Embed Size (px)
Citation preview

2003 Kluwer Academic Publishers. Printed in the Netherlands.
Vegetation:environment relationships and water management in SharkSlough, Everglades National Park
1, 1 1,2 1 1*M.S. Ross , D.L. Reed , J.P. Sah , P.L. Ruiz and M.T. Lewin1Southeast Environmental Research Center, Florida International University, University Park OE-148,
2Miami, Florida 33199, USA; Central Department of Botany, Tribhuvan University, Kirtipur, Kathmandu,*Nepal; Author for correspondence (e-mail: [email protected]; phone: 11-305-348-1420; fax: 11-305-348-
4096)
Received 25 January 2001; accepted in revised form 25 October 2002
Key words: Bayhead swamps, Hydrology, Long hydroperiod wetlands, Sawgrass marsh, Soils, South Florida,Spikerush marsh, Vegetation change
Abstract
The hydrologic regime of Shark Slough, the most extensive long hydroperiod marsh in Everglades National Park,is largely controlled by the location, volume, and timing of water delivered to it through several control structuresfrom Water Conservation Areas north of the Park. Where natural or anthropogenic barriers to water flow arepresent, water management practices in this highly regulated system may result in an uneven distribution of waterin the marsh, which may impact regional vegetation patterns. In this paper, we use data from 569 samplinglocations along five cross-Slough transects to examine regional vegetation distribution, and to test and describe theassociation of marsh vegetation with several hydrologic and edaphic parameters. Analysis of vegetation:environ-ment relationships yielded estimates of both mean and variance in soil depth, as well as annual hydroperiod, meanwater depth, and 30-day maximum water depth within each cover type during the 1990’s. We found that rankabundances of the three major marsh cover types (Tall Sawgrass, Sparse Sawgrass, and Spikerush Marsh) wereidentical in all portions of Shark Slough, but regional trends in the relative abundance of individual communitieswere present. Analysis also indicated clear and consistent differences in the hydrologic regime of three marshcover types, with hydroperiod and water depths increasing in the order Tall Sawgrass , Sparse Sawgrass ,
Spikerush Marsh. In contrast, soil depth decreased in the same order. Locally, these differences were quite subtle;within a management unit of Shark Slough, mean annual values for the two water depth parameters varied lessthan 15 cm among types, and hydroperiods varied by 65 days or less. More significantly, regional variation inhydrology equaled or exceeded the variation attributable to cover type within a small area. For instance, estimatedhydroperiods for Tall Sawgrass in Northern Shark Slough were longer than for Spikerush Marsh in any of theother regions. Although some of this regional variation may reflect a natural gradient within the Slough, a largeproportion is the result of compartmentalization due to current water management practices within the marsh. Weconclude that hydroperiod or water depth are the most important influences on vegetation within managementunits, and attribute larger scale differences in vegetation pattern to the interactions among soil development,hydrology and fire regime in this pivotal portion of Everglades.
NomenclatureGodfrey and Wooten (1979, 1981)
Introduction agement in South Florida represent one of the mostambitious ecological restoration efforts ever attempt-
Plans currently being developed to revise water man- ed. The primary objective of the plan is to benefit the
291Wetlands Ecology and Management 11: 291–303, 2003.

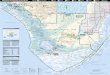
Greater Everglades ecosystem (USACE 1994), which 1). Since 1983, legally mandated concerns for theincludes an International Biosphere Reserve, i.e., restoration and maintenance of natural EvergladesEverglades National Park. The uniqueness of the ecosystems have stimulated a series of regulatoryEverglades lies in their combination of large size, changes within the existing structural network. How-subtropical climate, shallow oligotrophic waters, car- ever, in the southern Everglades, one managementbonaceous sediments, and proximity to a rapidly direction has remained consistent during the 1980’sexpanding urban population. The problems that have and 1990’s: the routing of southbound water to themotivated the Everglades restoration effort include: west at Highway 41, away from its primary pre-(a) sharp declines in populations of wading birds and development flow-way through Northeast Sharkother animals of special concern (Powell et al. 1989; Slough (Light and Dineen 1994;Van Lent et al. 1999).Ogden 1994; Curnutt et al. 1998); (b) deteriorating In this area, concentration of delivery through controlconditions in the Florida Bay estuary, the terminal structures west of the L-67 levee has eliminated thereceiving basin for Everglades surface water (Four- historic northeast-southwest flow of water in thequrean and Robblee 1999); (c) rapid expansion of northern portions of ENP (Figure 1). This distributioncattails (Typha domingensis Pers.), associated with pattern has generally served to decrease water levelthe outflow of phosphorus-enriched canal water into and flooding duration in Northeast Shark Slough,the Water Conservation Areas (WCA’s) and adjacent while creating wetter conditions in Northern Sharkmarshes north of Everglades National Park (Davis Slough. The spatial pattern of water delivery may1991; Craft and Richardson 1997; Doren et al. 1997; have had less hydrologic impact on the more remoteWu et al. 1997); (d) loss of tree islands due to southern portions of the Slough. Effects of theseexcessive flooding or fire in some parts of the WCA’s management activities on regional vegetation patterns(Brandt et al. 2000); (e) expansion of sawgrass at the have not been addressed in detail.expense of wet prairie communities (Davis et al. From a distance, the vegetation of the Slough1994); and (f) the rapid growth of exotic plant and appears rather homogeneous, but closer inspectionanimal populations throughout the Everglades reveals a complex mosaic of marsh assemblages,(Schmitz and Brown 1994; O’Hare and Dalrymple spotted liberally with distinct wooded parcels called1997). With the exception of (e) and (f), the problems tree islands. Within the marsh, mono-dominant standslisted above are primarily manifested in wetlands of the large sedge Cladium jamaicense Crantz (saw-peripheral to Shark Slough, the main pathway of grass) are interrupted by more open communitiessurface water drainage through Everglades National featuring a mixture of smaller aquatic plants andPark (ENP). However, as a result of its central loca- periphyton (an integrated mat community consistingtion, the Slough’s hydrology will be affected by water of algae, bacteria, and microheterotrophs). Thesemanagement measures undertaken to benefit other habitat units are frequently elongated in shape andportions of the Everglades ecosystem. Furthermore, oriented parallel to the historic NE-SW direction oflong-term alterations in the hydrologic regime of water flow, suggesting a strong functional connectionShark Slough may result in parallel biotic changes, between hydrology and vegetation. Recent investiga-especially in the distribution and abundance of native tions (Gunderson 1989; Olmsted and Armentanoplant and animal communities. 1997; Busch et al. 1998) documented small differ-
Hydrologic restoration activities in the Everglades ences in mean hydrologic regime among communityfollow on the heels of a long sequence of structural, types, e.g., flooding was usually deeper and moreoperational, and regulatory changes in water manage- persistent in open marsh types dominated by spiker-ment in South Florida, beginning long before the ush (Eleocharis cellulosa Torr.) or maidencanecreation of ENP in 1947. Rising population and (Panicum hemitomon Schult.) than sawgrass marsh,agricultural development throughout central Florida which in turn was wetter than forested cover types.provided the stimulus for state, federal, and privately However, these conclusions were based on data fromfunded diversion projects whose purposes were drain- a limited number of Shark Slough settings, andage, flood protection, and irrigation (McCally 1999). quantification of variation in the vegetation:hydrologyThese goals were achieved through construction and relationship at the broad landscape scale remainsoperation of an extensive network of pumps, canals, incompletely understood.and levees, which effectively compartmentalized the The relationship of hydrology and vegetation in thecentral Everglades north of US Highway 41 (Figure Everglades is complicated by interactions with other
292

Figure 1. Study area with location of sampling transects.
dynamic environmental factors, including soil charac- each point by referencing the nearest water levelteristics and disturbance regime (Duever et al. 1986). recorder, then control for local spatial effects and theFor instance, data presented by Olmsted and Armen- covariance among soil and hydrologic variables in thetano (1997) indicate that soils underlying sawgrass analytical phase.marsh were, on average, deeper than soils of spiker-ush /maidencane communities, though overlap wassubstantial. Everglades soil development reflects a Methodsbalance of accretion v. degradation processes, i.e.,root production, sedimentation, decomposition, and Vegetation and environmental samplingerosion rates; all of these processes may be func-tionally related to the overlying plant communities Five transects were established to examine the vari-(e.g., Craft et al. 1993; Day et al. 1999). These ability in vegetation, hydrology, and soils along therelationships are largely unquantified, but may ac-
Table 1. Transect and plot layout in four Shark Slough (SS)count for much of the variation in marsh surfacehydrologic regions.topography (and in the depth of the soil profile), withRegion Transect Transect Numberassociated feedbacks on hydrology.
length of plotsIn the research reported below, our objectives were:(km)
(1) to clarify the associations of hydrology, soil depth,Northeast SS 1 4.0 73and plant community structure in Shark Slough,
2E 1.4 22focusing especially on regional variation in theseNorthern SS 2W 5.0 91
relationships, and (2) to evaluate the possible role of 3 6.0 110water management in creating such variability, if Central SS 4E 3.0 55
4W 4.0 73present. Our approach was to sample vegetation, soils,Southern SS 5E 4.6 87and water depth at regular intervals along cross-
5W 3.3 58Slough transects, construct a hydrologic record for
293

length of Shark Slough in Everglades National Park scribed above, we also characterized vegetation cover(Figure 1, Table 1). Transects were oriented perpen- every five meters along the transects, based on adicular to the apparent direction of water flow, and visual assessment of several broad structural andranged in length from 4 to 12 km. Three transects compositional attributes. We assigned each point towere effectively divided into eastern and western one of six cover types defined in Table 2: spikerushportions, because they were interrupted for at least 1 marsh, sparse sawgrass, tall sawgrass, bayheadkm by impassable terrain or disturbance associated swamp, cattail marsh, and dead sawgrass /open water.with a canal or airboat trail. Sampling stations were The first four of these units were equivalent to typesestablished every 100 meters along each transect. To described by Olmsted and Armentano (1997).address smaller-scale variation, we sampled every 20 Concurrent with vegetation sampling, soil depthmeters within the section between meters 200 and 400 was measured at the SE corner of each plot and at thein each km. In all, 569 points were sampled. Vegeta- three herb subplots. Soils were probed to bedrocktion sampling was conducted when water levels were with a 1 cm diameter aluminum rod, and mean depthhigh in Shark Slough, i.e., during the periods was calculated from the four measured values. WaterNovember 1998 to January 1999 and July 1999 to depths were likewise recorded during vegetation sam-December 1999. Sampling within individual transects pling at the same four locations. Referencing thewas completed within six weeks or less. average of these values to the datum for the same day
A nested plot design was chosen for quantitative at the nearest long-term water recorder, daily watervegetation sampling, because of the range of plant level at each plot was estimated for the period Januarygrowth forms present in the Slough (e.g., Hedman and 1, 1990 to December 31, 1999; these data were usedVan Lear 1995). At each station, a PVC tube driven to calculate hydroperiod (number of days per yearinto the sediment marked the SE corner of both a 10 when the surface was inundated), mean water depth3 10 meter tree plot and a 5 3 5 meter shrub/herb (average of all days when water was above the sur-plot. In the former, we measured the DBH, crown face), and maximum water depth (highest 30-daylength and width of each tree (defined as woody stems running mean) for each year during the decade.$ 5 cm DBH), then calculated species cover assum- The assumption on which estimates of the aboveing elliptical crown form (Peek 1970). In the latter, hydrologic parameters are based is that daily changewe estimated the cover class of each species of shrub in water level is equal at the reference recorder and at(vines and woody stems $ 1 m height and , 5 cm all plots along a transect. Comparison of water levelsDBH), using a modified Braun Blanquet scale with along the transects and at the reference recorders forthe following categories: , 1%, 1–4%, 4–16%, 16– two days in October 1999 and February 2000 – a33%, 33–66%, and . 66% (Mueller-Dombois and recession period when water remained well above theEllenberg 1974). Species cover class of herbs and surface throughout the Slough – indicated that thewoody plants , 1 m height was estimated in similar 95% confidence limits for daily estimates were ap-
2fashion in three randomly located 1-m subplots proximately 6 1 cm. However, the assumption ofwithin the 5 3 5 m plot. Plot averages were calculated equal change along the transect is violated when waterusing the midpoints of the cover classes. Species at the recorder recedes below the soil surface, afterpresent in the plot but not found in any of the subplots which its level changes more rapidly than at anywere assigned a mean cover of 0.01%. Data analysis still-flooded plots, because water occupies porewas based on the total of tree, shrub, and herb stratum spaces only. Therefore, during periods when waterestimates for each species. level dropped below the surface at the water level
In addition to the quantitative observations de- recorder, we assumed that water level at the flooded
Table 2. Criteria used to distinguish major cover types in Shark Slough.
Vegetation type Criteria
Spikerush marsh Dominant species E. cellulosa, P. hemitomon, or P. geminatum; canopy height , 1.5 meters.Sparse sawgrass Dominant species C. jamaicense; canopy height , 2 meters.Tall sawgrass Dominant species C. jamaicense; canopy height 2 meters or more.Bayhead swamp Woody vegetation dominant or co-dominant with C. jamaicense.Cattail marsh Dominant species Typha domingensis; canopy height 2 meters or more.Dead sawgrass – open water Total live vegetation cover , 20%; evidence of large dead culms of C. jamaicense
294

Figure 2. Cover type distribution in four Shark Slough (SS) hydrologic regions.
21plots fell at a rate of 0.5 cm day until reaching the positional dissimilarities among sites, and thereforesurface; and rose, ahead of the water surface at the has advantages over parametric ordination techniquesrecorder, at the same rate. The above rate is a reason- in which the underlying assumptions are rarely satis-able maximum, based on inspection of recorded re- fied in field community data (Clarke 1993). The sitecession rates at several recorders, supported by esti- ordination was examined with respect to the distribu-mates of daily evapotranspiration in the Everglades tion of (a) the three hydrologic variables, (b) soil(German 2000; Gunderson, unpublished manuscript) depth, and (c) the vegetation categories defined inand an emergent aquatic macrophyte-dominated wet- Table 2. To test the strength of the relationshipsland in Central Florida (Dolan et al. 1984). The between plant species composition and the environ-situation described above constituted , 5 percent of mental variables while accounting for spatial struc-the data base, because most of the water level re- ture, we applied partial Mantel tests, using the R-corders were situated in open slough conditions where Package program (Legendre et al. 1996–99).water depths rarely dropped below the soil surface. In addition to the ordination approach described
above, in which vegetation was treated as a continu-Data analysis ous variable, we also tested for differences in hydro-
logic and edaphic measures among discrete manage-We examined compositional variation in Shark ment units, using two-way, Model 1 analysis ofSlough vegetation from nested plot data and its en- variance. Four management units, which also repre-vironmental correlates through ordination and other sent geographic regions within Shark Slough, weremultivariate analytical techniques, using data from all defined (Table 1, Figure 1) on the basis of proximityfive transects. To downweight the effects of very and position relative to the water delivery structures.abundant species, a square root transformation was Northeast Shark Slough (NESS), which included theapplied to the cover percentages, and the transformed water-deprived marshes east of the L-67 levee, wasdata were relativized within plots (Jongman et al. represented by Transect 1 and the east end of Transect1995). A site-by-site dissimilarity matrix was created 2. Northern Shark Slough (NSS), the water-enhancedbased on the Bray – Curtis metric, and the data were wetlands immediately west of the L-67 levee, wereordinated with the non-metric multidimensional sca- represented by points along Transect 3 and the westling (NMS) algorithm in PC-ORD version 4.10 side of Transect 2. Central Shark Slough (CSS) was(McCune and Mefford 1999). NMS is a non-paramet- represented by Transect 4, and Southern Shark Sloughric ordination technique based on the ranked com- (SSS) by Transect 5. Only plots characterized as
295

Figure 3. Contour plots of hydroperiod during a dry year (1991) and an average year (1997), superimposed on the NMS ordination ofvegetation at 164 Shark slough locations. Plots are categorized into five vegetation classes, following Olmsted and Armentano (1997).
spikerush marsh, sparse sawgrass, or tall sawgrass Resultswere considered in the analysis of variance, becauseother cover types were not sufficiently well repre- Based on characterization of points at five-metersented throughout the study area. Soil depth, hy- intervals along each transect, rank abundances of thedroperiod, mean water depth, and maximum water three major marsh cover types were identical in thedepth were the environmental variables considered. four management units: sparse sawgrass . spikerushWe analyzed data from 1991 and 1997, which were marsh . tall sawgrass (Figure 2). However, regionaldry and average water years, respectively, in com- trends in the relative abundance of individual com-parison to other years in the 1990’s. munities were noteworthy. Vegetation in NESS and
296

Table 3. P-values for partial Mantel statistics, indicating strength of association among plant species composition, soil depth, and threehydrologic variables in four regions, after removing effect of spatial contiguity among sampling points.
fIndependent variable tested Northeast SS (95) Northern SS (114) Central SS (114) Southern SS (113)a dAnalysis 1 Hydroperiod 0.113 0.001 0.001 0.007
eMean depth 0.085 0.001 0.001 0.002eMax depth 0.087 0.001 0.001 0.002
Soil 0.336 0.235 0.008 0.037bAnalysis 2 Mean depth 0.001 0.001 0.001 0.001
Max depth 0.001 0.001 0.001 0.001cAnalysis 3 Hydroperiod 0.026 0.040 0.020 0.140
Mean depth 0.009 0.110 0.067 0.026Max depth 0.011 0.030 0.015 0.056
a bassociations of species composition with environmental variables, associations of hydroperiod with other two hydrological parameters,c d eassociations of soil depth with three hydrologic variables, hydroperiod data is based on dry year (1991), mean and maximum water depth are
fbased on an average water year (1997), sample size.
SSS was relatively uniform, with sparse sawgrass sawgrass sites were inundated for 6–10 months andoccupying more than 60% of the landscape. Domi- bayhead swamp for 2–6 months, with concomitantlynance was more evenly distributed among cover types lower water depths. In 1997, a year in which waterin NSS and CSS, where open spikerush marsh cov- levels were about average for the decade, all pointsered nearly one-third of the surface. Tall sawgrass were inundated for at least eleven months except inwas least abundant in CSS and SSS, and never ex- bayhead swamp (range: 4–8 months). Dead sawgrass /ceeded 20%. Bayhead swamp, dead sawgrass /open open water was found at points intermediate in hy-water, and cattail marsh were minor components in droperiod between tall sawgrass and bayhead swamp.NESS. Application of partial Mantel tests to control for
Analysis of the compositional data within transects spatial effects within regions further supported theindicated considerable small-scale spatial variation in correlation of hydrology and plant species composi-marsh vegetation. For instance, along Transect 2, tion. Except in NESS, all three hydrologic parametersmean Bray-Curtis dissimilarity for pairs of points # were strongly correlated with vegetation when spatial40 meters apart was 0.42, while the mean for points effects were removed (Table 3: Analysis 1). Signifi-further than 40 meters apart was 0.56. Hydrology and cant Mantel associations between soil depth andsoil depth likewise exhibited spatial patterning. vegetation were also observed in CSS and SSS, but
To eliminate redundancy associated with spatial not in NSS or NESS. The weakness of the soil:vegeta-structure, ordination of the vegetation data was there- tion relationship in comparison to the relationshipfore based on points separated by 200 meters or more. between hydrology and vegetation is also evidentThe two-axis NMS site ordination derived from these when soil depths are superimposed on the NMSdata was characterized by a low stress value (13.1), ordination, without accounting for region (Figure 4).and explained about 96% of the total variation in the Partial Mantel tests were also applied to exploredissimilarity matrix. Plots within four major vegeta- associations among environmental variables. Thetion units (spikerush marsh, sparse sawgrass, tall three hydrological parameters were highly inter-corre-sawgrass, bayhead swamp) occupied distinct posi- lated (Table 3: Analysis 2), but the strength of theirtions in the ordination, with small areas of overlap relationships with soil depth was region-specific, withbetween tall and sparse sawgrass, and between the 1–3 significant associations per region (Table 3:latter and spikerush marsh (Figure 3). Superimposing Analysis 3).a contour plot based on calculated hydroperiods on The analysis of variance indicated highly signifi-the ordination diagram (Figure 3), it is clear that cant effects of both cover type (p , 0.01) and regionhydrology bears a strong influence on the Shark (p , 0.001) on all three hydrologic variables in bothSlough vegetation community mosaic. During 1991, a years, 1991 and 1997. Interaction effects were mostlydry year, spikerush marsh and sparse sawgrass were non-significant, except for hydroperiod in 1997, whengenerally inundated for nine months or more, which most of the non-forested portions of the Slough wereequated to maximum water depths of at least 40 cm, continuously inundated with the exception of severaland average water depths of 25 cm or more. Tall tall sawgrass stands in CSS (Figure 5). Inter-regional
297

Figure 4. Contour plot of soil depth superimposed on the NMS ordination of vegetation at 164 Shark Slough locations. Plots are categorizedinto five vegetation classes, following Olmsted and Armentano (1997).
differences were explored further by applying multi- was 10 to 31 cm deeper than in spikerush marsh andple comparison tests (Tukey’s HSD) to sparse saw- sparse sawgrass, depending on region. Multiple com-grass locations, which were abundant in all four parison tests among regions were applied to theregions. Differences between regions varied with year widely distributed sparse sawgrass type alone. Theyand with hydrologic variable. In general, hydroperiod indicated that soil depth in NESS exceeded that inwas longest and water level was deepest in NSS, NSS, but other regional differences were non-signifi-while CSS and SSS were characterized by shorter cant.hydroperiod and shallower water. Differences amongregions were comparable in magnitude to differencesamong cover types within a single region. For in- Discussionstance, in 1991, when mean hydroperiod for spikerushmarsh consistently exceeded that of tall sawgrass in The results described above support the contention,the same region by 10–60 days, tall sawgrass sites in expressed most comprehensively by Olmsted andNSS had a longer mean hydroperiod than spikerush Armentano (1997), that Shark Slough plant com-marsh in any of the other regions (Figure 5). munities are comprised of several widespread and
Two-way analysis of variance showed significant distinct cover types, arranged roughly along a hydro-effects of both cover type (p , 0.001) and region (p logic gradient. Owing to the extensive nature of the5 0.04) on soil depth also. Soil depth in tall sawgrass current data, our analyses go a step further in (a)
298

Figure 5. Mean hydroperiod in 1991 by cover type and region.
assessing the strength of relationships among vegeta- Vegetation:environment relationshipstion, hydrology, and soil depth, and (b) defining therange of temporal and spatial variation in hydrology Our examination of vegetation:environment relation-and soils within the major cover types, both for the ships consisted of graphical descriptions of Slough-Slough as a whole and for individual management wide patterns, and statistical analyses that also ac-units within it. Examination of broad spatial variation counted for regional effects. Statistics included bothindicates that water management has contributed to continuous and categorical approaches; both appear toregional hydrologic differences that are large in com- be justified, based on the clustering within the vegeta-parison to those that distinguish cover types in any tion ordination of sites assigned to specific wetlandone region within a single year. Given the observed community types according to the criteria of Olmstedstrength of the vegetation:hydrology association, and Armentano (1997).management that artificially maintains such regional A strong association of vegetation with hydrologypatterns in hydroperiod and water depth on a long- was evident in our data, whether viewed for Sharkterm basis is likely to have important effects on the Slough in its entirety (Figure 3) or within individualstructure of Everglades plant communities. regions of the marsh (three of four regions in Table
Figure 6. Conceptual model of processes underlying variation in Shark Slough wetlands.
299

3). Outside of the forested areas, sawgrass-dominated posed much more slowly than Eleocharis cellulosamarsh is most common where hydroperiod is shortest under several hydrologic conditions in Yucatan Penin-and water depth shallowest, and more open spikerush sula marshes. Vegetation-related differences in flowmarsh is characteristic of more hydric conditions. resistance among Shark Slough habitats are complexDramatic contrasts in vegetation are typically associ- and not yet completely understood, varying withated with slight differences in hydrologic regime, e.g., macrophyte density, periphyton structure, flow rate,5–10 cm water depth or 10–60 days annual inunda- and position in the water column (Carter et al. 1999).tion (Figures 3 and 5). Our results support earlier Nevertheless, it is reasonable to expect that the denserconclusions (Gunderson 1989; Olmsted and Armen- tall sawgrass community would be more resistant totano 1997; Busch et al. 1998) that hydroperiod and flow under many conditions, including extreme highwater depth variables are important determinants of water conditions when soils are more directly affectedmarsh community composition and structure in Shark by hydrology through erosion and deposition pro-Slough marshes. In the present study, however, we cesses. A net transport from spikerush marsh to tallalso found that in some management units soil depth sawgrass would result, enhancing soil accretion fur-was strongly associated with both hydrology and ther in the latter type. It is notable that, within thevegetation pattern. range of the model, the productivity and sedimenta-
In the southern Everglades, surface topography tion trends cited above would elicit a positive feed-may result from local variation in soil accretion rates, back loop, accentuating variability in surface topog-which are themselves based on the balance of bio- raphy, hydroperiod, and water depth until fire or otherlogical and physical processes. Soil depth was not disturbances intervened. In contrast, a hydrologicstrongly associated with vegetation over the Slough as mechanism cited by Craft and Richardson (1993) toa whole (Figure 4), but clear associations of soil depth explain patterns of soil accretion in their northernand vegetation were observed in two of four regions, Everglades study area, i.e., slower decomposition inand soil and hydrologic variables were generally wetter sites due to anaerobic conditions, would tend toassociated at the regional level (Table 3). Marsh soils reduce surface variability.were deepest in the tall sawgrass type (Figure 4), The above discussion concerns processes that resultwhich is characterized by relatively low water (Figure in small-scale topographic variation within regions.5). Similar results were reported by Olmsted and However, inter-regional variation in soil depth (Fig-Armentano (1997), who noted a statistically non-sig- ure 5) may be more a function of bedrock topographynificant tendency toward deeper soils in tall sawgrass than of soil development. Soils in Shark Sloughthan in neighboring plant communities. develop on uneven limestone bedrock, with micro-
The observed interactions between vegetation, hy- karst features prominent at the local scale. Regionally,drology, and soil depth suggest the simple model the Slough occupies a slightly inclined bedrock troughillustrated in Figure 6. The model postulates that bounded by the Miami Rock Ridge on the east and thehydrology’s influence on soil depth is primarily in- modestly elevated Big Cypress swamp on the west.direct, through its influence on fire and vegetation.Wu Within it, limestone outcroppings and narrow ridgeset al. (1996) modeled the effects of hydrology (as well oriented perpendicular to the predominant direction ofas vegetation) on the frequency and nature of Ever- flow (‘rock reefs’; Froehlich 1973) alternate withglades fires, which modify the surface and reduce soil more extensive areas of lower bedrock. One bedrockdepth by removing accumulated organic matter (e.g., depression occurs in the western portion of NESS,Arabas 2000). The pathway through vegetation may where all cover types are underlain by peat soils ofinvolve differential belowground product- more than 1 meter depth (Leighty and Hendersonion:decomposition and/or resistance to surface water 1958). Conversely, sections of NSS and smaller por-sheetflow, e.g., in tall sawgrass (deep soils) cf., spiker- tions of NESS and CSS are characterized by frequentush marsh (shallow soils). Daoust and Childers (1998) bedrock outcrops.found net aboveground primary production to beabout seven times higher in sawgrass than adjacent Regional variation and water management effectsspikerush marsh in the southern Everglades. Rootproduction was not estimated, but may also be rela- In light of the strong association between vegetationtively high in sawgrass marsh. Rejmankova et al. and hydrology discussed above, how are the observed(1996) observed that Cladium jamaicense decom- differences in hydrology among regions within the
300

same cover type to be interpreted? These regional (Herndon et al. 1991), but reinvasion by sawgrasspatterns may be summarized as follows: (1) NSS, may be exceedingly slow (Herndon, unpublisheddirectly below the primary water delivery points from manuscript). In contrast, the open water areas wethe Water Conservation Areas, experienced the encountered in southern Shark Slough represent a latelongest hydroperiods and deepest water levels during stage in a recurring phenomenon that Wade et al.1990–99; (2) More remote areas in CSS and SSS (1980) termed ‘sawgrass decadence’. The cause ofwere characterized by shorter hydroperiods and shal- sawgrass decadence, the successional sequence thatlower waters over the same period; but (3) the relative typically follows it, and the role of fire or hydrologyhydrologic positions of the major cover types were in the process are currently unknown.preserved throughout all regions (Figure 5). The The regional differences in hydrology (Figure 5)finding of strong regional variation may be taken as are those of a closely managed wetland ecosystem,evidence of some degree of imbalance between exist- rigidly compartmentalized at the upstream end buting vegetation pattern and recent hydrology. This open downstream. Within these regions, we foundimbalance is also suggested by the nature of the clear and consistent differences in the hydrologiccurrent vegetation mosaic (Figure 2). For instance, if regime of the three major marsh cover types, in thecurrent vegetation patterns were in equilibrium with direction expected based on the existing literature,hydrology of the last decade, then CSS would more i.e., hydroperiod and water depth increased in theclosely resemble the monotonous sawgrass marsh of order tall sawgrass marsh , sparse sawgrass marsh ,
NESS and SSS than the more heterogeneous mixture spikerush marsh. Locally, these differences were quiteof spikerush- and sawgrass-dominated communities subtle; mean annual values for the two water depthof NSS, as it currently does. parameters typically varied less than 15 cm among
The less-than-perfect concordance of vegetation types, and hydroperiod by 60 days or less. Thiswith recent water level and hydroperiod may reflect a narrow range is relevant in assessing the impact oflag in response, and/or the influence of interacting inter-regional variation in hydrology, whose mag-environmental factors, especially fire (Figure 6). Lit- nitude equaled or exceeded that attributable to covertle is currently known about the time frame of vegeta- type within a region.tion response to altered hydrology in the Everglades. We conclude that current water management prac-Busch et al. (1998) observed annual variation in plant tices have resulted in an artificial regionalization inspecies composition in several Shark Slough wet Shark Slough, with hydroperiod or water depth theprairies. Though these small changes did appear to be most important influences on vegetation within man-related to year-to-year variation in water level, they agement units. We attribute these large- scale changesdid not result in consistent changes in community type in vegetation pattern to the interactions among soilover the ten years of study. Based on an analysis of development, hydrology and fire regime, as depictedpairs of aerial photographs encompassing a 15–21 in the model. Changes in water management associ-year period from the mid-1960’s to the mid-1980’s, ated with the restoration will alter the conditions ofDavis et al. (1994) documented a general increase in the model, and perhaps the extent of regionalization insawgrass marshes at the expense of open slough the Everglades, but these fundamental ecologicalhabitats in portions of Shark Slough and the Water relationships will continue to operate.Conservation Areas. However, no attempt was madeto relate vegetation changes to hydrology at a localscale. In some cases, the response time of vegetation Acknowledgementschange to important forcing factors varies with thespatial scale considered. For instance, Williams et al. This work was supported by Everglades National(1999) found that small-scale effects of salt water Park, Critical Ecosystem Science Initiative, in col-intrusion in coastal forests (elimination of tree regene- laboration with the FCELTER (NSF Grant numberration) sometimes preceded coarse-scale effects 9910514). This is SERC contribution number 187.(death of established trees) by many decades. Only thelatter would have been detectable on aerial photos. InShark Slough, when fire is followed by a rapid rise in Referenceswater level, extensive areas of sawgrass marsh maychange to open water habitat within a matter of weeks Arabas K.B. 2000. Spatial and temporal relationships among fire
301

frequency, vegetation, and soil depth in an eastern North Ameri- Godfrey R.K. and Wooten J.W. 1979. Aquatic and Wetland Plantscan serpentine barren. Journal of the Torrey Botanical Society of Southeastern United States: Monocotyledons. University of127: 51–65. Georgia Press, Athens, USA, 712 pp.
Brandt L.A., Portier K.M. and Kitchens W.M. 2000. Patterns of Godfrey R.K. and Wooten J.W. 1981. Aquatic and Wetland Plantschange in tree islands in Arthur R. Marshall Loxahatchee Nation- of Southeastern United States: Dicotyledons. University of Geor-al Wildlife Refuge from 1950 to 1991. Wetlands 20: 1–14. gia Press, Athens, USA, 933 pp.
Busch D.E., Loftus W.F. and Bass O.L. Jr 1998. Long-term hydro- Gunderson L.H. 1989. Historical hydropatterns in wetland com-logic effects on marsh plant community structure in the southern munities in Everglades National Park. In: Sharitz R.R. andEverglades. Wetlands 18: 230–241. Gibbons J.W. (eds), Freshwater Wetlands and Wildlife. CONF-
Carter V., Reel J.T., Rybicki N.B., Ruhl H.A., Gammon P.T. and 8603101, DOE Symposium Series No. 61., pp. 1099–1111.Lee J.K. 1999. Vegetative Resistance to Flow in South Florida:
Hedman C.W. and Van Lear D.H. 1995. Vegetative structure andSummary of Vegetation Sampling at Sites NESRS3 and P33,
composition of Southern Appalachian riparian forests. BulletinShark River Slough, November, 1996. U. S. Geological Survey
of the Torrey Botanical Club 122: 134–144.Open File Report OFR-99-218. U.S. Department of the Interior.
Herndon A., Gunderson L. and Stenberg J. 1991. SawgrassClarke K.R. 1993. Non-parametric multivariate analyses of changes
(Cladium jamaicense) survival in a regime of fire and flooding.in community structure. Australian Journal of Ecology 18: 117–Wetlands 11: 17–27.143.
Jongman R.H.G., ter Braak C.J.F. and van Tongeren O.F.R. 1995.Craft C.B. and Richardson C.J. 1993. Peat accretion and N, P, andData Analysis in Community and Landscape Ecology. Cam-organic C accumulation in nutrient-enriched and unenrichedbridge University Press, UK, 299 pp.Everglades peatlands. Ecological Applications 3: 446–458.
Legendre P., Vaudor A. and Casgrain P. 1996–99. The R PackageCraft C.B. and Richardson C.J. 1997. Relationships between soil4.0. Departement de Sciences Biologiques, Universite de Mon-nutrients and plant species composition in Everglades peatlands.treal, Montreal, Canada.Journal of Environmental Quality 26: 224–232.
Leighty R.G. and Henderson J.R. 1958. Soil Survey (detailed-Craft C.B., Seneca E.D. and Broome S.W. 1993. Vertical accretionreconnaissance) of Dade County, Florida. Series 1947, No. 4.in microtidal regularly and irregularly flooded estuarine marshes.USDA Soil Conservation Service, Washington, DC, USA.Estuarine, Coastal and Shelf Science 37: 371–386.
Light S.S. and Dineen J.W. 1994. Water control in the Everglades: aCurnutt J.L., Mayer A.L., Brooks T.M., Manne L., Bass O.L. Jr,historical perspective. In: Davis S.M. and Ogden J.C. (eds),Fleming D.M. et al. 1998. Population dynamics of the en-Everglades: The Ecosystem and Its Restoration. St. Lucie Press,dangered Cape Sable seaside-sparrow. Animal Conservation 1:Boca Raton, Florida, USA, pp. 47–84.11–21.
McCally D. 1999. The Everglades: An Environmental History.Daoust R.J. and Childers D.L. 1998. Quantifying abovegroundUniversity Press of Florida, Gainesville, Florida, USA.biomass and estimating net aboveground primary production for
wetland macrophytes using a non-destructive phenometric tech- McCune B. and Mefford M.J. 1999. PC-ORD. Multivariate Analy-nique. Aquatic Botany 62: 115–133. sis of Ecological Data, Version 4. MjM Software Design,
Davis S.M. 1991. Growth, decomposition, and nutrient retention of Gleneden Beach, Oregon, USA.Cladium jamaicense Crantz and Typha domingensis Pers. in the Mueller-Dombois D. and Ellenberg H. 1974. Aims and Methods ofFlorida Everglades. Aquatic Botany 40: 203–224. Vegetation Ecology. John Wiley and Sons, New York, NY, USA,
Davis S.M., Gunderson L.H., Park W.A., Richardson J.R. and 574 pp.Matson J.E. 1994. Landscape dimension, composition, and func- Ogden J.C. 1994. A comparison of wading bird nesting colonytion in a changing Everglades ecosystem. In: Davis S.M. and dynamics (1931–1946 and 1974–1989) as an indication ofOgden J. (eds), Everglades: The Ecosystem and Its Restoration. ecosystem conditions in the southern Everglades. In: Davis S.M.St. Lucie Press, Boca Raton, Florida, USA, pp. 419–444. and Ogden J.C. (eds), Everglades: The Ecosystem and Its Resto-
Day J.W. Jr, Rybczyk J., Scarton F., Rismondo A., Are D. and ration. St. Lucie Press, Boca Raton, Florida, USA, pp. 533–570.Cecconi G. 1999. Soil accretionary dynamics, sea-level rise and O’Hare N.K. and Dalrymple G.H. 1997. Wildlife in southernthe survival of wetlands in Venice Lagoon: a field and modelling Everglades wetlands invaded by melaleuca (Melaleuca quin-approach. Estuarine, Coastal and Shelf Science 49: 607–628. quenervia). Bulletin of the Florida Museum of Natural History
Dolan T.J., Hermann A.J., Bayley S.E. and Zoltek J. Jr 1984. 41: 1–68.Evapotranspiration of a Florida, USA, freshwater wetland. Jour- Olmsted I. and Armentano T.V. 1997. Vegetation of Shark Slough,nal of Hydrology 74: 355–371. Everglades National Park. SFNRC Technical Report 97-001.
Doren R.F., Armentano T.V., Whiteaker L.D. and Jones R.D. 1997. South Florida Natural Resource Center, Everglades NationalMarsh vegetation patterns and soil phosphorus gradients in the Park, Homestead, Florida, USA.Everglades ecosystem. Aquatic Botany 56: 145–163. Peek J.M. 1970. Relation of canopy area and volume to production
Duever M.J., Carlson J.E., Meeder J.F., Dueverm L.C., Gunderson of three woody species. Ecology 51: 1098–1101.L.H., Riopelle L.A. et al. 1986. The Big Cypress National Powell G.V.N., Bjork R.D., Ogden J.C., Paul R.T., Powell A.H. andPreserve. National Audubon Society, New York, USA. Robertson W.B. Jr 1989. Population trends in some Florida Bay
Fourqurean J.W. and Robblee M.B. 1999. Florida Bay: a history of wading birds. Wilson Bulletin 101: 436–457.recent ecological changes. Estuaries 22: 345–357. Rejmankova E., Pope K.O., Post R. and Maltby E. 1996. Herbace-
Froehlich D.J. 1973. The Rock Reefs of the Everglades of South ous wetlands of the Yucatan Peninsula: communities at extremeFlorida, MS thesis, University of Miami, Miami, Florida, USA. ends of environmental gradients. Int. Revue ges. Hydrobiol. 81:
German E.R. 2000. Regional Evaluation of Evapotranspiration in 223–252.the Everglades. Report 00-4217. USGA Water-Resources In- Schmitz D.C. and Brown T.C. 1994. An Assessment of Invasivevestigations, Tallahassee, Florida, USA. Non-Indigenous Species in Florida’s Public Lands. Technical
302

Report No. TSS-94-100. Florida Department of Environmental ment Station, Forest Service, USDA, Asheville, North Carolina,Protection, Tallahassee, Florida, USA. USA.
USACE 1994. Central and Southern Florida Project Comprehen- Williams K., Ewel K.C., Stumpf R.P., Putz F.E. and Workman T.W.sive Review Study. U.S. Army Corps of Engineers, Jacksonville 1999. Sea-level rise and coastal forest retreat on the west coast ofDistrict, Jacksonville, Florida, USA. Florida, USA. Ecology 80: 2045–2063.
Van Lent T., Snow R.W. and James F.E. 1999. An Examination of Wu Y., Sklar F.H., Gopu K. and Rutchey K. 1996. Fire simulationsthe Modified Water Deliveries Project, the C-111 Project, and the in the Everglades landscape using parallel programming.Experimental Water Deliveries Project: Hydrologic Analyses and Ecological Modeling 93: 113–124.Effects on Endangered Species. South Florida Natural Resources Wu Y., Sklar F.H. and Rutchey K. 1997. Analysis and simulationsCenter, Everglades National Park, Homestead, Florida, USA. of fragmentation patterns in the Everglades. Ecological Applica-
Wade D.D., Ewel J.J. and Hofstetter R. 1980. Fire in South Florida tions 7: 268–276.Ecosystems. General Technical Report SE-17. SE Forest Experi-
303