
1
Membrane Bioinformatics SoSe 2009
Helms/Böckmann
2
Last Week:
Plasma Membrane:composition & function,
membrane models
Fats & Fatty Acids:Different Motor Protein: F1-ATP
Synthasepes of fatty acids, strange lipids,
composition of membranes
Membrane Electrostatics

3
Today:
Self-organization of membranes (self-assembly, stability of lipid bilayers, order parameters)
Elasticity of bilayers (theory, experiment, simulation)
4
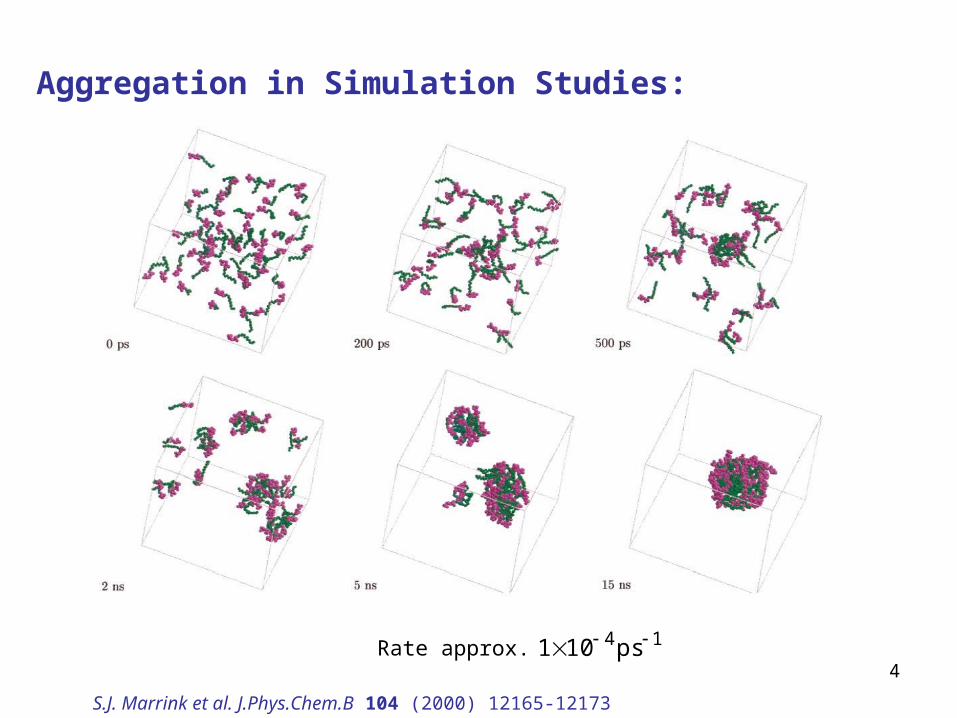
Aggregation in Simulation Studies:
S.J. Marrink et al. J.Phys.Chem.B 104 (2000) 12165-12173
Rate approx. 14ps101

5
Aggregation in Simulation Studies:
Fast initial aggregation of lipids, separation into lipid and aqueous domains (200ps)Formation of bilayer-like phase with defects (≈5ns)Defect lifetime ≈20ns
bilayer with defectS.J. Marrink et al. JACS 123 (2001) 8638-8639
6
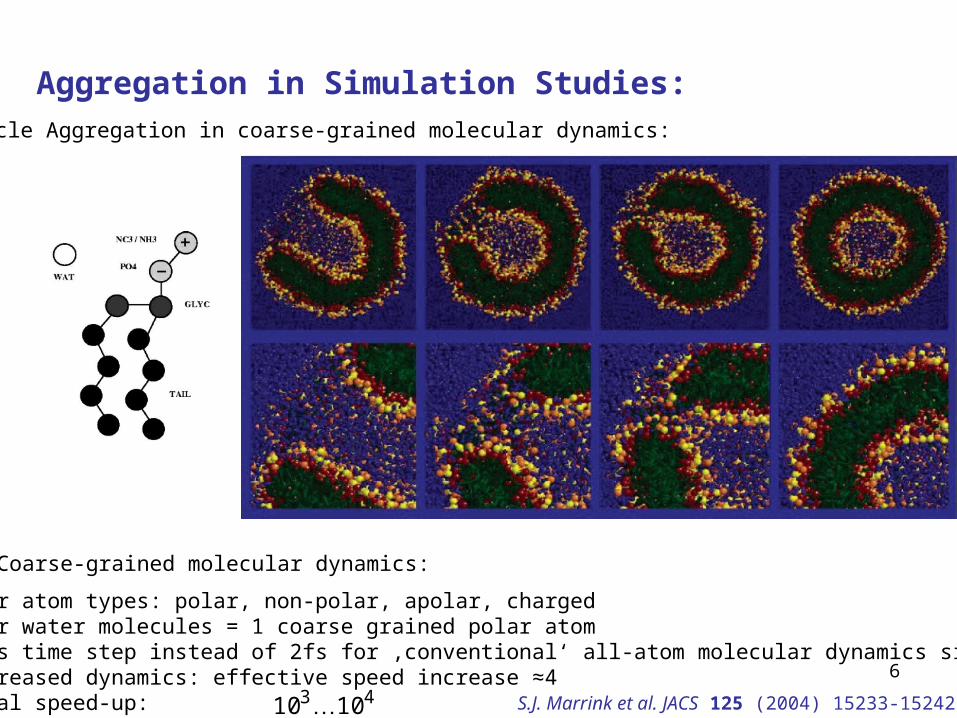
Aggregation in Simulation Studies:Vesicle Aggregation in coarse-grained molecular dynamics:
Coarse-grained molecular dynamics:
Four atom types: polar, non-polar, apolar, chargedFour water molecules = 1 coarse grained polar atom50fs time step instead of 2fs for ‚conventional‘ all-atom molecular dynamics simulationsIncreased dynamics: effective speed increase ≈4Total speed-up: 43 1010 S.J. Marrink et al. JACS 125 (2004) 15233-15242

7
Aggregation in Simulation Studies:Vesicle Aggregation in coarse-grained molecular dynamics:
What we can learn from simulation studies about aggregation (future):
Aggregation rates, dependency on temperature, pressure, ...Ab initio lipid distribution for mixed lipid systems, mixed micellesPore frequenciesEffect of detergent molecules... S.J. Marrink et al. JACS 125 (2004) 15233-15242
8
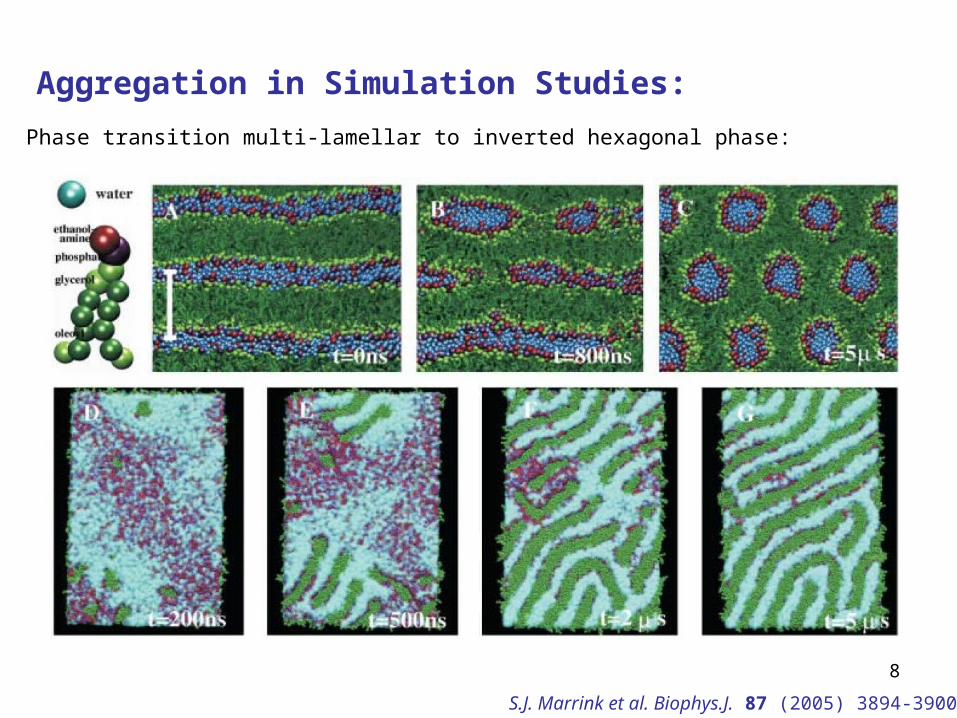
Aggregation in Simulation Studies:
Phase transition multi-lamellar to inverted hexagonal phase:
S.J. Marrink et al. Biophys.J. 87 (2005) 3894-3900

9
Aggregation in Simulation Studies:Hexagonal phase:
S.J. Marrink et al. Biophys.J. 87 (2005) 3894-3900

10
Aggregation in Simulation Studies:
rhombohedral phase:
S.J. Marrink et al. Biophys.J. 87 (2005) 3894-3900

11
Self-Organization of Membranes
Ebind : energy required to expose hydrophobic region of amphiphile to water
hydrophilic head
hydro-phobic tail cccln
hcR
cn : number of C-atoms
nm126.0lcc : average C-C bond length projected on chain
Area of hydrophobic chain: cchcc lRn2=A
Enthalpic change for exposure (energy required to create new water-hydrocarbon interface):
•lRn2=•A=E cchccbind
Free enthalpy change (free energy):
STESTHG bind
Define: for lipids aggregated in micelle or bilayer 0G

12
Statistical Physics: Entropy of an Ideal Gas
Canonical partition function: r
rc eZ
r : energy of state r
Tk1
B
r
: sum over all possible states r of the gas
Free Energy F=E-TS: cZln1
F
Entropy S: cB ZlnEkS
c
r
rr
c
Zlne
Z1
EAverage energy E of the system:

13
Statistical Physics: Entropy of an Ideal Gas
N1N1
p,...,p,r,...,rN3c pdpdrdrde
h
1!N
1Z N1N1
Partition function for a gas of undistinguishable particles:
N! different possibilities to arrange N identical atoms in the sum for the partition functionh3 phase space volume occupied by one state (normalization)
Energy of an ideal gas: N1
2N
21 r,,rV
m2p
m2p
E
kinetic energy potential energy
Rewrite the partition function as:space
N3momentumZZ
!N1
Z N
N1r,...,rV
space VrdrdeZ N1
2/1
m2p
momentumm2
h1
dpeh1
Z
2
with
No interaction between
particels (V=0)
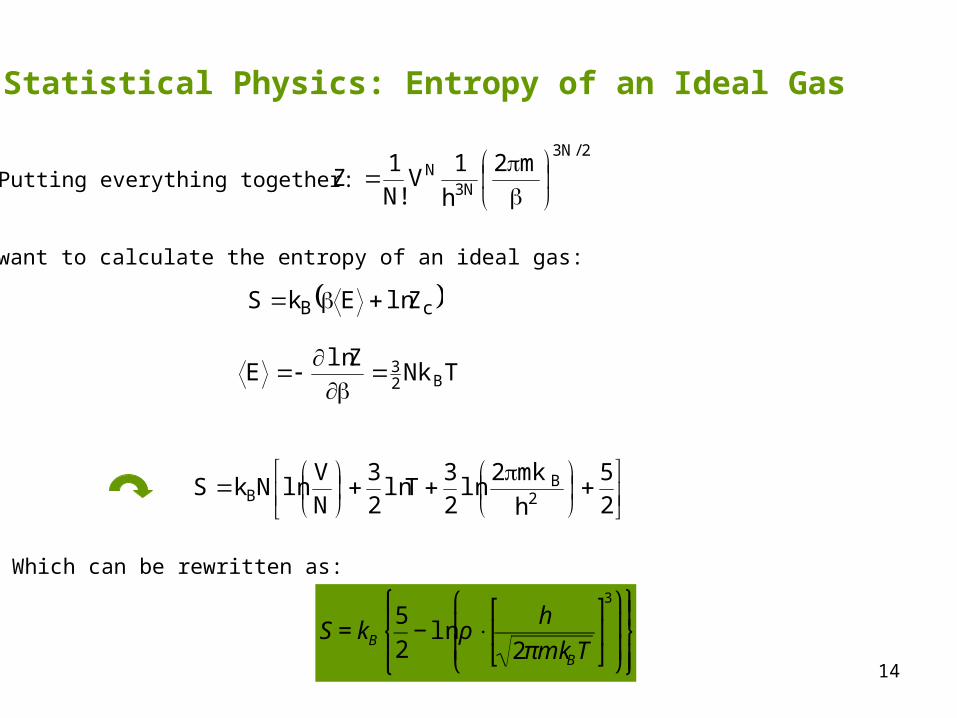
14
Statistical Physics: Entropy of an Ideal Gas
2/N3
N3N m2
h
1V
!N1
Z
Putting everything together:
cB ZlnEkS
TNkZln
E B23
We want to calculate the entropy of an ideal gas:
25
h
mk2ln
23
Tln23
NV
lnNkS2
BB
Which can be rewritten as:
€
S = kB5
2− ln ρ ⋅
h
2πmkBT
⎡
⎣ ⎢
⎤
⎦ ⎥
3 ⎛
⎝
⎜ ⎜
⎞
⎠
⎟ ⎟
⎧ ⎨ ⎪
⎩ ⎪
⎫ ⎬ ⎪
⎭ ⎪

15
Self-Organization of Membranes
Entropy S per molecule of an ideal gas at number density ρ:
3
BB Tmk2
hln
25
kS
Assumption 1: lipids in solution sufficiently dilute behaviour of lipids as ideal gas
Assumption 2: entropy of bulk water unchanged
- γ includes changes in entropy of close water molecules upon ordering
STEG bind dependent only on density of lipids ρ
-low ρ : entropy dominates, solution phase is dominated-large ρ : Ebind favors condensed phase

16
Self-Organization of Membranes
Cross-over between phases:
STE0G bind
•lRn2=•A=E cchccbind
3
BB Tmk2
hln
25
kS
=
TBkbindE
253
Bagg e
Tmk2h
-threshold for aggregation decreases as the binding energy of lipids increases

17
Self-Organization of Membranes
1st case: single chain phospholipid with 10 carbons (400 Dalton)
pm1.5m101.5Tmk2
h 12
B
Length scale:
Surface tension: 22
nmmol
kJ3.0J/m05.0
(for short alkanes)
Effective radius of single chain: 0.2nm
20Tk
E
B
bind M3.0agg
2nd case: double chain phospholipid with 10 carbons per chain (570 Dalton)
pm3.4m103.4Tmk2
h 12
B
Length scale:
Effective radius of double chain: 0.3nm 30Tk
E
B
bind
M102 5agg

18
Self-Organization of Membranes
single chain phospholipid with 10 carbons (400 Dalton) M3.0agg
double chain phospholipid with 10 carbons per chain (570 Dalton) M102 5agg
Experimental: M1010 23
Experimental: M1010 35
CMC = critical micelle concentration :
-cmc strongly depends also on the hydrophilic headgroup-computed numbers are very sensitive to the geometric properties (e.g. radius)
Single chain lipids uniformly higher cmc than double chain lipidsExponential decrease with number of chain carbons:
cmc decreases faster for double chain PC
TBkcclhcRcn2
253
Bagg e
Tmk2h
RnPC
RnRnPC

19
Molecular Packing in Different Aggregate Shapes
Important quantities:
Area per lipid
Volume of single, satu-rated hydrocarbon chain:
hc
hchc l
a
33chc nm10n9.264.27
I. Spherical Micelle:
3
2
R34
volume
R4surface
Number of molecules (area a0, volume vhc):
0
hc
hc3
02
a3R
/R34
or a/R4
If equal:
Condition:
31
la
lR
hc0
hc
hc
Spherical micelles are favored by large vaues for the area/lipid
2R

20
Molecular Packing in Different Aggregate Shapes
II. Cylindrical Micelle:
R
t
tRvolume
Rt2surface2
Number of molecules in the section:
0
hc
hc2
0
a2R
/tR or a/Rt2
If equal:
21
la31
lR
hc0
hc
hc
Condition for cylindrical micelles:

21
Molecular Packing in Different Aggregate Shapes
III. Bilayer:
hc0
0
lavolume
asurface
Ideal bilayer: 1la hc0
hc
1la2
1
hc0
hc
Condition for bilayers:
Typical area/lipid: 50...70Å2
Typical chain length: 16 carbon atoms ≈ 20ÅVolume: 916Å3
Double chain phospholipids:
92.065.0la hc0
hc
double chain phospholipids preferentially form lipid bilayers!

22
Molecular Packing in Different Aggregate Shapes
IV. Inverted Micelle:
volume > area x chain length(small headgroup area)
1la hc0
hc

23
Molecular Packing in Different Aggregate Shapes
A thermodynamics view:
N)p,T(pVTSE)N,p,T(G
pVE)N,p,S(H
TSE)N,V,T(F
NpVTS)N,V,S(E
Energy
Free Energy
Enthalpy
Free Enthalpy / Gibbs Free Energy
Thermodynamic Potentials:
dNVdpSdT)pV(d)TS(ddEdG
dNVdpTdS)pV(ddEdH
dNpdVSdT)TS(ddEdF
dNpdVTdSdE
total differentials:
The potentials are all extensive quantities, i.e.:
Thermodynamic potentials are state variables, i.e. they depend unambiguously on the state variables T,p,N,V,S
NV
,NS
eN)N,V,S(E

24
A thermodynamics view:
Entropy S is maximal for the equilibrium state of a closed system: (second law of thermodynamics)
Vp, given for min,G
VT, given for min,F
VE, given for max,S
Often the Free Enthalpy or the Gibbs Free Energy G is referred to as the Free Energy of a system
Thermodynamic Forces: derivatives of the thermodynamic potentials
STG
TSH
STF
TSE
N,p
N,p
N,V
N,V
VpG
VpH
pVF
pVE
N,T
N,S
N,T
N,S
T,pNG
: chemical potential
µ minimal at equilibrium!
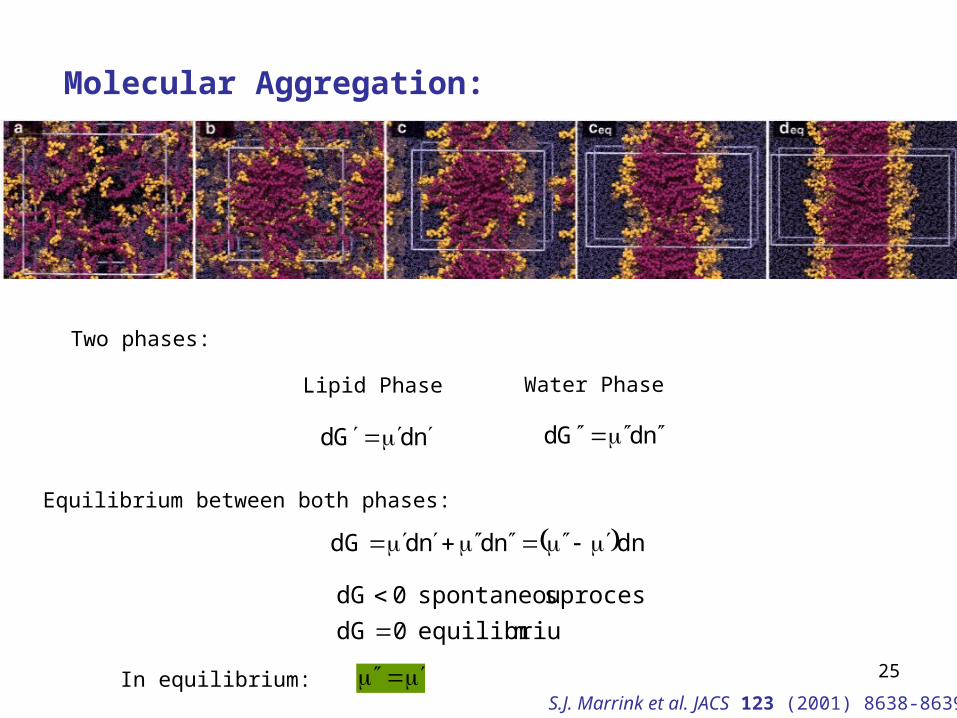
25
Molecular Aggregation:
Two phases:
Lipid Phase Water Phase
ndGd ndGd
Equilibrium between both phases:
dnndnddG
mequilibriu 0dG
process sspontaneou 0dG
In equilibrium: S.J. Marrink et al. JACS 123 (2001) 8638-8639

26
Molecular Aggregation:
Chemical potential for ideal gas:
Ideal gas(*): p
RTNV
NRTpV
o
*o
o*o
o*
p
op
pp
lnRT
plnplnRTpdp
d
pdp
RTd
Inserting (*): clnRT
VRTN
lnRT
o
*o
c=molar concentration of an ideal gas
e)temperatur constant (at dpNV
NdG
d
pVTSEN1
NG

27
Molecular Aggregation:
clnRT
clnRT
o
o
Equilibrium concentrations of lipids in lipid and in water phase:
RT/GRT/oo eecc
: distribution coefficient
Equilibrium constant for the transfer of lipids from bilayer/micelle to water phase:
RNk ,ec*C ABTBk/
o
Empirical rule for one chain amphiphiles:
kJ/mol n311 c
Lyso-DPPC: M 102*C 5
DPPC: M 10*C 12
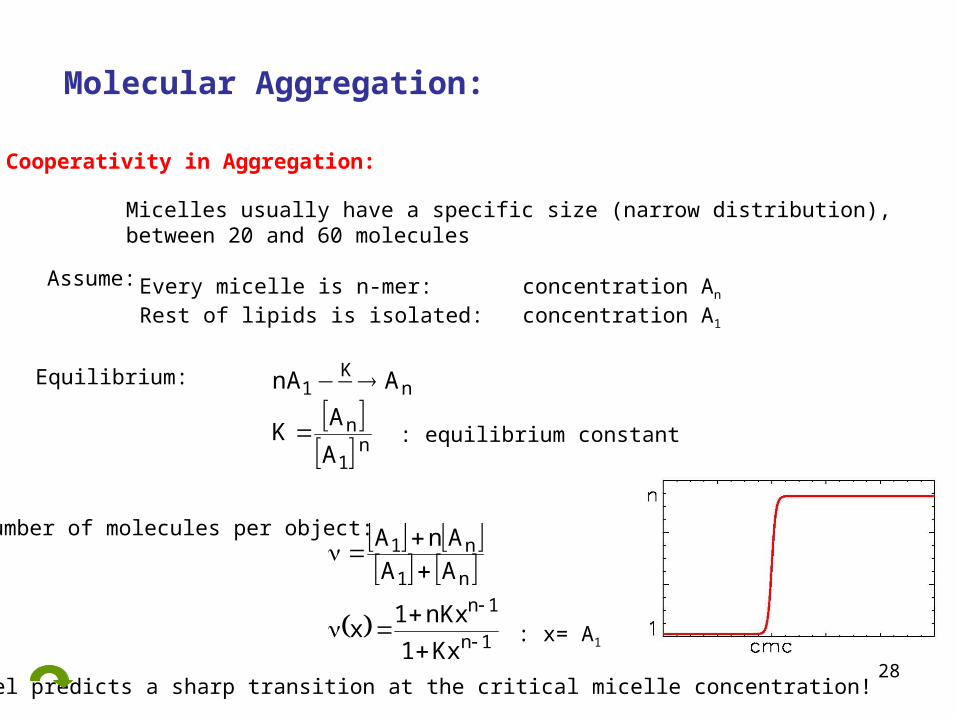
28
Molecular Aggregation:
Cooperativity in Aggregation:
Micelles usually have a specific size (narrow distribution), between 20 and 60 molecules
Assume: Every micelle is n-mer: concentration An
Rest of lipids is isolated: concentration A1
Equilibrium:
n
1
n
nK
1
A
AK
AnA
: equilibrium constant
Number of molecules per object:
1n
1n
n1
n1
Kx1
nKx1x
AAAnA
: x= A1
Model predicts a sharp transition at the critical micelle concentration!
Recommended








![SOSE Unit - We Are One[1]](https://img.pdfslide.us/doc/110x75/55275e7d550346d7358b47eb/sose-unit-we-are-one1.jpg)








