Embed Size (px)
Citation preview

17TH EUROPEAN NITROGEN-CYCLE MEETING (ENC2012)
2266--2288 SSeepptteemmbbeerr 22001122
OOsslloo,, NNoorrwwaayy
PROGRAMME AND ABSTRACT BOOK

1
17th European Nitrogen Cycle Meeting, Oslo 2012
Contents
Organisation ................................................................................................................................................ 2
Travel Information ...................................................................................................................................... 3
Programme ................................................................................................................................................. 4
Wednesday, 26 September 2012 ............................................................................................................ 4
Thursday, 27 September 2012 ................................................................................................................ 5
Friday, 28 September 2012 ..................................................................................................................... 8
Posters....................................................................................................................................................... 11
Abstracts ................................................................................................................................................... 13
Presentation abstracts .......................................................................................................................... 14
Poster abstracts .................................................................................................................................... 50
Delegate list .............................................................................................................................................. 70

2
17th European Nitrogen Cycle Meeting, Oslo 2012
Organisation
Local organising committee
Åsa Frostegård1
Natalie Lim1
Lars Bakken2
Peter Dörsch2
Norwegian University of Life Sciences 1 Department of Chemistry, Biotechnology and Food Science (IKBM)
Christian Magnus Falsens vei 1, N-1432 Ås, Norway
2 Department of Plant and Environmental Sciences (IPM)
Fougnerbakken 3, N-1430 Ås, Norway
Scientific committee
David Richardson (University of East Anglia / UK)
Rob van Spanning (VU University Amsterdam / NL)
Mike Jetten (Radboud University Nijmegen / NL)
Jeff Cole (University of Birmingham / UK)
Lars Bakken (Norwegian University of Life Sciences / NO)
Åsa Frostegård (Norwegian University of Life Sciences / NO)
Peter Dörsch (Norwegian University of Life Sciences / NO)

3
17th European Nitrogen Cycle Meeting, Oslo 2012
Travel Information
Arrival / Departure by air
Oslo has three airports (below). For information on
travel in the Oslo region:
http://www.visitoslo.com/en/transport/transport-
airport/
Oslo Airport Gardermoen (OSL)
Oslo’s main airport is located 47 km northeast of
Oslo. There are express trains with frequent
departures to the Oslo Central Station. Follow the
signs at the airport to find the express train.
Moss Airport Rygge (RYG)
Rygge airport is located 66 km south-east of Oslo.
Take “Rygge-ekspressen” to the main bus terminal
in Oslo, Oslo Bussterminal.
If you arrive on Wednesday afternoon, take either
“Rygge-ekspressen” or “Flybussekspressen”
(towards Oslo), and alight at the stop
“Korsegården” which is closest to the university.
Upon arrival, please contact one of the organisers
to be picked up there.
Sandefjord Airport Torp (TRF)
110 km south-west of Oslo, about 2 hours by
bus/train. Bus transport to the main bus terminal
in Oslo, Oslo Bussterminal, is available at the
airport.
Public transport
You can plan your travel by public transport
(including bus, train, tram, metro and ferry) on
http://ruter.no/en/. Connecting services are
shown on the website between the departure and
arrival point for any two points in Oslo and
Akershus. The website also has maps, timetables
and ticket fares.
Venue: Thon Hotel Opera
Thon Hotel Opera is situated across the street
from Oslo Central Station and 10 minutes’
walking distance from Oslo Bus Terminal (see
map below).
For more information about Thon Hotel Opera
(including driving directions), please see:
http://www.thonhotels.com/hotels/countrys/nor
way/oslo/thon-hotel-opera/

4
17th European Nitrogen Cycle Meeting, Oslo 2012
Programme
Wednesday, 26 September 2012
Location: Norwegian University of Life Sciences (UMB), Ås
1700 Shuttle bus from Thon Hotel Opera to UMB
1730 Registration, drinks and snacks
1900 Welcome address, Lars Bakken
1910
PT 01
The necessity of combining molecular biology / biochemistry with microbial ecology to
gain progress in the study of nitrogen cycling
Mary Firestone (UC Berkeley / US)
1940
PT 02
Nitrogen and atmosphere - perspectives from the European Nitrogen Assessment
(ENA)
Klaus Butterbach-Bahl (Karlsruhe Institute of Technology / DE)
2010 Dinner
2300 Departure by bus to Thon Hotel Opera, Oslo
- End of Day 1 -
5
17th European Nitrogen Cycle Meeting, Oslo 2012
Programme
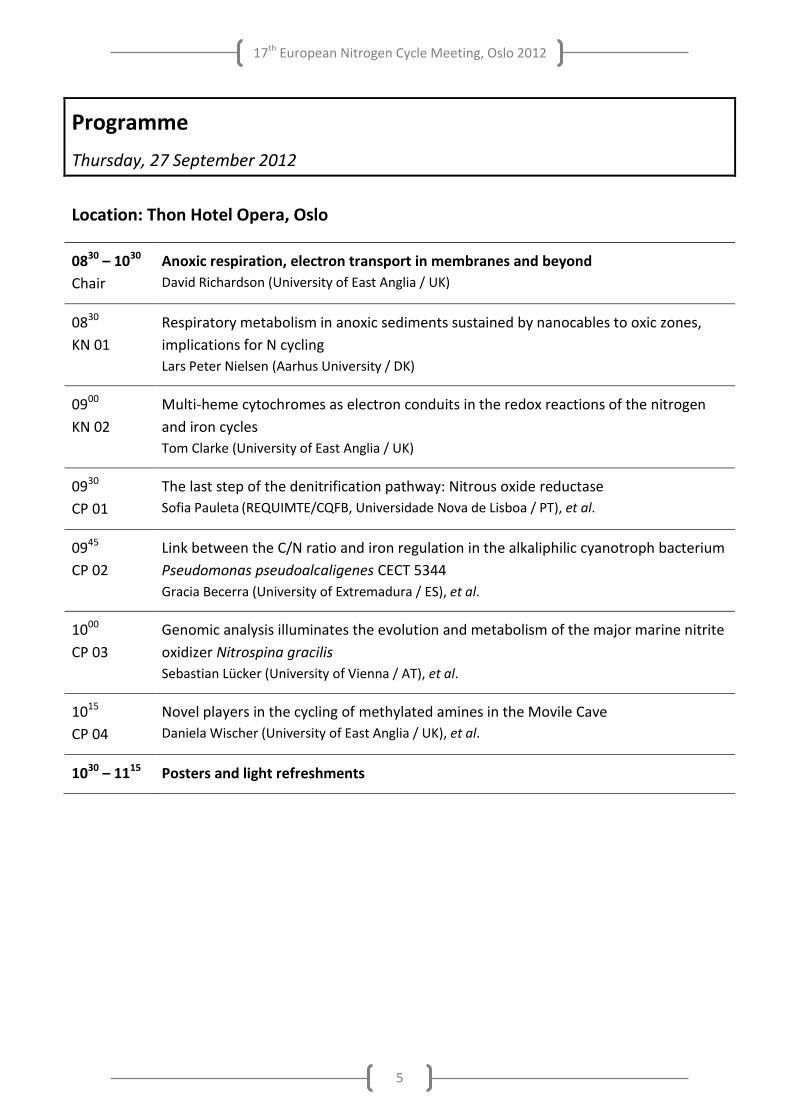
Thursday, 27 September 2012
Location: Thon Hotel Opera, Oslo
0830 – 1030
Chair
Anoxic respiration, electron transport in membranes and beyond
David Richardson (University of East Anglia / UK)
0830
KN 01
Respiratory metabolism in anoxic sediments sustained by nanocables to oxic zones,
implications for N cycling
Lars Peter Nielsen (Aarhus University / DK)
0900
KN 02
Multi-heme cytochromes as electron conduits in the redox reactions of the nitrogen
and iron cycles
Tom Clarke (University of East Anglia / UK)
0930
CP 01
The last step of the denitrification pathway: Nitrous oxide reductase
Sofia Pauleta (REQUIMTE/CQFB, Universidade Nova de Lisboa / PT), et al.
0945
CP 02
Link between the C/N ratio and iron regulation in the alkaliphilic cyanotroph bacterium
Pseudomonas pseudoalcaligenes CECT 5344
Gracia Becerra (University of Extremadura / ES), et al.
1000
CP 03
Genomic analysis illuminates the evolution and metabolism of the major marine nitrite
oxidizer Nitrospina gracilis
Sebastian Lücker (University of Vienna / AT), et al.
1015
CP 04
Novel players in the cycling of methylated amines in the Movile Cave
Daniela Wischer (University of East Anglia / UK), et al.
1030 – 1115 Posters and light refreshments
6
17th European Nitrogen Cycle Meeting, Oslo 2012
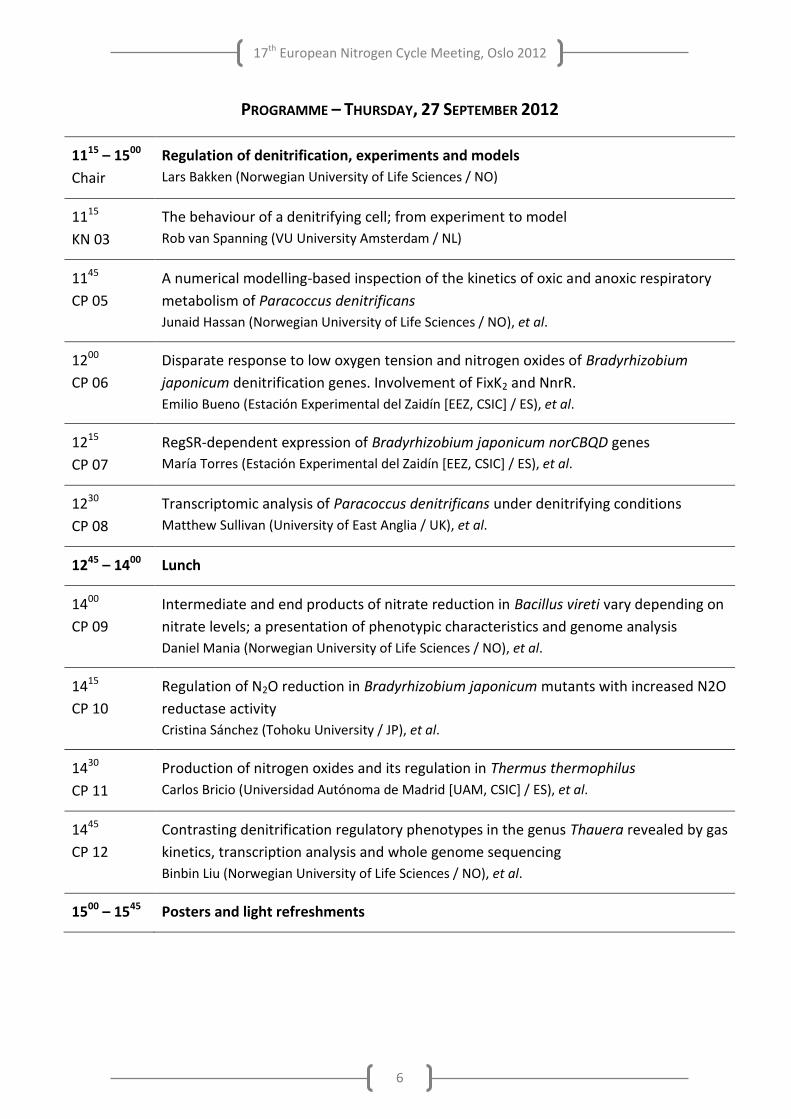
PROGRAMME – THURSDAY, 27 SEPTEMBER 2012
1115 – 1500
Chair
Regulation of denitrification, experiments and models
Lars Bakken (Norwegian University of Life Sciences / NO)
1115
KN 03
The behaviour of a denitrifying cell; from experiment to model
Rob van Spanning (VU University Amsterdam / NL)
1145
CP 05
A numerical modelling-based inspection of the kinetics of oxic and anoxic respiratory
metabolism of Paracoccus denitrificans
Junaid Hassan (Norwegian University of Life Sciences / NO), et al.
1200
CP 06
Disparate response to low oxygen tension and nitrogen oxides of Bradyrhizobium
japonicum denitrification genes. Involvement of FixK2 and NnrR.
Emilio Bueno (Estación Experimental del Zaidín [EEZ, CSIC] / ES), et al.
1215
CP 07
RegSR-dependent expression of Bradyrhizobium japonicum norCBQD genes
María Torres (Estación Experimental del Zaidín [EEZ, CSIC] / ES), et al.
1230
CP 08
Transcriptomic analysis of Paracoccus denitrificans under denitrifying conditions
Matthew Sullivan (University of East Anglia / UK), et al.
1245 – 1400 Lunch
1400
CP 09
Intermediate and end products of nitrate reduction in Bacillus vireti vary depending on
nitrate levels; a presentation of phenotypic characteristics and genome analysis
Daniel Mania (Norwegian University of Life Sciences / NO), et al.
1415
CP 10
Regulation of N2O reduction in Bradyrhizobium japonicum mutants with increased N2O
reductase activity
Cristina Sánchez (Tohoku University / JP), et al.
1430
CP 11
Production of nitrogen oxides and its regulation in Thermus thermophilus
Carlos Bricio (Universidad Autónoma de Madrid [UAM, CSIC] / ES), et al.
1445
CP 12
Contrasting denitrification regulatory phenotypes in the genus Thauera revealed by gas
kinetics, transcription analysis and whole genome sequencing
Binbin Liu (Norwegian University of Life Sciences / NO), et al.
1500 – 1545 Posters and light refreshments
7
17th European Nitrogen Cycle Meeting, Oslo 2012
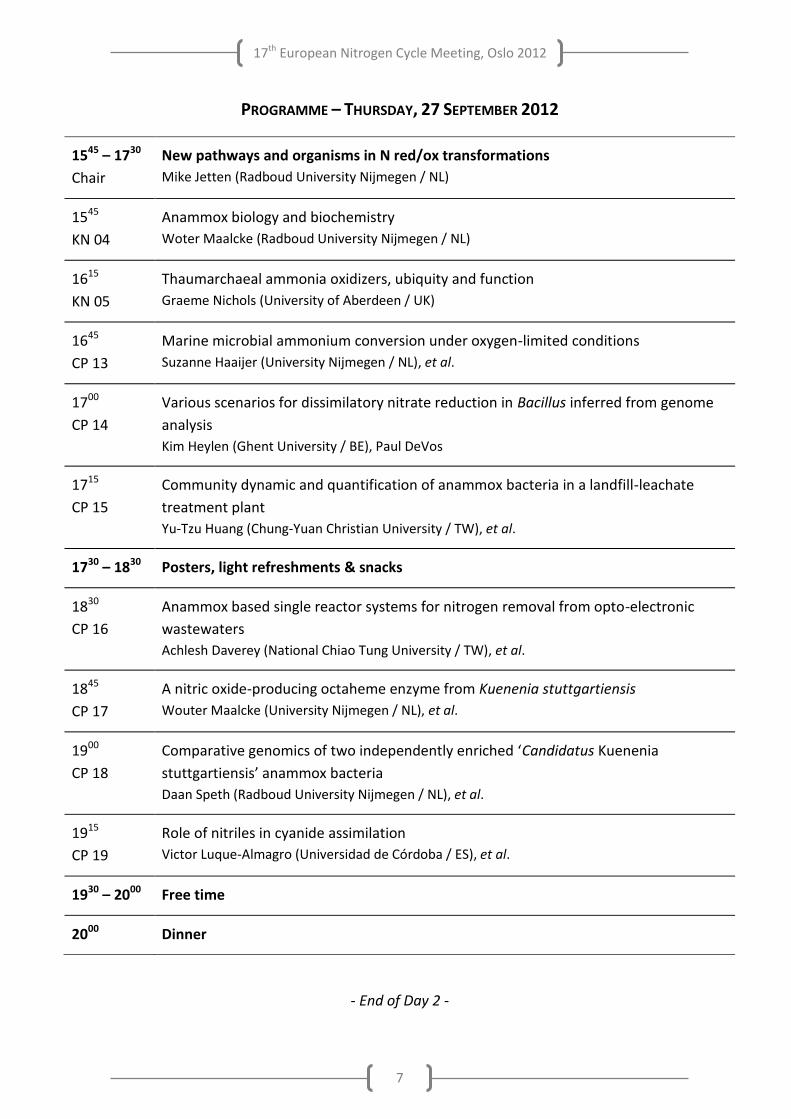
PROGRAMME – THURSDAY, 27 SEPTEMBER 2012
1545 – 1730
Chair
New pathways and organisms in N red/ox transformations
Mike Jetten (Radboud University Nijmegen / NL)
1545
KN 04
Anammox biology and biochemistry
Woter Maalcke (Radboud University Nijmegen / NL)
1615
KN 05
Thaumarchaeal ammonia oxidizers, ubiquity and function
Graeme Nichols (University of Aberdeen / UK)
1645
CP 13
Marine microbial ammonium conversion under oxygen-limited conditions
Suzanne Haaijer (University Nijmegen / NL), et al.
1700
CP 14
Various scenarios for dissimilatory nitrate reduction in Bacillus inferred from genome
analysis
Kim Heylen (Ghent University / BE), Paul DeVos
1715
CP 15
Community dynamic and quantification of anammox bacteria in a landfill-leachate
treatment plant
Yu-Tzu Huang (Chung-Yuan Christian University / TW), et al.
1730 – 1830 Posters, light refreshments & snacks
1830
CP 16
Anammox based single reactor systems for nitrogen removal from opto-electronic
wastewaters
Achlesh Daverey (National Chiao Tung University / TW), et al.
1845
CP 17
A nitric oxide-producing octaheme enzyme from Kuenenia stuttgartiensis
Wouter Maalcke (University Nijmegen / NL), et al.
1900
CP 18
Comparative genomics of two independently enriched ‘Candidatus Kuenenia
stuttgartiensis’ anammox bacteria
Daan Speth (Radboud University Nijmegen / NL), et al.
1915
CP 19
Role of nitriles in cyanide assimilation
Victor Luque-Almagro (Universidad de Córdoba / ES), et al.
1930 – 2000 Free time
2000 Dinner
- End of Day 2 -

8
17th European Nitrogen Cycle Meeting, Oslo 2012
Programme
Friday, 28 September 2012
Location: Thon Hotel Opera, Oslo
0830 – 1000
Chair
NO- biology, toxicity, mechanisms of reduction, role in regulation
Jeff Cole (University of Birmingham / UK)
0830
KN 06
Anaerobic bacterial response to NO generation, damage and repair
Charlene Bradley (University of Birmingham / UK), et al.
0900
CP 20
Nitric oxide detoxification and nitrate assimilation in Bradyrhizobium japonicum
Juan Cabrera (Estación Experimental del Zaidín [EEZ, CSIC] / ES), et al.
0915
CP 21
Nitrosative stress response in Paracoccus denitrificans
Katherine Hartop (University of East Anglia / UK), et al.
0930
CP 22
The production and detoxification of a potent cytotoxin, nitric oxide, by pathogenic
enteric bacteria
Sebastian Runkel (University of East Anglia / UK), et al.
0945
CP 23
Kinetics of NO scavenging in oxic and anoxic conditions, experimental approaches to
differentiate between chemical and enzymatic degradation
Shahid Nadeem (Norwegian University of Life Sciences / NO), et al.
1000 – 1100 Posters and light refreshments
9
17th European Nitrogen Cycle Meeting, Oslo 2012
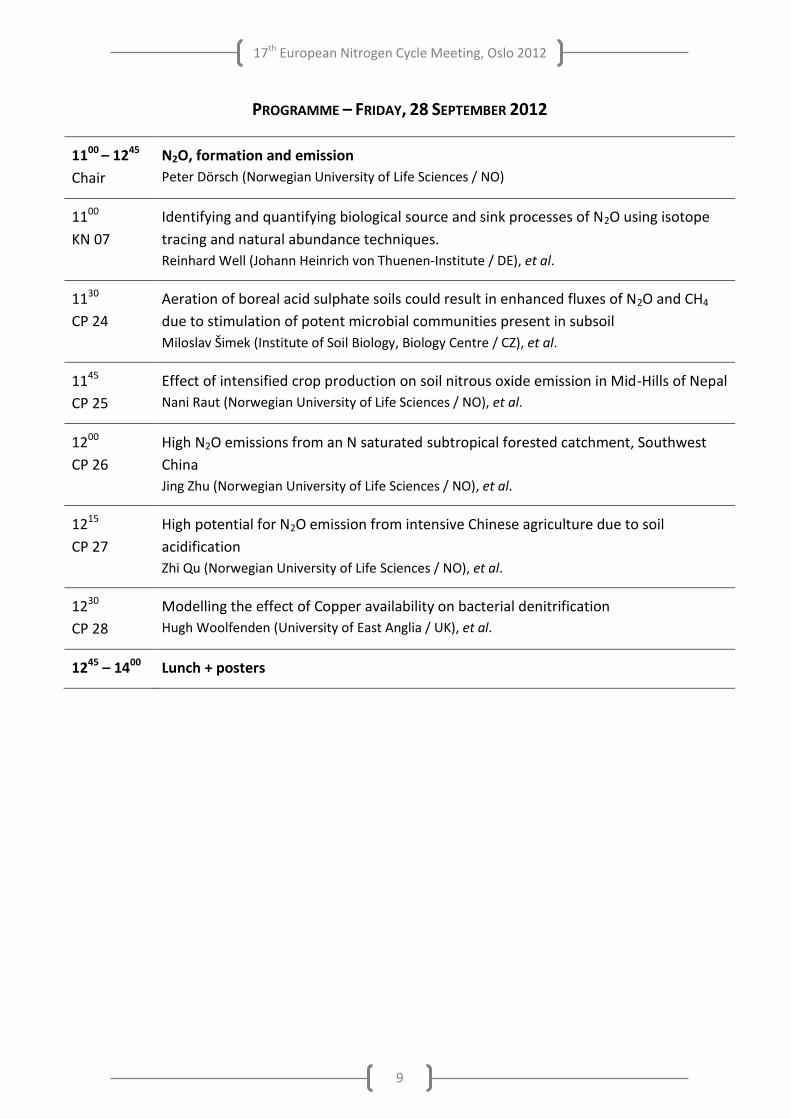
PROGRAMME – FRIDAY, 28 SEPTEMBER 2012
1100 – 1245
Chair
N2O, formation and emission
Peter Dörsch (Norwegian University of Life Sciences / NO)
1100
KN 07
Identifying and quantifying biological source and sink processes of N2O using isotope
tracing and natural abundance techniques.
Reinhard Well (Johann Heinrich von Thuenen-Institute / DE), et al.
1130
CP 24
Aeration of boreal acid sulphate soils could result in enhanced fluxes of N2O and CH4
due to stimulation of potent microbial communities present in subsoil
Miloslav Šimek (Institute of Soil Biology, Biology Centre / CZ), et al.
1145
CP 25
Effect of intensified crop production on soil nitrous oxide emission in Mid-Hills of Nepal
Nani Raut (Norwegian University of Life Sciences / NO), et al.
1200
CP 26
High N2O emissions from an N saturated subtropical forested catchment, Southwest
China
Jing Zhu (Norwegian University of Life Sciences / NO), et al.
1215
CP 27
High potential for N2O emission from intensive Chinese agriculture due to soil
acidification
Zhi Qu (Norwegian University of Life Sciences / NO), et al.
1230
CP 28
Modelling the effect of Copper availability on bacterial denitrification
Hugh Woolfenden (University of East Anglia / UK), et al.
1245 – 1400 Lunch + posters
10
17th European Nitrogen Cycle Meeting, Oslo 2012
PROGRAMME – FRIDAY, 28 SEPTEMBER 2012
1400 – 1600
Chair
N transforming communities; functions and environmental controls
Åsa Frostegård (Norwegian University of Life Sciences / NO)
1400
CP 29
Unexpected diversity of nitrite oxidoreductase genes (nxrB) in marine waters:
indications for novel NOB?
Andreas Schramm (Aarhus University / DK), et al.
1415
CP 30
Biogeochemical nitrogen cycling in Arctic snowpacks: generating hypotheses from
multi-disciplinary studies
Catherine Larose (Université de Lyon / FR), et al.
1430
CP 31
Is there a direct link between denitrifier community structure or size and
denitrification activity in arable soil?
Miriam Herold (University of Aberdeen / UK), et al.
1445
CP 32
Microbial composition of various components of closed aquaculture systems: analysis
of biofilters and indogenous microbiota of fish intestines
Maartje van Kessel (Radboud University Nijmegen / NL), et al.
1500 – 1530 Refreshments
1530
CP 33
Differences in composition of active and overall denitrifier communities in three
organic soils determine responses of denitrification to pH
Kristof Brenzinger (Max Planck Institute for Terrestrial Microbiology / DE), et al.
1545
CP 34
Unusual acid-tolerant denitrifier communities in arctic permafrost soils associated with
N2O fluxes
Katharina Palmer (University of Bayreuth / DE), Marcus Horn
1600 Summing up and closing remarks
End of conference
- End of Day 3 -
11
17th European Nitrogen Cycle Meeting, Oslo 2012
Posters
P 01 Impact of loss of norEF in Rhodobacter sphaeroides 2.4.1
Linda Bergaust (Norwegian University of Life Sciences / NO), et al.
P 02 Abundance and diversity of nitrifying prokaryotes in drinking water treatment plant
Mei-Chun Chen (National Cheng Kung University / TW), et al.
P 03 The nitrite:nitrate oxidoreductase system of the anammox bacterium Kuenenia
stuttgartiensis
Naomi de Almeida (Radboud University Nijmegen / NL), et al.
P 04 Genetic and symbiotic diversity of rhizobia isolated from Ethiopian soils
Tulu Degefu (Norwegian University of Life Sciences / NO), et al.
P 05 Diversity of nitrogen fixing microorganisms from a saline-alkaline former lake Texcoco,
México
Cristina Domínguez-Mendoza (Cinvestav / MX), et al.
P 06 Strong reduction of N2O emissions by long term liming in a drained organic soil
Peter Dörsch (Norwegian University of Life Sciences / NO), et al.
P 07 Identification of key player groups of denitrifiers in an organic agricultural soil by high-
throughput sequencing techniques
Binbin Liu (Norwegian University of Life Sciences / NO), et al.
P 08 The effect of pH and oxygen on nitrous oxide emissions from the denitrifying prokaryote
Paracoccus denitrificans 1222
George Giannopoulos (University of East Anglia / UK), et al.
P 09 Identification of controlling factors of Nitrous Oxide emission for a drained spruce forest on
peat soil, a modeling approach
Hongxing He (University of Gothenburg / SE), et al.
P 10 Study of nitrification inhibitors on ammonia oxidizing archaeal community
Yu-Zen Huang (National Cheng Kung University / TW), et al.
P 11 Regulatory effect of soil properties on N2O emission from wheat-growing season in five
soils: field and pot experiment
Ulrike Lebender (Research Center Hanninghof, YARA GmbH & Co. KG / DE), Mehmet Senbayram

12
17th European Nitrogen Cycle Meeting, Oslo 2012
POSTERS
P 12 Denitrifying bacteria produce much N2O at low pH, and we are beginning to understand
why
Åsa Frostegård (Norwegian University of Life Sciences / NO), et al.
P 13 Nitrous oxide turnover in the nitrate-ammonifying Epsilonproteobacterium Wolinella
succinogenes
Monique Luckmann (Technische Universität Darmstadt / DE), Jörg Simon
P 14 Spatial patterns of soil pH and N2O emissions in field experiments
Aina Lundon (Norwegian Institute for Agricultural and Environmental Research / NO), et al.
P 15 A nitric oxide-producing octaheme enzyme from Kuenenia stuttgartiensis
Wouter Maalcke (Radboud University Nijmegen, The Netherlands), et al.
P 16 The purple form of Marinobacter hydrocarbonoclasticus nitrous oxide reductase
Sofia Pauleta (REQUIMTE/CQFB, Universidade Nova de Lisboa, Portugal), et al.
P 17 Autotrophic carbon dioxide fixation via the Calvin-Benson-Bassham cycle by the
denitrifying anaerobic methanotroph Candidatus Methylomirabilis oxyfera
Olivia Rasigraf (Radboud University Nijmegen / NL), et al.
P 18 Isotopomer ratios of N2O produced during denitrification by fungal pure cultures and
associated oxygen exchange with water
Lena Rohe (Johann Heinrich Thuenen-Institute / DE), et al.
P 19 Regulation of denitrification: pathways to nitrous oxide and dinitrogen
Surinder Saggar (Landcare Research / NZ), et al.
P 20 Off-season N2O losses from clover-rich swards
Ievina Sturite (Norwegian Institute for Agricultural and Environmental Research / NO), et al.

13
17th European Nitrogen Cycle Meeting, Oslo 2012
Abstracts
Presentation abstracts Poster abstracts

14
17th European Nitrogen Cycle Meeting, Oslo 2012
Presentation abstracts
In order of presentation
CP 01
The last step of denitrification pathway: Nitrous oxide reductase
S.R. Pauleta1, S. Dell'Acqua1,2, J.J.G. Moura1, I. Moura1,* 1REQUIMTE/CQFB, Departamento de Química, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, 2829-
516 Caparica, Portugal 2Dipartimento di Chimica, Università di Pavia, Via Taramelli 12, 27100 Pavia, Italy
Email: [email protected]
The final step of bacterial denitrification, the two electron reduction of N2O to N2 is catalyzed by a
multicopper enzyme named nitrous oxide reductase (N2OR). The catalytic center of this enzyme is a
tetranuclear copper site called CuZ, which is unique in biological systems.
The structure of CuZ center opened a novel area of research in metallobiochemistry. In the last
decade, there has been progress in defining the structure of the CuZ center, characterizing the
mechanism of nitrous oxide reduction, and identifying intermediates of this reaction.
The crystal structure of N2OR from Marinobacter hydrocarbonoclasticus was solved to a resolution
of 2.4 Å. This enzyme contains one binuclear (CuA, the electron transfer center) and a tetranuclear
copper center (CuZ, the catalytic center). CuZ center is a new type of cluster, in which four copper
ions are coordinated by seven histidine residues. In addition, the determination of the structure of
the CuZ center allowed a structural interpretation of the spectroscopic data, which was supported by
theoretical calculations. The current knowledge of the structure, function, and spectroscopic
characterization of the nitrous oxide reductase will be addressed in this talk. Although many
questions have been answered about this enzyme it remains a scientific challenge, with many
hypothesis being formed.
Acknowledgements: We thank the many contributions from BIOIN and BIOPROT at REQUIMTE, and FCT-MCTES for
financial support (PTDC/QUI-BIQ/116481/2010).
References: Tavares et al. JIB 100 (2006) 2087-2100. Dell’Acqua et al. JBIC 16 (2011) 183-194. Dell’Acqua et al. JBIC 16
(2011) 1241-1254. Dell’Acqua et al. Philos. Trans. R. Soc. B 367 (2012) 1204-1212

15
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 02
Link between the C/N ratio and iron regulation in the alkaliphilic cyanotroph
bacterium Pseudomonas pseudoalcaligenes CECT 5344
Gracia Becerra, Rafael Blasco, Faustino Merchán, M. Isabel Igeño Departamento de Bioquímica y Biología Molecular y Genética. Facultad de Veterinaria. Universidad de Extremadura.
Avenida de la Universidad s/n.. 10003. Cáceres. Spain
Bacterial ferric uptake regulator (Fur) protein is the major sensor of cellular iron status. A gene
coding for a putative Fur has been identified in the genome of Pseudomonas pseudoalcaligenes CECT
5344. When iron is limiting for growth, Fur derepression increases the cellular capacity for iron
uptake and launches an iron-saving reaction mediated by the small noncoding RNA prrF, homolog to
RyhB, identified in Escherichia coli, where it has been demonstrated that negative regulation of
genes by RyhB is responsible for the positive regulation of some genes by Fur. RyhB RNA negatively
regulates some tricarboxylic acid cycle (TCA) genes, as Fe-S containing fumarase, aconitase and
succinate dehydrogenase, by pairing with their mRNA and causing rapid degradation of the mRNA
(Massé.et al, 2003 Genes Dev17 2374-2383). Computational prediction of PrrF targets in P.
pseudoalcaligenes CECT 5344 allowed to infer as potential targets gltD,B genes encoding for both
subunits of glutamate synthase iron–sulfur containing protein (GOGAT). α–ketoglutarate is a TCA
intermediate and substrate of GOGAT, thus serving as a central link between carbon and nitrogen
metabolism. In this work, we inactivated the fur gene and the resultant mutant showed a slow
growing phenotype and decreased fumarase and aconitase activities. Furthermore, prrF and fiuA
(ferrichrome receptor) expression was constitutive and independent of the iron concentration in the
culture media. Ammonia assimilatory enzymes were also analyzed in the context of Fur regulation.
Work funded by MICINN BIO2011-30026-C02-01, GR10165-CCV019, PRI07A097, FEDER 2007-2013.G. Becerra holds a
Gobierno Extremadura fellowship (Fondo Social Europeo).

16
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 03
Genomic analysis illuminates the evolution and metabolism of the major marine
nitrite oxidizer Nitrospina gracilis
Sebastian Lücker1, Boris Nowka2, Eva Spieck2, Michael Wagner1 and Holger Daims1 1Department of Microbial Ecology, University of Vienna, Austria
2Microbiology and Biotechnology, University of Hamburg, Germany
In marine systems, nitrate is the major reservoir for inorganic nitrogen. The only biological nitrate-
forming reaction is nitrite oxidation, but knowledge of the organisms catalyzing this key process of
the marine nitrogen cycle is limited. Even in oxygen minimum zones (OMZs) a large nitrification
potential is present and nitrite-oxidizing bacteria (NOB) are active and can compete for nitrite. This
recent finding was surprising since nitrite oxidation was believed to be a strictly oxygen-dependent
process. The key marine NOB is Nitrospina gracilis, an aerobic chemolithoautotrophic bacterium
isolated from ocean surface waters, but also abundant in OMZs. To date, few physiological and no
genomic data for this enigmatic organism were available and its phylogenetic affiliation was
uncertain.
To bridge this knowledge gap, we sequenced Nitrospina gracilis Nb-211. Unexpectedly, Nitrospina
lacks the classical genes for reactive oxygen defense mechanisms and utilizes the reverse carboxylate
cycle for carbon fixation. Genes for utilizing simple organic carbon sources were not identified, but
molecular hydrogen might serve as source of low potential electrons. Nitrospina further contains all
genes for gluconeogenesis, oxidative phosphorylation, and reverse electron transport. Interestingly,
comparative genomics revealed a strong evolutionary link to the genus Nitrospira and key genes
involved in nitrite oxidation are highly conserved between Nitrospina, Nitrospira, and anammox
bacteria. This indicates the occurrence of massive horizontal gene transfer at an early evolutionary
stage which spread the nitrite-oxidizing capability among these physiologically different organisms.
Further, detailed analyses using established marker genes finally shed light onto the phylogenetic
affiliation of Nitrospina within the Bacteria.

17
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 04
Novel players in the cycling of methylated amines in Movile Cave
Daniela Wischer1, Yin Chen2 and J. Colin Murrell1 1School of Environmental Sciences, University of East Anglia, Norwich, NR4 7TJ, UK
2School of Life Sciences, University of Warwick, Coventry CV4 7AL, UK
Movile Cave in Southern Romania is an unusual underground ecosystem discovered less than thirty
years ago. Sealed off for over 5.5 million years, the cave harbours a remarkably complex population
of microorganisms, fungi and cave-adapted invertebrates, including many endemic species. In the
absence of light, and with no fixed carbon entering, all life in the cave is sustained by non-
phototrophic, microbial carbon fixation. Extensive microbial mats grow at the redox interface
between the reduced, sulfidic waters and the oxygenated atmosphere. Methylated amines are key
intermediates in the Movile Cave food web, produced during decomposition processes. In addition
to being methylotrophic substrates, methylated amines are also nitrogen source for many non-
methylotrophic bacteria. Combining DNA stable isotope probing with cultivation, we identified active
methylated amine-utilising bacteria in Movile Cave, uncovering novel methylotrophs of the genera
Catellibacterium, Mesorhizobium and Cupriavidus, as well as a diversity of non-methylotrophs
including Acinetobacter, Azospirillum and Rhodobacter as key players. Presently available functional
gene markers for methylamine-utilising bacteria target the mauA gene, encoding methylamine
dehydrogenase. These primers do not detect bacteria using methylamine as a nitrogen source only.
We designed primer sets targeting gmaS, encoding gamma-glutamylmethylamide synthetase, the
first enzyme of the recently characterised indirect methylamine oxidation pathway. The indirect
pathway was prevalent amongst both methylotrophic and non-methylotrophic isolates, as revealed
by PCR-based screening. The mauA gene was detected in a number of methylotrophic isolates,
generally in addition to gmaS. The new primers have been used successfully for identification of
methylamine-utilising bacteria from a range of environments.

18
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 05
A numerical modelling-based inspection of the kinetics of oxic and anoxic
respiratory metabolism of Paracoccus denitrificans
Junaid Hassan1*, I. David Wheat2, Lars R. Bakken1 1 Department of Plant and Environmental Sciences, Norwegian University of Life Sciences, Ås, Norway
2 System Dynamics Group, University of Bergen, Norway
Denitrification sustains respiratory metabolism of some bacteria during anoxic spells. Since most
denitrifying organisms are non-fermentative, the denitrification enzymes must be synthesised using
the energy from oxic respiration. This implies that if the cells fail to synthesise these enzymes prior
to oxygen depletion, they may be entrapped in anoxia. This was hypothesised to be the case for the
majority of Paracoccus denitrificans cells in Bergaust et al’s (AEM 76:6387-6396) experiments. To
further investigate this, we set up a numerical model, which assumes that, once oxygen is low, the
transcription for denitrification enzymes is stochastic with a low probability for the cells to initiate it;
however, once initiated, it undergoes a fast autocatalytic process within each cell. The model
showed a good agreement between the simulated and experimental data for Pd, which lends strong
support to the Bergaust et al’s hypothesis.
The results have implications for interpretation of a number of experiments on regulatory biology of
Pd and possibly other denitrifying organisms. For instance, growth experiments with Pd have
demonstrated a long ‘lag-phase’ after rapid removal of oxygen, followed by a subsequent anoxic
growth (“diauxic lag”). Our simulations suggest that the ‘lag’ was a result of anoxic growth by a
minute fraction of the entire population.
Finally, the model-exercises generated a testable hypothesis that the initial levels of oxygen in a
medium should affect the onset as well as the rate of denitrification; if proven, this would explain
why oxygen is crucial for denitrifying organisms.
The observations indicate a ‘bet hedging strategy’ in P. denitrificans. A large part of the population
‘bets’ that the anoxic spell will be short. They save energy, but loose the bet if the anoxic spell lasts
for a long time.

19
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 06
Disparate response to low oxygen tension and nitrogen oxides of
Bradyrhizobium japonicum denitrification genes. Involvement of FixK2 and NnrR.
Emilio Bueno1, María J Torres1, Eloy F Robles1, Cristina Sánchez1, Tino Krell2, Eulogio J Bedmar1,
Socorro Mesa1, and María J Delgado1 1Departamento de Microbiología del Suelo y Sistemas Simbióticos
2Departamento de Protección Ambiental. Estación Experimental del Zaidín, CSIC, P.O. Box 419, 18080-Granada, España
Bradyrhizobium japonicum can reduce nitrate to molecular nitrogen by the denitrification process in
free-living conditions and in symbiotic association with soybean plants. In B. japonicum,
denitrification depends on napEDABC, nirK, norCBQD and nosRZDYFLX genes which encode the
enzymes nitrate, nitrite, nitric oxide and nitrous oxide reductases, respectively. It has been
previously reported that expression of nap, nirK, and nor genes under denitrifying conditions
depends on the FixLJ-FixK2-NnrR regulatory cascade. Moreover, a genome-wide transcription
profiling of a fixK2 mutant strain grown under free-living micro-oxic conditions showed that nap, nirK,
and nnrR genes, but not nor genes, are targets of the FixK2 regulatory protein. In support of these
observations, we show in this work that purified FixK2 activated transcription from nap and nirK
promoters but not from the nor promoter. In contrast, by using isothermal titration calorimetry,
NnrR bound specifically to the promoter region of the nor genes, but not to those of the nap or nirK
genes. Furthermore, -galactosidase activity of napE-lacZ, nirK-lacZ and norC-lacZ fusions, haem c-
staining analyses and nitrate and nitrite reductase activity assays of cells growing in complete
medium showed that, whereas micro-oxic conditions are sufficient to significantly induce nap and
nirK expression, nor requires low oxygen and the presence of a nitrogen oxide, presumably nitric
oxide. Taken together, these results suggest a disparate regulation of nap, nirK and nor genes of B.
japonicum with regard to their dependence on oxygen and nitrogen oxides. In this control, FixK2
activates expression of nap, nirK and nnrR genes in response to micro-oxia, and NnrR induces
expression of nor genes in response to NO.
Acknowledgements. This work was supported by Fondo Europeo de Desarrollo Regional co-financed grants from Junta
de Andalucía (Spain) [grant number CVI-3177] and from Ministerio de Economía y Competitividad (Spain) [grant
numbers AGL2010-18607 and AGL2011-23383]. Support from the Junta de Andalucıa to Group BIO-275 is also
acknowledged. Emilio Bueno was supported by a Postdoctoral contract from the Consejo Superior de Investigaciones
Cientificas, JAE Programme.

20
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 07
RegSR-dependent expression of Bradyrhizobium japonicum norCBQD genes
María J Torres, Socorro Mesa, Eulogio J Bedmar and María J Delgado
Estación Experimental del Zaidín, C.S.I.C., P. O. Box 419, 18080-Granada, Spain
Bradyrhizobium japonicum is a gram-negative soil bacterium associated symbiotically with soybean
plants which is also able to denitrify under free-living and symbiotic conditions. In this bacterium, the
napEDABC, nirK, norCBQD and nosRZDYFLX genes which encode reductases for nitrate, nitrite, nitric
oxide and nitrous oxide, respectively, are required for denitrification. B. japonicum RegSR proteins
belongs to the family of two-component regulatory systems present in a large number of
proteobacteria and they globally control gene expression, mostly in a redox-responsive manner. In
this work, we have performed transcription profiling of wild-type and regR mutant cells grown under
denitrifying conditions in minimal medium supplemented with succinate as carbon source. The
comparative analyses of WT and regR revealed that expression of nor and nos genes, among others,
is dependent on RegR. Furthermore, by using fluorescently labeled oligonucleotide extension, we
were able to identify two transcriptional start sites located at about 35 (P1) and 22 (P2) bp from the
putative translational start codon of NorC. The areas under the peaks let us to conclude that P2 is
the principal transcription start site and is modulated by RegR. Moreover, qRT-PCR experiments, β-
galactosidase activity assays of a norC-lacZ fusion and haem c staining analyses of regS and regR
mutants, revealed that the RegR control of nor genes is independent of the sensor protein RegS.
Taking together, these results clearly demonstrate the involvement of RegR in the control of nor
genes expression. Up to date, the exact mechanism how RegR exerts this control remains enigmatic.
Acknowledgements. This work was supported by Fondo Europeo de Desarrollo Regional co-financed grants from Junta
de Andalucía (Spain) [grant number CVI-3177] and from Ministerio de Economía y Competitividad (Spain) [grant numbers
AGL2010-18607 and AGL2011-23383]. Support from the Junta de Andalucıa to Group BIO-275 is also acknowledged.
Maria J. Torres was supported by a fellowship from the Consejo Superior de Investigaciones Cientificas JAE Programme.

21
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 08
Transcriptomic analysis of Paracoccus denitrificans under denitrifying conditions
Matthew J Sullivan1, Andrew J Gates1, Gary Rowley1, David J Richardson1 1 CMSB, School of Biological Sciences, University of East Anglia, Norwich Research Park, UK, NR4 7TJ
Molecule for molecule, the greenhouse gas nitrous oxide (N2O) is 300 times more effective at
insulating our climate than is CO2. N2O is very stable (~120 years) and contributes to ~ 9% of global
radiative forcing by greenhouse gases. Approximately 70% of anthropogenic loading of N2O arises
from agricultural practices, primarily through the use of nitrogenous fertilizers (e.g. NH4NO3) by soil
microbes. In the absence of oxygen the model denitrifier, Paracoccus denitrificans, converts nitrate
(NO3-) to di-nitrogen (N2) via four distinct reductases, the last of which is the Cu-containing nitrous
oxide reductase (NosZ). In copper limited conditions P. denitrificans will release gaseous N2O, since
Cu is required for synthesis of functional NosZ. Using batch culture experiments and microarray
analyses, we demonstrate that the structural genes required for N2O reduction, nosZDFLYX, are
subject to a novel form of transcriptional regulation, in addition to their control by the oxygen- and
nitric oxide-sensing FnrP and NnrP. Furthermore, we reveal a sub-set of gene clusters that are up-
regulated in the presence of N2O, some of which may impact significantly on growth in anaerobic
conditions.

22
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 09
Intermediate and end products of nitrate reduction in Bacillus vireti vary depending
on nitrate levels; a presentation of phenotypic characteristics and genome analysis
Daniel Mania1, Kim Heylen2, Ines Verbaendert2, Lars Bakken3 and Åsa Frostegård1 1 Dept Chemistry, Biotechnology and Food Science. Norwegian University of Life Sciences, Norway
2 Dept Biochemistry and Microbiology, Ghent University, Belgium
3 Dept Plant and Environmental Sciences. Norwegian University of Life Sciences, Norway
Endpoint analysis showed that Bacillus vireti LMG 21834Tproduces N2 when grown anaerobically in
presence of nitrate. A more detailed analysis of the gas kinetics using a robotized incubation system
demonstrated that the proportion of nitrate reduced to N2 increased with increasing initial nitrate
levels. At 5 mM initial nitrate, <10% of the nitrate was reduced to N2 while the main part was
reduced to ammonium. The highest proportion of nitrate reduced to N2 was 50%, found at 20 mM
initial nitrate concentration. The cultures reduced all available nitrate to nitrite (up to 50 mM NO2-)
before initiating a reduction of NO2-. In all cases, growth slowed down once all nitrate was respired to
nitrite. B. vireti has an apparently stringent control of NO production with a short peak of NO never
exceeding 800 nM in the liquid. Production of N2O was less controlled and the proportion of N2O
increased with NO3- concentration in the medium. N2O was the predominant endpoint product when
initial NO3- concentrations exceeded 30 mM. Whole genome sequencing confirmed the presence of
the general transcriptional regulator fnr, the narGHJI operon for nitrate reduction and narK encoding
nitrate uptake/nitrite excretion, and narX and narL encoding nitrate sensing regulators. None of the
typical denitrification genes for reduction of nitrate (nir) and NO (nor) were found. The genes
encoding the nitrite reductase subunits nrfA and nrfH, characteristic of DNRA organisms, may release
NO under stress conditions like high nitrite concentrations. An hmp gene was identified, and we
expect that it is involved in the reduction of NO to N2O. This is supposedly a non-energy yielding
process, which would explain the slowdown of growth after reduction of nitrate to nitrite. B. vireti
carries nosZ, encoding N2O reductase, and several genes involved in the maturation of this enzyme,
thereby allowing a kind of full denitrification from nitrate to N2. We are at present investigating to
what extent the N2O reduction may sustain growth by this organism.

23
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 10
Regulation of N2O reduction in Bradyrhizobium japonicum mutants with increased
N2O reductase activity
Cristina Sánchez, Manabu Itakura, Yusuke Uetake, Shima Eda, Hisayuki Mitsui and Kiwamu
Minamisawa
Graduate School of Life Sciences, Tohoku University, 2-1-1 Katahira, Aoba-ku, Sendai 980-8577, Japan
Nitrous oxide (N2O) is a key atmospheric green house effect gas that not only affects global warming but
also leads to the destruction of the ozone layer. Emissions from agricultural fields are a major source of
N2O through the microbial transformation of nitrogen in soil. Previous studies indicated that soybean
nodules emit N2O under field conditions in the late growth period.
In the strain USDA110 of the soybean symbiont Bradyrhizobium japonicum, the genes encoding N2O
respiration functions are clustered in a single locus: nosRZDFYLX. Previously, a B. japonicum mutant
(5M09) with increased N2O reductase (N2OR) activity was isolated by introducing a plasmid containing a
mutated B. japonicum dnaQ gene (impairing proofreading activity) and then performing enrichment
culture under selection pressure for N2O respiration.
Free-living and symbiotic cells of strain 5M09 showed higher N2O reductase activity (5- and 2-times,
respectively) than USDA110 strain under N2O-respiring conditions. Expression of nosZ (which encodes
N2OR) was analyzed in strain 5M09 under free-living aerobic and anaerobic conditions (anaerobic,
nitrate-denitrifying or N2O-respiring conditions). The results showed that constitutive expression of nosZ
gene (aerobic conditions) was increased in strain 5M09. Similarly, under anaerobic and N2O-respiring
conditions, expression of nosZ was higher in strain 5M09. These results suggest an alteration in the
regulatory mechanisms of nosZ expression in strain 5M09, perhaps by derepression. In cells grown
anaerobically in the presence of nitrate, no difference in nosZ expression was observed between strains
USDA110 and 5M09, suggesting a similar response to nitrogen-oxides.
The mechanisms underlying the induction of N2OR expression and activity in strain 5M09 are currently
being investigated.

24
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 11
Production of nitrogen oxides and its regulation in Thermus thermophilus
Carlos Bricio, Laura Álvarez, Alba Blesa, Carolina E. César, Aurelio Hidalgo, José Berenguer Centro de Biología Molecular Severo Ochoa, Universidad Autónoma de Madrid-CSIC. 28049-Madrid, Spain
Several strains of Thermus thermophilus are able to reduce nitrogen oxides under anaerobic
conditions to gain energy in a process that frequently ends up with the production of gaseous forms
of nitrogen that escapes to the atmosphere. Reduction of nitrate is encoded by a conjugative
element (NCE) (Cava et al., 2007), which contains the genes for a tetrameric nitrate reductase, one
or two nitrate/nitrite transporters, a NADH dehydrogenase and a sensory system that signals the cell
to switch between aerobic and anaerobic metabolism. Reduction of nitrite and nitric oxide is
managed by a cd1 type nitrite reductase (Nir) and a nitric oxide reductase (Nor) encoded
downstream of the NCE by a nor-nir cluster, which also includes a group of putative regulators of the
denitrification pathway (Alvarez et al., 2011; Bricio et al., 2011). When present, both the NCE and the
nor-nir clusters are unevenly associated to a highly variable region of a megaplasmid pTT27 like of
different T. thermophilus from which it can be mobilized to other strains of the species. Up to now,
no homologues to conventional nitrous oxide reductase (NosZ) has been found in the genome of
denitrificant strains of this species, suggesting that nitrous oxide is the likely final product of the
process.
For a better understanding of the whole denitrification pathway in the species T. thermophilus, 7
partial or complete denitrificant strains were sequenced. Production and consumption of nitric oxide
in living cells or with different cell extracts, and insertional mutagenesis have been developed to
analyze and confirm or sequence-based hypothesis.

25
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 12
Contrasting denitrification regulatory phenotypes in the genus Thauera revealed by
gas kinetics, transcription analysis and whole genome sequencing
Binbin Liua, Yuejian Maoa,d, Linda Bergausta,b, James P Shapleighc, Lars Reier Bakkenb and
Åsa Frostegårda a Department of Chemistry, Biotechnology and Food Sciences, Norwegian University of Life Sciences, Ås, Norway
b Department of Plant and Environmental Sciences, Norwegian University of Life Sciences, Ås, Norway
c Department of Microbiology, Cornell University, Ithaca, USA
d Key Laboratory of Microbial Metabolism, Ministry of Education, College of Life Science and Biotechnology, Shanghai
Jiaotong University, Shanghai, China
Denitrifiers differ in how they handle transition from oxic to anoxic respiration, with consequences
for NO and N2O emissions. Systematic studies of different organisms are scarce. To enable stringent
comparisons we defined parameters to describe the Denitrification Regulatory Phenotypes (DRP)
based on accumulation of NO2-, NO and N2O, oxic/anoxic growth and transcription of functional
genes. We here present a comparison of eight Thauera strains. The strains divided into two distinct
DRP types. Four strains were characterized by a rapid, complete onset (RCO) of all denitrification
genes and no detectable nitrite accumulation. The others showed progressive onset (PO) of the
different denitrification genes. The PO-group reduced practically all available nitrate (2 mM) to
nitrite prior to transcription of nirS (coding for nitrite reductase), suggesting that nitrate has a role in
the regulating transcription of nirS. Organisms with this phenotype are likely sources of N2O
emissions in natural environments, under conditions allowing aerobic denitrification. All eight strains
controlled NO at nanomolar concentrations, possibly reflecting the importance of strict control for
survival. Transient N2O accumulation differed by two orders of magnitude, indicating that control of
N2O is less essential. No correlation was seen between phylogeny (based on 16S rRNA and functional
genes) and DRP. Whole genomes were sequenced for three RCO strains and three PO strains. Both
paired end and mate pair libraries were constructed, which produced around 120 million reads. The
genome sizes of the 6 strains are in the range of 4.1~4.9 million base pairs. About 3700~4500 CDSs
(coding sequences) were identified for each strain based on the RAST (Rapid Annotation using
Subsystem Technology) pipeline. Some distinct differences were found between the RCO and PO
groups with respect to regulatory genes involved in denitrification, as well as genes involved in
metabolism of aromatic compounds.

26
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 13
Marine microbial ammonium conversion under oxygen-limited conditions
Suzanne C.M. Haaijer1, Jia Yan1, Jaap S. Sinninghe Damsté2, Mike S.M. Jetten1, Stefan Schouten2,
Huub J.M. Op den Camp1
1Microbiology IWWR Radboud University Nijmegen, NL
2Biogeochemistry, NIOZ Texel NL
This project focuses on marine microbial ammonium oxidation under oxygen-limited conditions.
Under these conditions ammonium may be partially converted by aerobic ammonium oxidizers,
yielding nitrite which may serve as a substrate for anaerobic ammonium oxidizers. Although
nitrification was previously assumed to be mediated by bacteria, archaeal ammonium oxidizers
(AOA) have been postulated as key players.
Interactions and coexistence of aerobic and anaerobic ammonium oxidizers were investigated using
bioreactor experiments under oxygen limitation which resulted in 2 stable cocultures. The first
culture [1] consisted of respectively aerobic ammonium-oxidizing bacteria (AOB, Nitrosomonas sp.),
nitrite-oxidizing bacteria (NOB, Nitrospira sp) and Scalindua [2] anammox bacteria. The second
consisted of AOB, AOA (N. maritimus strain SCM1 [3]) and anammox bacteria. These experiments
indicated that AOA are able to coexist with anammox bacteria as well as that competition between
AOA and AOB is likely substrate-level dependent. In addition, enrichments of marine nitrifiers from
the North Sea were established. Highly enriched (>80%) cultures of marine ammonium-oxidizing
bacteria (AOB; a new Nitrosomonas species) and nitrite-oxidizing bacteria (NOB, a new species of
Nitrospira sp) were obtained. In addition, a North Sea AOA enrichment was obtained containing N.
maritimus-like AOA. This batch flask enrichment served as the inoculum for a bioreactor culture
operated under low oxygen and ammonium conditions. This bioreactor is expected to yield sufficient
North Sea AOA biomass to allow more in-depth physiological characterization (e.g. pH, T optima,
affinity constants for oxygen and ammonium) and allow for comparison of this AOA species to N.
maritimus strain SCM1.
[1] Yan J, Op den Camp HJM, Jetten MSM, Hu YY, Haaijer SCM. (2010) Induced cooperation between marine nitrifiers and
anaerobic ammonium-oxidizing bacteria by incremental exposure to oxygen. Syst Appl Microbiol. 33: 407-415
[2] Van de Vossenberg J, Rattray JE, Geerts W, Kartal B, Van Niftrik L, Van Donselaar EG, Damste JSS, Strous M, Jetten
MSM. (2008) Enrichment and characterization of marine anammox bacteria associated with global nitrogen gas
production. Environ. Microbiol. 10, 3120-3129.
[3] Könneke M, Bernhard AE, de La Torre JR, Walker CB, Waterbury JB, Stahl DA (2005), Isolation of an autotrophic
ammonia-oxidizing marine archaeon. Nature 437: 543-546.

27
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 14
Various scenarios for dissimilatory nitrate reduction in Bacillus inferred from
genome analysis
Kim Heylen, Paul De Vos Laboratory of Microbiology, Ghent University, Belgium
Nitrous oxide continues to accumulate in the atmosphere as a product of anthropogenic activities
and land-use changes. Denitrification is considered to be the only ecologically relevant dissimilatory
nitrate reduction pathway responsible for nitrous oxide emission (next to aerobic nitrification).
Reports on nitrous oxide production during dissimilatory nitrate reduction to ammonium (DNRA)
already date back to several decades ago, but still relatively little consideration is given to this
pathway, its key genes/enzymes and its ecological role. We are currently investigating the role of
representative from the genus Bacillus in the nitrogen cycle. And because dissimilatory nitrate
reduction phenotype of completely sequenced strains is not always clear from annotation data or
literature, six strains were selected for genome sequencing (Illumina GAIIx, assembly with CLC
Genomics Workbench 4). Output from the RAST server, KAAS and NCBI’s PGAAP were compared and
used for functional annotation and reconstruction of the dissimilatory nitrate reduction pathways.
Two genomes (B. azotoformans LMG 9581T and B. bataviensis LMG 21833T) contained the
(complete) gene inventory for denitrification, but, in addition, also had nrfA genes, indicating the
potential to perform DNRA. In another genome (B. vireti LMG 21834T), only evidence for DNRA was
found, however together with a nosZ gene. And finally, in the genomes of three distinct strains of B.
licheniformis complete narGHIJ operons and a qnorB genes were found. In conclusion, these results
suggest a potentially complex and overlapping network of dissimilatory nitrate reduction scenarios
and indicate that nitrous oxide in Bacillus can be produced via either the classical denitrification
pathway or the one-electron reduction of nitrite by NarG to nitric oxide and further detoxification
with qNORB.

28
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 15
Community dynamic and quantification of anammox bacteria in a landfill-leachate
treatment plant
Yu-Tzu Huanga*, Shiou-Shiou Chen, Jih-Gaw Linb a
Department of Bioenvironmental Engineering, Chung-Yuan Christian University, 200 Chung Pei Road, Chung-Li 32023
Taiwan, ROC b
Institute of Environmental Engineering, National Chiao Tung University, 1001 University Road, Hsinchu City 30010,
Taiwan, ROC
Anaerobic ammonium-oxidizing (anammox) bacteria participate in a microbial nitrogen cycle
pathway that connects nitrite reduction and ammonium oxidation under anoxic conditions. To
detect anammox bacteria using molecular biology technique of polymerase chain reaction (PCR),
real-time quantitative PCR (qPCR) or fluorescence in situ hybridization (FISH), biomarkers such as 16S
rRNA, hzo and nirS were studied. In order to detect the specific species of anammox bacteria and
elucidate the dynamic shift of the anammox community corresponding to the operating condition of
the reactor, specific primers were designed to target two most common anammox species,
Candidatus Kuenenia stuttgartiensis (KS) and Candidatus Brocadia anammoxidans (BA).
Granular microbial sludge samples were collected and analyzed during a four-year period from a
simultaneous nitrification, annammox, and denitrification (SNAD) treatment plant treating landfill
leachate. Anammox bacteria were kept at relatively high concentration (1.59 x 107 to 6.58 × 108
copies μg-1 DNA). By applying the newly designed KS-specific primer (KSqF3/KSqR3) and BA-specific
primer (BAqF/BAqR), the estimated amount of KS and BA is in the range of 4.12 × 105 to 5.92 × 108
and 1.13 × 105 to 4.41 × 107 copies μg-1 DNA, respectively. KS was predominate species at all well
established anammox granules and played important role in formation of granules. KS/BA ratios
were positively correlated to the ammonia nitrogen removal efficiency of the treatment plant.

29
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 16
Anammox based single reactor systems for nitrogen removal from opto-electronic
wastewaters
Achlesh Davereya, Sin-Han Sua, Yu-Tzu Huangb, Jih-Gaw Lina* a
Institute of Environmental Engineering, National Chiao Tung University, 1001 University Road, Hsinchu City 30010,
Taiwan, ROC b
Department of Bioenvironmental Engineering, Chung-Yuan Christian University, 200 Chung Pei Road, Chung-Li 32023
Taiwan, ROC
Anammox is a nitrogen removal process with many advantages over traditional nitrification and
denitrification process. Anammox bacteria use nitrite as an electron acceptor to convert ammonia
into nitrogen gas and therefore, anammox process is generally combined with partial nitrification
process. Simultaneous partial nitrification, anammox and denitrification (SNAD) and completely
autotrophic nitrogen removal over nitrite (CANON) are two single reactor anammox based
processes. In this study, SNAD and CANON processes were developed to treat two different
optoelectronic industry wastewaters in lab scale sequential batch reactors.
The characteristics of wastewater treated by SNAD process were low C/N ratio (~0.2) with
100 mg-COD/L and 567 mg NH4+-N/L. The experiment was carried out over eight months, where
nitrogen loading rate was gradually increased to 230 g-N/m3·d. The results showed that COD and
nitrogen removal rates by the SNAD system reached to 28 g COD/m3·d and 197 g NH4+-N/m3·d,
respectively in final stage of the process.
The wastewater treated by CANON process had very high ammonium concentration
(3,600 mg NH4+-N/L) with no COD. Lab scale 18 L sequential batch reactor (SBR) was used to study
the wastewater treatment over 450 d (15 months). Around 95% NH4+-N removal and 90% total
nitrogen removal were observed at 4 d HRT with nitrogen loading rate of 909 g/m3/d.
Analysis of microbial communities (nitrifiers, anammox and denitrifiers bacteria) in the reactors was
also carried out by polymerase chain reaction.

30
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 17
A nitric oxide-producing octaheme enzyme from Kuenenia stuttgartiensis
WJ Maalcke1, A Dietl2, SJ Marritt3, MSM Jetten1, JTM Keltjens1, JN Butt3, TRM Barends2, B Kartal1 1 Department of Microbiology, Radboud University Nijmegen, Heijendaalseweg 135, 6525 AJ, Nijmegen, The Netherlands
2 Department of Biomolecular Mechanisms, Max Planck Institute for Medical Research, Jahnstrasse 29, D-69120,
Heidelberg, Germany 3 School of Chemistry, University of East Anglia, Earlham Park, Norwich, NR4 7TJ, United Kingdom
Anaerobic ammonium-oxidizing (anammox) bacteria combine ammonium and nitrite to form
dinitrogen gas. Anammox bacteria are abundantly present in the oxygen minimum zones of the
world’s oceans and have a significant role in global dinitrogen gas production. They are also
successfully applied in cost-effective and environment-friendly ways to treat industrial and municipal
wastewater. Although the physiology of anammox bacteria is quite well understood, the
biochemistry behind it is less clear. One of the persisting anammox enigmas is the presence of ten
copies of an open reading frame that resembles the octaheme enzyme hydroxylamine
oxidoreductase (HAO). HAO converts hydroxylamine to nitrite in aerobic ammonium oxidizing
bacteria. The function of one of these enzymes (kustc0694) is predicted to be hydrazine oxidation to
dinitrogen gas, whereas the functions of the other nine octaheme proteins remain to be established.
Here, a highly expressed octaheme protein was purified from native Kuenenia stuttgartiensis
biomass and characterized. MALDI-TOF mass spectrometry confirmed that the enzyme was encoded
by the open reading frame kustc1061. Enzyme kinetics, assessed by using cytochrome c-based assays
and gas chromatography coupled to mass spectrometry, showed kustc1061 to be a hydroxylamine
oxidizing enzyme that produces nitric oxide instead of nitrite. Protein film electrochemistry revealed
the midpoint potentials of the eight separate heme-groups, and elucidation of the crystal structure
enabled us to form a hypothetical mechanism of hydroxylamine oxidation to nitric oxide. As the
proposed hydrazine-oxidizing enzyme is inhibited by hydroxylamine, the physiological role of
kustc1061 is proposed to be hydroxylamine detoxification

31
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 18
Comparative genomics of two independently enriched ‘Candidatus Kuenenia
stuttgartiensis’ anammox bacteria
Daan R. Speth1, Baolan Hu2, Niek Bosch1, Jan G. Keltjens1, Henk G. Stunnenberg3, Mike S. M.
Jetten1, 4 1 Department of Microbiology, Institute for Water and Wetland Research, Radboud University Nijmegen, Nijmegen, The
Netherlands 2 Department of Environmental Engineering, Zhejiang University, Hangzhou, China.
3 Department of molecular biology, NCMLS, Radboud University Nijmegen, Nijmegen, The Netherlands
4 Department of Biotechnology, Delft University of Technology, Delft, The Netherlands
Bacteria capable of anaerobic oxidation of ammonium (anammox) have many unique features, such
as the conversion of hydrazine, an anammoxosome compartment and ladderane lipids. Although
their core metabolic pathway is largely resolved1, many questions still remain. Data mining of the
(meta)genomes of anammox bacteria is one of the powerful methods to address these questions or
identify targets for further study. The availability of high quality reference data greatly aids such
analysis. Currently, only a single high quality reference genome is available; that of model organism
‘Kuenenia stuttgartiensis’2.
We present a comparative genomic analysis of two ‘K. stuttgartiensis’ bacteria that were
independently enriched aiming to further improve the reference genome. The two bacteria used are
‘K. stuttgartiensis’ RU1, which was originally sequenced for the reference genome, and for the
present study resequenced after seven (2002-2009) years in continuous culture. Furthermore
‘K. stuttgartiensis’ CH1, enriched from a Chinese wastewater treatment plant was used as an
independent source of genomic information.
The two different ‘Kuenenia’ bacteria showed a very high sequence identity (> 99 % at nucleotide
level) over almost the entire genome, but 31 genomic regions (average size 11 kb) were absent from
strain CH1 and 220 kb of sequence was specifically found in the CH1 assembly. Moreover,
comparative analysis of the RU1 and CH1 assemblies led to identification 49 additional genes absent
from the reference genome, including two additional ammonium transporters. Future comparison
will include the metagenomes of Scalindua profunda3 and anammox strain KSU-1.
1. Kartal, B. et al. Molecular mechanism of anaerobic ammonium oxidation. Nature 479, 127–130 (2011).
2. Strous, M. et al. Deciphering the evolution and metabolism of an anammox bacterium from a community genome.
Nature 440, 790–794 (2006).
3. van de Vossenberg, J. et al. The metagenome of the marine anammox bacterium “Candidatus Scalindua profunda”
illustrates the versatility of this globally important nitrogen cycle bacterium. Environmental Microbiology no–no
(2012).doi:10.1111/j.1462-2920.2012.02774.x

32
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 19
Role of nitriles in cyanide assimilation
Victor M. Luque-Almagro, Isabel Manso, Jessica Estepa, M. Paz Escribano, Francisco Castillo,
Conrado Moreno-Vivián, M. Dolores Roldán Departamento de Bioquímica y Biología Molecular. Campus de Rabanales. Universidad de Córdoba. Córdoba, Spain
The alkaliphilic bacterium Pseudomonas pseudoalcaligenes CECT5344 is able to grow with cyanide as
the sole nitrogen source. Under cyanotrophic conditions, an inducible cyanide-insensitive alternative
oxidase is coupled to a malate:quinone oxidoreductase activity that oxidizes L-malate to
oxaloacetate. Oxaloacetate reacts chemically with cyanide to produce a cyanohydrin (2-
hydroxynitrile), which may be further converted into ammonium. Two cyanide-induced proteins of
unknown function identified by proteomic analysis are encoded by the nitB and nitG genes belonging
to the nit1C gene cluster in the strain CECT5344. In addition to NitB and NitG, the predicted products
of nit1C gene cluster are a Fis-like 54-dependent transcriptional activator (NitA), a nitrilase (NitC), an
S-adenosylmethionine superfamily member (NitD), an N-acyltransferase superfamily member (NitE),
a trifunctional polypeptide of the AIRS/GARS family (NitF), and an NADH-dependent oxidoreductase
(NitH). The P. pseudoalcaligenes CECT5344 nit1C gene cluster is involved in assimilation of free and
organic cyanides (nitriles) as deduced for the inability to grow with cyanide showed by the NitA, NitB
and NitC mutant strains. The wild-type strain CECT5344 shows nitrilase activity, which enables
growth on cyanide or hydroxynitriles, but the NitB and NitC mutants have only low basal levels of
nitrilase activity that are not enough to support growth on either free cyanide or nitriles. To
summarize, P. pseudoalcaligenes CECT5344 assimilates cyanide through a novel pathway that
includes the formation of 2-hydroxynitriles that are used as N-source with the involvement of the
nitrilase-encoding nit1C gene cluster.

33
17th European Nitrogen Cycle Meeting, Oslo 2012
KN 06
Anaerobic bacterial response to NO generation, damage and repair
Charlene Bradley, Vine C, Balseny B, Wang J, Cole J School of Biosciences, University of Birmingham, Birmingham B15 2TT, UK
Bacteria are exposed to NO generated as an immediate product of nitrite reduction by denitrifying
bacteria, from arginine by the mammalian NO synthetase, or as a by-product during nitrate
reduction to ammonia. NO binds to di-iron and iron-sulphur clusters, inactivating many enzymes
including aconitase and fumarase.
The Escherichia coli transcription factors NorR and NsrR bind NO specifically, triggering responses to
nitrosative stress. NsrR is a repressor of genes required for NO removal or damage repair, including
the flavohemoglobin, Hmp, the hybrid cluster protein, Hcp, and the di-iron repair protein, YtfE . Hmp
is an NO oxygenase. Its expression is repressed by FNR, implying that it is not required during
anaerobic growth. In contrast, Hcp expression is activated by FNR, suggesting it is important under
anaerobic conditions. Hcp contains a [4Fe-2S-2O] hybrid cluster that is so far unique in evolution.
Unique structure implies unique function and gene regulation usually reflects metabolic function: we
will demonstrate that Hcp plays a critical role in the response to nitrosative stress. Repair of
nitrosative damage inevitably focuses on how iron-sulphur cluster proteins are repaired. The repair
protein, YtfE, has been reported to repair iron-sulphur clusters damaged by nitrosative stress. We
will report experiments to establish the mechanism of repair. Data from isogenic mutants that lack
all reported NO reductases suggest that there are multiple responses to nitrosative stress in E. coli.
The responses of E. coIi to nitrosative stress during aerobic and anaerobic growth will be compared
with those of other organisms.

34
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 20
Nitric oxide detoxification and nitrate assimilation in Bradyrhizobium japonicum
Juan J Cabrera1, Cristina Sanchez1, Andrew Gates2, Eulogio J Bedmar1, David J Richardson2, Maria J
Delgado1
1 Estación Experimental del Zaidín, CSIC, P. O. Box 419, 18080-Granada, Spain
2 School of Biological Sciences, University of East Anglia, Norwich, NR4 7TJ, UK
Bradyrhizobium japonicum is a soil bacterium that establishes nitrogen-fixing symbiotic associations
with soybean plants and is also able to denitrify under free living and symbiotic conditions. In
B. japonicum, a single-domain haemoglobin (Bjgb) has been reported to be involved in NO
detoxification under free-living conditions (Sanchez et al., 2011 Biochem Soc Transac. 39 184-188).
This Bjgb is encoded by blr2807 that is located in a gene cluster (blr2803-09) where blr2803-05
encode an ABC-type nitrate transporter, blr2806 a nitrite extrusion protein, blr2808 a flavoprotein,
and blr2809 a assimilatory nitrate reductase. Functional and regulatory analyses of these genes are
presented this work. B. japonicum blr2807, blr2808 and blr2809 mutants showed a defect in their
ability to grow anaerobically with nitrate as sole nitrogen source. Furthermore, blr2808 and blr2809
mutants were also unable to grow with nitrate as sole nitrogen source under aerobic conditions and
they recovered growth when glutamic acid was added to the medium. In addition, blr2807- and
blr2808- mutants showed greater sensitivity to NO-generating compounds than the parental strain.
Reverse transcription-PCR analysis showed that blr2806-09 genes comprise a transcriptional unit.
-galactosidase activity of a transcriptional fusion of blr2806-09 promoter region to lacZ revealed
that nitrate is required to induce expression of these genes and the presence of glutamic acid
inhibits such expression. The presence of a NO donor increases expression of the blr2806-09 operon.
Taken together, these results suggest that the B. japonicum blr2803-09 gene cluster is involved in
nitrate assimilation and NO detoxification.
Acknowledgements. This work was supported by Fondo Europeo de Desarrollo Regional co-financed grants from Junta
de Andalucía (Spain) [grant number CVI-3177] and from Ministerio de Economía y Competitividad (Spain) [grant number
AGL2010-18607]. Support from the Junta de Andalucıa to Group BIO-275 is also acknowledged. Juan Cabrera was
supported by a fellowship from the Consejo Superior de Investigaciones Cientificas JAE Programme.

35
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 21
Nitrosative stress response in Paracoccus denitrificans
Hartop, K. R., Gates, A. J., Clarke, T. A. and Richardson, D. J. Centre for Molecular and Structural Biochemistry. School of Biological Sciences, University of East Anglia, Norwich
Research Park, Norwich, NR4 7TJ, U.K.
Nitrogen based compounds highly chemically reactive in solution, termed reactive nitrogen species
(RNS), generated in concentrations greater than can be neutralised by microorganisms induce
nitrosative stress in microorganisms leading to cytotoxic effects such as inhibition of cellular growth,
disruption of biofilm attachment and cell death. Increasing nitrite concentration changes the
phenotype of aerobic growth in Paracoccus denitrificans PD1222 causing an increase in lag phase, a
decrease in exponential growth rate and reduction of final OD600nm at stationary phase, until cell
growth is not observed. A pH effect is also present; exponential growth rate µmaxapp reduction as a
function of decreased pH, therefore reducing the concentration of nitrite at which toxicity occurs.
Toxicity is therefore postulated as linked to nitrite protonation: NO2- + H+ ↔ HNO2 (pka = 3.3),
forming free nitrous acid (FNA), where higher levels of FNA are present at low solution pH as defined
by the Henderson-Hasselbalch equation. With oxygen present, the anaerobic denitrification enzyme
pathway is down-regulated, thus the fate of nitrite under aerobic growth conditions is unclear. The
decomposition and disproportionation of FNA may generate cytotoxic reactive species such as nitric
oxide (NO) detrimental to cellular function. From which cellular detoxification may form nitrous
oxide (N2O). The focus of this work is to investigate aerobic nitrosative stress response on a microbial
and transcriptional level in P. denitrificans to assess its impact on FNA, NO and N2O generation and
cellular growth in both batch and continuous chemostat cultures.

36
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 22
The production and detoxification of a potent cytotoxin, nitric oxide, by pathogenic
enteric bacteria
Sebastian Runkel, Corinne Appia-Ayme, Anke Arkenberg, Elaine Patrick, Hannah Wells,
David J. Richardson and Gary Rowley School of Biological Sciences, University of East Anglia, Norwich Research Park, Norwich, Norfolk NR4 7TJ, U.K.
Escherichia coli and Salmonella are facultative anaerobes; using O2 as their preferred energy source.
However, during O2 shortage, they can switch to nitrate respiration to maintain their metabolism in a
process called denitrification. Unlike many soil bacteria, Salmonella and E. coli undergo only
truncated denitrification, where the alternative electron acceptor nitrate is converted into the
neuropharmacological agent and greenhouse gas nitrous oxide (N2O) via nitrite (NO2-) and nitric
oxide (NO). The ability to reduce NO to different compounds is crucial for their survival as NO is a
potent cytotoxin and used by macrophages to defeat invaders. Thus, E. coli and Salmonella have
evolved mechanisms to overcome the hosts’ attack and to defend themselves against their own toxic
by-products (Rowley et al., 2012 Biochemical Journal 441, 755-762). The three main NO
detoxification enzymes include: flavohaemoglobin (Hmp), flavorubredoxin (NorVW) and the
cytochrome c nitrite reductase NrfA (Mills et al., 2008 Microbiology 154, 1218-1228). Continuous
chemostat cultures and transcriptional analysis were used to investigate the importance of N2O
production and NO detoxification for the pathogenicity of enteric bacteria, using Salmonella and E.
coli as comparative pathogenic and non-pathogenic model systems (Arkenberg et al., 2011
Biochemical Society Transactions 39, 1876-1879). Results show that there are differences in N2O
production between Salmonella and selected E. coli strains, which are partially caused by differential
transcription of narG, nirB and hmp.

37
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 23
Kinetics of NO scavenging in oxic and anoxic conditions, experimental approaches to
differentiate between chemical and enzymatic degradation
Shahid Nadeem1, Andrew M. Stern4, Wenmin Long2, Peter Dörsch1, Åsa Frostegård1,
Jim Shapleigh3, Jun Zhu4 ,Binbin Liu1 and Lars R Bakken1 1
UMB Nitrogen Group, Norwegian University of Life Sciences, Norway. 2 Jiaotong Shanghai University, Shanghai, PRC,
3 Cornell University, Ithaca, NY USA,
4 Department of Microbiology, Perelman School of Medicine, University of Pennsylvania, Philadelphia, PA, USA
NO is a toxic signal molecule and an intermediate in denitrification, but NO is produced and
“scavenged” by a plethora of other enzymes and organisms. To understand its role as a toxic
compound and an inducer of denitrification in soil and N-transformations duirng host-pathogen
interactions, we need to determine the kinetics of NO scavenging by relevant organisms.
Experiments with NO are not trivial because NO is chemically unstable in the presence of oxygen due
to “autoxidation”, which is a “third order” process; V=k*[O2]*[NO]2. This happens both in the gas
phase and the liquid phase. The problem with chemical instability is escalating with high NO
concentrations, even at relatively low oxygen concentrations. We have developed an approach to
tackle these problems, by a combination of mathematical modeling and an incubation system that
monitors the concentrations of NO and O2 in a two phase system (liquid + headspace). The approach
allows a determination of the kinetic parameters for NO scavenging, be it under oxic or anoxic
conditions. The approach has been used to characterize oxic and anoxic NO scavenging in Vibrio
cholerae, Paracoccus denitrificans, Agrobacterium tumefaciens and mutants thereof, to elucidate the
role of different enzymes. The results indicate that oxic NO scavenging is very fast in some species,
and apparently absent in others, and the studies of mutants have identified some of the proteins
involved in NO scavenging.

38
17th European Nitrogen Cycle Meeting, Oslo 2012
KN 07
Identifying and quantifying biological source and sink processes of N2O using isotope
tracing and natural abundance techniques.
Reinhard Well1, Gesche Braker2, Anette Giesemann1, Dominika Lewicka-Szczebak1, Lena Rohe1,
Greta Roth1
1 Johann
Heinrich von Thuenen-Institute, Institute of Agricultural Climate Research, Braunschweig, Germany
2 Max Planck Institute for Terrestrial Microbiology, Marburg, Germany
To understand the control of N2O fluxes to the atmosphere it is essential to elucidate the various
processes of biological N2O production and consumption. This includes production by nitrification,
nitrifier-denitrification, bacterial or fungal denitrification, co-denitrification by fungi and bacterial
reduction to N2. 15N and 18O tracing techniques can be used to quantify the contribution of the
respective pathways. Moreover, natural abundance of N2O isotopologue signatures (δ18O, average
δ15N and 15N site preference, SP = difference in δ15N between the central and peripheral N positions
of the asymmetric N2O molecule) have recently received some attention as tools to elucidate the
contribution of nitrification, bacterial and fungal denitrification to N2O fluxes as well as to estimate
N2O reduction to N2. Isotopologue signatures of individual pathways have been studied with pure
cultures of fungal and bacterial denitrifiers and of autotrophic nitrifying bacteria. Moreover, N2O
isotopologues are increasingly used as indicators of process dynamics in soil flux studies. An
overview of the techniques as well as some examples from the literature and from our own studies
will be given.
To complement previous literature data, we determined the specific isotopic signature of N2O
production from pure cultures of bacterial and fungal denitrifiers. To investigate N2O reduction in
soil, we measured isotopologue values of soil-derived N2O produced under conditions favouring
denitrification. These experiments where combined with independent estimates of denitrification
obtained from 15N tracing or incubation under N2-free atmosphere. We used an isotope fractionation
model to check if N2 production can be calculated from isotopologue data.

39
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 24
Aeration of boreal acid sulphate soils could result in enhanced fluxes of N2O and CH4
due to stimulation of potent microbial communities present in subsoil
Miloslav Šimek1,2, Alica Chroňáková1, Dana Elhottová1, Seija Virtanen3, Asko Simojoki3,
Václav Krištůfek1, Markku Yli-Halla3
1 Biology Centre AS CR, v. v. i., Institute of Soil Biology, České Budějovice, Czech Republic
2 University of South Bohemia, Faculty of Science, České Budějovice, Czech Republic
3 University of Helsinki, Department of Food and Environmental Sciences, Helsinki, Finland
Boreal acid sulphate (AS) subsoil horizons contain significant amounts of organic carbon and
nitrogen. We hypothesized that large emissions of CO2, CH4, and N2O from AS soils could occur, if the
soil conditions, e.g. aeration status, change, provided relevant microorganisms are present. We
investigated this phenomenon in Helsinki region in Finland and sampled AS and non-AS soils to
depths of 165 - 180 cm. We confirmed large carbon and nitrogen stocks (110 Mg Corg ha-1 and 15 Mg
Ntot ha-1) in the permanently flooded C horizons of the AS soil but not in the control non-AS.
Moreover, in the subsoil, high ammonium concentrations were found. We also found very high basal
and glucose-induced respiration in the same Cg2 horizon of the AS soil. Examination of microbial
communities revealed much higher abundance of methanogens in the deepest C horizons, than in
the surface Ap horizons. Nitrous oxide production determined in laboratory incubations peaked in
the BCgc and BC horizons of the AS and non-AS soils, respectively, but in the AS soil the peak value
was tenfold higher than in the non-AS soil. We conclude that surprisingly high microbial abundance
and activity, together with the abundant carbon and mineral nitrogen in the deep layers of AS soils,
may result in large gas emissions. High emissions could occur for example when the generally high
water table would be lowered due to arable farming. Additional studies of boreal AS soils are needed
to assess their potential contribution to increases in greenhouse gas emissions.

40
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 25
Effect of intensified crop production on soil nitrous oxide emission in Mid-Hills of
Nepal
Nani Raut1, Bishal K Sitaula1, Lars R Bakken2
1 Department of International Environment and Development Studies, Norwegian University of Life Sciences PO box
5003, N-1432 Aas, Norway 2
Department of Plant and Environmental Sciences, Norwegian University of Life Sciences PO box 5003, N-1432 Aas,
Norway
Dramatic changes in agricultural practices have taken place in large part of South Asia. The
traditional low input farming is replaced by intensified production systems (high fertilizer levels and
more crops per year). The high N inputs will increase the emissions of N2O, but the prospects are
potentially worse than those predicted by IPCC-methodology which assumes that N2O emission
increase linearly with N-input. The reason is that crop intensification may acidify the soils which
could increase the N2O/(N2+N2O) product ratio of denitrification. We have investigated the
phenomenon by analyzing soil samples from seven different sites, each site containing both
traditional and adjacent intensified plots. The results showed that crop intensification invariably
lowered the soil pH and increases the N2O/(N2+N2O) product ratio under standardized anoxic
incubations (Raut et al., 2012, Soil Biology & Biochemistry in press). In theory, this would imply
escalating N2O emissions beyond that predicted by IPCC. This needs to be tested by measuring N2O
emissions in field experiment, and here we present such results: Regardless of the land use
categories, intensified agricultural system had significantly higher N2O emission than traditional
agricultural system. We can estimate the emission factor for intensification: EFint = ∆N2O-N year-1/ ∆
N-input year-1, where ∆N2O-N year-1 is the estimated increase in N2O-emission by intensification, and
∆ N input year-1 is the annual input of N to Intensified plots minus that in traditional plots. For the
site with permanently drained soil, EFint was 0.13, and for the periodically flooded soil, it was 0.03. In
comparison, the emission factor for N input in the IPCC formula is 0.01-0.02. These results confirm
the predictions from standardized anoxic incubations; soil acidification by N fertilization leads to
much higher N2O emission than that predicted by the IPCC formula.

41
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 26
High N2O emissions from an N saturated subtropical forested catchment, Southwest
China
Jing Zhu, Jan Mulder, Peter Dörsch Dept. Plant and Env. Sci. Norwegian University of Life Sciences, Aas, Norway
Subtropical forests in South China, after receiving long-term elevated nitrogen (N) deposition, have
become N saturated. Previous studies found that, despite low forest growth, N leaching via stream
water was moderate, indicating important unknown N sinks. Given the monsoonal climate with hot
and wet summers and the widespread acid soil in this region, denitrification may represent a major
sink of reactive N and result in considerable emission of nitrous oxide (N2O), a potent greenhouse
gas (GHG).
Field flux measurements of N2O with static chambers carried in an N saturated subtropical forested
catchment in southwest China, Tieshanping (TSP), found high N2O fluxes in summer. Fluxes of N2O
were significantly greater on hillslopes than in a hydrologically connected groundwater discharge
zone. Thus, landscape position, which integrates different soil characteristics, affected the N2O
emission. During summer, soil moisture was the determining factor, resulting in transient N2O peaks
(up to 1730 μg N2O-N m-1 h-1) on the hillslope after rain episodes. The extrapolated annual N2O fluxes
were 0.50 and 0.41 N2O-N m-2 in 2009 and 2010, respectively. An in situ 15N-NO3- labeling experiment
confirmed that denitrification rather than nitrification was the majority pathway of N2O production.
Results from lab incubations supported these findings: soils from hillslope have high denitrification
potential with impeded N2O reduction activity as well as low nitrification potential causing NO2 -
accumulation. The latter was shown to quickly dismutase to NO under acid condition, opening for
another important pathway for N removal.

42
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 27
High potential for N2O emission from intensive Chinese agriculture due to soil
acidification
Zhi Qu1, Jingguo Wang2, Lars R Bakken1
1 Department of Plant and Environmental Sciences, Norwegian University of Life Sciences
2 College of Resources and Environmental Sciences, China Agricultural University
Chinese farmers use large amount of mineral nitrogen (N) fertilizer and manure to sustain grain
production. Excessive N inputs result in gradual soil acidification all over China, which may impair
N2O reductase and hence induce an escalating emission of N2O. We investigated this by analyzing
soils from long-term fertilization experiments in different regions of China (Gongzhuling (GZL),
Shangzhuang (SZ), Qiyang (QY), Changwu (CW) and Shouguang (SG)). The soils were incubated after
adjusting their nitrate (NO3-) concentration to 2mM NO3
- -N by flooding and drainage. The NO, N2O
and N2 gas kinetics from denitrification were monitored during anaerobic incubations, to explore the
fertilization effects on the denitrification product stoichiometry (NO/N2O/N2) during such
standardized incubations. We also investigated if the product stoichiometry could be altered by
adjusting pH prior to such anoxic incubations. The results show that:
1- High levels of mineral N acidified the soils, but not in soils with high buffer capacity;
2- High levels of mineral N led to higher N2O/(N2O+N2) product ratios, but not in soils with high
buffer capacity;
3- Adjusting the soil pH in acidified soils restored a low N2O/(N2O+N2) ratio;
4- Manure had a variable effect on soil pH;
5- Overall, we found a strong negative correlation between soil pH and the N2O/(N2O+N2) ratio
(pH range 3.7-8.0, r2=0.75).
We conclude that the excessive N fertilization input in China resulted in soil acidification, which is a
direct cause of enhanced propensity for N2O emission.

43
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 28
Modelling the effect of Copper availability on bacterial denitrification
H.C. Woolfenden1, A.J. Gates2, G. Giannopoulos2, M.J. Sullivan2, D.J. Richardson2, V. Moulton1 1
School of Computing Sciences, University of East Anglia, Norwich Research Park, Norwich, NR4 7TJ, U.K. 2
School of Biological Sciences, University of East Anglia, Norwich Research Park, Norwich, NR4 7TJ, U.K.
When bacteria such as Paracoccus denitrificans respire anaerobically they convert nitrate (NO3- ) to
dinitrogen (N2) gas via a pathway which includes the potent greenhouse gas, nitrous oxide (N2O). The
copper-dependent enzyme Nitrous Oxide reductase (Nos) catalyses the reduction of N2O to N2.
Experiments have shown that in low-copper conditions Nos efficiency decreases, which can result in
significant emissions of N2O. A mathematical model is developed to describe the anaerobic
denitrification pathway based on Michaelis-Menten kinetics and published kinetic parameters.
Novelty is given to the model by adjusting the Nos concentration depending on the copper
concentration. The accuracy of the model can be checked by comparing the predictions with existing
experimental results from chemostats. Further, individual reductase concentrations are deduced
from simple algebraic expressions involving the kinetic parameters and the experimental data. The
relative expressions of the enzymes may then be compared to experiments. The model may also be
used as a prediction tool to estimate the output levels of the nitrogenous compounds in the
denitrification pathway, in particular nitrous oxide, for a prescribed set of initial conditions.

44
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 29
Unexpected diversity of nitrite oxidoreductase genes (nxrB) in marine waters:
indications for novel NOB?
Andreas Schramm1, Sandra Lopez1, and Sebastian Luecker2
1 Department of Bioscience, Microbiology, Aarhus University, Denmark
2 Department of Microbial Ecology, University of Vienna, Austria
Since the recent discovery of ammonia-oxidizing archaea (AOA), increasing evidence has
accumulated that AOA are the main ammonia oxidizers in the ocean, where they can be extremely
abundant and utilize minute concentrations of ammonia, which is oxidized to nitrite. However,
nitrite rarely accumulates in the ocean but is either further oxidized to nitrate or serves as important
link between aerobic and anaerobic N conversions in oxygen minimum zones (OMZ). Yet the
organisms mediating nitrite oxidation in the ocean are poorly studied, and the abundance of known
marine nitrite-oxidizing bacteria (NOB), i.e. Nitrospina, Nitrococcus, and Nitrospira, hardly correlates
with nitrite oxidation rates.
The objective of this study was to identify putative NOB in the OMZ off the coast of Chile and Peru
and to correlate their presence with chemical profiles in the water column. PCR-based approaches
were developed to target 16S rRNA genes of the known NOB genera, and the gene encoding the
beta subunit of nitrite oxidoreductase (nxrB). The only NOB-like 16S rRNA genes detected were
affiliated with Nitrospina and revealed a large diversity within this group, but also a lack of
correlation with the chemical profiles. In contrast, at least four distinct clusters of nxrB-like genes
were observed, showing conspicuous depth distributions. The possibility that these genes indicate
the presence of hitherto unknown NOB will be discussed.

45
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 30
Biogeochemical nitrogen cycling in Arctic snowpacks: generating hypotheses from
multi-disciplinary studies
Catherine Larose1, Sebastien Cécillon1, Emmanuel Prestat1, Aurélien Dommergue2,, Pascal
Simonet1 and Timothy M. Vogel1 1
Environmental Microbial Genomics, Ecole Centrale de Lyon, Université de Lyon, 36 avenue Guy de Collongue, 69134
Ecully, France 2
Université Joseph Fourier – Grenoble 1 / CNRS, LGGE, 54 rue Molière BP56, F-38402 Saint Martin d’Hères, France
Arctic snowpacks are often considered as chemical reactors for a variety of chemicals deposited
through wet and dry events, but are overlooked as potential sites for microbial metabolism of
reactive nitrogen species. The fate of deposited species is of utmost importance since melt leads to
the transfer of nutrients and contaminants to snowmelt-fed ecosystems. Here we examined the role
of microorganisms and the potential pathways involved in nitrogen cycling in the snow. Using a 16S
rRNA microarray to follow shifts in microbial community structure during a two-month springtime
field study at a high Arctic site, Svalbard, Norway (79°N) in addition to pyrosequencing, we showed
that despite the cold conditions and limited water supply, microbial communities inhabiting the
snow cover were dynamic and had the functional potential to carry out several different, but not all
pathways of the nitrogen cycle. Nitrogen cycling was shown to be a dominant process at the base of
the snowpack and seems to be among the most important functions in Arctic ecosystems.

46
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 31
Is there a direct link between denitrifier community structure or size and
denitrification activity in arable soil?
Miriam B Herold1,2, Liz M Baggs1, Tim J Daniell2
1 Institute of Biological and Environmental Sciences, University of Aberdeen, UK
2 The James Hutton Institute, UK
Denitrification is the biological process by which intermediates of the nitrogen cycle are reduced to
maintain respiration under oxygen limiting conditions. Several enzymes and their corresponding
genes have been identified in denitrifying bacteria. The reduction of nitrite to nitric oxide is catalysed
by the copper containing nitrite reductase (nirK) or the cd1 type nitrite reductase (nirS). Both nirK
and nirS were used in this study to investigate the effects of pH and cultivation management of
arable soil on the community structure and size of bacterial denitrifiers.
Soil was sampled from long-term pH plots under a cultivation gradient (SAC, Craibstone, UK). A
molecular finger printing technique (T-RFLP) was used to evaluate microbial community structure
and real-time PCR was used to estimate microbial community size. Potential denitrification rates
(PDR) were measured in order to compare denitrification activity and community structure/size.
Potential denitrification was strongly influenced by pH and cultivation with the highest PDR from
slightly acidic soil and soil under long-term ley grass. Both nirK and nirS community structures were
influenced by pH and changed significantly at acidic pH. The community structure of nirK but not the
community structure of nirS responded significantly to cultivation management (ploughing) which
suggested that bacteria with different nitrite reductases can follow different population dynamics.
Significant changes in community structures did not necessarily follow changes in gene abundance
and therefore we concluded that both community structure and size influenced denitrification
activity in arable soil.

47
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 32
Microbial composition of various components of closed aquaculture systems:
analysis of biofilters and indogenous microbiota of fish intestines
Maartje A.H.J. van Kessela,b, Arslan Arshada,b, Gert Flikb, Mike S.M. Jettena, Peter H.M. Klarenb,
Huub J.M. Op den Campa a
Department of Microbiology and b
Department of Organismal Animal Physiology, IWWR, Radboud University Nijmegen,
Heyendaalseweg 135, NL-6525 AJ Nijmegen, the Netherlands
The maintenance of an optimal water quality poses challenges for aquaculture worldwide. Especially
the accumulation of nitrogen compounds, which are excreted by fish, is a major problem in
aquaculture. Strict rules for concentrations of these compounds in the effluent water and toxicity of
these compounds force aquaculture operators to reduce nitrogen in the effluent water. This can be
accomplished in different manners: 1) Efficient removal of the nitrogenous compounds from the
water or 2) Reduction of the nitrogen input. Biofiltration is often used to remove nitrogen
compounds from the water. However, currently we know little about the contribution of various
groups of N-cycle bacteria to the removal of nitrogenous compounds in aquaculture systems. We
therefore investigated the composition of a biofilter which revealed the presence of various
members of nitrogen-cycling bacteria, including ammonium- and nitrite-oxidizing and anammox
bacteria. Reducing nitrogen accumulation by reducing the nitrogen input can be accomplished by
decreasing the protein concentration in food. Most fish are fed high protein content food to
maximize growth. Part of this protein can be exchanged for plant material, however it is important to
know if cultured fish can convert the plant components to usable metabolites. We therefore
investigated the intestinal microbiota of common carp (Cyprinus carpio) using culture-independent
methods. This revealed the presence of a large population of microorganisms able to ferment plant
components. Furthermore, members of the nitrogen cycle bacteria were found to reside in the fish’
gut pointing to new in situ possibilities to reduce nitrogen emissions in aquaculture.

48
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 33
Differences in composition of active and overall denitrifier communities in three
organic soils determine responses of denitrification to pH
Kristof Brenzinger1, Peter Dörsch2 and Gesche Braker1 1
Max Planck Institute for Terrestrial Microbiology, Karl-von-Frisch-Strasse 10, D-35043 Marburg, Germany 2
Norwegian University of Life Sciences, Aas, Norway
Denitrifier communities from three drained organic soils in Finland (FIN), Germany (GER) and
Sweden (SWE) were studied, differing in soil history and soil parameters. The soils showed marked
physiological differences in denitrification response to anoxia and low temperature. Moreover, a
direct effect of pH on denitrifier functioning had been observed when exposing bacterial consortia
extracted from these soils to two different pH levels (pH 5.4 and 7.1) (1). We hypothesized that the
denitrifier communities harbored by these soils were composed differently. Indeed, we found
differences in diversity, composition and abundance of denitrification genotypes between the soils
based on NO2--reductase (nirK/nirS) and N2O-reductase (nosZ) genes which appeared to congruent
with the communities’ ability to produce and reduce N2O (2).
In a subsequent study we explored the population dynamics and transcription of denitrification
genes in extracted communities during incubation at different pH and found both to be strongly
controlled by pH. Moreover, our results indicate that denitrifier activity may be additionally affected
by posttranslational modification of the enzymes.
We conclude that links exist between the genetic makeup of denitrifier communities and their
physiological responses to environmental triggers. Furthermore, we could show that links to
functional differences were mostly caused by differences in the composition of the active denitrifier
community in response to pH and we hypothesize that posttranslational regulation is an important
regulatory step impacting ecosystem functioning.
(1) Dörsch, P., Braker, G. and Bakken, L.R. (2012) Community specific pH response of denitrification: experiments with
cells extracted from organic soils. FEMS Microbiol. Ecol., 79, 530–541.
(2) Braker, G., Dörsch, P. and Bakken, L.R. (2012) Genetic characterization of denitrifier communities with contrasting
intrinsic functional traits. FEMS Microbiol. Ecol., 79, 542–554.

49
17th European Nitrogen Cycle Meeting, Oslo 2012
CP 34
Unusual acid-tolerant denitrifier communities in arctic permafrost soils associated
with N2O fluxes
Katharina Palmer and Marcus A. Horn
Peramfrost-affected peatlands were recentlly identified as sources of the greenhouse gas N2O.
Unvegetated tundra peat circles (PC) are nitrate-rich ‘hot spots’ of N2O emissions, while adjacent
vegetated tundra areas (UT), and moderately vegetated palsa peats (PP) emit only minor amounts of
N2O. Soil pH approximated 4 for all sites. N2O emission potentials of unsupplemented anoxic peat
soil slurries and apparent maximal nitrate-dependent maximal reaction velocities were highest for
PC, intermediate for PP, and lowest for UT. Stimulation of N2O production by nitrate-, nitrite-, and/or
acetylene suggested denitrification as major source of N2O in all soils. N2O/(N2+N2O) ratios were 30,
55, and 100% for PC, PP, and UT, respectively, when 10 µM of nitrate were supplied. Such dissimilar
physiological responses suggested dissimilar denitrifier communities in the soils. Barcoded amplicon
pyrosequencing of narG, nirK, nirS, and nosZ (encoding nitrate, nitrite, and N2O reductases,
respectively) revealed diverse denitrifiers including hitherto unknown species, and likewise
suggested that denitrifier diversity differed between the soils. Sequences affiliated with Actino-,
Alpha- and Betaproteobacteria as well as environmental sequences. Quantitative PCR of narG, nirK,
nirS, and nosZ indicated that detected nirS outnumbered nirK in all soils. Detected narG and nirK
were most abundant in PC. The collective data suggest that (i) Proteobacterial nirS-type rather than
nirK-type denitrifiers are abundant in acidic permafrost soils, and (ii) the contrasting N2O emission
patterns are associated with contrasting denitrifier communities.

50
17th European Nitrogen Cycle Meeting, Oslo 2012
Poster abstracts
Ordered alphabetically, according to first author
P 01
Impact of loss of norEF in Rhodobacter sphaeroides 2.4.1
Linda Bergaust1, Angela Hartsock2, Lars Bakken1, Åsa Frostegård1 and James P. Shapleigh2 1
UMB Nitrogen Group, Norwegian University of Life Sciences, Norway 2 Department of Microbiology, Cornell University, Ithaca, NY USA
During denitrification nitric oxide (NO) is reduced by NO reductase (Nor) to nitrous oxide (N2O).
There are two types of Nor found in denitrifying bacteria with one, qNor, receiving electrons from
quinol while the other, cNor, receives electrons from cytochrome c. Both are members of the
superfamily of heme-copper oxidases. Purified cNor is a two subunit complex. Many members of the
heme-copper superfamily contain a third subunit upon purification. A gene encoding a protein with
sequence similarity to the heme-copper subunit III is found in the denitrification clusters of many
denitrifiers. This gene has been designated norE and is nearly always associated with a second gene
designated norF. R. sphaeroides 2.4.1 contains the genes for a cNor but lacks norEF. This strain also
lacks genes for and nitrite reductase and nitrous oxide reductase but has a nitrate reductase. R.
sphaeroides 2.4.3 has both the cNor gene cluster and norEF but the latter do not form a tight genetic
cluster with the cNor operon. To learn more about the role of norEF we determined how the loss of
norEF impacted NO reduction in R. sphaeroides 2.4.1. Both 2.4.1 with nirK from 2.4.3 and 2.4.3
lacking norEF were able to reduce nitrate to N2O without accumulation of detrimentally high
concentrations of NO. However, reduction of nitrite lead to accumulation of NO consistent with an
impairment of Nor activity. Addition of norEF from 2.4.3 to the 2.4.1 strain with nirK reduced the
accumulation of NO that occurred during the reduction of nitrite. Taxis behavior of the various
strains to a nitrite source was consistent with norEF limiting nitrite toxicity, most likely because it
enables the cells to control NO accumulation. These results show that loss of norEF by 2.4.1 does
impact Nor activity. This suggests that despite the loss of norEF the Nor in 2.4.1 provides a selectable
advantage because this strain only periodically encounters exogenous NO. The rate of endogenous
NO production during denitrification prevents the loss of norEF in 2.4.3.

51
17th European Nitrogen Cycle Meeting, Oslo 2012
P 02
Abundance and diversity of nitrifying prokaryotes in drinking water treatment plant
Mei-Chun Chen1, Toshikazu Fukushima2, Sin-Ming Liu3, Pei-Chu Chen3, Liang-Ming Whang1 1
Department of Environmental Engineering, National Cheng Kung University, Tainan, Taiwan 2 Division of Environmental Engineering, Hokkaido University, Japan
3 Taiwan Water Corporation, Taichung, Taiwan
In order to better understand nitrification in drinking water treatment processes, this study was
motivated to investigate abundance and diversity of nitrifying prokaryotes in filtration processes in
three drinking water treatment plants (plant A, B-1 and B-2) using real-time PCR and terminal
restriction fragment length polymorphism. In general, real-time PCR results showed that AOA and
Nitrospira were dominant ammonia and nitrite oxidizers in the filtration processes, except for the
plant B-1 which practiced pre-chlorination.
The ratio of AOA to AOB in filter media in plants A and B-2 were 5-10 times higher in winter than
those in summer samples. It was presumed that temperature may be affect ammonia oxidizers
community in drinking water treatment process. Besides, the ratio of raw water and filtration was
difference. This implied that community of nitrifying in raw water did not affect community structure
in treatment process. Besides, Nitrobactor below the detect limit on raw water and media. Terminal
restriction fragment (T-RF) 220 bp was dominated ammonia-oxidizing archaea in our study, while
previous study reported that T-RF 169 was dominant in drinking water treatment plant in Japan, the
phenomenon would be interesting.
According to those finding, the contribution of nitrification suggest that ammonia-oxidizing archaea
and Nitrospira may account for most of the ammonia and nitrite oxidation. Furthermore, pre-
chlorination should be avoided to achieve good nitrification. The T-RF 220 bp necessary to do
sequencing in the future.

52
17th European Nitrogen Cycle Meeting, Oslo 2012
P 03
The nitrite:nitrate oxidoreductase system of the anammox bacterium
Kuenenia stuttgartiensis
Naomi M. de Almeida1, Hans J. C. T. Wessels2, Wouter J. Maalcke1, Jan T. Keltjens1,
Mike S. M. Jetten1, Boran Kartal1 1
IWWR, Microbiology, Radboud University Nijmegen, The Netherlands 2
Department of Pediatrics, Nijmegen Center for Mitochondrial Disorders, Radboud University Nijmegen Medical Centre,
Nijmegen, The Netherlands
The genome of the anaerobic ammonium-oxidizing (anammox) bacterium ‘Candidatus Kuenenia
stuttgartiensis’ encodes for a nitrite:nitrate oxidoreductase system that oxidizes nitrite to nitrate,
yielding two high potential electrons. Although the gene cluster kustd1699 – kustd1713 has initially
been annotated as a nitrate reductase, a closer look at the amino acid sequence suggests a more
elaborate role.
According to previous results, the gene products form two main subcomplexes. The soluble
subcomplex catalyzing the nitrite oxidation reaction comprises of three proteins, namely NarG, NarH
and a heme b containing protein. These together contain a molybdenum cofactor, 5 Fe-S clusters
and two heme b’s, all tuned on amino acid level to have the high redox potential needed for the
oxidation reaction of nitrite. In addition, the gene cluster contains a set of genes coding for a broad
repertoire of electron carriers. This includes six putative heme-containing proteins, two putative
blue-copper proteins and a membrane-bound polypeptide showing homology to the quinol-binding
subunit of a cytochrome bd oxidase.
These allow the electrons obtained from nitrite oxidation to be wired either towards the quinone
pool via the membrane bound subunit, or towards different reducing reactions in the cell via the
putative blue copper and heme containing proteins.
Current work includes the purification and characterization of the catalytic subcomplex and the
functional expression of the two blue-copper containing proteins. Further, we investigated the
possible interactions of the catalytic subcomplex with other protein complexes with protein
correlation profiling of purified membranes using LC-MS/MS data from consecutive blue native gel
slices.

53
17th European Nitrogen Cycle Meeting, Oslo 2012
P 04
Genetic and symbiotic diversity of rhizobia isolated from Ethiopian soils
Tulu Degefu1, Endalkachew Wolde-meskel1,2, Binbin Liu1, Anne Willems3, Ilse Cleenwick3 and
Åsa Frostegård1 1Department of Chemistry, Biotechnology and Food Science, Norwegian University of Life Sciences, P.O. Box 5003, NO-
1432 Ås, Norway 2School of Plant and Horticultural Sciences, Hawassa University, P.O. Box 5, Hawassa, Ethiopia
3Laboratorium voor Microbiologie (WE10), Universiteit Gent, KL Ledeganckstraat 35, B-9000
Gent, Belgium
Leguminous trees play an important role in agroforestry in Ethiopia partly due to their ability to
establish a symbiosis with rhizobia for biological nitrogen-fixation (BNF). BNF plays a substantial role
in linking the biological nitrogen cycle between the atmosphere and the earth and hence is a key
factor in agricultural sector and in the rehabilitation of degraded soil by improving nutrient cycling
and other soil physical conditions. By virtue of these, isolation and characterization of the legume
associated rhizobia is crucial to develop inocula and thereby utilize their use for sustainable
agriculture. As the first step to such an approach, 240 rhizobial strains were isolated from root
nodules of a large number of legumes species growing in diverse agro-ecological zones in southern
Ethiopia and characterized based on multilocus sequence analyses of several housekeeping and
symbiosis-related genes. Cross-inoculation experiments were also conducted on a range of legume
species to determine the host ranges. The core gene sequence-based phylogenetic analyses showed
that all test strains represent the branches designating the Rhizobium (5 genospecies),
Mesorhizobium (3 genospecies), Ensifer (7 genospecies) and Bradyrhizobium (5 genospecies), with
the majority of the test strains occupying distinct positions by excluding any defined species in each
of the genus. Further characterizations of a selected strains belonging to the Mesorhizobium genus
from the same collection using DNA-DNA hybridization, cellular fatty acid analyses and additional
housekeeping genes revealed the presence of novel lineages, hence we described three new species
within the genus. Our results while showing discordant phylogeny between housekeeping and
symbiotic gene in some genera like Ensifer and Rhizobium also revealed similar pattern of grouping
in some other genus like Mesorhizobium. The phylogenetic relationship based on the symbiosis-
related genes were not similar to those shown by core genes for the strains represented in Ensifer
and Rhizobium genus suggesting differences in the evolutionary history between the chromosomal
and symbiotic genes. The cross-inoculation experiment, conducted to evaluate the symbiotic
performance of the test strains on a range of legumes species, indicated the potential to improve N
fixation through host selection. Our study confirms a high diversity of rhizobia in Ethiopia and, while
contributing to the general knowledge of the biodiversity within the rhizobial genera, also highlights
the need to focus on previously less explored bio-geographical regions to unravel as yet unidentified
rhizobial resources.

54
17th European Nitrogen Cycle Meeting, Oslo 2012
P 05
Diversity of nitrogen fixing microorganisms from a saline-alkaline former lake
Texcoco, México
Cristina Domínguez-Mendoza, Carolina Castro-Silva, Rocío Alcántara-Hernández, Rodolfo Marsch,
Luc Dendooven* Laboratory of Soil Ecology, Cinvestav, México
*Author for correspondence: Luc Dendooven, Tel. +52 55 5747 33 19, Fax. +52 55 5747 33 13, E-mail:
Soil of the former lake Texcoco is an ‘extreme’ alkaline saline environment with pH>10 and
electrolytic conductivity >10 dS m-1. Although this environment is adverse for N2 fixing
microorganisms, earlier studies showed their presence in this soil. In this study nitrogenase activity
was measured and related with a nifH libraries with the objective to compare the diversity and
phylogenetic relationships of nitrogen fixing microorganisms.

55
17th European Nitrogen Cycle Meeting, Oslo 2012
P 06
Strong reduction of N2O emissions by long term liming in a drained organic soil
Peter Dörsch1, Linn Hovlandsdal1,2, Samson Øpstad2 and Lars Bakken1 1 Norwegian University of Life Sciences (Postboks 5003, 1432 Ås, [email protected])
2 Bioforsk West Fureneset (6967 Hellevik)
Drained peatlands have been reported to be among the strongest sources for soil-borne N2O.
Cultivated organic soils derived from peats are often limed or mixed with mineral soil to raise the
naturally low pH. We previously found that soil acidity strongly inhibits N2O reductase and
hypothesised that long-term liming would reduce N2O emissions by changing the product
stoichiometry of denitrification in favour of N2. Here we report N2O emission fluxes measured in a
liming experiment in Norway (Fjaler, Outer Sunnfjord) where different rates of mineral addition
(shell sand or moraine soil) had been applied more than 30 years ago resulting in distinct soil
pH(H2O) values (4.5, 5.3, 6.5, 7.2 and 7.4). N2O emission fluxes were measured by closed chamber
technique directly after fertilisation (90 kg N ha-1 as NPK) following the first harvest in 2010 and
throughout autumn when soil mineral N concentrations declined. Precipitation after fertilisation
elicited N2O emission fluxes of up to 7000 µg N m-2 h-1, which are among the highest fluxes reported
from land ecosystems ever. The control soil (no liming, pH 4.5) lost 6 kg N2O-N within the first 19
days after fertilisation equalling 6.4% of the applied nitrogen. N2O-N loss was significantly lower in
limed treatments (1.9 kg at pH 7.4), resulting in a significant negative correlation between soil pH
and N2O emission (r2 = 0.6, p<0.05). No such correlation was found after the fertiliser peak in mineral
N had levelled off and N2O emissions decreased to background fluxes. Our data suggest that long-
term liming of acid soils can reduce annual N2O emissions in crop production, supposedly by
increasing the N2/N2O ratio during periods of high denitrification activity.

56
17th European Nitrogen Cycle Meeting, Oslo 2012
P 07
Identification of key player groups of denitrifiers in an organic agricultural soil by
high-throughput sequencing techniques
Binbin Liu1, Xiaojun Zhang3, Lars R. Bakken2 and Åsa Frostegård1 1
Department of Chemistry, Biotechnology and Food Sciences, Norwegian University of Life Sciences, Ås, Norway 2
Department of Plant and Environmental Sciences, Norwegian University of Life Sciences, Ås, Norway 3
Key Laboratory of Microbial Metabolism, Ministry of Education, College of Life Science and Biotechnology, Shanghai
Jiaotong University,Shanghai, China
Our earlier studies of denitrification gene expression in soil show that transcription occurs only
during a few hours after oxygen depletion. DGGE analysis indicated sequential triggering of gene
expression by different populations. We used a robotized incubation system, and guided by the gas
kinetics we sampled soil for mRNA isolation at time intervals (1, 3 and 6 h after O2 depletion).
Functional genes (narG, napA, nirS, nirK, qnorB, nosZ) were amplified from both DNA and cDNA using
ID tagged primers. A total of 210,000 sequences were analyzed, mostly representing Proteobacteria.
The cDNA libraries confirmed that some groups initiated transcription earlier than others. The same
groups were detected both in cDNA and DNA libraries, but in different proportions; those
dominating the DNA library were often less abundant in the cDNA and vice versa. For example,
Dechloromonas sequences constituted 4 and 6% of the total sequences in nirS and napA DNA and
56% and 22% in the corresponding cDNA; Cupriavidus constituted 5% in nirS DNA and 22% in the
cDNA libraries; and Pseudomonas increased from 15% in nosZ DNA to >50% in cDNA while, in
contrast, Herbaspirillum decreased from 58% in DNA to 16% in cDNA. The findings reveal key-player
denitrifiers and are currently used for targeted isolation of these groups.

57
17th European Nitrogen Cycle Meeting, Oslo 2012
P 08
The effect of pH and Oxygen on nitrous oxide emissions from the denitrifying
prokaryote Paracoccus denitrificans 1222.
G. Giannopoulos1, N. Watmough1, P. Poole2 and D. Richardson1 1
School of Biological Sciences, University of East Anglia, Norwich, UK 2 John Innes Center, Norwich, UK
Denitrification forms a part of the bio-geo-chemical cycle in which nitrate is reduced to nitrite, nitric
oxide, nitrous oxide and finally to di-nitrogen. Prokaryotic organisms possess specific
metalloenzymes that reduce nitrogen species; a periplasmic and an inner membrane bound nitrate
reductase, a cytochrome or copper containing nitrite reductase, a cytochrome bc complex nitric
oxide reductase and a copper containing nitrous oxide reductase.
Paracoccus denitrificans PD1222 is a model heterotrophic denitrifying bacterium, capable of nitrate
reduction to di-nitrogen in anaerobic conditions, utilising nitrate as alternative electron acceptor and
succinate as an electron donor. However, in aerobic conditions with nitrate available, P.denitrificans
utilizes oxygen as the final electron acceptor with succinate as an electron donor.
Comparisons of aerobic and anaerobic denitrification as well as the effects of pH in anaerobic
denitrification were undertaken in continuous culture bioreactors, with focus on metabolic profiling,
enzymatic assay and transcriptional regulation. This was carried out using nitrate, nitrite, nitrous
oxide monitoring, immunoblotting, RNA extraction and microarray analysis. The postulated
mechanisms of the denitrification pathway will be discussed.

58
17th European Nitrogen Cycle Meeting, Oslo 2012
P 09
Identification of controlling factors of Nitrous Oxide emission for a drained spruce
forest on peat soil, a modeling approach
He, H.1; Kasimir Klemedtsson, Å.1; Jansson, P.-E.2; Nylinder, J.1; Meyer, A.3; Klemedtsson, L.3 1 Department of Earth Sciences, University of Gothenburg, Gothenburg
2 Land and Water Resources Engineering, Royal Institute of Technology (KTH), Stockholm 3 Department of Plant and
Environmental Sciences, University of Gothenburg, Gothenburg
High nitrous oxide (N2O) emission potential has recently been identified in hemiboreal forest on
drained Histosols (Maljanen et al. 2003; Von Arnold et al. 2005; Ernfors et al. 2007). However, the
environmental factors regulating the emissions in the ecosystems were unclear. To investigate the
importance of different factors on the N2O emission, a modeling approach was accomplished, using
the CoupModel together with Monti-Carlo based multi-criteria calibration method. The model was
made to represent a forest on drained peat soil in south-west Sweden where data of fluxes
combined with basic background information on soil and plant conditions were used. The model
outcome was consistent with measurements of abiotic (soil temperature, net radiation, groundwater
level and soil moisture) and biotic responses (net ecosystem exchange and soil respiration) from the
site. Also, both dynamics and magnitude of N2O emissions were well simulated compared to
measurements. A result for the robustness of the system was the reasonable nitrogen budget
simulated. The informal likelihoods for an ensemble of accepted simulations of N2O emission
dynamics (coefficient of determination, r2) and magnitudes (mean error, ME) were correlated to
calibrated parameters related to nitrification process and drainage depth. By applying Hornberger-
Spear-Young generalized sensitivity analysis, 12 out of 50 parameters were identified as sensitive
parameters. Parameters (Zp, gmfracN2O, nhrateNH4 and cnm) showed both correlation and
sensitivity to N2O emission within the behavior models selected. Nitrification process was identified
as the most important process regulating the N2O emissions for present forest soil. In addition,
drainage of fertile afforested peat soil had a potential increase of N2O emissions, which suggested
one practical mitigation option for N2O emission reduction is to lower drainage pipes or abandon
drainage.
Key words: N2O emission modeling; Peat soil, forest; CoupModel; GLUE
#Contact author: Hongxing He ([email protected])

59
17th European Nitrogen Cycle Meeting, Oslo 2012
P 10
Study of nitrification inhibitors on ammonia oxidizing archaeal community
Yu-Zen Huang1, Toshikazu Fukushima2, Liang-Ming Whang1
1 Department of Environmental Engineering, National Cheng Kung University, Tainan, Taiwan
2 Division of Environmental Engineering, Hokkaido University, Japan
Abstract category: Archaea: important players in diverse microbial ecosystems
Ammonia oxidation is one of the critical pathways in the global nitrogen cycle. It is believed that
ammonia oxidizing bacteria (AOB) is the major group responsible for ammonia oxidation. Recently,
new ammonia-oxidizing organisms belonged to the archaeal domain have been found also involving
in ammonia oxidation. Archaea has been known to have advantage over bacteria in extreme
environments, such as harsh temperature, pH, and has resistance to many inhibitory chemicals in the
environments. Therefore, it may be considered that ammonia oxidizing archaea (AOA) may play
more important role than AOB on ammonia oxidation. Many chemicals, presenting in wastewater
treatment processes, have been reported to inhibit AOB on ammonia oxidation activity. However,
there is no such information available for AOA.
In this study, two laboratory-scale bioreactors with AOA enrichments under different salinity levels
(2.5‰ and 34‰) were used for batch experiments. Although AOA community structures were
different in these two bioreactors, AOA populations were dominant over AOB in both bioreactors.
Batch experiments were conducted to evaluate the resistance of Nitrosomonas europaea pure
culture and AOA enriched cultures to allythiourea and dimethylsulfide. In addition, ampicillin was
added to inhibit AOB activity in batch experiments using AOA enriched cultures. The results
indicated that dimethylsulfide inhibited ammonia oxidation activity of Nitrosomonas europaea, as
well as AOA enrichments in both bioreactors. Addition of 10 mg/L of allythiourea totally inhibited
AOB activity, but resulted in only 50% and 84% inhibition on ammonia oxidation of AOA
enrichments in low and high salinity bioreactors, respectively. The results indicated that AOA
enrichments had a higher resistance to allythiourea than Nitrosomonas europaea, and suggested
that AOA may play an important role in bioreactors treating wastewaters containing chemicals that
are known as inhibitors of AOB.

60
17th European Nitrogen Cycle Meeting, Oslo 2012
P 11
Regulatory effect of soil properties on N2O emission from wheat-growing season in
five soils: field and pot experiment
Lebender, U.1, Senbayram M. 1,2 1Research Center Hanninghof, Yara International , Dülmen, Germany
2IAPN – Institute of Applied Plant Nutrition, Göttingen, Germany
Soils are a major source of N2O emission and the two biological processes (nitrification and
denitrification) are responsible for its production. Chemical and physical properties of soils such as
texture and total carbon (C) are important variables which control the formation of N2O from these
processes. However, the impact of these variables on regulatory mechanisms of N2O formation is still
poorly understood. The first objective of this study is to compare five German soil types for their N2O
emission during the wheat-growing season under field conditions. In addition to the field
experiment, the potential denitrification and respiration rate of the soils have been tested in
incubation experiments under standardized anoxic conditions.
Present field and incubation experiments showed that a higher level of available carbon content of
soils (rather than total soil organic matter content) induced higher N2O emission and denitrification
rate. There was a weak correlation between potential denitrification rate and cumulative N2O
emitted under field conditions over a whole season. However, maximum daily N2O fluxes correlated
significantly with the denitrification potential of soils which may indicate that N2O emissions during
high peak events were derived mainly from denitrification.

61
17th European Nitrogen Cycle Meeting, Oslo 2012
P 12
Denitrifying bacteria produce much N2O at low pH, and we are beginning to understand why
Åsa Frostegård1, Binbin Liu1and Lars Bakken2
1 Department of Chemistry, Biotechnology and Food Sciences, Norwegian University of Life Sciences, Ås, Norway
2 Department of Plant and Environmental Sciences, Norwegian University of Life Sciences, Ås, Norway
Denitrification in soil is a major source of atmospheric N2O, a potent greenhouse gas and ozone-
depleting compound. Denitrifying prokaryotes use NOx as terminal electron acceptors in response to
oxygen depletion. The process emits a mixture of NO, N2O and N2, depending on the relative activity
of the enzymes catalyzing the stepwise reduction of NO3- to N2O and finally to N2. Soil pH emerges as
a master variable determining the microbial community composition as well as its denitrification
product ratio (N2O/N2). Several studies demonstrate that this ratio is higher in acid than in alkaline
soil. It is therefore likely that emissions of N2O from agro-ecosystems will increase in large parts of
the world where soil pH is decreasing due to intensified management and increased use of chemical
fertilizers. Considering its immense implications, surprisingly few attempts have been made to
unravel the mechanisms involved in the pH-control of the product stoichiometry of denitrification.
We investigated the kinetics of gas transformations (O2, NO, N2O and N2) and transcription of
functional genes in intact soil samples from long-term liming experiments. Expression of nirS
(encoding nitrite reductase) and nosZ (encoding N2OR) was generally lower at pH6 compared to pH8,
but the nosZ /nirS transcript ratios were similar or even higher at pH6. These results were largely
corroborated in refined experiments using Nycodenz-extracted bacteria from the same soils, which
allowed us to better control the pH levels experienced by the cells. The findings indicated that the
higher N2O/N2 product ratios at pH6 were due to a post-transcriptional effect. Experiments with P.
denitrificans demonstrated N2O/N2 ratios ≤ 10-3 at pH7 while N2O reduction was severely inhibited
by suboptimal pH (~100% N2O production at pH=6). This inhibition occurred during protein
synthesis/assembly rather than at the transcription level since the relative transcription rate of nosZ
versus nirS and norB was unaffected by pH, and since low pH had a moderate effect on the N2OR
activity in cells with a denitrification proteome assembled at pH 7. Ongoing investigations of the
N2OR protein structure will show if this hypothesis is correct. The results are important for the
understanding of main factors affecting N2O emissions from terrestrial ecosystems.

62
17th European Nitrogen Cycle Meeting, Oslo 2012
P 13
Nitrous oxide turnover in the nitrate-ammonifying Epsilonproteobacterium
Wolinella succinogenes
Monique Luckmann1 and Jörg Simon1 1Microbial Energy Conversion and Biotechnology, Department of Biology, Technische Universität Darmstadt, Germany
Global warming is moving more and more to the public consciousness. Besides the commonly
mentioned carbon dioxide and methane, nitrous oxide (N2O) is one of the most important
greenhouse gases and accounts for about 10% of the anthropogenic greenhouse effect.
Several microbial energy converting processes appear to be the most important sources of N2O
emission and the responsible organisms comprise denitrifiers and ammonifiers of nitrate as well as
nitrifying bacteria and archaea. Interestingly, some respiratory nitrate-ammonifying
Epsilonproteobacteria are able to reduce nitrous oxide to dinitrogen via an unconventional
cytochrome c nitrous oxide reductase (cNosZ). The energy metabolism of one of these bacteria,
Wolinella succinogenes, has been characterized thoroughly in the past. The cells are able to use
formate or hydrogen gas as electron donors together with typical terminal electron acceptors like,
for example, fumarate, polysulfide, nitrate, nitrite or nitrous oxide.
Despite the presence of cNosZ there is neither a typical nitric oxide-producing nitrite reductase (NirS,
NirK) nor a nitric oxide reductase of the Nor-type present in W. succinogenes. It is speculated that
N2O might be produced during nitrite and/or NO detoxification by the flavodiiron protein Fdp. Using
different ratios of the electron acceptor nitrate and the donor formate the production and
consumption of nitrous oxide during nitrate respiration of W. succinogenes and various mutant
strains was characterized. Furthermore, a model of the electron transport chain of W. succinogenes
nitrous oxide respiration will be presented.

63
17th European Nitrogen Cycle Meeting, Oslo 2012
P 14
Spatial patterns of soil pH and N2O emissions in field experiments
Aina R. Lundon1, Lars Bakken2, Audun Korsæth1& Peter Dörsch2 1
Bioforsk – Norwegian Institute for Agricultural and Environmental Research. 2 Department of Plant and Environmental Sciences, Norwegian University of Life Sciences.
Emissions of N2O form agricultural soils represent a significant part of the anthropogenic emission of
nitrous oxide (N2O). pH might be an important factor regulating the magnitude of emission form
arable land. Three plots with wheat stubble with similar texture but different levels of pH in a field
trial were selected for measurements of N2O emissions in the off season between September 2010
and May 2011. The amount of N2O-N in µg m-2 h-1 were calculated from analysed gas samples
obtained from static flux chambers form September 2010 till May 2011. Four static flux chambers
were placed within each plot and the levels of pH in the total 12 chamber locations varied between
5.38 and 5.95 (pH measured in CaCl2 solution). The results showed a decrease of emissions in
samples from the plot with average pH 5.80 compared to the plot with average pH 5.48. The plot
with average pH 5.54 did not differ from the other two plots. The emissions varied throughout the
period of sampling with generally low emissions, but with a distinct peak during snowmelt in April.
The results suggest that even marginal increases in soil pH may indeed reduce the emission of N2O.

64
17th European Nitrogen Cycle Meeting, Oslo 2012
P 15
A nitric oxide-producing octaheme enzyme from Kuenenia stuttgartiensis
WJ Maalcke1, A Dietl2, SJ Marritt3, MSM Jetten1, JTM Keltjens1, JN Butt3, TRM Barends2, B Kartal1 1 Department of Microbiology, Radboud University Nijmegen, Heijendaalseweg 135, 6525 AJ, Nijmegen, The Netherlands
2 Department of Biomolecular Mechanisms, Max Planck Institute for Medical Research, Jahnstrasse 29, D-69120,
Heidelberg, Germany 3 School of Chemistry, University of East Anglia, Earlham Park, Norwich, NR4 7TJ, United Kingdom
Anaerobic ammonium-oxidizing (anammox) bacteria combine ammonium and nitrite to form
dinitrogen gas. Anammox bacteria are abundantly present in the oxygen minimum zones of the
world’s oceans and have a significant role in global dinitrogen gas production. They are also
successfully applied in cost-effective and environment-friendly ways to treat industrial and municipal
wastewater. Although the physiology of anammox bacteria is quite well understood, the
biochemistry behind it is less clear. One of the persisting anammox enigmas is the presence of ten
copies of an open reading frame that resembles the octaheme enzyme hydroxylamine
oxidoreductase (HAO). HAO converts hydroxylamine to nitrite in aerobic ammonium oxidizing
bacteria. The function of one of these enzymes (kustc0694) is predicted to be hydrazine oxidation to
dinitrogen gas, whereas the functions of the other nine octaheme proteins remain to be established.
Here, a highly expressed octaheme protein was purified from native Kuenenia stuttgartiensis
biomass and characterized. MALDI-TOF mass spectrometry confirmed that the enzyme was encoded
by the open reading frame kustc1061. Enzyme kinetics, assessed by using cytochrome c-based assays
and gas chromatography coupled to mass spectrometry, showed kustc1061 to be a hydroxylamine
oxidizing enzyme that produces nitric oxide instead of nitrite. Protein film electrochemistry revealed
the midpoint potentials of the eight separate heme-groups, and elucidation of the crystal structure
enabled us to form a hypothetical mechanism of hydroxylamine oxidation to nitric oxide. As the
proposed hydrazine-oxidizing enzyme is inhibited by hydroxylamine, the physiological role of
kustc1061 is proposed to be hydroxylamine detoxification

65
17th European Nitrogen Cycle Meeting, Oslo 2012
P 16 The purple form of Marinobacter hydrocarbonoclasticus nitrous oxide reductase
Sofia R. Pauleta1,*, Simone Dell'Acqua1,2, Ana Rute Nunes1, Susana Ramos1, Oliver Einsle3, José J.G.
Moura1, Isabel Moura1 1REQUIMTE/CQFB, Departamento de Química, Faculdade de Ciências e Tecnologia, Universidade Nova de Lisboa, 2829-
516 Caparica, Portugal. 2Dipartimento di Chimica, Università di Pavia, Via Taramelli 12, 27100 Pavia, Italy.
3Institute of Organic Chemistry and Biochemistry and BIOSS Centre for Biological Signalling Studies, Albert-Ludwigs-
Universität Freiburg, 79104 Freiburg, Germany.
Email: [email protected]
Nitrous oxide reductase catalyses the final step of the denitrification pathway, the reduction of
nitrous oxide to nitrogen. The catalytic centre, CuZ, is a unique tetranuclear copper centre bridged
by inorganic sulphur in a tetrahedron arrangement that can have different oxidation states.
Previously, Marinobacter hydrocarbonoclasticus nitrous oxide reductase was isolated with CuZ
centre as CuZ*, in the [1Cu2+:3Cu+] redox state, which is redox inert and requires a prolonged
incubation under reductive conditions to be activated. Here, we report the isolation of nitrous oxide
reductase from Marinobacter hydrocarbonoclasticus in the “purple” form, in which CuZ centre is in
the oxidized [2Cu2+:2Cu+] redox state and is redox active. This form of the enzyme was isolated in the
presence of oxygen from a microaerobic growth in the presence of nitrate and also from a strictly
anaerobic growth. The purple form of the enzyme was biochemically characterized and was shown
to be a redox active species, though it is still catalytically non-competent, as its specific activity is
lower than the one of the activated fully reduced enzyme and comparable to the one of the enzyme
with CuZ centre in either the [1Cu2+:3Cu+] redox state or in the redox inactive CuZ* state.
Acknowledgements: We thank FCT-MCTES for the financial support (PTDC/QUI-BIQ/116481/2010), and CRUP-DAAD
Luso-German Integrated Actions.
References: Dell’Acqua et al. JBIC 16 (2011) 183-194. Dell’Acqua et al. JBIC 16 (2011) 1241-1254. Dell’Acqua et al. Philos.
Trans. R. Soc. B 367 (2012) 1204-1212.

66
17th European Nitrogen Cycle Meeting, Oslo 2012
P 17
Autotrophic carbon dioxide fixation via the Calvin-Benson-Bassham cycle by the
denitrifying anaerobic methanotroph Candidatus Methylomirabilis oxyfera
O Rasigraf, DM Kool, MSM Jetten, KF Ettwig
Anaerobic oxidation of methane coupled to nitrite reduction has recently been shown to be
performed by a member of the NC10 candidate division, Methylomirabilis oxyfera. Though living
under anoxic conditions, M. oxyfera oxidizes methane via the 'conventional' aerobic pathway
starting with particulate methane monooxygenase. This is accomplished by the intracellular
dismutation of nitric oxide to molecular oxygen and nitrogen.
In contrast to proteobacterial methanotrophs, however, M. oxyfera does not encode complete
pathways to assimilate carbon from methane (serine and ribulose monophosphate pathways) in its
genome. In line with this, no significant incorporation of carbon from 13C-labelled methane into
biomass could be demonstrated. Like acidiphilic methanotrophs of the phylum Verrucomicrobia,
M. oxyfera possesses the whole gene set encoding the Calvin-Benson-Bassham (CBB) cycle, which is
also being expressed.
Here we provide evidence for autotrophic carbon dioxide fixation by M. oxyfera via the CBB cycle.
Cell-free extracts from the enrichment culture consisting of ~80% of M. oxyfera bacteria exhibited a
specific ribulose-1,5-bisphosphate carboxylase/oxygenase (RubisCO) activity of ~0.2 nmol CO2 mg-1
protein min-1. This activity accounted for up to 10% of the total methane oxidation activity. In
addition, stable isotope studies with whole cells in batch incubations supplied with 13C-labelled
methane and carbon dioxide were performed. The results showed a significant enrichment in 13C of
putatively specific M. oxyfera-like lipids after the incubation with labeled carbon dioxide and
unlabeled methane but not vice versa. Altogether, these results indicate that CBB cycle is active and
plays a major role in carbon assimilation by M. oxyfera bacteria, and show that autotrophy might be
more widespread among methanotrophs than previously believed.

67
17th European Nitrogen Cycle Meeting, Oslo 2012
P 18
Isotopomer ratios of N2O produced during denitrification by fungal pure cultures and
associated oxygen exchange with water
Rohe, L.1*, Braker, G2, Well, R.1, Giesemann, A.1, Anderson, T.-H.1, Wrage, N.3
1 Johann
Heinrich von Thuenen-Institute, Institute of Agricultural Climate Research, Braunschweig, Germany
2 Max Planck Institute for Terrestrial Microbiology, Marburg, Germany
3 Faculty of Life Sciences, Agricultural Sciences, Rhine-Waal University of Applied Sciences, Kleve, Germany
* Corresponding author: [email protected]; +49 (0)531 5962655
N2O from soil denitrification originates from bacteria and - to an unknown extent - also from fungi. In
pure culture studies, bacterial and fungal denitrification showed differences in isotopomer ratios of
N2O. Isotopomer ratios of N2O can be expressed as 15N site preference (SP), i.e. the difference
between δ15N of the central and terminal N-position of the asymmetric N2O molecule and might be
suitable to distinguish between bacterial and fungal N2O under denitrifying soil conditions.
Oxygen exchange takes place between H2O and intermediates during the denitrification process.
Several studies showed that O-exchange of bacterial denitrifiers varies, but information about fungal
O-exchanges is lacking.
The objectives of the study presented are i) to verify SP of fungal N2O reported for two strains from
pure culture studies and ii) to analyze the oxygen exchange between intermediates and H2O during
denitrification in fungal pure cultures. Six different fungal pure cultures known to be capable of
denitrification were incubated under anaerobic conditions, either with nitrite or nitrate. Gas samples
were analysed for concentration and isotopic signatures (SP, average δ15N, δ18O) of N2O. To
investigate the oxygen exchange, both treatments were also established in a tracer experiment
where 18O-labelled water was used in the medium.
The fungal strains evaluated so far indicate similar SP as reported previously. Concerning the amount
of N2O produced, the nitrite led to a higher yield. Results of O-exchange with 18O-labelled water
during fungal denitrification will be presented.

68
17th European Nitrogen Cycle Meeting, Oslo 2012
P 19
Regulation of denitrification: pathways to nitrous oxide and dinitrogen
S Saggar1, N Jha2, J Deslippe1, J Luo3, R Tillman2, D Giltrap1, H Jamali1 1Global Change Processes Group, Landcare Research, Private Bag 11052, Palmerston North 4442, New Zealand
2Soil & Earth Sciences Group, Institute of Natural Resources, Massey University, Private Bag 11222 Palmerston North
4442, New Zealand 3Climate, Land & Environment Section, AgResearch, Ruakura Research Centre, Private Bag 3123, Hamilton 3240, New
Zealand.
Denitrification is the primary process producing nitrous oxide (N2O) in temperate grassland soils and
it accounts globally for about 60% of total N2O emissions to the atmosphere. We lack knowledge of
reliable quantitative denitrification rates for grazed pasture soils and how these rates vary spatially
and temporally. Issues that hinder this understanding include: difficulties in measuring denitrification
rates, the large spatial and temporal heterogeneity of denitrification products, and the large
background atmospheric N2 concentration. An understanding of the biological processes, and the
soil and environmental factors that have the potential to reduce the production of N2O during
denitrification, is vital for the development of novel and effective N2O mitigation technologies. Our
objectives were to develop and test methods to identify pathways to N2O and N2 production during
denitrification such as enzyme activities, microbial biomass and microbial communities, and
determine the soil factors contributing to spatial variability in denitrification potential of dairy soils.
Experiments were conducted initially to standardise the techniques and develop the protocols for
the measurements of denitrification enzyme activity (DEA), in-situ denitrification rate (DR) in soils.
This followed collection of 10 surface (0-10 cm) and subsurface dairy pasture soils varying in texture,
mineralogy, pH, organic C, N, Olsen P and management history representing North and South Islands
of New Zealand. The DEA, DR, microbial biomass and the N2O to dinitrogen (N2) product ratio
determined using the standardised techniques showed large variations among soils. The amount of
N2O produced was correlated with soil explanatory and response variables to determine the key soil
variables contributing to denitrification potential. Characterisation of denitrifier community by
Terminal Restriction Fragment Length Polymorphism analysis (T-RFLP) of functional genes (nirS, nirK
and nosZ) and quantification of functional gene copy numbers in soils by real time quantitative
polymerase chain reaction (qPCR) has been estimated. Detailed statistical analysis is currently
underway to understand correlations between DR, DEA, soils physico-chemical properties with
denitrifier community structure and gene abundance.

69
17th European Nitrogen Cycle Meeting, Oslo 2012
P 20
Off-season N2O losses from clover-rich swards
Ievina Sturite1, Synnøve Rivedal1, Peter Dörsch2 1
Norwegian Institute for Agricultural and Environmental Research, Norway 2
Norwegian University of Life Sciences, Department of Plant and Environmental Sciences, NO-1432 Ås, Norway
In northern areas, the long winter period between cropping seasons is a challenge to keep nitrogen
(N) within the field, particularly in swards with high clover content. The hypothesis tested was that
freezing-thawing episodes cause N leakage from the standing plant biomass and clover-rich swards
therefore have higher nitrous oxide (N2O) emissions than pure grass swards during the winter.
Gaseous emissions from pure clover and grass stands and from mixed stands containing 30% clover
were measured in western and northern Norway. The sampling time lasted from October 2011 to
May 2012. In order to meet the most important periods of N2O emissions during the winter, the
sampling frequency varied at each site. Preliminary results indicate that stands with clover had
higher N2O emissions than pure grass stands before air and soil temperatures decreased below zero.
The first freezing-thawing cycle increased N2O emissions markedly for treatments containing clover.
Air temperatures around the freezing point resulting in unstable and heterogeneous snow cover led
to high but variable N2O emissions irrespective of treatment. In general, for clover-rich stands N2O
emissions were somewhat higher compared to pure grass stands over the entire seven-month
experiment. This indicates that clover-derived N from damaged plant tissue may be subject to N
pathways that lead to N2O emissions.
70
17th European Nitrogen Cycle Meeting, Oslo 2012
Delegate list
Tulu Abdi Norwegian University of Life Sciences Norway
[email protected] 4746277075
Liz Baggs University of Aberdeen UK
[email protected] 441 224 272 691
Lars Bakken Norwegian University of Life Sciences Norway
[email protected] 4797021822
Gracia Becerra University of Extremadura Spain
[email protected] 3492725700
Linda Bergaust Norwegian University of Life Sciences Norway
[email protected] 4764965819
Charlene Bradley University of Birmingham UK
[email protected] 7772852751
Kristof Brenziger Max Plank Institute for Terrestrial Microbiology Marburg Germany
[email protected] 496 421 178 732
Carlos Bricio CSIC Spain
[email protected] 341964525
Emilio Bueno CSIC Spain
[email protected] 958181600
Klaus Butterbach-Bahl Karlsruhe Institute of Technology Germany
[email protected] 498 821 183 136
Tom Clarke University of East Anglia UK
Jeff Cole University of Birmingham UK
[email protected] 441 214 145 440
Naomi de Almeida Radboud University Nijmegen The Netherlands
[email protected] 31 24 3652704
Helen Decleyre Ghent University Belgium
[email protected] 3292645134
Maria J. Delgado CSIC Spain
[email protected] 958181600 (ext. 327)
Christina Domínguez CINVESTAV Mexico
[email protected] 555 539 644 424
Peter Dörsch Norwegian University of Life Sciences Norway
[email protected] 4 791 663 372
Stuart Ferguson University of Oxford UK
[email protected] 441 869 613 299
Mary Firestone UC Berkeley USA
[email protected] 510-6423677
Åsa Frostegård Norwegian University of Life Sciences Norway
[email protected] 4764965880
Andrew Gates University of East Anglia UK
[email protected] 1603592931
71
17th European Nitrogen Cycle Meeting, Oslo 2012
George Giannopoulos University of East Anglia UK
[email protected] +44 (0) 7747336547
Suzanne Haaijer Radboud University Nijmegen The Netherlands
[email protected] 31243652568
Jon Fredrik Hanssen Norwegian University of Life Sciences Norway
[email protected] 4764965891
Katherine Hartop University of East Anglia UK
[email protected] 7999311294
Junaid Hassan Norwegian University of Life Sciences Norway
[email protected] 4746258806
Hongxing He University of Gothenburg Sweden
[email protected] 460 317 862 811
Miriam Herold University of Aberdeen UK
[email protected] 441 224 272 700
Kim Heylen Ghent University Belgium
[email protected] 3292645142
Marcus Horn University of Bayreuth Germany
[email protected] 49921555620
Yu-Tzu Huang Chung-Yuan Christian University Taiwan
[email protected] 886 3 265 4913
Yu-Zen Huang National Cheng Kung University Taiwan
[email protected] 886-6-2757575-65850-37
M. Isabel Igeño University of Extremadura Spain
[email protected] 3492725700 Ext:51333
Hizbullah Jamali Lancare Research New Zealand New Zealand
[email protected] 6 463 534 931
Mike Jetten Radboud University Nijmegen The Netherlands
[email protected] 31243652940
Erik Joner Bioforsk Norway
[email protected] 4 792 833 168
Melanie Kern Technische Universität Darmstad Germany
[email protected] 49-6151165239
Catherine Larose University of Lyons France
[email protected] 33669719019
Ulrike Lebender Research Center Hanninghof, YARA GmbH & Co. KG Germany
[email protected] +492594798122
Jih-Gaw Lin National Chiao Tung University Taiwan
[email protected] 886-3-5722681
Natalie Lim Norwegian University of Life Sciences Norway
[email protected] 4796839503
Binbin Liu Norwegian University of Life Sciences Norway
[email protected] 4764965815
Sebastian Lücker University of Vienna Austria
[email protected] +43 1 4277 57316
Monique Luckmann TU Darmstadt Germany
[email protected] 6 151 165 239
72
17th European Nitrogen Cycle Meeting, Oslo 2012
Aina R. Lundon Bioforsk Norway
[email protected] 4740483136
Victor M. Luque-Almagro Universidad de Córdoba Spain
[email protected] 34957218318
Wouter Maalcke Radboud University Nijmegen The Netherlands
[email protected] 0031-24-3652704
Daniel Mania Norwegian University of Life Sciences Norway
[email protected] 0047-46231353
Lars Molstad Norwegian University of Life Sciences Norway
[email protected] 0047-99791282
Nick Morley University of Aberdeen UK
[email protected] +44(0)1224 272698
Isabel Moura REQUIMTE-CQFB Portugal
[email protected] 351212948385
Shahid Nadeem Norwegian University of Life Sciences Norway
[email protected] 4746270730
Graeme Nichol University of Aberdeen UK
[email protected] 441 224 272 258
Lars Peter Nielsen Aarhus University Denmark
[email protected] 4 560 202 654
Sofia Pauleta REQUIMTE-CQFB Portugal
[email protected] 351212948385
Zhi Qu Norwegian University of Life Sciences Norway
[email protected] 0047-96876850
Olivia Rasigraf Radboud University Nijmegen The Netherlands
[email protected] 31617073948
Nani Raut Norwegian University of Life Sciences Norway
[email protected] 0047-96869118
David Richardson University of East Anglia UK
[email protected] 441 603 593 250
M. Dolores Roldán Universidad de Córdoba Spain
[email protected] 34957218318
Sebastian Runkel University of East Anglia UK
[email protected] 447 909 157 820
Cristina Sanchez Tohoku University Japan
[email protected] 81-22-217-5687
Andreas Schramm Aarhus University Denmark
[email protected] 4 560 202 659
Miloslav Simek Institute of Soil Biology, Biology Centre Czech Republic
[email protected] 420 723 734 722
Jörg Simon Technische Universität Darmstad Germany
[email protected] 491 704 354 951
Bishal Sitaula Norwegian University of Life Sciences Norway
[email protected] 0047-47888760
Daan Speth Radboud University Nijmegen The Netherlands
[email protected] 31243652569
73
17th European Nitrogen Cycle Meeting, Oslo 2012
Ievina Sturite Bioforsk Norway
[email protected] 4 793 499 484
Matthew Sullivan University of East Anglia UK
[email protected] 447 881 490 099
Maria J. Torres CSIC Spain
[email protected] 958181600 (ext. 327)
Rannei Tjåland Norwegian University of Life Sciences Norway
[email protected] 0047-64965904
Maartje van Kessel Radboud University Nijmegen The Netherlands
[email protected] +31 24 3652704
Rob van Spanning VU University Amsterdam The Netherlands
[email protected] 31650536015
Claire Vine University of Birmingham UK
[email protected] 441 214 145 435
Reinhard Well Institute of Agricultural Climate Research , Johann
Heinrich von Thünen-Institut
Germany
[email protected] 495 315 962 535
Liang-Ming Whang National Cheng Kung University Taiwan
[email protected] 011-886-6-2757575-65837
Daniela Wischer University of East Anglia UK
[email protected] 447 743 364 277
Hugh Woolfenden University of East Anglia UK
[email protected] 447 796 950 963
Xiaojun Zhang Shanghai Jiao Tong University China
[email protected] +86 21 34204878
Jing Zhu Norwegian University of Life Sciences Norway
[email protected] 0047-46259348





























