dimethylfor-mamide) in 200 ml of 0.05 M sodium acetate buffer(pH 5.0) containing 0.03% hydrogen peroxide. Cov-erslips were mounted with Aquamount (BDH Chem-icals Ltd., Poole, UK).
Immunofluorescence Staining
Frozen sections were hydrated for 30 min in PBSand incubated with the primary monoclonal antibod-
Fig. 1. Synaptophysin in human brain and ocular nerves. (A)The synapse-rich gray matter of normal human brain is labeleddiffusely in a granular pattern. Blood vessels (bv), perikarya ofneurones (n), and glia remain negative (immunoperoxidase, X290).(B) Beaded immunoreactivity (arrowheads) is present in nervesbetween ciliary muscle fibres. Pee, pigmented ciliary epithelium(immunoperoxidase, X290). (C) Similar nerve fibres around bloodvessels (bv) in the choroid (indirect immunofluorescence, X275).
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018
214 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / February 1989 Vol. 30
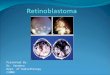
Fig. 2. Synaptophysin in humanretina (immunoperoxidase stain-ing). (A) Retina from an eye enucle-ated because of an orbital tumor.The plexiform layers are specificallylabeled. Neuronal perikarya are neg-ative, although there is slight reactiv-ity in inner processes of some pho-toreceptor cells (X175). (B) In a tan-gential section through the outerplexiform layer cone pedicles (arrow)and rod spherules (arrowheads) reactpositively (haematoxylin counter-stain, X38O). (C) Near ora serratacone perikarya and processes are la-beled adjacent to cystic spaces (c) ofage-related degeneration (X360). (D)Retina from an eye with an uvealmelanoma shows identical immuno-reactivity. The pigment epithelium(arrowhead) shows autofluorescence(indirect immunofluorescence offrozen sections, X280). (E) Retinafrom an eye with retinoblastoma.The perikarya and processes of manyphotoreceptor cells react with theantibody (X290). (F) In another reti-noblastoma eye also processes of bi-polar cells (arrowheads) are visual-ized (X220).
ies (protein concentrations as above) in a moistchamber for 60 min at 37 °C. After three washes inPBS, they were further incubated with fluoresceinisothioeyanate-coupled (FITC-coupled) rabbit anti-serum to mouse immunoglobulins (Lot 34F-8820;Sigma; diluted 1:100). After three washes in PBS, thecoverslips were mounted with PBS-glycerol contain-ing 0.1% (w/v) p-phenylenediamine (Sigma) to re-duce fading of the fluorochrome. The results wereevaluated under a Leitz Diaplan microscopeequipped with an incident light illuminator and thefilter block 12/3 for FITC-fluorescence.
Gel Electrophoresis and Western Blotting
Macroscopically normal-appearing undetachedhuman retina was obtained from an eye with a malig-
nant melanoma of the ciliary body. The eye globe wassectioned immediately after enucleation and a 1 cm2
piece of retina was taken from the posterior pole. Apiece of an undifferentiated retinoblastoma was simi-larly obtained from a 2-year-old girl having unilateraldisease without any family history of retinoblastoma.For control purposes, a piece of gray matter fromhuman brain was dissected from a frozen tissue blockbiopsied from a brownish brain area during an oper-ation for meningioma and originally processed forobtaining diagnostic frozen sections. These showednormal brain except for the presence of hemosiderin-laden phagocytes responsible for the discoloration.
The specimens were homogenized in 10-20 vol-umes of Laemmli's sample buffer,24 incubated for 5min in a boiling water bath, centrifuged for 20 min at12,000 rpm, and stored frozen in aliquots at -20°C.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018

No. 2 SYNAPTOPHYSIN IN RETINODLASTOMA / Kivelo er al 215
Table 2. Presence of tumor cells expressing synaptophysin in the retinoblastomas studied
Retinoblastoma type
Rosettes present
Rosettes absent
Unilateral tumors
Bilateral tumors
All tumors
* Only cells that were apparently
Many
52%(n = 14)
11%(n = 3)
24%(n = 8)
45%.(n = 9)
31%( n = 17)
Number of positive cells*
Moderate
33%(n = 9)
19%(n = 5)
26%(n = 9)
25%(n = 5)
26%(n = 14)
unrelated to infiltrated retina and toareas of necrosis were accepted as positive. Many = over 50%, Moderate
Few
15%(n = 4)
37%(n = 10)
32%( n = 11)
15%(n = 3)
26%(n = 14)
= 50-10%, and Few =physin.
None
0%(n = 0)
33%(n = 9)
18%(n = 6)
15%(n = 3)
17%(n = 9)
under 10% of cells reacting positi
Total
N = 27
N = 27
N = 34
N = 20
N = 54
vely for synapto-
Polyacrylamide gel electrophoresis in the presence ofsodium dodecyl sulphate (SDS-PAGE) was carriedout according to Laemmli24 by using 1.5 mm thick10% slab gels and a Protean II cell (Bio-Rad Labora-tories, Richmond, CA). After SDS-PAGE, the gelswere equilibrated in transfer buffer25 for 15 min andthe protein bands were electrically transferred25 ontoa nitrocellulose membrane (pore size 0.45 /*m;Schleicher & Schiill, Dassel, FRG) at a constant volt-age of 70 V for 3 hr in a Bio-Rad Trans-Blot cell. Themembranes were cut into strips and either stained fortotal protein with amido black25 (Bio-Rad) or im-munostained with the anti-synaptophysin antibody.For immunostaining, nonspecific binding of proteinwas blocked by treating the nitrocellulose strips for 2hr with 3% (w/v) BSA in PBS. Subsequently theywere incubated with the monoclonal anti-synapto-physin antibody at a protein concentration of 2 /ig/mlin a moist chamber for 60 min at 37 °C, washed inPBS, and incubated with peroxidase-coupled rabbitantiserum to mouse immunoglobulins (Lot 096; Da-kopatts a/s, Glostrup, Denmark; diluted 1:1000) for30 min. The peroxidase reaction was developed with10 mg of 4-chloro-l-naphthol (Sigma; dissolved in 3ml of methanol) in 15 ml of PBS containing 0.03%hydrogen peroxide.
Results
Immunohistochemical Stainings
The monoclonal antibody (SY38) to synaptophy-sin labeled in a granular pattern the gray matter of thehuman brain which is known to be rich in nerveterminals containing this antigen1-3 (Fig. 1A). A deli-cate, beaded positive reaction was seen between someciliary muscle fibers, probably residing in sympa-thetic nerves (Fig. IB). Similar positive reactivity wasalso observed accompanying blood vessels in the cho-roid, especially in frozen sections studied by indirectimmunofluoreseence microscopy (Fig. 1C). The
optic and larger ciliary nerves were negative. Omit-ting the primary antibody, the secondary antiserumor the ABC complex resulted in a negative reaction.The control monoclonal IgGl antibody to myelinbasic protein labeled only myelin sheaths andSchwann cells of the optic and ciliary nerves.
In light microscopically normal-appearing humanretinas from eyes enucleated because of orbitaltumors, the monoclonal SY38 antibody reacted withthe plexiform layers (Fig. 2A). The inner plexiformlayer was labeled in a granular pattern and showed, inmany specimens, a stratified arrangement (Fig. 2A).In the outer plexiform layer, both synaptic pedicles ofcones and rod spherules reacted positively (Fig. 2B).The perikarya of retinal neurons were negative forsynaptophysin, except for some photoreceptor cellsclose to the ora serrata, where some age-related cysticdegeneration was observed (Fig. 2C). The frozen sec-tions of retinas from intraocular melanoma patientsstudied by indirect immunofluorescence microscopygave entirely identical results (Fig. 2D).
In several retinoblastoma specimens synaptophy-sin-like immunoreactivity was more widely distrib-uted in the retina. In addition to the plexiform layers,positive reaction could often be seen in the perikaryaand outer processes of photoreceptor cells (Fig. 2E,F). Likewise, short cytoplasmic processes, compati-ble with those of bipolar cells, sometimes reachedfrom the inner plexiform to the inner nuclear layer(Fig. 2F). In one specimen, several neurons in theganglion cell layer had positive granules in their cyto-plasm. Remnants of plexiform layers entrappedwithin the tumor were easily observed in every caseand served as internal controls.
Neoplastic cells positive for synaptophysin wereseen in 45 of the 54 formalin-fixed and paraffin-em-bedded intraocular retinoblastomas studied. Theoverall number of positive cells tended to be greaterin differentiated retinoblastomas (Table 2) and onlynine undifferentiated tumors were entirely negative.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018
216 INVESTIGATIVE OPHTHALMOLOGY 6 VISUAL SCIENCE / February 1989 Vol. 30
. * . * •
• ^
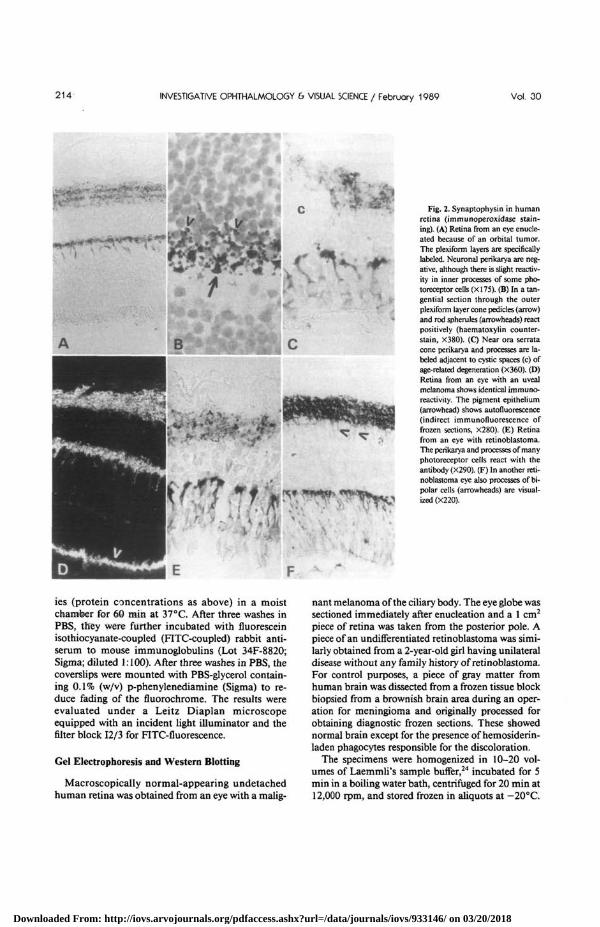
Fig. 3. Synaptophysin in formalin-fixed and paraffin-embedded retinoblastomas (immunoperoxidase staining). (A) Transition fromnormal retina to an undifferentiated retinoblastoma, variably positive for synaptophysin (X30). (B) Many tumor cells of an undifferentiatedretinoblastoma show uneven positive cytoplasmic reaction, whereas stromal areas (s) are negative (X145). (C). Some tumor cells distant fromany part of infiltrated retina have short cytoplasmic processes (arrowheads). A rosette (arrow) is nearby (X28O). (D) Most neoplastic cells of adifferentiated retinoblastoma, particularly those of Flexner-Wintersteiner rosettes, react for synaptophysin (X145). (E) The positive reactionis limited to the apical cytoplasm of rosettes in another differentiated tumor (X68O). (F) A tumor with fleurettes (arrowheads) demonstratespositivity for synaptophysin in their apical cytoplasmic processes (haematoxylin counterstain, X575).
V
There was also significant variation in the labelingintensity of individual neoplastic cells within a tumor(Fig. 3). Stromal elements did not react with theSY38 monoclonal antibody (Fig. 3B). The positivereaction was present as small to relatively coarsegranules, which were generally unevenly distributedin the tumor cell cytoplasm. In other cases the posi-
tive reaction was more evenly present throughout thecytoplasm of undifferentiated retinoblastoma cells.Short cytoplasmic processes reacting positively forsynaptophysin were sometimes seen (Fig. 3C).Flexner-Wintersteiner rosettes were consistently posi-tive (Fig. 3D, E) even when single ones were presentin otherwise undifferentiated tumors. The positive re-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018
No. 2 SYNAPTOPHYSIN IN RETINOBLASTOMA / Kivelo er ol 217
*Jt
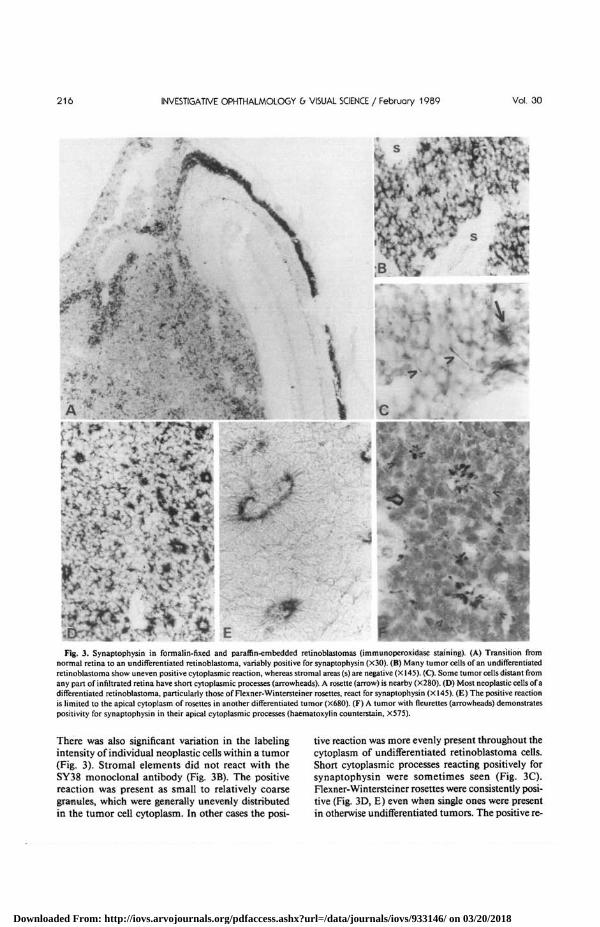
Fig. 4. Synaptophysin inretinoblastoma (indirectimmunofluorescence of fro-zen sections). Phase con-trast figures (A, D) showboth tumors to be undiffer-entiated. (B) Perinuclearimmunoreactivity for syn-aptophysin is observed inmany neoplastic cells(X200). (C) The controlmonoclonal antibodyagainst myelin basic proteindoes not label them (X200).(E) In the second tumor, thepositive reaction is un-evenly distributed in mostretinoblastoma cells (X200).(F) Higher magnificationfrom the same tumor(X510).
A
action was most intense in the apical cytoplasm im-mediately surrounding the lumen of the rosette (Fig.3E). The apical cytoplasmic processes of differen-tiated tumor cells forming fleurettes were also im-munostained (Fig. 3F).
The frozen retinoblastoma specimens studied byindirect immunofluorescence microscopy gaveequivalent results (Fig. 4). In two cases, almost everytumor cell was positive for synaptophysin, while inthe third case moderate numbers of positive tumorcells were observed.
Western Blotting
In homogenates from human retina and from anundifferentiated retinoblastoma, several polypeptidesof identical electrophoretic mobility were observed inprotein staining after transfer onto nitrocellulosemembranes (Fig. 5; Lanes 3 and 4). When parallelnitrocellulose strips were immunostained with theSY38 monoclonal anti-synaptophysin antibody, adistinct immunoreactive polypeptide was observed inthe sample from normal human retina (Fig. 5; Lane6) that comigrated with the band seen in homogenatefrom human brain gray matter (Fig. 5; Lane 5). Asimilar immunoreactive polypeptide with a slightlyslower electrophoretic mobility was revealed in thehomogenate from retinoblastoma (Fig. 5; Lane 7).
Discussion
Consistent with the ultrastructural localization ofsynaptophysin to presynaptic vesicles in the brain,13
synaptophysin was found in the plexiform layers ofthe human retina, which are formed of nerve pro-cesses and synapses between retinal neurones. Whilecone pedicles and rod spherules were unequivocallypositive for synaptophysin, it seems probable, due tothe difiuse granular labeling of the inner plexiformlayer, that most presynaptic terminals of amacrineand bipolar cells reacted positively as well. Identicaldistribution has been reported for the mouse retina.1
However, while synaptophysin-containing nerveswere not mentioned to be present in the mouse cho-roid,1 such elements accompanied human choroidalblood vessels. Some positively reacting nerves werealso present between extraocular and ciliary musclefibers, as has been reported for skeletal and heartmuscle.15
In several retinoblastoma specimens, where piecesof light microscopically normal-appearing retinawere present, the monoclonal antibody to synapto-physin reacted with the perikarya and processes ofphotoreceptors in addition to the plexiform layers.Since similar reactivity was normally seen near theora serrata with cystic degeneration, this could beinterpreted Ss a reactive secondary change. It mightalso be related to electron microscopic studies show-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018

218 INVESTIGATIVE OPHTHALMOLOGY & VISUAL SCIENCE / February 1989 Vol. 30
1 2 4 5
Fig. S. Western blotting of SDS-PAGE separated polypeptidesfrom crude homogenates of human brain gray matter (Lanes 2 and5), normal human retina (Lanes 3 and 6), and an undifferentiatedretinoblastoma (Lanes 4 and 7). Lanes 2-4: Amido black stainingfor protein. Lanes 5-7: Immunostaining for synaptophysin withthe SY38 monoclonal antibody. The major immunoreactive poly-peptide of human retina migrates identically with that detectedamong brain polypeptides, while the immunoreactive band in theretinoblastoma sample migrates slightly slower. Molecular weightstandards (Lane 1; from top to bottom) phosphorylase B (92.5 kD),bovine serum albumin (66.2 kD), ovalbumin (45 kD), carbonicanhydrase (31 kD), and soybean trypsin inhibitor (21.5 kD).
ing that photoreceptor cell synapses in retinoblas-toma eyes may be abnormal.26
Synaptophysin was found in over two-thirds of the54 formalin-fixed and paraffin-embedded retinoblas-toma specimens examined, and in all three retino-blastomas studied as frozen sections. In agreementwith previous results with antisera to neuron-specificenolase,23 there appeared to be a correlation betweentumor differentiation and the number of positivelylabeled retinoblastoma cells (Table 2). An immuno-reactive polypeptide compatible with synaptophysinwas also observed on immunoblots of homogenatesfrom an undifferentiated retinoblastoma. The slightdifference noted in its electrophoretic mobility rela-tive to retinal synaptophysin may be due to alteredglycosylation of this glycoprotein, as has been sug-gested for pheochromocytoma3-4 and paragan-glioma.6 It is worthy of notice that synaptophysin wasconcentrated to the apical cytoplasm of rosettes,which is thought to correspond to the outer limitingmembrane region of photoreceptors rather than totheir synaptic processes.27 Other antigens found inphotoreceptor cells, such as rhodopsin and retinal S-antigen, are distributed in rosettes in a way identical
to their compartmentalization in normal photore-ceptor cells.20-2'-28'29 Although the reason for this dis-crepancy remains unclear, it might result from defec-tive transport of synaptophysin-containing vesiclesdue to lack of processes corresponding to synapticpedicles in tumor cells forming rosettes.27
Synaptophysin is confined to small neurotransmit-ter vesicles with an electron-translucent interior bothin central nervous system neurons and neuroendo-crine cells,1'37 and it is not present in significantamounts in dense-core neurosecretory granules.1*3
While dense-core granules can be found in severalretinoblastomas,27'30"32 small, clear presynaptic vesi-cles are not mentioned in major ultrastructural stud-ies of retinoblastoma.30'31 Nevertheless, in several ret-inoblastomas electron microscopy has disclosed syn-aptic vesicles33"38 and even more-or-less typicalsynaptic lamellae in undifferentiated tumor cells,3536
rosettes34 and fleurettes.37'38 Several neurotransmittercandidates have also been implicated in retinoblas-toma.32'39'40 The current findings add new weight tothese observations and point to the possibility thatsynaptic elements may be more frequent in retino-blastoma than has hitherto been appreciated.
Given the specific expression of synaptophysin inneurons, neuroendocrine cells and their malignantcounterparts, particularly in medulloblastomas andneuroblastomas,7'8 its presence in retinoblastomalends further support for a primarily neuronal originfor this tumor.14'23 Synaptophysin has advantagesover other neuronal markers previously used to studyretinoblastoma. Unlike neuron-specific enolase,41'42
it has not yet been found in nonneuronal, nonneur-oendocrine tissues or tumors.1"10 It is also expressedin the majority of retinoblastomas, whereas antibod-ies to neurofilaments have rarely reacted even withdifferentiated tumors.1821 Neurofilament antibodiesdo label, however, cultured Y79 retinoblastoma cellstreated with differentiating agents.12 Synaptophysinmay prove to be a particularly valuable marker whenstudying neuronal properties of retinoblastoma cells.
Key words: retina, retinoblastoma, synaptophysin, presyn-aptic vesicles, immunohistochemistry
Acknowledgments
The authors are very grateful to docent Matti Haltia forkindly assisting in obtaining control material from humanbrain biopsies. The diligent and meticulous technical assis-tance of Mrs. Marjatta Koikkalainen is acknowledged withsincere gratitude.
References
1. Wiedenmann B and Franke WW: Identification and localiza-tion of synaptophysin, an integral membrane glycoprotein ofM, 38,000 characteristic of presynaptic vesicles. Cell 41:1017,1985.
2. Jahn R, Schiebler W, Ouimet C, and Greengard P: A 38,000-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018

No. 2 5YNAPTOPHYSIN IN RETINOBLASTOMA / Kivela er ol 219
dalton membrane protein (p38) present in synaptic vesicles.Proc Natl Acad Sci USA 82:4137, 1985.
3. Navone F, Jahn R, di Gioia G, Stukenbrok H, Greengard P,and de Camilli P: Protein p38: An integral membrane proteinspecific for small vesicles of neurons and neuroendocrine cells.J Cell Biol 103:2511, 1986.
4. Rehm H, Wiedenmann B, and Betz H: Molecular character-ization of synaptophysin, a major calcium-binding protein ofthe synaptic vesicle membrane. EMBO J 5:535, 1986.
5. Metz J, Gerstheimer FP, and Herbst M: Distribution of syn-aptophysin immunoreactivity in guinea pig heart. Histochem-istry 86:221, 1986.
6. Wiedenmann B, Franke WW, Kuhn C, Moll R, and GouldVE: Synaptophysin: A marker protein for neuroendocrine cellsand neoplasms. Proc Natl Acad Sci USA 83:3500, 1986.
7. Gould VE, Wiedenmann B, Lee I, Schwechheimer K, Dock-horn-Dworniczar B, Radosevich JA, Moll R, and Franke WW:Synaptophysin expression in neuroendocrine neoplasms asdetermined by immunocytochemistry. Am J Pathol 126:243,1987.
8. Schwechheimer K, Wiedenmann B, and Franke WW: Synap-tophysin: A reliable marker for medulloblastomas. VirchowsArch (Pathol Anat) 411:53, 1987.
9. Collins VP: Pineocytoma with neuronal differentiation dem-onstrated immunocytochemically: A case report. Acta PatholMicrobiol Immunol Scand (A) 95:113, 1987.
10. Miettinen M: Synaptophysin and neurofilament proteins asmarkers for neuroendocrine tumors. Arch Pathol Lab Med111:813, 1987.
11. Kyritsis AP, Tsokos M, Triche TJ, and Chader GJ: Retinoblas-toma—origin from a primitive neuroectodermal cell? Nature307:471, 1984.
12. Kyritsis AP, Tsokos M, Triche TJ, and Chader GJ: Retinoblas-toma: A primitive tumor with multipotential characteristics.Invest Ophthalmol Vis Sci 27:1760, 1986.
13. Jiang Q, Lim R, and Blodi FC: Dual properties of culturedretinoblastoma cells: Immunohistochemical characterizationof neuronal and glial markers. Exp Eye Res 39:207, 1984.
14. Lane JC and Klintworth GK: A study of astrocytes in retino-blastomas using the immunoperoxidase technique and anti-bodies to glial fibrillary acidic protein. Am J Ophthalmol95:197, 1983.
15. Terenghi G, Polak JM, Ballesta J, Cocchia D, Michetti F, DahlD, Marangos PJ, and Garner A: Immunocytochemistry of neu-ronal and glial markers in retinoblastoma. Virchows Arch(Pathol Anat) 404:61, 1984.
16. Molnar ML, Stefansson K, Marton LS, Tripathi RS, and Mol-nar GK: Immunohistochemistry of retinoblastomas inhumans. Am J Ophthalmol 97:301, 1984.
17. Messmer EP, Font RL, Kirkpatrick JB, and Hopping W: Im-munohistochemical demonstration of neuronal and astrocyticdifferentiation in retinoblastoma. Ophthalmology 92:167,1985.
18. Kivela T, Tarkkanen A, and Virtanen I: Intermediate fila-ments in the human retina and retinoblastoma: An immuno-histochemical study of vimentin, glial fibrillary acidic protein,and neurofilaments. Invest Ophthalmol Vis Sci 27:1075, 1986.
19. Kivela' T and Tarkkanen A: S-100 protein in retinoblastomarevisited. An immunohistochemical study. Acta Ophthalmol(Copenh) 64:664, 1986.
20. Rodrigues MM, Wiggert B, Shields J, Donoso L, BardensteinD, Katz N, Friendly D, and Chader G: Retinoblastoma: Im-munohistochemistry and cell differentiation. Ophthalmology94:378. 1987.
21. Perentes E, Herbort CP, Rubinstein LJ, Herman MM, Uffer S,
Donoso LA, and Collins VP: Immunohistochemical character-ization of human retinoblastomas in situ with multiplemarkers. Am J Ophthalmol 103:647, 1987.
22. Nakajima T, Kameya T, Tsumuraya M, Shimosato Y, andKato K: Enolase distribution in human brain tumors, retino-blastomas, and pituitary adenomas. Brain Res 308:215, 1984.
23. Kivela1 T: Neuron-specific enolase in retinoblastoma: An im-munohistochemical study. Acta Ophthalmol (Copenh) 64:19,1986.
24. Laemmli UK: Cleavage of structural proteins during the as-sembly of the head of bacteriophage T4. Nature 227:680,1970.
25. Towbin H, Staehelin T, and Gordon J: Electrophoretic transferof proteins from polyacrylamide gels to nitrocellulose sheets:Procedure and some applications. Proc Natl Acad Sci USA76:4350, 1979.
26. Nakao F and Uga S: Electron microscopic studies on retino-blastoma: 1. On the fine structure of the retina distant fromtumor masses. Nippon Ganka Gakkai Zasshi 77:1072, 1973.
27. Tso MOM: Clues to the cells of origin in retinoblastoma. IntOphthalmol Clin 20:2:191, 1980.
28. Donoso LA, Hamm H, Dietzschold B, Augsburger JJ, ShieldsJA, and Arbizo V: Rhodopsin and retinoblastoma: A mono-clonal antibody histopathologic study. Arch Ophthalmol104:111, 1986.
29. Mirshahi M, Boucheix C, Dhermy P, Haye C, and Faure J-P:Expression of the photoreceptor-specific S-antigen in humanretinoblastoma. Cancer 57:1497, 1986.
30. Popoff NA and Ellsworth RM: The fine structure of retinoblas-toma. In vivo and in vitro observations. Lab Invest 25:389,1971.
31. Albert DM, Craft J, and Sang DN: Ultrastructure of retinoblas-toma: Transmission and scanning electron microscopy. In Oc-ular and Adnexal Tumors, Jakobiec FA, editor. Birmingham,Aesculapius Publishing Company, 1978, pp. 157-171.
32. Sang DN and Albert DM: Catecholamine levels in retinoblas-toma. In Ocular and Adnexal Tumors, Jakobiec FA, editor.Birmingham, Aesculapius Publishing Company, 1978, pp.172-180.
33. Choux R, Tripier MF, Berard M, Hassoun J, and Toga M:Ultrastructure d'une tumeur de la retine. Bull Cancer (Paris)59:301, 1972.
34. Ohkuma M, Amemiya T, and Koyama S: Synapse-like struc-tures in retinoblastoma. Nippon Ganka Gakkai Zasshi78:1392, 1974.
35. Radnot M: Synaptic lamellae in retinoblastoma. Am J Oph-thalmol 79:393, 1975.
36. Radnot M: On the histogenesis of retinoblastoma. J PediatrOphthalmol 12:259, 1975.
37. Dickson DH, Ramsey MS, and Tonus JG: Synapse formationin retinoblastoma tumours. Br J Ophthalmol 60:371, 1976.
38. Radnot M: Photorezeptorelemente in Retinoblastomen.Graefes Arch Klin Exp Ophthalmol 202:63, 1977.
39. Sano T, Saito H, Saito S, Kagawa N, and Hizawa K: Somato-statin-immunoreactive cells in human retina and retinoblas-toma. Biomed Res 2:709, 1981.
40. Tarkkanen A, Tervo T, Tervo K, Eranko L, and Cuello AC:Substance P immunoreactivity in normal human retina and inretinoblastoma. Ophthalmic Res 15:300, 1983.
41. Haimoto H, Takahashi Y, Koshikawa T, Nagura H, and KatoK: Immunohistochemical localization of y-enolase in normalhuman tissues other than nervous and neuroendocrine tissues.Lab Invest 52:257, 1985.
42. Vinores SA, Bonnin JM, Rubinstein LJ, and Marangos PJ:Immunohistochemical demonstration of neuron-specific eno-lase in neoplasms of the CNS and other tissues. Arch PatholLab Med 108:536, 1984.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933146/ on 03/20/2018