Embed Size (px)
Citation preview

Deoxyribonucleic Acid
(DNA)


The double helix

Nitrogenous Bases and Pentose Sugars

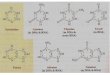
Purine and Pyrimidine Structure
(1) Pyrimidines are planar
(2) Purines are nearly planar
(3) Numbering is different

Numbering Is Different


Bases Have Tautomeric Forms
Uracil


Nucleosides vs. Nucleotides
Glycosidic bond

Nucleotides formed by condensation reactions


Monophosphates

Deoxyribonucleotides

Ribonucleotides

Only RNA Is Hydrolyzed by Base

Nucleoside Diphosphate and Triphosphate

Dinucleotides and Polynucleotides
Ester bonds

Watson-Crick Base Pairs
A=T
G=C

Hoogsteen Base Pairs

Other Base Pairs Are Possible
Homo Purines Hetero PurinesWatson-Crick,
Reverse Watson-Crick, Hoogsteen,
Reverse Hoogsteen, Wobble,
Reverse Wobble

Base Pairing Can Result in Alternative DNA Structures
Triplex Tetraplex
Hairpin Loop Cruciform

• Periodicity: A pair of strong vertical arcs (C & N atoms) indicate a very regular periodicity of 3.4 Å along the axis of the DNA fiber.
• Astbury suggested that bases were stacked on top of each other "like a pile of pennies".
• Helical nature: Cross pattern of electron density indicates DNA helix and angles show how tightly it is wound.
• Diameter: lateral scattering from electron dense P & O atoms.

DNase can only cleave external bond demonstrating periodicity

Watson and Crick Model (1953)
• 2 long polynucleotide chains coiled around a central axis
• Bases are 3.4 Å (0.34 nm) apart on inside of helix
• Bases flat & lie perpendicular to the axis
• Complete turn = 34 Å • 10 bases/turn• Diameter = 20 Å• Alternating major and
minor grooves
Hydrophobic
Hydrophilic
Complementarity

Base Pairing Results from H-Bonds
Only A=T and GC yield 20 Å Diameter

A:C base pair incompatibility

Bases Are Flat

Chains Are Antiparallel…

…Because of Base Pair Torsional Bond Angles

Base Pairs and Groove Formation

Base flipping can occur

Helix Is Right-Handed


Biologically Significant Form = B-DNA
Low Salt = Hydrated, 10.5 bp/turn

A- DNA Exists Under High Salt Conditions
Side-view Top-view
Base pairs tilted, 23 Å, 11bp/turn

Z-DNA Is a Left-Handed Helix
Zig-zag conformation, 18 Å, 12 bp/turn, no major groove


Propeller Twist Results from Bond Rotation

Sugar Conformations
Ideal B-DNA is C2'-endo (South)
Ideal A-RNA is C3'-endo (North)

anti and syn conformational ranges for glycosydic bonds in pyrimidine (left) and
purine (right) nucleosides
Source: Blackburn and Gait, Nucleic acids in chemistry and biology, Oxford University Press New York 1996.

Syn vs. Anti Conformations
Syn conformation causes left-handed helix

Syn-Anti Bond Rotation


Reassociation Kinetics

Denaturation of DNA Strands and the Hyperchromic Shift
• Denaturation (melting) is the breaking of H, but not covalent, bonds in DNA double helix duplex unwinds strands separate
• Viscosity decreases and bouyant density increases• Hyperchromic shift – uv absorption increases with
denaturation of duplex• Basis for melting curves because G-C pairs have three
H bonds but A-T pairs have only two H bonds• Duplexes with high G-C content have a higher melting
temperature because G-C pairs require a higher temperature for denaturation


Molecular Hybridization
• Reassociation of denatured strands• Occurs because of complementary base pairing • Can form RNA-DNA Hybrids• Can detect sequence homology between species• Basis for in situ hybridization, Southern and
Northern blotting, and PCR

Hybridization

Reassociation Kinetics• Derive information about the complexity of
a genome• To study reassociation, genome must first
be fragmented (e.g. by shear forces)• Next, DNA is heat-denatured• Finally, temperature is slowly lowered and
rate of strand reassociation (hybridization) is monitored

Data Analysis
• Pieces of DNA collide randomly and hybridize if complementary
• Plot the % reassociation versus the log of the product of the concentration of single-stranded (ss) DNA and time
• Reassociation follows second order kinetics: C/C0 = 1/1 + k C0t
• Initially, C = C0 that is [ssDNA] = 100%
• As time elapses, C approaches a [ssDNA] of 0%

• Initially there is a mixture of unique DNA sequence fragments so hybridization occurs slowly. As this pool shrinks, hybridization occurs more quickly
• C0t1/2 = half-reaction time or the point where one half of the DNA is present as ds fragments and half is present as ss fragments
• If all pairs of ssDNA hybrids contain unique sequences and all are about the same size, C0t1/2 is directly proportional to the complexity of the DNA
• Complexity = X represents the length in nucleotide pairs of all unique DNA fragments laid end to end
• Assuming that the DNA represents the entire genome and all sequences are different from each other, then X = the size of the haploid genome

The Tm

The Hyperchromic Shift (Melting Curve Profile)
Tm = temperature at which 50% of DNA is denatured
Maximum denaturation = 100% single stranded
Double stranded
50% double, 50% single stranded

High G-C Content Results in a Genome of Greater Bouyant Density

Ideal C0t Curve
100% ssDNA
100% dsDNA

Larger genomes take longer to reassociate because there are more DNA
fragments to hybridize
Largest genomeSmallest genome

C0t1/2 Is Directly Proportional to Genome Size

Genomes are composed of unique, moderately repetitive and highly repetitive
sequences
Highly repetitive DNA
Moderately repetitive DNA
10-4 10-2 100 102 104
Fra
ctio
n r
emai
nin
gsi
ngl
e-st
ran
ded
(C
/C0)
Unique DNA sequences
0
100
C0t (moles x sec/L)

More complex genomes contain more classes of DNA sequences

G-C Content Increases Tm

DNA Topology
Some of the following slides and text are taken from the DNA Topology lecture from Doug Brutlag’s January 7, 2000 Biochemistry 201 Advanced Molecular Biology Course at Stanford University

What Is Supercoiling & Why Should I Care?
• DNA forms supercoils in vivo • Important during replication and transcription• Topology only defined for a continuous strand - no strand
breakage• Numerical expression for degree of supercoiling:
Lk = Tw + Wr • L:linking number, # of times that one DNA strand winds
about the others strands - is always an integer • T: twist, # of revolutions about the duplex helix • W: writhe, # of turns of the duplex axis about the
superhelical axis is by definition the measure of the degree of supercoiling

• Supercoiling or writhing of circular DNA is a result of the DNA being underwound with respect to the relaxed form of DNA
• There are actually fewer turns in the DNA helix than would be expected given the natural pitch of DNA in solution (10.4 base pairs per turn)
• When a linear DNA is free in solution it assumes a pitch which contains 10.4 base pairs per turn
• This is less tightly wound than the 10.0 base pairs per turn in the Watson and Crick B-form DNA
DNA Topology

• DNA that is underwound is referred to as negatively supercoiled – The helices wind about each other in a right-
handed path in space• DNA that is overwound will relax and
become a positively supercoiled DNA helix– Positively coiled DNA has its DNA helices
wound around each other in a left-handed path in space

DNA topology

• Linking number - # times would have to pass cccDNA strand through the other to entirely separate the strands and not break any covalent bonds
• Twist - # times one strand completely wraps (# helical turns) around the other strand
• Writhe – when long axis of double helix crosses over itself (causes torsional stress)

Linking Defined
• Linking number, Lk, is the total number of times one strand of the DNA helix is linked with the other in a covalently closed circular molecule

1. The linking number is only defined for covalently closed DNA and its value is fixed as long as the molecule remains covalently closed.
2. The linking number does not change whether the covalently closed circle is forced to lie in a plane in a stressed conformation or whether it is allowed to supercoil about itself freely in space.
3. The linking number of a circular DNA can only be changed by breaking a phosphodiester bond in one of the two strands, allowing the intact strand to pass through the broken strand and then rejoining the broken strand.
4. Lk is always an integer since two strands must always be wound about each other an integral number of times upon closure.

Linking Number, Twists and Writhe

DNA tied up in knots• Metabolic events involving
unwinding impose great stress on the DNA because of the constraints inherent in the double helix
• There is an absolute requirement for the correct topological tension in the DNA (super-helical density) in order for genes to be regulated and expressed normally– For example, DNA must be
unwound for replication and transcription
Figure from Rasika Harshey’s lab at UT Austin showing an enhancer protein (red) bound to the DNA in a specific interwrapped topology that is called a transposition synapse. www.icmb.utexas.edu/.../47_Topology_summary.jpg

Knots, Twists, Writhe and Supercoiling
• Circular DNA chromosomes, from viruses for instance, exist in a highly compact or folded conformation

Twist
• The linking number of a covalently closed circular DNA can be resolved into two components called the twists, Tw and the writhes, Wr.
• Lk = Tw + Wr• The twists are the number of
times that the two strands are twisted about each other
• The length and pitch of DNA in solution determine the twist. [Tw = Length (bp)/Pitch (bp/turn)]

Writhe
• Writhe is the number of times that the DNA helix is coiled about itself in three-dimensional space
• The twist and the linking number, determine the value of the writhe that forces the DNA to assume a contorted path is space. [Wr = Lk - Tw ]

• Unlike the Twist and the Linking number, the writhe of DNA only depends on the path the helix axis takes in space, not on the fact that the DNA has two strands
• If the path of the DNA is in a plane, the Wr is always zero
• If the path of the DNA helix were on the surface of a sphere (like the seams of a tennis ball or base ball) then the total Writhe can also be shown to be zero

• Molecules that differ by one unit in linking number can be separated by electrophoresis in agarose due to the difference in their writhe (that is due to difference in folding).
• The variation in linking number is reflected in a difference in the writhe.
• The variation in writhe is subsequently reflected in the state of compaction of the DNA molecule.

Writhe of supercoiled DNA
Interwound Toroidal

Types of Supercoils

Supercoiling

Negative vs. Positive Supercoiling• Right handed supercoiling =
negative supercoiling (underwinding)
• Left handed supercoiling = positive supercoiling
• Relaxed state is with no bends
• DNA must be constrained: plasmid DNA or by proteins
• Unraveling the DNA at one position changes the superhelicity

Relaxed
Supertwisted

Unwinding DNA

Toposomerase

Topoisomerase II makes ds breaks

Topoisomerase I makes ss breaks







Ability of Uracil To Form Stable Base Pairs Enhances RNA’s Ability To Form Stem-
loop Structures

Intercalating Agents: Ethidium Bromide• By electrophoresing supercoiled
DNA in the presence of an intercalating agent such as ethidium bromide, one can distinguish negatively supercoiled DNA from positively supercoiled DNA
• When negatively supercoiled DNA binds an intercalating agent, the average pitch is reduced because the twist angle between adjacent base pairs on either side of the intercalating agent is reduced
• Reduction of twist causes a compensatory increase in writhe in a covalently closed molecule. Thus, a molecule that is initially negatively supercoiled will become more relaxed and a positively supercoiled molecule will become more twisted.

Histone Variants• Alter nucleosome function
– H2A.z often found in areas with transcribed regions of DNA
– prevents nucleosome from forming repressive structures that would inhibit access of RNA polymerase
• Mark areas of chromatin with alternate functions– CENP-A replaces H3– Associated with nucleosomes
that contain centromeric DNA– Has longer N-terminal tail
that may function to increase binding sites available for kinetochore protein binding

Unwrapping of DNA from nucleosome allows DNA-binding proteins access to their binding
sites• Many DNA-binding proteins
require histone-free DNA• DNA-histone interactions
dynamic: unwrapping is spontaneous and intermittent
• Accessibility to binding protein sites dependent on location in nucleosomal DNA– more central sites less accessible
than those near the ends decreasing probability of protein binding and hence regulating transcriptional activity
more central
more peripheral

Nucleosome remodeling complexes• Alter stability of DNA-
histone interaction to increase accessibility of DNA
• Change nucleosome location
• Require ATP• 3 mechanisms:1. Slide histone octamer
along DNA2. Transfer histone octamer
to another DNA3. Remodel to increase
access to DNA

DNA-binding protein dependent nucleosome positioning
• Nucleosomes are sometimes specifically positioned
• Keeps DNA-binding protein site in linker region (hence accessible)
• Can be directed by DNA-binding proteins or by specific sequences
• Usually involves competition between nucleosomes and binding proteins
• If proteins are positioned such that less than 147 bp exists between them, nucleosomes cannot associate

Positioning can be inhibitory• Some proteins can bind to
DNA and a nucleosome• By putting a tightly
bound binding protein next to a nucleosome, additional nucleosomes will assemble immediately adjacent to the protein preferentially

DNA sequences can direct positioning
• DNA sequences that position nucleosomes are A-T or G-C rich because DNA is bent in nucleosomes
• By alternating A-T or G-C rich sequences, can change the position in which the minor groove faces the histone octamer
• These sequences are rare

• Majority of nucleosomes are not positioned• Tightly positioned nucleosomes are usually
associated with areas for transcription initiation
• Positioned nucleosomes can prevent or enhance access to DNA sequences needed for binding protein attachment

Modification of N-terminal tails
• Results in increased or decreased affinity of nucleosome for DNA
• Modifications include acetylation, methylation and phosphorylation
• Combination of modifications may encode information for gene expression (positively or negatively

• Acetylated nucleosomes are associated with actively transcribed areas because reduces the affinity of the nucleosome for DNA
• Deacetylation associated with inactive transcription units
• Phosphorylation also increases transcription• Like acetylation, phosphorylation reduces
positive charge on histone proteins• Methylation represses transcription• Also affects ability of nucleosome array to
form higher order structures


HAT
Acetylation creates binding sites for bromo- and chromodomain protein binding

Chromatin remodeling complexes and histone modifying enzymes work together to make
DNA more accessible

Distributive inheritance of old histones
• Old histones have to be inherited to maintain histone modifications and appropriate gene expression
• H3▪H4 tetramers are randomly transferred to new daughter strand, never put into soluble pool
• H2A▪H2B dimers are put into pool and compete for association with H3▪H4 tetramers


Histone assembly requires chaperones
• Assembly of nucleosome is not spontaneous
• Chaperone proteins are needed to bring in free dimers and tetramers after replication fork has been passed
• Chaperones are associated with PCNA, the sliding clamp protein of eukaryotic replication, immediately after PCNA is released by DNA polymerase

Nucleotides and primer:template junction are essential substrates for DNA
synthesis