
Vol. 132, No. 2, 1985
October 30, 1985
BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Pages 780-786
THE MITOGENIC/ONCOGENIC ~21 Ki-RAS PROTEIN STIMIJLATES AUENYLATE CYCLASE ACTIVITY EARLY IN THE GI PHASE OF
NRK IUT KIDNEY CELLS
Douglas J. Franksl,P, James F. Whitfieldl and Jon P. DurkinI*
1 Cell Physiology Group Division of Biological Sciences
National Research Council of Canada Ottawa, Ontario Canada KlA OR6
2 Department of Pathology Faculty of Health Sciences
University of Ottawa Ottawa, Ontario KlH 8M5
Received September 18, 1985
SUMMARY: tsK-NRK rat cells infected with a temperature-sensitive mutant of the Kirxen murine sarcoma virus were arrested in the GO/G1 phase of their cell cycle by incubation in serum-deficient medium at a temperature (41'C) which inactivates the virus' abnormally thermolabile mitogenic/oncogenic 21 kDa (p 21) RAS protein product. Reactivating the viral RAS protein by lowering the temperature to a permissive 36'C rapidly (within 1 hour) stimulated adenylate cyclase, sensitized the enzyme to stimulation by GTP and forskolin and caused the tsK-NRK cells to transit G1 and start replicating their DNA about 10 hours later. The 41'C + 36°C shift did not affect adenylate cyclase or stimulate G1 transit in uninfected NRK cells. Thus, an oncogenic viral RAS protein was able to stimulate adenylate cyclase and G1 transit in a mammalian cell just as other RAS proteins appear to do in yeast cells. 0 1985 Academic
Press, Inc.
A transient burst of adenylate cyclase activity and an
increase in the cellular cyclic AMP content have been observed in the GI
phases of the growth-division cycles of a wide variety of normal
eukaryotic cells (reviewed in [l]). The importance of this seemingly
ubiquitous burst of adenylate cyclase activity for GI transit is now
widely accepted, but its cause is unknown.
Eukaryotic cells, ranging from the primitive (e.g., the
budding yeast Saccharomyces cerevisiae) to the advanced (e-g., mouse 3T3
cells, rat liver cells, human cancer cells) also have a set of highly
conserved genes, the RAS oncogenes, which code for GTP-binding/GTPase
proteins (2). These proteins attach to the cell membrane and appear to
be structurally and functionally similar to the a subunits of the
membrane-associated G/F proteins that control adenylate cyclase activity
*To whom reprint requests should be addressed.
N.R.C.C. No: 24963
0006-291X/85 $1.50 Copyright 0 1985 by Academic Press, Inc. All rights of reproduction in any form reserved. 780

Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
(3,4). Some RAS proteins, such as those produced by the Harvey and
Kirsten murine sarcoma viruses that have amino acid substitutions at
positions 12 or 61, neoplastically transform, or are involved in the
transformation of, human and rodent cells (5).
A clear link has been established between ras gene expression
and adenylate cyclase activity in yeast (6,7). Moreover G1 transit of
2. cerevisiae is dependent upon functioning ras genes (8). I f cellular
ras gene products also influence adenylate cyclase activity in mammalian
cells as has been suggested (9) they might be responsible for the burst
of adenylate cyclase activity that accompanies, and is necessary for,
the G1 transit of these cells (1). Here we present evidence suggesting
that a RAS protein, in this case the oncogenic, abnormally thermolabile
21 kDa (~21) product of a temperature-sensitive Kirsten murine sarcoma
virus mutant (10) indeed causes an early increase in adenylate cyclase
activity during the G1 transit of a mammalian cell, the NRK rat kidney
fibroblast.
MATERIALS AND METHODS
Cell Lines. Normal rat kidney (NRK) cells and tsK-NRK cells were generous gifts from Dr. E.M. Scolnick (Merck, Sharpe and Dhome, West Point. PA).
.The tsK-NRK cells were produced by infecting normal NRK cells with a temperature-sensitive, transformation-defective mutant (ts 371) of Kirsten sarcoma virus that produces an abnormally thermolabile ~21 RAS protein (11).
Cell Culture. Cells were routinely cultured in Dulbecco's modification of Eagle's medium (DMEM) containing gentamycin and supplemented with 15% (v/v> b ov ne i calf serum (Colorado Serumn Co., Denver, CO) and maintained at 36°C in a humidified atmosphere of 95% air and 5% CO*. Before experiments, cells were detached from the culture dishes by a brief exposure to 0.25% trypsin in phosphate-buffered saline (PBS). They were then plated in 60 mm plastic Petri dishes at a density of 1.5 x 103/cm2 in 5 ml of complete medium consisting of 85% (v/v) DMEM and 15% (v/v) serum and incubated at 4O'C. After 48 hrs, the cells were arrested at GO/G1 by incubation at 41°C in DMEM-F12 (1:l) medium containing 10 mM Hepes pH 7.0 and 0.2% calf serum. These quiescent serum deprived cultures were stimulated to resume cycling by simply lowering the temperature from 41'C to 36°C.
Cyclic AMP Synthesis. The intracellular ATP pool was labeled by adding 0.5 uCi of 2,8 ['HI-adenine (ICN, Irvine, CA) to the medium 12 hours before the start of an experiment. At time "zero" (i.e. 48 hours after the shift to serum-deficient DMEM-F12 medium), the quiescent tsK-NRK cells were stimulated by lowering the temperature from 41'C to36"C. Other identically treated cultures were maintained at the non-permissive 41°C. To
8 revent degradation of the [3H]-cyclic AMP newly synthesized
from the [ HI-labeled ATP pool 3-isobutyl-l-methylxanthine (MIX) was added to each culture 30 min. before harvesting the cells.
At the times indicated cultures were washed three times with PBS and the cells lysed in 0.5 ml of lysis buffer (150 mM NaCl, 10 mM Na*HPO,,, 1 mM EDTA, 0.1 mM cyclic AMP, 0.1 mM MIX, and 1% Nonidet P-40). The culture dishes were scraped, rinsed with a further 0.5 ml of lysis buffer and debris was removed by centrifugation. Labeled nucleotides
781

Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
were separated by ion exchange chromatography. Each lysate was applied to a 10 x 40 mm column of AG 50 X 4 (Bio-Rad, Richmond, CA) and ATP was eluted with 4 ml of H20. The remaining nucleotides were eluted with 8 ml of HZ0 directly onto a 15 x 10 mm column of neutral alumina. Cyclic AMP was then eluted with 5 ml of 50 mM Tris ( H 7.5). To monitor the cyclic AMP recovery, all extracts contained [ 5 PI-cyclic AMP. Replicate cultures were used to determine cell protein by the method of Bradford (12). The labeled ATP pool remained constant throughout the experimental period at a level of lOO,OOO-150,000 cpm/mg protein. All assays were carried out in triplicate.
Adenylate Cyclase Assay. Cultures were washed twice with PBS and the cells scraped off the dish in 2 ml of PBS. The cells were sedimented by centrifugation at 500 xg for 3 min, washed with a further 5 ml of PBS and centrifuged again. The pellet was frozen at -90°C. The thawed pellets were homogenized in 0.5 ml of buffer (50 u&f Tris [pH 7.41, 330 mM sucrose, 1 mM MgCl*, and 1 n@f dithiothreitol) in a motor-driven teflon/glass homogenizer (10 strokes, 10,000 rpm). The homogenate was centrifuged at 20,000 x g for 20 min, the supernatant fluid discarded and the pellet dispersed in a small volume of buffer. This cell-free preparation contained adenylate cyclase, determined from the conversion of [a-32
the activity of which was PI-ATP to [32P]-cyclic AMP as
previously described (13). The enzyme reaction was linear with time and protein concentration and all assays were carried out in triplicate.
DNA-Synthetic Activity. DNA-synthetic activity was measured auto- radiographically by exposing cultures to [3H]-thymidine (5 nCi/ml of medium; Sp. Act. 20 Ci/mmole; New England Nuclear Corp., Boston, MA) according to Durkin and WhitfIeld (14). The percentage of cells with [3H] thymidine labeled nuclei was determined by examining at least 1000 cells.
RESULTS AND DISCUSSION
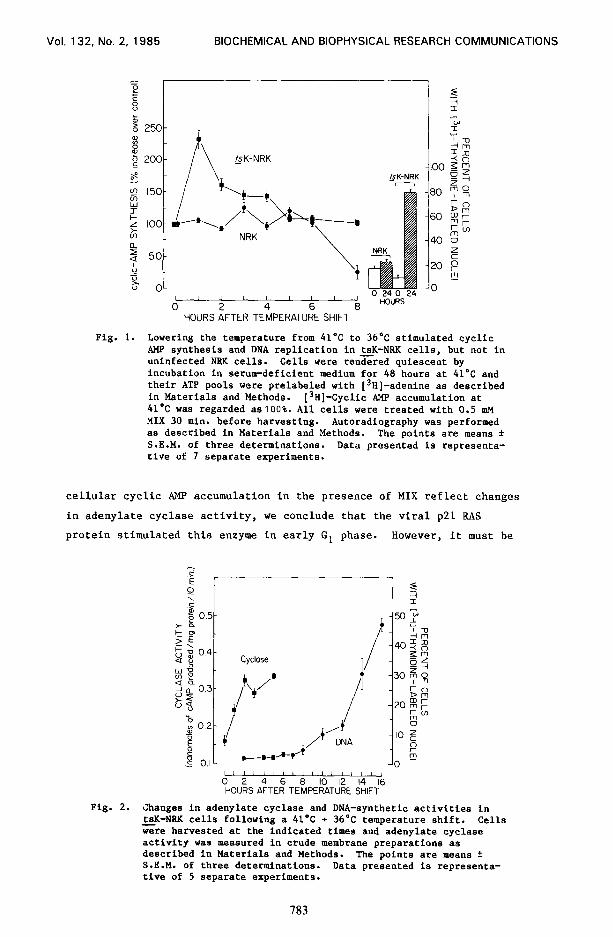
The oncogenic 21 kDa RAS protein made by the Ki-MSV mutant,
ts 371, carried by tsK-NRK cells is inactivated by temperatures above - - 39'C (15) and cells grown at this elevated temperature are
phenotypically untransformed. Thus, tsK-NRK cells, like uninfected and - untransformed NRK cells, were arrested in the GO/G1 phase of their cycle
when incubated in serum-deficient medium for 48 hours at 41°C (Figs. 1
and 2). Lowering the temperature to permissIve levels has been shown to
reactivate the thermolabile viral ~21 RAS protein (15). In particular,
lowering the temperature from 41°C to 36'C activated enough viral ~21
RAS protein to cause tsK-NRK cells to transit G1 and start replicating - DNA about 10 hrs later despite the serum deficiency (Figs. 1 and 2). By
contrast, the temperature shift did not cause uninfected NRK cells to
transit G1 (Figs. 1 and 2). The Ki-RAS protein's mitogenicity will be
characterized fully elsewhere.
Lowering the temperature from 41'C to 36'C in the presence of
MIX, a cyclic nucleotide phosphodiesterase inhibitor, increased the
accumulation of cyclic AMP in tsK-NRK cells, but did not affect cyclic - AMP accumulation in the still quiescent, uninfected NRK cells. The
results of a typical experiment are shown in Fig. 1. Since changes in
782
Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
100
-80
-60
-40
-20
-0
Fig. 1.
HOURS AFTER TEMPERATURE SHIFT
Lowering the temperature from 41’C to 36*C stimulated cyclic AMP synthesis and DNA replication in tsK-NKK cells, but not in uninfected NKK cells. Cells were rendered quiescent by incubation in serum-deficient medium for 48 hours at 41°C and their ATP pools were prelabeled with [3H]-adenine as described in Materials and Methods. [3H]-Cyclic AMP accumulation at 41°C was regarded aslOO%. All cells were treated with 0.5 m?l MIX 30 min. before harvesting. Autoradiography was performed as described in Materials and Methods. The points are means f S.E.M. of three determinations. Data presented is representa- tive of 7 separate experiments.
cellular cyclic AMP accumulation in the presence of MIX reflect changes
in adenylate cyclase activity, we conclude that the viral p21 RAS
protein stimulated this enzyme in early G1 phase. However, it must be
Fig. 2.
1111111111lII,I,
0 2 4 6 8 IO 12 14 16 HOURS AFTER TEMPERATURE SHIFT
Changes in adenylate cyclase and DNA-synthetic activities in tsK-NKK cells following a 41’C + 36°C temperature shift. Cells Gre harvested at the indicated times and adenylate cyclase activity was measured in crude membrane preparations as described in Materials and Methods. The points are means f S.E.M. of three determinations. Data presented is representa- tive of 5 separate experiments.
783

Vol. 132, No. 2. 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
noted that there was no detectable increase in cyclic AMP accumulation
in temperature shifted tsK-NRK cells in the absence of MIX, which - suggests that the viral mitogenic action did not rely on an overall
increase in cyclic AMP content. Hence, the viral RAS protein probably
caused a localized increase in cyclic AMP accumulation similar to those
that accompany and mediate various hormone responses (16).
To confirm that the viral ~21 protein did stimulate adenylate
cyclase, cells were harvested at various times after the p21 RAS-
activating temperature shift and the adenylate cyclase activities in
crude membrane preparations from these cells were measured. The results
of a typical experiment are shown in Fig. 2. The 41°C + 36OC shift
caused an early increase in adenylate cyclase activity as cells
transited the Gl phase. This burst of adenylate cyclase activity in
temperature-shifted tsK-NRK cells was probably due to the reactivated - viral RAS protein rather than the 5' temperature drop, because lowering
the temperature from 41'C to 36'C did not increase adenylate cyclase
activity in membranes from identically treated uninfected NRK cells
(data not shown).
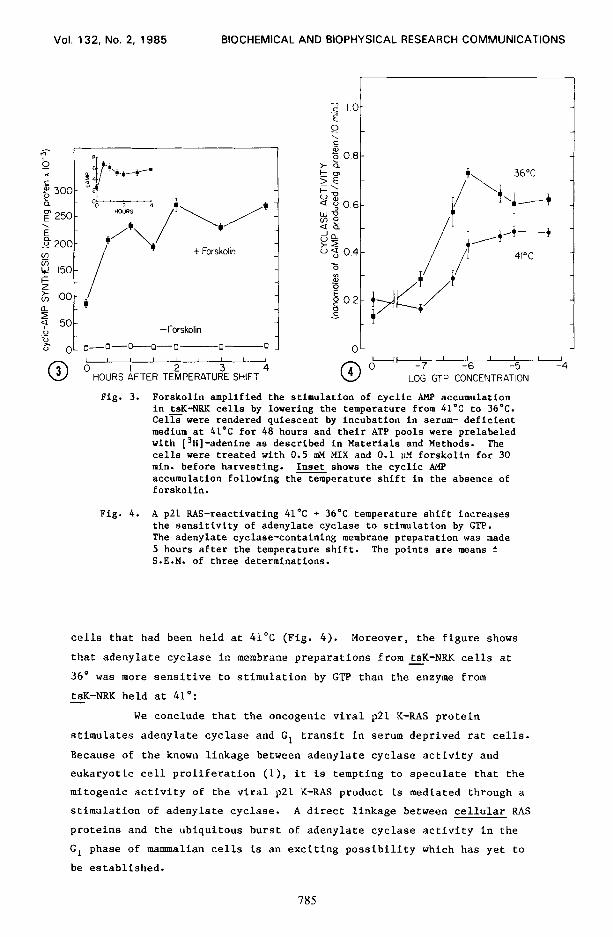
The diterpene, forskolin, is a potent adenylate cyclase
activator (17) and a component of its activation is dependent on the
presence of the regulatory G/F protein (18). I f the viral p21 RAS
protein functions as a G/F protein (3), then reactivation of this
protein by temperature shift in tsK-NRK cells should amplify the ability - of forskolin to stimulate cyclic AMP synthesis. Adding O.lnM forskolin
to quiescent tsK-NRK cultures at 4l”C, 30 min. before harvesting the - cells caused a 40-fold increase in cyclic AMP synthesis (Fig. 3).
However, the forskolin-induced amplification of adenylate cyclase
activity was far greater (i.e., almost loo-fold) in tsK-NRK cells which - had been shifted from 41'C to a ~21 activating 36°C (Fig. 3). This
large forskolin-induced increase in cyclic AMP accumulation in the
temperature-shifted tsK-NRK cells followed the same time course as the - smaller increase that occurred in these cells without forskolin (Fig. 3,
inset). Hence, the increase in cyclic AMP synthesis induced by p21 RAS
activation was greatly potentiated by forskolin, which is consistent
with the viral RAS protein functioning as a G/F protein.
For the above hypothesis to be correct, reactivation of the
membrane viral ~21 RAS protein in tsK-NRK cells should be marked by a - sensitization of adenylate cyclase in membrane preparations to
stimulation by GTP. The response of adenylate cyclase in membranes from
temperature-shifted tsK-NRK cells to increasing concentrations of GTP - was indeed greater than the response to GTP in membranes from tsK-NRK -
784
Vol. 132, No. 2. 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
0”“““’
I I I I 1 I 1
3 0 HOURS AFTER T&PERAT& SHIFT
4 0 4 O -’ -7 -6 -5 -4
LOG GTP CONCENTRATION
Fig. 3. Forskolin amplified the stimulation of cyclic AMP accumulation in tsK-NRK cells by lowering the temperature from 4L°C to 36OC. Cell were rendered quiescent by incubation in serum- deficient medium at 41’C for 48 hours and their ATP pools were prelabeled with [3H]-adenine as described in Materials and Methods. The cells were treated with 0.5 mM MIX and 0.1 PM forskolin for 30 min. before harvesting. Inset shows the cyclic AMP accumulation following the temperature shift in the absence of forskolin.
Fig. 4. A ~21 RAS-reactivating 41’C + 36’C temperature shift increases the sensitivity of adenylate cyclase to stimulation by GTP. The adenylate cyclase-containing membrane preparation was made 5 hours after the temperature shift. The points are means f S.E.M. of three determinations.
cells that had been held at 41°C (Fig. 4). Moreover, the figure shows
that adenylate cyelase in membrane preparations from tsK-NRK cells at - 36" was more sensitive to stimulation by GTP than the enzyme from
tsK-NRK held at 41': - We conclude that the oncogenic viral ~21 K-RAS protein
stimulates adenylate cyclase and GI transit in serum deprived rat cells.
Because of the known linkage between adenylate cyclase activity and
eukaryotic cell proliferation (l), it is tempting to speculate that the
mitogenic activity of the viral ~21 K-RAS product is mediated through a
stimulation of adenylate cyclase. A direct linkage between cellular RAS
proteins and the ubiquitous burst of adenylate cyclase activity in the
GI phase of mammalian cells is an exciting possibility which has yet to
be established.
785

Vol. 132, No. 2, 1985 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ACKNOWLEDGEMENTS
Supported in part by a grant (MA-5718) from the Medical
Research Council of Canada to D.J.F. The authors thank Ms. Karen Prokai
and Ms. Sylvia Soder for excellent technical assistance, D.J. Gillan for
preparing the illustrations and Mrs. K. Hamelin for processing the
manuscript.
1.
2. 3.
4. 5.
6.
7. 8.
9.
10.
11.
12. 13.
14.
15.
16.
17.
18.
REFERENCES
Boynton, A.L. and Whitfield, J.F., Adv. Cyclic Nucleotide Res. (1983) 15: 193-294. Shih, T.Y. and Weeks, M.O. (1984) Cancer Xnvest. 2: 109-123. Hurley, J.B., Simon, M.I., Teplow, D.B., Robishaw, J.D. and Gilman, A.G. (1984) Science, 226: 860-862. Gilman, A.G. (1984) Cell 36: 577-579. Ellis, R.W., Lowry, D.R. and Scolnick, E.M. (1982) Adv. Viral Oncol. 1: 107-126. Toda, T., Uno, I., Ishikawa, T., Powers, S., Kataoka, T., Broek, D., Cameron, S., Broach, J., Matsumoto, K. and Wigler, M. (1985) Cell 40: 27-36. Frankel, D.G. (1985) Proc. Natl. Acad. Sci. USA 82: 4740-4744. Kataoka, T., Powers, S., Cameron, S., Fasano, O., Goldfarb, M., Broach, J. and Wigler, M. (1985) Cell. 40: 19-26. Saltarelli, D., Fischer, S. and Gacon, G. (1985) Biochem. Biophys. Res. Comm. 127: 318-325. Scolnick, E.M., Stephenson, J.R. and Aaronson, A.A. (1972) J. Virol. 10: 653-657. Scolnick, E.M., Goldberg, R.J. and Parks, W.P. (1974) Cold Spring Harbor Symp. Quant. Biol. 39: 885-895. Bradford, M. (1979) Anal. Biochem. 72: 248-254. Franks, D.J., Plamondon, J. and Hamet, P. (1984) J. Cell. Physiol. 119: 41-45.. Durkin, J.P. and Whitfield, J.F. (1979) J. Cell. Physiol. 120: 135-145. Shih, T.Y., Weeks, M.O., Young, H-A. and Scolnick, E.M. (1979) J. Virol. 31: 546-556. Rasmussen, H. (1981) Calcium and cyclic AMP as synaptic messengers. John Wiley and Sons, New York. Seamon, K.B. and Daly, J.W. (1981) J. Cyclic Nucleotide Res. 7: 201-204. Insel, P., Stengel, D., Ferry, N. and Hanoune, J. (1982) J. Biol.
Chem. 257: 7485-7490.
786
Recommended

















