Embed Size (px)
Citation preview

DNA Replication, Mutation, Repair
M 211A MOLECULAR GENETICS
1ST SEMESTER, 2009
NABIL BASHIR

DNA Replication, Mutation, Repair
a). DNA replicationi). Cell cycle/ semi-conservative replicationii). Initiation of DNA replicationiii). Discontinuous DNA synthesisiv). Components of the replication apparatus
b). Mutationi). Types and rates of mutationii). Spontaneous mutations in DNA replicationiii). Lesions caused by mutagens
c). DNA repairi). Types of lesions that require repairii). Mechanisms of repair
Proofreading by DNA polymeraseMismatch repairExcision repair
iii). Defects in DNA repair or replication

Cell Cycle
• The S (for synthesis) phase of the cell cycle is when chromosomes are replicated. This requires DNA synthesis and histone synthesis (the latter to make the proteins that will package the newly replicated DNA).

The mammalian cell cycle
G1
S
G2M
G0
DNA synthesis and histone synthesis
Growth and preparation forcell division
Rapid growth and preparation forDNA synthesis
Quiescent cells
phase
phase
phase
phase
Mitosis

Semi-conservative DNA Replication
• DNA replication is said to be "semi-conservative" because each strand of the DNA double helix serves as a template for the synthesis of a new complementary DNA strand. Hence, the newly replicated chromosome consists of one old and one new DNA strand.

DNA replication is semi-conservative
Parental DNA strands
Daughter DNA strands
Each of the parental strands serves as a template for a daughter strand

Origins Of DNA Replication On Mammalian Chromosomes
• Each chromosome consists of a very long DNA strand, the complete length of which needs to be replicated in a relatively short (a few hours) period of time. This is accomplished by having multiple origins of DNA replication on each chromsome, which are spaced every ~150 kb. Replication initiates independently at each origin and proceeds bidirectionally as the new DNA strands (red) are synthesized. The resulting "replication bubbles" then fuse together or merge completing the synthesis of the daughter chromosomes.

origins of DNA replication (every ~150 kb)
replication bubble
daughter chromosomes
fusion of bubbles
bidirectional replication
Origins of DNA replication on mammalian chromosomes
5’3’
3’5’
5’3’
3’5’
3’5’
5’3’

Initiation Of DNA Synthesis At The E. Coli Origin (Ori)
• The single, circular E. coli chromosome has one origin of replication (ori). Initiation of replication begins with the binding of dnaA proteins to the ori sequence. As these proteins coalesce, the adjacent DNA is forced to undergo melting into single strands. This then allows the dnaB and dnaC proteins to bind the single-stranded DNA and further unwind the double helix, catalyzed by the dnaB protein which is a DNA "helicase" or DNA unwinding enzyme.

Initiation of DNA synthesis at the E. coli origin (ori)
5’3’
3’5’
origin DNA sequence
binding of dnaA proteins
A A A
dnaA proteins coalesce
DNA melting inducedby the dnaA proteinsA
AA
AA
A
AA
AA
A
A B C
dnaB and dnaC proteins bind to the single-stranded DNA
dnaB further unwinds the helix

• As further unwinding occurs, which displaces the dnaA proteins, the dnaG protein binds. This protein is a "primase" and synthesizes a short RNA primer of about 5 nucleotides. Because the primase synthesizes an RNA strand, it is an RNA polymerase. The primer provides a free 3' OH end to initiate DNA synthesis. As a rule, DNA polymerases cannot initiate DNA synthesis de novo, but can only add onto an existing 3' OH. Hence, the need for the RNA primer. In contrast to DNA polymerases, RNA polymerases can initiatate synthesis de novo.

A
A
A
AA
A B C
dnaB further unwinds the helix and displaces dnaA proteins
GdnaG (primase) binds...
A
A
A
AA
AB C
G...and synthesizes an RNA primer
RNA primer

Primasome
• The "primasome" consists of the dnaB, dnaC, and dnaG proteins.

B C
G
5’ 3’template strand
RNA primer(~5 nucleotides)
Primasome dna B (helicase) dna C dna G (primase)
OH3’ 5’

• Once the RNA primer has been synthesized, DNA polymerase can then bind and begin to synthesize DNA. DNA polymerase catalyzes an attack by the 3' OH on the alpha phosphate of the dGTP, forming a 3', 5'-phosphodiester bond, and releasing pyrophosphate, which is then hydrolyzed to two molecules of inorganic phosphate. All DNA polymerases require a primer (or a growing DNA chain) with a free 3' OH. The new strand of DNA grows in a 5' to 3' direction; however, some DNA polymerases also have a 3' to 5' proofreading activity, which can remove the 3' terminal nucleotide in case the polymerase makes a mistake.

3’
5’ 3’
RNA primer
newly synthesized DNA
5’
5’
DNA polymerase

Discontinuous Synthesis Of DNA
• Because DNA synthesis always proceeds in a 5' to 3' direction, and because the two DNA strands are arranged antiparallel with respect to each other, only one of the two newly synthesized strands can be made "continuously" (continuous red line) as the DNA polymerase moves away from the origin of replication and more DNA template is exposed. The other strand has to be made "discontinuously" in short pieces (short red lines). This latter strand is called the "lagging strand" while the continuously synthesized strand is called the "leading strand."

Discontinuous synthesis of DNA
3’5’
5’ 3’
3’ 5’
Because DNA is always synthesized in a 5’ to 3’ direction,synthesis of one of the strands...
5’3’ ...has to be discontinuous.
This is the lagging strand.
5’3’
3’5’
5’3’

Leading And A Lagging Strand
• The small pieces of DNA that comprise the lagging strand are called "Okazaki fragments." They are eventually ligated together forming a continuous DNA strand.

3’5’
5’ 3’
3’ 5’
5’3’
3’5’
5’3’
leading strand (synthesized continuously)
lagging strand (synthesized discontinuously)
Each replication fork has a leading and a lagging strand
• The leading and lagging strand arrows show the direction of DNA chain elongation in a 5’ to 3’ direction• The small DNA pieces on the lagging strand are called
Okazaki fragments (100-1000 bases in length)
replication fork replication fork

• The next series of figures shows the process of DNA synthesis at the so-called "replication fork." Note the direction (arrows) of leading strand synthesis and lagging strand synthesis (both are in a 5' to 3' direction). As stated previously, DNA polymerase moves continuously along the leading strand. As the template DNA unwinds, exposing the single-stranded template for the lagging strand, primase has to synthesize an RNA primer to which the DNA polymerase synthesizing the lagging strand can bind.

RNA primer
5’3’
3’5’
3’5’
direction of leading strand synthesis
direction of lagging strand synthesis
replication fork

Strand Separation At The Replication Fork Causes Positive
Supercoiling Of The Downstream Double Helix
• The strand separation process (unwinding the complementary Watson-Crick DNA strands) causes overwinding ahead of the fork. Any DNA that is overwound (or underwound) is said to be “supercoiled.” Overwound DNA is positively supercoiled. The increasing torsional stress needs to be dissipated in order for the fork to continue to unwind so that replication can proceed. This is accomplished by DNA topoisomerases, which cut the DNA strands, unwind them and reseal the strands. As they do so they introduce negative supercoiling into the DNA to compensate for the positive supercoiling. Gram-negative bacteria, such as E. coli, Klebsiella pneumoniae, and Pseudomonas aeruginosa, can be killed by fluoroquinolone antibiotics, which inhibit DNA gyrase, a topoisomerase II. Topoisomerase II cuts both strands of DNA, swivels them and rejoins them. Topoisomerase I cuts only one strand, and relaxes negative supercoils.

5’3’
3’5’
3’5’
Strand separation at the replication fork causes positivesupercoiling of the downstream double helix
• DNA gyrase is a topoisomerase II, which breaks and reseals the DNA to introduce negative supercoils ahead of the fork• Fluoroquinolone antibiotics target DNA gyrases in many gram-negative bacteria: ciprofloxacin and levofloxacin (Levaquin)

Movement Of The Replication Fork
• As the replication fork moves further to the left, opening up more DNA, another RNA primer has to be synthesized.

5’3’ 5’
3’
Movement of the replication fork

Movement Of The Replication Fork
• Each RNA primer (dashed red line) on the lagging strand then serves as a starting point for the initiation of DNA synthesis (Okazaki fragment; solid red line).

Movement of the replication fork
RNA primerOkazaki fragment
RNA primer
5’

• E. coli DNA polymerase III initiates at the RNA primer, synthsizing DNA to fill in the gap up to the next RNA primer. However, it falls off the template DNA once it reaches the next RNA primer. At this point, DNA polymerase I takes over. It contains a 5' to 3' exonuclease activity that can remove the RNA primer while it simultaneously adds DNA nucleotides to the 3' end of the Okazaki fragment.

3’
RNA primer5’
DNA polymerase III initiates at the primer andelongates DNA up to the next RNA primer
5’
5’3’
5’
newly synthesized DNA (100-1000 bases) (Okazaki fragment)
5’3’
DNA polymerase I inititates at the end of the Okazaki fragment and further elongates the DNA chain while simultaneously removing the RNA primer with its 5’ to 3’ exonuclease activity
pol III
pol I

• However, DNA polymerase I cannot seal the gap between the two adjacent Okazaki fragments. This job is carried out by DNA ligase, which catalyzes the formation of a 3', 5'-phosphodiester bond in an ATP-dependent reaction. Thus, it takes one RNA polymerase (primase), two DNA polymerases (III and I), and DNA ligase to synthesize the lagging strand. Once initiation occurs on the RNA primer, the leading strand only requires DNA polymerase III for its synthesis.

newly synthesized DNA (Okazaki fragment)5’
3’
5’3’
DNA ligase seals the gap by catalyzing the formationof a 3’, 5’-phosphodiester bond in an ATP-dependent reaction

Proteins At The Replication Fork In E. Coli
• This figure shows the proteins required for DNA synthesis at the replication fork. In addition to the proteins required for leading and lagging strand synthesis, there are several others that act upstream of the replication fork. The act of unwinding the DNA double helix puts torsional stress in the form of positive supercoils in the DNA upstream of the fork. To overcome this, DNA gyrase, which is a topoisomerase II, breaks and reseals the DNA in order to introduce negative supercoils in the DNA, thus overcoming the positive supercoils. The unwinding itself is carried out by the Rep protein, which is a helicase. Finally, to keep the unwound strands single-stranded, they bind SSB (single-strand binding protein).

5’3’
3’5’
Proteins at the replication fork in E. coli
Rep protein (helicase)
Single-strandbinding protein (SSB)
BC
G Primasome
pol I
pol III
pol III
DNA ligase
DNA gyrase - this is a topoisomerase II, whichbreaks and reseals double-stranded DNA to introducenegative supercoils ahead of the fork

Components Of The Replication Apparatus
• This table lists the proteins required for replication of the E. coli chromosome, and their activities.

Components of the replication apparatus
dnaA binds to origin DNA sequencePrimasome dnaB helicase (unwinds DNA at origin) dnaC binds dnaB dnaG primase (synthesizes RNA primer)DNA gyrase introduces negative supercoils ahead
of the replication forkRep protein helicase (unwinds DNA at fork)SSB binds to single-stranded DNADNA pol III primary replicating polymeraseDNA pol I removes primer and fills gapDNA ligase seals gap by forming 3’, 5’-phosphodiester bond

Properties Of DNA Polymerases
• The main DNA polymerases required for DNA replication in E. coli are DNA polymerases I and III. They both have 5' to 3' polymerizing activity and 3' to 5' proofreading activity. DNA polymerase I also removes the RNA primer and is also a DNA repair enzyme, and thus requires a 5' to 3' exonuclease activity.
• In contrast to just three DNA polymerases in E. coli, human cells have at least five DNA polymerases. The enzymes thought to be responsible for replication of nuclear DNA are DNA polymerases alpha, delta, and epsilon. Alpha is associated with an RNA primase and it is thought that these activities are responsible for synthesizing a short RNA-DNA primer. There is some uncertainty as to the function of alpha in lagging strand synthesis. Since it seems to lack a 3’ to 5’ exonuclease activity, it may not be able to synthesize DNA with high fidelity and thus it may not be the main lagging strand polymerase. If not, the main polymerase for the lagging strand may be delta, which is also specifically responsible for synthesis of the leading strand. DNA polymerase epsilon may function like the bacterial DNA polymerase I, by removing primers and extending the Okazaki fragment. In humans, there is also a requirement for DNA ligase.
• Proliferating cell nuclear antigen (PCNA) has a role in both replication and repair. It is a toroidal-shaped (donut) protein which encircles DNA and can slide bidirectionally along the duplex. One of its functions is to serve as a processivity factor for DNA polymerase delta and epsilon. PCNA holds the polymerase to the DNA template for rapid and processive DNA synthesis. Recently, it has been discovered that PCNA also interacts with proteins involved in cell-cycle progression.

Properties of DNA polymerases
DNA polymerases of E. coli_
pol I pol II pol III (core)Polymerization: 5’ to 3’ yes yes yesProofreading exonuclease: 3’ to 5’ yes yes yesRepair exonuclease: 5’ to 3’ yes no no
DNA polymerase III is the main replicating enzymeDNA polymerase I has a role in replication to fill gaps and excise primers on the lagging strand, and it is also a repair enzyme and is used in making recombinant DNA molecules
• all DNA polymerases require a primer with a free 3’ OH group• all DNA polymerases catalyze chain growth in a 5’ to 3’ direction• some DNA polymerases have a 3’ to 5’ proofreading activity

Mutation
• This slide shows the three basic types of mutational events and their frequencies. We will be concentrating on "gene mutations," which are base pair mutations or small deletions or insertions.

Types and rates of mutation
Type Mechanism Frequency________ Genome chromosome 10-2 per cell division mutation missegregation
(e.g., aneuploidy)
Chromosome chromosome 6 X 10-4 per cell division mutation rearrangement
(e.g., translocation)
Gene base pair mutation 10-10 per base pair per mutation (e.g., point mutation, cell division or
or small deletion or 10-5 - 10-6 per locus per insertion generation
Mutation

Mutation rates* of selected genes
Gene New mutations per 106 gametes
Achondroplasia 6 to 40Aniridia 2.5 to 5Duchenne muscular dystrophy 43 to 105Hemophilia A 32 to 57Hemophilia B 2 to 3Neurofibromatosis -1 44 to 100Polycystic kidney disease 60 to 120Retinoblastoma 5 to 12
*mutation rates (mutations / locus / generation) can varyfrom 10-4 to 10-7 depending on gene size and whetherthere are “hot spots” for mutation (the frequency at mostloci is 10-5 to 10-6).

Many Polymorphisms Exist In The Genome
• What is the frequency of new germline mutations? Consider the following: each sperm contains ~100 new mutations; a normal ejaculate has ~100 million sperm; 100 X 100 million = 10 billion new mutations; ~1 in 10 sperm carries a new deleterious mutation; at a rate of production of ~80 million sperm per day, a male will produce a sperm with a new mutation in the Duchenne muscular dystrophy gene approximately every 10 seconds.

Many polymorphisms exist in the genome
• the number of existing polymorphisms is ~1 per 500 bp• there are ~5.8 million differences per haploid genome• polymorphisms were caused by mutations over time• polymorphisms called single nucleotide polymorphisms
(or SNPs) are being catalogued by the HumanGenome Project as an ongoing project

Types Of Base Pair Mutations
• This slide illustrates the four basic types of base pair mutations. Two of them result in the conversion of one base pair to another (base pair substitution). The others result in removal (deletion) or addition (insertion) of one or more base pairs. (Note that a transition mutation results when a pyrimidine on one strand is converted to another pyrimidine on the same strand. The complementary strand would see a conversion from one purine to the other purine.)

Types of base pair mutations
CATTCACCTGTACCAGTAAGTGGACATGGT
CATGCACCTGTACCAGTACGTGGACATGGT
CATCCACCTGTACCAGTAGGTGGACATGGT
transition (T-A to C-G) transversion (T-A to G-C)
CATCACCTGTACCAGTAGTGGACATGGT
deletionCATGTCACCTGTACCAGTACAGTGGACATGGT
insertion
base pair substitutions transition: pyrimidine to pyrimidine transversion: pyrimidine to purine
normal sequence
deletions and insertions can involve one or more base pairs

Spontaneous Mutations Can Be Caused By Tautomers
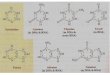
• There are many different causes of mutations. Spontaneous mutations (those that result from no external cause) can occur simply by rearrangement of bonds and by the repositioning of hydrogens in the purine and pyrimidine bases. The common forms of adenine and cytosine are the amino forms, which can rearrange to the imino forms. The repositioning of hydrogens changes their base pairing, hydrogen bonding chemistry.

Spontaneous mutations can be caused by tautomers
Tautomeric forms of the DNA bases
Adenine
Cytosine
AMINO IMINO

Tautomeric Forms Of The DNA Bases
• The common forms of guanine and thymine are the keto forms, which can rearrange to the enol forms.

Guanine
Thymine
KETO ENOL
Tautomeric forms of the DNA bases

Mutation Caused By Tautomer Of Cytosine
• This figure illustrates how a conversion from the amino to the imino form of cytosine changes the locations of hydrogen donor and acceptor groups, such that the imino form of cytosine base pairs with adenine instead of guanine. As shown in the next figure, this will ultimately lead to a transition mutation. As for the other tautomers, the imino form of adenine "looks sort of like" a guanine and should be able to form two hydrogen bonds with cytosine; the enol form of guanine looks like an adenine and should be able to form three hydrogen bonds with thymine; and the enol form of thymine looks like a cytosine and should be able to form three hydrogen bonds with guanine. In all cases, the wrong nucleotide will be inserted into the growing DNA chain.

Mutation caused by tautomer of cytosine
Cytosine
Cytosine
Guanine
Adenine
• cytosine mispairs with adenine resulting in a transition mutation
Normal tautomeric form
Rare imino tautomeric form

Mutation Is Perpetuated By Replication
• The conversion of a C-G base pair to a T-A base pair takes two steps. If the tautomeric form of cytosine is present during DNA replication, an adenosine will be inserted into the daughter DNA strand (instead of the normal guanosine). During the next round of DNA replication, the adenosine then serves as a template for the insertion of a thymidine in the new DNA strand, resulting in a transition mutation (the conversion of a C-G to a T-A).

Mutation is perpetuated by replication
• replication of C-G should give daughter strands each with C-G
• tautomer formation C during replication will result in mispairing and insertion of an improper A in one of the daughter strands
• which could result in a C-G to T-A transition mutation in the next round of replication, or if improperly repaired
C G C G
C G C A
AC T A

Chemical Mutagens
• Mutation can also occur by the action of chemical mutagens. This figure shows how oxidative deamination of cytosine converts it to uracil, and how the oxidative deamination of adenine converts it to hypoxanthine, both processes altering the hydrogen bonding specificities of these bases.

Chemical mutagens
Deamination by nitrous acid

Attack By Oxygen Free RadicalsLeading To Oxidative Damage
• The deoxyribose ring is also susceptible to damage, which can be cleaved by oxygen free radicals and break the phosphodiester backbone of DNA.

N
NH
NH
N
NH2
O
N
NH
NH
NH
NH2
O
O
Attack by oxygen free radicalsleading to oxidative damage
guanine
8-oxyguanine (8-oxyG)
• many different oxidative modifications occur• by smoking, etc.• 8-oxyG causes G to T transversions
• the MTH1 protein degrades 8-oxy-dGTP preventing misincorporation• mutation of the MTH1 gene causes increased tumor formation in mice

Ames test for mutagen detection
• named for Bruce Ames• reversion of histidine mutations by test compounds• His- Salmonella typhimurium cannot grow without histidine• if test compound is mutagenic, reversion to His+ may occur• reversion is correlated with carcinogenicity

Thymine Dimer Formation By UV Light
• Sunlight is particularly damaging to DNA. This figure shows the formation of a thymine dimer, catalyzed by UV light. The thymine dimer bridges two adjacent thymine residues on the same DNA strand

Thymine dimer formation by UV light

DNA Lesions
• As shown here, DNA is prone to many different kinds of damaging reactions, any of which can alter DNA function and cause mutations if not repaired.

Summary of DNA lesions
Missing base Acid and heat depurination (~104 purinesper day per cell in humans)
Altered base Ionizing radiation; alkylating agents
Incorrect base Spontaneous deaminationscytosine to uraciladenine to hypoxanthine
Deletion-insertion Intercalating reagents (acridines)
Dimer formation UV irradiation
Strand breaks Ionizing radiation; chemicals (bleomycin)
Interstrand cross-links Psoralen derivatives; mitomycin C
Tautomer formation Spontaneous and transient

Mechanisms of Repair
• Mutations that occur during DNA replication are repaired whenpossible by proofreading by the DNA polymerases
• Mutations that are not repaired by proofreading are repairedby mismatch (post-replication) repair followed byexcision repair
• Mutations that occur spontaneously any time are repaired byexcision repair (base excision or nucleotide excision)

Mismatch (Post-replication) Repair(Reduces DNA Replication Errors 1,000-fold)
• For DNA to be repaired properly following the misincorporation of a nucleotide into the newly synthesized DNA strand, the replication machinery must have a means by which to distinguish between the "old" (template) strand shown in black and the "new" (daughter) strand shown in red. After DNA replication takes place, the newly synthesized DNA is methylated on certain adenine bases. This, however, does not occur right away - there is a "window of time" in which the newly synthesized DNA (in red) is not methylated. Thus, if a mutation occurs in the new strand, the repair machinery can tell which is the template strand (presumable the correct strand) and the new strand (containing the mutation). It will then repair the mismatch by excision repair. Once some time has passed, the new strand will also become methylated; at that point it will not be possible to distinguish the correct nucleotide from the incorrect nucleotide at the site of a base pair mismatch. Defects in mismatch repair are a cause of hereditary nonpolyposis colon cancer (Thompson & Thompson, Case 13). There are six mismatch repair genes that can be affected in HNPCC.

Mismatch (post-replication) repair(reduces DNA replication errors 1,000-fold)
5’3’
CH3
CH3
CH3
CH3
• the parental DNA strands are methylated on certain adenine bases
• mutations on the newly replicated strand are identified by scanning for mismatches prior to methylation of the newly replicated DNA
• the mutations are repaired by excision repair mechanisms• after repair, the newly replicated strand is methylated

Excision Repair
• There are two types of excision repair: base excision repair (left) and nucleotide excision repair (right). While they differ in their initial steps (top), they are similar in the latter steps (bottom).
• If cytosine is deaminated forming uracil, the U can be recognized as being an improper base in DNA by the enzyme, uracil DNA glycosylase. This enzyme cleaves the uracil base from the phosphodiester backbone, and the space is opened up by repair nucleases that remove a number of nucleotides from one strand (the other strand has to be left intact to serve as the template for DNA repair). The repair polymerase, DNA polymerase beta, then fills in the gap and DNA ligase seals the last phosphodiester bond. The double strandedness of DNA makes possible both DNA replication and DNA repair, because the template strand always contains the information for the synthesis of a complementary strand.
• Nucleotide excision repair occurs when the DNA lesion is larger, for example when there is a thymine dimer. In this case, a special repair excinuclease removes about 30 nucleotides, including the lesion. The DNA is then resynthesized and ligated together as with base excision repair.

Excision repair
ATGCUGCATTGATAGTACGGCGTAACTATC
thymine dimer
AT AGTACGGCGTAACTATC
ATGCCGCATTGATAGTACGGCGTAACTATC
ATGCCGCATTGATAGTACGGCGTAACTATC
excinuclease
DNA polymerase
DNA ligase
(~30 nucleotides)
ATGCUGCATTGATACGGCGTAACT
ATGC GCATTGATACGGCGTAACT
AT GCATTGATACGGCGTAACT
deamination
ATGCCGCATTGATACGGCGTAACT
ATGCCGCATTGATACGGCGTAACT
uracil DNA glycosylase
repair nucleases
DNA polymerase
DNA ligase
Base excision repair Nucleotide excision repair

Deamination Of Cytosine Can Be Repaired
• Deaminated cytosine is fairly straightforward to repair because uracil is recognized as being "foreign" in the DNA molecule. However, many cytosines are reversibly methylated at CG sites (or CpG sites to emphasize that the C and G are adjacent nucleotides on the same DNA strand). It is believed that this methylation functions to regulate gene expression because 5-methylcytosine (5mC) residues are often clustered near the promoters of genes in so-called "CpG islands." The problem that arises from these methylations is that subsequent deamination of a 5mC results in the production of thymine, which is not foreign to DNA. Thus, while a base pair mismatch is seen in the DNA by the repair machinery, it does not know which of the two strands to repair (50% of the time it will make the right choice and 50% of the time it will make the wrong choice). As such, 5'-mCG-3' sites are "hot-spots" for mutation.

Deamination of cytosine can be repaired
More than 30% of all single base changes that have been detected as a cause of genetic disease have occurred at 5’-mCpG-3’ sites
Deamination of 5-methylcytosine cannot be repaired
cytosine uracil
thymine5’-methyl-cytosine

Correlation Between DNA RepairActivity And
The Life Span Of The Organism
• There is a direct correlation between DNA repair enzymatic activity and the life span of organisms, suggesting that DNA repair activity slows down cellular senescence and that cellular senescence is caused by mutations in DNA. Defects in DNA repair or replication can lead to a number of abnormalities. See Baynes & Dominiczak, pg. 601.

DNA repair activity
Life
spa
n
1
10
100 human
elephant
cow
hamsterratmouseshrew
Correlation between DNA repairactivity in fibroblast cells fromvarious mammalian species andthe life span of the organism

Defects In DNA Repair Or Replication
• As shown here, there are a number of defects of DNA replication and repair that are associated with a predisposition to cancer and other disorders. This highlights the importance of high fidelity DNA replication and for the presence of DNA repair mechanisms for normal cell function and longevity. See Baynes & Dominiczak, pg. 315.

Defects in DNA repair or replicationAll are associated with a high frequency of chromosome
and gene (base pair) mutations; most are also associated with a predisposition to cancer, particularly leukemias
• Xeroderma pigmentosum• caused by mutations in genes involved in nucleotide excision repair• associated with a >1000-fold increase of sunlight-induced skin cancer and with other types of cancer such as melanoma
• Ataxia telangiectasia• caused by gene that detects DNA damage• increased risk of X-ray• associated with increased breast cancer in carriers
• Fanconi anemia• caused by a gene involved in DNA repair• increased risk of X-ray and sensitivity to sunlight
• Bloom syndrome• caused by mutations in a a DNA helicase gene• increased risk of X-ray• sensitivity to sunlight
• Cockayne syndrome• caused by a defect in transcription-linked DNA repair• sensitivity to sunlight
• Werner’s syndrome• caused by mutations in a DNA helicase gene• premature aging