Embed Size (px)
Citation preview

IN ERNATIfN.\i. J„i uvnt. or Lrr'aosv^
V'i,Iun,c 67, Numbcr 1l'rint d in the USA.
(ISSN Ot-1S-91(,X)
Dermal Extracellular Matrix in CutaneousLeprosy Lesions'
Sergio Luiz Gomes Antunes, Maria Eugenia Gallo,Sonia Maria Rocha de Almeida, Ester Mota, Marcelo Pelajo, and
Henrique Leonel Lenzl
The extracellular matrix (ECM) has beenconsidered an environment of crucial im-portance, inlluencing cell behavior. It canwork as an adhesion substrate for migratingcells ('' "-'`') or for rcleased growth factors( 13 ). The ECM also functions as costimula-tory signals for cell proliferation and differ-entiation ( 2 ' 1 ').
The function of cell populations in thetissues can be inferred through the analysisof their extracellular matrix. Developn)ent,neoplasia and inflammation have commonfeatures related to cell-matrix interaction.Therefore, there is an intimate relationshipbetween the assembly of the tissue extracel-lular matrix and the biolosl ical activity. Theknowledge from research on cell-matrix in-teractions in development and in oncog ene-sis may be applied to the interpretation ofthe inflammatory process.
Experimental models of murine schisto-somiasis show granulomas with remodel-ling of the original hepatic extracellular ma-trix and with a stage-dependent preferentialaccumulation of collagen (type I and type
' Received for puhlication on 17 June 1997.Accepted for publication in revised forro on 6 January1999.
S. L. G. Antunes, NI.D., Ph.D., Laboratory ofLeprosy, Oswaldo Cruz Institute, Oswaldo Cruz Foun-dation, Av. Brasil 4365, Manguinhos 21045-900, Riode Janeiro, RJ, Brazil and Iguacu University, NovaIguacu, Brazil. NI. E. Gallo, M.D., Ph.D.; S. M. R. deAlmeida, M.D., Laboratory of Leprosy, Oswaldo CruzInstitute, Oswaldo Cruz Foundation, Rio de Janeiro,RJ, Brazil. E. Mota, M.Sc.; M. Pelajo, M.D.; Il. L.Lenzl, M.D., Ph.D., Department of Pathology, Os-waldo Cruz Institute, Oswaldo Cruz Foundation, Riode Janeiro, RJ, Brazil.
Reprint request to Dr. Antunes at the Oswaldo CruzInstitute address above or FAX 55-21-270-9997;e-mail: santunesOgcne.dbbm.fiocruz.hr
III) (". 0 ), proteoglycans (dermatan-sulfateand heparan sulfate) (5), and fibronectinduring the formation of the granulomatousstructure Cl. A concomitant proteolytic ac-tivity represented by collagenase is also de-tected (""). Maximal degradation and syn-thesis coincides with the peak of the mouseimmune response to the presence of Schis-tose/na nrunsoni ova in the tiver, linking thematrix remodelling occurring in granulo-mas to immunological factors ( 10 ).
ECM changes in leprosy have been stud-ied, emphasizing the presence of auto-anti-bodies to collagen in the serum of leproma-tous patients (''), the pattern of fibronectinin different polar granulomas ( 2') and thepresence of anti-elastic microfibril antibod-ies in the cera of patients with leprosy ( 21 ).
In this work, we studied the effects ofleprosy inflammatory lesions upon the der-mal ECM components (collagen, proteo-glycans, elastic fibers and fibronectin), cor-relating the findings with the correspondingpatient's clinicai and histopathological pic-ture according to the Ridley-Jopling classi-fication ("). An overview of dermal ECMstructural changes in leprosy lesions willraise some specific questions for each ma-trix component involved. Furthermore,pathogenetic implications can be drawnfrom the global comprehension of the al-tered cell-matrix interaction in the leprosylesions.
MATERIALS AND METHODSThe study WaS done with cutaneous le-
sion biopsies from 38 leprosy patients fromSouza Araújo Ambulatory (Laboratory ofLeprosy), Department of Tropical Medicine(Instituto Oswaldo Cruz), Rio de Janeiro,Brazil. The patients were grouped accord-
24
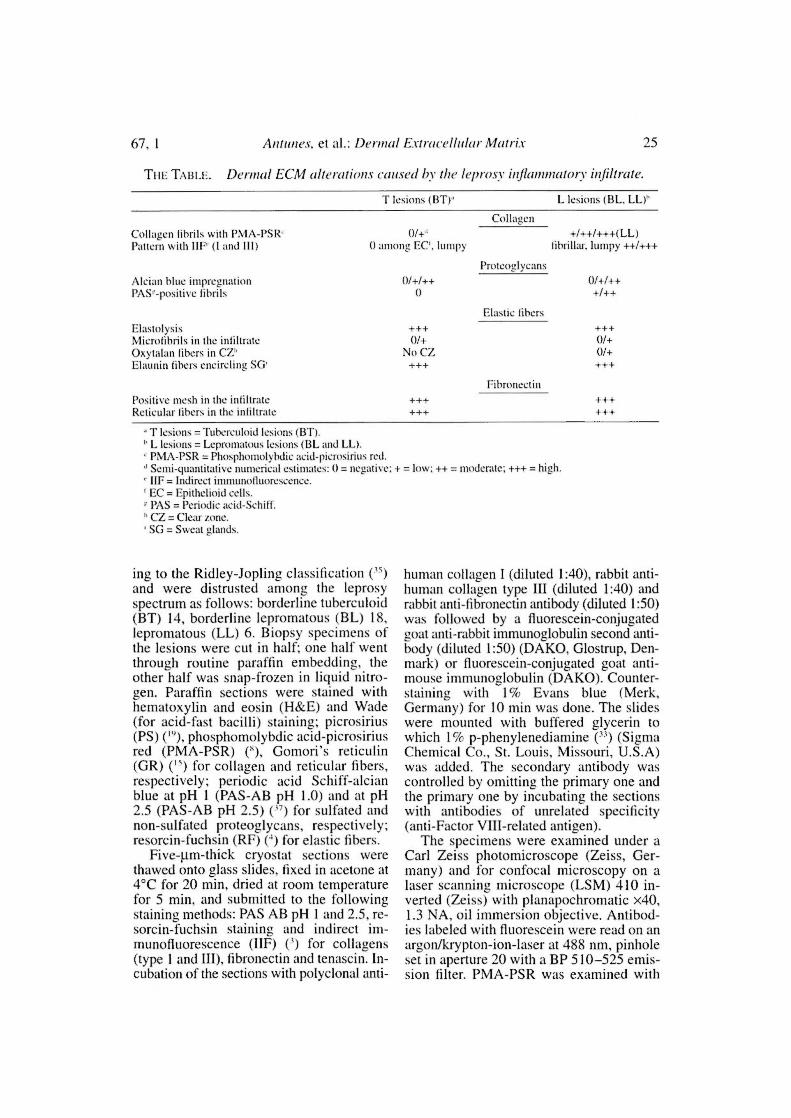
01+''^ +/++/+++(LL )0 among EC'. Ismpy^1ibriiIzu, 1unmpy ++/+++
Proteoglycans
Elastic libers
1ibronectia
67, 1^Antunes, et al.: !)erma/ Evtracellular Matar -^25
TABU:. Defra/ ECM alterations cause(' hv the /e/nc/sv mllUiWm a lnrv infiltrate.
T lesions (BT)'^ L lesions (BL, LL)''
CollagcnCoIiauen lihrils with PNIA-PSR'Pattern with 111 ,° (1 and III)
Alcian blue intpregn: tiosPAS'-positive lihrils
ElastolysisMicrolibrils in the inliltrateOxytalan libers in CZ"Elaunin libers encircling SG'
Positive mesh in the inliltraleReticular libers in the inliltrale
T lesions = Tuberculoid lesions (BT).L lesions = Lepromatous lesions (BL and LL).PMA-PSR = Phosphonrolyhdic acid-picrosirius red.Semi-quantitative numerical estimates: O= negative; + = low: ++ = moderate; +++ = high.IIF = Indirect inummolluorescence.
' EC = Epithelioid cclls.PAS = Periodie acid-Schiff.
" CZ = Clear fone.SG = Sweat alands.
ing to the Ridley-Jopling classification ('^)and were distrusted among the leprosyspectrum as follows: borderline tuberculoid(BT) 14, borderline lepromatous (BL) 18,lepromatous (LL) 6. Biopsy specimens ofthe lesions were cut in half; one half wentthrough routine paraffin embedding, theother half was snap-frozen in liquid nitro-gen. Paraffin sections were stained withhematoxylin and eosin (H&E) and Wade(for acid-fast bacilli) staining; picrosirius(PS) ('`'), phosphomolybdic acid-picrosiriusred (PMA-PSR) (`), Gomori's reticulin(GR) ('') for collagen and reticular libers,respectively; periodic acid Schiff-alcianblue at pH 1 (PAS-AB pH 1.0) and at pH2.5 (PAS-AB pH 2.5) (") for sulfated andnon-sulfated proteoglycans, respectively;resorcin-fuchsin (RF) ( 4 ) for elastic fibers.
Five-µm-thick cryostat sections werethawed onto glass slides, fixed in acetone at4°C for 20 min, dried at room temperaturefor 5 min, and submitted to the followingstaining methods: PAS AB pH 1 and 2.5, re-sorcin-fuchsin staining and indirect im-munofluorescence (IIF) (`) for collagens(type 1 and III), fibronectin and tenascin. In-cubation of the sections with polyclonal anti-
human collagen I (diluted 1:40), rabbit anti-human collagen type III (diluted 1:40) andrabbit anti-fibronectin antibody (diluted 1:50)was followed by a fiuorescein-conjugatedgoat anti-rabbit immunoglobulin second anti-body (diluted 1:50) (DAKO, Glostrup, Den-mark) or fiuorescein-conjugated goat anti-mouse immunoglobulin (DAKO). Counter-staining with 1% Evans blue (Merk,Germany) for lO min was done. The slideswere mounted with buffered glycerin towhich 1% p-phenylenediamine (") (SigmaChemical Co., St. Louis, Missouri, U.S.A)was added. The secondary antibody wascontrolled by omitting the primary one andthe primary one by incubating the sectionswith antibodies of unrelated specilicity(anti-Factor VIII-related antigen).
The specimens were examined under aCarl Zeiss photomicroscope (Zeiss, Ger-many) and for confocal microscopy on alaser scanning microscope (LSM) 410 in-verted (Zeiss) with planapochromatic x40,1.3 NA, oil immersion objective. Antibod-ies labeled with Iuorescein were read on anargon/krypton-ion-laser at 488 nm, pinholeset in aperture 20 with a BP 510-525 emis-sion filter. PMA-PSR was examined with
26^
International Journal r►/ Lel►rn.S_v^ 1999
the same laster at 543 um with an LP 570emission fìlter. Images were obtained usingslow scanning and line Kalman averaging.The confocal scanning optical microscopywas not applied to generate three-dimen-sional (313) ima es but to remove out-of-fo-cus blur from non-confocal tluorescent mi-croscopy. This instrument, due to the confo-cal principie, allows optical tomography(optical sections without touching the ob-ject) information collection only out of uneplane and 3-D reconstruction of the col-lected sections es). The original digital im-ages were printed on a Codonics NT 1600printer.
RESULTSRemarkable alterations of the derma'
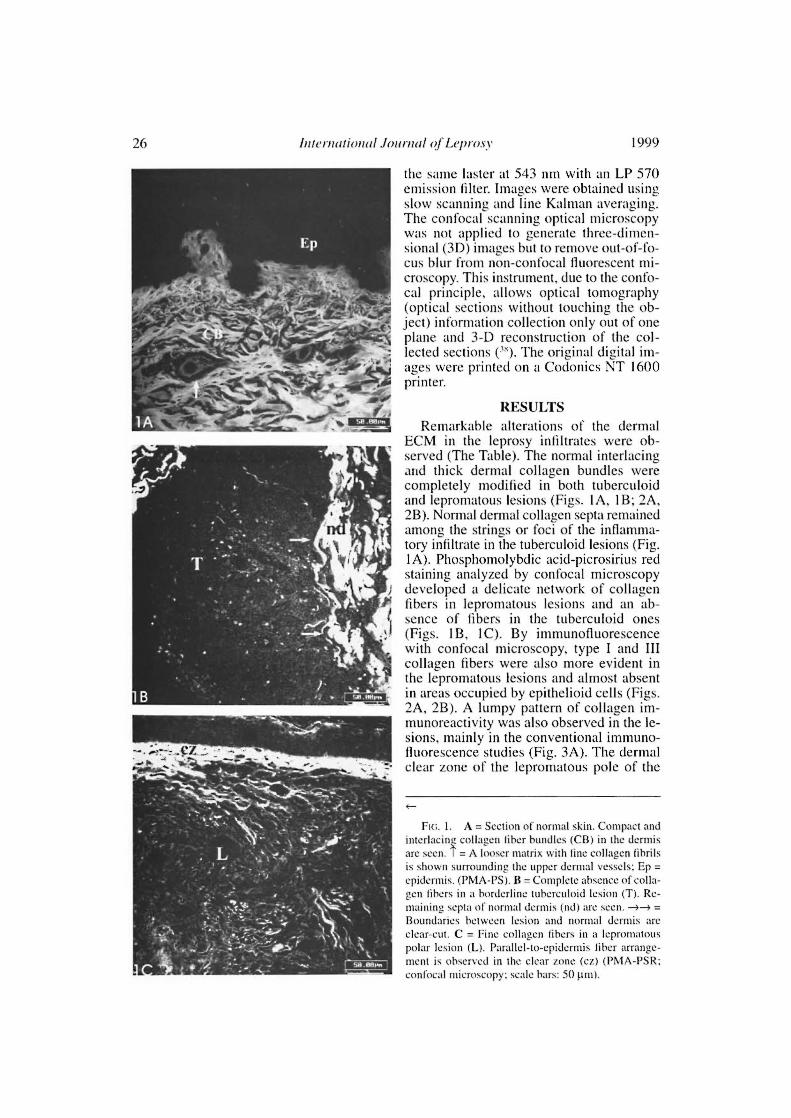
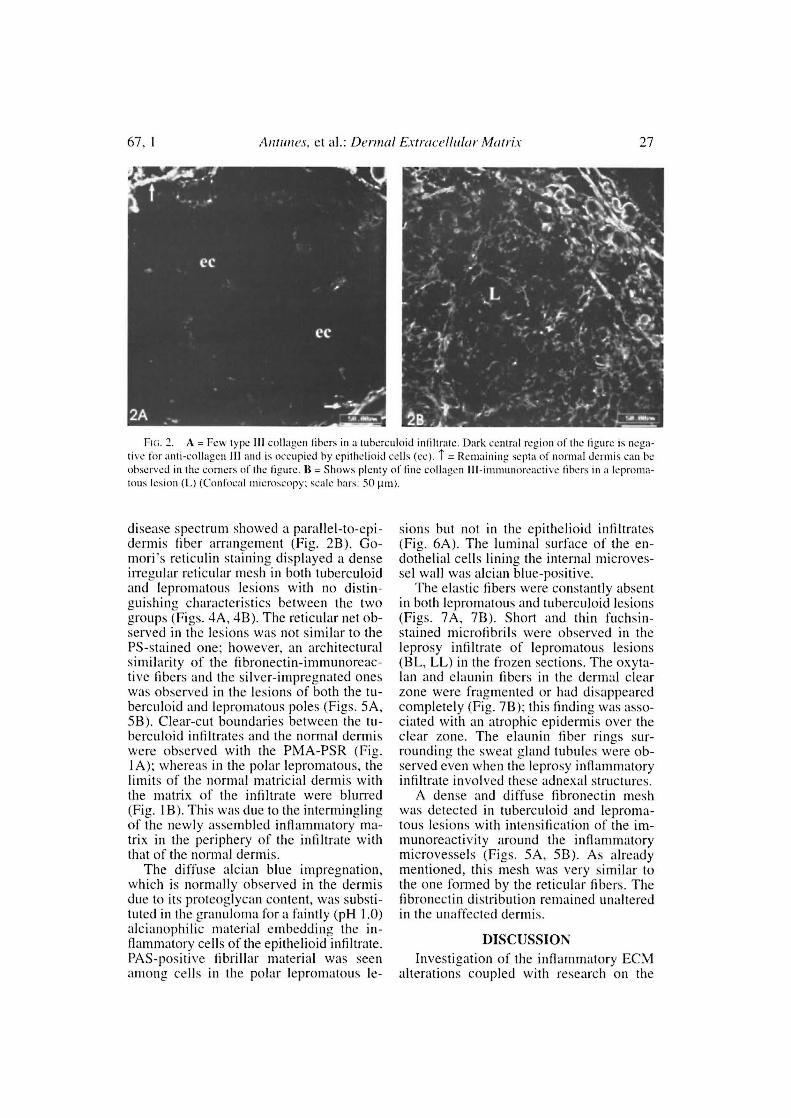
ECM in the leprosy intiltrates were ob-served (The Table). The normal interlacingand thick dermal collagen bundles werecompletely modified in both tuberculoidand lepromatous lesions (Figs. IA, 1B; 2A,2B). Normal dermal collagen septa remainedamong the strings or foci of the inflamma-tory infiltrate in the tuberculoid lesions (Fig.IA). Phosphomolybdic acid-picrosirius redstaining analyzed by confocal microscopydeveloped a delicate network of collagenfibers in lepromatous lesions and an ab-sence of fibers in the tuberculoid ones(Figs. 1B, 1C). By immunofluorescencewith confocal microscopy, type I and IIIcollagen fibers were also more evident inthe lepromatous lesions and almost absentin areas occupied by epithelioid cells (Figs.2A, 2B). A lumpy pattern of collagen im-munoreactivity was also observed in the le-sions, mainly in the conventional immuno-fluorescence studies (Fig. 3A). The dermalolear zone of the lepromatous pole of the
Fio. 1. A = Section of normal skin. Compact andinterlacing collagen liber bundles (CB) in the dermisare seco. T = A looser matriz with fine collagen tihrilsis shown surrounding the upper dermal vessels; Ep =epidermis. (I n1A-PS). B = Complete absence of colla-gen tìhers in a horderline tuherculoid lesion (T). Re-maining septa of normal derntis (nd) are seca. --s =Boundaries hctween lesion and normal derntis areolear-cut. C = Pine collagen fibers in a lepromatouspolar lesion (L). Partllel-to-epidermis tihcr arrange-ment is observed in the olear tone (cz) (PAMA-PSR;confocal nticroscopv; seale hars: 50 pai).
67, 1^Antunes, et al.: Derma! Ex!racellular Matrix^27
Ria. 2. A = Rew type III collagen libers in a tuberculoid infiltrate. Dark central region of the figure is nega-tive for anti-collagen III anel is occupied by epithelioid cells (cc). T = Remaining septa of normal dermis can beobserved in the corners of the figure. B = Shows plenty of fine collagen III-immunoreactive libers in a leproma-tous lesion (L) (Confocal microscopy; scale bars: 50 µm).
disease spectrum showed a parallel-to-epi-dermis fiber arrangement (Fig. 2B). Go-mori's reticulin staining displayed a denseirregular reticular mesh in both tuberculoidand lepromatous lesions with no distin-guishing characteristics between the twogroups (Figs. 4A, 4B). The reticular net ob-served in the lesions was not similar to thePS-stained one; however, an architecturalsimilarity of the fìbronectin-immunoreac-tive fibers and the silver-impregnated oneswas observed in the lesions of both the tu-berculoid and lepromatous poles (Figs. 5A,5B). Clear-cut boundaries between the tu-berculoid infiltrates and the normal dermiswere observed with the PMA-PSR (Fig.1A); whereas in the polar lepromatous, thelimits of the normal matricial dermis withthe matrix of the infiltrate were blurred(Fig. 1 B). This was due to the interminglingof the newly assembled inflammatory ma-trix in the periphery of the infiltrate withthat of the normal dermis.
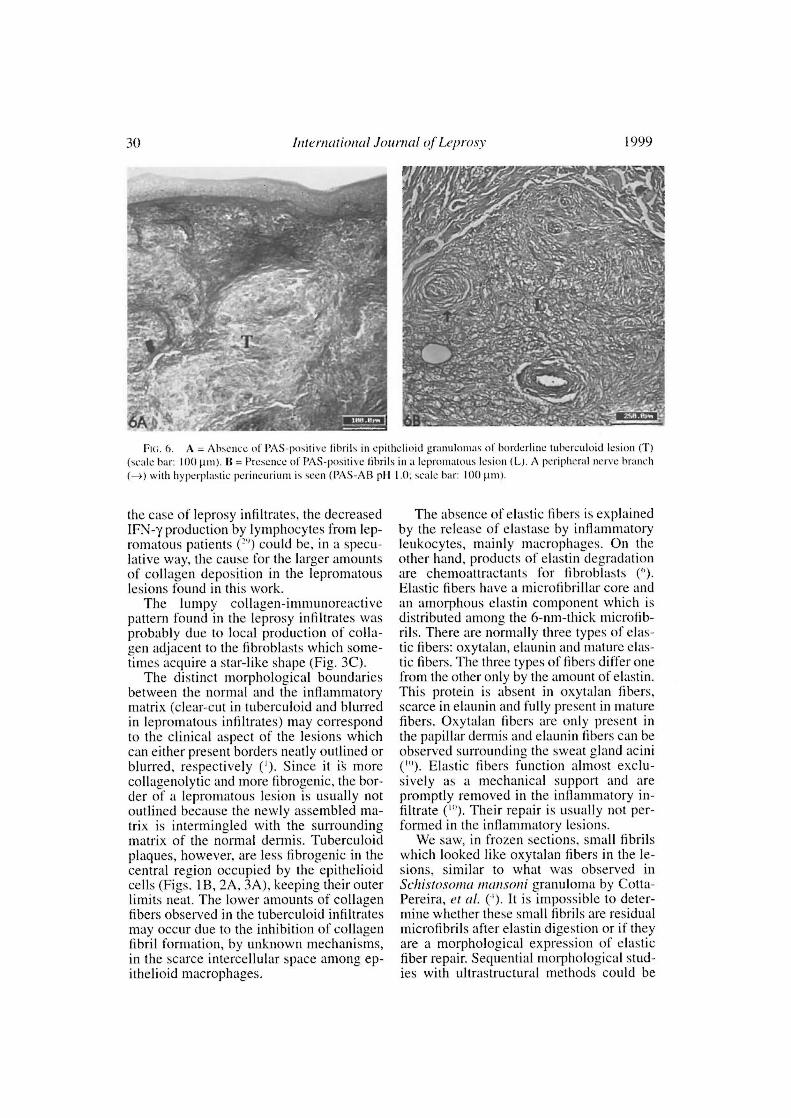
The diffuse alcian blue impregnation,which is normally observed in the dermisdue to its proteoglycan contem, was substi-tuted in the granuloma for a faintly (pH 1.0)alcianophilic material embedding the in-flammatory cells of the epithelioid infiltrate.PAS-positive fibrillar material was seenamong cells in the polar lepromatous le-
sions but not in the epithelioid infiltrares(Fig. 6A). The luminal surface of the en-dothelial cells lining the internai microves-sel wall was alcian blue-positive.
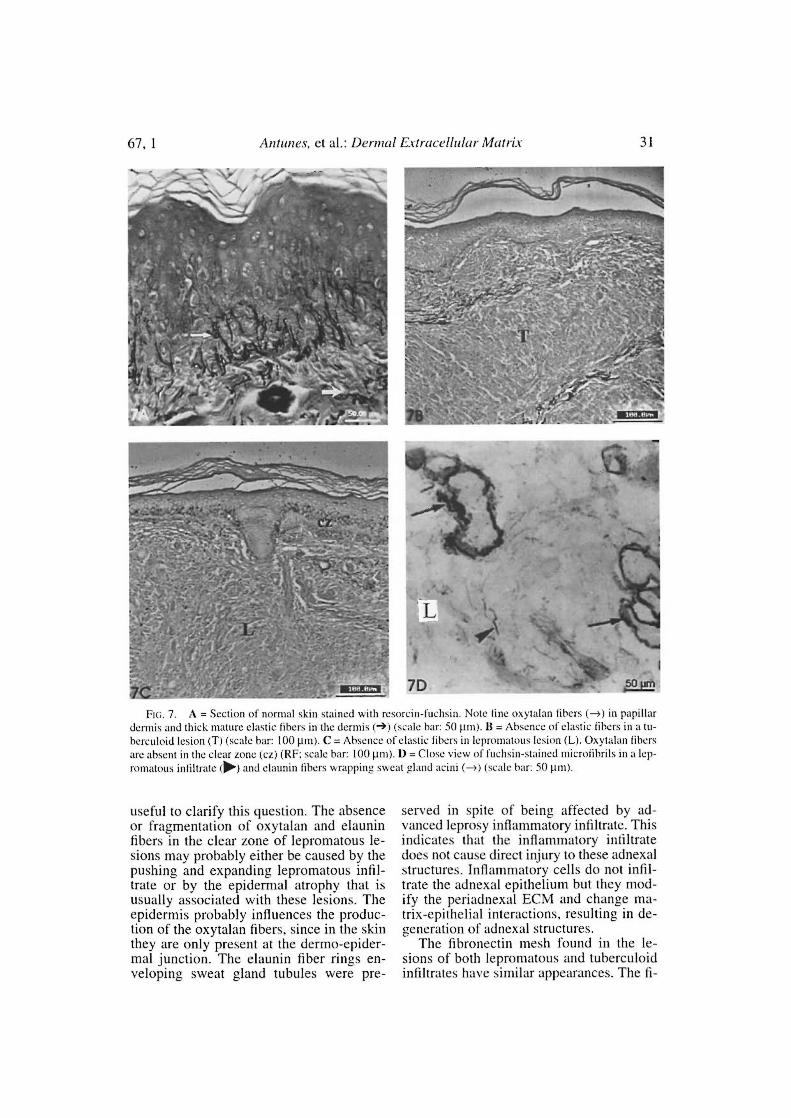
The elastic libers were constantly absentin both lepromatous and tuberculoid lesions(Figs. 7A, 7B). Short and thin fuchsin-stained microfibrils were observed in theleprosy infiltrate of lepromatous lesions(BL, LL) in the frozen sections. The oxyta-lan and elaunin fibers in the dermal clearzone were fragmented or had disappearedcompletely (Fig. 7B): this finding was asso-ciated with an atrophic epidermis over theclear zone. The elaunin fiber rings sur-rounding the sweat gland tubules were ob-served even when the leprosy inflammatoryinfiltrate involved these adnexal structures.
A dense and diffuse fibronectin meshwas detected in tuberculoid and leproma-tons lesions with intensification of the im-munoreactivity around the inflammatorymicrovessels (Figs. 5A, 5B). As alreadymentioned, this mesh was very similar tothe one formed by the reticular fibers. Thefibronectin distribution remained unalteredin the unaffected dermis.
DISCUSSIONInvestigation of the inflammatory ECM
alterations coupled with research on the
19992^ Internation al10111/1(1 1 n/' Leprosv
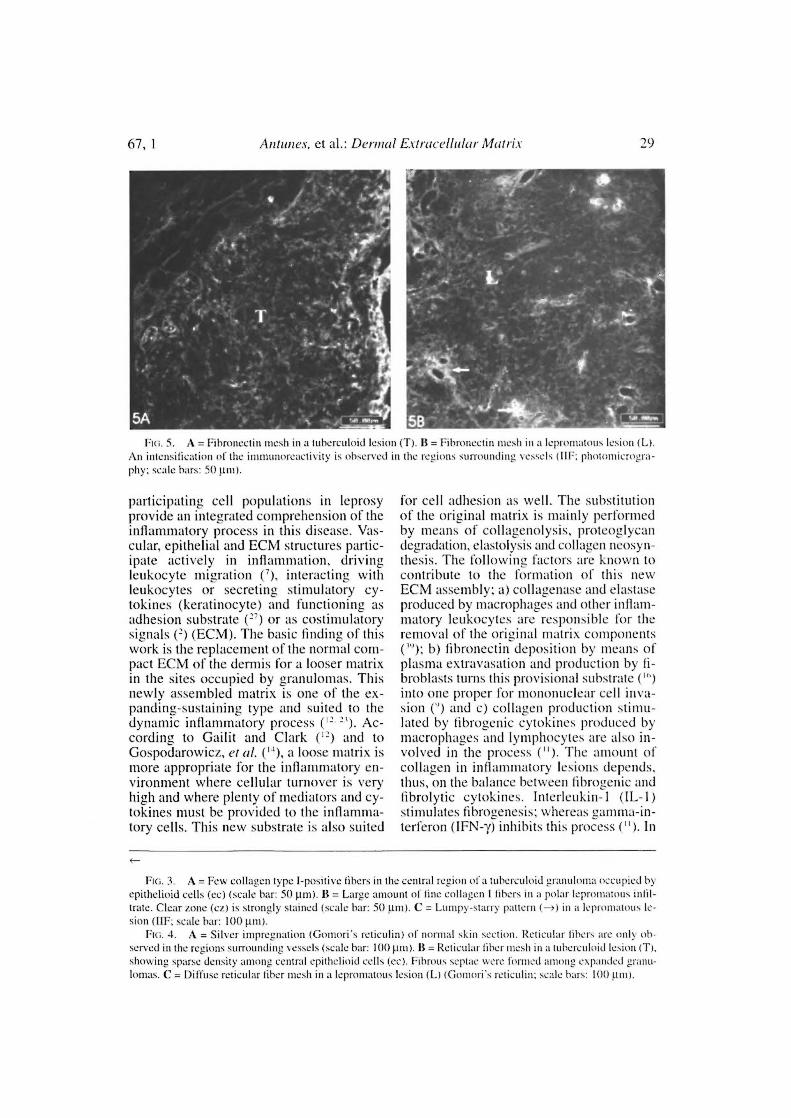
67, 1^Antunes, et al.: Derma[ Eviraceiluiar Mairix^2c)
Fui. 5. A = Fibruuectin mesh in a tubcrculoid lesion (T). 11= Fibrunectin mesh in a Icpromatous lesion (L).\n intensification of the inuuunorcactivity is observed in the regions surrouncling vcsscls I11F: photonticrogra-
phy: scale bars: 50 µ111).
participating cell populations in leprosyprovido an integrated cotnprehension of theinflanunatory process in this disease. Vas-cular, epithelial and ECM structures partic-ipate actively in inflammation, drivingleukocyte migration ('), interacting withleukocytes or secreting Stimulatory cy-tokines (keratinocyte) anel functioning asadhesion substrate ('') or as costimulatorysignals ( 2 ) (ECM). The basic finding of thiswork is the replacement of the normal com-pact ECM of the dormis for a looser matrixin the cites occupied by granulomas. Thisnewly assembled matriz is one of the ex-panding-sustaining type and suited to thedynamic inflammatory process ( 12 2 i). Ac-cording to Gailit and Clark ( 12 ) and toGospodarowicz, et al. ( 11 ), a loose matrix ismore appropriate for the inflaunmatory en-vironment where cellular turnover is veryhigh and wltere plenty of mediators and cy-tokines must be provided to the inflamma-tory cells. This new substrate is also suited
for cell adhesion as well. The substitutionof the original matrix is mainly perlormedby means of collagenolysis, proteoglycandegradation, elastolysis and collagen neosyn-thesis. The following factors are known tocontribute to the formation of this newECM assembly: a) collagenase and elastaseproduced by macrophages and other inflam-matory leukocytes are responsible for theremova! of the original matriz components("): b) fibronectin deposition by means ofplasma extravasation and production by fi-broblasts turns Chis provisional substrateroto one proper for mononuclear cell inva-sion (") and c) collagen production stimu-lated by fibrogenic cytokines produced bymacrophages and lymphocytes are also in-volved in the process ("). The antount ofcollagen in inflammatory lesions depeneis,thus, on the balance between lihrogenic andtibrolytic cytokines. Interleukin-1 (IL-1)stinudates librogenesis: whereas gamma-in-terferon (IFN-y) inhibits Chis process ("). In
Fie. 3. A = Fev.: collagen type I-positive libers in the central regam of a tuberculoid granuloma occupied byepithelioid edis (cc) (scale bar: 50 pint. II = Large amount of fine collagen 1 libers in a polar leproniatous infìltrate. Clear zonc (cz) is strongly stained (scale bar: 50 pit). C = Lumpy-starry panem (-) in a Icpromatous le-sion (110: scale bar: 100 )1m).
Fui. 4. A = Silvcr impreguation (Gomori's reticulin) of normal skin sectiou. Reticular libers are only oh-scrved in the regions surrouncling vessels (scalc bar: 100 µ11t). B = Reticular liber mesh in a tuberculoid [estou (T),showing sparse .lensity among central cpithelioid cens (cc). Fibrous septae were formed :mtong expandcil granu-tomas. C = Diffuse reticular liber mesh in a leprouuous lesion (L) (Gomori's reticulin: scale bars: 100 (tnt).
30^ International Jeunial of. Leprosy^ 1999
Fui. 6. A = Absence of PAS-positive fibrils in epithelioid granulomas of borderline tuberculoid lesion (T)(scale bar: 100µm). 13 = Presence of PAS-positive lihrils in a lepromatous lesion (L). A periplicral nerve brandi(-) with hyperplastic perincurium is seco (PAS-AB p11 1.0; scale bar: 100 µm).
the case of leprosy infiltrates, the decreasedIFN-y production by lymphocytes from lep-romatous patients ("') could be, in a specu-lative way, the cause for the larger amountsof collagen deposition in the lepromatouslesions found in this work.
The lumpy collagen-immunoreactivepattern found in the leprosy infiltrates wasprobably due to local production of colla-gen adjacent to the fibroblasts which some-times acquire a siar-like chape (Fig. 3C).
The distinct morphological boundariesbetween the normal and the inflammatorymatrix (clear-cut in tuberculoid and blurredin lepromatous infiltrates) may correspondto the clinicai aspect of the lesions whichcan either present borders neatly outlined orblurred, respectively ('). Since it i's morecollagenolytic and more fibrogenic, the bor-der of a lepromatous lesion is usually notoutlined because the newly assembled ma-trix is intermingled with the surroundingmatrix of the normal dermis. Tuberculoidplaques, however, are less fibrogenic in thecentral region occupied by the epithelioidcells (Figs. 1 B, 2A, 3A), keeping their outerlimits neat. The lower amounts of collagenfibers observed in the tuberculoid infiltratesmay occur due to the inhibition of collagenfibril formation, by unknown mechanisms,in the scarce intercellular space among ep-ithelioid macrophages.
The absence of elastic fibers is explainedby the release of elastase by inflammatoryleukocytes, mainly macrophages. On theother hand, products of elastin degradationare chemoattractants for fibroblasts (`').Elastic fibers have a microfibrillar core andan amorphous elastin component which isdistributed among the 6-nm-thick microfib-rils. There are normally three types of elas-tic fibers: oxytalan, elaunin and mature elas-tic fibers. The three types of fibers differ onefrom the other only by the amount of elastin.This protein is absent in oxytalan fibers,scarce in elaunin and fully present in maturefibers. Oxytalan fibers are only present inthe papillar dermis and elaunin fibers can beobserved surrounding the sweat gland acini("'). Elastic fibers function almost exclu-sively as a mechanical support and arepromptly removed in the inflammatory in-filtrate ( 1 "). Their repair is usually not per-formed in the inflammatory lesions.
We saw, in frozen sections, small fibrilswhich looked like oxytalan fibers in the le-sions, similar to what was observed inSchistesonra mincDoI granuloma by Cotta-Pereira, et al. (4). It is impossible to deter-mine whether these small fibrils are residualmicrofìbrils after elastin digestion or if theyare a morphological expression of elasticfiber repair. Sequencial morphological stud-ies with ultrastructural methods could be
67, 1^Antunes, et al.: Dernral E.vtracellular Mutrix^31
Fio. 7. A = Section of normal skin stained with resorcin-fuchsin. Note tine oxytalan tibers (-) in papillardormis and thick matute elastic fibers in the dormis (->) (scale bar: 50 pm). II = Absence of elastic libers in a tu-herculoid lesion (T) (scale bar: 10011m). C = Absence of elastic libers in lepromatous lesion (L). Oxytalan libersare ahsent in the clear tone (cz) (RF; scale bar: 100 µm). 1) = Close view of fuchsin-stained microtibrils in a lep-romatous infiltrate (li>.) and elaunin libers wrappine sweat gland acini (-) (scale bar: 50 pnt).
useful to clarify this question. The absenceor fragmentation of oxytalan and elauninfibers in the clear zone of lepromatous le-sions may probably either be caused by thepushing and expanding lepromatous infil-trate or by the epidermal atrophy that isusually associated with these lesions. Theepidermis probably influences the produc-tion of the oxytalan fibers, lince in the skinthey are only present at the dermo-epider-mal junction. The elaunin fiber rings en-veloping sweat gland tubules were pre-
served in spite of being affected by ad-vanced leprosy inflammatory infiltrate. Thisindicates that the inflammatory infiltratedoes not cause direct injury to these adnexalstructures. Inflammatory cells do not infil-trate the adnexal epithelium but they mod-ify the periadnexal ECM and change ma-trix-epithelial interactions, resulting in de-generation of adnexal structures.
The fibronectin mesh found in the le-sions of both lepromatous and tuberculoidinfiltrates have similar appearances. The fi-

32^ International Journal u/ Lcprosv^ 1999
bronectin immunoreactive pattern of bothtuberculoid and lepromatous lesions aresimilar to the silver-impregnated reticularmesh. No difference was observed in thereticular architectural pattern in the tuber-culoid and lepromatous lesions as well, ex-cept that reticular fibers were scarce amongthe central epithelioid cells of the tubercu-loid granulomas ( 2"). According to Wolmanand Kasten ( 4"), fibronectin could be a con-stituent of the reticular fibers. PAS-positivefibrillar material in the more organized po-lar lepromatous lesions may also corre-spond to fibronectin inflammatory mesh.Fibronectin is a multifunctional componentof the loose connective tissue suited for thedynamic interactions of inflammation. It isalso associated with opsonization ("•' 4 ),and with cell-matrix interaction by meansof integrin mediation ( 1 '). Mvcobactc'riumlepras presents a fibronectin-attachmentprotein (FAP) which is involved in the in-ternalization of the bacteria by Schwanncells and epithelial cells ("'). Antibodies tothe fibronectin binding 85 kDa ABC anti-genic complex of M. leprae were found tobe raised in lepromatous sera ("). This anti-gen complex can also elicit a Th 1 responseon the lymphocytes of leprosy contacts ( 22 ).Parti and Hõcik (") have used the acronymMSCRAMMs (microbial surface compo-nents recognizing adhesive matrix mole-cules) to identify the family of adhesinswhich recognize and bind amino-acid se-quences of matrix components. Diversebacterial tissue tropism and virulence canoccur because of specific MSCRAMMs.
In addition, the incidence of reactionalepisodes or changes in the activity of theleprosy lesion could, in a speculative way,be associated with changes in either the fi-bronectin or proteoglycan composition ofthe inflammatory matrix. These two com-ponents are closely associated with hapto-taxis and cell migration, respectively, whichundoubtedly occur in the early stages ofleprosy reaction.
The remarkable ECM alterations seen incutaneous leprosy lesions involve collagen,proteoglycans, elastic fibers and fibronec-tin. Although distinct cell populations areobserved in the inflammatory infiltrates cor-responding to the different forms of diseasepresentation, no striking differences on
ECM components were found between thetuberculoid and lepromatous goles insidethe lesions. Therefore, ECM cannot be anadditional aid parameter in the classifica-tion of leprosy. The striking ECM modifica-tions caused by the leprosy inflammatoryprocess affect the biological properties ofthe skin and, additionally, contribute to theclinicai manifestations of the disease.
SUMMARYThirty-eight biopsies of cutaneous lesions
from leprosy patients Iborderline tubercu-loid (BT) 14, borderline lepromatous (BL)18, lepromatous (LL) 6] were processed forstaining of some extracellular matrix (ECM)components (collagen, proteoglycans, elas-tic fibers and fibronectin). Specific histolog-ical staining and the indirect immunofluo-rescence method with antibodies to collagenand fibronectin were utilized. The ECM ofthe normal dermis was strikingly modifiedin the inflammatory infiltrate. By Gomori'sreticulin and anti-fibronectin immunostain-ing, replacement of the desse interlaced col-lagen fibers with a reticular mesh was ob-served in the infiltrate. The immunoreactiv-ity obtained with anti-type I and anti-type IIIcollagens showed positive fibrils and alumpy pattern in the lepromatous and tuber-culoid lesions with a higher amount in thelepromatous lesions. The lack of clear-cutboundaries between the normal dermis andthe inflammatory infiltrate in the leproma-tous (BL, LL) lesions was correlated withthe blurred limits of the clinicai lesions ofChis pole of the leprosy spectrum. Absenceof elastic fibers in the infiltrate was a con-stant finding, and fuchsin-positive microfib-rils were found in some infiltrates. The clearzone of lepromatous lesions was devoid ofoxytalan fibers. Elaunin fiber rings aroundsweat gland acini were present even whenthe leprosy infiltrate was seen envelopingthem. The original ECM is replaced by anewly assembled one, which is suited forthe dynamic nature of the inflammatory pro-cess. The trophic effects of the ECM uponthe cutaneous epithelial structures are modi-fied so that atrophy and late degenerationensues. These ECM modifications con-tribute, therefore, to the biological alter-ations of the skin functions in leprosy.

67, 1^Antunes, et t1.: Dernral Ertt•ac ellular Mcttri.r^33
RESUMENSe procesaron 38 biopsias de lesiones cutáneas de
pacientes con lepra (14 con lepra tuberculoide suhpo-lar, BT; 18 con lepra Iepromatosa suhpolar, BL; 6 conlepra Iepromatosa, LL) para la tinción de algunos com-ponentes de la matriz extracelular (CME): colágena,proteoglicanas, libras elásticas y libronectina. Se uti-lizaron tinciones histológicas específicas y Ia inmuno-
fluorescencia indirecta con anticucrpos contra colá-gena y fibronectina. Los CME dc la dermis normal re-sultaron ntarcadentente modificados en cl infiltradoinflamatorio. La tinción para reticulina de Gomori y lainmunotincion para libronectina, revelaron en el infil-trado la substitución de las libras de colágena densa-mente entrelazadas, por una trama de libras reticulares.La inntunoreactividad obtenida con anticucrpos anti-colágena tipo I y colágene tipo 111 mostró librillas pos-itivas y un patrón moteado en las lesiones lepro-matosas y tuberculoides, con predomínio en las le-siones lepromatosas. La falta de limites definidos entrela dermis normal y cl infiltrado inflamatorio en las le-siones lepromatosas (BL y LL) correlacionó con loslimites difusos de las lesiones clínicas observadas eneste polod de la enfermedad. La ausencia de libraselásticas en cl infiltrado fite wt hallazgo constante; en al-gunos infiltrados se encontraron uticrofibrillas fuchsina-positivas. La zune clara de las lesiones lepromatosas es-tuvo libre de libras de oxitalan. Los anillos de libras deelaunina alrededor de los acini de las glándulas sudorí-paras fueron evidentes a u n coando estuvieron envuel-tos por los infiltrados de la lepra. Los camhios en loscomponentes de la matriz extracelular son indicativosde lo dinástico del proceso inflamatorio. Los efectostróficos de los CME sobre las estructuras dc los epite-lios cutáneos son de tal magnitud que provocan laatrofia y la degeneración tardia de la piel. Estas modi-ficaciones en los CME tamhién contrihuycn a las al-teraciones en las functiones de la piei en la lepra.
RÉSUMÉ'frente huit hiopsies de lésions cutanécs provenant
de patients lépreux [ 14 tuberculoides horderlines (BT),18 lépromateux horderlines (BL), 6 lépromateux (LL)]furent étudiées pour la coloration des éléments de lamatrice extra-ccllulaire (MEC) tels que le col1agène,les protéoglycannes, les libres élastiques et la fìbronec-tine. Des colorations spécifiques et la méthode d'im-nwnofluorescence indirecte n partir d'anticorps anti-collagène et anti-fibronectine ont été utilisés pour cetteétude. L'infiltrat inflammatoire modifiait de façon frap-pante la MEC, telle que présente dans le derme nor-mal. Les libres denses et entrecroisées de collagèneétaient remplacées, dans les infiltrats, par un réseau delibres visihlcs par la coloration de Ia réticuline selouGomori et positives par inuminontarquage spécifiquede la fibronectine. Des fibrilles positives par i n tmmno-marquage par des anticorps anti-collagèae de type 1 etde type III, préscntaient un aspect en amas irrégulicrs
dans les lésions tant lépromateuses que tuberculoides,étant toutefois plus abondantes chez les premières. Iln'y avait pas de limites hien délinies entre le dermenormal et I'infiltrat inllanunatoire dans les lésionslépromateuses (BL, LL), ce qui corrélait bien avec leslimites indistinctes des lésions cliniques caractérisantce pOle du spectre de la lèpre. Il n'y avait pas de libresélastiques dans 1'infiltrat et ce, de façon constante. Desmicro-fibri1les positives à la fuchsine furem trouvéesdans certains infiltrais. La zune claire des lésionslépromateuses ne presentait pas de libres oxytalanes.Des anneaux de libres élaunines autour des acini deglandes sudoripares étaient détectables mcme lorsque1'infiltrat lépreux les enveloppait cotnplètement. LaMEC pré-existante est remplaçée par une matrice néo-formée, qui est adaptéc au caractère três dynamique duprocessus inflammatoire. Les actions trophiques de laMEC sur les structures cutanécs épithéliales sont mod-ifiées de tellc ntaniere qu'il s'cnsuit atrophie etdégénération de ces dernières. Ces modifications de laNIEC contribuem de ce fait ìt 1'altération des fonctionscutanées observées au cours de la Ièpre.
REFERENCES1. BttvcrsoN, A. and PrAt_ rrGRArr, R. F. Symptoms
and signs. In: Leprnsy.. Edinhurgh: Churchill Liv-ingstone, 1990, pp. 25-56.
2. CtANO, A. C., SALOVON, D. R., WADswoRTtt, S.,laxo, M. P., MoiciR, C. F., Orco, S., StuazACtt, E.
M. and Cot.to vN, J. E. a,(3,, a,(3, integrins medi-ate lantinin/merosin binding and function as co-stimulatory molecules for human thymocyte pro-liferation. J. Inummol. 154 (1995) 500-510.
3. Coo's, A. 11. Fluorescull antihody tnethods. In:General Cvtochentical dletlzods, Vol. I. Danielli,J. F.. cd. New York: Academie Press, 1958, pp.399-422.
4. CorrA-PEREIRA, G., LiNZI, H., Ot.tvtatt 1, D. N.,V1'Aist:NZoNi. J. R. and LtNZm, J. A. On the pres-ence of clastic microlibrils in tiver aranuloma ofmurine Sclti.stosomiasi.c^Mem. Inst. Os-waldo Cruz 86 Suppl. 111 (1991) 129-130.
5. DA Stt.vA, L. C., Mot nAo, P. A. and Boi:ott:vu', R.Patterns of sulfatcd glycosantinoglycan synthesisand accuntulation in hepatic grancdontas induccdhy schistosontal infcction. Exp. Mol. Pathol. 50(1989) 411-420.
6. DAvntsox, J. M. and Gnzo, M. G. Control ofelastin synthesis. In: Regulation c,¡ Alai i.r Acc•u-mulation. Orlando: Acadentie Press, Inc., 1986,w 178-217.
7. Dttcos, L. and KAItN, A. Adhésion cellulaire.Med. Sei. 3 (1987) 314-315.
8. Dourai:, P. C. and StAcn, M. S. Conventional andcoofocal ftuorescencc microscopy of collagenlibers in the heart. J. 1listochent. Cytochcm. 41(1993) 465-469.
9. DvoIAK, 11. F. Tumor wounds that do not tical. N.Eng1. J. Atcd. 315 (1986) 1650-1659.

34^ lnt('rnulion(tl .l (carnal (r/ I epros v^ 1999
10. FRANZIti.At , C. and FARIs, B. Elastin. In: ('cll /ti-olotu of . the Evirtu ellularNlutrrc. I lay, E. 1)., ed.New York, Plcnum Press, 1981, p. 65.
11. FRruNi)I.R li, i3., 13owAI.ASta, J. S., Nii i.soti, E.and J■rr:NI:/, S. A. Rcgulatiou of fihrohlast prolif-cration and collagen synthcsis hy cytokincs. !nt-ntunol. Today 7 (1986) 303-307.
12. GAn rr, J. and Ct.ARK, R. A. F. Wound repair inthe contcxt of extracellular matrix. Curr. Opi n .Cell Biol. 6 (1994) 717-725.
13. GoRDoN, M. Y., RILEY, G. P., WArr, S. M. and
GRIAVIS, M. F. Compartmunalization of a henw-poictic growth factor (GM-CSI) hy glycosai]nno-glycans in the honc marrow nticrocnvironmcnt.Nature 326 (1987) 403-405.
14. Gosl'ooARowRi, D., GRIa.NItDRC, G. and l3nt0-w'Ia.i., C. R. Detcrniination of ('('!!alar chape byextraccllular matrix and its correlation with thecontrol of cellular growth. Canccr Res. 38 (1978)4155-4171.
15. Gltiui.i:v, M. F. Stains for connective tissue. In:Manual of I1isrola ,gie and Special Stuiuir, Teeh-nies. New York: McGraw Ili!!. 1960, pp. 55-96.
16. GRI\1At:0, J. A., Boitos, D. L., TAKI\A, C.,MATiu:w, R. C. and E\1oNA130, II. Col!atzcn iso-types, laminin, and fibronectin in granulomas ofthe livcr and intcstincs of Sc histosonru marimari-infected mico. Am. J. Trop. Mcd. Hyg. 37 (1987)335-344.
17. I IAY, E. 1). Co!lagcn and cmhryonic developntcnt.In: Cell Biologv a/ Erhrarellnlar Mairix. I lay, E.D., cd. New York: Plcnum Press, 1982, pp.379-410.
18. HYNES, R. O. Integrins: a family of ccll surfaccreceptors. Cell 48 (1987) 549-554.
19. Jr a)tnttty, L. C. U., BuoNof.As, and BRtNTttit. R.R. Picrossirius staining plus polarization mi-croscopy, a spccific method for collagen dctcctionin tissuc sections. I listochem. J. 11 (1979)447-455.
20. KIIANOLKAR, V. R. Pathology in Icprosy. In: Lep-ru.t.v in Theor.v and Prac tic e. Cochranc. R. G., andDavey, T. F., cds. Bristol: John Wright. 1964, pp.125-151.
21. KLEIN, O. The extraccllular matrix of thehematopoictic microenvironment. Expericntia 51(1995) 914-926.
22. LAunois, P., N'Di:wu, M. N., CARITA., J. L.,MANE. 1.. DROwART, A., VAN VooRnN, J. P., SAR-Tifou, J. L. and Ht vcr:N, K. Fihronectin-bindineantigen 85 and the 10-kilodalton GroES-rclatcdheat shock protela are the prcdominant Th-1 re-sponse induccrs in Icprosy contacts. Infcct. Im-mun. 63 (1995) 88-93.
23. LE:Nri, H. L., CAflctt, V. L. G., Mn .:ui, M., SANO,
A., NISItt\n R:\ , K. and Bt Rclat, E. Fibrosis pat-tcrns of lesions devcloped hy athymic and eu-thymic mico infecicd with l'aracoc cidiodesbrasiliensis. Braz. Mcd. Biol. Res. 27 (1994)2301-2308.
24. LINDkit, E., 1.110 o, V. P., SrtNNLw, S., LINDI:vISF,
K., B1onvATN, 13. and 131RCNvtsr, R. Circulatingantibodies to conncctivc tissuc microfihrils anddermal imnwno,clohulin dcposits in Icprosy. Clin.Inun u nol. Inununopathol. 13 (1979) 1-8.
25. M(Ao.\s1, K. I'. W. J., Fuor.Nuku(i, II. II. andhlit ti \la.t, I). Antihodics to collagen in paticntswith Icprosy. Clin. In, nunol. Immunopathol. 9(1978) 16-21.
26. NARAYANAN, R. 13., Bntl:NI, L. K., Sii.toi,, A.K. and NAru, I. Fihroncctin in Icprosy lesions;observations using nwnoclonal antibodies to htt-tuan lihroncctin. Indian J. Lepr. 56 (1984)532-539.
27. Ni:w(iw r:N, 1). 'l'he role of the cxtraccllular matrixin the control of fletira! crest ccl! nugration. In: Ex-frocellaar Mairi.t. I lawkcs, S. and Wam" J. L., cds.Ncw York: Acadentie Press, 1982, pp. 141-146.
28. Nistnsn RA, M., Assai, M., Hav:tsut, M., TAhA-zotio, I., 'l',vN:tk:s, Y., KouoA, H. and Uo iti;, II.Extraccllular matrix in hcpatic granulomas ofmico infccicx l with Schi.tio.corna nrun.coni; qualita-tive and yt:mtitative analysis. Arch. Palitai. Lab.Med. 109 (19$5) 813-8I8.
29. Noat'It ntA, N., KAPI.AN, G., Lovv, E., SARNO, E.N., KtSIINifR, I., GRANI a .f.I-PmERN°, A., Vn:ntA,L., (30m.0, V. C., Lix is, W., SiiaNVANN, R., Ym,Y. K. and ColIN, Z. A. Detective y-inicr!cron pro-duction in Icprosy. J. Exp. Mcd. 158 (1983)2165-2170.
30. Oi.os, O. R., GRIITIN, A. and KRrsINA, T. F. Dy-namics of collagen accunmlations and polymor-phisnt in nwrinc Srhi.tto.'orrw japonicunr. Gas-trocntcrology 89 (19$5) 617-662.
3!. PArri, J. M. and Ilittlh, M. Microbial adhcsinsrccognizing extraccllular matrix macronwlccules.Curr. Opin. Cell Biol. 6 (1994) 752-758.
32. Pussot.ANt, M. C., PERALTA, J. M., Rt su \NEK, F.D., Go\u:s, II. M., MARQuEs, M. A., ALMEIDA, E.C., SAAD, M. II. and SARNO, E. N. Serology andlcprosy: inun u noassays comparing inun u noglobu-1w O antihody responses to 28- and 30-kilodaltonprotcins purificcl front Mveobucteriunr boiis BCG.J. Clin. Microbiol. 29 (1991) 2 2 85-22 90.
33. PI.Arr, J. L. and rs1ic ii,\iu., A. F. Rctardation offading and enh:tnccntcnt of intcnsity of in ununo-fluoresccncc by p-phcnilcncdiamine. J. Histochent.Cytochcnt. 157 (1983) 1339-1353.
34. PRoc -ioR, R. A., Pitr:NUu:RCAsr, E. and MosnoR, K.F. Opsonization of bactcria hy lihroncctin. 131ood59 (1982) 651-687.
35. Rini.r.v, D. S. and Jot'I.itia, W. II. Classification ofIcprosy according to inununity; a fivc-group sys-tem. Int. J. Lcpr. 34 (1966) 255-273.
36. Scnoui,v, J. S., Li, Q., M(Cot:R .r, D. W., Boxc-MASrr:h, M., CLARK-C'rirnss, J. E., R:crt.mr, T. L.and Biiowx, E. J. A ,tivc•obacterium lepra(' genecncoding a lihroncctin hinding protcin is uscd forcflicicnt invasion of cpithclial cells and Schwann
edis. Infcct. Inu n tin. 63 (1995) 2652-2657.

67, 1^Antunes, et al.: Derma! Ertracellular Matrix^ 35
37. Sir:r:ii , D. C. and 1Ii AI< ii,\h, M. S. M. Carbo-hydrates. In: Theorv aiul Pructiee n`Histotechnol-
og . and cd. Shechan, D. C., and Hrapchak, M. S.M., eds. St. Louis: The C. U. Moshy Company,1980, pp. 159-179.
38. SHEPAito, C. J. R. and SnorroN, D. M. Con%ocal
Laser Scanning Micro.vcopv. New York: Rios Sci-entilie Publishers and Springer Verlag, 1997, pp.1-10.
39. TAKAuAsln, S., DUNN, M. A. and SHt rIR, S.Liver collagenase in murine schistosomiasis. Gas-troenterology 78 (1980) 1425-1431.
40. WUI VAN, M. and KAsrr:N, P. Polarized light mi-croscopy in the study of the molecular structuresof collagen and reticulin. Histochemistry 85(1986) 41-49.





![Scanned by CamScanner › PDFFolder › LA2019-20 › la20 › Delhi-2020-pai… · I"iARYETA fiSVrftNru{EruT GEN MREL A#fu{ i|\IiSTR&T}#I\i SEPAHTi\lrNT N\,$CITIFICATISru l\': J$/il]i2s*5-3GSl](https://img.pdfslide.us/doc/110x75/5f0e3f267e708231d43e50e2/scanned-by-camscanner-a-pdffolder-a-la2019-20-a-la20-a-delhi-2020-pai.jpg)














![The Sun. (New York, N.Y.) 1900-08-18 [p 2]....wtoi iJDd rg nt lrnt-h bajr 1I ThIrinftthr-Jlmt hat dr h friendsbed 4oitts-PIon tht wotnn thI Pot aZd that nede n AW1 CArlT rnthawt setter](https://img.pdfslide.us/doc/110x75/5f6ea9e1f841f019553968e4/the-sun-new-york-ny-1900-08-18-p-2-wtoi-ijdd-rg-nt-lrnt-h-bajr-1i-thirinftthr-jlmt.jpg)








