Embed Size (px)
Citation preview

MOLECULAR GENETIC ANALYSIS AND ECOLOGICAL EVIDENCE REVEALS MULTIPLE CRYPTIC SPECIES AMONG THYNNINE WASP POLLINATORS OF SEXUALLY DECEPTIVE ORCHIDS
PREPARED BY:
NURFARAH AIN LIMIN (P74703)

INTRODUCTION
Thynnine wasps
Neozeleboria cryptoides
Male: strong & active flyers
Female: flightless & live underground
Sex pheromone: 2-ethyl-5-propylcyclohexan-1,3-
dione(chiloglottone 1)
Males showed equal preference
for synthetic compound
Mating:Females emerge from ground
& emit volatile sex pheromone to attract males.Males respond in seconds,
readily locate hidden females-proves chemical cues
are vital
Specific pollinator of
sexually deceptive
orchids

Chiloglottis trapeziformis
Sexually deceptive Orchids
Produce same novel compound to attract male N.cryptoides.
Chiloglottone 1: semiochemical involved in intra-/interspecies communication
Pollination occur during premating routine/
attempted to mate with flower
Pollination is highly specific with average of 1.1 pollinator species
per orchid
Pollination by sexual deception-Sexually lure male wasps by mimicry of sex pheromone

Bioassays with synthetic chiloglottones indicate 2 mechanisms to control orchid-pollinator specificity:
1. A single specific compound is required for pollinator attraction. Eg: C. trapeziformis & chiloglottone 1, with different compounds attract different specific pollinators
2. Two/more compounds in a particular blend trigger specific attraction. Eg: Allopatric Chiloglottis species known to use same semiochemical but attract different pollinator species

Prediction: the pollinator pheromones will be either identical to orchid semiochemical as in C.trapeziformis/N.cryptoides or will share same major components in similar proportions
Neozeleboria wasps that appear to be prime candidate for cryptic speciation associated with chemical rather than morphological change
Aim: To investigate this prospect among those species of
Neozeleboria genus that are involved in orchid pollination

Materials & methods The wasp genus Neozeleboria are divided into:
ProximaMonticol
aCryptoide
s
Impatiens
3 clades
Sub-clade

All wasp samples are associated with orchids in one three ways: Specific orchid pollinators whose behavior included attempted
copulation, pollen removal & pollen deposition Minor responders. They occasionally respond to orchids whose
behavior didn’t result in pollination (Eg: approach only) Responders to synthetic semiochemicals but not yet known to
be pollinators or minor responders

Samples represent 18 sites across South-Eastern Australia, from northern New South Wales to Tasmania
3 wasps chosen for sequencing from each site. Sequencing is performed at:
- 1 mitochondrial locus: Cytochrome c
oxidase 1 (CO1)
-2 nuclear loci: rhodopsin & wingless

DNA extraction, PCR &
Sequencing
Sequencing analysis
Samples and groups of interest
Statistical parsimony analysis
Population genetic analysis
Morphological identification
Consideration of candidate cryptic taxa

DNA extraction, PCR & sequencing
PCR reaction (40µl) contained approximately:
1-5ng of DNA template
PCR buffer
dNTPs
Forward & reverse primers
Taq Polymerase
Amplification:
Performed with 3 min denaturation at 94°C
30 cycles with annealing at 49°C followed by 10 min extension at 72°C
PCR products were purified with minor modification using ExoSAP-IT clean-up protocol

SEQUENCE ANALYSIS
The 3 molecular data sets were analyzed both separately and together. Maximum likelihood analyses & bootstrap tests are conducted
For comparison with maximum likelihood analyses, Bayesian and parsimony tree-estimation was performed

SAMPLES & GROUPS OF INTEREST
24 groups of interest are identified Based on inspection of combined 3 loci
maximum likelihood best tree and associated bootstrap values

Statistical parsimony analysis
To identify groups of interest by following method of Templeton et al.
In brief,the method estimates genealogical relationships among sequences & identifies sub-networks
CO1 data sets are compiled and trimmed to remove missing data before being analyzed separately for Proxima, Cryptoids & Monticola

Population genetic analysis
Performed Analysis of Molecular Variance (AMOVA) for mtDNA data set
To investigate the partitioning of genetic variation within and among various priori groupings of samples

Morphological identification
To assess whether groups of interest could be distinguished by current set of 40 morphological characteristics used to identify Neozeleboria wasp
One individual from each group was examined closely by 3rd author (Graham R.B.)
Emphasis on structure of male genitalia. A difference was defined as a minimum of three minor differences in the un-dissected male genitalia
Specimens were coded & submitted for identification without collection of biological data. As blind test, thus identification was based only on morphological

Consideration of candidate cryptic taxa
Probable cryptic taxa were identified by being morphologically similar but genetically distinct
With strong support from non-molecular evidence such as different orchids associations

RESULTS
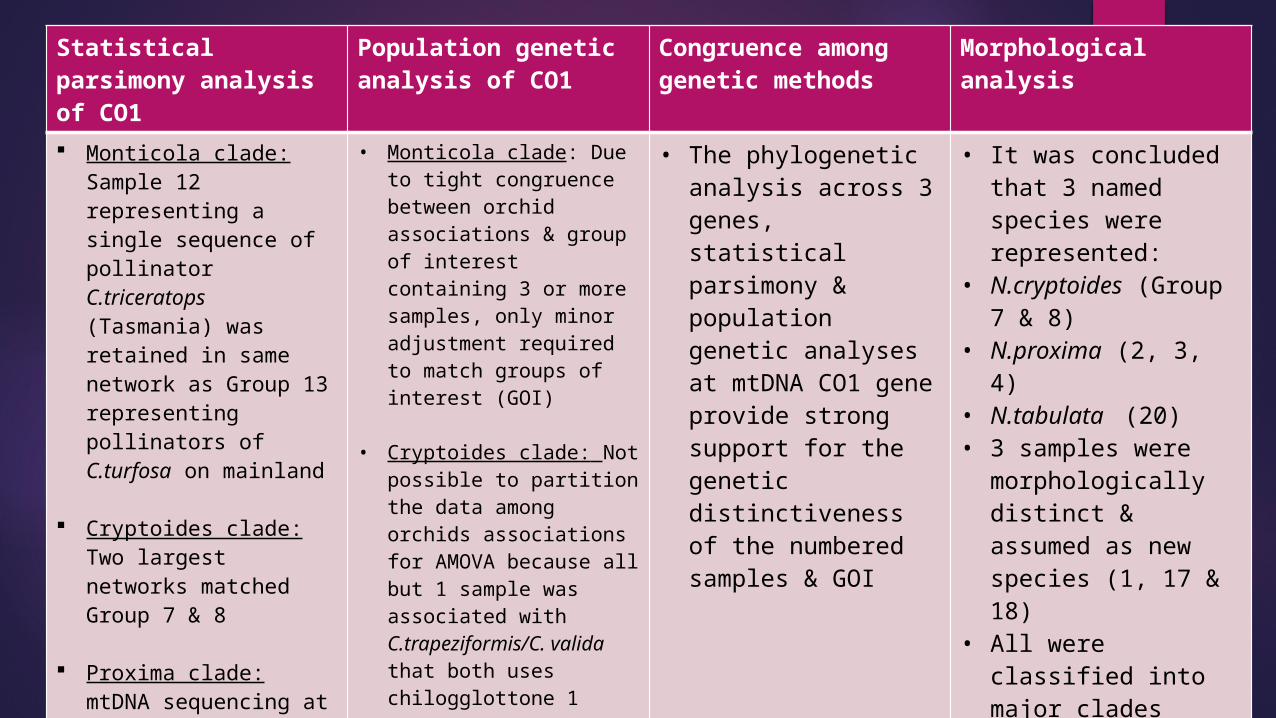
Statistical parsimony analysis of CO1
Population genetic analysis of CO1
Congruence among genetic methods
Morphological analysis
Monticola clade: Sample 12 representing a single sequence of pollinator C.triceratops (Tasmania) was retained in same network as Group 13 representing pollinators of C.turfosa on mainland
Cryptoides clade: Two largest networks matched Group 7 & 8
Proxima clade: mtDNA sequencing at CO1 failed for Group 3
• Monticola clade: Due to tight congruence between orchid associations & group of interest containing 3 or more samples, only minor adjustment required to match groups of interest (GOI)
• Cryptoides clade: Not possible to partition the data among orchids associations for AMOVA because all but 1 sample was associated with C.trapeziformis/C. valida that both uses chilogglottone 1
• Proxima clade: Due to a lack of sufficient replication, AMOVA was not feasible
• The phylogenetic analysis across 3 genes, statistical parsimony & population genetic analyses at mtDNA CO1 gene provide strong support for the genetic distinctiveness of the numbered samples & GOI
• It was concluded that 3 named species were represented:
• N.cryptoides (Group 7 & 8)
• N.proxima (2, 3, 4)• N.tabulata (20)• 3 samples were
morphologically distinct & assumed as new species (1, 17 & 18)
• All were classified into major clades (Proxima, Cryptoides or Monticola)

DISCUSSIONS Consistent with prediction made, a relationship between defined wasp
groups, their orchid associations & semiochemicals was found
Monticola clade Crypticoides clade Proxima clade
There is evidence for pheromone sharing among pollinators. Eg: Wasp groups 12-14 were associated with orchid taxa that use chiloglottone 4
All wasps appear to respond to chiloglottone 1only
Illustrate pheromone sharing among Crypticoides & Monticola
Wasp Group 4 associated with C . trilabra that produces chiloglottone1 & 3
Wasp Group 2 associated with C. seminuda (chiloglottone 1)

Majority of wasp groups were associated with 1 orchid species
Despite strong association with 1 orchid, there is pheromone sharing among wasp groups
The collective phylogenetic, statistical parsimony, population genetic, ecological & geographic evidences support the hypothesis that the wasp samples & GOI (Group 1-24) are taxonomically distinct

Are there cryptic species? Group 7 within Cryptoides clade. It was morphologically
identified as N.cryptoides but was characterized by a deep phylogenetic divergence from N.cryptoides (Group 8).
All 4 groups within Impatiens complex (Group 21-24) were morphologically indistinguishable
Within Proxima clade, Groups 2-4 were identified to be morphologically as N.proxima
Overall, up to 16 groups out of 24 may represent genetically distinct but cryptic species

IMPLICATIONS FOR EVOLUTION OF NEOZELEBORIA WASP
Genitalia are predicted to evolve rapidly & divergently under sexual selection. It may be the 1st morphological characters to define newly formed species
In this study, multiple cryptic thynnine wasp taxa that lacked diagnostic differences in male genitalia were uncovered
This is tally with the observation: chemical rather than visual cues are most important for wasp mate attraction.

IMPLICATIONS FOR EVOLUTION OF THE EVOLUTION AND DIVERSIFICATION OF ORCHIDS
The exploitation of Neozeleboria wasps by Chiloglottis orchids led to hypothesise that this strong association might limit the available pool of pollinators and therefore, orchid speciation
But the findings of multiple cryptic species in Neozeleboria suggest that the wasp genus maybe larger than previously thought
So rather than constraining evolution of orchid genus, species diverstiy within the wasp may have actually enabled diversification by providing diverse pool of species pollinator






























