4844
© 2013. Published by The Company of Biologists Ltd | Development (2013) 140, 4844-4849 doi:10.1242/dev.103010
ABSTRACTTerminally differentiated post-mitotic cells are generally consideredirreversibly developmentally locked, i.e. incapable of beingreprogrammed in vivo into entirely different cell types. We found thatbrief expression of a single transcription factor, the ELT-7 GATAfactor, can convert the identity of fully differentiated, highly specializednon-endodermal cells of the pharynx into fully differentiated intestinalcells in intact larvae and adult Caenorhabditis elegans. Stableexpression of intestine-specific molecular markers parallels loss ofmarkers for the original differentiated pharynx state; hence, there isno apparent requirement for a dedifferentiated intermediate duringthe transdifferentiation process. Based on high-resolutionmorphological characteristics, the transdifferentiated cells becomeremodeled to resemble typical intestinal cells at the level of both thecell surface and internal organelles. Thus, post-mitotic cells, thoughterminally differentiated, remain plastic to transdifferentiation acrossgerm layer lineage boundaries and can be remodeled to adopt thecharacteristics of a new cell identity without removal of inhibitoryfactors. Our findings establish a simple model to investigate how cellcontext influences forced transdifferentiation of mature cells.
KEY WORDS: Transdifferentiation, Cellular reprogramming, C. elegans
INTRODUCTIONCell fate is progressively restricted during development such thatmost cells ultimately become post-mitotic, highly specialized andterminally differentiated, with a fixed morphology and pattern ofgene expression. Early embryonic cells in C. elegans are capable ofbeing reprogrammed into cell types of all three germ layers byappropriate transcription factors, but lose this ability during mid-embryogenesis, as they commit to particular pathways ofdifferentiation (Horner et al., 1998; Zhu et al., 1998; Gilleard andMcGhee, 2001; Quintin et al., 2001; Fukushige and Krause, 2005).Eliminating polycomb complex (Yuzyuk et al., 2009) or Notchsignaling (Djabrayan et al., 2012) components extends the period formultipotency during embryogenesis; however, post-mitotic somaticcells in late embryos, larvae or adults cannot be reprogrammed bythe transcription factors reported to date (Horner et al., 1998; Zhu etal., 1998; Gilleard and McGhee, 2001; Quintin et al., 2001;Fukushige and Krause, 2005). Developing and proliferating
RESEARCH REPORT STEM CELLS AND REGENERATION
1Department of Molecular, Cellular and Developmental Biology, and NeuroscienceResearch Institute, University of California, Santa Barbara, CA 93106, USA.2Center for C. elegans Anatomy, Albert Einstein College of Medicine, Bronx, NY 10461, USA. 3School of Biological Sciences, University of Auckland, Auckland 1010, New Zealand.‡Present address: Department of Biology, Massachusetts Institute of Technology,Cambridge, MA 02139, USA.
*Author for correspondence ([email protected])
Received 29 August 2013; Accepted 29 September 2013
germline cells can prematurely adopt somatic cell fates uponremoval of translational regulators (Ciosk et al., 2006) or inhibitorychromatin remodeling factors (Tursun et al., 2011; Patel et al.,2012). However, germline cells are uncommitted pluripotent cellsthat are poised to differentiate shortly after gametogenesis, whereasmost somatic cells become terminally differentiated.
We report here that a transcription factor that regulates terminalintestine differentiation violates the embryonic multipotency tocommitment transition (MCT) and can reprogram and remodeldifferentiated cells in C. elegans larvae and adults into intestine-likecells without removal of inhibitory factors. We found that a briefpulse of the ELT-7 GATA transcription factor activates intestine-specific gene expression in diverse non-intestinal cells. Post-mitoticpharyngeal cells maintain intestine gene expression, lose pharynxgene expression, and become remodeled to resemble intestinal cellsat the fine ultrastuctural level. Our results suggests that post-mitotic,terminally differentiated cells can be reprogrammed and remodeledwithout prior removal of the initial cell identity and demonstrate thatsusceptibility to in vivo reprogramming is influenced bytranscription factor identity and cellular context.
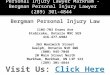
RESULTS AND DISCUSSIONELT-7 activates and maintains intestine-specific geneexpression in differentiated pharyngeal cells at all stages ofdevelopmentAlthough early embryonic cells in C. elegans are multipotent, cellsbecome refractory to reprogramming later in embryogenesis,following the MCT (Horner et al., 1998; Zhu et al., 1998; Gilleardand McGhee, 2001; Quintin et al., 2001; Fukushige and Krause,2005). In contrast to other regulators of cell fate specification thathave been reported, we found that ELT-7, a GATA transcriptionfactor that regulates terminal intestinal differentiation (Maduro andRothman, 2002; Sommermann et al., 2010), activates an intestinalmarker (elt-2::lacZ::GFP) in non-intestinal cells when brieflyectopically expressed via a heat-shock promoter at any embryonic,larval, or adult stage (Fig. 1A-C; supplementary material Fig. S1).We also detected IFB-2, an intermediate filament protein that is acomponent of the terminal web of the fully differentiated intestine(Bossinger et al., 2004), outside of the intestine in such animals(Fig. 1D; supplementary material Fig. S2). By contrast, the END-1GATA transcription factor, which specifies the endoderm progenitor(Zhu et al., 1997), does not activate widespread intestinal geneexpression after the MCT (Fig. 1; supplementary material Fig. S2).Thus, ELT-7 may violate the well-documented restriction todevelopmental reprogramming that occurs during embryogenesis.
In the endogenous intestine, ELT-7 and ELT-2 expression ismaintained throughout larval development and adulthood viacross- and auto-regulatory feedback mechanisms (Zhu et al., 1998;Fukushige et al., 1999; Sommermann et al., 2010). Althoughintestinal gene expression is never observed outside the intestinein non-heat-shocked hs-elt-7 larvae (n=128) or in heat-shocked
Transdifferentiation and remodeling of post-embryonic C. eleganscells by a single transcription factorMisty R. Riddle1, Abraham Weintraub1,‡, Ken C. Q. Nguyen2, David H. Hall2 and Joel H. Rothman1,3,*
Dev
elop
men
t
4845
RESEARCH REPORT Development (2013) doi:10.1242/dev.103010
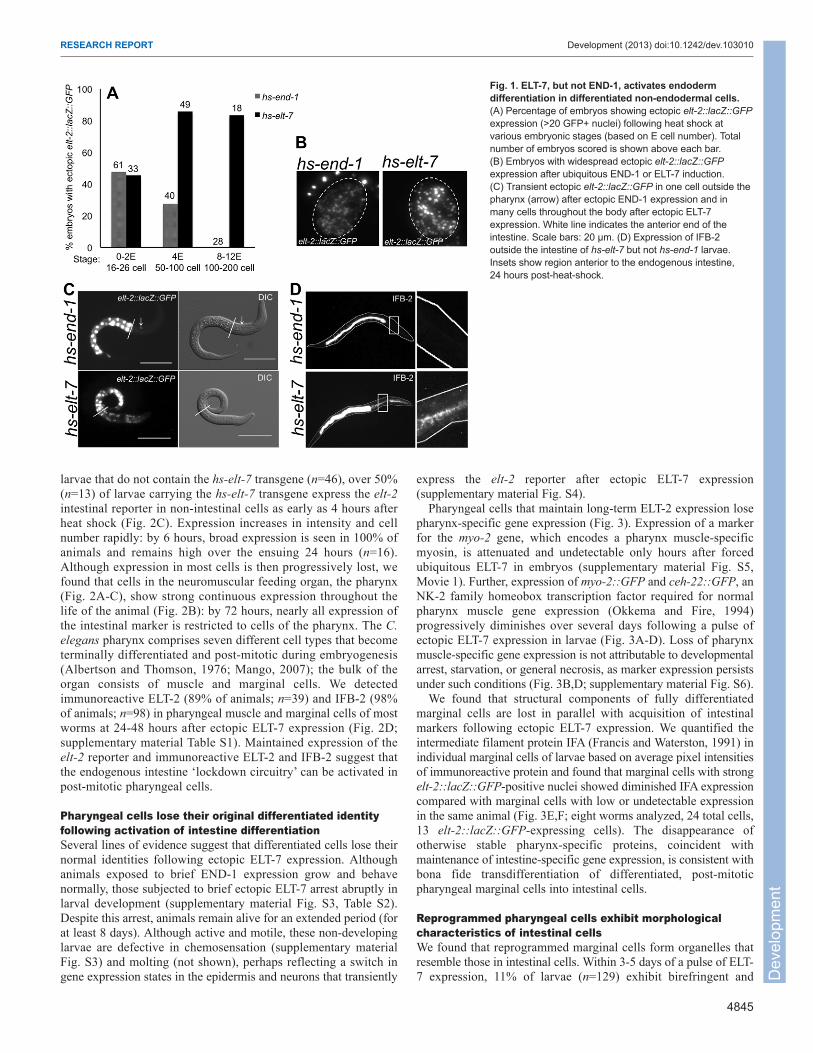
larvae that do not contain the hs-elt-7 transgene (n=46), over 50%(n=13) of larvae carrying the hs-elt-7 transgene express the elt-2intestinal reporter in non-intestinal cells as early as 4 hours afterheat shock (Fig. 2C). Expression increases in intensity and cellnumber rapidly: by 6 hours, broad expression is seen in 100% ofanimals and remains high over the ensuing 24 hours (n=16).Although expression in most cells is then progressively lost, wefound that cells in the neuromuscular feeding organ, the pharynx(Fig. 2A-C), show strong continuous expression throughout thelife of the animal (Fig. 2B): by 72 hours, nearly all expression ofthe intestinal marker is restricted to cells of the pharynx. The C.elegans pharynx comprises seven different cell types that becometerminally differentiated and post-mitotic during embryogenesis(Albertson and Thomson, 1976; Mango, 2007); the bulk of theorgan consists of muscle and marginal cells. We detectedimmunoreactive ELT-2 (89% of animals; n=39) and IFB-2 (98%of animals; n=98) in pharyngeal muscle and marginal cells of mostworms at 24-48 hours after ectopic ELT-7 expression (Fig. 2D;supplementary material Table S1). Maintained expression of theelt-2 reporter and immunoreactive ELT-2 and IFB-2 suggest thatthe endogenous intestine ‘lockdown circuitry’ can be activated inpost-mitotic pharyngeal cells.
Pharyngeal cells lose their original differentiated identityfollowing activation of intestine differentiationSeveral lines of evidence suggest that differentiated cells lose theirnormal identities following ectopic ELT-7 expression. Althoughanimals exposed to brief END-1 expression grow and behavenormally, those subjected to brief ectopic ELT-7 arrest abruptly inlarval development (supplementary material Fig. S3, Table S2).Despite this arrest, animals remain alive for an extended period (forat least 8 days). Although active and motile, these non-developinglarvae are defective in chemosensation (supplementary materialFig. S3) and molting (not shown), perhaps reflecting a switch ingene expression states in the epidermis and neurons that transiently
express the elt-2 reporter after ectopic ELT-7 expression(supplementary material Fig. S4).
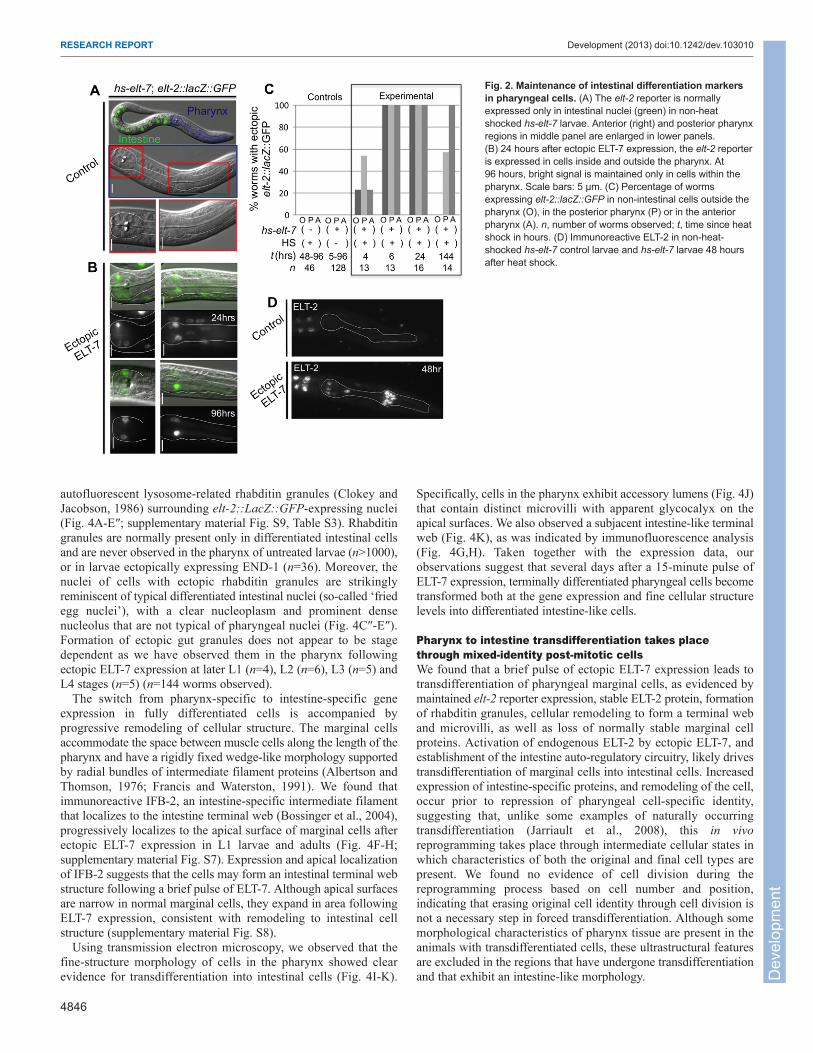
Pharyngeal cells that maintain long-term ELT-2 expression losepharynx-specific gene expression (Fig. 3). Expression of a markerfor the myo-2 gene, which encodes a pharynx muscle-specificmyosin, is attenuated and undetectable only hours after forcedubiquitous ELT-7 in embryos (supplementary material Fig. S5,Movie 1). Further, expression of myo-2::GFP and ceh-22::GFP, anNK-2 family homeobox transcription factor required for normalpharynx muscle gene expression (Okkema and Fire, 1994)progressively diminishes over several days following a pulse ofectopic ELT-7 expression in larvae (Fig. 3A-D). Loss of pharynxmuscle-specific gene expression is not attributable to developmentalarrest, starvation, or general necrosis, as marker expression persistsunder such conditions (Fig. 3B,D; supplementary material Fig. S6).
We found that structural components of fully differentiatedmarginal cells are lost in parallel with acquisition of intestinalmarkers following ectopic ELT-7 expression. We quantified theintermediate filament protein IFA (Francis and Waterston, 1991) inindividual marginal cells of larvae based on average pixel intensitiesof immunoreactive protein and found that marginal cells with strongelt-2::lacZ::GFP-positive nuclei showed diminished IFA expressioncompared with marginal cells with low or undetectable expressionin the same animal (Fig. 3E,F; eight worms analyzed, 24 total cells,13 elt-2::lacZ::GFP-expressing cells). The disappearance ofotherwise stable pharynx-specific proteins, coincident withmaintenance of intestine-specific gene expression, is consistent withbona fide transdifferentiation of differentiated, post-mitoticpharyngeal marginal cells into intestinal cells.
Reprogrammed pharyngeal cells exhibit morphologicalcharacteristics of intestinal cellsWe found that reprogrammed marginal cells form organelles thatresemble those in intestinal cells. Within 3-5 days of a pulse of ELT-7 expression, 11% of larvae (n=129) exhibit birefringent and
Fig. 1. ELT-7, but not END-1, activates endodermdifferentiation in differentiated non-endodermal cells.(A) Percentage of embryos showing ectopic elt-2::lacZ::GFPexpression (>20 GFP+ nuclei) following heat shock atvarious embryonic stages (based on E cell number). Totalnumber of embryos scored is shown above each bar.(B) Embryos with widespread ectopic elt-2::lacZ::GFPexpression after ubiquitous END-1 or ELT-7 induction.(C) Transient ectopic elt-2::lacZ::GFP in one cell outside thepharynx (arrow) after ectopic END-1 expression and inmany cells throughout the body after ectopic ELT-7expression. White line indicates the anterior end of theintestine. Scale bars: 20 μm. (D) Expression of IFB-2outside the intestine of hs-elt-7 but not hs-end-1 larvae.Insets show region anterior to the endogenous intestine,24 hours post-heat-shock.
Dev
elop
men
t
4846
RESEARCH REPORT Development (2013) doi:10.1242/dev.103010
autofluorescent lysosome-related rhabditin granules (Clokey andJacobson, 1986) surrounding elt-2::LacZ::GFP-expressing nuclei(Fig. 4A-E″; supplementary material Fig. S9, Table S3). Rhabditingranules are normally present only in differentiated intestinal cellsand are never observed in the pharynx of untreated larvae (n>1000),or in larvae ectopically expressing END-1 (n=36). Moreover, thenuclei of cells with ectopic rhabditin granules are strikinglyreminiscent of typical differentiated intestinal nuclei (so-called ‘friedegg nuclei’), with a clear nucleoplasm and prominent densenucleolus that are not typical of pharyngeal nuclei (Fig. 4C″-E″).Formation of ectopic gut granules does not appear to be stagedependent as we have observed them in the pharynx followingectopic ELT-7 expression at later L1 (n=4), L2 (n=6), L3 (n=5) andL4 stages (n=5) (n=144 worms observed).
The switch from pharynx-specific to intestine-specific geneexpression in fully differentiated cells is accompanied byprogressive remodeling of cellular structure. The marginal cellsaccommodate the space between muscle cells along the length of thepharynx and have a rigidly fixed wedge-like morphology supportedby radial bundles of intermediate filament proteins (Albertson andThomson, 1976; Francis and Waterston, 1991). We found thatimmunoreactive IFB-2, an intestine-specific intermediate filamentthat localizes to the intestine terminal web (Bossinger et al., 2004),progressively localizes to the apical surface of marginal cells afterectopic ELT-7 expression in L1 larvae and adults (Fig. 4F-H;supplementary material Fig. S7). Expression and apical localizationof IFB-2 suggests that the cells may form an intestinal terminal webstructure following a brief pulse of ELT-7. Although apical surfacesare narrow in normal marginal cells, they expand in area followingELT-7 expression, consistent with remodeling to intestinal cellstructure (supplementary material Fig. S8).
Using transmission electron microscopy, we observed that thefine-structure morphology of cells in the pharynx showed clearevidence for transdifferentiation into intestinal cells (Fig. 4I-K).
Specifically, cells in the pharynx exhibit accessory lumens (Fig. 4J)that contain distinct microvilli with apparent glycocalyx on theapical surfaces. We also observed a subjacent intestine-like terminalweb (Fig. 4K), as was indicated by immunofluorescence analysis(Fig. 4G,H). Taken together with the expression data, ourobservations suggest that several days after a 15-minute pulse ofELT-7 expression, terminally differentiated pharyngeal cells becometransformed both at the gene expression and fine cellular structurelevels into differentiated intestine-like cells.
Pharynx to intestine transdifferentiation takes placethrough mixed-identity post-mitotic cellsWe found that a brief pulse of ectopic ELT-7 expression leads totransdifferentiation of pharyngeal marginal cells, as evidenced bymaintained elt-2 reporter expression, stable ELT-2 protein, formationof rhabditin granules, cellular remodeling to form a terminal weband microvilli, as well as loss of normally stable marginal cellproteins. Activation of endogenous ELT-2 by ectopic ELT-7, andestablishment of the intestine auto-regulatory circuitry, likely drivestransdifferentiation of marginal cells into intestinal cells. Increasedexpression of intestine-specific proteins, and remodeling of the cell,occur prior to repression of pharyngeal cell-specific identity,suggesting that, unlike some examples of naturally occurringtransdifferentiation (Jarriault et al., 2008), this in vivoreprogramming takes place through intermediate cellular states inwhich characteristics of both the original and final cell types arepresent. We found no evidence of cell division during thereprogramming process based on cell number and position,indicating that erasing original cell identity through cell division isnot a necessary step in forced transdifferentiation. Although somemorphological characteristics of pharynx tissue are present in theanimals with transdifferentiated cells, these ultrastructural featuresare excluded in the regions that have undergone transdifferentiationand that exhibit an intestine-like morphology.
Fig. 2. Maintenance of intestinal differentiation markersin pharyngeal cells. (A) The elt-2 reporter is normallyexpressed only in intestinal nuclei (green) in non-heatshocked hs-elt-7 larvae. Anterior (right) and posterior pharynxregions in middle panel are enlarged in lower panels.(B) 24 hours after ectopic ELT-7 expression, the elt-2 reporteris expressed in cells inside and outside the pharynx. At96 hours, bright signal is maintained only in cells within thepharynx. Scale bars: 5 μm. (C) Percentage of wormsexpressing elt-2::lacZ::GFP in non-intestinal cells outside thepharynx (O), in the posterior pharynx (P) or in the anteriorpharynx (A). n, number of worms observed; t, time since heatshock in hours. (D) Immunoreactive ELT-2 in non-heat-shocked hs-elt-7 control larvae and hs-elt-7 larvae 48 hoursafter heat shock.
Dev
elop
men
t
4847
RESEARCH REPORT Development (2013) doi:10.1242/dev.103010
Transcription factor identity and cellular context influencesusceptibility to forced transdifferentiationAlthough transcription factors that regulate specification of each ofthe three germ layers can reprogram the fate of pre-gastrulationblastomeres in C. elegans, they are unable to reprogram cells in laterstage embryos without prior removal of inhibitory factors (reviewedby Joshi et al., 2010). We found that ELT-7 can initiate intestinegene expression in many differentiated cell types in embryos, larvae,and adults without removal of inhibitory factors, demonstrating thatnot all transcription factors adhere to the MCT. A growing numberof in vitro studies demonstrate that transcription factors can redirectdifferentiated cell identity without reversion to a pluripotentintermediate; however, the context and factors that result in efficientand stable transdifferentiation of any cell type remain relativelyunclear (reviewed by Ladewig et al., 2013).
We found that only pharynx cells appear to become fullytransdifferentiated and reprogrammed by ELT-7. We considered thepossibility that this reflects tissue-specific expression of the hsp-16-41/2 promoter used in the study, which is expressed at somewhathigher levels in the pharynx (Stringham et al., 1992). However, wefound that longer heat shocks (30 minutes and 60 minutes) alsofailed to result in cellular reprogramming outside the pharynx,
although such a regimen results in higher heat-shock promoteractivity in epidermal and muscle cells than in the pharynx under thebrief (15 minutes) heat-shock conditions used throughout this study.Thus, the cell-type specificity does not appear to be attributable todifferences in expression levels of the transgene.
Transdifferentiation of only pharynx marginal cells implies thatcellular context dictates susceptibility to reprogramming. With theexception of two cells in the posterior pharynx, the marginal cells areseparated in cell lineage from intestinal cells at the first cell division(Sulston et al., 1983); therefore, relatedness in cell lineage does notappear to explain their particular susceptibility to reprogramming byELT-7. The intestine and pharynx are both epithelial tubes that are apart of the digestive tract. It is conceivable that epithelial tube ordigestive tract identity has a role in establishing the context for directreprogramming into intestine per se; however, such an effect wouldnot extend to rectal epithelial cells, as we did not observe evidence fortransdifferentiation in that region. Although it is possible that marginalcells might be uniquely developmentally plastic, the results of Gilleardand McGhee (Gilleard and McGhee, 2001), Fukushige and Krause(Fukushige and Krause, 2005), and Turson and Patel (Turson andPatel, 2011) imply that differentiated marginal cells are not susceptibleto transdifferentiation into epidermis, muscle and neurons,
Fig. 3. Loss of pharynx-specific markers following apulse of ELT-7 expression. Expression of pharynxmuscle-specific ceh-22::GFP (A,B) or myo-2::GFP (C,D)reporters diminishes by 3 days after ectopic ELT-7expression compared with non-heat shocked hs-elt-7control larvae. Scale bars: 5 μm. Exposure time isindicated in the upper right-hand corner. (E) DecreasedIFA in cells expressing elt-2::lacZ::GFP compared withnon-GFP-expressing marginal cells within the samesection of the pharynx (boxed region shown in lowerpanels). Bottom panel, overlay of IFA (red) and elt-2::lacZ::GFP (green). Closed arrow indicates non-elt-2::lacZ::GFP-expressing marginal cell nucleus; openarrow indicates GFP-expressing nucleus. Faint greensignal over the non-expressing nucleus is the out-of-planesignal from the positive nucleus of the third marginal cell.(F) IFA average pixel intensity minus background wasnormalized to the cell with the highest pixel intensitywithin each worm. Normalized signal in anterior marginalcells with strong elt-2 reporter expression (greenhighlighted) was lower than in cells with weak or noexpression of elt-2 (white). Each row is an individualanimal and each column indicates the individual marginalcells ordered based on increasing GFP signal. Theaverage overall IFA signal is reduced in the elt-2::lacZ::GFP-positive marginal cells.
Dev
elop
men
t

4848
RESEARCH REPORT Development (2013) doi:10.1242/dev.103010
respectively. Rather, these cells may express a factor or set of factorsthat allow for transdifferentiation into intestine. Our currentobservations establish an in vivo model to investigate the role of cellcontext in stable transdifferentiation of somatic cells and revealremarkable plasticity in cellular differentiation in an organism with arigid pattern of cell division and identity.
MATERIALS AND METHODSC. elegans strains, maintenance, synchronization and heatshockTransgenic strains used were as follows: JR3410, wIs47 [hsp::end-1];rrIs1[elt-2::lacZ::GFP] (Fukushige et al., 1998; Zhu et al., 1998); JR3373,wIs125[hsp::elt-7] (Sommermann et al., 2010); rrIs1; JR3457, wIs125;CuIs2 V [ceh-22::gfp]; and JR3471, unc-119(ed3); ruIsIII[unc-119(+), myo-2::GFP; wIs125. Larvae were synchronized as described previously(Stiernagle, 2006) and heat shocked at the desired stage in M9 buffer at33°C for 15 minutes using a thermal cycler.
ImmunofluorescenceAntibodies MH33 (anti-IFB-2) (Bossinger et al., 2004) and anti-ELT-2 weregifts from J. McGhee (University of Calgary, Canada). Anti-IFA (Francisand Waterston, 1991) was a gift from G. Gunderson (Columbia University,NY, USA). Cy3 goat anti-rabbit and Cy3 goat anti-mouse were obtainedfrom Sigma. Fixation and permeabilization were carried out as describedpreviously (Sommermann et al., 2010). Animals were viewed with a ZeissAxioskop 2, Olympus BX60 or Nikon Eclipse Ti inverted microscope andimaged with a MicroFire camera or Hamamatsu flash Orca 2.8. Brightnessand contrast of some images were modified using NIS Elements software orPowerPoint to reveal relevant details more clearly in the figures.
IFA intensity measurementUsing NIS Elements software, individual marginal cells in the anteriorpharynx were outlined to create regions of interest (ROIs). Average intensityof the IFA signal was quantified within each of the three anterior marginal
cells in individual worms that contained one or two GFP-positive nuclei,bright staining, and non-elongated or abnormal nuclei. We compared onlycells within individual worms, in order to control for differences in stainingefficiency between worms. Background was determined by measuringaverage pixel intensity in region outside the worm.
Transmission electron microscopy (TEM)L1 larvae of strain JR3373 were heat shocked and inspected by fluorescencemicroscopy after 6 days. Worms with bright elt-2::lacZ::GFP-expressingnuclei and intestine-like morphology in the anterior pharynx were placed onice, shipped to the Hall lab, and subsequently transferred into a smallplanchette for the Bal-Tec HPM-010 High Pressure Freezing Machine usingE. coli as the surrounding matrix. Fixation and embedding proceduresgenerally followed Hall et al. (Hall et al., 2012). One or two animals werefast frozen per planchette, after which frozen samples were placed into anRMC FS-7500 Freeze Substitution unit, in a 1% osmium tetroxide solutionin 98% acetone, 2% dH2O. Samples were held at −90°C for 4 days, thenslowly warmed to 0°C, held for 3 days at 0°C, then rinsed in cold acetoneand gradually infiltrated with EmBed 812 resin. Samples were flatembedded between Aclar sheets, then cured at 60°C for 2 days. Singleworms were viewed under the dissecting microscope, cut out of the Aclarsandwich before re-embedding in fresh plastic resin and placed in a mold inprecise orientation before curing again at 60°C. The embedded sample wasserial thin-sectioned on an RMC PowerTome XL using a diamond knife,mounted on slot grids, post-stained with uranyl acetate and viewed with aPhilips CM10 electron microscope. Digital images were collected using anSIS camera system and viewed using iTEM or Photoshop softwareplatforms to analyze data and select images for illustrations.
AcknowledgementsWe thank Andrew Sumner for assistance with chemosensation experiments, andLeslie Gunther-Cummins and Geoff Perumal for help with the Bal-Tec HPF device.We thank G. Gunderson and J. McGhee for reagents. Some strains were providedby the CGC, which is funded by NIH Office of Research Infrastructure Programs(p40 O8010440).
Fig. 4. Reprogrammed pharyngeal cells haveintestine-specific morphology. Morphology of theposterior (A) and anterior (B) bulbs of the pharynx innon-heat shocked hs-elt-7 control animals andmorphology of the intestine (C), posterior bulb (D) andanterior bulb (E) after heat shock (D, 72 hours; E,96 hours) are seen using DIC. Boxed region (C′-E′)outlines elt-2::lacZ::GFP-expressing nuclei in C′′-E′′.Scale bars: 2 μm. (F-H) Overlay of IFB-2 (red) and elt-2::lacZ::GFP (green) signal in the endogenous intestine(F), and posterior (G) and anterior (H) pharynx 5 daysafter ectopic ELT-7. (I-K) Morphology of the intestine andremodeled pharynx by TEM. (Extensive images of theultrastructure of the wild-type pharynx for comparisoncan be viewed at wormatlas.org.) (I) Cross-section ofnormal L1 larva intestinal lumen. Microvilli (mv) on thetop are seen at nearly full length, whereas those at thebottom are at oblique angles. Black arrow indicates theterminal web. Scale bar: 1 μm. (J) Accessory lumen inthe anterior pharynx of hs-elt-7 L1 larva 6 days afterbrief ectopic ELT-7 expression. Arrows show remainingbuccal cuticle. Several long microvilli (mv) are seen andthe circular profiles in the middle of the side lumen areapparently additional microvilli extending at extremeangles from other edges of the side lumen. Some E. coli(Coli) lie within the buccal channel. Scale bar: 1 μm.(K) Higher magnification of ectopic microvilli (mv). Scalebar: 0.2 μm. Arrowheads indicate an electron-denseline, an apparent terminal web structure, runningsubjacent to the plasma membrane. The microvilliappear surrounded by diffuse glycocalyx. Nuc, nucleus.
Dev
elop
men
t

4849
RESEARCH REPORT Development (2013) doi:10.1242/dev.103010
Competing interestsThe authors declare no competing financial interests.
Author contributionsM.R.R. designed and performed experiments, and wrote the manuscript draft.A.W. performed experiments. K.C.Q.N. performed transmission electronmicroscopy experiments. D.H.H. interpreted the electron micrographs. J.H.R.conceived the project, and revised and approved the manuscript.
FundingThis work was supported by a training grant from the California Institute ofRegenerative Medicine, and grants from the National Institutes of Health [OD010943 to D.H.H. and HD062922 to J.H.R.]. Deposited in PMC for release after 12 months.
Supplementary materialSupplementary material available online athttp://dev.biologists.org/lookup/suppl/doi:10.1242/dev.103010/-/DC1
ReferencesAlbertson, D. G. and Thomson, J. N. (1976). The pharynx of Caenorhabditis elegans.
Philos. Trans. R. Soc. B 275, 299-325. Bossinger, O., Fukushige, T., Claeys, M., Borgonie, G. and McGhee, J. D. (2004).
The apical disposition of the Caenorhabditis elegans intestinal terminal web ismaintained by LET-413. Dev. Biol. 268, 448-456.
Ciosk, R., DePalma, M. and Priess, J. R. (2006). Translational regulators maintaintotipotency in the Caenorhabditis elegans germline. Science 311, 851-853.
Clokey, G. V. and Jacobson, L. A. (1986). The autofluorescent “lipofuscin granules” inthe intestinal cells of Caenorhabditis elegans are secondary lysosomes. Mech.Ageing Dev. 35, 79-94.
Djabrayan, N. J., Dudley, N. R., Sommermann, E. M. and Rothman, J. H. (2012).Essential role for Notch signaling in restricting developmental plasticity. Genes Dev.26, 2386-2391.
Francis, R. and Waterston, R. H. (1991). Muscle cell attachment in Caenorhabditiselegans. J. Cell Biol. 114, 465-479.
Fukushige, T., Hawkins, M. G. and McGhee, J. D. (1998). The GATA-factor elt-2 isessential for formation of the Caenorhabditis elegans intestine. Dev. Biol. 198, 286-302.
Fukushige, T., Hendzel, M. J., Bazett-Jones, D. P. and McGhee, J. D. (1999). Directvisualization of the elt-2 gut-specific GATA factor binding to a target promoter insidethe living Caenorhabditis elegans embryo. Proc. Natl. Acad. Sci. USA 96, 11883-11888.
Fukushige, T. and Krause, M. (2005). The myogenic potency of HLH-1 reveals wide-spread developmental plasticity in early C. elegans embryos. Development 132,1795-1805.
Gilleard, J. S. and McGhee, J. D. (2001). Activation of hypodermal differentiation inthe Caenorhabditis elegans embryo by GATA transcription factors ELT-1 and ELT-3.Mol. Cell. Biol. 21, 2533-2544.
Hall, D. H., Hartwieg, E. and Nguyen, K. C. (2012). Modern electron microscopymethods for C. elegans. Methods Cell Biol. 107, 93-149.
Horner, M. A., Quintin, S., Domeier, M. E., Kimble, J., Labouesse, M. and Mango,S. E. (1998). pha-4, an HNF-3 homolog, specifies pharyngeal organ identity inCaenorhabditis elegans. Genes Dev. 12, 1947-1952.
Jarriault, S., Schwab, Y. and Greenwald, I. (2008). A Caenorhabditis elegans modelfor epithelial-neuronal transdifferentiation. Proc. Natl. Acad. Sci. USA 105, 3790-3795.
Joshi P. M., Riddle M. R., Djabrayan N. J. and Rothman J. H. (2010).Caenorhabditis elegans as a model for stem cell biology. Dev. Dyn. 239, 1539-1554.
Ladewig, J., Koch, P. and Brustle, O. (2013). Leveling Waddington: the emergence ofdirect programming and the loss of cell fate hierarchies. Nat. Rev. Mol. Cell Biol. 14,225-236.
Maduro, M. F. and Rothman, J. H. (2002). Making worm guts: the gene regulatorynetwork of the Caenorhabditis elegans endoderm. Dev. Biol. 246, 68-85.
Mango S. E. (2007). The C. elegans pharynx: a model for organogenesis (ed. The C.elegans Research Community). WormBook, doi/10.1895/wormbook.1.129.1.
Okkema, P. G. and Fire, A. (1994). The Caenorhabditis elegans NK-2 classhomeoprotein CEH-22 is involved in combinatorial activation of gene expression inpharyngeal muscle. Development 120, 2175-2186.
Patel T., Tursun B., Rahe D. P. and Hobert O. (2012). Removal of Polycombrepressive complex 2 makes C. elegans germ cells susceptible to direct conversioninto specific somatic cell types. Cell Reports 2, 1178-1186.
Quintin, S., Michaux, G., McMahon, L., Gansmuller, A. and Labouesse, M. (2001).The Caenorhabditis elegans gene lin-26 can trigger epithelial differentiation withoutconferring tissue specificity. Dev. Biol. 235, 410-421.
Sommermann, E. M., Strohmaier, K. R., Maduro, M. F. and Rothman, J. H. (2010).Endoderm development in Caenorhabditis elegans: the synergistic action of ELT-2and -7 mediates the specification→differentiation transition. Dev. Biol. 347, 154-166.
Stiernagle T. (2006). Maintenance of C. elegans (ed. The C. elegans ResearchCommunity). WormBook, doi/10.1895/wormbook.1.101.1.
Stringham, E. G., Dixon, D. K., Jones, D. and Candido, E. P. (1992). Temporal andspatial expression patterns of the small heat shock (hsp16) genes in transgenicCaenorhabditis elegans. Mol. Biol. Cell 3, 221-233.
Sulston, J. E., Schierenberg, E., White, J. G. and Thomson, J. N. (1983). Theembryonic cell lineage of the nematode Caenorhabditis elegans. Dev. Biol. 100, 64-119.
Tursun, B., Patel, T., Kratsios, P. and Hobert, O. (2011). Direct conversion of C.elegans germ cells into specific neuron types. Science 331, 304-308.
Yuzyuk, T., Fakhouri, T. H., Kiefer, J. and Mango, S. E. (2009). The polycombcomplex protein mes-2/E(z) promotes the transition from developmental plasticity todifferentiation in C. elegans embryos. Dev. Cell 16, 699-710.
Zhu, J., Fukushige, T., McGhee, J. D. and Rothman, J. H. (1998). Reprogramming ofearly embryonic blastomeres into endodermal progenitors by a Caenorhabditiselegans GATA factor. Genes Dev. 12, 3809-3814.
Zhu, J., Hill, R. J., Heid, P. J., Fukuyama, M., Sugimoto, A., Priess, J. R. andRothman, J. H. (1997). end-1 encodes an apparent GATA factor that specifies theendoderm precursor in Caenorhabditis elegans embryos. Genes Dev. 11, 2883-2896.
Dev
elop
men
t
Recommended









![Chemical transdifferentiation: closer to regenerative medicine · 2019. 9. 24. · epigenetic modification. In this review, we briefly examine the history of cell ... factors [42]](https://img.pdfslide.us/doc/110x75/60f81fc29cd090799f0508df/chemical-transdifferentiation-closer-to-regenerative-2019-9-24-epigenetic.jpg)