Embed Size (px)
Citation preview

Vitamin C inhibits the enzymatic activity of Streptococcus
pneumoniae hyaluronate lyase
Songlin Li1, Kenneth B. Taylor2, Stephen J. Kelly1, and Mark J. Jedrzejas1*
1Department of Microbiology, 2Department of Biochemistry and Molecular Genetics
University of Alabama at Birmingham, Birmingham, AL 35294, USA
Running title: Hyaluronate lyase complexed with vitamin C
* To whom correspondence should be addressed at
Department of Microbiology
933 19th Street South, 545 CHSB-19
The University of Alabama at Birmingham
Birmingham, Alabama 35294-2041, USA
Tel./fax: (205)975-7627/(205)975-5424
email: [email protected]
Hyaluronate lyase complexed with vitamin C
1
Copyright 2001 by The American Society for Biochemistry and Molecular Biology, Inc.
JBC Papers in Press. Published on January 12, 2001 as Manuscript M011102200 by guest on M
ay 4, 2019http://w
ww
.jbc.org/D
ownloaded from

Summary
Enzyme activity measurement showed that L-ascorbic acid (vitamin C, Vc)
competitively inhibits the hyaluronan degradation by Streptococcus pneumoniae
hyaluronate lyase. The complex crystal structure of this enzyme with Vc was determined
at 2.0 Å resolution (PDB ID code: 1f9g). One Vc molecule was found to bind to the active
site of the enzyme. The Vc carboxyl group provides the negative charges that lead the
molecule into the highly positively charged cleft of the enzyme. The Vc ring system
forms hydrophobic interactions with the side chain of Trp292 which is one of the
aromatic patch residues of this enzyme responsible for the selection of the cleavage sites
on the substrate chain. The binding of Vc inhibits the substrate binding at HA1, HA2, and
HA3 catalytic positions. The high concentration of Vc in human tissues likely provides a
low level of natural resistance to the pneumococcal invasion. This is the first time that Vc
inhibition directly on the bacterial “spreading factor” was reported and Vc is also the
first chemical that had experimentally shown to have inhibitory effect on bacterial
hyaluronate lyase. These studies also highlight the possible structural requirement for the
design of a stronger inhibitor of bacterial hyaluronate lyase.
Keywords: Streptococcus pneumoniae, hyaluronate lyase, vitamin C, enzymatic activity
inhibition, antibacterial activity, protein-inhibitor complex
Hyaluronate lyase complexed with vitamin C
2
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Introduction
L-ascorbic acid, also known as vitamin C (Vc1), is synthesized in plants and
almost all animals except primates, guinea pig, Indian fruit bat, and some insects. Vc is
necessary in the diet of these animals and usually exists in significantly large
concentrations in their tissues (1). Prolonged lack of Vc in the diet of humans results in
scurvy, characterized by the skin lesions, blood vessel fragility, and poor wound healing.
Less severe deficiency of Vc produces alterations in connective tissue structure and may
also cause decreased resistance to some infections (2). Vc is a multi-functional molecule
in tissues. It usually acts as antioxidant (3), free radical scavenger (4), neuroprotectant
and neuromodulator (5). It also plays an important physiological function in activating
peptide hormones (2) and regulating cell division and growth (6). Vc is the single
synthetic chemical manufactured and consumed in the greatest amount in the world (1).
Although the importance of Vc in the normal function of animal tissues has long been
known, the detailed molecular basis of Vc action, especially the mechanisms of its
interactions with proteins and enzymes, are still largely unknown.
Protein-ligand interaction is an important aspect of modern biochemistry. It
provides information for the understanding of the essence of the molecular interactions,
enzyme action mechanism, protein activity control, and of the drug design. In regards to
the important function of Vc in so many life processes, the protein-Vc interfaces have
________________________1 Vc: vitamin C, L-ascorbic acid. SpnHL: Streptococcus pneumoniae hyaluronate lyase. GbsHL: S. agalactiae hyaluronate lyase.
Hyaluronate lyase complexed with vitamin C
3
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

not been fully characterized to date. The structural basis of the protein-Vc interface and
the possible influence of Vc on enzyme activities are certainly issues of significant
interest for the understanding of the functions and principles of Vc action. The protein-
Vc interaction was first seen in the crystal structure of D-xylose isomerase (PDB ID
code: 1xid) where Vc is present between two tryptophan residues (7). It clearly
emphasizes the importance of hydrophobic interactions in the protein-Vc interface. Here
we present the crystal structure of Streptococcus pneumoniae hyaluronate lyase (SpnHL)
cocrystallized with Vc which provides another opportunity to examine the protein-Vc
interactions at the atomic level.
S. pneumoniae is a Gram-positive bacterial pathogen which causes pneumonia,
bacteremia, meningitis, sinusitis, and otitis media in humans world wide, especially in
neonates and children, and often leads to significant rates of mortality and morbidity. It
secretes hyaluronate lyase to catalyze the degradation of hyaluronan (HA), one of the
main components of connective tissues in animals, to expose tissue cells to bacterial
toxins. Therefore, SpnHL is also being called “spreading factor” (8,9). The inhibition of
hyaluronate lyase is expected to reduce the spreading of this pathogen in the most early
stages of the pneumococcal invasion. The action mechanism of hyaluronan degradation
by SpnHL revealed recently provides a unique opportunity to target hyaluronate lyase in
the prevention of the pneumococcal invasion. The Vc complexed crystal structure of
SpnHL is an attempt on this line of research.
The enzymatic activity of SpnHL at the presence of various concentrations of Vc
were measured. The crystal structure of this enzyme cocrystallized with Vc was
determined at 2.0 Å resolution. The structural basis of the inhibitory effect of Vc on
Hyaluronate lyase complexed with vitamin C
4
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

SpnHL enzymatic activity was established.
Experimental Procedures
Crystallization and data collection L-Ascorbic acid (Sigma) was cocrystallized with
the S. pneumoniae hyaluronate lyase at conditions similar to the native SpnHL
crystallization condition with additional 10 to 100 mM ascorbic acid. A dataset at 2.0 Å
resolution was collected at 100 K using one crystal at 50 mM ascorbic acid, synchrotron
radiation and cryocooling technique at conditions similar to the native crystal (10, 11).
Diffraction data were processed and scaled using HKL program package (12). The crystal
belongs to the orthorhombic space group P212121 with cell parameter a = 84.264 Å, b =
102.666 Å, c = 103.253 Å. The data set is 90.1% complete (60% in the last shell) with
Rsym 0.087.
Structure refinement and validation Coordinates and B-factors of the protein part of
the native SpnHL crystal structure (8) were used directly as primary model in the
SpnHL-Vc complex structure refinement. All waters and solution molecules were
omitted from the model. X-plor package (13) was used to refine the structure against
54,217 reflections at 2.0 to 45 Å resolution range (87.8% completeness) and 1%
reflections were used in Rfree calculation to monitor the refinement progress and the
model improvement (14). Rigid body, position, and simulated annealing (3000 K)
protocols were employed. The model was manually fitted into the electron density maps
on graphics using program O (15) between each round of refinement calculations. The
Hyaluronate lyase complexed with vitamin C
5
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

electron densities for Vc were observed from the beginning, but Vc structure was only
included into the structural refinement until the R factor dropped below 25% and water
molecules were incorporated thereafter. And only after this point the B-factors
refinement was introduced.
Microplate assay of SpnHL activity The SpnHL enzymatic activity was measured
using a modification of previously described protocols using either cetylpyridinium
chloride (16) or cetrimide (17). Compounds assayed for inhibitory effects on the SpnHL
enzyme activity were ascorbic acid, epinephrine, apigenin, salicylic acid, and histamine
(Sigma).
Volumes of 90 µl of inhibitor solutions at concentrations of 1, 2, 4, 6, 8, 10, 12,
14, 16, 18, and 20 mM containing 50 mM sodium acetate and 10 mM calcium chloride at
pH 6.0 (apigenin was dissolved in 10% DMSO water solution) were added along one row
of a 96 well microtiter plate, leaving the first row as blank control. 10 µl of the wild-type
SpnHL at a concentration of 3.3 µg/ml was added to each well and incubated at room
temperature for 1 hour. Reactions using 40 units of bovine hyaluronidase (Sigma) at the
same conditions were run in parallel. 25 µl of 1 mg/ml HA were added to each well to
start the reaction which was proceeded at room temperature for 15 minutes. Undigested
HA was precipitated using freshly made 50 µl of 10 % (w/v) aqueous cetylpyridinium
chloride with additional 2.0% NaOH to stop the enzyme activity. The absorbance was
measured at 630 nm for cetylpyridinium chloride using an automated microplate reader
(EL808; Bio-Tek Instruments, Inc., Winooski, VT). A higher absorbance reading
corresponds to a higher remaining substrate HA concentration and, therefore, lower
enzyme activity. The O.D. 595 absorbance readings were then converted to percent
Hyaluronate lyase complexed with vitamin C
6
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

inhibition by subtracting the O.D. 595 value for the pure enzyme without inhibitor from
the reading with inhibitor divided by the difference between the 100% inhibition (average
values at 12.8 mM, 14.4 mM, and 16.0 mM ascorbic acid) and 0% inhibition (O.D. 595
reading for the pure enzyme).
To determine if this inhibition is reversible, a sample of SpnHL enzyme activity
was measured and designated as 100% activity. After dialyzing the sample against 20
mM Vc in buffer, the enzyme activity dropped to 0%. The sample was then dialyzed
against the reaction buffer to remove the Vc, and 94% of the enzyme activity was
recovered. Therefore, this inhibition is reversible. The slightly drop in activity was caused
by the volume changes during dialysis.
The reaction initial velocity was measured in quadruplicate using a modified
microplate assay (16). Initial hyaluronan concentrations were 0.2, 0.1, 0.07, 0.05 and
0.04 mg/ml in 50 mM acetate buffer at pH 6.0 and 10 mM calcium chloride and 8%
agarose at 55 ∞C. 100 µl of each of the 5 concentrations of the hyaluronan-agarose gel
were added to the microplate columns in quadruplicate and left at room temperature for
an hour to set. Solutions of SpnHL (0.33 µg/ml) with 0, 1, 5, 10 mM Vc in 10 mM
calcium chloride and 50 mM acetate buffer at pH 6.0 were pre-incubated at 37 ∞C for an
hour and 100 µl of each enzyme preparations were added to the first four lines of each
column of the microplate. Reaction buffer was added to the remaining four lines of the
microplate as control. The plates were then incubated at 37 ∞C for 14 hours. The enzyme
samples were removed and each well was washed three times with the buffer. Each well
was then filled with 100 µl of 2% aqueous cetrimide (hexadecyltrimethylammonium
bromide) and incubated at 37 ∞C for two hours. The absorbance was measured at 405 nm
Hyaluronate lyase complexed with vitamin C
7
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

using an automated microplate reader (EL808; Bio-Tek Instruments, Inc., Winooski,
VT).
Data deposition Structural factors and coordinates of SpnHL-Vc complex have been
deposited in the Brookhaven Protein Data Bank with accession number 1f9g.
Results
Overall structure of SpnHL-Vc complex The crystal structure of S. pneumoniae
hyaluronate lyase in complex with Vc (SpnHL-Vc complex) was determined at 2.0 Å
resolution. In total, 725 out of 731 residues were modeled and 303 waters were
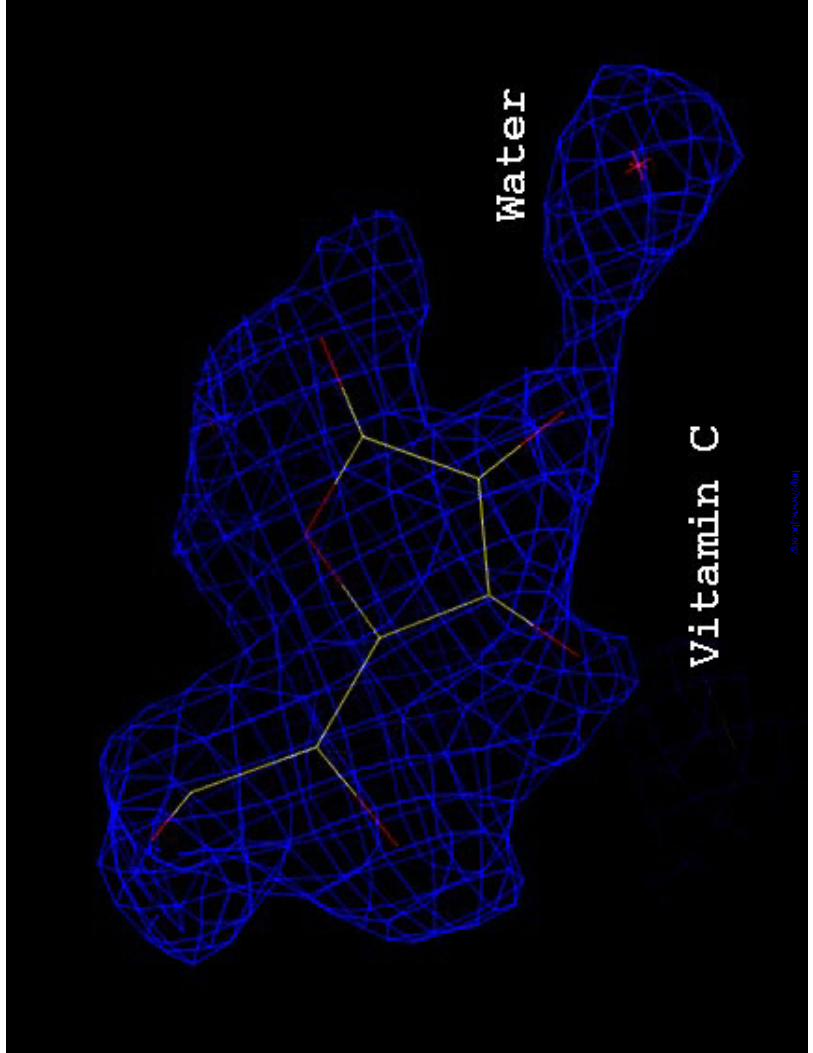
incorporated. One Vc molecule was clearly seen in the electron density map (Figure 1).
The final crystallographic R factor is 0.208 and Rfree 0.253. The protein part of the
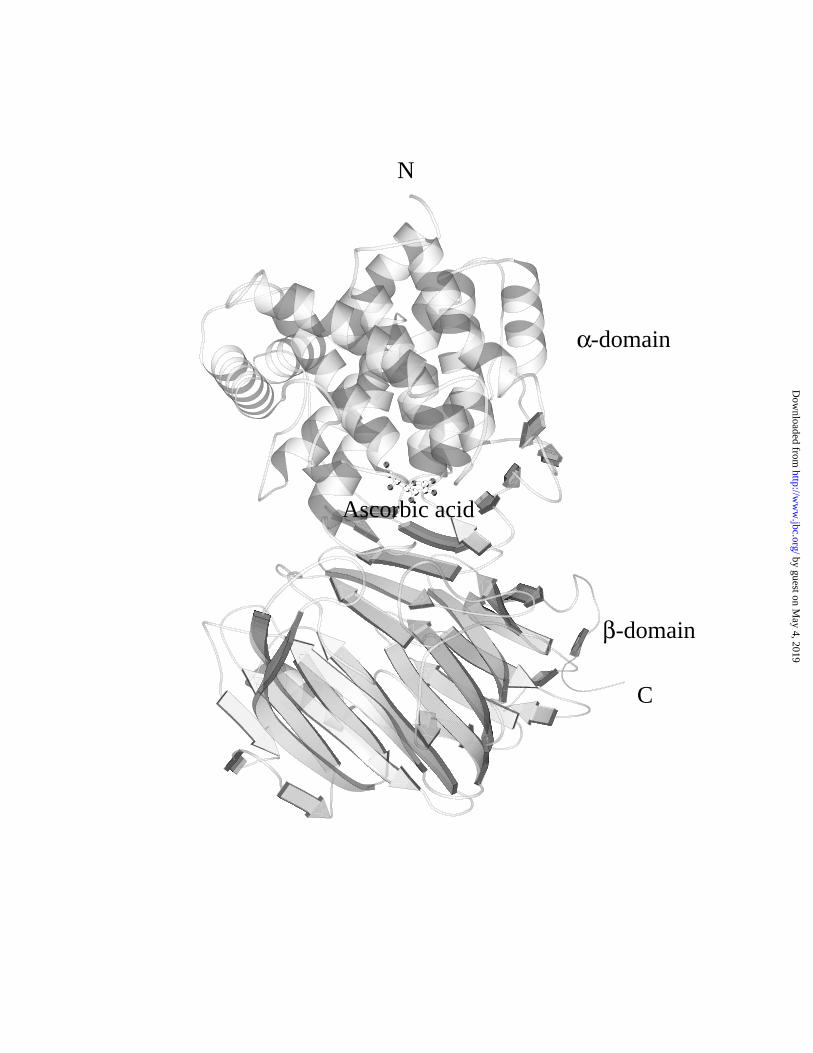
complex structure contains an N-terminal α-domain and a C-terminal β-domain
connected by a ten residue linker. The active site of this enzyme lies in the middle of the
molecule where a predominant cleft is formed between these two structural domains
(Figure 2). The cleft is about 30∞10 Å in dimension, enough to accommodate three
disaccharide units of the hyaluronan substrate chain simultaneously, which were named
HA1, HA2, and HA3 respectively from the reducing end to the non-reducing end of the
hyaluronan chain (19). The active site is located at one end of the cleft, corresponding to
the reducing end of the bound hyaluronan chain and is composed of two parts, an
aromatic patch responsible for the cleavage site selection on the substrate chain and a
catalytic group responsible for the cleavage of the β-1,4 glycosidic linkage between HA1
Hyaluronate lyase complexed with vitamin C
8
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

and HA2 disaccharide units in the hyaluronan chain (8, 19, 20).
The protein part of the SpnHL-Vc complex structure has only slight changes
when compared to the native SpnHL crystal structure. The r.m.s. deviation is only 0.538 Å
for all protein atoms. In the cleft region, all corresponding atoms in the protein-Vc
complex structure are located at the outer side, leaving the cleft about 0.2 Å wider than it
is in the native SpnHL structure (8). The position of Vc relative to the active site residues
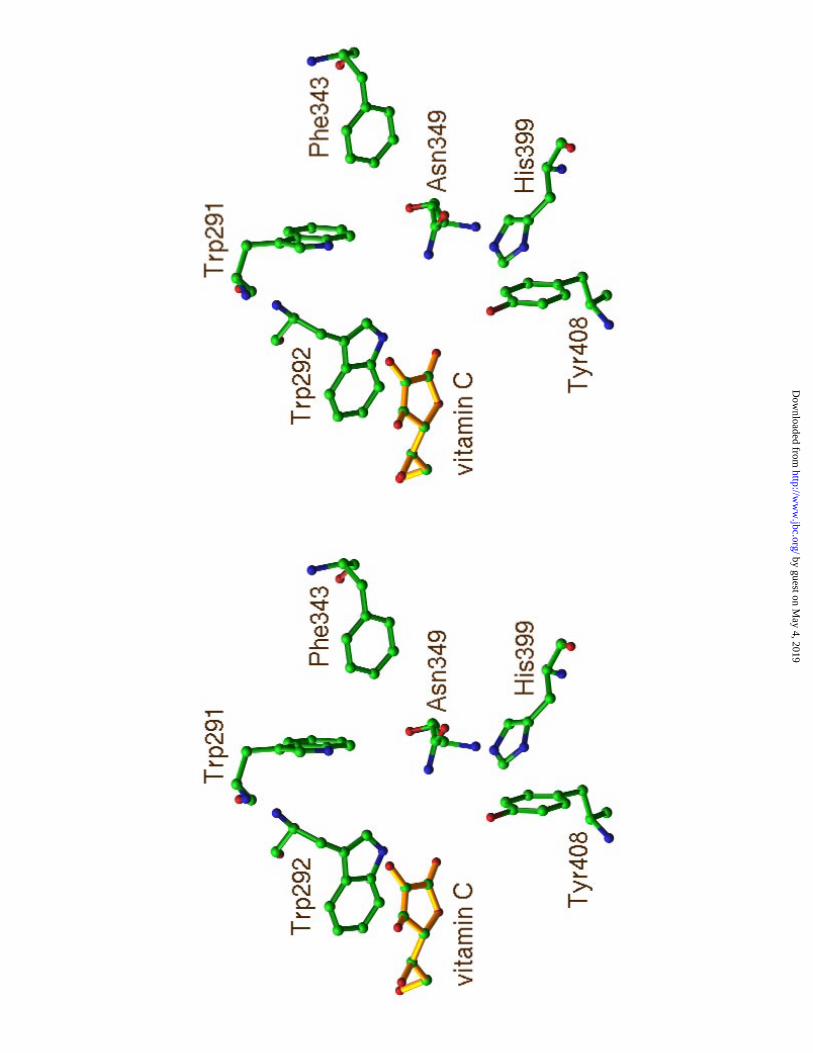
was shown in Figure 3a.
The aromatic patch of the enzyme active center is composed of three aromatic
residues, Trp291, Trp292, and Phe343 (8) (Figure 3a). The side chains of Trp292 and
Phe343 form hydrophobic interactions with the hydrophobic patches on the hyaluronan
chain. Through this matching, the cleavage sites are selected. Trp292 hydrophobically
interacts with HA2 disaccharide unit and accurately anchor HA2 into catalytic position.
In the hyaluronan PAD degradation model (19), the enzyme catalyses the degradation of
the β-1,4 glycosidic linkage between HA1 and HA2 and produces 4,5-unsaturated HA1.
Vc in the SpnHL-Vc complex structure is found to bind to Trp292 (Figure 3a) indole
group and occupies the HA2 position.
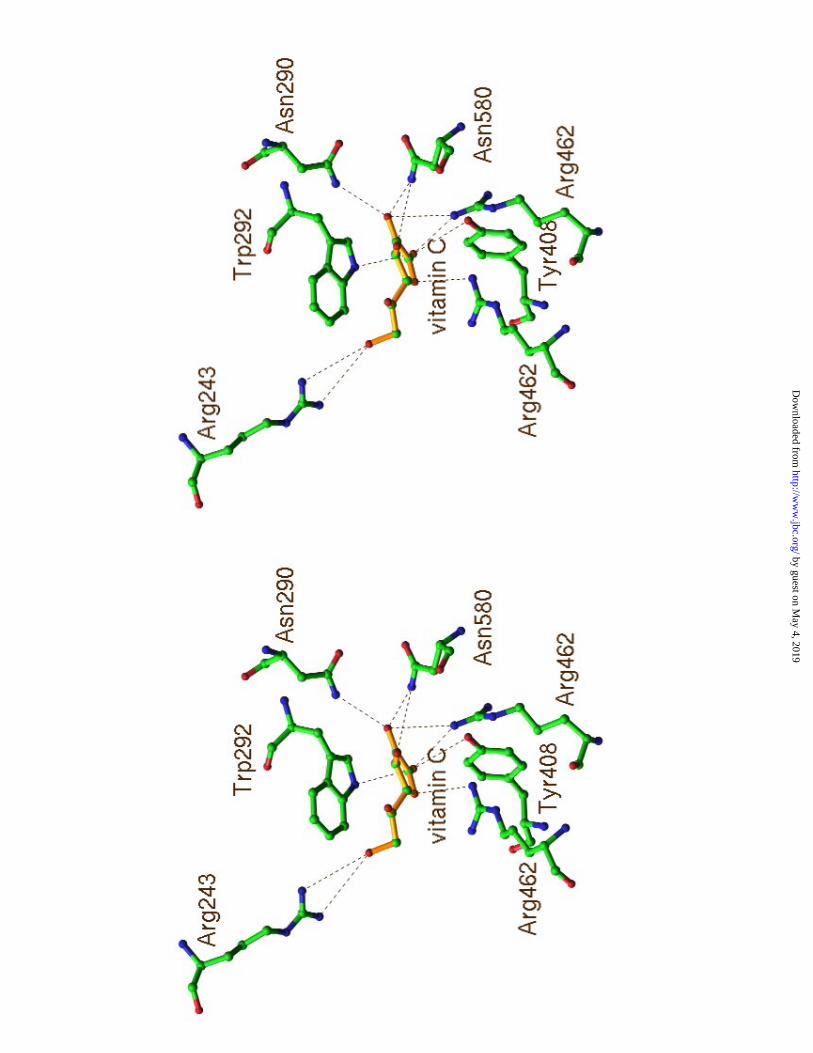
The binding of Vc to SpnHL The bound Vc forms 25 interactions with 7 residues of
the enzyme (Table 1). These residues were also shown to interact with the substrate (8).
The relative positions of these residues to the bound Vc were shown in Figure 3b. Five
out of the seven interface residues, Arg243, Asn290, Trp292, Tyr408, and Asn580, were
extensively studied in our previous biochemical and structural studies and were shown to
play important roles in the normal function of this enzyme (8, 20, 21).
Trp292 forms 8 interactions with Vc which accounts for the most interactions
Hyaluronate lyase complexed with vitamin C
9
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

among all these 7 residues in the protein-Vc interface. The indole group of Trp292 is in
parallel to the five member ring of Vc (Figure 3a). This structural arrangement provides
the main hydrophobic interaction that stabilizes the Vc molecule inside the cleft. Trp292
is one of the aromatic patch residues and is responsible for the selection of the cleavage
sites on the substrate chain. The binding of Vc, therefore, likely competes with the
binding of hyaluronan substrate at the HA2 position which is located at the middle of the
cleft.
Tyr408 is one of the three key catalytic residues in the hyaluronan degradation. It
donates one proton to the glycosidic oxygen to break the β-1,4 glycosidic linkage
between HA1 and HA2 disaccharide units (8). In the complex structure, Tyr408 forms
one salt bridge with the Vc O1 oxygen (refer to Figure 4 for Vc atom labelings).
Therefore, both the aromatic patch and the catalytic group of the active center of SpnHL
are involved in the binding of Vc. The binding of Vc blocks both the aromatic patch and
the catalytic group.
Residues Asn290 and Asn580 form the narrowest part across the cleft. Asn580 is
the only residue from the β-domain which is involved in the SpnHL-substrate and the
SpnHL-Vc interface. The mutation N580G causes a small increase (about 15%) in the
enzyme activity because the wider cleft opening allows for easier substrate entry (8, 21).
SpnHL-Vc complex structure showed that Vc is also in contact with Asn580 and
Asn290.
The cleft is highly positively charged by the accumulation of lysine and arginine
residues inside the cleft. There are 9 conserved arginine residues present in the cleft.
Three of them are involved in the interaction with Vc. Arg462 and Arg466 are
Hyaluronate lyase complexed with vitamin C
10
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

extensively involved in Vc binding (Table 1). Arg243 forms one salt bridge with Vc
carboxyl group. When the substrate is bound into the cleft, Arg243 interacts with HA2
and HA3 disaccharide units and with the oxygen atom of the second glycosidic linkage
which is suspected to be the next glycosidic linkage to be degraded. Mutation R243V
surprisingly decreased the enzyme activity by 33% (8). In our previous studies, we
proposed that Arg243 plays an important role in the substrate translocation after the
initial glycosidic bond is degraded (9, 19). The glycosidic linkage in contact with Arg243
is the next linkage to be degraded. The interaction with Arg243 suggest that the Vc
binding also inhibits substrate binding at HA3 position. It was recently reported that one
arginine residue is believed to be involved in two Vc binding sites in the Vc-peroxidase
complex structure (22). The presence of one or more arginine residues is likely one of the
characteristic features in the protein-Vc interface.
The 25 interactions in the SpnHL-Vc interface (Table 1) can be classified into
two groups, hydrophobic and ionic interactions. Trp292 contributes mostly to the
hydrophobic interactions with Vc. Three arginine residues, Arg243, Arg462, and Arg466
form several salt bridges, while Tyr408, Asn290, and Asn580 form hydrogen bonds with
the ligand. In comparison with the protein-Vc interface observed in the D-xylose
isomerase where hydrophobic interactions play the dominant role, hydrophobic and ionic
interactions are almost equally important in the SpnHL-Vc interface. Therefore, the
SpnHL-Vc interface represents a novel type of protein-Vc interface.
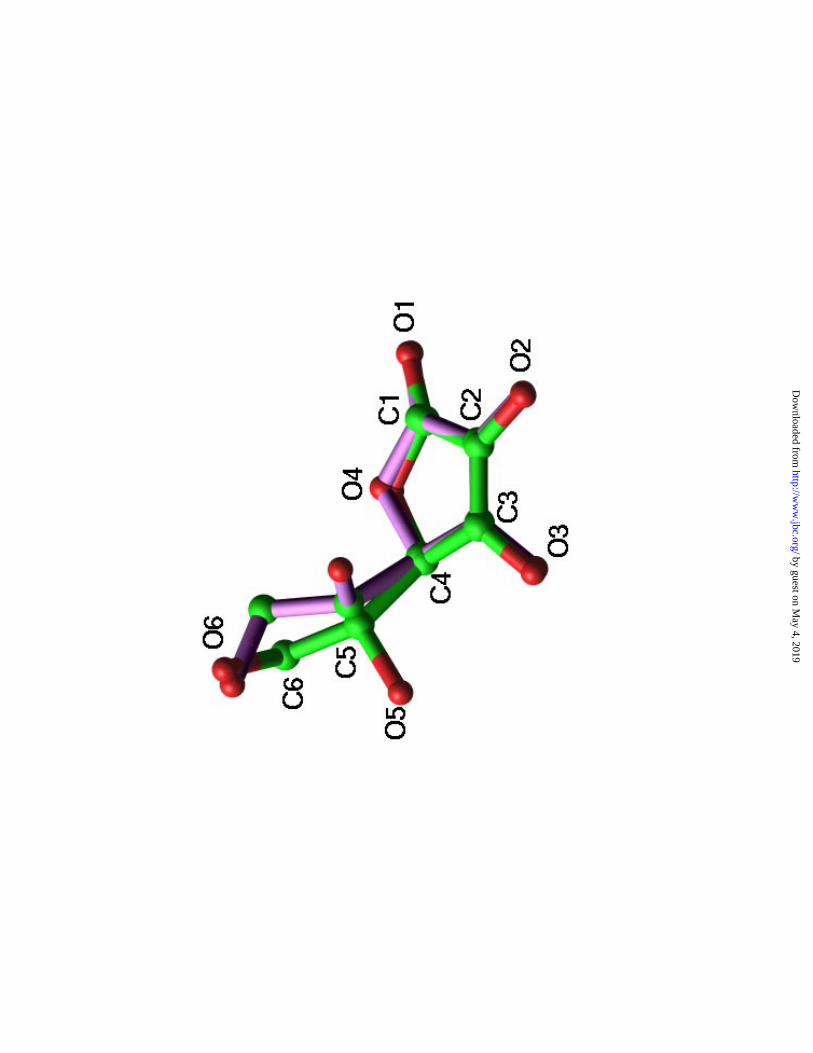
The protein-Vc interactions cause minor structural changes at both parts of the
interface. In comparison with the native SpnHL structure, the side chain displacement of
these interface residues are very small. On the contrary, Vc itself has relatively significant
Hyaluronate lyase complexed with vitamin C
11
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

structural changes compared to its native crystal structure (23) used for the modeling and
refinement. The carboxyl group is forced to bend towards the plane of the Vc ring (Figure
4). The negative charges caused by the carboxyl group at physiological condition are
apparently important in leading the Vc molecule into the highly positively charged cleft.
The actual binding geometry shows that the five member ring of the Vc molecule
provides the most hydrophobic binding interface, whereas the carboxyl group interferes
with the Trp292 indole group which is not in favor of the Vc binding.
Discussion
Inhibitory effect of Vc on SpnHL enzyme activity Vitamin C, salicylate, and
flavonoids had been reported to have certain inhibitory effects on the enzyme activity of
hyaluronidases (18) which are a group of hydrolases employed by mammals for the
hyaluronan degradation. Bacteria usually produce hyaluronate lyases to degrade
hyaluronan. The search for the inhibitors of bacterial hyaluronate lyases was started from
these chemicals. Our activity measurements clearly showed that Vc inhibits the
hyaluronan degradation by SpnHL. Vc is structurally similar to one of the sugar units of
hyaluronan, the main substrate of SpnHL. Hyaluronan is composed of linear repeats of
the disaccharide unit β-1,4-glucuronic-β-1,3-glucosamine. One of the main
components of hyaluronan, glucuronic acid, is also the precursor in the Vc biosynthesis.
Therefore, Vc can be regarded as a substrate analogue of hyaluronate lyase.
The effects of these compounds on the enzyme activity of SpnHL and bovine
Hyaluronate lyase complexed with vitamin C
12
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

hyaluronidase were investigated and measured by the microplate enzyme activity essay.
The results showed that none of these compounds had any significant influence on the
activity of bovine hyaluronidase at our experimental condition (data not shown).
However, Vc inhibited the SpnHL activity (Figure 5). At our experimental condition, the
IC50 of this inhibition is about 5.8 mM.
The initial velocity of the degradation at various concentrations of hyaluronan was
measured in the presence of three concentrations of Vc and in its absence. The results
were fit to both the competitive model and to the noncompetitive model by nonlinear
regression. The fitting attempts with the noncompetitive model repeatedly resulted in an
unreasonably high value (>50,000 mM) for the parameter Ki, which is not present in the
competitive model. Since this parameter indicates the concentration of Vc required for
the binding to the enzyme-substrate complex, it is apparent from this analysis that Vc
binds only to the free enzyme (Ki = 53 mM), not to the enzyme-substrate complex in the
experiments described here. Therefore, the substrate competes successfully for the
binding of Vc and this inhibition is competitive.
Physiological significance The degradation of hyaluronan in the host connective
tissues is an important step in the pneumococcal invasion. The bacterial strains that
produce more hyaluronate lyase were shown to be more virulant than those strains
producing less (24). S. pneumoniae strains with hyaluronate lyase and cell toxin
pneumolysin double mutations showed significant additive attenuation in virulence (25).
Therefore, the inhibition of hyaluronate lyase activity is likely important in the control of
the pneumococcal invasion. And since animals usually use hydrolases to degrade
Hyaluronate lyase complexed with vitamin C
13
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

hyaluronan, pneumococcal hyaluronate lyase becomes a potential target for developing
novel antibacterial agent. Vc is the first chemical shown to have inhibition effect on the
activity of this pneumococcal enzyme.
The adult minimum daily requirement for Vc is approximately 10 mg. Vc exists
in human tissues at the level of 0.2 mM to over 10 mM concentrations and has an
unusually varied distribution compared with other vitamins (26, 27). Large
concentrations of Vc were detected in the adrenal gland and the aqueous humor of the
eye. Human corneal epithelium normally contains about 1.33 mg Vc per gram of wet
weight tissue (27), which corresponds to about 7.5 mM Vc. In the activated human
neutrophils, internal Vc concentrations as high as 14 mM were detected when external
vitamin is kept at physiological concentrations (28). Therefore, the inhibitory effect of Vc
on the SpnHL enzymatic activity may have a physiological meaning. It has long been
known that deficiency of Vc may cause decreased resistance to some bacterial infections.
One explanation to this is that animal cells (such as neutrophils) generate oxidants to kill
bacteria and using Vc to quench and control the extra oxidants released (28). The
inhibitory effect of Vc on SpnHL activity provides an additional possible explanation to
the Vc function as an antibacterial agent. The large concentration of Vc in human tissues
makes the tissue environment more unfavorable to the pneumococcal invasion, therefore,
providing a low level of natural resistance to such bacterial invasion. The infections and
diseases caused by pneumococci are thus significantly reduced. Therefore, Vc is likely a
natural constituent of the biochemical defense system against the pneumococcal invasion
in the host tissues. Pneumococcal invasions usually occur in tissues with relatively low
concentrations of Vc. The normal Vc concentration in plasma is around 0.1-0.2 mM, and
Hyaluronate lyase complexed with vitamin C
14
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

about 10 times higher in lung, brain, kidney, lymph glands, and small intestinal mucosa
(29) which is about 10 times less than the highest Vc concentrations detected in human
tissues.
The SpnHL-Vc complex structure also provides some clues for the design of a
more efficient hyaluronate lyase inhibitor. Based on the interface characteristics, it can be
expected that a stronger hyaluronate lyase inhibitor should have a larger ring system to
benefit the hydrophobic binding to the Trp292 indole group. At least one negative charge
provider, for example, a carboxyl group, is required in the inhibitor structure to provide
the negative charges to lead the inhibitor into the cleft region. In summary, a stronger
inhibitor can be expected to have an increased area of hydrophobic interactions and to
have more properly placed negatively charged substituents such as carboxyl groups.
Molecular properties of Vc are closely related to its structural characteristics. The
widely studied free radical scavenger and antioxidant properties of Vc are directly related
to the active redox chemical characteristics of this molecule. Our studies emphasized the
significance of this structural similarities of Vc, a sugar derivative, to polysaccharides.
This structural similarity confers Vc the capacity of protecting hyaluronan, the main
component of connective tissues, from being degraded by bacterial hyaluronate lyases.
Any destructive factors of polysaccharides, oxidants or hyaluronate lyases, may be
buffered by the existence of the large amount of Vc in tissues. The Vc structural
similarities to sugars, its interaction patterns with proteins revealed from the SpnHL-Vc
complex structure, and the importance of both hydrophobic and ionic contacts in the
protein-Vc interface might lead to the reevaluation of the structure and function
relationships of Vc.
Hyaluronate lyase complexed with vitamin C
15
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Conclusions and general implications Vc may compress or retard bacterial invasion
by directly inhibiting bacterial “spreading factor” such as hyaluronate lyase, through
binding to the enzyme active site and competing with the binding of the hyaluronan
substrate. All seven protein interface residues interacting with Vc are strictly conserved
among all known bacterial hyaluronate lyases (8). The studies on the SpnHL-Vc
interface are thus significantly relevant to all these bacterial hyaluronate lyases. For
example, S. agalactiae hyaluronate lyase (GbsHL) crystal structure was recently
determined (19, 30). Its active center construction and geometry is nearly the same as it is
in SpnHL. Therefore, the results shown may be applicable to GbsHL, which means that
Vc might also provide the host with the ability to resist the S. agalactiae invasion to a
certain extent.
This is the first time that the direct action of Vc on a bacterial “spreading factor”
has been observed. The structural basis of this inhibition is due to the structural
similarity of Vc to the glucuronate residues in hyaluronan, the substrate of hyaluronate
lyases. The inhibitory effect, confirmed by our enzyme activity measurements and the
SpnHL-Vc complex structure studies, shows that Vc is likely directly involved in the
inhibition of bacterial invasion, in addition to its antioxidant and free radical scavenger
properties.
Hyaluronate lyase complexed with vitamin C
16
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

References
1. Khurana, S., Powers, D.B., Anderson, S., and Blaber, M. (1998) Proc. Natl. Acad. Sci.
USA 95, 6768-73
2. Levine, K. (1986) New Engl. J. Med. 314, 892-902
3. Hwang, J., Peterson, H., Hodis, H.N., Choi, B., and Sevanian, A. (2000) Atheros. 150,
275-284
4. Kalka, K., Mukhtar, H., Turowski-Warke, A., and Merk, H. (2000) Skin Pharmacol.
Appl. Skin Physiol. 13, 143-149
5. Rice, M.E. (2000) Trends Neurosci. 23, 209-216
6. Smirnoff, N. (2000) Curr. Opin. Plant Biol. 3, 229-235
7. Carrell, H.L., Glusker, J.P., Burger, V., Manfre, F., and Tritsch, D. (1998) Proc. Natl.
Acad. Sci. USA 86, 4440-4444
8. Li, S., Kelly, S.J., Lamani, E., Ferraroni, M., and Jedrzejas, M.J. (2000) EMBO J. 19,
1128-1140
9. Jedrzejas, M.J. (2000) Crit. Rev. Biochem. Mol. Biol. 35, 221-251
10. Jedrzejas, M.J., Mewbourne, R.B., Chantalat, L., and McPherson, D.T. (1998) Protein
Expr. Purif. 13, 83-89
11. Jedrzejas, M.J., Chantalat, L., and Mewbourne, R.B. (1998) J. Struct. Biol. 121, 73-
75
12. Otwinowski, Z., and Minor, W. (1997) Mothods Enzymol. 276, 307-326
13. Brunger, A.T., and Rice, L.M. (1997) Methods Enzymol. 277B, 243-269
14. Brunger, A.T. (1997) Methods Enzymol. 272B, 366-396
Hyaluronate lyase complexed with vitamin C
17
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

15. Jones, T.A., Zhou, J.Y., Cowan, S.W., and Kjeldgaard, M. (1991) Acta Crystallogr.
A47, 110-119
16. Li, M.W., Yudin, A.I., VandeVoot, C.A., Sabeur, K., Primakoff, P., and Overstreet,
J.W. (1997) Biol. Reprod. 56, 1383-1389
17. Berry, A.M., Lock, R.A., Thomas, S.M., Rajan, D.P., Hansman, D., and Paton J.C.
(1994) Infect. Immun. 62, 1101-1108
18. Menzel, E.J., and Farr, C. (1998) Cancer Lett. 131, 3-11
19. Li, S., and Jedrzejas, M.J. (2000) J. Biol. Chem. submitted
20. Ponnuraj, K., and Jedrzejas, M.J. (2000) J. Mol. Biol. 299, 885-895
21. Kelly, S.J., Taylor, K.B., Li, S., and Jedrzejas, M.J. (2000) Glycobiol. submitted
22. Bursey, E.H., and Poulos, T.L. (2000) Biochem. 39, 7374-7379
23. Hvoslef, J.(1968) Acta Crystallogr. B24, 1431-1440
24. Rollend, K., Marois, C. Siquier, V., Cattier, B., and Quentin, R. (1999)J. Clin.
Microbiol. 37, 1892-1898
25. Berry A.M., and Paton J.C. (2000) Infect. Immun. 68, 133-140
26. Bergsten, P., Amitai, G., Kehrl, J., Dhariwal, K., Klein, H.G., and Levine, M. (1990)
J. Biol. Chem. 265, 2584-2587
27. Brubaker, R.F., Bourne, W.M., Bachman, L.A., Mclaren, J.W. (2000) Invest.
Ophthalmol. Vis. Sci. 41, 1681-1683
28. Washko, P., Wang, Y., and Levine, M. (1993) J Biol. Chem. 268, 15531-15535
29. Lester, P., and Fuchs, J. (1997)Vitamin C in health and disease. Marcel Dekker, Inc.
30. Jedrzejas, M.J., and Chantalat, L. (2000) Acta Crystallogr. D 56, 460-463
Hyaluronate lyase complexed with vitamin C
18
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Footnotes:
Acknowledgments: Diffraction data were collected at the Brookhaven National
Laboratory, National Synchrotron Light Source at the beamline X25. This work was
supported by NIH grant AI 44079 (MJJ).
Figure legends:
Figure 1. 2Fo-1Fc electron density map (1 σ) for the refined Vc bound to the active site
of SpnHL. One water molecule hydrogen bonded to Vc O2 atom can be identified.
Figure 2. The Vc binding position in the SpnHL structure. The positions of the N- and
C- terminals, the α- and β-domains were shown. One Vc molecule is bound to the cleft
between two structural domains.
Figure 3. The environment of Vc in the cleft of the S. pneumoniae hyaluronate lyase. (a)
The relative position of Vc to the active center residues. (b) All residues interacting with
Vc.
Figure 4. Structural comparison of protein-bound Vc overlapped with the Vc crystal
structure. Hydrogen atoms were not shown. Vc atoms were labeled as conventional (23).
SpnHL bound Vc is in green color whereas the Vc crystal structure is in lavender color.
Large atomic displacement in the carboxyl group atoms can be seen, whereas the ring
atoms change less.
Hyaluronate lyase complexed with vitamin C
19
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Figure 5. Enzyme activity of SpnHL at the presence of ascorbic acid. The final enzyme
concentration was 0.33 mg/ml. The final Vc concentrations were 0.8 to 16 mM. Each
point represents the averaged value of seven parallel measurements. Vc inhibited SpnHL
activity shows in a dose-dependent manner with 50% inhibition (IC50) at around 5.8
mM.
Hyaluronate lyase complexed with vitamin C
20
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
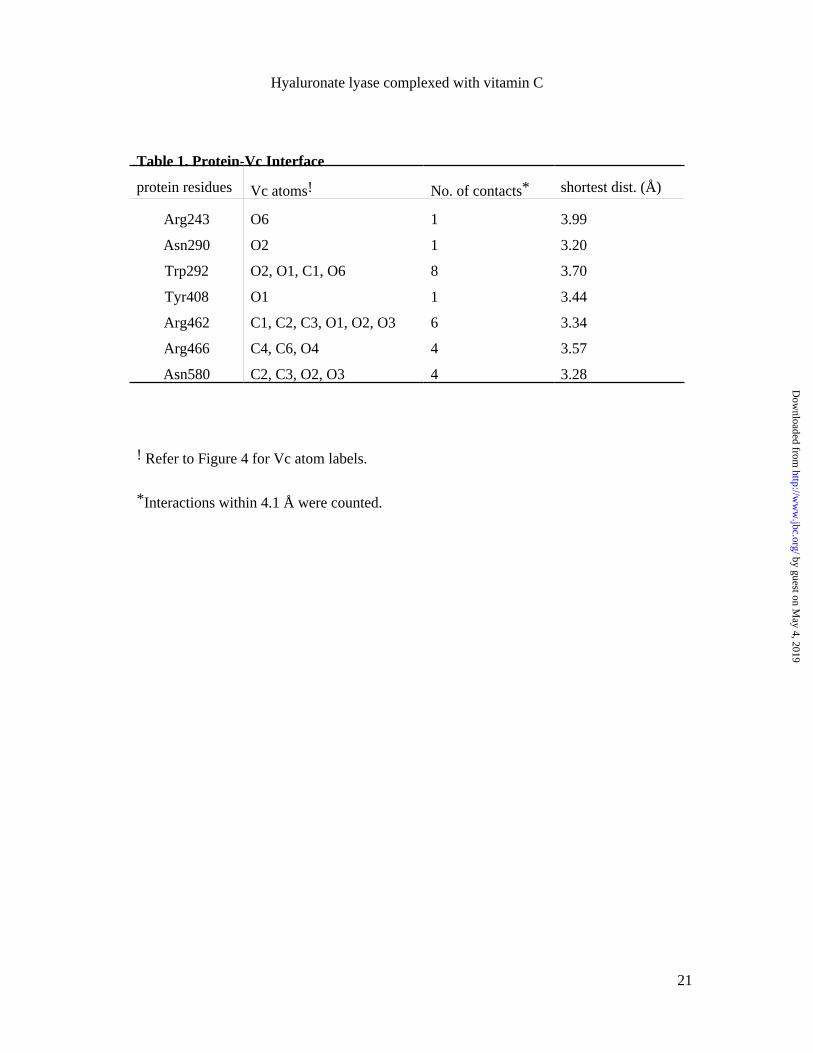
Table 1. Protein-Vc Interface
protein residues Vc atoms! No. of contacts* shortest dist. (Å)
Arg243 O6 1 3.99
Asn290 O2 1 3.20
Trp292 O2, O1, C1, O6 8 3.70
Tyr408 O1 1 3.44
Arg462 C1, C2, C3, O1, O2, O3 6 3.34
Arg466 C4, C6, O4 4 3.57
Asn580 C2, C3, O2, O3 4 3.28
! Refer to Figure 4 for Vc atom labels.
*Interactions within 4.1 Å were counted.
Hyaluronate lyase complexed with vitamin C
21
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from
C
Ascorbic acid
-domainβ
-domainα
N
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from

Songlin Li, Kenneth B. Taylor, Stephen J. Kelly and Mark J. Jedrzejaslyase
Vitamin C inhibits the enzymatic activity of streptococcus pneumoniae hyaluronate
published online January 12, 2001J. Biol. Chem.
10.1074/jbc.M011102200Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on May 4, 2019
http://ww
w.jbc.org/
Dow
nloaded from





![i-Visc · i-Visc VISCOELASTIC SOLUTION FOR INTRAOCULAR USE based on SODIUM HYALURONATE Specification Sodium hyaluronate Molecular weight [mio. Daltons] Viscosity* [mPas] Osmolality](https://img.pdfslide.us/doc/110x75/5f8d228ac639c80bb7041471/i-visc-i-visc-viscoelastic-solution-for-intraocular-use-based-on-sodium-hyaluronate.jpg)