Embed Size (px)
Citation preview

Widespread occurrence and genetic diversity of marineparasitoids belonging to Syndiniales (Alveolata)
L. Guillou,1* M. Viprey,1 A. Chambouvet,1
R. M. Welsh,2 A. R. Kirkham,3 R. Massana,4
D. J. Scanlan3 and A. Z. Worden2
1Station Biologique de Roscoff, UMR7144, CNRS etUniversité Pierre et Marie Curie, BP74, 29682 RoscoffCedex, France.2Monterey Bay Aquarium Research Institute, 7700Sandholdt Road, Moss Landing, CA 95039, USA.3Department of Biological Sciences, University ofWarwick, Coventry CV4 7AL, UK.4Institut de Ciències del Mar, CMIMA, Passeig Marítimde la Barceloneta 37-49, 08003 Barcelona, Catalonia,Spain.
Summary
Syndiniales are a parasitic order within the eukaryoticlineage Dinophyceae (Alveolata). Here, we analysedthe taxonomy of this group using 43655 18S rRNAgene sequences obtained either from environmentaldata sets or cultures, including 6874 environmentalsequences from this study derived from Atlantic andMediterranean waters. A total of 5571 out of the 43655sequences analysed fell within the Dinophyceae.Both bayesian and maximum likelihood phylogeniesplaced Syndiniales in five main groups (I–V), as amonophyletic lineage at the base of ‘core’ dinoflagel-lates (all Dinophyceae except Syndiniales), althoughthe latter placement was not bootstrap supported.Thus, the two uncultured novel marine alveolategroups I and II, which have been highlighted previ-ously, are confirmed to belong to the Syndiniales.These groups were the most diverse and highlyrepresented in environmental studies. Within each,8 and 44 clades were identified respectively.Co-evolutionary trends between parasitic Syndinialesand their putative hosts were not clear, suggestingthey may be relatively ‘general’ parasitoids. Based onthe overall distribution patterns of the Syndiniales-affiliated sequences, we propose that Syndiniales areexclusively marine. Interestingly, sequences belong-ing to groups II, III and V were largely retrieved from
the photic zone, while Group I dominated samplesfrom anoxic and suboxic ecosystems. Nevertheless,both groups I and II contained specific clades prefer-entially, or exclusively, retrieved from these latterecosystems. Given the broad distribution of Syndini-ales, our work indicates that parasitism may be amajor force in ocean food webs, a force that isneglected in current conceptualizations of the marinecarbon cycle.
Introduction
The Alveolata, one of the major eukaryotic lineages, iscomposed of four protist classes: the Ciliophora, theApicomplexa, the Perkinsea and the Dinoflagellata.Alveolata have adopted a large range of trophic modesand habitats. They can be important marine primaryproducers. For instance, about half of dinoflagellates arephotosynthetic (Lessard and Swift, 1986), with somebeing responsible for toxic algal blooms. Ciliates anddinoflagellates can also be active predators; others arecoral symbionts (e.g. the dinoflagellate Symbiodiniumspp.) and some ciliates even reside in mammalian guts(Williams and Coleman, 1992). Finally, a large number ofspecies are parasites, broadly distributed throughoutthese Alveolata classes. For instance, Apicomplexa iscomposed solely of obligate parasites. In addition to phy-logenetic relatedness based on gene sequence compari-sons, Alveolata are unified by specific morphologicalcharacters. These include the presence of membrane-bound flattened vesicles, termed alveoli (Cavalier-Smith,1993; Patterson, 1999), distinct pores called microporesthat pierce the outer membrane and the presence of amore or less developed apical complex apparatus usedby alveolate parasites to enter their host (reviewed byLeander and Keeling, 2003).
The discovery of novel marine alveolate (MALV) lin-eages in marine planktonic communities by culture-independent techniques, specifically MALV groups I and II(Díez et al., 2001a; López-García et al., 2001; Moon-vander Staay et al., 2001), has raised questions regardingfunctional roles of these diverse populations. Phyloge-netic analyses showed MALV Group II belongs to theSyndiniales, a dinoflagellate order exclusively composedof marine parasites. This assignment was based on theclose phylogenetic relatedness of the environmental 18S
Received 17 April, 2008; accepted 29 June, 2008. *Forcorrespondence. E-mail [email protected]; Tel. (+33) 2 98 29 2379; Fax (+33) 2 98 29 23 24.
Environmental Microbiology (2008) 10(12), 3349–3365 doi:10.1111/j.1462-2920.2008.01731.x
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing LtdNo claim to original French government works

rRNA gene sequences (Moon-van der Staay et al., 2001;Skovgaard et al., 2005) with three previously describedgenera: Amoebophrya spp., Hematodinium spp. and Syn-dinium spp. More recently, 18S rRNA gene sequencesderived from Ichthyodinium chaberladi (Gestal et al.,2006) and Duboscquella sp. (Harada et al., 2007), twoparasitic Syndiniales genera, showed an affiliation withMALV Group I.
MALV groups I and II have been retrieved from variousmarine habitats, mainly from the picoplankton fraction (< 2or < 3 mm size fractionated samples). These groups fre-quently form the majority of sequences within marineenvironmental clone libraries (see López-García et al.,2001; Moon-van der Staay et al., 2001; Massana et al.,2004; Romari and Vaulot, 2004; Not et al., 2007). This hasled to a large increase in the number of MALV sequencesdeposited in GenBank over the last few years. In a previ-ous study, Groisillier and colleagues (2006) detected fivedistinct clades within MALV Group I, and 16 clades withinMALV Group II. However, in that study, many sequencescould not be clearly assigned to a specific clade, suggest-ing further discrete lineages might exist. Interestingly,some clades comprised sequences retrieved only fromvery specific habitats, such as anoxic environments ordeep sea hydrothermal vents, while other clades con-tained sequences obtained from widely varying habitats.
In the present work our aims were (i) to broaden repre-sentation of the different environments in which membersof the Alveolata are potentially encountered by construct-ing and sequencing 18S rRNA gene libraries; (ii) to clarifythe phylogeny of the Alveolata using recently publishedand the new 18S rRNA gene sequences for this group; (iii)to undertake a rigorous review of clade organizationwithin the above mentioned MALV groups I and II; (iv)to compare the genetic diversity of environmentalsequences derived from culture-independent PCRsurveys (targeting the 18S rRNA gene) to our presentknowledge of the taxonomy of Syndiniales; and (v) toextract general information on the preferred habitats anddistribution of specific members of the Syndiniales.
Results
Sample sites and environmental conditions
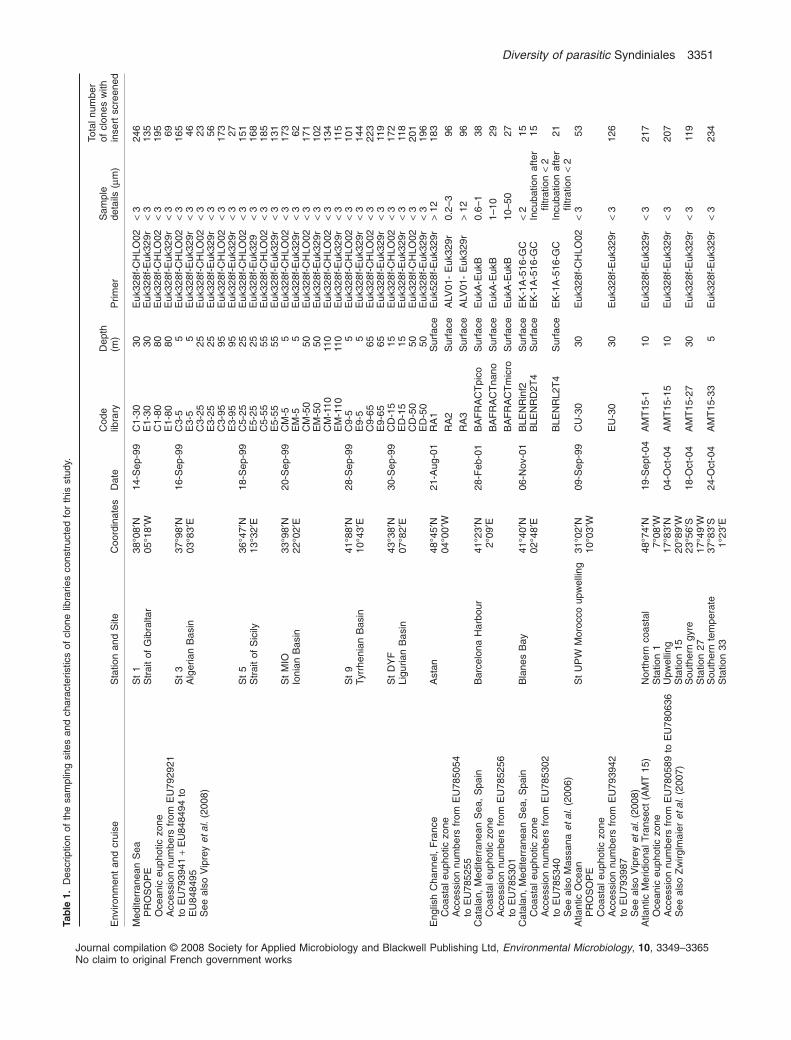
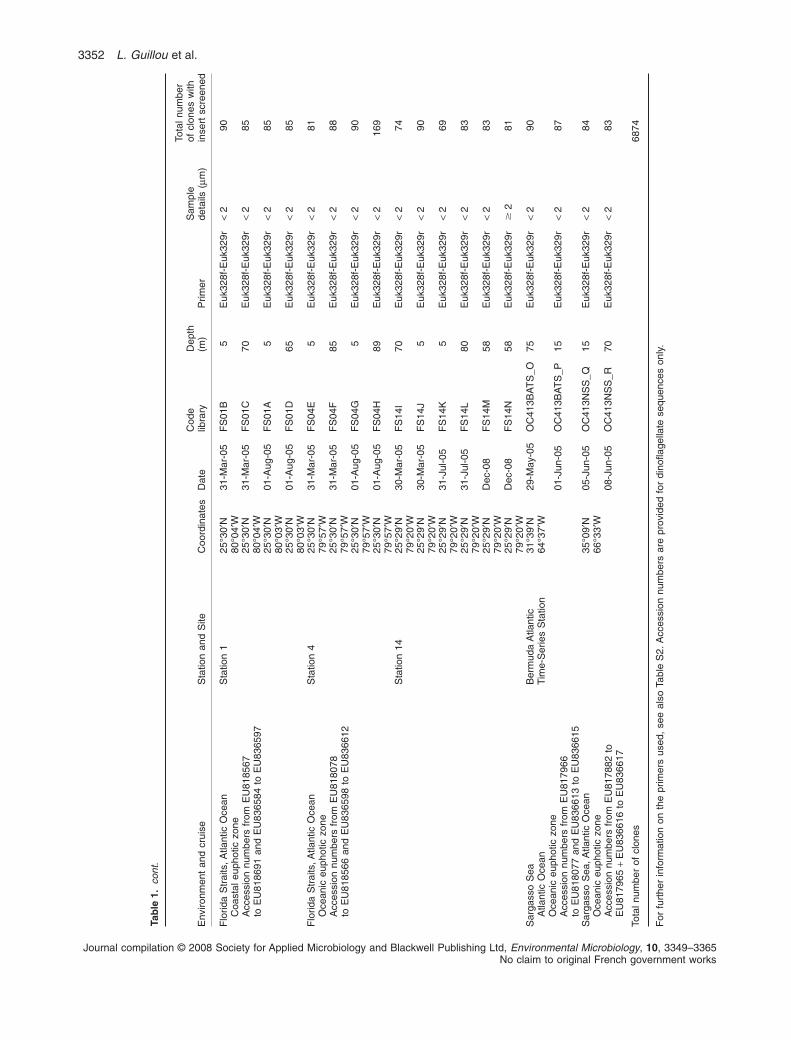
Most of the novel environmental sequences obtained inthis study were retrieved from coastal and oceanic watersin the Atlantic Ocean and Mediterranean Sea (Table 1).The collection sites varied in terms of season, depth andlevel of oligotrophy. For example, the Bermuda AtlanticTime-series Station (BATS) is relatively oligotrophic, andat the time of sampling was already strongly stratified. Incontrast, the northern Sargasso Sea Station, althoughrelatively close to BATS, was likely still influenced by deep
winter mixing that occurs in this area, bringing nutrients tosurface waters (see Cuvelier et al., 2008 for furtherdiscussion). The Florida Straits sites represent three dis-tinct water types. Station 1, on the western side of theFlorida Straits, was relatively coastal. Station 4 representsthe core of the Gulf Stream Current-forming waters, whichare highly oligotrophic, while Station 14 is also olig-otrophic but a shallow setting on the eastern side of theFlorida Straits (see Cuvelier et al., 2008). Other environ-mental sequences from the Atlantic Ocean were obtainedduring the Atlantic Meridional Transect (AMT 15, see alsoZwirglmaier et al., 2007), which extended from 48°N[south-west (SW) of the UK] to 40°S (SW of Cape Town,South Africa). Environmental sequences from the Medi-terranean Sea were collected in late summer along aMediterranean transect sampled during the PROSOPEcruise in 1999 (Garczarek et al., 2007), from high nutrientlevels in the Morocco upwelling to strong phosphoruslimitation in the eastern most basin. Although half of thegenetic libraries were built using a PCR approach biasedtowards green algae, dinoflagellate sequences wereretrieved in both data sets (the majority of them wereretrieved using general eukaryotic primers). We also useda PCR approach biased towards MALV Group II (usingthe specific primer ALV01) that we compared with the useof general eukaryotic primers from a coastal site (EnglishChannel, France). Most of genetic libraries were built onvery small size fractions (less than 2–3 mm), althoughsome were processed on larger size fractions and evenafter incubations (see genetic libraries from coastal sitesfrom the Mediterranean Sea). This heterogeneous dataset offered us a very large range of environmentalsequence origins. In total, 6874 new environmentalsequences were generated and screened for dinoflagel-late sequences.
Alveolate 18S rRNA gene sequence data set
The completed data set comprised 43655 eukaryotic 18SrRNA gene sequences obtained either from GenBank orfrom the environmental clone libraries described above.From marine environments, 351 environmental clonelibraries were analysed with major contribution from sites(Table 2) in the Atlantic (Díez et al., 2001a; Countwayet al., 2007; Not et al., 2007), Indian (Not et al., 2008),Arctic (Lovejoy et al., 2006; Stoeck et al., 2007) andPacific (Moon-van der Staay et al., 2001) Oceans, Medi-terranean Sea (Viprey et al., 2008), Antarctic (López-García et al., 2001), as well as several coastal sites(Massana et al., 2004; Romari and Vaulot, 2004; Medlinet al., 2006; Worden, 2006). These encompassed a rangeof marine habitats including the photic zone, sediments,hydrothermal vents and anoxic ecosystems (Table 2). Ter-restrial and continental ecosystems were also included
3350 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works
Tab
le1.
Des
crip
tion
ofth
esa
mpl
ing
site
san
dch
arac
teris
tics
ofcl
one
libra
ries
cons
truc
ted
for
this
stud
y.
Env
ironm
ent
and
crui
seS
tatio
nan
dS
iteC
oord
inat
esD
ate
Cod
elib
rary
Dep
th(m
)P
rimer
Sam
ple
deta
ils(m
m)
Tota
lnum
ber
ofcl
ones
with
inse
rtsc
reen
ed
Med
iterr
anea
nS
eaP
RO
SO
PE
Oce
anic
euph
otic
zone
Acc
essi
onnu
mbe
rsfr
omE
U79
2921
toE
U79
3941
+E
U84
8494
toE
U84
8495
See
also
Vip
rey
etal
.(2
008)
St
1S
trai
tof
Gib
ralta
r38
°08′
N05
°18′
W14
-Sep
-99
C1-
3030
Euk
328f
-CH
LO02
<3
246
E1-
3030
Euk
328f
-Euk
329r
<3
135
C1-
8080
Euk
328f
-CH
LO02
<3
195
E1-
8080
Euk
328f
-Euk
329r
<3
69S
t3
Alg
eria
nB
asin
37°9
8′N
03°8
3′E
16-S
ep-9
9C
3-5
5E
uk32
8f-C
HLO
02<
316
5E
3-5
5E
uk32
8f-E
uk32
9r<
346
C3-
2525
Euk
328f
-CH
LO02
<3
23E
3-25
25E
uk32
8f-E
uk32
9r<
356
C3-
9595
Euk
328f
-CH
LO02
<3
173
E3-
9595
Euk
328f
-Euk
329r
<3
27S
t5
Str
ait
ofS
icily
36°4
7′N
13°3
2′E
18-S
ep-9
9C
5-25
25E
uk32
8f-C
HLO
02<
315
1E
5-25
25E
uk32
8f-E
uk32
9<
316
8C
5-55
55E
uk32
8f-C
HLO
02<
318
5E
5-55
55E
uk32
8f-E
uk32
9r<
313
1S
tM
IOIo
nian
Bas
in33
°98′
N22
°02′
E20
-Sep
-99
CM
-55
Euk
328f
-CH
LO02
<3
173
EM
-55
Euk
328f
-Euk
329r
<3
62C
M-5
050
Euk
328f
-CH
LO02
<3
171
EM
-50
50E
uk32
8f-E
uk32
9r<
310
2C
M-1
1011
0E
uk32
8f-C
HLO
02<
313
4E
M-1
1011
0E
uk32
8f-E
uk32
9r<
311
5S
t9
Tyrr
heni
anB
asin
41°8
8′N
10°4
3′E
28-S
ep-9
9C
9-5
5E
uk32
8f-C
HLO
02<
310
1E
9-5
5E
uk32
8f-E
uk32
9r<
314
4C
9-65
65E
uk32
8f-C
HLO
02<
322
3E
9-65
65E
uk32
8f-E
uk32
9r<
311
9S
tD
YF
Ligu
rian
Bas
in43
°38′
N07
°82′
E30
-Sep
-99
CD
-15
15E
uk32
8f-C
HLO
02<
317
2E
D-1
515
Euk
328f
-Euk
329r
<3
118
CD
-50
50E
uk32
8f-C
HLO
02<
320
1E
D-5
050
Euk
328f
-Euk
329r
<3
196
Eng
lish
Cha
nnel
,F
ranc
eC
oast
aleu
phot
iczo
neA
cces
sion
num
bers
from
EU
7850
54to
EU
7852
55
Ast
an48
°45′
N04
°00′
W21
-Aug
-01
RA
1S
urfa
ceE
uk52
8f-E
uk32
9r>
1218
3
RA
2S
urfa
ceA
LV01
-E
uk32
9r0.
2–3
96
RA
3S
urfa
ceA
LV01
-E
uk32
9r>
1296
Cat
alan
,M
edite
rran
ean
Sea
,S
pain
Coa
stal
euph
otic
zone
Acc
essi
onnu
mbe
rsfr
omE
U78
5256
toE
U78
5301
Bar
celo
naH
arbo
ur41
°23′
N2°
09′E
28-F
eb-0
1B
AF
RA
CT
pico
Sur
face
Euk
A-E
ukB
0.6–
138
BA
FR
AC
Tna
noS
urfa
ceE
ukA
-Euk
B1–
1029
BA
FR
AC
Tm
icro
Sur
face
Euk
A-E
ukB
10–5
027
Cat
alan
,M
edite
rran
ean
Sea
,S
pain
Coa
stal
euph
otic
zone
Acc
essi
onnu
mbe
rsfr
omE
U78
5302
toE
U78
5340
See
also
Mas
sana
etal
.(2
006)
Bla
nes
Bay
41°4
0′N
02°4
8′E
06-N
ov-0
1B
LEN
Rin
f2S
urfa
ceE
K-1
A-5
16-G
C<
215
BLE
NR
D2T
4S
urfa
ceE
K-1
A-5
16-G
CIn
cuba
tion
afte
rfil
trat
ion
<2
15
BLE
NR
L2T
4S
urfa
ceE
K-1
A-5
16-G
CIn
cuba
tion
afte
rfil
trat
ion
<2
21
Atla
ntic
Oce
anP
RO
SO
PE
Coa
stal
euph
otic
zone
Acc
essi
onnu
mbe
rsfr
omE
U79
3942
toE
U79
3987
See
also
Vip
rey
etal
.(2
008)
St
UP
WM
oroc
coup
wel
ling
31°0
2 ′N
10°0
3′W
09-S
ep-9
9C
U-3
030
Euk
328f
-CH
LO02
<3
53
EU
-30
30E
uk32
8f-E
uk32
9r<
312
6
Atla
ntic
Mer
idio
nalT
rans
ect
(AM
T15
)O
cean
iceu
phot
iczo
neA
cces
sion
num
bers
from
EU
7805
89to
EU
7806
36S
eeal
soZ
wirg
lmai
eret
al.
(200
7)
Nor
ther
nco
asta
lS
tatio
n1
48°7
4′N
7°08
′W19
-Sep
t-04
AM
T15
-110
Euk
328f
-Euk
329r
<3
217
Upw
ellin
gS
tatio
n15
17°8
3′N
20°8
9′W
04-O
ct-0
4A
MT
15-1
510
Euk
328f
-Euk
329r
<3
207
Sou
ther
ngy
reS
tatio
n27
23°5
6′S
17°4
9′W
18-O
ct-0
4A
MT
15-2
730
Euk
328f
-Euk
329r
<3
119
Sou
ther
nte
mpe
rate
Sta
tion
3337
°83′
S1°
23′E
24-O
ct-0
4A
MT
15-3
35
Euk
328f
-Euk
329r
<3
234
Diversity of parasitic Syndiniales 3351
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works
Tab
le1.
cont
.
Env
ironm
ent
and
crui
seS
tatio
nan
dS
iteC
oord
inat
esD
ate
Cod
elib
rary
Dep
th(m
)P
rimer
Sam
ple
deta
ils(m
m)
Tota
lnum
ber
ofcl
ones
with
inse
rtsc
reen
ed
Flo
rida
Str
aits
,Atla
ntic
Oce
anC
oast
aleu
phot
iczo
neA
cces
sion
num
bers
from
EU
8185
67to
EU
8186
91an
dE
U83
6584
toE
U83
6597
Sta
tion
125
°30′
N80
°04′
W31
-Mar
-05
FS
01B
5E
uk32
8f-E
uk32
9r<
290
25°3
0′N
80°0
4′W
31-M
ar-0
5F
S01
C70
Euk
328f
-Euk
329r
<2
85
25°3
0′N
80°0
3′W
01-A
ug-0
5F
S01
A5
Euk
328f
-Euk
329r
<2
85
25°3
0′N
80°0
3′W
01-A
ug-0
5F
S01
D65
Euk
328f
-Euk
329r
<2
85
Flo
rida
Str
aits
,Atla
ntic
Oce
anO
cean
iceu
phot
iczo
neA
cces
sion
num
bers
from
EU
8180
78to
EU
8185
66an
dE
U83
6598
toE
U83
6612
Sta
tion
425
°30′
N79
°57′
W31
-Mar
-05
FS
04E
5E
uk32
8f-E
uk32
9r<
281
25°3
0′N
79°5
7′W
31-M
ar-0
5F
S04
F85
Euk
328f
-Euk
329r
<2
88
25°3
0′N
79°5
7′W
01-A
ug-0
5F
S04
G5
Euk
328f
-Euk
329r
<2
90
25°3
0′N
79°5
7′W
01-A
ug-0
5F
S04
H89
Euk
328f
-Euk
329r
<2
169
Sta
tion
1425
°29′
N79
°20′
W30
-Mar
-05
FS
14I
70E
uk32
8f-E
uk32
9r<
274
25°2
9′N
79°2
0′W
30-M
ar-0
5F
S14
J5
Euk
328f
-Euk
329r
<2
90
25°2
9′N
79°2
0′W
31-J
ul-0
5F
S14
K5
Euk
328f
-Euk
329r
<2
69
25°2
9′N
79°2
0′W
31-J
ul-0
5F
S14
L80
Euk
328f
-Euk
329r
<2
83
25°2
9′N
79°2
0′W
Dec
-08
FS
14M
58E
uk32
8f-E
uk32
9r<
283
25°2
9′N
79°2
0′W
Dec
-08
FS
14N
58E
uk32
8f-E
uk32
9r�
281
Sar
gass
oS
eaA
tlant
icO
cean
Oce
anic
euph
otic
zone
Acc
essi
onnu
mbe
rsfr
omE
U81
7966
toE
U81
8077
and
EU
8366
13to
EU
8366
15
Ber
mud
aA
tlant
icTi
me-
Ser
ies
Sta
tion
31°3
9′N
64°3
7′W
29-M
ay-0
5O
C41
3BA
TS
_O75
Euk
328f
-Euk
329r
<2
90
01-J
un-0
5O
C41
3BA
TS
_P15
Euk
328f
-Euk
329r
<2
87
Sar
gass
oS
ea,A
tlant
icO
cean
Oce
anic
euph
otic
zone
Acc
essi
onnu
mbe
rsfr
omE
U81
7882
toE
U81
7965
+E
U83
6616
toE
U83
6617
35°0
9′N
66°3
3′W
05-J
un-0
5O
C41
3NS
S_Q
15E
uk32
8f-E
uk32
9r<
284
08-J
un-0
5O
C41
3NS
S_R
70E
uk32
8f-E
uk32
9r<
283
Tota
lnum
ber
ofcl
ones
6874
For
furt
her
info
rmat
ion
onth
epr
imer
sus
ed,
see
also
Tabl
eS
2.A
cces
sion
num
bers
are
prov
ided
for
dino
flage
llate
sequ
ence
son
ly.
3352 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works
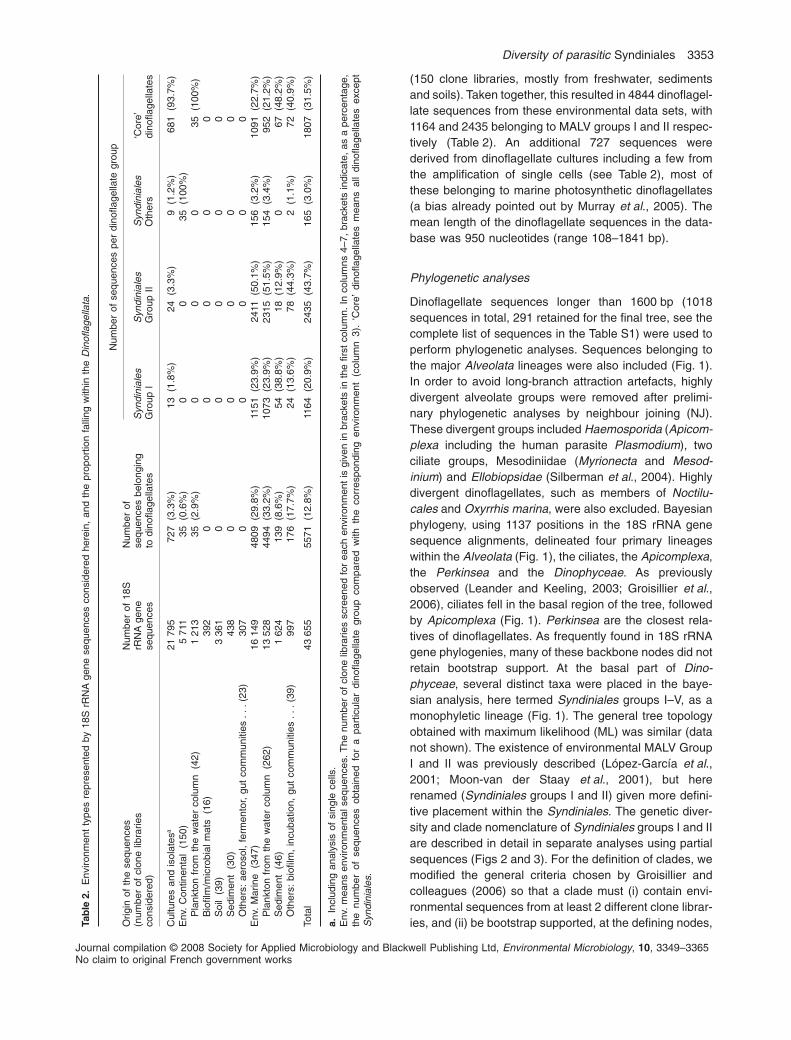
(150 clone libraries, mostly from freshwater, sedimentsand soils). Taken together, this resulted in 4844 dinoflagel-late sequences from these environmental data sets, with1164 and 2435 belonging to MALV groups I and II respec-tively (Table 2). An additional 727 sequences werederived from dinoflagellate cultures including a few fromthe amplification of single cells (see Table 2), most ofthese belonging to marine photosynthetic dinoflagellates(a bias already pointed out by Murray et al., 2005). Themean length of the dinoflagellate sequences in the data-base was 950 nucleotides (range 108–1841 bp).
Phylogenetic analyses
Dinoflagellate sequences longer than 1600 bp (1018sequences in total, 291 retained for the final tree, see thecomplete list of sequences in the Table S1) were used toperform phylogenetic analyses. Sequences belonging tothe major Alveolata lineages were also included (Fig. 1).In order to avoid long-branch attraction artefacts, highlydivergent alveolate groups were removed after prelimi-nary phylogenetic analyses by neighbour joining (NJ).These divergent groups included Haemosporida (Apicom-plexa including the human parasite Plasmodium), twociliate groups, Mesodiniidae (Myrionecta and Mesod-inium) and Ellobiopsidae (Silberman et al., 2004). Highlydivergent dinoflagellates, such as members of Noctilu-cales and Oxyrrhis marina, were also excluded. Bayesianphylogeny, using 1137 positions in the 18S rRNA genesequence alignments, delineated four primary lineageswithin the Alveolata (Fig. 1), the ciliates, the Apicomplexa,the Perkinsea and the Dinophyceae. As previouslyobserved (Leander and Keeling, 2003; Groisillier et al.,2006), ciliates fell in the basal region of the tree, followedby Apicomplexa (Fig. 1). Perkinsea are the closest rela-tives of dinoflagellates. As frequently found in 18S rRNAgene phylogenies, many of these backbone nodes did notretain bootstrap support. At the basal part of Dino-phyceae, several distinct taxa were placed in the baye-sian analysis, here termed Syndiniales groups I–V, as amonophyletic lineage (Fig. 1). The general tree topologyobtained with maximum likelihood (ML) was similar (datanot shown). The existence of environmental MALV GroupI and II was previously described (López-García et al.,2001; Moon-van der Staay et al., 2001), but hererenamed (Syndiniales groups I and II) given more defini-tive placement within the Syndiniales. The genetic diver-sity and clade nomenclature of Syndiniales groups I and IIare described in detail in separate analyses using partialsequences (Figs 2 and 3). For the definition of clades, wemodified the general criteria chosen by Groisillier andcolleagues (2006) so that a clade must (i) contain envi-ronmental sequences from at least 2 different clone librar-ies, and (ii) be bootstrap supported, at the defining nodes,Ta
ble
2.E
nviro
nmen
tty
pes
repr
esen
ted
by18
SrR
NA
gene
sequ
ence
sco
nsid
ered
here
in,
and
the
prop
ortio
nfa
lling
with
inth
eD
inofl
agel
lata
.
Orig
inof
the
sequ
ence
s(n
umbe
rof
clon
elib
rarie
sco
nsid
ered
)
Num
ber
of18
SrR
NA
gene
sequ
ence
s
Num
ber
ofse
quen
ces
belo
ngin
gto
dino
flage
llate
s
Num
ber
ofse
quen
ces
per
dino
flage
llate
grou
p
Syn
dini
ales
Gro
upI
Syn
dini
ales
Gro
upII
Syn
dini
ales
Oth
ers
‘Cor
e’di
nofla
gella
tes
Cul
ture
san
dis
olat
esa
2179
572
7(3
.3%
)13
(1.8
%)
24(3
.3%
)9
(1.2
%)
681
(93.
7%)
Env
.C
ontin
enta
l(1
50)
571
135
(0.6
%)
00
35(1
00%
)P
lank
ton
from
the
wat
erco
lum
n(4
2)1
213
35(2
.9%
)0
00
35(1
00%
)B
iofil
m/m
icro
bial
mat
s(1
6)39
20
00
00
Soi
l(3
9)3
361
00
00
0S
edim
ent
(30)
438
00
00
0O
ther
s:ae
roso
l,fe
rmen
tor,
gut
com
mun
ities
...(
23)
307
00
00
0E
nv.
Mar
ine
(347
)16
149
4809
(29.
8%)
1151
(23.
9%)
2411
(50.
1%)
156
(3.2
%)
1091
(22.
7%)
Pla
nkto
nfr
omth
ew
ater
colu
mn
(262
)13
528
4494
(33.
2%)
1073
(23.
9%)
2315
(51.
5%)
154
(3.4
%)
952
(21.
2%)
Sed
imen
t(4
6)1
624
139
(8.6
%)
54(3
8.8%
)18
(12.
9%)
067
(48.
2%)
Oth
ers:
biofi
lm,
incu
batio
n,gu
tco
mm
uniti
es..
.(39
)99
717
6(1
7.7%
)24
(13.
6%)
78(4
4.3%
)2
(1.1
%)
72(4
0.9%
)
Tota
l43
655
5571
(12.
8%)
1164
(20.
9%)
2435
(43.
7%)
165
(3.0
%)
1807
(31.
5%)
a.In
clud
ing
anal
ysis
ofsi
ngle
cells
.E
nv.
mea
nsen
viro
nmen
tals
eque
nces
.The
num
ber
ofcl
one
libra
ries
scre
ened
for
each
envi
ronm
ent
isgi
ven
inbr
acke
tsin
the
first
colu
mn.
Inco
lum
ns4–
7,br
acke
tsin
dica
te,
asa
perc
enta
ge,
the
num
ber
ofse
quen
ces
obta
ined
for
apa
rtic
ular
dino
flage
llate
grou
pco
mpa
red
with
the
corr
espo
ndin
gen
viro
nmen
t(c
olum
n3)
.‘C
ore’
dino
flage
llate
sm
eans
all
dino
flage
llate
sex
cept
Syn
dini
ales
.
Diversity of parasitic Syndiniales 3353
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

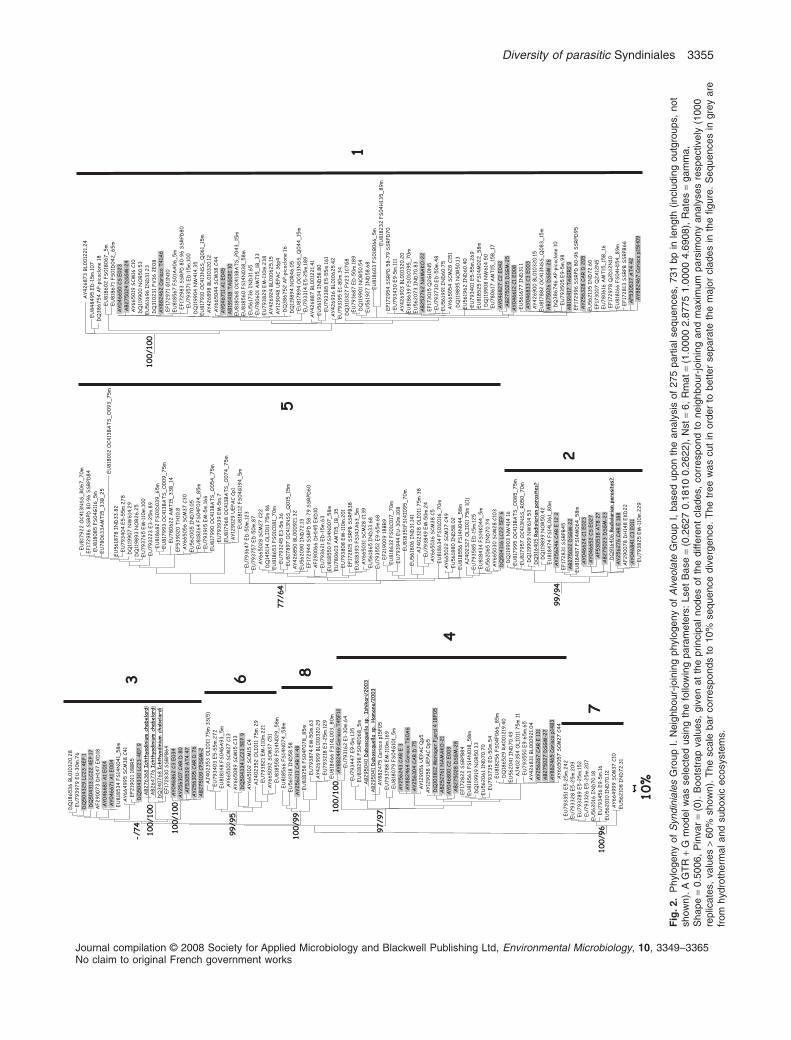
by values greater than 60% in NJ and maximum parsi-mony (MP) analysis. Some exceptions to these rules weremade as detailed below. The mean sequence identitywithin a clade was 87% for Syndiniales Group I (rangingfrom 76.6% for clade 5 to 91.9% for clade 7) and 93.5%for Syndiniales Group II (ranging from 80.8% for clade 8 to99.4% for clades 35 and 43, see also Fig. S1). Syndini-ales Group I contained eight different clades (Fig. 2).Clades 1–5 were described previously by Groisillier andcolleagues (2006), while three new clades emerged fromthis study. All of these eight clades are supported bybootstrap values > 60%, except for Clade 3, which is onlysupported by MP, bootstrap analyses (74%). Neverthe-less, the tree topology is identical with the two differentmethods (NJ and MP) and minimal sequence identitieswithin this clade are inside the range of other clades
belonging to this group (see Fig. S1). Ichthyodinium chab-erladi, a parasitoid of fish eggs, belongs to Group I Clade3, while Duboscquella spp., a parasitoid of tintinnides,belongs to Group I Clade 4 (Harada et al., 2007). Syndini-ales sequences retrieved by single-cell PCR on radiolar-ian isolates (Dolven et al., 2007) belong to Clade 1(DQ916408, though not in the tree because this sequencecontained several nucleotide ambiguities) and Clade 2(DQ916404–DQ916407 and DQ916410). Clades 5–8 areonly composed of environmental sequences. Somesequences within clades 1–4 (highlighted in grey in Fig. 2)were retrieved exclusively from suboxic and anoxicecosystems. Together, clades 1 and 5 were the mostcommonly retrieved from environmental clone libraries,representing � 75% of sequences belonging to Syndini-ales Group I.
DQ504327 LC22-5EP-44
DQ925237 CsH1AF421184 Hematodinium sp.EF065718 Hematodinium pereziEF065717 Hematodinium pereziEF172791 SSRPB76
DQ504356 LC22-5EP-37EU793192 E1-80m.27DQ146406 Syndinium sp.DQ146404 Syndinium turboDQ146403 Syndinium turboDQ146405 Syndinium turbo
AF286023 Hematodinium sp.
IV
EU562105 IND72.28EU818223 FS04H087_89mEU793790 EM-110m.171EU793574 ED-15m.111
EU562107 IND72.30EU780615 AMT15_15B_12EU818001 OC413BATS_O020_75m
EU793839 EM-50m.113EU818407 FS14JA31_5m
V
EU793465 E9-5m.27EU793466 E9-5m.29
EU561787 IND31.121DQ344732 TWS091
EU562079 IND72.02EU817912 OC413NSS_Q043_15m
EU818259 FS04F028_85mEU817911 OC413NSS_Q031_15m
EU793593 ED-15m.135EU793324 E5-25m.204DQ918792 ENVP36162.00253
AY664993 SCM15 C23EU818113 FS04G196_5mEU818541 FS14N090_58m
EU818542 FS14N072_58mEU561731 IND31.60EU818431 FS14K050_5mEU818080 FS04G102_5m
EU780604 AMT15_1B_36EF539059 MB01.33
EU818673 FS01D005_65mAY295467 RA000609.43EU818141 FS04GA80_5mAY426937 BL010625.44EU818430 FS14K031_5m
EU818672 FS01D055_65mEU793987 EU-30m.99
EU818014 OC413BATS_O095_75mEU793515 E9-65m.113
EU818655 FS01D056_65mDQ918274 ENVP21819.00021
EU818152 FS04H034_89mAJ402349 OLI11005
DQ918653 ENVP223.00266EF526769 NIF 4C5
DQ310226 FV23 1E1DQ918940 ENVP366.00147
DQ918434 ENVP21819.00364EU818349 FS14I039_70mEU793615 ED-15m.57
EU793164 E1-30m.67EU793978 EU-30m.74EU793475 E9-5m.53EU793818 EM-110m.215
EF539065 TH07.15DQ647514 CD8.10EU562048 IND70.56EU780607 AMT15_1B_40
EU793959 EU-30m.3DQ918980 ENVP366.00230
EF527026 SIF 3A10EF527085 SIF 4E8EF527091 SIF 4G12
2%
III
Otherdinoflagellates
II
Syndinium turboHematodinium spp.
III
IV
Perkinsea
Colpodellidae
CoccidiaPiroplasmida
Cryptosporidium spp.
Ciliates
Apicomplexa
IV
10%
AY665063 SCM38 C14AF133909 Parvilucifera
Und. Apicomplexa 1
Gregarinia
Colpodellidae Und. Apicomplexa 2
Outgroups
100/100
100/100
80/-
98/95
100/99
64/-
100/100
100/100
100/100100/100
99/82
100/99
Syndiniales
Dinophyceae
Fig. 1. Phylogeny of dinoflagellates using near-complete 18S rRNA gene sequences. Left: Bayesian phylogeny of alveolates based onanalysis of 291 near full-length 18S rRNA gene sequences. Five sequences of Bolidophyceae (stramenopiles) were used as an outgroup.Gblock retained 1137 positions for the phylogenetic analyses. Bootstrap values, given at the principal nodes of the tree, correspond toneighbour-joining and maximum parsimony analyses respectively (1000 replicates, values > 60% shown). For neighbour-joining bootstrapping,a GTR + G + I model was selected with the following parameters: Lset Base = (0.2757 0. 1787 0.2494), Nst = 6, Rmat = (1.0746 2.96001.2514 1.0820 4.7079), Rates = gamma, Shape = 0.7091, Pinvar = 0.2330. The scale bar corresponds to 10% sequence divergence. Inset (atthe right): Details of groups III to VI, based upon analysis of partial (500 bp) sequences (59 sequences including outgroups). The scale barcorresponds to 2% sequence divergence. The two sequences in grey belonging to Group IV are from hydrothermal vents.
3354 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works
DQ
186
53
6 B
L0
103
20
.28
EU
79
39
79
EU
-30
m.7
6D
Q5
04
32
9 L
C2
3 4
EP
1
DQ
50
43
13 L
C2
2 4
EP
17
AF
29
00
73
DH
147
EK
D18
AY
04
66
81
A1
E0
14
AY
04
66
71
A1
E0
04
EU
818
53
4F
S14
N0
41_
58
m
AY
66
49
95
SC
M3
8 C
41
EF
219
011
18
BR
5
DQ
50
43
30
LC
23
-4E
P 9
AB
27
63
68
Ich
thyo
dinium
chab
elar
diA
B2
64
77
6 Ich
thyo
dinium
chab
elar
diD
Q3
40
76
8 Ich
thyo
dinium
chab
elar
diE
F17
28
30
SS
RPB
64
AY
04
68
62
C3
E0
34
AY
25
63
07
CA
R D
80
AF
53
05
39
AT
4 4
7
AY
25
63
05
CA
R D
76
AB
27
510
9 C
PSG
M-2
AJ
40
23
53
OLI0
11 7
5m
33
(5)
EU
79
34
03
E5
-55
m.2
72
EU
818
148
FS
04
GA
93
_5
m
AY
66
510
0 S
CM
27
C13
AY
66
50
89
SC
M15
C3
3
DQ
50
43
34
LC
23
5E
P 9
AY
66
510
2 S
CM
15 C
4A
J4
02
35
2 O
LI0
11 7
5m
29
EU
79
38
21
EM
-110
m.2
21
AY
66
50
92
SC
M3
7 C
51
EU
818
55
8F
S14
N0
29
_5
8m
EU
818
56
6F
S14
N0
74
_5
8m
EU
56
1918
IN
D5
8.5
8
AY
25
62
13 C
AR
H 4
8
EU
818
25
8F
S0
4F
07
8_
85
m
EU
79
38
74
EM
-50
m.6
3
AY
42
69
09
BL0
103
20
.29
EU
79
32
18E
3-2
5m
.12
9
EU
818
46
6 F
S14
L0
03
_8
0m
EU
79
37
88
EM
-110
m.1
69
EU
818
07
9F
S0
4G
101_
5m
AY
25
62
43
CA
R E
3
AY
88
24
64
Car
iaco
T6
0A
6
AY
25
63
04
CA
R D
75
AY
129
05
6 U
EPA
C Q
p5
AY
129
05
8 U
EPA
C D
p5
DQ
103
812
Mar
iager
Fjo
rd 4
18
F0
5
AB
25
27
61
NA
MA
KO
-21
AY
04
68
00
C2
E0
09
AB
27
50
28
DS
GM
-28
EF
172
82
2 S
SR
PB8
4
EU
818
56
3 F
S14
N0
18_
58
m
DQ
119
919
NO
R5
0.3
3
EU
56
20
61
IN
D7
0.7
0
EU
79
37
35
ED
-50
m.5
4E
U8
182
56
FS
04
F0
66
_8
5m
DQ
186
52
9 R
A0
012
19.4
0
EU
56
20
41
IN
D7
0.4
9
AJ
40
23
54
OLI0
11 7
5m
11
EU
79
35
50
E9
-65
m.6
5A
Y4
26
83
1 B
L0
00
92
1.0
4
AY
25
62
67
CA
R E
13
1
AB
27
50
27
DS
GM
-27
AY
88
24
65
Car
iaco
p14
B3
AY
66
50
57
SC
M2
7 C
44
AY
88
24
49
Car
iaco
T4
5F
12
EU
79
316
2 E
1-3
0m
.64
EU
79
34
47
E9
-5m
.13
5E
U8
182
98
FS
04
E0
68
_5
m
AB
29
50
40
Dub
oscq
uella
sp.
Ishika
ri/2
003
AB
29
50
41 Dub
oscq
uella
sp.
Ham
ana/
2003
AY
88
24
75
Car
iaco
p15
F0
5
EU
817
92
2O
C4
13N
SS
_R
06
7_
70
m
EF
172
98
6 S
SR
PD 8
0-9
6 S
SR
PD8
4E
U8
180
85
FS
04
G11
6_
5m
EU
78
06
33
AM
T15
_3
3B
_2
5
EU
818
012
OC
413
BA
TS
_O
09
3_
75
m
EU
56
187
8 I
ND
33
.82
EU
79
34
04
E5
-55
m.2
78
DQ
119
90
7 N
W6
14.2
9
DQ
119
89
3 N
OR
26
.25
EU
79
37
65
EM
-110
m.1
00
EU
79
32
23
E3
-25
m.8
9
EU
818
68
2 F
S0
1D0
39
_6
5m
EU
817
99
3 O
C4
13B
AT
S_
O0
09
_7
5m
EU
78
06
22
AM
T15
_3
3B
_14
EF
53
90
20
TH
10.8
AY
66
50
56
SC
M3
7 C
30
EU
56
20
05
IN
D7
0.0
5E
U8
182
64
FS
04
F0
24
_8
5m
EU
79
39
15 E
M-5
m.1
66
EU
817
99
0 O
C4
13B
AT
S_
O0
54
_7
5m
EU
79
39
39
EM
-5m
.7
EU
817
96
8 O
C4
13B
AT
S_
O0
74
_7
5m
AY
129
02
9 U
EPA
C C
p1
EU
818
112
FS
04
G19
4_
5m
EU
79
36
47
ED
-50
m.1
23
EU
79
37
57
ED
-50
m.8
7
AY
66
50
28
SC
M2
7 C
22
DQ
145
114
OLI0
11 7
5m
86
EU
818
65
3 F
S0
1C0
81_
70
m
EU
79
32
45
E3
-5m
.36
EU
817
89
7 O
C4
13N
SS
_Q
015
_15
m
AY
42
68
50
BL0
00
92
1.3
2
EU
56
20
90
IN
D7
2.1
3
EF
172
94
4 S
SR
PD 5
8-7
9 S
SR
PD6
0
AF
29
00
66
DH
145
EK
D10
EU
79
36
21
ED
-15
m.6
3
EU
818
55
0 F
S14
N0
07
_5
8m
EU
78
06
03
AM
T15
_1B
_3
5
EU
79
38
08
EM
-110
m.2
01
EF
172
815
SS
RPB
SS
RPB
38
EU
818
39
3 F
S14
JA
63
_5
m
AY
66
50
31
SC
M2
8 C
139
EU
56
186
5 I
ND
33
.68
EU
79
35
52
E9
-65
m.6
8
EF
219
00
9 1
8B
R9
EU
818
63
2 F
S0
1C0
17_
70
m
EU
79
39
44
EU
-30
m.1
08
EU
818
35
1FS
14I0
95
_7
0m
EU
56
180
6 I
ND
31.
141
AJ
40
23
28
OLI0
11 7
5m
38
EU
79
38
49
EM
-50
m.2
4
AY
66
50
36
SC
M3
8 C
5E
U8
186
34
FS
01C
016
_7
0m
AY
66
50
22
SC
M3
7 C
49
EU
56
188
0 I
ND
58
.02
EU
818
55
6 F
S14
N0
44
_5
8m
AJ
40
23
27
OLI0
11 7
5m
1(1
)E
U7
93
58
5 E
D-1
5m
.12
5
EU
818
43
4 F
S14
K0
44
_5
m
EU
56
20
65
IN
D7
0.7
4
AY
66
50
30
SC
M2
8 C
103
DQ
50
43
16 L
C2
2 5
EP
6D
Q11
99
03
NW
414
16
EU
817
99
5 O
C4
13B
AT
S_
O0
85
_7
5m
EU
817
95
7 O
C4
13N
SS
_R
05
0_
70
m
DQ
119
90
9 N
W6
14 5
3
DQ
916
40
5 R
adiolarian
para
site
s?D
Q11
98
99
NO
R5
0.4
2
EU
818
47
4 F
S14
L0
61_
80
m
AY
25
62
46
CA
R E
12
2
EF
172
83
1 S
SR
PB4
5
AB
27
50
22
DS
GM
-22
EU
818
48
7 F
S14
M0
64
_5
8m
AY
04
66
24
C1
E0
23
AY
04
66
52
CS
E0
27
AF
53
05
38
AT
8 2
7
AB
27
50
23
DS
GM
-23
DQ
916
40
6 R
adiolarian
para
site
s?A
Y2
56
27
6 C
AR
E 1
88
AF
29
00
78
DH
148
EK
D2
2
AY
04
68
41
C3
E0
11
EU
79
38
25
EM
-110
m.2
29
AY
42
68
73
BL0
012
21.
24
EU
84
84
95
ED
-15
m.1
07
DQ
38
67
54
AP-p
icoc
lone
18
EU
818
60
2F
S0
1B0
67
_5
m
EU
818
67
1 F
S0
1D0
42
_6
5m
AY
04
66
50
CS
E0
23
AB
27
50
24
DS
GM
-24
AY
66
50
19 S
CM
16 C
10
DQ
119
90
0 N
OR
50
.53
EU
56
169
6 I
ND
31.
23
DQ
310
23
1 F
V3
6 2
E0
8
AY
88
24
62
Car
iaco
T4
1A6
EF
172
97
1 G
11N
10
EU
818
56
7 F
S0
1AA
06
_5
m
EF
172
98
3 S
SR
PD 8
0-9
6 S
SR
PD8
0
EU
79
35
71
ED
-15
m.1
00
DQ
119
90
4 N
W4
14.3
5E
U8
179
10 O
C4
13N
SS
_Q
06
1_15
m
AY
42
68
98
BL0
103
20
.12
AY
66
50
44
SC
M3
8 C
44
AY
04
67
11 A
1 E
04
5
AB
1914
18 T
AG
IR
I 1
0E
U8
180
68
OC
413
BA
TS
_P0
43
_15
m
EU
818
56
0 F
S14
N0
38
_5
8m
EU
56
173
6 I
ND
31.
65
EU
78
06
01
AM
T15
_1B
_3
3
EU
79
38
29
EM
-110
m.2
38
AY
42
69
24
BL0
106
25
.15
AY
129
04
8 U
EPA
C 3
6p4
DQ
38
67
52
AP-p
icoc
lone
16
DQ
119
89
4 N
OR
46
.05
EU
817
89
4 O
C4
13N
SS
_Q
04
4_
15m
EU
79
33
14 E
5-2
5m
.18
9
AY
42
68
87
BL0
012
21.
41
EU
56
193
4 I
ND
58
.80
EU
79
33
85
E5
-55
m.1
61
AY
42
69
36
BL0
106
25
.42
EU
79
319
5 E
1-8
0m
.34
DQ
310
32
7 F
V2
3 1
C7
G8
EU
79
36
87
ED
-50
m.1
89
DQ
119
90
1 N
OR
50
.54
EU
56
192
7 I
ND
58
.68
EU
818
60
3 F
S0
1B0
66
_5
m
EU
818
23
2 F
S0
4H
135
_8
9m
EF
172
95
4 S
SR
PD 5
8-7
9 S
SR
PD7
0
EU
79
34
30
E9
-5m
.111
AY
42
69
03
BL0
103
20
.20
EU
818
63
9 F
S0
1C0
95
_7
0m
EU
56
20
73
IN
D7
0.8
3
AB
25
27
62
NA
MA
KO
-22
EF
173
015
Q2
G10
N5
EU
79
37
30
ED
-50
m.4
8E
U5
619
91
IN
D6
0.7
5
AY
66
50
54
SC
M2
8 C
151
DQ
119
89
5 N
OR
50
.13
EU
56
196
2 I
ND
60
.40
EU
79
34
01
E5
-55
m.2
69
EU
818
52
5 F
S14
M0
22
_5
8m
DQ
119
90
8 N
W6
14 5
0E
U7
80
617
AM
T15
_15
B_
17
AY
04
68
27
C2
E0
42
AB
27
50
25
DS
GM
-25
AY
04
66
12 C
1 E
00
8
EU
56
167
7 I
ND
31.
1
AY
04
68
33
C3
E0
03
AY
42
69
00
BL0
103
20
.15
EU
817
88
2 O
C4
13N
SS
_Q
08
3_
15m
AB
27
50
26
DS
GM
-26
DQ
38
67
46
AP-p
icoc
lone
10
EU
79
35
05
E9
-5m
.98
AB
1914
17 T
AG
IR
I 9
EF
172
99
6 S
SR
PD 8
0-9
6 S
SR
PD9
5
AY
25
63
28
CA
R D
20
5
EU
56
213
5 I
ND
72
.60
EF
173
00
7 Q
2A
12N
5E
U7
80
616
AM
T15
_15
B_
16
EF
172
97
8 Q
2D
12N
10
EU
818
166
FS
04
H15
4_
89
m
EF
172
813
SS
RPB
SS
RPB
66
AF
53
05
37
AT
4 4
2A
Y8
82
46
7 C
aria
cop15
H0
9
10%
3 6 8
4
EU
79
33
51
E5
-25
m.2
42
EU
79
33
28
E5
-25
m.2
09
EU
79
32
89
E5
-25
m.1
52
EU
79
33
26
E5
-25
m.2
07
EU
56
20
16 I
ND
70
.18
EU
79
34
56
E9
-5m
.16
EU
56
20
10 I
ND
70
.12
AY
66
49
99
SC
M3
7 C
10
EU
56
210
8 I
ND
72
.31
7
5
2
1
-/74 100/
100
100/
100
100/
96
99/9
4
100/
100
99/9
5 100/
100
97/9
7
100/
99
77/6
4
Fig
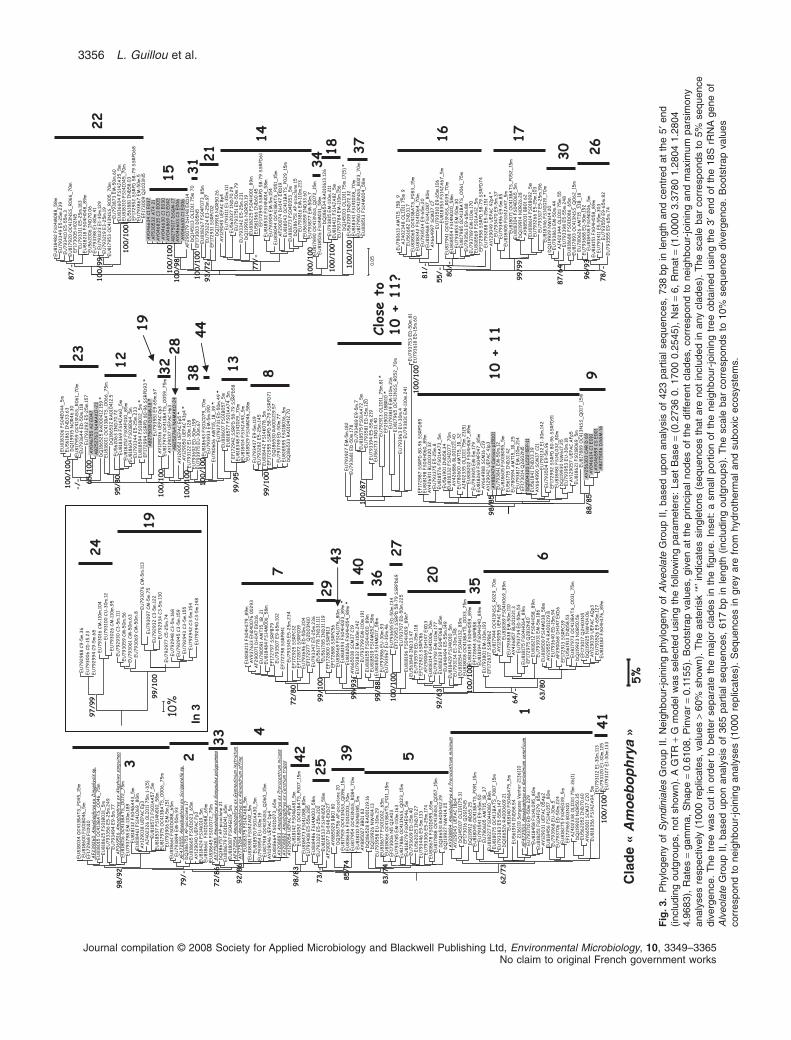
.2.
Phy
loge
nyof
Syn
dini
ales
Gro
upI.
Nei
ghbo
ur-jo
inin
gph
ylog
eny
ofA
lveo
late
Gro
upI,
base
dup
onth
ean
alys
isof
275
part
ials
eque
nces
,73
1bp
inle
ngth
(incl
udin
gou
tgro
ups,
not
show
n).A
GT
R+
Gm
odel
was
sele
cted
usin
gth
efo
llow
ing
para
met
ers:
Lset
Bas
e=
(0.2
627
0.18
100.
2622
),N
st=
6,R
mat
=(1
.000
02.
8775
1.00
004.
6908
),R
ates
=ga
mm
a,S
hape
=0.
5006
,P
inva
r=
(0).
Boo
tstr
apva
lues
,gi
ven
atth
epr
inci
paln
odes
ofth
edi
ffere
ntcl
ades
,co
rres
pond
tone
ighb
our-
join
ing
and
max
imum
pars
imon
yan
alys
esre
spec
tivel
y(1
000
repl
icat
es,
valu
es>
60%
show
n).
The
scal
eba
rco
rres
pond
sto
10%
sequ
ence
dive
rgen
ce.
The
tree
was
cut
inor
der
tobe
tter
sepa
rate
the
maj
orcl
ades
inth
efig
ure.
Seq
uenc
esin
grey
are
from
hydr
othe
rmal
and
subo
xic
ecos
yste
ms.
Diversity of parasitic Syndiniales 3355
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works
EU
79
36
92
ED
-50
m.1
99
E
U7
93
97
5E
U-3
0m
.67
100/
100
62/7
3
98/8
3
92/8
6
72/8
6
79/-
98/9
2
73/-
85/7
4
83/7
4
EU
818
03
4O
C4
13B
AT
S_
P0
45
_15
mE
U8
186
00
FS
01B
04
6_
5m
EF
172
96
8 F
11N
10A
F2
39
26
0 A
moe
bop
hry
aex D
inop
hys
issp
.E
U8
180
15 O
C4
13B
AT
S_
O0
46
_7
5m
EU
818
62
5 F
S0
1B0
31_
5m
EU
79
33
56
E5
-25
m.2
49
EU
79
34
94
E9
-5m
.77
AF
06
95
16 A
moe
bop
hry
aex A
kash
iwo
sang
uine
aE
U8
180
16 O
C4
13B
AT
S_
O0
72
_7
5m
EU
79
37
98
EM
-110
m.1
89
EU
818
132
FS
04
GA
43
_5
mE
U8
184
61
FS
14L0
69
_8
0m
AY
129
03
8 U
EPA
C E
p3
AJ
40
23
26
OLI0
11 7
5m
1-1
(5)
EU
818
57
3 F
S0
1AA
57
_5
mE
U8
185
37
FS
14N
010
_5
8m
EU
817
97
5 O
C4
13B
AT
S_
O0
06
_7
5m
EU
79
39
06
EM
-5m
.15
1E
U8
186
61
FS
01D
08
8_
65
mE
U7
93
89
4 E
M-5
0m
.93
AF
47
25
55
Am
oeb
ophry
aex S
crip
psie
llasp
. D
Q3
86
75
3 A
P-p
icoc
lone
17E
U8
186
65
FS
01D
02
3_
65
mA
Y12
90
40
UE
PAC
Fp3
EU
818
42
4 F
S14
K0
19_
5m
EU
818
66
1 F
S0
1D0
88
_6
5m
EU
818
64
5 F
S0
1C0
72
_7
0m
AY
77
52
85
Am
oeb
ophry
aex G
onya
ulax
poly
gram
ma
DQ
38
67
57
AP-p
icoc
lone
21
EU
818
67
9 F
S0
1D0
41_
65
mE
U8
180
87
FS
04
G11
9_
5m
AF
47
25
54
Am
oeb
ophry
aex G
ymno
din
ium
inst
riat
umA
Y7
75
28
4 A
moe
bop
hry
aex A
lexan
dri
umaf
fine
EU
818
37
5 F
S14
JA
68
_5
mE
U8
183
81
FS
14J
A0
2_
5m
EU
818
57
0 F
S0
1AA
30
_5
mE
U7
93
95
4 E
U-3
0m
.19
EU
817
918
OC
413
NS
S_
Q0
60
_15
mA
Y12
90
46
UE
PAC
Dp4
EU
818
66
4 F
S0
1D0
73
_6
5m
AY
20
88
93
Am
oeb
ophry
aex P
roro
cent
rum
mic
ans
AY
20
88
92
Am
oeb
ophry
aex C
erat
ium
trip
osA
Y12
90
54
UE
PAC
APp
5E
U8
1814
2 F
S0
4G
A8
1_5
mE
U8
180
52
OC
413
BA
TS
_P0
07
_15
mE
U8
184
73
FS
14L0
88
_8
0m
EU
79
34
08
E5
-55
m.2
85
EU
818
48
6 F
S14
M0
03
_5
8m
EU
79
33
22
E5
-25
m.2
02
EU
818
53
3 F
S14
N0
52
_5
8m
AF
29
00
77
DH
148
-EK
D2
7A
Y8
85
02
9 B
B0
1 8
0D
Q3
86
75
6 A
P-p
icoc
lone
20
EU
817
919
OC
413
NS
S_
Q0
96
_15
mE
U8
186
36
FS
01C
00
3_
70
mE
U8
179
54
OC
413
NS
S_
R0
84
_7
0m
EU
818
110
FS
04
G18
5_
5m
AY
88
50
27
BB
01
83
DQ
186
52
8 R
A0
012
19.1
6D
Q11
99
26
NW
414
.13
EU
79
39
58
EU
-30
m.2
9E
U8
186
77
FS
01D
012
_6
5m
EU
818
06
6 O
C4
13B
AT
S_
P0
11_
15m
EF
173
00
6 N
5 Q
2A
03
EU
79
37
85
EM
-110
m.1
63
EU
817
88
6 O
C4
13N
SS
_Q
02
3_
15m
EU
818
27
9 F
S0
4E
04
6_
5m
EU
79
36
09
ED
-15
m.4
0E
U5
62
02
5 I
ND
70
.27
EU
79
34
46
E9
-5m
.13
3E
U7
93
30
6 E
5-2
5m
.17
4E
U8
186
78
FS
01D
09
5_
65
mE
U8
178
88
OC
413
NS
S_
Q0
57
_15
mD
Q18
65
31
RA
010
412
.29
DQ
119
92
7 N
W4
14.2
5A
Y2
08
89
4 A
moe
bop
hry
aex P
roro
cent
rum
min
imum
AY
129
04
3 U
EPA
C 1
8p4
DQ
145
107
OLI0
117
5.3
1E
F17
30
16 Q
2G
12N
5D
Q11
99
12 M
D6
5 2
5E
U8
180
45
OC
413
BA
TS
_P0
91_
15m
EU
79
35
41
E9
-65
m.5
0E
U7
80
60
5 A
MT
15_
1B_
37
EU
818
22
7 F
S0
4H
08
0_
89
mE
U8
180
75
OC
413
BA
TS
_P0
87
_15
mE
U7
93
38
3 E
5-5
5m
.14
7D
Q18
65
26
RA
00
06
09
.21
EU
818
140
FS
04
GA
75
_5
mE
U5
619
14 I
ND
58
.54
213
611
0 S
argas
soV
ent
er
213
611
0A
F4
72
55
3 A
moe
bop
hry
aex K
arlo
din
ium
vene
ficu
mE
U7
93
710
ED
-50
m.2
29
EU
818
45
8 F
S14
L0
95
_8
0m
EU
818
25
1 F
S0
4F
07
9_
85
mA
Y12
90
31
UE
PAC
05
Ap2
EU
818
45
6 F
S14
L0
57
_8
0m
EU
79
39
68
EU
-30
m.5
EU
79
37
06
ED
-50
m.2
24
EU
818
27
5 F
S0
4E
03
9_
5m
EF
172
96
5 B
03
N10
AJ
40
23
38
OLI0
11 7
5m
26
(1)
DQ
119
92
0 N
OR
50
.35
EU
56
20
52
IN
D7
0.6
0A
Y6
65
00
1 S
CM
37
C2
0E
U8
183
56
FS
14J
A9
4_
5m
EU
79
311
2 E
1-3
0m
.113
EU
79
312
0 E
1-3
0m
.12
5E
U7
93
127
E1-
30
m.1
33
3 2
33
4
4225
39 5
1
41
EU
79
29
88
C9
-5m
.36
EU
79
30
06
CD
-15
.23
EU
79
29
94
C9
-5m
.68
EU
79
30
16 C
M-1
10m
.10
4E
U7
93
100
CU
-30
m.1
2E
U7
93
02
6 C
M-1
10m
.95
EU
79
29
30
C3
-5m
.111
E
U7
93
05
9 C
M-5
0m
.56
EU
79
30
61
CM
-50
m.6
3E
U7
93
06
9 C
M-5
0m
.8
EU
79
30
76
CM
-5m
.113
EU
79
30
97
CM
-5m
.75
EU
79
29
32
C3
-5m
.12
2E
U7
92
93
4 C
3-5
m.1
30
EU
79
29
77
C5
-25
m.7
4E
U7
92
94
8 C
3-5
m.1
68
EU
79
29
45
C3
-5m
.15
9E
U7
92
94
3C
3-5
m.1
55
EU
79
29
42
C3
-5m
.15
4E
U7
92
94
0 C
3-5
m.1
48
99/1
00
97/9
9
10%
24
19
In 3
í10
0/10
0
100/
100
99/1
00
99/9
5
95/8
0
-/-
EU
818
30
6F
S0
4E
04
4_
5m
EU
56
186
1IN
D3
3.6
3D
Q11
99
15 N
OR
46
.30
EU
817
95
9O
C4
13N
SS
_R
06
1_7
0m
EU
79
36
44
ED
-50
m.1
18E
U7
93
29
2E
5-2
5m
.15
7A
Y0
46
75
5 A
2 E
04
7A
B2
52
76
3 N
AM
AK
O-2
3D
Q18
65
25
RA
00
04
12.1
59
*E
U8
180
11O
C4
13B
AT
S_
O0
66
_7
5m
DQ
186
53
2 R
A0
104
12.5
EU
56
20
63
IN
D7
0.7
2E
U8
183
69
FS
14J
A6
0_
5m
EU
818
25
2F
S0
4F
010
_8
5m
EU
818
60
4 F
S0
1B0
64
_5
mE
U7
93
34
4 E
5-2
5m
.23
3E
U8
185
94
FS
01B
012
_5
m*
EF
172
99
4 S
SR
PD 8
0-9
6 S
SR
PD9
3 *
AB
27
50
18 D
SG
M-1
8 *
EU
79
35
51
E9
-65
m.6
7A
Y12
90
36
UE
PAC
Cp3
EU
817
97
4 O
C4
13B
AT
S_
O0
59
_7
5m
AY
129
03
7 U
EPA
C H
p3
AB
25
27
64
NA
MA
KO
-24
AY
129
05
2 U
EPA
C E
p5
*A
Y12
90
50
UE
PAC
43
p4
*E
U7
93
122
E1-
30
m.1
28
EU
818
64
3 F
S0
1C0
79
_7
0m
EU
79
39
33
EM
-5m
.19
0E
U7
80
60
6 A
MT
15_
1B_
39
*E
U7
93
73
1E
D-5
0m
.49
*E
U8
186
07
FS
01B
05
7_
5m
EU
818
57
7F
S0
1AA
74
_5
mE
F17
29
42
SS
RPD
58
-79
SS
RPD
58
EU
818
34
8F
S14
I0
29
_7
0m
EU
818
414
FS
14J
A4
3_
5m
EU
818
52
9F
S14
M0
46
_5
8m
EU
79
34
37
E9
-5m
.119
EU
79
32
82
E5
-25
m.1
45
EU
818
44
2F
S14
K0
78
_5
mE
F17
29
55
SS
RPD
58
-79
SS
RPD
71
DQ
186
53
0 R
A0
012
19.5
7E
U7
93
74
9E
D-5
0m
.77
EU
818
59
5F
S0
1B0
36
_5
mD
Q18
65
33
RA
010
412
.70
100/
100
23 1219
3228
3844
13 8
AB
27
50
16 D
SG
M-1
6
100/
100
88/8
5
EU
79
39
27
EM
-5m
.18
2E
U7
93
68
1E
D-5
0m
.17
8E
U7
93
48
9E
9-5
m.7
EU
818
57
5F
S0
1AA
72
_5
mE
U7
93
58
1E
D-1
5m
.12
0E
U7
93
39
4E
5-5
5m
.219
EU
56
1713
IN
D3
1.4
0E
U7
92
913
OLI0
11_
75
m.8
1 *
EF
219
010
18
BR
10 *
EU
79
38
19E
M-1
10m
.216
EU
817
96
5O
C4
13N
SS
_R
05
2_
70
mE
U7
93
96
3E
U-3
0m
.4E
U7
93
83
1E
M-1
10m
.24
1E
U7
93
75
3E
D-5
0m
.81
EU
79
36
18E
D-1
5m
.60
EF
172
98
7 S
SR
PD 8
0-9
6 S
SR
PD8
5E
U8
1819
8F
S0
4H
06
0_
89
mA
Y4
26
911
BL0
103
20
.32
EU
79
33
75
E5
-25
m.8
EU
818
57
6F
S0
1AA
73
_5
mE
U5
619
01
IN
D5
8.3
4E
U8
183
32
FS
14I0
77
_7
0m
AY
42
68
86
BL0
012
21.
40
EU
78
06
00
AM
T15
_1B
_3
2A
J4
02
33
5 O
LI0
11 7
5m
23
(5)
EU
818
20
2F
S0
4H
06
7_
89
mE
U7
93
92
5E
M-5
m.1
79
EU
818
26
9F
S0
4F
04
7_
85
mA
Y6
64
98
8 S
CM
16 C
19A
Y12
90
42
UE
PAC
41p
3A
Y8
82
47
4 C
aria
cop15
D0
1E
U7
93
84
1E
M-5
0m
.12
EU
818
60
5F
S0
1B0
55
_5
mE
U5
617
15IN
D3
1.4
2E
U7
80
59
4 A
MT
15_
1B_
25
EU
79
38
17E
M-1
10m
.214
EF
173
014
Q2
G0
2N
5D
Q5
04
32
0 L
C2
2-5
EP-1
1E
U5
618
41
IN
D3
3.3
9A
Y6
64
99
0 S
CM
37
C2
1E
U7
93
132
E1-
30
m.1
42
EU
79
35
04
E9
-5m
.96
EF
172
99
2 S
SR
PD 8
0-9
6 S
SR
PD9
1E
U8
184
81
FS
14L0
32
_8
0m
DQ
119
93
0 N
W6
17.2
5E
U7
93
95
3E
U-3
0m
.18
AY
129
05
7 U
EPA
C A
Fp5
EU
818
62
1 F
S0
1B0
69
_5
mE
U8
178
89
OC
413
NS
S_
Q0
17_
15m
AY
25
63
03
CA
R D
69
AY
04
66
14 C
1 E
010
AY
04
65
99
CS
E0
01
10 +
11
9
Clos
e to
10
+ 1
1?
EU
818
213
FS
04
H17
9_
89
mA
Y9
37
63
3 E
NI4
00
76
00
29
3A
F2
90
07
1 D
H14
7 E
KD
16E
U7
80
59
1 A
MT
15_
1B_
21
EU
818
48
4F
S14
M0
06
_5
8m
EF
172
79
7 S
SR
PB7
9E
U7
93
50
7 E
9-6
5m
.10
2E
F17
27
98
SS
RPB
41
EU
79
33
45
E5
-25
m.2
34
EF
172
92
5 S
SR
PC3
2E
F17
28
72
SS
RPC
09
EU
79
36
96
ED
-50
m.2
04
EF
172
97
7
Q2
D0
3N
10E
U7
93
34
7 E
5-2
5m
.23
6E
U5
617
78
IN
D3
1.11
1E
U5
617
85
IN
D3
1.11
9E
F17
28
00
SS
RPB
13E
F17
28
88
SS
RPC
56
EU
818
46
9 F
S14
L0
76
_8
0m
EU
818
193
FS
04
H0
23
_8
9m
*E
U8
182
06
FS
04
H0
54
_8
9m
*A
Y6
65
03
8 S
CM
37
C19
EU
79
38
32
EM
-110
m.2
42
EU
79
37
99
EM
-110
m.1
91
EU
818
155
FS
04
H0
10_
89
mE
U8
185
19 F
S14
M0
52
_5
8m
EU
818
52
0 F
S14
M0
17_
58
mE
U7
93
98
6 E
U-3
0m
.93
EU
79
39
67
EU
-30
m.4
6E
U7
93
65
5 E
D-5
0m
.13
4E
F17
29
53
SS
RPD
58
-79
SS
RPD
69
EU
79
37
07
ED
-50
m.2
25
EU
818
196
FS
04
H0
19_
89
m*
EU
56
184
8 I
ND
33
.47
EU
79
35
79
ED
-15
m.1
18E
F17
29
23
SS
RPC
86
AF
29
00
69
DH
147
EK
D3
EU
818
314
FS
14I0
06
_7
0m
EU
79
37
92
EM
-110
m.1
77
EU
818
23
9 F
S0
4F
06
8_
85
mE
U8
48
49
4 E
5-5
5m
.14
9E
U8
184
29
FS
14K
010
_5
mE
U7
93
134
E1-
30
m.1
48
EU
818
154
FS
04
H13
2_
89
mE
U8
180
18O
C4
13B
AT
S_
O0
33
_7
5m
EU
818
195
FS
04
H0
59
_8
9m
EF
172
84
4 S
SR
PB5
8E
U8
1819
4 F
S0
4H
04
0_
89
mE
U7
93
80
1 E
M-1
10m
.19
3E
F17
28
37
SS
RPB
62
EU
817
94
4 O
C4
13N
SS
_R
02
9_
70
mA
Y12
90
55
UE
PAC
Fp5
EU
818
189
FS
04
H0
69
_8
9m
EU
79
38
38
EM
-50
m.1
10A
Y4
26
85
7 B
L0
012
21.
3E
U7
93
73
4 E
D-5
0m
.53
EU
818
175
FS
04
H0
64
_8
9m
EF
172
97
5 Q
2B
12N
10E
U8
182
12 F
S0
4H
158
_8
9m
EU
79
39
31
EM
-5m
.18
6E
U8
185
00
FS
14M
03
8_
58
mA
Y2
95
57
4 R
A0
012
19.8
EU
79
36
40
ED
-15
m.9
4A
F2
90
06
8 D
H14
7 E
KD
6E
F17
29
13 S
SR
PC3
9E
U5
619
13IN
D5
8.5
3E
U8
179
73
OC
413
BA
TS
_O
03
1_7
5m
DQ
119
92
3 N
OR
50
.56
EF
173
017
Q2
H0
3N
5D
Q11
99
25
NW
414
.06
AY
129
03
9 U
EPA
C A
Qp3
EU
79
35
26
E9
-65
m.1
27
EU
818
182
FS
04
H17
6_
89
m
100/
100
100/
100
99/1
00
99/9
3
99/8
8
72/8
0
92/6
3
64/-
7
29
4043
203627
35
6
EU
78
06
13 A
MT
15_
15B
_9
AJ
40
23
48
OLI0
11 7
5m
9E
U5
616
82
IN
D3
1.7
EU
818
06
5O
C4
13B
AT
S_
P0
83
_15
mE
U8
183
42
FS
14I0
64
_7
0m
EU
79
35
34
E9
-65
m.2
0E
U8
184
68
FS
14L0
87
_8
0m
AY
66
49
97
SC
M3
7 C
7E
U7
93
510
E9
-65
m.1
06
EU
818
35
3F
S14
JA
67
_5
m
EU
79
32
88
E5
-25
m.1
51 *
EU
817
94
1O
C4
13N
SS
_R
03
9_
70
mE
U8
185
54
FS
14N
02
0_
58
mE
U7
93
85
3E
M-5
0m
.30
EU
817
98
5O
C4
13B
AT
S_
O0
61_
75
mE
U7
93
56
3E
9-6
5m
.88
EU
79
37
89
EM
-110
m.1
70
EU
818
56
9F
S0
1AA
12_
5m
EF
172
95
8 S
SR
PD 5
8-7
9 S
SR
PD7
4
AY
129
04
9 U
EPA
C 4
2p4
*E
U7
93
47
0E
9-5
m.3
7E
U7
93
49
6E
9-5
m.8
5E
U8
184
05
FS
14J
A4
7_
5m
EU
818
07
1O
C4
13B
AT
S_
P0
52
_15
mE
U8
183
04
FS
04
E0
65
_5
mD
Q11
99
14 N
OR
26
.28
AY
88
50
31
BB
01
62
EU
818
614
FS
01B
09
2_
5m
EU
79
32
66
E5
-25
m.1
03
EU
79
33
17E
5-2
5m
.19
6E
U8
183
46
FS
14I0
19_
70
mD
Q11
99
29
NW
614
.20
EU
79
38
63
EM
-50
m.4
4A
J4
02
34
4 O
LI0
11 7
5m
55
EU
79
33
98
E5
-55
m.2
48
EU
818
68
8F
S0
1D0
86
_6
5m
EU
817
89
3O
C4
13N
SS
_Q
06
2_
15m
EU
78
06
26
AM
T15
_3
3B
_18
EU
79
36
88
ED
-50
m.1
92
EU
818
43
8F
S14
K0
06
_5
mE
U8
1815
1F
S0
4H
05
8_
89
mE
U7
93
411
E5
-55
m.3
03
EU
79
35
59
E9
-65
m.8
2E
U7
93
55
5E
9-6
5m
.74
99/9
9
80/-
87/6
4
78/-
16 17
30 26
EU
818
48
2F
S14
M0
88
_5
8m
EU
79
33
49
E5
-25
m.2
39
EU
79
34
10E
5-5
5m
.3E
U8
179
20
OC
413
NS
S_
R0
66
_7
0m
EF
173
00
5 H
02
N5
EU
79
33
11E
5-2
5m
.18
3E
U8
182
18 F
S0
4H
04
8_
89
mE
U5
62
00
6IN
D7
0.0
6E
U7
93
198
E1-
80
m.4
1E
U7
93
123
E1-
30
m.1
29
EU
79
32
19E
3-2
5m
.19
EU
817
95
3O
C4
13N
SS
_R
00
5_
70
mE
U7
93
87
3E
M-5
0m
.60
EU
83
66
04
FS
14J
A3
5_
5m
EU
818
310
FS
14I0
45
_7
0m
EU
56
188
1IN
D5
8.0
3E
F17
29
52
SS
RPD
58
-79
SS
RPD
68
EU
79
38
67
EM
-50
m.5
EF
173
00
9 Q
2C
03
N5
AY
04
66
34
C1
E0
37
AF
53
05
32
AT
4 2
1A
Y0
46
83
5 C
3 E
00
5A
Y0
46
63
0 C
1 E
03
0A
Y0
46
86
9 C
3 E
04
2A
B2
75
017
DS
GM
-17
AY
04
66
03
CS
E0
06
AY
04
66
10 C
1 E
00
6A
F2
90
07
9 D
H14
8 E
KD
14D
Q14
511
3O
LI0
11 7
5m
70
EF
173
00
3 D
05
N5
EU
818
26
7 F
S0
4F
07
3_
85
mE
U7
93
22
4E
3-2
5m
.97
EF
172
93
1 S
SR
PC0
2D
Q11
99
18 N
OR
50
.26
AY
129
04
5 U
EPA
C B
p4
EU
79
311
1E
1-3
0m
.111
EU
56
20
27
IN
D7
0.2
9E
U7
93
75
1E
D-5
0m
.79
EU
79
33
42
E5
-25
m.2
31
DQ
119
916
NO
R5
0.1
9E
U8
182
25
FS
04
H0
01_
89
mE
U5
619
66
IN
D6
0.4
5E
F17
29
45
SS
RPD
58
-79
SS
RPD
61
EU
818
53
2F
S14
N0
14_
58
mE
U7
93
90
9E
M-5
m.1
54
EU
818
04
4O
C4
13B
AT
S_
P0
51_
15m
AF
29
00
74
DH
147
EK
D19
EU
818
04
3O
C4
13B
AT
S_
P0
29
_15
mE
U8
182
73
FS
04
E0
53
_5
mD
Q3
86
75
1 A
P-p
icoc
lone
15E
U7
93
816
EM
-110
m.2
13E
U5
618
59
IN
D3
3.6
0E
U7
93
87
7E
M-5
0m
.7E
U8
178
90
OC
413
NS
S_
Q0
72
_15
mE
U8
185
26
FS
14M
03
1_5
8m
DQ
186
53
4 R
A0
106
13.1
26
EU
79
38
15E
M-1
10m
.211
EU
818
413
FS
14J
A8
2_
5m
EU
79
37
84
EM
-110
m.1
56
DQ
145
103
OLI0
11 7
5m
17
(5) *
EU
56
210
9IN
D7
2.3
2E
U8
183
41
FS
14I0
28
_7
0m
EU
817
95
0O
C4
13N
SS
_R
02
3_
70
mE
U8
185
13F
S14
M0
04
_5
8m
100/
100
100/
100
100/
100
100/
100
100/
100
100/
98 77/-
93/7
2
87/-
0.0
5
22
15 3121
14
18 37
34
Cla
de
«A
mo
ebo
ph
rya
»5%
63/8
0
100/
99
81/-
55/-
98/8
5
100/
87
100/
100
65/1
00
100/
100
96/9
3
Fig
.3.
Phy
loge
nyof
Syn
dini
ales
Gro
upII.
Nei
ghbo
ur-jo
inin
gph
ylog
eny
ofA
lveo
late
Gro
upII,
base
dup
onan
alys
isof
423
part
ials
eque
nces
,73
8bp
inle
ngth
and
cent
red
atth
e5′
end
(incl
udin
gou
tgro
ups,
not
show
n).A
GT
R+
Gm
odel
was
sele
cted
usin
gth
efo
llow
ing
para
met
ers:
Lset
Bas
e=
(0.2
736
0.17
000.
2545
),N
st=
6,R
mat
=(1
.000
03.
3780
1.28
041.
2804
4.96
83),
Rat
es=
gam
ma,
Sha
pe=
0.61
08,
Pin
var
=0.
1155
).B
oots
trap
valu
es,
give
nat
the
prin
cipa
lnod
esof
the
diffe
rent
clad
es,
corr
espo
ndto
neig
hbou
r-jo
inin
gan
dm
axim
umpa
rsim
ony
anal
yses
resp
ectiv
ely
(100
0re
plic
ates
,va
lues
>60
%sh
own)
.T
heas
teris
k‘*
’ind
icat
essi
ngle
tons
(seq
uenc
esth
atar
eno
tin
clud
edin
any
clad
es).
The
scal
eba
rco
rres
pond
sto
5%se
quen
cedi
verg
ence
.T
hetr
eew
ascu
tin
orde
rto
bette
rse
para
teth
em
ajor
clad
esin
the
figur
e.In
set:
asm
allp
ortio
nof
the
neig
hbou
r-jo
inin
gtr
eeob
tain
edus
ing
the
3′en
dof
the
18S
rRN
Age
neof
Alv
eola
teG
roup
II,ba
sed
upon
anal
ysis
of36
5pa
rtia
lseq
uenc
es,
617
bpin
leng
th(in
clud
ing
outg
roup
s).
The
scal
eba
rco
rres
pond
sto
10%
sequ
ence
dive
rgen
ce.
Boo
tstr
apva
lues
corr
espo
ndto
neig
hbou
r-jo
inin
gan
alys
es(1
000
repl
icat
es).
Seq
uenc
esin
grey
are
from
hydr
othe
rmal
and
subo
xic
ecos
yste
ms.
3356 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

By comparison, Syndiniales Group II is genetically morediverse than Group I. In addition to clades 1–16 (seeGroisillier et al., 2006), 28 new clades emerged from thepresent study (clades 17–44, Fig. 3). Some of theseclades are not supported by bootstrap analyses obtainedwith one of the two phylogenetic methods tested (clades2, 6, 10+11, 14, 16, 23, 25 and 26), but in such cases thetree topologies are identical between the two methods.Clade 10 and Clade 11 described by Groisillier and col-leagues (2006) are now merged into a single clade(herein named Clade 10+11), which includes cloneBL10320.32 (a singleton in the work of Groisillier et al.,2006). However, Clade 10+11 is not supported by boot-strap analyses. Numerous environmental sequences dis-playing long branches (labelled ‘close to 10+11?’ in Fig. 3)also appear to be closely related to Clade 10+11.
Only a single genus within Syndiniales Group II hasbeen formally described, the Amoebophrya. AllAmoebophrya sequences obtained from cultures andsingle-cell analyses formed a monophyletic group as rec-ognized by Groisillier and colleagues (2006). This mono-phyletic group now also includes clades 1–5 as well asclades 25, 33, 39, 41 and 42 (Fig. 3). Recently, Kim andcolleagues (2008) recognized nine different subgroupswithin Amoebophrya. Subgroup 1 (from the analysis ofKim and colleagues) corresponds primarily with our Clade1 (except sequence AF290077 which belongs to the novelClade 25) while subgroups 3, 4, 5 and 6 correspond toclades 4, 3, 5 and 2 respectively. The recognition of sub-group 2 (identified in Kim et al., 2008) is probably skewedby inclusion of a sequence we identified to be a chimera(DQ186527). Thus, sequence AY260468, obtained fromthe direct amplification of Ceratium tripos infected withAmoebophrya, likely remains a singleton. SequenceAY295690 (subgroup 8) is also a probable chimera,whereas sequences included within subgroups 7 and 9belong to Clade 33 in the present study. SequenceDQ916402, obtained from the direct amplification of a cellof spumellarida Hexacontium gigantheum (Radiolaria), isclosely related to Clade 6. This sequence was removedfrom our global analyses due to the high number of nucle-otide ambiguities it contained. Clades 15 and 9 are pri-marily composed of environmental sequences fromanoxic or suboxic ecosystems (highlighted in grey inFig. 3). Both clades also include sequences retrievedfrom deep-sea methane cold seeps (DSGM-16 and 17,Takishita et al., 2007) and deep oceanic waters (Count-way et al., 2007), but these sequences were not includedin this analysis due to their short length. Clades originallycontaining only sequences from coastal systems (i.e.clades 2, 4, 5, 8, 12 and 13; see Groisillier et al., 2006) areshown here to also include clones from other oceanicregions, including oligotrophic waters. This highlights theimportance of gaining sequence representation from a
broader array of geographical locations. Finally, in termsof the number of sequences, the most commonly foundclades in rank order were clades 10+11, 7, 6, 1, 19, 3, 22and 16.
Other Syndiniales groups (III–V) also emerged from thepresent study (detailed in Fig. 1). Syndiniales Group IIIcontains 71 environmental sequences, including cloneOLI11005 (AJ402349) that was previously placed outsideGroup II (see Groisillier et al. 2006), as well as numerousenvironmental sequences retrieved from various oceanicsurface waters (Mediterranean Sea, Indian Ocean, Sar-gasso Sea), coastal waters [Southern Taiwan Strait,Blanes Bay (Spain) and the English Channel] and from asupersulfidic anoxic fjord (DQ310226). Syndiniales GroupIV contains the genera Syndinium and Hematodinium,five closely related environmental sequences, a sequenceretrieved from the Mediterranean Sea, E1–80m.27,closely related to Syndinium turbo and a clade allied withthe genus Hematodinium and composed of sequencesfrom the Sargasso Sea (EF172791 and DQ918583)and from deep sea hydrothermal vents (DQ504327 andDQ504356). Syndiniales Group V contains 67 environ-mental sequences, all collected within the euphotic zone,but includes clones retrieved from very different oceanicecosystems (Indian Ocean, Atlantic Ocean, Mediterra-nean Sea and Sargasso Sea). Sequence BL000921.23from Blanes also belongs to Syndiniales Group V (notincluded here due to sequence length dissimilarities).Finally, two partial environmental sequences could not beassigned to any specific group: EU793524 (E9–65m.123)and EU818559 (F14DEC+), but phylogenetic analysisplaced them closely related, although separated fromthese Syndiniales groups.
Ecological distributions
Dinoflagellate sequences represent less than 1% ofenvironmental sequences from continental ecosystems(aquatic and terrestrial), whereas they represent close to30% of the sequences obtained from marine ecosystems.With respect to aquatic environments (both marine andfreshwater), dinoflagellates sequences as a whole wereobtained primarily from the plankton, as opposed to otherenvironments such as sediments (Table 2). Sequencesbelonging to Syndiniales groups I and II were absent insamples collected to date from continental ecosystems,however, they represent the largest portion of dinoflagel-late sequences from marine systems. Syndiniales GroupII alone represented about half of all environmentaldinoflagellate sequences (Table 2).
We divided marine ecosystems included in oursequence database into various categories: (i) anoxic andsuboxic ecosystems, (ii) ecosystems with hydrothermalactivities, (iii) sediments, or (iv) the water column
Diversity of parasitic Syndiniales 3357
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works
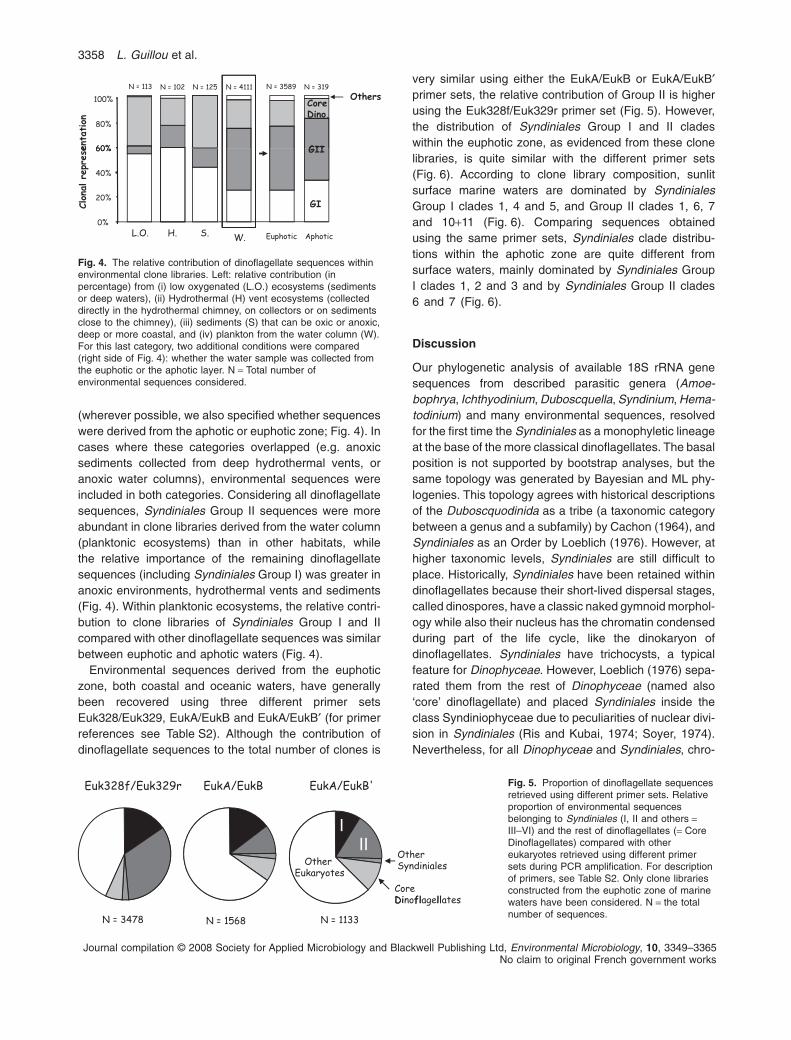
(wherever possible, we also specified whether sequenceswere derived from the aphotic or euphotic zone; Fig. 4). Incases where these categories overlapped (e.g. anoxicsediments collected from deep hydrothermal vents, oranoxic water columns), environmental sequences wereincluded in both categories. Considering all dinoflagellatesequences, Syndiniales Group II sequences were moreabundant in clone libraries derived from the water column(planktonic ecosystems) than in other habitats, whilethe relative importance of the remaining dinoflagellatesequences (including Syndiniales Group I) was greater inanoxic environments, hydrothermal vents and sediments(Fig. 4). Within planktonic ecosystems, the relative contri-bution to clone libraries of Syndiniales Group I and IIcompared with other dinoflagellate sequences was similarbetween euphotic and aphotic waters (Fig. 4).
Environmental sequences derived from the euphoticzone, both coastal and oceanic waters, have generallybeen recovered using three different primer setsEuk328/Euk329, EukA/EukB and EukA/EukB′ (for primerreferences see Table S2). Although the contribution ofdinoflagellate sequences to the total number of clones is
very similar using either the EukA/EukB or EukA/EukB′primer sets, the relative contribution of Group II is higherusing the Euk328f/Euk329r primer set (Fig. 5). However,the distribution of Syndiniales Group I and II cladeswithin the euphotic zone, as evidenced from these clonelibraries, is quite similar with the different primer sets(Fig. 6). According to clone library composition, sunlitsurface marine waters are dominated by SyndinialesGroup I clades 1, 4 and 5, and Group II clades 1, 6, 7and 10+11 (Fig. 6). Comparing sequences obtainedusing the same primer sets, Syndiniales clade distribu-tions within the aphotic zone are quite different fromsurface waters, mainly dominated by Syndiniales GroupI clades 1, 2 and 3 and by Syndiniales Group II clades6 and 7 (Fig. 6).
Discussion
Our phylogenetic analysis of available 18S rRNA genesequences from described parasitic genera (Amoe-bophrya, Ichthyodinium, Duboscquella, Syndinium, Hema-todinium) and many environmental sequences, resolvedfor the first time the Syndiniales as a monophyletic lineageat the base of the more classical dinoflagellates. The basalposition is not supported by bootstrap analyses, but thesame topology was generated by Bayesian and ML phy-logenies. This topology agrees with historical descriptionsof the Duboscquodinida as a tribe (a taxonomic categorybetween a genus and a subfamily) by Cachon (1964), andSyndiniales as an Order by Loeblich (1976). However, athigher taxonomic levels, Syndiniales are still difficult toplace. Historically, Syndiniales have been retained withindinoflagellates because their short-lived dispersal stages,called dinospores, have a classic naked gymnoid morphol-ogy while also their nucleus has the chromatin condensedduring part of the life cycle, like the dinokaryon ofdinoflagellates. Syndiniales have trichocysts, a typicalfeature for Dinophyceae. However, Loeblich (1976) sepa-rated them from the rest of Dinophyceae (named also‘core’ dinoflagellate) and placed Syndiniales inside theclass Syndiniophyceae due to peculiarities of nuclear divi-sion in Syndiniales (Ris and Kubai, 1974; Soyer, 1974).Nevertheless, for all Dinophyceae and Syndiniales, chro-
60%
80%
100%
N = 113 N = 102 N = 3589
GII
CoreDino.
Othersen
tation
0%
20%
40%
60%
GI
GII
Clon
al r
epre
se
L.O. H. S. W. Euphotic Aphotic
N = 125 N = 4111 N = 319
Fig. 4. The relative contribution of dinoflagellate sequences withinenvironmental clone libraries. Left: relative contribution (inpercentage) from (i) low oxygenated (L.O.) ecosystems (sedimentsor deep waters), (ii) Hydrothermal (H) vent ecosystems (collecteddirectly in the hydrothermal chimney, on collectors or on sedimentsclose to the chimney), (iii) sediments (S) that can be oxic or anoxic,deep or more coastal, and (iv) plankton from the water column (W).For this last category, two additional conditions were compared(right side of Fig. 4): whether the water sample was collected fromthe euphotic or the aphotic layer. N = Total number ofenvironmental sequences considered.
Fig. 5. Proportion of dinoflagellate sequencesretrieved using different primer sets. Relativeproportion of environmental sequencesbelonging to Syndiniales (I, II and others =III–VI) and the rest of dinoflagellates (= CoreDinoflagellates) compared with othereukaryotes retrieved using different primersets during PCR amplification. For descriptionof primers, see Table S2. Only clone librariesconstructed from the euphotic zone of marinewaters have been considered. N = the totalnumber of sequences.
EukA/EukB'Euk328f/Euk329r EukA/EukB
Core Di fl ll
OtherSyndiniales
III
Dinoflagellates
N = 3478 N = 1133N = 1568
OtherEukaryotes
3358 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

mosomes stay anchored to the persistant nuclear enve-lope during mitotic division, but their segregation is drivenby microtubules through the nuclear envelope in Dino-phyceae while centrioles participate in the formation of themitotic apparatus in Syndiniales (Hollande, 1974; Ris andKubai, 1974). Another Syndiniales peculiarity is their lowchromosome number, 4–10 for Syndiniales compared with>100 for some Dinophyceae members such as Gonyaulaxand Ceratium (see Loeblich, 1976). Taking all this together,it seems reasonable that these last two features (peculiari-ties in nuclear division and low chromosome number) arederived from an initial dinoflagellate model (Xiao-Ping andJing-Yan, 1986), being forced by the obligately parasiticlifestyle of Syndiniales. Controversially, some recent phy-logenies using 28S rRNA gene sequences, have placedthe Syndiniales Ichthyodinium chaberladi (Group I Clade3) within dinoflagellates (Gestal et al., 2006) and an envi-ronmental sequence belonging to Group I clade 1 close toPerkinsus (Massana et al., 2008). More sequences areneeded to obtain a LSU rRNA phylogeny robust enough toprovide accurate affiliations for the Syndiniales. Proteincoding genes, such as actin and b- and g-tubulins may alsobe a better alternative to resolve basal branches of thedinoflagellate lineage (Saldarriaga et al., 2003), includingseveral deep genera such as Perkinsus, Oxyrrhis andNoctiluca, which are almost impossible to place usingphylogenies based upon ribosomal genes.
Within Syndiniales, five main families have beendescribed: the Sphaeriparidae and the Coccidinidae (nosequences available to date for both), the Amoebophry-idae, the Duboscquellidae and the Syndinidae (Table S3).As Ichthyodinium (Syndinidae) and Duboscquella(Duboscquellidae) are clearly allied within Group I by 18S
rRNA gene analyses, while Hematodinium and Syndinium(Syndinidae) are members of Group IV and quite distinctfrom Group I, it is likely that the taxonomy of Syndinidaewill soon be modified. Nevertheless, it is possible thatAmoebophryidae will remain synonymous with Syndini-ales Group II. To date, seven different species of Amoe-bophrya have been described (see Table S3). However,this clearly does not reflect the genetic diversity observedin Group II; herein we find at least 44 distinct clades.Unfortunately, only one of the described species,A. ceratii, has been genetically characterized to date.Amoebophrya ceratii is a parasitoid of many (if not all)dinoflagellates, including heterotrophic species (for areview see Park et al., 2004) and was most recentlyreported as being a ‘complex species’ rather than a realspecies (see Coats et al., 1996). Currently, available SSUrRNA gene sequences from this complex species arefrom strains able to infect photosynthetic dinoflagellates,in accordance with the fact that this group is largelyretrieved from the euphotic zone in environmental studies.Nevertheless, A. ceratii was also described as being ableto parasitize other Amoebophrya species (e.g. it is fre-quently observed in A. leptodisci) or other Syndiniales(such as the genus Keppenodinium; see Cachon 1964).Consequently, despite recent additions of a substantialnumber of new sequences, all Amoebophrya SSU rRNAgene sequences fall within a monophyletic lineage, whichcontains at least 10 different clades. However, the taxo-nomic correspondence of each clade is still not clear.Within Hematodinium and Syndinium, SSU rRNA genesequences from different species are almost identical,and species can only be distinguished using more vari-able genetic regions (ITS, see Hudson and Adlard, 1996;
Fig. 6. Syndiniales clades retrieved usingdifferent primer sets, in the euphotic andaphotic layers of marine waters. The relativeproportion of clades belonging to Syndiniales(groups I and II) using different sets ofprimers is indicated. Only clone librariesconstructed from the euphotic zone of marinewaters have been included (as in Fig. 5).N = the total number of sequences.
88
Euphotic Euphotic Aphotic
15
67
1
4
5
67
Group I
1
23
4
56 7 8
23
4
23
4
1
2345
1617
2226
20 1
2345
616
20222327
Group II
69
10152021
29 1
Euk328f/Euk329r EukA/EukB +
6
7
89
10+11
1213141516 6
7
89
10+111213
141516Group II
Euk328f/Euk329r +EukA/EukB +
7
Euk328f/Euk329r EukA/EukB +EukA/EukB’
EukA/EukB +EukA/EukB’
Diversity of parasitic Syndiniales 3359
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

Skovgaard et al., 2005). In the case of Amoebophrya,genetic variability is much more evident, even using con-served markers such as the SSU rRNA gene.
All described Syndiniales are parasitoids, and obli-gately kill their host. A potential exception is Sphaeriparacatenata, which in some cases only causes castration ofits host, but not death (Cachon and Cachon-Enjumet,1964). Described Syndiniales generally complete their lifecycle in less than 3 days, leading to the release of hun-dreds of free-living dispersive dinospores. Environmentalsequences obtained from genetic libraries likely resultfrom such dinospores. In general, Syndiniales producetwo different types of dinospores, the micro and mac-rospores that can both be very small (from 1 to 12 mm indiameter depending on the species). There is a generalagreement that dinospores can only survive a few daysafter their release (e.g. Coats and Park, 2002). Hence, thefact that they systematically dominate clone libraries fromcoastal and oceanic waters, whatever the season consid-ered, suggests that their production is fairly constant.Alternatively, it could mean that other more resistantplanktonic stages are produced. To our knowledge, cystproduction has never been reported within Syndiniales,with the notable exception of cyst-like cells described innatural populations of Duboscquella cachoni infecting thetintinnid Eutintinnus pectinis (Coats, 1988). In this particu-lar case, cyst-like formation was completely independentof dinospore production, and the cysts formed were non-motile. Within dinoflagellates, permanent cysts are pro-duced by the fusion of two gametes. Sexuality, issuedfrom different individuals (anisogamous and heterothal-lic), has only been observed once in Coccidinidae, withthe production of a planozygote-like body (Chatton andBiecheler, 1936). Nevertheless, cyst production was notobserved.
The recent reports of Amoebophrya spp. in freshwatersystems (Lefèvre et al., 2008) are based on placement oftwo environmental sequences incorrectly labelled uncul-tured Amoebophrya Clone E and F (Di Giuseppe and Dini,2004). These sequences are in fact closely related to thegenus Cryptocaryon (Ciliates, Protosmatea). Thus, todate, environmental sequences of Syndiniales have beenretrieved solely from marine ecosystems. This supportsthe ‘marine-ness’ of this group, together with the fact thatall described Syndiniales species are marine. In contrast,other unicellular alveolate parasites, such as Perkinsea,Colpodellids or even other parasitic dinoflagellates, haveboth marine and continental species. This is interestinggiven that almost all known Syndiniales hosts havespecies well adapted to continental ecosystems (withinciliates, dinoflagellates, cercozoa and crabs).
Although the distribution of Syndiniales is relativelyhomogeneous at the group level (note their similar con-tribution to both euphotic and aphotic zones), communi-
ties are quite different at the clade level. Some cladesare uniformly distributed vertically through the watercolumn (e.g. Group I clade 1, Group II clade 6), whereasothers (e.g. Group I clades 2 and 3, Group II clade 7)are more prevalent in aphotic layers. We speculate thatthese ‘deeper’ clades are able to parasitize well-knowndeep planktonic organisms such as Phaeodarea (Cerco-zoa), Acantharea (Radiolaria) and Polycystinea (Radi-olaria). Recent analyses of environmental sequencesbelonging to these groups highlight the increasing con-tribution of radiolarian environmental sequences withdepth (Not et al., 2007), in particular sequences belong-ing to Spumellarida (Polycystinea). Interestingly, radi-olarians are also exclusively marine, and well known tobe oceanic, or blue-water, organisms. Hence, both Syn-diniales and radiolarians may have shared a commonevolutionary history inhabiting similar ecosystems asdiscussed by Not and colleagues (2007). Sequencesretrieved from single radiolarian cells (Dolven et al.,2007), likely infected by Syndiniales, belong to bothGroup I and Group II (Fig. 3). Nevertheless, it is difficultto assign an identity to these sequences as Syndinialesable to infect radiolarians are described within three ofthe five described families (the Amoebophryidae, theDuboscquellidae and the Syndinidae). Much more infor-mation on host specificity is provided by data from thegenus Amoebophrya. Phylogenies of Amoebophryaclades (Fig. 3) indicate that host–parasite co-evolution isnot a central feature driving the observed diversity. Para-sites of the same host genus, but of different species,belong to completely separate clades. In fact, within the‘A. ceratii complex’, host specificity as assessed in cul-tures, is highly strain dependent, with some beingextremely specific (Coats and Park, 2002), while othershave broader host ranges (Kim, 2006). This fact couldmask clear co-evolutionary patterns. The same conclu-sion can be drawn when the entire Syndiniales order isconsidered. For example, all described Syndiniales thatoperate as parasitoids of metazoans are not closelyrelated. The genus Ichthyodinium, a parasite of fisheggs, is more closely related to parasites that infect cili-ates (i.e. Duboscquella) belonging to Group I than toGroup IV, which contains Syndinium and Hematodinium(two genera which infect metazoans). Thus, evenbetween groups, Syndiniales and their hosts do notseem to co-evolve. Consequently, we expect that para-sites within Syndiniales are generalists or opportunistswith respect to host range. This is also congruent withthe detection of clades widely distributed in all exploredmarine ecosystems. Nevertheless, the presence of morespecialized clades or subclades, related with depth orspecifically retrieved from extreme environments, suchas Group I clades 3 and 4, and Group II clades 9 and15, confirms that specific interactions may exist.
3360 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

Conclusions
Based on the integration of a large number of environ-mental sequences, we propose a new phylogeny forSyndiniales. This order should be considered a mono-phyletic sister group of the rest of Dinophyceae basedupon 18S rRNA gene phylogenies. The Syndiniales arecomposed of five main groups, which include environ-mental sequences formerly referred to as novel marinealveolate groups I and II. All Syndiniales speciesdescribed to date are parasitic marine organisms. Theexclusively marine lifestyle of this order as a whole,including sequences from uncultured organisms, is con-firmed by the fact that their 18S rRNA gene sequenceshave not been retrieved from terrestrial or freshwaterPCR surveys, but rather come solely from marine eco-systems. Syndiniales appear to colonize all the marinehabitats investigated thus far, from oceanic surfacewaters to sediments. Some clades are likely adapted tofairly specific habitats (and/or hosts that reside only inspecific habitats). However, these parasites and theirhosts do not appear to share co-evolutionary trends.This suggests that at least some Syndiniales are highlyopportunistic, with a capacity to infect very differentmarine hosts, from many trophic levels within marinefood webs. Even so, we cannot completely exclude thatsome members of the Syndiniales may have retainedspecific ancestral traits still found among the differentalveolate lineages, e.g. phagotrophy and/or the inherit-ance of photosynthetic genes. In fact, proteins charac-teristic of secondary endosymbiont plastids haverecently been detected from both the non-photosyntheticgenera Oxyrrhis and Perkinsus (Matsuzaki et al., 2008;Slamovits and Keeling, 2008). Hence, the ecologicalsuccess of the Syndiniales may yet have a more sur-prising and complex explanation. That said, it is certainlystill the case that the overall role of parasitism in marinesystems requires further investigation and incorporationinto food web and carbon flow models.
Experimental procedures
Environmental sampling and library processing
Materials and methods for each of the libraries publishedherein varied by site. Thus, details are provided separately foreach of the regions sampled. See also Table 1 for additionaldetails on the clone libraries constructed.
English Channel, France (RA). Ten litres of surface seawater was sampled weekly at the ASTAN site (SOMLITstation, English Channel, France, see http://www.domino.u-bordeaux.fr/somlit_national/) from the end of June throughto the middle of September 2004. Water samples were size-fractionated using a peristaltic pump (with the flow rate fixed
at 100 ml min-1) through sequential 47-mm-diameter poly-carbonate filters (12 and 3 mm, Osmonic Poretics Products)in filter housings connected in series, terminated by a 0.2-mm-pore-size Sterivex unit (Millipore). Following sample col-lection, filters were then submerged in DNA lysis buffer(0.75 M sucrose, 40 mM EDTA, 50 mM Tris-HCl, pH 8),immediately frozen in liquid nitrogen and stored at -80°C.DNA was extracted using a 3% (w/v) CTAB (Cetyltrimethy-lammonium bromide) extraction procedure (Doyle andDoyle, 1987). PCR conditions for amplifying the 18S rRNAgene were as described in Moon-van der Staay and col-leagues (2001). PCR products were cloned using theTOPO-TA cloning kit (Invitrogen). Clones (96 randomlyselected) were partially sequenced using the primerEuk528f for RA1 and the primer ALV01 (for primer refer-ences see Table S2) for both RA2 and RA3 on an ABIPrism 3100 (Applied Biosystems).
Barcelona Harbour, Mediterranean Sea, Spain (BAFRACT).Fifty litres of sea water was collected on 28 February 2001from Barcelona harbour (Spain) and pre-filtered through a50 mm mesh. On return to the laboratory (less than one hour),the sample was then fractionated through seven differentpore-sizes (20, 14, 10, 5, 3, 1 and 0.6 mm), using similar filterhousings connected in series as described above. Samplestorage and DNA extraction were as described above.Genetic diversity in each size fraction was determined byDGGE (not shown), as described by Díez and colleagues(2001b). Clone libraries were performed for size fractionsdisplaying the most ‘contrasting’ DGGE band patterns. PCRconditions were as in Díez and colleagues (2001a). Cloningwas as described above. Restriction fragment length poly-morphism (RFLP) analysis was used to detect polymor-phisms among the retrieved clones. Briefly, PCR productswere digested with 1 U ml-1 of restriction enzyme HaeIII(Gibco BRL) for 6–12 h at 37°C. The digested products wereseparated by electrophoresis at 80 V for 2–3 h in a 2.5%low-melting-point agarose gel. One clone per unique RFLPpattern was partially sequenced using the internal primerEuk528f. Sequencing was performed by Qiagen GenomicsSequencing Services (Germany).
Blanes Bay. Surface water was collected in Blanes Bay(Catalan coast, north-western Mediterranean Sea). Seawater was rapidly transported to the laboratory (less than2 h), and gently filtered by gravity through 2- and 5-mm-pore-size polycarbonate filters. One litre of each size fraction wasincubated both in the light [around 100 (mmol quanta) m-2 s-1]and in the dark (four bottles in total). All the incubations werecarried out at 20°C, close to in situ temperature (18°C).Samples (100 ml) were filtered through 0.2-mm-pore-sizeDurapore filters for DNA analysis over a course of 5 days.DNA was then extracted following the protocol described inDíez and colleagues (2001b). Abundance and genetic diver-sity of eukaryotes were monitored by epifluorescence micros-copy and DGGE (using the same protocol as describedabove, data not show). 18S rRNA genes were amplified fromthe initial day and after 4 days of incubation both in the lightand dark. PCR products were cloned using the TOPO-TAcloning kit. Clones were sequenced by the Qiagen GenomicsSequencing Services (Germany).
Diversity of parasitic Syndiniales 3361
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

Atlantic Meridional Transect (AMT15). Both the samplingstrategy, DNA extraction procedure and the PCR amplifica-tion protocol used have been previously published (Zwirgl-maier et al., 2007). Cloning was as described above.Restriction fragment length polymorphism analysis was usedto screen for distinct inserts by digesting with HaeIII in a totalvolume of 20 ml using 5 U of HaeIII at 37°C for 2 h. Fragmentswere resolved by electrophoresis in a 2.5% agarose gel at60 V for 3 h. At least two representatives of each RFLPpattern were sequenced in entirety following purification ofthe PCR product using the QIAquick PCR Purification Kit(Qiagen), on an ABI 3130xl Genetic Analyser (AppliedBiosystems).
Florida Straits and Sargasso Sea. Sea water was collectedfrom both the surface and deep chlorophyll maximum onthree transect cruises across the Straits of Florida in 2005 asdescribed in Cuvelier and colleagues (2008). Briefly, 1 lsamples were first gravity filtered through 47 mm diameter,2-mm-pore-size filters (GE Osmonics, Minnetonka, Minne-sota, USA) and then onto a 0.2-mm-pore-size Supor filter (PallGelman, Ann Arbor, Michigan, USA) using approximately 5psi vacuum. Filters were placed in cryovials and frozen inliquid nitrogen. DNA was extracted using the DNeasy kitaccording to the manufacturer’s recommendations (Qiagen,Germantown, MD, USA). PCR conditions have been pub-lished elsewhere (Cuvelier et al., 2008). The PCR productwas ligated into vector pCR2.1 (Invitrogen, Carlsbad, CA,USA) and transformed. Clones were then sequenced on anABI 3730xl sequencer (Applied Biosystems).
Samples were also collected on a cruise from Woods Hole,MA USA to the BATS. Two regions were sampled, BATS anda site north-west of BATS. Samples were collected and pro-cessed as for the Florida Straits clone libraries (as reported inCuvelier et al., 2008).
Development of sequence database
The 18S rRNA gene database was built using all publishedenvironmental sequences deposited in GenBank up toDecember 2007. We also integrated the data sets generatedherein and a number of other libraries which are beingpublished independently, but without specific attention tothe analysis of Alveolata sequences. The complete list ofsequences used in this study and their annotation is availableupon request (Excel file).
Sequence group assignments
The software package, KeyDNAtools, was used to (i) detectsequences belonging to dinoflagellates within a large data-base including more than 42000 eukaryotic SSU rRNA genesequences; (ii) remove potential chimeras; and (iii) automati-cally assign sequences to a specific clade. Details of thesoftware development will be described elsewhere. Briefly,environmental sequences were annotated using small oligo-nucleotides ‘probes’ (15 bp length), named ‘keys’, which weregenerated in silico. The specificity of each key, from the Orderto the clade level (where possible) was automaticallydeduced from a reference database containing as many
eukaryotic lineages as possible as outgroup and at leasttwo sequences from each taxa created within marinedinoflagellates. In this study, more than 110000 keys weregenerated, each being specific at least to the level of Order.Annotations of 18S rRNA genes were deduced from thespecificity of each key when they matched a sequence. As anexample, sequence EF172938 was annotated by 175 differ-ent keys, providing converging annotation at the order leveland covering positions 1–1289 along the sequences(Table 3). Thus, this sequence was considered as a Syndini-ales, Dino-Group II, and belonging to Dino-Group II-Clade 7(this last annotation is provided by 47 keys). The 18S rRNAgene sequences belonging to the majority of Dinophyceaewere not variable enough to allow a clear separation ofsequences even at the Order level. Thus, keys specific tomost of the Dinophyceae are labelled as Dinophyceae at theClass and the Order levels (see annotation of sequenceEU087283). Wherever possible, more precise information isgiven (e.g. at the Family, Genus, Species and Clade levels;see annotation of sequence EU087283). New groups/cladeswere often detected by conflicting annotation (caused byincorrect deduction of specificity by the keys) in a particulartaxonomic field. Chimeras were detected when annotationsconflicted for separate parts of an individual sequence. As anexample (Table 3), sequence AJ965100 was detected as achimera at the class level, a false gene sequence combiningpart of a Ciliophora (annotation provided by 25 keys coveringpositions 44–155) gene sequence with part of a Dinophyceae(31 keys covering positions 213–474) gene sequence. Auto-matic annotation of eukaryotic 18S rRNA environmental genesequences using the final generated keys can be freelytested on the following web site: http://KeyDNAtools.com.
Phylogenetic analyses
Sequences were aligned using the slow and iterative refine-ment method FFT-NS-i with Mafft 5.8 software (Katoh et al.,2007). Using the BioEdit sequence editor (Hall, 1999) forvisualization of the alignment, we deduced the secondarystructures by hand using previously published studies (seefor example Lange et al., 1996). For complete sequencealignments only, poorly aligned and highly variable regions ofthe alignments were automatically removed using Gblocks(Castresana, 2000) with the following parameters: allowinggaps in half positions and the minimum length of a block = 5.Different nested models of DNA substitution and associatedparameters were tested using Modeltest v.3.06 (Posada andCrandall, 1998). Settings given by Modeltest were used toperform the NJ and the ML analyses. The NJ, ML and MPanalyses were performed using PAUP 4.0b10 (Swofford,2002). A heuristic search procedure using the tree bisection/reconnection branch swapping algorithm (setting as in MP)was performed to find the optimal ML tree topology (with70 000 re-arrangements). Bootstrap values for NJ and MPwere estimated from 1000 replicates. For MP, the number ofre-arrangements was limited to 5000 for each bootstrapreplicate. Starting trees were obtained by randomized step-wise addition (number of replicates = 20). Additionally, weused Bayesian reconstruction for the analysis of completesequences with MrBayes, v.3.0b4 (Huelsenbeck and Ron-quist, 2001). The GTR model of substitution was used, taking
3362 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

into account a gamma-shaped distribution of the rates ofsubstitution among sites. The chains were run for 1000000generations. Trees were sampled every 100 generations. Thefirst 5000 sampled trees, corresponding to the initial phasebefore the chains became stationary, were discarded(burn in).
Acknowledgements
We thank R. Cowen, B.J. Binder and E. Goetze for cruisespace as well as the captain and crews of the R/V Oceanus,R/V Walton Smith; R/V Atalante, R/V Melville, RRS Discov-ery; M.L. Cuvelier, A. Engman, C. Guigand, J.A. Hilton and F.Not for cruise assistance; J. Heidelberg, A. Ortiz, M. Peren-nou, C. Cormier-Caillault, J. Winkler and A. Gobet for helpwith clone libraries and sequencing; and P. Zimmermann forhelp with sequence annotation. This work was supported bythe GIS génomique marine and the ANR AQUAPARADOXprojects (to L.G.), the Gordon and Betty Moore Foundationand National Science Foundation grant OCE-0836721 (toA.Z.W.) and by the NERC (to A.R.K., D.J.S.).
References
Cachon, J. (1964) Contribution à l’étude des péridiniensparasites. Cytologie, cycles évolutifs. Ann Sc Nat ZoolParis VI: 1–158.
Cachon, J., and Cachon-Enjumet, M. (1964) Cycle évolutif etcytologie de Neresheimeria catenata Neresheimer, péridi-nien parasite d’appendiculaires. Ann Sc Nat Zool Paris 12ème série: 779–800.
Castresana, J. (2000) Selection of conserved blocks frommultiple alignments for their use in phylogenetic analysis.Mol Biol Evol 17: 540–552.
Cavalier-Smith, T. (1993) Kingdom Protozoa and its 18 phyla.Microbiol Rev 57: 953–994.
Chatton, E., and Biecheler, B. (1936) Documents nouveauxrelatifs aux Coccidinides (Dinoflagellés parasites). Lasexualité du Coccidinium meslini n.sp. C R Acad Sc Paris203: 573–576.
Coats, D.W. (1988) Duboscquella cachoni n. sp., a parasiticdinoflagellate lethal to its tintinnine host Eutintinnuspectinis. J Protozool 35: 607–617.
Coats, D.W., and Park, M.G. (2002) Parasitism of photosyn-thetic dinoflagellates by three strains of Amoebophrya(Dinophyta): parasite survival, infectivity, generation time,and host specificity. J Phycol 38: 520–528.
Coats, D.W., Adam, E.J., Gallegos, C.L., and Hedrick, S.(1996) Parasitism of photosynthetic dinoflagellates in ashallow subestuary of Chesapeake Bay, USA. AquatMicrob Ecol 11: 1–9.
Countway, P.D., Gast, R.J., Dennett, M.R., Savai, P., Rose,J.M., and Caron, D.A. (2007) Distinct protistan assem-blages characterize the euphotic zone and deep sea (2500m) of the Western North Atlantic (Saragasso Sea and GulfStream). Environ Microbiol 9: 1219–1232.
Cuvelier, M.L., Ortiz, A., Kim, E., Moehlig, H., Richardson,D.E., Heidelberg, J.F., et al. (2008) Widespread distribu-tion of a unique marine protistan lineage. Environ Microbiol10: 1621–1634.Ta
ble
3.E
xam
ples
ofth
ree
diffe
rent
auto
mat
ican
nota
tions
gene
rate
dby
the
Key
DN
Ato
ols.
IDLe
ngth
Sup
erG
roup
Div
isio
nC
lass
Ord
erF
amily
Gen
usS
peci
esC
lade
Cas
e1:
EF
1729
3812
88C
hrom
alve
olat
a–>
175
(1–1
289)
Alv
eola
ta–>
175
(1–1
289)
Din
ophy
ceae
–>17
5(1
–128
9)S
yndi
nial
es–>
175
(1–1
289)
Din
o-G
roup
II–>
174
(1–1
289)
Din
o-G
roup
II-C
lade
7–>
47(1
–119
6)
Cas
e2:
EU
0872
8388
6C
hrom
alve
olat
a–>
53(3
8–71
6)A
lveo
lata
–>53
(38–
716)
Din
ophy
ceae
–>53
(38–
716)
Din
ophy
ceae
–>53
(38–
716)
Sue
ssia
les
–>11
(38–
656)
Sym
biod
iniu
m–>
7(1
42–6
56)
Sym
biod
iniu
mcl
ade
B+
B′+
D–>
3(5
21–6
56)
Cas
e3:
AJ9
6510
049
9C
hrom
alve
olat
a–>
56(4
4–47
4)A
lveo
lata
–>56
(44–
474)
Din
ophy
ceae
–>31
(213
–474
)/C
iliop
hora
–>25
(44–
155)
Syn
dini
ales
–>31
(213
–474
)/O
ligot
riche
a–>
25(4
4–15
5)
Din
o-G
roup
II–>
31(2
13–4
74)
Str
ombi
dium
grou
p–>
2(7
5–99
)
Diversity of parasitic Syndiniales 3363
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

Di Giuseppe, G., and Dini, F. (2004) Identita dei dinoflagellatiproliferanti in strutture sperimentali in situ nel Lago di Tovel(Nord Italia). Studi Trentini Sci. Nat. Acta Biol 81: 427–438.
Díez, B., Pedrós-Alió, C., and Massana, R. (2001a) Study ofgenetic diversity of eukaryotic picoplankton in differentoceanic regions by small-subunit rRNA gene cloning andsequencing. Appl Environ Microbiol 67: 2932–2941.
Díez, B., Pedrós-Alió, C., Marsh, T.L., and Massana, R.(2001b) Application of denaturing gradient gel electro-phoresis (DGGE) to study the diversity of marine picoeu-karyotic assemblages and comparison of DGGE with othermolecular techniques. Appl Environ Microbiol 67: 2942–2951.
Dolven, J.K., Lindqvist, C., Albert, V.A., Bjørklund, K.R.,Yuasa, T., Takahashi, O., and Mayama, S. (2007) Molecu-lar diversity of alveolates associated with neritic NorthAtlantic radiolarians. Protist 158: 65–76.
Doyle, J.J., and Doyle, J.L. (1987) A rapid DNA isolation forsmall quantities of fresh leaf tissue. Phytochem Bull 19:11–15.
Garczarek, L., Dufresne, A., Rousvoal, S., West, N.J.,Mazard, S., Marie, D., et al. (2007) High vertical and lowhorizontal diversity of Prochlorococcus ecotypes in theMediterranean Sea in summer. FEMS Microbiol Ecol 60:189–206.
Gestal, C., Novoa, B., Posada, D., Figueras, A., andAzevedo, C. (2006) Perkinsoide chabelardi n. gen., a pro-tozoan parasite with an intermediate evolutionary position:possible cause of the decrease of sardine fisheries?Environ Microbiol 8: 1105–1114.
Groisillier, A., Massana, R., Valentin, K., Vaulot, D., andGuillou, L. (2006) Genetic diversity and habitats of twoenigmatic marine alveolate lineages. Aquat Microb Ecol42: 277–291.
Hall, T.A. (1999) BioEdit: a user-friendly biological sequencealignment editor and analysis program for windows 95/98/NT. Nucleic Acids Symp Ser 41: 95–98.
Harada, A., Ohtsuka, S., and Horiguchi, T. (2007) Species ofthe parasitic genus Duboscquella are members of the enig-matic marine alveolate Group I. Protist 158: 337–347.
Hollande, A. (1974) Etude comparée de la mitose syndini-enne et de celle des péridiniens libres et deshypermastigines. Infrastructure et cycle évolutif des Syn-dinides parasites de radiolaires. Protistologica X: 413–451.
Hudson, D.A., and Adlard, R.D. (1996) Nucleotide sequencedetermination of the partial SSU rDNA gene and ITS1region of Hematodinium cf perezi and Hematodinium-likedinoflagellates. Dis Aquat Organ 24: 55–60.
Huelsenbeck, J.P., and Ronquist, F. (2001) MrBayes: baye-sian inference of phylogenetic trees. Bioinformatics 17:754–755.
Katoh, K., Kuma, K.-i., Toh, H., and Miyata, T. (2007) MAFFTversion 5: improvement in accuracy of multiple sequencealignment. Nucleic Acids Res 33: 511–518.
Kim, S. (2006) Patterns in host range for two strains ofAmoebophrya (Dinophyta) infecting thecate dinoflagel-lates: Amoebophrya spp. ex Alexandrium affine and exGonyaulax polygramma. J Phycol 42: 1170–1173.
Kim, S., Park, M.G., Kim, K.-Y., Kim, C.-H., Yih, W., Park,J.S., and Coats, D.W. (2008) Genetic diversity of parasiticdinoflagellates in the genus Amoebophrya and its relation-
ship to parasite biology and biogeography. J EukaryotMicrobiol 55: 1–8.
Lange, M., Guillou, L., Vaulot, D., Simon, N., Amann, R.I.,Ludwig, W., and Medlin, L.K. (1996) Identification of theclass Prymnesiophyseae and the genus Phaeocystis withribosomal RNA-targeted nucleic acid probes detected byflow cytometry. J Phycol 32: 858–868.
Leander, B.S., and Keeling, P.J. (2003) Morphostasis inalveolate evolution. Trends Ecol Evol 18: 395–402.
Lefèvre, E., Roussel, B., Amblard, C., and Sime-Ngando, T.(2008) The molecular diversity of freshwater picoeukary-otes reveals high occurrence of putative parasitoids in theplankton. PLoS ONE 3: e2324.
Lessard, E.J., and Swift, E. (1986) Dinoflagellates from theNorth Atlantic classified as phototrophic or heterotrophic byepifluorescence microscopy. J Plankton Res 8: 1209–1215.
Loeblich, A.R. III (1976) Dinoflagellate evolution: speculationand evidence! J Protozool 23: 13–28.
López-García, P., Rodríguez-Valera, F., Pedrós-Alió, C., andMoreira, D. (2001) Unexpected diversity of small eukary-otes in deep-sea Antarctic plankton. Nature 409: 603–607.
Lovejoy, C., Massana, R., and Pedrós-Alió, C. (2006) Diver-sity and distribution of marine microbial eukaryotes in theArctic Ocean and adjacent seas. Appl Environ Microbiol72: 3085–3095.
Massana, R., Balagué, V., Guillou, L., and Pedrós-Alió, C.(2004) Picoeukaryotic diversity in an oligotrophic coastalsite studied by molecular and culturing approaches. FEMSMicrobiol Ecol 50: 231–243.
Massana, R., Guillou, L., Terrado, R., Forn, I., and Pedrós-Alió, C. (2006) Growth of uncultured heterotrophic flagel-lates in unamended seawater incubations. Aquat MicrobEcol 45: 171–180.
Massana, R., Karniol, B., Pommier, T., Bodaker, I., and Béjà,O. (2008) Metagenomic retrieval of a ribosomal DNArepeat array from an uncultured marine alveolate. EnvironMicrobiol 10: 1335–1343.
Matsuzaki, M., Kuroiwa, H., Kuroiwa, T., Kita, K., and Nozaki,H. (2008) A cryptic algal group unveiled: a plastid biosyn-thesis pathway in the oyster parasite Perkinsus marinus.Mol Biol Evol 25: 1167–1179.
Medlin, L.K., Metfies, K., Mehl, H., Wiltshire, K., and Valentin,K. (2006) Picoeukaryotic plankton diversity at the Helgo-land time series site as assessed by three molecularmethods. Microb Ecol 52: 53–71.
Moon-van der Staay, S.Y., Watcher, R.D., and Vaulot, D.(2001) Oceanic 18S rDNA sequences from picoplanktonreveal unsuspected eukaryotic diversity. Nature 409: 607–610.
Murray, S., Jørgensen, M.F., Ho, S.Y.W., Patterson, D.J.,and Jermiin, L.S. (2005) Improving the analysis ofdinoflagellate phylogeny based on rDNA. Protist 156: 269–286.
Not, F., Gausling, R., Penn, K., Heidelberg, J., and Worden,A.Z. (2007) Vertical distribution of picoeukaryotic diversityin the Sargasso Sea. Environ Microbiol 9: 1233–1252.
Not, F., Latasa, M., Scharek, R., Viprey, M., Karleskind, P.,Balagué, V., et al. (2008) Protistan assemblages acrossthe Indian Ocean: a specific emphasis on picoeukaryotes.Deep Sea Res Part I (in press).
3364 L. Guillou et al.
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works

Park, M.G., Yih, W., and Coats, D.W. (2004) Parasites andphytoplankton, with special emphasis on dinoflagellateinfections. J Eukaryot Microbiol 51: 145–155.
Patterson, D.J. (1999) The diversity of eukaryotes. Am Nat154: 96–124.
Posada, D., and Crandall, K.A. (1998) Modeltest: testing themodel of DNA substitution. Bioinformatics 14: 817–818.
Ris, H., and Kubai, D.F. (1974) An unusual mitotic mecha-nism in the parasitic protozoan Syndinium sp. J Cell Biol60: 702–720.
Romari, K., and Vaulot, D. (2004) Composition and temporalvariability of picoeukaryote communities at a coastal site ofthe English channel from 18S rDNA sequences. LimnolOceanogr 49: 784–798.
Saldarriaga, J.F., McEwan, M.L., Fast, N.M., Taylor, F.J.R.,and Keeling, P.J. (2003) Multiple protein phylogenies showthat Oxyrrhis marina and Perkinsus marinus are earlybranches of the dinoflagellate lineage. Int J Syst EvolMicrobiol 53: 355–365.
Silberman, J.D., Collins, A.G., Gershwin, L.-A., Johnson,P.J., and Roger, A.J. (2004) Ellobiopsidae of the genusThallasomyces are alveolates. J Eukaryot Microbiol 51:246–252.
Skovgaard, A., Massana, R., Balagué, V., and Saiz, E. (2005)Phylogenetic position of the copepod-infesting parasiteSyndinium turbo (Dinoflagellata, Syndinea). Protist 156:413–423.
Slamovits, C.H., and Keeling, P.J. (2008) Plastid-derivedgenes in the non-photosynthetic alveolate Oxyrrhis marina.Mol Biol Evol 25: 1297–1306.
Soyer, M.-O. (1974) Etude ultrastructurale de Syndinium sp.Chatton parasite coelomique de copépodes pélagiques.Vie Milieu XXIV: 191–212.
Stoeck, T., Kasper, J., Bunge, J., Leslin, C., Ilyin, V., andEpstein, S. (2007) Protistan diversity in the Arctic: a case ofpaleoclimate shaping modern biodiversity? PLoS ONE 2:e728.
Swofford, D.L. (2002) PAUP*. Phylogenetic Analysis UsingParsimony (*And Others Methods). Sunderland, MA, USA:Sinauer Associates.
Takishita, K., Yubuki, N., Kakizoe, N., Inagaki, Y., andMaruyama, T. (2007) Diversity of microbial eukaryotes insediment at a deep-sea methane cold seep: surveys ofribosomal DNA libraries from a raw sediment samples andtwo enrichment cultures. Extremophiles 11: 563–576.
Viprey, M., Guillou, L., Férréol, M., and Vaulot, D. (2008)Wide genetic diversity of picoplanktonic green algae (Chlo-roplastida) uncovered in the Mediterranean Sea by aphylum-biased PCR approach. Environ Microbiol 10:1804–1822.
Williams, A.G., and Coleman, G.S. (1992) The RumenProtozoa. New York, USA: Springer-Verlag.
Worden, A.Z. (2006) Picoeukaryote diversity in coastalwaters of the Pacific Ocean. Aquat Microb Ecol 43: 165–175.
Xiao-Ping, G., and Jing-Yan, L. (1986) Nuclear division in themarine dinoflagellate Oxyrrhis marina. J Cell Sci 85: 161–175.
Zwirglmaier, K., Heywood, J.L., Chamberlain, K., Woodward,E.M.S., Zubkov, M.V., and Scanlan, D.J. (2007) Basin-scale distribution patterns of picocyanobacterial lineages inthe Atlantic Ocean. Environ Microbiol 9: 1278–1290.
Supporting information
Additional Supporting Information may be found in the onlineversion of this article:Fig. S1. Mean sequence identities within the different cladesbelonging to Syndiniales groups I and II.Table S1. Details of the sequences used for the phyloge-netic analyses described in Fig. 1.Table S2. Primer sequences used or cited in this study.Table S3. Taxonomy of Syndiniales (= Syndinida) and theirhosts modified from Loeblich (1976).
Please note: Blackwell Publishing are not responsible for thecontent or functionality of any supporting materials suppliedby the authors. Any queries (other than missing material)should be directed to the corresponding author for the article.
Diversity of parasitic Syndiniales 3365
Journal compilation © 2008 Society for Applied Microbiology and Blackwell Publishing Ltd, Environmental Microbiology, 10, 3349–3365No claim to original French government works





























